
Market Access of Papua New Guinea Bananas (Musa spp.)...
187
Market Access of Papua New Guinea Bananas (Musa spp.) with Particular Respect to Banana Fly (Bactrocera musae (Tryon)) (Diptera: Tephritidae) Amanda Mararuai B.Sc Agriculture, Graduate Diploma in Applied Science A thesis submitted in fulfilment of the requirements for the degree of Doctor of Philosophy School of Natural Resource Sciences Queensland University of Technology Brisbane Australia April 2010
Transcript of Market Access of Papua New Guinea Bananas (Musa spp.)...

Market Access of Papua New Guinea Bananas (Musa spp.)
with Particular Respect to Banana Fly (Bactrocera
musae (Tryon)) (Diptera: Tephritidae)
Amanda Mararuai
B.Sc Agriculture, Graduate Diploma in Applied Science
A thesis submitted in fulfilment of the requirements for the degree of
Doctor of Philosophy
School of Natural Resource Sciences
Queensland University of Technology
Brisbane Australia
April 2010

ii
Keywords
Bactrocera musae, banana fly, bananas, biosecurity, host availability, invasion biology,
invasive, market access, Musa spp., novel environment, Papua New Guinea, pest risk
analysis, population distribution

iii
Abstract
International market access for fresh commodities is regulated by international
accepted phytosanitary guidelines, the objectives of which are to reduce the biosecurity
risk of plant pest and disease movement. Papua New Guinea (PNG) has identified
banana as a potential export crop and to help meet international market access
requirements, this thesis provides information for the development of a pest risk
analysis (PRA) for PNG banana fruit. The PRA is a three step process which first
identifies the pests associated with a particular commodity or pathway, then assesses
the risk associated with those pests, and finally identifies risk management options for
those pests if required.
As the first step of the PRA process, I collated a definitive list on the organisms
associated with the banana plant in PNG using formal literature, structured interviews
with local experts, grey literature and unpublished file material held in PNG field
research stations. I identified 112 organisms (invertebrates, vertebrate, pathogens and
weeds) associated with banana in PNG, but only 14 of these were reported as
commonly requiring management. For these 14 I present detailed information
summaries on their known biology and pest impact.
A major finding of the review was that of the 14 identified key pests, some research
information occurs for 13. The single exception for which information was found to be
lacking was Bactrocera musae (Tryon), the banana fly. The lack of information for this
widely reported ‘major pest on PNG bananas’ would hinder the development of a PNG
banana fruit PRA. For this reason the remainder of the thesis focused on this organism,
particularly with respect to generation of information required by the PRA process.
Utilising an existing, but previously unanalysed fruit fly trapping database for PNG, I
carried out a Geographic Information System analysis of the distribution and
abundance of banana in four major regions of PNG. This information is required for a
PRA to determine if banana fruit grown in different parts of the country are at different
risks from the fly. Results showed that the fly was widespread in all cropping regions
and that temperature and rainfall were not significantly correlated with banana fly
abundance. Abundance of the fly was significantly correlated (albeit weakly) with host

iv
availability. The same analysis was done with four other PNG pest fruit flies and their
responses to the environmental factors differed to banana fly and each other. This
implies that subsequent PRA analyses for other PNG fresh commodities will need to
investigate the risk of each of these flies independently.
To quantify the damage to banana fruit caused by banana fly in PNG, local surveys and
one national survey of banana fruit infestation were carried out. Contrary to
expectations, infestation was found to be very low, particularly in the widely grown
commercial cultivar, Cavendish. Infestation of Cavendish fingers was only 0.41% in a
structured, national survey of over 2 700 banana fingers. Follow up laboratory studies
showed that fingers of Cavendish, and another commercial variety Lady-finger, are
very poor hosts for B. musae, with very low host selection rates by female flies and
very poor immature survival.
An analysis of a recent (within last decade) incursion of B. musae into the Gazelle
Peninsula of East New Britain Province, PNG, provided the final set of B. musae data.
Surveys of the fly on the peninsular showed that establishment and spread of the fly in
the novel environment was very rapid and thus the fly should be regarded as being of
high biosecurity concern, at least in tropical areas. Supporting the earlier impact
studies, however, banana fly has not become a significant banana fruit problem on the
Gazelle, despite bananas being the primary starch staple of the region.
The results of the research chapters are combined in the final Discussion in the form of
a B. musae focused PRA for PNG banana fruit. Putting the thesis in a broader context,
the Discussion also deals with the apparent discrepancy between high local abundance
of banana fly and very low infestation rates. This discussion focuses on host utilisation
patterns of specialist herbivores and suggests that local pest abundance, as determined
by trapping or monitoring, need not be good surrogate for crop damage, despite this
linkage being implicit in a number of international phytosanitary protocols.

v
Table of Contents
Keywords .................................................................................................................................................. ii
Abstract .................................................................................................................................................... iii
List of Tables ......................................................................................................................................... viii
List of Figures ........................................................................................................................................... x
Supplementary material ....................................................................................................................... xiv
Statement of original authorship .......................................................................................................... xv
Acknowledgements ................................................................................................................................ xvi
Chapter 1. Literature review ................................................................................................................... 1
1.1 Introduction ..................................................................................................................................... 1 1.1.1 Agriculture in Papua New Guinea ........................................................................................... 1 1.1.2 Limitations of market access ................................................................................................... 2 1.1.3 Invasive species and export systems ........................................................................................ 3 1.1.4 Overview of chapter ................................................................................................................ 4
1.2 Invasion biology and its role in biosecurity .................................................................................... 5
1.3 Fruit flies as invasive organisms ..................................................................................................... 8
1.4 The study system: banana and banana fly ..................................................................................... 10
1.5 Thesis structure ............................................................................................................................. 13
Chapter 2. Review of banana (Musa spp.) pests in Papua New Guinea ............................................ 17
2.1 Introduction ................................................................................................................................... 17
2.2 Materials & Methodology ............................................................................................................. 19
2.3 Results ........................................................................................................................................... 22 2.3.1 Pest List ................................................................................................................................. 22 2.3.2 Pest Summaries ...................................................................................................................... 27 2.3.3 Pathogens - Bacteria .............................................................................................................. 28
2.3.3.1 Erwinia chrysanthemi Burkholder et al. ................................................................. 28 2.3.4 Pathogens - Fungi .................................................................................................................. 29
2.3.4.1 Cordana musae (Zimm.) ......................................................................................... 29 2.3.4.2 Mycosphaerella fijiensis Morelet ............................................................................ 31 2.3.4.3 Phyllachora musicola Booth & Shaw ..................................................................... 33 2.3.4.4 Ramichloridium musae de Hoog ............................................................................ 34
2.3.5 Insects & Mites - Coleoptera ................................................................................................. 34 2.3.5.1 Cosmopolites sordidus (Germar) ............................................................................ 34 2.3.5.2 Papuana species ...................................................................................................... 35 2.3.5.3 Rhyparida sobrina Bryant ....................................................................................... 37 2.3.5.4 Scapanes australis grossepunctatus Sternberg ........................................................ 38
2.3.6 Insects & Mites - Diptera ....................................................................................................... 39 2.3.6.1 Bactrocera musae (Tryon)....................................................................................... 39
2.3.7 Insects & Mites - Lepidoptera ............................................................................................... 41 2.3.7.1 Erionata thrax (L.) ................................................................................................... 41 2.3.7.2 Nacoleia octasema (Meyrick) ................................................................................. 43
2.3.8 Nematodes ............................................................................................................................. 44 2.3.8.1 Pratylenchus coffeae (Zimmermann) ...................................................................... 44

vi
2.3.8.2 Radopholus similis Thorne ..................................................................................... 46
2.4 Discussion ..................................................................................................................................... 47
2.5 Thesis Progress ............................................................................................................................. 49
Chapter 3. Distribution and abundance of five economically important fruit fly species in Papua New Guinea ............................................................................................................................................. 51
3.1 Introduction ................................................................................................................................... 51
3.2 Materials and methods .................................................................................................................. 53 3.2.1 Trapping ................................................................................................................................ 53 3.2.2 Databases and analysis .......................................................................................................... 56 3.2.3 Fly species ............................................................................................................................. 57
3.3 Results ........................................................................................................................................... 59 3.3.1 General Patterns ..................................................................................................................... 59 3.3.2 Bactrocera bryoniae (Tryon) .................................................................................................. 62 3.3.3 Bactrocera cucurbitae (Coquillett) ......................................................................................... 67 3.3.4 Bactrocera frauenfeldi (Schiner) ............................................................................................ 71 3.3.5 Bactrocera musae (Tryon) ..................................................................................................... 75 3.3.6 Bactrocera umbrosa (Fabricius) ............................................................................................. 80
3.4 Discussion ..................................................................................................................................... 85
3.5 Thesis Progress ............................................................................................................................. 87
Chapter 4. Infestation of bananas by Bactrocera musae (Tryon) in Papua New Guinea ................. 88
4.1 Introduction ................................................................................................................................... 88
4.2 Materials and methods .................................................................................................................. 90 4.2.1 Differences in varietal susceptibility ..................................................................................... 90 4.2.2 National Cavendish survey .................................................................................................... 92
4.3 Results ........................................................................................................................................... 96 4.3.1 Differences in varietal susceptibility ..................................................................................... 96 4.3.2 National Cavendish survey .................................................................................................. 102
4.4 Discussion ................................................................................................................................... 103
4.5 Thesis progress ............................................................................................................................ 104
Chapter 5. Host selection and utilisation by Bactrocera musae (Tryon) on two banana varieties at different ripening stages ...................................................................................................................... 106
5.1 Introduction ................................................................................................................................. 106
5.2 Materials and Methods ................................................................................................................ 109 5.2.1 Adult host choice and utilisation ......................................................................................... 109 5.2.2 Larval host utilisation .......................................................................................................... 110 5.2.3 Host use of green banana ..................................................................................................... 110 5.2.4 Host data .............................................................................................................................. 111 5.2.5 Analysis ............................................................................................................................... 111
5.3 Results ......................................................................................................................................... 112 5.3.1 Adult Choice ........................................................................................................................ 112 5.3.2 Adult no choice .................................................................................................................... 112 5.3.3 Larval performance.............................................................................................................. 114 5.3.4 Green Cavendish as a host of Bactrocera musae ................................................................. 116 5.3.5 Banana attributes ................................................................................................................. 116
5.4 Discussion .................................................................................................................................. 119
5.5 Thesis progress ............................................................................................................................ 121

vii
Chapter 6. Bactrocera musae (Tryon) in a novel environment: banana fly as an invasive organism on the Gazelle Peninsula, Papua New Guinea ................................................................................... 122
6.1 Introduction ................................................................................................................................. 122
6.2 Materials and Methods ................................................................................................................ 124 6.2.1 Distribution and spread of Bactrocera musae (Tryon) on the Gazelle Peninsula................. 124 6.2.2 Population abundance and phenology ................................................................................. 126 6.2.3 Impact Studies ..................................................................................................................... 127
Market Surveys (2000-2001) ................................................................................................... 127 Bagging trial (2001-2003) ........................................................................................................ 127 Current damage ........................................................................................................................ 127
6.3 Results ......................................................................................................................................... 128 6.3.1 Distribution and spread of Bactrocera musae (Tryon) on the Gazelle Peninsula................. 128 6.3.2 Population abundance and phenology ................................................................................. 130 6.3.3 Impact studies ...................................................................................................................... 131
6.3.3.1 Fruit rearing (2000-2001) ..................................................................................... 131 6.3.3.2 Bagging trial (2001-2002) .................................................................................... 131 6.3.3.3 Current status ........................................................................................................ 133
6.4 Discussion ................................................................................................................................... 134
Chapter 7. Discussion ........................................................................................................................... 136
7.1 Thesis summary ........................................................................................................................... 136 7.1.1 Introduction ......................................................................................................................... 136 7.1.2 Summary .............................................................................................................................. 136
7.2 PRA for PNG Banana .................................................................................................................. 137 7.2.1 Definitive statement of IPPC PRA process ......................................................................... 137 7.2.2 Summary on PNG banana PRA ........................................................................................... 138 7.2.3 Detailed PRA response for banana fly ................................................................................. 139
7.3 Implications of thesis for wider fruit fly market access issues ..................................................... 141 7.3.1 Trap abundance and host use by fruit flies .......................................................................... 142 7.3.2 How does this relate to Market access? ............................................................................... 144
References ............................................................................................................................................. 145
Appendices ............................................................................................................................................ 169
Appendix 1 Fruit flies in Papua New Guinea ................................................................................... 169
Appendix 2 Distribution and biogeography of Bactrocera and Dacus species (Diptera: Tephritidae) in Papua New Guinea ........................................................................................................................ 182
Appendix 3 Introduction and distribution of Bactrocera musae (Tryon) (Diptera: Tephritidae) in East New Britain, Papua New Guinea .............................................................................................. 192

viii
List of Tables
Table 1.1 Host plants for Bactrocera musae (Tryon) in Australia (Distribution: Torres Strait islands and northeast Queensland, as far north as Townsville). Taken from - The Distribution and Host Plants of Fruit Flies (Diptera: Tephritidae) in Australia (Hancock et al., 2000). ................................................................................................. 12
Table 2.1 Names and history on professional background of formally interviewed agriculture and quarantine field officers ......................................................................................... 20
Table 2.2 Pathogens, arthropods, nematodes and weeds reported associated with banana in Papua New Guinea ....................................................................................................... 23
Table 3.1 Location and number of fruit fly trap sites in four study areas in Papua New Guinea in relation to altitude (m.a.s.l) and annual rainfall (mm) levels used in the Papua New Guinea Resource Information System (PNGRIS) ........................................................ 55
Table 3.2 The monthly trap catch (June 1998 – September 2001) of five economically important fruit fly Bactrocera species in four study areas in Papua New Guinea ....... 60
Table 3.3 Linear regression analysis of the influence of: (i) altitude or (ii) rainfall on the abundance of five Bactrocera species within and across four study areas in Papua New Guinea, or (iii) banana in local cropping systems for B. musae only. Results are R2 values (and probability values in brackets). Note: Analysis not applicable (NA) for altitude in Central because 20 of 21 traps are located at one altitude level, nor for B.musae on the Gazelle due to inconsistent and sporadic sampling (* = P≤0.05). ...... 61
Table 3.4 Multiple regression analysis of the influence of both altitude and rainfall on fly distribution and abundance (P≤0.05) (* = P≤0.05). ..................................................... 61
Table 4.1 Sampling details for the national Cavendish survey ................................................ 94
Table 4.2 Damage assessment records of miscellaneous host records of banana varieties sampled between 1998 and 2000 in Western Highlands, Central, East New Britain, Madang, and Morobe provinces in Papua New Guinea ............................................... 97
Table 4.3 continued…Damage assessment records of miscellaneous host records of banana varieties sampled between 1998 and 2000 in Western Highlands, Central, East New Britain, Madang, and Morobe provinces in Papua New Guinea .................................. 98
Table 4.4 continued…Damage assessment records of miscellaneous host records of banana varieties sampled between 1998 and 2000 in Western Highlands, Central, East New Britain, Madang, and Morobe provinces in Papua New Guinea .................................. 99
Table 4.5 Infestation of Cavendish banana samples for 22 localities in Papua New Guinea. Each sample consisted of about 30 individual fingers collected at mature green stage of ripeness ....................................................................................................................... 103
Table 5.1 Summary two-way ANOVA output table for Cavendish and Ladyfinger fruit attributes at three ripening stages ............................................................................... 117
Table 5.2 continued…Summary two-way ANOVA output table for Cavendish and Ladyfinger fruit attributes at three ripening stages ....................................................................... 118
Table 6.1 Tephritid fruit flies reared from bananas purchased from markets (March to June 2000) or collected from gardens (May to June 2001) on the Gazelle Peninsula, East New Britain, Papua New Guinea ............................................................................... 132
Table 6.2 Infestation of mature green Cavendish banana by Bactrocera musae (Tryon) at four locations on the Gazelle Peninsula, East New Britain, Papua New Guinea, in 2007 . 133

ix
Table 7.1 Checklist of information available and necessary for carrying out a pest risk analysis of PNG bananas; a pathway initiated analysis ............................................................ 140

x
List of Figures
Figure 1.1 (A) The multi-step process of non-native species invasion (Lockwood et al., 2005); and (B) Transitions that non-invasive species must overcome to continue in the invasion process (Kolar & Lodge, 2001) ....................................................................... 6
Figure 1.2 Conceptual model of the process that invasive species go through in their movement from endemic to non-endemic areas ............................................................. 7
Figure 1.3 Pest Risk Analysis process ...................................................................................... 13
Figure 2.1 Report generated by Papua New Guinea Pest List Database in December 2006 of the pests recorded on banana and plantains in Papua New Guinea .............................. 18
Figure 2.2 (A) Fallen banana, cause may be due to rotting of basal pseudostem (sometimes termed ‘tip-over’ by field officers in PNG) (Source: CTAHR Hawaii); and (B) Rotting xylem vessels referred to as internal pseudostem necrosis (Source: CTAHR Hawaii www.ctahr.hawaii.edu/nelsons/banana) ....................................................................... 28
Figure 2.3 Cordana leaf spot on banana leaves (Source: CTAHR Hawaii web site: www.ctahr.hawaii.edu/nelsons/banana) ....................................................................... 29
Figure 2.4 (A) Black Sigatoka on banana leaves (Source: DAFF Australia – AQIS); and (B) Leaf infested with black Sigatoka has yellow transition zones between infected and green uninfected leaf area (Source: CTAHR Hawaii www.ctahr.hawaii.edu/nelsons/banana/) ...................................................................... 31
Figure 2.5 Black-cross on underside of banana leaf of local cultivar. Photo taken at Chanel College, Kokopo, ENB (Source: A. Mararuai) ............................................................ 33
Figure 2.6 (A) Lifecycle and damage caused by banana weevil borer (Source: Cook Islands Biodiversity Database); and (B) Adult banana weevil borer (Source: G McCormack. Cook Islands Biodiversity Database- http://entnem.ufl.edu/creatures/fruit/borers/banana_root_borer.htm) .......................... 34
Figure 2.7 (A) Taro beetle burrows in damaged corm of banana sword sucker (Source: A Mararuai); (B) Adult Papuana woodlarkiana Montrouzier (Source: A Carmichael, PaDIL); and (C) Larvae, pupae and adult taro beetle collected at LAES Keravat, 2005 (Source: A. Mararuai) .................................................................................................. 36
Figure 2.8 Sketch of Rhyparidella sobrina (Bryant) (Source: Gressit (1974)) ........................ 37
Figure 2.9 (A) Pseudostem bore-hole caused by Scapanes australis grossepunctatus (Source: A. Mararuai); (B) S grossepunctatus boring into banana pseudostem (Source: A. Mararuai); and (C) Banana bunch emerging through a hole made by S. grossepunctatus in banana pseudostem (Source: A Mararuai) .................................... 38
Figure 2.10 (A) Adult Bactrocera musae (Source SPC PaciFly); (B) Fruit fly larvae feeding tracks in ripe Cavendish banana variety (Bundun, Morobe Province) (Source: A. Mararuai); and (C) Fruit fly larvae feeding tracks in mature green Cavendish (Kaiapit, Morobe Province) (Source: A. Mararuai) .................................................................... 39
Figure 2.11 (A) Adult Erionota thrax butterfly (Source: K Walker, PaDIL); (B) Caterpillar feeding on leaf (Gazelle Peninsula, ENB) (Source: A Mararuai); (C) Leaves of banana rolled up by banana skipper (Bubia, Morobe province) (Source: A Mararuai); and (D) Pupae in leaf roll (Gazelle, ENB) (Source: A Mararuai) ............................................. 41
Figure 2.12 (A) Adult Nacoleia octasema (NAIC Kilakila, Port Moresby) (Source: A Mararuai); and (B) N. octasema damage on banana fingers (Source: A Mararuai) ..... 43

xi
Figure 2.13 (A) Female nematode (Source: http://nematode.unl.edu): and (B) Symptoms of Pratylenchus coffeae feeding on banana root (Source: CAB Crop Protection Compendium Module 1) ............................................................................................... 45
Figure 2.14 (A) Illustration of burrowing nematode (Source: http://plpnemweb.ucdavis.edu/Nemaplex); (B) Damage to banana roots caused by the burrowing nematode (Source: http://www.ctahr.hawaii.edu); and (C) Toppled banana (black head) (Source: http://www.ctahr.hawaii.edu/nelsons/banana) ........................... 46
Figure 2.15 Pest Risk Analysis process; with arrow indicating which step in process information generated in this thesis has been collected for .......................................... 50
Figure 3.1 Fruit fly trapping sites used to study the influence of site variables on the distribution and abundance of five economically important fruit fly species in Papua New Guinea .................................................................................................................. 55
Figure 3.2 Fruit fly study species: (A) Bactrocera bryoniae (Tryon) (approximate magnification x5), (B) Bactrocera cucurbitae (Coquillett) (x6), (C) Bactrocera frauenfeldi (Schiner) (x6), (D) Bactrocera musae (Tryon) (x6), and (E) Bactrocera umbrosa (Fabricius) (x5). ............................................................................................. 59
Figure 3.3 Seasonal abundance of Bactrocera bryoniae (Tryon) in four areas in Papua New Guinea between June 1998 and September 2001 .......................................................... 63
Figure 3.4 Mean monthly abundance of Bactrocera bryoniae at different altitude levels in Papua New Guinea between June 1998 and September 2001. ..................................... 64
Figure 3.5 Mean monthly abundance of Bactrocera bryoniae against annual rainfall levels in Papua New Guinea between June 1998 and September 2001 ...................................... 65
Figure 3.6 Seasonal abundance of Bactrocera cucurbitae (Coquillett) in four areas in Papua New Guinea between June 1998 and September 2001 ................................................. 68
Figure 3.7 Mean monthly abundance of Bactrocera cucurbitae at different altitude levels in Papua New Guinea between June 1998 and September 2001 ...................................... 69
Figure 3.8 Mean monthly abundance of Bactrocera cucurbitae against annual rainfall levels in Papua New Guinea between June 1998 and September 2001 ...................................... 70
Figure 3.9 Seasonal abundance of Bactrocera frauenfeldi (Schiner) in four areas in Papua New Guinea between June 1998 and September 2001 ................................................. 72
Figure 3.10 Mean monthly abundance of Bactrocera frauenfeldi at different altitude levels in Papua New Guinea between June 1998 and September 2001 ...................................... 73
Figure 3.11 Mean monthly abundance of Bactrocera frauenfeldi plotted against annual rainfall levels in Papua New Guinea between June 1998 and September 2001 ........................ 74
Figure 3.12 Seasonal abundance of Bactrocera musae in three areas in PNG between June 1998 and September 2001 ............................................................................................. 76
Figure 3.13 Mean monthly abundance of Bactrocera musae at different altitude levels in Papua New Guinea between June 1998 and September 2001 ...................................... 77
Figure 3.14 Mean monthly abundance of Bactrocera musae plotted against annual rainfall levels in Papua New Guinea between June 1998 and September 2001 ........................ 78
Figure 3.15 Mean monthly abundance between June 1998 and September 2001 of Bactrocera musae plotted against the relative importance of banana as a food crop in cropping systems in Papua New Guinea ...................................................................................... 79
Figure 3.16 Seasonal abundance of Bactrocera umbrosa in four areas in PNG between June 1998 and September 2001 ............................................................................................. 81

xii
Figure 3.17 Mean monthly abundance of Bactrocera umbrosa at different altitude levels in Papua New Guinea between June 1998 and September 2001 ...................................... 82
Figure 3.18 Mean monthly abundance of Bactrocera umbrosa plotted against annual rainfall levels in Papua New Guinea between June 1998 and September 2001 ....................... 83
Figure 3.19 Mean monthly abundance between June 1998 and September 2001 of Bactrocera umbrosa in Papua New Guinea where breadfruit (Artocarpus spp.) is grown ............. 84
Figure 3.20 Pest Risk Analysis process; with arrow indicating which step in process information generated in this chapter has been collected for ....................................... 87
Figure 4.1 Map of harvest spots for Cavendish banana during the fruit fly damage assessment survey (November 2007-January 2008) carried out in five provinces in Papua New Guinea .......................................................................................................................... 93
Figure 4.2 Mean (± SE) fruit fly infestation in finger samples collected from three locations on a banana bunch (top, middle, bottom). Finger samples were from three banana varieties; Kekiau, Vudu Papua, and Tukuru ............................................................... 100
Figure 4.3 Bactrocera species infestation of bunches from four banana varieties (Daru, Kurisa, Kalapua and Babi) grown in a common garden at Laloki, Papua New Guinea ......... 101
Figure 5.1 Number of oviposition events and subsequent number of emergent flies from single ripe fingers of two banana varieties when offered in a choice arena to single female Bactrocera musae (n=16 for each variety) ................................................................. 113
Figure 5.2 Number of oviposition events (darker shade) and subsequent number of emergent flies (lighter shade) from single fingers of Cavendish and Ladyfinger banana varieties at three stages of ripeness when offered in a choice arena to single female Bactrocera musae (n=16 per variety/ ripeness combination) ....................................................... 114
Figure 5.3 Number of emergent flies from single fingers of two banana varieties at three stages of ripeness when offered in a no-choice arena to single female Bactrocera musae (n=16 banana fingers per variety/ ripeness combination) ............................... 114
Figure 5.4 Mean (± SE) number of flies reared from individual fingers of two banana varieties at three ripeness stages when inoculated with 20 Bactrocera musae eggs (n=16 inoculated banana fingers per variety/ ripeness combination, 60 eggs per banana) ... 115
Figure 5.5 Mean (± SE) number of Bactrocera musae emerging from green Cavendish bananas following the exposure of 500gm of banana to 25 gravid female flies (n = 5 replicates). Emergence of flies from ripe fruit (n = 1 replicate) is a positive control, demonstrating that the flies used to run the trial were gravid. It should not be used to compare yield of flies from green versus ripe fruit .................................................... 116
Figure 5.6 Mean (± SE) fruit attributes for two banana varieties at three stages of ripeness. Letters above columns denote significant difference in the fruit attribute between ripeness stages within the one banana variety (based on 1-way ANOVA with a Tukey’s post-hoc test at p < 0.05) .............................................................................. 119
Figure 6.1 The Gazelle Peninsula, East New Britain Province, Papua New Guinea. The three highlighted localities are where impact trials were carried out .................................. 126
Figure 6.2 Distribution of Bactrocera musae (Tryon) on the Gazelle Peninsula, East New Britain, Papua New Guinea in mid 2000. Source: Mararuai et al. (2001) ................. 129
Figure 6.3 Distribution of Bactrocera musae (Tryon) on the Gazelle Peninsula in December 2000. Source: Mararuai et al. (2001) ......................................................................... 129
Figure 6.4 Distribution of Bactrocera musae (Tryon) indicated by red pins on the Gazelle Peninsula in June 2009; pins mark village residential areas surrounded by vegetables gardens, plantations, secondary or primary rainforest ................................................ 130

xiii
Figure 6.5 Phenology curve of Bactrocera musae (Tryon) on the Gazelle Peninsula (East New Britain, Papua New Guinea (PNG)) (from April 2002 to July 2003) compared with curves in three areas on the PNG mainland (from 1999 to 2001) ............................... 131
Figure 6.6 Infestation of banana bunches protected (bagged) or unprotected (unbagged) from Bactrocera musae (Tryon) on the Gazelle Peninsula, East New Britain, Papua New Guinea ......................................................................................................................... 132

xiv
Supplementary material
Attached is a CD-Rom that includes three supplementary appendices: Appendix 1
contains pdf versions of all formally published literature used in developing the list of
organisms associated with banana in PNG; Appendix 2 has pdf versions of all the
informal ‘grey’ information used in that study, while Appendix 3 contains the
questionnaire.

xv
Statement of original authorship
The work contained in this thesis has not been previously submitted to meet
requirements for an award at this or any other higher education institution.
Data and text extracts of the following publication:
Mararuai, A., Allwood, A.J., Balagawi, S., Dori, F., Kalamen, M., Leblanc, L.,
Putulan, D., Sar, S., Schuhbeck, A., Tenakanai, D., & Clarke, A.R. (2001). Introduction
and distribution of Bactrocera musae (Tryon) (Diptera: Tephritidae) in East New
Britain, Papua New Guinea. Papua New Guinea Agricultural Journal, 45, 59-65
have been used in Chapter 6 of this thesis because the paper contains information
which is an integral component of the larger research area that chapter covers. I am the
first author on the publication and took part in all activities reported in it.
To the best of my knowledge and belief, the rest of the thesis contains no material
previously published or written by another person except where due reference is made.
Signature
Date

xvi
Acknowledgements
My journey through the making and the telling of this story has not been a ‘magical red carpet’ ride. But like all veteran ships of the seven seas you need a good crew and a wise captain to take it through all the rough and stormy weather. I am not a veteran ship but I have had the privilege of having such a supportive crew and a good captain; if I can put it that way. They’ve held me together and I’ve weathered the storms to reach this port. I am grateful to many for the time, consideration, support and understanding you have all given during my journey. Thank you to the Institutions I have worked with and their staff: PNG National Agriculture Research Institute, Australian Centre for International Agriculture Research, Queensland University of Technology & School of Natural Resource Sciences, PNG National Agriculture and Quarantine Inspection Authority, Fresh Produce Development Authority, Queensland Department of Primary Industries & Fisheries (Indooroopilly & Cairns), and Secretariat of the Pacific Community. To ACIAR, thank you for your faith in me and for the considerable support and understanding along the way, I hope I have fulfilled your aspirations in selecting me for the John Allwright Fellowship Award. To my supervisors: Tony, Grant, Ian, and help from Raghu - I don’t know how to say thank you. There are no words so I simply say ‘bikpela tenkyu tumas’ To my family, thank you, know that I am still the same person. A big thank you to my friends, in particular: Amy, Trish, Helen, ‘Boss man’ David, Fred, Rosa, Roy, Tim, Anna, Sol, Eilish, Aunty Janet and Howard. To my godmother Ena and Tom, thank you for listening and helping me find my wings. And a special thank you to my son Laurence: I wish I had the same courage as you at the age of four to be so brave and understanding in supporting me by living apart at this time.

1
Chapter 1. Literature review
1.1 Introduction
1.1.1 Agriculture in Papua New Guinea
Papua New Guinea (PNG), the largest of the Pacific Island Nations and Territories,
relies heavily on its agricultural sector for both internal food security and export
earnings. Agriculture at the subsistence, semi-commercial and commercial level is
practiced throughout Papua New Guinea (Hanson et al., 2001). Over 80% of the
population of PNG live through subsistence agriculture, while a large percentage of the
remainder practice or partake in commercial agriculture for domestic and international
markets (Allen et al., 1995; Asafu-Adjaye, 1996; Bourke, 2001; Allen et al., 2005).
Agriculture is a main economic activity for rural people (Bourke & Vlassak, 2004),
providing income generation from the sale of crops (fruit, nuts, vegetables, leafy
vegetables) and/or livestock (poultry, pigs) (Thompson, 1986; Allen et al., 1995; Coelli
& Fleming, 2004).
Improvement in production and management activities in the agriculture sector,
particularly that which leads to greater farmer cash incomes, can be a catalyst for
broad-based economic growth and development including enhanced access to health,
education and information (Shack et al., 1990; Duncan & Temu, 1995; Benjamin et al.,
2001; Gwaiseuk, 2001; Allen et al., 2005). Unfortunately, as with many developing
countries, PNG lacks much of the underpinning scientific research which is needed to
drive significant increases in rural agricultural production (Allen et al., 1995; Ohtsuka
et al., 1995). Pest management in agriculture production is an example of one area.
Standardised quantification of pest damage on crops and the proper identification and
collation of pest information is generally lacking, despite this information being
essential in improving harvestable yields and market quality. The country also does not
have an appropriate and recognised pathway through which commodities may be
produced and suitably prepared for market (Wamala, 2001). This is especially true for
fresh commodities.
In January 2006, a national workshop on fresh commodity pathways called
Commodity Pathways for Main Exported Commodities (ginger, taro, banana,
asparagus, etc) was held in Port Moresby, the capital of PNG. The workshop identified

2
problems in commodity supply and infrastructure affecting rural industries in the
country. Through the support of current PNG National and Regional initiatives, it is
considered that long term industry goals targeting international fresh commodity trade
at a regional level are achievable. Biosecurity standards for potential export
commodities are yet to be established and commodity pathways do not exist, but
establishment of fresh commodity export will enhance socio-economic levels in the
country. There is a large range of potential fresh commodity export crops in PNG,
including mango, pawpaw and banana. As part of the commodity pathway workshop it
was decided that a single commodity should be used as a case study upon which to
focus and guide future research and development. The commodity that was chosen was
banana and the research presented in this thesis is directed at supporting that national
initiative.
Banana (Musa spp.) is an important staple food crop in PNG (Bourke et al., 1998;
Gibson, 2001b; Gwaiseuk, 2001; Bourke & Vlassak, 2004). It is cultivated from sea
level to elevations above 2000 meters and is grown in over 300 of PNG’s agricultural
cropping systems (Bellamy & McAlpine, 1995; Bourke et al., 1998; Hartemink &
Bourke, 2001; Gunther et al., 2003). The fruit is sold in almost all rural and urban
markets, with an annual production of about 413,000 tonnes which generates an
estimated PNG Kina 150 million (Gibson, 2001a); hence it is the second most
important national food crop after sweet potato (Ipomoea batatas) (Gibson, 2001a;
Bauer et al., 2003). Banana is a commodity produced and marketed largely in its fresh
state and, therefore, as a potential export commodity it will be subjected to more
stringent trade protocols than those imposed on exported processed commodities (e.g.
coffee and cocoa beans). International market access for PNG bananas is reliant on
knowledge of the biosecurity threats posed by its associated pests. There is, however,
very limited formal literature on the pests of PNG bananas, with most information
being available through unpublished expert knowledge or in non-refereed (and hence
non-verified) “grey literature”.
1.1.2 Limitations of market access
Provision of information on commodity pests by an exporting country is an essential
and compulsory part of trade negotiations prior to gaining export approval for a fresh
commodity (Follett & Neven, 2006). The information provided should include the

3
identification of pest(s) of the commodity and pathways through which the pest(s) may
travel; the assessment and categorisation of a pests’ status; the probability of
introduction, spread and economic impact of the pest(s); and finally risk management
of the pest(s) (IPPC, 2004a, 2006a). Under guidelines of the International Plant
Protection Convention (IPPC, 2004a, 2008), this collection of information is referred
to as the Pest Risk Analysis (PRA). PRA information should be generated using
standardised analytical approaches that produce scientifically credible data and be
reported in a transparent manner to convey accurate understanding (Alvarez-Coque &
Bautista, 1994; Gray et al., 1998; Zepeda et al., 2001; Lugard & Smart, 2006).
Inability to provide sufficient PRA information to trading partners and the lack of
knowledge, awareness and effectiveness in the monitoring and management of pests is
a major limitation for many developing countries (Duncan & Lutz, 1983; Markelova et
al., 2009). This limitation causes difficulty in accessing and maintaining market access
to industrial country markets. Other reasons for poor market access of developing
country fresh commodities include central governing support systems inflexible to
market changes, poorly developed financial systems, high costs in legal or non-legal
transactions and a lack of trust in banking and financial transactions (Margolis et al.,
2005; Schillhorn van Veen, 2005). Papua New Guinea is one country which has yet to
establish some of these systems for fresh agricultural crop commodities (Kannapiran,
2000; Manning, 2001).
1.1.3 Invasive species and export systems
Successfully established invasive pest species can result in negative social (Drew,
1996), economic (Perrings, 2005) and environmental (Usher, 1988; Zavaleta et al.,
2001) impacts in the countries or regions where they establish (Hennessy, 2008).
Biological security (= biosecurity) protocols exist to monitor and regulate trade
pathways to prevent the spread of such potentially invasive pests. International
standards (IPPC, 2006a) exist which, for signatory countries, describe quarantine
protocols that must be followed in order to minimise the risk of movement of plant
pests through trade, while allowing that international commodity trade to continue.
Regulatory biosecurity standards describe a set of procedures or measures established
to safeguard the flora and fauna (both agricultural and native) of a country, or a
particular area within a country, against exotic pests. These standards, in part, draw

4
from research from the scientific field of invasion biology; a field which studies the
[generally human aided] movement and establishment of invasive organisms from
endemic to non-endemic areas. There are generally considered to be four stages in the
invasion process; introduction, establishment, reproduction and spread (Lockwood et
al., 2005). Management of invasives at the introduction phase is identified as being the
most effective, both practically and cost wise (Myers et al., 2000; Maynard et al.,
2004; Hennessy, 2008).
A wide array of diverse fields are encompassed by biosecurity, including higher
education, scientific research, administration and policy, media, politics, trade and
industry (Whittle, 2004). Market access is one component of biosecurity and focuses
on establishing commodity export markets in the presence of biosecurity requirements
(e.g. within-field or post-harvest controls) imposed by an importing nation (Duncan &
Lutz, 1983). Countries may impose different equivalent conditions and systems in
order for an exporter to meet market access requirements.
1.1.4 Overview of chapter
In the absence of biosecurity protocols for market access, PNG will not be able to
market its fresh agriculture produce at an international level. As part of the process for
developing appropriate biosecurity protocols for PNG commodities, especially banana,
this thesis covers and provides components of the total information required for a PNG
banana PRA. The components vary from reviews of existing, but poorly documented
(i.e. “grey”) knowledge, through to the generation of new scientific data. To establish
the context of the thesis, this chapter provides an overview of the biosecurity
requirements required for market access of fresh commodities and the reasons why
they are necessary. It introduces and reviews pertinent aspects of the invasion biology
theory, which provides the scientific underpinning for many biosecurity protocols.
Following from that, fruit flies (Diptera: Tephritidae) are introduced and the reasons
why they are of such high global biosecurity concern and why fruit fly susceptible
commodities are stringently monitored. The review then focuses in on the study
system, introducing PNG bananas and their pests, and explains why particular focus
within the thesis is given to Bactrocera musae (Tryon) (Diptera: Tephritidae: Dacinae),
the banana fly. The last section of this chapter formally outlines the thesis structure.

5
1.2 Invasion biology and its role in biosecurity
Invasion biology covers the processes which occur when an organism moves from an
endemic to a non-endemic area, establishes in that new area, and then spreads out from
that area. The organism involved in such a process is called an invasive species and
there is often a need for its management as many invasive species often have negative
environmental or economic impacts in the new area (Higgins et al., 1999; Parker et al.,
1999; Park, 2004). Biosecurity protocols aim for entry prevention, monitoring, post-
entry management and/or eradication of a potential or actual invasive organism and
only through understanding the processes involved in biological invasions can
appropriate international protocols be established (Myers et al., 2000).
Several descriptions of the process of biological invasion have been published: all have
in common an initial movement of an invasive species into a new area, where the
invasive subsequently settles and, if successful, establishes and spreads. I cover three
such descriptions of biological invasions to illustrate their commonalities and relatively
minor differences. Lockwood et al. (2005) proposed what they termed ‘the multi-step’
invasion process (Figure 1.1A). This model identifies an initial phase of the invasion
where the invasive organism is transferred by a vector from its native habitat into a
non-native habitat. During the second phase of ‘establishment’ the invasive organism
acclimatises to the new local environment and likewise the environment to the
invasive. Successful establishment then leads to the third phase, ‘population growth’.
As the population increases and secondary natural and/or anthropogenic dispersal
occurs, the fourth phase may be reached when invasive individuals disperse from the
initial area of introduction. Vermeij (1996) divided the invasion process into three
successive stages: arrival, establishment, and integration. Arrival (analogous to the
Transfer and Release stages in the Lockwood et al. (2005) model) is defined by the
dispersal of individuals to the recipient region, occurring naturally or with the aid of
humans. Establishment implies that the new population can sustain itself through local
reproduction and/or recruitment. Integration occurs when, as the invading species
forges ecological links with other species in the recipient region, local adaptive
evolution occurs, reflecting a changed selective regime in the recipient community.
The invasion model of Kolar & Lodge (2001) has similarities to the above two models,
but additionally describes the invasion process as having ‘transition’ periods and the
authors assert that each stage is discrete (Figure 1.1B). Invasive species must

6
successfully pass through the different transition stages of transportation, release,
establishment and spread to continue through the invasion process. Incorporating all
three of the invasion process models into a single consensus model confirms the
following four basic stages: arrival, establishment, population growth and range
expansion (Figure 1.2). A conceptual model which breaks the invasion process into
discrete successive stages has both theoretical and applied benefits. From a
management perspective, targeting the early stages of arrival and establishment has
been deemed most effective when developing management strategies against invasive
species (Ruesink et al., 1995; Carey, 1996; Kolar & Lodge, 2001).
Species entrained in transport pathway
Fails in transport
Survives transport and introduction
Fails to establish
Establishment
Non-invasive
Invasive
Uptake from native range
Transfer via vector
Population increase and range expansion
Establishment
Release; arrival
A B
Species entrained in transport pathway
Fails in transport
Survives transport and introduction
Fails to establish
Establishment
Non-invasive
Invasive
Species entrained in transport pathway
Fails in transport
Survives transport and introduction
Fails to establish
Establishment
Non-invasive
Invasive
Uptake from native range
Transfer via vector
Population increase and range expansion
Establishment
Release; arrival
Uptake from native range
Transfer via vector
Population increase and range expansion
Establishment
Release; arrival
A B
Figure 1.1 (A) The multi-step process of non-native species invasion (Lockwood et
al., 2005); and (B) Transitions that non-invasive species must overcome to
continue in the invasion process (Kolar & Lodge, 2001)

7
Arrivaldispersal routes, human transport
Establishmentacclimatisation, breaking down ecological resistance
Population growthhave a minimum viable population, reproduction increases
Expansionspread and in the process influences the new environment for support
Arrivaldispersal routes, human transport
Establishmentacclimatisation, breaking down ecological resistance
Population growthhave a minimum viable population, reproduction increases
Expansionspread and in the process influences the new environment for support
Figure 1.2 Conceptual model of the process that invasive species go through in their
movement from endemic to non-endemic areas
Within the invasion process, arrival and establishment have been found to be the two
areas which, when targeted, are most likely to give positive outcomes in management
(Drew, 1996; Williamson & Fitter, 1996; Maynard et al., 2004; Hochberg & Gotelli,
2005). This is because populations are generally small and can be controlled or
potentially eradicated. The arrival stage describes the natural or anthropogenic
movement of an organism from an endemic area to the non-endemic area (Vermeij,
1996), while establishment refers to the persistence of an immigrant population by
means of local reproduction and recruitment (Vermeij, 1996). Establishment may be
accompanied by sporadic spreading of the population in the recipient region
(Ebenhard, 1989).
Successful establishment of an invasive organism is commonly related to positive
propagule pressure (Kolar & Lodge, 2001; Lockwood et al., 2005). Propagule pressure
is a measure of the number of individuals released into a region to which they are not
native (Carlton, 1996) (also termed ‘introduction effort’, (Blackburn & Duncant,
2001)) and incorporates estimates of the absolute number of individuals involved in
any one release event (propagule size) and the number of discrete release events
(propagule number). Propagule pressure increases as the number of releases and/or the

8
number of individuals released increases (Lockwood et al., 2005; Memmott et al.,
2005). Many invasive species have low establishment success, but once introduction
succeeds then there may be rapid population growth (Shigesada & Kawasaki, 1997;
Christian & Wilson, 1999; Sax & Brown, 2000). Reducing the chance of entry or
establishment by reducing propagule pressure and/or initial population size is the key
objective of trade related biosecurity measures.
The spread of invasive species is related to international trade and travel and the likely
continuity of new invasions is linked to the increasing demand, efficiency and ever
shortening travel times of the global transport industry (McAusland & Costello, 2004;
Perrings, 2005; Cook & Fraser, 2008). Management and monitoring of invasive
organisms in such situations becomes difficult, but the utilisation of the invasion
process identifies optimum stages where control and monitoring activities are best
applied (Everett, 2000; Hall & Hastings, 2007). Identification of pests and the likely
pathways through which they can move is therefore an essential investigatory
component in the management of invasive species.
Numerous global invasions have occurred in the area of agriculture and horticulture
which have negatively affected agricultural communities and associated stakeholders
(Allwood & Leblanc, 1996; Drew, 1996; Maynard et al., 2004). The trade of export
commodities in agriculture requires the establishment of globally accepted standards of
pest management before acquiring, and then while maintaining, market access (Follett
& Neven, 2006). Such standards often require regulating local biosecurity measures,
including pre-harvest control methods such as pesticide use (Vargas et al., 2002; Wang
et al., 2005; Stonehouse et al., 2007; Burrack et al., 2008) and integrated pest
management options (Meats et al., 2003; Follett & Hennessey, 2007; Narrod et al.,
2009), and post-harvest control methods such as packaging and storage of commodities
(Moy & Wong, 2002; Hofman et al., 2003; Follett, 2004; Birla et al., 2005). The more
serious the known impacts of the pest, the more stringent such biosecurity measures
tend to be.
1.3 Fruit flies as invasive organisms
Global monitoring and management of one particular group of crop pests, the true fruit
flies (Diptera: Tephritidae) is especially stringent (Aluja & Mangan, 2008). Within the

9
Tephritidae, members of the subfamily Dacinae are particularly important as one of the
most globally recognised groups of insect pests attacking fruit and vegetable crops
(Fletcher, 1987; Aluja & Mangan, 2008). Dacine fruit flies are frugivorous insects, the
immature life stages of which feed and develop in fruit and seed pods (Fletcher, 1987).
Most dacine species are monophagous (feed on one host species) or oligophagous
(narrow range of related hosts), while a few, commonly the serious pest species, are
polyphagous (wide range of hosts) (Christenson & Foote, 1960). The Dacinae are
recognized for their invasiveness in tropical, sub-tropical and temperate regions of the
world, although not all species are invasive (Hardy, 1991; Carey, 1996; Drew, 1996).
For the remainder of this thesis, unless otherwise specifically referred to, the term
“fruit fly” refers to members of the Tephritidae, sub-family Dacinae.
Fruit flies possess morphological and biological characteristics that are thought to
increase their invasive potential. For example, some species (e.g. Bactrocera dorsalis
(Hendel), B. carambolae Drew & Hancock, B. philippinensis Drew & Hancock, and B.
papayae Drew & Hancock) have longer and stronger ovipositors than other related
species (Drew et al., 2008), enabling female flies to penetrate thicker and denser
epidermal layers of host fruits, resulting in a broader host range than related flies
(Drew & Hancock, 1994; Allwood et al., 1999). Aggressive territorial behaviour also
allows populations to dominate the areas they invade (Christenson & Foote, 1960),
creating aggressive competition for resources that have also been speculated to drive
fruit fly invasions (Duyck et al., 2004). Flexibility in temperature requirements for life
stage development may also enable adaptation and survival of fruit flies in different
regions with different temperature regimes (Zhou et al., 1994). For example,
Bactrocera cucurbitae (Coquillett) and B. dorsalis are able to reproduce and survive in
both low and high temperature environments (Vargas et al., 2000b; Vayssieres et al.,
2008).
Commodities susceptible to fruit fly (generally fresh fruits and some vegetables) are
internationally traded on a routine basis and the detection of infested commodities
often only occurs after the commodity has entered the area into which it is being
imported. Non-commercial carriage of fresh fruits (i.e. fruit carried by passengers on
boats or planes) poses the same potential risk (Putulan et al., 2004). Establishment of
exotic fruit flies in new agricultural production areas are costly. For example, the
detection of Bactrocera papayae (Asian papaya fruit fly) in Cairns, North Queensland,

10
was detected about two years after introduction. Eradication was achieved but the
exercise involved entire community efforts and direct eradication costs of about
AUD$33M (Drew, 1996; Maynard et al., 2004). This case emphasises the necessity of
ensuring that incursions be prevented and detections be immediately managed.
Another, but slightly different example is the continued presence of invasive Ceratitis
capitata (Weidemann) populations in orchard areas in California. Studies have
determined that the continued presence of C. capitata in closely managed fruit and
vegetable growing areas is due to isolated re-introductions or pockets where
populations have evaded control efforts (Carey, 1996). Management of this pest in
California, which has a US$17 billion fruit industry, requires and drives continued
research into finding solutions to maintain low infestation levels and international
market access (Headrick & Goeden, 1996). In the Pacific, fresh commodity exports of
mango and papaya from countries such as Fiji and Samoa are acceptable only with the
continuous management of invasive fruit fly pests such as B. passiflorae (Froggatt) and
B. xanthodes (Broun) (McGregor, 1996; McLeod, 2005). Such management requires
the use of control methods such as fruit fly trapping networks that keep populations at
low levels, or provide ongoing evidence of ‘area freedom’ status (Allwood, 1996b;
Papadopoulos et al., 2001). Orchard areas in the southern states of Australia also
maintain market access based on area-free zones, which are tightly regulated and
actively managed (Fletcher, 1974; Maelzer et al., 2004).
1.4 The study system: banana and banana fly
Bananas (Musa spp.) are endemic to Southeast Asia and the Pacific. Papua New
Guinea is geographically located at the centre of that diversity range and has the
highest banana diversity of any nation in the Pacific region (Kingwell et al., 2001). The
diversity of banana in PNG consists of over 200 genetic variations identified to be pure
or hybrid varieties of Musa acuminata and other wild Musa species. There are diploid,
triploid, tetraploid and fe’i varieties cultivated throughout the country (Arnaud &
Horry, 1997).
Cultivation and production of banana in PNG is continuously and negatively affected
by pests. There is little formal information available on the pests of bananas in PNG.
Informal pest reports commonly document the following: Nacoleia octasema
(Meyrick) (banana scab moth), Erionota thrax (Linnaeus) (banana skipper),

11
Bactrocera species (fruit flies), nematodes and numerous fungal pests such as
Mycosphaerella musae (leaf speckle), Cordana musae, Deightoniella torulosa,
Guignardia musae (freckle disease of banana), Mycosphaerella fijiensis (black leaf
streak) and Phyllachora musicola (Pone, 1994; Waterhouse, 1997; Kambuou, 2003).
The generation of an entire list of pests of PNG bananas is essential in helping define
which are major or minor, exotic or endemic, and will indicate areas for further
research. Currently there is a lack of research into the quantitative damage levels for
banana pests, their spatial and temporal distribution, and economic impact. Field
observations and reports show the presence of pests in different parts of the country but
evaluation of damage or economic impact with the use of non-standardised survey
methods do not allow for national or regional comparison.
Substantial informal knowledge (e.g. (Pone, 1994; Waterhouse, 1997; Leblanc et al.,
2001; Kambuou, 2003) and limited formal research (Smith, 1977b; Fletcher, 1998)
indicate one fruit fly species, B. musae, as perhaps the most important insect pest of
cultivated bananas in PNG. Bactrocera musae occurs in far northern Queensland,
Australia, and PNG (Drew, 1989). The species is not uniformly distributed in PNG
(pers. obs.) and it is invasive in some PNG island provinces (Mararuai et al., 2001).
Detailed spatial and temporal distribution information for the fly is limited, but fruit fly
surveys in PNG show that it is widespread on the mainland (Clarke et al., 2004).
Distribution of the fly in the island provinces of PNG is less clear, but it has been
trapped and bred from bananas in East New Britain where it is becoming widespread
over most of the Gazelle Peninsula; this population is recently invasive (Mararuai et
al., 2001). Banana fly specimens have been collected on Lihir Island (New Ireland
Province) and Manus, but it is not confirmed whether permanent breeding populations
occur there (Leblanc et al., 2001).
In its natural range, the primary host of B. musae is banana, with a single record only
from papaya. In PNG the fly causes commercially significant damage to eating and
cooking bananas. In Australia, B. musae has been recorded from 12 host species, from
10 genera and nine families (Hancock et al., 2000), but the majority of records are
from banana (Hancock et al., 2000; Leblanc et al., 2001) (Table 1.1). Banana fly has
been demonstrated in the laboratory to show a high preference for bananas over other
fruit (Fitt, 1986) and field officers in PNG have observed that there may be preferential
selection of banana varieties, based on varying infestation levels (Smith, 1977b).

