
Preventing Gasoline Burn Injuries American Burn Association Burn Prevention Committee.

Ionic Nature of Burn-Induced Variation Potential inWheat LeavesLyubov Katicheva, Vladimir Sukhov, Elena Akinchits and Vladimir Vodeneev*Department of Biophysics, Lobachevsky State University of Nizhni Novgorod, Gagarina Av., 23, Nizhni Novgorod, Russia*Corresponding author: E-mail, [email protected]; Fax, 8(831) 462-32-02.(Received July 18, 2013; Accepted June 9, 2014)
Variation potential (VP) in higher plants cells is a transitorydepolarization of the plasma membrane occurring in re-sponse to external damage. The effects of VP on differentphysiological processes are actively studied, but little isknown about their ionic nature, which limits the interpret-ation of VP-induced functional changes. It is thought thatVP generation is based on transient inactivation of plasmamembrane proton pumps and is not connected to passiveionic fluxes. To study burn-induced VP in wheat seedlings,we measured membrane electric potential and cell inputresistance. Cell input resistance decreased during VP gener-ation, indicating that ionic channels were activated. In add-ition, VP amplitude decreased when the extracellularcalcium concentration was lowered. When anion channelswere blocked by ethacrynic acid addition, the VP had poordepolarization speed and amplitude. A decrease in thechlorine gradient by extracellular chlorine concentrationshift leads to lowering of the VP amplitude and depolariza-tion speed. This result indicates the role of chlorine efflux indepolarization phase formation. The VP repolarization isconnected to potassium ion efflux, that is confirmed byrepolarization suppression under addition of the potassiumchannel blocker tetraethylammonium (TEA) and an increasein the extracellular potassium concentration. We alsoshowed that the addition of a proton pump inhibitorleads to membrane potential depolarization and inhibitionof VP generation. These results suggest that the VP may beformed not only by transient suppression of proton pumpsbut also by passive ionic fluxes through the membrane.
Keywords: Cell input resistance � Microelectrode technique� Passive ionic fluxes � Proton pump � Variation potential �
Wheat.
Abbreviations: AP, action potential; APW, artificial pondwater; Avp, amplitude of variation potential; EA, ethacrynicacid; Em, transmembrane electrical potential; Em
r, transmem-brane electrical potential at rest; Rin, cell input resistance;TEA, tetraethylammonium; VD, velocity of depolarization;VP, variation potential; VR, velocity of repolarization; WS,wound signal.
Introduction
Electrical signals in higher plants have been reported to play akey role in stress sensing and systemic response development.Plants show two types of electrical reactions depending on thenature of the stimulation: action potential (AP) and variationpotential (VP). A plethora of physiological responses inducedby electrical signals have been observed in plants, includingchanges in photosynthetic processes, increase in the respirationrate and gene expression activation (Trebacz et al. 2006, Frommand Lautner 2007, Baluska and Mancuso 2009, Grams et al.2009, Sukhov et al. 2012). Though little is known about themechanisms of induction of functional changes by AP or VP,it is considered that ionic concentration shifts in cells duringelectrical reaction development play the main role in suchtransformation (Trebacz et al. 2006, Pyatygin et al. 2008). Theionic nature of the AP is currently better described; its forma-tion starts with Ca2+ influx and the opening of chlorine chan-nels, which depolarizes the plasma membrane. Shortly after Cl�
efflux has begun, the AP peaks, and K+ efflux starts processesleading to repolarization (Trebacz et al. 2006, Fromm andLautner 2007).
Unlike APs, VPs are not self-propagated electrical signals.The prevalent hypothesis of VP propagation relates to theirinduction by localized damaged hydraulic waves, which causegeneration of an electrical response in plant cells. The possibilityof such a propagation mechanism has been proved experimen-tally in a number of studies (Malone et al. 1994, Stahlberg andCosgrove 1997, Mancuso, 1999). However, according to anotherhypothesis, suggested by Ricca (1916), VPs might also beinduced by migration of a wound chemical substance fromthe zone of damage (Rhodes et al. 1999, Vodeneev et al. 2012).
The mechanism of VP generation also differs from that ofAPs and seems to reflect only a transient inactivation of protonpumps at the plasma membrane without a contribution frompassive ionic fluxes (Julien et al. 1991, Stahlberg et al. 2006).Proton pump participation in VP generation was identifiedby modulation of H+-ATPase activity and membrane perme-ability to H+, and measurement of apoplastic pH changesduring VP generation (Julien et al. 1991, Stahlberg et al. 2006,Vodeneev et al. 2011). In some studies, the absence of passive
Plant Cell Physiol. 55(8): 1511–1519 (2014) doi:10.1093/pcp/pcu082, available online at www.pcp.oxfordjournals.org! The Author 2014. Published by Oxford University Press on behalf of Japanese Society of Plant Physiologists.All rights reserved. For permissions, please email: [email protected]
1511Plant Cell Physiol. 55(8): 1511–1519 (2014) doi:10.1093/pcp/pcu082 ! The Author 2014.
