
Le tissu conjonctif / soutien - epathologies.comepathologies.com/acad/intros/Tissus de...
22
Le tissu conjonctif / soutien Introduction Le tissu conjonctif est le terme traditionnellement appliqué à un type de base du tissu d'origine mésodermique qui fournit un support structural et métabolique pour d'autres tissus et organes dans le corps. En plus d'un rôle mécanique structurel, les tissus conjonctifs constitue un milieu de médiation de l'échange des nutriments, des métabolites et des produits de déchets entre les tissus et le système circulatoire. Les tissus conjonctifs contiennent généralement des vaisseaux sanguins et lymphatiques. «Tissu conjonctif» Le terme traditionnel donc guère rend justice à la vaste gamme de fonctions de ce type de tissu et il est maintenant sans doute plus approprié d'utiliser le terme tissu de soutien. Tous les tissus de soutien / conjonctifs sont composés d'une population de cellules de soutien spécialisés, dont certains produisent une matrice extracellulaire abondante. La matrice extracellulaire est le composant dominant de certains tissus de soutien qui détermine les propriétés physiques de chaque type. La matrice extracellulaire est constituée d'un gel dit substance fondamentale dans laquelle est intégrés une variété de fibres. Le tissu de soutien existe dans de nombreuses formes différentes avec diverses propriétés physiques. Dans la plupart des organes, du tissu lâche de support agi en tant que matériau d'emballage biologique entre les cellules et les tissus autres fonctions plus spécifiques. Les formes denses de tissus de soutien fournissent un soutien physique de consistance ferme dans le derme de la peau, et forment les capsules solides d'organes tels que le foie et la rate, et sont la source d'une grande résistance à la traction dans les ligaments et les tendons. LE cartilage et les os, les deux principales composantes du squelette, sont des formes hautement spécialisés de soutien du tissu qui sont considérés séparément lors de l’étude de l’appareil locomoteur. Certain tissu de soutien ont des fonctions métaboliques importantes telles que le stockage des graisses (tissus adipeux blanc) et la régulation de la température corporelle du nouveau- né (tissu adipeux brun). Les cellules du système immunitaire pénètrent dans les tissus de soutien où ils aident à la défense contre les microorganismes pathogènes. En réponse à une lésion tissulaire, les processus de réparation des tissus sont en grande partie fonction des tissus de soutien. Les cellules du tissu de soutien / conjonctif Les cellules du tissu de soutien sont dérivées de cellules précurseurs de tissus de soutien primitif (mésenchyme) et peut être divisé en plusieurs types, chacun avec différentes fonctions. Une fonction commune dominante est la synthèse et l'entretien du matériel de la matrice extracellulaire. • La cellule de soutien la plus courante est appelée fibroblaste qui est responsable de la sécrétion de la matrice extracellulaire dans la plupart des tissus. • Les chondrocytes et les ostéocytes sont responsables de la sécrétion de la matrice extracellulaire du cartilage et de l'os respectivement. • Les myofibroblastes ont une fonction contractile et un rôle dans la sécrétion de la matrice extracellulaire. • Un groupe de cellules de soutien hautement modifié sont responsables du stockage et du métabolisme des graisses. Ce sont les adipocytes qui forment collectivement le tissu adipeux. Les cellules ayant des fonctions de défense et immunitaires sont couramment rencontrées dans les tissus de soutien. Cela inclus les mastocytes, les macrophages tissulaires, tous les types de globules blancs et de cellules plasmatiques sécrétant les immunoglobulines. Certaines de ces cellules migrent dans les tissus de soutien et reste sur place, et exerce leur fonction localement. D'autres cellules immunitaires migrent à travers le tissu de soutien et sont en route pour remplir leur fonction ailleurs.
-
Upload
truongxuyen -
Category
Documents
-
view
225 -
download
0
Transcript of Le tissu conjonctif / soutien - epathologies.comepathologies.com/acad/intros/Tissus de...

Le tissu conjonctif / soutien Introduction Le tissu conjonctif est le terme traditionnellement appliqué à un type de base du tissu d'origine mésodermique qui fournit un support structural et métabolique pour d'autres tissus et organes dans le corps. En plus d'un rôle mécanique structurel, les tissus conjonctifs constitue un milieu de médiation de l'échange des nutriments, des métabolites et des produits de déchets entre les tissus et le système circulatoire. Les tissus conjonctifs contiennent généralement des vaisseaux sanguins et lymphatiques. «Tissu conjonctif» Le terme traditionnel donc guère rend justice à la vaste gamme de fonctions de ce type de tissu et il est maintenant sans doute plus approprié d'utiliser le terme tissu de soutien. Tous les tissus de soutien / conjonctifs sont composés d'une population de cellules de soutien spécialisés, dont certains produisent une matrice extracellulaire abondante. La matrice extracellulaire est le composant dominant de certains tissus de soutien qui détermine les propriétés physiques de chaque type. La matrice extracellulaire est constituée d'un gel dit substance fondamentale dans laquelle est intégrés une variété de fibres. Le tissu de soutien existe dans de nombreuses formes différentes avec diverses propriétés physiques. Dans la plupart des organes, du tissu lâche de support agi en tant que matériau d'emballage biologique entre les cellules et les tissus autres fonctions plus spécifiques. Les formes denses de tissus de soutien fournissent un soutien physique de consistance ferme dans le derme de la peau, et forment les capsules solides d'organes tels que le foie et la rate, et sont la source d'une grande résistance à la traction dans les ligaments et les tendons. LE cartilage et les os, les deux principales composantes du squelette, sont des formes hautement spécialisés de soutien du tissu qui sont considérés séparément lors de l’étude de l’appareil locomoteur. Certain tissu de soutien ont des fonctions métaboliques importantes telles que le stockage des graisses (tissus adipeux blanc) et la régulation de la température corporelle du nouveau-né (tissu adipeux brun). Les cellules du système immunitaire pénètrent dans les tissus de soutien où ils aident à la défense contre les microorganismes pathogènes. En réponse à une lésion tissulaire, les processus de réparation des tissus sont en grande partie fonction des tissus de soutien. Les cellules du tissu de soutien / conjonctif Les cellules du tissu de soutien sont dérivées de cellules précurseurs de tissus de soutien primitif (mésenchyme) et peut être divisé en plusieurs types, chacun avec différentes fonctions. Une fonction commune dominante est la synthèse et l'entretien du matériel de la matrice extracellulaire. • La cellule de soutien la plus courante est appelée fibroblaste qui est responsable de la sécrétion de
la matrice extracellulaire dans la plupart des tissus. • Les chondrocytes et les ostéocytes sont responsables de la sécrétion de la matrice extracellulaire
du cartilage et de l'os respectivement. • Les myofibroblastes ont une fonction contractile et un rôle dans la sécrétion de la matrice
extracellulaire. • Un groupe de cellules de soutien hautement modifié sont responsables du stockage et du
métabolisme des graisses. Ce sont les adipocytes qui forment collectivement le tissu adipeux. Les cellules ayant des fonctions de défense et immunitaires sont couramment rencontrées dans les tissus de soutien. Cela inclus les mastocytes, les macrophages tissulaires, tous les types de globules blancs et de cellules plasmatiques sécrétant les immunoglobulines. Certaines de ces cellules migrent dans les tissus de soutien et reste sur place, et exerce leur fonction localement. D'autres cellules immunitaires migrent à travers le tissu de soutien et sont en route pour remplir leur fonction ailleurs.
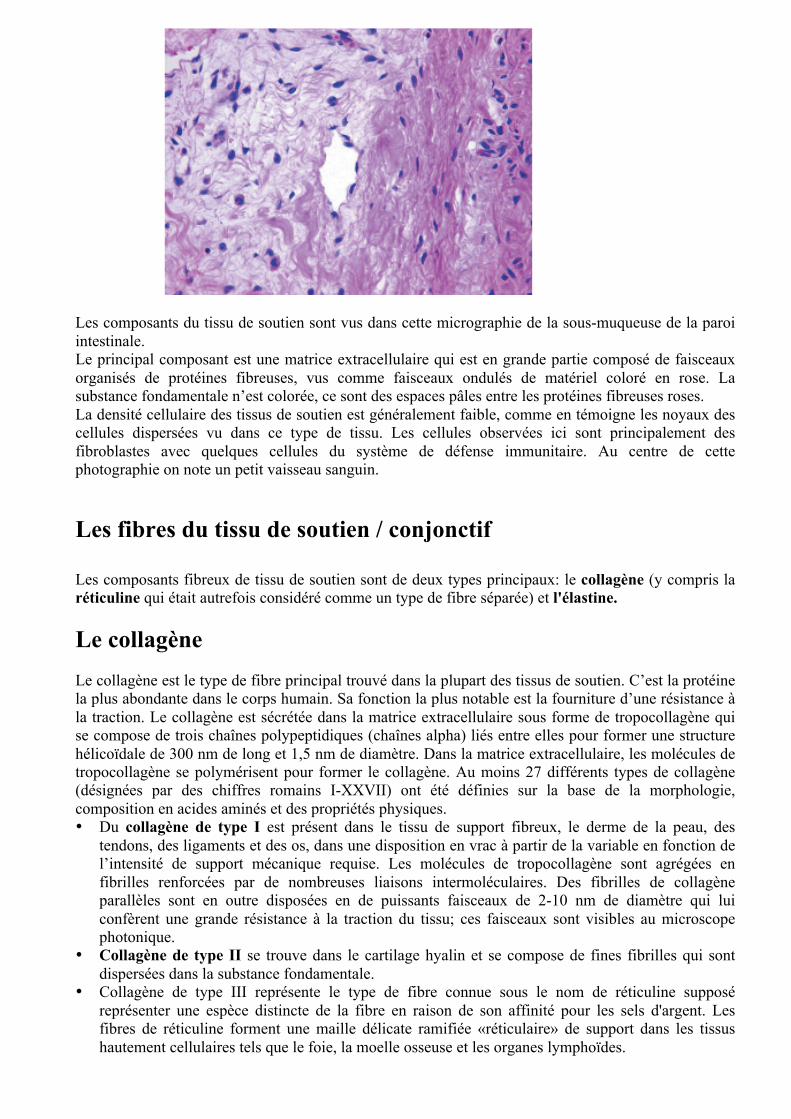
Les composants du tissu de soutien sont vus dans cette micrographie de la sous-muqueuse de la paroi intestinale. Le principal composant est une matrice extracellulaire qui est en grande partie composé de faisceaux organisés de protéines fibreuses, vus comme faisceaux ondulés de matériel coloré en rose. La substance fondamentale n’est colorée, ce sont des espaces pâles entre les protéines fibreuses roses. La densité cellulaire des tissus de soutien est généralement faible, comme en témoigne les noyaux des cellules dispersées vu dans ce type de tissu. Les cellules observées ici sont principalement des fibroblastes avec quelques cellules du système de défense immunitaire. Au centre de cette photographie on note un petit vaisseau sanguin. Les fibres du tissu de soutien / conjonctif Les composants fibreux de tissu de soutien sont de deux types principaux: le collagène (y compris la réticuline qui était autrefois considéré comme un type de fibre séparée) et l'élastine. Le collagène Le collagène est le type de fibre principal trouvé dans la plupart des tissus de soutien. C’est la protéine la plus abondante dans le corps humain. Sa fonction la plus notable est la fourniture d’une résistance à la traction. Le collagène est sécrétée dans la matrice extracellulaire sous forme de tropocollagène qui se compose de trois chaînes polypeptidiques (chaînes alpha) liés entre elles pour former une structure hélicoïdale de 300 nm de long et 1,5 nm de diamètre. Dans la matrice extracellulaire, les molécules de tropocollagène se polymérisent pour former le collagène. Au moins 27 différents types de collagène (désignées par des chiffres romains I-XXVII) ont été définies sur la base de la morphologie, composition en acides aminés et des propriétés physiques. • Du collagène de type I est présent dans le tissu de support fibreux, le derme de la peau, des
tendons, des ligaments et des os, dans une disposition en vrac à partir de la variable en fonction de l’intensité de support mécanique requise. Les molécules de tropocollagène sont agrégées en fibrilles renforcées par de nombreuses liaisons intermoléculaires. Des fibrilles de collagène parallèles sont en outre disposées en de puissants faisceaux de 2-10 nm de diamètre qui lui confèrent une grande résistance à la traction du tissu; ces faisceaux sont visibles au microscope photonique.