12
Table 1.1 Host plants for Bactrocera musae (Tryon) in Australia (Distribution: Torres
Strait islands and northeast Queensland, as far north as Townsville). Taken
from - The Distribution and Host Plants of Fruit Flies (Diptera: Tephritidae) in
Australia (Hancock et al., 2000).
Host species Common name
Reference Comments
CAPPARACEAE Capparis lucida coast caper Hardy 1951 one specimen only CARICACEAE Carica papaya pawpaw Hardy 1951, QDPI Papaya fruit
fly database (1995-1999) occasional host
MUSACEAE Musa banksii native banana Hardy 1951, May 1953, QDPI
Papaya fruit fly database (1995-1999)
major host
Musa x paradisiaca (=acuminata)
banana Hardy 1951, QDPI Papaya fruit fly database (1995-1999)
not known from hard green
Musa x paradisiaca dwarf banana May 1953, QDPI Papaya fruit fly database (1995-1999)
Musa x paradisiaca lady finger banana
QDPI Papaya fruit fly database (1995-1999)
Musa x paradisiaca sugar banana QDPI Papaya fruit fly database (1995-1999)
Musa x paradisiaca plaintain May 1953, QDPI Papaya fruit fly database (1995-1999)
MYRTACEAE Psidium guajava guava Hardy 1951,May 1960, QDPI
Papaya fruit fly database (1995-1999)
occasional host
Syzygium bamagense Drew 1989 OLACACEAE Ximenia amerciana putit QDPI Torres Strait database
(1993-1995)
PASSIFLORACEAE Passiflora edulis passionfruit QDPI Papaya fruit fly database
(1995-1999) two records
RUBIACEAE Lasianthus strigosus blue rubi QDPI Papaya fruit fly database
(1995-1999) one specimen only
RUTACEAE Citrus paradisi grapefruit QDPI Papaya fruit fly database
(1995-1999) one specimen only
Citrus reticulata mandarin QDPI Papaya fruit fly database (1995-1999)
one record from ripe fruit
SOLANACEAE Solanum lycopersicum
tomato QDPI Papaya fruit fly database (1995-1999)
one record from ripe fruit

13
1.5 Thesis structure
The information presented in this thesis aims to provide some of the underpinning
scientific knowledge required to set up biosecurity standards for PNG bananas,
information which is essential for gaining international market access for one or more
varieties of this commodity. The term ‘PNG bananas’ is used throughout the thesis and
where it is used it refers to the fruit of the banana, unless specified otherwise. One
thesis alone can never supply all that information, for there are a large number of
potential pest species associated with this commodity. This thesis does, however,
initially identify and categorise the status of all pests associated with the banana plant
in PNG, before focusing towards one major insect pest, the banana fly. The research
approach used here follows the Pest Risk Analysis (PRA) process as defined by the
International Standards for Phytosanitary Measures presented by the Food and
Agricultural Organisation of the United Nations (IPPC, 2006a) (Figure 1.3). While I
will not address all components of a PRA, I have identified key components, within
each of the three stages that make up the PRA process, thus helping focus my research.
The three stages in the PRA process are: (i) Risk Initiation [Identify pest(s) and
pathways of concern, and consider whether the pest should be considered for risk
analysis in relation to the area at risk]; (ii) Risk Assessment [Assess (a) the risk of
entry; (b) the risk of establishment; and (c) the risk of economic and other impacts];
and (iii) Risk Management [Review risk management options (e.g. import
restrictions, invasion tactics)].
RISK MANAGEMENTReview risk management options (e.g. import
restrictions, invasion tactics)
RISK ASSESSMENTAssess (a) the risk of entry; (b) the risk of establishment; and (c)
the risk of economic and other impacts
RISK INITIATIONIdentify pest(s) and pathways of concern, and consider whether the pest should
be considered for risk analysis in relation to the area at risk
3
2
1
RISK MANAGEMENTReview risk management options (e.g. import
restrictions, invasion tactics)
RISK ASSESSMENTAssess (a) the risk of entry; (b) the risk of establishment; and (c)
the risk of economic and other impacts
RISK INITIATIONIdentify pest(s) and pathways of concern, and consider whether the pest should
be considered for risk analysis in relation to the area at risk
3
2
1
Figure 1.3 Pest Risk Analysis process

14
In addressing the initial step in the PRA process, Risk Initiation, Chapter 2 of this
thesis is a review of the pests of PNG bananas. While the thesis subsequently focuses
on B. musae, as a major pest of PNG banana fruit, there are no available collations,
reports, or lists of all other known banana pests. This chapter is therefore aimed at
identifying and collating as much information as is possible on banana pests in PNG.
The chapter is more than a simple, desk-based literature review, as it utilises, in
addition to refereed literature, structured interviews with PNG researchers and
“investigative” research sorting through personal unpublished files and grey-literature
kept by individuals or PNG field research-station libraries. The outcome of this chapter
showed that while there are over 100 organisms associated with the banana plant in
PNG, only 14 are of sufficient concern that they are regularly managed. Of these 14,
13 already have significant existing information which can be used towards a PNG
banana fruit PRA. In contrast to those 13, required information for the banana fly,
Bactrocera musae, is almost entirely lacking. As a result of this review, the decision
was made to focus the remainder of the thesis on banana fly.
Thesis chapters’ three to six provide new information on B. musae pertinent to
providing information required by a PRA. Information covered includes distribution
and abundance, crop impacts and invasive potential (detailed more fully below). The
information provided in any one chapter does not easily slot into one particular step of
the PRA as does Chapter 2. Rather, chapters’ three to six provide information which
can be applied to different facets of both steps two and three of the PRA. The pulling
together of this information for a PRA is undertaken in the final Discussion chapter.
While Bactrocera musae is endemic to PNG, this need not imply that banana fly is
uniformly distributed or abundant within PNG. Knowledge of the distribution and
abundance of a pest species is important within a PRA both to gauge potential risk.(e.g.
assessing areas where crop infestation is most likely) and also for risk management
(e.g. growing crops within areas of low pest pressure). Chapter 3 uses an existing, but
previously unanalysed data set to investigate the spatial and temporal distribution of B.
musae and four other pest fruit flies in PNG and factors influencing that distribution.
The detailed local distribution of banana fly in PNG is unknown and without this
information identifying production areas which may be at a higher or lower risk from
the fly is impossible. The other pest fruit fly species are included as they allow a

15
comparative study, of broader ecological and applied interest, about factors impacting
on the abundance of fruit flies in tropical regions.
Following from Chapter 3, Chapter 4 investigates if the same banana variety,
Cavendish, grown in different geographic areas of PNG has higher or lower risk of
banana fly infestation. Cavendish is an economically important banana variety in PNG
and is a variety likely to be considered for export from PNG. As such, it is a basic
requirement for Stages 1 and 2 of the PRA process that banana fly infestation levels
suffered by fruit of this variety in PNG are documented. Further, if in different PNG
agro-ecological regions Cavendish fruit suffer significantly less from banana fly
infestation, then export only from such regions could be used as a risk mitigation step
(PRA Step 3). The Cavendish data is supported by other PNG field data, on the
infestation levels of other banana varieties.
The results of Chapter 4 showed very low field levels of banana fly infestation in
Cavendish fruit, a particularly unexpected result given the general abundance of
banana fly in PNG (Chapter 3) and the perceived pest status of the insect (Chapter 2).
To further investigate host usage by banana fly, and thus better quantify the biological
and economic threat posed by the insect, detailed laboratory studies were undertaken.
Better knowledge of host use is not only important for assessing risk, but also for risk
management. Commodities which are non-hosts, or poor hosts, may have reduced, or
even no risk reduction treatments imposed upon them. Host status, however, can vary
depending on ripening stage of a commodity, or vary between cultivars/varieties of a
commodity, and this needs to be known. In Chapter 5, I use laboratory trials to
investigate host choice and utilisation of B. musae for two economically important
banana varieties, Cavendish and Ladyfinger. I also investigated the influence of fruit
ripening stages on host-use. Key fruit traits (e.g. peel thickness, fruit toughness, etc)
were recorded to investigate if such traits correlate with host use.
A Pest Risk Analysis is an attempt to identify the risks associated with transporting a
commodity, particularly with respect to the likely introduction, establishment, spread
and impact of pests associated with that commodity. By the nature of biosecurity, i.e.
aimed as preventing incursive events, most PRAs can only hypothesise about the likely
impact of an exotic pest. A real incursion, however, gives an opportunity to study a
pest’s post-incursion distribution and abundance, that insect’s economic impact in a

16
novel area, and options for management. In PNG, B. musae was detected as a new
incursive on the Gazelle Peninsula of East New Britain Province in mid 2000. In
Chapter 6 I present information on this incursion, including the flies changing
distribution and, abundance over time, it’s phenology in the novel environment
compared to its phenology in its endemic range, and its crop impacts. The study
provides new information, or supporting information, for all three steps of the PRA
process.
The final discussion chapter (Chapter 7) brings together the results obtained throughout
the thesis by summarising them within the PRA framework, specifically with respect to
a PNG banana fruit PRA. The apparently unusual observation of very high pest
abundance, but very low crop infestation, is also discussed in detail. I conclude that for
specialist herbivores, such as B. musae, such a result should not be unexpected. The
implications of this conclusion for international market access protocols are raised,
specifically that pest trap catch should not be automatically used as a surrogate for crop
risk.

17
Chapter 2. Review of banana (Musa spp.) pests in Papua New Guinea
2.1 Introduction
Banana is an important food and commodity crop in Papua New Guinea (PNG), being
widely cultivated from sea level to above 2000 meters above sea level (m.a.s.l)
(Hartemink & Bourke, 2001; Gunther et al., 2003). Production, however, is often
affected by damage to plant parts caused by pests, or through competition with weeds.
This is a major concern when trying to obtain cross-border market access for PNG
banana because the unrestricted presence of pests of quarantine concern will stop trade.
Globally, plant pathogens are considered the major pests of banana production,
followed by nematodes, insects and mites (Gold et al., 2002). In PNG, there is a range
of pathogens, arthropods and nematodes reported on banana (Kumar, 2001), but only a
few are commonly observed and reported as having an impact on the amount and
quality of bananas produced (Pone, 1994; Waterhouse, 1997; ACNARS, 2003;
Kambuou, 2003). Some of these are endemic while others are not (Shivas & Philemon,
1996; Davis et al., 2000). Schuhbeck (1996), from his surveys in PNG’s island
provinces in the Bismarck Archipelago (i.e. West New Britain, East New Britain,
Manus, New Ireland, and Bougainville), further reported that some PNG banana pest
species endemic to the mainland, are exotic to the islands. Earlier surveys and
publications on PNG banana pests are dominated by insects (O'Connor, 1949; Szent-
Ivany & Barrett, 1956; Szent-Ivany & Catley, 1960; Ostmark, 1974; Smith, 1976) and
some of those insects remain important (Wilkie et al., 1993; Lubulwa & McMeniman,
1998; Mararuai et al., 2001). In more recent reports, pathogens and nematodes have
been added to the national banana pest list (Bridge & Page, 1982; Tomlinson, 1984;
Philemon, 1986; King et al., 1988; Tomlinson et al., 1988). Most of the publications
cited here that list putative pest species on banana are informal (e.g. unpublished
reports of limited circulation), often scarce and difficult to access in either paper or
electronic format. Further, the literature on PNG banana pests is scattered and the few
available summative lists are out dated, incomplete, or both. For example, the work
done by Szent-Ivany and his colleagues is half a century old and was not exhaustive
even when published, but is the most recently prepared formal summative publication
of the insects of banana in PNG.

18
Research conducted as part of this chapter showed that official national records on the
pests found on banana are incomplete and do not even reflect known research done on
banana crop management and plant protection within PNG, let alone representing an
authorative national pest list. Official national records are maintained on the PNG Pest
List Database (PLD). The PLD is an information system that records pest occurrences
within a country, provides reports on their occurrences and is held by the National
Agriculture and Quarantine Inspection Authority (NAQIA) (Masamdu, 2006).
Extraction of records from the PLD, however, found that the database held only six
records, five of these were fungal pathogens, and did not include even well
documented PNG banana pests (Figure 2.1). It was therefore of little use, even as a
starting point, for developing a PRA.
Page 1 of 1 Papua New Guinea Pest Lists Database 22-Dec-06 Hosts with Recorded Pest
Host Pest Literature
.../ ..common Order / common name reference
Musa sp / banana and f Mycosphaerella musae Leaf speckle PNG Plant Protection DB
plantain a Bactrocera bryoniae (Tryon) Drew, 1989
f Cordana musae PNG Plant Protection DB
f Deightoniella torulosa Hyde KD & Philemon E, 1994
f Guignardia musae freckle disease of banana Hyde KD & Philemon E, 1994
f Mycosphaerella fijiensis Black leaf streak PNG Plant Protection DB
f Phyllachora musicola PNG Plant Protection DB
Total number of host-pest: 7
Prepared in collaboration with the:
Plant Protection Service, Secretariat of the Pacific Community
This information is provided in good faith from the best records available at the time. The Secretariat of
the Pacific Community cannot accept responsibility for an consequences arising from the use of this
data. For further information please contact the Government Department and/or the SPC Plant Protection Service.
By request from users, where more than one occurrence of a
Pest has been recorded on the same host,
only a single occurrence is displayed
Figure 2.1 Report generated by Papua New Guinea Pest List Database in December
2006 of the pests recorded on banana and plantains in Papua New Guinea
The obvious incompleteness of official records will seriously impede the processes
involved in acquiring market access for PNG bananas. With PNG’s intention to
investigate the potential of banana as an export commodity, it is an international
requirement to supply an authorative list of the pests of the commodity within the

19
country: defining the pests of a commodity is the first step in the Pest Risk Analysis
(PRA) process, a key element of market access protocols. The primary goal of this
chapter was therefore to document all the pests reported and recorded on banana in
PNG. Further, I review the known biological research and economic impact data of the
most commonly reported organisms as a mechanism of prioritising where further
research is required if market access for PNG bananas is to be gained.
2.2 Materials & Methodology
Sources of Information. Refereed literature on pests of PNG bananas is limited,
restricted in the taxa covered and most of the papers document aspects of the biology
and management of individual pest species. A comprehensive list would require
information from additional sources and thus a search for literature was done and was
able to find formal literature, informal ‘grey’ information and knowledge from local
experts.
The informal ‘grey’ information was not electronically accessible and was collected by
individually visiting and searching the small libraries associated with agricultural
research stations across PNG. Station specific reports were numerous but cataloguing
systems were different in each of the libraries in their level of comprehensiveness and
the individual knowledge of the local librarian proved to be the most useful “search
tool”. As a PNG scientist, I am aware that a great deal of expert knowledge has not
been captured on paper and to overcome this I developed a questionnaire and formally
interviewed agriculture and quarantine field officers (Table 2.1). The surveyed officers
were all internationally and/or locally recognised scientists with research experience in
PNG bananas and most of the grey literature that I accessed in the research station
libraries was authored by individuals from this group. Individual questioning of these
scientists thus not only allowed me to gather new information, but also to clarify any
issues or questions I may have had arising from the reports. The information gathered
from these various sources is presented as a comprehensive list of organisms
associated with banana in PNG. To be included in that list, an organism’s association
with banana had to be confirmed through a primary research paper, research report, or
through consensus with one or more of local scientists.

20
Table 2.1 Names and history on professional background of formally interviewed
agriculture and quarantine field officers
Name Research experience John Bokosou
Over ten years of collective experience in entomological research with the Department of Agriculture and Primary Industry in PNG. A Senior Research Associate with the Entomology section at the National Agriculture Research Institute. Currently based in Keravat (East New Britain).
Fred Dori
Over twenty years of collective experience in entomological research with the Department of Agriculture, Primary Industry and the Coconut Research Institute in PNG. A Senior Entomologist specialised in integrated bio control and pest control research. Is currently retired.
Elick Guaf
Over ten years of experience in food crop agronomy with the Department of Agriculture and Primary Industry in PNG. A Senior Agronomist with the National Agriculture Research Institute based in Lae, Morobe Province. Has acted in the role of Research Programme Leader on several occasions.
Tony Gunua Over ten years of experience in plant protection and pathology research with the Department of Agriculture and Primary Industry in PNG. Now a Senior Plant Pathologist with the PNG National Agriculture and Quarantine Inspection Authority. Was interviewed while on study leave at University of Queensland.
Rosa Kambuou
Over ten years of collective experience in entomological research with the Department of Agriculture and Primary Industry in PNG. A Senior Agronomist specialised in biological science and germplasm conservation. Currently based at the National Agriculture Research Institute at Laloki (Central Province). Extensive knowledge and experience on agronomy, genetic and plant protection aspects of banana research in PNG.
Annastasia Kawi
Over ten years of experience in applied technical plant protection research with the Department of Agriculture and Primary Industry in PNG. At time of interview was on study leave at University of Queensland (Gatton Campus). Now a research Entomologist with the Entomology section at the National Agriculture Research Institute.
Tim Knox and Peter Sale
Tim is a food crop Agronomist with over 20 years experience in semi-commercial farming of fruit and vegetables in PNG. Is currently the Farm manager of a semi-commercial fruit and vegetable farm at the Pacific Adventist University (PAU) located at 12 mile outside Port Moresby on road to Sogeri, Central Province). Peter has a bachelor’s degree in Agriculture from PAU and specialises in food crop agronomy. He has worked at the PAU farm for 10 years.
Kiteni Silvia Kurika
Over ten years of collective experience in entomological research with the Department of Agriculture, Primary Industry and the Coconut Research Institute in PNG. A Senior Research Associate with the Entomology section at the National Agriculture Research Institute and based in Keravat (East New Britain).
Table continued overleaf

21
Table 2.1 continued - Names and history on professional background of formally
interviewed agriculture and quarantine field officers
Name Research experience Louis Kurika
Over ten years of collective experience in entomological research with the Department of Agriculture, Primary Industry and the Coconut Research Institute in PNG. A Senior Agronomist with the National Agriculture Research Institute. Currently based in Keravat (East New Britain). Knowledgeable on island and atoll farming systems.
Gadi Ling
Over ten years of experience in food crop agronomy with the Department of Agriculture and Primary Industry in PNG. Specialised in integrated farming systems particularly the intercropping of cocoa with dominant staple crops such as banana, fruit trees and spice crops. A Senior Agronomist with the National Agriculture Research Institute. Based in Keravat (East New Britain) where he has acted in the role of Research Programme Leader on several occasions.
Meli Lolo
Over ten years of collective experience in entomological research with the Department of Agriculture, Primary Industry and the Coconut Research Institute in PNG. An Agronomist working on a European Union funded project on Atoll agriculture. Based at the National Agriculture Research Institute at Keravat (East New Britain). Extensive experience in germplasm collection on food crops such as banana and green leafy vegetables
Sim Sar
Over ten years of collective experience in entomological research with the Department of Agriculture and Primary Industry in PNG. Senior Entomologist with the PNG National Agriculture Research Institute.
David Tenakanai
Over ten years of experience in entomology research with the Department of Agriculture and Primary Industry in PNG. Now a Senior Entomologist with the PNG National Agriculture and Quarantine Inspection Authority. Was on study leave at University of Queensland at time of interview
Pest Summaries. A comprehensive pest list for PNG bananas has not been previously
generated but there have been partial lists published (Pone, 1994; ACNARS, 2003;
Kambuou, 2003). I have used the “consensus” outcomes of these publications
(specifically if a specific organism is referred to in at least two of these publications),
and the reporting of taxa by multiple local experts during interview, to generate a list
of 14 taxa which are the most commonly observed and managed organisms on banana
in PNG. Investigative research and reporting of these 14 would have been due to them
causing damage to the banana plant or the fruit bunch which would have caused a
reduction in yield and raised farmer concerns. I specifically do not use the term most
“pestiferous”, as in a number of cases the true pest status of an organism is unknown
and guilt is by association, rather than through documentation of impact. Even if some
of these organisms are not true pests, their repeated mention in the literature means

22
they will be priority organisms to be dealt with in any PRA. For these 14 taxa I provide
“pest summaries”, which detail the taxonomy, distribution, accomplished national
biological research and economic impact of these species. This process helps identify
priority areas for further research, as well as providing the type of information needed
to be supplied within a PRA. Economic impact is a crucial section in a PRA however
little information is provided for the 14 taxa and therefore this section is not included
in the pest summaries.
2.3 Results
Supporting Information. The information collected during this review is unlikely to
be gathered again due to the difficulty in accessing the majority of reports and re-
interviewing individuals (some of whom are near retirement age). Due to their
importance as supporting material for the summary information provided in this
section and their importance in the PNG banana PRA and for trade negotiation, the raw
“data” on literature and interview information are provided on a CD-Rom (attached
inside back-cover) as electronic appendices. The CD includes three appendices folders:
Appendix 1 contains pdf versions of all formally published literature; Appendix 2 has
pdf versions of all the informal ‘grey’ information, while Appendix 3 contains the
questionnaire; its format, list and technical qualifications of each interviewed officer,
and recipient answers. The appendices amount to over 150 pages of new, “lost”, or
hard to access information on PNG pests of bananas collated into a single source.
2.3.1 Pest List
There are 112 species of pathogens, arthropods, nematodes and weeds reported to be
associated with banana in PNG. The majority are fungal pathogens, Coleoptera,
Lepidoptera and nematodes. The taxa list is presented in detail in Table 2.2. Some of
the 112 species are known pests of bananas in countries other than PNG (e.g.
Bactrocera papayae Drew and Hancock (Asian papaya fruit fly) (Allwood et al.,
1999)), but their pest status on bananas in PNG is still to be determined. Diseases have
only in more recent years become identified as a problem but banana genetic diversity
may be providing resistance against many pathogens such as the serious pathogen pest
Mycosphaerella fijiensis Morelet (black Sigatoka). The inclusion of nematodes as an
organism associated with PNG bananas originates from a survey by Bridge and Page
(1982). The only nematode species they indicate to be important are Pratylenchus

23
coffeae Goodey (lesion nematode) and Radopholus similis Thorne (burrowing
nematode). Weeds have been included but these have not often been observed as being
a problem for fully established banana plants, particularly in managed gardens
(interviewed officer comments from T. Gunua and D. Tenakanai). They do however
pose problems during the establishment phase of banana plants. Such weeds include:
Mimosa invisa Mart (giant sensitive plant), Rottboellia cochinchinensis (itch grass),
Imperata cylindrical Beauv. (blady grass) and Piper aduncum Linnaeus. Mimosa
invisa and P. aduncum are invasive species in PNG (Orapa, 2006).
Table 2.2 Pathogens, arthropods, nematodes and weeds reported associated with
banana in Papua New Guinea
Pest/ Pathogen Common name Information source
Bacteria Enterobacteriales Entobacteriaceae
Erwinia chrysanthemi Burkholder et al rhizome rot (Tomlinson, 1984)
Erwinia carotovora carotovora (Pectobacterium carotovorum)
(Tomlinson, 1984)
Chromista Pythiaceae
Trachysphaera fructigena Abor & Bunting
cigar end rot Expert opinion (T Gunua)
Fungi
Agaricales Marasmiaceae
Marasmiellus inoderma Singer Marasmius semiustus Berk. & Curtis
Marasmiellus rot Expert opinion (T Gunua)
Botryosphaeriales Botryosphaeriaceae Guignardia musae Racib. banana freckle Hyde & Philemon 1994, PNG
Pest List Database (NAQIA)
Lasiodiplodia theobromae (Pat.) Griffon & Maubl. (syn. Botryodiplodia theobromae Pat.)
finger rot Expert opinion (T Gunua)
Phyllostictina musarum freckle (ACNARS, 2003), Shaw 1984,
Kokoa 1991
Capnodiales Mycosphaerellaceae
Mycosphaerella musae (Speg.) speckle, leaf speckle
PNG Pest List Database (NAQIA)
Ceratobasidiales Ceratobasidiaceae
Ceratocystis paradoxa (Dade) crown rot Expert opinion (T Gunua)
Hypocreales Nectriaceae
Fusarium oxysporum f. sp. cubense Schltdl
Panama disease, Fusarium wilt
(Shivas & Philemon, 1996), Expert opinion (T Gunua)
Incertae sedis Incertae sedis
Cordana musae (Zimmermann) Cordana leaf spot PNG Pest List Database (NAQIA)
Deightoniella torulosa Ellis Deightoniella fruit speckle
Hyde & Philemon 1994, PNG Pest List Database (NAQIA)
Ramichloridium musae (de Hoog) tropical speckle (ACNARS, 2003; Kambuou, 2003)
Uredo musae Cummins leaf rust (ACNARS, 2003)
Mycosphaerellales Mycosphaerella (Genus)
Mycosphaerella fijiensis Morelet black Sigatoka PNG Pest List Database (NAQIA)
Mycosphaerella musicola R. Leach ex J.L. Mulder
yellow Sigatoka Expert opinion (T Gunua)
Table continued overleaf

24
Table 2.2 continued - Pathogens, arthropods, nematodes and weeds reported associated
with banana in Papua New Guinea
Pest/ Pathogen Common name Information source
Phyllachorales Phyllachoraceae
Colletotrichum musae (Berk. & Curtis) anthracnose Expert opinion (T Gunua)
Phyllachora musicola Booth & Shaw black-cross PNG Pest List Database (NAQIA)
Sordariomycetes incertae sedis
Magnaporthaceae
Magnaporthe grisea Barr Johnson spot Expert opinion (T Gunua)
Virus VI: RNA Reverse Transcribing Viruses, Caulimoviridae (Fam), Badnavirus (Genus)
banana streak virus (BSV)
(Davis et al., 2000)
Arthropods Aracnida Protstigmata
Tetranychidae Tetranychus spp. spider mites (Schuhbeck, 1996; ACNARS, 2003)
Tetranychus lambi Pritchard & Baker banana spider mite
(Waterhouse, 1997)
Coleoptera
Chrysomelidae Lema papuana Jac. (Szent-Ivany & Catley, 1960) (on Musa sapientum & M. textilis)
Curculionidae Apirocalus cornutus Pasc horned polyphagous weevil
(Szent-Ivany & Barrett, 1956)
Cosmopolites sordidus (Germar) banana weevil borer
(Froggatt, 1941; Szent-Ivany & Barrett, 1956; Greve & Ismay, 1983; Thistleton & Masamdu, 1985; Issacson & King, 1987)
Odoiporus longicollis Oliver banana stem borer
Expert opinion (D Tenakanai)
Oribius sp (Bourke et al., 1973; Thistleton & Masamdu, 1985; French, 1986)
Oribius cruciatus Marshall (French, 1986)
Oribius inimicus Marshall (Greve & Ismay, 1983)
Trochorhopalus strangulatus Gyllenhal (Ismay & Dori, 1985)
Geometridae Ectropris sabulosa Walker (Lamb & Johnston, 1976)
Scarabaeidae Dermolepida nigrum Nonfrid (Szent-Ivany & Barrett, 1956; Lamb & Johnston, 1976; French, 1986)
Lepidiota spp Expert opinion (D Tenakanai)
Oryctes centaurus Sternberg Rhinoceros beetle (Smee, 1965)
Oryctes rhinoceros(Linnaeus) Asiatic Rhinoceros beetle
(Smee, 1965; Lamb & Johnston, 1976)
Papuana spp taro beetle Expert opinion (J Bokosou)
Papuana woodlarkiana laevipennis Arrow taro beetle (Froggatt, 1941; Szent-Ivany & Barrett, 1956)
Papuana woodlarkiana Montrouzier taro beetle (Lamb & Johnston, 1976)
Rhyparida sobrina Bryant Rhyparid beetle (Waterhouse, 1997)
Scapanes australis australis (Boisduval) Melanesian rhinoceros beetle
(Szent-Ivany & Barrett, 1956; Lamb & Johnston, 1976)
Scapanes australis grossepunctatus Sternberg
coconut rhinoceros beetle
(Szent-Ivany & Barrett, 1956; Bourke et al., 1973; Lamb & Johnston, 1976)
Trichogomphus semilinki Ritz (Smee, 1965)
Xyloptrupes spp. (Smee, 1965)
Diptera
Table continued overleaf

25
Table 2.2 continued - Pathogens, arthropods, nematodes and weeds reported associated
with banana in Papua New Guinea
Pest/ Pathogen Common name Information source
Muscidae Myospila argentata Walker (Greve & Ismay, 1983)
Tephritidae Bactrocera bryoniae (Tryon) Byron’s fly (Drew, 1989), PNG Pest List
Database (NAQIA)
Bactrocera dorsalis complex (CABI, 1998)
Bactrocera frauenfeldi (Schiner) mango fruit fly (Drew, 1989; Dori et al., 1993)
Bactrocera musae (Tryon) banana fruit fly (Szent-Ivany & Barrett, 1956)
Bactrocera papayae Drew & Hancock Asian papaya fruit fly
Expert opinion (S Balagawi, A Kawi, D Tenakanai)
Enoplopteron hieroglyphicum Meijere (Szent-Ivany & Barrett, 1956; Greve & Ismay, 1983)
Dermaptera
Chelisochidae Chelisoches morio (Fabricius) black earwig (Greve & Ismay, 1983; Ismay & Dori, 1985)
Hemiptera
Aleyrodidae Aleurodicus disperses Russel spiralling whitefly
Expert opinion (F Dori, R Kambuou)
Coreidae Amblypelta lutescens (Distant) banana-spotting bug
(CABI, 1998)
Miridae Prodromus spp. (Ismay & Dori, 1985)
Pentatomidae Acanthotyla Stål spp. (Greve & Ismay, 1983)
Tingidae Stephanitis typical (Distant) (Szent-Ivany & Catley, 1960) (on Musa sapientum)
Homoptera
Aphididae Pentolonia nigronervosa Coquerel banana aphid (Schuhbeck, 1996; CABI, 1998)
Rhopalosiphum maidis (Fitch) maize aphid (Waterhouse, 1997)
Coccidae Ferrisia virgata (Cockerell) (CABI, 1998)
Parasaissetia nigra (Nietner) nigra scale (CABI, 1998)
Saissetia coffea (Walker) hemispherical scale
(CABI, 1998)
Diaspididae Aonidiella aurantii (Maskell) California red scale
(CABI, 1998)
Aspidiotus destructor Sign (CABI, 1998)
Pseudaulcapsis pentagona (Targioni-Tozzetti)
(CABI, 1998)
Pseudococcidae Dysmococcus neobrevipes (Cockerell) grey pineapple mealybug
(CABI, 1998)
Lepidoptera
Amanthusiidae Taenaris dimona Hewitson Myops owl butterfly
(Greve & Ismay, 1983)
Arctidae Diacrisia papuana Rothschild (Greve & Ismay, 1983)
Geometridae Hyposidra talaca (Walker) coffee ring borer (Lamb & Johnston, 1976)
Hesperiidae Erionota thrax (Linnaeus) banana leaf roller (Issacson & King, 1987; Waterhouse & Norris, 1989; Sands et al., 1991; Sands et al., 1993)
Limocodidae Limacodidae gen.et.sp. indet slug moths, cup moths
(Ismay & Dori, 1985)
Scopelodes dinawa Bethune-Baker (Szent-Ivany, 1955; Szent-Ivany & Catley, 1960) (on Musa textilis)
Scopelodes nitens Bethune-Baker (Szent-Ivany & Catley, 1960) (on Musa sapientum)
Noctuidae Agrotis ipsilon (Huf) (CABI, 1998)
Helicoverpa armigera (Hübner) (Greve & Ismay, 1983)
Table continued overleaf

26
Table 2.2 continued - Pathogens, arthropods, nematodes and weeds reported associated
with banana in Papua New Guinea
Pest/ Pathogen Common name Information source
Tirocola plagiata Walker (Catley, 1962; Smee, 1964)
Psychidae Psychidae species Boisduval bagworm Expert opinion (J Bokosou)
Pyralidae Nacoleia sp? (Walker) (Bourke et al., 1973)
Nacoleia octasema (Meyrick) banana scab moth
(O'Connor, 1949; Szent-Ivany & Barrett, 1956)
Tirathaba rufivena Walker (Smee, 1965)
Tineidae Opogona spp. (Greve & Ismay, 1983)
Zygaenidae Artona spp. (Ismay & Dori, 1985; Thistleton & Masamdu, 1985)
Homophylotis sp. nr albicilia Hampson (Greve & Ismay, 1983)
Orthoptera
Acrididae Locusta migratoria Linnaeus locusts, migratory locust
(Anonymous, 1969)
Tettigonidae Eumossula gracilis Willemse (Bourke et al., 1973)
Segestidea defoliaria defoliaria Uvarov (Greve & Ismay, 1983)
Segestidea montana Willemse (Greve & Ismay, 1983)
Segestidea novaguineae (Brancsik) (Greve & Ismay, 1983)
Sexava spp. (Smee, 1965)
Thysanoptera
Thripidae Chaetanaphothrips signipennis (Bangall) banana rust thrips
(CABI, 1998)
Thrips hawaiiensis (Morgan) (CABI, 1998)
Nematodes Aphelenchoides besseyi Christie (Bridge & Page, 1982)
Aphelenchoides spp. (Bridge & Page, 1982)
Aphelenchus avenae Bastian (Bridge & Page, 1982)
Criconematid n.g., n. sp. (Bridge & Page, 1982)
Gracilacus spp. (Bridge & Page, 1982)
Helicotylenchus dihystera Sher (Bridge & Page, 1982)
Helicotylenchus microcephalus Sher causes root/corm necrosis
(Bridge & Page, 1982)
Helicotylenchus mucronatus Siddiqi causes root/corm necrosis
(Bridge & Page, 1982)
Helicotylenchus multicinctus Golden banana spiral nematode
(Bridge & Page, 1982)
Hoplolaimus seinhorsti Luc (Bridge & Page, 1982)
Meloidogyne incognita Chitwood root-knot nematode
(Bridge & Page, 1982)
Meloidogyne javanica Chitwood root-knot
nematode (Bridge & Page, 1982)
Paratrichodorus minor Siddiqi (Bridge & Page, 1982)
Pratylenchus coffeae Goodey lesion nematode (Bridge & Page, 1982)
Radopholus similis Thorne burrowing nematode
(Bridge & Page, 1982)
Rotylenchulus reniformis Linford & Oliveira Reniform nematode
(Bridge & Page, 1982)
Scutellonema cf. minutum Sher (Bridge & Page, 1982)
Trichodorus cylindricus Hooper (Bridge & Page, 1982)
Tylenchus spp. sensu lato (Bridge & Page, 1982)
Xiphinema elongatum Schuurmans Stekhoven & Teunissen
(Bridge & Page, 1982)
Xiphinema ensiculiferum Thorne (Bridge & Page, 1982)
Xiphinema guirani apud Lamberti & Bleue-Sacheo (Bridge & Page, 1982)
Xiphinema insigne Loos (Bridge & Page, 1982)
Xiphinema orthotenum Cohn & Sher (Bridge & Page, 1982)
Table continued overleaf

27
Table 2.2 continued - Pathogens, arthropods, nematodes and weeds reported associated
with banana in Papua New Guinea
Pest/ Pathogen Common name Information source
Weeds
Cyperales Rottboellia cochinchinensis W.D Clayton itchgrass Expert opinion (T Gunua)
Fabales Mimosa diplotricha C. Wright or Mimosa invisa Mart giant sensitive grass
Expert opinion (T Gunua)
Piperales Piper aduncum Linnaeus false kava, false Matico tree
Expert opinion (T Gunua)
Poales Imperata cylindrical Beau cogongrass, blady grass
Expert opinion (T Gunua)
Polygonales Persicaria perfoliata H. Gross mile-a-minute weed
Expert opinion (D Tenakanai)
Miscellaneous
Birds Cockatoos,
parrots, starlings Expert opinion (F Dori, E Guaf, L Kurika, G Ling, S Sar)
Flying foxes or Bats Expert opinion (F Dori, L
Kurika, G Ling, S Sar)
Achatina achatina (Linné) giant African
snail Expert opinion (L Kurika)
Rats long tailed rat Expert opinion (E Guaf)
2.3.2 Pest Summaries
Of the 112 species in Table 2.2, the following pest summaries cover fourteen of the
most commonly observed and managed species. Nine have been consistently reported
in formal and informal literature as being important species on PNG banana: Erwinia
chrysanthemi Burkholder et al (causes rhizome rot), Mycosphaerella fijiensis Morelet
(black Sigatoka), Ramichloridium musae (de Hoog) (tropical speckle), Cosmopolites
sordidus (Germar) (banana weevil borer), Bactrocera musae (Tryon) (banana fly),
Erionota thrax (Linnaeus) (banana leaf roller), Nacoleia octasema (Meyrick) (banana
scab moth), Pratylenchus coffeae Goodey (lesion nematode) and Radopholus similis
Thorne (burrowing nematode). An additional five taxa, in addition to being reported in
the literature, were also selected as being of importance by a majority of local experts
interviewed: Cordana musae (Zimmermann) (Cordana), Phyllachora musicola Booth
& Shaw (black-cross), Papuana spp. (taro beetles), Rhyparida sobrina (Bryant)
(Rhyparid beetle), and Scapanes australis grossepunctatus Sternberg (coconut
rhinoceros beetle). Please note that some figures may not have a measuring scale but
have instead been provided to show what the pest/pathogen looks like.

28
2.3.3 Pathogens - Bacteria
2.3.3.1 Erwinia chrysanthemi Burkholder et al.
A B Figure 2.2 (A) Fallen banana, cause may be due to rotting of basal pseudostem
(sometimes termed ‘tip-over’ by field officers in PNG) (Source: CTAHR
Hawaii); and (B) Rotting xylem vessels referred to as internal pseudostem
necrosis (Source: CTAHR Hawaii www.ctahr.hawaii.edu/nelsons/banana)
Taxonomic information: Order: Enterobacteriales; Family: Enterobacteriaceae;
Genus: Erwinia. (Note: information on phylum and class have been provided in Table
2.2 and therefore not provided here)
Common name(s): Erwinia rot. Symptoms of damage caused by E. chrysanthemi are
also referred to as rhizome rot, bacteria head rot, or tip-over (Figure 2.2A)
Part of banana plant affected: Roots and pseudostem
Distribution: Erwinia rot is a serious disease causing root and stem rot, it is mostly
recorded in lowland plantain crops (interviewed officer comments from T. Gunua).
The pathogen was isolated from banana samples in Laloki (Central Province)
(Tomlinson, 1984; Pone, 1994). Although present in the country the local distribution
of this pathogen is largely unknown (EPPO/CABI, 1997).
Symptoms: Erwinia chrysanthemi is a soft rot pathogen that degrades succulent and
fleshy plant organs such as roots, tubers, stem cuttings and thick leaves. It is also a
vascular wilt pathogen, colonizing the xylem vessels (Figure 2.2B) and becoming
systemic within the plant (EPPO/CABI, 1997). Rhizome rot describes the damage
caused by nematode infestation of the roots that cause the rhizomes to rot (Tomlinson,

29
1984). Symptoms also include wilting and death of leaves before fruit has ripened. The
centre of the pseudostem rots and there is some discolouration of the vascular tissues
of the outer leaf sheaths which can extend into the stalk of the fruit (Kohler et al.,
1997).
Literature from PNG on pest: Erwinia chrysanthemi was not previously recorded on
banana until an investigatory trial was done in 1983 (Tomlinson et al., 1988). Banana
test plants failed to mature due to failure of newly planted corms establishing and the
falling over of older plants. These symptoms were not previously recorded on banana.
The bacterium E. chrysanthemi was consistently isolated from diseased stem tissue of
suckers and artificial inoculation was shown to produce rotting in 12 weeks
(Tomlinson et al., 1988). There were ten banana cultivars tested and these represented
3 genomes (AA, AAA, and ABB). The AA genome cultivars were more susceptible
than the AAA and ABB genome cultivars. The disease is often not recognised due to
the many causes of poor growth of cultivated banana plants. The damage it causes such
as ‘tip-over’ is often attributed to the effects of strong windy conditions or beetle
damage on bananas (Philemon, 1986). High range in banana genetic diversity and
cultivation of more than one variety in subsistence and semi-commercial farming
systems may provide resistance between varieties lessening the damage caused
(Tomlinson et al., 1988).
Economic impact: Unknown.
2.3.4 Pathogens - Fungi
2.3.4.1 Cordana musae (Zimm.)
Figure 2.3 Cordana leaf spot on banana leaves (Source: CTAHR Hawaii web site:
www.ctahr.hawaii.edu/nelsons/banana)

30
Taxonomy information: Order: Incertae sedis; Family: Incertae sedis; Genus:
Cordana
Common name: Cordana leaf spot
Part of banana plant affected: Leaves
Distribution: Local distribution is not known. The leaf spot disease has been observed
in the lowlands (mainland or island provinces or both uncertain) attacking both
plantain and dessert varieties (interviewed officer comments from T. Gunua). Present
on the Gazelle Peninsula, East New Britain Province (ENB) (interviewed officer
comments from M. Lolo) and observed on Buka Island (Bougainville) (interviewed
officer comments from L. Kurika). Surveys are being carried out in PNG to determine
local distribution of pathogens (interviewed officer comments from R. Kambuou, E.
Guaf).
Symptoms: Large leaf spots, up to 100mm, pale brown or yellow, oval or diamond-
shaped, usually surrounded by a yellow halo (Figure 2.3). Note: Figure 2.3 does not
have a scale but is provided to show what the pathogen looks like on banana leaf. The
spots occur on and between the veins. Often entire edge of leaf may be infected with an
uneven, zigzag, yellow band separating diseased from green tissues. Infections often
occur on leaf spots caused by the black cross fungus (Phyllachora musicola), or leaf
blotches associated with Deightoniella torulosa (Kohler et al., 1997).
Literature from PNG on pest: Cordana is reported observed as being a minor
pathogen pest (ACNARS, 2003; Kambuou, 2003).
Economic impact: Unknown.

