
Mitochondrial phylogeography of European pond turtles (Emys ...
9
Mitochondrial phylogeography of European pond turtles (Emys orbicularis, Emys trinacris) – an update ∗ Uwe Fritz 1 , Daniela Guicking 2,3 , Hajigholi Kami 4 , Marine Arakelyan 5 , Markus Auer 1 , Dinçer Ayaz 6 , César Ayres Fernández 7 , Andrey G. Bakiev 8 , Antonia Celani 9 , Georg Džuki´ c 10 , Soumia Fahd 11 , Peter Havaš 12 , Ulrich Joger 13 , Viner F. Khabibullin 14 , Lyudmila F. Mazanaeva 15 , Pavel Široký 16 , Sandro Tripepi 9 , Aitor Valdeón Vélez 17 , Guillermo Velo Antón 7 , Michael Wink 2 Abstract. Based on more than 1100 samples of Emys orbicularis and E. trinacris, data on mtDNA diversity and distribution of haplotypes are provided, including for the first time data for Armenia, Georgia, Iran, and the Volga, Ural and Turgay River Basins of Russia and Kazakhstan. Eight mitochondrial lineages comprising 51 individual haplotypes occur in E. orbicularis, a ninth lineage with five haplotypes corresponds to E. trinacris. A high diversity of distinct mtDNA lineages and haplotypes occurs in the south, in the regions where putative glacial refuges were located. More northerly parts of Europe and adjacent Asia, which were recolonized by E. orbicularis in the Holocene, display distinctly less variation; most refuges did not contribute to northern recolonizations. Also in certain southern European lineages a decrease of haplotype diversity is observed with increasing latitude, suggestive of Holocene range expansions on a smaller scale. 1 - Museum of Zoology (Museum für Tierkunde), Natural History State Collections Dresden, A. B. Meyer Build- ing, D-01109 Dresden, Germany e-mail: [email protected] 2 - Institute of Pharmacy and Molecular Biotechnology, Heidelberg University, INF 364, D-69120 Heidelberg, Germany 3 - Current address: University of Kassel, FB1, Systematik und Morphologie der Pflanzen, Heinrich-Plett-Str. 40, D-34132 Kassel, Germany 4 - Department of Biology, Faculty of Sciences, Gorgan University of Agricultural Sciences and Natural Re- sources, Golestan Province, Iran 5 - Department of Biology, Yerevan State University, Alek Maukyan 1, Yerevan, 375025, Armenia 6 - Ege University, Faculty of Science, Department of Biology, Zoology Section, TR-35100 Bornova-Izmir, Turkey 7 - Grupo de Ecoloxía Evolutiva, Departamento de Ecoloxía e Bioloxía Animal, Universidade de Vigo, E.U.E.T. Forestal, Campus Universitario, E-36005 Pontevedra, Spain 8 - Institute of Ecology of the Volga River Basin, Russian Academy of Sciences, Komzina 10, Togliatti, 445003, Russia 9 - Dipartimento di Ecologia, Università degli Studi della Calabria, I-87030 Arcavacata di Rende, Italy 10 - Institute for Biological Research “Siniša Stankovi´ c”, Bulevar Despota Stefana 142, 11060 Belgrade, Serbia 11 - Département de Biologie, Faculté des Sciences, Université Abdelmalek Essaâdi, BP. 2121, Tétouan, Morocco 12 - Fauna Carpatica, Titogradská 18, SK-040 23 Košice, The mitochondrial cytochrome b gene (cyt b) became a frequently used marker for infer- ring phylogeography in reptiles (e.g. Brown and Pestano, 1998; Burbrink et al., 2000; Carranza et al., 2000, 2002; Surget-Groba et al., 2001; Harris et al., 2002; Austin et al., 2003; Pod- nar et al., 2005; Poulakakis et al., 2005; Fritz et al., 2006a) and several studies on Euro- pean pond turtles (Emys orbicularis, Emys tri- nacris) were based on this marker gene. While Lenk et al. (1999) provided a nearly range- wide phylogeography, their study suffered from a patchy sampling for many parts of the range Slovakia 13 - Staatliches Naturhistorisches Museum, Pockelsstr. 10, D-38106 Braunschweig, Germany 14- Faculty of Biology, Bashkir State University, Frunze Street 32, Ufa, 450074, Bashkortostan, Russia 15- Department of Zoology, Dagestan State University, 37a M. Gadyeva st., Makhachkala, 367025, Dagestan, Russia 16 - Department of Biology and Wildlife Diseases, Faculty of Veterinary Hygiene and Ecology, University of Veterinary and Pharmaceutical Sciences, Palackého 1-3, CZ-612 42 Brno, Czech Republic 17 - Departamento de Vertebrados, Aranzadi Society of Sciences, Zorroagagaina 11, E-20014 Donostia-San Sebastián, Gipuzkoa, Spain *This study is dedicated to the late Peter Lenk © Koninklijke Brill NV, Leiden, 2007. Amphibia-Reptilia 28 (2007): 418-426 Also available online - www.brill.nl/amre
Transcript of Mitochondrial phylogeography of European pond turtles (Emys ...

