
El Computador Isabela Montoya Monsalve Yulieth Fernanda Moreno Martínez.

EVALUACIÓN DE LA DIVERSIDAD GENÉTICA EN UNA COLECCIÓN
DE GERMOPLASMA DE FRÍJOL COMÚN (Phaseolus vulgaris L.) DE
RUANDA (ÁFRICA)
LAURA FERNANDA GONZÁLEZ MARTÍNEZ
PONTIFICIA UNIVERSIDAD JAVERIANA
FA S CULTA ENCIAD DE CICARRERA BIOLOGÍA
Bogotá 18 de Febrero de 2008

EVALUACIÓN DE LA DIVERSIDAD GENÉTICA EN UNA COLECCIÓN
DE GERMOPLASMA DE FRÍJOL COMÚN (Phaseolus vulgaris L.) DE
RUANDA (ÁFRICA)
LAURA FERNANDA GONZÁLEZ MARTÍNEZ
TRABAJO DE GRADO
Presentado como requisito parcia para optar al título de: l
BIÓLOGA
PONTIFICIA UNIVERSIDAD JAVERIANA FA S CULTA IENCIAD DE CCARRERA BIOLOGÍA
Bogotá 18 de Febrero de 2008
ii

iii
Artículo 23 de la Resolución Nº 13 de Julio de 1946
“La Universidad no se hace responsable por los conceptos emitidos por sus alumnos en sus trabajos de tesis. Sólo velará por que no se publique nada contrario al dogma y a la moral católica y por que las tesis no contengan ataques personales contra persona alguna, antes bien se vea en ellas el anhelo de buscar la verdad y la justicia”.

EVALUACIÓN DE LA DIVERSIDAD GENÉTICA EN UNA COLECCIÓN DE GERMOPLASMA DE FRÍJOL COMÚN (Phaseolus vulgaris L.)
DE RUANDA (África).
LAURA FERNANDA GONZÁLEZ MARTÍNEZ
APROBADO
_________________________ ___________________________ Matthew W. Blair, Ph.D María del Pilar Márquez, MSc.
Director Codirectora
_________________________ ________________________ Manuel Ruíz García, Ph.D Wilson Terán, Ph.D
Jurado Jurado
ii

EVALUACIÓN DE LA DIVERSIDAD GENÉTICA EN UNA COLECCIÓN DE
GERMOPLASMA DE FRÍJOL COMÚN (Phaseolus vulgaris L.)
DE RUANDA (África).
LAURA FERNANDA GONZÁLEZ MARTÍNEZ
APROBADO
______________________ _______________________
ANGELA UMAÑA ANDREA FORERO Decana Académico Directora de Carrera
iii

Bogotá, 6 de Febrero 2008 Señores PONTIFICIA UNIVERSIDAD JAVERIANA Cuidad Estimados Señores: Yo Laura Fernanda González Martínez, identificada con C.C. No. 52966299 de Bogotá , autora del trabajo de grado titulado “Evaluación de la diversidad genética en una colección de germoplasma de fríjol común (Phaseolus vulgaris L.) de Ruanda (África)”, presentado como requisito para optar al título de Bióloga en el año de 2008; autorizo a la Pontificia Universidad Javeriana a: a) Reproducir el trabajo en medio digital o electrónico con el fin de ofrecerlo para la consulta en la Biblioteca General SI .. b) Poner a disposición para la consulta con fines académicos, en la página web de la Facultad, de la Biblioteca General y en redes de información con las cuales tenga convenio la Universidad Javeriana No . c) Enviar el trabajo en formato impreso o digital, en caso de que sea seleccionado para participar en concursos de trabajos de grado SI . d) Distribuir ejemplares de la obra, para la consulta entre las entidades educativas con las que la facultad tenga convenio de intercambio de información, para que este sea consultado en las bibliotecas y centros de documentación de las respectivas entidades SI EN TEXTO, NO EN DOCUMENTO ELECTRÓNICO .. e) Todos los usos, que tengan finalidad académica SI EN TEXTO, NO EN DOCUMENTO ELECTRÓNICO Los derechos morales sobre el trabajo son de los autores de conformidad con lo establecido en el artículo 30 de la Ley 23 de 1982 y el artículo 11 de la Decisión Andina 351 de 1993, los cuales son irrenunciables, imprescriptibles, inembargables e inalienables. Atendiendo lo anterior, siempre que se consulte la obra, mediante cita bibliográfica se debe dar crédito al trabajo y a su(s) autor(es). Este documento se firma, sin perjuicio de los acuerdos que el autor(es) pacte con la Unidad Académica referentes al uso de la obra o a los derechos de propiedad industrial que puedan surgir de la actividad académica. __________________________ Laura Fernanda González M C.C 52.966.299
iv

FORMATO DESCRIPCIÓN TRABAJO DE GRADO AUTOR Apellidos: González Martínez Nombres: Laura Fernanda DIRECTOR Apellidos: Blair Nombres: Matthew W. ASESOR Apellidos: Márquez Nombres: María del Pilar TRABAJO PARA OPTAR POR EL TÍTULO DE: Bióloga TÍTULO COMPLETO DEL TRABAJO: Evaluación de la diversidad genética en una colección de germoplasma de fríjol común (Phaseolus vulgaris L.) de Ruanda (África). FACULTAD: Ciencias PROGRAMA: Carrera: Biología CIUDAD: Bogotá 2008 NÚMERO DE PÁGINAS: 130 TIPO DE ILUSTRACIONES: - Ilustraciones - Mapas - Tablas - Gráficos y diagramas
DESCRIPTORES O PALABRAS CLAVES: Diversidad genética, Microsatélites,
Fríjol común.
v

Dedico este trabajo a Dios, porque fueron sus manos las que lo
elaboraron; por ser mi guía y mi fortaleza, por haber puesto los mejores
regalos en mi camino.
Al regalo mas hermoso que Dios me ha dado, Mi Familia, especialmente a mis
padres, José y Julia y mis abuelitos Belén y Mardoqueo por su infinito anhelo por
verme feliz siempre; por entregarme su sabiduría y sus consejos; y simplemente
porque a ustedes les debo mi vida. Las lágrimas, el sacrificio y las mil risas que lleva
este trabajo son dedicadas a ustedes por ser los artífices de la mujer que ahora soy.
vi

vii
AGRADECIMIENTOS
Al doctor Matthew Blair por esta magnífica oportunidad de dejarme hacer parte de su equipo
de trabajo y por haber depositado su confianza en mi.
A Tito Sandoval por su felicidad e inmensa entrega; por darme tanta tranquilidad y hacerme
brotar una sonrisa cada día… por hacer esto parte de su vida. Gracias por ser mi ángel y mi
fuerza durante esta travesía de la tesis.
A Myriam Cristina Duque por su INCONDICIONAL apoyo en el desarrollo de la parte
estadística y en general en la elaboración del documento; por sus consejos llenos de
sabiduría y en especial por su amistad.
A Héctor Fabio Buendía, por sus invaluables consejos y por el inmenso aporte de
conocimientos académicos y de la vida. A Lucy Milena Díaz por su confianza y su esfuerzo
depositados en este trabajo. A Natalia Moreno por la colaboración en todas las etapas de mi
paso por CIAT.
A Louis Butare por el inmenso aporte de conocimientos de Ruanda, por las apreciaciones
hechas y sus buenos consejos; por su alegría y su apoyo.
A los demás compañeros de laboratorio, Marcela, Asrat, Teshale, Juliana, Hernán, Carolina A, Carlos, Gina, Carolina Ch, Gloria, Alejandra, Aura, y Alejandro, por su compañía, sus risas y en especial por las oportunidades de crecimiento profesional y personal brindadas en el ambiente que compartimos.
A los trabajadores del patio de fríjol, Don Luis, Alcides y en especial a Agobargo Hoyos por
su paciencia y su ánimo en los momentos difíciles.
A la doctora Ingrid Schuler y a María del Pilar Márquez por creer siempre en mí y por su
ayuda para mi vinculación en el CIAT.
A Piedad Medina y toda la familia por su inmenso apoyo emocional y porque en realidad me
hicieron sentir parte de su hogar.
A Clara Yalexy Delgado por brindarme su amistad sincera durante tantos años, por ser mi
alegría y calma en los momentos más difíciles.
A mis amigas Natalia Galindo, Andrea Barrera, Ivonne Salamanca, Martica Melo, Xamara
Albarán y Luisa Castellanos, por su paciencia, alegría y compañía durante todos estos años.

TABLA DE CONTENIDO
1 INTRODUCCIÓN ________________________________________ 3
2 MARCO TEÓRICO Y REVISIÓN DE LITERATURA _____________ 5
2.1 GENERALIDADES DEL FRÍJOL __________________________________________ 5 2.1.1 Características Taxonómicas y Botánicas _____________________________________ 5 2.1.2 Origen, distribución y domesticación ________________________________________ 7 2.1.3 Cultivo, producción y Usos _______________________________________________ 10 2.1.4 Composición Química y Calidad nutricional __________________________________ 11
2.1.4.1 Composición Química _______________________________________________ 11 2.1.4.2 Calidad Nutricional _________________________________________________ 11
2.2 SITUACIÓN DEL FRÍJOL EN ÁFRICA _____________________________________ 13 2.2.1 Generalidades Ruanda __________________________________________________ 15 2.2.2 Fríjol en Ruanda _______________________________________________________ 17
2.2.2.1 Producción y consumo ______________________________________________ 18 2.2.2.2 Problemática Nutricional ____________________________________________ 19 2.2.2.3 Estrategias para combatir la problemática nutricional _____________________ 21
2.2.3 Perspectivas del cultivo de Fríjol ___________________________________________ 23
2.3 DIVERSIDAD GENÉTICA Y ANALISIS MOLECULAR MEDIANTE MICROSATLITES __ 23 2.3.1 Marcadores Moleculares ________________________________________________ 23 2.3.2 Microsatélites analizados mediante fluorescencia y Análisis secuenciador automático 26 2.3.3 Estudios de diversidad genética en Fríjol común ______________________________ 27 2.3.4 Estudios de diversidad en fríjol Africano ____________________________________ 30
2.4 CUANTIFICACION DEL CONTENIDO DE MINERALES EN FRÍJOL ______________ 31 2.4.1 Cuantificación por Absorción Atómica (AA) __________________________________ 31 2.4.2 Estudios de cuantificación de minerales en Fríjol Común _______________________ 32
3 FORMULACIÓN DEL PROBLEMA ________________________ 34
3.1 FORMULACIÓN DEL PROBLEMA ______________________________________ 34
3.2 JUSTIFICACIÓN ____________________________________________________ 35
4 OBJETIVOS ___________________________________________ 37
4.1 Objetivo General __________________________________________________ 37
4.2 Objetivos Específicos _______________________________________________ 37
5 MATERIALES Y MÉTODOS ______________________________ 38
5.1 Población de Estudio _______________________________________________ 38
5.2 MÉTODOS ________________________________________________________ 40
viii

5.2.1 Procesamiento del material vegetal: _______________________________________ 40 5.2.1.1 Germinación de la semilla: ___________________________________________ 40 5.2.1.2 Procesamiento Post‐germinación _____________________________________ 40
5.2.2 Extracción y cuantificación de ADN ________________________________________ 41 5.2.3 Amplificación mediante PCR de microsatélites analizados con fluorescencia ________ 42 5.2.4 Análisis en secuenciador Analítico ABI 3730xl ________________________________ 44 5.2.5 Cuantificación de hierro y zinc ____________________________________________ 46 5.2.6 Identificación de características de grano ___________________________________ 46
5.3 ANÁLISIS DE DATOS ________________________________________________ 47 5.3.1 Análisis de Diversidad Genética ___________________________________________ 47
5.3.1.1 Parámetros de uso común de diversidad para población total _______________ 48 5.3.1.2 Parámetros de uso común de diversidad para grupos formados a partir de PCoA 49 5.3.1.3 Coeficiente de similaridad ___________________________________________ 49
5.3.2 Análisis de cuantificación de minerales _____________________________________ 50 5.3.3 Características morfológicas ______________________________________________ 51
6 RESULTADOS ________________________________________ 52
6.1 ANÁLISIS DE DIVERSIDAD GENÉTICA ___________________________________ 52 6.1.1 Diversidad Genética y estructura Poblacional ________________________________ 52
6.1.1.1 Determinación de alelos ____________________________________________ 52 6.1.1.2 Población Total ____________________________________________________ 53 6.1.1.3 Análisis de Ordenación ______________________________________________ 56 6.1.1.4 Grupos definidos mediante PCoA _____________________________________ 64
6.1.2 Posibles eventos de introgresión entre acervos _______________________________ 65 6.1.3 Agrupación mediante índice de disimilaridad ________________________________ 69
6.2 ANÁLISIS DE MINERALES ____________________________________________ 71 6.2.1 Contenido de minerales en semilla de fríjol __________________________________ 71 6.2.2 Asociación contenido de minerales y estructura genética _______________________ 74
6.3 CARACTERISTICAS DEL GRANO DE FRÍJOL ______________________________ 76
7 DISCUSIÓN DE RESULTADOS ___________________________ 79
7.1 ANÁLISIS DE DIVERSIDAD GENÉTICA ___________________________________ 79 7.1.1 Población Total ________________________________________________________ 79 7.1.2 Grupos definidos mediante PCoA __________________________________________ 83 7.1.3 Agrupación mediante índice de disimilaridad ________________________________ 85 7.1.4 Posibles eventos de introgresión en la población estudiada _____________________ 86
7.2 ANÁLISIS DE MINERALES ____________________________________________ 87
8 CONCLUSIONES ______________________________________ 90
9 RECOMENDACIONES Y PERSPECTIVAS __________________ 92
10 ANEXOS ____________________________________________ 100
ix

INDICE DE TABLAS
Tabla 1. Contenido nutricional de semilla de fríjol. Contenido total en semilla y por 100g de semilla (Geil y Anderson, 1994 En: (Sandoval, 2006). ......................... 12
Tabla 2. Áreas de producción de Fríjol en 20 países de África, modificado de “Atlas of common bean production in Africa” (Wortmann et al, 1998). ......................... 15
Tabla 3. Comparación de los indicadores poblacionales y económicos de Ruanda y África Sub-Sahariana (ISAR, 2000). .................................................................. 17
Tabla 4. Concentraciones de Hierro, Zinc y proteína en cultivares producidos en África Oriental, Central y del sur (CIAT, 2005a) ................................................. 22
Tabla 5. Controles Diversidad utilizados en este estudio con su respectivo origen (acervo al que pertenecen) ................................................................................ 39
Tabla 6. Marcadores microsatélite fluorescentes utilizados en la caracterización de la diversidad genética de 355 genotipos de una colección de germoplasma de Ruanda. .............................................................................................................. 42
Tabla 7. Reactivos necesarios para coctel PCR, condiciones necesarias y concentraciones originales (stock). .................................................................... 43
Tabla 8. Parámetros de descripción para características de la semilla de fríjol; basado en el catalogo de líneas avanzadas. ..................................................... 47
Tabla 9. Rango de valores medidos en ppm para interpretación de resultados de cuantificación de minerales (hierro y zinc). ........................................................ 50
Tabla 10. No de Alelos, Diversidad Genética y Heterocigocidad observada de los 30 microsatélites (16 Genómicos y 14 Génicos) utilizados en este estudio. .......... 54
Tabla 11. Prueba t para comparar parámetros de diversidad (Heterocigocidad Observada y Diversidad Genética) entre marcadores genómicos y génicos. .... 56
Tabla 12. Descriptores de diversidad para 6 grupos definidos por PCoA. ................ 64
x

Tabla 13. Descriptores de diversidad (Número de alelos y Diversidad genética de Nei) para 30 loci SSR en población definida a partir de PCoA. ......................... 65
Tabla 14. Análisis de los parámetros de diversidad (Número de alelos y Diversidad genética de Nei) para 7 grupos formados incluyendo el nuevo grupo considerado como introgresado (Mesi-Intro)...................................................... 68
Tabla 15. Estadística descriptiva para contenido de (ppm) de hierro y zinc en 355 accesiones de frijol provenientes de Ruanda..................................................... 71
Tabla 16. Estadística descriptiva para contenido en ppm de hierro y zinc en las poblaciones generadas mediante PCoA. ........................................................... 75
xi

INDICE DE FIGURAS
Figura 1. Mapa de Ruanda, organización política (UnitedNations, 2007) ................. 16
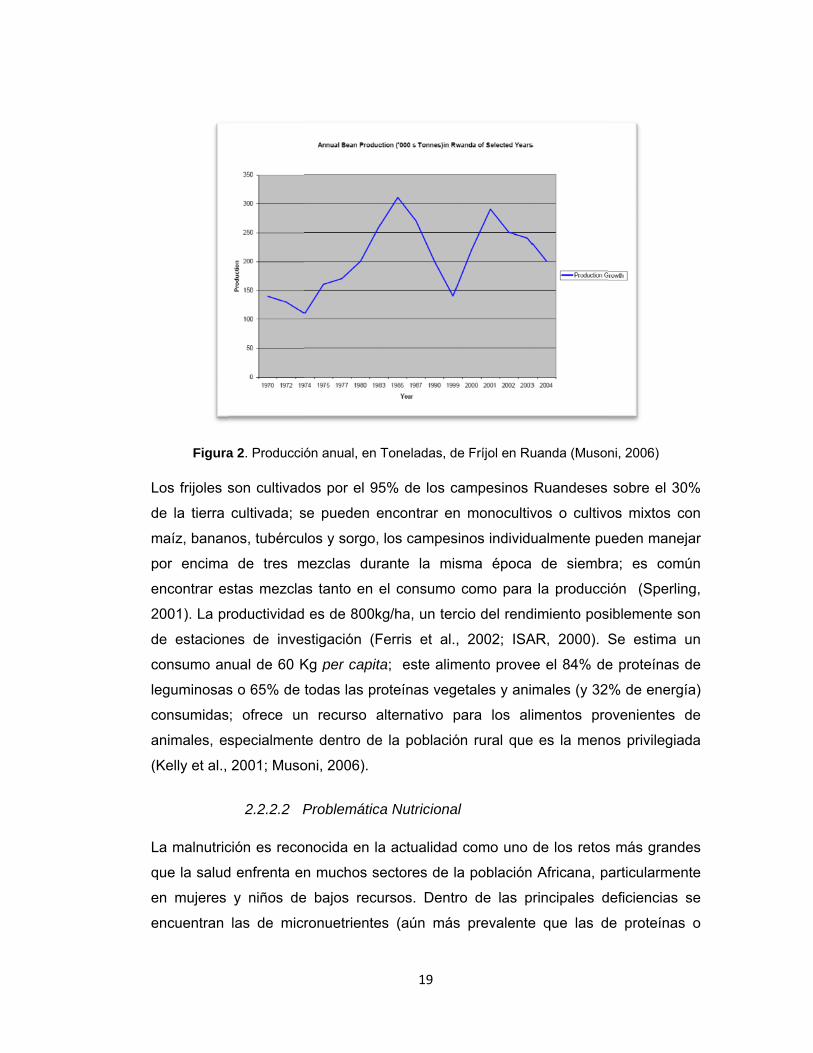
Figura 2. Producción anual, en Toneladas, de Fríjol en Ruanda (Musoni, 2006) ..... 19
Figura 3. Sistema de flujo de la información obtenida a partir del archivo de la muestra dentro del software de auto-análisis. Modificado de (Joe et al., 2004). ........................................................................................................................... 27
Figura 4. Localización geográfica de Ruanda en el continente Africano ................... 38
Figura 5. Diagrama del procedimiento que sigue la muestra a analizar (líneas azules gruesas señalan cada paso). Modificado de (Joe et al., 2004). ......................... 45
Figura 6. Electroferograma obtenido a partir del análisis por ABI 3730xl y generado por el software GeneMapper v.3.7. .................................................................... 52
Figura 7. Electroferograma obtenido a partir del análisis por secuenciador ABI 3730xl y generado por software GeneMapper v.3.7. ......................................... 53
Figura 8. Representación gráfica en tres dimensiones de la distribución y ubicación espacial de los genotipos de Ruanda a partir del Análisis de Coordenadas Principales (PCoA), teniendo en primer plano horizontal la dimensión 2. ......... 57
Figura 9. Representación gráfica en tres dimensiones de la distribución y ubicación espacial de los genotipos de Ruanda a partir del Análisis de Coordenadas Principales (PCoA), teniendo en primer plano horizontal las dimensiones 1 y 2 ........................................................................................................................... 58
Figura 10. Individuos considerados como raros a partir del análisis de coordenadas principales. ......................................................................................................... 59
Figura 11. Dendrograma de acervos genéticos obtenido por distancia euclidiana aplicada a coordenadas obtenidas por PCoA. ................................................... 60
Figura 12A. Dendrograma de acervos genéticos obtenido por distancia euclidiana aplicada a coordenadas obtenidas por PCoA. Muestra dos grupos formados en el acervo Mesoamericano (MI y MIII). ................................................................ 61
xii

xiii
Figura 13. Individuos que se presentan como posibles eventos de introgresión dentro del estudio ............................................................................................... 66
Figura 14. Individuos pertenecientes a Meso-Intro (agrupados por PCoA como Meso II pero que posiblemente presenten eventos de introgresión) ........................... 69
Figura 15. Dendrograma basado en el índice de disimilaridad de DICE para la colección de germoplasma de Ruanda.. ............................................................ 70
Figura 16. Histogramas de cuantificación de minerales mediante la metodología de AA. ..................................................................................................................... 72
Figura 17. Genotipos con mayor y menor contenido de hierro y zinc ....................... 73
Figura 18. Gradiente de concentración de zinc encontrado en grupos mesoamericanos indicado por las flechas azules. ............................................. 75
Figura 19. Distribución porcentual de color, tamaño y patrón de coloración de semilla en la población de estudio. ................................................................................ 76
Figura 20. Distribución del color de la semilla dentro de cada grupo poblacional formado a partir de PCoA................................................................................... 77

RESUMEN
La presente investigación, adelantada mediante la realización de un trabajo
interinstitucional entre la Pontificia Universidad Javeriana y el Centro Internacional
de Agricultura Tropical (CIAT) ubicados en Colombia, es una contribución de la
ciencia para enfrentar uno de los más importantes problemas mundiales
relacionados con el hambre en países en desarrollo. El fríjol común Phaseolus
vulgaris L. es una de las leguminosas más comunes a nivel mundial, siendo uno de
los alimentos más consumidos en los países en vías de desarrollo; en Ruanda
(África) éste es el principal alimento en la dieta diaria. Debido a estos hechos
trascendentales esta planta es el foco de muchos estudios de diversidad genética
alrededor del mundo. El objetivo de este estudio fue caracterizar la diversidad
genética de una colección de fríjol común proveniente de Ruanda. Un total 355
genotipos y 4 individuos (pertenecientes a los dos acervos principales del fríjol)
usados como controles, fueron analizados mediante el uso de 30 marcadores
moleculares tipo microsatélites fluorescentes. Se utilizaron dos tipos de marcadores
genómicos (16) y génicos (14). Un total de 301 bandas fueron generadas con un
promedio de 10.16 alelos por marcador y una diversidad total de 0.62. El análisis de
coordenadas principales identificó la presencia de los dos acervos principales del
fríjol (Andino y mesoamericano) y un posible grupo de genotipos introgresados. Se
presentó mayor diversidad dentro del grupo de los mesoamericanos. Adicional a
esto se realizó un análisis de datos minerales (hierro y zinc), mediante la técnica de
absorción atómica (AA), encontrando valores altos para estos dos minerales. La
caracterización de la diversidad genética de este grupo de genotipos de Ruanda
permitió observar altos niveles de variabilidad genética y un importante contenido de
minerales lo cual puede ser muy útil en futuros planes de mejoramiento.
1

ABSTARCT
This research work, carried out through an interinstitutional work between Pontificia
Javeriana University and The International Center of Tropical Agriculture –CIAT-
located in Colombia, is a contribution of science to face one of the most important
global issues dealing with hunger in developing countries. Common bean Phaseolus
vulgaris L. is one of the most important legumes worldwide, belonging to the most
consumed food in developing countries. In Rwanda (Africa), beans provide the
centre-piece of the daily diet. Due to its great significance, common bean forms a
focal point in many studies of genetic diversity throughout the world. The objective of
this thesis is the genetic diversity characterization of a common bean collection
originating from Rwanda. A total of 355 genotypes and 4 individuals used as controls
(belonging to the two major gene pools of common bean) were analyzed through the
use of 30 molecular markers which were fluorescent microsatellites. Two types of
markers were used: genomic (16) and cDNA markers (14). A total of 301 bands were
generated with an average of 10.16 alleles per marker and a total diversity of 0.62.
Principal component analysis identified the presence of two major gene pools of
common bean (Andean and Meso-American) and a possible group of introgressed
genotypes. Most diversity was found in the group of the Meso-American genotypes.
Additionally, an analysis of mineral content (iron and zinc) was realized using the
method of atomic absorption (AA). High amounts were found for both minerals. The
characterization of genetic diversity of this genotype collection of Rwanda allowed to
observe high levels of genetic variability and high contents of minerals. This
information may be very useful in future breeding projects.
2

1 INTRODUCCIÓN
La desnutrición es uno de los grandes problemas que actualmente aqueja a la
humanidad. Alrededor de 800 millones de personas en todo el mundo la padecen,
esto ha llevado a buscar soluciones encaminadas al mejoramiento y conservación
de las especies y hacía una mayor producción de las mismas, especialmente, en
países en vía de desarrollo.
Una consideración importante para iniciar esta investigación, radica en el hecho de
que tres de los principales centros de estudio en el mundo son África, el
subcontinente Asiático y América Latina; estos con frecuencia basan su dieta en
alimentos como yuca, maíz, arroz, trigo, entre otros, que son ricos en carbohidratos,
pero pobres en proteínas y minerales. Por tanto, especies vegetales como las
leguminosas, que son el grupo de plantas que produce grano más rico en proteínas
y minerales, juegan un papel crucial en la dieta de sus habitantes.
Dentro de este grupo de plantas se encuentra una de las especies mas importantes
en la que la humanidad fundamenta su alimentación, el fríjol común Phaseolus
vulgaris L. América y África son los principales productores y consumidores de esta
leguminosa. Más de 100 millones de personas en África, la mayoría de bajos
recursos pertenecientes tanto a áreas rurales como urbanas, consumen fríjoles
dentro de su dieta diaria, por ser estos una fuente importante y económica de
proteínas, energía y micronutrientes. Específicamente en Ruanda el fríjol es un
cultivo de gran relevancia, alrededor de 8 millones de Ruandeses consumen fríjol y
se estima un consumo anual de 60 Kilogramos per capita. Este alimento provee el
84% de proteínas de leguminosas o 65% de todas las proteínas vegetales y
animales (y 32% de energía) consumidas. Adicional a esto, el fríjol es un cultivo
tradicional y de importancia cultural ya que se usa como ofrenda o regalo.
Sólo en África, cerca de 4 millones de hectáreas son cultivadas anualmente con una
producción de 2 millones de toneladas. A pesar de que en sus países la producción
es alta, debido a la gran demanda que existe de este alimento, se presentan
problemas fitosanitarios o ambientales que afectan el rendimiento de los cultivos.
3

Por todo lo anteriormente expuesto, y por su alta variabilidad genética, el fríjol
común P. vulgaris L se constituye en una especie de gran importancia dentro de
estudios de mejoramiento, lo que generó el interés por adelantar esta investigación
realizada en el Laboratorio de Caracterización de Germoplasma de Fríjol (LCGF),
ubicado en el CIAT, en torno al siguiente problema: El fríjol común (P. vulgaris L.),
como renglón importante de la economía y alimentación de la mayoría de países en
vía de desarrollo de África específicamente Ruanda, requiere la profundización en
estudios a nivel molecular que permitan caracterizar su diversidad genética con
miras a apoyar su mejoramiento nutricional.
El presente estudio tiene como objetivo: Caracterizar la diversidad genética de 355
genotipos de fríjol común provenientes de diferentes regiones de Ruanda (África)
mediante el uso de marcadores moleculares tipo microsatélites fluorescentes. El uso
de los microsatélites en estudios de diversidad es muy importante debido a que
ofrecen gran cantidad de información por su naturaleza multialélica, son
codominantes, tienen alto poder discriminatorio, son reproducibles, tienen herencia
mendeliana y presentan fácil detección dentro de sistemas automatizados. Adicional
a la caracterización de la diversidad se realizó un análisis del contenido de hierro y
zinc a partir de la técnica de Absorción Atómica.
En este trabajo se logró encontrar altos niveles de diversidad genética y de
contenidos de minerales para los genotipos en estudio, lo cual despierta la atención
debido a las necesidades alimentarias por las que atraviesa Ruanda, siendo esto un
paso importante para el inicio de programas que favorezcan la producción de la
especie en este país.
4

2 MARCO TEÓRICO Y REVISIÓN DE LITERATURA
El sustento teórico para la realización de esta investigación se basa en aportes de
autores reconocidos e investigaciones acerca de las generalidades de fríjol,
características de Ruanda, los cuales se contemplan a continuación:
2.1 GENERALIDADES DEL FRÍJOL
El fríjol común (Phaseolus vulgaris L.) es una de las especies más importantes en el
mundo pues hace parte del grupo de plantas en las cuales la humanidad encuentra
la principal fuente de proteína para su alimentación. La mayor parte de su
producción se presenta en los países de bajos recursos, como los pertenecientes a
los continentes Africano y Americano (Broughton et al., 2003). Sin embargo, esta
fuente alimenticia se ve amenazada por problemas que afectan la producción,
debido a proliferación de plagas y enfermedades que generan pérdidas del 10 y
100% (Beebe et al., 2000b).Esto hace del fríjol, una especie interesante para
estudios de diversidad enfocados hacia mejoramiento.
2.1.1 Características Taxonómicas y Botánicas
El fríjol común Phaseolus vulgaris L. es una planta leguminosa dicotiledónea. La
superfamilia Leguminosae, familia Fabaceae que comprende casi 10.000 especies
agrupadas en 643 géneros encontrados en diferentes ambientes y temperaturas. El
género Phaseolus comprende aproximadamente 35 especies, entre las que se
encuentran Phaseolus vulgaris, Phaseolus lunatus, Phaseolus coccineus, Phaseolus
polyanthus y Phaseolus acutifolius, las cuales corresponden a las cinco cultivadas
comercialmente (Betancour & Dávila, 2002; Fonnegra, 1996). El genero Phaseolus
se caracteriza por la presencia de la Faseolina, una proteína de almacenamiento en
el tejido cotiledonal de la semilla; determina la cantidad y calidad nutricional de las
proteínas en las semillas de frijol (Gepts et al., 1986); existen diferentes tipos de
faseolinas asociados con el tamaño de la semilla, por ejemplo la de tipo ‘S’ (Sanilac)
se relaciona con semilla pequeña, mientras que la ‘T’ (Tendergreen) y ‘C’ (Chile) con
mediana y grande (Islam et al., 2002b; Singh et al., 1988).
5

Como planta anual y herbácea se cultiva esencialmente para obtener y consumir las
semillas, que son las que más aporte nutricional hacen, al presentar alto contenido
de proteínas (22% en materia seca). En cuanto a la morfología, presenta un sistema
radicular que desarrolla raíces secundarias y terciarias, es superficial ya que el
mayor volumen de la raíz se encuentra en los primeros 20 cm de profundidad del
suelo; presenta nódulos distribuidos en las raíces laterales de la parte superior
radical, estos nódulos son colonizados por bacterias del género Rhizobium las
cuales fijan nitrógeno atmosférico (Debouck & Hidalgo, 1989).
El tallo es cilíndrico y sub-glabro o pubescente, puede ser verde, morado o rosado.
Tiene un incremento progresivo en la longitud de internodos y se pueden identificar
4 tipos de hábitos de crecimiento según la terminación apical del tallo y de las
ramas, dependiendo de si se forma un racimo o un meristema apical,
respectivamente (Betancour & Dávila, 2002):
Los dos primeros tipos corresponden a los arbustivos, TIPO I: determinado
arbustivo, sus ramas terminan en racimos; la inflorescencia desarrollada es terminal,
es decir detiene el crecimiento de la rama; carecen de la habilidad de trepar. TIPO
II: indeterminado arbustivo, tienen tallos cortos erectos de 30 a 50 internodos y
ramas; las plantas terminan en guías cortas y son semi-trepadoras (Beebe et al.,
2000b; Debouck & Hidalgo, 1989; Schoonhoven & Oswaldo, 1994). Los otros dos
son: TIPO III indeterminado postrado, con ramificación bien desarrollada y altura
superior a los 80cm; el hábito TIPO IV, se le conoce como indeterminado trepador,
el cual desarrolla la capacidad de torsión, sus ramas son poco desarrolladas,
pueden alcanzar alturas de más de dos metros (Schoonhoven & Oswaldo, 1994).
Las hojas son trifoliadas con pecíolo y raquis acanalado; la inflorescencia es
racimosa, en el caso del tipo determinado es terminal y para los indeterminados es
axilar; la flor es la típica de la familia Fabaceae es decir, con forma de mariposa,
bilateralmente simétrica, encerrada por bractéolas verdes, la corola es estándar, dos
alas y una quilla asimétrica, los colores de la colora son usualmente verde, blanco,
rosado o morado; el fruto es una vaina con dos valvas, las cuales provienen del
ovario comprimido; presenta placentación marginal (Beebe et al., 2000b; Debouck &
Hidalgo, 1989; Schoonhoven & Oswaldo, 1994).
6