31
2.3.4.2 Mycosphaerella fijiensis Morelet
A B Figure 2.4 (A) Black Sigatoka on banana leaves (Source: DAFF Australia – AQIS);
and (B) Leaf infested with black Sigatoka has yellow transition zones between
infected and green uninfected leaf area (Source: CTAHR Hawaii
www.ctahr.hawaii.edu/nelsons/banana/)
Taxonomic information: Order: Mycosphaerellales; Genus: Mycosphaerella
Common name(s): Black Sigatoka, black leaf streak disease
Part of banana plant affected: Leaves
Distribution: Detailed distribution in PNG is uncertain, informal observation has
noted its presence in various parts of the country but formal verification is yet to be
done. Observed in the central Province (interviewed officer comments from R.
Kambuou, T. Knox, P. Sale) and also observed in Madang (King et al., 1988) but
Schuhbeck (1996) does not mention it when reporting on banana pests in Island
provinces (Manus, New Ireland, New Britain and Bougainville) (Schuhbeck, 1996).
However, symptoms have been observed on the Gazelle Peninsula (interviewed officer
comments from L. Kurika, M. Lolo) and seen in Manus (Manus Province) and
Kavieng (New Ireland Province) (interviewed officer comments from M. Lolo). Not
much seen on atolls in the Island provinces (interviewed officer comments from M.
Lolo). It has also been observed in Morobe and Madang on Kalapua banana variety
(interviewed officer comments from S. Sar). Observations however need verification
through methodological sampling and expert identification.

32
Symptoms: The first symptoms of black Sigatoka are small, chlorotic flecks that
appear on the under surface of the third and fourth fully expanded leaves. The flecks
develop into narrow rusty brown streaks (up to 2 mm wide and 20 mm long) and often
have truncated ends and sides that are sharply limited by the leaf veins. During early
stages, streaks are only visible from the lower surface. The colour of streaks intensifies
to red, brown or black, sometimes with a purple tinge. The streaks enlarge, becoming
fusiform and elliptical, and darken to give the characteristic black streaking of the
leaves (Figure 2.4A). Note: Figure 2.4 does not have a scale but is provided to show
what the pathogen looks like on banana leaf. Adjacent tissues are often water-soaked,
especially under humid conditions. Central tissues of the lesions eventually collapse.
Lesions dry to a light grey with dark brown or black borders and often have narrow,
yellow transition zones between the borders and the green leaf tissue (Figure 2.4B).
Streaks on juvenile leaves are often oval and surrounded by a yellow margin. Fruit
losses occur due to a lack of functional leaf surface area (Stansbury et al., 2000).
Literature from PNG on pest: The worst fungal cases are when black Sigatoka is
present in combination with black cross and Cordana (interviewed officer comments
from R. Kambuou). Comments from local expertise suggest that wide varietal diversity
in PNG bananas may be playing a role in neutralising the spread and thus reducing the
intensity of the disease (interviewed officer comments from R. Kambuou). Of all
common fungal diseases M. fijiensis is however the most commonly observed
(interviewed officer comments from R. Kambuou) particularly on dessert cultivars
(interviewed officer comments from T. Gunua). At the Pacific Adventist University
(PAU) Farm outside Port Moresby, Sigatoka is the main problem on Cavendish banana
variety (interviewed farm managers’ comments from T. Knox & P. Sale).
Currently collaborative efforts with NARI have enabled the planting of black Sigatoka
resistant varieties developed at NARI (Kambuou, 2001; NARI, 2005, 2007b) at the
PAU farm (interviewed farm managers’ comments from T. Knox & P. Sale).
Economic impact: Unknown.

33
2.3.4.3 Phyllachora musicola Booth & Shaw
Figure 2.5 Black-cross on underside of banana leaf of local cultivar. Photo taken at
Chanel College, Kokopo, ENB (Source: A. Mararuai)
Taxonomy information: Order: Phyllachorales; Family: Phyllachoraceae; Genus:
Phyllachora
Common name: Black-cross
Part of banana plant affected: Leaves
Distribution: Not known. Observed causing damage in Madang province (King et al.,
1988). Observed in banana plantation at Chanel College in Kokopo (ENB)
(interviewed officer comments from J. Bokosou, K Kurika) and observed on Buka
Island (interviewed officer comments from L. Kurika).
Symptoms: Black four-pointed stars, up to 60 mm long, most prominent on the lower
surface of older leaves, with long axis of the star parallel to the leaf veins (Figure 2.5).
Note: Figure 2.5 does not have a scale but is provided to show what the pathogen looks
like on banana leaf. The spots are scattered or sometimes occur in large groups. Spores
develop on the dark lines. Sometimes C. musae leaf spots are centred on the black-
cross lesions (Kohler et al., 1997).
Literature from PNG on pest: Reported observed as a minor pathogen pest
(ACNARS, 2003).
Economic impact: Unknown.

34
2.3.4.4 Ramichloridium musae de Hoog
Taxonomy information: Order: Incertae sedis; Family: Incertae sedis; Genus;
Ramichloridium
Common name: Tropical speckle
Synonyms: Periconiella musae, Veronaea musae
Part of banana plant affected: Leaves
Distribution: Local distribution not known
Symptoms: The fungus penetrates leaf stomata and, since spread in leaf is restricted to
an air chamber and the palisade cells immediately surrounding it, only a small pin
point, brown or black lesion results. These speckles are more prominent on the upper
surface; they are very numerous and become aggregated into large (up to 4 cm
diameter) circular blotches which are chlorotic amongst the necrotic speckles.
Literature from PNG on pest: Pathogen has been reported as a minor pest
(ACNARS, 2003).
Economic impact: Unknown
2.3.5 Insects & Mites - Coleoptera
2.3.5.1 Cosmopolites sordidus (Germar)
A B Figure 2.6 (A) Lifecycle and damage caused by banana weevil borer (Source: Cook
Islands Biodiversity Database); and (B) Adult banana weevil borer (Source: G
McCormack. Cook Islands Biodiversity Database-
http://entnem.ufl.edu/creatures/fruit/borers/banana_root_borer.htm)

35
Taxonomic information: Order: Coleoptera; Family: Curculionidae
Common name(s): Banana root borer, banana weevil borer
Part of banana plant affected: Roots, corm
Distribution: Detailed local distribution in PNG is not known but it has been reported
to be present in the country (Szent-Ivany & Barrett, 1956; Waterhouse, 1997).
Symptoms: Damage done by extensive tunnelling of larvae in the corm weakening the
plant (Figure 2.6A) and causing plants to fall over light winds. Complete life cycle is
about 30-40 days (Woodruff, 1969; CSIRO, 2004). Eggs are white, about 1 mm in
diameter and are laid at the leaf base. Within 5-8 days larvae hatch out. After 3-6
weeks they pupate within the stems they bore into and after a period of one week
emerge as adults (French, 2006) (Figure 2.6B). Note: Figure 2.6B does not have a scale
but is provided to show what the insect looks like.
Literature from PNG on pest: The report by Waterhouse (1997) rated C. sordidus as
one of six major invertebrate pests of agriculture in the Southern and Western Pacific
but the relative importance of the beetle in PNG was very low. Cosmopolites sordidus
has been reported as an insect pest on banana in the Highlands provinces (Gunther et
al., 2003) when it co-occurs with Papuana spp. (taro beetle). Damage is often wrongly
attributed to wind damage or Papuana species (ACNARS, 2003). No quantitative work
has been carried out to determine the level of damage carried out by C. sordidus
(ACNARS, 2003).
Economic impact: Unknown.
2.3.5.2 Papuana species
A B C

36
Figure 2.7 (A) Taro beetle burrows in damaged corm of banana sword sucker (Source:
A Mararuai); (B) Adult Papuana woodlarkiana Montrouzier (Source: A
Carmichael, PaDIL); and (C) Larvae, pupae and adult taro beetle collected at
LAES Keravat, 2005 (Source: A. Mararuai)
Taxonomy information: Order: Coleoptera; Family: Scarabaeoidae
Common name: Taro beetle
Part of banana plant affected: Corm
Distribution: Taro beetle is found in PNG. There are nine Papuana species found in
PNG but local distribution is dependent on species. While some species may overlap in
distribution not all do (Masamdu & Simbiken, 2001).
Symptoms: Adult beetles burrow tunnels into the taro corm (Figure 2.7A).
Literature from PNG on pest: Adult taro beetle are about 25 mm long and 12 mm
wide (Figure 2.7B). Note: Figure 2.7B does not have a scale but is provided to show
what the insect looks like. Males have a horn on their head. Females sometimes have a
small horn. Newly emerged beetles are brown in colour but turn black as they get older
(Carmichael et al., 2007) (Figure 2.7C). Field based surveys and trials show that
physical corm damage of 15% and above renders taro corms unmarketable in most
areas in PNG (Masamdu & Simbiken, 2001).
Banana is a primary host of adult taro beetles (Masamdu & Simbiken, 2001). Studies
however into the damage caused by taro beetles on banana requires more study
(ACNARS, 2003). Extensive research on taro beetles has been done in PNG through
national (DAL, DPI, NAQIA, NARI) and international (ACIAR, FAO, PRAP, SPC)
collaboration. Research on control and management which spans over 20 years
covering: taxonomy (MacFarlane, 1987; Masamdu & Simbiken, 2001), biology (Perry,
1977; Onwueme, 1999), damage assessments (Arura & Akus, 1988), and biological
control (Aloalii et al., 1993; Theunis & Aloali'i, 1999; Schuhbeck & Bokosou, 2006;
Simbiken, 2006); to produce field guides (Carmichael et al., 2007).
Economic impact: Unknown.

37
2.3.5.3 Rhyparida sobrina Bryant
Figure 2.8 Sketch of Rhyparidella sobrina (Bryant) (Source: Gressit (1974))
Taxonomy information: Order: Coleoptera; Family: Chrysomelidae
Common name: Rhyparid beetle
Part of banana plant affected: Leaves
Distribution: Detailed national distribution not known.
Symptoms: Adult beetles feed on young unrolled leaves and scar newly opened
fingers. Sketch diagram shows adult beetle (Figure 2.8) (Gressit, 1974). Note: Figure
2.8 does not have a scale but is provided to show what the insect looks like.
Literature from PNG on pest: Known to be present in the Markham valley (Morobe
Province) where sporadic outbreaks have occurred (Masamdu et al., 1988, 1989). In
the Markham valley it is a damaging but seasonal pest (interviewed officer comments
from J. Bokosou, E. Guaf). Pest management trials in the valley during the 1988
outbreak found practical non-chemical solutions of control through bagging bunches
using hessian and banana leaves. It also found that there was variation in varietal
susceptibility. Banana cultivars; Wamu and Garab II were severely attacked by beetles.
Kalapua I, Gampu and Amoa experienced medium to high levels of attack; Bansim,
Araduran and Mangas varieties experienced medium levels of attack; Grukrang I and
Yawa experienced slight to medium; Stifang, Rupis and Tsitsutsitsu slight; and
Yabimu suffered no damage (Masamdu et al., 1988, 1989).
Economic impact: Unknown.

38
2.3.5.4 Scapanes australis grossepunctatus Sternberg
A A B C Figure 2.9 (A) Pseudostem bore-hole caused by Scapanes australis grossepunctatus
(Source: A. Mararuai); (B) S grossepunctatus boring into banana pseudostem
(Source: A. Mararuai); and (C) Banana bunch emerging through a hole made
by S. grossepunctatus in banana pseudostem (Source: A Mararuai)
Taxonomy information: Order: Coleoptera; Family: Scarabaeoidae
Common name: Coconut rhinoceros beetle
Part of banana plant affected: Pseudostem
Distribution: Scapanes australis grossepunctatus is found on New Britain and New
Ireland islands (Bedford, 1980; Gende et al., 2006).
Symptoms: Scapanes grossepunctatus bores through the pseudostem of banana
varieties (Figure 2.9A) and beetles can often be found boring in the pseudostem
(Figure 2.9B) Note: Figure 2.9B does not have a scale but is provided to show what the
insect looks like. This causes the growing meristem to grow out through the puncture
(Figure 2.9C). The boring position may occur from any where between ground level
and the base of the leaf stems at the top of the banana plant (interviewed officer
comments from J. Bokosou, F. Dori, K. Kurika, L. Kurika).
Literature from PNG on pest: On New Britain Island, particularly on the Gazelle
Peninsula, S. grossepunctatus is a major pest of coconut (Moxon, 1988) and also
attacks banana varieties (interviewed officer comments from G. Ling). The planting of
banana gardens near existing or within coconut plantations or near breeding sites
(glyricidia and coconut groves) may be drawing Scapanes into the gardens ((Gende et
al., 2006) and interviewed officer comments from K Kurika). Considerable research

39
has been done on the biology (Bedford, 1976; Beaudoin-Olliviera et al., 2000) and
ecology (Beaudoin-Ollivier et al., 2001) of Scapanes and its control on coconut but not
on banana (Bedford, 1986; Kakul et al., 2000; Rochat et al., 2002; Rowland et al.,
2005).
Economic impact: Unknown.
2.3.6 Insects & Mites - Diptera
2.3.6.1 Bactrocera musae (Tryon)
A B C Figure 2.10 (A) Adult Bactrocera musae (Source SPC PaciFly); (B) Fruit fly larvae
feeding tracks in ripe Cavendish banana variety (Bundun, Morobe Province)
(Source: A. Mararuai); and (C) Fruit fly larvae feeding tracks in mature green
Cavendish (Kaiapit, Morobe Province) (Source: A. Mararuai)
Taxonomic information: Order: Diptera, Family: Tephritidae
Common name(s): Banana fruit fly, banana fly
Synonyms: Strumeta musae Tryon, Dacus musae (Tryon)
Part of banana plant affected: bunch fingers, fruit pulp
Distribution: Widespread and very common on mainland Papua New Guinea, where it
is as common in the Highlands as at low elevations (Leblanc et al., 2001). In early
1999, it was trapped and bred from bananas on the Gazelle (Mararuai et al., 2001). It
may have been introduced with infested bananas brought from mainland PNG as food
relief after the devastating 1994 volcanic eruption but this has not been confirmed.
Breeding populations also occur on Lihir Island (New Ireland Province). A few
specimens have been occasionally trapped on Manus, but it is not confirmed whether

40
breeding populations occur there (Drew & Romig, 1996; Leblanc et al., 2001).
Bactrocera musae is an exotic species to the Gazelle and spatial distribution is
expanding further inland and along the coastline from initial detection sites in Kokopo
(Mararuai et al., 2001).
Symptoms: Adult (Figure 2.10A) female fruit flies lay their eggs in the fruit and the
maggots develop as the fruit matures, destroying the flesh. Note: Figure 2.10A does
not have a scale but is provided to show what the insect looks like. They oviposit into
ripe banana (Figure 2.10B) and can oviposit in mature green bananas (Figure 2.10C).
Oviposition sites can provide entry for rot producing organisms. The maggots destroy
the flesh, and secondary rots, which enter through the oviposition puncture, cause pulp
breakdown. The damage is usually concentrated on bunches suffering from physical
damage or where mixed ripening of bunches is occurring because of poor plantation
management (Pinese & Piper, 1994). Oviposition stings caused by egg laying female
flies are often invisible to the untrained eye. Feeding of larval stages occurs in the fruit
pulp beneath the peel and damage is only seen when the peel is removed.
Literature from PNG on pest: Very little research has been done on the biology and
ecology of banana fly in PNG. Research on damage levels of banana fly on banana
varieties was done in Northern province including work on biology and control options
against infestation in the same area (Smith, 1977b). Smith (1977b) observed different
levels of infestation between three banana varieties: Giant Cavendish, Dwarf
Cavendish and Tui. Peel toughness and plant height were considered as the factors
influencing the variation. On the Gazelle, fruit fly larvae damage has been noticed by
banana farmers particularly in the Vudal and Vunapalding area (interviewed officer
comments from J. Bokosou, K. Kurika). Infestation in fruit on banana varieties such as
Tukuru at Vudal is higher than that observed in other areas on the Gazelle (interviewed
officer comments from J. Bokosou).
Economical impact: Unknown.

41
2.3.7 Insects & Mites - Lepidoptera
2.3.7.1 Erionata thrax (L.)
A B
C D Figure 2.11 (A) Adult Erionota thrax butterfly (Source: K Walker, PaDIL); (B)
Caterpillar feeding on leaf (Gazelle Peninsula, ENB) (Source: A Mararuai); (C)
Leaves of banana rolled up by banana skipper (Bubia, Morobe province)
(Source: A Mararuai); and (D) Pupae in leaf roll (Gazelle, ENB) (Source: A
Mararuai)
Taxonomic information: Order: Lepidoptera; Family: Hesperiidae
Common name(s): Banana leaf roller, banana skipper
Part of banana plant affected: Leaves
Distribution: Erionata thrax is a hesperiid butterfly native to SE Asia (Waterhouse &
Norris, 1989) (Figure 2.11A). Note: Figure 2.11A does not have a scale but is provided
to show what the insect looks like. It is an exotic to PNG. The butterfly was first
observed in Vanimo (Sandaun Province) on the PNG mainland in 1983 (Dori, 1988)
and spread rapidly, recently observed in ENB (Schuhbeck, 1996; Waterhouse, 1997). It
has been observed in the Sepik since 1986; Madang, Eastern Highlands and Morobe
since 1987 and Port Moresby since September 1988 (Arura et al., 1988, 1989;
Waterhouse & Norris, 1989; Sands et al., 1991). A detailed description of the
establishment and spread of E. thrax in PNG is given by Waterhouse & Norris (1989).

42
It has spread eastward to invade New Britain, Duke of York and New Ireland islands,
and possibly Bougainville (Dori, 1988; PaDIL, 2008).
Symptoms: Caterpillars roll the banana leaves (Figure 2.11B) as they feed reducing
leaf surface area (Figure 2.11C) and eventually pupate in them (Figure 2.11D).
Literature from PNG on pest: The life cycle length of E. thrax from egg to adult is
about 5-6 weeks long (French, 2006). Banana skipper is reported as a minor pest on
banana. It has been observed to be a seasonal pest in the Markham valley (Morobe
province) (interviewed officer comments from E. Guaf), in Central (interviewed officer
comments from R. Kambuou) and also on the Gazelle Peninsula (interviewed officer
comments from J. Bokosou, K. Kurika).
Research on the management of E. thrax in PNG has found that of eight studied
banana varieties; Dwarf Cavendish, Tall Cavendish, Babi Yadefana, Kuriva, Small
Kalapua, Wudupataten, and Brown River, the dwarf and tall Cavendish varieties were
the least of the eight varieties infested (Dori, 1988). Immature stages of E. thrax were
found on cultivated and wild banana species but none were found on closely related
species (e.g. E. glaucum or palms (coconuts, betel nut, oil palm or other palms) (Arura
et al., 1988). In the Markham valley (Mutzing) banana skipper damage observed on
Yawa and Tukuru banana varieties was very bad (interviewed officer comments from
F. Dori).
Management of E thrax through biological control agents has been successful (Arura et
al., 1988; Dori, 1988; Sands et al., 1988; Lubulwa & McMeniman, 1998; Bauer et al.,
2003). There were three eggs parasites (Ooencyrtus erionotae, Ooencryrtus sp. and
Anastatus sp.) and two pupal parasites (a Tachinid and a Chalcidid) studied (Arura et
al., 1988, Dori, 1988). The release of egg parasite O. erionotae (Dori, 1988) and the
larval parasite Cotesia erionotae achieved reasonable control (Sands et al., 1993;
ACNARS, 2003). Cotesia erionotae was released in the Island provinces, Highlands,
Central and Oro with F Dori from 1990-1993 (interviewed officer comments from K.
Kurika). In the Highlands parasitoids were released in Kainantu (Eastern Highlands
Province) and Mt Hagen (Western Highlands Province) (interviewed officer comments
from F. Dori).

43
Assessment of the impact of projects, particularly by ACIAR, carried out for the
control of E. thrax indicate that proper management can lead to better production
levels and subsequently the income generated from the sale of more produce (Lubulwa
& McMeniman, 1998; Waterhouse et al., 1998; Bauer et al., 2003).
Economic impact: Unknown.
2.3.7.2 Nacoleia octasema (Meyrick)
A B Figure 2.12 (A) Adult Nacoleia octasema (NAIC Kilakila, Port Moresby) (Source: A
Mararuai); and (B) N. octasema damage on banana fingers (Source: A
Mararuai)
Taxonomic information: Order: Lepidoptera, Family: Pyralidae
Common name: Banana scab moth
Part of banana plant affected: Bunch fingers, fruit peel
Distribution: The moth is found throughout PNG (O'Connor, 1949; Schuhbeck, 1996)
(Figure 2.12A). Note: Figure 2.12A does not have a scale but is provided to show what
the insect looks like. Banana scab moth (BSM) was not observed in Manus on the
islands of Lou, Ahus and Andra during surveys in early 2007 (interviewed officer
comments from K. Kurika, M. Lolo). On Buka Island some BSM damage was seen at
Nova but none at Selau and Chaba villages (Buka Island, Bougainville) (interviewed
officer comments from K. Kurika, L. Kurika). Not much BSM damage was seen in
Kavieng (interviewed officer comments from M. Lolo).
Symptoms: On the islands of New Britain, New Ireland and Bougainville the moth
causes fruit damage by scarring the peel (Wilkie et al., 1993; Schuhbeck, 1996).
Damage inhibits full maturity of the fingers (Figure 2.12B)

44
Literature from PNG on pest: Only moths on the island provinces of New Britain
and New Ireland cause cosmetic damage to banana fingers (Schuhbeck, 1996). The
moths on the island provinces have morphological features identical to populations on
the mainland but these do not cause damage on banana fingers in the same way,
particularly those on the Gazelle Peninsula (Schuhbeck, 1996; ACNARS, 2003). On
the Gazelle Peninsula, BSM damage is extensive and widespread throughout the
peninsula and common on Cavendish banana variety. Abandoned banana gardens are
breeding grounds for BSM (interviewed officer comments from J. Bokosou, F. Dori,
K. Kurika, L. Kurika, G. Ling, M. Lolo). On the Gazelle scab moth damage is seen on
diploid varieties of banana (although some are noted to be less infested) and Cavendish
but few on triploid varieties (interviewed officer comments from G. Ling). At the PAU
Farm outside Port Moresby, some BSM damage is observed but not as severe as that
seen on the Gazelle and is managed with chemical sprays (interviewed farm managers’
comments from T. Knox & P. Sale). Studies on chemical control have shown to control
damage levels and the application method has been innovatively re-designed to allow
local farmer usage (NARI, 2007b). An imitation of the proper application apparatus, a
bell-injector commonly used in Queensland banana farms was built using old umbrella
parts, rubber tubing, a small sprayer and a wooden pole (NARI, 2007b).
The scarring stops growth and maturity in the fingers resulting in a loss of crop (Wilkie
et al., 1993; Schuhbeck, 1996). The brown scarring and consequential cracks reduce
surface quality and market value. Formal economic evaluation has not been done but is
observed to have a major impact.
Economic impact: Unknown.
2.3.8 Nematodes
2.3.8.1 Pratylenchus coffeae (Zimmermann)
A B

45
Figure 2.13 (A) Female nematode (Source: http://nematode.unl.edu): and (B)
Symptoms of Pratylenchus coffeae feeding on banana root (Source: CAB Crop
Protection Compendium Module 1)
Taxonomic information: Order: Tylenchida; Family: Pratylenchidae: Genus:
Pratylenchus
Common name(s): Banana root nematode, lesion nematode
Synonyms: Pratylenchus musicola, Tylenchus coffeae Zimmermann, 1898, Tylenchus
mahogani Cobb, 1920, Anguillulina mahogani (Cobb, 1920) Goodey, 1932,
Pratylenchus mahogani (Cobb, 1920) Filipjev, 1936, Tylenchus musicola Cobb, 1919
Part of banana plant affected: Roots
Distribution: A survey by Bridge et al (1982) indicated that P. coffea was present in
surveyed locations in the PNG provinces of Southern Highlands Western Highlands,
East Sepik, Morobe and East New Britain (Bridge & Page, 1982); but current
distribution is not known.
Symptoms: The lesion nematode causes severe damage to roots
Literature from PNG on pest: Results from the survey show that the most important
of nematodes is the lesion nematode, Pratylenchus coffeae (Zimmermann) (Bridge &
Page, 1982) (Figure 2.13A). Note: Figure 2.13A does not have a scale but is provided
to show what the nematode looks like. It was found causing severe root damage
(Figure 2.13B) at most sample sites in the five surveyed provinces. Current
observations in the field indicate that nematode damage has an impact on plants
particularly when there is a build up of nematode population particularly in long-term
banana gardens or if an infested stand of bananas remains for a while ((Fooks, 1989,
2002) comments from R Kambuou, and farm managers T. Knox & P. Sale).
Economic impact: Unknown

46
2.3.8.2 Radopholus similis Thorne
A B C Figure 2.14 (A) Illustration of burrowing nematode (Source:
http://plpnemweb.ucdavis.edu/Nemaplex); (B) Damage to banana roots caused by the
burrowing nematode (Source: http://www.ctahr.hawaii.edu); and (C) Toppled banana
(black head) (Source: http://www.ctahr.hawaii.edu/nelsons/banana)
Taxonomy: Order: Tylenchida, Family: Pratylenchidae, Genus: Radopholus
Common name(s): Burrowing nematode, root rot nematode (Figure 2.14A). Note:
Figure 2.14A does not have a scale but is provided to show what the nematode looks
like.
Part of banana plant affected: Roots, corm
Distribution: Survey by Bridge et al (1982) indicate that Radopholus similis Thorne
was present in the five provinces they surveyed (Southern Highlands, Western
Highlands, East Sepik, Morobe and East New Britain) (Bridge & Page, 1982;
Philemon, 1986). Current distribution however is not known.
Symptoms: Radopholus similis causes Radopholus root rot. Symptoms: reddish-brown
or black rots (Figure 2.14B), often several cm long, on the root, sometimes with cracks.
The areas of rot are at first outside the vascular tissues; later, they spread throughout
the root, causing total decay. As the nematode burrows into the corm, black spots with
red margins develop. These rots, known as ‘blackheads’, may extend up to 20 mm into
the corms. Plants are weakened by the root attack and are readily blown over during
storms (Kohler et al., 1997) (Figure 2.14C).

47
Literature from PNG on pest: The damage caused by R. similis in PNG was reported
to be not as damaging as P. coffea (Bridge & Page, 1982). It is not as widespread or
serious a pest in survey locations as P. coffeae (Bridge & Page, 1982; Philemon, 1986).
Economic impact: Unknown.
2.4 Discussion
Formal publications on banana pests in PNG are few. Possibly the first formal
publication was on the basic biology, distribution and management of banana scab
moth (O'Connor, 1949). Szent-Ivany & Barrett (1956) shortly after produced the first
list on banana pests recording 14 insect pests from banana, subsequently adding to that
four years later (Szent-Ivany & Catley, 1960). These 14 are not the 14 ‘taxa’ identified
in this thesis. Subsequent pest lists have been provided by Pone (1994), Schuhbeck
(1996), Waterhouse (1997), ACNARS (2003) and Kambuou (2003). The quality of
these lists vary, some being based on local surveys (e.g. Schuhbeck, Kambuou), while
others have been external reviews (e.g. Waterhouse 1997). None are comprehensive;
each dealing with different taxa groups (e.g. insects or pathogens) while others are
spatially restricted to a certain part of the country. The list of 112 organisms provided
in this chapter (Table 2.2) is thus the first and most comprehensive list to date of all
organisms associated with banana in PNG.
While searching through publications I have noted that publications may list species as
being minor or major pests on banana but most lack the assessment methodologies
which were used to classify those organisms as pests. The nematodes are a group
example. The commonly cited Bridge & Page (1982) survey report lists 14 nematode
species as being important on PNG banana but when Pone (1994) refers to that report
he lists 24 species. Another more improved and recent example is the ACNARS (2003)
report that mentions four nematode genera of importance present in the country but
specifies that damage assessment studies are needed to determine if they cause
significant problems to banana. The reason why the species are included as pests or
non-pests in PNG is also not stated. The economic impacts of many of the organisms
listed in Table 2.2 are not documented and I conclude that formal verification of pest
status is required for most species. Crop impact studies for even major crop pests in

48
PNG is generally lacking, but can be achieved (Wesis et al., in press). Thus,
economical impact is largely unknown for all groups of pathogens and pests discussed.
There is also the issue on pest status of a recently introduced or non-endemic pest of
banana. When non-endemic banana pests are confirmed as occurring in PNG, their pest
status based on known importance in other countries cannot be used as a measure of
their current impact in PNG. Two severe pathogen diseases on banana, Fusarium
oxysporum f. sp. cubense Schltdl (Fusarium wilt) and banana streak virus (BSV),
illustrate this point. Fusarium oxysporum f. sp. cubense was detected in PNG in the
mid-1990s at three locations along the PNG/Indonesian border (Shivas & Philemon,
1996; Davis, 2004; Kokoa, 2006) and surveys are currently being done to find out its
local distribution. However, despite now being on the national pest list, its limited
distribution means it is not a current production problem for most local growers. BSV
has been detected in Alotau (in 1997, Milne Bay Province), while streak symptoms
were found on many banana cultivars (AAB, Mysore subgroup) on the coast of
Western Province in June 1999. In May 2000, BSV-Mys was confirmed from a
Mysore cultivar at Green River (Sandaun Province), BSV-Onne from a Mysore
suspected cultivar at Telefomin (Sandaun Province) and BSV-GF from Pisang Raja
(AAB) at Niokamban (Western Province) (Davis et al., 2000). Davis et al (2000)
suggest that BSV may be widespread in PNG, but observations of field officers suggest
it is not causing a concern in production systems for growers. For the PRA process,
however, all potential pests need to be identified.
Kambuou (2003) reported that the main pests of banana in PNG were: B. musae, E.
thrax, N. octasema, and the banana leaf spot complex [C. musae, M. fijiensis, P.
musicola and R. musae] (Kambuou, 2003). This list, prior to that produced in this
chapter, was the most current for PNG banana pests. The author of that paper, Rosa
Kambuou, is an internationally respected banana specialist and one of PNG’s most
senior agricultural researchers. Her list might thus be considered as having some
authority. If so, then it is these species which are most likely to impact on commodity
production and of greatest importance in managing if PNG bananas are to be exported.
Of the seven, the biology, ecology and control of E. thrax and N. octasema have been
documented. Sporadic outbreaks of E. thrax caterpillars may occur due to seasonal
weather conditions reducing foliage cover, but this occurs only occasionally and the
species is regarded and known as a seasonal pest (interviewed officer comments from

49
J. Bokosou, E. Guaf, R. Kambuou, K. Kurika, D. Tenakanai). Nacoleia octasema is
found throughout the country, but only the moths found on the Gazelle Peninsula
(ENB) cause economic levels of cosmetic damage to banana fingers. Control options
have been studied and applied practically, and the application method adapted for local
use (NARI, 2007b). With the pathogens, M. fijiensis in combination with other
pathogens can be quite damaging (interviewed officer comments from R. Kambuou),
but research is currently multiplying resistant banana varieties for cultivation and this
approach is showing success (NARI, 2005, 2007a). Surveys are also being done to
determine the local distribution of the pathogens.
For only a few species in my list of 14 is there sufficient information available to meet
all the requirements of a PNG banana PRA. It is not possible within the course of one
PhD to gather all this information, so a focus upon just one pest, as a case study, was
considered an appropriate way forward. Bactrocera musae, the banana fly, was
consistently regarded in the literature and by interview participants as the major insect
pest of bananas in PNG, yet there is very little available information on this pest. For
this reason banana fly was chosen as the focus for the remainder of the thesis. It would
also be the major pest of concern for a PNG banana PRA because B. musae affects the
fruit of banana, the commodity to be traded. Banana fly is considered a common fruit
fly species on the mainland of PNG (Tenakanai, 1996; Clarke et al., 2004). However
there is little knowledge and research done on the biology and ecology of this fly in
PNG. There is no documentation on local levels of abundance which will be important
because there are different cropping systems in PNG (Bourke et al., 1998; Bourke,
2001). In the 1970s, Stuart Smith published three papers on banana fly in Northern
(now Oro) Province detailing results of a trapping programme which show seasonal
changes in abundance in that area (Smith, 1977a). He also carried out research on the
fly’s biology and methods on how to control infestation on its host (Smith, 1977b).
While these provide a stepping stone from which to continue research, they are
insufficient for developing a national management perspective appropriate for
international commodity trade for the pest.
2.5 Thesis Progress
The aim of this thesis is to provide information for the development of a pest risk
analysis (PRA) for PNG bananas. There are three steps in the PRA (Figure 2.16) and

50
this chapter has carried out an essential part of the first step in identifying pests
associated with the commodity. I have identified 112 organisms associated with
banana in PNG and in doing so I have also identified a smaller group of 14 organisms
that are commonly identified as needing management. Of these 14, Bactrocera musae
is recorded and regarded as the major pest of banana in PNG but there is limited
knowledge available on the species to further progress through the PRA process. In
subsequent chapters I focus on this fly so as to provide information required by the
PRA. Specifically, in the next chapter, I provide detailed analysis of the local
distribution of banana fly in PNG. Outcomes of that research are pertinent for both the
“Risk Assessment” and “Risk Management” steps of the PRA.
RISK MANAGEMENTReview risk management options (e.g. import
restrictions, invasion tactics)
RISK ASSESSMENTAssess (a) the risk of entry; (b) the risk of establishment; and (c)
the risk of economic and other impacts
RISK INITIATIONIdentify pest(s) and pathways of concern, and consider whether the pest should
be considered for risk analysis in relation to the area at risk
3
2
1
Review of banana (Musa spp.)
pests in Papua New Guinea [Chapter 2]
RISK MANAGEMENTReview risk management options (e.g. import
restrictions, invasion tactics)
RISK ASSESSMENTAssess (a) the risk of entry; (b) the risk of establishment; and (c)
the risk of economic and other impacts
RISK INITIATIONIdentify pest(s) and pathways of concern, and consider whether the pest should
be considered for risk analysis in relation to the area at risk
3
2
1
Review of banana (Musa spp.)
pests in Papua New Guinea [Chapter 2]
RISK MANAGEMENTReview risk management options (e.g. import
restrictions, invasion tactics)
RISK ASSESSMENTAssess (a) the risk of entry; (b) the risk of establishment; and (c)
the risk of economic and other impacts
RISK INITIATIONIdentify pest(s) and pathways of concern, and consider whether the pest should
be considered for risk analysis in relation to the area at risk
3
2
1
RISK MANAGEMENTReview risk management options (e.g. import
restrictions, invasion tactics)
RISK ASSESSMENTAssess (a) the risk of entry; (b) the risk of establishment; and (c)
the risk of economic and other impacts
RISK INITIATIONIdentify pest(s) and pathways of concern, and consider whether the pest should
be considered for risk analysis in relation to the area at risk
3
2
1
Review of banana (Musa spp.)
pests in Papua New Guinea [Chapter 2]
Figure 2.15 Pest Risk Analysis process; with arrow indicating which step in process
information generated in this thesis has been collected for

51
Chapter 3. Distribution and abundance of five economically important fruit fly species in Papua New Guinea
3.1 Introduction
Accurate information on the distribution and seasonal abundance of fruit flies, and
knowing and understanding the factors which influence distribution and abundance, is
a key element in the strategies employed in fruit fly management and market access
(Allwood, 1996a; Leweniqila et al., 1996; Sutherst et al., 2000; Yonow et al., 2004;
Dhillon et al., 2005; Khan et al., 2005). Growers producing commodities in areas free
of fruit flies, or with very low fruit fly abundance, receive favourable treatment with
respect to market access (Maelzer et al., 2004; Follett & Neven, 2006). Even in areas
where flies are endemic and normally abundant, seasonal periods of low fruit fly
numbers can be used as part of a systems approach management strategy for market
access (IPPC, 2002c). For example, both citrus and strawberries are exported from
Queensland during winter with access to interstate markets based, in part, on a winter
window of low pest pressure (H. Gu, Queensland Primary Industries & Fisheries, pers.
comm.).
Fruit fly distribution is influenced by abiotic and biotic environmental factors such as
temperature, rainfall and the availability of hosts (Christenson & Foote, 1960;
Bateman, 1972; Fletcher, 1987). A suite of biological and environmental factors also
influence fruit fly abundance, including daily nutritional requirements (Aluja et al.,
2001; Carey et al., 2002), light intensity and day length (Brieze-Stegeman et al., 1978;
Brevault & Quilici, 2000), natural enemies (Duan & Messing, 1997; Bautista et al.,
2004), and fly movement (Kovaleski et al., 1999; Bonizzoni et al., 2004). The relative
importance of these factors may, however, vary in different regions of the world, for
example between tropical and temperate regions (Muthuthantri, 2008).
In the Pacific, research indicates that the major factor influencing fruit fly distribution
and abundance is the spatial and seasonal availability of susceptible hosts (Leweniqila
et al., 1996; Vueti et al., 1996). Most Pacific Island Countries and Territories (PICTs)
are, however, small to medium sized islands, islets and atolls which have little
variation in climatic attributes (e.g. temperature, rainfall) because altitude variation is

52
minimal and climate is largely dominated by maritime influence. In contrast, with
regions which vary significantly in climate Papua New Guinea (PNG) in the Western
Pacific has a much larger land mass than neighbouring PICTs with a significantly
greater altitudinal and tropical climate range. How these factors influence fruit fly
distribution in this particular environmental setting is largely unknown.
The geography of PNG is highly diverse and influences the tropical weather patterns
experienced in the country. Annual rainfall levels in PNG range from 1000 to >5000
mm, with variation due primarily to topographical variation (McGregor, 1989;
Bellamy & McAlpine, 1995; Macfarlane, 2000). Temperature ranges in the country are
largely affected by altitude. In general, lowland coastal areas have warm temperatures
that range from 23->35 °C, in comparison to cooler temperatures of 0-28 °C
experienced at altitudes above 1200 m.a.s.l in the Highland provinces (Macfarlane,
2000). The combined influence of geography, the southeast trade winds and the
northwest monsoon, produces seasonal patterns which are variable in their timing and
intensity in different parts of the country. For example, the wet season occurs at two
different times based on locality. Most of the country experiences maximum monthly
rainfall from January to April, but mainland areas in Milne Bay, much of Gulf
Province, the coastal region and Finschhafen area of Morobe Province, the southern
coast of New Britain, and the southern most tip of the main island of Bougainville
experience their major rainfall from May to August (Macfarlane, 2000).
Geographic diversity also contributes toward the diversity in local cropping systems
influencing the cultivation and availability of susceptible hosts. The influence of this
type of tropical cropping system on population dynamics is also largely unknown.
There are 287 discrete agricultural systems identified in PNG, each practicing a
particular cultivation system and growing a certain range of crops (Bourke et al.,
1998). A PNG specific geographical dataset known as the Papua New Guinea
Resource Information System (PNGRIS) (Bellamy & McAlpine, 1995) provides GIS
surface layer maps of these cropping systems, as well as other site specific information
such as elevation and annual rainfall which, if combined with fruit fly abundance data,
allows questions to be asked about the impact of these variables on fly distribution.
PNG’s geography has an influence the regional biodiversity of the country’s fruit fly
fauna (Michaux & White, 1999; Clarke et al., 2004; Kitching et al., 2004), but whether
climatic factors and/or cultivation practices influence the local distribution and

53
abundance of fruit fly species is largely unknown. Presence/absence distribution
records at the provincial (i.e. local government) level for all PNG Dacinae have been
published (Clarke et al., 2004), but nothing is known of the fine scale spatial
distribution and temporal abundance of fruit flies within each PNG province.
The general lack or absence of formal scientific documentation on pest fruit fly species
and their susceptible hosts in PNG is a factor which will contribute to reducing the
likelihood of market access acquirement for susceptible commodities (Allwood,
1996a). Many of the potential fresh fruit and vegetables commodities which could be
exported from PNG, including banana, are hosts of the fruit flies dealt with in this
chapter. In the management of economically important species for which detailed
ecological information is lacking readily available weather or geographic data on
factors such as rainfall and temperature, help contribute to providing an initial and
general perspective of pest distribution and abundance (Carey, 1996; Yonow et al.,
2004). Given the diverse geographical environment, the different agricultural systems,
and the large fruit fly fauna, PNG is also an ideal area to investigate the influence and
impact that tropical environmental factors may have on the distribution and abundance
of pest fruit fly species. The objectives of this study are thus twofold. Firstly, as an aid
to local pest management and market access, to detail the fine-scale distribution and
abundance of Bactrocera musae (Tryon) (banana fly) as well as four other pest fruit fly
species in major cropping regions of PNG. Secondly, so as to further our general
knowledge of fruit flies, to determine factors influencing the distributions of those
flies. In the context of the overall thesis, this chapter includes information on the main
target species: banana fly. The inclusion of the other fly species, while not directly
related to banana market access, provides an understanding on how these
environmental factors influence the population dynamics of other economically
important fruit fly species. Information of which is limited in tropical fruit fly ecology
and management.
3.2 Materials and methods
3.2.1 Trapping
The fruit flies used in this study were collected as part of fruit fly trapping surveys
carried out in PNG between June 1998 and September 2001. While not enrolled as a
student at the time, I was personally involved in that trapping program as a scientist,

54
with responsibility for the component of the trapping network covering the PNG island
provinces. In total, 167 pairs of cue-lure (4(para-acetophenyl)-2-butanone) and methyl
eugenol (4-allyl-1,2-dimethoxybenzene) baited modified Steiner traps were used.
Traps were hung, wherever possible, in trees at approximately 1.8m above the ground
and were cleared of flies every 2-3 weeks. Samples were subsequently forwarded to
Griffith University, Brisbane, where they were identified to species level using the
taxonomic keys in Drew (1989). Further details of the traps and trapping program are
presented in Leblanc et al. (2001) and Clarke et al. (2004), which are supplied as
Appendices 1 and 2. I am a co-author of both of these publications.
For this chapter, detailed analysis is undertaken for five economically important fruit
fly species: Bactrocera bryoniae (Tryon), B. cucurbitae (Coquillett), B. frauenfeldi
(Schiner), B. musae and B. umbrosa (Fabricius). Only a subset of the total trapping
data-base is used for this chapter. Trap sites providing data were chosen based on the
duration that traps were serviced. Only traps serviced for a minimum time of eight to
nine months were included to determine population fluctuations over a period of time.
These traps occurred in districts which were consistently serviced by more than one
trap so as to allow comparative analysis within an area. This reduced the study to 69
trap sites in four selected areas of PNG (Table 3.1 & Figure 3.1): (i) seventeen traps in
the Highlands Provinces (six in Eastern Highlands [EHP], three in Chimbu, four in
Western Highlands [WHP], two in Enga and two in Southern Highlands [SHP]); (ii)
sixteen traps in Morobe Province; (iii) twenty-one in Central Province; and (iv) fifteen
traps on the Gazelle Peninsula in East New Britain (ENB). Even with reduced use of
the total available data set, the tallied recordings of 891,970 trapped fruit fly specimens
are analysed here. The trap site locations cover a broad range of agricultural systems
and environment types (Bellamy & McAlpine, 1995; Bourke et al., 1998; NARI,
2008). Excluded data provided little more additional information than already provided
in Clarke et al. (2004).
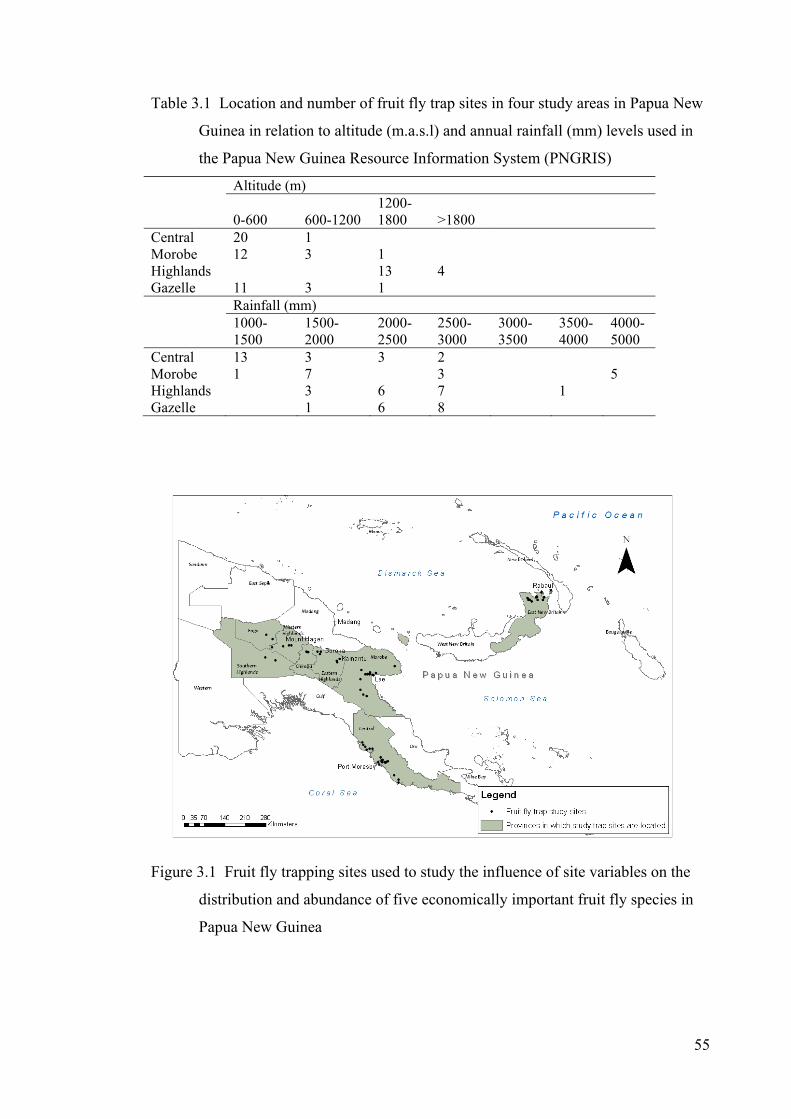
55
Table 3.1 Location and number of fruit fly trap sites in four study areas in Papua New
Guinea in relation to altitude (m.a.s.l) and annual rainfall (mm) levels used in
the Papua New Guinea Resource Information System (PNGRIS)
Altitude (m)
0-600 600-1200 1200-1800 >1800
Central 20 1 Morobe 12 3 1 Highlands 13 4 Gazelle 11 3 1 Rainfall (mm)
1000-1500
1500-2000
2000-2500
2500-3000
3000-3500
3500-4000
4000-5000
Central 13 3 3 2 Morobe 1 7 3 5 Highlands 3 6 7 1 Gazelle 1 6 8
Figure 3.1 Fruit fly trapping sites used to study the influence of site variables on the
distribution and abundance of five economically important fruit fly species in
Papua New Guinea

