
Lymphocyte-Mediated Cytotoxicity Dogs Infected Virulent ... · CELLULAR IMMUNITY IN CANINE...
9
Vol. 37, No. 2 INFECTION AND IMMUNITY, Aug. 1982, p. 592-600 0019-9567/82/080592-09$02.00/0 Lymphocyte-Mediated Immune Cytotoxicity in Dogs Infected with Virulent Canine Distemper Virus MAX J. G. APPEL,'* WILLIAM R. SHEK,tt AND BRIAN A. SUMMERS2 James A. Baker Institute for Animal Health, Department of Microbiology,1 and Department of Pathology,2 New York State College of Veterinary Medicine, Cornell University, Ithaca, New York 14853 Received 8 January 1982/Accepted 12 April 1982 Immune lymphocyte-mediated cytotoxicity (ILMC) was evaluated in dogs after intranasal exposure to one of the following three virulent strains of canine distemper virus: Cornell A75/17, Ohio R252, and Snyder Hill. Cytotoxicity was tested with peripheral blood lymphocytes as effector cells and primary dog testicle cells that were matched for histocompatibility as target cells. A strong correlation was found between ILMC and the course of the infection. Dogs that succumbed to encephalitis with any of the strains had little or no ILMC, whereas dogs that recovered had the highest activity. In the intermediate range were dogs with a delayed or reduced ILMC which developed persistent but subclinical central nervous system infections. A significant difference in onset, peak, and duration of ILMC was observed in dogs infected with different strains of canine distemper virus. ILMC responses began at 14 days postinfection (p.i.), reached a peak at 21 to 28 days p.i., and returned to preinoculation levels by 63 to 70 days p.i. in canine distemper virus A75/17- and R252-infected dogs. In contrast, ILMC in canine distemper virus Snyder Hill-infected dogs began at 10 days p.i., peaked by 14 to 17 days p.i., and approached preinoculation levels by 28 days p.i. Antiviral immunity as measured by ILMC appears to be a critical factor in determining the outcome in canine distemper virus-infected hosts. Furthermore, for certain viral biotypes, a delayed ILMC response correlated with persistent infection of the central nervous system. Canine distemper virus (CDV), a morbillivirus closely related to measles virus (3), causes a fatal infection with central nervous system (CNS) involvement in dogs and other carni- vores. The Snyder Hill (SH) strain (2) causes an acute encephalomyelitis with predominantly gray-matter changes, whereas strains Ohio R252 (18, 19) and Cornell A75/17 (24) cause delayed encephalomyelitis with predominantly white- matter lesions and demyelination reminiscent of CNS lesions in multiple sclerosis. Regardless of the virus strain, some dogs recover spontane- ously, whereas others succumb. In addition, strain R252 or A75/17 may induce persistent infection without clinical signs of CNS disease (18, 19, 24). Thus, the outcome of infection depends upon both viral and host factors. A crucial host factor that limits the spread of virus is the immune response. In canine distem- per, a correlation has been found between sur- vival and the capability of dogs to produce CDV- neutralizing antibody (2, 15). However, both humoral and cellular immune activity are impor- t Present address: Department of Avian and Aquatic Ani- mal Medicine, New York State College of Veterinary Medi- cine, Cornell University, Ithaca, NY 14853. tant for effective resistance to viral diseases. In a previous communication, we demonstrated that dogs vaccinated with attenuated CDV mount a strong virus-specific, lymphocyte-mediated cy- totoxic response against 1Cr-labeled, CDV-in- fected dog testicle cells. Using the same system, we report here a strong correlation between suppressed immune lymphocyte-mediated cyto- toxicity (ILMC) and fatal or persistent infection of dogs exposed to virulent CDV. MATERIALS AND METHODS Virus strains. Three virulent strains of CDV were used for dog inoculations; SH and Cornell A75/17 strains were isolated in the James A. Baker Institute for Animal Health, Ithaca, N.Y. The Ohio R252 strain was kindly supplied by A. Koestner. All three strains were maintained in frozen spleen suspensions that were harvested from specific-pathogen-free dogs 5 days postinfection (p.i.). The attenuated Rockborn strain of CDV was used for the infection of target cells. For virus neutralization tests, the Vero-cell-adapted Onderstepoort strain of CDV was employed. Animal inoculation, experimental design, and prepa- ration of target and effector cells. Specific-pathogen- free male beagles from the Baker Institute colony were used. Peripheral blood leukocytes were typed for dog leukocyte antigens (DLA) by Frances D. Cannon (20). 592 on June 6, 2020 by guest http://iai.asm.org/ Downloaded from
Transcript of Lymphocyte-Mediated Cytotoxicity Dogs Infected Virulent ... · CELLULAR IMMUNITY IN CANINE...