Mitochondrial phylogeography of European pond turtles(Emys orbicularis, Emys trinacris) – an update∗
Uwe Fritz1, Daniela Guicking2,3, Hajigholi Kami4, Marine Arakelyan5, Markus Auer1,
Dinçer Ayaz6, César Ayres Fernández7, Andrey G. Bakiev8, Antonia Celani9, Georg Džukic10,
Soumia Fahd11, Peter Havaš12, Ulrich Joger13, Viner F. Khabibullin14, Lyudmila F. Mazanaeva15,
Pavel Široký16, Sandro Tripepi9, Aitor Valdeón Vélez17, Guillermo Velo Antón7, Michael Wink2
Abstract. Based on more than 1100 samples of Emys orbicularis and E. trinacris, data on mtDNA diversity and distributionof haplotypes are provided, including for the first time data for Armenia, Georgia, Iran, and the Volga, Ural and Turgay RiverBasins of Russia and Kazakhstan. Eight mitochondrial lineages comprising 51 individual haplotypes occur in E. orbicularis,a ninth lineage with five haplotypes corresponds to E. trinacris. A high diversity of distinct mtDNA lineages and haplotypesoccurs in the south, in the regions where putative glacial refuges were located. More northerly parts of Europe and adjacentAsia, which were recolonized by E. orbicularis in the Holocene, display distinctly less variation; most refuges did notcontribute to northern recolonizations. Also in certain southern European lineages a decrease of haplotype diversity isobserved with increasing latitude, suggestive of Holocene range expansions on a smaller scale.
1 - Museum of Zoology (Museum für Tierkunde), NaturalHistory State Collections Dresden, A. B. Meyer Build-ing, D-01109 Dresden, Germanye-mail: [email protected]
2 - Institute of Pharmacy and Molecular Biotechnology,Heidelberg University, INF 364, D-69120 Heidelberg,Germany
3 - Current address: University of Kassel, FB1, Systematikund Morphologie der Pflanzen, Heinrich-Plett-Str. 40,D-34132 Kassel, Germany
4 - Department of Biology, Faculty of Sciences, GorganUniversity of Agricultural Sciences and Natural Re-sources, Golestan Province, Iran
5 - Department of Biology, Yerevan State University, AlekMaukyan 1, Yerevan, 375025, Armenia
6 - Ege University, Faculty of Science, Department ofBiology, Zoology Section, TR-35100 Bornova-Izmir,Turkey
7 - Grupo de Ecoloxía Evolutiva, Departamento deEcoloxía e Bioloxía Animal, Universidade de Vigo,E.U.E.T. Forestal, Campus Universitario, E-36005Pontevedra, Spain
8 - Institute of Ecology of the Volga River Basin, RussianAcademy of Sciences, Komzina 10, Togliatti, 445003,Russia
9 - Dipartimento di Ecologia, Università degli Studi dellaCalabria, I-87030 Arcavacata di Rende, Italy
10 - Institute for Biological Research “Siniša Stankovic”,Bulevar Despota Stefana 142, 11060 Belgrade, Serbia
11 - Département de Biologie, Faculté des Sciences,Université Abdelmalek Essaâdi, BP. 2121, Tétouan,Morocco
12 - Fauna Carpatica, Titogradská 18, SK-040 23 Košice,
The mitochondrial cytochrome b gene (cyt b)became a frequently used marker for infer-ring phylogeography in reptiles (e.g. Brown andPestano, 1998; Burbrink et al., 2000; Carranzaet al., 2000, 2002; Surget-Groba et al., 2001;Harris et al., 2002; Austin et al., 2003; Pod-nar et al., 2005; Poulakakis et al., 2005; Fritzet al., 2006a) and several studies on Euro-pean pond turtles (Emys orbicularis, Emys tri-nacris) were based on this marker gene. WhileLenk et al. (1999) provided a nearly range-wide phylogeography, their study suffered froma patchy sampling for many parts of the range
Slovakia13 - Staatliches Naturhistorisches Museum, Pockelsstr. 10,
D-38106 Braunschweig, Germany14 - Faculty of Biology, Bashkir State University, Frunze
Street 32, Ufa, 450074, Bashkortostan, Russia15 - Department of Zoology, Dagestan State University,
37a M. Gadyeva st., Makhachkala, 367025, Dagestan,Russia
16 - Department of Biology and Wildlife Diseases, Facultyof Veterinary Hygiene and Ecology, University ofVeterinary and Pharmaceutical Sciences, Palackého1-3, CZ-612 42 Brno, Czech Republic
17 - Departamento de Vertebrados, Aranzadi Society ofSciences, Zorroagagaina 11, E-20014 Donostia-SanSebastián, Gipuzkoa, Spain
*This study is dedicated to the late Peter Lenk
© Koninklijke Brill NV, Leiden, 2007. Amphibia-Reptilia 28 (2007): 418-426Also available online - www.brill.nl/amre

Short Notes 419
(North Africa, Iberian and Apennine Peninsu-las, France, East Europe, Kazakhstan, Turkey,Caucasus, Iran and Turkmenistan). Subsequentpapers used a much denser sampling but co-vered only small regions (Fritz et al., 2004,2005a, b, 2006b; Kotenko et al., 2005). Here weprovide a range-wide update on haplotype di-versity and distribution, and present for the firsttime data on pond turtles from Armenia, Geor-gia, Iran, and the Volga, Ural and Turgay RiverBasins of Russia and Kazakhstan. This note isbased on more than 1100 cyt b sequences ofknown-locality samples and specimens of un-known geographic origin and is likely to repre-sent the largest data set ever published for west-ern Palaearctic vertebrates. Our paper is aimedas a starting-point for further research, indicat-ing still badly-sampled regions and suggestingfuture research directions.
