
Interaction DNA ox/,3-Type Small, Acid-Soluble Spore ... · NEWCLASS OF DNA-BINDING PROTEINS 2313...
11
Vol. 174, No. 7 JOURNAL OF BACTERIOLOGY, Apr. 1992, p. 2312-2322 0021-9193/92/072312-11$02.00/0 Copyright X 1992, American Society for Microbiology Interaction between DNA and ox/,3-Type Small, Acid-Soluble Spore Proteins: a New Class of DNA-Binding Protein BARBARA SETLOW, DONGXU SUN, AND PETER SETLOW* Department of Biochemistry, University of Connecticut Health Center, Farmington, Connecticut 06030 Received 15 October 1991/Accepted 16 January 1992 DNA in spores of BaciUlus and Clostridium species is associated with small, acid-soluble proteins (SASP) of the cz/13 type; the presence of these proteins is a major factor in causing spore resistance to UV light. cV/,3-type SASP did not bind to single-stranded DNA, single- or double-stranded RNA, or DNA-RNA hybrids in vitro. However, these proteins bound a variety of double-stranded DNAs and conferred protection against DNase cleavage. The binding of at/J-type SASP to DNA saturated at a protein/DNA ratio (wt/wt) of 4:1 to 5:1, which is -1 SASP per 4 bp. a/3-type SASP-DNA interaction did not require divalent cations, was independent of pH between 6 and 8, and, for some SASP-DNA pairs, was relatively insensitive to salt up to 0.3 M. The relative affinity of a/,8-type SASP for different DNAs was poly(dG) poly(dC) > poly(dG-dC) poly(dG-dC) > plasmid pUC19 > poly(dA-dT) poly(dA-dT), with poly(dA) poly(dT) giving no detectable binding. This order in a/18-type SASP-DNA affinities parallels the facility with which the DNAs adopt an A-like conformation, the conformation in ao1$-type SASP-DNA complexes. An oligo(dG) oligo(dC) of 12 bp was bound by a/p-type SASP. While a 26-bp oligo(dG) oligo(dC) bound more tightly than the 12-mer, there was no significant increase in affinity for or/n-type SASP with further increase in size of oligo(dG) oligo(dC). In contrast, binding of ot/j-type SASP to oligo(dA-dT) oligo(dA-dT) was minimal up to at least a 70-mer, and binding to poly(dA-dT) poly(dA-dT) was very cooperative. In addition to blocking DNase digestion, binding of CV/I3-type SASP to DNA blocked (i) cleavage of the DNA backbone by hydroxyl radicals and orthophenanthroline-Cu21, (ii) DNA cleavage by restriction enzymes, in particular those with specificity for GC-rich sequences; and (iii) in vitro transcription of some but not all genes. However, methylation of dG residues by dimethyl sulfate was not affected by cv/13-type SASP binding. Approximately 5 to 10% of the protein in dormant spores of Bacillus and Clostridium species is a group of small, acid-soluble proteins (SASP) of the a/P type, which are coded for by multiple genes in all species examined (5, 11, 35). The amino acid sequences of a/e-type SASP are very highly conserved both within and across species (5, 11, 35). a/p-type SASP are synthesized at about the third hour of sporulation only in the forespore compartment (37), and their synthesis parallels the acquisition of UV light resis- tance by the developing spore as well as a 50% increase in the number of negative supertwists in forespore plasmids (23, 36). During spore germination, a/o3-type SASP are rapidly degraded to amino acids, with degradation initiated by a protease termed GPR, which is specific for SASP (35, 41). Over the past few years, much evidence has accumulated which indicates that oa/P-type SASP are DNA-binding pro- teins and that their binding to DNA in vivo causes spore UV resistance by altering the spore DNA's UV photochemistry. In both sporulating Bacillus subtilis and Escherichia coli, in which a/P-SASP synthesis is artificially induced, a/0-type SASP are associated only with the nucleoid (7, 30, 32). B. subtilis spores lacking ao/a-type SASP (termed at--) are UV sensitive, and thymine dimers (TT) are formed in DNA of at - spores upon UV irradiation. In contrast, no TT is produced in wild-type spores, but a different photoproduct, termed spore photoproduct, is formed (12, 33). The in- creased negative superhelicity of spore plasmid DNA noted above is also greatly reduced in ao-V- spores (23). These * Corresponding author. altered properties of ao-- spores can be restored to those of wild-type spores by sufficient levels of any a/3-type SASP (13, 42). However, mutant a/L-type SASP with alterations in residues highly conserved throughout evolution are ineffec- tive in this regard (42). The binding of ot/i-type SASP to DNA in vitro has been shown by a variety of procedures (19, 21, 22). This binding results in a structural change in the DNA from the B conformation to an A-like conformation (19), as well as an increase in the apparent negative superhelicity of covalently closed plasmids (21). There is also a change in the DNA's UV photochemistry upon o/Pi-type SASP binding in vitro, going from production of 1T in DNA alone to spore photo- product in SASP-DNA complexes (22). It has been noted that the changes in DNA properties brought about by x/3-type SASP binding are different from those caused by any other DNA-binding protein (19, 21, 22). Indeed, the amino acid sequences of o/IP-type SASP do not match the consensus motif for any DNA-binding protein (or any other protein, for that matter) in currently available data bases. Because it appears that ot/a-type SASP may represent a new class of DNA-binding protein, it seemed of significant value to study oa/n-type SASP-DNA binding in vitro in some detail-in particular, with respect to the specificity for the DNA moiety and the effects on various DNA properties. In this communication, we report the relative affinity of a number of a/n-type SASP for different nucleic acids as well as some characteristics of SASP-DNA binding. We also report the effect of o/t3-type SASP-DNA binding on the DNA's cleavage or modification by chemicals or enzymes and its transcriptional activity. 2312 on June 27, 2020 by guest http://jb.asm.org/ Downloaded from
Transcript of Interaction DNA ox/,3-Type Small, Acid-Soluble Spore ... · NEWCLASS OF DNA-BINDING PROTEINS 2313...