Vol. 37, No. 2INFECTION AND IMMUNITY, Aug. 1982, p. 592-6000019-9567/82/080592-09$02.00/0
Lymphocyte-Mediated Immune Cytotoxicity in Dogs Infectedwith Virulent Canine Distemper Virus
MAX J. G. APPEL,'* WILLIAM R. SHEK,tt AND BRIAN A. SUMMERS2James A. Baker Institute for Animal Health, Department of Microbiology,1 and Department of Pathology,2
New York State College of Veterinary Medicine, Cornell University, Ithaca, New York 14853
Received 8 January 1982/Accepted 12 April 1982
Immune lymphocyte-mediated cytotoxicity (ILMC) was evaluated in dogs afterintranasal exposure to one of the following three virulent strains of caninedistemper virus: Cornell A75/17, Ohio R252, and Snyder Hill. Cytotoxicity was
tested with peripheral blood lymphocytes as effector cells and primary dog testiclecells that were matched for histocompatibility as target cells. A strong correlationwas found between ILMC and the course of the infection. Dogs that succumbed toencephalitis with any of the strains had little or no ILMC, whereas dogs thatrecovered had the highest activity. In the intermediate range were dogs with adelayed or reduced ILMC which developed persistent but subclinical centralnervous system infections. A significant difference in onset, peak, and duration ofILMC was observed in dogs infected with different strains of canine distempervirus. ILMC responses began at 14 days postinfection (p.i.), reached a peak at 21to 28 days p.i., and returned to preinoculation levels by 63 to 70 days p.i. in caninedistemper virus A75/17- and R252-infected dogs. In contrast, ILMC in caninedistemper virus Snyder Hill-infected dogs began at 10 days p.i., peaked by 14 to 17days p.i., and approached preinoculation levels by 28 days p.i. Antiviral immunityas measured by ILMC appears to be a critical factor in determining the outcome incanine distemper virus-infected hosts. Furthermore, for certain viral biotypes, adelayed ILMC response correlated with persistent infection of the central nervoussystem.
Canine distemper virus (CDV), a morbillivirusclosely related to measles virus (3), causes afatal infection with central nervous system(CNS) involvement in dogs and other carni-vores. The Snyder Hill (SH) strain (2) causes anacute encephalomyelitis with predominantlygray-matter changes, whereas strains Ohio R252(18, 19) and Cornell A75/17 (24) cause delayedencephalomyelitis with predominantly white-matter lesions and demyelination reminiscent ofCNS lesions in multiple sclerosis. Regardless ofthe virus strain, some dogs recover spontane-ously, whereas others succumb. In addition,strain R252 or A75/17 may induce persistentinfection without clinical signs of CNS disease(18, 19, 24). Thus, the outcome of infectiondepends upon both viral and host factors.A crucial host factor that limits the spread of
virus is the immune response. In canine distem-per, a correlation has been found between sur-vival and the capability of dogs to produce CDV-neutralizing antibody (2, 15). However, bothhumoral and cellular immune activity are impor-
t Present address: Department of Avian and Aquatic Ani-mal Medicine, New York State College of Veterinary Medi-cine, Cornell University, Ithaca, NY 14853.
tant for effective resistance to viral diseases. In aprevious communication, we demonstrated thatdogs vaccinated with attenuated CDV mount astrong virus-specific, lymphocyte-mediated cy-totoxic response against 1Cr-labeled, CDV-in-fected dog testicle cells. Using the same system,we report here a strong correlation betweensuppressed immune lymphocyte-mediated cyto-toxicity (ILMC) and fatal or persistent infectionof dogs exposed to virulent CDV.
MATERIALS AND METHODS
Virus strains. Three virulent strains of CDV wereused for dog inoculations; SH and Cornell A75/17strains were isolated in the James A. Baker Institutefor Animal Health, Ithaca, N.Y. The Ohio R252 strainwas kindly supplied by A. Koestner. All three strainswere maintained in frozen spleen suspensions thatwere harvested from specific-pathogen-free dogs 5days postinfection (p.i.). The attenuated Rockbornstrain ofCDV was used for the infection of target cells.For virus neutralization tests, the Vero-cell-adaptedOnderstepoort strain of CDV was employed.Animal inoculation, experimental design, and prepa-
ration of target and effector cells. Specific-pathogen-free male beagles from the Baker Institute colony wereused. Peripheral blood leukocytes were typed for dogleukocyte antigens (DLA) by Frances D. Cannon (20).
592
on June 6, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

CELLULAR IMMUNITY IN CANINE DISTEMPER 593
Testicles were surgically removed when dogs were 3months of age and used to prepare CDV Rockborn-infected fibroblast target cells for ILMC assays. Be-tween 70 and 90%o of the target cells were infected asshown by immunofluorescence. Infected target cellsand uninfected testicle cells were stored in liquidnitrogen as described previously (22).
After DLA typing and preparation of target cells, alldogs were inoculated intranasally at 4 months of agewith approximately 5 x 103 dog lung macrophage(DLM) infectious doses of virulent virus in dog spleensuspensions in minimal essential medium with Earlesalts. A total of 7 dogs were exposed to CDV SH, 12were exposed to A75/17, and 6 were exposed to R252.Body temperature was recorded daily, and diseaseprogress was monitored by weight loss, peripheralblood lymphocyte counts, and observation of clinicalsigns.Heparinized blood samples for the preparation of
peripheral blood lymphocytes to be used as effectorcells in ILMC assays were taken twice before infec-tion, on days 7, 10, 14, 17, and 21 PI, and in weeklyintervals thereafter until ILMC was detectable atinsignificant levels (<10%o). Effector lymphocyteswere prepared from 10 to 30 ml of heparinized blood,the volume depending upon the level of lymphopeniaof CDV-infected dogs. Phagocytic cells were removedwith carbonyl-iron powder, and lymphocytes werebanded in Ficoll-Hypaque (Pharmacia, Fine Chemi-cals, Inc., N.J.) as described previously (22).ILMC assay. The ILMC assay was described in
detail previously (22). Briefly, peripheral blood lym-phocyte effector cells were tested for their ability tolyse CDV Rockborn-infected target cells at the timesspecified above. Target cells were removed from liq-uid nitrogen, thawed, washed, suspended in MLAmedium (22) supplemented with fetal calf serum, andplaced into flat-bottomed, 96-well microplates (Lin-bro, Div. of Flow Laboratories, Inc., Rockville, Md.)for attachment overnight. Cell concentration wasadjusted to approximately 5 x 103 attached target cellsper well. Target cells were labeled with Na[51Cr]04 asdescribed previously (22). Lymphocyte preparationswere added to target cells in microplate wells ateffector/target cell ratios of 100:1 in 0.2 ml of medium.Cells were incubated for 6 h at 37°C and 5% CO2 in air.Immediately thereafter, supernatant fluids were com-pletely removed with the Titertek collection system(Flow Laboratories). The remaining cells were dis-solved in 1% sodium dodecyl sulfate and also harvest-ed with Titertek. Radioactivity was counted in aBeckman 4000 Gamma Counting System (BeckmanInstruments, Inc., Fullerton, Calif.).Measurement of lymphocyte-mediated cytotoxicity.