• Collagène de type II se trouve dans le cartilage hyalin et se compose de fines fibrilles qui sont dispersées dans la substance fondamentale.
• Collagène de type III représente le type de fibre connue sous le nom de réticuline supposé représenter une espèce distincte de la fibre en raison de son affinité pour les sels d'argent. Les fibres de réticuline forment une maille délicate ramifiée «réticulaire» de support dans les tissus hautement cellulaires tels que le foie, la moelle osseuse et les organes lymphoïdes.
• Collagène de type IV ne forme pas de fibrilles, mais plutôt un réseau en maille et est un constituant important des membranes basales.
• Collagène de type VII forme des fibrilles d'ancrage qui pointent vers la membrane basale.
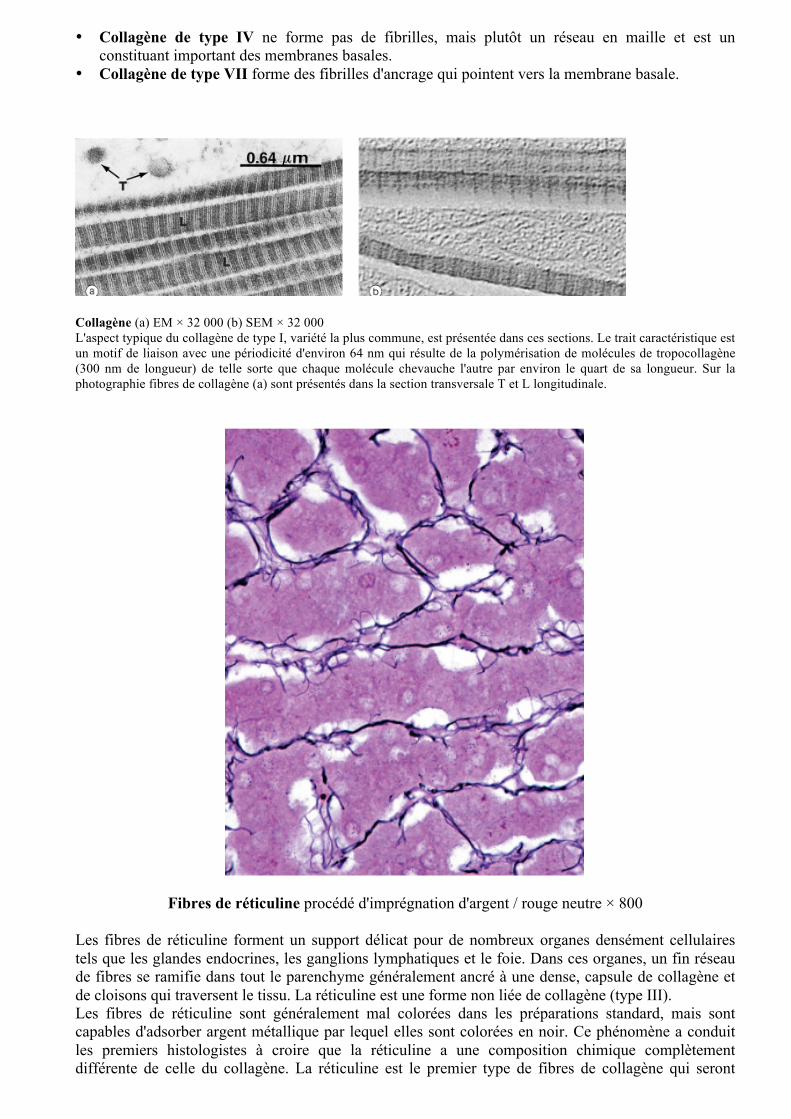
Collagène (a) EM × 32 000 (b) SEM × 32 000 L'aspect typique du collagène de type I, variété la plus commune, est présentée dans ces sections. Le trait caractéristique est un motif de liaison avec une périodicité d'environ 64 nm qui résulte de la polymérisation de molécules de tropocollagène (300 nm de longueur) de telle sorte que chaque molécule chevauche l'autre par environ le quart de sa longueur. Sur la photographie fibres de collagène (a) sont présentés dans la section transversale T et L longitudinale.
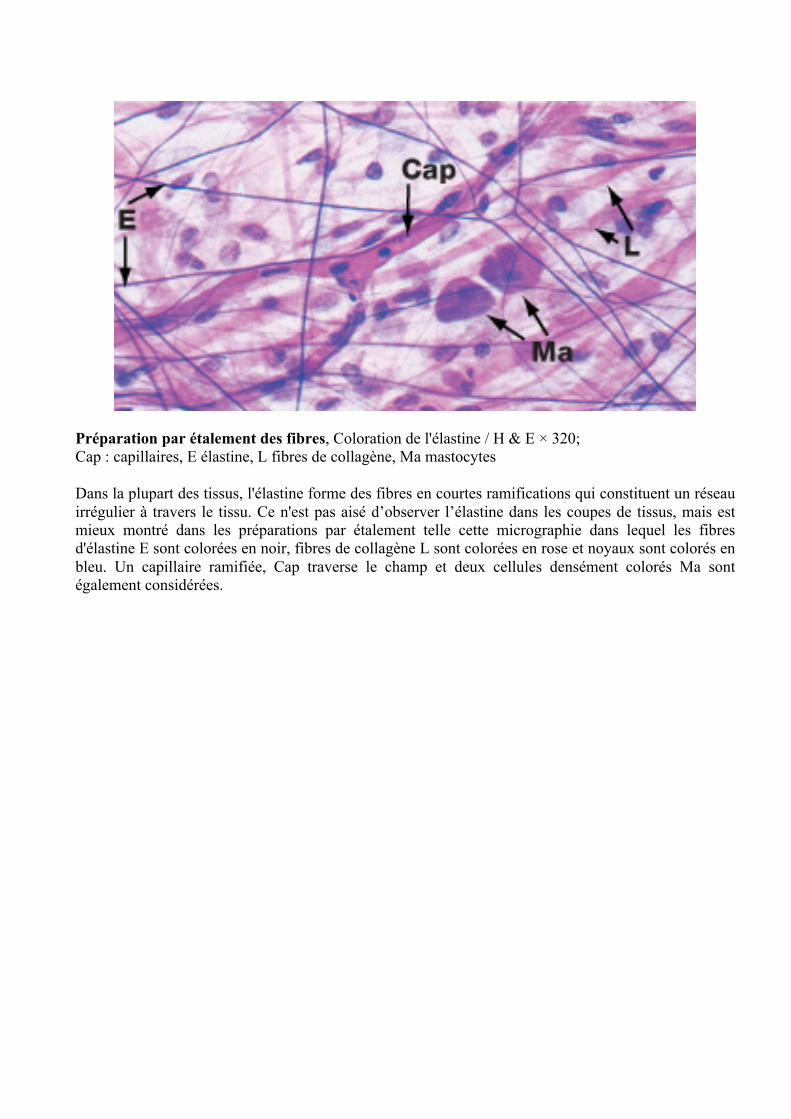
Fibres de réticuline procédé d'imprégnation d'argent / rouge neutre × 800
Les fibres de réticuline forment un support délicat pour de nombreux organes densément cellulaires tels que les glandes endocrines, les ganglions lymphatiques et le foie. Dans ces organes, un fin réseau de fibres se ramifie dans tout le parenchyme généralement ancré à une dense, capsule de collagène et de cloisons qui traversent le tissu. La réticuline est une forme non liée de collagène (type III). Les fibres de réticuline sont généralement mal colorées dans les préparations standard, mais sont capables d'adsorber argent métallique par lequel elles sont colorées en noir. Ce phénomène a conduit les premiers histologistes à croire que la réticuline a une composition chimique complètement différente de celle du collagène. La réticuline est le premier type de fibres de collagène qui seront

produites durant le développement de tous les tissus de soutien et est également présent en quantités variables dans la plupart des tissus de soutien matures. Cette micrographie montre l'échafaudage réticulinique fin dans le foie, le réseau offre un soutien pour les hépatocytes (les lames violettes de cellules), et les sinusoïdes par lequel le sang s'écoule.
Elastine L'élastine est une protéine structurale importante qui est agencée sous forme de fibres et / ou de feuilles discontinues de la matrice extracellulaire en particulier de la peau, du poumon et du sang qui confère les propriétés d'étirement et de rappel élastique. L'élastine est synthétisée par les fibroblastes dans une forme de précurseur connue sous le nom tropoélastine qui subit une polymérisation dans les tissus extracellulaires. Le dépôt d'élastine dans la forme de fibres nécessite la présence de microfibrilles d’une glycoprotéine de structure, la fibrilline qui s'incorporent autour et à l'intérieur des fibres élastiques.
Élastine EM × 50 000
Cette micrographie montre l’élastine E dans le tissu délicat de support de l'épithélium de la trachée de souris. Le champ contient aussi fibrilles de collagène T (coupé en coupe transversale) et les extensions cytoplasmiques fines des fibroblastes F responsables de l'élaboration des constituants extracellulaires. L'élastine n'est pas constituée de fibrilles mais plutôt une masse amorphe polymérisé de tropoélastine. Les microfibrilles de fibrilline M, la glycoprotéine structurelle, qui est impliqué dans le processus de dépôt d'élastine, peuvent être discerné à ce grossissement (en coupe transversale) située à l'intérieur et autour de l'élastine. Les microfibrilles sont également visibles dans la partie inférieure de la coupe en section longitudinale ML, en association avec de petites quantités d'élastine.