Vol. 174, No. 7JOURNAL OF BACTERIOLOGY, Apr. 1992, p. 2312-23220021-9193/92/072312-11$02.00/0Copyright X 1992, American Society for Microbiology
Interaction between DNA and ox/,3-Type Small, Acid-Soluble SporeProteins: a New Class of DNA-Binding Protein
BARBARA SETLOW, DONGXU SUN, AND PETER SETLOW*Department ofBiochemistry, University of Connecticut Health Center,
Farmington, Connecticut 06030
Received 15 October 1991/Accepted 16 January 1992
DNA in spores of BaciUlus and Clostridium species is associated with small, acid-soluble proteins (SASP) ofthe cz/13 type; the presence of these proteins is a major factor in causing spore resistance to UV light. cV/,3-typeSASP did not bind to single-stranded DNA, single- or double-stranded RNA, or DNA-RNA hybrids in vitro.However, these proteins bound a variety of double-stranded DNAs and conferred protection against DNasecleavage. The binding of at/J-type SASP to DNA saturated at a protein/DNA ratio (wt/wt) of 4:1 to 5:1, whichis -1 SASP per 4 bp. a/3-type SASP-DNA interaction did not require divalent cations, was independent ofpHbetween 6 and 8, and, for some SASP-DNA pairs, was relatively insensitive to salt up to 0.3 M. The relativeaffinity of a/,8-type SASP for different DNAs was poly(dG) poly(dC) > poly(dG-dC) poly(dG-dC) > plasmidpUC19 > poly(dA-dT) poly(dA-dT), with poly(dA) poly(dT) giving no detectable binding. This order ina/18-type SASP-DNA affinities parallels the facility with which the DNAs adopt an A-like conformation, theconformation in ao1$-type SASP-DNA complexes. An oligo(dG) oligo(dC) of 12 bp was bound by a/p-typeSASP. While a 26-bp oligo(dG) oligo(dC) bound more tightly than the 12-mer, there was no significantincrease in affinity for or/n-type SASP with further increase in size of oligo(dG) oligo(dC). In contrast, bindingof ot/j-type SASP to oligo(dA-dT) oligo(dA-dT) was minimal up to at least a 70-mer, and binding topoly(dA-dT) poly(dA-dT) was very cooperative. In addition to blocking DNase digestion, binding of CV/I3-typeSASP to DNA blocked (i) cleavage of the DNA backbone by hydroxyl radicals and orthophenanthroline-Cu21,(ii) DNA cleavage by restriction enzymes, in particular those with specificity for GC-rich sequences; and (iii)in vitro transcription of some but not all genes. However, methylation of dG residues by dimethyl sulfate wasnot affected by cv/13-type SASP binding.
Approximately 5 to 10% of the protein in dormant sporesof Bacillus and Clostridium species is a group of small,acid-soluble proteins (SASP) of the a/P type, which arecoded for by multiple genes in all species examined (5, 11,35). The amino acid sequences of a/e-type SASP are veryhighly conserved both within and across species (5, 11, 35).a/p-type SASP are synthesized at about the third hour ofsporulation only in the forespore compartment (37), andtheir synthesis parallels the acquisition of UV light resis-tance by the developing spore as well as a 50% increase inthe number of negative supertwists in forespore plasmids(23, 36). During spore germination, a/o3-type SASP arerapidly degraded to amino acids, with degradation initiatedby a protease termed GPR, which is specific for SASP (35,41).Over the past few years, much evidence has accumulated
which indicates that oa/P-type SASP are DNA-binding pro-teins and that their binding to DNA in vivo causes spore UVresistance by altering the spore DNA's UV photochemistry.In both sporulating Bacillus subtilis and Escherichia coli, inwhich a/P-SASP synthesis is artificially induced, a/0-typeSASP are associated only with the nucleoid (7, 30, 32). B.subtilis spores lacking ao/a-type SASP (termed at--) are UVsensitive, and thymine dimers (TT) are formed in DNA ofat - spores upon UV irradiation. In contrast, no TT isproduced in wild-type spores, but a different photoproduct,termed spore photoproduct, is formed (12, 33). The in-creased negative superhelicity of spore plasmid DNA notedabove is also greatly reduced in ao-V- spores (23). These
* Corresponding author.
altered properties of ao-- spores can be restored to those ofwild-type spores by sufficient levels of any a/3-type SASP(13, 42). However, mutant a/L-type SASP with alterations inresidues highly conserved throughout evolution are ineffec-tive in this regard (42).The binding of ot/i-type SASP to DNA in vitro has been
shown by a variety of procedures (19, 21, 22). This bindingresults in a structural change in the DNA from the Bconformation to an A-like conformation (19), as well as anincrease in the apparent negative superhelicity of covalentlyclosed plasmids (21). There is also a change in the DNA'sUV photochemistry upon o/Pi-type SASP binding in vitro,going from production of 1T in DNA alone to spore photo-product in SASP-DNA complexes (22). It has been notedthat the changes in DNA properties brought about byx/3-type SASP binding are different from those caused byany other DNA-binding protein (19, 21, 22). Indeed, theamino acid sequences of o/IP-type SASP do not match theconsensus motif for any DNA-binding protein (or any otherprotein, for that matter) in currently available data bases.Because it appears that ot/a-type SASP may represent a newclass of DNA-binding protein, it seemed of significant valueto study oa/n-type SASP-DNA binding in vitro in somedetail-in particular, with respect to the specificity for theDNA moiety and the effects on various DNA properties. Inthis communication, we report the relative affinity of anumber of a/n-type SASP for different nucleic acids as wellas some characteristics of SASP-DNA binding. We alsoreport the effect of o/t3-type SASP-DNA binding on theDNA's cleavage or modification by chemicals or enzymesand its transcriptional activity.