La semilla puede ser redonda, esférica o arriñonada, de acuerdo con la variedad de
fríjol, al igual que el hilum y el micrópilo. La semilla no es endospermica sino
consiste en embrión con dos cotiledones simétricos, su germinación es epigea y
consume estos cotiledones que sirven de almacenamiento para el crecimiento. Las
hojas primarias son simples, opuestas, cordiformes y acuminadas y caen antes de
que la planta esté completamente desarrollada; mientras que las hojas secundarias
son compuestas, trifoliadas con venación reticular, pueden ser de forma ovalada a
triangular (Debouck & Hidalgo, 1989; Debouck & Tohme, 1988).
2.1.2 Origen, distribución y domesticación
El fríjol común Phaseolus vulgaris L. (2n=2x=22) fue domesticado en el Nuevo
mundo aproximadamente hace 7000 a 10000 años atrás El fríjol ha evolucionado a
través de su domesticación, desde las formas silvestres ancestrales como
Phaseolus vulgaris var. aborigineus [Burk] Baudet, una enredadera anual que se
distribuía en altitudes medias 1500-2000 msnm, en bosques claros o en las regiones
del neotrópico en un rango superior a 8000km, desde el norte de México hasta el
norte de Argentina (Beebe et al., 1997; Chrispeels & Savada, 2003; Koenig & Gepts,
1989).
Estudios bioquímicos asociados con los patrones geográficos (Koenig & Gepts,
1989), evidencias de proteínas de la semilla (Gepts & Bliss, 1986), rasgos
morfológicos (Singh et al., 1991b) y marcadores moleculares (Beebe et al., 2001;
Beebe et al., 2000b), incluyendo los microsatélites (Blair et al., 2007; Blair et al.,
2006; Díaz & Blair, 2006), han revelado que existen dos centros de diversidad
primarios, un origen Mesoamericano y un origen Andino. En fríjol silvestre, según
Koenig y Gepts (1989), Gepts y Bliss (1986), Beebe et al (1997), se ha hecho
referencia a un tercer acervo en el norte de los Andes. Un estudio de la colección
núcleo de CIAT por Islam (2002) quien evaluó 1072 accesiones, dio soporte a la
evidencia de la existencia de este otro acervo. Hasta ahora éste no ha sido
caracterizado como recurso potencial para el mejoramiento del fríjol y tampoco se
ha realizado una evaluación nutricional de concentraciones de sus elementos (Islam
et al., 2002b).
7

Estos centros de diversidad, actualmente llamados acervos, reflejan múltiples
eventos de domesticación dentro de las distintas poblaciones silvestres (Gepts &
Bliss, 1986). El acervo Mesoamericano se caracteriza por presentar genotipos con
semillas pequeñas a medianas, con faseolinas tipo ‘S’, ‘CH’, ‘Sb’, ‘Sd’ y ‘M’, mientras
que en el acervo Andino se encuentran semillas medianas a grandes con tipo de
faseolina ‘T’, ‘H’ y ‘C’ (Islam et al., 2004; Kami et al., 1995; Singh et al., 1991a).
Dentro de cada acervo se pueden distinguir razas, cuatro para Mesoamericano y
tres para Andino, las cuales han surgido debido a procesos de domesticación (Singh
et al., 1991a); para su clasificación y caracterización se han basado en las
diferencias morfológicas de la planta (Hábito de crecimiento), de la semilla (Tamaño,
forma, color) y a partir de estudios moleculares como Beebe et al 2000 quienes
utilizaron RAPD (Random Amplified Polymorphic DNA) y posteriormente Beebe et al
2001 con la técnica de AFLP (Amplified Fragment Length Polymorphism) o por
medio de microsatélites (Blair et al., 2007; Blair et al., 2006; Díaz & Blair, 2006;
Gomez et al., 2004).
Los genotipos del acervo Mesoamericano predominan en México, América central
(Guatemala, El Salvador, Honduras, Nicaragua y Costa Rica) y Brasil; se pueden
distinguir las razas Mesoamérica (M), Durango (D), Jalisco (J) y Guatemala (G).
(Beebe et al., 2000b; Broughton et al., 2003; Islam et al., 2004).
La raza Mesoamérica es común en México y América Central, se caracteriza por
tener un tamaño de semilla relativamente pequeño, por su adaptación a tierras bajas
y por presentar principalmente los hábitos de crecimiento Tipo II o III, y en algunos
casos Tipo IV. Las clases comerciales de esta raza incluyen semilla pequeña negra,
semilla roja pequeña y semilla blanca. La raza Durango está compuesta
principalmente por plantas con tipo de crecimiento III con hojas pequeñas, semillas
de tamaño medio y con adaptación a ambientes secos de tierras altas en México;
las clases comerciales incluyen pinto, grande del norte y rojo mexicano. La Raza
Jalisco, se encuentra en ambientes más húmedos de las tierras altas de México y
está compuesta principalmente por plantas trepadoras (Tipo IV), los genotipos
presentan tamaño medio de semilla (Beebe et al., 2000b). Y por último la Guatemala
8

formada por especies trepadoras en regiones húmedas montañosas (Islam et al.,
2004).
Análisis mediante AFLP (de las siglas en Inglés Amplified Fragment Length
Polymorphism) revelan que las poblaciones de fríjol silvestre en la zona Andina han
sido aisladas la una de la otra, resultando poblaciones discretas en Ecuador y el
norte de Perú; en el sur de Perú, Bolivia y el norte de Argentina (Tohme et al.,
1996). Con base en estudios morfológicos y criterios ecológicos, el acervo Andino
puede ser subdividido en tres razas; Raza Nueva Granada (NG), Perú (P) y Chile
(C). La primera representa a los genotipos con tamaño de semilla medio a grande y
con hábito de crecimiento arbustivo, e incluye la mayoría de cultivares comerciales
de semilla grande. La raza NG es la más cultivada dentro del acervo Andino,
prosperando principalmente en alturas medias de África y América; en ambientes de
tierras bajas de Brasil, México y el Caribe y en climas templados de Norte América y
Europa. A la raza Perú pertenecen las plantas trepadoras, la mayoría de las cuales
están adaptadas a tierras altas por encima de los 2000 msnm, mientras que la raza
Chile se característica por presentar habito de crecimiento tipo III, semilla de tamaño
medio, de forma redonda u ovalada, y usualmente colores pálidos que se
encuentran en latitudes altas en Turquía, Irán y China (Beebe et al., 2001).
Hallazgos arqueológicos en Perú y en el sur occidente de los Estados Unidos,
determinaron que el Nuevo mundo es el centro de origen del fríjol; sin embargo en la
actualidad este se encuentra distribuido mundialmente (Diaz, 2005a; Singh, 2001).
Los centros secundarios de diversidad están en África, Brasil, Europa, Medio
Oriente, así como América del Norte (Broughton et al., 2003). Es posible que el fríjol
halla sido introducido a África por los exploradores Árabes y Europeos y por los
comerciantes de Brasil y el sur de los Andes en los siglos XVI y XVII (Debouck &
Hidalgo, 1989; Musoni, 2006).
Ruanda es considerado como un centro secundario de diversidad de fríjol. Existe
gran diversidad de genotipos pertenecientes a los dos acervos; mezclas de plantas
de diferentes granos crecen normalmente en las fincas de este país. Algunos
materiales considerados como extintos en los centros de origen de América Latina
existen en Ruanda (Musoni, 2006).
9

La alta variedad de ambientes en los cuales se cultiva el fríjol ha llevado a
incrementar la variabilidad fenotípica, especialmente de caracteres como el hábito
de crecimiento, el tipo de grano, la fenología y la sensibilidad al fotoperiodo. Los
altos niveles de polimorfismo, la diversidad fisiológica y amplia distribución
geográfica hacen difícil el establecimiento de patrones de domesticación y relación
entre genotipos (Becerra & Gepts, 1994).
El fríjol común tiene varios centros posibles de domesticación en América latina. De
las especies conocidas del género Phaseolus, 5 de ellas se domesticaron durante la
época precolombina; el fríjol común fue de gran importancia en los imperios azteca
e inca siendo utilizado como elemento para el pago de tributos (Diaz, 2005a)
El progenitor del fríjol y los descendientes cultivados generalmente dan una
progenie fértil y viable, muestran diferencias contrastantes constituyendo así el
síndrome de la domesticación; los genotipos cultivados, muestran un hábito de
crecimiento más compacto comparado con los parentales silvestres, menos nodos
vegetativos, hojas de mayor tamaño e internodos más largos. En cuanto a las
semillas, éstas son más grandes y tienen menor pigmentación por antocianina.
Algunos genotipos cultivados no poseen sensibilidad a la longitud del día (su
progenitor solo florecía en días cortos), estos florecen más temprano (Koinange et
al., 1996). Dos de los atributos más importantes en el proceso de domesticación
son: pérdida de la habilidad de dispersión de semilla y la dormancia de la semilla
(fundamentales para la adaptación a cultivo) (Broughton et al., 2003). La
domesticación, fue un proceso rápido lo que puede indicar que la adaptación a las
condiciones ambientales cambiantes involucró genes con mayores efectos
genotípicos (Koinange & Gepts, 1992).
2.1.3 Cultivo, producción y Usos
El fríjol es extremadamente diverso en términos de métodos de cultivo, usos, rangos
de ambientes a los cuales ha sido adaptado y variabilidad morfológica. Puede
cultivarse en terrenos ubicados desde la altura del nivel de mar hasta 3000
m.s.n.m, es cultivado en monocultivos, en asociaciones o en rotaciones. Los frijoles
son consumidos como granos maduros, también como semilla inmadura, tanto
como vegetales, hojas y vainas (Broughton et al., 2003).
10

Su recurso genético existe como un arreglo complejo de acervos genéticos mayores
y menores, razas y tipos intermedios con introgresión ocasional entre tipos salvajes
y domésticos. El cultivo de fríjol es entonces uno de los que mejor se adapta a
diferentes ambientes; la producción mundial de fríjol alcanza los 23 millones de
toneladas, de los cuales 7 millones se producen en países tropicales de
Latinoamérica y África (Pastor-Corrales & Schwartz, 1994).
Las condiciones donde se produce el fríjol son muy variables, van desde zonas muy
húmedas a zonas semi-desérticas como el noreste de Brasil, el centro y el altiplano
nororiental de México, el valle de Rift del oriente de África y entre las zonas
montañosas de Estados Unidos (Singh, 2001). Sin embargo, la producción de fríjol
está reducida a pequeños terrenos con agricultores de bajos recursos y en
ambientes no óptimos para la especie con sequías estacionales (Broughton et al.,
2003; López et al., 1985). El fríjol es uno de los principales cultivos, después del
maíz, en el centro y este de África. Cerca de 4 millones de hectáreas de fríjol son
cultivadas anualmente en éste continente, con una producción de dos millones de
toneladas (Chrispeels & Savada, 2003).
2.1.4 Composición Química y Calidad nutricional
2.1.4.1 Composición Química
Al 11 % del contenido de humedad, la semilla tiene 17-30% de proteína,
generalmente bajos en amino ácidos azufrados (metionina y cisteína) pero alto en
lisina. También tiene 57.8% del complejo de azucares, 1.6% de grasa, 4% de fibra y
grandes cantidades de ácido fólico; en cuanto a los micronutrientes esenciales,
presenta cerca de 34-89 ppm de hierro y 21-54 ppm de zinc(Beebe et al., 2000a;
Musoni, 2006). Las hojas frescas son ricas en hierro y vitamina A. En África del este
ESCABREN identificó ciertas variedades como ‘Ngwinurare’, ‘Gofta’, ‘Maharagi
Soya’, con altos contenidos de minerales en hojas y semillas (Kimani et al., 2000)
2.1.4.2 Calidad Nutricional
Más de 300 millones de personas basan su dieta diaria en la leguminosa comestible
más importante del mundo, el Fríjol común, por ende hace una contribución
11

importante a la nutrición (Beebe et al., 2000a). Los fríjoles proveen proteína a la
dieta y son alimento de gran importancia por complementar cereales que son fuente
primaria de carbohidratos; como en otras leguminosas, los fríjoles contienen
grandes cantidades de hierro y otros minerales (Beebe et al., 2000b; Broughton et
al., 2003).
El valor nutricional del fríjol radica en que su semilla es una fuente importante de
calorías, vitaminas, proteínas, carbohidratos y grandes concentraciones de
minerales esenciales, el contenido nutricional de la semilla se puede detallar en la
tabla 1. En América Latina y en el continente Africano, el fríjol es utilizado
totalmente para consumo humano y mundialmente el fríjol es conocido como la
“carne de los pobres”, por ser fuente muy importante de proteína y de bajo costo
(MINAGRI/MINEDUC, 1993). El contenido de proteína del fríjol varía según la
variedad pero en general es de un 24% superando a la papa y al maíz (Betancour &
Dávila, 2002).
Tabla 1. Contenido nutricional de semilla de fríjol. Contenido total en semilla y por 100g de
semilla (Geil y Anderson, 1994 En: (Sandoval, 2006).
Contenido Total en Semilla
Contenido por 100g de semilla
Calorías (kcal)
Proteínas (%)
Carbohidratos (%)
Fibra (%)
Grasa (%)
21-25
60-65
3-7
0.8-1.5
110-143
Vitaminas (% RDA)
Ácido fólico 30
Tiamina 25
Piridoxina 10-12
Niacina 10
Riboflavina 10
Minerales (% RDA) Hierro 29-55
Fósforo 20-25
Calcio 10
Zinc 10
% RDA: porcentaje que aporta de acuerdo a los requerimientos mínimos recomendados
12

El hierro es uno de los minerales vitales en la dieta del hombre, ya que participa en
importantes procesos de oxido-reducción, está presente en numerosas enzimas
involucradas en el mantenimiento de la integridad celular, tales como las catalasas,
peroxidasas y oxigenasas y a su vez forma parte estructural y/o funcional de
algunas enzimas como la hemoglobina. La forma de absorción en el organismo
depende de la forma química presente en la dieta, ya sea como hierro hemo (en
alimentos de origen animal) o hierro inorgánico (en alimentos de origen vegetal), en
este último caso su absorción no es proporcional a su contenido ya que se ve
afectada por factores químicos presentes en los alimentos, tales como oxalatos,
taninos y fitatos (Barrios et al., 2000).
El zinc es importante para la formación de enzimas, se encuentra directamente
involucrado con el crecimiento y desarrollo, la insuficiencia de este mineral impide el
crecimiento normal; adicional a esto se ha observado su importancia en el
mantenimiento de la función y la estructura de las biomembranas; inhiben el daño
oxidativo al atrapar radicales libres a través de su enlace con la metalotioneina o
bien mediante la unión a las membranas en sitios que pueden ser ocupados por
metales con potencial redox, estabiliza los grupos “tio” (O´Dell, 1981 En: (Moreno,
2007).
2.2 SITUACIÓN DEL FRÍJOL EN ÁFRICA
En términos de producción y consumo, el fríjol común es la leguminosa de grano
más importante en el oriente, centro y sur de África, mas de 100 millones de
personas en este continente lo consumen; se estima que éste es el segundo recurso
de proteína, y el tercero de calorías más importante de la región; los agricultores
cultivan muchas variedades de fríjol. Típicamente 6 cultivares cuentan para el 95%
de la producción en muchas comunidades, pero la diversidad es más alta en las
regiones de los grandes lagos y áreas adyacentes a Uganda, donde las mezclas
son la norma.
13

Las mezclas son también importantes en parte de Malawi, Mozambique y Tanzania
(Martin & Adams, 1987). Sin embargo, hay una tendencia a producir cultivares más
orientadas hacia el mercadeo, en poblaciones urbanas que demandan mayor
uniformidad para el consumo de fríjoles (Broughton et al., 2003; CIAT, 2005a).
Millones de hectáreas son cultivadas en más de 20 países de África (Tabla 2), en
estos lugares los agricultores de bajos recursos e ingresos, producen fríjoles
principalmente en escala pequeña; las mujeres siembran mucho más que los
hombres. Gran cantidad de cultivos se pierden debido a enfermedades, pestes de
insectos, sequía, baja fertilidad en los suelos u otros factores de estrés abiótico
(David & Sperling, 1999).
Gran cantidad de variedades de fríjol se cultivan en África, siendo notorio en la
diversidad del tipo de semilla y su adaptación. La elección de las variedades más
populares para el mercado, pueden estar dictadas por la resistencia a variabilidad
climática y a condiciones agronómicas, en algunos casos dicha elección puede
producirse por el tamaño grande de la semilla, pero la mayoría de agricultores
especialmente del oriente y del centro de África hacen mezclas de todas las formas
y colores de las semillas (Broughton et al., 2003; Musoni, 2006).
Las semillas pequeñas y medianas suelen ser seleccionadas con mayor frecuencia
debido a que se puede sembrar mayor área y además estas confieren mayor
resistencia a factores ambientales y a enfermedades (Comunicación Personal B.
Lewis investigador programa fríjol Ruanda CIAT, 2007).
El comercio de la semilla de esta planta es muy importante, y puede ocurrir
localmente, entre ciudades o informalmente entre países, usualmente se establecen
rutas de comercio hacia áreas urbanas dentro del mismo país de producción; las
hojas de esta planta también hacen parte del comercio(David & Sperling, 1999).
14

Tabla 2. Áreas de producción de Fríjol en 20 países de África, modificado de “Atlas of
common bean production in Africa” (Wortmann et al, 1998).
Región Países Área (%) Área (ha x 10-3)
Africa Oriental- tierras altas y alturas medias
Burundi, DR Congo, Etiopia, Kenia, Ruanda,
Tanzania, Uganda
62 2490
Africa del Sur Lesotho, Madagascar, Malawi, Mozambique, Sur
África, Swaziland, Tanzania, Zambia,
Zimbabue
31 1290
Africa Occidental Angola, Cameroon, Cape Verde, Togo
3 135
Tierras bajas- estación de invierno
Algeria, DR Congo, Egipto, Mali, Malawi, Mauritius, Morocco,
Nigeria, Sudan, Tunisi
4 200
TOTAL 100 4025
Se considera que en el oriente y sur de África el consumo de fríjol es mayor que el
de Latinoamérica, las estadísticas muestran que 66kg por persona son consumidos
en algunas zonas rurales de Kenia mientras que en Ruanda y Burundi el consumo
promedio nacional excede 40kg por persona por año (Broughton et al., 2003). Los
dos acervos principales de Phaseolus vulgaris L. están representados en África. El
61% de las cultivares presentes corresponden a semillas grandes típicas del acervo
Andino; el resto pertenecen al acervo Mesoamericano, caracterizado por las
semillas pequeñas o medianas (CIAT, 2005b).
2.2.1 Generalidades Ruanda
Ruanda se encuentra localizado en una región conocida como África Subsahariana
(término utilizado para describir a los países del continente africano ubicados al sur
del desierto del Sahara y que no forman parte de la región conocida como Noráfrica)
específicamente en la parte oriental de África Central (MINAGRI/MINEDUC, 1993).
El país tiene una población de 8.6 millones de personas viviendo en un área de
26,368 Km2, haciendo de Ruanda uno de los países más densamente poblados en
África, 322 habitantes por Km2 (ISAR, 2000). En la Figura 1 se muestra el mapa de
organización política de Ruanda.
15

L
m
9
l
(
q
c
p
s
L
c
c
p
d
s
Fig
La mayoría
montañosa.
90%), cuya
los twa (1%
(ISAR, op c
que crezca
calcula que
país ha enc
su gente (Ng
La agricultur
como el pri
contribuye e
principal pro
de las gana
segundo pro
gura 1. Mapa
de los hab
La població
lengua es e
%), pueblos p
it). En la act
a 11.4 millo
la població
contrando m
girumwami,
ra aporta el
incipal recu
en un 36.6%
oducto de ex
ancias nacio
oducto más
a de Ruanda,
bitantes vive
ón está form
el bantú; los
pigmeos y p
tualidad la t
ones de per
ón urbana se
uchos retos
1992)
91.1% de lo
rso de crec
% al PIB (P
xportación e
onales, mie
importante d
16
organización
en en núcleo
mada por tres
s tutsis (9%)
presuntamen
tasa de crec
rsonas en e
e duplique y
s para satisf
os empleos
cimiento eco
Producto Int
es el café, e
entras que e
de exportac
n política (Uni
os familiares
s grupos étn
), destacado
nte habitante
cimiento anu
el año 2010;
y que la rura
facer las nec
para la pobl
onómico. El
terno Bruto)
l cual en 19
en el mism
ión, aportó u
tedNations, 2
s dispersos
nicos: los hu
os criadores
es originales
ual es de 2.9
; para ese m
al crezca a 9
cesidades a
lación activa
l sector de
) (Ferris et
997 contribu
mo periodo e
un 24% de l
2007)
por la regi
utus (cerca d
de ganado
s de la regi
9%, se espe
mismo año
9.8 millones;
alimenticias
ón
del
, y
ón
era
se
el
de
a y permane
la agricultu
al., 2002).
yó en un 52
el té como
las ganancia
ece
ura
El
2%
el
as;

sin embargo es necesario tener en cuenta que más del 40% de la comida es
importada (ISAR, op cit). Ruanda es un país con menor PIB que otros en áfrica,
pero desde la guerra civil de 1994 ha comenzado a recuperarse económicamente
(Ferris et al., 2002; Sperling, 2001), en la Tabla 3 se muestran los principales
indicadores poblacionales de Ruanda y de África Subsahariana, teniendo en cuenta
el porcentaje de población activa dentro de cada sector económico.
Tabla 3. Comparación de los indicadores poblacionales y económicos de Ruanda y África
Sub-Sahariana (ISAR, 2000).
Indicador Ruanda África Subsahariana Tamaño poblacional 7.7 millones 596 millones Densidad Poblacional 303 por km2 26 por km2 . Población Activa:
• Sector Agrícola • Sector Industrial • Sector de
servicios
• 91.1 % • 1.7% • 7.2 %
• 70.0 % • 7.5 % • 22.5 %
PIB US$ 240 US$ 490
2.2.2 Fríjol en Ruanda
El fríjol común (Phaseolus vulgaris L.) puede ser llamado una comida casi perfecta,
por ser rico en proteínas carbohidratos, vitaminas y minerales como hierro, zinc y
ácido fólico, componentes esenciales para el crecimiento y desarrollo del cuerpo
humano y para el buen mantenimiento de la salud y desarrollo cognoscitivo (Ferris
et al., 2002; ISAR, op cit).
Los fríjoles son el principal componente en la nutrición de la vida de los Ruandeses;
la mayoría de familias consumen al menos una vez al día esta leguminosa y no sólo
en la forma más común como es el grano seco sino también utilizan en sus platos
las hojas, las vainas verdes y las semillas verdes. El cultivo del fríjol es el más
tradicional y de mayor importancia cultural, con frecuencia se usa como ofrenda o
regalo (Sperling, 2001).
17

A pesar de su importancia se puede reconocer algunos problemas en la producción
del fríjol y en gran parte se debe a que las tierras destinadas para los cultivos no
presentan procesos de rotación o tratamientos adecuados que ayuden a mejorar la
calidad del suelo. Adicional a esto el suelo carece de nutrientes esenciales tales
como nitrógeno, fósforo y potasio (Kelly et al., 2001). Algunos suelos son ácidos,
contribuyendo a la toxicidad de elementos minerales como aluminio y manganeso.
Los problemas bióticos también son de resaltar, dentro de los cuales se encuentran
enfermedades por patógenos como hongos, bacterias y virus, que atacan hojas,
tallos, raíces y el grano (Ferris et al., 2002; ISAR, 2000).
Las pérdidas se estiman hasta 305,700 toneladas por año en África oriental
(Musoni, 2006). Las enfermedades (mancha angular de la hoja, antracnosis,
pudrición de raíz) que atacan al fríjol causan pérdidas anuales de 761,900 toneladas
en el oriente de África (Wortmann et al., 1999). En Ruanda estas enfermedades
causan pérdidas de 219,575 toneladas, equivalente a 89 millones de dólares por
año (Musoni, op cit).
2.2.2.1 Producción y consumo
La producción de fríjol en Ruanda fluctuaba entre 150mil y 200mil toneladas al inicio
de la década de los 70’s; a partir del año 1974 hubo un rápido incremento que llegó
a tener su máximo valor en año de 1985 cuando se produjo más de 300mil
toneladas; en el periodo comprendido entre 1998 y 2000 se encontró un total de
200mil y 300mil toneladas. Las epidemias de la enfermedad de pudrición de raíz y
la guerra civil contribuyeron a la caída en la producción a mediados de los 90’s
(Sperling, 2001); durante el periodo del 2000 al 2001 la producción tuvo un
incremento, sin embargo los siguientes años hasta el 2004 por causa de problemas
climáticos (aumento de lluvias) la producción se tuvo una amplia reducción
(Comunicación Personal L. Butare investigador programa fríjol Ruanda CIAT,
2007). La figura 2 muestra las fluctuaciones de la producción anual de frijol entre
1979 y el 2004.
18
L
d
m
p
e
2
d
c
l
c
a
(
L
q
e
e
Figura
Los frijoles s
de la tierra
maíz, banan
por encima
encontrar es
2001). La pr
de estacion
consumo an
leguminosas
consumidas
animales, es
(Kelly et al.,
La malnutric
que la salud
en mujeres
encuentran
a 2. Produccióón anual, en Toneladas, dde Fríjol en Ruuanda (Musonni, 2006)
son cultivad
cultivada; s
nos, tubércu
de tres m
stas mezcla
roductividad
nes de inve
nual de 60 K
s o 65% de
s; ofrece un
specialment
2001; Muso
dos por el 95
se pueden e
los y sorgo,
mezclas dur
as tanto en e
es de 800k
stigación (F
Kg per capit
todas las pr
n recurso a
te dentro de
oni, 2006).
5% de los c
encontrar en
los campes
rante la mis
el consumo
kg/ha, un ter
Ferris et al.
ta; este alim
roteínas veg
alternativo p
e la població
campesinos
n monocultiv
sinos individu
sma época
como para
rcio del rend
, 2002; ISA
mento prove
etales y ani
para los ali
ón rural que
Ruandeses
vos o cultivo
ualmente pu
de siembr
la producci
dimiento pos
AR, 2000). S
e el 84% de
males (y 32
imentos pro
e es la meno
sobre el 30
os mixtos c
ueden manej
ra; es com
ión (Sperlin
siblemente s
Se estima
e proteínas
% de energ
ovenientes
os privilegia
0%
on
jar
ún
ng,
on
un
de
ía)
de
da
2.2.2.2 PProblemática Nutricionaal
ción es reco
d enfrenta en
y niños de
las de mic
onocida en la
n muchos se
e bajos recu
ronuetriente
a actualidad
ectores de l
ursos. Dentr
es (aún más
d como uno
a población
ro de las p
s prevalente
de los retos
Africana, p
rincipales d
e que las d
s más grand
articularmen
deficiencias
e proteínas
es
nte
se
s o
19

carbohidratos), las cuales incluyen las de hierro, zinc, vitaminas y aminoácidos
azufrados (Comunicación Personal M.Blair mejorador de fríjol CIAT, 2007). La
principal causa de esto es que las dietas son abundantes en energía pero pobres en
proteínas, minerales y vitaminas; dicha situación adquiere mayor complejidad en
estos países en donde la mayoría de personas no tienen acceso a los alimentos
provenientes de animales ya que son muy costosos y además el conocimiento del
valor nutricional del alimento que se encuentra a su disponibilidad es limitado (CIAT,
2005b; Donovan & Bailey, 2005).
Debido a este desconocimiento muchas personas de bajos recursos en África basan
su alimentación en los cereales, papas blancas y yuca, estos alimentos
generalmente presentan bajos niveles de micronutrientes (CIAT, 2005b). La
deficiencia en hierro afecta a millones de individuos durante todo su ciclo de vida,
en especial a los lactantes, niños pequeños y las mujeres embarazadas, pero
igualmente a los niños mayores, los adolescentes y las mujeres en edad
reproductiva; los organismos vivos requieren hierro para que sus células funcionen
normalmente (Donovan & Bailey, op cit; OPS, 2002).
El hierro es necesario para el desarrollo de tejidos vitales —incluido el cerebro— y
para transportar y almacenar oxígeno en la hemoglobina y la mioglobina muscular.
La anemia ferropénica es la forma grave de carencia de hierro, puede dar lugar a
una baja resistencia a infecciones, limitaciones en el desarrollo psicomotor y la
función cognoscitiva en los niños, bajo rendimiento académico, así como fatiga y
una baja resistencia física y bajo rendimiento en el trabajo (OPS, 2002). La
prevalencia de esta anemia por falta de hierro varia en África desde un 8% de la
población en Etiopía, 67% en Tanzania hasta el 69% en Burundi (CIAT, 2005b).
Dietas deficientes en hierro frecuentemente son deficientes en zinc. Las
consecuencias de la deficiencia de zinc pueden incluir crecimiento y desarrollo
anormal, inmadurez sexual, complicaciones en el embarazo, mortalidad maternal e
infantil, baja defensa inmunológica, por lo que hace necesario su consumo
especialmente en personas que han sido diagnosticadas positivas para VIH/SIDA
(Donovan & Bailey, 2005; Islam et al., 2002a). Sin embargo, la deficiencia de zinc es
20

un problema que hasta ahora se empieza a evidenciar como un serio tema de salud
pública en los países de África (CIAT, 2005b).
2.2.2.3 Estrategias para combatir la problemática nutricional
Existen tres estrategias de intervención que pueden contribuir en la prevención y
disminución del problema de deficiencia de micronutrientes en África:
abastecimiento con productos farmacéuticos a poblaciones vulnerables
(micronutrientes), la fortificación de alimentos y el mejoramiento de la dieta
(Biofortificación) (OPS, 2002).
La primera estrategia, es efectiva para lograr un acceso con facilidades médicas en
grupos vulnerables. Sin embargo solo sería efectiva en un grupo pequeño y requiere
de un gran capital y una elaborada y costosa red de distribución; por lo que dejaría
por fuera a muchos grupos de riesgo que no podrían recibir dicho abastecimiento
(CIAT, 2005b). La fortificación de alimentos ha tenido un grado limitado de éxito en
África debido a que la industria de alimentos se encuentra subdesarrollada y carece
de una efectiva legislación. Hasta el momento los programas de fortificación se
encuentran operando en dos países de África oriental y central: Kenya y Uganda.
Este programa es más efectivo en áreas urbanas, dejando a un lado a comunidades
pobres del área rural (HarvestPlus, 2006).
El mejoramiento de la dieta es probablemente la estrategia más efectiva y sostenible
para reducir la deficiencia de micronutrientes en África. Ésta ayuda a incrementar la
disponibilidad de alimentos, permite que exista consumo de alimentos ricos en
minerales en poblaciones de alto riesgo (CIAT, 2005b). La Biofortificación
(incremento de concentraciones de los principales micronutrientes), se logra con la
creación de variedades superiores agronómicamente hablando, es decir con valor
nutritivo mejorado o variedades que presenten características más atractivas para
los agricultores, tales como tolerancia a sequía o baja fertilidad de suelo. La
Biofortificación del fríjol común podría producir los mejores resultados en áreas
donde el fríjol es el suplemento de una proporción significativa de nutrientes en la
dieta. Estas áreas incluyen partes de África Oriental, Central y del Sur; y América
Central y Brasil (HarvestPlus, 2006).
21
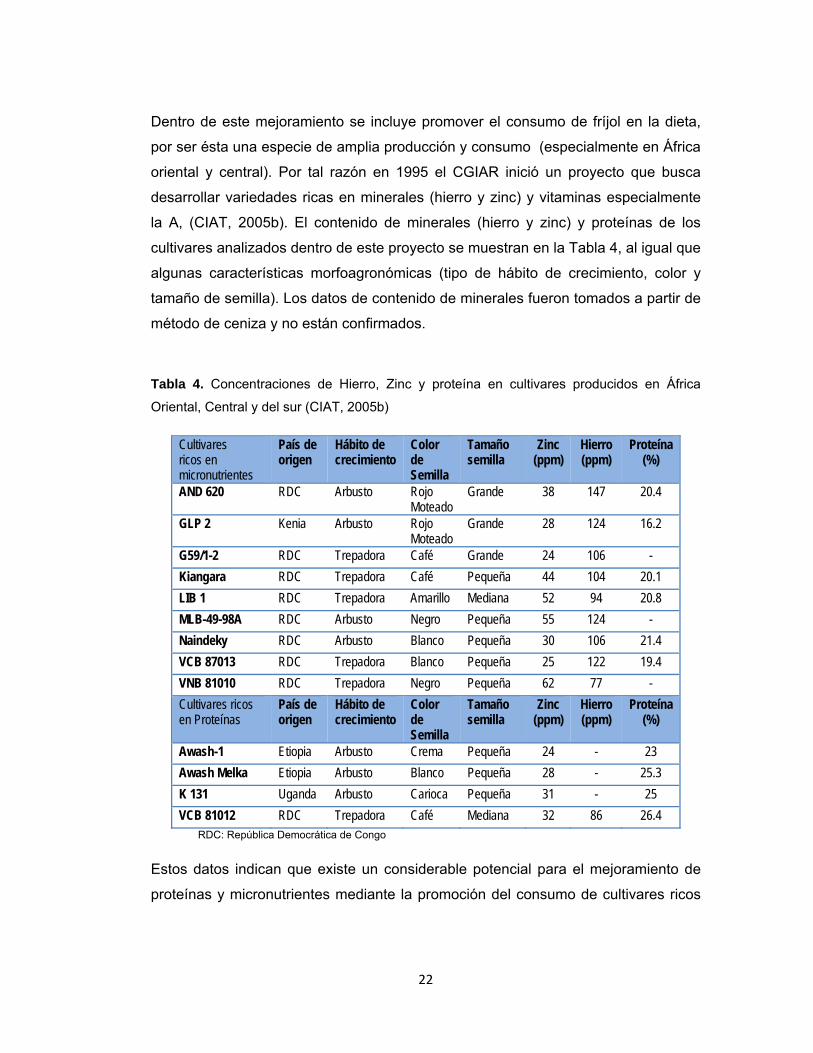
Dentro de este mejoramiento se incluye promover el consumo de fríjol en la dieta,
por ser ésta una especie de amplia producción y consumo (especialmente en África
oriental y central). Por tal razón en 1995 el CGIAR inició un proyecto que busca
desarrollar variedades ricas en minerales (hierro y zinc) y vitaminas especialmente
la A, (CIAT, 2005b). El contenido de minerales (hierro y zinc) y proteínas de los
cultivares analizados dentro de este proyecto se muestran en la Tabla 4, al igual que
algunas características morfoagronómicas (tipo de hábito de crecimiento, color y
tamaño de semilla). Los datos de contenido de minerales fueron tomados a partir de
método de ceniza y no están confirmados.
Tabla 4. Concentraciones de Hierro, Zinc y proteína en cultivares producidos en África
Oriental, Central y del sur (CIAT, 2005b)
Cultivares ricos en micronutrientes
País de origen
Hábito de crecimiento
Color de Semilla
Tamaño semilla
Zinc (ppm)
Hierro (ppm)
Proteína (%)
AND 620 RDC Arbusto Rojo Moteado
Grande 38 147 20.4
GLP 2 Kenia Arbusto Rojo Moteado
Grande 28 124 16.2
G59/1-2 RDC Trepadora Café Grande 24 106 - Kiangara RDC Trepadora Café Pequeña 44 104 20.1 LIB 1 RDC Trepadora Amarillo Mediana 52 94 20.8 MLB-49-98A RDC Arbusto Negro Pequeña 55 124 - Naindeky RDC Arbusto Blanco Pequeña 30 106 21.4 VCB 87013 RDC Trepadora Blanco Pequeña 25 122 19.4 VNB 81010 RDC Trepadora Negro Pequeña 62 77 - Cultivares ricos en Proteínas
País de origen
Hábito de crecimiento
Color de Semilla
Tamaño semilla
Zinc (ppm)
Hierro (ppm)
Proteína (%)
Awash-1 Etiopia Arbusto Crema Pequeña 24 - 23 Awash Melka Etiopia Arbusto Blanco Pequeña 28 - 25.3 K 131 Uganda Arbusto Carioca Pequeña 31 - 25 VCB 81012 RDC Trepadora Café Mediana 32 86 26.4
RDC: República Democrática de Congo
Estos datos indican que existe un considerable potencial para el mejoramiento de
proteínas y micronutrientes mediante la promoción del consumo de cultivares ricos
22