Sampling techniques, PCR and sequencingare described in Lenk et al. (1999) and Fritzet al. (2005a). Besides samples from nativewild-caught turtles or known-locality speci-mens from captive breeding projects, samples ofconfiscated, pet trade, or wild-caught allochtho-nous turtles were studied. Sequences were ap-proximately 970-1125 bp long and aligned man-ually for haplotype determination. Haplotypenomenclature follows Lenk et al. (1999) andFritz et al. (2004, 2005a, b, 2006b): Roman nu-merals designate major clades of haplotypes asrevealed by phylogenetic analyses (=mtDNAlineages); within each lineage individual haplo-types are distinguished by consecutive letters.
In addition to the previously identified 48haplotypes in nine lineages (I-IX; Lenk et al.,1999; Fritz et al., 2004, 2005a, b, 2006b) wefound eight new haplotypes, belonging to al-ready known lineages. For many haplotypes de-scribed in earlier studies, distributional data areprovided here for the first time. The geographicorigin of lineage IX (with haplotype IXa only),discovered in a pet trade turtle (Fritz et al.,2004), is still unknown however (table 1).
Based on an alignment of 1031 bp, rela-tionships of haplotypes are illustrated using a
TCS 1.21 parsimony network (Clement et al.,2000) and a MP strict consensus tree rootedwith the closely related Nearctic taxa Actinemysmarmorata and Emydoidea blandingii (PAUP*4.0b10, TBR branch-swapping algorithm, allcharacters with equal weight, stepwise randomaddition of 10 sequences; Swofford, 2002). Forthe ingroup taxa 957 characters were constant,30 were variable but parsimony-uninformativeand 44 were parsimony-informative; for all taxa877 characters were constant, 33 variable char-acters were parsimony-uninformative and 121were parsimony-informative. Maximum likeli-hood estimates for genetic differences of allnine lineages and most haplotypes were de-picted in a recently published ML phylogram(Fritz et al., 2005a); uncorrected p distanceswere reported in the same study.
The network (fig. 1) is flock-like and hap-lotypes cluster into eight major branches. Onebranch splits into lineages I and II (E. o. orbic-ularis and related subspecies); the other eightbranches correspond with lineages III (E. tri-nacris) and lineages IV to IX (remaining E. or-bicularis subspecies). Lineages I and II are in-terconnected over two loops; a further loop oc-curs within lineage IV. Under MP phylogeneticanalysis (fig. 2), monophyly of haplotypes oflineages II, III, IV, V and VII is well-supported,while monophyly for lineage I and VI haplo-types is only weakly supported. Lineages VIIIand IX are represented by only one haplotypeeach that is located outside of all other lineages.Lineage III (E. trinacris) constitutes the sistergroup of a weakly supported clade containinglineages I-II and IV-IX (E. orbicularis); a well-supported clade of lineages I and II is the sistergroup of the remaining lineages IV-IX within E.orbicularis. Bootstrap support for monophylyof the clade comprising lineages IV-IX is below50% however. Within lineage VI, North Africanhaplotypes (VIc, VIf) are paraphyletic with re-spect to European haplotypes (VIa, VIb, VId,VIe).
Regarding geographic distribution and phylo-geography, our new data confirm the findings of
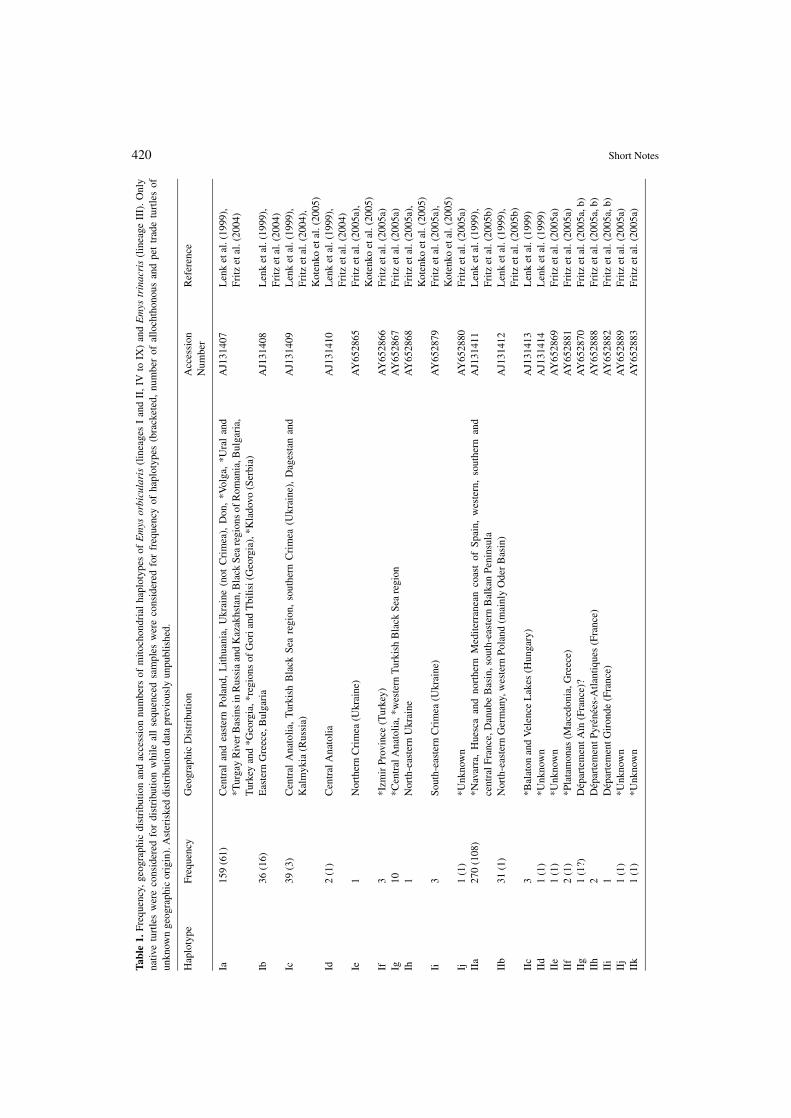
420 Short Notes
Tabl
e1.