56
3.2.2 Databases and analysis
The geographical dataset PNGRIS (Bellamy & McAlpine, 1995) was used to supply
GIS surface layers for rainfall, altitude and land use to create respective maps in
ArcGIS 9.0. Categorisation of altitude ranges and annual rainfall used in PNGRIS were
used to map and analyse the trapped fly abundance for the five studied fruit fly species
collected from traps at different levels.
Note. Altitude ranges in map legend are presented as categorised in the PNGRIS
dataset. Majority of trap sites were located from 0-1800m and therefore ranges above
1800m were grouped and similarly shaded for easier presentation. The annual rainfall
is similarly done but this time distinct shading has been given for ranges 1000-
2500mm. Rainfall ranges above 2500mm are shaded in various shades of blue.
The landuse surface layers had information on cropping systems such as the type of
agricultural crops cultivated in an area, how often they were planted, and how
commonly they were consumed, etc. Banana is one of four major food crops cultivated
in PNG and therefore there was substantial information on the crop to be able to query
PNGRIS to providing information on banana grown as a dominant (crop consumed on
a weekly basis) or subdominant (crop consumed at a lesser rate than weekly) crop. It
was therefore feasible to create a map of the spatial distribution of banana in ArcGIS
and this was labelled and referred to as a map indicating the importance of banana (the
cultivated and staple food crop) in an area. This map was used to determine the
relationship between fly abundance and host distribution. I was not able to get detailed
data on the importance and distribution of susceptible hosts for the other fly species
studied in PNGRIS. Consequently it was not possible to investigate the effect of host
distribution on the distribution of those species. For example, chillie is a host for B.
bryoniae but a spice crop not cultivated extensively and not as a staple crop like
banana. The PNGRIS surveys collected information basically on staple crops and
grouped other cultivated plants under a collective label and therefore I was not able to
query PNGRIS to extract information specifically on chillie to be able to map the crops
distribution across PNG in ArcGIS.
The fruit fly trapping data set was summarised into mean monthly trap catches and was
similarly spatially mapped within ArcGIS. The trap catch per month for each trap in an
area were totalled and monthly averages calculated for each study fruit fly species for

57
that area. These mean abundance data were also plotted over time to generate fruit fly
seasonal phenology curves for all five study species. Linear and multiple regression,
and descriptive statistics were done in SPSS Vs 15.0 to study the relationships between
fruit fly distribution and abundance and altitude and rainfall. Linear regression was
also done for B. musae abundance versus the relative importance of banana at trapping
sites.
3.2.3 Fly species
Bactrocera bryoniae (Bryon’s fruit fly) (Figure 3.2A) is a pest of capsicums and
chillies and is widely distributed across PNG except for Bougainville and Manus
(Clarke et al., 2004). Leblanc et al. (2001) regard it as one of the few economic species
common in the Highlands (along with B. musae and Bactrocera papayae Drew and
Hancock). Bactrocera bryoniae is widely distributed down the east coast of Australia,
but is not regarded as a pest (Raghu & Clarke, 2001).
Bactrocera cucurbitae (melon fly) (Figure 3.2B) is native to tropical Asia and is an
introduced fruit fly species in PNG. The fly may have been introduced into the country
during the Second World War and has now spread and is reported to be present in
virtually all provinces, but it is absent from Manus and it was collected from Western
Highlands Province only once. It is less common in the Highlands than at lower
altitude (Leblanc et al., 2001). The fly is a major pest of cucurbits and has been
extensively researched in Hawaii, where it is also a long established invader (Stark,
1995; Vargas et al., 1997; Vargas et al., 2000a).
Bactrocera frauenfeldi (mango fly) (Figure 3.2C) is a widespread, polyphagous pest
species throughout PNG and very common throughout lowland areas. It has been
recorded in all provinces, but only a few specimens have been collected in the
Highlands where it is not considered economically important (Leblanc et al., 2001).
Bactrocera frauenfeldi is a species of major importance to PNG and other Pacific
nations (Allwood & Leblanc, 1996). The fly is invasive in Australia after having been
introduced via Cape York in the mid 1970s (Drew & Romig, 1996).
Bactrocera musae (banana fly) (Figure 3.2D) is reported to be widespread and
common on mainland PNG, where it is as common in the Highlands as at low
elevations. It is a primary insect pest of bananas in PNG, causing up to 20% fruit

58
infestation (Leblanc et al., 2001). In early 1999 it was trapped and bred from bananas
on the Gazelle Peninsula, ENB (Mararuai et al., 2001). Breeding populations also
occur on Lihir Island (New Ireland Province). A few specimens have been occasionally
trapped on Manus, but it is not confirmed whether breeding populations occur there
(Leblanc et al., 2001). Bactrocera musae’s endemic range also includes far-north
Queensland (Drew, 1989).
Bactrocera umbrosa (breadfruit fly) (Figure 3.2E) is reported to be widespread and
very common in PNG, but much less abundant in the Highlands (Clarke et al., 2004). It
has been collected in all provinces except Gulf, Enga and Southern Highlands (Leblanc
et al., 2001); it undoubtedly occurs in these provinces as well. Bactrocera umbrosa
breeds in Artocarpus species (Allwood et al., 1999) and is one the few flies whose
native range, like that of its host (Walter & Sam, 2002), extends west from the Pacific
across Wallace’s line into south-east Asia.
A B
C D

59
Figure 3.2 Fruit fly study species: (A) Bactrocera bryoniae (Tryon) (approximate
magnification x5), (B) Bactrocera cucurbitae (Coquillett) (x6), (C) Bactrocera
frauenfeldi (Schiner) (x6), (D) Bactrocera musae (Tryon) (x6), and (E)
Bactrocera umbrosa (Fabricius) (x5).
3.3 Results
3.3.1 General Patterns
Species specific patterns (developed below) in fruit fly abundance were apparent but
(with the exception of B. musae) in general numbers declined above 1200m (Table
3.2). Trapping results show that most B. bryoniae were trapped in Morobe (45% of
trapped flies) and Central (32%) provinces, B. cucurbitae was most abundant in
Central Province (64%), B. frauenfeldi (65%) and B. musae (75%) were both most
common in Morobe Province, while B. umbrosa was equally abundant on the Gazelle
(45%) and in Morobe (42%). In many cases these figures do not support the general
distribution and abundance statements supplied in Leblanc et al. (2001).
The spatial population distributions and seasonal abundances of four of the five fruit
fly species were significantly influenced by altitude and annual rainfall at the “whole
country” or national level. The influence was however variable (Table 3.3); detailed
results for each fly are reported in Sections 3.3.2 to 3.3.6. The spatial distribution and
seasonal abundance of B. musae was not significantly influenced by either altitude or
rainfall (Table 3.3). Multiple regression analysis of the effect of altitude and rainfall on
distribution and abundance also show a weak relationship (Table 3.4). Seasonal
abundance of B. bryoniae, B. cucurbitae, B. frauenfeldi and B. umbrosa at the national
level suggest that fluctuations in abundance were related to the onset and duration of
E

60
the wet and/or dry season in PNG (detailed results for each fly are in Sections 3.3.2 to
3.3.6).
Agricultural cropping systems are diverse in the country but in comparing the spatial
distribution of B. musae and B. umbrosa against the cultivated spatial distribution of
their major host, results show a close relationship between the two (see details Section
3.3.5 (Figure 3.15) and 3.3.6 (Figure 3.19) respectively). For both species abundance
was high in areas where their respective hosts are an important food source (see details
Section 3.3.5 and 3.3.6 respectively). Information on the spatial distribution of
susceptible hosts for B. bryoniae and B. cucurbitae was very restricted in PNGRIS and
so not amenable to plotting. Similarly, B. frauenfeldi is a polyphagous fly with many
potential hosts and not suitable for studying if cropping system and fly abundance
visually correlate; it was also therefore not done.
Table 3.2 The monthly trap catch (June 1998 – September 2001) of five economically
important fruit fly Bactrocera species in four study areas in Papua New Guinea
Mean monthly trap catch (± SE)
Fly species Highlands Morobe Central Gazelle All regions
B. bryoniae 14.3 ± 4.6 118.9 ± 34.3 86.0 ± 15.7 49.5 ± 13.5 68.0 ± 10.6
B. cucurbitae 3.9 ± 2.0 9.2 ± 8.4 119.5 ± 43.8 67.7 ± 25.0 54.2 ± 15.5
B. frauenfeldi 0.1 ± 0.1 714.5 ± 236.9 139.2 ± 44.6 326.8 ± 78.8 279.1 ± 66.0
B. musae 123.6 ± 42.0 505.8 ± 177.8 44.0 ± 20.3 12.6 ± 11.8 163.9 ± 48.0
B. umbrosa 0.3 ± 0.1 83.9 ± 37.5 17.8 ± 11.1 131.3 ± 28.2 53.5 ± 12.5

61
Table 3.3 Linear regression analysis of the influence of: (i) altitude or (ii) rainfall on
the abundance of five Bactrocera species within and across four study areas in
Papua New Guinea, or (iii) banana in local cropping systems for B. musae only.
Results are R2 values (and probability values in brackets). Note: Analysis not
applicable (NA) for altitude in Central because 20 of 21 traps are located at one
altitude level, nor for B.musae on the Gazelle due to inconsistent and sporadic
sampling (* = P≤0.05).
Table 3.4 Multiple regression analysis of the influence of both altitude and rainfall on
fly distribution and abundance (P≤0.05) (* = P≤0.05).
Fly species Central Gazelle Highlands Morobe Papua New Guinea
Altitude (m.a.s.l)
B. bryoniae NA +0.10 (0.24) -0.11 (0.20) +0.01 (0.77) -0.08 (0.02)*
B. cucurbitae NA -0.16 (0.13) +0.04 (0.44) -0.02 (0.61) -0.08 (0.02)*
B. frauenfeldi NA -0.34 (0.02)* +0.20 (0.07) -0.14 (0.16) -0.12 (0.003)*
B. musae NA NA -0.16 (0.11) -0.13 (0.18) -0.02 (0.28)
B. umbrosa NA -0.28 (0.05)* -0.13 (0.16) -0.08 (0.28) -0.16 (0.004)*
Annual Rainfall (mm)
B. bryoniae -0.00 (0.79) +0.19 (0.11) -0.21 (0.06) -0.19 (0.095) -0.06 (0.04)*
B. cucurbitae -0.01 (0.62) -0.16 (0.14) -0.08 (0.28) -0.13 (0.17) -0.08 (0.02)*
B. frauenfeldi -0.01 (0.76) -0.02 (0.60) +0.04 (0.46) +0.25 (0.05) +0.13 (0.002)*
B. musae -0.02 (0.53) NA -0.11 (0.20) -0.16 (0.12) -0.01 (0.35)
B. umbrosa -0.01 (0.71) -0.33 (0.02)* +0.001 (0.89) +0.31 (0.03)* +0.10 (0.01)*
Banana importance in local cropping system
B. musae +0.10 (0.16) +0.06 (0.39) +0.06 (0.34) +0.19 (0.09) +0.08 (0.02)*
Fly species Central Gazelle Highlands Morobe Papua New Guinea
Altitude (m.a.s.l) and Annual rainfall (mm)
B. bryoniae 0.05 (0.63) 0.20 (0.26) 0.21 (0.18) 0.19 (0.25) 0.11 (0.02)*
B. cucurbitae 0.02 (0.80) 0.22 (0.23) 0.28 (0.10) 0.15 (0.35) 0.13 (0.01)*
B. frauenfeldi 0.02 (0.81) 0.36 (0.07) 0.21 (0.19) 0.39 (0.04)* 0.36 (0.00)*
B. musae 0.02 (0.80) NA 0.18 (0.26) 0.28 (0.11) 0.02 (0.45)
B. umbrosa 0.01 (0.92) 0.41 (0.04)* 0.23 (0.17) 0.40 (0.04)* 0.32 (0.00*)

62
3.3.2 Bactrocera bryoniae (Tryon)
Bactrocera bryoniae was most abundant in Morobe Province (45% of total sample)
and least common in the Highlands (4% of sample) (Table 3.2). (Note: percentage
figures shown here refer mean monthly trap abundance results presented in Table 3.2
were calculated). At four of the 69 trap sites no B. bryoniae were caught (Figure 3.4),
but the fly was present in the Highlands provinces, Morobe, Central and on the Gazelle
Peninsula. As previously reported (Clarke et al. 2004) the species is present in the
Highlands, but contrary to the report of Leblanc et al. (2001) it is not a common fly in
that area. The mean monthly abundance of B. bryoniae in trap catches show numbers
are low in the Highlands (above 1200m) and along the coastline (below 600m)
suggesting B. bryoniae is a mid-altitude (600-1200m), rather than low or high altitude,
species (Figure 3.4). Rainfall, at the national level, has a significant influence on B.
bryoniae abundance (Table 3.3). This effect of rainfall on B. bryoniae is particularly
clear in Morobe where numbers from trap sites along the coastline where annual
rainfall levels are above 2000mm are low in comparison to numbers from traps situated
inland (Figure 3.5). Seasonal abundance patterns of B. bryoniae were dependent on
locality. However, in general abundance in Morobe, Central and Gazelle is lower
between January-April than it is between August and December (Figure 3.3). In the
Highlands numbers are fairly static but there is an evident decline in abundance in
June/July (Figure 3.3). The occurance of the wet season between January-April may
explain the decline in seasonal abundance in Central and Gazelle. During the wet
season in Morobe this trend is not observed (Figure 3.3). The cause may be due to the
high trap catchs from traps situated inland from Lae where less rainfall occurs (Figure
3.5).

63
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
Ave
rage
pop
ulat
ion
abun
dan
ce
0
50
100
150
200
250
300
HighlandsMorobeCentralGazelle
Figure 3.3 Seasonal abundance of Bactrocera bryoniae (Tryon) in four areas in Papua
New Guinea between June 1998 and September 2001

64
Figure 3.4 Mean monthly abundance of Bactrocera bryoniae at different altitude levels in Papua New Guinea between June 1998 and September
2001.
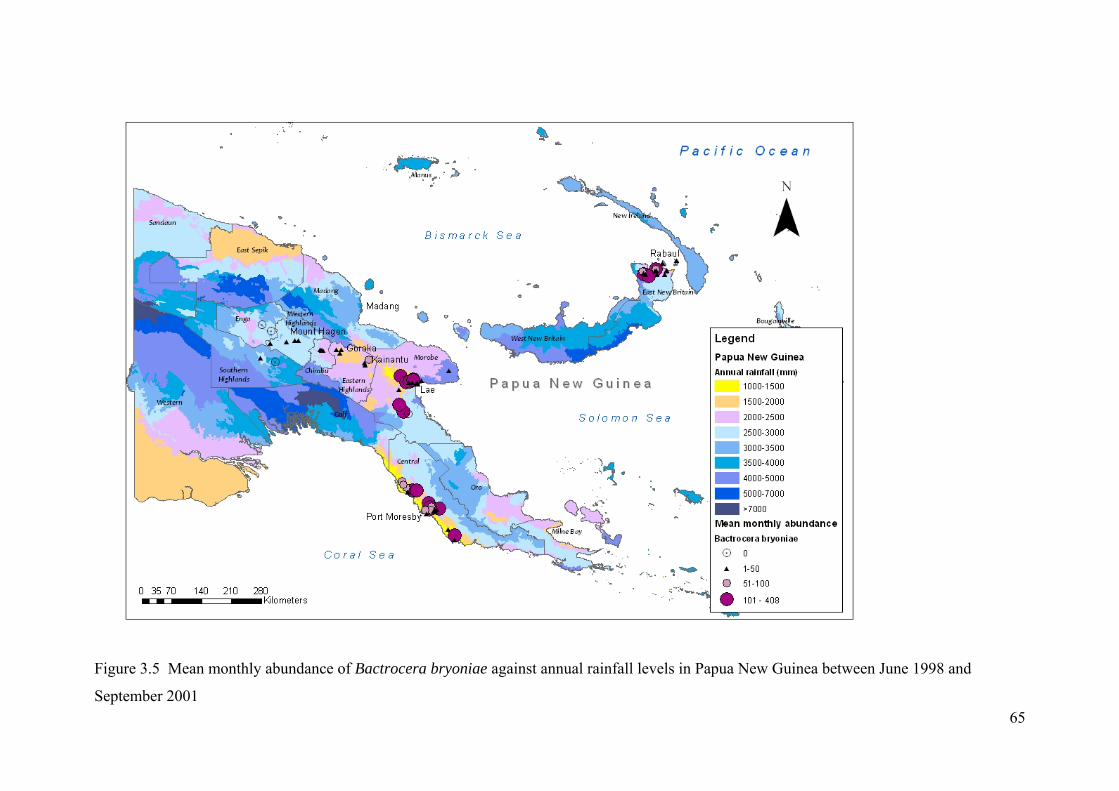
65
Figure 3.5 Mean monthly abundance of Bactrocera bryoniae against annual rainfall levels in Papua New Guinea between June 1998 and
September 2001


67
3.3.3 Bactrocera cucurbitae (Coquillett)
Contrary to existing assumptions, abundance of B. cucurbitae was not uniformly
distributed across lowland PNG (Table 3.2). The highest trap catches were caught in
Central (64% of sample) while the lowest were in the Highlands (1% of sample) (Table
3.2). At the national level temperature and rainfall had a significant influence on B.
cucurbitae distribution and abundance. Eighty-six percent of flies were trapped
between 0-600m; indicated by large green circles on map (Figure 3.7) and abundance
was found to decrease significantly as altitude increased. Between the Highlands and
Morobe only one trap site stands out with the highest mean monthly abundance catch
in both areas (Figure 3.7). Rainfall also has a significant influence at that level (Table
3.3). Seventy-four percent of flies were collected in areas that had 1000-2000 mm
annual rainfall. A single trap between Kainantu and Lae in the 1000-1500 mm rainfall
area had the largest abundance compared to others in the same area and this area
experiences <1500mm of annual rainfall. An effect indicating that B. cucurbitae spatial
distribution is limited to elevations below 600m (Figure 3.7) and to areas that
experience less than 1500mm annual rainfall (Figure 3.8). Seasonal abundance of B.
cucurbitae in each study area is different (Figure 3.6). While the patterns are different,
in general abundance in Central and on the Gazelle fall between January-June and
increase between July-December. In Morobe and the Highalnds seasonal abundance is
higher during January-June than between June-December (Figure 3.6).

68
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
Ave
rag
e p
opu
latio
n ab
und
anc
e
0
50
100
150
200
250
300
HighlandsMorobeCentralGazelle
Figure 3.6 Seasonal abundance of Bactrocera cucurbitae (Coquillett) in four areas in
Papua New Guinea between June 1998 and September 2001

69
Figure 3.7 Mean monthly abundance of Bactrocera cucurbitae at different altitude levels in Papua New Guinea between June 1998 and
September 2001

70
Figure 3.8 Mean monthly abundance of Bactrocera cucurbitae against annual rainfall levels in Papua New Guinea between June 1998 and
September 2001

71
3.3.4 Bactrocera frauenfeldi (Schiner)
The distribution and abundance of B. frauenfeldi in PNG is significantly influenced by
altitude and rainfall (Table 3.3). Results show that this species is largely limited in
distribution to areas below 1200m because it is particularly common in lowland areas
(0-600m) (Figure 3.10). Within this altitude range abundance is related to local rainfall
levels for numbers increase in areas that receive more than 1500 mm of annual rainfall
(Figure 3.11). The high numbers caught in traps in and around Port Moresby are
indicative of this, more so to the availability of water; similar to a trap located further
south of the city (Figure 3.11). The highest numbers of B. frauenfeldi were collected in
Morobe (65%), Gazelle (24%), and Central and virtually none in the Highlands (12%)
(Table 3.2). Rainfall appears to be the major influential factor of both population
distribution and abundance for this species, with the common feature of the lowland
areas where the fly is abundant being above average (> 2000mm) annual rainfall. In
this context it is important to note that there are many dry lowland sites (esp. in
Central) where the fly is rare or absent. In multiple regression analysis, the
combination of rainfall and altitude explained 36% of the variation in the trapping data
(Table 3.3). The seasonal abundance of B. frauenfeldi is notably dependent on rainfall
levels. During the wet season from January-April numbers are high in dry rain-shadow
areas like Central and also high in Morobe (it is the dry season there) (Figure 3.9). In
relatively wet areas like the Gazelle numbers are consistent throughout the year
increasing slightly between July to December.

72
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
Ave
rage
pop
ulat
ion
abun
danc
e
0
500
1000
1500
2000
2500
HighlandsMorobeCentralGazelle
Figure 3.9 Seasonal abundance of Bactrocera frauenfeldi (Schiner) in four areas in
Papua New Guinea between June 1998 and September 2001

73
Figure 3.10 Mean monthly abundance of Bactrocera frauenfeldi at different altitude levels in Papua New Guinea between June 1998 and
September 2001
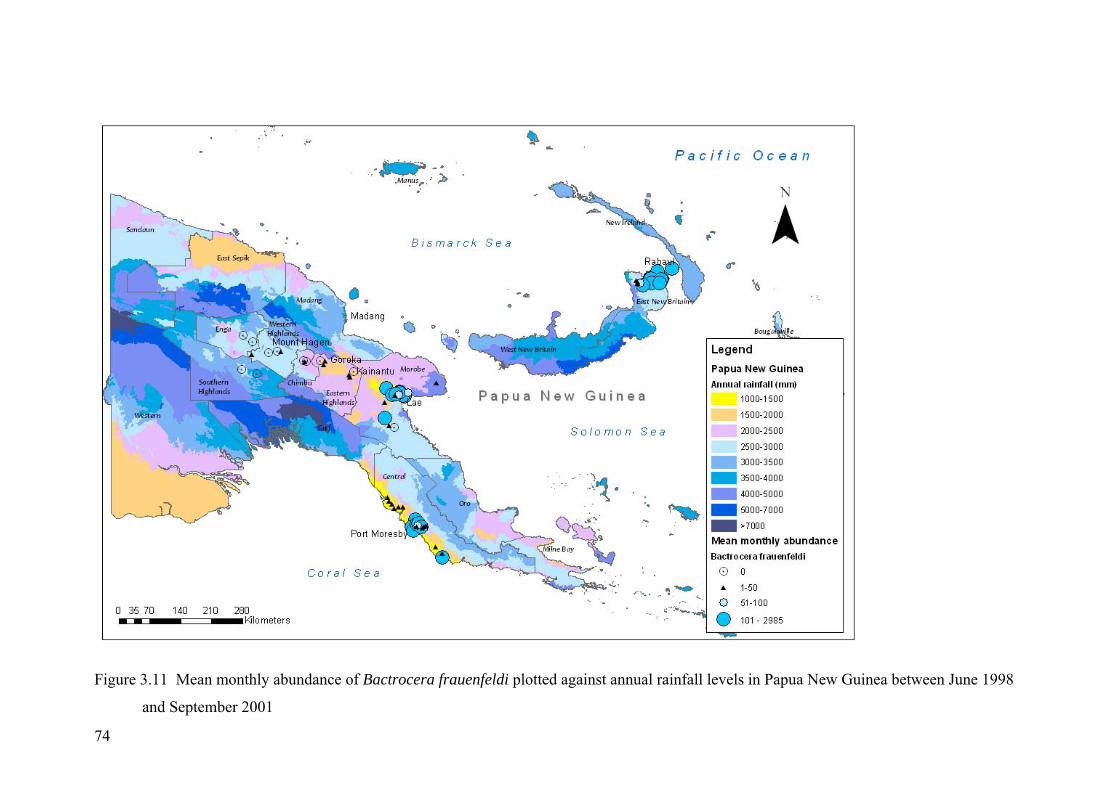
74
Figure 3.11 Mean monthly abundance of Bactrocera frauenfeldi plotted against annual rainfall levels in Papua New Guinea between June 1998
and September 2001

75
3.3.5 Bactrocera musae (Tryon)
All 54 traps located on the mainland of PNG had samples of B. musae, but the fly was
not uniformly abundant. The greatest abundance of B. musae was in Morobe (75%),
followed by the Highlands (17%), and then Central (7%) (Table 3.2). Unlike the other
four fly species, spatial abundance was not significantly related to altitude or rainfall
(Table 3.3). The fly was sampled from sea level to above 1800m (Figure 3.13) and in
areas that experienced between 1000-3500 mm of annual rainfall (Figure 3.14).
Seasonal abundance appeared very much dependent on locality. In Morobe populations
increased January-April and again May-July. In the Highlands population peaks
occurred between January-March and a degree longer July-October. In comparison
there was no real difference in Central (Figure 3.12). Distribution of banana fly was
not correlated with the relative importance of banana in local cropping systems within
Morobe, the Highlands, Central or Gazelle. However, when abundance was correlated
against the relative importance of banana across all four areas, abundance did rise as
the importance of banana as a food crop increased in an area (r= 0.285, p = 0.02)
(Table 3.3, Figure 3.15). On the Gazelle B. musae is known to be invasive and, at the
time data was collected, the fly was still spatially and temporally sporadic. Seven of 15
traps on the Gazelle Peninsula did not trap banana fly and due to this inconsistency and
low numbers seasonal phenology patterns could not be generated (but see data
presented in Chapter 6: Bactrocera musae (Tryon) in a novel environment: banana fly
as an invasive organism on the Gazelle Peninsula, Papua New Guinea). Observations
made during field work for this thesis indicate that the fly is still expanding its range
on the Gazelle.

76
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
Ave
rage
pop
ulat
ion
abun
danc
e
0
200
400
600
800
1000
1200
1400
1600
1800
HighlandsMorobeCentral
Figure 3.12 Seasonal abundance of Bactrocera musae in three areas in PNG between
June 1998 and September 2001

77
Figure 3.13 Mean monthly abundance of Bactrocera musae at different altitude levels in Papua New Guinea between June 1998 and September
2001

78
Figure 3.14 Mean monthly abundance of Bactrocera musae plotted against annual rainfall levels in Papua New Guinea between June 1998 and
September 2001

79
Figure 3.15 Mean monthly abundance between June 1998 and September 2001 of Bactrocera musae plotted against the relative importance of
banana as a food crop in cropping systems in Papua New Guinea

80
3.3.6 Bactrocera umbrosa (Fabricius)
Bactrocera umbrosa is most abundant on the Gazelle (49% of total sample) and in
Morobe (42%) but much less common elsewhere (Table 3.2, Figure 3.17). Abundance
is significantly negatively correlated with altitude (Table 3.3) and this is reflected in
the very low numbers trapped in the Highlands (Figure 3.17). Rainfall has a significant
influence on trap catch (Table 3.3). Traps situated in areas with >1500mm of annual
rainfall have high numbers (Figure 3.18). In Morobe trap catch in Lae is higher than
those situated away from the city and on the Gazelle most traps catch high numbers
(Figure 3.18). Seasonal abundance of B. umbrosa in the two main areas where it was
trapped (i.e. Morobe and Gazelle) were similar, with peaks coinciding with the wet
season (Figure 3.16). In these localities wet seasons are, coincidently, when breadfruit
(B. umbrosa’s major host) comes into season. It is thus unclear if rainfall pattern, host
fruiting cycle, or a combination of both, is driving annual cycles. The seasonal
abundance of B. umbrosa seen here mirrors the seasonality for breadfruit in the Pacific
(Nix, 1986; Houlder et al., 2001). The host of B. umbrosa is breadfruit a predominantly
low altitude plant (<1800m). The insignificance of breadfruit as an important food in
the Highlands region may influence the lack of breadfruit trees and fruit which may
explain the low abundance of flies at higher altitudes (Bourke et al., 1998) (Figure
3.19). Low abundance levels represented by orange circles are found in areas where
breadfruit is not important while green and blue circles showing higher abundance are
found in areas where breadfruit is part of the local diet (Figure 3.19). Where breadfruit
is part of the local diet it was recorded (in PNGRIS) as a nut in combination with other
‘nuts’. The combinations vary considerably across PNG and breadfruit was also not
categorised as was banana; as a dominant, subdominant or a minor ‘nut’. A visual
comparison was therefore done and presented (Figure 3.19). Map shows area where
breadfruit is grown. Statistical analysis was not done to verify the visual results
because of the difficulties of extracting detailed breadfruit data (as was done for
banana; dominant and subdominant food crop) from PNGRIS. Thus while rainfall and
altitude explained over 30% of the variation in the B. umbrosa trapping data it may be
that both of these variables are indirectly acting on the fly through host plant
abundance and fruiting season.

81
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
Ave
rage
pop
ulat
ion
abun
danc
e
0
100
200
300
400
500
HighlandsMorobeCentralGazelle
Figure 3.16 Seasonal abundance of Bactrocera umbrosa in four areas in PNG between
June 1998 and September 2001

82
Figure 3.17 Mean monthly abundance of Bactrocera umbrosa at different altitude levels in Papua New Guinea between June 1998 and
September 2001

83
Figure 3.18 Mean monthly abundance of Bactrocera umbrosa plotted against annual rainfall levels in Papua New Guinea between June 1998
and September 2001

84
Figure 3.19 Mean monthly abundance between June 1998 and September 2001 of Bactrocera umbrosa in Papua New Guinea where breadfruit
(Artocarpus spp.) is grown

85
3.4 Discussion
Current distribution records of fruit fly species in different parts of PNG either make
no comment about relative abundance (e.g. Clarke et al. 2004), or make statements
about distribution and abundance with no supporting data (e.g. Leblanc et al. 2001).
The data presented in this chapter has analysed and presented trapping data against
temperature (as related to altitude) and rainfall levels to identify the differential
influence of both on the fine scale spatial and temporal distribution of five fruit fly
species. It has also been able to show that host availability for two species (B. musae
and B. umbrosa) is an influential factor in their distributions but one which requires
further study. All species, with the exception of B. musae, are not common in the
Highlands provinces and there are studied areas like the provinces of Enga and
Southern Highlands where species are absent.
Attempts to use this spatial data to develop predictive distribution models (for example
to predict where B. musae might establish if introduced to another regional country)
were unsuccessful. Environmental matching programs such as Climex and BioClim are
predictive modelling tools which use information on climatic data to predict the
potential distribution range of a pest species (Yonow & Sutherst, 1998; Sutherst et al.,
1999; Vera et al., 2002; Paul et al., 2005; Stephens et al., 2007). This information is
vital for setting up appropriate and effective management measures (Baker, 1996;
Wright et al., 2005; Worner & Gevrey, 2006). I studied Climex and Dymex programs
to investigate the potential distribution of B. musae but found they were not applicable
for PNG. The country does not have adequate numbers of weather stations and the kind
of long-term consistently collected detail on weather variables such as temperature and
rainfall required to run the programmes. This problem was accentuated by the
topographical diversity of PNG. There are programmes available with which weather
variables between weather stations can be fitted but such programs still require either a
relatively even terrain between stations or a higher density of stations. The terrain of
PNG is not relatively even and the number of stations is low. Given the large number
of plant pests and diseases which are native to PNG and could invade neighbouring
countries, or conversely the number of offshore pests which might enter PNG and
spread, the inability to use ecological distribution models is a potentially serious
problem for both PNG and its neighbours.

86
With respect to B. musae, while temperature and rainfall were not found to have a
significant influence on the fly’s spatial and temporal distribution, both may play a role
on distribution and abundance at the ‘within province’ level. In low rainfall areas, such
as Central province, abundance of B. musae in dry areas outside of Port Moresby are
quite low for an area where banana is an important food and commodity crop (Hanson
et al., 2001). These abundance levels are also much lower than those in the Morobe
and Highlands study areas. The positive correlation between the relative importance of
banana in an area and the abundance of B. musae seen at the national level shows that
host availability is an important factor for further study. Specifically, there is need for
more targeted research to determine if host availability does indeed drive and sustain
banana fly population dynamics. Here host availability information is related to the
importance of the crop in an area but availability and importance may not be the same
because a host may be widely cultivated in an area but its importance as a staple food
may be low. It is therefore necessary to clarify whether trapping data can relate
abundance to host distribution and relate this measurement to the impact caused by a
study species population on susceptible hosts.
The low population abundance of four fruit fly species at higher altitudes provides
some resolution to the debate that the pest status of different fruit fly species may not
be the same at all localities within PNG, specifically when comparing between lowland
and highland production areas. These areas of low fruit fly population densities can be
monitored and managed to be classified as low-risk areas for commodity production.
Pre- and post-harvest management levels of pest species at higher altitude levels may
therefore not need to be as stringent as those required of production areas below
1200m where fly abundance is higher. Due to the different effects that temperature and
rainfall has on the five studied fruit fly species, control strategies will have to be
designed for individual species and the use of generic management models would be
inappropriate. The management of economically important fruit fly species in PNG
must incorporate long-term strategies that monitor infestation levels to help ensure that
the marketability of potential susceptible commodities is not hindered by the lack of or
inadequate number of required pest fruit fly control measures. This also highlights the
necessity to improve local quarantine measures to monitor and maintain the current
distribution of pest species and areas with low-pest-risk status.

87
3.5 Thesis Progress
The study of the distribution and abundance of B. musae and four other economically
important fruit fly species in PNG provides essential information toward the
development of the PNG banana PRA. Knowing how altitude (which can be a
surrogate for temperature), rainfall and host availability influences fly distribution can
be used to model and predict the likely risk of establishment of the fly in a novel
environment (a component of Step 2 of the PRA process), while determining possible
sites of low pest pressure for production is of relevance in developing risk management
options (Step 3 of the PRA). The following chapter specifically studies the impact of
natural B. musae populations on banana in PNG. The results pursue the question of
identifying if local abundance of the fly is linked to banana infestation rates. This
information is needed to identify the levels of risk associated with banana fly should it
establish in a novel area and for identifying potential low risk productions areas, again
relevant to Steps 2 and 3 of the PRA process.
RISK MANAGEMENTReview risk management options (e.g. import
restrictions, invasion tactics)
RISK ASSESSMENTAssess (a) the risk of entry; (b) the risk of establishment; and (c)
the risk of economic and other impacts
RISK INITIATIONIdentify pest(s) and pathways of concern, and consider whether the pest should
be considered for risk analysis in relation to the area at risk
3
2
1
Distribution and abundance of five economically important fruit fly species in Papua New Guinea [Chapter 3]
RISK MANAGEMENTReview risk management options (e.g. import
restrictions, invasion tactics)
RISK ASSESSMENTAssess (a) the risk of entry; (b) the risk of establishment; and (c)
the risk of economic and other impacts
RISK INITIATIONIdentify pest(s) and pathways of concern, and consider whether the pest should
be considered for risk analysis in relation to the area at risk
3
2
1
RISK MANAGEMENTReview risk management options (e.g. import
restrictions, invasion tactics)
RISK ASSESSMENTAssess (a) the risk of entry; (b) the risk of establishment; and (c)
the risk of economic and other impacts
RISK INITIATIONIdentify pest(s) and pathways of concern, and consider whether the pest should
be considered for risk analysis in relation to the area at risk
3
2
1
Distribution and abundance of five economically important fruit fly species in Papua New Guinea [Chapter 3]
Figure 3.20 Pest Risk Analysis process; with arrow indicating which step in process
information generated in this chapter has been collected for

88
Chapter 4. Infestation of bananas by Bactrocera musae (Tryon) in Papua New Guinea
4.1 Introduction
Banana (Musa spp.) is the fourth most important food crop in the world and in
developing countries it is the fourth most important commodity (Gold et al., 2002;
Ploetz, 2004). The cultivars most commonly produced for regional and international
trade are dessert eating Cavendish varieties (Stover, 1986). In Papua New Guinea
(PNG), there is a large range of banana varieties farmed at the subsistence and semi-
commercial farming level. The cultivars grown include both cooking varieties and
dessert types, including Cavendish (Bourke et al., 1998; Hartemink & Bourke, 2001;
Gunther et al., 2003). Bananas are cultivated from sea level up to 2200 meters above
sea level (Bourke et al., 1998). There are 235 banana accessions in PNG and over 100
varieties are farmed (Arnaud & Horry, 1997). With one exception, however, most
banana varieties have localised areas of production; only Cavendish is commonly
cultivated throughout the country (Gunther et al., 2003; Kambuou, 2005).
Banana production levels in PNG are often constrained by pest problems (Smith, 1976,
1977b; Tomlinson, 1984; King et al., 1988; Masamdu et al., 1989). One-hundred and
twelve organisms have been associated with banana in PNG (Chapter 2: Review of
banana (Musa spp.) pests in Papua New Guinea) and, of these, fourteen are reported to
be commonly observed and managed. Bactrocera musae (Tryon) (banana fly) is one of
these and is reported to be the major pest (Pone, 1994; Leblanc et al., 2001), or one of
several major pests (Kambuou, 2004), on banana in PNG. The actual infestation rate of
B. musae on PNG bananas is poorly documented, despite its apparent importance.
Leblanc et al. (2001) report 0-75% B. musae infestation on banana in PNG, but provide
no information on sample size or banana variety. Sar et al. (2001) report 25%
infestation on Kalapua variety in East New Britain Province (ENB) and 30% on
Cavendish in Central Province, but again show no assessment methodology or
information on sample size.
Leblanc et al. (2001), if read uncritically, could be used as evidence that all banana
varieties cultivated in PNG have high infestation levels and that infestation is common

89
across all parts of PNG. This would be unfortunate as Leblanc et al. (2001) give no
measure of banana varietal susceptibility to B. musae (a critical issue given how many
varieties there are) and restricted data on locality differences (again important given
known information on the distribution of the fly [Chapter 3: Distribution and
abundance of five economically important fruit fly species in PNG]). Formal
documentation on B. musae infestation levels on different banana varieties in PNG is
rare. Smith (1977, 1976) described higher infestation levels caused by B. musae on
short varieties of banana compared to a taller variety. He described Giant Cavendish as
the tall variety and the shorter varieties to be Tui/Robusta and Dwarf Cavendish.
Reports done for the commercial production of Giant Cavendish in PNG state that B.
musae infestation is a problem, particularly when the fruit is harvested ripe (Fooks,
1989, 2002).
The small amount of documentation available on B. musae infestation rates on PNG
banana is not of a standard which complies with international trade and phytosanitary
requirements. No detailed impact assessment has been carried out on Cavendish,
geographically the most widely planted banana in PNG and a variety likely to be
considered for international trade. Nor is there, with the exception of Smith (1977), any
quantified field information on susceptibility of different banana varieties. The main
objectives of this chapter were therefore twofold. The first was to carry out a national
level damage assessment survey of B. musae on Cavendish, so as to assess infestation
levels in different regions of PNG. I wanted particularly to determine if differences in
abundance of B. musae (as determined in Chapter 3) translated to lower or higher
levels of fruit infestation. If so, then cropping district could potentially be used as a
component of a banana market access tool (i.e. by picking production regions with low
infestation rates). Because it is so widely planted, Cavendish was the only variety for
which a uniform national survey could be done. The second objective investigated
differences in varietal susceptibility to banana fly infestation when bananas were
grown in a common-garden trial. Should evidence for varietal difference be confirmed,
then future market access work could concentrate on searching for commercial
varieties of low susceptibility, a recognized risk management tool (Armstrong, 2001;
Rattanapun et al., 2009).

90
The chapter falls into four parts. All investigate levels of infestation caused by B.
musae on banana varieties. The initial three parts assess damage impact on more than
one variety while the last carries out the extensive damage assessment survey on a
single variety. The initial component shows results from general host surveys of
banana carried out in different parts of PNG showing infestation levels of B. musae on
a number of cultivated banana varieties. The second part is a methodology trial, the
results of which were used to design an appropriate sampling strategy for the
Cavendish survey. The third section reports on a common-garden experiment which
looked at infestation levels on four different banana varieties, while the fourth section
reports on a national damage assessment survey of banana fly infestation in Cavendish.
4.2 Materials and methods
4.2.1 Differences in varietal susceptibility
General banana host surveys. Numerous host surveys of banana were carried out in
different parts of PNG. These samples were not collected in any methodological
manner and therefore were not analysed but are presented here to show the varieties of
banana sampled and having fruit fly infestation in different provinces. Recorded
banana varieties are labelled using common names or for varieties in Central and ENB
using germplasm identification. Banana germplasm collections in Central are kept at
the NARI’s dry-lowlands agriculture station at Laloki and in ENB at the lowlands
agriculture experiment station at Kerevat in ENB where germplasm collections are
kept. These samples of cultivated banana varieties were collected at different maturity
stages and were incubated as single finger samples or in multiple finger samples.
Records were made of the number of pupae developing and the number of adult flies
emerging from each sample.
Methodology trial. The trial was run on the Gazelle Peninsula, East New Britain
province (ENB) from November to December 20021. Three varieties of cooking
banana were used in the trial: Kekiau (PNG101 Musa Eumusa AA), Vudu Papua
(PNG004 Musa Eumusa AA), and Tukuru (PNG118 Musa Eumusa ABB). The
morphological and taxonomic characteristics of each variety are described in the PNG
1 The field work for this trial was undertaken prior to my PhD candidacy. However, I fully designed and implemented this trial, the data of which remained unanalysed until I started my PhD. All data analyses reported here were undertaken while I was an enrolled PhD candidate.