The total 51Cr released during the assay period wascomputed as follows: (total 51Cr released) = [superna-tant - (sodium dodecyl sulfate-dissolved cells + su-pernatant)] x 100. The percentage of spontaneousrelease was the total 51Cr released in the absence ofeffector lymphocytes. Cytotoxicity caused by thepresence of lymphocytes was computed as follows:(lymphocyte-mediated cytotoxicity) = [(total 5 Cr re-leased) - (spontaneous release)]. If the lymphocyte-mediated cytotoxicity was caused by nonimmmunelymphocytes, it was termed natural lymphocyte-medi-ated cytotoxicity. After exposure to CDV, the per-centage of ILMC was computed as follows: ILMC =
[(lymphocyte-mediated cytotoxicity) - (natural lym-phocyte-mediated cytotoxicity)].
Virus neutralization. Virus neutralization tests weremade with the Onderstepoort strain of CDV in Verocells in flat-bottomed, 96-well microplates (Linbro) asdescribed previously (5).
Virus isolations. Dogs were euthanized either whenmoribund or electively up to 70 days p.i. The lattergroup, which was clinically stable, included both per-sistently infected and recovered dogs. A few dogswere kept for clinical observation beyond 70 days andwere not necropsied (see Table 1). Brain tissues weresterilely removed from euthanized dogs. Virus isola-tion attempts from brain tissues were made in thefollowing two ways. (i) Brain tissues were ground andsuspended in minimal essential medium with Earlesalts. Serial dilutions were inoculated into primaryDLM cultures as described previously (4). Virus-induced syncytial formation 5 days after inoculationand direct immunofluorescence were used as criteriafor the presence of CDV. (ii) Brain tissues wereminced in minimal essential medium with Earle saltsand mechanically stirred for 1 h at 20°C. Free cellswere filtered through cheesecloth, washed three timesin minimal essential medium with Earle salts, andincubated at 37°C in 5% C02-95% air in 40%o McCoymedium-40o Leibovitz medium-20% fetal calf serum(GIBCO, Grand Island, N.Y.)-antibiotics-29.2% glu-tamine. Between 6 and 8 days after initiation, culturesusually had formed monolayers. They were then pas-saged, and cover slips were seeded with secondarycells. At 5 to 7 days later, when monolayers wereformed again, cover slips were stained for directimmunofluorescence, and the presence of free viruswas tested by isolation in DLM cultures as describedpreviously (4). If results were negative at that time,cultures were passaged a second time and monitoredfor presence of virus as described.CNS pathology. One-half of the brain (sagittal sec-
tion) was fixed in 10%o buffered Formalin and routinelyprocessed for light microscopy. Sections were takenjust rostral to the corpus callosum, through the inter-thalamic adhesion, at the rostral colliculus, obliquelythrough the optic cortex, and medially (sagittal plane)through the cerebellum and medulla. Slides werestained with hematoxylin-eosin.
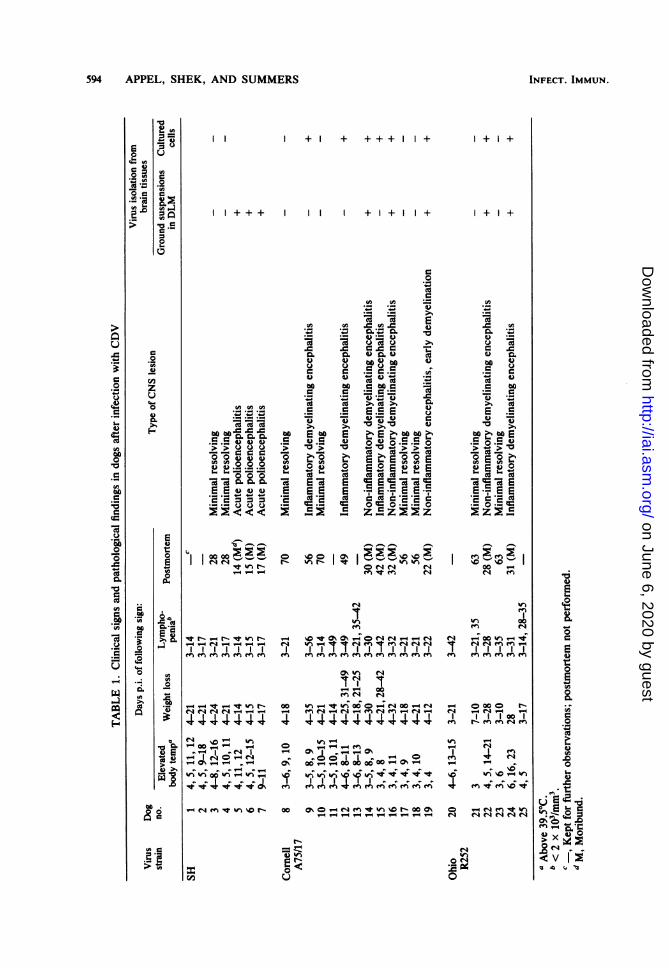
RESULTSClinical signs. Dogs with elevated body tem-
perature (above 39.5°C) were generally de-pressed, and anorexia or diarrhea or both wasfollowed by weight loss. Fever, weight loss, andlymphopenia are listed in Table 1. All infecteddogs became depressed and anorexic by 3 or 4days p.i. They became clinically stable between14 and 21 days p.i., with the exception of dogsfatally infected with CDV SH, which deteriorat-ed rapidly, became moribund 14 to 17 days p.i.,and were euthanized. Dogs that succumbed toCDV A75/17 or R252 infection between 22 and42 days p.i. became depressed and anorectic 5 to10 days earlier. CNS signs (convulsions, ataxia,spasms, myoclonus, paresis, paralysis, or blind-ness) occurred between one and several days