2312
on June 27, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

NEW CLASS OF DNA-BINDING PROTEINS 2313
MATERIALS AND METHODS
Proteins and nucleic acids. The a/,-type SASP SspCwtfrom B. subtilis, SASP-A from Bacillus megaterium, SASP-Ifrom Bacillus cereus, and SASP-a and -P from Clostridiumbifennentans as well as the -y-type SASP SASP-B from B.megatenium were purified to homogeneity as describedpreviously (4, 21, 34, 44). Two mutant derivatives of SspCwt,SspCAla and SspCTYr, were also purified as described previ-ously (42). SSpCMa has an alanine residue replacing glycineat position 52 in SspCwt; this residue is a glycine in alla/13-type SASP which have been analyzed, and SspCAla hasno detectable DNA-binding activity in vivo or in vitro (5, 11,35, 42). SspCTYr has a tyrosine residue replacing leucine atposition 29 in SspCwt; this residue is a tyrosine in a numberof other oa/1-type SASP, and the DNA-binding activity ofSsPCTYr is identical to that of SspCwt in vivo and in vitro(42). Consequently, we have used SspCTYr and SspCwtinterchangeably in this work, and these will be referred to inthe text jointly as SspC.RNA polymerase containing predominantly or (Eor) was
purified from vegetative cells of B. subtilis 168 through theheparin-agarose column step (24). RNA polymerase contain-ing a' (EcrG) was purified similarly from vegetative cells ofB. subtilis PS766 [trpC2 (pDG298 spac-spoIIIG Kmr)], inwhich a'0 synthesis had been induced with isopropyl-p-D-thiogalactopyranoside (24, 40). DNases were purchasedfrom Sigma Chemical Corporation, and restriction enzymes
were from New England BioLabs.All synthetic RNA and DNA copolymers, homopolymers,
or homopolymer pairs and p(T)g were purchased from Phar-macia or Sigma Chemical Corporation. Note that the syn-
thetic polynucleotides are polydisperse. Other oligodeoxyri-bonucleotides were synthesized on an Applied Biosystemssynthesizer; after cleavage from the column and deblocking,they were used without further purification. The oligodeoxy-nucleotides made were 5'-AT(G)12, 5'-AT(C)12, and 5'-GC(AT)6-3'. Annealing of all oligo- or polynucleotides togenerate double-stranded structures was routinely carriedout at -50 ,ug of total nucleic acid (equimolar in nucleotidefrom complementary nucleic acids) per ml in 25 mM Tris-HCl (pH 7.4)-i mM EDTA-0.1 M NaCl. Samples were
heated at -95°C for 10 min and then incubated for 15 mineach at 70, 50, and 30°C before use. Double-stranded oligo-deoxyribonucleotides were treated with polynucleotide ki-nase and ATP before use; aliquots were treated with phageT4 DNA ligase at 16°C for 4 h to generate multimers, whichwere purified by phenol extraction and ethanol precipitation.
Plasmid pBSM was obtained from P. Stabach and S.Padula (Department of Medicine, University of ConnecticutHealth Center). This plasmid is a derivative of plasmid pBS(17) carrying a 510-bp EcoRI fragment derived from a cDNAencompassing the 3' end, including the poly(A) tail, of an
mRNA for a murine T-cell receptor P chain. The EcoRIinsert contains a run of 58 A residues and then a G adjacentto one EcoRI site. The plasmid was cleaved with EcoRI, andthe 510-bp insert was isolated, end labeled with [32P]dATPand reverse transcriptase, and recut with PvuII, which givesa 270-bp fragment containing the poly(dA) region, and thelabeled 270-bp fragment was isolated. Plasmid pUC19 was
isolated from E. coli JM83 and purified by two CsCl densitygradient centrifugations. Restriction enzyme fragments ofpUCi9 were isolated by gel electrophoresis and labeled withreverse transcriptase and an appropriate aL-32P-labeled de-oxynucleoside triphosphate; in some cases, the labeledfragment was cut with a second restriction enzyme, and a
labeled subfragment was isolated. [methyl-3H]thymidine-labeled pUC19 was prepared and purified as describedpreviously (22). Plasmid pUB110 was isolated from B. sub-tilis 168 (23) and purified as described above. Synthesis oflabeled polydeoxynucleotides was routinely carried out atpH 7.3 in 200 ,ul of 40 mM Tris-HCl-160 mM HEPES(N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid)-4mM MgCl2-1 mM dithiothreitol with -2 ,ug of unlabeledpolymer, except for synthesis of poly(dA) poly(3H-dT), forwhich 5 ,ug of poly(dA) plus 0.25 ,ug of p(T)g was used.Nucleotide additions for labeling various polymers were:poly(dA). poly(dT), [methyl-3H]TTP (50 ,uCi) was used at10 ,uM; poly(dA-dT). poly(dA-dT), dATP was used at 50,uM and [methyl-3HflTP (50 ,uCi) was used at 10 FiM;poly(dG-dC) poly(dG-dC), [5-3H]dCTP (50 ,uCi) was usedat 10 ,uM and dGTP was used at 50 ,uM; and poly(dG).poly(dC), [5-3H]dCTP (50 ,uCi) was used at 10 ,uM. Labelingwas initiated by addition of the large fragment of E. coliDNA polymerase (10 U), and the mixture was incubated at37°C for 1 to 2 h until incorporation had leveled off. Gener-ally, 15 to 60% of the input label was incorporated intoacid-insoluble material. The solution was then diluted 1:1with 0.6 M NaCl, 20 to 50 ,ug of unlabeled polymer wasadded, and DNA was precipitated with 2 volumes of ethanol.The pellet was rinsed with 1 ml of 70% ethanol, air dried, anddissolved in 100 to 400 [lI of 10 mM Tris-acetate (pH 7.0).Assay of SASP-nucleic acid binding. SASP-nucleic acid
binding was assessed by measuring the ability of a SASP toprotect a nucleic acid from nuclease digestion. Four differentprocedures were used to monitor protein-nucleic acid inter-action; two measured the interaction of a SASP with onenucleic acid (direct assays), while the other two measuredthe ability of two different nucleic acids to compete for oneSASP (competitive assays). In the direct electrophoreticassay, nucleic acid (3 ,ug unless otherwise stated) wasincubated in 25 ,u of 10 mM Tris-acetate (pH 7.0)-i mMEDTA with various amounts of SASP. After 1 h at 37°C, 2.5Il' of MgCl2 (30 mM) was added, followed by 2.5 ,ul of DNaseI (1 mg/ml). For analysis of RNAs in this assay, only 2.5 ,ulof pancreatic RNase (1 mg/ml) was added. The solution wasincubated a further 15 min at 37°C, and then 100 ,ul of 1.25%sodium dodecyl sulfate (SDS)-25 mM EDTA was added,followed by 8 ,ul of 5 M NaCl and 280 ,ul of ethanol toprecipitate the DNA. The precipitated DNA was rinsed with300 ,ul of 70% ethanol, air dried, and dissolved in 10 [L ofwater prior to analysis by agarose gel (polynucleotides) oracrylamide gel (oligonucleotides) electrophoresis. In a fewexperiments in which we wished to minimize the time forDNase treatment, DNase I at 15 mg/ml was used as de-scribed above, and the subsequent incubation was restrictedto 1 min. In the competitive electrophoretic assay, each25-pul incubation mix described above contained 3 ,ug ofEcoRI-linearized pUC19 as well as various amounts of othernucleic acids and oa/,-type SASP. After processing as de-scribed above, the precipitated nucleic acid was analyzed byagarose gel electrophoresis. This assay was most useful inidentifying nucleic acids which bound a/a-type SASP eithermuch more strongly than pUC19 or much more weakly (ornot at all). In the direct acid precipitation assay, 25-pAincubation mixes as described above for the direct electro-phoretic assay contained 3 or 5 jig of a 3H-labeled DNA(-104 cpm) as well as various amounts of SASP. Afterincubation and DNase treatment as described above, 100 pulof 1 N NaOH was added, and the solution was incubated for10 min at room temperature to dissociate protein-DNAcomplexes. One milliliter of 7% perchloric acid was added to
VOL. 174, 1992
on June 27, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