en estos nutrientes. Otros cultivares pueden ser mejorados a través de cruzas y
programas de Fitomejoramiento.
2.2.3 Perspectivas del cultivo de Fríjol
Se requiere la adquisición y la adopción de nuevas tecnologías para el cultivo de
fríjol, con el fin de mejorar su producción y elevar el consumo y comercio. Las
investigaciones se enfocan hacía la creación de nuevas tecnologías generadas por
los estudios en patología, agronomía y aspectos socio-económicos. Algunos de los
avances se han centrado en generar nuevas variedades por programas de
entrecruzamiento e investigaciones en colección (Ferris et al., 2002; ISAR, 2000).
Un avance adicional es una caracterización molecular de colecciones con el fin de
conocer la base genética del germoplasma de estudio que permitan la adecuada
selección de genotipos para cruzar con fuentes de alto contenido de minerales,
proteínas o vitaminas en semilla.
2.3 DIVERSIDAD GENÉTICA Y ANALISIS MOLECULAR MEDIANTE
MICROSATLITES
2.3.1 Marcadores Moleculares
Los marcadores moleculares son definidos como “todo y cualquier fenotipo
molecular oriundo de la expresión de un gen o estado alélico” (Ferreira &
Grattapaglia, 1998); son herramientas que permiten conocer la variabilidad genética
de un germoplasma. Existen diferentes tipos de marcadores: morfológicos, se
refieren a rasgos fenotípicos; bioquímicos, incluyen variantes alélicas de enzimas
llamadas isoenzimas; y marcadores de ADN que revelan sitios de variación del
ADN. Las principales desventajas de los marcadores bioquímicos y morfológicos es
que ellos están limitados en número y son influenciados por los factores ambientales
o de la etapa de desarrollo de la planta. Sin embargo estos han sido de gran utilidad
para los mejoradores (Winter & Kahl, 1995).
Los marcadores de ADN son el tipo de marcador más ampliamente usado debido a
su abundancia. Ellos surgen de diferentes clases de mutaciones de ADN tales como
23

mutaciones por sustitución (mutaciones puntuales), rearreglos (inserciones o
deleciones) o errores en la replicación de tandems de repetición del ADN. Estos
marcadores son selectivamente neutrales ya que ellos usualmente están localizados
en regiones no codificantes del ADN. Dentro de los usos de este tipo de marcadores
se encuentran, la elaboración de mapas de ligamiento y la evaluación del nivel de
diversidad genética (Collard et al., 2005).
Los marcadores moleculares pueden clasificarse en tres grupos de acuerdo a su
fundamento: Marcadores basados en la hibridación del ADN, marcadores basados
en la secuenciación de fragmentos de ADN y marcadores basados en la
amplificación de fragmentos mediante la Reacción en Cadena de la Polimerasa
(PCR) (Ferreira & Grattapaglia, 1998).
Estos últimos son generalmente menos costosos y revelan mayores cantidades de
polimorfismo. La PCR está diseñada para amplificar ADN en procedimiento cíclico y
automatizado el cual resulta en un incremento exponencial en la cantidad de una
secuencia específica del ADN. La selección del fragmento de ADN para la
amplificación es el resultado del alineamiento, en la cual un “primer” o cebador (5 a
30 bases de largo) se une a una cadena simple de ADN genómico presentada en la
reacción. El complejo primer-ADN se convierte en el punto de inicio para la
replicación de la secuencia de ADN adyacente por la intervención de una
polimerasa termoestable en la reacción de esta mezcla (Reisch, 1998).
Uno de los marcadores más importantes basados en PCR son los microsatélites,
fundamentados en el descubrimiento de secuencias simples repetidas (SSR) en el
genoma; también se puede llamar ocasionalmente sitios de microsatélite de
secuencia etiquetada (STMS) o polimorfismo de repeticiones de secuencia simple
(SSRP) (Hajeer et al., 2000; Reisch, 1998). Estos hacen parte de los marcadores
que permiten visualizar las diferencias genéticas entre individuos, organismos o
especies. Generalmente ellos no representan genes “blanco” pero, sí actúan como
señales o banderas. Estos marcadores están localizados cerca de los genes
“blanco” y no afectan el fenotipo del rasgo de interés porque ellos están
simplemente localizados cerca de los genes que controlan el rasgo. Todos los
24

marcadores genéticos ocupan una posición específica dentro de los cromosomas
(como los genes), llamados “loci” (en singular locus) (Collard et al., 2005)
Los microsatélites son repeticiones cortas en serie cuya secuencia básica tiene una
longitud entre 1 y 10 pb, los más típicos de 2 a 4 pb. Son altamente variables y
están distribuidos por igual en todo el genoma. Este tipo de ADN repetitivo es
común en organismos eucariotas, y el número de unidades repetidas varía
ampliamente entre los organismos, hallándose en algunos hasta 50 copias o más de
la unidad repetida. Para identificar estos polimorfismos, se construyen cebadores o
primers para la amplificación mediante PCR de la región del ADN que flanquea el
microsatélite. Las regiones adyacentes a los microsatélites tienden a conservarse
dentro de las especies, aunque a veces se conservan también en niveles
taxonómicos mayores (Hajeer et al., 2000)
La variación en tamaño de los productos de la PCR para un microsatélite se debe a
las diferencias en el número de las unidades repetidas en el locus. El polimorfismo
de los microsatélites es generado por la pérdida o ganancia de repeticiones, aunque
se cree que se deba en mayor medida a la ganancia. Éste fenómeno no es conocido
completamente, pero se piensa que esta expansión es debida a procesos de
mutación durante la replicación (Roizès, 2000).
Los polimorfismos de SSR se pueden visualizar mediante electroforesis en geles de
agarosa o de poliacrilamida. Los alelos del microsatélite se detectan usando
diversos métodos: tinción con bromuro de etidio, nitrato de plata, radioisótopos o
fluorescencia. Si se usan cebadores marcados con fluorescencia, y los productos
son suficientemente diferentes en tamaño y no se sobreponen, se pueden generar
varios productos de forma simultánea, lo que aumenta enormemente la eficiencia de
estos marcadores (Dean et al., 1999).
Debido a que estos marcadores presentan grandes ventajas fueron seleccionados
para este estudio. Dentro de estas características se encuentran: alto poder
discriminatorio, ofrecen gran cantidad de información por su naturaleza multialélica,
son codominantes, reproducibles, tienen herencia mendeliana, tienen relativa
abundancia con una cobertura uniforme del genoma y presentan fácil detección
25

dentro de sistemas automatizados (Ferreira & Grattapaglia, 1998; Gaitan-Solis et al.,
2002).
2.3.2 Microsatélites analizados mediante fluorescencia y Análisis
secuenciador automático
Las soluciones que sean altamente eficientes y a bajo costo, son esenciales en la
actualidad en todos los campos de la investigación. La implementación de nuevas
tecnologías y metodologías de trabajo proveen dichas soluciones; tal es el caso de
los secuenciadores analíticos de ADN, los cuales permiten el estudio de
microsatélites de una forma más eficiente, y vienen acompañados del uso de
software que aumenta la precisión, sensibilidad, resolución y eficiencia en el
genotipaje (Joe et al., 2004).
Los microsatélites pueden ser marcados con etiquetas fluorescentes (fluorocromos)
de diferentes colores por ejemplo verde, azul, amarillo y rojo, y posteriormente
detectados automáticamente en secuenciadores de ADN; este método es
económico y eficiente. Se pueden realizar combinaciones (multiplex) de marcadores
en un mismo panel, lo cual puede incrementar la eficiencia ya que tiene en cuenta
muchos fragmentos de ADN polimórficos y representa diferentes loci a lo largo del
genoma.
En los paneles, los marcadores etiquetados con el mismo fluorocromo deben tener
diferente tamaño, mientras que los marcadores que se sobrelapan en tamaño deben
ser diferenciados mediante el uso de colores diferentes. Muchos marcadores
pueden ser mezclados después de la amplificación o bien amplificados juntos en
una reacción común de PCR (Blair et al., 2002). Estos productos de PCR son leídos
por un secuenciador analítico de ADN, por ejemplo el ABI serie 3730xl.
El procesamiento de la información dentro del software funciona como se muestra
en la figura 3. Los datos de la muestra leída mediante el secuenciador analítico de
ADN, son convertidos en un archivo y éste es auto-analizado directamente por un
software que interpreta el lenguaje análogo de las lecturas que hace el
26

secuenciador, realiza la identificación de picos y tamaños. Este software combina la
información del archivo de la muestra con análisis preconfigurados y calibraciones
estándar; las muestras procesadas pueden ser vistas, editadas y analizadas en una
ventana generada por el software (Joe et al., 2004).
Método de
análisis
Detección de pico
Confirmación de tamaño
Etiquetado de tamaño
Asignación de Auto‐Bin
Etiquetado de Alelo
Tabla de genotipo
Figura 3. Sistema de flujo de la información obtenida a partir del archivo de la muestra
dentro del software de auto-análisis. Modificado de (Joe et al., 2004).
2.3.3 Estudios de diversidad genética en Fríjol común
Debido a la importancia económica y nutricional del fríjol se ha llevado a cabo gran
cantidad de estudios de diversidad genética en distintas partes del mundo,
principalmente en el continente americano por ser centro de origen. Los estudios en
el continente Africano se presentan en menor número pero han tomado mayor
fuerza, al ser este continente un centro secundario. Las metodologías varían desde
descriptores morfológicos, bioquímicos (isoenzimas y faseolinas) o técnicas
moleculares con marcadores de ADN.
Gepts & Bliss en 1986 estudiaron material silvestre y mostraron la presencia de
faseolina (principal proteína de almacenamiento de la semilla de fríjol) tipo ‘S’,
además un nuevo patrón denominado ‘CH’ (Chibcha); mientras que para materiales
cultivados presentaron ‘S’, ‘T’ y ‘C’ y un nuevo tipo de faseolina denominada ‘B’
(Boyacá).
27

Estudios complementarios confirmaron la diversidad de los acervos genéticos, pues
adicional a los tipos de faseolinas ya encontrados, se registraron nuevos tipos para
las accesiones Andinas cultivadas: H (Huevo de Huanchaco), mientras que en
materiales silvestres de Mesoamérica se encontró el tipo M (Mesoamérica) (Gepts
et al., 1986).
Koenig et al. 1989 consideraron que las accesiones de fríjol de Mesoamérica y de
los Andes tienen diferentes tipos de faseolinas, en el primer grupo se distinguen las
faseolinas tipo ‘S’ y ‘M’ mientras que en el grupo Andino predominaron las 'T', 'C',
'H', 'A', 'J' y la 'I'. Además encontraron una relación entre el tamaño de la semilla y el
tipo de faseolina.
Estudios como el realizado por Koenig y Gepts en 1989, en poblaciones silvestres
analizadas mediante aloenzimas y características morfológicas, definen los dos
grandes centros de diversidad. Sus resultados fueron consistentes con la diversidad
encontrada en estudios previos con faseolinas.
En otro estudio, se evaluaron accesiones pertenecientes a Centroamérica y
Suramérica (desde México hasta Argentina y Chile) mediante la evaluación de
patrón de aloenzimas; en este estudio fue posible distinguir dos grupos mayores con
formas Andinas y Mesoamericanas. Las aloenzimas encontradas en los genotipos
cultivados son un reflejo de las encontradas en cada uno de los dos grupos de
genotipos silvestres analizados en las mismas áreas (Singh et al., 1991b).
La técnica RFLP (Restriction fragment length Polymorphism) fue utilizada en 1994
por Becerra y Gepts (1994), quienes confirmaron la divergencia entre genotipos
Mesoamericanos y Andinos. Sugieren la existencia de un aislamiento reproductivo,
confirmando que en los cruces entre genotipos del acervo Mesoamericano y Andino
hay debilidad del híbrido F1.
Análisis mediante AFLP (de las siglas en Inglés Amplified Fragment Length
Polymorphism) revelan que las poblaciones de fríjol silvestre en la zona Andina han
sido aisladas la una de la otra, resultando poblaciones discretas en Ecuador y el
norte de Perú; en el sur de Perú, Bolivia y el norte de Argentina(Tohme et al., 1996).
28

Beebe et al (2000), analizaron 269 genotipos de fríjol común implementando la
técnica RAPD (Random Amplified Polymorphic DNA) con el fin de determinar la
estructura del acervo genético de fríjol cultivado en América central y México. Los
análisis permitieron relacionar la agrupación en razas que se había realizado
previamente mediante criterios morfológicos y agronómicos, con las agrupaciones
formadas a partir de este estudio molecular. Este germoplasma resultó ser más
complejo de lo que se pensaba anteriormente, se conservaron las agrupaciones de
las razas Mesoamérica, Jalisco y Durango. Pero las accesiones del germoplasma
de Guatemala no se agruparon con ninguna de las razas mencionadas previamente
definiéndose así como una nueva raza.
Análisis reportados por Beebe et al. (2001) determinaron la estructura genética de
una muestra de 182 genotipos Andinos, analizados mediante el uso de 189 primers
polimórficos AFLPs. Dentro de su estudio incluyeron genotipos Mesoamericanos y
silvestres; un análisis de correspondencia múltiple permitió separar los grupos
Andinos de los Mesoamericanos; dentro de los silvestres los genotipos
Mesoamericanos y del sur del Perú se separaron de los del sur de los andes. El
Acervo Andino no hubo una clasificación de accesiones como la encontrada por
Tohme et al 1996, y la tendencia a formar razas discretas no se detectó. Las razas
Nueva Granada, Perú y Chile del acervo Andino fueron definidas esencialmente
basadas en la morfología, los resultados de Beebe y colaboradores sugieren que las
tres razas pueden tener un origen común, estas razas pudieron haber resultado de
la intensiva intervención humana y de la selección para estos rasgos.
Un estudio reciente, por Blair et al en 2006, estima la diversidad alélica y la
Heterocigocidad de 129 microsatélites en un panel de 44 genotipos de fríjol. Fueron
evaluados marcadores génicos y genómicos; el análisis del contenido de
información polimórfica (PIC) permitió reconocer un grupo de marcadores con gran
capacidad de discriminación que pueden ser útiles en estudios de diversidad
genética. Se evaluó la distribución y el rango del tamaño de alelos. Este estudio
sugirió que los marcadores genómicos proveen mayor polimorfismo que los génicos
arrojando valores en número de alelos de 6.0 vs 9.2 y valores de PIC de 0.446 vs
0.594.
29

Los microsatélites fueron útiles para distinguir genotipos Andinos de
Mesoamericanos, para revelar las razas de cada acervo y para separar las
accesiones cultivadas de las silvestres. Se encontró mayor polimorfismo (53%
Andino y 33.4% Mesoamericano) y estructura de razas dentro del acervo Andino y la
tasa de polimorfismo entre los genotipos fue consistente con el acervo y la identidad
de razas. La diversidad intrapoblacional fue mayor dentro del acervo Andino que
dentro del Mesoamericano y este patrón fue observado para los dos tipos de
marcadores evaluados (génicos y genómicos) (Blair et al., 2006).
Uno de los últimos estudios realizado con el fin de describir la estructura de razas
del acervo Mesoamericano usando microsatélites es el de Díaz & Blair en el 2006,
en cual evaluaron 60 genotipos con 52 marcadores; el análisis de correspondencia
identificaron dos grupos principales equivalentes a la raza Mesoamérica y un grupo
que contenía las razas Durango y Jalisco. El análisis con estos marcadores permitió
identificar estos subgrupos, los cuales coinciden con las divisiones de clases
comerciales.
En una investigación realizada por Blair y colaboradores en el 2007, se buscó
caracterizar la estructura de razas de una colección de 123 genotipos
representantes del acervo Andino con 33 marcadores; adicional a esto se evaluó
una colección de individuos provenientes de Colombia. El análisis de
correspondencia múltiple de los genotipos Andinos identificó dos grupos
predominantes correspondientes a las razas Nueva Granada y Perú; algunos
individuos representantes de la raza Chile se agruparon de forma separada pero
muchos individuos definidos en estudios anteriores dentro de esta raza fueron
incluidos dentro de las otras razas.
2.3.4 Estudios de diversidad en fríjol Africano
Se destacan los trabajos de Martin & Adams (1987) y Khairallah et al. (1990), ambos
desarrollados en Malawi, para los que se emplearon dos técnicas importantes para
el estudio de la diversidad. En el primero se recurrió al estudio de datos
morfológicos, fenológicos y agronómicos, mientras que en el segundo se emplearon
marcadores mitocondriales. Estos estudios permitieron encontrar una gran
variabilidad morfológica en genotipos cultivados (variedades criollas y líneas
30

mejoradas) de fríjol Africano, aunque estos sólo se agruparon en los dos grandes
acervos en términos de variabilidad genética: Andino y Mesoamericano.
Por otra parte, en un reciente trabajo llevado a cabo por Moreno en el 2007, en el
CIAT, se evaluó la diversidad genética existente en 221 variedades comerciales
africanas de fríjol común, siendo este análisis una base para un programa de
mejoramiento nutricional en donde la meta final será aumentar el contenido de
hierro y zinc.
2.4 CUANTIFICACION DEL CONTENIDO DE MINERALES EN FRÍJOL
2.4.1 Cuantificación por Absorción Atómica (AA)
La Absorción Atómica es una de las principales técnicas utilizadas en la
cuantificación del contenido de minerales en muestras orgánicas. Ésta técnica
espectroscópica tiene como fundamento la absorción de radiación de una longitud
de onda determinada por parte de átomos libres, por lo tanto existe la necesidad de
liberar el analito de la matriz que lo contiene. Esta radiación es absorbida
selectivamente por átomos que tengan niveles energéticos cuya diferencia en
energía corresponda a la energía de los fotones incidentes (Rocha, 2000; Skoog et
al., 2001).
El principio en que se fundamenta la absorción atómica se puede describir de la
siguiente forma: cuando se hace pasar una disolución por una llama, los elementos
presentes son parcialmente llevados a su forma atómica. Y si a través de la llama se
hace pasar un haz de luz proveniente de una lámpara cuyo cátodo contiene el
elemento que se desea determinar y que, por tanto, emite las líneas características
de ese elemento, los átomos del elemento en cuestión presentes en la llama
absorberán esa radiación obedeciendo leyes cuantitativas de forma análoga a la
espectrofometría molecular (Muñiz, 1980).
Para llevar a cabo esta técnica se requiere que la muestra esté de forma líquida o
en solución. Las muestras sólidas son preparadas para el análisis mediante su
disolución en un solvente apropiado, en el caso de tejido vegetal se usa ácido
31

perclórico o nítrico; cuando la muestra no es soluble, ésta puede ser digerida a altas
temperaturas (Harvey, 2000).
La muestra se hace pasar en la forma de una dispersión de gotas muy finas a un
atomizador de llama, la disolución de la muestra es nebulizada mediante un flujo de
gas oxidante, mezclado con gas combustible, y se transporta a una llama donde se
produce la atomización. En la llama se produce una desolvatación, se evapora el
disolvente hasta obtener un aerosol rico en moléculas y luego la disociación de la
mayoría de estas moléculas produce un gas atómico. La mayoría de los átomos
formados así se ionizan originando cationes y electrones (Skoog et al., 2001).
Existen dos formas de atomización dentro de la técnica de AA, bien sea por llama o
atomización electrotérmica; la elección de cualquiera de estos dos métodos está
determinada principalmente por la concentración del analito en la muestra a
analizar. La principal ventaja de la absorción atómica con atomizador de llama es la
reproducibilidad con la cual la muestra es introducida dentro del espectrofotómetro,
y su desventaja significativa es que la eficiencia de la atomización puede ser
bastante pobre.
Independientemente del método de atomización, la AA es una técnica ideal para
cuantificación de trazas y ultratrazas de metales, debido a su sensibilidad,
selectividad, precisión y exactitud; adicionalmente, su análisis es rápido y no implica
mayor costo.
2.4.2 Estudios de cuantificación de minerales en Fríjol Común
Uno de los principales estudios que se han realizado para el análisis del contenido
de minerales en el fríjol común, es el de Beebe et al. (2000), en el que se analizaron
mas de mil accesiones de la colección núcleo de fríjol del CIAT con el fin de
determinar el grado de variabilidad genética del fríjol por contenido mineral, usando
la tecnología de Plasma Acoplado Inductivamente ó ICP (Inductive Coupling
Plasma), para la cuantificación de hierro y zinc. Encontraron un rango de 34 a 89
ppm con un promedio de 55 ppm para hierro, sin que hubiera evidencia de
correlación entre el contenido de hierro y la distribución geográfica; sin embargo, las
accesiones Andinas tendían a presentar valores más altos que las Mesoamericanas.
32

El contenido de Zinc en los fríjoles fue uno de los más altos dentro de las plantas; en
la evaluación de la colección núcleo se encontraron rangos de 21 a 45 ppm en el
contenido de Zinc , con un valor promedio de 35 ppm (Beebe et al., 2000a).
Islam et al (2002), analizaron cinco elementos nutricionales (Ca, P, S, Fe y Zn), en
la colección núcleo de CIAT, 1072 accesiones, mediante la implementación del
método ICP. El producto del ICP fue usado para Espectroscopía de emisión atómica
(AES). Obtuvieron un valor mínimo para hierro de 34,60 y máximo de 91,90 ppm y
un valor promedio de 54,81 ppm. Para zinc, el valor mínimo fue de 20,7 y valor
máximo de 59,40 ppm; el promedio correspondió a 34,40 ppm.
En otro estudio se analizaron los contenidos de hierro y zinc en una población de
líneas recombinantes, cruce entre G21242 (fríjol trepador colombiano con alto
contenido de hierro y zinc) y G21078 (fríjol trepador Argentino con bajo contenido de
hierro y zinc). Dos métodos de análisis de minerales fueron implementados, Plasma
acoplado inductivamente (ICP) y Espectroscopia de Absorción Atómica (AA). La
población fue analizada en dos localizaciones en Colombia (Popayán y Darién), el
contenido promedio de hierro y zinc calculado en ppm en la progenie del cruce de
poblaciones (G21242 x G21078). Se encontró correlación entre minerales y entre
ambientes (Blair et al., 2005).
33

3 FORMULACIÓN DEL PROBLEMA 3.1 FORMULACIÓN DEL PROBLEMA
El fríjol común (Phaseolus vulgaris L.), es un renglón importante de la economía y
alimentación de la mayoría de países en vía de desarrollo; Ruanda (África) es un
país con uno de los índices más altos de consumo de ésta leguminosa, por lo tanto
se requiere de la profundización en estudios a nivel molecular que permitan
caracterizar la diversidad genética de esta especie con miras a apoyar su
mejoramiento nutricional.
Ruanda, como la mayoría de los países en vía de desarrollo de África Central basa
sus ingresos, oportunidades de empleo y en general su economía en la agricultura,
ya que cerca del 90% de sus habitantes se encuentra en las áreas rurales (Ferris et
al., 2002). La perspectiva económica de Ruanda es un gran reto. El país
experimenta un aumento en las tasas de crecimiento de la población, lo que afecta
directamente en la disminución de terreno útil para el uso de la agricultura per cápita
y a su vez genera un aumento en la demanda de alimentos, lo que trae como
consecuencia que la seguridad alimentaria y el alivio de la pobreza estén en su
punto más crítico. También se responsabiliza esta presión por tierra como causa de
la guerra civil de 1994 que encajó un genocidio de más de un millón de personas en
ese país (Sperling, 2001).
Para garantizar seguridad alimentaria y que los ingresos rurales aumenten es
necesario incrementar la productividad en el sector agronómico. Dicho incremento
es posible con el mejoramiento de los principales cultivos del país. Dentro de éstos,
se encuentra el fríjol (Phaseolus vulgaris L.), el cual forma parte integral del sistema
de producción y consumo en Ruanda; principalmente es apreciado por su larga vida
durante el almacenamiento y buenas propiedades nutricionales. Es uno de los
alimentos centrales de la dieta en Ruanda, aportando cerca del 60% y 30% de todas
las proteínas y calorías ingeridas respectivamente, siendo el segundo en
importancia después del maíz (Ferris et al., 2002).
34

Lo anterior indica la necesidad de adelantar procesos de caracterización y
evaluación de la diversidad genética de una colección de fríjol de Ruanda, con el fin
de establecer un punto de partida en el camino de la implementación de un
programa eficiente de mejoramiento, ya sea este nutricional o meramente genético,
de esta especie y así poder contar en un futuro cercano con materiales promisorios
que suplan las necesidades de consumo. Por otra parte, las accesiones africanas
han sido poco estudiadas a nivel molecular lo que genera mayor interés al realizar
este estudio.
3.2 JUSTIFICACIÓN
El fríjol común (Phaseolus vulgaris L.) es la leguminosa de grano más importante en
el mundo siendo uno de los principales productos de la dieta de los países en vía de
desarrollo. Es especialmente importante en países en desarrollo de América y en
África, regiones donde se produce el 47% y 24% respectivamente, de la producción
mundial de fríjol seco (Becerra & Gepts, 1994). El valor nutritivo del fríjol se centra
en el alto contenido de proteínas, carbohidratos, grasas, vitaminas y minerales
(Bazel & Anderson, 1994).
Aún cuando el fríjol es una de las especies más importantes del sistema de
producción y consumo en el continente Africano, específicamente en el país de
Ruanda, su consumo sobrepasa la producción siendo necesaria su importación;
además éste cultivo presenta diversos problemas, por ejemplo la baja fertilidad del
suelo, plagas y enfermedades (Ferris et al., 2002).
Teniendo en cuenta estos factores que han afectado la economía y la seguridad
alimentaria de sus habitantes, se hace imperiosa la necesidad de emprender
estudios del conocimiento de la diversidad genética, encaminados hacía un futuro
fitomejoramiento de la especie. La diversidad de los recursos genéticos facilita la
adaptación a necesidades y condiciones siempre cambiantes y hace posible la
agricultura sostenible en muchos de los diferentes ambientes de producción. Los
recursos genéticos de una especie vegetal constituyen la materia prima con la que
los fitomejoradores producen nuevas variedades (FAO, 1996).
35

Estas son las principales razones por las que se motivó la realización de un estudio
de caracterización de la diversidad genética de la colección de germoplasma del
fríjol de Ruanda, como apoyo a los procesos de mejoramiento de esta especie en
ese país, especialmente dirigido a un mejoramiento nutricional.
La caracterización y evaluación de la colección de fríjol se realizará mediante el uso
de marcadores genéticos, lo que permitirá el conocimiento de la composición
genotípica de estos materiales. Por lo tanto, la presente propuesta se centra en
determinar la variabilidad genética mediante marcadores microsatélites y a su vez la
composición de minerales (hierro y zinc) de una colección de fríjol de Ruanda.
Se recurre a las técnicas moleculares como los microsatélites, ya que se han
utilizado con mucho éxito en la estimación de la variabilidad genética dentro y entre
poblaciones, así como en muestras de accesiones de especies cultivadas o
silvestres, de manera efectiva y rápida (Mitchell et al., 1997) y específicamente se
utiliza un marcaje fluorescente, por ser una técnica automatizada que garantiza
mayor precisión, sensibilidad, resolución y eficiencia en el genotipaje (Joe et al.,
2004).
36

4 OBJETIVOS
4.1 Objetivo General
Caracterizar la diversidad genética de 355 genotipos de fríjol común provenientes de
Ruanda (África), mediante el uso de marcadores moleculares tipo microsatélites
marcados con fluorescencia.
4.2 Objetivos Específicos
i) Evaluar el polimorfismo alélico para 355 genotipos de fríjol de Ruanda
(África) mediante el uso de marcadores moleculares tipo microsatélites
analizados con una técnica de fluorescencia.
ii) Analizar la relación que presentan los genotipos entre sí mediante su
similaridad evaluada con criterios de diversidad genética y el uso de
métodos estadísticos.
iii) Relacionar por medio de estadística descriptiva los caracteres de grano
(color y tamaño) y el contenido de minerales de los genotipos, con las
agrupaciones generadas a partir de la evaluación del polimorfismo
alélico.
37

5 MATEERIALES YY MÉTODOSS
L
y
C
d
5
P
e
e
S
4
L
p
e
3
d
La elaborac
y análisis
Caracterizac
Internaciona
departamen
5.1 Poblac
Para el des
específicam
está localiza
Subsaharian
4′ E, en la fig
Fi
Las semillas
pertenecient
en Palmira,
355 genotip
diversidad, e
ión de este t
estadístico
ción de G
al de Agri
to del Valle
ción de Estu
arrollo de e
ente de ISA
ado en la pa
na o la de “g
gura 4 se m
igura 4. Loca
s se encontr
te al progra
Valle. La m
pos y adici
en la tabla 5
trabajo (el p
de los da
Germoplasma
icultura (C
del Cauca, C
udio
este proyecto
AR (Instituto
rte oriental d
grandes lago
uestra la ub
alización geog
raban almac
ama de Cara
muestra para
onalmente
5 se muestra
38
rocesamien
atos), se lle
a de Fríjo
IAT), situa
Colombia.
o se utilizó
de Ciencias
de África Ce
os”, con coor
icación de R
gráfica de Rua
cenadas en e
acterización
a la evaluac
se utilizaro
an los genoti
to de las mu
evó a cabo
ol (LCGF),
do en el
semilla de f
s Agronómic
entral en una
rdenadas ge
Ruanda en e
anda en el co
el cuarto de
de Germop
ción de la d
on 4 materi
pos y su res
uestras, aná
o en el L
ubicado e
municipio
lisis molecu
aboratorio
en el Cent
de Palm
lar
de
tro
ira
fríjol proven
cas de Ruan
a región con
eográficas de
el continente
iente Ruand
nda); este pa
nocida como
e 1° 57′ S 3
e Africano.
da,
aís
o la
30°
ontinente Africcano
conservació
plasma de F
iversidad ge
iales como
spectivo orig
ón de semill
Fríjol del CIA
enética fue
controles
gen.
las
AT
de
de

Tabla 5. Controles Diversidad utilizados en este estudio con su respectivo origen (acervo al
que pertenecen)
CONTROLES ACERVO GENÉTICO
FOTO
DOR 364 Mesoamericano
ICA-PIJAO Mesoamericano
CALIMA Andino
G19833 Andino
El modelo de tamaño de muestra propuesto por Leung et al. (1993), predice que con
una confiabilidad del 95%, son necesarios 29 genotipos de un acervo genético para
capturar o incluir alelos con una frecuencia de hasta el 10%, lo que se cumpliría
para el caso de las accesiones provenientes de Ruanda escogidas para este estudio
y es conforme con lo propuesto por Li y Nei (1987) quienes aconsejan utilizar un
mínimo de 20 a 30 individuos por locus sí el número de loci a examinar es 25. Para
este estudio se tiene una muestra de 355 individuos, lo que indica que se tiene un
buen número para el análisis de diversidad.
39