VOL. 37, 1982
on June 6, 2020 by guesthttp://iai.asm
.org/D
ownloaded from
594 APPEL, SHEK, AND SUMMERS INFECT. IMMUN.
+ + +++ +
+++ + + +
0)
C)
Co
0=
* *-
E~~.E.E
G 0
H00 . -a*Au"4
-e- re _t %
0
CISac
0) IL) -
.eF.e A.
bcW z P4 4)S 2= L
8) .02. 44)* _4)
.W C)CeW> s
Co OCo0
.2 2.EmE~ 0,.5gE=>< o~ 4)-*2 Z Z2
00 tn_ W tn0eW rs1 I4
-~~~~~~
Cs \04 en q m _I "
o r--tO 00 _
0o°s - 0
+ I+
+ I+
._
0 .04) s.
0 C0
2 ._
00 r m
en en en
T'- 0000 N-
cd-
_ _
en %O4en
r r'
00 0 m *'n 0 0 f fi" nT-~ - V- -4 t1 V1 V- 1- r- eq 4 e4 el l
N-
Et u
0
42.(0c
0 U24)-
c
00
la2=
u4)u)
c:0.)2
=a.=-0
0
C0._
.2
00
zu
4)
I-
.03
r.
0
4)
w
'4-
ce10
Co
00C_10c
-Cw
.200.6a
10
w
0)
oocw
-a.
m-,F:
24)
0
0O4-
0,oML.*VF.2
04)
)>
0
- .@
0 S
v 2
.2
C-p
0
._
-6
4)
a
0
.4)A0
0)c
0.0)
0
F.4)6
s
0)0.
0
*t4).0_D .
4E w
Co.
(A 004 -g>02
0)°c
.e .g
on June 6, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

CELLULAR IMMUNITY IN CANINE DISTEMPER 595
before dogs became moribund. One dog (no. 12)developed a facial myoclonus on day 35 p.i.,which remained unchanged until day 49 p.i.,when he was euthanized (Table 1).ILMC. The level and duration of ILMC de-
pended on the clinical progression of disease andthe strain of CDV. Dogs that recovered and hadminimal resolving CNS lesions mounted muchgreater ILMC responses than those that suc-cumbed to infection. Little or no ILMC wasdetected in the latter group (see Fig. 1A, 2A, and3A). Of six dogs infected with A75/17 that hadno or low levels of ILMC response on day 21p.i., three (no. 14, 16, and 19) were moribundbetween days 22 and 32 p.i. One dog (no. 15)that became moribund on day 42 p.i. had onlylow ILMC responses on days 14, 21, 28, and 42p.i. In one clinically stable dog (no. 9) that
706
60 0
50 S
*40z }\
0
*2300
:5.20/
developed inflammatory demyelinating enceph-alitis, low levels of ILMC were found on days28, 35, and 56 p.i., when he was euthanized. Onedog that developed a facial myoclonus on day 35p.i. but that was otherwise clinically stable be-gan to respond on day 28 and had increasingILMC levels up to day 49 p.i., when he waseuthanized (Fig. 1A). Two CDV R252-infecteddogs that became moribund at days 28 and 31p.i. and three CDV SH-infected dogs that be-came moribund between days 12 and 17 p.i. hadno measurable ILMC.Dogs which stabilized after infection with the
strains ofCDV that cause delayed encephalomy-elitis (A75/17 and R252) had ILMC responsesthat persisted significantly lower (P < 0.05) thanthose detected in survivors of CDV SH infection(Table 2). Six dogs infected with CDV A75/17
A
S
S
.
Days Post Infection
0
*° 21 -
zi
l 14 21 28 35 42 49 56 63 70
Days Pos rInection
FIG. 1. (A) ILMC response of 12 dogs after intranasal exposure to CDV A75/17. Upper line, Mean valuesfrom six dogs (0, 0) that recovered; lower line, mean values from six dogs (O, *) with encephalitis that becamemoribund or that had persistent CDV in the CNS between 28 and 56 days p.i. (B) Neutralizing-antibody titers inthe sera of 12 dogs after intranasal exposure to CDV A75/17. Upper line, Mean values from six dogs (0, 0) thatrecovered; lower line, mean values from six dogs (l, *) with encephalitis that became moribund or that hadpersistent CDV in the CNS between 28 and 56 days p.i.