Freq
uenc
y,ge
ogra
phic
dist
ribu
tion
and
acce
ssio
nnu
mbe
rsof
mito
chon
dria
lha
plot
ypes
ofE
mys
orbi
cula
ris
(lin
eage
sI
and
II,I
Vto
IX)
and
Em
ystr
inac
ris
(lin
eage
III)
.Onl
yna
tive
turt
les
wer
eco
nsid
ered
for
dist
ribu
tion
whi
leal
lse
quen
ced
sam
ples
wer
eco
nsid
ered
for
freq
uenc
yof
hapl
otyp
es(b
rack
eted
,nu
mbe
rof
allo
chth
onou
san
dpe
ttr
ade
turt
les
ofun
know
nge
ogra
phic
orig
in).
Ast
eris
ked
dist
ribu
tion
data
prev
ious
lyun
publ
ishe
d.
Hap
loty
peFr
eque
ncy
Geo
grap
hic
Dis
trib
utio
nA
cces
sion
Ref
eren
ceN
umbe
r
Ia15
9(6
1)C
entr
alan
dea
ster
nPo
land
,L
ithua
nia,
Ukr
aine
(not
Cri
mea
),D
on,
*Vol
ga,
*Ura
lan
d*T
urga
yR
iver
Bas
ins
inR
ussi
aan
dK
azak
hsta
n,B
lack
Sea
regi
ons
ofR
oman
ia,B
ulga
ria,
Tur
key
and
*Geo
rgia
,*re
gion
sof
Gor
iand
Tbi
lisi(
Geo
rgia
),*K
lado
vo(S
erbi
a)
AJ1
3140
7L
enk
etal
.(19
99),
Fritz
etal
.(20
04)
Ib36
(16)
Eas
tern
Gre
ece,
Bul
gari
aA
J131
408
Len
ket
al.(
1999
),Fr
itzet
al.(
2004
)Ic
39(3
)C
entr
alA
nato
lia,
Tur
kish
Bla
ckSe
are
gion
,so
uthe
rnC
rim
ea(U
krai
ne),
Dag
esta
nan
dK
alm
ykia
(Rus
sia)
AJ1
3140
9L
enk
etal
.(19
99),
Fritz
etal
.(20
04),
Kot
enko
etal
.(20
05)
Id2
(1)
Cen
tral
Ana
tolia
AJ1
3141
0L
enk
etal
.(19
99),
Fritz
etal
.(20
04)
Ie1
Nor
ther
nC
rim
ea(U
krai
ne)
AY
6528
65Fr
itzet
al.(
2005
a),
Kot
enko
etal
.(20
05)
If3
*Izm
irPr
ovin
ce(T
urke
y)A
Y65
2866
Fritz
etal
.(20
05a)
Ig10
*Cen
tral
Ana
tolia
,*w
este
rnT
urki
shB
lack
Sea
regi
onA
Y65
2867
Fritz
etal
.(20
05a)
Ih1
Nor
th-e
aste
rnU
krai
neA
Y65
2868
Fritz
etal
.(20
05a)
,K
oten
koet
al.(
2005
)Ii
3So
uth-
east
ern
Cri
mea
(Ukr
aine
)A
Y65
2879
Fritz
etal
.(20
05a)
,K
oten
koet
al.(
2005
)Ij
1(1
)*U
nkno
wn
AY
6528
80Fr
itzet
al.(
2005
a)II
a27
0(1
08)
*Nav
arra
,H
uesc
aan
dno
rthe
rnM
edite
rran
ean
coas
tof
Spai
n,w
este
rn,
sout
hern
and
cent
ralF
ranc
e,D
anub
eB
asin
,sou
th-e
aste
rnB
alka
nPe
nins
ula
AJ1
3141
1L
enk
etal
.(19
99),
Fritz
etal
.(20
05b)
IIb
31(1
)N
orth
-eas
tern
Ger
man
y,w
este
rnPo
land
(mai
nly
Ode
rB
asin
)A
J131
412
Len
ket
al.(
1999
),Fr
itzet
al.(
2005
b)II
c3
*Bal
aton
and
Vel
ence
Lak
es(H
unga
ry)
AJ1
3141
3L
enk
etal
.(19
99)
IId
1(1
)*U
nkno
wn
AJ1
3141
4L
enk
etal
.(19
99)
IIe
1(1
)*U
nkno
wn
AY
6528
69Fr
itzet
al.(
2005
a)II
f2
(1)
*Pla
tam
onas
(Mac
edon
ia,G
reec
e)A
Y65
2881
Fritz
etal
.(20
05a)
IIg
1(1
?)D
épar
tem
entA
ïn(F
ranc
e)?