5.2 MÉTODOS
5.2.1 Procesamiento del material vegetal:
5.2.1.1 Germinación de la semilla:
Se seleccionaron 10 semillas de cada uno de los 355 genotipos las cuales se
germinaron en condiciones de laboratorio, de la siguiente manera: Las diez semillas
de cada genotipo fueron lavadas superficialmente con agua bidestilada y
posteriormente escarificadas (corte longitudinal de la testa en el lado opuesto del
micrópilo de la semilla) para asegurar una rápida y uniforme germinación, ya que
mediante este proceso se facilita la absorción de agua a la semilla.
Las semillas se colocaron de forma lineal y espaciada en el centro de un rectángulo
de papel de germinación de 30 x 25cm, debidamente rotulado con el número de
identificación de cada genotipo. Cada rectángulo de papel se enrolló y se amarró
con un alambre con el fin de asegurar las semillas. Cada rollo se colocó en
imbibición en una bandeja con una solución de Sulfato de Calcio (Ca2SO4) 0.5 mM
para evitar contaminación con patógenos.
Las bandejas con los rollos de papel de germinación se almacenaron bajo
condiciones de oscuridad, para evitar el desarrollo de tejidos fotosintéticos ya que
éstos pueden afectar la calidad del ADN por la acumulación de almidones. Después
de 8 días de crecimiento las hojas cotiledonares habían emergido; se escogió un
trifolio etiolado de cada planta, se colectaron en total 10 trifolios por genotipo y se
distribuyeron en tres tubos Eppendorf de 2mL (3-3-4 trifolios).
5.2.1.2 Procesamiento Post-germinación
Los tubos marcados con la identificación de cada genotipo, fueron almacenados en
un congelador a -80°C hasta el momento de su liofilización. La liofilización es un
método para deshidratar el tejido por medio de choques congelantes y secado en el
estado de congelación bajo condiciones de vacío, haciendo que el hielo pase
directamente a vapor de agua (sublimación). El material celular y las
macromoléculas secas mantienen su actividad biológica y pueden almacenarse por
40

este estado por periodos largos (Ferreira & Grattapaglia, 1998). Este proceso se
llevó a cabo durante dos días en un equipo Dryer MODULYoD-115 marca Thermo.
Una vez secas las muestras, se procedió a su maceración con morteros especiales
para tubos de microcentrífuga y con la ayuda de un motor manual.
5.2.2 Extracción y cuantificación de ADN
Una vez macerados los trifolios se combinó el material de los tres tubos de cada
genotipo en un solo tubo Eppendorf de 1,5mL; es necesario aclarar que debido a
que la semilla llevaba aproximadamente dos años almacenada en el cuarto de
conservación de semillas del CIAT, durante el proceso de germinación se
presentaron problemas y en muchos casos los trifolios colectados eran muy
pequeños, por lo que la cantidad macerada de los tres tubos cabían perfectamente
en un solo tubo de 1,5mL.
El proceso de extracción, se realizó empleando el método de Mahuku (2004), que
utiliza la acción combinada de la Proteinasa K y del EDTA para inhibir o degradar
completamente a las nucleasas (Anexo 1), este método funciona muy bien con poco
tejido (Ferreira & Grattapaglia, 1998; Mahuku, 2004). El producto de la extracción se
almacenó a -20°C. Con el fin de determinar la calidad del ADN, se sembraron 2 µL
de ADN total de cada muestra en un gel de agarosa al 1%, teñido con Bromuro de
etidio. La cuantificación del ADN extraído se realizó mediante electroforesis en gel
de agarosa utilizando el software Quantity one® v 4.0.3, el cual calcula la cantidad
de ADN en cada una de las bandas, de acuerdo a las extrapolaciones que realiza
con los patrones de concentración de ADN conocidos (Bio-Rad, 1998), para este
caso se utilizaron ADN lambda de [50ng], [100ng], [200ng] y [400ng].
La electroforesis, se realizó en gel de agarosa al 1% y se tiñó con bromuro de etidio;
en cada pozo del gel se sembró una mezcla de 2µL de ADN, 6µL de agua grado
HPLC y 4µL de Blue Juice (Glicerol + Azul de Bromofenol) y se sometió a
electroforesis durante 20 minutos aproximadamente a 120 V. Posteriormente se
tomó una fotografía del gel con el equipo GelDoc 2000 de BioRad acoplado a un
trasniluminador de luz U.V.
41

Con el fin de corroborar los datos obtenidos a partir de la cuantificación realizada
con el software Quantity one®, se realizó una segunda cuantificación del material
mediante el uso del Fluorometro DyNA Quant, (Hoefer Pharmacia Biotech Inc.,
1995) con base en la solución de colorante HoeschtH 33258
5.2.3 Amplificación mediante PCR de microsatélites analizados con
fluorescencia
La caracterización de los genotipos se realizó a partir del uso de 30 marcadores tipo
microsatélites marcados con fluorocromos (tabla 6) seleccionados de 6 paneles
según Blair et al 2007, de los cuales 14 eran génicos y 16 genómicos; este grupo de
marcadores fue seleccionado por ser altamente polimórfico (Blair et al., 2003; Diaz,
2005a; Diaz, 2005b; Gaitan-Solis et al., 2002):
Tabla 6. Cebadores fluorescentes para la amplificación de microsatélites utilizados en la
caracterización genética de la colección de germoplasma de Ruanda.
Marcador Motivo Fluorescencia Panel Genómicos
AG01 (GA)8GGTA(GA)5GGGGACG(AG)4 6-FAM 7 BM139 (CT)25 PET 6 BM143 (GA)35 6-FAM 8 BM156 (CT)32 6-FAM 6 BM160 (GA)15(GAA)5 NED 6 BM165 (TC)14 TET 3 BM172 (GA)23 PET 5 BM175 (AT)5(GA)19 VIC 5 BM183 (TC)14 TET 7 BM187 (CT)10T(CT)14 HEX 7 BM188a (CA)18(TA)7 NED 5 BM188b (CA)18(TA)7 NED 5 BM201 (GA)15 6-FAM 7 BM205 (GT)11 PET 5 GATs54 (GA)5AACAGAGT(GA)8 HEX 7 GATs91 (GA)17 6-FAM 5
Génicos BMd01 (AT)9 6-FAM 6 BMd02 (CGG)8 PET 6 BMd08 (CT)7 NED 5 BMd16 (CATG)4 VIC 6 BMd17 (CGCCAC)6 TET 7 BMd18 (TGAA)3 HEX 9 BMd20 (TA)5 VIC 5 BMd45 (AG)5 TET 3 BMd46 (TCT)4 TET 8 BMd47 (AT)5 HEX 8 Pv-ctt001 (AT)9 6-FAM 6 Pv-ag003 (AG)8 NED 6 Pv-gaat001 (AT)4(T)2 TET 9 Pv-cct001 (CTT)3(T)3(CTT)6 VIC 6
PET: rojo, VIC: verde, 6-FAM: azul, NED: amarillo TET: verde
42

Los marcadores fueron diseñados a partir de regiones codificantes (génicos) y no
codificantes (genómicos) de Phaseolus vulgaris (Blair et al., 2003; Gaitan-Solis et
al., 2002; Yu et al., 2002). Dicha amplificación fue hecha en el laboratorio de
Caracterización de Germoplasma de Fríjol del CIAT; los modelos de los
termocicladores usados para tal fin fueron PTC-100TM y PTC-200TM (Programmable
Thermal Controller) del fabricante MJ Research, INC. Los pasos básicos para
realizar una PCR se muestran el la figura 5.
Amplificaciones individuales de PCR de cada microsatélite fueron llevadas a cabo
en un volumen final de 15 µL por reacción con 15ng de ADN; los reactivos
necesarios y condiciones requeridas para realizar el coctel o mezcla para la
amplificación de los microsatélites marcados con fluorocromos se muestran en la
tabla 7. El extremo 5’ (Forward), de los primers seleccionados, estaba marcado con
los siguientes fluorocromos: hexachloro-6-carboxyfluorescein (HEX), 6-
carboxyflourescein (6-FAM), (NED), (PET), y (VIC), y sintetizada en Applied
Biosystem 3730xl (Applied Biosystem, Foster City, Calif), facilitado por el centro de
recursos biotecnológicos (BRC) de la universidad de Cornell, USA.
Tabla 7. Reactivos necesarios para coctel PCR, condiciones necesarias y concentraciones
originales (stock).
Reactivo Condiciones Requeridas
[ ] soluciones Originales
Buffer PCR 1X 10X
MgCl2 1.5mM 25mM
dNTP's 0.2mM 20mM
Primer 3pmol/rxn 2µL
taq [1:10] 0.15µL
ADN 25ng/µL
Volumen final = 15µL (5µL ADN + 10µL mix)
La composición del buffer PCR (100 mM Tris HCl pH 7.2; 500 mM KCl)
43

El programa de PCR para la amplificación consistió en una denaturacion fuerte
durante 2 minutos, seguida de 30 ciclos de denaturación a 95°C cada uno de 30
segundos; una etapa de hibridación a 55°C durante 30 segundos; extensión a 72°C
durante 50 segundos; y una extensión final de 72°C durante 45 minutos. Se tomaron
2µL del producto de PCR de cada reacción de cada microsatélite y 9µL de agua
HPLC, se realizó una mezcla en una placa nueva de 96 pocillos, teniendo en cuenta
que la mezcla de cada reacción debía tener producto de los cuatro primers teñidos
con fluorocromos (PET: Rojo; VIC: Verde; 6-FAM: Azul; NED: Amarillo) (Blair et al.,
2002).
Se establecieron 5 paneles (5, 6, 7, 8 y 9) cada uno con la presencia de 4
marcadores (Comunicación Personal M.W Blair mejorador de fríjol, CIAT 2006),
teniendo en cuenta que para hacer esta mezcla los marcadores deben tener
diferente tamaño y sí éstos se sobrelapan en tamaño, deben ser diferenciados
mediante el uso de colores diferentes (Joe et al., 2004). En el Anexo 2 se muestran
los marcadores utilizados para este estudio y la forma de organización de estos
dentro de cada panel. Se determinó que se debían agregar dos marcadores más
(BM165 y BMd45), por lo que se hizo el arreglo de un panel adicional.
De esta mezcla se tomaron 0.5 µL para adicionarlo con 9 µL de Formamida Hi Di,
0.06µL de Liz 500 estándar, 0.44µL de agua, es decir un volumen final de 10µL para
cada reacción; cada placa de 96 pozos se sometió a denaturación a 94°C durante 3
minutos para finalmente ser analizada en el secuenciador ABI 3730xl.
5.2.4 Análisis en secuenciador Analítico ABI 3730xl
Los microsatélites marcados con etiquetas fluorescentes (fluorocromos) de colores
verde, azul, amarillo y rojo, fueron analizados automáticamente en un secuenciador
de ADN Applied Biosystem 3730xl (Applied Biosystem, Foster City, Calif) facilitado
por el centro de recursos biotecnológicos (BRC) de la universidad de Cornell USA,
el cual detecta la muestra a través de un sistema de electroforesis capilar; en el
anexo 3 se puede observar el ciclo que sigue la muestra en el secuenciador.
44

Los datos de la muestra leída mediante el secuenciador ABI 3730xl, fueron
analizados automáticamente por el software GeneMapper v.3.7, el cual interpreta el
lenguaje análogo de las lecturas que hace el secuenciador y realiza la identificación
de picos y tamaños en los electroferograma generados por la evaluación de
unidades fluorescentes por tiempo de elución. Este software combina la información
del archivo de la muestra con análisis preconfigurados y calibraciones estándar; las
muestras procesadas luego fueron vistas, editadas y analizadas en una ventana
generada por el software (AppliedBiosystems, 2004; Joe et al., 2004).
En la figura 5 se observa el procedimiento que sigue la muestra una vez se han
efectuado las amplificaciones y se ha realizado la mezcla. La muestra entra al
secuenciador, posteriormente un software colecta la información analizada y
procede a ser leída en otro software autoanalizador el cual puede encontrarse en
otro computador con conexión remota, el cual recoge los datos mediante un
servidor.
Muestra Preparada
Secuenciador ABI 3730
Software colector de datos
Servidor
Software Auto‐analizador GeneMapper
Figura 5. Diagrama del procedimiento que sigue la muestra a analizar (líneas azules
gruesas señalan cada paso): entra al secuenciador ABI 3730xl, es colectada por un software
y se analiza con GeneMapper v.3.7. Modificado de (Joe et al., 2004).
45

5.2.5 Cuantificación de hierro y zinc
Los genotipos analizados en este estudio corresponden a la colección de frijol de
Ruanda, que incluye semilla de color blanco, rojo, rojo moteado, café, crema y
crema moteado, amarillo y amarillo moteado, rosado, púrpura y negro; con tamaños
de semilla que se encontraban desde pequeña, mediana y grande. El contenido de
minerales, hierro y zinc, fue evaluado mediante la técnica de espectroscopía por
absorción atómica (AA). El proceso de obtención de datos para la realización del
análisis de minerales, fue realizado previamente a este estudio en el laboratorio de
caracterización de germoplasma de fríjol de CIAT.
Para llevar a cabo este procedimiento, se pesaron aproximadamente 3 gramos de
semillas por cada genotipo, las cuales fueron lavadas y limpiadas con agua
destilada y gasa, posteriormente se secaron en un horno a 40°C durante dos días y
finalmente se molieron en un molino Retsch modificado en cámaras de teflón y con
balines de zirconio con volumen de 25 ml. El polvo obtenido se envasó y etiquetó en
frascos de plástico herméticamente sellados.
Una vez obtenido el material, las muestras fueron enviadas al Departamento de
Servicios Analíticos del CIAT para analizarlas por AA. Las ecuaciones de calibración
para predecir el contenido de hierro y zinc fueron obtenidas mediante el software
winISI III Project Manager v 1.60, con el modelo matemático de Mínimos Cuadrados
Parciales (PLS).
5.2.6 Identificación de características de grano
La semilla fue registrada fotográficamente mediante el escaneo digital de 4 semillas,
esto con el fin de tener la información visual del color, tamaño y forma de cada
genotipo. Se realizó una clasificación de la semilla de acuerdo con su patrón de
coloración siguiendo el Catálogo de líneas avanzadas de fríjol del CIAT 1995, ver
tabla 8, en donde cada color se representa con un número (1-9), al igual que el brillo
de la semilla (1-3) y el patrón de coloración con una letra (R, J, M, P, V), Anexo 4:
46

Tabla 8. Parámetros de descripción para características de la semilla de fríjol; basado en el
catalogo de líneas avanzadas.
Color de Semilla Brillo de Semilla 1 Blanco 1 Opaco 2 Bayo, Crema 2 Semi‐Brillante 3 Amarillo 3 Brillante 4 Café, Marrón Patrón de Color 5 Rosado M Moteado 6 Rojo J Jaspeado 7 Morado, Púrpura R Rayado 8 Negro P Punteado 9 Gris o Raros V Venación
5.3 ANÁLISIS DE DATOS
5.3.1 Análisis de Diversidad Genética
Como primer paso para el análisis de la diversidad se llevó a cabo la determinación
de alelos mediante el software GeneMapper v.3.7 (AppliedBiosystems, 2004), el
cual generó un archivo en Excel que registró para cada genotipo los alelos en cada
uno de los loci.
A continuación se construyó una matriz de similaridad genética basada en la
proporción de alelos compartidos (PS) calculado con el módulo IML/SAS, esta es
una medida adecuada para datos de individuos diploides analizados con
microsatélites; este proceso realizado de esta manera es una herramienta útil para
los análisis de similaridad genética cuando existe una pequeña porción de datos
faltantes por la no amplificación de determinados loci (Comunicación Personal M.C.
Duque consultora estadística CIAT, 2007). La elaboración de esta nueva matriz de
similaridad se llevó a cabo mediante el paquete estadístico SAS el cual utiliza la
siguiente fórmula:
/
i locus número total de loci 30.
Psi proporción de alelos compartidos en locus i.
47

A partir de la construcción de la matriz de similaridad fue posible efectuar dos tipos
de análisis: un análisis multivariado de Coordenadas Principales (PCoA) y la
obtención de dos representaciones gráficas: una gráfica de 3D y un dendrograma
utilizando distancia Euclidiana.
El PCoA es una técnica descriptiva de análisis multivariado que representa en un
espacio multidimensional la asociación de las bandas alélicas derivadas de los
genotipos estudiados; el nivel de asociación se determina de acuerdo a la cercanía
que presenten los genotipos en el gráfico de tres dimensiones resultante. Resuelve
relaciones complejas en interacciones de factores más simples y menos numerosos
(Ferreira & Grattapaglia, 1998).
Las coordenadas de ubicación obtenidas para cada genotipo fueron graficadas
mediante SAS-GRAPH modulo G3D para gráficas, Proc cluster para generación de
grupos, lo cual permitió determinar conjuntos de genotipos de acuerdo con el perfil
molecular obtenido; en NTSYS se hizo una descomposición espectral mediante el
módulo EIGEN, se crearon las coordenadas de cada punto en el nuevo espacio
definido por las coordenadas principales; se realizó un proceso de agrupamiento en
SAS y con el criterio de R2 y se eligió el número de grupos (Comunicación Personal
M.C Duque consultora estadística, CIAT 2007). Esta representación gráfica refleja la
similaridad genética entre genotipos en términos de distancias geométricas, definida
por la proporción de alelos compartidos; esto permitió establecer la estructura del
conjunto de individuos estudiados (Ayad et al., 1997).
El siguiente paso fue la obtención del dendrograma el cual tiene las coordenadas
del PCoA utilizando distancia euclidiana y el método de agrupamiento UPGMA
mediante los módulos GRAPHICS y TREE PLOT del programa NTSYS v 2.10
(Rohlf, 2002).
5.3.1.1 Parámetros de uso común de diversidad para población total
Se midieron los parámetros de uso común tales como: Heterocigocidad Observada,
Diversidad Genética de Nei (1987) y número de alelos encontrados para cada
marcador, mediante el programa estadístico Powermarker V3.25 (Liu & Muse,
2005),
48

La Heterocigocidad observada corresponde a la proporción de individuos
heterocigotos en la población. Para un locus es estimada como (Weir, 1996):
1
locus l‐ésimo Frecuencia del genotipo en el locus é en la población
La diversidad genética se define en términos de frecuencias génicas y usualmente
se refiere a la Heterocigocidad esperada; es definida como la probabilidad de que
dos alelos que sean escogidos aleatoriamente de la población sean diferentes.
Según Weir en 1996 un estimador insesgado de la diversidad genética en el locus
th es (Liu & Muse, 2005):
1 / 11
alelos en el locus l locus lth
th en la población
= Frecuencia del alelo en
Número de individuos mu coeficiente de endogamia
el locus estreados
5.3.1.2 Parámetros de uso común de diversidad para grupos formados a partir de PCoA
Los parámetros tenidos en cuenta fueron los mismos que se utilizaron en la
caracterización de la diversidad genética polimorfismo, Heterocigocidad observada,
Diversidad genética, entre los grupos definidos a partir de PCoA, para este fin se
utilizó el software Popgene v1.32 (Yeh et al., 1997).
5.3.1.3 Coeficiente de similaridad
Con el fin de observar otra forma de agrupación de los datos se utilizó un método
diferente al de PCoA, usando el software Darwin v. 5.0.149. A partir de una matriz
binaria de los alelos presentes en el estudio, se estimó el índice de disimilaridad de
49

DICE, que utiliza la fórmula a continuación expuesta. El módulo para dibujar el árbol
fue NeighborJoining.
2
: disimilaridad entre unidades i y j
: número de variables donde xi presencia y xj presenci: número de variables donde xi presencia y xj ausencia: número de variables donde xi ausencia
a
y xj presencia
Este método fue propuesto por Saitou y Masatoshi Nei (1987), basado en el
principio de parsimonia y consiste en la unión de los genotipos más cercanos
(vecinos) tratando de minimizar la longitud total del árbol.
5.3.2 Análisis de cuantificación de minerales
Con base en estudios previos de cuantificación de minerales realizados para la
colección núcleo del CIAT (Beebe et al., 2000a; Islam et al., 2002a), se consideró lo
siguiente (tabla 9):
Tabla 9. Rango de valores medidos en ppm para interpretación de resultados de
cuantificación de minerales (hierro y zinc).
Hierro Zinc Bajo <50ppm <20ppm Medio 51-60ppm 21-30ppm Alto >61ppm >31ppm
Se analizaron los datos mediante estadística descriptiva en el programa SAS, este
análisis incluyó valores máximos, mínimos, promedio, coeficiente de variación,
desviación estándar, varianza y curtosis. Se elaboraron histogramas con el fin de
determinar la distribución de datos en la población de estudio. Se elaboraron
pruebas ANOVA con el fin de comparar los grupos generados a partir del análisis de
diversidad (PCoA) y poder reconocer si había o no diferencias significativas,
50

posteriormente se realizó una prueba para ajustar medias llamada Least Square
Means y se hizo una prueba de separación de medias de rango múltiple (Ryan-
Einot-Gabriel-Welsch), mediante el paquete estadístico SAS.
5.3.3 Características morfológicas
Se realizaron gráficas que muestran la distribución porcentual de cada característica
morfológica presentada por los individuos dentro de la población total. Adicional a
esto se realizaron tablas de contingencia que permitieron describir la distribución de
cada característica dentro de los grupos formados a partir de PCoA
51

6 RESULTADOS
6.1 ANÁLISIS DE DIVERSIDAD GENÉTICA
6.1.1 Diversidad Genética y estructura Poblacional
6.1.1.1 Determinación de alelos
Una vez se obtuvieron los electroferogramas se realizó la lectura de alelos
presentes para cada marcador. En la figura 6 se muestra la amplificación de 4
marcadores para un mismo individuo, (GATs91, BMd20, BMd08 y BM172)
pertenecientes al panel 5A. Como se puede observar cada marcador fue etiquetado
con un fluorocromo de color diferente: 6-FAM (azul), NED (amarillo), PET (rojo), y
VIC (verde).
MARCADOR
Figura 6. Electroferograma obtenido a partir del análisis por ABI 3730xl y generado por el
software GeneMapper v.3.7. Se muestra la amplificación de 4 microsatélites marcados con
fluorocromos 6-FAM (Azul), PET (Rojo), VIC (Verde) y NED (Amarillo; en la gráfica con color
negro), para un mismo individuo. Los picos representan la intensidad de fluorescencia (eje
vertical) y tamaños estimados en nucleótidos (eje horizontal).
52

En la figura 7 se presentan los mismos 4 marcadores de la figura anterior; en este
caso se sitúan en un solo plano con el marcador estándar denominado LIZ 500, el
cual fue efectivo para estimar los pesos moleculares de todos los productos de PCR
amplificados a lo largo de los paneles.
Figura 7. Electroferograma obtenido a partir del análisis por secuenciador ABI 3730xl y
generado por software GeneMapper v.3.7. Se muestra la distribución de los picos (zapote)
del marcador estándar LIZ-500 junto con los marcadores del panel 5A. Nuevamente los
picos representan la intensidad de fluorescencia (eje vertical) y tamaños estimados en
nucleótidos (eje horizontal).
6.1.1.2 Población Total
Para el análisis de la población de 359 genotipos (incluyendo los 4 controles de
diversidad genética) mediante el uso de 30 microsatélites marcados con
fluorescencia, fue posible encontrar 301 alelos en total para toda la población en
estudio, un valor alto sí es comparado con otros estudios también de microsatélites
(Blair et al., 2007; Díaz & Blair, 2006) en los que no se encontró un valor superior a
270 alelos. Uno de los promedios de alelos más altos fue el encontrado por Blair y
53
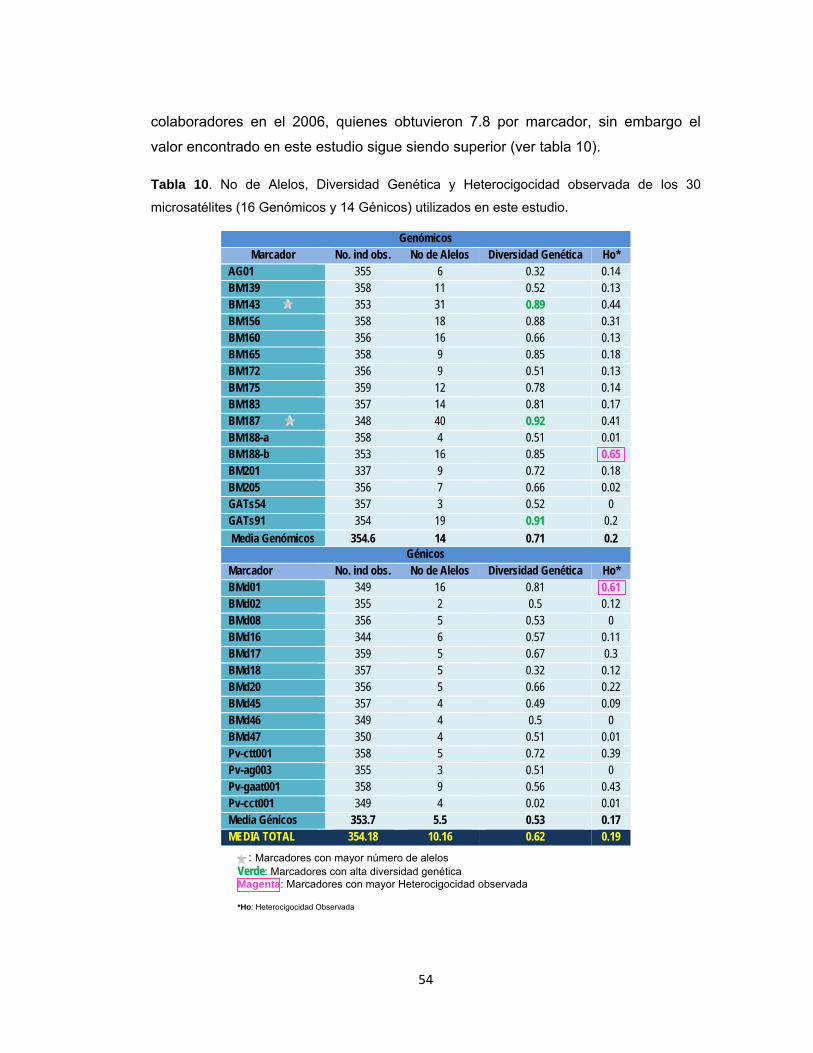
colaboradores en el 2006, quienes obtuvieron 7.8 por marcador, sin embargo el
valor encontrado en este estudio sigue siendo superior (ver tabla 10).
Tabla 10. No de Alelos, Diversidad Genética y Heterocigocidad observada de los 30
microsatélites (16 Genómicos y 14 Génicos) utilizados en este estudio.
Genómicos Marcador No. ind obs. No de Alelos Diversidad Genética Ho*
AG01 355 6 0.32 0.14 BM139 358 11 0.52 0.13 BM143 353 31 00..8899 0.44 BM156 358 18 0.88 0.31 BM160 356 16 0.66 0.13 BM165 358 9 0.85 0.18 BM172 356 9 0.51 0.13 BM175 359 12 0.78 0.14 BM183 357 14 0.81 0.17 BM187 348 40 00..9922 0.41 BM188-a 358 4 0.51 0.01 BM188-b 353 16 0.85 0.65 BM201 337 9 0.72 0.18 BM205 356 7 0.66 0.02 GATs54 357 3 0.52 0 GATs91 354 19 00..9911 0.2 Media Genómicos 354.6 14 0.71 0.2
Génicos Marcador No. ind obs. No de Alelos Diversidad Genética Ho* BMd01 349 16 0.81 0.61 BMd02 355 2 0.5 0.12 BMd08 356 5 0.53 0 BMd16 344 6 0.57 0.11 BMd17 359 5 0.67 0.3 BMd18 357 5 0.32 0.12 BMd20 356 5 0.66 0.22 BMd45 357 4 0.49 0.09 BMd46 349 4 0.5 0 BMd47 350 4 0.51 0.01 Pv-ctt001 358 5 0.72 0.39 Pv-ag003 355 3 0.51 0 Pv-gaat001 358 9 0.56 0.43 Pv-cct001 349 4 0.02 0.01 Media Génicos 353.7 5.5 0.53 0.17 MEDIA TOTAL 354.18 10.16 0.62 0.19
: Marcadores con mayor número de alelos V Veerrddee: Marcadores con alta diversidad genética Magenta: Marcadores con mayor Heterocigocidad observada *Ho: Heterocigocidad Observada
54

Todos los marcadores fueron polimórficos; se destacaron BM143 y BM187 por
presentar el número más alto de alelos dentro de la población de estudio, lo cual
coincide con el estudio de razas andinas (Diaz, 2005a), en el que se reporta al
marcador BM143 como muy polimórfico. Estos dos marcadores junto con GATs91
presentan también los valores más altos de diversidad genética.
Los marcadores BM188b y BMd01 presentaron alto número de individuos
heterocigotos (tabla 10), lo que indica que estos marcadores son menos útiles para
estudios en un futuro por la posible presencia de bandas inesperadas que pueden
inflar el resultado de la Heterocigocidad observada, ya que la mayoría de
marcadores presentan este valor por debajo de 0.20; otros marcadores
problemáticos pueden ser: BM143, BM156, BM187, BMd17, BMd20, Pv-ctt001 y Pv-
gaat001, por presentar altos valores para este índice.
Al hacer la comparación del promedio de número de alelos entre los dos tipos de
marcadores utilizados, es posible observar que los genómicos presentan valores
más altos que los marcadores génicos. De la misma forma el índice de diversidad
genética fue mayor para los genómicos. Estos resultados son coherentes y
corresponden con los presentados por Blair y colaboradores y Díaz & Blair en 2006,
quienes reportan que los marcadores genómicos presentan mayor número de alelos
y diversidad genética. Por otra parte se observa que el número de alelos en los
marcadores genómicos fluctúa entre 3 y 40, mientras que en los génicos presentan
valores muy similares, es decir no están variando de la misma forma que los
genómicos.
Para los marcadores Pv-cct001 (génico) y AG01 (genómico) se encontraron los
valores más bajos de diversidad genética, sin embargo es posible observar una
amplia diferencia numérica entre estos dos valores, mostrando que los marcadores
genómicos tienen diversidad mucho más alta. Por otra parte, los marcadores BM187
(genómico) y BMd01 (génico) presentaron el número máximo de alelos encontrados
para un locus dado dentro del conjunto completo de genotipos; se aprecia una
amplia diferencia numérica entre los dos tipos de marcadores.
Se realizó una prueba F para comparar las varianzas de los parámetros de
diversidad para los dos tipos de marcadores; esta prueba demostró que las
55

varianzas no son estadísticamente diferentes. Luego se realizó una prueba t de
comparación de medias para los mismos parámetros y se encontró que la
diversidad genética promedio para los marcadores génicos difiere significativamente
del promedio encontrado para los genómicos. En la tabla 11 se muestran los
resultados para cada parámetro dentro de cada tipo de marcador, la prueba
completa se muestra en el anexo 5.
Tabla 11. Prueba t para comparar parámetros de diversidad (Heterocigocidad Observada y
Diversidad Genética) entre marcadores genómicos y génicos.
Tipo de marcador
N Het. Obs D. Genética
Prom Var Prom* Var Genómicos 16 0.2 0.02 0.7 0.034 Génicos 14 0.17 0.03 0.52 0.035
* : Promedios significativamente diferentes P<0.013 NOTA: en cada prueba realizada, hubo homogeneidad de varianza
6.1.1.3 Análisis de Ordenación
El análisis de ordenación permitió determinar de forma contundente la existencia y
separación de los dos acervos principales del fríjol común, en la colección de
germoplasma de Ruanda. Mediante el método de ordenación conocido como
Análisis de Coordenadas Principales (PCoA) y su representación gráfica en tres
dimensiones (Figuras 8 y 9), se mostró la presencia de agrupamientos dentro de los
genotipos analizados, los cuales tienen correspondencia con la estructura de
acervos Andino y Mesoamericano propuesta en diversos estudios morfológicos
(Singh et al., 1991b), bioquímicos (Gepts & Bliss, 1986; Koenig & Gepts, 1989) y
moleculares (Beebe et al., 2001; Beebe et al., 2000b; Blair et al., 2006; Díaz & Blair,
2006). La representación gráfica en tres dimensiones permite observar la estructura
y las distancias geométricas del conjunto de los genotipos estudiados, definidas por
la proporción de alelos compartidos, siendo esto un reflejo de la similaridad genética
existente entre los mismos. Una de las grandes ventajas de este tipo de gráficos en
3D es que se puede observar las relaciones de cada genotipo con todos los demás
del estudio, siendo ésta representación mucho más didáctica y menos lineal que los
dendrogramas en los cuales no se permite observar todas las diferentes relaciones
que ocurren entre uno y todos los genotipos; es posible decir que las distancias que
manejan los dendrogramas pueden estar solapando otras importantes ya que
56

muestran una agrupación de un individuo a un grupo específico pero teniendo en
cuenta solo un criterio de distancia (Comunicación Personal M.C Duque consultora
estadística, CIAT 2007).
DOR 364
ICA‐PIJAO
CALIMA
G19833
CONTROLESAndino
Meso I
Meso II
Meso III
Meso IV
Introgresados
Individuo Raro 1
Individuo Raro 2
Figura 8. Representación gráfica en tres dimensiones de la distribución y ubicación espacial
de los genotipos de Ruanda a partir del Análisis de Coordenadas Principales (PCoA),
teniendo en primer plano horizontal la dimensión 2. Controles de cada acervo encerrados en
círculos. El grupo señalado corresponde un posible grupo de individuos introgresados.
Las accesiones utilizadas como controles, andinas (círculos azules) y
Mesoamericanas (círculos rojos), se ubicaron respectivamente como se esperaba
57

en los diferentes grupos, Calima y G19833 dentro del grupo Andino y en el grupo de
los Mesoamericanos DOR364 e ICA-PIJAO.
Figura 9. Representación gráfica en tres dimensiones de la distribución y ubicación espacial
de los genotipos de Ruanda a partir del Análisis de Coordenadas Principales (PCoA),
teniendo en primer plano horizontal las dimensiones 1 y 2
De acuerdo a lo visualizado en la figura 8 en la que se tiene como primer plano la
dimensión 2, se puede describir la forma en la cual se distribuye la variabilidad de la
siguiente manera: la dimensión 2 separa por completo los acervos Andino y
Mesoamericano dejando en el centro algunos individuos que posiblemente sean
candidatos de introgresión ya que tienden a fluctuar entre los dos grupos
principales, algunos de estos genotipos corresponden al grupo de Mesoamericanos
II (trébol verde encerados por una línea azul) y los otros son los genotipos
representados con cruces azules.
58