Regu
larP
aper
at FMR
P/USP/B
IBL
IOT
EC
A C
EN
TR
AL
on June 9, 2016http://pcp.oxfordjournals.org/
Dow
nloaded from

ionic fluxes through membrane channels during VP develop-ment was shown by several parameters: stable membrane re-sistance at the VP (Stahlberg and Cosgrove 1996, Stahlberg et al.2006), no measurable changes in membrane permeability to K+,Cl–, Ca2+ and Na+ during VP development (Julien et al. 1991)and no changes in VP parameters with ionic channel activityinhibitors (Stahlberg and Cosgrove 1996, Rousset et al. 2002).
Conversely, the complex kinetics of VPs with fast potentialchanges at the VP front slope and the occurrence of impulsesmay be related to the activation of ion channels during VP for-mation, and the involvement of passive ionic fluxes in VP gen-eration is still under discussion. Calcium and chlorine ionconcentration shifts during development of the electrical reac-tion (Zimmerman and Felle 2009) are in good agreement with apossible role for passive ionic fluxes in the VP. It has also beenthought that calcium ions may cause an efflux of anions into theapoplast to induce fast depolarization during VPs (Vodeneevet al. 2011). Moreover, theoretical analysis of VP generationmechanisms also indicates the possibility that passive ionicfluxes participate in this process (Sukhov et al. 2013).
The aim of the present study was to analyze the possibleinvolvement of passive ionic fluxes, i.e. Ca2+, Cl� and K+, inthe generation of VPs. For the recognition of the ionic diffusionduring VPs, cell input resistance was recorded simultaneouslywith electrical activity. Changes in VP parameters with the add-ition of different inhibitors served as indicators of the participa-tion of calcium, chlorine or potassium channels in VP generation.
Results
VP characteristics
The transmembrane electrical potential (Em) of parenchymalcells recorded without leaf tip burning (Em
r) was �98 ± 3 mV.Leaf tip burning led to the generation of a VP; transient mem-brane depolarization with an amplitude (AVP) of 75 ± 4 mV, andspeeds of membrane depolarization (VD) of 9.09 ± 0.09 mV s�1
and repolarization (VR) of 0.54 ± 0.09 mV s�1 (Fig. 2). Changesin these parameters under various inhibitors and ionic concen-trations were used to identify the ionic flows that participate inVP generation.
VP generation and resistance changes
The stability of Em shifts induced by an applied current waschecked before VP initiation. As seen in Fig. 3A, without per-turbation the amplitude of Em shifts was constant, i.e. currentpulses with the chosen parameters did not affect voltage-dependent processes at the plasma membrane and cell inputresistance (Rin). Fig. 3B shows a typical recording of VP inducedby leaf burning with simultaneous measurement of resistancechanges. It should be noted that the recorded VPs had param-eters close to those of one recorded without the application ofcurrent pulses. As seen in Fig. 3B, there was a decrease in Rin
during VP generation. Rin dropped at the beginning of depolar-ization and slowly recovered in the repolarization phase. Such
changes in Rin may indicate the appearance of passive ionicfluxes through activated channels.
Inhibitor and ion concentration effects on VPgeneration
To reveal the transport systems taking part in VP formation,metabolic inhibitors and channels blockers were used.
The role of H+-ATPase in VP generation was studied by theaddition of sodium orthovanadate (0.5 mM). As seen in Fig. 4A,the suppression of the proton pump activity led to a membranedepolarization of 32 ± 0.8 mV and the leaf injury resulted onlyin a very weak electrical reaction without an expressed repolar-ization phase.
The participation of calcium ions in VP generation was esti-mated by the addition of the calcium chelator EGTA (1 mM) tothe artificial pond water (APW). The absence of free calcium hadno effect on Em values but AVP decreased dramatically (Fig. 4B).
Fig. 4C shows that the addition of an anion channels blocker[ethacrynic acid (EA), 0.5 mM] also led to VP amplitude de-crease. Moreover, the depolarization speed was also decreased.
Analysis of potassium ion participation in VP generation wasperformed by adding a potassium channel blocker [tetraethy-lammonium (TEA), 0.5 mM] to the medium. The electric reac-tion to the leaf injury in the presence of TEA exhibited differentbehavior to that of the control VP; in particular, the amplitude ofthe depolarization and membrane repolarization decreased at aslower rate (Fig. 4D). Moreover, the addition of both TEA and EAto the medium resulted in a small Em depolarization and con-siderable suppression of the electric reaction to burning (Fig. 4E).
In order to provide additional validation of the VP experi-ments carried out using ionic channel blockers, we performedmeasurements of the changes in VP vs. ionic concentration inthe extracellular space, with the results summarized in Table 1.Both the amplitude and depolarization speed were increased, asthe extracellular calcium molar concentration was increased ina range from 0.05 to 5 mM, which we interpret to be due to theincrease of the calcium influx to the cell. At the same time, theincrease of the extracellular chlorine concentration in the rangeof 1–10 mM resulted in a notable drop in the decrease of the VPamplitude and depolarization speed, as the efflux of chlorineions from the cell was suppressed. At the same time, the in-crease of the potassium ion concentration suppressed the repo-larization speed and had a negligible effect on the amplitudeand speed of depolarization of VP. No essential changes in Em
r
were detected following 1 h incubation of the plant in the solu-tions with different ionic concentrations. The presented evi-dence indicates that the passive ion fluxes play an importantrole in VP generation, as discussed in the following section.