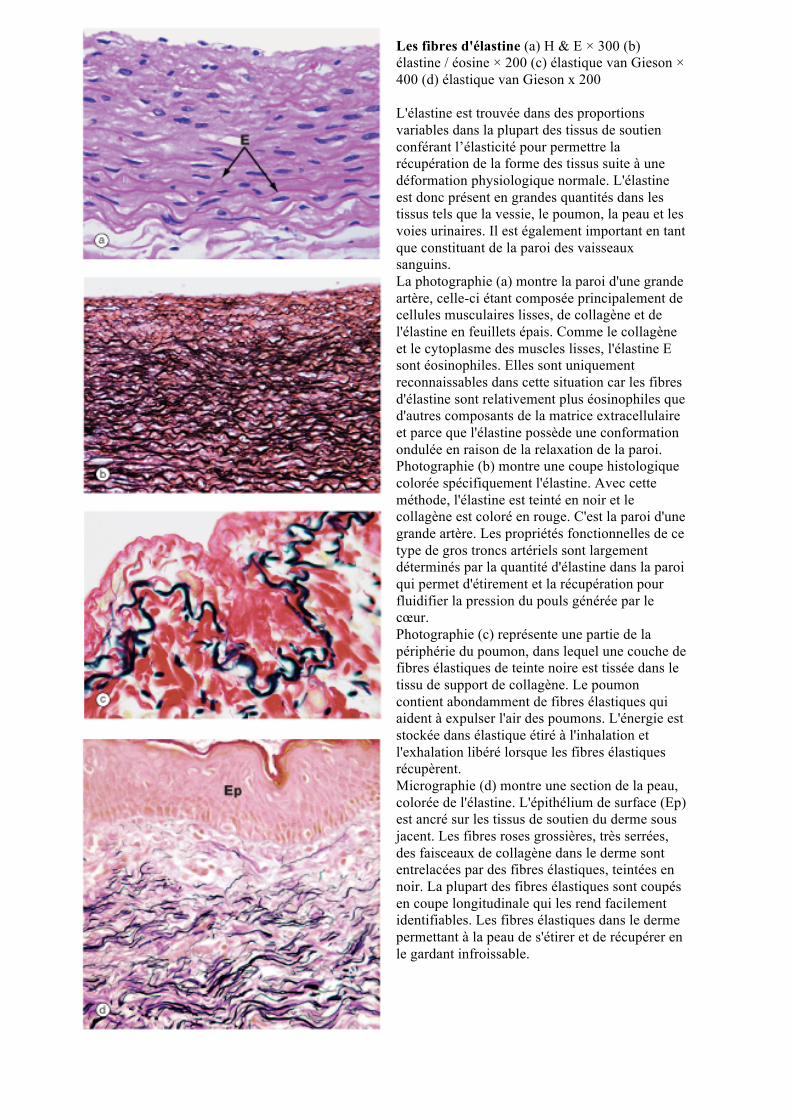
Préparation par étalement des fibres, Coloration de l'élastine / H & E × 320; Cap : capillaires, E élastine, L fibres de collagène, Ma mastocytes Dans la plupart des tissus, l'élastine forme des fibres en courtes ramifications qui constituent un réseau irrégulier à travers le tissu. Ce n'est pas aisé d’observer l’élastine dans les coupes de tissus, mais est mieux montré dans les préparations par étalement telle cette micrographie dans lequel les fibres d'élastine E sont colorées en noir, fibres de collagène L sont colorées en rose et noyaux sont colorés en bleu. Un capillaire ramifiée, Cap traverse le champ et deux cellules densément colorés Ma sont également considérées.
Les fibres d'élastine (a) H & E × 300 (b) élastine / éosine × 200 (c) élastique van Gieson × 400 (d) élastique van Gieson x 200 L'élastine est trouvée dans des proportions variables dans la plupart des tissus de soutien conférant l’élasticité pour permettre la récupération de la forme des tissus suite à une déformation physiologique normale. L'élastine est donc présent en grandes quantités dans les tissus tels que la vessie, le poumon, la peau et les voies urinaires. Il est également important en tant que constituant de la paroi des vaisseaux sanguins. La photographie (a) montre la paroi d'une grande artère, celle-ci étant composée principalement de cellules musculaires lisses, de collagène et de l'élastine en feuillets épais. Comme le collagène et le cytoplasme des muscles lisses, l'élastine E sont éosinophiles. Elles sont uniquement reconnaissables dans cette situation car les fibres d'élastine sont relativement plus éosinophiles que d'autres composants de la matrice extracellulaire et parce que l'élastine possède une conformation ondulée en raison de la relaxation de la paroi. Photographie (b) montre une coupe histologique colorée spécifiquement l'élastine. Avec cette méthode, l'élastine est teinté en noir et le collagène est coloré en rouge. C'est la paroi d'une grande artère. Les propriétés fonctionnelles de ce type de gros troncs artériels sont largement déterminés par la quantité d'élastine dans la paroi qui permet d'étirement et la récupération pour fluidifier la pression du pouls générée par le cœur. Photographie (c) représente une partie de la périphérie du poumon, dans lequel une couche de fibres élastiques de teinte noire est tissée dans le tissu de support de collagène. Le poumon contient abondamment de fibres élastiques qui aident à expulser l'air des poumons. L'énergie est stockée dans élastique étiré à l'inhalation et l'exhalation libéré lorsque les fibres élastiques récupèrent. Micrographie (d) montre une section de la peau, colorée de l'élastine. L'épithélium de surface (Ep) est ancré sur les tissus de soutien du derme sous jacent. Les fibres roses grossières, très serrées, des faisceaux de collagène dans le derme sont entrelacées par des fibres élastiques, teintées en noir. La plupart des fibres élastiques sont coupés en coupe longitudinale qui les rend facilement identifiables. Les fibres élastiques dans le derme permettant à la peau de s'étirer et de récupérer en le gardant infroissable.

Substance fondamentale La substance fondamentale tire son nom du fait qu’elle est constituée d'un matériau amorphe transparent qui présente les propriétés d'un gel semi-fluide. Les fluides tissulaires sont plus ou moins associés à une substance fondamentale, en formant ainsi le moyen de passage de molécules à travers les tissus de support et pour l'échange des métabolites avec le système circulatoire. La substance fondamentale est constitué d'un mélange de longues chaînes polysaccharidiques linéaires de sept types différents, chacun composé d'unités répétitives de disaccharides. L'une des unités disaccharidiques est généralement un acide uronique et l'autre un sucre aminé (soit Nacetyl glucosamine ou la galactosamine Nacetyl) donnant ainsi naissance à des glycosaminoglycanes (GAG dénomination actuelle), qui ont été anciennement appelé mucopolysaccharides. Les glycosaminoglycanes sont acides (chargé négativement) en raison de la présence de groupes hydroxyle, carboxyle latéraux et de sulfate sur les unités disaccharidiques. L'acide hyaluronique est le GAG prédominant dans les tissus lâches de support et est le seul des groupes latéraux sans sulfate, les autres GAG (chondroitin4 sulfate, chondroitin6 sulfate, le dermatane sulfate, l'héparane sulfate, l'héparine sulfate et le kératane sulfate) différent de l'acide hyaluronique en ce qu'ils sont de manière covalente lié à une variété de molécules de protéines pour former des protéoglycanes (anciennement connu sous le nom mucoprotéines); ces protéoglycanes sont des molécules volumineuses composées à 90-95% de glucides. En outre, les protéoglycanes peuvent former des liaisons non covalentes avec des chaînes d'acide hyaluronique pour former encore des molécules plus complexes. Contrairement à de nombreuses protéines, les molécules GAG sont pas assez souples pour former des agrégats globulaires, mais restent sous une forme expansive, occupant ainsi un énorme volume, de masse relativement faible. En outre, leurs groupes latéraux fortement chargées les rendent extrêmement hydrophile, attirant ainsi un grand volume d'eau et d'ions positifs, en particulier de sodium, ce qui constitue le fluide extracellulaire. Le liquide extracellulaire donne la turgescence caractéristique des tissus de soutien. En résumé, la substance fondamentale est essentiellement composée de glycosaminoglycanes sous la forme de l'acide hyaluronique et de protéoglycanes. Ces molécules sont volumineuses et enchevêtrés, électrostatiquement lié l’une à l’autre, et par leur hydratation, forment un gel flexible à travers laquelle peuvent diffuser les métabolites. La taille des espaces entre les molécules GAG et la nature des changements électrostatiques déterminent les caractères de perméabilité des tissus de soutien. Ce fait est d'une importance particulière dans la structure des membranes basales. Les propriétés mécaniques de la substance fondamentale sont renforcées par les protéines fibreuses du tissu extracellulaire, auxquelles les composants de la substance fondamentale sont également liés. Les glycoprotéines de structure Les glycoprotéines de structure sont un groupe de molécules composées principalement de chaînes de protéines liées à des polysaccharides ramifiés; Il reste encore beaucoup à apprendre de leur rôle dans la fonction de matériel extracellulaire. Les glycoprotéines de structure comprennent deux molécules formatrice de fibrille : la fibronectine, et la fibrilline, ainsi qu'un certain nombre de protéines non filamenteuses comprenant la laminine, l'entactine et la ténascine qui fonctionnent comme des liens entre les cellules et la matrice extracellulaire. La fibrilline forme des microfibrilles de 8-12 nm de diamètre qui, dans certaines situations particulières, par exemple le mésangium du rein, semblent améliorer l'adhérence avec les autres constituants extracellulaires. La fibrilline est un constituant des fibres élastiques, où il semble jouer un rôle dans le dépôt ordonné des fibres.