2314 SETLOW ET AL.
precipitate DNA, and after 5 min at room temperature, theprecipitate was collected on a Whatman GFC filter, the filterwas washed four times with 1 ml of 7% perchloric acid andthen with ethanol, and finally the filter was dried andradioactivity was counted in a toluene-based scintillationfluid. In the competitive acid precipitation assay, 25-,uincubation mixes as described above contained 3 or 5 ,g ofa 3H-labeled DNA (-104 cpm), a/n-type SASP at a 4:1weight/weight ratio with the labeled DNA, and variousamounts of an unlabeled nucleic acid. Samples were proc-essed and counted as described above for the direct acidprecipitation assay.
In the competitive acid precipitation assay, <10% reduc-tion in acid-precipitable counts by an unlabeled competitornucleic acid was scored as no binding of the competitor. Inthe competitive electrophoretic assay, no observable reduc-tion in the size of the DNase-protected fragments uponcompetitor addition was scored as no binding of the compet-itor (see Fig. 4). In the two direct assays, no binding wasdefined as either no nuclease-resistant fragments larger thanin a no-SASP control (direct electrophoretic assay) or <3%of input counts remaining acid precipitable at a protein/DNA(wt/wt) ratio of 10:1 (direct acid precipitation assay). Itshould be emphasized that the DNase protection given DNAby a/,B-type SASP is due to its binding to the DNA, as theproteins alone do not inhibit DNase (42).
In vitro transcription. In vitro transcription reactions withEoA or Ec0 and various templates were carried out with[ac-32P]UTP without heparin, and transcripts were separatedby acrylamide gel electrophoresis under denaturing condi-tions and detected by autoradiography (24, 39, 40). In mostcases, a/,3-type SASP were incubated with templates inreaction buffer at 37°C for 30 min prior to addition of RNApolymerase to initiate transcription. In one experiment,a/P-type SASP was added just after RNA polymerase.Templates used for in vitro transcription were: B. subtilisans, pPS1521 cut with EcoRI (this gives a 121-nucleotide [nt]transcript with EoA [39]); B. subtilis gerA, pPS964 cut withXbaI (this gives a 114-nt transcript with Eo0 [6]); B. subtilissspA, pPS706 cut with XbaI (this gives a 139-nt transcriptwith EoG [24]); B. subtilis sspB, pPS706 cut with PstI (thisgives a 230-nt transcript with EorG [24]); B. subtilis sspE,pPS591 cut with either EcoRI or PvuII (these give either a194-nt transcript [EcoRI] or an 82-nt transcript [PvuII] withEuG [24]); B. subtilis veg, pMS109 cut with BamHI (thisgives a 105-nt transcript with ErA [20]).Other procedures. Cleavage of DNA with hydroxyl radi-
cals or by orthophenanthroline-Cu2" was carried out asdescribed previously (38, 43). The DNAs used in theseexperiments were restriction fragments of pUC19 whichwere labeled with an appropriate a-32P-labeled deoxynucle-otide triphosphate and reverse transcriptase. Methylation ofG residues in 32P-end-labeled DNA fragments prepared asdescribed above was carried out with and without SspC asdescribed by Maxam and Gilbert (14). The methylatedDNAs were then cleaved by procedures giving strand break-age only at G residues or at both A and G residues (G > A)(14). The cleaved fragments were analyzed by acrylamide gelelectrophoresis under denaturing conditions, and fragmentswere detected by autoradiography.
ot/3-type SASP were preincubated with the restrictionenzyme template (1 ,ug of pUC19 linearized with eitherEcoRI or SspI) for 1 h at 37°C in 25 RI of the buffer specifiedfor the restriction enzyme to be tested. Digestion was carriedout for 30 min at 37°C with 2 to 10 U of enzyme, and digestswere analyzed by electrophoresis on 2% agarose gels. The
A4
B_2.7 _
_1.3 _.
_0.6 4
c3 4 5
_4.5
_2.3
_ 1.3
-0.6
FIG. 1. Protection of pUC19 and pUB110 against nuclease diges-tion by SspC binding. SspClyr was incubated with pUC19 (A and B)or pUB110 (C), digested with various nucleases, and analyzed byelectrophoresis on a 2% agarose gel as described for the directelectrophoretic assay. (A) Samples were digested with DNase I andhad been incubated with SspClYr/pUC19 ratios (wt/wt) of 0:1 (lane1), 0.4:1 (lane 2), 1:1 (lane 3), 2:1 (lane 4), 4:1 (lane 5), or 10:1 (lane6). (B) Samples were incubated with SspCTYr/pUC19 ratios of 1:1(lanes 1 and 4), 2:1 (lanes 2 and 5), or 4:1 (lanes 3 and 6) and thendigested with either DNase I (lanes 1 to 3) or S. aureus nuclease(lanes 4 to 6). (C) Samples were digested with DNase I and had beenincubated with SspC "'/pUB110 ratios of 0:1 (lanes 1 and 2), 0.4:1(lane 3), 1:1 (lane 4), 2:1 (lane 5), 4:1 (lane 6), or 7:1 (lane 7). Thesample in lane 1 was not treated with DNase. The numberedhorizontal arrowheads denote the migration positions of DNA sizemarkers (in kilobases).
extent of inhibition of restriction enzyme digestion wasestimated by inspection of these gels. However, this willunderestimate the degree of inhibition in highly inhibitedsamples, as an excess of some restriction enzymes was used.