Changes of membrane resistance under the effectof inhibitors on VP
To identify the ionic channels whose activation results in re-sistance changes, the VP and Rin were recorded simultaneouslyafter the addition of channels blockers.
1512 Plant Cell Physiol. 55(8): 1511–1519 (2014) doi:10.1093/pcp/pcu082 ! The Author 2014.
L. Katicheva et al.
at FMR
P/USP/B
IBL
IOT
EC
A C
EN
TR
AL
on June 9, 2016http://pcp.oxfordjournals.org/
Dow
nloaded from

Wheat leaf Wheat leaf
Microelectrode current/voltage clamp amplifier, PC
ErE Cuvette with: artificial pond water (APW) or APW with inhibitors
Wheat leaf
Burn
in out
E I = 40pA E Er
Er
Wheat roots
Objective
Light source
Microscope stage
V, mV
in out
Rin =
Fig. 1 Scheme of the experimental system for measuring Em and Rin. A wheat leaf is placed on the microscope stage, part of it is fixed in thecuvette, and the measuring electrode is inserted in a parenchymal cell. The site of burning is indicated by the arrow. The separate scheme on theleft illustrates registration of the membrane potential (Em); the separate scheme on the right illustrates registration of the cell input resistance(Rin). For the Rin measurement, current pulses with an amplitude of 40 pA alternated with frequencies of 2 Hz injected through the electrode tocells. The amplitude of the membrane potential deflections was then used to calculate the Rin by Ohm’s law equation. E, measuring electrode; Er,reference electrode; I, amplitude of the current pulses injected into the cells; �V, amplitude of the membrane potential deflections.
Fig. 2 Variation potential in wheat induced by leaf tip burning (arrow). The measuring electrode was located in the leaf parenchymal cell of thesecond to third layers. The reference electrode was immersed in APW. The distance between the burning site and recording zone wasapproximately 5 cm. (A) A typical record of the variation potential (VP). (B) The parameters of the electric reaction: Em
r, membrane voltageat rest (mV); AVP, amplitude of the VP (mV); VD, speed of VP depolarization (mV s�1); VR, speed of VP repolarization (mV s�1).
1513Plant Cell Physiol. 55(8): 1511–1519 (2014) doi:10.1093/pcp/pcu082 ! The Author 2014.
Ionic nature of variation potential
at FMR
P/USP/B
IBL
IOT
EC
A C
EN
TR
AL
on June 9, 2016http://pcp.oxfordjournals.org/
Dow
nloaded from

-100
-80
-60
-40
-20
0
-110
-90
-70
-50
-30
-10
-80
-60
-40
-20
0
-100
-80
-60
-40
-20
0Em, mV Em, mV
Em, mV Em, mV
100s
BA
C D
E
-100
-80
-60
-40
-20
0Em, mV
0
20
40
60
80
100
1 2 3 4
0
20
40
60
80
100
1 2 3 4 Emr AVP VD
0
20
40
60
80
100
1 2 3 4
0
20
40
60
80
100
1 2 3 4
0
20
40
60
80
100
1 2 3 4
%
%
%
Emr AVP VD
Emr AVP VD
Emr AVP VD
Emr AVP VD
%
%
Fig. 4 Changes in parameters of VP induced by leaf tip burning (arrow) with addition of inhibitors. Inhibitors were added separately into thecuvette where the 4 cm area of the leaf was fixed. Plants were incubated in the presence of the inhibitors for at least 1 r. In the diagrams, themeasured VP parameters are presented as a percentage of the control values. (A) The effect of proton pump inactivation by sodiumorthovanadate (0.1 mM) on VP parameters. (B) The effect of the calcium chelator EGTA (0.5 mM) on VP parameters. (C) The effect of theanion channel blocker EA (0.5 mM) on VP parameters. (D) The effect of the potassium channel blocker TEA (0.5 mM) on VP parameters. (E) Theeffect of addition of both EA (0.5 mM) and TEA (0.5 mM) on VP parameters.
Fig. 3 Changes in cell input resistance during VP generation. For calculation of Rin, current clamp mode was used and square current pulses withfrequencies of 2 Hz and an amplitude of 40 pA were applied. (A) A typical record of the membrane voltage with transient induction by currentpulses and calculated Rin at rest. (B) Changes in membrane voltage with transient induction by current pulses and calculated membraneresistance after leaf tip burning (arrow). 1, Em; 2, Rin.