VOL. 37, 1982
on June 6, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

5% APPEL, SHEK, AND SUMMERS
TABLE 2. ILMC in clinically stable dogs after CDV infectionILMC in dogs infected with following CDV virus strain:
Days SH A75/17 R-252r--- No. positive/ % ILMC ± SEMb No. positive/ % ILMC + SEM No. positive/ % ILMC ± SEM
totala %IM±SEtotal % LC±SMtotal
%IM E
10 2/4 11.8 ± 5.08 0/6 <3 0/4 <314 4/4 28.6 ± 6.81c 2/6 11.2 ± 3.89d 0/4 5.25 ± 1.11d21 4/4 14.63 ± 0.88c 6/6 42.92 ± 5.89d 4/4 35.48 ± 6 90d28 0/4 5.88 ± 1.26c 6/6 36.50 ± 7.07" 4/4 37.48 ± 4.93da Number of dogs with ILMC .10%/number of dogs tested.b Mean ILMC for all dogs tested.c.d Different superscripts within a row denotes statistically significant differences in the levels of ILMC (P <
0.05 by the unpaired Student's t test).
that became clinically stable after 2 to 3 weeksp.i. had initial ILMC responses on day 14 p.i.;maximal levels occurred on day 21 p.i., followedby a gradual decline thereafter until 70 days p.i.,when levels had reached or approached preinoc-ulation values (Fig. 1A). Similarly, four of sixCDV R252-inoculated dogs that clinically recov-ered developed peak ILMC levels on day 28,with a gradual decline until day 63 p.i., whenthey approached preinoculation values (Fig.2A). In contrast, four dogs infected with CDVSH that had clinically recovered between 2 and 3weeks p.i. had ILMC responses that began onday 10 p.i., reached a peak on days 14 and 17,were greatly reduced by day 21, and approachedpreinoculation levels by day 28 p.i. (Fig. 3A).
Virus neutralization. Onset of CDV antibodyin dogs that became moribund was delayed, andtiters remained low or absent (Fig. 1B, 2B, and3B). CDV SH-infected dogs that later recoveredhad measurable virus-neutralizing antibody by10 days that became maximal at 17 to 21 days(Fig. 3B). Dogs that recovered from CDV A75/17 or R252 infection had initial CDV antibody by14 days; titers increased up to 42 days p.i. (Fig.1B, and 2B) and remained constant thereafter.Virus isolation. Virus was isolated in DLM
cultures from brain suspensions of dogs thatbecame moribund with acute polioencephalitissubsequent to infection with CDV SH and non-inflammatory demyelinating encephalitis subse-quent to infection with CDV A75/17 and R252.Brain explant cultures from these same dogsbecame virus positive. Virus was also detectedin brain explant cultures from dogs infected withCDV A75/17 and R252 that had inflammatoryencephalitis. However, brain suspensions fromthese dogs were, with one exception, virus nega-tive in DLM culture. Virus isolation from recov-ered dogs remained negative, irrespective of thetechnique used (see Table 1).
Histopathology. Neuropathological findings indogs which developed clinical disease or persist-
ent infection of the CNS varied with the viralbiotype. Dogs which succumbed to CDV SHinfection had an encephalitis centered on gray-matter areas (cerebral cortex, basal ganglia,thalamus, and hypothalamus). The reaction re-vealed microgliosis, neuronal satellitosis andneuronophagia, and narrow perivascular cuffs oflymphoid cells in the immediate vicinity of neu-ronal injury. Specific viral inclusions were infre-quently observed, mainly in neuronal cyto-plasm. Occasional small glial foci occurred inmyelinated tissue, but demyelination did notoccur.
Lesions in myelinated areas induced by theA75/17 and R252 strains of CDV were predomi-nantly subependymal or subpial and most severein the metencephalon. Myelin loss was typicallyseen as a vacuolar change with an accompanyingastrocytosis. Sometimes syncytial astrocyteswere present, and viral inclusions were mostreadily found within the nuclei and cytoplasm ofastrocytes. In some animals, this reaction wasaccompanied by an infiltrate of lymphocytes andmacrophages into the overlying leptomeningesand into the demyelinative lesions beginning asperivascular aggregates. Accordingly, thesechanges were recorded as non-inflammatory orinflammatory demyelinating encephalitis (Table1).Minimal resolving lesions were found in dogs
that had recovered from experimental exposure,regardless of biotype. These lesions were char-acterized by sporadic small glial nodules whichwere scattered randomly through the neuraxis.They were not present in all sections examinedand, iffewer levels had been studied, could havebeen missed.DLA in relation to disease. A distinct pattern
between DLA and disease could not be estab-lished. Most dogs that became moribund or thathad persistent infection had DLAs that wereidentical to DLAs of dogs that recovered (Table3). However, dog no. 19, which was the only dog
INFECT. IMMUN.
on June 6, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

CELLULAR IMMUNITY IN CANINE DISTEMPER 597
A
I
0
0
14 ii i8 35 4-2Days Post Infedion
B
0
0
I
0
Days Post Infeon
FIG. 2. (A) ILMC response of six dogs after intranasal exposure to CDV R252. Upper line, Mean values fromfour dogs (0, *) that recovered; lower line, mean values from two dogs (O, *) with encephalitis that becamemoribund 28 and 31 days p.i. (B) Virus-neutralizing-antibody titers in the sera of six dogs after intranasalexposure to CDV R252. Upper line, Mean values from four dogs (0, 0) that recovered; lower line, mean valuesfrom two dogs (l, *) with encephalitis that became moribund 28 and 31 days p.i.
in this study without g and but with m antigen,was the first dog that became moribund (day 22p.i.) after exposure to a chronic strain of CDV.