RESULTS
Protection of plasmid DNAs against DNase. Incubation ofincreasing amounts of SspC with plasmid pUB110 or pUC19resulted in protection of the plasmid against pancreaticDNase I, as discrete DNA fragments remained after DNasetreatment (Fig. 1A and C). Use of DNase from Staphylococ-cus aureus instead of DNase I gave a similar pattern ofprotected fragments from SspC-pUC19 complexes (Fig. 1B).DNase protection of these plasmid DNAs was maximal at aprotein/DNA ratio (wt/wt) of 4:1 to 5:1 (Fig. 1), which issimilar to the value obtained for the maximum binding ofSspC (SspCwt or SSpCTYr) to DNA by other methods (21, 22)
J. BACTERIOL.
on June 27, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

NEW CLASS OF DNA-BINDING PROTEINS 2315
AA-) 4 5- 6
411.3
m0.9
40.6
40.2
B1 2 3 4 5 6 7 8
4 2.7 _
< 1.3 _
40.6 _
FIG. 2. DNase digestion of SspC-pUC19 complexes as a func-tion of time. Plasmid pUC19 (35 ,ug) was preincubated in 175 ,ul of 10mM Tris-acetate (pH 7.0)-i mM EDTA with (A) 140 ,ug of SspClyror (B) 35 ,ug of SspCTYr. After 2 h at 37°C, 2 mM MgCl2 and 0.1 mgof DNase I per ml were added. At various times after DNaseaddition, aliquots (25 ,ul) were added to 100 p.l of 1.25% SDS-25 mMEDTA, and DNA was isolated and analyzed by electrophoresis on a2% agarose gel. The different lanes are samples isolated afterdifferent times of DNase digestion and are: lane 1, before DNaseaddition; lane 2, 2 min; lane 3, 5 min; lane 4, 15 min; lane 5, 45 min;and lane 6, 120 min. Control experiments (not shown) demonstratedthat the DNase digestion conditions used left no fragments largerthan 100 bp after 1 min of digestion. The numbered horizontalarrowheads denote the migration positions of DNA size markers (inkilobases).
(see below). The presence of discrete DNA bands afterDNase treatment of SspC-plasmid complexes suggests thatthere is some specificity in this protein's binding to DNA.However, all regions of these plasmids can be protectedfrom DNase by SspC binding, as significant amounts ofintact pUB110 and pUC19 remained in digests of saturatedSspC-plasmid complexes under conditions in which intactplasmid was degraded completely (Fig. 1). The sizes ofDNase-resistant bands increased significantly as the SspC/DNA ratio increased, suggesting that the sizes of DNAregions completely covered by SspC increase concurrently(Fig. 1A and B; see below). One possible reason for this maybe that at a high SspC/DNA ratio, the protein binds toregions of DNA between areas to which the protein binds atlow SspC/DNA ratios. When DNase digestion of SspC-pUC19 complexes was followed over time under conditionsin which free pUC19 was digested completely in <1 min,full-length DNase-resistant pUC19 in the complex formedwith a 4:1 (wt/wt) ratio disappeared quite rapidly, whileDNase-protected bands from complexes formed with a 1:1ratio of SspC to pUC19 were more resistant to furtherdigestion (Fig. 2). This suggests that at low SspC levels, theDNA bound has a greater affinity for the protein than doessome of the DNA bound at higher SspC levels. Again, thesedata suggest that SspC has significant specificity in itsbinding to DNA.
Analysis of the DNase protection of pUC19 by four otherot/f-type SASP indicated that these proteins also conferDNase resistance to this plasmid (Fig. 3). The two a/a-typeSASP from Bacillus species gave patterns of protected bandssimilar to those given by SspC, although the patterns withthe two clostridial proteins were different (Fig. 3). Thebinding of these other a11-type SASP to pUC19 also satu-rated at a protein/DNA ratio (wt/wt) similar to that found forSspC (i.e., 4:1 to 5:1), with the exception of SASP-A from B.megaterium, which required a higher ratio for good DNase
C
2 3 4 5 6
-1.3
00.6
FIG. 3. DNase protection of pUC19 by different o/n-type SASP.Different a/03-type SASP were tested for DNase protection ofpUC19 in the direct electrophoretic assay on a 2% agarose gel. (A)Lanes 1 to 3, SspCwt/pUC19 ratios of 2:1 (lane 1), 3:1 (lane 2), and6:1 (lane 3); lanes 4 to 7, B. cereus SASP-I/pUC19 ratios of 1:1 (lane4), 2:1 (lane 5), 3:1 (lane 6), and 6:1 (lane 7). (B) Lanes 1 to 4, Cbifermentans SASP-aIpUC19 ratios of 1:1 (lane 1), 2:1 (lane 2), 3:1(lane 3), and 6:1 (lane 4); lanes 5 to 8, C bifermentans SASP-P/pUC19 ratios of 1:1 (lane 5), 2:1 (lane 6), 3:1 (lane 7), and 6:1 (lane8). (C) Lanes 1 to 3, SspCTYr/pUC19 ratios of 1:1 (lane 1), 2.5:1 (lane2), and 5:1 (lane 3); lanes 4 to 6, B. megaterium SASP-A/pUC19ratios of 2:1 (lane 4), 5:1 (lane 5), and 12:1 (lane 6). The numberedhorizontal arrowheads denote the migration positions of DNA sizemarkers (in kilobases).
protection (Fig. 3C). This appears to be because B. megate-num SASP-A binds more weakly to pUC19 than does SspC.When these latter two proteins were tested with a moretightly bound nucleic acid [poly(dG) poly(dC); see below],both had similar protein/DNA ratios at saturation. Becausethe DNA-binding specificity of SspC appeared to be similarto that of other ca/p-type SASP from Bacillus species (Fig. 3,and see below), and because SspCs can be produced in andpurified from E. coli in high yield (21), most other work on
a/4-type SASP-DNA binding was done with SspC (SspCwtor SspCTYr). These two SspC derivatives were used inter-changeably, because previous work has shown that theyhave identical DNA-binding specificities (42). In contrast tothe DNase protection given pUC19 upon binding to SspCand other a/p-type SASP, addition of a large amount (10:1,wt/wt) of a -y-type SASP (SASP-B ofB. megaterium) gave nodetectable DNase protection in the two direct assays (datanot shown). This is not surprising, because -y-type SASPhave a different conserved primary sequence than ot/p3-typeSASP and are not preferentially associated with DNA invivo (7, 35). Previous work has shown that the SspC variantsSspCMO and SspCGIn, in which amino acid residues con-served throughout evolution have been altered, also do notbind to pUC19 (42).