1514 Plant Cell Physiol. 55(8): 1511–1519 (2014) doi:10.1093/pcp/pcu082 ! The Author 2014.
L. Katicheva et al.
at FMR
P/USP/B
IBL
IOT
EC
A C
EN
TR
AL
on June 9, 2016http://pcp.oxfordjournals.org/
Dow
nloaded from
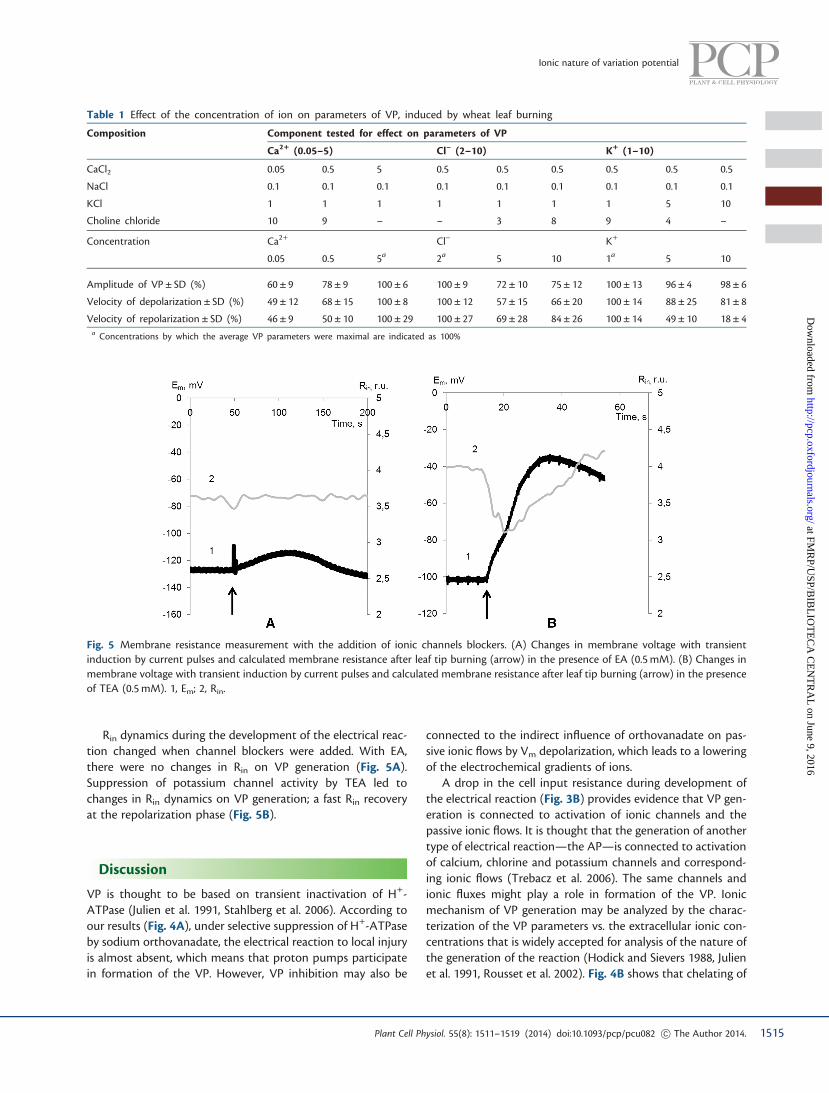
Rin dynamics during the development of the electrical reac-tion changed when channel blockers were added. With EA,there were no changes in Rin on VP generation (Fig. 5A).Suppression of potassium channel activity by TEA led tochanges in Rin dynamics on VP generation; a fast Rin recoveryat the repolarization phase (Fig. 5B).
Discussion
VP is thought to be based on transient inactivation of H+-ATPase (Julien et al. 1991, Stahlberg et al. 2006). According toour results (Fig. 4A), under selective suppression of H+-ATPaseby sodium orthovanadate, the electrical reaction to local injuryis almost absent, which means that proton pumps participatein formation of the VP. However, VP inhibition may also be
connected to the indirect influence of orthovanadate on pas-sive ionic flows by Vm depolarization, which leads to a loweringof the electrochemical gradients of ions.
A drop in the cell input resistance during development ofthe electrical reaction (Fig. 3B) provides evidence that VP gen-eration is connected to activation of ionic channels and thepassive ionic flows. It is thought that the generation of anothertype of electrical reaction—the AP—is connected to activationof calcium, chlorine and potassium channels and correspond-ing ionic flows (Trebacz et al. 2006). The same channels andionic fluxes might play a role in formation of the VP. Ionicmechanism of VP generation may be analyzed by the charac-terization of the VP parameters vs. the extracellular ionic con-centrations that is widely accepted for analysis of the nature ofthe generation of the reaction (Hodick and Sievers 1988, Julienet al. 1991, Rousset et al. 2002). Fig. 4B shows that chelating of
Fig. 5 Membrane resistance measurement with the addition of ionic channels blockers. (A) Changes in membrane voltage with transientinduction by current pulses and calculated membrane resistance after leaf tip burning (arrow) in the presence of EA (0.5 mM). (B) Changes inmembrane voltage with transient induction by current pulses and calculated membrane resistance after leaf tip burning (arrow) in the presenceof TEA (0.5 mM). 1, Em; 2, Rin.