La fibronectine joue un rôle dans le contrôle de l'orientation et le dépôt de collagène dans la matrice extracellulaire, et la liaison de la cellule à la matière extracellulaire. Les membranes cellulaires incorporent un groupe de complexes protéiques transmembranaires appelés intégrines qui agissent comme des molécules d'adhésion cellulaire. L'un d'eux, le récepteur de la fibronectine, crée des liens au sein de la cellule aux filaments d'actine du cytosquelette à la fibronectine extérieure. La fibronectine, à son tour se lie au collagène et à la glycosaminoglycane, la héparine sulfate, établissant ainsi une continuité structurelle entre le cytosquelette et la matrice extracellulaire. La laminine est un composant majeur des membranes basales. Elle se lie à certaines molécules d'adhésion cellulaire de manière à former des liens entre les membranes cellulaires et d'autres constituants de la membrane basale. L’entactine, une autre protéine non fibrillaire, a pour fonction, la liaison de la laminine au collagène de type IV de la membrane basale. La ténascine se lie également aux intégrines, et est importante dans l'embryon, où elle semble être impliqué dans le contrôle de la croissance des cellules nerveuses. Les membranes basales Les membranes basales sont des arrangements lamellaires de protéines de la matrice extracellulaire qui agissent comme une interface entre les tissus de soutien et les cellules parenchymateuses. Ces membranes basales sont associés à des cellules épithéliales et musculaires, et forment aussi une membrane de séparation dans le système nerveux central. Le terme provient du fait que les membranes basales premières à être reconnus étaient celles situées sous les cellules basales de l'épithélium de surface. Pour les tissus musculaires et nerveux, la dénomination de lamina externa peut également être appliqué. Les épithéliums sont composés en particulier de cellules étroitement entourées de matériau intercellulaire minime entre eux. La membrane basale fournit un soutien métabolique ainsi que la liaison de l'épithélium au tissu de soutien sous-jacent. La membrane basale est également impliquée dans le contrôle de la croissance et de la différenciation épithéliale, formant une barrière à la croissance épithéliale vers le bas. Elle est rompue si l'épithélium subi une transformation maligne. L’épithélium est dépourvu de vaisseaux sanguins. La membrane basale doit donc permettre le passage des nutriments, des métabolites et autres molécules de, et vers l'épithélium. Lorsque l'épithélium agit comme une barrière sélective pour le passage des molécules d'un compartiment à un autre (par exemple entre la lumière des vaisseaux sanguins et les tissus environnants), la membrane basale joue un rôle critique dans la régulation de la perméabilité. Celle-ci atteint un extrême de sophistication au niveau du rein, où la membrane basale glomérulaire fait partie des filtres très sélectifs pour les molécules passant du sang dans l'urine. Les principaux constituants des membranes basales et des lamina externa sont le glycosaminoglycane héparane sulfate, la fibre de protéine de collagène de type IV, la fibronectine et les glycoprotéines de structure, la laminine et l’entactine. La fibronectine semble être produite par les fibroblastes du tissu de soutien, mais les autres sont au moins en partie, voire exclusivement, élaboré par les tissus soutenus. Au microscope électronique, la membrane basale est considéré sont constitués de trois couches. Une couche relativement électron claire, la lamina lucida (allant de 10 à 50 nm de largeur), vient en butée contre la membrane des cellules basales du tissu parenchymateux. La couche intermédiaire est électron dense et est donc connue comme la lamina densa; selon le tissu, cela varie de 20 à 300 nm d'épaisseur. Au-delà de la lamina densa est une vaste couche relativement électron claire connu sous le nom de lame fibroreticularis qui se confond avec le tissu de soutien sous-jacent. La structure de la lamina lucida et la lamina densa est une maille fine de collagène de type IV qui est exclusif aux membranes basales. Un autre constituant majeur de ces couches est la laminine qui se lie au collagène de type IV et aux autres constituants de la membrane basale, et au récepteur de laminine des membranes cellulaires basales parenchymateuses. L’entactine intervient dans la liaison de la laminine au collagène de type IV. La couche fibroreticularis représente probablement une condensation du tissu de soutien sous-jacent. Sa teneur en collagène est principalement de type III (réticuline) qui, par l'intermédiaire de la glycoprotéine fibrillaire, la fibronectine, est également liée aux intégrines de la membrane cellulaire basale du parenchyme.
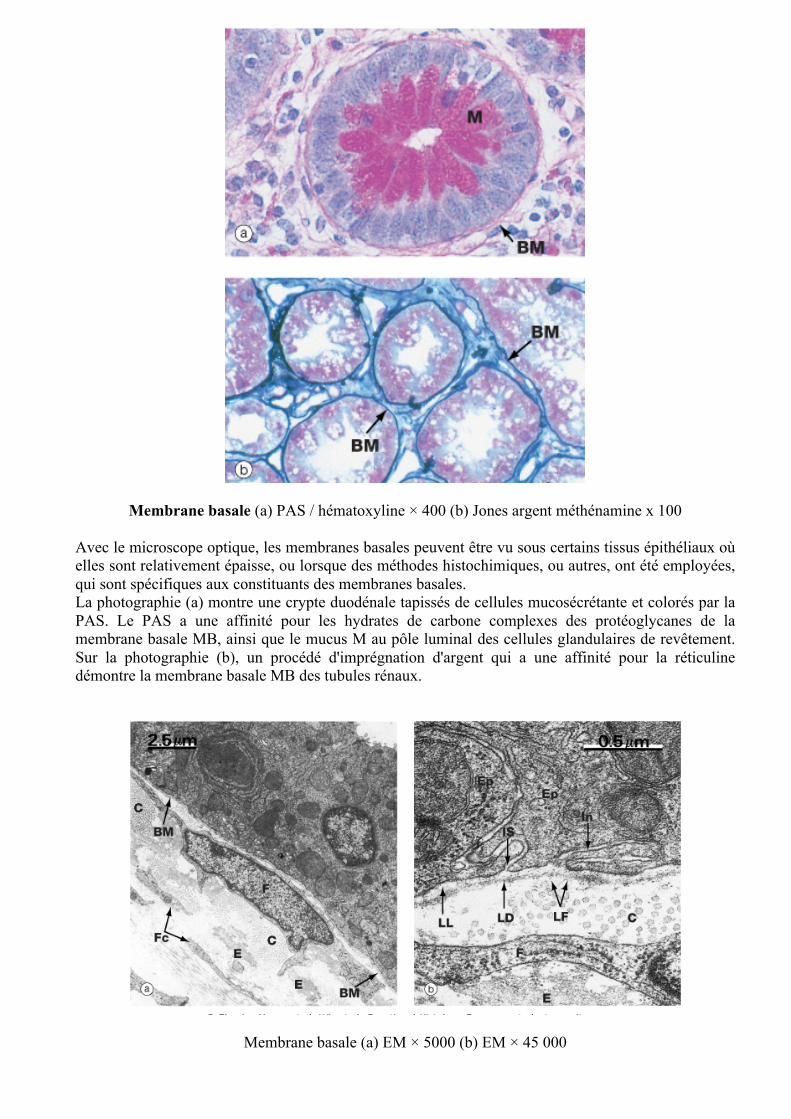
Membrane basale (a) PAS / hématoxyline × 400 (b) Jones argent méthénamine x 100 Avec le microscope optique, les membranes basales peuvent être vu sous certains tissus épithéliaux où elles sont relativement épaisse, ou lorsque des méthodes histochimiques, ou autres, ont été employées, qui sont spécifiques aux constituants des membranes basales. La photographie (a) montre une crypte duodénale tapissés de cellules mucosécrétante et colorés par la PAS. Le PAS a une affinité pour les hydrates de carbone complexes des protéoglycanes de la membrane basale MB, ainsi que le mucus M au pôle luminal des cellules glandulaires de revêtement. Sur la photographie (b), un procédé d'imprégnation d'argent qui a une affinité pour la réticuline démontre la membrane basale MB des tubules rénaux.
Membrane basale (a) EM × 5000 (b) EM × 45 000

La photographie (a) montre l'épithélium de la trachée de souris. Une membrane basale MB peut être clairement vu le séparant du tissu de soutien sous-jacent qui contient le corps cellulaire et le noyau d'un fibroblaste F, de nombreuses fines expansions cytoplasmiques fibroblastiques Fc, des faisceaux de fibrilles de collagène C (en coupe transversale) et les fibres d'élastine E. A plus fort grossissement, sur la photographie (b), les trois couches de la membrane basale peuvent être vues. La relative électron claire lamina lucida LL vient buter contre la membrane cellulaire basale épithéliale. La couche intermédiaire est électron dense et représente la lamina densa LD. Au-delà de la lamina densa est la large lame relativement électron claire fibroreticularis LF qui se confond avec le tissu fibreux et fibrillaire (réticulaire) des composants du tissu de soutien sous-jacent. Dans ce champ, on note les fibrilles de collagène C, une partie d’un fibroblaste F et d'élastine E. La membrane basale passe généralement sans interruption sous l'espace intercellulaire IS, entre deux cellules épithéliales Ep, et sous une invagination basale de l'une de ces cellules. La lamina densa a été autrefois connu comme la lame basale. Les termes, lame basale et membrane basale étaient souvent utilisés de façon interchangeable jusqu'à ce qu'il soit rendu compte que les trois couches observées au microscope électronique représente la seule couche observée avec le microscope optique. Cela a conduit à une confusion terminologique considérable et, le cas échéant, le terme de lame basale devrait être limité à la lamina densa. Troubles des membranes basales Les membranes basales sont impliquées dans des processus pathologiques. La fonction rénale Dans le rein, les membranes basales fusionnées des cellules endothéliales et les podocytes forme la barrière de filtration pour l'ultrafiltrat dans le glomérule. Si la membrane basale glomérulaire devient anormale alors la fonction rénale est altérée. Chez les patients atteints de diabète sucré il y a épaississement de la membrane basale glomérulaire qui devient anormalement perméable à de petites protéines. Cancer Les tissus épithéliaux prolifère, régénère et sont ancrés, mais séparée du tissu de soutien par une membrane basale. Les changements génétiques dans les cellules épithéliales entraînent une croissance anormale (néoplasie) formant le cancer. Une tumeur maligne se développe dans un organe et se propage dans les tissus locaux (invasion). L‘invasion est la conséquence de la sécrétion de facteurs par les cellules cancéreuses qui facilitent la destruction de la membrane basale, permettant aux cellules de se développer dans la matrice extracellulaire. Les maladies génétiques Des mutations dans les gènes codant pour les composants de la membrane basale et avérés de la maladie rénale (maladie d'Alport), maladie du muscle (dystrophie musculaire congénitale), et la maladie de la peau (épidermolyse bulleuse jonctionnelle). Maladie auto-immune Dans l'état appelé syndrome de Goodpasture, des auto-anticorps sont produits à un composant de la membrane basale commune aux glomérules et du poumon, rénal et conduisant à une maladie pulmonaire.
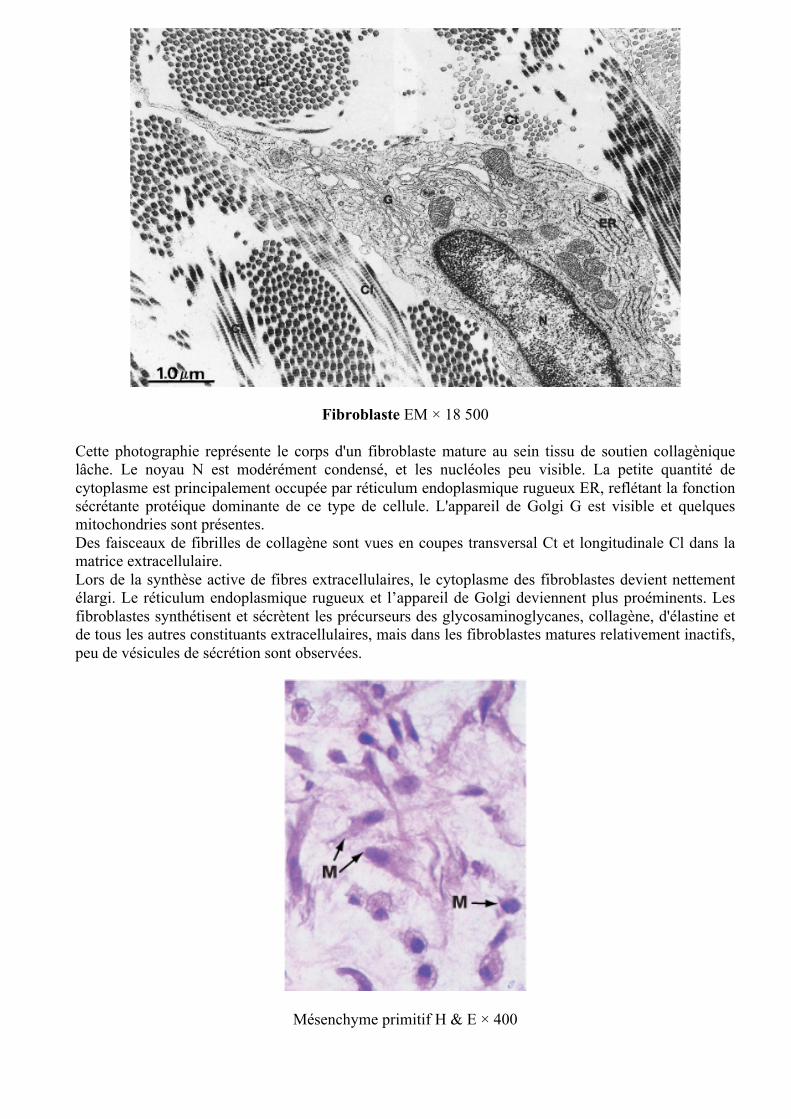
Fibroblaste EM × 18 500
Cette photographie représente le corps d'un fibroblaste mature au sein tissu de soutien collagènique lâche. Le noyau N est modérément condensé, et les nucléoles peu visible. La petite quantité de cytoplasme est principalement occupée par réticulum endoplasmique rugueux ER, reflétant la fonction sécrétante protéique dominante de ce type de cellule. L'appareil de Golgi G est visible et quelques mitochondries sont présentes. Des faisceaux de fibrilles de collagène sont vues en coupes transversal Ct et longitudinale Cl dans la matrice extracellulaire. Lors de la synthèse active de fibres extracellulaires, le cytoplasme des fibroblastes devient nettement élargi. Le réticulum endoplasmique rugueux et l’appareil de Golgi deviennent plus proéminents. Les fibroblastes synthétisent et sécrètent les précurseurs des glycosaminoglycanes, collagène, d'élastine et de tous les autres constituants extracellulaires, mais dans les fibroblastes matures relativement inactifs, peu de vésicules de sécrétion sont observées.