DISCUSSIONCell-mediated immune responses in CDV-in-
fected dogs that have been studied earlier in-cluded skin tests (11), lymphocyte blastogenesis(13), lymphocyte-mediated inhibition of CDV-induced syncytial formation (16), antibody-de-pendent cellular cytotoxicity (12), and naturallymphocyte-mediated cytotoxicity and ILMC invaccinated dogs (22).We previously reported (22) that ILMC in
response to vaccination was mediated by T cellsand not by NK or antibody-dependent, cell-
mediated cytotoxicity for the following reasons.(i) The level of ILMC was unaffected by deple-tion of B cells or Fc receptor-bearing cells. (ii)CDV antibody persisted in the sera of vaccinat-ed dogs long after ILMC decreased to insignifi-cant levels. Although ILMC was CDV specific,since uninfected and SV5-infected target cellswere not lysed, antibody neither blocked norenhanced ILMC, suggesting recognition of lym-phocyte-defined CDV antigens. (iii) Interferonenhances NK cell activity (26), but Tsai (S. Tsai,Ph.D. thesis, Cornell University, Ithaca, N.Y.)could not detect interferon in the sera of vacci-nated dogs. (iv) Finally, ILMC was geneticallyrestricted, as is known for virus-specific, T-cell-mediated cytotoxicity (8).
60.
50*
a
o
so
, 40u
O 30
u
E 20EE
10,
0
2'0
0z
VOL. 37, 1982
14 56 63
on June 6, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

598 APPEL, SHEK, AND SUMMERS
TABLE 3. Serologically defined DLA of experimental dogs
Virus strain Dog DLA Resultsno.
SH 1 b c g k 1 n o Clinically stable2 b c e g k 1 n o Clinically stable3 bcefgkl o Recovered4 bce gkl o Recovered5 bc fgkl o Moribund at day 14 p.i.6 bcefgkl no Moribund at day 15 p.i.7 bc gkl no Moribund at day 17 p.i.
Cornell A75/17 8 b c e g k 1 n o Recovered9 b c e g k 1 n o Persistently infected10 b c g k l n o Recovered11 b c e g k 1 n o Clinically stable12 c e g 1 n Persistently infected13 c e g 1 n Clinically stable14 b c e g k l n o Moribund at day 30 p.i.15 b g I n o Moribund at day 42 p.i.16 b g 1 n o Moribund at day 32 p.i.17 b g 1 n o Recovered18 b g I n o Recovered19 b c f k m Moribund at day 22 p.i.
Ohio R252 20 c g 1 n Clinically stable21 c g 1 n Recovered22 c g 1 n Moribund at day 28 p.i.23 bc gkl n Recovered24 c g 1 n Moribund at day 31 p.i.25 c g 1 n Clinically stable
sa
0unx
uc0
EE
A
0
*
DAYS POST INFECTION
0
a
Days Post Infecion
FIG. 3. (A) ILMC response of seven dogs after intranasal exposure to CDV SH. Upper line, Mean valuesfrom four dogs (0, 0) that recovered; lower line, mean values from three dogs (O, *) that died between 14 and17 days p.i. with acute encephalitis. (B) Virus-neutralizing-antibody titers in the sera of seven dogs afterintranasal exposure to CDV SH. Upper line, Mean values from four dogs (0, 0) that recovered; lower line, meanvalues from three dogs (l, U) that died between 14 and 17 days p.i. with acute encephalitis.
ILMC elicited by virulent CDV, like thatelicited by the vaccine strain CDV Rockborn,was found to be genetically restricted (data notshown) and to decrease to insignificant levels at
times p.i. when CDV-neutralizing antibody waspresent at high levels (Fig. 1B, 2B, and 3B).Transient, genetically restricted cytotoxicitywas also reported in humans acutely infected
INFECT. IMMUN.
on June 6, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

CELLULAR IMMUNITY IN CANINE DISTEMPER 599
with measles virus (17). Unlike sera from vacci-nated dogs, sera from dogs exposed to virulentCDV has interferon activity (S. Tsai, B. A.Summers, and M. J. G. Appel, Arch. Virol., inpress). However, serum interferon activitypeaks early and, in contrast to ILMC, is negligi-ble by 2 weeks p.i. Thus, ILMC detected in theperipheral blood lymphocytes of dogs exposedto CDV is probably T-cell mediated. To our
knowledge, this and our previous publication(22) are the only reports of genetically restricted,antiviral T-cell-mediated cytotoxicity in dogs.ILMC responses in dogs after exposure to
subacute (A75/17 and R252) and acute (SH) viralbiotypes of CDV were assayed at various timesp.i. to gain further insight into the host-virusinteraction. Data presented in this paper clearlydemonstrate that the outcome of infection withCDV is determined by the ability of the host tomount a virus-specific immune response and thevirus strain.A strong correlation was found between sup-
pressed ILMC and fatal or persistent infection.Dogs that succumbed to CDV SH did not have a
measurable ILMC at any time. Similarly, dogsthat became moribund with non-inflammatoryencephalitis after CDV A75/17 or R252 infectionhad little or no ILMC (see Table 1). Dogs withinflammatory encephalitis had a delayed or
greatly reduced or both levels of ILMC in com-parison with those of recovering dogs (see Fig.1A). It appeared that CDV A75/17- or R252-infected dogs that had a strong ILMC response
by 21 days p.i. recovered, whereas dogs withlittle or no ILMC at that time subsequentlybecame moribund or remained persistently in-fected.The association of strong early ILMC and
neutralizing-antibody responses (Fig. 1, 2, and3) with the absence of morbidity support theimportance of virus-specific immunity in recov-ery from canine distemper. This is not surpris-ing, since cytotoxic T-cells have been shown tobe highly effective at limiting virus spread (1,27). Alternatively, in certain viral disease, e.g.,lymphocytic choriomeningitis in mice (9), virus-specific immunity is believed to be the cause ofpathological changes. It is similarly possible thatdemyelination associated with the inflammatoryencephalitis that occurred in dogs infected withCDV A75/17 and R252 was exacerbated byinfiltrating cytotoxic lymphocytes at a timewhen ILMC became positive. However, demy-elination was not dependent on lymphocyte infil-tration of the CNS, since four of eight dogs withdemyelination did not have inflammatory en-cephalitis. Moreover, Summers et al. (24) re-cently demonstrated that primary demyelinationin CDV-infected dogs was independent of lym-phocyte infiltration.