91
Musalogue (Arnaud & Horry, 1997). The banana bunches were collected from gardens
in the Vudal and Vunapalading areas northwest from Kokopo and Rabaul and an area
into which B. musae populations were still spreading (Mararuai et al., 2001). Whole
banana bunches were hung at three different locations on the eastern side of the
Gazelle Peninsula between Kokopo (04° 20’S, 152° 18’E) and Rabaul (04° 11’S, 152°
08’E), an area where banana fly is present (Mararuai et al., 2001 and Chapter 3). At
each location a mature, harvested banana bunch of each variety were simultaneously
hung uncovered on posts about two meters off the ground and exposed to wild flies for
a week. After this time, individual fingers were removed and setup over moist sawdust
for the recording of fly emergence (and hence infestation levels). Details on the
methods used for rearing-out fruit flies from harvested fruit (for this and subsequent
trials) are described and illustrated in Leblanc et al. (2001) (see Appendix 1). The
selection procedure for finger samples was as follows: the top two hands on the bunch
were referred to as the top part of the bunch and from these two hands five fingers
were randomly selected. Sample selection was concentrated on the top two hands
because field observation showed more flies sitting on the top half of the banana bunch
and maturity in banana fingers began at this location. The next two to three hands
below were referred to as the middle part and from here three fingers were randomly
selected. From the remaining hands below the middle part, two fingers were randomly
selected. Six bunches of each of the three banana varieties were exposed and sampled,
leading to a total sample of 180 banana fingers. A two-way ANOVA was carried out to
determine if there was any significant differences in banana fly infestation rates at
different sample locations within the bunch. The intention was that if such a difference
was found, subsequent surveys would sample hands within a banana bunch which had
the highest probability of being infested.
Common garden experiment. A banana garden was planted in mid-2007 at the [PNG]
National Agricultural Research Station Laloki (09°23’S, 147°17’E), in the Central
province of PNG, just outside the capital city of Port Moresby. Three fields (3
replicates) were planted using a randomised planting allocation design with ten plants
of each of four banana varieties: Kokopo 1 (PNG030); Kalapua [dwarf] (PNG171);
Daru (PNG131); and Kurisa. The characteristics of PNG030, PNG171 and PNG131
are supplied in Musalogue (Arnaud & Horry, 1997), but Kurisa is not included in that
catalogue. These four are economically important banana varieties PNG. Banana

92
bunch harvest began in February 2007 and sampling continued until March 2009.
When bunches were mature and ready for harvest, multiple fingers were setup in single
incubation containers over moist sawdust as previousy described. The entire bunch was
setup for observation as single fingers, not a subset of fingers from the bunch as in the
Methodology trial. A record was made of the number of pupae recovered and the
number of adult flies that emerged from each sample/container. The date of submission
for this thesis fell prior to the completion of the trial. In addition, complete sampling of
the all varieties was not done due to labour constraints at the garden but what data was
recorded is presented here. Low banana fly infestation levels and incomplete sampling
meant that formal analysis of data could not be undertaken, rather results are
summarized and presented graphically. The purpose of this experiment was to
investigate B. musae infestation levels on economically important banana varieties
other than Cavendish.
4.2.2 National Cavendish survey
I carried out the national survey between November 2007 and January 2008 with the
assistance of technical staff from the National Agricultural Research Institute (NARI)
and Fresh Produce Development Authority. There were three reasons why this survey
was done at this time of the year. Firstly, during the course of my PhD study this time
frame was the most logistically fesible during which I could carry out the survey.
Secondly, it is during this time of the year when the fluctuations in seasonal abundance
of the study fly, B. musae, are at their lowest in Morobe and the Highalnds but are
uniform to Central (Section 3.3.5 - Chapter 3). This is important because it can be
assumed that the natural B. musae populations in samping areas are similar and one
area will not have a higher population number which may consequently have an effect
on infestation levels and prevent comparative analysis between surveyed areas.
Thirdly, the resultant information gathered during this time period of low seasonal
abundance is certainly a factor that can be incorporated into management systems and
approaches into fruit fly management and be used as a banana market access tool.
There were 22 areas or sampling locations, evenly spread in number over the four PNG
production areas which were analysed in Chapter 3. Five locations were situated in the
Central province; seven were located in the Highlands zone (covering three locations
in Chimbu province and four in Eastern Highlands province (EHP)), six in Morobe
province, and four on the Gazelle Peninsula, ENB (Figure 4.1). At each location five

93
mature bunches of Cavendish banana were harvested. Within a location the bunches
may have all come from one site (a single farm for example), or neighbouring sites
(e.g. adjoining farms) depending on availability: in total there were 57 local collection
sites (11 in Morobe, 18 in EHP, 12 in Chimbu, nine in Central, and seven on the
Gazelle). Detailed collection information for each bunch, including site coordinates,
weight and number of fruit sampled are provided in Table 4.1. The sampling
methodology used to select bananas from a bunch was that determined from the
Methodology trial. Altogether there were 111 banana samples collected; 31 in Morobe,
20 in EHP, 15 in Chimbu, 26 in Central and 19 on the Gazelle. All subsamples were
incubated over moist sawdust and a record was made of pupal and adult emergence
numbers. Again, because of the very low banana fly infestation levels recorded, formal
analysis of data was not undertaken, rather results are summarized and presented
graphically.
Figure 4.1 Map of harvest spots for Cavendish banana during the fruit fly damage
assessment survey (November 2007-January 2008) carried out in five provinces
in Papua New Guinea

94
Table 4.1 Sampling details for the national Cavendish survey
Province Sample location
Sample site GPS reading Stage of Maturity
*
Weight (g)
No of banana fingers (B-
Bunch) Latitude (S) Longitude (E)
Morobe Kaiapit Mutzing 06° 20΄48˝ 146°13΄51˝ MG/R 4572.1 32 Nasuapum Orockangku 06°35΄18˝ 146°49΄22˝ MG 3198.7 24 Ngarefe 06°35΄36˝ 146°48΄41˝ MG 2411.4 18 Ngarefe 06°35΄36˝ 146°48΄41˝ MG 3603.6 27 Gomamos 06°35΄48˝ 146°48΄31˝ MG 1916.4 15 Gomamos 06°35΄48˝ 146°48΄31˝ MG 1950.8 15 Gabensis SDA Primary Sch 06°41΄51˝ 146°48΄00˝ MG 3865.5 29 SDA Primary Sch 06°41΄42˝ 146°48΄02˝ MG 941.4 7 SDA Primary Sch 06°41΄42˝ 146°48΄02˝ MG 1325.7 10 SDA Primary Sch 06°41΄42˝ 146°48΄02˝ MG 1960.4 15 SDA Primary Sch 06°41΄42˝ 146°48΄02˝ MG 2417.9 18 Bundun Training Centre 06°51΄16˝ 146°37΄06˝ GM 3474.2 26 Training Centre 06°51΄16˝ 146°37΄06˝ MG 1741 13 Training Centre 06°51΄16˝ 146°37΄06˝ MG 18614.9 14 Training Centre 06°51΄16˝ 146°37΄06˝ MG 3001.2 23 Training Centre 06°51΄16˝ 146°37΄06˝ MG 1176.7 9 Bukawa Poahum 06°39΄22˝ 147°02΄17˝ MG 1355 10 Poahum 06°39΄22˝ 147°02΄17˝ MG 2021.3 15 Didiman station 06°38΄59˝ 147°02΄14˝ MG/R 3814.5 29 Liklik Rot, Situm 06°39΄50˝ 147°03΄44˝ MG 1646.2 12 Liklik Rot, Situm 06°39΄50˝ 147°03΄44˝ MG 1983.9 15 Liklik Rot, Situm 06°39΄50˝ 147°03΄44˝ MG 1900.9 14 Bubia NARI Bubia 06°40΄04˝ 146°54΄75˝ MG 2187.3 26 NARI Bubia 06°40΄04˝ 146°54΄75˝ MG 3078.3 23 NARI Bubia 06°40΄04˝ 146°54΄75˝ MG 5022.2 38 NARI Bubia 06°40΄04˝ 146°54΄75˝ MG 2347.9 18 NARI Bubia 06°40΄04˝ 146°54΄75˝ MG 3372.7 33 NARI Bubia 06°40΄04˝ 146°54΄75˝ MG 4430.1 34 NARI Bubia 06°40΄04˝ 146°54΄75˝ MG 3702 26 NARI Bubia 06°40΄04˝ 146°54΄75˝ MG 1881.7 30
Eastern Highlan
ds Watabung Top - Sharp Kona 06°04΄19˝ 145°12΄41˝ MG 3574.2 30
Top - Sharp Kona 06°04΄19˝ 145°12΄41˝ MG 4313.9 41 Fionoku 06°04΄06˝ 145°12΄24˝ MG/R 3028.9 30 Watabung station 06°04΄53˝ 145°12΄28˝ MG/R 3536.7 35 Efaka 06°04΄39˝ 145°12΄19˝ MG/R 4853.7 38 Goroka Kofa, Korepa 06°02΄08˝ 145°15΄39˝ MG/R 7199.3 43 Kerefa 06°01΄40˝ 145°23΄36˝ MG 4145.6 36 Massy, Okiufa 06°03΄05˝ 145°23΄07˝ MG 2661.4 26 Kabiufa village 05°59΄45˝ 145°22΄20˝ R 5127.7 36 Fanifa 06°05΄53˝ 145°23΄45˝ R 5819.6 32 Henganofi Kapitina primary 06°17΄28˝ 145°42΄55˝ MG 6405.9 50 Avani, Kompri 06°18΄00˝ 145°41΄00˝ MG 6433.3 32 Agafa 06°18΄17˝ 145°38΄59˝ R 6359.2 35 Seganafamo 06°14΄54˝ 145°35΄20˝ MG 3524.8 25 Manicon 06°14΄59˝ 145°33΄45˝ MG 5125.9 47 Table continued overleaf
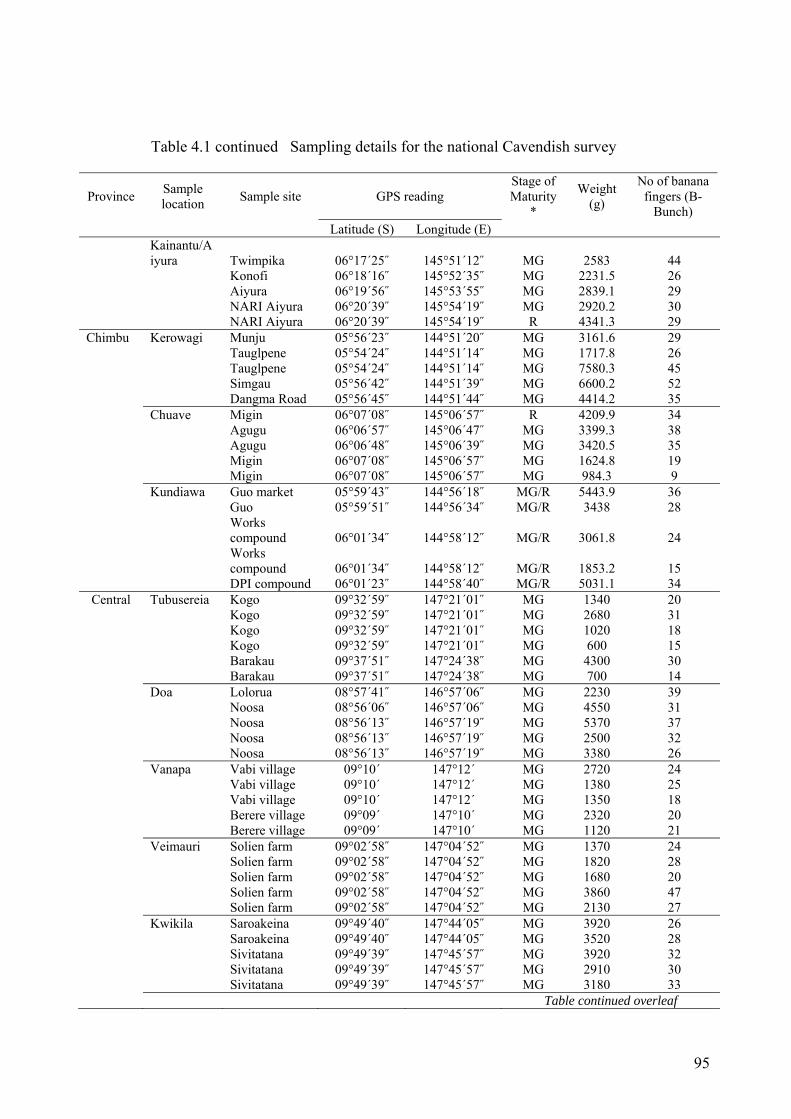
95
Table 4.1 continued Sampling details for the national Cavendish survey
Province Sample location
Sample site GPS reading Stage of Maturity
*
Weight (g)
No of banana fingers (B-
Bunch) Latitude (S) Longitude (E)
Kainantu/Aiyura Twimpika 06°17΄25˝ 145°51΄12˝ MG 2583 44
Konofi 06°18΄16˝ 145°52΄35˝ MG 2231.5 26 Aiyura 06°19΄56˝ 145°53΄55˝ MG 2839.1 29 NARI Aiyura 06°20΄39˝ 145°54΄19˝ MG 2920.2 30 NARI Aiyura 06°20΄39˝ 145°54΄19˝ R 4341.3 29
Chimbu Kerowagi Munju 05°56΄23˝ 144°51΄20˝ MG 3161.6 29 Tauglpene 05°54΄24˝ 144°51΄14˝ MG 1717.8 26 Tauglpene 05°54΄24˝ 144°51΄14˝ MG 7580.3 45 Simgau 05°56΄42˝ 144°51΄39˝ MG 6600.2 52 Dangma Road 05°56΄45˝ 144°51΄44˝ MG 4414.2 35 Chuave Migin 06°07΄08˝ 145°06΄57˝ R 4209.9 34 Agugu 06°06΄57˝ 145°06΄47˝ MG 3399.3 38 Agugu 06°06΄48˝ 145°06΄39˝ MG 3420.5 35 Migin 06°07΄08˝ 145°06΄57˝ MG 1624.8 19 Migin 06°07΄08˝ 145°06΄57˝ MG 984.3 9 Kundiawa Guo market 05°59΄43˝ 144°56΄18˝ MG/R 5443.9 36 Guo 05°59΄51˝ 144°56΄34˝ MG/R 3438 28
Works compound 06°01΄34˝ 144°58΄12˝ MG/R 3061.8 24
Works compound 06°01΄34˝ 144°58΄12˝ MG/R 1853.2 15
DPI compound 06°01΄23˝ 144°58΄40˝ MG/R 5031.1 34 Central Tubusereia Kogo 09°32΄59˝ 147°21΄01˝ MG 1340 20
Kogo 09°32΄59˝ 147°21΄01˝ MG 2680 31 Kogo 09°32΄59˝ 147°21΄01˝ MG 1020 18 Kogo 09°32΄59˝ 147°21΄01˝ MG 600 15 Barakau 09°37΄51˝ 147°24΄38˝ MG 4300 30 Barakau 09°37΄51˝ 147°24΄38˝ MG 700 14 Doa Lolorua 08°57΄41˝ 146°57΄06˝ MG 2230 39 Noosa 08°56΄06˝ 146°57΄06˝ MG 4550 31 Noosa 08°56΄13˝ 146°57΄19˝ MG 5370 37 Noosa 08°56΄13˝ 146°57΄19˝ MG 2500 32 Noosa 08°56΄13˝ 146°57΄19˝ MG 3380 26 Vanapa Vabi village 09°10΄ 147°12΄ MG 2720 24 Vabi village 09°10΄ 147°12΄ MG 1380 25 Vabi village 09°10΄ 147°12΄ MG 1350 18 Berere village 09°09΄ 147°10΄ MG 2320 20 Berere village 09°09΄ 147°10΄ MG 1120 21 Veimauri Solien farm 09°02΄58˝ 147°04΄52˝ MG 1370 24 Solien farm 09°02΄58˝ 147°04΄52˝ MG 1820 28 Solien farm 09°02΄58˝ 147°04΄52˝ MG 1680 20 Solien farm 09°02΄58˝ 147°04΄52˝ MG 3860 47 Solien farm 09°02΄58˝ 147°04΄52˝ MG 2130 27 Kwikila Saroakeina 09°49΄40˝ 147°44΄05˝ MG 3920 26 Saroakeina 09°49΄40˝ 147°44΄05˝ MG 3520 28 Sivitatana 09°49΄39˝ 147°45΄57˝ MG 3920 32 Sivitatana 09°49΄39˝ 147°45΄57˝ MG 2910 30 Sivitatana 09°49΄39˝ 147°45΄57˝ MG 3180 33 Table continued overleaf

96
Table 4.1 continued Sampling details for the national Cavendish survey
Province Sample location
Sample site GPS reading Stage of Maturity
*
Weight (g)
No of banana fingers (B-
Bunch) Latitude (S) Longitude (E)
East New Kokopo Ulapea 04°20΄38˝ 152°18΄25˝ MG 5300 38 Britain Ulapea 04°20΄38˝ 152°18΄25˝ MG 3950 29 Ulapea 04°20΄38˝ 152°18΄25˝ MG 3430 30 Ulapea 04°20΄38˝ 152°18΄25˝ MG 3180 34 Ulapea 04°20΄38˝ 152°18΄25˝ MG 4730 39 Rabaul Pilapila 04°11΄29˝ 152°08΄20˝ MG 3180 28 Vuvu 04°12΄41˝ 152°07΄35˝ MG 2940 27 Vuvu 04°12΄41˝ 152°07΄35˝ MG 5530 34 Karakakaul 04°12΄35˝ 152°07΄12˝ MG 4130 24 Keravat NARI Keravat 04°19΄ 152°01΄ MG 2910 28 NARI Keravat 04°19΄ 152°01΄ MG 4460 29 NARI Keravat 04°19΄ 152°01΄ MG 4650 46 NARI Keravat 04°19΄ 152°01΄ MG 3600 30 NARI Keravat 04°19΄ 152°01΄ MG 5060 41 Induna Induna Plantation 04°35΄56˝ 152°22΄06˝ MG 4840 34 Induna Plantation 04°35΄56˝ 152°22΄06˝ MG 2960 30 Induna Plantation 04°35΄56˝ 152°22΄06˝ MG 2890 37 Induna Plantation 04°36΄13˝ 152°21΄48˝ MG 6800 39 Induna Plantation 04°36΄13˝ 152°21΄48˝ MG 3200 22
* Stage of maturity: MG – mature green, R – ripe
4.3 Results
4.3.1 Differences in varietal susceptibility
General banana host survey. Cultivated banana varieties were sampled in five
provinces (Central, East New Britain, Madang, Morobe, and Western Highlands)
(Table 4.2). In the Western Highlands mature green Kalapua samples did not have fruit
fly infestation (Table 4.2). In the Central province twenty-one varieties were sampled
but only five varieties were infested. Mature-green to ripe samples of Cavendish,
Kalapua, OBP21, OBY6 and Yawa were infested by B. musae (Table 4.2). In East
New Britain, twenty-one varieties were collected but four of these had infestation. Ripe
samples of KMC15, Ramarama and Tukuru had B. frauenfeldi infestations while ripe
samples of Yawa had both B. frauenfeldi and B. musae. In Madang two varieties were
sampled but only one variety, Kalapua had B. musae infestation. In Morobe three
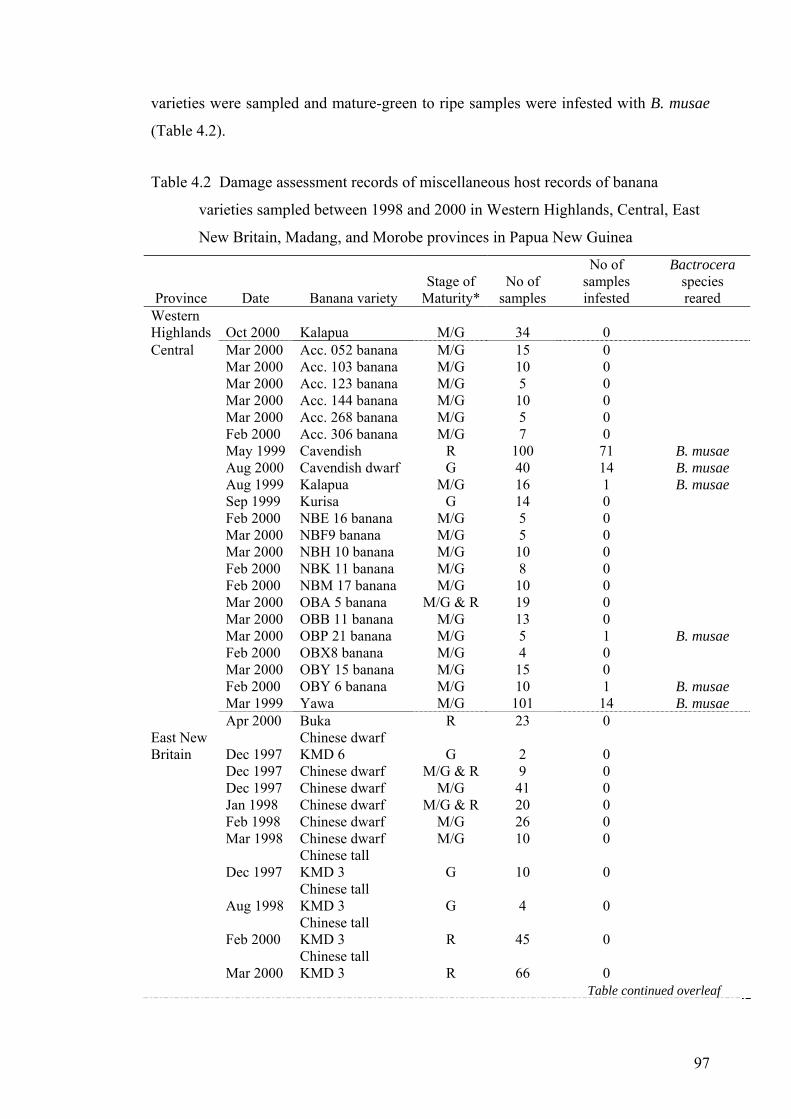
97
varieties were sampled and mature-green to ripe samples were infested with B. musae
(Table 4.2).
Table 4.2 Damage assessment records of miscellaneous host records of banana
varieties sampled between 1998 and 2000 in Western Highlands, Central, East
New Britain, Madang, and Morobe provinces in Papua New Guinea
Province Date Banana variety Stage of
Maturity*No of
samples
No of samples infested
Bactrocera species reared
Western Highlands Oct 2000 Kalapua M/G 34 0 Central Mar 2000 Acc. 052 banana M/G 15 0 Mar 2000 Acc. 103 banana M/G 10 0 Mar 2000 Acc. 123 banana M/G 5 0 Mar 2000 Acc. 144 banana M/G 10 0 Mar 2000 Acc. 268 banana M/G 5 0 Feb 2000 Acc. 306 banana M/G 7 0 May 1999 Cavendish R 100 71 B. musae Aug 2000 Cavendish dwarf G 40 14 B. musae Aug 1999 Kalapua M/G 16 1 B. musae Sep 1999 Kurisa G 14 0 Feb 2000 NBE 16 banana M/G 5 0 Mar 2000 NBF9 banana M/G 5 0 Mar 2000 NBH 10 banana M/G 10 0 Feb 2000 NBK 11 banana M/G 8 0 Feb 2000 NBM 17 banana M/G 10 0 Mar 2000 OBA 5 banana M/G & R 19 0 Mar 2000 OBB 11 banana M/G 13 0 Mar 2000 OBP 21 banana M/G 5 1 B. musae Feb 2000 OBX8 banana M/G 4 0 Mar 2000 OBY 15 banana M/G 15 0 Feb 2000 OBY 6 banana M/G 10 1 B. musae Mar 1999 Yawa M/G 101 14 B. musae
East New Britain
Apr 2000 Buka R 23 0
Dec 1997 Chinese dwarf KMD 6 G 2 0
Dec 1997 Chinese dwarf M/G & R 9 0 Dec 1997 Chinese dwarf M/G 41 0 Jan 1998 Chinese dwarf M/G & R 20 0 Feb 1998 Chinese dwarf M/G 26 0 Mar 1998 Chinese dwarf M/G 10 0
Dec 1997 Chinese tall KMD 3 G 10 0
Aug 1998 Chinese tall KMD 3 G 4 0
Feb 2000 Chinese tall KMD 3 R 45 0
Mar 2000 Chinese tall KMD 3 R 66 0
Table continued overleaf

98
Table 4.3 continued…Damage assessment records of miscellaneous host records of
banana varieties sampled between 1998 and 2000 in Western Highlands, Central, East
New Britain, Madang, and Morobe provinces in Papua New Guinea
Province Date Banana variety Stage of
Maturity*No of
samples
No of samples infested
Bactrocera species reared
Apr 2000 Chinese tall KMD 3 R 13 0
Dec 1997 Gunth M/G 14 0 Apr 2000 Kalapua R 16 0 Dec 1999 Katkatur R 7 0 Jan 1998 Kekiau M/G 12 0 Feb 1998 Kekiau G 6 0 Feb 2000 Kekiau R 38 0 Mar 2000 Kekiau R 57 0 Apr 2000 Kekiau M/G & R 68 0 Dec 1997 KMC 2 R 12 0 Dec 1997 KMC 11 M/G 25 0 Dec 1997 KMC 15 R 12 1 B. frauenfeldi Dec 1997 Maram R 40 0 Jan 1998 Maram M/G & R 19 0 Feb 1998 Maram R 22 0
Jan 1998 Marnaiar KMC 13 M/G & R 49 0
Dec 1997 Pitu KMC 12 G & M/G 32 0 Feb 1998 Poro KMD 7 M/G & R 51 0 Feb 2000 Ramarama R 18 0 Mar 2000 Ramarama R 19 1 B. frauenfeldi Apr 2000 Talauba R 7 7 ? Dec 1997 Touben KMC 9 M/G & R 4 0 Dec 1997 Tukuru M/G 16 0 Feb 1998 Tukuru M/G 12 0 Feb 2000 Tukuru KMC 10 R 11 4 B. frauenfeldi Mar 2000 Tukuru R 50 1 B. frauenfeldi Apr 2000 Tukuru M/G & R 34 0 May 2000 Tukuru R 17 0
Jan 1998 Vabokor (minintina) M/G & R 28 0
Dec 1997 Vudu vok KMC 16 G 47 0
Dec 1997 Vudu vok KMC 16 R 6 0
Dec 1997 Vudu vok KMC 16 M/G 17 0
Dec 1997 Yawa KMD 1 M/G 28 0 Dec 1997 Yawa KMD 1 R 54 0 Jan 1998 Yawa KMD 1 R 29 0 Feb 1998 Yawa KMD 1 R 13 0 Mar 1998 Yawa KMD 1 R 11 0 Table continued overleaf

99
Table 4.4 continued…Damage assessment records of miscellaneous host records of
banana varieties sampled between 1998 and 2000 in Western Highlands, Central, East
New Britain, Madang, and Morobe provinces in Papua New Guinea
Province Date Banana variety Stage of
Maturity*No of
samples
No of samples infested
Bactrocera species reared
Feb 2000 Yawa KMD 1 R 38 1 B. frauenfeldi & B. musae
Mar 2000 Yawa KMD 1 R 52 0 Madang Jun 2000 Kalapua R 17 7 B. musae Jun 2000 Cavendish M/G 121 0 Morobe Oct 1998 Kalapua R 12 0 May 1999 Kalapua G 74 1 B. musae Aug 1999 Kalapua R 68 4 B. musae Oct 1999 Kalapua M/G 311 34 B. musae Oct 1999 Kalapua R 33 0 Nov 1999 Kalapua R 22 0 Dec 1999 Kalapua M/G 20 0 Apr 2000 Kalapua R 16 15 B. musae Apr 2000 Kalapua M/G 34 16 B. musae Apr 2000 Kekiau R 22 1 B. musae Dec 1998 Wild banana M/G 287 0 May 1999 Wild banana R 15 1 B. musae Sep 1999 Wild banana R 100 0 Nov 1999 Wild banana R 105 3 B. musae Jan 2000 Wild banana R 101 0 Apr 2000 Wild banana R 291 2 B. musae Sep 1998 Yawa F 13 1 B. musae Jan 1999 Yawa R 61 0 Jan 1999 Yawa M/G 12 0 Jan 1999 Yawa G 10 0 Apr 1999 Yawa R 13 2 B. musae Apr 1999 Yawa M/G 60 7 B. musae Jun 1999 Yawa R 10 0 Jun 1999 Yawa M/G 23 0 Aug 1999 Yawa R 14 2 B. musae Aug 1999 Yawa M/G 13 1 B. musae Nov 1999 Yawa R 37 2 B. musae Nov 1999 Yawa M/G 15 0 Apr 2000 Yawa R 16 0
* Stage of maturity: G – green, MG – mature green, R – ripe, F - fallen
Methodology trial. Data from the trial, when tested for homogeneity of variance using
Levene’s test was found to be non-homogenous, even after standard transformations.
Analysis was thus done using the nonparametric Kruskall-Wallis test. Infestation rates
(and total flies yielded) of Kekiau, Vudu Papua and Tukuru varieties were found,
respectively, to be 25% (606), 20% (495) and 37% (343). The mean yield of flies was
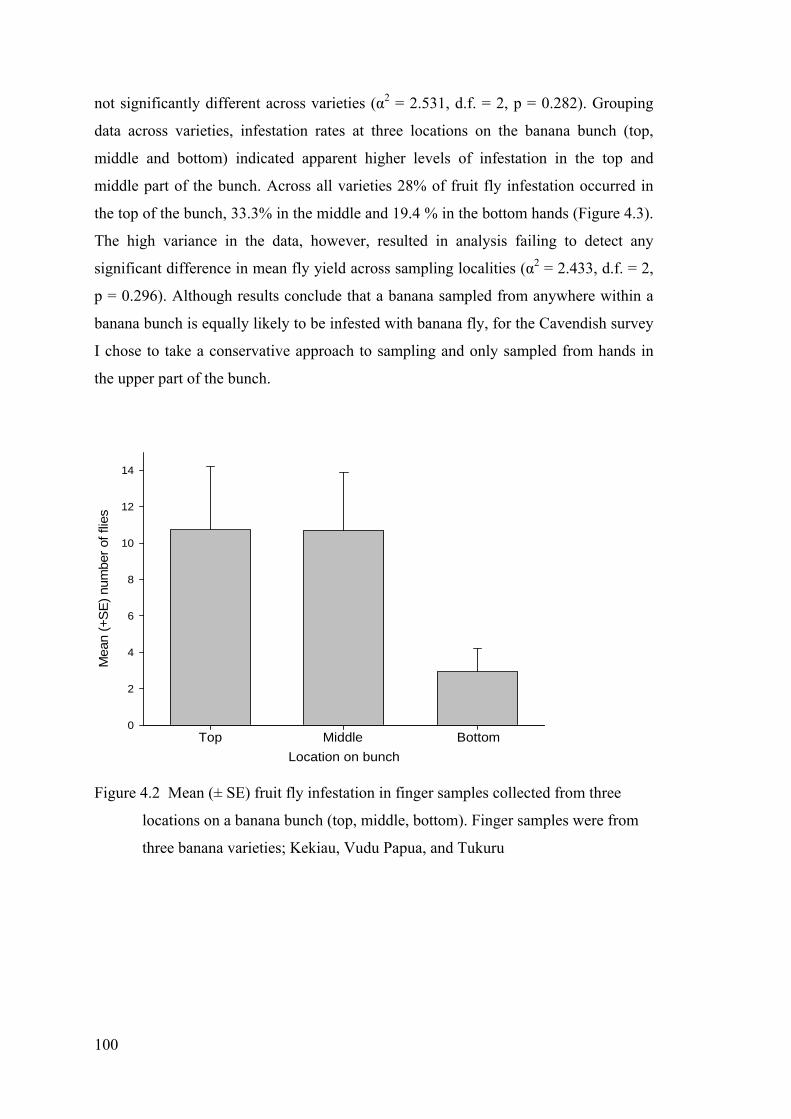
100
not significantly different across varieties (α2 = 2.531, d.f. = 2, p = 0.282). Grouping
data across varieties, infestation rates at three locations on the banana bunch (top,
middle and bottom) indicated apparent higher levels of infestation in the top and
middle part of the bunch. Across all varieties 28% of fruit fly infestation occurred in
the top of the bunch, 33.3% in the middle and 19.4 % in the bottom hands (Figure 4.3).
The high variance in the data, however, resulted in analysis failing to detect any
significant difference in mean fly yield across sampling localities (α2 = 2.433, d.f. = 2,
p = 0.296). Although results conclude that a banana sampled from anywhere within a
banana bunch is equally likely to be infested with banana fly, for the Cavendish survey
I chose to take a conservative approach to sampling and only sampled from hands in
the upper part of the bunch.
Location on bunch
Mea
n (+
SE
) nu
mbe
r of
flie
s
0
2
4
6
8
10
12
14
Top Middle Bottom
Figure 4.2 Mean (± SE) fruit fly infestation in finger samples collected from three
locations on a banana bunch (top, middle, bottom). Finger samples were from
three banana varieties; Kekiau, Vudu Papua, and Tukuru
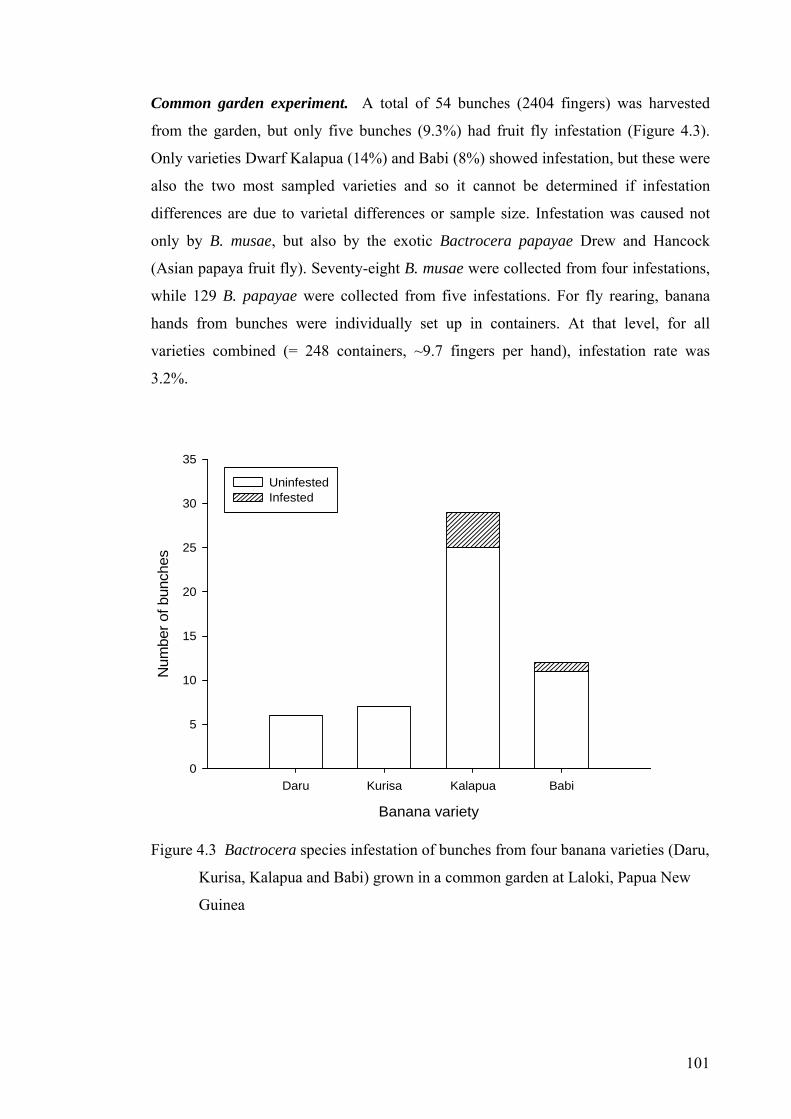
101
Common garden experiment. A total of 54 bunches (2404 fingers) was harvested
from the garden, but only five bunches (9.3%) had fruit fly infestation (Figure 4.3).
Only varieties Dwarf Kalapua (14%) and Babi (8%) showed infestation, but these were
also the two most sampled varieties and so it cannot be determined if infestation
differences are due to varietal differences or sample size. Infestation was caused not
only by B. musae, but also by the exotic Bactrocera papayae Drew and Hancock
(Asian papaya fruit fly). Seventy-eight B. musae were collected from four infestations,
while 129 B. papayae were collected from five infestations. For fly rearing, banana
hands from bunches were individually set up in containers. At that level, for all
varieties combined (= 248 containers, ~9.7 fingers per hand), infestation rate was
3.2%.
Banana variety
Daru Kurisa Kalapua Babi
Num
ber
of b
unch
es
0
5
10
15
20
25
30
35
UninfestedInfested
Figure 4.3 Bactrocera species infestation of bunches from four banana varieties (Daru,
Kurisa, Kalapua and Babi) grown in a common garden at Laloki, Papua New
Guinea

102
4.3.2 National Cavendish survey
The national survey showed that B. musae infestation levels on Cavendish banana were
generally low throughout PNG. Sample infestation was 15.4% in Central, 8.3% for the
Highlands, 7.4% for Morobe and 10.5% for the Gazelle. While at least some bananas
from all agroecological zones were infested, large areas within each zone yielded no
infested fruit (Table 4.3). For example, in the Highlands, 35 samples were collected but
the only infested samples collected were from the Watabung area in the Eastern
Highlands; between Chuave and Goroka. No flies were reared from samples collected
in Chimbu; from Kerowagi to Chuave, nor from sites between Goroka and Aiyura. In
other regions infestation was similarly patchy.
While sampled bananas were not set up as single fingers (logistics precluded this), it is
none-the-less possible to estimate infestation rate at the individual fruit level. For
example, using the data in Table 4.3 and assuming an average of 30 fingers per sample,
it can be seen that for Morobe 19 flies were reared from approximately 900 fingers, an
infestation level of only 2%. This analysis assumes one fly is emerging per banana, but
in practice, however, multiple flies will generally emerge from an infested banana as B.
musae lays in clutches (see results in preceding section and next chapter). At the level
of the individual finger, therefore, banana fly infestation of Cavendish in PNG is
probably well less than 1% during the period November 2007 to January 2008. It is a
time frame when B. musae seasonal abundance is low.
Identification of adult flies was only done for samples collected in Central Province.
Of the four infested samples from Central sample 1 (from Doa) had 1 B. musae, sample
2 (from Veimauri) has 65 B. musae and 42 B. papayae, sample 3 (from Saroakeina)
had 26 B. musae and two B. papayae, and sample 4 from Sivitatana had nine B. musae.
It is unknown if this level of co-infestation occurs in other regions where B. papayae is
established, but it does clearly show that in future work that the invasive B. papayae
has to be considered equally with B. musae as a PNG banana pest.
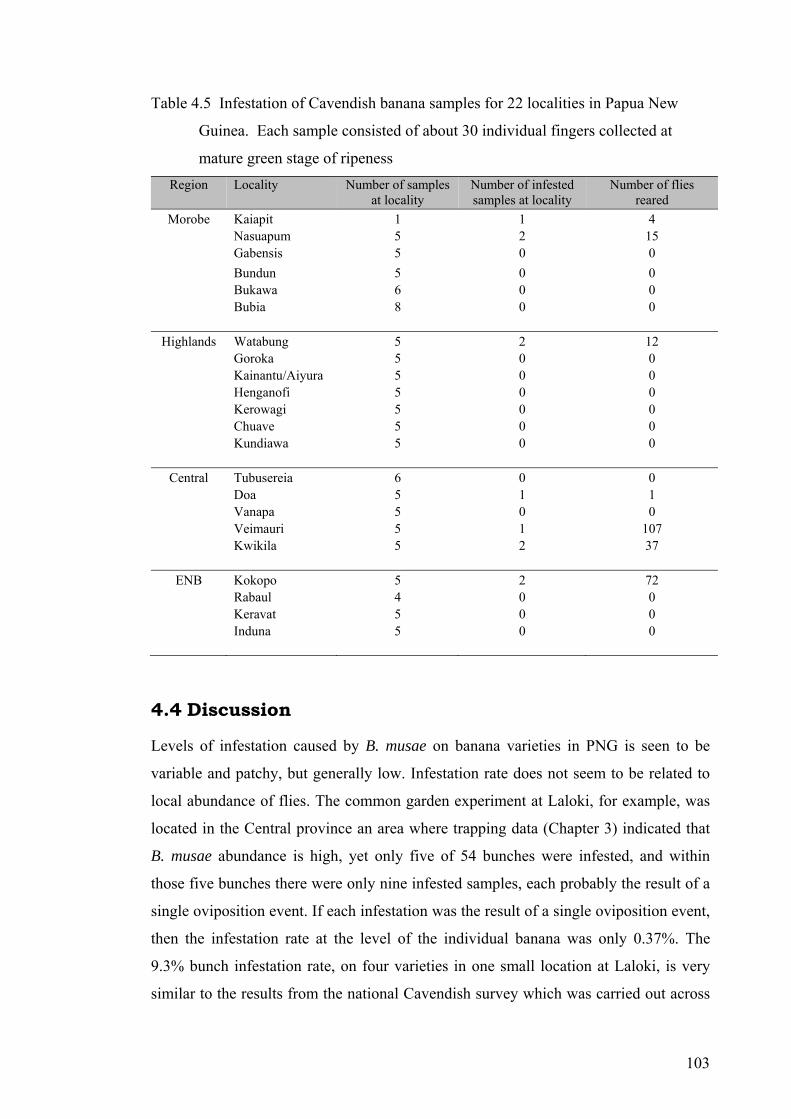
103
Table 4.5 Infestation of Cavendish banana samples for 22 localities in Papua New
Guinea. Each sample consisted of about 30 individual fingers collected at
mature green stage of ripeness
Region Locality Number of samples at locality
Number of infested samples at locality
Number of flies reared
Morobe Kaiapit 1 1 4 Nasuapum 5 2 15 Gabensis 5 0 0
Bundun 5 0 0 Bukawa 6 0 0 Bubia 8 0 0
Highlands Watabung 5 2 12 Goroka 5 0 0 Kainantu/Aiyura 5 0 0 Henganofi 5 0 0 Kerowagi 5 0 0 Chuave 5 0 0 Kundiawa 5 0 0
Central Tubusereia 6 0 0 Doa 5 1 1 Vanapa 5 0 0 Veimauri 5 1 107 Kwikila 5 2 37
ENB Kokopo 5 2 72 Rabaul 4 0 0 Keravat 5 0 0 Induna 5 0 0
4.4 Discussion
Levels of infestation caused by B. musae on banana varieties in PNG is seen to be
variable and patchy, but generally low. Infestation rate does not seem to be related to
local abundance of flies. The common garden experiment at Laloki, for example, was
located in the Central province an area where trapping data (Chapter 3) indicated that
B. musae abundance is high, yet only five of 54 bunches were infested, and within
those five bunches there were only nine infested samples, each probably the result of a
single oviposition event. If each infestation was the result of a single oviposition event,
then the infestation rate at the level of the individual banana was only 0.37%. The
9.3% bunch infestation rate, on four varieties in one small location at Laloki, is very
similar to the results from the national Cavendish survey which was carried out across

104
a much more diverse environmental landscape. Such data reinforces the results for B.
musae in Chapter 3, which suggests that banana fly populations are probably not
driven by local site factors, but are present wherever bananas are grown (resulting in a
standard level of fruit infestation). Such data suggests that trying to identify areas of
low pest pressure for commodity production will not be a particularly useful strategy
for B. musae.
The quality of data was consistently low due to sampling problems and consequently
there were no obvious data sets in this chapter to suggest banana varietal differences in
infestation by B. musae. Infestation level did not differ for three varieties on the
Gazelle Peninsula, and while two out of four varieties at Laloki were not infested, this
could simply have been due to the low sampling effort on those varieties. Such
findings are at odds, however, with Smith (1977) and observations of PNG banana
researchers who believe different varieties are differentially attacked (R. Kambuou per.
comm.).
Additional to variation between varieties, are still questions pertaining to general low
host usage. The Cavendish survey was carried out when seasonal abundance of B.
musae in all surveyed locations was low. Low infestation of collected banana hand
samples by B. musae during this time may be an indication of a ‘low pest population
pressure’.
4.5 Thesis progress
Banana fly is very abundant in PNG (Chapter 3), yet infestation was so infrequent in a
comprehensive survey of Cavendish that no formal analysis of data could be done
because of the excess of zero values. This seems very unusual and, from the point of
view of understanding the risk posed by banana fly as a biosecurity threat, needs to be
pursued. In the following chapter I study in detail host use by banana fly on two
commercial banana varieties, Cavendish and Ladyfinger. In the laboratory I look at
both host selection (i.e. oviposition) and utilization (larval survival) on these two
varieties at three stages of ripeness; green, colour-break and ripe. Understanding and
documenting such behaviour is critical for understanding the factors which influence
B. musae infestation rates on banana. This, in turn, is important in assessing the
potential risk involved in the transportation of the commodity and pre-harvest

105
management of the fruit fly. Both are essential to the second and third part of the PRA
process; Risk Assessment and Risk Management.

106
Chapter 5. Host selection and utilisation by Bactrocera musae (Tryon) on two banana varieties at different ripening stages
5.1 Introduction
Market access for certain crops can be based on non-host status (Follett & Hennessey,
2007) and for fruit flies, non-host or conditional non-host status at a particular stage of
harvest maturity is an internationally recognised phytosanitary measure (FAO, 2005).
While most market access protocols for fruit fly susceptible commodities rely on post-
harvest disinfestation treatments (e.g. heat treatments and irradiation (Jacobi et al.,
1993; Moy & Wong, 2002; Follett, 2004)), more recent research has begun to
investigate the concept of varietal susceptibility, particularly in commercial hosts for
which more than one variety are produced (Follett & Neven, 2006).
Differences in fruit fly infestation levels were found in commercial avocado varieties,
with some varieties more susceptible than others (Hennessey et al., 1995a). Similar
studies have shown that different levels of infestation, and thus susceptibility to fruit
fly attack, also occurs in commercial apple varieties (Bower, 1977), citrus varieties
(Staub et al., 2008), star fruit (Hennessey et al., 1995b), tomato (Balagawi et al., 2005)
and mango (Rattanapun et al., 2009). Subtle differences between varieties of a fruit
may influence fruit fly host utilisation as host selection behaviours are influenced by a
variety of host cues including colour (Dalby-Ball & Meats, 2000b; Brevault & Quilici,
2007), fruit shape and size (Sugayama et al., 1997), smell (Dalby-Ball & Meats,
2000b) and pericarp thickness (Eisemann & Rice, 1989; Balagawi et al., 2005).
Bananas are an international commodity for which several varieties are marketed. The
susceptibility of different varieties to fruit fly infestation is not known. In PNG,
research has indicated Bactrocera musae (Tryon) (banana fly) infestation can be high
on some varieties and low in others (Smith, 1977b). Field observations by local
agricultural officers in PNG also suggest that B. musae infestation, which is most
commonly attributed to the presence of larvae and larval feeding tracks in banana
fingers, is not the same across different cultivars. Smith (1977) suggested plant height
and peel thickness as indicative of the preferential fly behaviour he observed.

107
There are direct implications of banana varietal differences for trying to understand the
biosecurity risk posed by B. musae to PNG bananas. The studies on B. musae
infestation on PNG bananas in Chapter 4 indicate that field infestation levels are much
lower than might be expected for such an abundant, unmanaged fly. One reason for
this may be that the “primary” host(s) of B. musae is not the banana varieties I sampled
or, to put it another way, banana fly may simply not “like” Cavendish and the other
varieties I sampled. While it may initially seem odd to say that banana fly doesn’t like
bananas, it needs to be kept in mind that despite the human applied label of “banana
fly”, we actually know almost nothing about host use of B. musae. The fly is
considered largely host specific to Musa species (Drew & Romig, 1996) and will
attack fruit of both cultivated and wild Musa species (Gold et al., 2002), but it is
crucial to note that available host records show Musa banksii is the major host specie
for banana fly, not Musa x paradisiaca cultivars (May, 1953; Rowe, 1981; Simmonds
& Weatherup, 1990; Hancock et al., 2000). Even the assumption that B. musae is
restricted to Musa species is a largely untested assumption, although laboratory studies
on adult oviposition preferences showed that B. musae had a strong preference for its’
“usual” host, i.e. Musa x paradisiaca over other fruits (Fitt, 1986). Whether that
preference is displayed within its native geographic range is largely unknown, although
host surveys in Australia indicate that Musa banksii is the major host for banana fly
(Hancock et al., 2000), while in PNG the fly has been collected from unidentified
Musa sp. in lowland rainforest (Novotny et al., 2005). If the primary host preference of
B. musae is for wild, non-cultivated banana species, then infestation of cultivated
varieties may only occur because they (both the flies and bananas) are so abundant in
the environment. Whatever the reason may be, the unexpected low levels of infestation
in the field prompts the need for a more detailed understanding of host selection and
utilisation by B. musae.
In addition to preferences between banana varieties or species, another factor may well
be important in influencing host choice in banana fly. Across many species of
Bactrocera, ripeness of fruit is a factor which has been shown to influence the number
of visits to fruit (Messina & Jones, 1990), the number of eggs laid and larval survival
(Rull & Prokopy, 2004), and development rates (Souza-Filho et al., 2009). Generally
all of these attributes are more favourable in ripe fruit, versus green or colour-break
fruit maturity stages (Jang & Light, 1991; Rattanapun et al., 2009). As most

108
Bactrocera exhibit such preferences, harvesting fruit at mature green stage is an
accepted quarantine management strategy (Armstrong, 1983; Armstrong, 2001).
Banana fly seems to be something of an exception to this general rule, as it has been
reported that the fly will infest green bananas. The literature about this, however, is
contradictory. Vijaysegaran (1996) and Drew et al. (1982) report that banana fly will
attack green fruit, with Drew et al. 1982 specifically stating “they can sting even young
fruit as it appears on the bunch.” Other authors, however, qualify this position,
recording that while B. musae is able to oviposit into both immature and mature
bananas, eggs in immature fruit will not develop unless fruit ripens very soon after
oviposition (Smith, 1977b; Fitt, 1986; Gold et al., 2002). Smith (1977), for example,
reports that gravid females were able to oviposit into green bananas at the ‘full’ stage,
but only if the bananas began to colour within the following three days. The
description of ‘full’ that Smith uses may resemble a maturity stage close to what
current industry colour charts (Dadzie & Orchard, 1997; Banana Colour chart,
websites) refer to as the colour-break stage (in particular “Stage 2”)2. This is not the
green and hard maturity stage when bunches are harvested for international trade,
referred to as ‘three quarters full’ (Dadzie & Orchard, 1997). The studies of Smith, and
also May (1953), are applied in commercial banana production in Australia with the
harvesting of green banana bunches an accepted practise for banana fly management
(Fooks, 1989; Pinese & Piper, 1994; Fooks, 2002; Gold et al., 2002). Given this
conflict in the literature, and its impact on biosecurity/pest management practice, a
definitive study on B. musae host use at different ripeness stages is required.
Given the above discussions concerning banana fly host use, both across banana
varieties and within a variety across ripening stages, in this chapter I study both the
impact of variety and maturity stage on B. musae host use. Specifically, in choice and
no-choice arenas I study adult host selection and larval host utilisation of B. musae for
two banana varieties, Cavendish and Ladyfinger, at three ripening stages, green,
colour-break and ripe. I also carry out a trial, following internationally accepted
2 See also standard banana colour chart websites:
http://www.globalfruit.ie/index.php?option=com_content&task=view&id=14&Itemid=21, http://www.catalyticgenerators.com/service.html, http://lib.store.yahoo.net/lib/catalyticgenerators/bananacc.pdf, http://www.bananaland.com.au/info/facts/banana_details_ripening_stages.php.