Table 1 Effect of the concentration of ion on parameters of VP, induced by wheat leaf burning
Composition Component tested for effect on parameters of VP
Ca2+ (0.05–5) Cl– (2–10) K+ (1–10)
CaCl2 0.05 0.5 5 0.5 0.5 0.5 0.5 0.5 0.5
NaCl 0.1 0.1 0.1 0.1 0.1 0.1 0.1 0.1 0.1
KCl 1 1 1 1 1 1 1 5 10
Choline chloride 10 9 – – 3 8 9 4 –
Concentration Ca2+ Cl– K+
0.05 0.5 5a 2a 5 10 1a 5 10
Amplitude of VP ± SD (%) 60 ± 9 78 ± 9 100 ± 6 100 ± 9 72 ± 10 75 ± 12 100 ± 13 96 ± 4 98 ± 6
Velocity of depolarization ± SD (%) 49 ± 12 68 ± 15 100 ± 8 100 ± 12 57 ± 15 66 ± 20 100 ± 14 88 ± 25 81 ± 8
Velocity of repolarization ± SD (%) 46 ± 9 50 ± 10 100 ± 29 100 ± 27 69 ± 28 84 ± 26 100 ± 14 49 ± 10 18 ± 4a Concentrations by which the average VP parameters were maximal are indicated as 100%
1515Plant Cell Physiol. 55(8): 1511–1519 (2014) doi:10.1093/pcp/pcu082 ! The Author 2014.
Ionic nature of variation potential
at FMR
P/USP/B
IBL
IOT
EC
A C
EN
TR
AL
on June 9, 2016http://pcp.oxfordjournals.org/
Dow
nloaded from

calcium ions leads to VP suppression. An increase in the extra-cellular calcium concentration from 0.05 to 5 mM gives rise tothe increase in the VP amplitude and depolarization speed, asshown in Table 1. It should also be noted that a decrease in theextracellular calcium concentration leads to lowering of therepolarization speed (Fig. 4B; Table 1). Such a result mightbe connected to an ability of calcium ions on the extracellularside to activate the proton pump (Qiu and Su 1998). Generallythese results convincingly prove that the calcium flux is essen-tial for VP generation in wheat leaves and corroborate theresults reported for other plants (Julien et al. 1991, Vodeneevet al. 2011).
For identification of participation of ionic channels in electricreactions, inhibition of transport system activity is also widelyused (Tyerman 1992, Shimmen 2002, Lautner et al. 2005). Inparticular, EA was used as chlorine channels blocker in manystudies on higher plants and algae (Marten et al. 1992, Johanneset al. 1998, Shimmen 2002, Tavares et al. 2011). The effect of TEAon potassium channels in higher plant cells was also widelyshown previously (Stahlberg and Cosgrove 1992, Grabov et al.1997, Lautner et al. 2005, Demidchik et al. 2010). The results weobtained when channels blockers were added indicate the pos-sibility that VP is formed by ionic channels (Fig. 4). In particular,the recorded drop in VP amplitude with EA addition indicatesthe participation of anion channels in the development of theburn-induced electrical reaction. The anions that make a con-tribution to depolarization could be represented by chlorineions. This supposition is based on the significant concentrationgradient of the chlorine ions and their participation in the APdepolarization (Opritov et al. 1991, Trebacz et al. 2006). A de-crease in the chlorine gradient by an extracellular chlorine con-centration shift leads to AVP and depolarization speed lowering(Table 1). Moreover, changes of apoplastic pCl which corres-ponds to chlorine ion efflux to the apoplast during VP gener-ation, were noticed (Zimmerman and Felle 2009, Vodeneev et al.2011). Hence, chlorine ions make a contribution to the mem-brane depolarization of VP.
The repolarization phase of VP in the wheat leaf is possiblyconnected not only with proton pump reactivation but alsowith potassium ion efflux. A decrease of potassium gradients byan increase in extracellular potassium concentration decreasesthe repolarization phase (Table 1). Addition of the potassiumchannel blocker TEA also leads to a decrease in repolarizationspeed (Fig. 4D). Under TEA, a small decrease in depolarizationspeed was also registered (Fig. 4D) that may be connected to anon-specific effect of the channel blocker.
It is known that changes in membrane resistance of the APcorrespond exactly to activation of chlorine and potassium chan-nels (Samejima and Sibaoka 1982). The fast drop in Rin at thestart of VP generation may also be connected to chlorine channelactivation, and the low values of Rin at repolarization may beconnected to potassium channel activation. This was supportedby the stable Rin values on VP generation when an anion channelblocker was added and by the fast Rin value recovery at therepolarization phase with TEA addition (Fig. 5A).
Our results show the participation of not only protonpumps, but also of passive ionic fluxes during VP generation.However, these results do not agree with previous studies inwhich the absence of passive ionic flows on VP generationwas reported. There are data showing stable levels of mem-brane resistance during VP formation (Stahlberg andCosgrove 1992, Stahlberg and Cosgrove 1996). It is worthnoting that the authors do not exclude the participationof passive ionic flows in the development of VP; in theirexperiments, membrane resistance may have remainedstable because the opening of ionic channels was compen-sated for by the closure of other channels (Stahlberg andCosgrove 1992). There are also results indicating that thereare no measurable changes in membrane permeability toCa2+, Cl– or K+ during VP development (Julien et al.1991). It should be noted that these permeability changeswere recorded during the slow repolarization phase and,according to our results, membrane resistance changesfrom the very beginning of the electrical reaction.