The basis for the variation in the immuneresponses of recovered and moribund or persis-tently infected dogs is presently unknown. Theability to respond immunologically to certainantigens and susceptibility to a variety of dis-eases has been linked in various species to themajor histocompatibility complex (MHC) (25).The susceptibility of murine retrovirus-infectedcells to T-cell-mediated cytolysis is affected bythe target cell haplotype (6, 21). Resistance ofchickens to Marek's disease is associated withcertain MHC haplotypes (7). On the other hand,we were unable to demonstrate a correlationbetween DLA and susceptibility to canine dis-temper. However, these results are inconclusivebecause the reagents used were polyspecific anddetected only serologically defined antigenic de-terminants (20). Furthermore, non-MHC geneticfactors might be more important in determiningthe outcome of infection with CDV. Gallatin andLongenecker (10) recently demonstrated thatgenetically determined non-MHC-linked resist-ance to Marek's disease was determined by thelevel of susceptibility of lymphocytes to viralinfection. Both B- and T-cells in lymphatic tis-sues and in buffy coat cells are infected duringthe early viremic phase in all CDV-infecteddogs. Infection of lymphocytes, lymphoid deple-tion, and lymphopenia result in immunosuppres-sion (14). The degree of immunosuppression, theability to mount a virus-specific immune re-sponse, and consequently, the clinical outcomecould in part be determined by the susceptibilityof lymphocytes or lymphocyte subpopulationsto CDV infection.A significant difference in onset, peak, and
duration of ILMC was observed in dogs thatwere infected with different strains of CDV. Theearliest onset and the shortest duration wereobserved in a previous study in dogs that wereimmunized with modified live CDV Rockbornstrain (22). In dogs that survived CDV SHinfection, the onset and peak occurred severaldays later than in vaccinated dogs, and theduration was approximately 2 weeks longer (Fig.3A). In contrast, dogs infected with either CDVA75/17 or R252 had a delayed onset and peakwith a gradual decline that lasted for more than 2months (Fig. 1A and 2A).The delayed ILMC in CDV A75/17- or R252-
infected dogs may be responsible for the persist-ent CNS infection that is known to occur. Thetime of onset of ILMC may be critical forclearance of virus from the CNS or even forpreventing infection of the CNS. A mild enceph-alitis with CDV invasion into the CNS is usuallyseen by 8 to 10 days p.i. (23). If CDV persists inthe canine CNS beyond days 14 to 21 p.i., thedog seems committed to develop gray- or white-matter disease. If, on the other hand, a prompt
VOL. 37, 1982
on June 6, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

600 APPEL, SHEK, AND SUMMERS
response develops at this period, then virusspread within the CNS is inhibited, the infectionis aborted, and the animal recovers with minimalresolving CNS lesions that support our conten-tion of early CNS infection (23). Dogs withinsignificant ILMC develop polioencephalitis af-ter CDV SH exposure and non-inflammatorydemyelinating encephalitis after CDV A75/17 orR252 exposure. Dogs with a delayed, low-levelILMC become persistently infected, clinicallystable and, if necropsied, show a pattern ofinflammatory demyelinating disease.
Virus isolation results were influenced by theimmune response. Isolation of infectious virusfrom brain suspensions in DLM cultures wasaccomplished from moribund dogs with acute ornon-inflammatory encephalitis and with little orno ILMC or virus-neutralizing antibody. Thistechnique failed for virus isolation from dogswith inflammatory encephalitis and with positiveILMC and virus-neutralizing antibody. Persist-ent virus was probably neutralized in brain sus-pensions from dogs with inflammatory encepha-litis. Once cells from these dogs were freed ofthe immune environment and grown in vitro,free infectious virus developed. Conversely,brain cell cultures from recovered dogs re-mained CDV negative.
ACKNOWLEDGMENTSThis work was supported by Public Health Service grant NS
14342 from the National Institutes of Health.The assistance of Mary Beth Metzgar and Ann Signore and
the DLA typing of dog leukocytes by Frances D. Cannon aregreatly appreciated.
LITERATURE CITED1. Ada, G. L., D. C. Jackson, R. V. Blanden, R. Tha Hla,
and N. A. Bowern. 1976. Changes in the surface of virusinfected cells recognized by cytotoxic T cells. I. Minimalrequirements for lysis of ectromelia-infected P-815 cells.Scand. J. Immunol. 5:23-30.