1.3 o
0.9 o
0.61
0.3-
VOL. 174, 1992
on June 27, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

2316 SETLOW ET AL.
TABLE 1. Binding of SspC to synthetic polynucleotides
SspC Polynucleotidesabound A B C D
No Poly(A), poly(C), poly(G), poly(U), Poly(A) * poly(U), poly(G) poly Poly(dI) poly(dC), poly(dI-dC). Poly(dA) poly(dT)poly(dA), poly(dC), poly(dT) (C), poly(I) * poly(C) poly(dI-dC), poly(G) poly(dC)
Yesb Poly(dA-dT) * poly(dA-dT), poly(dG-dC) poly(dG-dC), poly(dG). poly(dC)
a The binding assays used are indicated as follows: A, competitive acid precipitation assay with [3H]pUC19 and competitive electrophoretic assay (for anexample of the latter, see Fig. 4A); B, competitive acid precipitation assay with [3H]pUC19 and direct electrophoretic assay for RNase protection (an exampleof the latter is given in Fig. 4C); C, competitive acid precipitation assay with both poly(dA-[3H]dT) poly(dA-[3H]dT) and [3HjpUC19 and the direct andcompetitive electrophoretic assays; D, competitive acid precipitation assay with both poly(dA-[3H]dT) poly(dA-[3H]dT) and [3H]pUC19, competitiveelectrophoretic assay (see Fig. 4A), direct electrophoretic assay (see Fig. 4B), and direct acid precipitation assay. For examples of the polynucleotides to whichSspC bound, see Fig. 5.
b Note that a number of mixed-sequence nucleic acids (i.e., plasmids and chromosomal DNAs) bind SspC (19, 21).
Characteristics of cd,1-type SASP-DNA binding. The inter-action between SspC and pUC19 which resulted in DNaseprotection was not instantaneous, taking -40 and 10 min at4 and 37°C, respectively, to reach maximum binding in 10mM Tris-acetate (pH 7.0)-i mM EDTA with 80 ,ug ofEcoRI-cut pUC19 and 240 ,ug of SspCTYr per ml (data notshown). A slow step in the binding of SspC to DNA has beenseen with several other analytical procedures (21). While theevent responsible for the slow step in SspC-DNA interactionis not clear, it could be some part of the change in DNA fromthe B form to an A-like structure which is induced by SspCbinding (19, 21, 22). Not only was SspC-pUC19 complexformation more rapid at 37 than at 4°C, but more complexwas formed at the higher temperature, as larger DNase-resistant fragments were obtained (data not shown). How-ever, at both temperatures, the overall pattern of DNase-resistant bands from SspC-pUC19 complexes was similar(data not shown).As shown by analysis of the pattern of DNase-resistant
bands in 10 mM Tris-acetate (pH .7.0)-i mM EDTA, thebinding of SspC (SspCTYr, 720 ,ug/ml) to pUC19 (EcoRI cut,120 ,ug/ml) was almost unaffected by salt (potassium acetate)up to -50 mM (data not shown). As the salt concentrationwas increased, larger protected DNA bands disappeared andsmaller ones became more prominent; however, SspC pro-vided significant (>20%) DNase resistance to pUC19 at 300mM salt (data not shown). In contrast to the relative resis-tance of SspC-pUC19 binding to 50 mM salt, the binding ofB. megaterium SASP-A to pUC19 under similar conditionswas almost completely abolished (>95%) at 50 mM salt (datanot shown). This finding is consistent with the weakerbinding of B. megaterium SASP-A to pUC19 noted above.The SspC-pUC19 interaction giving DNase protection wasunaffected by the presence or absence of 5 mM divalentcation (Ca2', Mg2+, or Mn2+) and appeared to be similarbetween pH 6 and 8 (data not shown); this result has beenfound previously with a different assay to measure a/a-typeSASP-DNA binding (21).
Specificity of a/'j-type SASP for different nucleic acids. Thepresence of discrete DNA fragments after DNase digestionof a/I-type SASP-pUC19 complexes suggested that theseproteins have some sequence or base specificity in theirbinding to DNA. Consequently, we determined the locationof the ends of some of the predominant DNase-protectedfragments obtained from a saturated SspC-pUC19 complex.In all cases, the ends of these fragments were in AT-richregions, invariably regions with runs of A's or T's (data notshown). While this suggests that a/13-type SASP binding may
be weakest to AT-rich DNA, it was difficult to show thisconclusively by using pUC19. Consequently, we used nucle-ase protection assays to measure the binding of a/3-typeSASP to various synthetic copolymers, homopolymers, andhomopolymer pairs. Nuclease protection of al/,-type SASPbinding was monitored directly by agarose gel electrophore-sis or acid precipitation of 3H-labeled nucleic acid (directassays) or by competition between two different nucleicacids for a limiting amount of SASP (competitive assays).Where tested, the results from all four assays were consis-tent with each other.SspC exhibited no detectable binding to any single-
stranded nucleic acid tested, including both ribo- and deoxy-ribonucleic acids (Table 1, Fig. 4A). Similarly, no bindingwas observed to any double-stranded RNA and the oneRNA. DNA hybrid tested (Table 1, Fig. 4C). In contrast, anumber of double-stranded DNAs tested gave significantbinding, with the exception of dI-containing polymers andpoly(dA) poly(dT) (Table 1, Table 2, Fig. 4A and B, Fig. 5).The SspC/nucleic acid ratio giving maximal DNase protec-tion of poly(dA-dT) poly(dA-dT), poly(dG-dC) poly(dG-dC), and poly(dG) poly(dC) was 4:1 to 5:1 (wt/wt) (Table 2,Fig. 5), as was found above for pUC19. Four other a/3-typeSASP (including B. megaterium SASP-A) also had a 4:1 to5:1 ratio of protein to poly(dG) poly(dC) at saturation (datanot shown). However, a -y-type SASP gave no binding to thispolymer (data not shown). The lack of any detectablebinding of SspC to poly(dA) poly(dT)- was confirmed byanalysis of the nuclease protection afforded to a DNAfragment containing a long run of dA residues (Fig. 6). Whilethe fragment's sequences other than the large oligo(dA)region were generally protected against DNase by SspCbinding, the oligo(dA) region exhibited little if any protection(Fig. 6). Note, however, that there was significant DNaseprotection provided to the end of the oligo(dA) region, aswell as to an adjacent dA3 (Fig. 6, arrows a and b). Thus, atleast short stretches of oligo(dA) in a DNA can bind SspC(see Discussion).Use of the two competitive assay procedures showed that
the affinity of SspC and C. bifennentans SASP-3 for variousDNAs was in the order poly(dG) .poly(dC) > poly(dG-dC).poly(dG-dC) > pUC19 > poly(dA-dT) poly(dA-dT) >>>poly(dA) poly(dT) (Table 1, Table 3; data not shown).Three other oa/3-type SASP tested, B. cereus SASP-I, B.megaterium SASP-A, and C. bifermentans SASP-ac, exhib-ited a similar specificity for double-stranded nucleic acids(data not shown). The hierarchy of SspC binding to syntheticand natural DNAs was reflected in the salt sensitivity of the