There are also some contradictions with our data on theinfluence of channel blockers on VP generation. Our resultsshow dramatic changes of VP parameters with the additionof inhibitors, whereas in other studies channel blockers hadno influence on VP (Stahlberg and Cosgrove 1996). At thesame time, in several works, the possible participation of cal-cium ions in VP generation has been shown by the reduced AVP
in calcium-free extracellular medium (Julien et al. 1991,Stahlberg and Cosgrove 1996) and with inactivation of calciumchannels by lanthanum (Julien et al. 1991).
Based on our results and data from the literature, thefollowing sequence of events might occur during VP gener-ation (Fig. 6). External damage induces formation of awound signal (WS) which might be manifested as a hydrau-lical wave (Malone et al. 1994, Mancuso 1999) or a chemicalagent (Rhodes et al. 1996, Vodeneev et al. 2012). The WStriggers stretch-activated (Stahlberg et al. 2006) or ligand-operated (Sukhov et al. 2013) calcium channels, and calciumions enter the cell. It is known that calcium ions can induceproton pump inactivation, that causes proton influx, andaffect Ca2+-dependent chlorine channels and chlorine ionefflux (Fromm and Lautner 2007, Vodeneev et al. 2011).Both processes lead to membrane depolarization. The startof the membrane repolarization is probably based on potas-sium ion diffusion out of the cell. Such consequent activa-tion of anionic and potassium channels leads to a transientdecrease in the cell input resistance, which has been regis-tered in our work. The reactivation of proton pumps makesa significant contribution to the subsequent repolarization,because a slow repolarization phase develops under morenegative voltage values than the Nernst potential for potas-sium ions (Stahlberg et al. 2006).
This suggested mechanism is similar to the model for APgeneration in higher plants (Vodeneev et al. 2006). The maindifference is probably in the initial activation of different typesof calcium channels; ligand-operated or stretch-activated in VP
1516 Plant Cell Physiol. 55(8): 1511–1519 (2014) doi:10.1093/pcp/pcu082 ! The Author 2014.
L. Katicheva et al.
at FMR
P/USP/B
IBL
IOT
EC
A C
EN
TR
AL
on June 9, 2016http://pcp.oxfordjournals.org/
Dow
nloaded from

instead of voltage-dependent in AP (Stahlberg et al. 2006,Sukhov et al. 2013). This difference might result in a morecontinuous increase in the concentration of calcium ions incells, causing a slower electrical reaction in comparison withthe development of AP.
Materials and Methods
Plant materials
Seedlings of wheat (Triticum aestivum L.) were cultivatedhydroponically in a Binder KBW 240 (Germany) plant growthchamber at 24�C under a 16/8 h (light/dark) photoperiod.Seedlings that were 14–21 d old with a leaf blade length of15–20 cm were used in experiments (Vodeneev et al. 2012).
Electrical measurements
For measurements of the intracellular membrane potential(Em) and resistance (Rin) dynamics, the microelectrode tech-nique [Olympus BX51microscope; Luigs and Neumann auto-matic manipulators; Multiclamp 700B amplifier (Axon Inst.;Molecular Devices); and a PC] was used. The entire plant was
placed at the side of the microscope, and the leaf was fixedhorizontally. Part of the leaf was placed in a cuvette filled withAPW, which comprised 0.1 mL NaCl, 0.1 mL KCl and 0.5 mL
CaCl2 (Fig. 1). For separate experiments, APW in the cuvettewas changed to a solution with inhibitors or different ioniccompositions as described below.
Single-barreled microelectrodes were fabricated from1.2 mm (OD) borosilicate glass tubing. Capillaries were pulledin a horizontal pipette puller and filled with 100 mM KCl. Thetip diameter was approximately 0.5 mm. The electrode wasmoved by motorized manipulators and its location waschecked visually using a microscope. Electrical measurementswere carried out in parenchymal cells of the second to thirdlayer. The reference electrode was immersed into a cuvette withAPW. The VP was induced by open-flame burning of the leaf tip(apical 2–3 mm) for 2 s (Vodeneev et al. 2012). The distancebetween the registration zone and the wounded area equaled5 cm. VP parameters such as amplitude (AVP), and depolariza-tion (VD) and repolarization (VR) speeds were estimated. TheVR was measured as the average speed of the membrane po-tential recovering from the VP peak to the potential values athalf of the AVP.
Ca2+
H+ H+
Ca2+ K+ Cl–
Em
in out
Burn
[Ca]in
Wheat leaf
WS
WS
Fig. 6 Scheme of the possible mechanism of the variation potential generation in wheat leaf. WS, wound signal; Em, membrane potential; [Ca]in,concentration of intracellular calcium.
1517Plant Cell Physiol. 55(8): 1511–1519 (2014) doi:10.1093/pcp/pcu082 ! The Author 2014.