A diferencia del grupo de Andinos, dentro del acervo Mesoamericano se puede
observar que hay una subdivisión a nivel de la dimensión 3 que parte a este grupo
en cuatro: Mesoamericanos I (círculo azul), Mesoamericanos II (trébol verde),
Mesoamericanos III (Rombo fucsia) y Mesoamericanos IV (estrella negra). Sin
embargo comparten elementos en las dimensiones 1 y 2 razón por la cual se
encuentran agrupados dentro del gran grupo Mesoamericano, reflejando similaridad
en su composición alélica.
El grupo de las cruces constituye individuos muy diversos que pueden estar en parte
relacionados con el grupo de Mesoamericanos I y con los Andinos puesto que están
a la misma altura y forma un puente a través de la dimensión 2. Además, estos
individuos pueden estar compartiendo características con los grupos de los
Mesoamericanos III y nuevamente los Andinos, ya que se observa en la dimensión 2
la aparición de parte de estos individuos dentro de estos grupos. Por lo tanto, de
acuerdo a esta distribución en estos diagramas este grupo (cruces azules) puede
corresponder a eventos de introgresión
En la figura 9 los individuos candidatos a ser determinados como introgresados se
muestran dispersos en el diagrama, entrando al conjunto de los dos acervos
principales. Por otra parte, se puede observar que los dos individuos RW226 y
RW269 se consideran como raros debido a que en esta perspectiva se muestran
como individuos completamente separados en el estudio; en la figura 10 se muestra
su registro fotográfico, se caracterizan por ser de grano pequeño de color rojo,
características típicas del acervo mesoamericano.
RW226 RW269
Figura 10. Individuos considerados como raros a partir del análisis de coordenadas principales.
59

Por otra parte, a partir del análisis de coordenadas principales se elaboró un
dendrograma (figura 11) en el que se muestran la separación y definición de dos
grupos principales con una similaridad promedio de 0.85. De acuerdo con los
controles de diversidad que se tuvieron en cuenta fue posible establecer a que
acervo pertenecían los genotipos en estudio. Es posible determinar la
predominancia del acervo Mesoamericano con respecto al Andino, con un total de
208 genotipos contra 132; el resto de individuos son los que posiblemente sean
introgresados, sin embargo no se ubican en esta figura ya que a la distancia de
similaridad de 0.85 solo se dividen los dos acervos principales.
Coeficiente de distancia euclidiana
0.00 0.25 0.50 0.75 1.00
1 0 3 MW
ICA
DOR
G19
CAL
Me
so
am
eri
can
os
An
din
os
Figura 11. Dendrograma de acervos genéticos obtenido por distancia euclidiana aplicada a
coordenadas obtenidas por PCoA. En la parte superior izquierda en rojo se encuentra el
grupo Mesoamericano con sus respectivos controles en azul y en la inferior el grupo Andino
en verde y sus controles en amarillo.
60

En los dendrogramas de las figuras 12A, 12B y 12C se determina de manera más
detallada las agrupaciones formadas dentro de cada acervo, se observa que el
grupo de los Andinos no sufrió una subdivisión mientras que el Mesoamericano tuvo
4. Con fines ilustrativos la gráfica se dividió en tres partes.
0.00 0.25 0.50 0.75 1.00
103 438
596
257 595
134 400
175
266 465
500 586
151
258 415
152 158
606
273 181
211 556
582
238 518
207 77
557
59 563
110 119
145
96 440
112
122 121
124 128
126
143 185
240 267
328
147 605
270 87
166
187 429
436 242
95
156 572
21 550
23
39 421
38 530
408
445 284
285 101
136
492 140
196 153
186
102 441
682 73
76
69 71
133 188
434
679 61
157 277
64
180 176
182
235 72
244 74
177
66 184
62 70
428
150 444
410 234
281
676 105
107 142
287
137 149
209 141
146
192
M I
M III
Coeficiente de distancia euclidiana
MI: Mesoamericanos grupo 1 MII: Mesoamericanos grupo 2 MIII: Mesoamericanos grupo 3 MIV: Mesoamericanos grupo 4 A: Grupo de Andinos I: Grupo de Introgresados
Figura 12A. Dendrograma de acervos genéticos obtenido por distancia euclidiana aplicada a
coordenadas obtenidas por PCoA. Muestra dos grupos formados en el acervo
Mesoamericano (MI y MIII).
En la figura 12A se observan algunas discrepancias para genotipos de un grupo
determinado que fueron asignados en otro diferente; por ejemplo dos individuos
(marcados con color fucsia) del MIII se ubican dentro de MI (marcados con color
azul claro). Algunos individuos que aparecieron como posibles candidatos a ser
61

introgresados en las figuras 8 y 9 se ubicaron dentro de los grupos de
Mesoamericanos I y IV, (fig 12A se señalan con líneas azules oscuras gruesas).
0.00 0.25 0.50 0.75 1.00
103MW
199 144
109
139 154
278 459
575
127 16
243 155
ICA
104 274
259 12
597
170 DOR
114 115
275
116 118
130 319
320
236 117
129 131
312
58 570
159
81 94
212 82
80
93 88
271 24
260
25 564
461 647
272
509 255
507 171
172
568 174
92 10
17
15 574
11 554
165
224 539
567 90
19
1 27
28 540
269
603 26
3 601
37
536 522
573 20
620
667 247
498 279
655
426 613
604
268 611
357 678
226
162 44
36 41
40
193 386
227 231
4
89 204
32 31
45
229 526
5 35
552
523
M II
M IV
A
I
Coeficiente de distancia euclidiana
M-I?
MI: Mesoamericanos grupo 1 MII: Mesoamericanos grupo 2 MIII: Mesoamericanos grupo 3 MIV: Mesoamericanos grupo 4 A: Grupo de Andinos I: Grupo de Introgresados Individuos raros: Líneas rojas en MIV y en I.
Figura 12B. Dendrograma de acervos genéticos obtenido por distancia euclidiana aplicada a
coordenadas obtenidas por PCoA. Muestra los grupos formados en el acervo
Mesoamericano (MII y MIV), al igual que un grupo de introgresados y parte de los andinos y
un posible grupo nuevo de introgresados que se separa de MII.
En el dendrograma 12B se ve delimitado el grupo de los MII (color verde) dentro del
cual se ubican los dos controles; al igual que en las gráficas 8 y 9, se puede
observar que un grupo de individuos de MII se separó del gran grupo y se ubican
hacía la posición de los Andinos muy cerca de los introgresados; lo que podría
sugerir que las dos gráficas muestran a estos individuos como un posible nuevo
62

grupo de introgresados, sugiriendo flujo de genes entre acervos. Por otra parte en la
representación del dendrograma 12B se ubican los individuos raros como líneas
rojas dentro del grupo de MIV y dentro de un grupo de posibles Introgresados.
0.00 0.25 0.50 0.75 1.00
103MW
475 201
220
6 253
325 479
78
55 7
9 219
609
630 385
397 404
221
538 249
290 303
298
299 625
481 216
302
516 310
454 316
468
389 640
514
205 223
333 218
206
297 54
8 214
373
33 340
34 215
232
217 321
383 375
335
542 359
336 371
288
291 368
341 50
345
338 296
47 G19
191
476 399
246 289
480
245 393
474 636
631
637 634
295 194
497
311 56
329 377
46
49 478
283
292 464
48 384
672
282 332
376 374
CAL
370 390
394 379
301
164 230
674 30
189
43 326
163 528
642
A
Coeficiente de distancia euclidiana
MI: Mesoamericanos grupo 1 MII: Mesoamericanos grupo 2 MIII: Mesoamericanos grupo 3 MIV: Mesoamericanos grupo 4 A: Grupo de Andinos I: Grupo de Introgresados
Figura 12C. Dendrograma de acervos genéticos obtenido por distancia euclidiana aplicada a
coordenadas obtenidas por PCoA. Muestra dos grupos formados en el acervo
Mesoamericano (MI y MIII).
El grupo de los genotipos pertenecientes al acervo Andino, líneas de color rosado
pálido, se ubica en parte de la figura 12B y se completa este grupo en la figura 12C;
para este estudio este acervo no tiene subdivisión; los controles están mucho más
separados que los controles Mesoamericanos lo que puede sugerir la presencia de
razas diferentes. Mientras tanto hay una agrupación de individuos mayormente
introgresados que se ubican dentro de los andinos al final de la figura 12C.
63
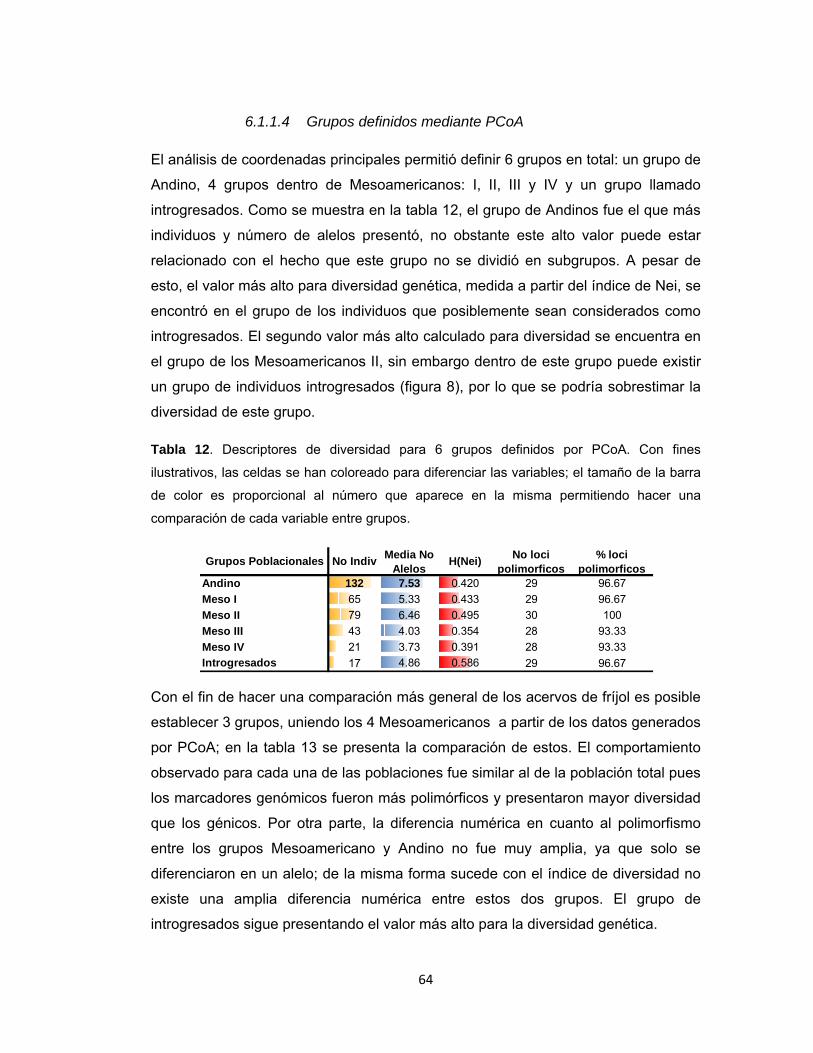
6.1.1.4 Grupos definidos mediante PCoA
El análisis de coordenadas principales permitió definir 6 grupos en total: un grupo de
Andino, 4 grupos dentro de Mesoamericanos: I, II, III y IV y un grupo llamado
introgresados. Como se muestra en la tabla 12, el grupo de Andinos fue el que más
individuos y número de alelos presentó, no obstante este alto valor puede estar
relacionado con el hecho que este grupo no se dividió en subgrupos. A pesar de
esto, el valor más alto para diversidad genética, medida a partir del índice de Nei, se
encontró en el grupo de los individuos que posiblemente sean considerados como
introgresados. El segundo valor más alto calculado para diversidad se encuentra en
el grupo de los Mesoamericanos II, sin embargo dentro de este grupo puede existir
un grupo de individuos introgresados (figura 8), por lo que se podría sobrestimar la
diversidad de este grupo.
Tabla 12. Descriptores de diversidad para 6 grupos definidos por PCoA. Con fines
ilustrativos, las celdas se han coloreado para diferenciar las variables; el tamaño de la barra
de color es proporcional al número que aparece en la misma permitiendo hacer una
comparación de cada variable entre grupos.
Grupos Poblacionales No Indiv Media No Alelos H(Nei) No loci
polimorficos% loci
polimorficosAndino 132 7.53 0.420 29 96.67Meso I 65 5.33 0.433 29 96.67Meso II 79 6.46 0.495 30 100Meso III 43 4.03 0.354 28 93.33Meso IV 21 3.73 0.391 28 93.33Introgresados 17 4.86 0.586 29 96.67
Con el fin de hacer una comparación más general de los acervos de fríjol es posible
establecer 3 grupos, uniendo los 4 Mesoamericanos a partir de los datos generados
por PCoA; en la tabla 13 se presenta la comparación de estos. El comportamiento
observado para cada una de las poblaciones fue similar al de la población total pues
los marcadores genómicos fueron más polimórficos y presentaron mayor diversidad
que los génicos. Por otra parte, la diferencia numérica en cuanto al polimorfismo
entre los grupos Mesoamericano y Andino no fue muy amplia, ya que solo se
diferenciaron en un alelo; de la misma forma sucede con el índice de diversidad no
existe una amplia diferencia numérica entre estos dos grupos. El grupo de
introgresados sigue presentando el valor más alto para la diversidad genética.
64

Tabla 13. Descriptores de diversidad (Número de alelos y Diversidad genética de Nei) para
30 loci SSR en población definida a partir de PCoA. Con fines ilustrativos, las celdas se han
coloreado para diferenciar las variables; el tamaño de la barra de color es proporcional al
número que aparece en la misma y permite hacer una comparación de cada variable entre
marcadores génicos y genómicos.
Marcador No de Alelos H(Nei) No de Alelos H(Nei) No de Alelos H(Nei)AG01 6 0.0982 3 0.4954 3 0.4239BM139 7 0.0801 8 0.7836 7 0.4446BM143 20 0.8181 24 0.8845 7 0.7949BM156 10 0.7818 13 0.8462 8 0.8183BM160 9 0.2086 15 0.8788 9 0.6522BM165 8 0.8155 7 0.5212 6 0.7889BM172 6 0.0613 8 0.7876 6 0.4014BM175 10 0.6158 7 0.5605 7 0.7093BM183 11 0.7301 11 0.5437 4 0.6436BM187 17 0.8706 35 0.9320 9 0.8547BM188-a 4 0.5454 2 0.1400 3 0.6016BM188-b 15 0.8604 12 0.7775 12 0.84BM201 9 0.6602 6 0.1145 5 0.6348BM205 6 0.4284 5 0.4955 5 0.7266GATs54 3 0.4355 3 0.3585 2 0.3599GATs91 16 0.8516 12 0.7799 10 0.8564BMd01 15 0.7353 13 0.5996 7 0.8244BMd02 2 0.2523 2 0.0301 2 0.4922BMd08 5 0.5746 2 0.2829 3 0.6367BMd16 6 0.3910 5 0.1940 3 0.4941BMd17 5 0.6146 4 0.1288 4 0.6436BMd18 5 0.3981 3 0.1783 3 0.2145BMd20 5 0.5491 3 0.1168 3 0.6228BMd45 2 0.0959 4 0.0965 2 0.4844BMd46 3 0.3085 2 0.2159 3 0.5547BMd47 3 0.2320 3 0.0543 2 0.4983Pv-ctt001 5 0.6460 4 0.4873 4 0.6436Pv-ag003 3 0.2584 1 0.0000 2 0.4297Pv-gaat001 7 0.4253 7 0.5162 4 0.5069Pv-cct001 4 0.0239 2 0.0239 1 0Suma Total alelos 227 226 146Media Genómicos 9.813 0.554 10.688 0.619 6.438 0.659Media Génicos 5.000 0.393 3.929 0.209 3.071 0.503MEDIA TOTAL 7.567 0.479 7.533 0.427 4.867 0.587
Mesoamericanos Andinos Introgresados
6.1.2 Posibles eventos de introgresión entre acervos
A partir del análisis de coordenadas principales se puede observar que existe un
grupo de individuos que pueden describirse como eventos de introgresión. En las
figuras 8 y 9 se muestran con un ícono azul en forma de cruz y se ven claramente
distribuidos en la mitad de cada gráfico, es decir fluctuando entre los dos grupos
principales.
65

Aunque en este estudio no se tengan los datos morfoagronómicos suficientes que
permitan establecer de manera definitiva rasgos aceptables para hablar de
introgresión, es posible observar que a pesar de que algunos individuos presenten
ciertas características morfológicas de semillas típicas de un acervo, estos son
clasificados o agrupados dentro de otro como respuesta a su perfil molecular
evaluado en 30 loci. Por ejemplo, en la figura 13a los individuos muestran
caracteres de semilla propios del acervo Mesoamericano (tamaño pequeño), sin
embargo en el dendrograma mostrado en la figura 12C se agrupan dentro del
acervo Andino.
RW 105 RW 107
RW 142
RW 284
RW 285 RW 426RW 268 RW 357 RW 604
RW 611 RW 613 RW 678
RW 30 RW 43 RW 230
RW 301 RW 674a
c
b
d
Figura 13. Individuos que se presentan como posibles eventos de introgresión dentro del
estudio; se divide la imagen de acuerdo con la agrupación observada en el dendrograma. a)
Individuos agrupados dentro de acervo Andino; b) Individuos agrupados dentro de acervo
Mesoamericano grupo I; c) Individuos agrupados dentro de acervo Mesoamericano grupo II;
d) Individuos agrupados dentro de acervo Mesoamericano pero no incluido dentro de ningún
grupo de Mesoamericano.
66

En el caso de los demás individuos presuntos de ser introgresados, se puede
observar (figura 13 b, c y d) que morfológicamente su semilla tiene características
típicas del acervo Mesoamericano (semilla de tamaño mediano-pequeño) y en
efecto en el dendrograma (figura 12A y B) se agrupan dentro de algunos grupos de
este acervo; no obstante se puede presumir a partir de los datos moleculares
obtenidos, que estos genotipos presentan algún tipo de introgresión ya que al
realizar el análisis de ordenación PCoA, éstos no se encuentran definidos dentro de
un grupo en particular sino que fluctúan entre los dos acervos principales (como se
muestra en las figuras 8 y 9).
Dentro de este mismo contexto, es de gran importancia resaltar la posible aparición
de un segundo grupo de introgresión, señalado en la gráfica 8 con un circulo azul.
Algunos individuos que se agruparon dentro del grupo de Mesoamericano II (a partir
del análisis de coordenadas principales), se encuentran formando un puente entre
éste y los andinos, como se señala en la misma gráfica, razón que permite pensar
que existe un posible flujo de genes entre acervos.
Se llevó a cabo un nuevo análisis de los parámetros de diversidad (tabla 14)
teniendo en cuenta el nuevo grupo de posibles introgresados, al cual se le ha
denominado Meso-Intro; como era de esperase, los valores no cambiarían dentro
de los grupos anteriormente analizados, excepto para el grupo de Mesoamericanos
II, debido a que se separaron 17 individuos de éste. El nuevo grupo formado (Meso-
Intro) tiene el segundo valor más alto de diversidad en comparación con los
Mesoamericanos y Andinos, lo cual resulta interesante ya que los dos grupos con
diversidad más alta son los que posiblemente sean considerados como
Introgresados.
La diversidad y el promedio de número de alelos en el grupo Mesoamericano II
disminuyen cuando se separa el grupo de posibles introgresados, esto se muestra
en la parte inferior de la tabla 14, al igual que la comparación de valores entre el
grupo general de los Mesoamericanos antes de la separación de Meso-Intro y
después de esta.
67

Tabla 14. Análisis de los parámetros de diversidad (Número de alelos y Diversidad genética de Nei) para 7 grupos formados incluyendo el nuevo grupo considerado como introgresado (Mesi-Intro). Se resaltan las columnas en azul claro para identificar los cambios obtenidos con respecto a los análisis realizados para 6 grupos. Se muestra una tabla adicional en la parte inferior que muestra los valores obtenidos en el grupo Meso II y Meso general, para los mismos parámetros, antes y después de la separación del grupo Meso-Intro.
Meso II Meso General
Separación Meso‐Intro Antes Después Antes Después
No Alelos 6.46 5.17 7.57 6.77
Div genética 0.49 0.44 0.48 0.46
Marcador No de Alelos
H(Nei) No de Alelos
H(Nei) No de Alelos
H(Nei) No de Alelos
H(Nei) No de Alelos
H(Nei) No de Alelos
H(Nei) No de Alelos
H(Nei)
AG01 4 0.089 3 0.078 2 0.045 1 0.000 3 0.495 3 0.424 4 0.395BM139 3 0.031 3 0.078 1 0.000 3 0.092 8 0.784 7 0.445 4 0.393BM143 14 0.835 12 0.735 13 0.833 4 0.364 24 0.885 7 0.795 8 0.649BM156 7 0.758 6 0.777 5 0.686 4 0.609 13 0.846 8 0.818 8 0.808BM160 5 0.145 4 0.122 3 0.246 2 0.047 15 0.879 9 0.652 7 0.666BM165 6 0.751 6 0.801 4 0.570 4 0.693 7 0.521 6 0.789 6 0.803BM172 3 0.030 2 0.032 1 0.000 3 0.092 8 0.788 6 0.401 3 0.348BM175 6 0.483 7 0.548 3 0.152 3 0.176 7 0.561 7 0.709 8 0.803BM183 9 0.695 7 0.667 5 0.602 6 0.594 11 0.544 4 0.644 8 0.796BM187 13 0.853 14 0.844 9 0.767 5 0.523 35 0.932 9 0.855 9 0.784BM188-a 3 0.480 4 0.541 4 0.112 3 0.177 2 0.140 3 0.602 2 0.360BM188-b 12 0.840 11 0.864 8 0.657 9 0.862 12 0.778 12 0.840 10 0.822BM201 7 0.580 8 0.670 5 0.102 3 0.395 6 0.115 5 0.635 5 0.662BM205 4 0.368 5 0.416 4 0.389 4 0.458 5 0.496 5 0.727 5 0.621GATs54 2 0.401 2 0.271 3 0.511 2 0.172 3 0.359 2 0.360 2 0.484GATs91 12 0.839 9 0.840 8 0.604 8 0.750 12 0.780 10 0.856 12 0.884BMd01 10 0.696 11 0.712 9 0.715 12 0.795 13 0.600 7 0.824 8 0.760BMd02 2 0.281 2 0.121 2 0.110 2 0.387 2 0.030 2 0.492 2 0.500BMd08 3 0.555 4 0.599 4 0.522 3 0.390 2 0.283 3 0.637 3 0.519BMd16 5 0.367 5 0.253 4 0.394 4 0.359 5 0.194 3 0.494 2 0.430BMd17 4 0.522 5 0.562 4 0.395 3 0.595 4 0.129 4 0.644 3 0.638BMd18 4 0.309 4 0.368 2 0.427 4 0.581 3 0.178 3 0.215 3 0.337BMd20 4 0.560 4 0.511 2 0.392 3 0.520 3 0.117 3 0.623 4 0.545BMd45 2 0.030 1 0.000 2 0.023 2 0.278 4 0.097 2 0.484 2 0.438BMd46 2 0.346 2 0.228 2 0.093 3 0.335 2 0.216 3 0.555 2 0.469BMd47 2 0.142 2 0.180 3 0.068 2 0.408 3 0.054 2 0.498 2 0.498Pv-ctt001 3 0.601 4 0.581 3 0.641 4 0.472 4 0.487 4 0.644 4 0.647Pv-ag003 3 0.119 3 0.177 2 0.357 2 0.091 1 0.000 2 0.430 2 0.469Pv-gaat001 5 0.301 2 0.453 2 0.206 3 0.526 7 0.516 4 0.507 4 0.548Pv-cct001 1 0.000 3 0.063 2 0.023 1 0.000 2 0.024 1 0.000 1 0.000Suma Total alelos 160 155 121 112 226 146 143Media Genómicos 6.875 0.511 4.875 0.392 4.000 0.375 10.688 0.619 6.438 0.659Media Génicos 3.571 0.345 3.071 0.312 3.429 0.410 3.929 0.209 3.071 0.503
MEDIA TOTAL 5.333 0.434 4.033 0.355 3.733 0.391 7.533 0.427 4.867 0.587
Meso I Andinos Introgresados Meso-IntroMeso II Meso III Meso IV
6.438 0.518 6.313 0.6423.714 0.343 3.000 0.4865.167 0.436 4.767 0.569
68

A continuación se muestra el registro fotográfico de los 17 individuos pertenecientes
al grupo denominado como Meso-Intro, se puede observar que dentro de este
registro es posible encontrar individuos con colores raros (grises) o semillas con
colores mezclados; estos rasgos podrían esperarse en un grupo de genotipos
introgresados.
RW3
RW247
RW26
RW279
RW20
RW498
RW536 RW601
RW667
RW37
RW573
RW522
RW655
RW620
RW163
RW528
RW642
Figura 14. Individuos pertenecientes a Meso-Intro (agrupados por PCoA como Meso II pero que posiblemente presenten eventos de introgresión)
6.1.3 Agrupación mediante índice de disimilaridad
En la figura 15 se muestra claramente la subdivisión de la población de estudio en
los dos acervos principales y posiblemente dos grupos de introgresados, lo cual
coincide con lo encontrado a partir del análisis de coordenadas principales. Se
encontró un total de 205 genotipos para el grupo Mesoamericano, 127 para el grupo
Andino, 15 para Introgresados I y 12 para Introgresados II; de tal manera es posible
decir que aunque no se recurra a la elaboración de una minuciosa comparación de
69

las dos metodologías (matriz de similaridad e índice de disimilaridad), si es posible
decir que la cantidad de individuos asignados para cada grupo es comparable entre
las dos formas de asociación, ya que sigue siendo mayor el número de individuos
para Mesoamericano. El agrupamiento de los controles también fue similar en las
gráficas generadas por ambas metodologías, ya que en el caso de los Meso, éstos
aparecen más unidos que los andinos.
Figura 15. Dendrograma basado en el índice de disimilaridad de DICE para la colección de
germoplasma de Ruanda. Muestra en verde (parte superior) al grupo de los
mesoamericanos y sus dos respectivos controles en azul; en la parte inferior en rojo el grupo
de los andinos y sus controles en amarillo. Los dos posibles grupos de introgresados se
observan en la mitad de la gráfica.
70

6.2 ANÁLISIS DE MINERALES
6.2.1 Contenido de minerales en semilla de fríjol
En la evaluación de minerales de los 355 genotipos de la colección de germoplasma
de fríjol de Ruanda, mediante la técnica de Absorción Atómica, fue posible observar
que los valores para el contenido de hierro estuvieron en el rango esperado en
semilla de fríjol de acuerdo con estudios previos (Beebe et al., 2000a; Blair et al.,
2005; Islam et al., 2002a); sin embargo al comparar el contenido promedio se
encuentra que para este estudio los valores de hierro resultaron más altos que los
encontrados en los trabajos mencionados, excepto por el de Blair et al. (2005)
quienes obtuvieron un valor de 83.3 ppm. En este caso se hallaron 68.24 ppm,
como se muestra en la tabla 15, mientras que en el trabajo de Islam et al. 2002 el
contenido de hierro correspondió a 54.8 ppm y en Beebe et al. (2000) fue de 55ppm.
Tabla 15. Estadística descriptiva para contenido de (ppm) de hierro y zinc en 355
accesiones de frijol provenientes de Ruanda.
Fe (ppm) Zn (ppm)
Contenido Medio 68.24 34.46
Desviación Estándar 8.41 3.39
Varianza 70.74 11.50
Mínimo 45.35 25.12
Máximo 97.55 49.05
Rango 52.20 23.92
Coeficiente de Variación 12.33 9.84
Curtosis -0.0047 0.7471
En la tabla 15 se observa que los resultados de zinc obtenidos para este estudio
(34.5ppm) fueron congruentes con los reportados en los estudios anteriores. En su
investigación Blair y colaboradores (2005) encontraron un promedio de 32.5 ppm,
71
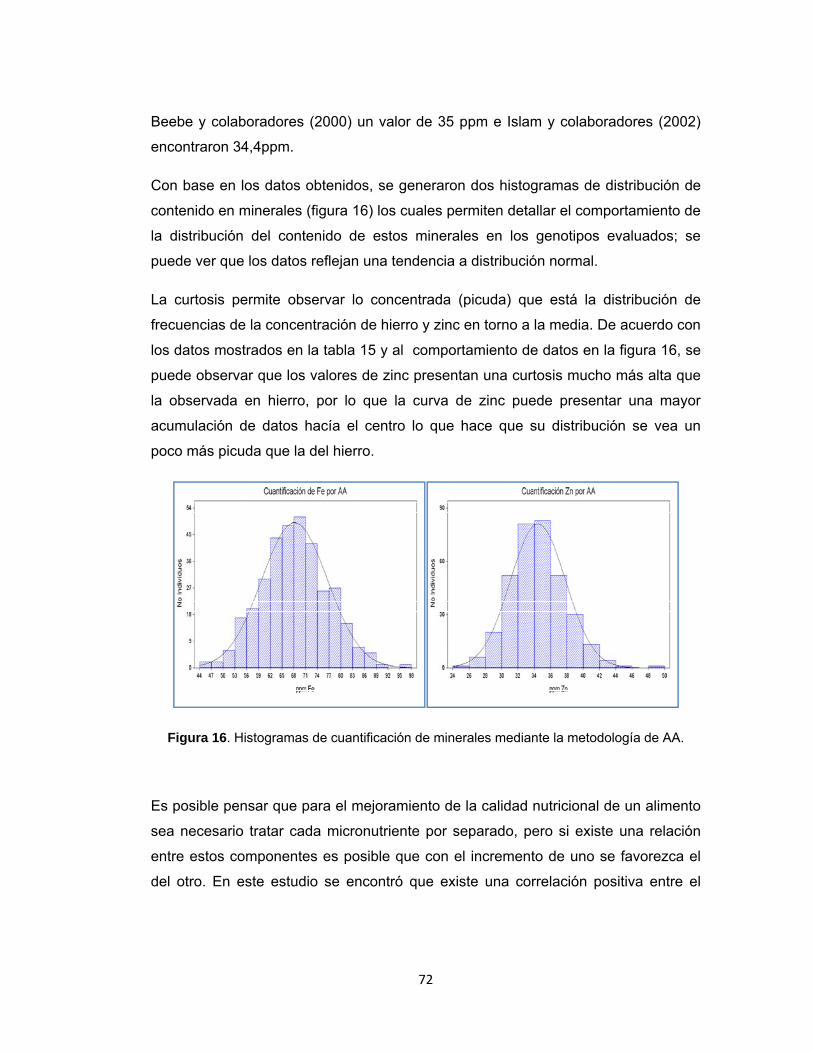
Beebe y colaboradores (2000) un valor de 35 ppm e Islam y colaboradores (2002)
encontraron 34,4ppm.
Con base en los datos obtenidos, se generaron dos histogramas de distribución de
contenido en minerales (figura 16) los cuales permiten detallar el comportamiento de
la distribución del contenido de estos minerales en los genotipos evaluados; se
puede ver que los datos reflejan una tendencia a distribución normal.
La curtosis permite observar lo concentrada (picuda) que está la distribución de
frecuencias de la concentración de hierro y zinc en torno a la media. De acuerdo con
los datos mostrados en la tabla 15 y al comportamiento de datos en la figura 16, se
puede observar que los valores de zinc presentan una curtosis mucho más alta que
la observada en hierro, por lo que la curva de zinc puede presentar una mayor
acumulación de datos hacía el centro lo que hace que su distribución se vea un
poco más picuda que la del hierro.
Figura 16. Histogramas de cuantificación de minerales mediante la metodología de AA.
Es posible pensar que para el mejoramiento de la calidad nutricional de un alimento
sea necesario tratar cada micronutriente por separado, pero si existe una relación
entre estos componentes es posible que con el incremento de uno se favorezca el
del otro. En este estudio se encontró que existe una correlación positiva entre el
72

hierro y el zinc, la cual es estadísticamente significativa a lo largo de los diferentes
genotipos, su valor es de 0.52 (P=0.0000).
A partir del rango de valores determinados para la interpretación de resultados de
cuantificación de minerales (ver tabla 9), se estableció que los genotipos (figura 17a)
con el contenido más alto de hierro fueron: RW 582, RW 375, RW 221, RW 500, RW
216, RW 28 y RW 298; de la misma forma se encontraron que los que presentaban
valores más bajos fueron RW 77, RW 32 y RW 40, RW 95 y RW 76 (figura 17c).
48.43 ppm
RW 40
45.81 ppm
RW 32
45.35 ppm
RW 77
50.45 ppm
RW 76
48.88 ppm
RW 95
RW 582
97.55ppm
RW 375
89.45ppm
RW 221
87.46ppm
RW 500
86.7ppm
RW 216
86.55ppm
RW 28
86.48ppm
RW 298
86.36ppm
RW 582
49.05ppm
RW 6
44.94ppm
RW 257
43.56ppm
RW 88
42.91ppm
RW 149
42.70ppm
RW 492
42.54ppm
RW 500
41.91ppm
RW 345
25.12ppm
RW 542
26.30ppm
RW 8
26.56ppm
a b
c d
Figura 17. Genotipos con mayor y menor contenido de hierro y zinc: (a) Genotipos con
contenido más alto de Fe; (b) Genotipos con contenido más alto de Zn; (c) Genotipos con
contenido más bajo de Fe; (d) Genotipos con contenido más bajo de Zn.
73