Mésenchyme primitif H & E × 400
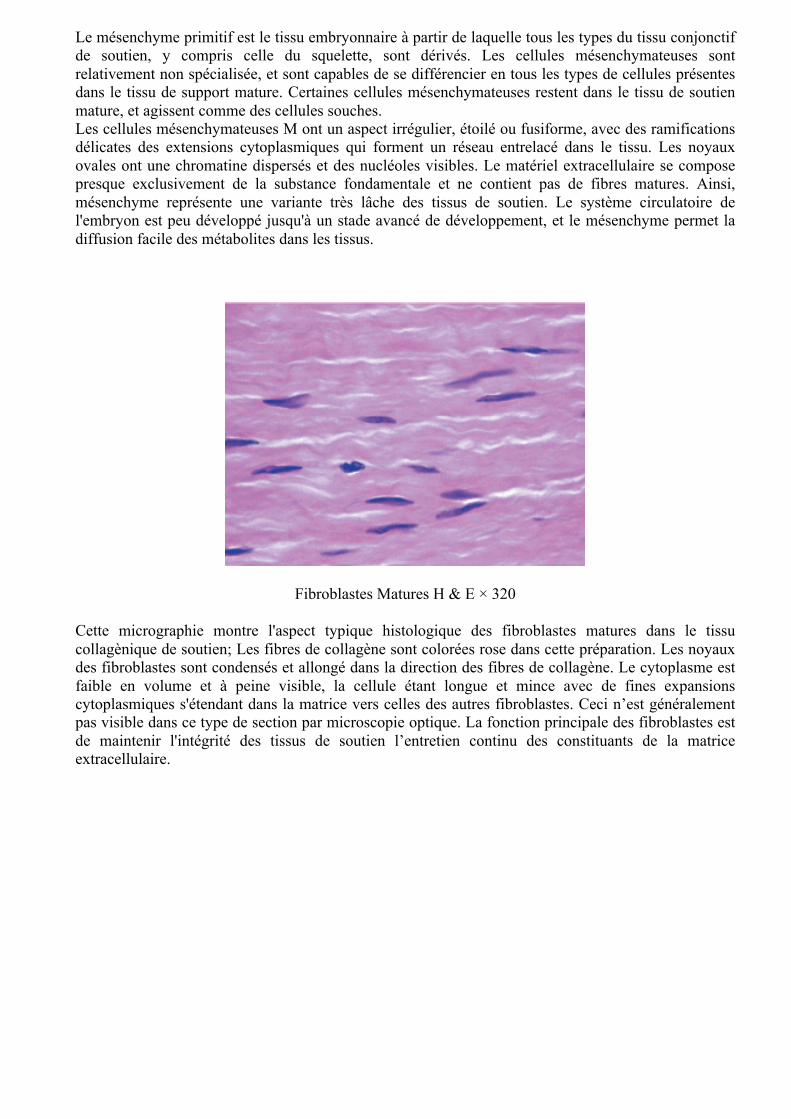
Le mésenchyme primitif est le tissu embryonnaire à partir de laquelle tous les types du tissu conjonctif de soutien, y compris celle du squelette, sont dérivés. Les cellules mésenchymateuses sont relativement non spécialisée, et sont capables de se différencier en tous les types de cellules présentes dans le tissu de support mature. Certaines cellules mésenchymateuses restent dans le tissu de soutien mature, et agissent comme des cellules souches. Les cellules mésenchymateuses M ont un aspect irrégulier, étoilé ou fusiforme, avec des ramifications délicates des extensions cytoplasmiques qui forment un réseau entrelacé dans le tissu. Les noyaux ovales ont une chromatine dispersés et des nucléoles visibles. Le matériel extracellulaire se compose presque exclusivement de la substance fondamentale et ne contient pas de fibres matures. Ainsi, mésenchyme représente une variante très lâche des tissus de soutien. Le système circulatoire de l'embryon est peu développé jusqu'à un stade avancé de développement, et le mésenchyme permet la diffusion facile des métabolites dans les tissus.
Fibroblastes Matures H & E × 320
Cette micrographie montre l'aspect typique histologique des fibroblastes matures dans le tissu collagènique de soutien; Les fibres de collagène sont colorées rose dans cette préparation. Les noyaux des fibroblastes sont condensés et allongé dans la direction des fibres de collagène. Le cytoplasme est faible en volume et à peine visible, la cellule étant longue et mince avec de fines expansions cytoplasmiques s'étendant dans la matrice vers celles des autres fibroblastes. Ceci n’est généralement pas visible dans ce type de section par microscopie optique. La fonction principale des fibroblastes est de maintenir l'intégrité des tissus de soutien l’entretien continu des constituants de la matrice extracellulaire.
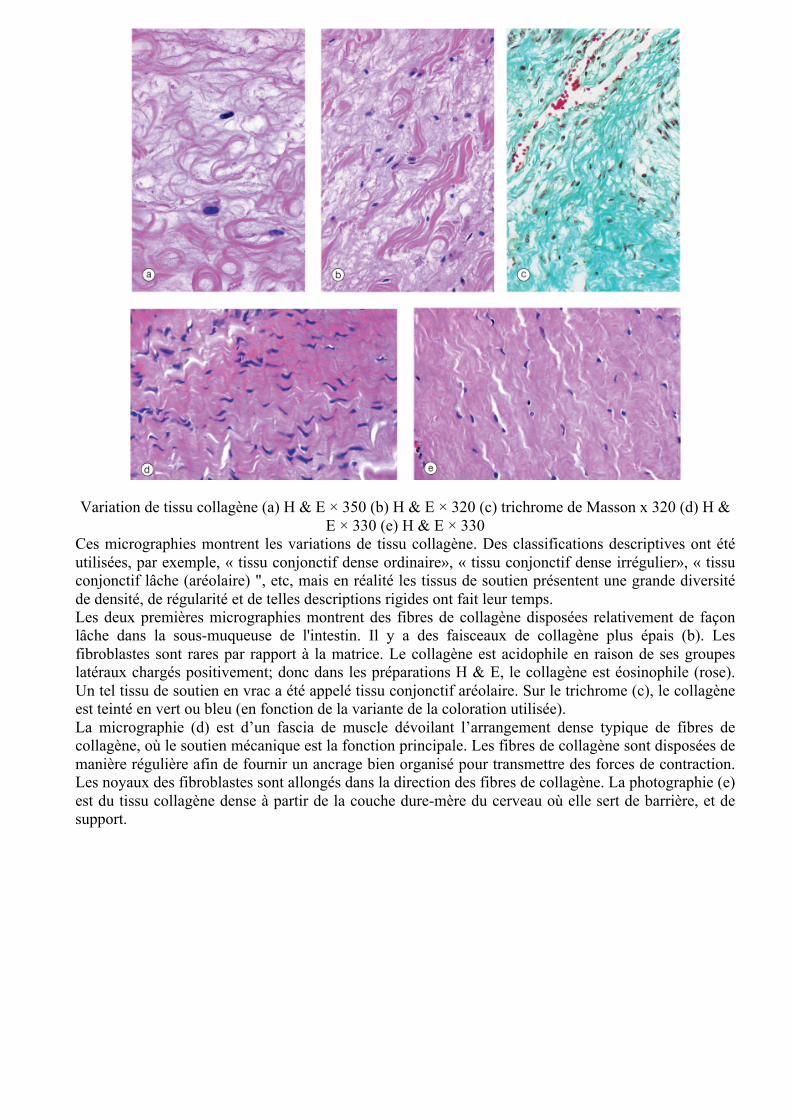
Variation de tissu collagène (a) H & E × 350 (b) H & E × 320 (c) trichrome de Masson x 320 (d) H &
E × 330 (e) H & E × 330 Ces micrographies montrent les variations de tissu collagène. Des classifications descriptives ont été utilisées, par exemple, « tissu conjonctif dense ordinaire», « tissu conjonctif dense irrégulier», « tissu conjonctif lâche (aréolaire) ", etc, mais en réalité les tissus de soutien présentent une grande diversité de densité, de régularité et de telles descriptions rigides ont fait leur temps. Les deux premières micrographies montrent des fibres de collagène disposées relativement de façon lâche dans la sous-muqueuse de l'intestin. Il y a des faisceaux de collagène plus épais (b). Les fibroblastes sont rares par rapport à la matrice. Le collagène est acidophile en raison de ses groupes latéraux chargés positivement; donc dans les préparations H & E, le collagène est éosinophile (rose). Un tel tissu de soutien en vrac a été appelé tissu conjonctif aréolaire. Sur le trichrome (c), le collagène est teinté en vert ou bleu (en fonction de la variante de la coloration utilisée). La micrographie (d) est d’un fascia de muscle dévoilant l’arrangement dense typique de fibres de collagène, où le soutien mécanique est la fonction principale. Les fibres de collagène sont disposées de manière régulière afin de fournir un ancrage bien organisé pour transmettre des forces de contraction. Les noyaux des fibroblastes sont allongés dans la direction des fibres de collagène. La photographie (e) est du tissu collagène dense à partir de la couche dure-mère du cerveau où elle sert de barrière, et de support.