109
protocols, to confirm the status (either positively or negatively) of green Cavendish as
a host of B. musae.
5.2 Materials and Methods
There were three areas investigated in this study. The first was adult oviposition choice
and subsequent offspring emergence. Tests were done to investigate host selection
behaviour by female flies given choice or no-choice between different banana varieties
and ripeness stages. The second area was on larval host utilisation. The impact of
different banana hosts on juvenile development was calculated by inoculating fertile
eggs under the peel of two banana varieties at three ripeness stages. The third area
looked at was banana fly use of green banana, using standard international protocols.
The test was setup using the specified requirements for testing the host status of a fruit
or vegetable variety at a defined stage of maturity to a given fruit fly species (FAO,
2005).
Experiments were carried out in a controlled environment room (RH 70%, 26°C) at the
Queensland Primary Industries and Fisheries laboratories in Cairns, Queensland. The
banana varieties used were organically produced Cavendish (Williams) and
Ladyfinger, obtained from commercial growers in Tully and Mareeba. Three maturity
stages of banana were used, green, colour break and ripe, and these were identified
using an industry standard maturity colour chart (Dadzie & Orchard, 1997; QDPI,
2004). The B. musae colonies used were 3rd-4th generation flies from locally collected
wild populations (sourced from infested Musa x paradisiaca banana varieties). Flies
were cultured on protein, water and sugar, with oviposition into banana and artificial
egging domes, and larvae reared on fruit fly carrot medium. Flies used in the
experiments were mature, mated 20-25 day old female flies. Most flies were subjected
to a single exposure to artificial and banana egging domes prior to exposure to the test
fruit to encourage oviposition. All observation trials were carried out in 30 x 30 x 30
cm observation cages and flies had access to sugar and water during observation
periods.
5.2.1 Adult host choice and utilisation
Choice tests. Three choice experiments were done. Experiment 1 observed oviposition
choice by a single female fly in a cage when provided with a single ripe finger of

110
Cavendish and Ladyfinger. Experiment 2 observed oviposition choice by a female fly
in a cage with a green, colour-break and ripe finger of Cavendish. Experiment 3
observed the same but of green, colour-break and ripe Ladyfinger. All experiments
were observed at 3 min intervals from 1000-1400 hrs (i.e. an observation was done at
1000am, the next at 1003am, the next at 1006am, etc). There were 16 replicates for all
experiments and fingers were individually incubated for subsequent adult fly
emergence (i.e. 128 containers were used to incubate the samples). Incubation period
was a minimum of two weeks and numbers of emergent adults, plus the number of
observed oviposition events, were the measures of host use analysed.
No-choice tests. In the no-choice tests, a single finger of each ripeness stage of each
variety was placed in a separate cage and left exposed to a female fly for one day
between 10.00am and 3.00pm. There were 16 replicates for each ripeness stage of each
variety. Samples were then individually incubated and adult emergence was recorded
after a minimum incubation period of two weeks. Oviposition was not recorded.
5.2.2 Larval host utilisation
Fertile eggs were collected from B. musae cultures and then artificially inserted into
each variety and ripeness stage tested (as above). Methodology used to collect the eggs
is as described in Balagawi et al. (2005). To insert the eggs under the peel, a 1 cm slit
was made about 3 cm from the flowering end of the banana finger and then pulled back
gently to deposit 60 eggs before resealing the wound with parafilm. Sixteen replicates
were done for each variety and ripeness stage. Fingers were then incubated
individually as for the preceding trials and the number of adult flies emerging from
each recorded.
5.2.3 Host use of green banana
This trial tested the status of green Cavendish fruit as a host of B. musae using
internationally accepted host status tests (FAO, 2005). The protocol prescribed 500gms
of fruit (the nearest whole fruit load was four green banana fingers, slightly more than
500gms) was placed inside a 30 x 30 x 30 cm cage for a period of 24 hours. During
that time the fruit were exposed to 25 mature, female B. musae flies randomly selected
from a culture colony. After 24 hours fingers were removed and individually
incubated. Adult emergence was recorded after two weeks. The trial was replicated

111
five times, with an additional single replicate using ripe Cavendish (a formally
recognised known host) to act as a control (to ensure flies used were gravid and
fertile).
5.2.4 Host plant attributes
Measurements were taken of banana peel toughness, peel thickness, pulp thickness,
peel colour, and sugar content in pulp of Cavendish and Ladyfinger in ripe, colour-
break and mature-green ripeness stages. Procedures for the measurement of each
biological feature were obtained from procedures used in an INIBAP technical
guideline (Dadzie & Orchard, 1997). A Chatillon digital force gauge LTCM-6 and a
hand-held penetrometer using a 0.5mm needle were used to measure peel toughness. A
digital vernier calliper was used to measure peel and pulp thickness. Sugar content,
measured in Brix (%), was recorded using methodology described in the INIBAP
technical guideline (Dadzie & Orchard, 1997) using a hand-held Brix Refractometer
(model: MT-032ATC, measuring range: 0-32% Brix, accuracy: ± 0.2%). The sugar
content is described by a rank for which, in banana, a reading of 8-10 is poor, 10-12 is
average, 12-14 good and 16 excellent3. A Minolta CR-300 colour meter was used to
measure peel colour. Peel colour was measured using the CIE LCH(uv) chromatic
colour system: L stands for luminance (lightness) and is measured vertically from 0
(black) to 10 (white); C stands for chroma (colour purity or colourfulness) and is
measured radially outward from the neutral (gray) vertical axis; and measures the
lightness and “purity” or strength of a colour; lower chroma being less pure, more
washed out, as in pastels); and H is for hue and is measured in degrees around
horizontal circles and measures which colour is most complemented (Wright, 1984).
5.2.5 Analysis
Choice and no-choice trials were initially designed to be analysed using one-way and
two-way ANOVA. Large numbers of zeros, however, and non-homogeneously
distributed errors, which could not be made homogeneous following transformation,
meant that parametric data analysis was not possible. Rather, non-parametric analysis
was carried out to determine if there were significant differences in observed
3 http://crossroads.ws/brix/index-page6.html http://www.honeycreek.us/brix.php.

112
oviposition behaviour and adult emergence numbers between varieties and ripeness
stage. Depending on the test the analyses included the Kruskall-Wallis or the Mann-
Whitney U-test, with the Games-Howell test used as a post-hoc for the Kruskall-
Wallis. Where there were simply too many zeros to allow analysis, data is presented
visually (i.e. it is summarised and graphed). Differences between banana varieties in
individual plant attributes were analysed using one and two-way analysis of variance
tests.
5.3 Results
5.3.1 Adult host choice and utilization
Direct observation of oviposition behaviour under laboratory conditions showed very
few flies attempted to oviposit in either Cavendish or Ladyfinger. Observations
showed that flies did move between fingers within a cage and so were able to make a
choice, but oviposition was still rare. Of the few oviposition events made, not all
attempts (if eggs were laid) were successful in maturing into adult flies. In a choice
arena between ripe fingers of Cavendish and Ladyfinger, no oviposition was made into
Cavendish and only a very small number into Ladyfinger (Figure 5.1). When flies were
given a choice across three ripeness stages within a single variety, overall oviposition
was higher on Cavendish than Ladyfinger, and flies emerged from all ripeness stages
of Cavendish (Figure 5.2). For both these trials the large number of zero values made
statistical analysis inappropriate.
5.3.2 Adult no choice
In the adult no-choice experiment, the Kruskall-Wallis test detected no effect of banana
variety on adult emergence (2 = 0.867, df =1, P=0.360). There was, however, an
effect of fruit ripeness stage (2 = 9.811, df =2, P=0.007), with the post hoc Games-
Howell test detecting that significantly fewer flies were bred from colour-break fruit
than from ripe fruit. Green fruit was intermediate between the other two and not
significantly different from either (Figure 5.3).

113
Banana variety
Num
ber
of o
vipo
sitio
n e
vent
s or
flie
s
0
1
2
3
4
5
Number of oviposition eventsNumber of emergent flies
Cavendish Ladyfinger
Figure 5.1 Number of oviposition events and subsequent number of emergent flies
from single ripe fingers of two banana varieties when offered in a choice arena
to single female Bactrocera musae (n=16 for each variety)
Banana variety and ripeness stage
Num
ber
of o
vipo
stio
n ev
ents
or
flies
0
5
10
15
20
25
GreenGreen Colour-break Colour-break RipeRipe
Cavendish Ladyfinger

114
Figure 5.2 Number of oviposition events (darker shade) and subsequent number of
emergent flies (lighter shade) from single fingers of Cavendish and Ladyfinger
banana varieties at three stages of ripeness when offered in a choice arena to
single female Bactrocera musae (n=16 per variety/ ripeness combination)
Ripeness stage
Num
ber
of f
lies
0
10
20
30
40
50
CavendishLadyfinger
Green Colour break Ripe
Figure 5.3 Number of emergent flies from single fingers of two banana varieties at
three stages of ripeness when offered in a no-choice arena to single female
Bactrocera musae (n=16 banana fingers per variety/ ripeness combination)
5.3.3 Larval host utilization
Non-homogenous errors meant that data from the egg inoculation trials also had to be
analysed using nonparametric techniques. Combining data across varieties, when eggs
were inoculated into fruit of three ripeness stages, adult emergence from ripe and
colour-break fruit was significantly better than from green fruit (Kruskall-Wallis test,
2 = 13.048, df =2, P<0.001, Games-Howell post hoc test). Combining data across
ripening stages, mean adult emergence was greater from Ladyfinger than from
Cavendish (Mann-Whitney U = 677.00, Z = -3.985, P<0.001) (Figure 5.4).

115
Banana variety
Num
ber
of f
lies
0
2
4
6
8
Stage of ripeness
Num
ber
of f
lies
0
2
4
6
8
Green Colour break Ripe
Cavendish Ladyfinger
A
B
A
BB
Figure 5.4 Mean (± SE) number of flies reared from individual fingers of two banana
varieties at three ripeness stages when inoculated with 20 Bactrocera musae
eggs (n=16 inoculated banana fingers per variety/ ripeness combination, 60
eggs per banana)

116
Ripeness stage
Nu
mbe
r of
flie
s
0
20
40
60
80
100
Green Ripe
Figure 5.5 Mean (± SE) number of Bactrocera musae emerging from green Cavendish
bananas following the exposure of 500gm of banana to 25 gravid female flies
(n = 5 replicates). Emergence of flies from ripe fruit (n = 1 replicate) is a
positive control, demonstrating that the flies used to run the trial were gravid.
It should not be used to compare yield of flies from green versus ripe fruit
5.3.4 Host use of green bananas – green Cavendish as a host of Bactrocera musae
Following international protocols for assessing fruit fly host status, green Cavendish
should be considered a host of B. musae as flies were reared from it (Figure 5.5).
5.3.5 Host plant attributes
Six attributes of Cavendish and Ladyfinger banana fingers were measured at mature-
green, colour-break and ripe stages: peel toughness, peel thickness, pulp thickness, peel
colour, and pulp sugar content. The analysis of the importance of variety, ripeness
stage and the Variety x Ripeness interaction for each attribute is presented in Table 5.1
and presented graphically in Figure 5.6.
Peel toughness varied significantly between varieties (greater in Ladyfinger) and
ripening stages (softer in ripe banana). There was a significant interaction effect
between variety and ripening stage, most likely due to a very large drop in peel

117
toughness between colour-break and ripe stage in Ladyfinger, an effect which was
present, but less marked, in Cavendish. Fruit pulp thickness does vary between
varieties or across ripeness stages. Peel thickness does not vary significantly but
changes across ripening stage, generally decreasing as fruit matures. The decline in
peel thickness in ripe fruit was more marked in Ladyfinger than Cavendish. Cavendish
is a heavier fruit than Ladyfinger, the weight of which does not change across ripening
stage. The weight of Ladyfinger, however, declined slightly as fruit ripened. Cavendish
was a sweeter variety overall than Ladyfinger, but what was most dramatic was the
change in Brix across ripening stage. Mature green and colour-break bananas of both
varieties had similar, low Brix percentages, but Brix increased dramatically when the
fruit ripened. Colour measurements (Hue, Chroma and Luminescence) clearly indicate
that each ripeness stage has a distinct colour (Figure 5.6).
Table 5.1 Summary two-way ANOVA output table for Cavendish and Ladyfinger fruit
attributes at three ripening stages
Attribute Source df F P value Peel toughness Variety 1 46.57 <0.00 Ripeness 2 353.71 <0.00 Variety x ripeness 2 160.39 <0.00 Pulp thickness Variety 1 0.96 0.331 Ripeness 2 1.06 0.350 Variety x ripeness 2 3.74 0.228 Peel thickness Variety 1 1.36 0.247 Ripeness 2 131.94 <0.00 Variety x ripeness 2 18.90 <0.00 Chroma Variety 1 12.56 0.001 Ripeness 2 307.98 <0.00 Variety x ripeness 2 79.11 <0.00 Luminance Variety 1 35.02 <0.00 Ripeness 2 426.31 <0.00 Variety x ripeness 2 3.37 0.039 Hue Variety 1 0.13 0.724 Ripeness 2 200.48 <0.00 Variety x ripeness 2 2.44 0.093 Weight Variety 1 204.33 <0.00 Ripeness 2 0.94 0.394 Variety x ripeness 2 5.91 0.004 Table continued overleaf

118
Table 5.2 continued…Summary two-way ANOVA output table for Cavendish and
Ladyfinger fruit attributes at three ripening stages
Attribute Source df F P value Brix (%) Variety 1 26.78 <0.00 Ripeness 2 1785.52 <0.00 Variety x ripeness 2 9.99 <0.00
Gra
ms
0
200
400
600
800
Green C-B Ripe Green C-B Ripe Cavendish Lady-finger
mm
26
27
28
29
30
31
Green C-B Ripe Green C-B Ripe Cavendish Lady-finger
Chr
oma
(C)
x
0
10
20
30
40
50
60
70
Green C-B Ripe Green C-B Ripe Cavendish Lady-finger
mm
2.0
2.5
3.0
3.5
4.0
4.5
Green C-B Ripe Green C-B Ripe Cavendish Lady-finger
Hue
h (
y)
80
90
100
110
120
Green C-B Ripe Green C-B Ripe Cavendish Lady-finger
Lum
inan
ce L
(Y
)
40
50
60
70
80
Green C-B Ripe Green C-B Ripe Cavendish Lady-finger
Peel toughness Pulp thickness
Peel thickness Chroma
Luminance Hue
aa
b
a a
b
aa
b
a
a
b
a
b
c
a
b
c
a
b
c
a
b
ca
b
c
a
b
c
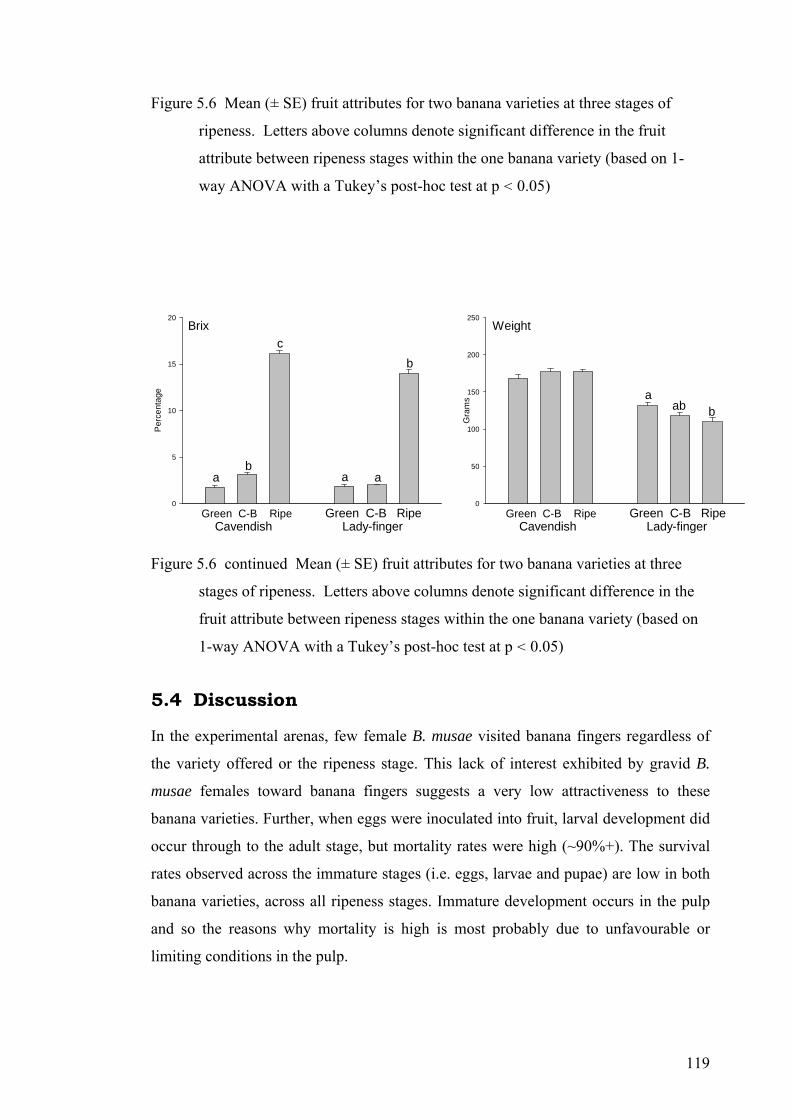
119
Figure 5.6 Mean (± SE) fruit attributes for two banana varieties at three stages of
ripeness. Letters above columns denote significant difference in the fruit
attribute between ripeness stages within the one banana variety (based on 1-
way ANOVA with a Tukey’s post-hoc test at p < 0.05)
Gra
ms
0
50
100
150
200
250
Per
cent
age
0
5
10
15
20
Green C-B Ripe Green C-B Ripe Cavendish Lady-finger
Green C-B Ripe Green C-B Ripe Cavendish Lady-finger
Brix Weight
ab
c
a a
b
aab b
Figure 5.6 continued Mean (± SE) fruit attributes for two banana varieties at three
stages of ripeness. Letters above columns denote significant difference in the
fruit attribute between ripeness stages within the one banana variety (based on
1-way ANOVA with a Tukey’s post-hoc test at p < 0.05)
5.4 Discussion
In the experimental arenas, few female B. musae visited banana fingers regardless of
the variety offered or the ripeness stage. This lack of interest exhibited by gravid B.
musae females toward banana fingers suggests a very low attractiveness to these
banana varieties. Further, when eggs were inoculated into fruit, larval development did
occur through to the adult stage, but mortality rates were high (~90%+). The survival
rates observed across the immature stages (i.e. eggs, larvae and pupae) are low in both
banana varieties, across all ripeness stages. Immature development occurs in the pulp
and so the reasons why mortality is high is most probably due to unfavourable or
limiting conditions in the pulp.

120
Different experiments gave inconsistent patterns with respect to the preferred banana
variety. Of the two varieties studies, adult choice oviposition trials suggest that
Cavendish was the more preferred host, but this pattern is not repeated in the no-choice
trials, while the inoculation trial suggests Ladyfinger is a better host. The simple
interpretation of this data is that under no-choice conditions the flies will lay into any
banana, when given a choice between varieties they show varietal preference for
Cavendish, but once eggs are in fruit larvae do best in Ladyfinger. A preferential
selection by fruit flies between fruit varieties in a choice arena, but which is then lost in
a no-choice arena, is known for fruit flies (Ero, 2009; Rattanapun et al., 2009) and may
be happening with banana fly. However, the generally low oviposition rates and poor
offspring survival makes any definitive interpretation of the data difficult. However,
further studies are needed to provide definite evidence to confirm this. One piece of
hard evidence which cannot be argued against, however, is that in both individual fly
arenas, and following international protocols, green Cavendish fingers harvested at
maturity are stung by B. musae and deposited eggs can develop through to adult flies.
The low oviposition rates and poor larval development rates observed in this chapter
on Cavendish appear to reflect the very low field infestation results obtained in the
PNG Cavendish survey (Chapter 4). This poses the question: Is Cavendish an
unattractive variety to B. musae? If so then this may explain the low damage
assessment levels found during the PNG Cavendish survey. Infestation was also low in
other varieties studied in Chapter 4, as it was in Ladyfinger in this chapter. Such
consistent results, from both the field and lab, suggest that very low levels of host
utilisation is the ‘normal’ behaviour exhibited by banana fly toward Musa x
paradisiaca varieties, i.e. M. x paradisiaca varieties are not major hosts for B. musae.
Infestation, where it does occur, may be because these varieties are simply so abundant
in the PNG environment that even very low levels of host use, infestation will still be
picked up.
Fruit attributes and characteristics are known to influence host selection and utilisation.
For Cavendish and Ladyfinger, fruit attributes of both show similar peel characteristics
which may explain the similar responsiveness or attractiveness received from B.
musae. Varietal characteristics are different in peel toughness, weight and Brix
measure but these differences are much greater between the three ripeness stages.

121
Mature-green and colour break characteristics are more similar to each other in
comparison to the peel attributes of ripe fruit.
For potential export banana production in PNG, there is a large number of cultivated
banana varieties which might be considered. For each of these economically important
varieties, susceptibility to B. musae will be necessary in helping determine those
selected for commercial production. Even at low levels of infestation, commercial
production will still require management protocols and strategies that ensure harvested
bunches are not exposed to B. musae infestation. Nevertheless, evidence accumulating
in this thesis suggests that M. x paradisiaca varieties are not inherently highly
susceptible to banana fly, which will help make other control strategies more effective.
Harvesting bananas at mature green, however, is unlikely to be utilised as a lone
control method, because female flies are able to oviposit into mature green bananas,
but one used with two or more methods in risk reduction strategies.
5.5 Thesis progress Population distribution and abundance of B. musae is related to the availability of its
host (Chapter 3). While B. musae is documented to be attracted to Musa species, within
this genus studies on host selection and utilisation now indicate that female flies are
likely to be selective with respect to species and variety. Such behaviour may explain
the low infestation levels obtained on cultivated Musa x paradisiaca cultivars studied
in this thesis thus far. In the following chapter (Chapter 6), while host selection is not
the chapter’s aim, I do provide further information on how host utilisation impacts on
the fly’s pest status. Chapter 6 studies the impact of B. musae on banana in the field
from an invasive biology perspective. In PNG, the Gazelle Peninsula of East New
Britain province is a novel environment where B. musae is a recently introduced exotic
fruit fly species, and it is in this area that the fly’s distribution, population abundance,
phenology and impact on banana are studied.

122
Chapter 6. Bactrocera musae (Tryon) in a novel environment: banana fly as an invasive organism on the Gazelle Peninsula, Papua New Guinea
Note concerning chapter contents Approximately half of this chapter reports work accomplished and published (by me) prior to my PhD candidacy. I acknowledge that this component of the chapter does not meet the requirements of a PhD, in that the work was not done whilst I was an enrolled student. The chapter, including the previously published material, is still included here, however, because it presents a case-study about an invasive potential of B. musae in a non-endemic environment. The chapter ties the thesis together by providing an example which pulls together the independent studies presented in earlier chapters. The published components of this chapter are presented as Appendix 3 and should not be considered as part of the “original contribution” of this thesis.
6.1 Introduction
Perhaps the single most important aspect of a Pest Risk Analysis (PRA) is predicting
the consequences of how a pest organism will behave in a novel environment and the
impact that it will have on local crops (IPPC, 2006a). While an organism may be
transported along a commodity pathway, simply knowing that transport may occur is
not, of itself, sufficient to then regard that organism as being of biosecurity concern.
Rather, the organism has to have the capacity to establish in the novel environment,
reproduce and spread, and then negatively impact on crops or the environment in that
new range (Vermeij, 1996; Kolar & Lodge, 2001; Maynard et al., 2004; Lockwood et
al., 2005). Knowing the likelihood of these factors allows managers to setup effective
control strategies (Carey, 1996; Simberloff, 2003a; Simberloff, 2003b; Maynard et al.,
2004; Clarke et al., 2005).
Establishment probability is dependent on whether or not ecological factors in the
novel area are suitable. Often invasive organisms are not able to reproduce successfully
and do not establish. However, if they are able to establish the impact they may have
on the novel environment becomes of concern. Much has been documented on the
continuous negative effect that invasive incursions can have on the social, economical

123
and natural environment they invade (Levine, 2008). With respect to tephritids, the
effect that invasive fruit flies have had on the social and economic livelihood of
affected agricultural communities is commonly negative and with management costly
(Drew, 1996; Headrick & Goeden, 1996; Maynard et al., 2004).
Prevention or very rapid management of incursions is the primary aim of biosecurity
organisations around the world, because management of the invasive at the earliest
opportunity is the most efficient and cost effective solution (Perrings et al., 2002;
McAusland & Costello, 2004; Perrings, 2005). As part of this, studies on the behaviour
and impact of invasives in novel environments are useful (Andersen et al., 2004).
Finding out the likely chance of establishment and spread, phenology in the new
environment and the impact that an invasive may have on local production of a
susceptible commodity are all important elements for study. This chapter looks at just
such an example, where a non-endemic fruit fly species has been detected in a novel
environment and has subsequently been tracked post-establishment. Relevant to this
thesis is that the invasive organism is Bactrocera musae (Tryon) (banana fly) and the
invaded region is a locality where banana plays a significant social and economic role,
the Gazelle Peninsula, East New Britain Province (ENB), Papua New Guinea (PNG).
Bactrocera musae was traditionally considered as being absent from the Gazelle and
other PNG island provinces (Drew 1989). It was, however, detected on the Gazelle in
mid-2000 (author’s data). The pathway for entry was not confirmed, but may have
been linked to either food relief supplied after the 1994 Rabaul volcano explosion, or
simply through carriage of infested bananas by travellers (Putulan et al., 2004). I was
the locally based fruit fly entomologist on the Gazelle at that time and subsequently
carried out research on the fly up until the time I commenced my PhD studies. All data
presented in this chapter was gathered before my studies commenced but, with one
exception, had not been analysed. The exception is work on the establishment and
spread of banana fly on the Gazelle which was published prior to commencing studies
(Mararuai et al., 2001) (Appendix 3). Sections from that paper are included in this
chapter because of the insights they provide on B. musae as a biosecurity threat.
The presence of B. musae in a novel environment, such as the Gazelle, provides an
ideal opportunity to provide data on various aspects of its potential invasion biology,
including spread potential and likely impacts. The Gazelle Peninsula is an area where

124
banana, the major and [presumed] preferred host of B. musae, is widely cultivated in
semi-plantation stands; an ideal setting to study the fly from an invasive species point
of view. The impact that this fruit fly species may have on banana in the Gazelle is
unknown, but a priori would be considered serious given the fly’s perceived pest status
on the crop.
In this chapter I collate information which has been gathered on banana fly in the new
environment of the Gazelle. The peninsular has a wet-tropical, lowland environment
similar to most northern coastal areas of mainland PNG (where banana fly is endemic).
Environmental and cropping information for the Gazelle is provided as part of the
analyses carried out in Chapter 3 (Distribution and abundance of five economically
important fruit fly species in Papua New Guinea). Since 2000, B. musae has moved
from being a newly arrived and still spreading incursive on the Gazelle, to being an
established part of the local environment. Information I have collated for this chapter
covers the fly’s distribution and spread, population abundance and phenology of the
established population, and then host infestation levels. All such information is a
crucial and necessary requirement in carrying out a PRA for PNG bananas.
6.2 Materials and Methods
6.2.1 Distribution and spread of Bactrocera musae (Tryon) on the Gazelle Peninsula
Two sets of delimiting surveys were carried out soon after banana fly was detected (i.e.
in 2000) to determine the distribution and population levels of banana fly. These
surveys were: (i) an initial set of three “snapshot” surveys (i.e. very short term) in the
Rabaul and Kokopo areas to identify and confirm incursion; and (ii) a second,
intensive delimiting survey six months later. All trapping was done with modified
Steiner traps (Drew et al. 1982) baited with a mixture of male fruit fly attractant
methyl-eugenol and the insecticide malathion. Trapped flies were sorted to species
level, identified at NARI’s Lowlands Agricultural Experiment Station, Keravat, and
then sent for confirmation of identity to Prof. R.A.I. Drew, Griffith University,
Brisbane. Additional material was screened in the genetics laboratory of Prof J.
Hughes, Griffith University, and confirmed to be genetically similar to known material
of B. musae from North Queensland.

125
The initial set of surveys was carried out between late 1999 and mid 2000 to confirm
the incursion of B. musae. They covered areas from the coast to the hinterland of the
Gazelle Peninsula (Figure 6.1). Survey one ran from November to December 1999,
covering the Kokopo town area, and had 23 trap locations setup in both residential and
commercial areas. Survey two ran from May to June 2000 along the northern coastline
of the peninsular from Rabaul to Tavilo. Traps were set at 11 locations in village
residential areas and vegetable gardens. Survey three ran from June to July 2000, and
covered the mountain plateau areas of Malmaluan, Nangananga and Raluan about 5km
inland from the coastline between Rabaul and Kokopo towns. Traps were established
at 13 trap locations covering village residential areas with many areas under vegetable
cultivation.
The delimiting survey was an intensive survey of the Gazelle Peninsula which was run
in December 2000. The aim of the survey was to determine, as far as possible, the
distribution of banana fly at that time. Sixty-one traps were distributed covering the
major road networks on the peninsular at approximately 10km intervals. The trap
network extended along the northern coastline from Tavui to Lassul (04°13’S,
151°43’E), along the southern coastline from Kokopo to Gar in the Sum Sum Bay
(04°42’S, 151°21’E) and further inland from Kokopo toward Warangoi (04°29’S,
152°09’E) and as far as Riet (04°34’S, 152°05’E) at the base of the Baining Mountains
(Figure 6.1).
In addition to presenting the data from these two surveys, I also present the current
known distribution of banana fly on the Gazelle. This information is not based on a
single formal trapping program, but is a collation of information from different sources
including growers, research colleagues and personal observations made during survey,
experimental and farmer out-reach work.
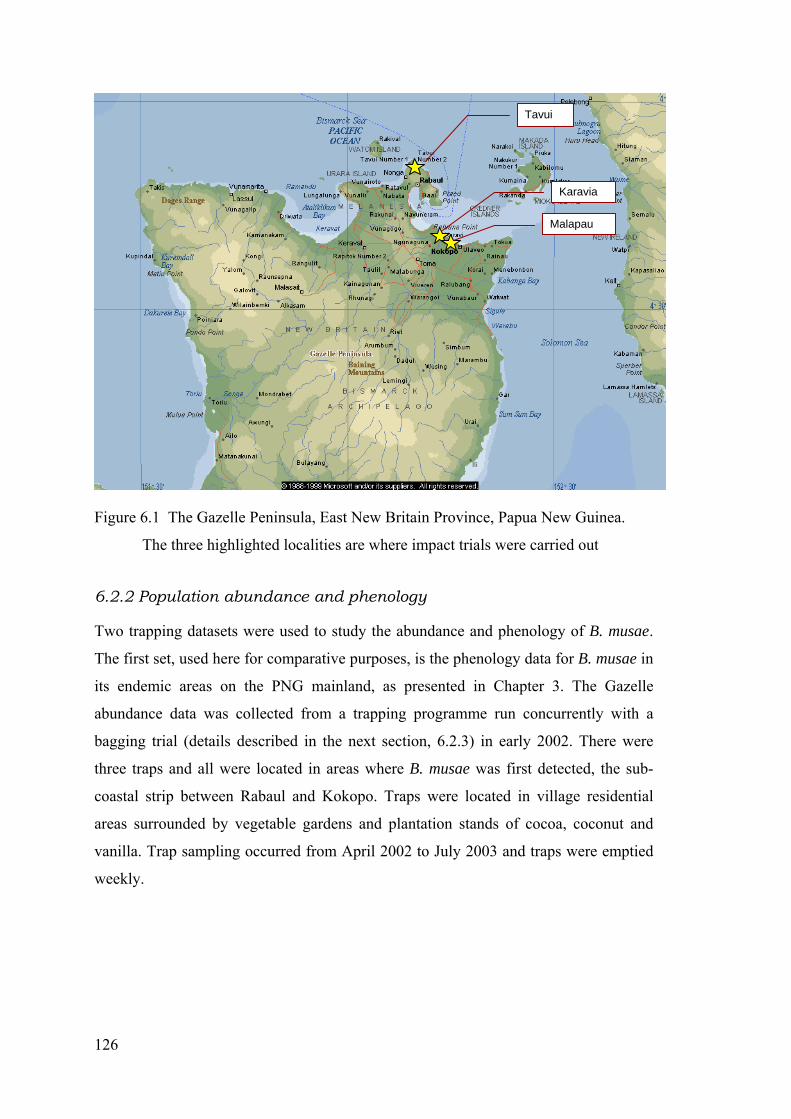
126
Figure 6.1 The Gazelle Peninsula, East New Britain Province, Papua New Guinea.
The three highlighted localities are where impact trials were carried out
6.2.2 Population abundance and phenology
Two trapping datasets were used to study the abundance and phenology of B. musae.
The first set, used here for comparative purposes, is the phenology data for B. musae in
its endemic areas on the PNG mainland, as presented in Chapter 3. The Gazelle
abundance data was collected from a trapping programme run concurrently with a
bagging trial (details described in the next section, 6.2.3) in early 2002. There were
three traps and all were located in areas where B. musae was first detected, the sub-
coastal strip between Rabaul and Kokopo. Traps were located in village residential
areas surrounded by vegetable gardens and plantation stands of cocoa, coconut and
vanilla. Trap sampling occurred from April 2002 to July 2003 and traps were emptied
weekly.
Tavui
Karavia
Malapau

127
6.2.3 Impact Studies
Market Surveys (2000-2001)
To measure the level of infestation being caused by the banana fly incursion at the time
of the initial incursion, bananas were purchased from local roadside markets and
collected from the Lowlands Agricultural Experiment Station banana plots and from
gardens in the Rabaul and Kokopo areas. Fruit were set in individual containers and
kept to assess the level of fruit fly infestation as described in Leblanc et al. (2001)
(Appendix 3). Rearing occurred at intervals over a twelve-month period from June
2000 onwards. All reared flies were identified by R.A.I. Drew.
Bagging trial (2001-2003)
Host infestation levels were assessed from a banana field control trial carried out
between March 2001 and July 2003. The trial established three banana plots in separate
locations, but all in areas with high B. musae abundance. The trial was a collaborative
effort with local farmers aimed at demonstrating the procedure and benefits of bagging
against fruit fly infestation. Plots were established at Tavui, Karavia and Malapau
(Figure 6.1). The banana variety chosen for study, Tukuru (PNG118 Musa Eumusa
ABB) (Arnaud & Horry, 1997), is a commonly consumed and marketed variety on the
Gazelle and one of important cultural value.
At each plot, 20 banana bunches were bagged (i.e. physically protected from banana
fly) and 20 were not bagged (i.e. exposed to banana fly). The type of bags used were
polythene bags used in the packaging of fresh or dried cocoa beans, a widely available
and non-costly option for subsistence and semi-commercial farmers. All bagged
bunches were bagged at the mature green stage, about six weeks after the appearance
of the inflorescence bulb. Bagged and un-bagged bunches were then harvested at
maturity and banana samples were setup over moist sawdust. Incubation period was a
minimum of two weeks during which pupae and adult emergence were observed and
recorded.
Current damage
To get an estimate of current levels of damage this chapter uses information from the
Cavendish survey (Chapter 4: Infestation of bananas by Bactrocera musae (Tryon) in

128
Papua New Guinea). It also presents some informal information gathered from
colleagues.
6.3 Results
6.3.1 Distribution and spread of Bactrocera musae (Tryon) on the Gazelle Peninsula
On the Gazelle, B. musae was first collected from sites just behind Kokopo (e.g. 40
flies from 15 clearances of a trap hanging at the Vunamami Farmer Training Centre
(04°21’S, 152°13’E)). At the same time isolated individuals were trapped at more
distant localities, such as Keravat, and these probably represented dispersing flies. The
absence of flies in surveys prior to its detection (Leblanc et al., 2001) and the lack of
local cultural control methods against fruit fly infestation on banana supported the
conclusion that B. musae is an invasive on the Gazelle Peninsula.
Snapshot surveys confirmed the presence of B. musae on the Gazelle. The flies
distribution when first surveyed in mid-2000 showed the population to be radiating
from Rabaul and Kokopo (Figure 6.2). The initial distribution was limited to areas
along the coastline with high abundance areas in and around Rabaul and Kokopo. By
late 2000 the distribution had spread further inland and along coastal areas (Figure
6.3). Traps were able to catch few flies in areas further along the northern coastline
toward Lassul and along the eastern coastline toward Gar. Toward the hinterland,
samples were collected in areas such as Warangoi and Reit. To my knowledge the
current distribution of banana fly on the Gazelle covers all parts of the Gazelle up to
the Baining Mountains (Figure 6.4).

129
Figure 6.2 Distribution of Bactrocera musae (Tryon) on the Gazelle Peninsula, East
New Britain, Papua New Guinea in mid 2000. Source: Mararuai et al. (2001)
Figure 6.3 Distribution of Bactrocera musae (Tryon) on the Gazelle Peninsula in
December 2000. Source: Mararuai et al. (2001)
0 flies
Keravat
RABAUL Lungalunga
Tavui No. 2
KOKOPO
Karavia No. 2
Toma
Gazelle Peninsula 11-50 flies
1-10 flies
>50 flies
Tokua
0km 20km
0 flies
1-10 flies
11-50 flies
>50 flies
Toma
Lungalunga
Riet
Vunamarita
Keravat Kokopo
Watwat
Gar
Gazelle Peninsula
0km 20km

130
Figure 6.4 Distribution of Bactrocera musae (Tryon) indicated by red pins on the
Gazelle Peninsula in June 2009; pins mark village residential areas surrounded
by vegetables gardens, plantations, secondary or primary rainforest
6.3.2 Population abundance and phenology
On the Gazelle, B. musae populations fell in January and remained low until July,
when the population began to rise, with a nearly continual increase through to
December (Figure 6.5). This trend is somewhat similar to that shown by banana fly in
the Highlands between January and July. However between August and December
seasonal abundance remains high while the curves in Morobe and the Highlands fall
distinctly. Analysis in Chapter 3 suggests that weather variables do not have a major
influence on the population dynamics of B. musae and so cultivation practices for
banana may be responsible for the differences observed. If this is the case for the
mainland sites, it may also be the case for the Gazelle. The mean monthly abundance
levels of B. musae on the Gazelle between April 2002 and July 2003 were quite large
and similar to levels in areas on the PNG mainland.

131
0
200
400
600
800
1000
1200
1400
1600
1800
Jan
Feb
Mar
Apr
May Ju
n
Jul
Aug Sep Oct
Nov
Dec
Time (Months)
Ave
rag
e m
on
thly
tra
p c
atc
h
Highlands
Morobe
Central
Gazelle
Figure 6.5 Phenology curve of Bactrocera musae (Tryon) on the Gazelle Peninsula
(East New Britain, Papua New Guinea (PNG)) (from April 2002 to July 2003)
compared with curves in three areas on the PNG mainland (from 1999 to 2001)
6.3.3 Impact studies
6.3.3.1 Fruit rearing (2000-2001)
From fruit collected from 28 roadside markets along the north and south coasts of the
Gazelle, 6% infestation was recorded, caused equally by B. musae and Bactrocera
frauenfeldi (Schiner). Fruit collected from 18 field locations in May and June 2001
yielded fewer flies, with less than 1% infestation rate (Table 6.1). However, comments
from village farmers during this collection were pertinent; for example “Bananas have
to be harvested earlier than usual to prevent them getting damaged”. When infested
fruit was collected, infestation of individual fingers was found to be heavy. One sample
of seven ripe fingers, weighing 1.76 kg and collected from Tavui No 3 village (behind
Rabaul town) on 12 April 2000, yielded 418 B. musae pupae.
6.3.3.2 Bagging trial (2001-2002)
The infestation impact of B. musae on bagged and un-bagged Tukuru was almost the
same. There were 12 bagged and 12 un-bagged samples collected from each plot at
Tavui, Karavia, and Malapau (i.e. a total of 36 bagged and 36 un-bagged bunches).
Only one un-bagged bunch harvested from Malapau was infested (Figure 6.6). These

132
infestation levels are despite the presence of a large wild population in the
environment.
Table 6.1 Tephritid fruit flies reared from bananas purchased from markets (March to
June 2000) or collected from gardens (May to June 2001) on the Gazelle
Peninsula, East New Britain, Papua New Guinea
Figure 6.6 Infestation of banana bunches protected (bagged) or unprotected
(unbagged) from Bactrocera musae (Tryon) on the Gazelle Peninsula, East
New Britain, Papua New Guinea
Market Surveys Garden Surveys Number of banana varieties sampled 15 11 Number of markets/gardens visited 28 18 Fruit fly species collected Bactrocera frauenfeldi
Bactrocera musae Bactrocera frauenfeldi
Bactrocera musae % infestation by both fruit fly species 6% 0.8% % infestation by Bactrocera musae 3% 0.2% Most common varieties sampled Yawa Kiakiau, Yawa,
Tukuru, Katkatur, Chinese Tall
Total weight of banana sampled 19.6 kg 36.8 kg Number of banana fingers set up Not recorded 393 Average development stage of bananas sampled mature green mature green

133
6.3.3.3 Current status
The current levels of damage caused by B. musae across the whole peninsular have not
been formally documented. While on a field trip to PNG in July 2007, I learned from
local sources that areas of the Gazelle along the northern coastline around Vudal and
Vunapalading may be hot spots for B. musae. Farmers in those areas had begun to
notice fruit fly larval damage on banana and were enquiring at the Lowlands
Agriculture Experiment Station (Keravat) for staff to determine the cause and advise of
control options. Farmers were advised to bag their bananas and demonstrations were
provided in gardens and/or at the agriculture station. That farmers are now seeking
advice to control the pests implies that, for at least some districts, damage levels are of
sufficient level to concern growers. In contrast to this, however, is the Cavendish data
(Table 6.2 and Chapter 4), for which only two samples from 19 yielded any flies. This
low level of infestation much better reflects the earlier results of the bagging
experiment and market survey.
Table 6.2 Infestation of mature green Cavendish banana by Bactrocera musae (Tryon)
at four locations on the Gazelle Peninsula, East New Britain, Papua New
Guinea, in 2007
Location Sample Weight of sample (gms)
Number of fingers Number of flies
Kokopo 1 5300 38 0 2 3950 29 3 3 3430 30 0 4 3180 34 69 5 4730 39 0 Rabaul 1 3180 28 0 2 2940 27 0 3 5530 34 0 4 4130 24 0 Keravat 1 2910 28 0 2 4460 29 0 3 4650 46 0 4 3600 30 0 5 5060 41 0 Induna 1 4840 34 0 2 2960 30 0 2 2890 37 0 4 6800 39 0 5 3200 22 0 Total 19 77,740 619 72

134
6.4 Discussion
This chapter, on an invasion by B. musae, identifies a species that has arrived in a
novel area, become established and spread from the original point of arrival.
Ecological factors in the environment have obviously been favourable for B. musae
establishment, while the initial population was also obviously large enough to allow
establishment. Sampling for B. musae on the Gazelle shows banana fly populations are
as large as endemic mainland sites, but the impact of those flies on local banana
production is apparently minimal. Almost ten years post incursion (at least seven since
first detection), it is only recently that I have heard of banana farmers observing
damage and enquiring for appropriate control measures; and then only in certain areas
on the peninsular. Whether impact has been low due to limitation by local cultivation
practices is uncertain, but that may have an influential role. On the Gazelle farmers
harvest at maturity and consumption is often immediate, meaning that the fruit may be
picked and consumed before they are at the stage most vulnerable to fly infestation. Or,
again reinforcing the point made in both Chapters 4 and 5 (Infestation of bananas by
Bactrocera musae (Tryon) in Papua New Guinea), commercial banana varieties may
be poor hosts of banana fly and generally little affected by it (at least in an
environment where more preferred Musa species or cultivars may be apparent). Further
studies into the biology and ecology of B. musae in this novel environment would
further improve our understanding of the invasibility of B. musae in this novel
environment. It would also provide information on the invasiveness of such a
Bactrocera species, one with a narrow host range, in a novel tropical environment.
With respect to information for developing a PRA, what does the incursion into East
New Britain tell us? Firstly, that flies can be transported along some pathway, in
numbers sufficient to lead to local establishment. What that pathway was is uncertain,
but Putulan et al. (2004) argue that simple carriage of infested bananas was a likely
pathway. Data from their work identified that one in every 100 airline passengers
landing on the Gazelle (i.e. about one person a day) would be carrying an infested
banana. That the flies must have been carried to the Gazelle, rather than dispersing
naturally, is evidenced by the fact that West New Britain Province, much closer to the
mainland, remains free of the fly (Mararuai et al., 2001), inferring that the fly has
“hopped” (via a pathway) to the Gazelle. Having established a bridgehead, the fly very
rapidly expanded its range, occupying substantially larger areas in only six months

135
between the first phase and second phase delimiting surveys. This rapid spread meant
that eradication plans for the fly were abandoned (A. Allwood 2001, unpublished
report), and poses a serious biosecurity concern for regions where the fly might
potentially enter. In short, this data suggests that if the fly does enter and establish in a
new area, eradication needs to be attempted immediately if hoped to be successful. The
final issue for a PRA is to do with impact. While banana fly is a recognised pest of
banana fruit, on the Gazelle infestation levels are low in experimental trials, surveys
and gardens. Part of this may be due to banana varietal effects and management
practices, but it again reinforces the data from other chapters that banana fly is not of
major concern in PNG.