Para el zinc los genotipos que se muestran en la figura 17b corresponden a los de
mayor contenido: RW 582, RW 6, RW 257, RW 88, RW 149, RW 492 y RW 500, y
los de la figura 17d, son los de contenido más bajo: RW 345, RW 542 y RW 8. Sin
embargo, de acuerdo con el rango para interpretación, ningún valor de
concentración de zinc se encontró clasificado como bajo, ya que dicho rango
sugiere que los genotipos se deben determinar con contenido bajo cuando
presentan valores menores a 20 ppm y para este estudio todos los genotipos
contenían concentraciones superiores a 25 ppm.
Es importante resaltar que el individuo RW582 presentó los valores más altos para
estos dos minerales, por lo tanto es un genotipo digno de tener en cuenta en
programas de mejoramiento. Lo mismo ocurre con el individuo RW500 que estuvo
dentro de los genotipos con altos valores para ambos minerales.
6.2.2 Asociación contenido de minerales y estructura genética
De acuerdo con los datos de contenido de minerales obtenidos y con las
agrupaciones formadas a partir de PCoA, se puede decir que el grupo de los
genotipos introgresados presentó el valor numérico de contenido medio de hierro
más alto de toda la colección, aunque el genotipo de mayor contenido se localizó en
el grupo de Mesoamericanos IV. En la comparación entre grupos Andino y
Mesoamericano se observa que el primero tiene mayor valor numérico en contenido
de hierro que cualquiera de las agrupaciones formadas para el segundo. Sin
embargo se encontró que estos datos no son significativamente distintos entre sí
(ver prueba ANOVA en anexo 6).
Por el contrario en el caso del zinc se encontró que los valores entre grupos difieren
significativamente entre sí (tabla 16 y anexo 6); al realizar las pruebas de separación
de medias de rango múltiple se encontró que los introgresados no son diferentes de
ningún grupo de los mesoamericanos pero sí es diferente del grupo andino. En
general los andinos resultaron muy diferentes de los demás lo cual era de esperarse
debido que los acervos genéticos pueden diferir bastante en características
fenotípicas como genotípicas; sin embargo como contraste a esto se encontró que
no son tan diferentes con el grupo de Mesoamericanos IV.
74

En general todos los grupos presentan individuos de alto y bajo hierro y de alto y
medio contenido de zinc. Dentro del grupo de los Andinos se presentaron los
genotipos con contenido más bajo de zinc y el promedio general de este grupo fue
el más bajo con respecto a los demás, lo cual está de acuerdo con lo reportado por
Islam 2004 quien encontró el mismo patrón.
Tabla 16. Estadística descriptiva para contenido en ppm de hierro y zinc en las poblaciones
generadas mediante PCoA.
ANDINO MESO I MESO II MESO III MESO IV INTRO ANDINO MESO I MESO II MESO III MESO IV INTROn 132 65 79 43 21 17 132 65 79 43 21 17
Media 69.19 68.59 67.43 66.65 65.49 70.96 35.7733.02 35.54 35.48 33.98 35.11Desv estd 8.39 8.66 7.07 10.35 7.99 7.64 3.10 3.25 2.82 3.83 3.00 3.50Varianza 70.39 75.05 49.99 107.08 63.88 58.41 9.62 10.55 7.93 14.68 9.00 12.25Mínimo 45.81 45.35 51.86 50.45 52.04 59.04 25.12 29.90 28.42 31.07 29.15 27.93Máximo 89.45 86.73 84.10 97.55 86.48 84.40 44.94 43.56 42.91 49.05 39.51 39.56Rango 43.64 41.38 32.24 47.10 34.44 25.36 19.81 13.66 14.49 17.97 10.36 11.63
CV 12.13 12.63 10.49 15.53 12.20 10.77 9.39 9.14 7.93 10.71 8.83 9.97*: Promedios significativamente diferentes p<0.0001
HIERRO ZINC*
A partir de las pruebas de separación de medias de rangos múltiples se pudo
observar que existe un gradiente de distribución de concentración de zinc a lo largo
de la dimensión 3, como se observa en la figura 18.
35.77
35.53
35.48
35.10
Figura 18. Gradiente de concentración de zinc encontrado en grupos mesoamericanos
indicado por las flechas azules.
75

66.3 CARAACTERISTICCAS DEL GRRANO DE FFRÍJOL
L
t
d
q
p
ú
Las caracter
tamaño y pa
de la semilla
que pudo h
puede ver
únicamente
rísticas de la
atrón de colo
a se pudo v
haber gener
reflejado en
bajo criterio
a semilla ten
oración. Sin
ver afectada
rando un po
n el color o
o visual.
nidas en cue
n embargo e
a por el largo
osible proce
observado.
enta para es
es necesario
o tiempo qu
eso de oxid
El tamaño
ste estudio fu
o aclarar que
ue llevaba a
dación que
de semilla
ueron el colo
e la coloraci
lmacenada,
finalmente
fue estima
or,
ón
lo
se
do
S
c
g
v
s
r
p
Fp
L
f
e
g
t
e
g
Se encontró
color café el
granos de c
vio fuerteme
semillas de
representac
para la sem
Figura 19. D
población de
La distribuc
formado a p
el anexo 7)
grupos, mie
tamaño med
estuvo hom
grande fue e
14%
1
ó gran varie
l que se pres
color rojo no
ente marcad
e un solo c
ión del 14%
illa de peque
Distribución po
estudio.
ción de colo
partir de PCo
). Los color
entras que e
diano y peq
mogéneamen
exclusiva de
26%
25%17%
12%
2% 2% 1%
dad de colo
sentó en ma
o fue muy am
do por semill
color, segui
%. En cuant
eña como pa
orcentual de
or, tamaño
oA se mues
res café, am
el color mora
queño se pre
nte distribuid
e los Andino
1% Color de
Café
Rojo
Ama
Crem
Neg
76
ores dentro
ayor proporc
mplia. El pa
las sin patró
ido por el
to al tamaño
ara la media
de la colecc
ción, sin emb
trón de colo
ón de colora
patrón mot
o se encont
ana (46%) y
ción (figura
bargo la dife
oración en la
ción definida
teado, el c
tró igual pro
solo un 8%
19), siendo
erencia con l
a colección
a, es decir p
cual tuvo u
oporción tan
de la grande
el
os
se
por
na
nto
e.
color, tamaño
y patrón d
stra en la fig
marillo y cre
ado fue exc
esentó en to
do en los M
os. Todos los
e semilla
é/Marrón
o
arillo
ma
ro
o y patrón de
e coloració
ura 20 (las
ema fueron
clusivo de lo
odos los gru
Mesoamerica
s grupos pre
11
3%
57%
Patrón de
e coloración d
n dentro de
tablas de co
constantes
os Andinos.
upos; el tam
anos I, II y
esentaron se
9%
14%
6%1%
Coloración
Jaspeado
Moteado
Punteado
Rayado
Venación
Nde semilla enP: Sin Patrón
n la
e cada gru
ontingencia
en todos l
La semilla
maño peque
III; la semi
emillas que
po
en
os
de
ño
illa
no

t
m
tenían un pa
moteado fue
atrón de colo
e constante y
oración es d
y el rayado e
decir eran de
exclusivo de
e un solo co
el grupo And
olor, sin emb
dino.
bargo el patrrón
0
20
40
60
80
100
120
140
No de Individu
os
0
20
40
60
80
100
120
140
No de
Individu
os
111
No de
Individu
os
Grupos pobla
Figura 20. D
acionales: MI: Meso
Distribución d
oamericanos grupo 1;
grupo 4; A: Gru
del color de la
MI MII M
Grup
MI MII
Grup
020406080100120140
MI MII
G
77
MII: Mesoamericanos
po de Andinos; I: Grup
a semilla dent
partir de PCo
MIII MIV A
po Poblacional
Color
MIII MIV A
po Poblacional
Tamaño
MIII MIV A
rupo Poblacional
Patrón
s grupo 2; MIII: Mesoa
po de introgresados
tro de cada gr
oA.
I
I
I
mericanos grupo 3; M
Amarillo
Rojo
Rosado
Negro
Morado
Gris
Crema
Café
Blanco
Pequeña
Mediana
Grande
NP
Venación
Rayado
Punteado
Moteado
Jaspeado
MIV: Mesoamericanos
rupo poblacioonal formado a

En los grupos Mesoamericanos hubo predominio de semilla pequeña de color café
con patrón de coloración moteado, con excepción del grupo cuatro que presentó un
mayor número de individuos con patrón de coloración jaspeado. Sin embargo el
tamaño de semilla mediano también tuvo una gran presencia dentro de estos grupos
Mesoamericanos, lo cual es congruente con estudios morfológicos que resaltan que
el tamaño típico de las semillas Mesoamericanas puede ser pequeño o mediano
(Gepts & Bliss, 1986).
Por otra parte las semillas pertenecientes al grupo Andino se caracterizaron por
presentar semillas de tamaño mediano (primordialmente) y grande, lo que
concuerda con estudios previos en semilla de fríjol (Singh et al., 1991a). El color
predominante fue el rojo moteado y tuvieron prevalencia de semilla mediana a
grande. En el grupo de los introgresados el color de semilla que más se encontró
fue el amarillo con patrón moteado y los tamaños mediano y pequeño fueron los que
se presentaron en este grupo.
78

7 DISCUSIÓN DE RESULTADOS
7.1 ANÁLISIS DE DIVERSIDAD GENÉTICA
El conocimiento de la diversidad permite tener una perspectiva amplia de la
estructura de las especies y ayuda a los mejoradores a encontrar genotipos con
características óptimas para la obtención de cultivos mejorados.
7.1.1 Población Total
Para el nivel de polimorfismo (número de alelos), se encontró en su mayoría valores
más altos que los obtenidos en estudios previos (Blair, 2007; Blair et al., 2006; Díaz
& Blair, 2006), lo cual podría estar indicando el buen desempeño de la técnica. Por
otra parte, es relevante tener en cuenta que tanto para el presente estudio como en
el de Moreno (2007), ambos realizados mediante la técnica de microsatélites
fluorescentes, se obtuvieron altos valores de número de alelos (301 y 429
respectivamente). Este factor junto con el hecho de que todos los loci sean
polimórficos podría verse explicado por los niveles de divergencia entre los
genotipos estudiados o por la sensibilidad, precisión y resolución de la técnica al
momento de reconocer polimorfismos; el secuenciador ABI 3730xl permite identificar
polimorfismos de hasta un solo par de bases, los cuales ocurren por mutaciones
puntuales en los extremos de la región microsatélite amplificada (Smith et al., 1997).
Los marcadores BM143, BM187 y GATs91 presentaron los números de alelos mas
altos para la población, lo que podría indicar que estos tienen alto poder de
discriminación, este hallazgo se apoya en la caracterización realizada por Gaitán et
al. (2002) quienes reportaron que dos de estos mismos loci (BM143 y GATs91) son
bastante útiles en el genotipaje de accesiones de germoplasma de P. vulgaris. Los
marcadores genómicos en promedio presentan este valor más alto que los génicos
lo que sugeriría que con los primeros se podría llevar a cabo una mejor
caracterización en colecciones similares a la del presente estudio.
La diversidad total del conjunto de genotipos evaluados, fue alta (0.62) al
compararse con los resultados obtenidos en estudios anteriores con otras clases de
79

marcadores. Por ejemplo con RFLPs se encontró un total de 0.38 (Becerra & Gepts,
1994), en este mismo se halló que mediante marcadores tipo isoenzima el valor de
la diversidad fue de 0.19. En otro estudio también realizado con isoenzimas el valor
correspondió a 0.159 (Paredes-Lopez & Gepts, 1995) y con CAP se encontró 0.37
(Chacón et al., 2005).
Un aspecto importante a resaltar es que este alto valor de diversidad genética,
posiblemente se deba al poder de resolución y gran cantidad de información que
poseen los microsatélites, ya que en general los altos valores en cuanto a dicho
parámetro, se encuentran para este tipo de marcadores; tal y como se muestran en
otras investigaciones en las que se evaluó la diversidad, por ejemplo Díaz J.M en el
2005, encontró un valor de 0.49 para 125 genotipos Andinos evaluados con 30
marcadores y Díaz, L. en el mismo año halló una diversidad de 0.47 para
accesiones Mesoamericanas. Sin embargo se sigue encontrando un valor mas alto
para estudios realizados con microsatélites fluorescentes.
Otro posible argumento del cual se podría inferir el hecho de que estos resultados
tuvieran altos valores en diferentes parámetros de diversidad fue que se usó un
subconjunto de primers con altos valores de PIC a partir de los trabajos realizados
por Gaitán y colaboradores (2002) y Blair y colaboradores (2006). El PIC
corresponde al contenido de información polimórfica y es un índice importante
debido a que permite determinar la capacidad discriminatoria de un grupo de
marcadores, y de esta forma poder reproducirlo en otros estudios en donde el
conocimiento y caracterización de la población sea el principal objetivo, como ocurre
en este estudio.
El número máximo de alelos encontrado en todo el conjunto de genotipos fue de 40
y correspondió al marcador BM187, este valor es alto comparado con estudios
previos como el elaborado por Blair et al (2006), en el que el número más alto que
se presentó fue 23 alelos. Al igual ocurre en el estudio de Díaz y Blair en el 2006,
quienes encontraron un número de alelos máximo de 15. Es posible hacer la misma
comparación para los valores promedio de número de alelos, ya que en este estudio
se presentaron 10 alelos en promedio para cada marcador, mientras que en
80

estudios previos fueron inferiores: 7.8 en Blair et al (2006) y 5.1 en Díaz & Blair
(2006).
Estos resultados son muy favorables si este estudio se encamina hacia objetivos de
mejoramiento genético, ya que la alta variabilidad confiere mayor probabilidad de
responder favorablemente a presiones de selección ambiental, como las que
comúnmente se presentan en Ruanda entre las que se encuentran enfermedades,
cambios climáticos drásticos, variación en la composición química del suelo entre
otros.
Por otra parte, se observó que en cuanto a la clase de marcadores evaluados los
genómicos permitieron identificar mayores niveles de diversidad y mayor número de
alelos. Las diferencias en el polimorfismo detectado entre los tipos de marcadores
también fue observada en varios estudios de diversidad genética en fríjol (Blair et
al., 2006; Blair et al., 2003; Diaz, 2005a; Díaz & Blair, 2006; Moreno, 2007), y puede
justificarse en el origen de cada clase de marcador, pues los microsatélites génicos
al provenir de secuencias codificantes, podrían no transmitir una modificación en su
secuencia a nuevas generaciones en el caso que dicha variante corresponda a un
cambio drástico o mutación ya que haría menos viable al individuo. En el caso de
los marcadores genómicos, que por estar principalmente en secuencias no
codificantes, pueden albergar mayor cantidad de variantes en la secuencia y no
comprometer la supervivencia o adaptación del organismo mutante (Diaz, 2005a).
En este estudio se encontró un valor alto de Heterocigocidad observada (Ho)
correspondiente a 0.19 comparado con el valor (0.038) obtenido para Díaz y Blair en
el 2006. Los marcadores que especialmente presentaron alta Heterocigocidad
observada son: el marcador genómico BM188b (0.65) y el génico BMd01 (0.61).
Este alto valor probablemente corresponde a una fuerte endogamia dentro del
objeto de estudio, lo que supone un enorme exceso de homocigotos en el conjunto
de accesiones y eso está en consonancia con fuerte endogamia dentro de las
accesiones (Comunicación Personal, M. Ruíz).
81

Sin embargo es necesario resaltar que los microsatélites fluorescentes permiten
identificar más fácilmente los polimorfismos, debido a la ya mencionada alta
sensibilidad; es posible que el nivel de Heterocigocidad observado, se haya visto
incrementado por esta causa, por lo que es necesario prestar mayor atención a
estos marcadores, en futuros estudios para no confundir el mérito de la técnica con
características de la población.
En general estos resultados reflejan valores altos en cuanto a parámetros de
diversidad y nivel de Heterocigocidad y pueden estar ocurriendo debido a las
mezclas de semillas que comúnmente se ve en las regiones en Ruanda. Aunque en
ciertos lugares y situaciones se haga selección de grano, en la mayoría de los casos
el consumo y producción de fríjol no se ve marcado por el tipo y tamaño de semilla
(Comunicación Personal L. Butare investigador programa fríjol Ruanda, CIAT 2007).
Las mezclas pueden ocurrir en diferentes momentos después de la cosecha, por
ejemplo cuando se ponen a secar los fríjoles bajo la incidencia de la luz solar,
resulta ser más atractivo para los campesinos ver mezcla de fríjoles, por lo tanto
ellos se encargan de organizarlas. También puede darse en el momento de la
colecta cuando recogen los granos pues hacen este proceso sin una selección
determinada; ó simplemente las mezclas pueden originarse por actividades
culturales como juegos, regalos o deudas (Martin & Adams, 1987).
El fríjol en Ruanda es producido tanto para la venta como para la alimentación
mediante sistemas de cultivos en mezcla durante dos estaciones al año. Las
semillas son cultivadas y cosechadas principalmente para consumo por parte de los
propios campesinos, sin embargo su venta es común especialmente cuando se
presentan problemas ambientales graves que afectan los cultivos o debido a los
pobres sistemas de almacenamiento de semilla que se tienen (David & Sperling,
1999).
En este proceso de venta y adquisición es cuando se maneja con más detalle el
tema de las mezclas de variedades. Los campesinos que venden la semilla se
encargan de escoger la que presente mejor calidad, lo cual garantiza el
mejoramiento del recurso genético pero no existe preferencia por un tipo de grano
(David & Sperling, op cit.). El conocimiento de los campesinos acerca de la
82

efectividad de las mezclas en la producción es amplio, ellos saben que este tipo de
método confiere mayor resistencia por parte de los genotipos a enfermedades o
fuertes cambios ambientales, siendo esta una posible causa del alto valor de la
variabilidad genética encontrado en la colección de este estudio.
Por otra parte, aunque el intercambio de semilla en forma de regalo o como red
comercial entre países ha disminuido en la actualidad (David & Sperling, op cit.),
este es un factor muy importante que podría estar afectando la diversidad, ya que
podría existir una recombinación de material genético confiriendo mayor variabilidad
en un mismo conjunto.
7.1.2 Grupos definidos mediante PCoA
Mediante el análisis de ordenación (PCoA) se mostró la presencia de grupos dentro
de los genotipos analizados, los cuales tienen correspondencia con la estructura de
acervos propuesta, esto permite inferir nuevamente la utilidad de la técnica para
diferenciar entre estos dos grupos.
El comportamiento observado para cada grupo definido a partir de PCoA fue similar
al de la población total ya que los marcadores genómicos presentaron mayor
polimorfismo que los génicos, lo que podría sugerir que las fuerzas existentes que
modelan cada patrón de cada grupo son similares (Diaz, 2005a). Sin embargo al
encontrar una mayor diversidad de los loci genómicos en el grupo de los
Introgresados insinúa que la composición alélica de los genotipos que lo conforman
puede ser diferente o tener diferentes frecuencias.
La alta diversidad total encontrada para todos los grupos generados por PCoA, en
especial para el grupo de los introgresados, podría demostrar que los agricultores
pueden estar contribuyendo al mantenimiento e incluso aumento de la diversidad a
través de entrecruzamientos intencionales o in intencionados, como ya ha sido
reportado por Martin y Adams (1987). El manejo post-cosecha en África juega un
papel importante en el mantenimiento y aumento de la variabilidad genética, ya que
en la mayoría de los casos los pequeños agricultores mezclan las semillas, lo que
podría aumentar las probabilidades de polinización cruzada.
83

Aunque en este estudio no se puede hablar de una estructura de razas debido a la
ausencia de datos que permitan confirmarlo, se puede tener en cuenta la
subdivisión del grupo Mesoamericano, infiriendo dentro de este gran grupo una
posible estructura de razas, que será resuelta en la medida en que sean utilizados
controles moleculares definidos para tal fin y que sea posible la obtención de
características, morfológicas, agronómicas y bioquímicas que permitan hacerlo.
Contrario a lo que se muestra en otros estudios en cuanto a representación de
semilla de los dos acervos principales (Blair et al., 2006; Islam et al., 2002b), el
mayor porcentaje de genotipos pertenecen al grupo de los Mesoamericanos (58.3%)
mientras que la representación andina correspondió al 37% y el resto hizo parte de
los introgresados. Es posible pensar que esto se deba a la implementación de
estrategias para combatir las amenazas a las que se ha visto enfrentada la
biodiversidad en Ruanda; dichas estrategias van encaminadas hacía la utilización
de los recursos genéticos para incrementar la productividad. Por ejemplo han
aumentado la siembra de variedades de semillas medianas y pequeñas que
presentan mayor tolerancia a las enfermedades de raíz (CIAT, 2005a) y a la sequía
(Martin & Adams, 1987).
Lo anterior puede ser confirmado por la incidencia de un alto porcentaje de semilla
de tamaño mediano y pequeño en toda la colección. De acuerdo con las
experiencias de los campesinos Ruandeses (Comunicación Personal L. Butare
investigador programa fríjol Ruanda, CIAT 2007), la tendencia de los campesinos,
que comercializan la semilla y que dependen de ella para su alimentación, es a
sembrar muchas áreas con fríjol y la semilla de menor tamaño confiere la ventaja
de poder sembrar mas semillas en una menor área, razón por la cual se pudo ver
reflejada esta incidencia. Además se produce mayor cantidad de semillas pequeñas
o medianas en una misma planta que las que se pueden producir de tamaño grande
(Martin & Adams, 1987).
La presencia de gran cantidad de tipos de fríjol (variedad de colores y tamaños)
sugiere que se mantiene la existencia de redes de distribución y que estos fríjoles
han tenido la aceptación de los consumidores (o una amplia adaptación ambiental)
(Martin & Adams, 1987). La acción de las preferencias de consumo o factores
84

ambientales, o las dos en conjunto, ha sido fundamental para la existencia de la
variabilidad existente en esta colección de germoplasma de fríjol de Ruanda.
Los colores que más se comercializan en Ruanda son rojo (moteado) y café por lo
tanto hacen parte de la gran producción de este país, esto podría explicar el porque
estos dos tipos de semilla fueron predominantes en la colección (Comunicación
Personal L. Butare investigador programa fríjol Ruanda, CIAT 2007); los fríjoles de
colores blanco y amarillo se comercializan principalmente en las ciudades y son
destinados a los restaurantes, por lo tanto son los que mayor valor comercial
presentan, sin embargo la baja incidencia de los de color blanco para este estudio,
puede verse explicada por el hecho de que muchos campesinos no siembran estos
genotipos por su difícil cocción o por tradiciones culturales. Los fríjoles de color
negro nos son muy apreciados en el mercado sin embargo éste lo producen para
consumo en mezcla (Butare op cit.).
En cuanto a los individuos que se les ha denominado como raros, debido a que no
se agruparon dentro de ningún acervo, se puede decir que son excéntricos dentro
de la población estudiada, ya que aparentemente no comparten muchas
características moleculares con los dos acervos encontrados e inclusive tampoco
se encuentran relacionados con los individuos candidatos a introgresión. Por lo que
se sugiere que se realice un seguimiento de estos, ya que su aislamiento molecular
se pudo haber originado por problemas de contaminación, errores en el
procedimiento llevado a cabo en el laboratorio, entre muchas otras causas.
La variabilidad encontrada en la colección de germoplasma de Ruanda puede ser el
resultado del conjunto de diferentes fuerzas, dentro de las que se incluye las
prácticas agronómicas, la selección humana, la selección natural, el intercambio
local y nacional de semilla y los eventos naturales de polinización cruzada.
7.1.3 Agrupación mediante índice de disimilaridad
Se observa que en este caso el grupo es dividido en los dos acervos principales y
dos posibles grupos de introgresión, lo cual coincide con el análisis de PCoA. La
agrupación de individuos en cada metodología no es muy diferente la una de la otra
ya que agrupan un número similar de individuos dentro de cada grupo; sin embargo
85

debido a que utilizan índices diferentes y sistema de graficas es muy diferente no es
posible hacer una comparación directa de estas dos metodologías, ya que esto
implicaría realizar una compleja comparación de software que no compete en este
estudio. Sin embargo se quiso mostrar esta otra forma de agrupación para
evidenciar a través de otro método las agrupaciones por acervos dadas para este
estudio.
7.1.4 Posibles eventos de introgresión en la población estudiada
De acuerdo con la caracterización molecular realizada y al análisis de ordenación
implementado, fue posible encontrar un primer grupo de 17 individuos que
presentan características moleculares que los hacen candidatos de ser
introgresados, estos fluctúan en sentido Mesoamericano-Andino. Dentro de este
grupo 5 individuos presentan características morfológicas típicas del acervo
Mesoamericano y en el dendrograma mostrado en la figura 13C se agrupan dentro
del acervo Andino. Adicionalmente se encontró un segundo grupo que presenta
características moleculares propias de introgresión; estos dos grupos son los que
presentan valores más altos de diversidad, lo cual podría ser un indicativo de que la
introgresión entre acervos tiende a aumentar la diversidad (Beebe et al., 2001)
La formación de estos grupos no es sorprendente en una región como Ruanda a la
que, al igual que el resto de África, llegaron los dos acervos principales de fríjol y a
partir de ese entonces las mezclas de estos se han realizado continuamente (Martin
& Adams, 1987). Esta formación de grupos, puede deberse a una posible
hibridación entre acervos, un evento que ha sido postulado en estudios como el de
Beebe et al (1997) quienes encontraron en mezclas de cultivares sembradas en
Colombia y en Perú, la posible ocurrencia de un proceso de alogamia. En la
producción de semillas en la que es indiferente la elección de algún tipo de grano
específico, es frecuente encontrar que los campesinos finalmente escogen todas las
semillas producidas, sin importar que estas provengan de una recombinación
genética (Beebe et al., 2001); por lo tanto en esta situación los tipos introgresados
son más probables de mantenerse en las mezclas de los campesinos que lo que
ocurriría en los cultivadores orientados hacia el mercado. Esto podría explicar la
presencia de los individuos posiblemente introgresados dentro de la colección
86

ruandesa; como se sabe la selección por color es laxa y las mezclas son aceptadas
por campesinos.
Ruanda es considerado como un centro secundario de diversidad de fríjol, por lo
que la diversidad encontrada refleja en la influencia de los dos acervos introducidos
a este país. Por lo tanto encontrar genotipos que presentan características
genéticas de los dos acervos puede representar el flujo genético entre estos o la
introgresión desde el acervo Mesoamericano al andino o viceversa; sin embargo se
sugiere confrontar estos datos moleculares con mayores evidencias morfológicas y
bioquímicas que permitan llegar a una conclusión más contundente.
7.2 ANÁLISIS DE MINERALES
Las diferencias encontradas entre este estudio y otros de minerales (Beebe et al.,
2000a; Blair et al., 2005; Islam et al., 2002a) pueden ser de origen genético y/o
ambiental; se encontraron valores altos de hierro y zinc dentro de la colección, lo
cual es de gran importancia debido a que estos minerales son fundamentales para
la nutrición de la población Ruandesa.
Es pertinente resaltar que los genotipos que presentaron los contenidos más altos
de hierro se encuentran por encima de 85ppm siendo este un valor alto teniendo en
cuenta los estudios mencionados anteriormente. Estos resultados son muy
importantes ya que el hierro es un componente esencial de la semilla de fríjol pues
interviene en importantes procesos metabólicos en el cuerpo humano siendo
esencial en la prevención de enfermedades graves como lo es la anemia
ferropénica; por ende encontrar genotipos con alto contenido de este mineral en
países en vía de desarrollo como lo es Ruanda facilita los planes de mejoramiento
de cultivos en los que se buscan incrementar los valores nutricionales y así generar
variedades de fríjol con mayor contenido de nutrientes.
Todos los valores hallados para zinc fueron considerados entre medianos y altos, es
decir fueron mayores a 25 ppm, lo que resulta ser muy interesante ya que encontrar
valores altos de zinc pueden garantizar que en futuros estudios de mejoramiento del
fríjol común en Ruanda se utilicen estos genotipos y de esta forma incrementar el
87

consumo en esta población que presenta graves problemas de malnutrición y salud
pública especialmente el VIH/SIDA. Es bien sabido que el zinc ayuda a incrementar
la defensa inmunológica del organismo por lo que es importante en el tratamiento de
enfermedades tales como VIH/SIDA.
Otro punto importante a tratar es el caso de los individuos RW582 y RW500 que
presentaron los valores más altos para hierro y para zinc; es importante resaltar que
estos genotipos no son morfológicamente atractivos para producción y/o consumo,
debido a la coloración negra que presentan, por lo tanto podrían ser importantes en
un plan de mejoramiento ya que podrían servir de progenitores donantes de
nutrientes.
Al comparar el dato de la desviación estándar encontrada con el estudio de Beebe
et al. 2005, se puede ver que la variación es similar (8.3 Beebe y 8.4 tabla 14). La
desviación estándar y el coeficiente de variación pueden ser un reflejo de la
variabilidad genética que existe en el material estudiado. El valor de la desviación
era de esperarse debido a que los genotipos provienen de diferentes regiones de
Ruanda y existe alto grado de combinación de material.
Se encontró un valor más alto de curtosis en zinc que en hierro lo que podría sugerir
que existe mayor varianza para este mineral, esto podría deberse a que existen
desviaciones infrecuentes en los extremos, que se oponen a desviaciones comunes
de medidas menos pronunciadas.
Al comparar este estudio con el de Beebe et al. 2002 se encontró el mismo valor
para el coeficiente de correlación de los dos minerales (0.52). Es posible proponer
que esta correlación sea un reflejo de la cosegregación de genes para estos dos
rasgos. La implicación de estas correlaciones es que algunos factores genéticos
para los minerales están cosegregando y que la selección de uno de estos
elementos puede resultar simultáneamente en un incremento en el otro, de esta
forma se multiplica el impacto y se facilita el mejoramiento.
A pesar que la diferencia entre grupos para la variable de hierro no fue significativa
es posible decir que, de acuerdo con la asociación de datos de contenido de
minerales obtenidos y las agrupaciones formadas a partir del análisis de diversidad
88

genética se observó que el grupo de los genotipos introgresados presentó el
contenido medio de hierro más alto de toda la colección. Esto podría deberse a la
influencia que ha comenzado a tener la introgresión en la evolución dentro de los
acervos del fríjol, algunos rasgos importantes como alto contenido de minerales o
resistencia a una enfermedad puede deberse a eventos de introgresión (Islam et al.,
2004).
Encontrar un gradiente de zinc en los grupos mesoamericanos podría representar
un factor importante en el conocimiento de procesos de selección para este estudio.
Se podría hablar de un proceso de domesticación o que la posible causa de la
aparición de este gradiente sea debida a procesos de selección humana, sin
embargo no se tienen argumentos claros para explicar la aparición de este
gradiente.
La baja diferencia existente entre los valores de zinc del grupo de Mesoamericanos
IV y el grupo de los andinos puede explicarse en la posible influencia de factores
ambientales o genéticos que han ayudado a moldear los genotipos permitiendo así
encontrar valores muy similares entre estos dos grupos. También se puede argüir
que posiblemente exista introgresión por parte de alguno de estos grupos pero que
esta no sea detectada a nivel molecular.
89