Fibroblastes actifs: la cicatrisation des plaies H & E × 400
Ces cellules sont des fibroblastes et les myofibroblastes dans une plaie en cicatrisation. Les noyaux sont grands et avec des nucléoles proéminents reflétant la synthèse protéique active. Le cytoplasme est abondant et rose, l’aspect granuleux est la preuve d'un vaste système de réticulum endoplasmique rugueux impliquée dans la synthèse des protéines. L'absence relative de fibres de collagène formé dans la matrice extracellulaire permet l’observation facile des extensions cytoplasmiques des fibroblastes. Dans les tissus de soutien moins actifs, ces derniers sont moins visibles. Les fibroblastes avec une fonction contractile (myofibroblastes) jouent un rôle important dans la contraction et le rétrécissement du tissu cicatriciel résultant. Guérison et la réparation Suite à des altérations des cellules et des tissus, il y a une réponse inflammatoire qui est responsable de l'élimination de l'agent d’agression et le déblayement des tissus lésés. La réparation des tissus endommagés est principalement réalisée par des cellules de soutien du tissu conjonctif. En bref, il y a une prolifération locale des cellules mésenchymateuses à partir du tissu normal résiduel périphérique et se forme des fibroblastes et les myofibroblastes. Ceux-ci comblent la zone de la lésion tissulaire. Cette prolifération est également associée à la croissance de nouveaux vaisseaux sanguins capillaires pour fournir des nutriments. Les fibroblastes et les myofibroblastes sécrètent la matrice extracellulaire, le collagène et la substance fondamentale, afin de remplacer la partie endommagée par un tissu fibro-collagénique. C’est la cicatrice fibreuse. Avec l’évolution, le remodelage du collagène permet de maximiser la résistance du collagène et de l'associer aux tissus adjacents. Ce processus est appelé la guérison par la réparation. Les tissus spécialisés lésés, comme les muscles ou les poumons, sont remplacés par une cicatrice collagènique dense, mais non fonctionnel.

Le tissu adipeux La plupart des tissus de soutien contient des cellules qui sont adaptées pour le stockage de la graisse; ces cellules, appelées adipocytes, sont dérivés de mésenchyme primitif où ils se développent en tant que lipoblastes. Les adipocytes sont trouvés isolément ou en amas lâches à travers les tissus de soutien ou peuvent constituer le principal type de cellule constituant le tissu adipeux. Les graisses stockées dans les adipocytes proviennent de trois sources principales: les graisses alimentaires circulant dans le sang comme les chylomicrons, les triglycérides synthétisés dans le foie et transporté dans le sang, et les triglycérides synthétisés à partir du glucose dans les adipocytes. Le tissu adipeux est souvent considéré comme un accumulateur d'énergie inactif, mais il est un participant très important dans les processus métaboliques généraux. Il agit comme une réserve temporaire de substrat pour les processus énergétique dans presque tous les tissus. Le tissu adipeux, donc, a généralement un approvisionnement en sang riche. Le taux de dépôt de lipides et de l'utilisation du tissu adipeux est largement déterminée par l'apport alimentaire et la dépense énergétique. Un certain nombre d'hormones et le système nerveux sympathique influent profondément sur le métabolisme des lipides des adipocytes. En plus de leur rôle de stockage énergétique, les adipocytes ont un rôle endocrine. Grâce à la sécrétion de plusieurs protéines, les adipocytes modulent le métabolisme énergétique et influencent le métabolisme général en coordination avec les hormones telles que l'insuline, pour réguler la masse corporelle. Les protéines sécrétées sont connues sous le nom adipocytokines. Il s'agit notamment de la leptine, la dipsine, la résistine, l’adiponectine, le facteur de nécrose tumorale alpha, et l’inhibiteur de l’activateur du plasminogene de type 1. Il existe deux principaux types de tissu adipeux: • Le tissu adipeux blanc : Ce type de tissu adipeux représente jusqu'à 20% du poids total du corps
dans des conditions normales de l’homme adulte et jusqu'à 25% chez les femmes. Il est distribué dans tout le corps en particulier dans les couches profondes de la peau. En plus d'être une réserve d'énergie importante, le tissu adipeux blanc agit comme un isolant thermique sous la peau et agit comme un coussin contre les chocs mécaniques dans des sites tels que les loges des reins.
• Le tissu adipeux brun : Ce type hautement spécialisée du tissu adipeux se trouve chez les mammifères, les nouveau-nés et des animaux hibernants, où il joue un rôle important dans la régulation de la température du corps. Seules de petites quantités de tissu adipeux brun se trouvent chez l'adulte.

Le tissu adipeux blanc (a) H & E × 240 (b) H & E × 480 (c) EM × 6000; C capillaire, Co fibrilles de collagène, D petite gouttelette lipidique, F cytoplasme des fibroblastes, L grande gouttelette lipidique,
M mitochondrie, N noyau, P cytoplasme des adipocytes
Le tissu adipeux blanc (a) H & E × 240 (b) H & E × 480 (c) EM × 6000 L'aspect typique du tissu adipeux blanc est illustré dans micrographies (a) et (b). Au niveau du tissu adipeux au faible grossissement, la coloration est pâle, car la quasi-totalité de la cellule est occupée par des lipides, qui a été dissout dans des préparations de tissus. La membrane cellulaire et une mince bande de cytoplasme périphérique donne un aspect grillagé. Les lipides stockées dans les adipocytes s'accumule sous forme de gouttelettes lipidiques qui fusionnent pour former une seule goutte importante qui distend et occupe la majeure partie du cytoplasme. L'adipocyte noyau N est comprimé et déplacé d'un côté de la gouttelette lipidique stockée et le cytoplasme se réduit à un petit rebord autour de la périphérie. Dans certaines cellules, la coupe tangentielle du haut ou en bas d'une cellule, le cytoplasme P apparaît comme une bande rosâtre. Notez les dimensions minimes capillaires sanguins C par rapport à la taille des adipocytes environnants. La micrographie électronique (c) montre la périphérie de deux adipocytes adjacentes. Contrairement à l'impression donnée par microscopie optique, la vacuole lipidique L dans chaque cellule a un contour irrégulier, avec de nombreuses petites gouttelettes D à la périphérie en voie de fusion avec la vacuole principale. La goutte de lipide n'est pas délimité par une membrane. La bordure mince de cytoplasme contient des organites habituels, notamment les mitochondries M. Dans le tissu extracellulaire adjacent l’extension d’un fibroblaste F et des fibrilles de collagène Co sont vu. Les adipocytes ont des récepteurs à l'insuline, aux glucocorticoïdes, à l'hormone de croissance et à la noradrénaline (norépinephrine) qui modulent l'absorption et la libération des lipides. Les adipocytes sécrètent la leptine, hormone impliquée dans la régulation de l'appétit.
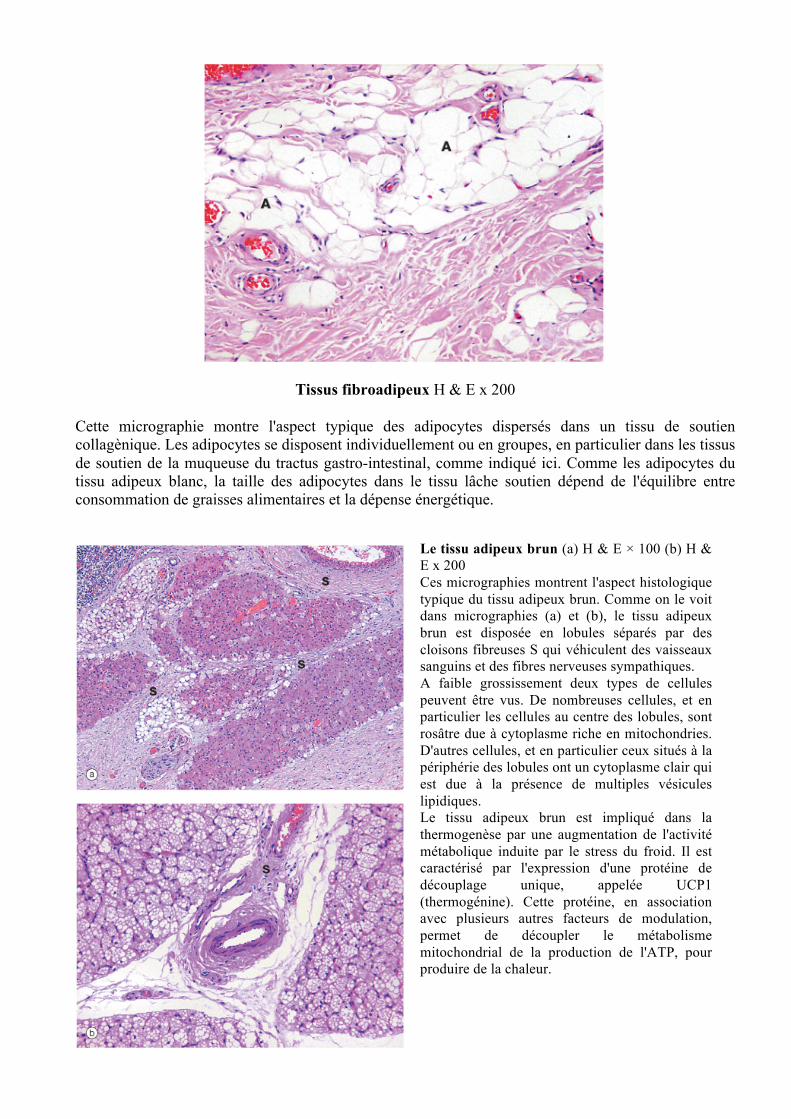
Tissus fibroadipeux H & E x 200
Cette micrographie montre l'aspect typique des adipocytes dispersés dans un tissu de soutien collagènique. Les adipocytes se disposent individuellement ou en groupes, en particulier dans les tissus de soutien de la muqueuse du tractus gastro-intestinal, comme indiqué ici. Comme les adipocytes du tissu adipeux blanc, la taille des adipocytes dans le tissu lâche soutien dépend de l'équilibre entre consommation de graisses alimentaires et la dépense énergétique.
Le tissu adipeux brun (a) H & E × 100 (b) H & E x 200 Ces micrographies montrent l'aspect histologique typique du tissu adipeux brun. Comme on le voit dans micrographies (a) et (b), le tissu adipeux brun est disposée en lobules séparés par des cloisons fibreuses S qui véhiculent des vaisseaux sanguins et des fibres nerveuses sympathiques. A faible grossissement deux types de cellules peuvent être vus. De nombreuses cellules, et en particulier les cellules au centre des lobules, sont rosâtre due à cytoplasme riche en mitochondries. D'autres cellules, et en particulier ceux situés à la périphérie des lobules ont un cytoplasme clair qui est due à la présence de multiples vésicules lipidiques. Le tissu adipeux brun est impliqué dans la thermogenèse par une augmentation de l'activité métabolique induite par le stress du froid. Il est caractérisé par l'expression d'une protéine de découplage unique, appelée UCP1 (thermogénine). Cette protéine, en association avec plusieurs autres facteurs de modulation, permet de découpler le métabolisme mitochondrial de la production de l'ATP, pour produire de la chaleur.