Ionic nature of variation potential
at FMR
P/USP/B
IBL
IOT
EC
A C
EN
TR
AL
on June 9, 2016http://pcp.oxfordjournals.org/
Dow
nloaded from

For the measurement of Rin, changes in the membranevoltage under applied current pulses were estimated. Currentpulses with an amplitude of 40 pA alternated with frequenciesof 2 Hz depolarized cells by a few milliVolts. The amplitude ofthe membrane potential deflections was then used to calculatethe Rin (Stahlberg and Cosgrove 1992). A scheme of the experi-mental system for Em and Rin measurement is shown in Fig. 1.Calculated Rin includes membrane resistance and the resistanceof the electrode, which remains constant during themeasurements.
Inhibition of transport system activity
This analysis was based on measurements of VP parameterchanges under the influence of inhibitors. A direct inhibitorof the plasma membrane H+-ATPase activity, sodium orthova-nadate (Lew 1991), a calcium chelator, EGTA (Julien et al. 1991,Stahlberg and Cosgrove 1992), an anion channel blocker, EA(Tyerman 1992, Tavares et al. 2011), and a potassium channelblocker, TEA (Stahlberg and Cosgrove 1992, Lautner et al. 2005),were used. The concentrations of the chemical agents are indi-cated in the Results and in the figure legends. Plants wereincubated in the presence of the inhibitors for at least 1 h.The measured parameters of variation potentials are presentedas percentages of the control values.
Change in the ionic composition
Parameters of VP were measured under different ionic con-centrations in extracellular medium (Hodick and Sievers1988). For the preparation of different compositions, CaCl2,NaCl, KCl and choline chloride were used in several combin-ations of concentrations. To define the effect of each separ-ate ion by its concentration variation, the concentrations ofthe other active ions were maintained constant (Table 1).Plants were incubated in the compositions for at least 1 h.The final concentrations of each tested ions are presented inTable 1.
Statistical analysis
Each series of experiments comprised 5–8 repetitions, andevery measurement was performed on a separate plant.Representative records obtained in individual measurements,mean values and standard errors are presented.
Funding
This work was supported by the Ministry of Education andScience of the Russian Federation [contract No 6.2050.2014/K] and by the Russian Foundation for Basic Research [researchproject No 13-04-97152-r-a.].
Disclosures
The authors have no conflicts of interest to declare.
References
Baluska, F. and Mancuso, S. (2009) Plant neurobiology: from stimulusperception to adaptive behavior of plants, via integrated chemicaland electrical signaling. Plant Signal. Behav. 4: 475–476.
Demidchik, V., Cuin, T.A., Svistunenko, D., Smith, S.J., Miller, A.J.,Shabala, S., Sokolik, A. and Yurin, V. (2010) Arabidopsis root K+-efflux conductance activated by hydroxyl radicals: Single-channelproperties, genetic basis and involvement in stress-induced celldeath. J. Cell Sci. 123: 1468–1479.
Fromm, J. and Lautner, S. (2007) Electrical signals and their physio-logical significance in plants. Plant Cell Environ. 30: 249–257.
Grabov, A., Leung, J., Giraudat, J. and Blatt, M.R. (1997) Alteration ofanion channel kinetics in wild-type and abi1-1 transgenic Nicotianabenthamiana guard cells by abscisic acid. Plant J. 12: 203–213.
Grams, T.E.E., Lautner, S., Felle, H.H., Matyssek, R. and Fromm, J. (2009)Heat-induced electrical signals affect cytoplasmic and apoplastic pHas well as photosynthesis during propagation through the maizeleaf. Plant Cell Environ. 32: 319–326.
Hodick, D. and Sievers, A. (1988) The action potential of Dionaeamuscipula Ellis. Planta 174: 8–18.
Johannes, E., Crofts, A. and Sanders, D. (1998) Control of Cl� efflux inChara Corallina by cytosolic pH, free Ca2+, and phosphorylationindicates a role of plasma membrane anion channels in cytosolicpH regulation. Plant Physiol. 118: 173–181.
Julien, J.L., Desbiez, M.O., de Jaeger, G. and Frachisse, J.M. (1991)Characteristics of the wave of depolarization induced by woundingin Bidens pilosa L. J. Exp. Bot. 42: 131–137.
Lautner, S., Grams, T.E., Matyssek, R. and Fromm, J. (2005)Characteristics of electrical signals in poplar and responses inphotosynthesis. Plant Physiol. 138: 2200–2209.
Lew, R.R. (1991) Electrogenic transport properties of growingArabidopsis root hairs. Plant Physiol. 97: 1527–1534.
Malone, M., Alarcon, J.-J. and Palumbo, L. (1994) An hydraulic inter-pretation of rapid, long-distance wound signaling in the tomato.Planta 193: 181–185.
Mancuso, S. (1999) Hydraulic and electrical transmission of wound-induced signals in Vitis vinifera. Aust. J. Plant Physiol. 26: 55–61.
Marten, I., Zeilinger, C., Redhead, C., Landry, D.W., AI-Awqatil, Q. andHedrich, R. (1992) Identification and modulation of a voltage-dependent anion channel in the plasma membrane of guard cellsby high-affinity ligands. The EMBO Journal. 11: 3569–3575.
Opritov, V.A., Pyatygin, S.S. and Retivin, V.G. (1991) Bioelectrogenesisin higher plants. Nauka, Moskow [in Russian].