8 CONCLUSIONES
En la caracterización de la diversidad genética para 355 genotipos de fríjol común
de Ruanda (África), mediante el uso de marcadores moleculares analizados con
fluorescencia se observó que esta colección presentó un alto polimorfismo y altos
índices de variabilidad genética. El valor para diversidad total encontrado en este
estudio se consideró como alto en comparación con estudios en otras regiones del
mundo, lo que indica la importancia de Ruanda como centro secundario de
diversidad del fríjol común.
Los 30 marcadores utilizados presentaron un alto nivel de información y un
excelente poder de discriminación, lo que permitió dividir a la población de estudio
en los dos acervos genéticos principales para fríjol común.
La presencia de posibles eventos de introgresión permitió ver efectos relacionados
con flujo de genes o recombinación genética, siendo este un factor importante en la
conservación e incremento de la variabilidad genética.
Las características morfológicas de la semilla como el color y tamaño se pudieron
relacionar con los grupos derivados a partir de los análisis de diversidad y se
observó que cada grupo se encuentra específicamente caracterizado por alguno de
los niveles de estas dos variables. Adicional a esto se observa la presión que ha
ejercido selección humana en la caracterización de cada grupo.
El alto contenido de minerales encontrado en este estudio evidencia la importancia
que la presión de selección humana está ejerciendo sobre la colección de
germoplasma de fríjol en este país.
La correlación encontrada en el contenido de los dos minerales permite esperar que
en futuros planes de mejoramiento los dos minerales tengan un incremento
simultáneo.
90

El acervo predominante en cuanto a número de individuos en este estudio fue el
mesoamericano, siendo este resultado contrario a lo que hasta el momento se ha
observado en estudios de diversidad genética (mayor número de individuos para
andinos).
Los marcadores moleculares tipo microsatélites analizados mediante fluorescencia,
por su alta sensibilidad, permitieron obtener el mayor número de alelos descritos
para algún marcador microsatélite de P. vulgaris.
91

9 RECOMENDACIONES Y PERSPECTIVAS
Se recomienda hacer futuros estudios de diversidad utilizando genotipos controles
pertenecientes a las diferentes razas en que se dividen los acervos ya que de esta
forma sería posible establecer una estructura poblacional más completa.
El principal factor tenido en cuenta para este estudio de diversidad fue el molecular
por lo que se sugiere ampliar el estudio con datos morfoagronómicos que permitan
definir de manera contundente las relaciones moleculares acá encontradas.
Realizar una comparación de las técnicas utilizadas para la evaluación de
polimorfismos en microsatélites, la tradicional tinción en plata y la fluorescente a
través del ABI 3730xl.
Este estudio se constituye como el primero en la caracterización de la colección de
germoplasma de fríjol de Ruanda, por lo que es imperante desarrollar
investigaciones más profundas que contemplen otro tipo de marcadores y la
experiencia directa de los campesinos para poder llegar a conclusiones que vayan
más allá de una evaluación molecular.
El haber encontrado valores altos de diversidad genética y de características
nutricionales, abre la posibilidad para la realización de futuros estudios enfocados
hacía el mejoramiento.
Existen dos individuos importantes para un plan de mejoramiento al servir de
progenitores donantes de nutrientes, estos podrían ir junto con otro genotipo que
presente características morfológicas atractivas para el consumo en este país, por
ejemplo un fríjol rojo moteado.
92

10. REFERENCIAS
Applied Biosystems. 2004. GeneMapper® Software v.3.7 User Guide. Applied Biosystems, USA.
Ayad, W., T. Hodkin, A. Jaradat, and V. Rao. 1997. Molecular genetic techniques for plant genetic resources, In I. P. G. R. Institute, (ed.) Report of an IPGRI Workshop, 9-11 Octubre, Rome, Italy.
Barrios, M.F., H.G. du Défaix, and N. Fernandez. 2000. Metabolismo del Hierro. Rev Cubana Hematol Inmunol Hemoter. 16:149-160.
Bazel, P., and J. Anderson. 1994. Nutrition And Health Implication of Dry Beans: A Review. Journal of American College of Nutrition. 13:549-558.
Becerra, V., and P. Gepts. 1994. RFLP Diversity of common bean (Phaseolus vulgaris L.) in its centres of origin. Genome. 37:256-263.
Beebe, S., A.V. Gonzalez, and J. Rengifo. 2000a. Research on trace minerals in the common bean. Food and Nutrition Bull. 21:387-391.
Beebe, S., O. Toro, A.V. Gonzalez, M.I. Chacón, and D.G. Debouck. 1997. Wild-weed.crop complexes of common bean (Phaseolus vulgaris L., Fabaceae) in the Andes of Peru and Colombia, and their implications for conservation and breeding. Genetic Resources and Crop Evolution. 44:73-91.
Beebe, S., J. Rengifo, E. Gaitán-Solís, M.C. Duque, and J. Tohme. 2001. Diversity and origin of Andean landraces of common bean. Crop Sci. 41:854-862.
Beebe, S., P.W. Skroch, J. Tohme, M.C. Duque, F. Pedraza, and J. Nienhuis. 2000b. Structure of Genetic Diversity among Common Bean Landraces of Middle American Origin Based on Correspondence Analysis of RAPD. Crop Sci. 40:264-273.
Betancour, M.J., and J.E. Dávila. 2002. El fríjol (Phaseolus vulgaris L.): Cultivo, beneficio y variedades. FENALCO., Medellín. Colombia.
Bio-Rad. 1998. Quantity One: User Guide for version 4.0.3 for Windows and Macintosh. Versión 1999. Bio-Rad, USA.
Blair, M., J. Díaz, R. Hidalgo, L. Díaz, and M. Duque. 2007. Microsatellite characterization of Andean races of common bean (Phaseolus vulgaris L.). TAG Theoretical and Applied Genetics. in press.
Blair, M.W. 2007. Organización de marcadores fluorescentes en paneles.
93

Blair, M.W., V. Hedetale, and S.R. McCouch. 2002. Fluorescent-labeled microsatellite panels useful for detecting allelic diversity in cultivated rice (Oryza sativa L.). Theor Appl Genet. 105:449–457.
Blair, M.W., C. Astudillo, and S. Beebe. 2005. Analysis of Nutritional Quality traits in an Andean Recombinant inbred Line Population, En B. I. C, ed. Annual Report of the Bean Improvement Cooperative, Michigan.
Blair, M.W., M.C. Giraldo, H.F. Buendía, E. Tovar, M.C. Duque, and S.E. Beebe. 2006. Microsatellite marker diversity in common bean (Phaseolus vulgaris L.). Theor Appl Genet. 113:100–109.
Blair, M.W., F. Pedraza, H.F. Buendia, E. Gaitan-Solis, S.E. Beebe, P. Gepts, and J. Tohme. 2003. Development of a genome-wide anchored microsatellite map for common bean ( Phaseolus vulgaris L.). Theor Appl Genet. 107:1362–1374.
Broughton, W.J., G. Hernández, M. Blair, S. Beebe, P. Gepts, and J. Vanderleyden. 2003. Beans (Phaseolus spp.) – model food legumes. Plant and Soil. 252:55-128.
CIAT. 2005a. Utlisation of bean genetic diversity in Africa Highlights CIAT in Africa, Vol. 21. CIAT, Univesity of Nairobi, PABRA, SABRN, ESCABREN.
CIAT. 2005b. Fast tracking of nutritionally-rich bean varieties Highlights CIAT in Africa, Vol. 24. CIAT, Univesity of Nairobi, PABRA, SABRN, ESCABREN.
Collard, B., M.Z. Jahufer, J.B. Brouwe, and E.C. Pang. 2005. An introduction to markers, quantitative trait loci (QTL) mapping and marker-assisted selection for crop improvement: The basic concepts. Euphytica. 142:169 - 196.
Chacón, M.I., S. Pickersgill, and D.J. Debouck. 2005. Domestication patterns in common bean (Phaseolus vulgaris L.) and the origin of Mesoamerican and Andean cultivated races. Theor Appl Genet. 110:432-444.
Chrispeels, M.J., and D.E. Savada. 2003. Plants Genes and Crop Biotechnology. Jones and Bartlett Publishers, Inc, USA.
David, S., and L. Sperling. 1999. Improving technology delivery mechanisms: Lessons from bean seed systems research in eastern and central Africa. Agriculture and Human Values. 16:381-388.
Dean, R.E., J.A. Dahlberg, M.S. Hopkins, S.E. Mitchell, and S. Kresovich. 1999. Genetic redundancy and diversity among 'Orange' accessions in the U.S. national sorghum collection as assessed with Simple Sequence Repeat (SSR) markers. Crop Sci. 39:1215-1221.
Debouck, D., and R. Hidalgo. 1989. Morfología de la planta de fríjol común, p. 7-41, En M. Lopez, et al., eds. Investigación y producción, Cali, Colombia.
94

Debouck, D.J., and J. Tohme. 1988. Implications for bean breeders of studies on the origins of common beans, Phaseolus vulgalis L., p. 2-42, En S. Beebe, ed. Current Topics in Breeding of Common Bean. Proceedings of the International Bean Breeding Workshop, Cali, Colombia
Diaz, J.M. 2005a. Diversidad Genética y Estructura Poblacional de Germoplasma Andino De Fríjol Común Phaseolus vulgaris L. Tesis, Universidad Nacional De Colombia Sede Palmira, Colombia.
Diaz, L. 2005b. Diversidad Genética de Accesiones Representativas del Acervo Mesoamericano de Phaseolus Vulgaris L. Tesis, Universidad Nacional De Colombia Sede Palmira, Colombia.
Díaz, L.M., and M.W. Blair. 2006. Race structure within the Mesoamerican gene pool of common bean (Phaseolus vulgaris L.) as determined by microsatellite markers. TAG Theoretical and Applied Genetics. 114:143-154.
Donovan, C., and L. Bailey. 2005. Understanding Rwandan agricultural households’ strategies to deal with prime age illness and death: A propensity score matching approach IFPRI/Renewal Conference on HIV/AIDS and Food and Nutrition Security, Durban South Africa.
FAO (ed.) 1996. Conferencia Técnica Internacional sobre los recursos Fitogenéticos, Leipzing, Alemania. 17-23 junio.
Ferreira, M., and D. Grattapaglia. 1998. Introducción al uso de marcadores moleculares en el análisis genético. Brasilia, DF.
Ferris, R.S.B., J. Tuyisenge, M. Rucibango, D. Munkankubana, C. Ngagoyisonga, C. Gatarayiha, L. Butare, C. Kanyange, C. Uwantege, B. Kagiraneza, and K. Wanda. 2002. Atdt-Ciat/Isar/Iita-Foodnet And Pearl Project – Ruanda. International Institute Of Tropical Agricultura (IITA - Foodnet), Institute Des Sciences Agronomique Du Ruanda (ISAR), PEARL– Project., Rwanda, Africa.
Fonnegra, R. 1996. Taxonomía de plantas vasculares. 2a edición ed. Editorial Universidad de Antioquia, Medellín, Colombia.
Gaitan-Solis, E., M.C. Duque, K.J. Edwards, and J. Tohme. 2002. Microsatellite Repeats in Common Bean (Phaseolus vulgaris): Isolation, Characterization, and Cross-Species Amplification in Phaseolus ssp. Crop Sci. 42:2128-2136.
Gepts, P., and F.A. Bliss. 1986. Phaseolin variability among wild and cultivated common beans (Phaseolus vulgaris) from Colombia. Econ. Bot. 40:469-478.
Gepts, P., T.C. Osborn, K. Rashka, and F.A. Bliss. 1986. Phaseolin-protein variability in wild forms and landraces of the common bean (Phaseolus vulgaris L.) evidence for multiple centers of domestication. Econ. Bot. 40:451-468.
95

Gomez, O.J., M.W. Blair, B.E. Frankow-Lindberg, and U. Gullberg. 2004. Molecular and Phenotypic Diversity of Common Bean Landraces from Nicaragua. Crop Sci. 44:1412-1418.
Hajeer, A., J. Worthington, and S. John, (eds.) 2000. SNP and microsatellite genotyping: Markers for genetic analysis. Eaton Publishing, Manchester, U.K.
HarvestPlus. 2006. Biofotified Beans. Breeding Crops for Better Nutrition.
Harvey, D. 2000. Modern Analytical Chemistry. McGraw-Hill, New York.
ISAR. 2000. Rwanda Brief introduction to the Country [Online] http://www.isar.cgiar.org/Isarprog/Bean.htm (verified 15 Enero).
Islam, F.M.A., K.E. Basford , C. Jara, R.J. Redden, and S. Beebe. 2002a. Seed compositional and disease resistance differences among gene pools in cultivated common bean. Genetic Resources and Crop Evolution. 49:285-293.
Islam, F.M.A., K.E. Basford, R.J. Redden, A.V. Gonzalez, P.M. Kroonenberg, and S. Beebe. 2002b. Genetic variability in cultivated common bean beyond the two major gene pools. Genetic Resources and Crop Evolution. 49:271-283.
Islam, F.M.A., S. Beebe, M. Muñoz, J. Tohme, R.J. Redden, and K.E. Basford 2004. Using molecular markers to assess the effect of introgression on quantitative attributes of common bean in the Andean gene pool. Theor Appl Genet. 108:243-252.
Joe, L.K., A.M. Wheaton, S.J. Bay, L.E. Krotzer, P.A. Baybayan, O.S. Ostadan, S.R. Berosik, T.H. Yang, S.A. Pistacchi, S.-S. Feng, and S.-S. XLou. 2004. Improved Solutions for High Throughput Microsatellite Analysis. Applied Biosystems, Foster.
Kami, J., V. Becerra, D. Debouck, and P. Gepts. 1995. Identification of presumed ancestral DNA sequences of phaseolin in Phaseolus vulgaris. Proc. Natl. Acad. Sci. 92:1101-1104.
Kelly, V., E. Mpyisi, E. Shingiro, and J.B. Nyarwaya. 2001. Agricultural Intensification in Rwanda:An Elusive Goal Fertilizer Use and Conservation Investments, In Rwanda, (ed.), Vol. 3. FSRP/DSA Publication.
Khairallah, M.M., M.W. Adams, and B.B. Sears. 1990. Michondrial DNA polymorphisms of Malawian bean lines: further evidence for two major genepools. Theor Appl Genet. 80:753-761.
Kimani, P.M., R. Chirwa, and M.W. Blair. 2000. Bean breeding for east and central África: Strategy and Plan East and central Africa Research Network. Revised draft.
Koenig, R., and P. Gepts. 1989. Allozyme diversity in wild Phaseolus vulgaris: further evidence for two major centers of genetic diversity. Theor Appl Genet. 78:809-817.
96

Koinange, E.M.K., and P. Gepts. 1992. Hybrid weakness in wild Phaseolus vulgaris L. J Hered. 83:135–139.
Koinange, E.M.K., S.P. Singh, and P. Gepts. 1996. Genetic control of the domestication syndrome in common-bean. Crop Sci. 36:1037–1045.
Liu, K., and S.V. Muse. 2005. PowerMarker: Integrated analysis environment for genetic marker data. Bioinformatics. 21:2128-2129.
López, M., F. Fernández, and A.v. Schoonhoven, (eds.) 1985. Frijol: investigación y producción, pp. 1-417. Programa de las Naciones Unidas (PNUD); Centro Internacional de Agricultura Tropical (CIAT), Cali, CO.
Mahuku, G.S. 2004. A simple extraction method suitable for PCR based analysis of plant, fungal and bacterial DNA. Plant Mol Biol Rep. 22:71-81.
Martin, G.B., and M.W. Adams. 1987. Landraces of Phaseolus vulgaris (Fabaceae) in Northen Malawi I. Regional variation. Econ. Bot. 41:190-203.
MINAGRI/MINEDUC. 1993. Plan directeur Nationale de la recherche agricole 1993-2003. République Rwandaise, Rwuanda.
Mitchell, S.E., S. Kresovich, C.A. Jester, C.J. Hernandez, and M.A.K. Szewc. 1997. Application Of Multiplex-PCR And Fluorescence-Based, Semi-Automated Allele Sizing Technology For Genotyping Plant Genetic Resources. Crop Science 37:617-624.
Moreno, N. 2007. Caracterización molecular y nutricional de 221 variedades Africanas de fríjol común (Phaseolus vulgaris L.) postuladas como parentales en un programa de mejoramiento nutricional. Tesis, Biologa. Pontificia Universidad Javeriana, Bogotá D.C.
Muñiz, O. 1980. Espectrofotometria de absorción atómica. Su aplicación a la determinación de elementos metalicos en la Agroquímica. Boletín de Reseñas Suelos y Agroquímica:5-35.
Musoni, A. 2006. Inheritance of resistance to Fusarium wilt (F. oxysporum f.sp. phaseoli) and selection of marketable climbing bean varieties with multiple disease resistance. Thesis, Master of Science. University of Nairobi, Rwanda.
Ngirumwami, J.L. 1992. Analysis of food crop production and marketing trends in Rwnada with emphasis on dry beans. Thesis, Master of Science. Michigan State University.
OPS. 2002. Compuestos de hierro para la fortificación de alimentos: guías para América Latina y el Caribe. Washington, D.C.
97

Paredes-Lopez, O., and P. Gepts. 1995. Extensive introgression of middle American germplasm into Chilean common bean cultivars. Genetic Resources and Crop Evolution. 42:29-41.
Pastor-Corrales, M.A., and H.F. Schwartz, (eds.) 1994. Problemas de producción del frijol en los trópicos. . pp. 1-734 p. + Figuras a color (p. 735-805). Centro Internacional de Agricultura Tropical (CIAT), Cali, CO.
Reisch, B.I. 1998. Molecular markers: The foundation for grapevine genetic mapping, DNA fingerprinting and genomics Proc. 7th Intern. Symp. Grapevine Genetics and Breeding., Montpellier, France.
Rocha, E. 2000. Principios Básicos de Espectroscopía. UACh, México.
Rohlf, F. 2002. NTSys-pc v.2.10: Numerical Taxonomy System. Exter Publishing, NY.
Roizès, G. 2000. Identification of Microsatellite Markers: Screening for Repeat Sequences and Mapping Polymorphisms, p. 152, En A. Hejeer, et al., eds. SNP and microsatellite genotyping: markers for genetic analysis. Biotechniques Books, USA.
Sandoval, T. 2006. Cuantificación de Fitatos por espectroscopia visible en dos poblaciones de fríjol común (Phaseolus vulgaris L.) e identificación de QTL´s asociados a su contenido, Quimico. Universidad del Valle, Santiago de Cali.
Schoonhoven, A.A., and V.V. Oswaldo. 1994. El fríjol común en América Latina y sus limitaciones. Centro Internacional de Agricultura Tropical. Cali, Colombia.
Singh, S., P. Gepts, and D. Debouck. 1991a. Races of common bean (Phaseolus vulgaris Fabaceae). Econ. Bot. 45:379-396.
Singh, S.P. 2001. Broadening the Genetic Base of Common Bean Cultivars: A Review. Crop Sci. 41:1659-1675.
Singh, S.P., D.G. Debouck, and P. Gepts. 1988. Races of Common Bean, Phaseolus vulgalis, p. 75-89, En S. Beebe, ed. Current Topics in Breeding of Common Bean. Proceedings of International Bean Breeding Workshop, Cali, Colombia.
Singh, S.P., R.O. Nodari, and P. Gepts. 1991b. Genetic diversity in cultivated common bean: I. Allozymes. Crop Sci. 31:19-23.
Skoog, D.A., F.J. Holler, and T.A. Nieman. 2001. Principios de Análisis Instrumental. 5a ed. McGraw-Hill, Madrid.
Smith, J.S.C., S.L. Chin, H. Shu, O.S. Smith, J.S. Wall, and M.L. Senior. 1997. An Evaluation of the utility of SSR loci as molecular markers in Maize (Zea mays L.): comparisons with data from RFLPs and Pedigree. Theor Appl Genet. 95:163-173.
98

Sperling, L. 2001. The effect of the civil war on Rwanda’s bean seed systems and unusual bean diversity. Biodiversity and Conservation. 10:989–1009.
Tohme, J., D.O. Gonzalez, S. Beebe, and M.C. Duque. 1996. AFLP analysis of gene pools of a wild bean core collection. Crop Sci. 36:1375-1384.
UnitedNations. 2007. Maps and Geographic Information Resources [Online]. Available by Cartographic Section, DPKO http://www.un.org/Depts/Cartographic/map/profile/rwanda.pdf (verified 27 Agosto).
Weir, B.S. 1996. Genetic data analysis II. Sunderland, M.A: Sinauer Associates, Inc.
Winter, P., and G. Kahl. 1995. Molecular marker technologies for plant improvement. World Journal of Microbiology and Biotechnology. 11:438-448.
Wortmann, C.S., R.A. Kirkby, E. C.A, and D.J. Allen. 1999. Atlas of Common Bean [Phaseolus vulgalis L.] Produciton in Africa. CIAT Pan-African Bean Research Alliance.
Yeh, F., R. Yang, T. Boyle, Z. Ye, and J. Mao. 1997. POPGENE, the User-Friendly Shareware for Population Genetics Analysis. Molecular Biology and Biotechnology Centre, University of Alberta, Canada.
Yu, K., S. Park, V. Poysa, and P. Gepts. 2002. Integration of simple sequence repeat (SSR) markers into a molecular linkage map of common bean (Phaseolus vulgaris L.). J Hered. 91:429-434.
99

10 ANEXOS
Anexo 1. Protocolo de Extracción de ADN de fríjol utilizando proteinasa K (Mahuku 2004)
1. Agregar en un tubo de micro centrífuga de 2 µl: 800 μl de buffer de extracción (NaCl 5M, Tris 1M, pH 7.5, EDTA 0.5M pH 8, SDS 10%, proteinasa K 10mg/ml agua) y 0.03g de tejido. Se deja a baño maria 1 hora a 65ºC agitando cada 15 min.
2. Terminada la hora agitar el tubo y agregar 200 μl de acetato de Amonio 7.5 M, agitar y se dejar por 15 min a temperatura ambiente.
3. Centrifugar a 12000 r.p.m. por 15 min. 4. Transferir el sobrenadante a otro tubo y agregar 500 μl de cloroformo:octanol (24:1),
mezclar bien y centrifugar a 12000 r.p.m. por 10 min. 5. Transferir el sobrenadante y agregar ½ volumen de isopropanol frío y dejar
precipitando a 20ºC mínimo dos horas, pero preferiblemente toda la noche. 6. Posteriormente centrifugar a 12000 r.p.m. por 15 min, eliminar el sobrenadante lavar
el pellet con 500 μl de etanol frío al 70%. 7. Deja evaporar el etanol totalmente 8. Preparar el agua de resuspensión (6 μl de RNAasa/ 1 mL de TE 10:1 (Tris HCL 1M
pH 8.0, EDTA 0.5 M, pH 8.0)), y agregar entre 100 a 200 μL dependiendo del tamaño del pellet.
9. Dejar el ADN a 37ºC por 30 min y se observa su calidad en un gel de agarosa 1.0%.
100
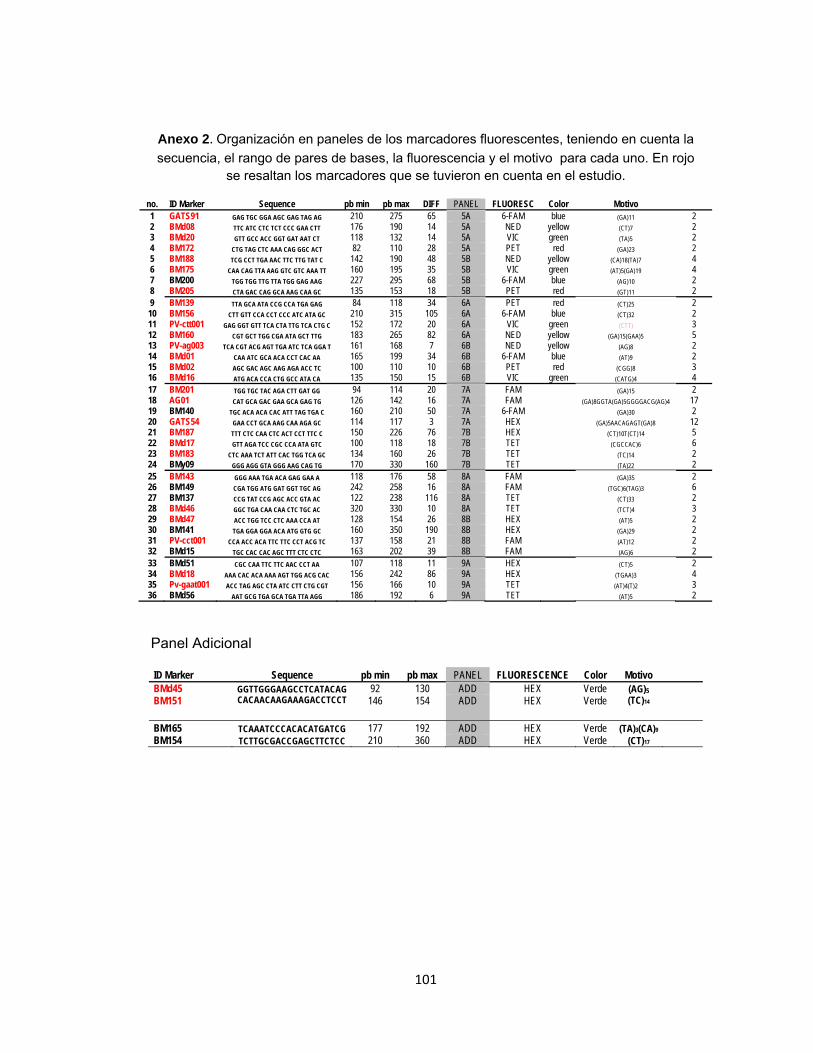
Anexo 2. Organización en paneles de los marcadores fluorescentes, teniendo en cuenta la secuencia, el rango de pares de bases, la fluorescencia y el motivo para cada uno. En rojo
se resaltan los marcadores que se tuvieron en cuenta en el estudio.
no. ID Marker Sequence pb min pb max DIFF PANEL FLUORESC Color Motivo 1 GATS91 GAG TGC GGA AGC GAG TAG AG 210 275 65 5A 6-FAM blue (GA)11 2 2 BMd08 TTC ATC CTC TCT CCC GAA CTT 176 190 14 5A NED yellow (CT)7 2 3 BMd20 GTT GCC ACC GGT GAT AAT CT 118 132 14 5A VIC green (TA)5 2 4 BM172 CTG TAG CTC AAA CAG GGC ACT 82 110 28 5A PET red (GA)23 2 5 BM188 TCG CCT TGA AAC TTC TTG TAT C 142 190 48 5B NED yellow (CA)18(TA)7 4 6 BM175 CAA CAG TTA AAG GTC GTC AAA TT 160 195 35 5B VIC green (AT)5(GA)19 4 7 BM200 TGG TGG TTG TTA TGG GAG AAG 227 295 68 5B 6-FAM blue (AG)10 2 8 BM205 CTA GAC CAG GCA AAG CAA GC 135 153 18 5B PET red (GT)11 2 9 BM139 TTA GCA ATA CCG CCA TGA GAG 84 118 34 6A PET red (CT)25 2 10 BM156 CTT GTT CCA CCT CCC ATC ATA GC 210 315 105 6A 6-FAM blue (CT)32 2 11 PV-ctt001 GAG GGT GTT TCA CTA TTG TCA CTG C 152 172 20 6A VIC green (CTT) 3 12 BM160 CGT GCT TGG CGA ATA GCT TTG 183 265 82 6A NED yellow (GA)15(GAA)5 5 13 PV-ag003 TCA CGT ACG AGT TGA ATC TCA GGA T 161 168 7 6B NED yellow (AG)8 2 14 BMd01 CAA ATC GCA ACA CCT CAC AA 165 199 34 6B 6-FAM blue (AT)9 2 15 BMd02 AGC GAC AGC AAG AGA ACC TC 100 110 10 6B PET red (CGG)8 3 16 BMd16 ATG ACA CCA CTG GCC ATA CA 135 150 15 6B VIC green (CATG)4 4 17 BM201 TGG TGC TAC AGA CTT GAT GG 94 114 20 7A FAM (GA)15 2 18 AG01 CAT GCA GAC GAA GCA GAG TG 126 142 16 7A FAM (GA)8GGTA(GA)5GGGGACG(AG)4 17 19 BM140 TGC ACA ACA CAC ATT TAG TGA C 160 210 50 7A 6-FAM (GA)30 2 20 GATS54 GAA CCT GCA AAG CAA AGA GC 114 117 3 7A HEX (GA)5AACAGAGT(GA)8 12 21 BM187 TTT CTC CAA CTC ACT CCT TTC C 150 226 76 7B HEX (CT)10T(CT)14 5 22 BMd17 GTT AGA TCC CGC CCA ATA GTC 100 118 18 7B TET (CGCCAC)6 6 23 BM183 CTC AAA TCT ATT CAC TGG TCA GC 134 160 26 7B TET (TC)14 2 24 BMy09 GGG AGG GTA GGG AAG CAG TG 170 330 160 7B TET (TA)22 2 25 BM143 GGG AAA TGA ACA GAG GAA A 118 176 58 8A FAM (GA)35 2 26 BM149 CGA TGG ATG GAT GGT TGC AG 242 258 16 8A FAM (TGC)6(TAG)3 6 27 BM137 CCG TAT CCG AGC ACC GTA AC 122 238 116 8A TET (CT)33 2 28 BMd46 GGC TGA CAA CAA CTC TGC AC 320 330 10 8A TET (TCT)4 3 29 BMd47 ACC TGG TCC CTC AAA CCA AT 128 154 26 8B HEX (AT)5 2 30 BM141 TGA GGA GGA ACA ATG GTG GC 160 350 190 8B HEX (GA)29 2 31 PV-cct001 CCA ACC ACA TTC TTC CCT ACG TC 137 158 21 8B FAM (AT)12 2 32 BMd15 TGC CAC CAC AGC TTT CTC CTC 163 202 39 8B FAM (AG)6 2 33 BMd51 CGC CAA TTC TTC AAC CCT AA 107 118 11 9A HEX (CT)5 2 34 BMd18 AAA CAC ACA AAA AGT TGG ACG CAC 156 242 86 9A HEX (TGAA)3 4 35 Pv-gaat001 ACC TAG AGC CTA ATC CTT CTG CGT 156 166 10 9A TET (AT)4(T)2 3 36 BMd56 AAT GCG TGA GCA TGA TTA AGG 186 192 6 9A TET (AT)5 2
Panel Adicional
ID Marker Sequence pb min pb max PANEL FLUORESCENCE Color Motivo BMd45 GGTTGGGAAGCCTCATACAG 92 130 ADD HEX Verde (AG)5 BM151 CACAACAAGAAAGACCTCCT 146 154 ADD HEX Verde (TC)14
BM165 TCAAATCCCACACATGATCG 177 192 ADD HEX Verde (TA)3(CA)9 BM154 TCTTGCGACCGAGCTTCTCC 210 360 ADD HEX Verde (CT)17
101

Anexo 3. Diagrama en el que se observa el ciclo que sigue la muestra dentro del secuenciador ABI 3730xl
102