136
Chapter 7. Discussion
7.1 Thesis summary
7.1.1 Introduction
Banana is an important staple food crop in PNG, as well as being an important revenue
generating crop. Improved crop production methods and the setting up of pathways for
crop marketing are recognised mechanisms by which improved socio-economic
development can be achieved for PNG’s subsistence and semi-commercial farming
population. With the PNG National Government’s intentions of promoting banana fruit
as an export commodity, international phytosanitary guidelines on market access
protocols must be adhered to if export is to occur (IPPC, 2005, 2006a, 2008). The Pest
Risk Analysis (PRA), a three step process used for identifying phytosanitary risk and
risk mitigation treatments, is a key element of those protocols (IPPC, 2006b, 2007). A
PRA evaluates scientifically generated information to determine whether an organism
is a pest of quarantine concern for a commodity importing country or region, defines
the probability of introduction, establishment and spread of the pest in that region, and
risk management options that can reduce the risk to an acceptable level (IPPC, 2007).
This thesis carried out work to help assist the PRA process for PNG banana fruit
exports. A summary of the thesis results follow, while a full description of how the
results fit within a PNG banana PRA after that is presented.
7.1.2 Summary
Evaluation of agriculture research reports, published literature and expert opinion for
pests of banana in PNG found that there are 112 organisms associated with this crop;
consisting of pathogens, insects, nematodes and weeds. Fourteen of these species are
commonly managed. Reports on the local research done on these 14 cover aspects in
biology, damage assessment, and applicable methods for their control however there is
little information in comparison available for B. musae, a fruit fly species documented
as the major pest of banana fruit. Information supplied in this thesis fills key aspects of
the previously missing information about this pest.
Analysis of trapping surveys carried out in PNG show that the detailed distribution of
B. musae in PNG is contrary to that previously reported (e.g. Drew 1989). The fly

137
species is widely distributed across mainland PNG, but is absent (except as a recent
incursive) on the PNG island provinces of the Bismarck Archipelago (i.e. New Britain,
New Ireland, Manus and Bougainville Islands). On the mainland the fly is found from
sea level to above 1800 m.a.s.l. Host availability is weakly, but significantly correlated
with population abundance, while the abiotic factors of altitude and rainfall are not
significantly correlated with banana fly population abundance.
Previously reported as a major (often “the major”) pest of PNG banana fruit,
information reported in this thesis contradicts this assessment. Results gathered from
damage assessment surveys, field trials and detailed laboratory work, present a
consistent outcome that unambiguously point to the fact that cultivated banana
varieties (at least of the varieties assessed here) are not the preferred or primary host of
B. musae. This judgement is based on the uniformly low levels of field infestation, in
both endemic and introduced parts of the species’ range, and very poor host selection
outcomes in lab based studies. Laboratory trials found the fly has a preference for ripe
fruit over mature-green and colour-break fruit and, while able to oviposit and develop
in mature green fruit, probably does so only rarely in the field. Similar, but very low
rates of oviposition and adult emergence from Cavendish and Ladyfinger banana,
strongly suggests that both these varieties are poor hosts for the fly. Very high
abundances of banana fly in PNG must be generated from Musa species, or Musa x
paradisiaca varieties, not studied in this thesis.
7.2 PRA for PNG Banana
7.2.1 Definitive statement of IPPC PRA process
Organisms associated with a commodity intended for international commodity trade
are studied to determine if they may pose a social, economical or environmental threat
if transported from the site of production and establish in the region of sale (IPPC,
2004a, b). The studies must adhere to standardised requirements and be reported as
specified by international guidelines (IPPC, 2002a). The study of each organism of
interest is important because their biology will describe the method through which the
organism attacks or causes damage, while ecological information on factors
influencing survival and reproduction is essential information in their management
(Allwood, 1996a). Due to the continuous threat of invasive organisms transported via
commodity trade, such information enables the setting up of processes and approaches

138
to minimise pest movement, and to predict and prevent their establishment (Cook,
2008).
Reporting of information for a PRA has standardised outlines, with specific
requirements for necessary elements (IPPC, 2002a, 2007). The framework must
include verification of the purpose of the PRA, stating the pathway of concern that may
harbour and transport potential risk organisms to the destined import market
environment (IPPC, 2007). Main elements for documentation require identification of
the target organism, or in my case the banana variety selected for trade, and the area
where production will occur. Required information on biological attributes of
organisms associated with the commodity is their ability to cause damage, their host
range, and method and mode of infestation. Information on the evidence and detailed
analysis on social, economic and environmental impact and the sources from which
such information is obtained is important. The report should also contain the
conclusion obtained from the completed PRA and the decisions and justifications
made. It should clearly state the pest risk management measures identified, evaluated
and recommended and the dates of when the PRA was done, by whom and reviewed
by which authoritative individual(s) and reviewer(s).
7.2.2 Summary on PNG banana PRA
The purpose of a PNG banana PRA would be to determine risks involved in moving
unwanted organisms via the production, storage and transportation of banana from
PNG to an international market destination. The PRA would be initiated for the
assessment of a commodity, or pathway, as referred to in the IPPC guideline not a
certain pest and thus is recognised as a ‘pathway initiated analysis’ (IPPC, 2007). The
report requires certification of the processes carried out, verification of the supervising
national quarantine authority and names of authors, contributors and reviewers
responsible and associated in producing the report.
The PNG banana PRA would require a morphological and taxonomical description of
the banana variety(s) selected for commodity production. Other necessary information
required include: areas identified for commodity production, distribution of and
cultivation practices for the variety, non-commodity varieties present in the producing
area, and use of the commodity variety in the producing area. A categorised pest list
would require the current 112 organisms identified associated with banana to be sorted

139
by pest status. The fourteen commonly reported organisms may be reported as pests,
but they also require categorisation as to whether pest status is that of a quarantine
pest. This list, however, is not specific to a certain banana variety and therefore is best
utilised as a reference point when studying organisms associated with the variety(s)
selected for commodity production. Verification of areas where production would
occur is important and a detailed study in the locality would be required. Research in
PNG on those fourteen species provides information on biological, ecological and
management details, but some of the reported work is not current and an update would
be appropriate. Studies would need authentic investigation, be formally reported and
should be readily accessible to the international community. Economic impact
assessment is an area which needs more focus. Documentation and the use of
standardised methodologies and processes are important to verify the impact of
damage on produce quantity and quality even for those species that do not cause
serious problems and to allow for comparison across different banana varieties.
Assessment procedures and methodologies are important for transparency in report
writing and for trade partners and is an area which future assessment reports must
provide. It is also important to note that the collecting of information on the organisms
associated with banana is a continuous activity. Investigations carried out in this thesis
provide the following information for the PRA and shows areas that need attention
(Table 7.1).
7.2.3 Detailed PRA response for banana fly
Within the context of a full PNG banana PRA as described above, and using the three
step outline of the PRA, this thesis has found the following for banana fly:
Step 1 Risk Initiation. Banana fly is a presumed oligophagous fruit fly species on
Musa species. Reported as a major pest of commercial banana (Musa x paradisiaca
varieties), this thesis has shown that due to the consistently low infestation levels on
cultivated varieties reported pest status of banana fly is incorrect and that it should be
regarded as a minor pest of cultivated banana varieties in PNG. In the tropical
environment of PNG, the importance of banana in local cropping systems is weakly,
but significantly correlated with banana fly population abundance, while temperature
and rainfall have no significant influence. The distributional range of the species is
from sea level to above 1800 meters in elevation and endemic to the PNG mainland.

140
The inference of results presented here are that bananas grown anywhere in PNG are at
equal, albeit low, risk of banana fly infestation.
Table 7.1 Checklist of information available and necessary for carrying out a pest risk
analysis of PNG bananas; a pathway initiated analysis
Requirements Information available Information needed Commodity description Banana varieties in PNG
currently documented Selected variety(s) for commodity production
Categorised Pest list 112 organism identified, 14/112 commonly managed
Categorisation list – quarantine pest, non-quarantine pest
Evidence of economic impact, which includes environmental impact
No detailed or specific information available for economic and environmental impact
Update needed for 14 commonly managed pests, Environmental impact studies
Conclusions of pest risk assessment (probabilities and consequences)
Not yet done
Decisions and justifications to stop the PRA process
Not yet done
Pest risk management: phytosanitary measures identified, evaluated and recommended
Not yet done
Step 2 Risk Assessment. Fly larvae can be transported via banana and establishment in
novel areas will be influenced by the presence of susceptible hosts. Based on the
known incursion in East New Britain province, establishment of the fly in non-endemic
areas may readily occur in the presence of hosts, with spread from the point of
establishment rapid. Ripe banana fingers are the preferred maturity stage for
oviposition, but mature-green and colour-break fingers are able to sustain banana fly
immature development through to the adult stage. Wild banana fly populations can
occur in relatively large numbers, but the impact that such populations may have on the
commodity will be dependent on the banana variety and stage of ripeness at harvest.
Further study is required to determine the impact of banana fly on a wider range of
cultivated varieties because studied varieties are not heavily infested, suggesting these
varieties may not be the primary host. Other banana varieties may be heavily infested.
This is a particular risk as export of banana from PNG may focus on less common
banana varieties which target specialist and gourmet markets.

141
Step 3 Risk Mitigation. The research done in this thesis suggests that there will only
be a few possible risk mitigation techniques. The very high abundance of flies, but
very low infestation rates, means that traditional pre-harvest control techniques (e.g.
cover-sprays, male annihilation, protein bait spraying, crop hygiene) (Allwood, 1996b)
are unlikely to substantially influence infestation rates, as flies are obviously breeding
in non-managed areas. Management of flies in the immediate crop area is thus likely to
cause little or no reduction in the total background fly populations and, hence, no
change in the already low infestation rates. Harvesting bananas at mature green, a
currently recognised protocol, should be maintained, although I show that this cannot
be regarded as a definitive, single-step treatment as flies can oviposit into, and develop
through, on mature green bananas. Bagging of banana bunches does appear an
effective management strategy, with no infestation of bagged bunches in the trial run
on the Gazelle Peninsula. Low levels of infestation in unbagged controls, however,
mean that further trials would need to be run before this approach could be used with
confidence. Nonetheless, fruit bagging is a widely used fruit fly management tool,
particularly in countries where labour is relatively inexpensive (Allwood, 1996b).
Given the field infestations are so low, single step post-harvest treatments (e.g.
insecticide dipping, heat treatments) (Armstrong, 1996; Neven & Drake, 2000; Shellie
& Mangan, 2000) are likely to be the most effective phytosanitary protocol for banana
fly, as there is little or no need for in-field, pre-harvest controls..
7.3 Implications of thesis for wider fruit fly market access issues
The abundance of an insect pest is often assumed to be directly and positively
correlated to the crop damage caused by that pest (Drew et al., 1984; Alyokhin et al.,
2001; Meats et al., 2003). The international phytosanitary protocols of “areas of low
pest pressure” (IPPC, 1995, 1997, 1998, 1999, 2002b) operate under the assumption
that low levels of pest abundance will result in low levels of crop infestation.
Alternatively, areas where a pest is well established require commodity treatment
protocols (Armstrong, 1996; Carey, 1996; Armstrong, 2001; Follett, 2004; Stice et al.,
2007), based on the assumption that commodities will be infested. However, the results
obtained in this thesis for B. musae on banana in PNG shows that levels of fly
abundance need not relate to host infestation levels. Low banana finger infestation

142
levels in areas with very large banana fly populations raises questions about the
certainty of predicting crop damage levels from population abundance (e.g. trap) data.
7.3.1 Trap abundance and host use by fruit flies
Detection and monitoring (leading to subsequent management) of pest Bactrocera
species, such as B. musae, is almost entirely done through adult trapping (Cowley,
1990; Rössler et al., 1998; Broumas et al., 2002; Hollingsworth et al., 2003; Meats et
al., 2003; Meats & Clift, 2005; Burrack et al., 2008). Fruit fly traps utilise two lures
that are recognised by adult male flies, with a particular fruit fly species responsive to
only one lure type (Metcalf et al., 1975; Brieze-Stegeman et al., 1978; Hooper, 1978).
Maturing and mature adult male flies forage for, and are positively attracted to,
naturally occurring plant chemicals and these chemicals are identical, or very similar,
to the lures used in traps (Fletcher et al., 1975; Chuah et al., 1997; Nishida et al.,
2004). Female flies do not generally respond to these lures and are rarely caught in
fruit fly traps (Metcalf et al., 1975; Hill, 1986). Traps for most Bactrocera species are
very effective and are able to attract adult male flies within a radius of 500m (Metcalf
& Metcalf, 1992; Jang & Light, 1996). Such traps allow for the collection of
information on male fruit fly distribution and abundance in different habitats and
cropping systems. It is a general tacit assumption of fruit fly trapping that measures of
male abundance are positively correlated with assumed female abundance.
Abiotic factors such as temperature, rainfall and host availability all influence the
population dynamics of tephritid species (Bateman, 1972; Allwood, 1996a). Biotic
environmental factors such as amount of canopy cover (Dalby-Ball & Meats, 2000a;
Raghu et al., 2004), natural sources of male parapheromones (Jang & Light, 1996),
olfactory chemicals (Flath et al., 1990; Meats & Osborne, 2000), and food resources
(Drew et al., 1983; Fletcher, 1987; Raghu, 1998) also influence total population
abundance, as well as the local densities of flies within a larger population. While all
such factors may influence the abundance of flies, the effect that each may have on a
local population of a particular fruit fly species may vary. This is illustrated with the
correlation analysis in Chapter 3, where the correlation of altitude and rainfall on
population levels of five fruit fly species all varied. Most importantly, of all the factors
which may influence the abundance of a local fly population, it is only one of these,
host availability, which is directly related to crop impact (i.e. flies utilising a

143
commodity for breeding will impact upon it). It is equally important to note that fruit
fly traps attract males, yet is the unstudied females which cause crop damage. Thus is it
perhaps not surprising that there need be no obvious relationship between fruit fly trap
catch and crop infestation, as in the case of banana fly.
For fruit fly susceptible commodities, host impact is the damage resulting from feeding
by fruit fly larvae. The larvae hatch from eggs oviposited by female flies. Host
selection is a complex and time-consuming process of search and investigation, before
selection and oviposition, and is influenced by olfactory, semiochemical and physical
cues present in the habitat in which the parental female forages (West & Cunningham,
2002; Bruce et al., 2005). Having said that, with respect to banana fly in PNG, why
does an abundant and widespread fly not routinely use a common and widespread
host?
Banana fly is a narrowly oligophagous fruit fly species (Fletcher, 1987) and has a
preferential host utilisation and selection behaviour (for Musa spp) characteristic of
specialist herbivores (Fitt, 1986). Specialist herbivores are more sensitive to their
environment and are more affected by host-plant resistance mechanisms and variable
host quality than are generalists (Tscharntke et al., 2001). Plant attributes which are
known to influence tephritid host use include characteristics such as colour, sugar
content, and peel hardness or toughness (Bateman, 1972; Fletcher, 1987), all of which
are highly variable across banana species and cultivars (Osuji et al., 1997; Lucas et al.,
2000). It is thus possible (likely?) that particular banana varieties may be highly
susceptible to banana fly, just not the varieties studied here.
Population dynamics of specialist herbivores are also considered to be less influenced
by abiotic conditions and more by their host’s population characteristics, such as host
population size (Scheidel et al., 2003). With respect to the PNG banana fly situation,
the germplasm diversity of banana (Musa spp.) in PNG is very large and multiple M. x
paradisiaca varieties are cultivated in banana growing areas at any one time, while
wild Musa species can be highly abundant in contiguous and remnant rainforest areas.
The low levels of banana crop utilisation observed for banana fly in PNG may simply
be the normal behaviour of a specialist herbivore which, in the presence of an abundant
food source, utilises only a small portion of potential larval food resources (Singer et
al., 1989; Debouzie et al., 2002). This behaviour may be exacerbated if the main

144
commercial varieties grown, such as Cavendish and Ladyfinger, are inherently non-
preferred or poor quality hosts, as seems to be the case based on laboratory results
(Chapter 5).
7.3.2 How does this relate to Market access?
The nature of developing international protocols (in any field) means that issues are
often highly simplified so to address as wide a range as possible of different
contingencies and stake-holder issues and sensitivities. With respect to phytosanitary
protocols, one of the most common simplifications is that given a pest (insect or
pathogen), and given a reported host crop, infestation is going to occur and needs to be
managed. While the logic for such a simplification is obvious, the science
underpinning the simplification is less obvious. Even for highly polyphagous pest
insects host preference rankings occur (Bravo et al., 2001; Clarke et al., 2005), a very
well documented trait among herbivorous insects (Fry, 1996; Carriere, 1998;
Steinbauer, 2002). Preferences are not static, but can change based on the physiological
status of the insect, the quality of the host plant(s) and a suite of other traits both
internal and external to the insect. Herbivore preferences are such that in the presence
of a preferred host, less preferred hosts are unlikely to be utilised (Fitt, 1986). In my
banana fly case, literature records that bananas are hosts for banana fly means that
export restrictions for getting banana out off PNG will be very stringent, despite
evidence from the field that banana usage by banana fly is very low. This is because
the published records make little or no acknowledgement of the fact that banana fly is
likely to infest different banana species and cultivars differently, even when in high
abundance. If international phytosanitary protocols are to be based on sound science,
as international agreements require, then greater emphasis must be placed on
understanding the biological link between herbivores and host use, so as to allow better
links to be made between pest abundance, crop infestation and phytosanitary risk.

145
References
ACNARS (2003). 7 Pests and Diseases. In Research and Development Issues Related to Banana
Production in PNG. URS Sustainable Development Project & the Australian Contribution to a
National Agricultural Research System in PNG (ACNARS), Adelaide.
Allen, B., Bourke, R.M., & Gibson, J. (2005). Poor rural places in Papua New Guinea. Asia Pacific
Viewpoint, 46, 201-217.
Allen, B.J., Bourke, R.M., & Hide, R.L. (1995). The sustainability of Papua New Guinea agricultural
systems: the conceptual background. Global Environmental Change, 5, 297-312.
Allwood, A.J. (1996a). Biology and Ecology: Prerequisites for Understanding and Managing Fruit Flies.
In Management of Fruit Flies in the Pacific (eds A.J. Allwood & R.A.I. Drew), pp. 95-101.
ACIAR Proceedings No 76, Canberra.
Allwood, A.J. (1996b). Control Strategies for Fruit Flies (Family Tephritidae) in the South Pacific. In
Management of Fruit Flies in the Pacific (eds A.J. Allwood & R.A.I. Drew), pp. 171-178.
ACIAR Proceedings No 76, Canberra.
Allwood, A.J., Chinajariyawong, A., Drew, R.A.I., Hamacek, E.L., Hancock, D.L., Hengsawad, C.,
Jipanin, J.C., Jirasurat, M., Kong Krong, C., Kritsaneepaiboon, S., Leong, C.T.S., &
Vijaysegaran, S. (1999). Host plant records for fruit flies (Diptera: Tephritidae) in Southeast
Asia. The Raffles Bulletin of Zoology, Supplement No 7, 1-92.
Allwood, A.J. & Leblanc, L. (1996). Losses Caused by Fruit Flies (Diptera: Tephritidae) in Seven
Pacific Island Countries. In Management of Fruit Flies in the Pacific (eds A.J. Allwood &
R.A.I. Drew), pp. 208-211. ACIAR Proceedings No 76, Canberra.
Aloalii, I., Masamdu, R., Theunis, W., & Thistleton, B. (1993). Prospects for biological control of taro
beetles, Papuana spp. (Coleoptera: Scarabaeidae), in the South Pacific. In Proceedings of the
Sustainable Taro Culture for the Pacific Conference (Sept 24-25, 1992) (ed L. Ferentinos), pp.
66-70. University of Hawaii, Honolulu, Hawaii.
Aluja, M., Diaz-Fleischer, F., Papaj, D.R., Lagunes, G., & Sivinski, J. (2001). Effects of age, diet,
female density, and the host resource on egg load in Anastrepha ludens and Anastrepha obliqua
(Diptera: Tephritidae). Insect Physiology, 47, 975-988.
Aluja, M. & Mangan, R.L. (2008). Fruit fly (Diptera: Tephritidae) host status determination: critical
conceptual, methodological, and regulatory considerations. Annual Review of Entomology, 53,
473-502.
Alvarez-Coque, J.M.G. & Bautista, R.M. (1994). Sources of EC horticultural import growth from
developing countries. Agricultural Economics, 10, 125-141.

146
Alyokhin, A.V., Messing, R.H., & Duan, J.J. (2001). Abundance and mating behaviour of oriental fruit
flies (Diptera: Tephritidae) near methyleugenol-baited traps. Pan-Pacific Entomologist, 77,
161-167.
Andersen, M.C., Adams, H., Hope, B., & Powell, M. (2004). Risk analysis for invasive species: General
framework and research needs. Risk Analysis, 24, 893-900.
Anonymous (1969). Insect pest survey for the year ending 30th June 1968. Papua New Guinea
Agricultural Journal, 22, 179-201.
Armstrong, J.W. (1983). Infestation biology of three fruit fly (Diptera: Tephritidae) species on Brazilian,
Valery, and William's cultivars of banana in Hawaii. Economic Entomology, 76, 539-543.
Armstrong, J.W. (1996). Quarantine Treatment Options for Fruit Fly Host Commodities for Pacific
Island Countries. In Management of Fruit Flies in the Pacific (eds A.J. Allwood & R.A.I.
Drew), pp. 222-224. ACIAR Proceedings No 76, Canberra.
Armstrong, J.W. (2001). Quarantine security of bananas at harvest maturity against Mediterranean and
Oriental fruit flies (Diptera: Tephritidae) in Hawaii. Economic Entomology, 94, 302-314.
Arnaud, E. & Horry, J.P. (1997). Musalogue: A Catalogue of Musa Germplasm. Papua New Guinea
Collecting Missions, 1988-1989 International Network for the Improvement of Banana and
Plantain, Montpellier, France.
Arura, M. & Akus, W.L. (1988). Physical Yield Loss Due to Taro Beetle. Papua New Guinea
Department of Agriculture and Livestock, Annual Research Report 1984-1988, 68.
Arura, M., Sands, M.C., & Sands, D.P.A. (1988). Banana Skipper Butterfly Survey. In Department of
Agriculture & Livestock Annual Research Report 1984-1988, pp. 67-68.
Arura, M., Sands, M.C., & Sands, D.P.A. (1989). Banana Skipper Butterfly Survey in Papua New
Guinea. In Department of Agriculture & Livestock - Bubia. Annual Research Report 1989, pp.
23-26.
Asafu-Adjaye, J. (1996). Revitalising growth in Papua New Guinea's agricultural sector. Agricultural
Systems, 51, 299-316.
Baker, R.H.A. (1996). Developing a European pest risk mapping system. EPPO Bulletin, 26, 485-494.
Balagawi, S., Vijaysegaran, S., Drew, R.A.I., & Raghu, S. (2005). Influence of fruit traits on oviposition
preference and offspring performance of Bactrocera tryoni (Froggatt) (Diptera: Tephritidae) on
three tomato (Lycopersicon lycopersicum) cultivars. Australian Journal of Entomology, 44, 97-
103.
Banana Colour chart (websites).
http://www.globalfruit.ie/index.php?option=com_content&task=view&id=14&Itemid=21,
http://www.catalyticgenerators.com/service.html,
http://lib.store.yahoo.net/lib/catalyticgenerators/bananacc.pdf,
http://www.bananaland.com.au/info/facts/banana_details_ripening_stages.php.

147
Bateman, M.A. (1972). The ecology of fruit flies. Annual Review of Entomology, 17, 493-518.
Bauer, M., Pearce, D., & Vincent, D. (2003). Saving A Staple Crop: Impact of Biological Control of the
Banana Skipper on Poverty Reduction in Papua New Guinea. ACIAR project CS2/1988/002-
C. In ACIAR Impact Assessment Series No. 22, pp. 23. ACIAR, Canberra.
Bautista, R.C., Harris, E.J., Vargas, R.I., & Jang, E.B. (2004). Parasitization of melon fly (Diptera:
Tephritidae) by Fopius arisanus and Psyttalia fletcheri (Hymenoptera: Braconidae) and the
effect of fruit substrates on host preference by parasitoids. Biological Control, 30, 156-164.
Beaudoin-Ollivier, L., Prior, R.N.B., P, M.J., Nanguai, R., & Kakul, T. (2001). The larval development
sites of the Melanesian coconut beetle Scapanes australis Boisduval (Coleoptera: Scarabaeidae:
Dynastinae) in Papua New Guinea. Science in New Guinea, 26, 52-60.
Beaudoin-Olliviera, L., Prior, R.N.B., & Laup, S. (2000). Simplified field key to identify larvae of some
rhinoceros beetles and associated Scarabs (Coleoptera: Scarabaeoidea) in Papua New Guinea
coconut developments. Annals of the Entomological Society of America, 93, 90-95.
Bedford, G.O. (1976). Observations on the biology and ecology of Oryctes rhinoceros and Scapanes
australis (Coleoptera: Scarabaeidae: Dynastinae) pests of coconut palms in Melanesia. Journal
of Austustralian Entomology Society, 15, 241-251.
Bedford, G.O. (1980). Biology, ecology, and control of palm rhinoceros beetles. Annual Review of
Entomology, 25, 309-339.
Bedford, G.O. (1986). Biological control of the rhinoceros beetle (Oryctes rhinoceros) in the South
Paciic by Baculovirus. Agriculture, Ecosystems & Environment, 15, 141-147.
Bellamy, J.A. & McAlpine, J.R. (1995). Papua New Guinea Inventory of Natural Resources, Population
Distribution and Land Use Handbook, Natural Resources Series No 6, 2nd edn.
Commonwealth Scientific and Industrial Research Organisation, Australian Agency for
International Development, Canberra.
Benjamin, A.K., Mopafi, I., & Duke, T. (2001). A Perspective on Food and Nutrition in the PNG
Highlands. In Food Security For Papua New Guinea (eds R.M. Bourke, B.J. Allen & J.G.
Salisbury), Vol. 99, pp. 94-99. ACIAR Proceedings, Lae.
Birla, S.L., Wang, S., Tang, J., Fellman, J.K., Mattinson, D.S., & Lurie, S. (2005). Quality of oranges as
influenced by potential radio frequency heat treatments against Mediterranean fruit flies.
Postharvest Biology and Technology, 38, 66-79.
Blackburn, T.M. & Duncant, R.P. (2001). Determinants of establishment success in introduced birds.
Nature, 414, 195-197.
Bonizzoni, M., Guglielmino, C.R., Smallridge, C.J., Gomulski, M., Malacrida, A.R., & Gasperi, G.
(2004). On the origins of Medfly invasion and expansion in Australia. Molecular Ecology, 13,
3845-3855.

148
Bourke, R.M. (2001). Intensification of Agricultural Systems in Papua New Guinea. Asia Pacific
Viewpoint, 42, 219-235.
Bourke, R.M., Allen, B.J., Hobsbawn, P., & Conway, J. (1998). Papua New Guinea: Text Summaries
Department of Human Geography, Research School of Pacific and Asian Studies. Australian
National University, Canberra.
Bourke, R.M., Fenner, T.L., Sticbick, J.N.L., Baker, G.L., Hassan, E., O'Sullivan, D.F., & Li, C.S.
(1973). Insect pest survey for the year ending 30th June 1969 Department of Agriculture &
Fisheries, Port Moresby.
Bourke, R.M. & Vlassak, V. (2004). Estimates of Food Crop Production in Papua New Guinea Land
Management Group, The Australian National University, Canberra.
Bower, C.C. (1977). Inhibition of larval growth of the Queensland fruit fly, Dacus tryoni (Diptera:
Tephritidae) in apples. Annals of the Entomological Society of America, 70, 97-100.
Bravo, I.S.J., Fernandes, O.A., deBortoli, S.A., & Zucoloto, F.S. (2001). Oviposition behavior of
Ceratitis capitata Wiedemann (Diptera, Tephritidae): Association between oviposition
preference and larval performance in individual females. Neotropical Entomology, 30, 559-564.
Brevault, T. & Quilici, S. (2000). Diel patterns of reproductive activities in the tomato fruit fly,
Neoceratitis cyanescens. Physiological Entomology, 25, 233-241.
Brevault, T. & Quilici, S. (2007). Visual response of the tomato fruit fly, Neoceratitis cyanescens, to
colored fruit models. Entomologia Experimentalis et Applicata, 125, 45-54.
Bridge, J. & Page, S.L.J. (1982). Plant Nematodes of Papua New Guinea: Their Importance as Crop
Pests. Report of a Plant Nematode Survey in Papua New Guinea. CAB Commonwealth
Institute of Parasitology, St. Albans, UK.
Brieze-Stegeman, R., Rice, M.J., & Hooper, G.H.S. (1978). Daily periodicity in attraction of male
tephritid fruit flies to synthetic chemical lures. Australian Journal of Entomology, 17, 341-346.
Broumas, T., Haniotakis, G., Liaropoulos, C., Tomazou, T., & Ragoussis, N. (2002). The efficacy of an
improved form of the mass-trapping method, for the control of the olive fruit fly, Bactrocera
oleae (Gmelin) (Dipt., Tephritidae): pilot-scale feasibility studies. Journal of Applied
Entomology, 126, 217-223.
Bruce, T.J.A., Wadhams, L.J., & Woodcock, C.M. (2005). Insect host location: A volatile situation.
Trends in Plant Science, 10, 269-274.
Burrack, H.J., Connell, J.H., & Zalom, F.G. (2008). Comparison of olive fruit fly (Bactrocera oleae
(Gmelin)) (Diptera: Tephritidae) captures in several commercial traps in California.
International Journal of Pest Management, 54, 227 - 234.
CABI (1998). Crop Protection Compendium. Global Module 2nd Edition CAB International,
Wallingford, OXON, OX10 8DE, UK.

149
Carey, J.R. (1996). The future of the Mediterranean fruit fly Ceratitis capitata invasion of California: A
predictive framework. Biological Conservation, 78, 35-50.
Carey, J.R., Liedo, P., Harshman, L., Zhang, Y., Muller, H., Partridge, L., & Wang, J. (2002). Life
history response of Mediterranean fruit flies to dietary restriction. Aging Cell, 1, 140-148.
Carlton, J.T. (1996). Pattern, process, and prediction in marine invasion ecology. Biological
Conservation, 78, 97-106.
Carmichael, A.E., Harding, R., Jackson, G., Kumar, S., Lal, S.N., Masamdu, R., Wright, J., & Clarke,
A.R. (2007). TaroPest: A guide to pests and diseases of taro in the South Pacific. Field Manual
& CD-Rom Queensland University of Technology, Brisbane.
Carriere, Y. (1998). Constraints on the evolution of host choice by phytophagous insects. Oikos, 82,
401-406.
Catley, A. (1962). Tiracola plagiata Walk. (Lepidoptera, Noctuidae) A serious pest of cacaocin Papua
New Guinea. Papua New Guinea Agricultural Journal, 15, 105-109.
Christenson, L.D. & Foote, R.H. (1960). Biology of fruit flies. Annual Review of Entomology, 5, 171-
192.
Christian, J.M. & Wilson, S.D. (1999). Long-term ecosystem impacts of an introduced grass in the
Northern Great Plains. Ecology, 80, 2397-2407.
Chuah, C.H., Yong, H.S., & Goh, S.H. (1997). Methyl eugenol, a fruit-fly attractant, from the browning
leaves of Proiphys amboinensis (Amaryllidaceae). Biochemical Systematics and Ecology, 25,
391-393.
Clarke, A.R., Armstrong, K.F., Carmichael, A.E., Milne, J.R., Raghu, S., Roderick, G.K., & Yeates,
D.K. (2005). Invasive phytophagous pests arising through a recent tropical evolutionary
radiation: The Bactrocera dorsalis complex of fruit flies. Annual Review of Entomology, 50,
293-319.
Clarke, A.R., Balagawi, S., Clifford, B., Drew, R.A.I., Leblanc, L., Mararuai, A., McGuire, D., Putulan,
D., Romig, T., Sar, S., & Tenakanai, D. (2004). Distribution and biogeography of Bactrocera
and Dacus species (Diptera: Tephritidae) in Papua New Guinea. Australian Journal of
Entomology, 43, 148-156.
Coelli, T. & Fleming, E. (2004). Diversification economies and specialisation efficiencies in a mixed
food and coffee smallholder farming system in Papua New Guinea. Agricultural Economics,
31, 229-239.
Cook, D.C. (2008). Benefit cost analysis of an import access request. Food Policy, 33, 277-285.
Cook, D.C. & Fraser, R.W. (2008). Trade and invasive species risk mitigation: Reconciling WTO
compliance with maximising the gains from trade. Food Policy, 33, 176-184.
Cowley, J.M. (1990). A new system of fruit fly surveillance trapping in New Zealand. New Zealand
Entomologist, 13, 81-84.

150
CSIRO (2004). Cosmopolites sordidus (Germar). http://www.ento.csiro.au/aicn/name_s/b_1159.htm.
Dadzie, B.K. & Orchard, J.E. (1997). Routine Post Harvest Screening of Banana/Plantain Hybrids:
Criteria and Methods. In INIBAP Technical Guidelines 2, pp. 80. International Plant Genetic
Resources Institute, Rome, Italy.
Dalby-Ball, G. & Meats, A. (2000a). Effects of fruit abundance within a tree canopy on the behaviour of
wild and cultured Queensland fruit flies, Bactrocera tryoni (Froggatt) (Diptera: Tephritidae).
Australian Journal of Entomology, 39, 201-207.
Dalby-Ball, G. & Meats, A. (2000b). Influence of the odour of fruit, yeast and cue-lure on the flight
activity of the Queensland fruit fly, Bactrocera tryoni (Froggatt) (Diptera: Tephritidae).
Australian Journal of Entomology, 39, 195-200.
Davis, R. (2004). Fusarium Wilt (Panama Disease) of Banana. In Pest Advisory Leaflet No.42, Vol.
Reprinted March 2005. Secretariat of the Pacific Community, Suva, Fiji.
Davis, R.I., Geering, A.D.W., Thomas, J.E., Gunua, T.G., & Rahamma, S. (2000). Disease notes or new
records: First records of banana streak virus on the island of New Guinea. Australaslan Plant
Pathology, 29, 281.
Debouzie, D., Desouhant, E., Oberli, F., & Menu, F. (2002). Resource limitation in natural populations
of phytophagous insects. A long-term study case with the chestnut weevil. Acta Oecologica, 23,
31-39.
Dhillon, M.K., Singh, R., Naresh, J.S., & Sharma, H.C. (2005). The melon fruit fly, Bactrocera
cucurbitae: A review of its biology and management. Journal of Insect Science, 5.
Dori, F. (1988). Biology and Field Population of the Banana Skipper, Erionota thrax. In Department of
Agriculture & Livestock Annual Research Report 1984-1988, pp. 50-51.
Dori, F., Tenakanai, D., & Kurika, K. (1993). The current status of fruit flies (Tephritidae) in Papua
New Guinea. Pan-Pacific Journal of Agriculture Extension, 15, 22-25.
Drew, R.A.I. (1989). The tropical fruit flies (Diptera: Tephritidae: Dacinae) of the Australasian and
Oceanian regions. Memoirs of the Queensland Museum, 26, 1-521.
Drew, R.A.I. (1996). The Economic and Social Impact of the Bactrocera papayae Drew and Hancock
(Asian Papaya fruit fly) Outbreak in Australia. In Management of Fruit Flies in the Pacific (eds
A.J. Allwood & R.A.I. Drew), pp. 205-207. ACIAR Proceedings No 76, Canberra.
Drew, R.A.I., Courtice, A.C., & Teakle, D.S. (1983). Bacteria as a natural source of food for adult fruit
flies (Diptera: Tephritidae). Oecologia, 60, 279-284.
Drew, R.A.I. & Hancock, D.L. (1994). The Bactrocera dorsalis complex of fruit flies (Diptera:
Tephritidae: Dacinae) in Asia. In Bulletin of Entomology Research Supplement, 2, 1-68.
Drew, R.A.I., Raghu, S., & Halcoop, P. (2008). Bridging the morphological and biological species
concepts: studies on the Bactrocera dorsalis (Hendel) complex (Diptera : Tephritidae :
Dacinae) in South-east Asia. Biological Journal of the Linnean Society, 93, 217-226.

151
Drew, R.A.I. & Romig, M. (1996). Overview - Tephritidae in the Pacific and Southeast Asia. In
Management of Fruit Flies in the Pacific (eds A. Allwood & R.A.I. Drew), pp. 46-53. ACIAR
Proceedings No 76, Canberra.
Drew, R.A.I., Zalucki, M.P., & Hooper, G.H.S. (1984). Ecological studies of eastern Australian fruit
flies (Diptera: Tephritidae) in their endemic habitat. I. Temporal variation in abundance.
Oecologia, 64, 267-272.
Duan, J.J. & Messing, R.H. (1997). Effect of two Opiine parasitoids (Hymenoptera: Braconidae)
introduced for fruit fly control on a native Hawaiian Tephritid, Trupanea dubautiae (Diptera:
Tephritidae). Biological Control, 8, 177-184.
Duncan, R. & Lutz, E. (1983). Penetration of industrial country markets by agricultural products from
developing countries. World Development, 11, 771-786.
Duncan, R. & Temu, I. (1995). Papua New Guinea: longer term developments and recent economic
problems. Asian-Pacific Economic Literature, 9, 36-54.
Duyck, P. F., David, P., & Quilici, S. (2004). A review of relationships between interspecific
competition and invasions in fruit flies (Diptera: Tephritidae). Ecological Entomology, 29, 511-
520.
Ebenhard, T. (1989). Bank vole Clethrionomys glareolus (Schreber, 1780) propagules of different sizes
and island colonization. Journal of Biogeography, 16, 173-180.
Eisemann, C.H. & Rice, M.J. (1989). Behavioural evidence for hygro - and mechanoreception by
ovipositor sensilla of Dacus tryoni (Diptera: Tephritidae). Physiological Entomology, 14, 273-
277.
EPPO/CABI (1997). Data Sheets on Quarantine Pests: Erwinia chrysanthemi. In Quarantine Pests for
Europe 2nd edition (eds I.M. Smith, D.G. McNamara, P.R. Scott & M. Holderness), pp. 1425.
CABI International, Wallingford, UK.
Ero, M.M. (2009). Host searching behaviour of Diachasmimorpha krausii (Fullaway) (Hymenoptera:
Braconidae: Opiinae), a polyphagous parasitoid of Dacinae fruit flies (Diptera: Tephritidae).
Unpublished PhD thesis, Queensland University of Technology, Brisbane.
Everett, R.A. (2000). Patterns and pathways of biological invasions. Trends in Ecology & Evolution, 15,
177-178.
FAO (2005). Regional Standards for Phytosanitary Measures; Guidelines for the Confirmation of Non-
host Status of Fruit and Vegetables to Tephritid Fruit Flies Report of the Twenty-Fourth
Session of the Asia and Pacific Plant Protection Commission. Asia and Pacific Plant Protection
Commission, Bangkok.
Fitt, G.P. (1986). The roles of adult and larval specialisations in limiting the occurrence of five species
of Dacus (Diptera: Tephritidae) in cultivated fruits. Oecologia, 69, 101-109.

152
Flath, R.A., Light, D.M., Jang, E.B., Mon, T.R., & John, J.O. (1990). Headspace examination of volatile
emissions from ripening papaya (Carica papaya L., Solo Variety). Journal of Agriculture Food
Chemistry, 38, 1060-1063.
Fletcher, B.S. (1974). The ecology of a natural population of the Queensland fruit fly, Dacus tryoni. V.
The dispersal of adults. Australian Journal of Zoology, 22, 189-202.
Fletcher, B.S. (1987). The biology of Dacine fruit flies. Annual Review of Entomology, 32, 115-144.
Fletcher, B.S. (1998). Dacine fruit flies collected during the dry season in the lowland rainforest of
Madang Province, Papua New Guinea (Diptera: Tephritidae). Australian Journal of
Entomology, 37, 315-318.
Fletcher, B.S., Bateman, M.A., Hart, N.K., & Lamberton, J.A. (1975). Identification of a fruit fly
attractant in an Australian plant, Zieria smithii, as O-Methyl eugenol. Economic Entomology,
68, 815-816.
Follett, P.A. (2004). Irradiation to control insects in fruits and vegetables for export from Hawaii.
Radiation Physics and Chemistry, 71, 163-166.
Follett, P.A. & Hennessey, M.K. (2007). Confidence limits and sample size for determining nonhost
status of fruits and vegetables to Tephritid fruit flies as a quarantine measure. Economic
Entomology, 100, 251-257.
Follett, P.A. & Neven, L.G. (2006). Current trends in quarantine entomology. Annual Review of
Entomology, 51, 359-385.
Fooks, L.D. (1989). Commercial Production of Cavendish Bananas for Papua New Guinea Markets.
Department of Agriculture and Livestock Technical Bulletin, revised Nov 1989, 23pp.
Fooks, L.D. (2002). Commercial Production of Giant Cavendish Bananas in the Lowlands of Papua
New Guinea Rural Resources P/L, Lae.
French, B.R. (1986). Food plants of Papua New Guinea: A compendium Australian Pacific Science
Foundation.
French, B.R. (2006). Insect Pests of Food Plants of Papua New Guinea: A compendium
http://foodplantsinternational.com/resources/3Insects%20on%20food%20plants%20in%20PNG.pdf.
Froggatt, J.L. (1941). Entomological notes. The New Guinea Agriculture Gazette, 7, 298-300.
Fry, J.D. (1996). The evolution of host specialization - are trade-offs overrated. American Naturalist,
148, S 84-S 107.
Gende, P., Kakul, T., Laup, S., & Embupa, S. (2006). Breeding Sites of Major Coconut Beetle Pest
Scapanes australis Boisd. (Coleoptera: Scarabaeidea, Dynastinae) in East New Britain, Papua
New Guinea. In Pest and Disease Incursions: Risks, Threats and Management in Papua New
Guinea. Papers presented at the 2nd Papua New Guinea Plant Protection Conference, Kokopo,
East New Britain Province, 8-10 November 2004 (ed T.V. Price), pp. 57-59. Canberra, ACIAR
Technical Report No, 62.

153
Gibson, J. (2001a). The Economic and Nutritional Importance of Household Food Production. In Food
Security For Papua New Guinea (eds R.M. Bourke, M.G. Allen & J.G. Salisbury), pp. 37-44.
ACIAR Proceedings No 99, Lae.
Gibson, J. (2001b). Food Demand in the Rural and Urban Sectors of PNG. In Food Security For Papua
New Guinea (eds R.M. Bourke, M.G. Allen & J.G. Salisbury), pp. 45-53. ACIAR Proceedings
No 99, Lae.
Gold, C.S., Pinese, B., & Peña, J.E. (2002). Pests of Banana. In Tropical Fruit Pests and Pollinators;
Biology, Economic Importance, Natural Enemies and Control (eds J.E. Peña, J.L. Sharp & M.
Wysoki), pp. 13-56. CABI Publishing.
Gray, G.M., Allen, J.C., Burmaster, D.E., Gage, S.H., Hammitt, J.K., Kaplan, S., Keeney, R.L., Morse,
J.G., North, D.W., Nyrop, J.P., Stahevitch, A., & Williamslau, R. (1998). Principles for conduct
of pest risk analyses: report of an expert workshop. Risk Analysis, 18, 773-780.
Greve, J.E.S. & Ismay, J.W. (1983). Crop insect survey of Papua New Guinea, from July 1st 1969 to
Dec 31st 1978. Papua New Guinea Agricultural Journal, 32, 1-120.
Gunther, M., Wiles, G.C., & Laraki, J. (2003). Banana. In Proceedings of the Highlands Horticulture
Workshop (eds M. Gunther, G.C. Wiles & J. Laraki), Vol. 3, pp. 23-24. National Agricultural
Research Institute, Mt Hagen, Western Highlands. September 8-9, 1999.
Gwaiseuk, W.R. (2001). The Role of Agriculture in the PNG Economy. In Food Security For Papua
New Guinea (eds R.M. Bourke, B.J. Allen & J.G. Salisbury), pp. 30-36. Proceedings of the
Papua New Guinea Food And Nutrition 2000 Conference, PNG University of Technology, Lae,
26-30 June 2000. ACIAR Proceedings No 99.
Hall, R.J. & Hastings, A. (2007). Minimizing invader impacts: Striking the right balance between
removal and restoration. Journal of Theoretical Biology, 249, 437-444.
Hancock, D.L., Hamacek, E., Lloyd, A.C., & Elson-Harris, M.M. (2000). The distribution and host
plants of fruit flies (Diptera: Tephritidae) in Australia. Queensland Department of Primary
Industries - Information Series QI99067, 75pp.
Hanson, L.W., Allen, B.J., Bourke, R.M., & McCarthy, T.J. (2001). Papua New Guinea Rural
Development Handbook The Australian National University, Canberra.
Hardy, D.E. (1991). Contribution of Taxonomic Studies to the Integrated Pest Management of Fruit
Flies, with Special Emphasis on the Asia-Pacific Region. In Proceedings of the First
International Symposium of Fruit Flies in the Tropics (eds S. Vijaysegaran & A.G. Ibrahim),
pp. 430, Kuala Lumpur.
Hartemink, A.E. & Bourke, R.M. (2001). Nutrient Deficiencies in Export Tree and Food Crops:
Literature Review and Field Observations. In Food Security for Papua New Guinea (eds R.M.
Bourke, M.G. Allen & J.G. Salisbury), pp. 316-327. ACIAR Proceedings No 99, Canberra.