Pyatygin, S.S., Opritov, V.A. and Vodeneev, V.A. (2008) Signaling roleof action potential in higher plants. Russ. J. Plant Physiol. 55:285–291.
Qiu, Q.-S. and Su, X.-F. (1998) The influence of extracellular side Ca2+
on activity of the plasma membrane H+-ATPase from wheat roots.Aust. J. Plant Physiol. 25: 923–928.
Rhodes, J.D., Thain, J. and Wildon, D.C. (1996) The pathway for sys-temic electrical signal conduction in the wounded tomato plant.Planta 200: 50–57.
Rhodes, J.D., Thain, J. and Wildon, D.C. (1999) Evidence for physicallydistinct systemic signaling pathways in the wounded tomato plant.Ann. Bot. 84: 109–116.
Ricca, U. (1916) Soluzione d’un problema di fisiologia: la propagazionedi stimulo nella Mimosa. Nuovo G. Bot. Ital. 23: 51–170.
Rousset, M., de Roo, M., Guennecb, J.Y.L. and Pichon, O. (2002)Electrophysiological characterization of tomato hypocotyl putative
1518 Plant Cell Physiol. 55(8): 1511–1519 (2014) doi:10.1093/pcp/pcu082 ! The Author 2014.
L. Katicheva et al.
at FMR
P/USP/B
IBL
IOT
EC
A C
EN
TR
AL
on June 9, 2016http://pcp.oxfordjournals.org/
Dow
nloaded from

action potentials induced by cotyledon heating. Physiol. Plant. 115:197–203.
Samejima, M. and Sibaoka, T. (1982) Membrane potentials and resist-ances of excitable cells in the petiole and main pulvinus of Mimosapudica. Plant Cell Physiol. 23: 459–465.
Shimmen, T. (2002) Electrical perception of “Death Message” in chara:analysis of rapid component and ionic process. Plant Cell Physiol. 43:1575–1584.
Stahlberg, R., Cleland, R.E. and van Volkenburgh, E. (2006) Slow wavepotentials—a propagating electrical signal unique to higher plants.In Communication in Plants. Neuronal Aspects of Plant Life. Editedby Baluska, F., Mancuso, S. and Volkmann, D. pp. 291–309. Springer-Verlag, Berlin.
Stahlberg, R. and Cosgrove, D.J. (1992) Rapid alteration in growth rateand electric potentials upon stem excision in pea seedlings. Planta187: 523–531.
Stahlberg, R. and Cosgrove, D.J. (1996) Induction and ionic basis ofslow wave potentials in seedlings of Pisum sativum L. Planta 200:416–425.
Stahlberg, R. and Cosgrove, D.J. (1997) The propagation of slow wavepotentials in pea epicotyls. Plant Physiol. 113: 209–217.
Sukhov, V., Akinchits, E., Katicheva, L. and Vodeneev, V. (2013)Simulation of variation potential in higher plant cells. J. Membr.Biol. 246: 287–296.
Sukhov, V., Orlova, L., Mysyagin, S., Sinitsina, J. and Vodeneev, V. (2012)Analysis of the photosynthetic response induced by variation po-tential in geranium. Planta 235: 703–712.
Tavares, B., Domingos, P., Nuno Dias, P., Feijo, J.A. and Bicho, A. (2011)The essential role of anionic transport in plant cells: the pollen tubeas a case study. J. Exp. Bot. 62: 2273–2298.
Trebacz, K., Dziubinska, H. and Krol, E. (2006) Electrical signals in long-distance communication in plants. In Communication in Plants.Neuronal Aspects of Plant Life. Edited by Baluska, F., Mancuso, S.and Volkmann, D. pp. 277–290. Springer, Berlin.
Tyerman, S.D. (1992) Anion channels in plants. Annu. Rev. PlantPhysiol. Plant Mol. Biol. 43: 351–373.
Vodeneev, V.A., Akinchits, E.K., Orlova, L.A. and Sukhov, V.S. (2011)The role of Ca2+, H+, and Cl– ions in generation of variation po-tential in pumpkin plants. Russ. J. Plant Physiol. 58: 974–981.
Vodeneev, V.A., Opritov, V.A. and Pyatygin, S.S. (2006) Reversiblechanges of extracellular pH during action potential generation ina higher plant Cucurbita pepo. Russ. J. Plant Physiol. 53: 481–487.
Vodeneev, V., Orlova, A., Morozova, E., Orlova, L., Akinchits, E.,Orlova, O. et al. (2012) The mechanism of propagation of variationpotentials in wheat leaves. J. Plant Physiol. 169: 949–954.
Zimmermann, M.R. and Felle, H.H. (2009) Dissection of heat inducedsystemic signals: superiority of ion fluxes to voltage changes insubstomatal cavities. Planta 229: 539–547.
1519Plant Cell Physiol. 55(8): 1511–1519 (2014) doi:10.1093/pcp/pcu082 ! The Author 2014.
Ionic nature of variation potential
at FMR
P/USP/B
IBL
IOT
EC
A C
EN
TR
AL
on June 9, 2016http://pcp.oxfordjournals.org/
Dow
nloaded from


