J. BACTERIOL.
on June 27, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

NEW CLASS OF DNA-BINDING PROTEINS 2317
A B C Av 7 ad
B1 9 I 4
C-, 3 4
b -
C o
d -
a _
c _
d -
FIG. 4. Demonstration of lack of binding of single-strandedDNA, poly(dA) * poly(dT), and poly(A) poly(U) by SspC. (A) Thecompetitive electrophoretic assay was carried out as described inMaterials and Methods with 5 ,ug (lanes 1, 3, 4, and 5) or 15 ptg (lane2) ofEcoRI-cleaved pUC19 and 10 pLg of SspCWt. Competitor nucleicacids added: lanes 1 and 2, none; lane 3, 10 pLg of poly(dA); lane 4,10 ,ug of poly(dT); lane 5, 10 ,ug of poly(dA) poly(dT). Nuclease-resistant products were analyzed on a 2% agarose gel. (B) The directelectrophoretic assay was carried out as described in Materials andMethods with 5 ,ug of poly(dA) poly(dT). Additions to samplesanalyzed in different lanes: lanes 1 and 2, no additions; lane 3, 10 ,ugof SspCwt; lane 4, 25 Vg of SspCwt. The sample in lane 1 was nottreated with DNase, while those in lanes 2 through 4 were. Finalsamples were analyzed by electrophoresis on a 2% agarose gel. (C)The direct electrophoretic assay was carried out as described inMaterials and Methods with 5 ,ug of poly(A) poly(U). Additions tosamples analyzed: lanes 1 and 2, no additions; lane 3, 10 ,ug ofSspCw'; lane 4, 25 ,ug of SspCwl. The sample in lane 1 was nottreated with RNase, while the others were. Final samples wereanalyzed by electrophoresis on a 2% agarose gel. The letteredarrowheads denote the migration positions of DNA size markers of4.5 kb (a), 2.7 kb (b), 1.1 kb (c), and 0.2 kb (d).
complex formed with these different polymers. While arather low salt concentration (100 mM) completely dissoci-ated the complex between SspC and poly(dA-dT) poly(dA-dT), a much higher concentration (>300 mM) was requiredto completely remove pUC19 or poly(dG) poly(dC) fromSspC (data not shown). In contrast to the 10 and 40 min ittook for pUC19 to become DNase resistant upon SspCbinding at 37 and 4°C, respectively, acquisition of full DNaseprotection upon binding of SspC to poly(dG) poly(dC) orpoly(dA-dT) poly(dA-dT) at 4 or 37°C took less than 20 s(data not shown).There was little if any cooperativity between SspC mole-
TABLE 2. Protection of nucleic acids fromDNase by SspC binding'
DNase-resistant nucleic acid' (% ofNucleic acid maximum) at SspC/DNA ratio:
1 1.25 2.5 4 5 6 8
Poly(dA-dT) poly(dA-dT)c 27 56 87 89pUC19d 16 63 93 96Poly(dG-dC) poly(dG-dC)d 14 44 96 93Poly(dG) poly(dC)d 25 59 98
a DNase protection assays were carried out with the appropriate labelednucleic acid and SspC as described in Materials and Methods.
b All values are averages of duplicates; in the absence of SspC, <6% of thenucleic acid was resistant to DNase.
c SspC`' used with 5 pug of DNA per assay.d SspCTYr used with 3 ,ug of DNA per assay.
s, a _
v b _
_ c _
_ d _
FIG. 5. DNase resistance of poly(dA-dT)- poly(dA-dT), poly(dG-dC). poly(dG-dC), and poly(dG) poly(dC) upon binding toSspC. The direct electrophoretic assay was carried out with (A) 5 ,ugof poly(dA-dT) poly(dA-dT), (B) 5 ,ug of poly(dG-dC). poly(dG-dC), or (C) 5 ,ug of poly(dG) poly(dC). Additions to samples: lanes1 and 2, no additions; lane 3, 10 SLg of SspCwl; lane 4, 25 ,ug ofSspCwl. The samples in lane 1 were not treated with DNase, whilethose in the other lanes were. DNase-resistant fragments were thenanalyzed by electrophoresis on 2% agarose gels. The lettered arrowsrefer to the migration positions of DNA size markers of 9.6 kb (a),1.3 kb (b), 0.3 kb (c), and 0.12 kb (d).