AY
6528
70Fr
itzet
al.(
2005
a,b)
IIh
2D
épar
tem
entP
yrén
ées-
Atla
ntiq
ues
(Fra
nce)
AY
6528
88Fr
itzet
al.(
2005
a,b)
IIi
1D
épar
tem
entG
iron
de(F
ranc
e)A
Y65
2882
Fritz
etal
.(20
05a,
b)II
j1
(1)
*Unk
now
nA
Y65
2889
Fritz
etal
.(20
05a)
IIk
1(1
)*U
nkno
wn
AY
6528
83Fr
itzet
al.(
2005
a)

Short Notes 421Ta
ble
1.(C
ontin
ued)
.
Hap
loty
peFr
eque
ncy
Geo
grap
hic
Dis
trib
utio
nA
cces
sion
Ref
eren
ceN
umbe
r
IIIa
12(1
or2)
Sici
ly,p
erha
psC
alab
ria
(Ita
ly)
AJ1
3141
5L
enk
etal
.(19
99),
Fritz
etal
.(20
05a)
IIIb
1(1
)U
nkno
wn
AJ1
3141
6L
enk
etal
.(19
99)
IIIc
34(2
)Si
cily
AY
6528
90Fr
itzet
al.(
2005
a)II
Id1
Sici
lyA
M23
0632
Fritz
etal
.(20
06b)
IIIe
1Si
cily
AM
2306
33Fr
itzet
al.(
2006
b)IV
a14
9(8
8)A
pulia
and
Adr
iatic
coas
tof
Ital
y,w
est
coas
tof
Bal
kan
Peni
nsul
a(n
otC
epha
loni
aan
dPe
lopo
nnes
us),
Cor
fu,E
vvia
(Gre
ece)
AJ1
3141
7L
enk
etal
.(19
99),
Fritz
etal
.(20
05a)
IVb
3C
epha
loni
a(G
reec
e)A
J131
418
Len
ket
al.(
1999
)IV
c7
(2)
Pelo
ponn
esus
(Gre
ece)
AJ1
3141
9L
enk
etal
.(19
99)
IVd
22(8
)So
uthe
rnA
pulia
(Ita
ly),
*Tiv
at(M
onte
negr
o)A
Y65
2871
Fritz
etal
.(20
05a)
IVe
1(1
)U
nkno
wn
AY
6528
84Fr
itzet
al.(
2005
a)IV
f1
(1)
Unk
now
nA
Y65
2872
Fritz
etal
.(20
05a)
IVg
1Pe
lopo
nnes
us(G
reec
e)A
Y65
2873
Fritz
etal
.(20
05a)
IVh
8C
alab
ria,
sout
hern
Apu
lia(I
taly
)A
Y65
2874
Fritz
etal
.(20
05a)
IVi
2So
uthe
rnA
pulia
(Ita
ly)
AY
6528
85Fr
itzet
al.(
2005
a)IV
j2
Sout
hern
Apu
lia(I
taly
)A
Y65
2886
Fritz
etal
.(20
05a)
IVk
1(1
)*U
nkno
wn
AM
1179
37T
his
stud
yV
a18
1(5
2)N
orth
ern
Med
iterr
anea
nco
asto
fSp
ain,
sout
hern
Fran
ce,w
estc
oast
ofA
penn
ine
Peni
n-su
la,C
alab
ria,
Bas
ilica
ta,s
outh
ern
Apu
lia(I
taly
),C
orsi
ca,S
ardi
nia
AJ1
3142
0L
enk
etal
.(19
99),
Fritz
etal
.(20
05a)
Vb
3C
alab
ria,
sout
hern
Apu
lia(I
taly
)A
Y65
2875
Fritz
etal
.(20
05a)
Vc
7*C
alab
ria,
Bas
ilica
ta(I
taly
)A
Y65
2876
Fritz
etal
.(20
05a)
Vd
5*C
alab
ria
(Ita
ly)
AM
2698
87T
his
stud
yV
Ia46
(1or
2)*A
ltoT
rás-
os-M
onte
s(P
ortu
gal)
,*G
alic
ia,
Hue
lva,
*Mad
rid
and
Ciu
dad
Rea
l(S
pain
),M
edite
rran
ean
coas
tof
Spai
n,pe
rhap
sD
épar
tem
entP
yrén
ées-
Atla
ntiq
ues
(Fra
nce)
AJ1
3142
1L
enk
etal
.(19
99),
Fritz
etal
.(20
05b)
VIb
1A
lgar
ve(P
ortu
gal)
AJ1
3142
2L
enk
etal
.(19
99)
VIc
1M
iddl
eA
tlas
(Mor
occo
)A
J131
423
Len
ket
al.(
1999
)V
Id5
Ale
ntej
o(P
ortu
gal)
,*L
eón,
Hue
sca
and
Ebr
oD
elta
(Spa
in)
AJ1
3142
4L
enk
etal
.(19
99)
VIe
2*G
alic
ia(S
pain
)A
Y65
2877
Fritz
etal
.(20
05a)
VIf
3*R
ifM
ts.(
Mor
occo
)A
M26
9888
Thi
sst
udy

422 Short Notes
Tabl
e1.
(Con
tinue
d).