Anexo 4. Descripción de características morfológicas de la semilla de fríjol
Cod Color Principal Color Secundario Patrón de Color Tamaño Brillo Cod Color Principal Color Secundario Patrón de Color Tamaño Brillo
1 Blanco ‐ ‐ Mediana Opaco 101 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
3 Blanco ‐ Venación Mediana Semi‐Brillante 102 Amarillo ‐ Venación Pequeña Semi‐Brillante
4 Café/Marrón Café/Marrón Rayado Mediana Semi‐Brillante 103 Amarillo ‐ Venación Pequeña Semi‐Brillante
5 Café/Marrón Rojo Rayado Mediana Opaco 104 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
6 Amarillo Rojo Jaspeado Mediana Semi‐Brillante 105 Café/Marrón ‐ Venación Pequeña Semi‐Brillante
7 Amarillo Rojo Jaspeado Mediana Semi‐Brillante 107 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
8 Crema Negro Rayado Grande Semi‐Brillante 109 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
9 Amarillo Negro Rayado Grande Semi‐Brillante 110 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
10 Café/Marrón Negro Jaspeado Pequeña Opaco 112 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
11 Café/Marrón Negro Jaspeado Mediana Semi‐Brillante 114 Café/Marrón ‐ ‐ Pequeña Opaco
12 Café/Marrón Negro Jaspeado Pequeña Opaco 115 Café/Marrón ‐ ‐ Pequeña Opaco
15 Café/Marrón Negro Jaspeado Pequeña Opaco 116 Café/Marrón ‐ ‐ Pequeña Opaco
16 Café/Marrón Negro Jaspeado Pequeña Opaco 117 Café/Marrón ‐ ‐ Pequeña Opaco
17 Café/Marrón Negro Jaspeado Pequeña Opaco 118 Café/Marrón ‐ ‐ Pequeña Opaco
19 Amarillo Café/Marrón Jaspeado Mediana Opaco 119 Café/Marrón ‐ ‐ Pequeña Opaco
20 Amarillo ‐ ‐ Pequeña Semi‐Brillante 121 Café/Marrón ‐ ‐ Pequeña Opaco
21 Amarillo ‐ ‐ Pequeña Semi‐Brillante 122 Amarillo ‐ ‐ Pequeña Semi‐Brillante
23 Amarillo ‐ ‐ Pequeña Semi‐Brillante 124 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
24 Amarillo ‐ ‐ Mediana Semi‐Brillante 126 Rosado ‐ ‐ Pequeña Semi‐Brillante
25 Amarillo ‐ ‐ Pequeña Semi‐Brillante 127 Café/Marrón ‐ ‐ Pequeña Opaco
26 Negro ‐ ‐ Pequeña Opaco 128 Café/Marrón ‐ ‐ Pequeña Opaco
27 Amarillo ‐ ‐ Mediana Opaco 129 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
28 Amarillo ‐ ‐ Mediana Semi‐Brillante 130 Café/Marrón ‐ ‐ Pequeña Opaco
30 Amarillo Café/Marrón Punteado Mediana Semi‐Brillante 131 Café/Marrón ‐ ‐ Pequeña Opaco
31 Amarillo Café/Marrón Punteado Mediana Semi‐Brillante 133 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
32 Amarillo Café/Marrón Punteado Mediana Semi‐Brillante 134 Negro ‐ ‐ Pequeña Opaco
33 Morado/Púrpura ‐ ‐ Mediana Semi‐Brillante 136 Negro ‐ ‐ Pequeña Opaco
34 Rojo Crema Moteado Grande Semi‐Brillante 137 Negro ‐ ‐ Pequeña Opaco
35 Amarillo Café/Marrón Punteado Mediana Semi‐Brillante 139 Negro ‐ ‐ Pequeña Opaco
36 Amarillo Café/Marrón Punteado Mediana Semi‐Brillante 140 Negro ‐ ‐ Mediana Opaco
37 Amarillo ‐ ‐ Pequeña Semi‐Brillante 141 Negro ‐ ‐ Mediana Opaco
38 Amarillo ‐ ‐ Pequeña Semi‐Brillante 142 Negro ‐ ‐ Mediana Semi‐Brillante
39 Amarillo ‐ ‐ Pequeña Semi‐Brillante 143 Negro ‐ ‐ Mediana Opaco
40 Amarillo Café/Marrón Punteado Mediana Semi‐Brillante 144 Negro ‐ ‐ Mediana Opaco
41 Amarillo Café/Marrón Punteado Mediana Semi‐Brillante 145 Negro ‐ ‐ Pequeña Opaco
43 Amarillo Café/Marrón Punteado Mediana Semi‐Brillante 146 Negro ‐ ‐ Pequeña Semi‐Brillante
44 Amarillo Rojo Rayado Mediana Semi‐Brillante 147 Negro ‐ ‐ Pequeña Opaco
45 Amarillo Negro Rayado Mediana Semi‐Brillante 149 Negro ‐ ‐ Mediana Semi‐Brillante
46 Crema Negro Rayado Mediana Semi‐Brillante 150 Negro ‐ ‐ Pequeña Semi‐Brillante
47 Crema Negro Rayado Mediana Semi‐Brillante 151 Negro ‐ ‐ Pequeña Semi‐Brillante
48 Crema Negro Rayado Mediana Semi‐Brillante 152 Negro ‐ ‐ Pequeña Semi‐Brillante
49 Crema Negro Rayado Mediana Semi‐Brillante 153 Negro ‐ ‐ Pequeña Opaco
50 Crema Negro Rayado Mediana Semi‐Brillante 154 Negro ‐ ‐ Mediana Semi‐Brillante
54 Crema Negro Rayado Mediana Semi‐Brillante 155 Negro ‐ ‐ Mediana Semi‐Brillante
55 Crema ‐ ‐ Mediana Semi‐Brillante 156 Negro ‐ ‐ Pequeña Opaco
56 Crema ‐ ‐ Mediana Semi‐Brillante 157 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
58 Rojo ‐ ‐ Pequeña Semi‐Brillante 158 Café/Marrón Café/Marrón Jaspeado Pequeña Semi‐Brillante
59 Rojo ‐ ‐ Pequeña Semi‐Brillante 159 Gris/Raros Café/Marrón Moteado Pequeña Opaco
61 Rosado ‐ ‐ Pequeña Semi‐Brillante 162 Café/Marrón ‐ ‐ Mediana Semi‐Brillante
62 Rojo ‐ ‐ Pequeña Semi‐Brillante 163 Café/Marrón Café/Marrón Punteado Mediana Semi‐Brillante
64 Rojo ‐ ‐ Pequeña Semi‐Brillante 164 Crema Negro Rayado Mediana Semi‐Brillante
66 Rojo ‐ ‐ Pequeña Semi‐Brillante 165 Café/Marrón Negro Moteado Mediana Opaco
69 Rojo ‐ ‐ Pequeña Semi‐Brillante 166 Amarillo ‐ ‐ Mediana Opaco
70 Rojo ‐ ‐ Pequeña Semi‐Brillante 170 Blanco Negro Jaspeado Pequeña Opaco
71 Rojo ‐ ‐ Pequeña Semi‐Brillante 171 Crema Negro Jaspeado Pequeña Semi‐Brillante
72 Rojo ‐ ‐ Pequeña Semi‐Brillante 172 Café/Marrón Negro Jaspeado Pequeña Semi‐Brillante
73 Rojo ‐ ‐ Pequeña Semi‐Brillante 174 Café/Marrón Negro Jaspeado Mediana Semi‐Brillante
74 Rojo ‐ ‐ Pequeña Semi‐Brillante 175 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
76 Rojo ‐ ‐ Pequeña Semi‐Brillante 176 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
77 Rosado ‐ ‐ Pequeña Semi‐Brillante 177 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
78 Negro Crema Moteado Grande Semi‐Brillante 180 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
80 Amarillo ‐ Venación Mediana Opaco 181 Café/Marrón Café/Marrón Moteado Pequeña Semi‐Brillante
81 Amarillo ‐ ‐ Pequeña Semi‐Brillante 182 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
82 Amarillo ‐ ‐ Pequeña Semi‐Brillante 184 Café/Marrón Negro Moteado Pequeña Semi‐Brillante
87 Amarillo ‐ ‐ Pequeña Semi‐Brillante 185 Crema ‐ Venación Pequeña Semi‐Brillante
88 Amarillo ‐ Venación Pequeña Semi‐Brillante 186 Crema ‐ Venación Pequeña Semi‐Brillante
89 Rosado Café/Marrón Rayado Mediana Semi‐Brillante 187 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
90 Amarillo ‐ Venación Mediana Semi‐Brillante 188 Amarillo ‐ ‐ Pequeña Semi‐Brillante
92 Café/Marrón Negro Jaspeado Pequeña Opaco 189 Rojo ‐ ‐ Mediana Semi‐Brillante
93 Crema Negro Jaspeado Mediana Opaco 191 Rojo ‐ ‐ Mediana Semi‐Brillante
94 Amarillo ‐ Venación Pequeña Opaco 192 Amarillo ‐ ‐ Pequeña Semi‐Brillante
95 Amarillo ‐ Venación Mediana Semi‐Brillante 193 Rojo ‐ ‐ Mediana Semi‐Brillante
96 Amarillo ‐ Venación Mediana Semi‐Brillante 194 Rojo ‐ ‐ Mediana Semi‐Brillante
103

Anexo 4. (cont.) Descripción de características morfológicas de la semilla de fríjol
Cod Color Principal Color Secundario Patrón de Color Tamaño Brillo Cod Color Principal Color Secundario Patrón de Color Tamaño Brillo
196 Negro ‐ ‐ Pequeña Semi‐Brillante 301 Amarillo ‐ ‐ Mediana Semi‐Brillante
199 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante 302 Amarillo ‐ ‐ Mediana Semi‐Brillante
201 Crema Rojo Rayado Grande Semi‐Brillante 303 Amarillo ‐ ‐ Mediana Semi‐Brillante
204 Crema Rojo Rayado Mediana Semi‐Brillante 310 Amarillo ‐ ‐ Mediana Semi‐Brillante
205 Crema Rojo Rayado Mediana Semi‐Brillante 311 Café/Marrón Negro Rayado Grande Semi‐Brillante
206 Crema Rojo Rayado Mediana Semi‐Brillante 312 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
207 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante 316 Amarillo ‐ ‐ Mediana Semi‐Brillante
209 Café/Marrón ‐ ‐ Mediana Semi‐Brillante 319 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
211 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante 320 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
212 Café/Marrón ‐ ‐ Mediana Semi‐Brillante 321 Rojo Crema Jaspeado Mediana Semi‐Brillante
214 Café/Marrón ‐ ‐ Mediana Semi‐Brillante 325 Negro Crema Moteado Grande Semi‐Brillante
215 Rojo Crema Moteado Mediana Semi‐Brillante 326 Amarillo ‐ ‐ Mediana Semi‐Brillante
216 Rojo Crema Moteado Mediana Semi‐Brillante 328 Crema Café/Marrón Jaspeado Pequeña Semi‐Brillante
217 Rojo Crema Moteado Mediana Semi‐Brillante 329 Crema Rojo Rayado Grande Semi‐Brillante
218 Rojo Crema Moteado Mediana Semi‐Brillante 332 Crema Rojo Rayado Mediana Semi‐Brillante
219 Rojo Crema Moteado Mediana Semi‐Brillante 333 Crema Rojo Rayado Mediana Semi‐Brillante
220 Rojo Crema Moteado Mediana Semi‐Brillante 335 Crema Rojo Rayado Mediana Semi‐Brillante
221 Rojo Crema Moteado Mediana Semi‐Brillante 336 Crema Rojo Rayado Mediana Semi‐Brillante
223 Rojo Crema Moteado Mediana Semi‐Brillante 338 Crema Negro Rayado Mediana Semi‐Brillante
224 Café/Marrón Negro Jaspeado Mediana Semi‐Brillante 340 Crema Negro Rayado Mediana Semi‐Brillante
226 Rojo ‐ ‐ Pequeña Semi‐Brillante 341 Crema Negro Rayado Mediana Semi‐Brillante
227 Rojo ‐ ‐ Mediana Semi‐Brillante 345 Crema Negro Rayado Mediana Semi‐Brillante
229 Amarillo Café/Marrón Punteado Pequeña Semi‐Brillante 357 Amarillo ‐ ‐ Pequeña Semi‐Brillante
230 Amarillo Café/Marrón Moteado Mediana Semi‐Brillante 359 Amarillo ‐ ‐ Mediana Semi‐Brillante
231 Rojo Crema Moteado Mediana Semi‐Brillante 368 Rojo Crema Moteado Mediana Semi‐Brillante
232 Café/Marrón Rojo Punteado Mediana Semi‐Brillante 370 Rojo Crema Moteado Mediana Semi‐Brillante
234 Crema ‐ ‐ Pequeña Semi‐Brillante 371 Rojo Crema Moteado Mediana Semi‐Brillante
235 Café/Marrón Café/Marrón Venación Pequeña Opaco 373 Rojo Crema Moteado Mediana Semi‐Brillante
236 Rojo ‐ ‐ Pequeña Opaco 374 Rojo Crema Moteado Mediana Semi‐Brillante
238 Rojo ‐ ‐ Mediana Opaco 375 Rojo Crema Moteado Mediana Semi‐Brillante
240 Rojo ‐ ‐ Mediana Semi‐Brillante 376 Rojo Crema Moteado Mediana Semi‐Brillante
242 Negro ‐ ‐ Pequeña Opaco 377 Rojo Crema Moteado Mediana Semi‐Brillante
243 Negro ‐ ‐ Mediana Opaco 379 Rojo Crema Moteado Mediana Semi‐Brillante
244 Rojo ‐ ‐ Pequeña Semi‐Brillante 383 Rojo Crema Moteado Mediana Semi‐Brillante
245 Rojo Café/Marrón Jaspeado Mediana Semi‐Brillante 384 Rojo Crema Moteado Mediana Semi‐Brillante
246 Rojo ‐ ‐ Mediana Semi‐Brillante 385 Rojo ‐ ‐ Mediana Semi‐Brillante
247 Café/Marrón Rojo Jaspeado Mediana Semi‐Brillante 386 Rojo ‐ ‐ Mediana Semi‐Brillante
249 Rojo Crema Moteado Mediana Semi‐Brillante 389 Rojo ‐ ‐ Grande Semi‐Brillante
253 Rojo ‐ ‐ Mediana Semi‐Brillante 390 Rojo ‐ ‐ Mediana Semi‐Brillante
255 Gris/Raros Café/Marrón Moteado Pequeña Opaco 393 Rojo ‐ ‐ Mediana Semi‐Brillante
257 Amarillo ‐ ‐ Pequeña Semi‐Brillante 394 Rojo ‐ ‐ Grande Semi‐Brillante
258 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante 397 Rojo ‐ ‐ Grande Brillante
259 Café/Marrón ‐ ‐ Mediana Semi‐Brillante 399 Rojo ‐ ‐ Grande Brillante
260 Café/Marrón ‐ ‐ Mediana Semi‐Brillante 400 Negro ‐ ‐ Pequeña Opaco
266 Amarillo Café/Marrón Moteado Mediana Semi‐Brillante 404 Rojo Crema Moteado Mediana Semi‐Brillante
267 Crema ‐ Moteado Mediana Semi‐Brillante 408 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
268 Rojo ‐ Punteado Pequeña Semi‐Brillante 410 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
269 Rojo ‐ Punteado Pequeña Semi‐Brillante 415 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
270 Amarillo ‐ Moteado Mediana Semi‐Brillante 421 Rojo ‐ ‐ Pequeña Semi‐Brillante
271 Café/Marrón ‐ Punteado Pequeña Semi‐Brillante 426 Crema Negro Moteado Pequeña Semi‐Brillante
272 Crema ‐ Punteado Pequeña Semi‐Brillante 428 Rojo ‐ ‐ Pequeña Brillante
273 Gris/Raros Café/Marrón Punteado Pequeña Semi‐Brillante 429 Rojo ‐ ‐ Pequeña Brillante
274 Gris/Raros Café/Marrón Moteado Mediana Semi‐Brillante 434 Negro ‐ ‐ Pequeña Opaco
275 Café/Marrón Crema Moteado Mediana Semi‐Brillante 436 Negro ‐ ‐ Pequeña Opaco
277 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante 438 Negro ‐ ‐ Pequeña Opaco
278 Amarillo ‐ ‐ Pequeña Semi‐Brillante 440 Negro ‐ ‐ Pequeña Opaco
279 Amarillo ‐ ‐ Pequeña Semi‐Brillante 441 Negro ‐ ‐ Pequeña Opaco
281 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante 444 Negro ‐ ‐ Pequeña Semi‐Brillante
282 Amarillo ‐ ‐ Mediana Semi‐Brillante 445 Negro ‐ ‐ Pequeña Opaco
283 Gris/Raros Crema Moteado Mediana Semi‐Brillante 454 Amarillo ‐ ‐ Pequeña Semi‐Brillante
284 Blanco ‐ ‐ Mediana Opaco 459 Crema Negro Moteado Pequeña Semi‐Brillante
285 Blanco ‐ ‐ Pequeña Semi‐Brillante 461 Café/Marrón Negro Jaspeado Mediana Semi‐Brillante
287 Blanco ‐ ‐ Mediana Opaco 464 Café/Marrón ‐ ‐ Grande Semi‐Brillante
288 Crema Negro Rayado Grande Semi‐Brillante 465 Café/Marrón Negro Jaspeado Pequeña Semi‐Brillante
289 Crema Negro Rayado Grande Semi‐Brillante 468 Rojo ‐ ‐ Grande Semi‐Brillante
290 Crema Negro Rayado Grande Semi‐Brillante 474 Rojo ‐ ‐ Mediana Semi‐Brillante
291 Crema Negro Rayado Grande Semi‐Brillante 475 Amarillo Café/Marrón Punteado Pequeña Semi‐Brillante
292 Café/Marrón Negro Rayado Grande Semi‐Brillante 476 Rojo ‐ ‐ Mediana Semi‐Brillante
295 Café/Marrón Negro Rayado Mediana Semi‐Brillante 478 Rojo ‐ ‐ Mediana Semi‐Brillante
296 Café/Marrón Negro Rayado Grande Semi‐Brillante 479 Rojo ‐ ‐ Mediana Semi‐Brillante
297 Crema Negro Rayado Grande Semi‐Brillante 480 Rojo ‐ ‐ Mediana Semi‐Brillante
298 Café/Marrón Negro Rayado Grande Semi‐Brillante 481 Rojo ‐ ‐ Mediana Semi‐Brillante
299 Café/Marrón Negro Rayado Grande Semi‐Brillante 492 Rojo ‐ ‐ Pequeña Semi‐Brillante
104

Anexo 4. (cont.) Descripción de características morfológicas de la semilla de fríjol
Cod Color Principal Color Secundario Patrón de Color Tamaño Brillo
497 Crema ‐ ‐ Mediana Semi‐Brillante
498 Crema ‐ ‐ Mediana Semi‐Brillante
500 Rojo ‐ ‐ Pequeña Semi‐Brillante
507 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
509 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
514 Café/Marrón Negro ‐ Grande Semi‐Brillante
516 Rojo ‐ ‐ Mediana Semi‐Brillante
518 Rojo ‐ ‐ Pequeña Semi‐Brillante
522 Amarillo ‐ ‐ Pequeña Semi‐Brillante
523 Amarillo Café/Marrón Punteado Mediana Semi‐Brillante
526 Amarillo Café/Marrón Punteado Mediana Semi‐Brillante
528 Amarillo Café/Marrón Punteado Mediana Semi‐Brillante
530 Amarillo Café/Marrón Punteado Mediana Semi‐Brillante
536 Gris/Raros Café/Marrón ‐ Mediana Semi‐Brillante
538 Rojo ‐ ‐ Mediana Semi‐Brillante
539 Café/Marrón Negro Jaspeado Pequeña Semi‐Brillante
540 Crema Negro Jaspeado Pequeña Semi‐Brillante
542 Rojo Crema Moteado Grande Semi‐Brillante
550 Café/Marrón Café/Marrón Moteado Pequeña Semi‐Brillante
552 Crema Rojo Jaspeado Mediana Semi‐Brillante
554 Café/Marrón Café/Marrón Jaspeado Pequeña Semi‐Brillante
556 Crema ‐ ‐ Pequeña Semi‐Brillante
557 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
563 Crema ‐ ‐ Pequeña Semi‐Brillante
564 Crema Negro Moteado Pequeña Semi‐Brillante
567 Café/Marrón Negro Jaspeado Mediana Semi‐Brillante
568 Café/Marrón Negro Moteado Mediana Semi‐Brillante
570 Crema Negro Moteado Pequeña Semi‐Brillante
572 Crema Negro Moteado Mediana Semi‐Brillante
573 Crema Negro Jaspeado Mediana Semi‐Brillante
574 Café/Marrón Negro Jaspeado Mediana Semi‐Brillante
575 Crema Negro Jaspeado Mediana Semi‐Brillante
582 Café/Marrón Crema Moteado Pequeña Semi‐Brillante
586 Rojo ‐ ‐ Pequeña Semi‐Brillante
595 Negro ‐ ‐ Pequeña Semi‐Brillante
596 Negro ‐ ‐ Pequeña Opaco
597 Negro ‐ ‐ Pequeña Opaco
601 Negro ‐ ‐ Pequeña Semi‐Brillante
603 Negro ‐ ‐ Pequeña Semi‐Brillante
604 Negro ‐ ‐ Pequeña Opaco
605 Negro ‐ ‐ Mediana Semi‐Brillante
606 Café/Marrón Negro Rayado Mediana Semi‐Brillante
609 Rojo ‐ ‐ Grande Semi‐Brillante
611 Rosado ‐ ‐ Mediana Semi‐Brillante
613 Rojo ‐ ‐ Mediana Semi‐Brillante
620 Rojo ‐ ‐ Mediana Semi‐Brillante
625 Rojo ‐ ‐ Mediana Semi‐Brillante
630 Café/Marrón ‐ Moteado Mediana Semi‐Brillante
631 Rojo ‐ ‐ Mediana Semi‐Brillante
634 Rojo ‐ ‐ Mediana Semi‐Brillante
636 Morado/Púrpura ‐ ‐ Mediana Semi‐Brillante
637 Café/Marrón ‐ ‐ Mediana Semi‐Brillante
640 Café/Marrón ‐ ‐ Mediana Semi‐Brillante
642 Crema ‐ ‐ Pequeña Semi‐Brillante
647 Crema ‐ ‐ Mediana Semi‐Brillante
655 Rojo Crema Moteado Mediana Semi‐Brillante
667 Negro ‐ ‐ Pequeña Semi‐Brillante
672 Crema ‐ ‐ Pequeña Semi‐Brillante
674 Rojo ‐ ‐ Mediana Semi‐Brillante
676 Rojo ‐ ‐ Pequeña Semi‐Brillante
678 Rojo ‐ ‐ Mediana Semi‐Brillante
679 Rojo ‐ ‐ Pequeña Semi‐Brillante
682 Café/Marrón ‐ ‐ Pequeña Semi‐Brillante
CALIMA Rojo Crema Moteado Grande Brillante
DOR364 Rojo ‐ ‐ Mediana Semi‐Brillante
G19833 Café/Marrón Rojo Jaspeado Grande Brillante
ICA‐PIJAO Negro ‐ ‐ Pequeña Opaca
105

Anexo 5. Prueba F y t de comparación de varianzas y medias para los parámetros de diversidad analizados en la población de estudio
Two Sample Test for Variances of Het__Observada_ and Het__Observada_1 1
15:13 Wednesday, October 31, 2007
Sample Statistics
Group N Mean Std. Dev. Variance
‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐
Het__Observada_ 16 0.2025 0.1729 0.029887
Het__Observada_1 14 0.172143 0.1925 0.037064
Hypothesis Test
Null hypothesis: Variance 1 / Variance 2 = 1
Alternative: Variance 1 / Variance 2 ^= 1
‐ Degrees of Freedom ‐
F Numer. Denom. Pr > F
‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐
0.81 15 13 0.6832
R// varianzas no diferentes
distribución prueba f
106

Anexo 5 (cont). Prueba F y t de comparación de varianzas y medias para los parámetros de diversidad analizados en la población de estudio
Prueba t
Two Sample t‐test for the Means of Het__Observada_ and Het__Observada_1 3
Sample Statistics
Group N Mean Std. Dev. Std. Error
‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐
Het__Observada_ 16 0.2025 0.1729 0.0432
Het__Observada_1 14 0.172143 0.1925 0.0515
Hypothesis Test
Null hypothesis: Mean 1 ‐ Mean 2 = 0
Alternative: Mean 1 ‐ Mean 2 ^= 0
If Variances Are t statistic Df Pr > t
‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐
Equal 0.455 28 0.6525
Not Equal 0.452 26.42 0.6551
R/// Medias no diferentes, igual se toma la primera por la prueba f que dijo que no sondiferentes
Distribucion t
107

Anexo 5 (cont). Prueba F y t de comparación de varianzas y medias para los parámetros de diversidad analizados en la población de estudio
Para diversidad
Two Sample Test for Variances of Diversidad_Gen_tica and Diversidad_Gen_tica1 1
Sample Statistics
Group N Mean Std. Dev. Variance
‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐
Diversidad_Gen_tica 16 0.706875 0.1842 0.033916
Diversidad_Gen_tica1 14 0.526429 0.1885 0.035517
Hypothesis Test
Null hypothesis: Variance 1 / Variance 2 = 1
Alternative: Variance 1 / Variance 2 ^= 1
‐ Degrees of Freedom ‐
F Numer. Denom. Pr > F
‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐
0.95 15 13 0.9224
R// varianzas no diferentes distribución prueba f
108

Anexo 5 (cont). Prueba F y t de comparación de varianzas y medias para los parámetros de diversidad analizados en la población de estudio
Two Sample t‐test for the Means of Diversidad_Gen_tica and Diversidad_Gen_tica1 2
Sample Statistics
Group N Mean Std. Dev. Std. Error
‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐
Diversidad_Gen_tica 16 0.706875 0.1842 0.046
Diversidad_Gen_tica1 14 0.526429 0.1885 0.0504
Hypothesis Test
Null hypothesis: Mean 1 ‐ Mean 2 = 0
Alternative: Mean 1 ‐ Mean 2 ^= 0
If Variances Are t statistic Df Pr > t
‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐‐
Equal 2.649 28 0.0131
Not Equal 2.644 27.29 0.0134
R// medias diferentes
distribución prueba t
109

Anexo 6. ANOVA para contenidos de minerales (hierro y zinc) a partir de las poblaciones generadas mediante PCoA.
The GLM Procedure
Class Level Information
Class Levels Values grupo_ 6 1 2 3 4 5 6
Number of observations 353
NOTE: All dependent variables are consistent with respect to the presence or absence of missing values. However only 343 observations can be used in this analysis.
09:39 Wednesday, November 14, 2007 2
The GLM Procedure Dependent Variable: Fe_ Fe Sum of Source DF Squares Mean Square F Value Pr > F Model 5 549.41626 109.88325 1.57 0.1676 Error 337 23570.50801 69.94216 Corrected Total 342 24119.92427 R-Square Coeff Var Root MSE Fe_ Mean 0.022779 12.25295 8.363143 68.25414 Source DF Type III SS Mean Square F Value Pr > F grupo_ 5 549.4162648 109.8832530 1.57 0.1676
09:39 Wednesday, November 14, 2007 3The GLM Procedure
Dependent Variable: Zn_ Zn Sum of Source DF Squares Mean Square F Value Pr > F Model 5 496.840452 99.368090 9.88 <.0001 Error 337 3389.091097 10.056650 Corrected Total 342 3885.931549 R-Square Coeff Var Root MSE Zn_ Mean 0.127856 9.196127 3.171222 34.48432 Source DF Type III SS Mean Square F Value Pr > F grupo_ 5 496.8404522 99.3680904 9.88 <.0001
110

Anexo 6 (cont). ANOVA para contenidos de minerales (hierro y zinc) a partir de las poblaciones generadas mediante PCoA.
09:39 Wednesday, November 14, 2007 4
The GLM Procedure Least Squares Means
LSMEAN grupo_ Fe_ LSMEAN Number 1 68.5490573 1 2 69.1892573 2 3 67.4307669 3 4 66.6529650 4 5 65.4870095 5 6 70.9563438 6
Least Squares Means for effect grupo_ Pr > |t| for H0: LSMean(i)=LSMean(j)
Dependent Variable: Fe_ i/j 1 2 3 4 5 6 1 0.6202 0.4379 0.2644 0.1480 0.3054 2 0.6202 0.1497 0.0944 0.0607 0.4257 3 0.4379 0.1497 0.6359 0.3479 0.1272 4 0.2644 0.0944 0.6359 0.6052 0.0829 5 0.1480 0.0607 0.3479 0.6052 0.0496 6 0.3054 0.4257 0.1272 0.0829 0.0496
LSMEAN
grupo_ Zn_ LSMEAN Number
1 35.5382718 1 2 33.0242438 2 3 35.4809358 3 4 35.7669388 4 5 33.9813857 5 6 35.1076594 6
Least Squares Means for effect grupo_ Pr > |t| for H0: LSMean(i)=LSMean(j)
Dependent Variable: Zn_ i/j 1 2 3 4 5 6 1 <.0001 0.9164 0.7224 0.0527 0.6285 2 <.0001 <.0001 <.0001 0.2003 0.0136 3 0.9164 <.0001 0.6461 0.0567 0.6697 4 0.7224 <.0001 0.6461 0.0374 0.4827 5 0.0527 0.2003 0.0567 0.0374 0.2853 6 0.6285 0.0136 0.6697 0.4827 0.2853 NOTE: To ensure overall protection level, only probabilities associated with pre-planned comparisons should be used.
111

Anexo 6 (cont). ANOVA para contenidos de minerales (hierro y zinc) a partir de las poblaciones generadas mediante PCoA.
9:39 Wednesday, November 14, 2007 5
The GLM Procedure
Ryan-Einot-Gabriel-Welsch Multiple Range Test for Fe_
NOTE: This test controls the Type I experimentwise error rate.
Alpha 0.05 Error Degrees of Freedom 337 Error Mean Square 69.94216 Harmonic Mean of Cell Sizes 34.79191
NOTE: Cell sizes are not equal.
Number of Means 2 3 4 5 6 Critical Range 4.8120927 5.2402928 5.4729326 5.4993403 5.7471385
Means with the same letter are not significantly different.
REGWQ Grouping Mean N grupo_
A 70.956 16 6 A
A 69.189 130 2 A
A 68.549 62 1 A
A 67.431 74 3 A
A 66.653 40 4 A
A 65.487 21 5 09:39 Wednesday, November 14, 2007 6
The GLM Procedure
Ryan-Einot-Gabriel-Welsch Multiple Range Test for Zn_
NOTE: This test controls the Type I experimentwise error rate.
Alpha 0.05 Error Degrees of Freedom 337 Error Mean Square 10.05665 Harmonic Mean of Cell Sizes 34.79191
NOTE: Cell sizes are not equal.
Number of Means 2 3 4 5 6 Critical Range 1.8246986 1.9870679 2.0752826 2.0852962 2.1792588
Means with the same letter are not significantly different.
112

Anexo 6 (cont). ANOVA para contenidos de minerales (hierro y zinc) a partir de las poblaciones generadas mediante PCoA.
REGWQ Grouping Mean N grupo_
A 35.7669 40 4
A A 35.5383 62 1
A A 35.4809 74 3
A A 35.1077 16 6
A B A 33.9814 21 5
B B 33.0242 130 2
113
Anexo 7. Tablas de contingencia que describen la distribución de cada característica morfológica (color, patrón de coloración y tamaño) dentro de cada grupo de PCoA formado.
vember 14, 2007 1
The FREQ Procedure
Table of clustert by color
clustert color
Frequency|Amarillo|Blanco |Cafe/Mar|Crema |Gris/Rar|Morado/P|Negro |Rojo |Rosado | Total | | |ron | |os |urpura | | | | ---------+--------+--------+--------+--------+--------+--------+--------+--------+--------+ 1 | 14 | 1 | 19 | 5 | 0 | 0 | 16 | 8 | 2 | 65 ---------+--------+--------+--------+--------+--------+--------+--------+--------+--------+ 2 | 23 | 0 | 17 | 31 | 1 | 2 | 2 | 55 | 1 | 132 ---------+--------+--------+--------+--------+--------+--------+--------+--------+--------+ 3 | 14 | 2 | 28 | 11 | 4 | 0 | 15 | 5 | 0 | 79 ---------+--------+--------+--------+--------+--------+--------+--------+--------+--------+ 4 | 2 | 0 | 14 | 2 | 1 | 0 | 8 | 15 | 1 | 43 ---------+--------+--------+--------+--------+--------+--------+--------+--------+--------+ 5 | 4 | 1 | 14 | 2 | 0 | 0 | 0 | 0 | 0 | 21 ---------+--------+--------+--------+--------+--------+--------+--------+--------+--------+ 6 | 5 | 2 | 2 | 1 | 0 | 0 | 2 | 4 | 1 | 17 ---------+--------+--------+--------+--------+--------+--------+--------+--------+--------+ 7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 ---------+--------+--------+--------+--------+--------+--------+--------+--------+--------+ 8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 ---------+--------+--------+--------+--------+--------+--------+--------+--------+--------+ Total 62 6 94 52 6 2 43 89 5 359
Table of clustert by patron
clustert patron
Frequency|Jaspeado|Moteado |NP |Punteado|Rayado |Venacion| Total ---------+--------+--------+--------+--------+--------+--------+ 1 | 3 | 5 | 51 | 1 | 1 | 4 | 65 ---------+--------+--------+--------+--------+--------+--------+ 2 | 6 | 29 | 48 | 11 | 38 | 0 | 132 ---------+--------+--------+--------+--------+--------+--------+ 3 | 8 | 9 | 54 | 4 | 0 | 4 | 79 ---------+--------+--------+--------+--------+--------+--------+ 4 | 0 | 2 | 37 | 1 | 0 | 3 | 43 ---------+--------+--------+--------+--------+--------+--------+ 5 | 15 | 2 | 3 | 0 | 0 | 1 | 21 ---------+--------+--------+--------+--------+--------+--------+ 6 | 0 | 2 | 11 | 3 | 0 | 1 | 17 ---------+--------+--------+--------+--------+--------+--------+ 7 | 0 | 0 | 1 | 0 | 0 | 0 | 1 ---------+--------+--------+--------+--------+--------+--------+ 8 | 0 | 0 | 0 | 1 | 0 | 0 | 1 ---------+--------+--------+--------+--------+--------+--------+ Total 32 49 205 21 39 13 359
The FREQ Procedure
Table of clustert by tamano
clustert tamano
Frequency|Grande |Mediana |Pequena | Total ---------+--------+--------+--------+ 1 | 0 | 13 | 52 | 65 ---------+--------+--------+--------+ 2 | 28 | 100 | 4 | 132 ---------+--------+--------+--------+ 3 | 0 | 29 | 50 | 79 ---------+--------+--------+--------+ 4 | 0 | 1 | 42 | 43 ---------+--------+--------+--------+ 5 | 0 | 12 | 9 | 21 ---------+--------+--------+--------+ 6 | 0 | 10 | 7 | 17 ---------+--------+--------+--------+ 7 | 0 | 0 | 1 | 1 ---------+--------+--------+--------+ 8 | 0 | 0 | 1 | 1 ---------+--------+--------+--------+ Total 28 165 166 359
114
115
Anexo 8. Comparación de agrupación de genotipos entre el índice de Disimilaridad de DICE utilizado por el software Darwin y las agrupaciones por PCoA.
ggd ( GRANDES GRUPOS Darwin) A_FGHI I____E I___J M_ABCD Total
Pcoa
M 1 65 65 A 2 126 1 5 132
M+I 3 7 3 69 79 M 4 43 43 M 5 21 21
M+I 6 1 6 4 6 17 raro 7 1 1 raro 8 1 1
Total 127 15 12 205 359


