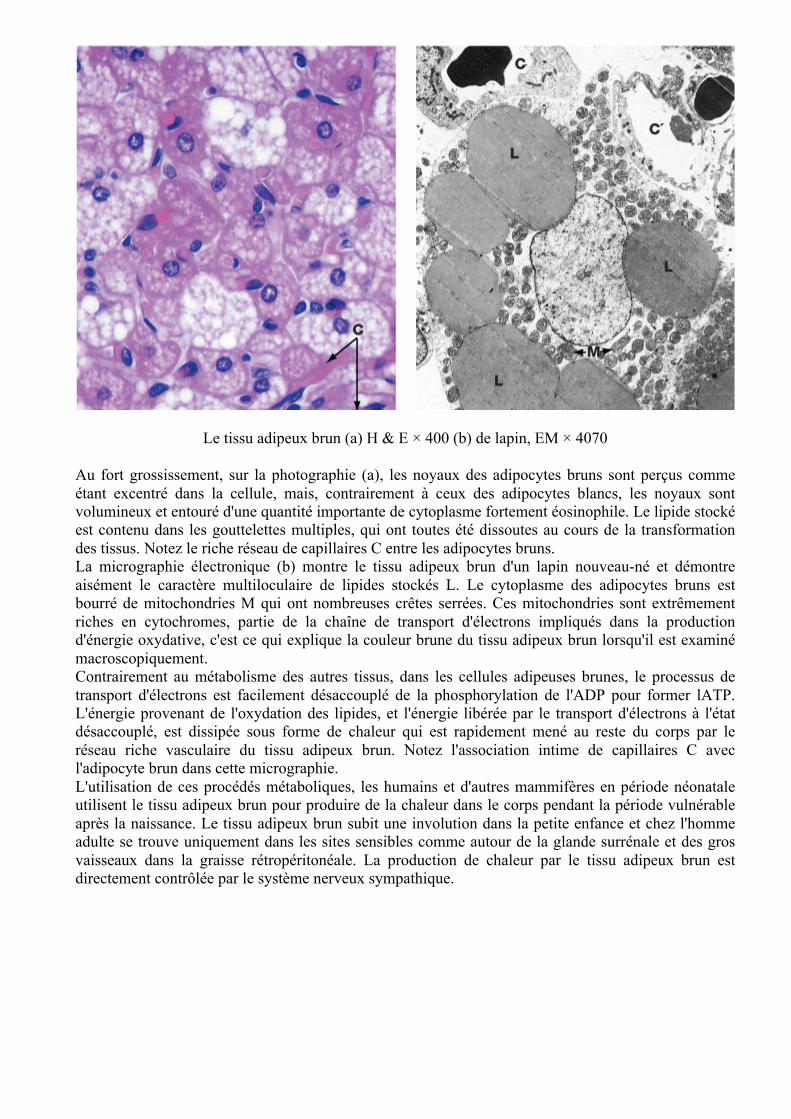
Le tissu adipeux brun (a) H & E × 400 (b) de lapin, EM × 4070
Au fort grossissement, sur la photographie (a), les noyaux des adipocytes bruns sont perçus comme étant excentré dans la cellule, mais, contrairement à ceux des adipocytes blancs, les noyaux sont volumineux et entouré d'une quantité importante de cytoplasme fortement éosinophile. Le lipide stocké est contenu dans les gouttelettes multiples, qui ont toutes été dissoutes au cours de la transformation des tissus. Notez le riche réseau de capillaires C entre les adipocytes bruns. La micrographie électronique (b) montre le tissu adipeux brun d'un lapin nouveau-né et démontre aisément le caractère multiloculaire de lipides stockés L. Le cytoplasme des adipocytes bruns est bourré de mitochondries M qui ont nombreuses crêtes serrées. Ces mitochondries sont extrêmement riches en cytochromes, partie de la chaîne de transport d'électrons impliqués dans la production d'énergie oxydative, c'est ce qui explique la couleur brune du tissu adipeux brun lorsqu'il est examiné macroscopiquement. Contrairement au métabolisme des autres tissus, dans les cellules adipeuses brunes, le processus de transport d'électrons est facilement désaccouplé de la phosphorylation de l'ADP pour former lATP. L'énergie provenant de l'oxydation des lipides, et l'énergie libérée par le transport d'électrons à l'état désaccouplé, est dissipée sous forme de chaleur qui est rapidement mené au reste du corps par le réseau riche vasculaire du tissu adipeux brun. Notez l'association intime de capillaires C avec l'adipocyte brun dans cette micrographie. L'utilisation de ces procédés métaboliques, les humains et d'autres mammifères en période néonatale utilisent le tissu adipeux brun pour produire de la chaleur dans le corps pendant la période vulnérable après la naissance. Le tissu adipeux brun subit une involution dans la petite enfance et chez l'homme adulte se trouve uniquement dans les sites sensibles comme autour de la glande surrénale et des gros vaisseaux dans la graisse rétropéritonéale. La production de chaleur par le tissu adipeux brun est directement contrôlée par le système nerveux sympathique.

Les cellules de défense du tissu de soutien Les tissus de soutien non seulement contiennent des cellules responsables de la synthèse, de l'entretien et de l'activité métabolique, mais aussi une variété de cellules avec la défense et les fonctions immunitaires. Traditionnellement, ces cellules ont été divisées en deux catégories: les cellules fixes (intrinsèque) et cellules de passage (extrinsèque). Les cellules intrinsèques de défense des tissus de soutien sont les macrophages fixes (histiocytes) et les mastocytes. Ces macrophages sont maintenant généralement considérés comme provenant de monocytes circulants, qui sont devenus au moins temporairement résident dans les tissus de soutien. Les mastocytes sont fonctionnellement analogue aux basophiles sanguins, mais il existe des différences structurelles qui suggèrent que les mastocytes ne sont pas simplement des basophiles résidant dans les tissus. Les cellules immunitaires de défense et les cellules de passage sont tous les membres restants de la série de globules blancs. Bien que les leucocytes (globules blancs) sont généralement considérés comme un constituant du sang, leur principal lieu d'activité est en dehors de la circulation sanguine, en particulier dans les tissus de soutien. Les leucocytes sont normalement présents seulement en quantités relativement faibles, mais en réponse à une lésion tissulaire et d'autres processus pathologiques, leur nombre augmente considérablement. Les tissus de soutien de ces régions du corps qui sont sous la menace constante d'une invasion pathogène, tels que les voies gastro-intestinales et respiratoires, contiennent une grande population de leucocytes, et assure le maintien d'une surveillance constante. Le concept du système réticulo-endothélial Le système réticulo-endothélial terme longtemps utilisé pour décrire un groupe diversifié de cellules dans de nombreux tissus, mais en particulier la moelle osseuse, le foie, la rate, les ganglions lymphatiques et le thymus. La principale caractéristique fonctionnelle de ces cellules est leur capacité à phagocyter les particules, micro-organismes, et cellules (usées ou mortes), par exemple, globules âgés. Une autre fonction importante est le stockage du fer et de certains produits métaboliques. Ces cellules phagocytaires se trouvent bordant les capillaires sanguins et espaces lymphatiques, tels que les sinusoïdes du foie, la moelle osseuse et la rate, et dans ce contexte, ils ont certaines caractéristiques communes avec les cellules endothéliales qui tapissent ces vaisseaux. Certains tissus hautement cellulaires, comme les ganglions lymphatiques et les cordons hématopoïétiques de la moelle osseuse, disposent d'un support en fibres de réticuline sur lesquels sont étalées ces cellules avec de longs prolongements cytoplasmiques morphologiquement similaires à des cellules mésenchymateuses primitives. Ces cellules sont traditionnellement décrites comme des cellules réticulaires. Certains, sinon la totalité de ces cellules sont probablement responsables de la synthèse du support de réticuline, étant donc analogue à des fibroblastes; bon nombre de ces cellules, sinon toutes, peuvent également présenter une forte activité phagocytaire. En raison de l'étroite association structurelle et fonctionnelle de ces types cellulaires avec les systèmes hématopoïétiques, macrophage-monocyte, et le système immunitaire, ils semblent représenter un même système fonctionnel. Par conséquent, «réticulo-endothélial» et termes apparentés sont devenus largement et souvent appliquée sans discrimination aux tissus et cellules, au sein de ces systèmes. Les progrès scientifiques ont rendu le concept imprécis. Toutefois, il est important de comprendre la signification de ce terme. A la recherche sur PubMed pour les documents écrits en 2005 va révéler continuant à utiliser le terme système réticulo-endothélial pour décrire un réseau de cellules phagocytaires distribué principalement situées dans la moelle osseuse, la rate, le foie et les ganglions lymphatiques. Dans certaines maladies héréditaires anormales de stockage des produits métaboliques peuvent également s'accumuler dans les cellules du système réticulo-endothélial conduisant à l'élargissement du foie et de la rate.