Hap
loty
peFr
eque
ncy
Geo
grap
hic
Dis
trib
utio
nA
cces
sion
Ref
eren
ceN
umbe
r
VII
a27
(2or
3)*A
rmen
ia(A
raxe
sR
iver
),A
zerb
aija
n,*G
ilan,
*Maz
anda
ran
and
*Gol
esta
n(I
ran)
;a
doub
tful
reco
rdex
ists
for
the
regi
onbe
twee
nV
olga
and
Ura
lR
iver
s(n
orth
ern
Cas
pian
Sea,
Kaz
akhs
tan)
,see
text
AJ1
3142
5L
enk
etal
.(19
99)
VII
b1
Unk
now
nA
J131
426
Len
ket
al.(
1999
)V
IIc
2*A
zerb
aija
n,*M
azan
dara
n(I
ran)
AM
2698
89T
his
stud
yV
IId
1*M
azan
dara
n(I
ran)
AM
2698
90T
his
stud
yV
IIe
1*A
zerb
aija
nA
M26
9891
Thi
sst
udy
VII
f2
*Aze
rbai
jan,
*Gol
esta
n(I
ran)
AM
2698
92T
his
stud
yV
IIg
1*G
ilan
(Ira
n)A
M26
9893
Thi
sst
udy
VII
Ia2
Ana
mur
(sou
ther
nT
urke
y)A
Y65
2878
Fritz
etal
.(20
04,2
005a
)IX
a1
(1)
Unk
now
nA
Y65
2887
Fritz
etal
.(20
04,2
005a
)
Tota
l11
07(3
57to
361)

Short Notes 423
Figure 1. Parsimony network (TCS, spring tree) of all 56known mtDNA haplotypes of Emys orbicularis (haplotypesof lineages I-II, IV-IX) and Emys trinacris (haplotypes oflineage III). Branch lengths correspond to inferred numberof nucleotide changes along each branch; open circles anddots, identified or hypothesized haplotypes, respectively.Each line between circles or dots indicates one substitution;circle size corresponds to approximate frequency of haplo-types (table 1); size classes: 1, 2-9, 10-25, 26-50, 51-150,151-200, >200 recorded haplotypes.
Lenk et al. (1999) and Fritz et al. (2005a) in thata high diversity of distinct mtDNA lineages andhaplotypes occurs in southern regions where pu-tative glacial refuges were located (fig. 3, ta-ble 1). With the advent of Holocene warming,more northerly parts of Europe and adjacentAsia were rapidly recolonized by E. orbicu-laris from few refuges in the Balkans and thenorthern Black Sea/northern Caucasus region,resulting in the respective mtDNA lineages indecreasing haplotype diversity with increasingdistance to the refuge (long distance dispersalmodel of Hewitt, 1996); most refuges did notcontribute to northern recolonizations.
With exception of a doubtful record of haplo-type VIIa, the northernmost parts of the range ofE. orbicularis are occupied only by three hap-lotypes (Ia, IIa and IIb), while lineages I andII exhibit a much higher diversity in the south.The same pattern, suggestive of Holocene range
Figure 2. Strict consensus of six equally parsimonioustrees of all Emys haplotypes using sequences of Actinemysmarmorata (accession numbers AJ131430, U81344) andEmydoidea blandingii (AF258869, AJ131432) as outgroup(CI = 0.7585, RI = 0.9120, RC = 0.6917; 207 steps). Num-bers above nodes are bootstrap values (1000 resamplings)greater than 50. The six equally parsimonious trees differonly in the branching pattern of lineage IV haplotypes.
expansions on a smaller scale, is observed inlineages IV and V with several endemic haplo-types in the southernmost parts of their ranges;northern portions harbour only haplotypes IVaor Va. A previous record of haplotype VIIa forthe northern Caspian Sea region (Lenk et al.,1999) should be treated with care. It was basedon a turtle kept in the zoological garden of Al-maty (Kazakhstan), and we cannot exclude lo-

424 Short Notes
Figure 3. Geographic distribution of mtDNA lineages in Emys orbicularis (I-II, IV-IX) and Emys trinacris (III). Neighbouringlocalities combined; overlapping symbols indicate syntopic occurrence of distinct lineages; question marks, doubtful recordsfor lineage III in Calabria (southern Italy) and lineage VII in the northern Caspian region. Evidently allochthonous specimensnot considered.
cality confusion. Otherwise, lineage VII occursfar away, in the eastern central Caucasus andalong the south coast of the Caspian Sea (fig. 3,table 1).
The wide distribution of lineage I haplotypesaround the Black Sea suggests that the entireBlack Sea region served as glacial refuge. Onthe other hand, the localized distribution of en-demic lineage I haplotypes (table 1) indicatesthat distinct microrefuges existed there. An un-expected finding was haplotype Ia in easternGeorgia (regions of Gori and Tbilisi: one recordeach from Kareli, Gldanula River and Udabno).These sites are situated in the Kura River sys-tem that drains into the Caspian Sea. In the sameriver system occurs another mitochondrial lin-eage (VII) farther east, suggesting that lineagesI and VII meet somewhere in the central Cauca-sus. This situation is echoed in the phylogeogra-phy of Testudo graeca. Also in this species twodistinct mitochondrial lineages meet or occur in
closest neighbourhood in the central Caucasus(Fritz et al., 2007).
Besides the wide-spread haplotype IIa, themost differentiated haplotype of lineage II(IIf) occurs on the south-eastern Balkan Penin-sula (table 1), suggesting the refuge for lin-eage II was located there. From the south-eastern Balkans, lineage II most probably spreadalong the valleys of the Vardar and JužnaMorava Rivers northward to the Danube catch-ment basin from where central and western Eu-rope (Fritz et al., 2005b) and, via the MoravianGate, the Oder River Basin (Germany, Poland)were reached.