154
Hayden, H.L. & Carlier, J. (2003). Genetic structure of Mycosphaerella fijiensis populations from
Australia, Papua New Guinea and the Pacific islands. Plant Pathology, 52, 703-712.
Headrick, D.H. & Goeden, R.D. (1996). Issues concerning the eradication or establishment and
biological control of the Mediterranean fruit fly, Ceratitis capitata (Wiedemann) (Diptera:
Tephritidae), in California. Biological Control, 6, 412-421.
Hennessey, M.K., Knight Jr, R.J., & Schnell, R.J. (1995a). Antibiosis to Caribbean fruit fly in avocado
germplasm. HortScience, 30, 5, 1061-1062.
Hennessey, M.K., Knight, R.J., Jr, & Schnell, R.J. (1995b). Antibiosis to Caribbean fruit fly (Diptera:
Tephritidae) immature stages in Carambola germplasm. The Florida Entomologist, 78, 354-
357.
Hennessy, D.A. (2008). Biosecurity incentives, network effects, and entry of a rapidly spreading pest.
Ecological Economics, 68, 230-239.
Higgins, S.I., Richardson, D.M., Cowling, R.M., & Trinder-Smith, T.H. (1999). Predicting the
Landscape-Scale Distribution of Alien Plants and Their Threat to Plant Diversity. Conservation
Biology, 13, 303-313.
Hill, A.R. (1986). Reduction in trap captures of female fruit flies (Diptera: Tephritidae) when synthetic
male lures are added. Australian Journal of Entomology, 25, 211-214.
Hochberg, M.E. & Gotelli, N.J. (2005). An Invasions Special Issue. Trends in Ecology & Evolution, 20,
211.
Hofman, P.J., Stubbings, B.A., Adkins, M.F., Corcoran, R.J., White, A., & Woolf, A.B. (2003). Low
temperature conditioning before cold disinfestation improves `Hass' avocado fruit quality.
Postharvest Biology and Technology, 28, 123-133.
Holliday, P. (1980). Veronaea. In Fungus Diseases of Tropical Crops, pp. 538-539.
Hollingsworth, R.G., Drew, R.A., Allwood, A.J., Romig, M., Vagalo, M., & Tsatsia, F. (2003). Host
plants and relative abundance of fruit fly (Diptera: Tephritidae) species in the Solomon Islands.
Australian Journal of Entomology, 42, 95-108.
Hooper, G.H.S. (1978). Effect of combining methyl eugenol and cuelure on the capture of male
Tephritid fruit flies. Australian Journal of Entomology, 17, 189-190.
Houlder, D., Hutchinson, M., Nix, H., & McMahon, J. (2001). ANUCLIM. Centre for Resource and
Environmental Studies, Canberra.
IPPC (1995). Requirements for the Establishment of Pest Free Areas. International Standards For
Phytosanitary Measures (ISPM) No. 4, 8p.
IPPC (1997). Guidelines for Surveillance. International Standards For Phytosanitary Measures (ISPM)
No. 6, 8p.

155
IPPC (1998). Determination of Pest Status in an Area. International Standards For Phytosanitary
Measures (ISPM) No. 08, 12p.
IPPC (1999). Requirements for the Establishment of Pest Free Places of Production and Pest Free
Production Sites. International Standards For Phytosanitary Measures (ISPM) No. 10, 10p.
IPPC (2002a). Pest Reporting. International Standards For Phytosanitary Measures (ISPM) No. 17, 10p.
IPPC (2002b). Regulated Non-Quarantine Pests: Concept and Application. International Standards For
Phytosanitary Measures (ISPM) No. 16, 10p.
IPPC (2002c). The use of Integrated Measures in a Systems Approach for Pest Risk Management.
International Standards For Phytosanitary Measures (ISPM) No. 14, 12p.
IPPC (2004a). Pest Risk Analysis for Quarantine Pests, including Analysis of Environmental Risks and
Living Modified Organisms. International Standards For Phytosanitary Measures (ISPM) No.
11, 38p.
IPPC (2004b). Pest Risk Analysis for Regulated Non-quarantine Pests. International Standards For
Phytosanitary Measures (ISPM) No. 21, 16p.
IPPC (2005). Guidelines for the Export, Shipment, Import and Release of Biological Agents and other
Beneficial Organisms. International Standards For Phytosanitary Measures (ISPM) No. 3, 12p.
IPPC (2006a). International Standards for Phytosanitary Measures - 1 to 27. International Standards For
Phytosanitary Measures (ISPM) 2006 edition, 353p.
IPPC (2006b). Phytosanitary Principles for the Protection of Plants and the Application of Phytosanitary
Measures in International Trade. International Standards For Phytosanitary Measures (ISPM)
No. 1, 10p.
IPPC (2007). Framework for Pest Risk Analysis. International Standards For Phytosanitary Measures
(ISPM) No. 2, 15p.
IPPC (2008). International Standards for Phytosanitary Measures - 1 to 31. International Standards For
Phytosanitary Measures (ISPM) 2008 edition, 420p.
Ismay, J.W. & Dori, F. (1985). Crop insect survey of Papua New Guinea from 1st January 1979 to
December 1981. Research Bulletin, 38, 65.
Issacson, B. & King, G. (1987). Report of a rapid rural appraisal of farming systems in the Bubia area,
Morobe Province. 9-19, August 1987, section I & II.
Jacobi, K.K., Wong, L.S., & Giles, J.E. (1993). Lychee (Litchi chinensis Sonn.) fruit quality following
vapour heat treatment and cool storage. Postharvest Biology and Technology, 3, 111-119.
Jang, E.B. & Light, D.M. (1991). Behavioral responses of female Oriental fruit flies to the odor of
papayas at three ripeness stages in a laboratory flight tunnel (Diptera: Tephritidae). Journal of
Insect Behavior, 4, 751-762.

156
Jang, E.B. & Light, D.M. (1996). Olfactory semiochemical of Tephritids. In Fruit Fly Pests: A World
Assessment Of Their Biology And Management (eds B.A. McPheron & G.J. Steck), pp. 73-90.
St. Lucie Press, Delvery Beach, Florida.
Kakul, T., Laup, S., Ollivier, L., Morin, J.P., & Rochat, D. (2000). Control of Scapanes australis a major
pest of coconut using semiochemicals. In XXI International Congress of Entomology (ed D.L.
Gazzoni), Vol. 21, pp. 172. International Congress of Entomology., Londrina : EMBRAPA
N.682. 21, 2000-08-20/2000-08-26, Foz do Iguaçu, Brésil.
Kambuou, R. (2001). Review of Germplasm Collections and Agronomic Research on Bananas in Papua
New Guinea. In Food Security For Papua New Guinea (eds R.M. Bourke, M.G. Allen & J.G.
Salisbury), pp. 809-815. ACIAR Proceedings No 99, Canberra.
Kambuou, R. (2003). Current situation in banana R&D in Papua New Guinea. In Proceedings of the 2nd
BAPNET Steering Committee Meeting. BAPNET, Jakarta, 6-19 Oct 2003.
Kambuou, R. (2005). Highlights of banana R&D in Papua New Guinea. In Advancing banana and
plantain R&D in Asia and the Pacific - Vol. 13 (eds A.B. Molina, L.B. Xu, V.N. Roa, I. Van
den Bergh & K.H. Borromeo), Vol. 13, pp. 103-108. Proceedings of the 3rd BAPNET Steering
Committee. International Network for the Improvement of Banana and Plantain, Guangzhou,
China. 23-26 November 2004.
Kambuou, R.N. (2004). Current Situation in Banana R&D in Papua New Guinea. In Advancing Banana
and Plantain R&D in Asia and the Pacific - Vol 12 (eds A.B. Molina, V.N. Roa, I. Van den
Bergh, M.A.G. Maghuyop & K. Borromeo), Vol. 12, pp. 95-100. Proceedings of the 2nd
BAPNET Steering Committee Meeting. International Network for the Improvement of Banana
and Plantain - Asia Pacific, Jakarta, 6-11 October 2003.
Kannapiran, C.A. (2000). Commodity price stabilisation: macroeconomic impacts and policy options.
Agricultural Economics, 23, 17-30.
Khan, M.A., Ashfaq, M., Akram, W., & Lee, J. (2005). Management of fruit flies (Diptera: Tephritidae)
of the most perishable fruits. Entomological Research, 35, 79-84.
King, G.A., Banag, J., Kambuou, R., Ovia, K., Ovia, A., Heywood, P.F., & Hide, R.L. (1988). Banana
production in Amele, Madang Province and the Vanapa River and Kabadi, Central Province.
Papua New Guinea Department of Agriculture and Livestock, Annual Research Report 1984-
1988, 18-19.
Kingwell, R., Godden, D., Kambuou, R., & Jackson, G. (2001). Managing and funding germplasm
preservation in Papua New Guinea - for improved indigenous foods. Food Policy, 26, 265-280.
Kitching, R.L., Bickel, D., Creagh, A.C., Hurley, K., & Symonds, C. (2004). The biodiversity of Diptera
in Old World rain forest surveys: A comparative faunistic analysis. Biogeography, 31, 1185-
1200.
Kohler, F., Pellegrin, F., Jackson, G., & McKenzie, E. (1997). Diseases of Cultivated Crops in Pacific
Island Countries South Pacific Commision, Noumea, New Caledonia.

157
Kokoa, P. (2006). Impact of Some Food-crop Disease Outbreaks in Papua New Guinea. In Pest And
Disease Incursions: Risks, Threats And Management In Papua New Guinea (ed T.V. Price), pp.
137-143. ACIAR Technical Report No.62, Canberra.
Kolar, C.S. & Lodge, D.M. (2001). Progress in invasion biology: predicting invaders. Trends in Ecology
& Evolution, 16, 199-204.
Kovaleski, A., Sugayama, R.L., & Malavasi, A. (1999). Movement of Anastrepha fraterculus from
native breeding sites into apple orchards in Southern Brazil. Entomologia Experimentalis et
Applicata, 91, 459-465.
Kumar, R. (2001). Insect Pests of Agriculture in Papua New Guinea. Part 1. Principles and Practice.
Pests of Tree Crops and Stored Products Science in New Guinea.
Lamb, K.P. & Johnston, A.D. (1976). Some insect pests of Papua New Guinea Department of Biology,
University of Papua New Guinea, Port Moresby.
Leblanc, L., Balagawi, S., Mararuai, A., Putulan, D., Tenakanai, D., & Clarke, A.R. (2001). Fruit flies in
Papua New Guinea. In Pest Advisory Leaflet No. 37, pp. 12. Secretariat of the Pacific
Community - Plant Protection Service, Suva.
Levine, J.M. (2008). Biological invasions. Current Biology, 18, 57-60.
Leweniqila, L., Heimoana, V., Purea, M., Munro, L., Allwood, A., Ralulu, L., & Vueti, E.T. (1996).
Seasonal abundance of Bactrocera facialis (Coquilletta), B. passiflorae (Froggatt), B.
xanthodes (Braun) and B. melanotus (Coquillett) in orchard and forest habitats. In Management
of Fruit Flies in the Pacific (eds A.J. Allwood & R.A.I. Drew), pp. 121-124. ACIAR
Proceedings No 76, Canberra.
Lockwood, J.L., Cassey, P., & Blackburn, T. (2005). The role of propagule pressure in explaining
species invasions. Trends in Ecology & Evolution, 20, 223-228.
Lubulwa, G. & McMeniman, S. (1998). ACIAR-supported biological control projects in the South
Pacific (1983-1996): An economic assessment. Biocontrol News and Information, 19, 91N-
98N.
Lucas, P.W., Turner, I.M., Dominy, N.J., & Yamashita, N. (2000). Mechanical Defences to Herbivory.
Annals of Botany, 86, 913-920.
Lugard, M. & Smart, M. (2006). The role of science in international trade law. Regulatory Toxicology
and Pharmacology, 44, 69-74.
MacFarlane, B. (1987). Papuana Beetles. In Pest Advisory Leaflet No.21, pp. 4. Plant Protection Service,
South Pacific Commission, Suva, Fiji.
Macfarlane, D. (2000). Climate and Agro-ecological Zones. In Papua New Guinea Country Pasture/
Forage Resource Profiles (eds J.M. Suttie & S.G. Reynolds), pp. 16. Report for the Food and
Agriculture Organization of the United Nations.

158
Maelzer, D.A., Bailey, P.T., & Perepelicia, N. (2004). Factors supporting the non-persistence of fruit fly
populations in South Australia. Australian Journal of Experimental Agriculture, 44, 109-126.
Manning, M. (2001). Food Security in PNG. In Food Security For Papua New Guinea (eds R.M.
Bourke, B.J. Allen & J.G. Salisbury), pp. 15-22. Proceedings of the Papua New Guinea Food
And Nutrition 2000 Conference, PNG University of Technology, Lae, 26-30 June 2000.
ACIAR Proceedings No 99.
Mararuai, A., Allwood, A.J., Balagawi, S., Dori, F., Kalamen, M., Leblanc, L., Putulan, D., Sar, S.,
Schuhbeck, A., Tenakanai, D., & Clarke, A.R. (2001). Introduction and distribution of
Bactrocera musae (Tryon) (Diptera: Tephritidae) in East New Britain, Papua New Guinea.
Papua New Guinea Agricultural Journal, 45, 59-65.
Margolis, M., Shogren, J.F., & Fischer, C. (2005). How trade politics affect invasive species control.
Ecological Economics, 52, 305-313.
Markelova, H., Meinzen-Dick, R., Hellin, J., & Dohrn, S. (2009). Collective action for smallholder
market access. Food Policy, 34, 1-7.
Masamdu, R. (2006). The PNG Pest List Database and its Uses in Quarantine Surveillance and Pest
Management. In Pest And Disease Incursions: Risks, Threats And Management In Papua New
Guinea (ed T.V. Price), pp. 144-145. ACIAR Technical Reports No.62, Canberra.
Masamdu, R., Bokosou, J., & Niangu, B. (1988). Rhyparid beetle. Papua New Guinea Department of
Agriculture and Livestock, Annual Research Report 1984-1988, 49-50.
Masamdu, R., Bokosou, J., & Niangu, B. (1989). Banana Rhyparid beetle. In Lowlands Farming
Systems Annual Report 1989, pp. 22-23. Department of Agriculture & Livestock, Bubia.
Masamdu, R. & Simbiken, N. (2001). Effect of Taro Beetles on Taro Production in PNG. In Food
Security For Papua New Guinea (eds R.M. Bourke, M.G. Allen & J.G. Salisbury), pp. 752-
757. ACIAR Proceedings No 99, Canberra.
May, A.W.S. (1953). Queensland host records for the Dacinae (Family: Trypetidae). Queensland
Journal of Agricultural Science, 10, 36-79.
Maynard, G.V., Hamilton, J.G., & Grimshaw, J.F. (2004). Quarantine - Phytosanitary, sanitary and
incursion management: An Australian entomological perspective. Australian Journal of
Entomology, 43, 318-328.
McAusland, C. & Costello, C. (2004). Avoiding invasives: Trade-related policies for controlling
unintentional exotic species introductions. Journal of Environmental Economics and
Management, 48, 954-977.
McGregor, A.M. (1996). An Economic Evaluation of Fruit Fly Research in the South Pacific. In
Management of Fruit Flies in the Pacific (eds A.J. Allwood & R.A.I. Drew), pp. 212-221.
ACIAR Proceedings No 76, Canberra.

159
McGregor, G.R. (1989). An assessment of the annual variability of rainfall: Port Moresby, Papua New
Guinea. Singapore Journal of Tropical Geography, 10, 43-54.
McLeod, R. (2005). Management of fruit flies in the Pacific. ACIAR, Impact Assessment Series, Report
No. 37, 56.
Meats, A. & Clift, A.D. (2005). Zero catch criteria for declaring eradication of tephritid fruit flies: the
probabilities. Australian Journal of Experimental Agriculture, 45, 1335-1340.
Meats, A. & Osborne, A. (2000). Dose-related upwind anemotaxis and movement up odour gradients in
still air in the presence of methyl eugenol by the wild tobacco fly, Bactrocera cacuminata.
Physiological Entomology, 25, 41-47.
Meats, A.W., Clift, A.D., & Robson, M.K. (2003). Incipient founder populations of Mediterranean and
Queensland fruit flies in Australia: The relation of trap catch to infestation radius and models
for quarantine radius. Australian Journal of Experimental Agriculture, 43, 397-406.
Memmott, J., Craze, P.G., Harman, H.M., Syrett, P., & Fowler, S.V. (2005). The effect of propagule size
on the invasion of an alien insect. Journal of Animal Ecology, 74, 50-62.
Messina, F.J. & Jones, V.P. (1990). Relationship between fruit phenology and infestation by the apple
maggot (Diptera: Tephritidae) in Utah. Annals of the Entomological Society of America, 83,
742-752.
Metcalf, R.L. & Metcalf, E.R. (1992). Volatile kairomes as lures for insects. In Plant Kairomones In
Insect Ecology And Control (eds T.A. Miller & H.F. van Emden), pp. 153-161. Chapman and
Hall, New York.
Metcalf, R.L., Mitchell, W.C., Fukuto, T.R., & Metcalf, E.R. (1975). Attraction of the oriental fruit fly,
Dacus dorsalis, to methyl eugenol and related olfactory stimulants. Proceedings of the National
Academy of Science USA, 72, 2501-2505.
Michaux, B. & White, I.M. (1999). Systematics and biogeography of southwest Pacific Bactrocera
(Diptera: Tephritidae: Dacini). Palaeogeography, Palaeoclimatology, Palaeoecology, 153, 337-
351.
Moxon, J.E., (1988). Scapanes and Rhyncophorus. In Department of Agriculture & Livestock Annual
Research Report 1984-1988, pg. 67.
Moy, J.H. & Wong, L. (2002). The efficacy and progress in using radiation as a quarantine treatment of
tropical fruits: A case study in Hawaii. Radiation Physics and Chemistry, 63, 397-401.
Muthuthantri, W.S.N. (2008). Population biology of the tropical fruit fly, Bactrocera tryoni (Froggatt)
(Diptera: Tephritidae), in Queensland, Australia. In Unpublished M.Sc. thesis, pp. 177.
Queensland University of Technology, Brisbane, Australia.
Myers, J.H., Simberloff, D., Kuris, A.M., & Carey, J.R. (2000). Eradication revisited: dealing with
exotic species. Trends in Ecology & Evolution, 15, 316-320.

160
NARI (2005). High Yielding and Disease Tolerant Banana Varieties. In Papua New Guinea National
Agricultural Research Institute - 2003-2004 Biennial Report, pp. 25-26.
NARI (2007a). 1. High Yielding and Disease Tolerant Banana Varieties. In National Agricultural
Research Institute - Recently Released Technologies for Papua New Guinean Farmers, pp. 1-3.
NARI (2007b). 14. Banana Pest Control Technology Package. In National Agricultural Research
Institute - Recently Released Technologies for Papua New Guinean Farmers, pp. 14.
NARI (2008). National Agricultural Research Institute: Research programmes & Agro-ecological zones.
http://www.nari.org.pg/res/res.html, source dated: June 2008.
Narrod, C., Roy, D., Okello, J., Avendaño, B., Rich, K., & Thorat, A. (2009). Public–private
partnerships and collective action in high value fruit and vegetable supply chains. Food Policy,
34, 8-15.
Neven, L.G. & Drake, S.R. (2000). Comparison of alternative postharvest quarantine treatments for
sweet cherries. Postharvest Biology and Technology, 20, 107-114.
Nishida, R., Tan, K.-H., Wee, S.-L., Hee, A.K.-W., & Toong, Y.-C. (2004). Phenylpropanoids in the
fragrance of the fruit fly orchid, Bulbophyllum cheiri, and their relationship to the pollinator,
Bactrocera papayae. Biochemical Systematics and Ecology, 32, 245-252.
Nix, H. (1986). A biogeographic analysis of Australian Elapid snakes. In Snakes: Atlas of Elapid snakes
of Australia (ed R. Longmore), pp. 4–10. Bureau of Flora and Fauna, Canberra.
Novotny, V., Clarke, A.R., Drew, R.A.I., Balagawi, S., & Clifford, B. (2005). Host specialization and
species richness of fruit flies (Diptera: Tephritidae) in a New Guinea rain forest. Journal of
Tropical Ecology, 21, 67-77.
O'Connor, B.A. (1949). The banana scab moth, Nacoleia octasema Meyer., and its control. Fiji
Agricultural Journal, 20, 84-86.
Ohtsuka, R., Inaoka, T., Umezaki, M., Nakada, N., & Abe, T. (1995). Long-term subsistence adaptation
to the diversified Papua New Guinea environment. Global Environmental Change, 5, 347-353.
Onwueme, I. (1999). 4. Diseases and Pests. In Taro Cultivation In Asia And The Pacific. RAP
Publication 1999/16. Food and Agriculture Organization of the United Nations. Regional
Office for Asia and the Pacific, Bangkok, Thailand.
Orapa, W. (2006). Invasive Weeds, Impacts, Prevention, Detection and Responses. In Pest and Disease
Incursions: Risks, Threats and Management in Papua New Guinea. Papers presented at the 2nd
Papua New Guinea Plant Protection Conference, Kokopo, East New Britain Province, 8-10
November 2004 (ed T.V. Price), pp. 124-131. Canberra, ACIAR Technical Report No, 62.
Ostmark, H.E. (1974). Economic insect pests of bananas. Annual Review of Entomology, 19, 161-176.
Osuji, J.O., Okoli, B.E., Vuylsteke, D., & Ortiz, R. (1997). Multivariate pattern of quantitative trait
variation in triploid banana and plantain cultivars. Scientia Horticulturae, 71, 197-202.

161
PaDIL (2008). Erionota thrax distribution. http://www.padil.gov.au/viewPestDistribution.aspx?id=77.
Papadopoulos, N.T., Katsoyannos, B.I., Kouloussis, N.A., Hendrichs, J., Carey, J.R., & Heath, R.R.
(2001). Early detection and population monitoring of Ceratitis capitata (Diptera: Tephritidae)
in a mixed-fruit orchard in northern Greece. Economic Entomology, 94, 971-978.
Park, K. (2004). Assessment and management of invasive alien predators. Ecology and Society, 9.
Parker, I.M., Simberloff, D., Lonsdale, W.M., Goodell, K., Wonham, M., Kareiva, P.M., Williamson,
M.H., Von Holle, B., Moyle, P.B., Byers, J.E., & Goldwasser, L. (1999). Impact: Toward a
Framework for Understanding the Ecological Effects of Invaders. Biological Invasions, 1, 3-19.
Paul, I., van Jaarsveld, A.S., Korsten, L., & Hattingh, V. (2005). The potential global geographical
distribution of Citrus Black Spot caused by Guignardia citricarpa (Kiely): likelihood of disease
establishment in the European Union. Crop Protection, 24, 297-308.
Perrings, C. (2005). Mitigation and adaptation strategies for the control of biological invasions.
Ecological Economics, 52, 315-325.
Perrings, C., Williamson, M., & Dalmazzone, S. (2002). The Economics of Biological Invasions Edward
Elgar, Cheltenham, UK & Northampton, MA.
Perry, C.H. (1977). The Ecology and Control of Some Taro Pests in Papua New Guinea. In Agriculture
In The Tropics (eds B.C. Enyi & T. Varghese), pp. 319-322. University of Papua New Guinea,
Port Moresby.
Philemon, E.C. (1986). Radopholus similis: The Burrowing Nematode of Banana. In Papua New Guinea
Department of Primary Industries - Plant Pathology Note: No.31, pp. 2. Department of Primary
Industries, Port Moresby.
Pinese, B. & Piper, R. (1994). Banana & Queensland fruit flies in Bananas.
http://www2.dpi.qld.gov.au/horticulture/5517.html.
Ploetz, R. (2004). Diseases and pests: A review of their importance and management. InfoMusa, 13, 11-
16.
Pone, S. (1994). Status of Nematode and Weevil Borer Problems Affecting Banana in Some of the
Pacific Islands. In Workshop on Nematodes and Weevil Borers Affecting Bananas in Asia and
the Pacific (eds R.V. Valmayor, R.G. Davide, J.M. Stanton, N.L. Treverrow & V.N. Roa), pp.
90-104. Interntaional Network for the Improvement of Banana and Plantain (INIBAP),
Serdang, Selangor, Malaysia.
Putulan, D., Sar, S., Drew, R.A.I., Raghu, S., & Clarke, A.R. (2004). Fruit and vegetable movement on
domestic flights in Papua New Guinea and the risk of spreading pest fruit-flies (Diptera:
Tephritidae). International Journal of Pest Management, 50, 1, 17-22.
QDPI (2004). Subtropical Banana Grower's Handbook. http://www2.dpi.qld.gov.au/agrilink/17868.html,
218pp.

162
Raghu, S. (1998). Geographical distribution, seasonal abundance and habitat preference of fruit fly
species (Diptera: Tephritidae) in South-East Queensland, with special reference to Bactrocera
tryoni (Froggatt) and Bactrocera neohumeralis (Hardy). Australian Journal of Entomology, 37,
371-371.
Raghu, S. & Clarke, A.R. (2001). Distribution and abundance of Bactrocera bryoniae (Tryon) in three
different habitat types in South-East Queensland, Australia. International Journal of Ecology
and Environment Science, 27, 179-183.
Raghu, S., Drew, R., & Clarke, A. (2004). Influence of host plant structure and microclimate on the
abundance and behavior of a tephritid fly. Journal of Insect Behavior, 17, 179-190.
Rattanapun, W., Amornsak, W., & Clarke, A.R. (2009). Bactrocera dorsalis preference for and
performance on two mango varieties at three stages of ripeness. Entomologia Experimentalis et
Applicata, 131, 243-253.
Rochat, D., Morin, J.-P., Kakul, T., Beaudoin-Ollivier, L., Prior, R., Renou, M., Malosse, I., Stathers, T.,
Embupa, S., & Laup, S. (2002). Activity of male pheromone of Melanesian Rhinoceros Beetle
Scapanes australis. Chemical Ecology, 28, 479-500.
Rössler, Y., Epsky, N., & Heath, R.R. (1998). Trapping females of the Mediteranean fruit fly (Diptera:
Tephritidae) in Israel: Comparison of lures and trap type. Economical Entomology, 91, 1355–
1359.
Rowe, P. (1981). Breeding Bananas and Plantains 1. In Plant Breeding Reviews, pp. 135-153. Working
papers for the Genetic Engineering for Crop Improvement. Rockefeller Foundation.
Rowland, J.M., Qualls, C.R., & Beaudoin-Ollivier, L. (2005). Discrimination of alternative male
phenotypes in Scapanes australis (Boisduval) (Coleoptera: Scarabaeidae: Dynastinae).
Australian Journal of Entomology, 44, 22–28.
Ruesink, J.L., Parker, I.M., Groom, M.J., & Kareiva, P.M. (1995). Reducing the risks of nonindigenous
species introductions. Guilty until proven innocent. Bioscience, 45, 465-477.
Rull, J. & Prokopy, R.J. (2004). Host-finding and ovipositional-boring responses of Apple maggot
(Diptera: Tephritidae) to different apple genotypes. Environmental Entomology, 33, 1695-1702.
Sands, D., Baker, P., Dori, F., & Arura, M. (1988). Biological Control of Banana Skipper. In
Department of Agriculture & Livestock Annual Research Report 1984-1988, pp. 51-52.
Sands, D.P.A., Bakker, P., & Dori, F. (1993). Cortesia erionatae (Wilkinson) (Hymenoptera:
Braconidae) for biological control of banana skipper, Erionota thrax (L) (Lepidoptera:
Hesperiidae) in Papua New Guinea. Micronesica Supplementary, 4, 99-105.
Sands, D.P.A., Sands, M.C., & Arura, M. (1991). Banana skipper, Erionota thrax (L.)(Lepidoptera:
Hesperiidae) in Papua New Guinea: a new pest in the South Pacific region. Micronesica -
University of Guam, 1991 (No. 3 (suppl.)) 93-98
http://www.cababstractsplus.org/google/abstract.asp?AcNo=19916777502.

163
Sar, S., Balagawi, S., Mararuai, A., & Putulan, D. (2001). Fruit Fly Research and Development in PNG.
In Food Security For Papua New Guinea (eds R.M. Bourke, M.G. Allen & J.G. Salisbury), pp.
571-576. ACIAR Proceedings No 99, Canberra.
Sax, D.F. & Brown, J.H. (2000). The paradox of invasion. Global Ecology and Biogeography, 9, 361–
371.
Scheidel, U., Rohl, S., & Bruelheide, H. (2003). Altitudinal gradients of generalist and specialist
herbivory on three montane Asteraceae. Acta Oecologica, 24, 275-283.
Schillhorn van Veen, T.W. (2005). International trade and food safety in developing countries. Food
Control, 16, 491-496.
Schuhbeck, A. (1996). Banana Pest Research in New Guinea Islands Region: A Situation Report.
Prepared for first Banana Working Group Meeting. National Agricultural Research Institute -
LAES, Keravat, East New Britain Province, Papua New Guinea.
Schuhbeck, A. & Bokosou, J. (2006). The Distribution of Oryctes Baculovirus in Different Species of
Scarabaeidae on New Britain Island, Papua New Guinea. In Pest and Disease Incursions: Risks,
Threats and Management in Papua New Guinea. Papers presented at the 2nd Papua New
Guinea Plant Protection Conference, Kokopo, East New Britain Province, 8-10 November 2004
(ed T.V. Price), pp. 184-193. Canberra, ACIAR Technical Report No, 62.
Shack, K.W., Grivetti, L.E., & Dewey, K.G. (1990). Cash cropping, subsistence agriculture, and
nutritional status among mothers and children in lowland Papua New Guinea. Social Science &
Medicine, 31, 61-68.
Shellie, K.C. & Mangan, R.L. (2000). Postharvest disinfestation heat treatments: response of fruit and
fruit fly larvae to different heating media. Postharvest Biology and Technology, 21, 51-60.
Shigesada, N. & Kawasaki, K. (1997). Biological Invasions Theory and Practice Oxford University
Press, Oxford.
Shivas, R.G. & Philemon, E. (1996). Disease notes or new records: First record of Fusarium oxysporum
f. sp. cubense on banana in Papua New Guinea. Australasian Plant Pathology, 25, 260.
Simberloff, D. (2003a). Eradication-preventing invasions at the outset. Weed Science, 51, 247-253.
Simberloff, D. (2003b). How much information on population biology is needed to manage introduced
species? Conservation Biology, 17, 83-92.
Simbiken, N. (2006). Incidence of pathogens naturally associated with taro beetles in sugarcane fields in
Papua New Guinea. In Pest And Disease Incursions: Risks, Threats And Management In Papua
New Guinea (ed T.V. Price), pp. 20-23. ACIAR Technical Reports No.62, Canberra.
Simmonds, N.W. & Weatherup, S.T.C. (1990). Numerical taxonomy of the wild bananas (Musa). New
Phytologist, 115, 567-571.
Singer, M.C., Thomas, C.D., Billington, H.L., & Parmesan, C. (1989). Variation among conspecific
insect populations in the mechanistic basis of diet breadth. Animal Behaviour, 37, 751-759.

164
Smee, L. (1964). Insect pests of Hevea brasiliensis in the territory of Papua New Guinea; Their habits
and control. Papua New Guinea Agricultural Journal, 17, 21-28.
Smee, L. (1965). Insect pests of sweet potato and taro in the territory of Papua New Guinea; Their habits
and control. Papua New Guinea Agricultural Journal, 17, 99-101.
Smith, E.S.C. (1976). The Banana Fruit Fly, Dacus musae (Tryon) in Papua New Guinea. In
Proceedings of the 10th Waigani Seminar (eds B.C. Enyi & T. Verghese), pp. 323-329, Lae.
Smith, E.S.C. (1977a). A fruit fly trapping programme in the Northern Province. Science in New
Guinea, 5, 38-42.
Smith, E.S.C. (1977b). Studies on the biology and commodity control of the banana fruit fly, Dacus
musae (Tryon), in Papua New Guinea. Papua New Guinea Agricultural Journal, 28, 47-56.
Souza-Filho, M.F., Raga, A., Azevedo-Filho, J.A., Strikis, P.C., Guimarães, J.A., & Zucchi, R.A.
(2009). Diversity and seasonality of fruit flies (Diptera: Tephritidae and Lonchaeidae) and their
parasitoids (Hymenoptera: Braconidae and Figitidae) in orchards of guava, loquat and peach.
Brazilian Journal of Biology, 69, 31-40.
SPC Pacifly (2002). Banana fly (Bactrocera musae (Tryon)).
http://www.spc.int/pacifly/Species_profiles/B_musae.htm.
Stansbury, C., McKirdy, S., & Greg Power, G. (2000). Black sigatoka (Mycosphaerella fijiensisi):
Exotic threat to Western Australia, Rep. No. Factsheet No. 14/2000. ISSN 1443-7783.
HortguardTM Initiative AGWEST, Bentley, Western Australia.
Stark, J.D. (1995). Nocturnal behaviour of Oriental fruit flies and melon flies (Diptera: Tephritidae) and
associated parasitoids in a commercial papaya growing region on Kaua'i, Hawai'i. Proceedings
of the Hawaiian Entomology Society, 32, 149-151.
Staub, C.G., De Lima, F., & Majer, J.D. (2008). Determination of host status of citrus fruits against the
Mediterranean fruit fly, Ceratitis capitata (Wiedemann) (Diptera: Tephritidae). Australian
Journal of Entomology, 47, 184-187.
Steinbauer, M.J. (2002). Oviposition preference and neonate performance of Mnesampela privata in
relation to heterophylly in Eucalyptus dunii and E. globulus. Agricultural and Forest
Entomology, 4, 245-253.
Stephens, A.E.A., Kriticos, D.J., & Leriche, A. (2007). The current and future potential geographical
distribution of the Oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae). Bulletin of
Entomological Research, 97, 369-378.
Stice, K.N., McGregor, A.M., & Kumar, S.N. (2007). A review of fresh breadfruit exports from Fiji.
Acta Horticulture (ISHS), 757, 243-249.
Stonehouse, J.M., Mumford, J.D., Verghese, A., Shukla, R.P., Satpathy, S., Singh, H.S., Jiji, T.,
Thomas, J., Patel, Z.P., Jhala, R.C., Patel, R.K., Manzar, A., Shivalingaswamy, T.M.,
Mohantha, A.K., Nair, B., Vidya, C.V., Jagadale, V.S., Sisodiya, D.B., & Joshi, B.K. (2007).

165
Village-level area-wide fruit fly suppression in India: Bait application and male annihilation at
village level and farm level. Crop Protection, 26, 788-793.
Stover, R.H. (1986). Disease management strategies and the survival of the banana Industry. Annual
Review of Phytopathology, 24, 83-91.
Sugayama, R.L., Branco, E.S., Malavasi, A., Kovaleski, A., & Nora, I. (1997). Oviposition behavior of
Anastrepha fraterculus in apple and diel pattern of activities in an apple orchard in Brazil.
Entomologia Experimentalis et Applicata, 83, 239-245.
Sutherst, R.W., Collyer, B.S., & Yonow, T. (2000). The vulnerability of Australian horticulture to the
Queensland fruit fly, Bactrocera (Dacus) tryoni, under climate change. Australian Journal of
Agricultural Research, 51, 467-480.
Sutherst, R.W., Maywald, G.F., Yanow, T., & Stevens, P.M. (1999). CLIMEX: Predicting the effects of
climate on plants and animals, Rep. No. 0 643 06361 7. CSIRO Publishing, Collingwood,
Victoria.
Szent-Ivany, J.J.H. (1955). The cupmoth Scolelodes dinawa B.Bak (Family: Limacodidae) as a pest of
manila hemp & mango. Papua New Guinea Agricultural Journal, 9, 1-5.
Szent-Ivany, J.J.H. & Barrett, J.H. (1956). Some insects of banana in the territory of Papua and New
Guinea. Papua New Guinea Agricultural Journal, 11, 40-44.
Szent-Ivany, J.J.H. & Catley, A. (1960). Host plant and distribution records of some insects in New
Guinea and adjacent islands. Pacific Insects, 2, 255-261.
Tenakanai, D. (1996). Fruit Fly Fauna in Papua New Guinea. In Management of Fruit Flies in the
Pacific (eds A.J. Allwood & R.A.I. Drew), pp. 87-94. ACIAR Proceedings No 76, Canberra.
Theunis, W. & Aloali'i, I. (1999). Susceptibility of the taro beetle, Papuana uninodis (Coleoptera,
Scarabaeidae) to two new Bacillus popilliae isolates from Papuana spp. Journal of Invertebrate
Pathology, 73, 255-259.
Thistleton, B.M. & Masamdu, R.T. (1985). Surveys of insects associated with food crops in three study
areas in the Southern Highlands Province Department of Primary Indistry - Research Bulletin,
Port Moresby.
Thompson, H. (1986). Subsistence agriculture in Papua New Guinea. Journal of Rural Studies, 2, 233-
243.
Tomlinson, D.L. (1984). Bacterial head rot of banana. Harvest, 8, 141-143.
Tomlinson, D.L., King, G.A., & Ovia, A. (1988). Bacterial Corm and Rhizome Rot. In Department of
Agriculture & Livestock Annual Research Report 1984-1988, pp. 80-81.
Tscharntke, T., Thiessen, S., Dolch, R., & Boland, W. (2001). Herbivory, induced resistance, and
interplant signal transfer in Alnus glutinosa. Biochemical Systematics and Ecology, 29, 1025-
1047.

166
Usher, M.B. (1988). Biological invasions of nature reserves: A search for generalisations. Biological
Conservation, 44, 119-135.
Vargas, R.I., Miller, N.W., & Prokopy, R.J. (2002). Attraction and feeding responses of Mediterranean
fruit fly and a natural enemy to protein baits laced with two novel toxins, phloxine B and
spinosada. Entomologia Experimentalis et Applicata, 102, 273-282.
Vargas, R.I., Stark, J.D., Kido, M.H., Ketter, H.M., & Whitehand, L.C. (2000a). Methyl eugenol and
cue-lure traps for suppression of male Oriental fruit flies and Melon flies (Diptera: Tephritidae)
in Hawaii: Effects of lure mixtures and weathering. Economic Entomology, 93.
Vargas, R.I., Walsh, W.A., Kanehisa, D., Jang, E.B., & Armstrong, J.W. (1997). Demography of four
Hawaiian fruit flies (Diptera: Tephritidae) reared at five constant temperatures. Annals of the
Entomological Society of America, 90, 162-168.
Vargas, R.I., Walsh, W.A., Kanehisa, D., Stark, J.D., & Nishida, T. (2000b). Comparative demography
of three Hawaiian fruit flies (Diptera: Tephritidae) at alternating temperatures. Annals of the
Entomological Society of America, 93, 75-81.
Vayssieres, J.F., Carel, Y., Coubes, M., & Duyck, P.F. (2008). Development of immature stages and
comparative demography of two cucurbit-attacking fruit flies in Reunion Island: Bactrocera
cucurbitae and Dacus ciliatus (Diptera Tephritidae). Environmental Entomology, 37, 307-314.
Vera, M.T., Rodriguez, R., Segura, D.F., Cladera, J.L., & Sutherst, R.W. (2002). Potential geographical
distribution of the Mediterranean fruit fly, Ceratitis capitata (Diptera: Tephritidae), with
emphasis on Argentina and Australia. Environmental Entomology, 31, 1009-1022.
Vermeij, G.J. (1996). An agenda for invasion biology. Biological Conservation, 78, 3-9.
Vueti, E.T., Ralulu, L., Walker, G.P., Allwood, A.J., Leweniqila, L., & Balawakula, A. (1996). Host
Availability - its Impact on Seasonal Abundance of Fruit Flies. In Management of Fruit Flies in
the Pacific (eds A.J. Allwood & R.A.I. Drew), pp. 105-110. ACIAR Proceedings No 76,
Canberra.
Walter, A. & Sam, C. (2002). Fruits of Oceania ACIAR Monograph No. 85, Canberra [trs P. Ferrar
from Fruits d'Oceanie].
Wamala, M.H. (2001). PNG's Food Production Constraints and Analysis: Agronomy, Agricultural
Extension and Socioeconomic Points of View. In Food Security For Papua New Guinea (eds
R.M. Bourke, B.J. Allen & J.G. Salisbury), pp. 516-519. Proceedings of the Papua New Guinea
Food And Nutrition 2000 Conference, PNG University of Technology, Lae, 26-30 June 2000.
ACIAR Proceedings No 99.
Wang, X.-G., Jarjees, E.A., McGraw, B.K., Bokonon-Ganta, A.H., Messing, R.H., & Johnson, M.W.
(2005). Effects of spinosad-based fruit fly bait GF-120 on tephritid fruit fly and aphid
parasitoids. Biological Control, 35, 155-162.

167
Waterhouse, D.F. (1997). The Major Invertebrate Pests and Weeds of Agriculture and Plantation
Forestry in the Southern and Western Pacific. ACIAR Monograph No, 44, 99.
Waterhouse, D.F., Dillon, B., & Vincent, D. (1998). Economic benefits to Papua New Guinea and
Australia from the biological control of banana skipper (Erionota thrax). In Impact Assessment
Series No. 12, pp. 36. ACIAR, Canberra.
Waterhouse, D.F. & Norris, K.R. (1989). Erionota thrax (Linnaeus). In Biological control: Pacific
prospects - supplement 1, pp. 88-100. ACIAR Monograph No. 12, Canberra.
Wesis, P., Niangu, B., Ero, M., Masamdu, R., Autai, M., Elmouttie, D., & Clarke, A.R. (in press). Host
use and crop impacts of Oribius Marshall species (Coleoptera: Curculionidae) in Eastern
Highlands Province, Papua New Guinea. Bulletin of Entomological Research.
West, S.A. & Cunningham, J.P. (2002). A general model for host plant selection in phytophagous
insects. Theoretical Biology, 214, 499-513.
Whittle, P. (2004). Invasive arthropods in agriculture: problems and solutions. Australian Journal of
Entomology, 43, 208-209.
Wilkie, L., Hela, F., & Moxon, J. (1993). Banana scab moth. Harvest, 15.
Williamson, M.H. & Fitter, A. (1996). The characters of successful invaders. Biological Conservation,
78, 163-170.
Woodruff, R.E. (1969). The banana root borer (Cosmopolites sordidus (Germar)) (Coleoptera:
Curculionidae) in Florida. Entomology Circular, 48, 2.
Worner, S.P. & Gevrey, M. (2006). Modelling global insect pest species assemblages to determine risk
of invasion. Journal of Applied Ecology, 43, 858-867.
Wright, M.G., Hoffmann, M.P., Kuhar, T.P., Gardner, J., & Pitcher, S.A. (2005). Evaluating risks of
biological control introductions: A probabilistic risk-assessment approach. Biological Control,
35, 338-347.
Wright, W.D. (1984). The basic concepts and attributes of colour order systems. Color Research &
Application, 9, 229-233.
Yonow, T. & Sutherst, R.W. (1998). The geographical distribution of the Queensland fruit fly,
Bactrocera (Dacus) tryoni, in relation to climate. Australian Journal of Agricultural Research,
49, 935-953.
Yonow, T., Zalucki, M.P., Sutherst, R.W., Dominiak, B.C., Maywald, G.F., Maelzer, D.A., & Kriticos,
D.J. (2004). Modelling the population dynamics of the Queensland fruit fly, Bactrocera
(Dacus) tryoni: A cohort-based approach incorporating the effects of weather. Ecological
Modelling, 173, 9-30.
Zavaleta, E.S., Hobbs, R.J., & Mooney, H.A. (2001). Viewing invasive species removal in a whole-
ecosystem context. Trends in Ecology & Evolution, 16, 454-459.

168
Zepeda, C., Salman, M., & Ruppanner, R. (2001). International trade, animal health and veterinary
epidemiology: challenges and opportunities. Preventive Veterinary Medicine, 48, 261-271.
Zhou, C.Q., Wu, K.K., Chen, H.D., Yang, P.J., & Dowell, R.V. (1994). Effect of Temperature on the
Population Growth of Bactrocera tau (Walker) (Dipt, Tephritidae). Journal of Applied
Entomology-Zeitschrift Fur Angewandte Entomologie, 117, 332-337.

Appendices
Appendix 1 Fruit flies in Papua New Guinea
Leblanc, L., Balagawi, S., Mararuai, A., Putulan, D., Tenakanai, D., & Clarke, A.R.
(2001). Fruit flies in Papua New Guinea. In Pest Advisory Leaflet No. 37, pp. 12. Plant
Protection Service, Secretariat of the Pacific Community, Suva.
halla
Due to copyright restrictions, this article is not available here. Please consult the hardcopy thesis available from QUT Library

182
Appendix 2 Distribution and Biogeography of Bactrocera and Dacus species (Diptera: Tephritidae) in Papua New Guinea
Clarke, A. R., S. Balagawi, B. Clifford, R. A. I. Drew, L. Leblanc, A. Mararuai, D.
McGuire, D. Putulan, T. Romig, S. Sar, and D. Tenakanai. 2004. Distribution and
biogeography of Bactrocera and Dacus species (Diptera: Tephritidae) in Papua New
Guinea. Australian Journal of Entomology 43:148-156.
halla
Due to copyright restrictions, this article is not available here. Please consult the hardcopy thesis available from QUT Library

192
Appendix 3 Introduction and Distribution of Bactrocera musae (Tryon) (Diptera: Tephritidae) in East New Britain, Papua New Guinea
Mararuai, A., Allwood, A.J., Balagawi, S., Dori, F., Kalamen, M., Leblanc, L., Putulan,
D., Sar, S., Schuhbeck, A., Tenakanai, D., & Clarke, A.R. (2001). Introduction and
distribution of Bactrocera musae (Tryon) (Diptera: Tephritidae) in East New Britain,
Papua New Guinea. Papua New Guinea Agricultural Journal, 45, 59-65.
halla
Due to copyright restrictions, this article is not available here. Please consult the hardcopy thesis available from QUT Library


