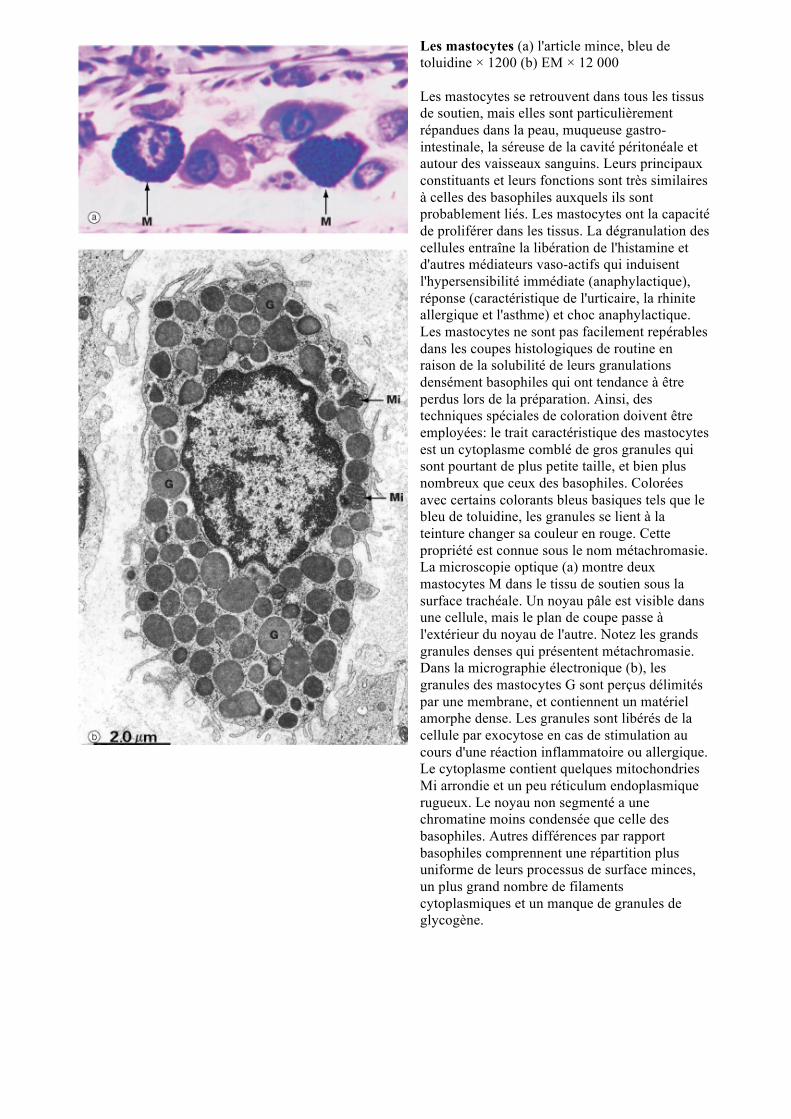
Les mastocytes (a) l'article mince, bleu de toluidine × 1200 (b) EM × 12 000 Les mastocytes se retrouvent dans tous les tissus de soutien, mais elles sont particulièrement répandues dans la peau, muqueuse gastro-intestinale, la séreuse de la cavité péritonéale et autour des vaisseaux sanguins. Leurs principaux constituants et leurs fonctions sont très similaires à celles des basophiles auxquels ils sont probablement liés. Les mastocytes ont la capacité de proliférer dans les tissus. La dégranulation des cellules entraîne la libération de l'histamine et d'autres médiateurs vaso-actifs qui induisent l'hypersensibilité immédiate (anaphylactique), réponse (caractéristique de l'urticaire, la rhinite allergique et l'asthme) et choc anaphylactique. Les mastocytes ne sont pas facilement repérables dans les coupes histologiques de routine en raison de la solubilité de leurs granulations densément basophiles qui ont tendance à être perdus lors de la préparation. Ainsi, des techniques spéciales de coloration doivent être employées: le trait caractéristique des mastocytes est un cytoplasme comblé de gros granules qui sont pourtant de plus petite taille, et bien plus nombreux que ceux des basophiles. Colorées avec certains colorants bleus basiques tels que le bleu de toluidine, les granules se lient à la teinture changer sa couleur en rouge. Cette propriété est connue sous le nom métachromasie. La microscopie optique (a) montre deux mastocytes M dans le tissu de soutien sous la surface trachéale. Un noyau pâle est visible dans une cellule, mais le plan de coupe passe à l'extérieur du noyau de l'autre. Notez les grands granules denses qui présentent métachromasie. Dans la micrographie électronique (b), les granules des mastocytes G sont perçus délimités par une membrane, et contiennent un matériel amorphe dense. Les granules sont libérés de la cellule par exocytose en cas de stimulation au cours d'une réaction inflammatoire ou allergique. Le cytoplasme contient quelques mitochondries Mi arrondie et un peu réticulum endoplasmique rugueux. Le noyau non segmenté a une chromatine moins condensée que celle des basophiles. Autres différences par rapport basophiles comprennent une répartition plus uniforme de leurs processus de surface minces, un plus grand nombre de filaments cytoplasmiques et un manque de granules de glycogène.
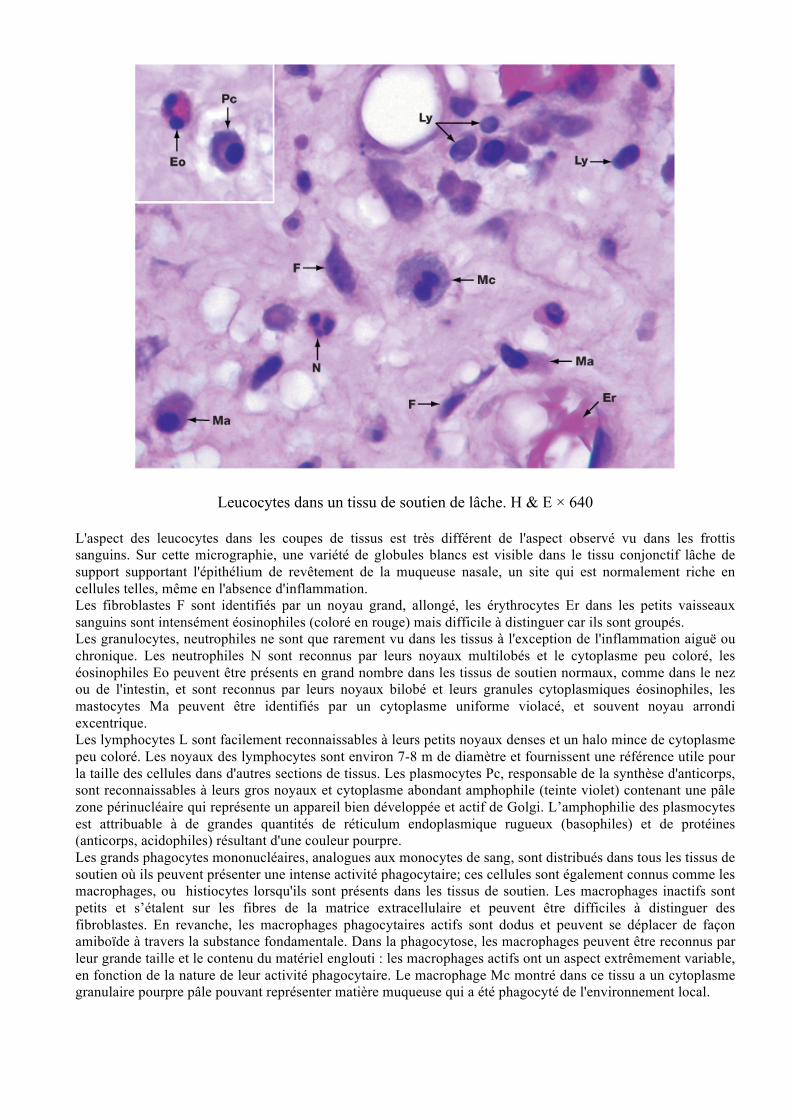
Leucocytes dans un tissu de soutien de lâche. H & E × 640
L'aspect des leucocytes dans les coupes de tissus est très différent de l'aspect observé vu dans les frottis sanguins. Sur cette micrographie, une variété de globules blancs est visible dans le tissu conjonctif lâche de support supportant l'épithélium de revêtement de la muqueuse nasale, un site qui est normalement riche en cellules telles, même en l'absence d'inflammation. Les fibroblastes F sont identifiés par un noyau grand, allongé, les érythrocytes Er dans les petits vaisseaux sanguins sont intensément éosinophiles (coloré en rouge) mais difficile à distinguer car ils sont groupés. Les granulocytes, neutrophiles ne sont que rarement vu dans les tissus à l'exception de l'inflammation aiguë ou chronique. Les neutrophiles N sont reconnus par leurs noyaux multilobés et le cytoplasme peu coloré, les éosinophiles Eo peuvent être présents en grand nombre dans les tissus de soutien normaux, comme dans le nez ou de l'intestin, et sont reconnus par leurs noyaux bilobé et leurs granules cytoplasmiques éosinophiles, les mastocytes Ma peuvent être identifiés par un cytoplasme uniforme violacé, et souvent noyau arrondi excentrique. Les lymphocytes L sont facilement reconnaissables à leurs petits noyaux denses et un halo mince de cytoplasme peu coloré. Les noyaux des lymphocytes sont environ 7-8 m de diamètre et fournissent une référence utile pour la taille des cellules dans d'autres sections de tissus. Les plasmocytes Pc, responsable de la synthèse d'anticorps, sont reconnaissables à leurs gros noyaux et cytoplasme abondant amphophile (teinte violet) contenant une pâle zone périnucléaire qui représente un appareil bien développée et actif de Golgi. L’amphophilie des plasmocytes est attribuable à de grandes quantités de réticulum endoplasmique rugueux (basophiles) et de protéines (anticorps, acidophiles) résultant d'une couleur pourpre. Les grands phagocytes mononucléaires, analogues aux monocytes de sang, sont distribués dans tous les tissus de soutien où ils peuvent présenter une intense activité phagocytaire; ces cellules sont également connus comme les macrophages, ou histiocytes lorsqu'ils sont présents dans les tissus de soutien. Les macrophages inactifs sont petits et s’étalent sur les fibres de la matrice extracellulaire et peuvent être difficiles à distinguer des fibroblastes. En revanche, les macrophages phagocytaires actifs sont dodus et peuvent se déplacer de façon amiboïde à travers la substance fondamentale. Dans la phagocytose, les macrophages peuvent être reconnus par leur grande taille et le contenu du matériel englouti : les macrophages actifs ont un aspect extrêmement variable, en fonction de la nature de leur activité phagocytaire. Le macrophage Mc montré dans ce tissu a un cytoplasme granulaire pourpre pâle pouvant représenter matière muqueuse qui a été phagocyté de l'environnement local.

Macrophage EM × 11 600
Les caractéristiques ultrastructurales des macrophages varient largement en fonction de leur état d'activité et la localisation tissulaire. Cette micrographie montre un macrophage actif obtenue à partir du péritoine d'un rat qui avait déjà été injecté par voie intrapéritonéale avec des particules de latex, un certain nombre de particules P ont été englouties par les macrophages. Le noyau des macrophages est irrégulier avec une hétérochromatine généralement agglutinée sous l'enveloppe nucléaire. Le cytoplasme contient peu de mitochondries M et une quantité variable de ribosomes libres et réticulum endoplasmique rugueux RER. Dans les macrophages quiescents, les lysosomes L sont abondants mais leur nombre est très réduit dans les cellules phagocytaires actives. Les lysosomes sont régénérés par l'appareil de Golgi. Le cytoplasme des macrophages contient un assortiment de phagosomes et les organes résiduels R. matières résiduelles peuvent être libérés par les macrophages par exocytose. Un tel matériel peut rester séquestré dans les tissus, comme c'est le cas avec les colorants utilisés dans le tatouage de la peau. Les cellules phagocytaires actives présentent des projections cytoplasmiques irrégulières ou pseudopodes Pp qui sont impliqués dans le mouvement amiboïdes et la phagocytose. En plus de leur rôle de nettoyage des tissus, les macrophages jouent un rôle important dans les mécanismes immunitaires, car elles sont souvent les premières cellules à entrer en contact avec des antigènes. Les macrophages traite le matériel antigénique avant de le présenter aux lymphocytes. Les lymphocytes sont ensuite stimulés a fournir des réponses immunitaires spécifiques. Les macrophages impliqués dans cette voie sont décrits en tant que cellules présentatrices d'antigène. En raison de divers mécanismes immunitaires, le matériel antigénique peut être combinés ou recouverts de substances telles que des anticorps et du complément qui sont phagocytés et connus sous le nom d’opsonines. Les opsonines sont reconnues par des récepteurs de surface sur la surface des macrophages, ce qui améliore grandement la capacité phagocytaire des macrophages et autres phagocytes comme les neutrophiles. Ce processus qui est connu comme d’opsonisation. D'autres substances comme les lymphokines, qui sont libérés lors de la réponse immunitaire, agissent directement sur les macrophages pour augmenter considérablement leur métabolisme et leur activité phagocytaire. Les macrophages sécrètent également une variété de cytokines qui agissent pour améliorer les réponses immunitaires locales et systémiques. Dans certains états pathologiques, les macrophages peuvent développer un cytoplasme abondant où ils sont appelés macrophages épithélioïdes. Dans d'autres situations pathologiques macrophages peuvent fusionner pour former des cellules géantes multinucléées.


