As for lineage IV, displaying a classic circum-Adriatic distribution, two distinct refuges ex-isted in southern Italy and the southernmostBalkan Peninsula. The lack of the common hap-lotype IVa, distributed in Italy, Istria, Dalma-tia, Corfu and Evvia, and the occurrence of en-demic haplotypes in the Peloponnesus and on

Short Notes 425
Cephalonia (IVb, IVc, IVg) provide evidencefor the colonization of the west coast of theBalkans, Corfu and Evvia from southern Italyand not from the geographically closer southernBalkanic refuge (Fritz et al., 2005a).
Like in Mauremys leprosa (Fritz et al.,2006a), mountain chains constitute major bio-geographic barriers for mtDNA lineages in E.orbicularis (fig. 3). The Pyrenees separate theIbero-Maghrebinian lineage VI from lineages IIand V in the north, and the Apennines sepa-rate lineages IV and V in Italy. In the BalkanPeninsula, the Dinarid and Pindos Mts. are abarrier between lineage IV along the west coastand lineages I and II in the east, and south ofthe Alps occur lineages IV and V while in thenorth lineage II is distributed. In East Europethe Carpathians act as barrier between lineageII in the south and lineage I in the north; far-ther eastward the Greater Caucasus separateslineages I and VII and the Taurus Mts. (Turkey)divide lineages I and VIII. Syntopic occurrencesof distinct mtDNA lineages, most probably asresult of Holocene range expansions, are con-fined to narrow contact zones in close proximityof these mountain chains in the northern IberianPeninsula, France and the southern Apennineand Balkan Peninsulas. Obviously, marshlandsalong sea coasts and river courses were used ascolonization corridors during range expansions,enabling development of contact zones.
The basal position of the North African hap-lotypes VIc and VIf in the MP tree and inthe network branch of lineage VI (figs 1, 2)suggests that the Iberian haplotypes VIa, VIb,VId and VIe are derived from North Africanhaplotypes. This implies a complicated colo-nization history, from Europe to North Africaand back to Europe, as recently proposed forthe ribbed newt Pleurodeles waltl (Veith etal., 2004). Denser sampling is still needed inNorth Africa however, especially in Algeria andTunisia. Some Moroccan amphibians and rep-tiles that occur in habitats resembling those of E.orbicularis (Pleurodeles waltl, Mauremys l. lep-rosa) are replaced in eastern Algeria and Tunisia
by other taxa (P. nebulosus, P. poireti, M. l. sa-harica, Carranza and Arnold, 2003; Carranzaand Wade, 2004; Veith et al., 2004; Fritz et al.,2006a; distinct mitochondrial lineages of Natrixmaura, Guicking et al., 2006), suggestive of asimilar pattern in E. orbicularis. Further sam-pling is also needed in Turkey, Turkmenistan, inthe Caucasus and in the northern Black Sea re-gion to reveal haplotype diversity and potentialcontact zones there; one of these regions mustharbour lineage IX.
Besides mtDNA haplotyping, future researchshould focus on gene flow along contact zonesof distinct mitochondrial lineages. A promisingapproach will be using microsatellites as mark-ers.
Acknowledgements. Thanks for lab work and curation ofsamples go to A. Hundsdörfer, Ch. Kehlmaier, A. Müller,and H. Sauer-Gürth. Blood or tissue samples were pro-vided by B. Andreas, M. Barata, J. Budó, H. Bringsøe, X.Capalleras, R. Castilho, J.-M. Ducotterd, O. Homeier, M.Korn, M. Kuprian, M. Meeske, K.D. Milto, S. Mitrus, D.Mosimann, M. Nembrini, N.L. Orlov, J.F. Parham, R. Pod-loucky, N. Schneeweiß, J. Schwarzendrube, P. Segurado,R. Stampfer, D. Tarknishvili, and R. Wicker; M. Fischerand J.-M. Lange assisted with the production of the map.Georg Džukic’s work for this study was supported by theSerbian Ministry of Science and Environmental Protection(“Patterns of amphibian and reptile diversity on the BalkanPeninsula”, Grant 143052).
References
Austin, J.J., Arnold, E.N., Bour, R. (2003): Was there a sec-ond adaptive radiation of giant tortoises in the IndianOcean? Using mitochondrial DNA to investigate spe-ciation and biogeography of Aldabrachelys. MolecularEcology 12: 1415-1424.
Brown, R.P., Pestano, J. (1998): Phylogeography of skinks(Chalcides) in the Canary Islands inferred from mito-chondrial DNA sequences. Molecular Ecology 7: 1183-1191.
Burbrink, F.T., Lawson, R., Slowinski, J.B. (2000): Mito-chondrial DNA phylogeography of the polytypic NorthAmerican rat snake (Elaphe obsoleta): a critique of thesubspecies concept. Evolution 54: 2107-2118.
Carranza, S., Arnold, E.N. (2003): History of West Mediter-ranean newts, Pleurodeles (Amphibia, Salamandridae)inferred from old and recent DNA sequences. Systemat-ics and Biodiversity 1: 327-337.

426 Short Notes
Carranza, S., Wade, E. (2004): Taxonomic revision ofAlgero-Tunisian Pleurodeles (Caudata: Salamandridae)using molecular and morphological data. Revalidationof the taxon Pleurodeles nebulosus (Guichenot, 1850).Zootaxa 488: 1-24.
Carranza, S., Arnold, E.N., Mateo, J.A., López-Jurado,L.F. (2000): Long-distance colonization and radiation ingekkonid lizards, Tarentola (Reptilia: Gekkonidae), re-vealed by mitochondrial DNA sequences. Proceedingsof the Royal Society B 267: 637-649.