2. Appel, M. J. G. 1969. Pathogenesis of canine distemper.Am. J. Vet. Res. 30:1167-1182.
3. Appel, M. J. G., E. P. J. Gibbs, S. J. Martin, V. terMeulen, B. K. kima, J. R. Stephenson, and W. P. Taylor.1981. Morbillivirus diseases of animals and man p. 235-297. In E. Kurstak (ed.), Comparative diagnosis of viraldiseases, vol. IV: vertebrate animal and related viruses.Academic Press, Inc., New York.
4. Appel, M. J. G., and 0. R. Jones. 1967. Use of alveolarmacrophages for cultivation of canine distemper virus.Proc. Soc. Exp. Biol. Med. 126:571-574.
5. Appel, M. J. G., and D. S. Robson. 1973. A microneutrali-zation test for canine distemper virus. Am. J. Vet. Res.34:1459-1463.
6. Bubbers, J. E., S. Chen, and F. Lilly. 1978. Nonrandominclusion of H-2K and H-2D antigens in Friend virusparticles from mice of various strains. J. Exp. Med.147:340-351.
7. Calnek, B. W. 1980. Marek's disease virus and lympho-ma, p. 103-144. In S. Rapp (ed.), Oncogenic herpesvirus-es. CRC Press, Boca Raton, Fla.
8. Doherty, P. C., R. V. Blanden, and R. M. Zinkernagel.1976. Specificity of virus-immune effector T cells for H-2K or H-2D compatible interactions: implication for H-antigen diversity. Transplant. Rev. 29:89-124.
9. Doherty, P. C., and R. M. Zinkernagel. 1974. T-cell-medi-ated immunopathology in viral infections. Transplant.Rev. 19:89-120.
10. Gallatin, W. M., and B. M. Longenecker. 1979. Expres-sion of genetic resistance to an oncogenic herpesvirus atthe target cell level. Nature (London) 280:587-589.
11. Gerber, J. D., and A. E. Marron. 1976. Cell-mediatedimmunity and age at vaccination associated with measlesinoculation and protection of dogs against canine distem-per. Am. J. Vet. Res. 37:133-138.
12. Ho, C.-K., and L. A. Babiuk. 1979. Immune mechanismsagainst canine distemper. I. Identification of K cellagainst canine distemper virus infected target cells invitro. Immunology 37:231-239.
13. Krakowka, S., G. Cockerell, and A. Koestner. 1975.Effects of canine distemper virus infection on lymphoidfunction in vitro and in vivo. Infect. Immun. 11:1069-1078.
14. Krakowka, S., R. J. Higgins, and A. Koestner. 1980.Canine distemper virus: review of structural and function-al modulations in lymphoid tissues. Am. J. Vet. Res.41:284-292.
15. Krakowka, S., R. Olsen, A. Confer, A. Koestmer, and B.McCullough. 1975. Serologic response to canine distem-per viral antigens in gnotobiotic dogs infected with caninedistemper virus. J. Infect. Dis. 132:384-392.
16. Krakowka, S., and A. L. Wallace. 1979. Lymphocyte-associated immune responses to canine distemper andmeasles viruses in distemper-infected gnotobiotic dogs.Am. J. Vet. Res. 40:669-672.
17. Kreth, H. W., V. ter Meulen, and G. Eckert. 1979. Dem-onstration of HLA restricted killer cells in patients withacute measles. Med. Microbiol. Immunol. 165:203-214.
18. McCullough, B., S. Krakowka, and A. Koestner. 1974.Experimental canine distemper virus-induced demyelin-ation. Lab. Invest. 31:216-222.
19. McCullough, B., S. Krakowka, A. Koestner, and J. Shad-duck. 1974. Demyelinating activity of canine distempervirus isolate in gnotobiotic dogs. J. Infect. Dis. 130:343-350.
20. Mollen, N., D. St. John, F. D. Cannon, and J. W. Ferre-bee. 1968. Lymphocyte typing in allografted beagles.Transplantation 6:939.
21. Pfizeninaer, K., G. Trinchieri, D. Solter, and B. B.Knowles. 1978. Mapping of H-2 genes associated with T-cell-mediated cytotoxic responses to SV40-tumor-associ-ated specific antigens. Nature (London) 274:691-693.
22. Shek, W. R., R. D. Schultz, and M. J. G. Appel. 1980.Natural and immune cytolysis of canine distemper virus-infected target cells. Infect. Immun. 28:724-734.
23. Summers, B. A., H. A. Greisen, and M. J. G. Appel. 1978.Possible initiation of viral encephalomyelitis in dogs bymigrating lymphocytes infected with distemper virus.Lancet il:187-189.
24. Summers, B. A., H. A. Grelsen, and M. J. G. Appel. 1979.Early events in canine distemper demyelinating encepha-lomyelitis. Acta Neuropathol. 46:1-10.
25. Svejgaard, A., M. Hauge, C. Jersild, P. Platz, L. P. Ryder,L. Staub-Nielson, and M. Thomsen. 1979. The HLA sys-tem. Monogr. Hum. Genet. 7:66-77.
26. Welsh, R. M. 1981. Natural cell-mediated immunity dur-ing viral infections, p. 83-103. In 0. Haller (ed.), Currenttopics in microbiology and immunology. Natural resist-ance to tumors and viruses. Springer-Verlag, Berlin.
27. Zlnkernagel, R. M., and P. C. Doherty. 1977. Major trans-plantation antigens, viruses and specificity of surveillanceT cells. Contemp. Top. Immunobiol. 7:170-220.
INFECT. IMMUN.
on June 6, 2020 by guesthttp://iai.asm
.org/D
ownloaded from


