Carranza, S., Arnold, E.N., Mateo, J.A., Geniez, P. (2002):Relationships and evolution of the North Africangeckos, Geckonia and Tarentola (Reptilia: Gekkonidae),based on mitochondrial and nuclear DNA sequences.Molecular Phylogenetics and Evolution 23: 244-256.
Clement, M., Posada, D., Crandall, K.A. (2000): TCS: acomputer program to estimate gene genealogies. Molec-ular Ecology 9: 1657-1660.
Fritz, U., Guicking, D., Lenk, P., Joger, U., Wink, M.(2004): When turtle distribution tells European history:mtDNA haplotypes of Emys orbicularis reflect in Ger-many former division by the Iron Curtain. Biologia 59(Supplement 14): 19-25.
Fritz, U., Fattizzo, T., Guicking, D., Tripepi, S., Pennisi,M.G., Lenk, P., Joger, U., Wink, M. (2005a): A newcryptic species of pond turtle from southern Italy, thehottest spot in the range of the genus Emys. ZoologicaScripta 34: 351-371.
Fritz, U., Cadi, A., Cheylan, M., Coïc, C., Détaint, M.,Olivier, A., Rosecchi, E., Guicking, D., Lenk, P., Joger,U., Wink, M. (2005b): Distribution of mtDNA haplo-types (cyt b) of Emys orbicularis in France and implica-tions for postglacial recolonization. Amphibia-Reptilia26: 231-238 [Erratum: Amphibia-Reptilia 27 (2006):157].
Fritz, U., Barata, M., Busack, S.D., Fritzsch, G., Castilho,R. (2006a): Impact of mountain chains, sea straits andperipheral populations on genetic and taxonomic struc-ture of a freshwater turtle, Mauremys leprosa. ZoologicaScripta 35: 97-108.
Fritz, U., d’Angelo, S., Pennisi, M.G., Lo Valvo, M.(2006b): Variation of Sicilian pond turtles, Emys tri-nacris – What makes a species cryptic? Amphibia-Reptilia 27: 513-529.
Fritz, U., Hundsdörfer, A.K., Široký, P., Auer, M., Kami,H., Lehmann, J., Mazanaeva, L.F., Türkozan, O., Wink,M. (2007): Phenotypic plasticity leads to incongruencebetween morphology-based taxonomy and genetic dif-ferentiation in western Palaearctic tortoises (Testudograeca complex; Testudines, Testudinidae). Amphibia-Reptilia 28: 97-121.
Guicking, D., Lawson, R., Joger, U., Wink, M. (2006):Evolution and phylogeny of the genus Natrix. BiologicalJournal of the Linnean Society 87: 127-143.
Harris, D.J., Carranza, S., Arnold, E.N., Pinho, C., Ferrand,N. (2002): Complex biogeographical distribution of ge-netic variation within Podarcis wall lizards across theStrait of Gibraltar. Journal of Biogeography 29: 1257-1262.
Hewitt, G.M. (1996): Some genetic consequences of iceages, and their role in divergence and speciation. Bio-logical Journal of the Linnean Society 58: 247-276.
Kotenko, T., Zinenko, O., Guicking, D., Sauer-Gürth, H.,Wink, M., Fritz, U. (2005): First data on the geographicvariation of Emys orbicularis in Ukraine: mtDNA haplo-types, coloration, and size. In: Herpetologica Petropoli-tana. Proceedings of the 12th Ordinary General Meetingof the Societas Herpetologica Europaea, August 12-16,2003, St. Petersburg. Ananjeva, N., Tsinenko, O., Eds.Russian Journal of Herpetology 12 (Supplement): 43-46.
Lenk, P., Fritz, U., Joger, U., Wink, M. (1999): Mito-chondrial phylogeography of the European pond turtle,Emys orbicularis (Linnaeus, 1758). Molecular Ecology8: 1911-1922.
Podnar, M., Mayer, W., Tvrtkovic, N. (2005): Phylogeogra-phy of the Italian wall lizard, Podarcis sicula, as revealedby mitochondrial DNA sequences. Molecular Ecology14: 575-588.
Poulakakis, N., Lymberakis, P., Valakos, E., Pafilis, P.,Zouros, E., Mylonas, M. (2005): Phylogeography ofBalkan wall lizard (Podarcis taurica) and its relativesinferred from mitochondrial DNA sequences. MolecularEcology 14: 2433-2443.
Surget-Groba, Y., Heulin, B., Guillaume, C.-P., Thorpe,R.S., Kupriyanova, L., Vogrin, N., Maslak, R., Maz-zotti, S., Venczel, M., Ghira, I., Odierna, G., Leontyeva,O., Monney, J.C., Smith, N. (2001): Intraspecific phy-logeography of Lacerta vivipara and the evolution ofviviparity. Molecular Phylogenetics and Evolution 18:449-459.
Swofford, D.L. (2002): PAUP*. Phylogenetic Analysis Us-ing Parsimony (*and Other Methods), Version 4.0b10.Sunderland, Massachusetts, Sinauer Associates.
Veith, M., Mayer, C., Samraoui, B., Donaire Barroso, B.,Bogaerts, S. (2004): From Europe to Africa and viceversa: evidence for multiple intercontinental dispersalin ribbed salamanders (genus Pleurodeles). Journal ofBiogeography 31: 159-171.
Received: May 29, 2006. Accepted: November 17, 2006.


