cules in binding to poly(dG) poly(dC) or poly(dG-dC) poly(dG-dC), as the average size of the DNase-resistant frag-ments increased with increasing SspC/DNA ratio, and ma-terial smaller than 100 bp was protected (Fig. SB and C). Inaddition, when small (<100-bp) DNase-protected fragmentsof poly(dG) poly(dC) were isolated, incubated with saturat-ing levels of SspC, and re-treated with DNase, they were stillfully protected against digestion (data not shown). In con-trast, binding of SspC to poly(dA-dT) poly(dA-dT) ap-peared to be rather cooperative, as the average size ofDNase-resistant fragments was large even at low protein/DNA ratios, and no protected fragments of < 250 bp in lengthwere seen (Fig. SA). The results of this analysis withpolymers were borne out by analysis of SspC binding todouble-stranded oligonucleotides, as a 12-bp oligo(dG).oligo(dC) was protected against DNase by SspC (Fig. 7).While this 12-mer was bound with less affinity than a 26-mer,there was no major discrimination between the 26-mer andlarger oligo(dG). oligo(dC) (Fig. 7). In contrast, oligo(dA-dT). oligo(dA-dT) oligonucleotides from 12 to 68 bp inlength were not protected from DNase by SspC, whichpresumably bound weakly if at all to these oligomers (datanot shown).Other effects of a/3-type SASP-DNA binding. Since a/,13-
type SASP-DNA binding provided strong DNase protection,it seems likely that these proteins are interacting with theDNA backbone. If this is the case, then attack of otherreagents on the DNA backbone might be slowed by ao/3-typeSASP binding. Indeed, DNA cleavage induced by eitherhydroxyl radicals or incubation with orthophenanthroline-Cu21 was significantly slowed by SspC binding but not byaddition of the SspC variant SspCA"a, which has lost allDNA-binding ability (Fig. 8A and B). However, the bindingof SspC to DNA had little effect on the susceptibility ofpurines to methylation, as treatment of DNA fragments withdimethyl sulfate gave essentially identical patterns of purine
VOL. 174, 1992
on June 27, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

2318 SETLOW ET AL.
1 2 3 4 c e
_ 165nt
_b
FIG. 6. DNase protection of a poly(dA)-containing DNA frag-ment. The 32P-end-labeled 270-bp EcoRI-PvuII fragment of pBSMcontaining 58 A residues at the EcoRI end was preincubated in 25 plof 10mM Tris-Acetate (pH 7.0) with (lanes 1 and 3) or without (lanes2 and 4) a 10:1 (wt/wt) ratio of SspCTYr. After 2 h at 37°C, MgCl2 wasadded to 2 mM and either 0.1 ,ug (lanes 1 and 2) or 0.03 ,ug (lanes 3and 4) of DNase was added. After 15 min at 37°C, DNase digestionwas halted by addition of SDS-EDTA, and DNA was isolated,electrophoresed on an 8% polyacrylamide sequencing gel, and thenautoradiographed. Arrowhead b denotes the end of the 58-bp dAtract at the end of the fragment; arrowhead a denotes the position ofa dA3 region near the large dA tract; the arrowhead labeled 165 ntdenotes the migration position of a size marker.
methylation in both naked DNA and a saturated SspC-DNAcomplex (Fig. 8C and D). Control experiments with theDNA fragments used to assess the effect of SspC on chem-ical cleavage or modification of DNA showed that SspCbinding provided complete DNase resistance to these frag-ments at SspC/DNA ratios (wt/wt) of 8:1 (125-bp TaqI-Hinflfragment) or 5:1 (396-bp Hinfl fragment) (data not shown).
-_54bp
_140bp
_*26bp
1' 2bp
FIG. 7. Protection of oligo(dG) oligo(dC) from DNase digestionby SspC binding. Aliquots (1 ,ug) of AT(dG)12. (dC)12AT which hadbeen self-ligated and then purified as described in Materials andMethods were incubated in 25 pd of 10 mM Tris-acetate (pH 7.0)-imM EDTA and increasing amounts of SspClyr. After 2 h at 37°C,the samples in lanes a to g were digested with DNase, and nucleicacid was isolated as described for the direct electrophoretic assay.Remaining oligonucleotides were then analyzed by electrophoresison a nondenaturing 12% polyacrylamide gel. The amounts of SspC-Tyr added in the samples run in the different lanes were none (lanesa and h), 0.5 ,ug (lane b), 1 ,ug (lane c), 2 ,ug (lane d), 4 ,ug (lane e),7 jig (lane f), and 10 ,ug (lane g). The lettered horizontal arrowheadsgive the sizes of these oligonucleotides (in base pairs).
The strong protection of the DNA backbone by SspCfurther suggested that attack by other enzymes on DNAwould be blocked in an SspC-DNA complex. Indeed, SspCblocked cleavage of pUC19 by a number of restrictionenzymes, with less of an effect on others (Table 4). Notsurprisingly, those enzymes whose action was inhibitedmost were generally ones recognizing GC-rich sequences,while those recognizing AT-rich sequences tended to beinhibited least (Table 4). In all cases, the SspC variantSSpCMa, which has lost all DNA-binding activity (42), hadno effect on restriction enzyme cleavage (Table 4 and datanot shown).As a further test of the effect of SspC on DNA properties,
we measured in vitro transcription from SspC-DNA com-
TABLE 3. Competition between different nucleic acids for SASP binding'
DNase-resistant nucleic acid (% of maximum)
Competitor nucleic Poly(dA-dT) pUC19 Poly(dG-dC) Poly(dG) poly(dC)
SspC SASP-lb SspC SASP-4 SspC SASP-P SspC SASP-(3Nonec 100 100 100 100 100 100 100 100Poly(dA-dT) poly(dA-dT) (10) 36 40 103 97 101 d 101pUC19 (5) <2 <2 27 31 92 94 98Poly(dG-dC) poly(dG-dC) <2 <2 11 9 41 31 67 74Poly(dG) poly(dC) (10) <2 <2 <2 <2 4 8 21 30
a Competitive acid precipitation assays were carried out as described in Materials and Methods with a 4:1 ratio of protein to DNA and either 5 pg[poly(dA-dT) poly(dA-dT)] or 3 ,ug [pUC19, poly(dG-dC) poly(dG-dC), poly(dG)- poly(dC)] of labeled DNA. SspC't was used with poly(dA-[3H]dT).poly(dA-[3H]dT) and [3H]pUC19; SspCT'r was used with poly(dG-[3H]dC). poly(dG-[3HjdC) and poly(dG) poly([3H]dC). All values are averages of at least twodeterminations.
b C bifermentans SASP-1.C These values were set at 100% and represent retention of 85 to 93% of total input counts in acid-insoluble form.d, not done.
J. BACTERIOL.
on June 27, 2020 by guesthttp://jb.asm
.org/D
ownloaded from
NEW CLASS OF DNA-BINDING PROTEINS 2319
A1 2 3 4 1 2 3 1 2 3 1 2 3
150O
95 D
125 _
35 _
_150
_oQ95
32 _-32
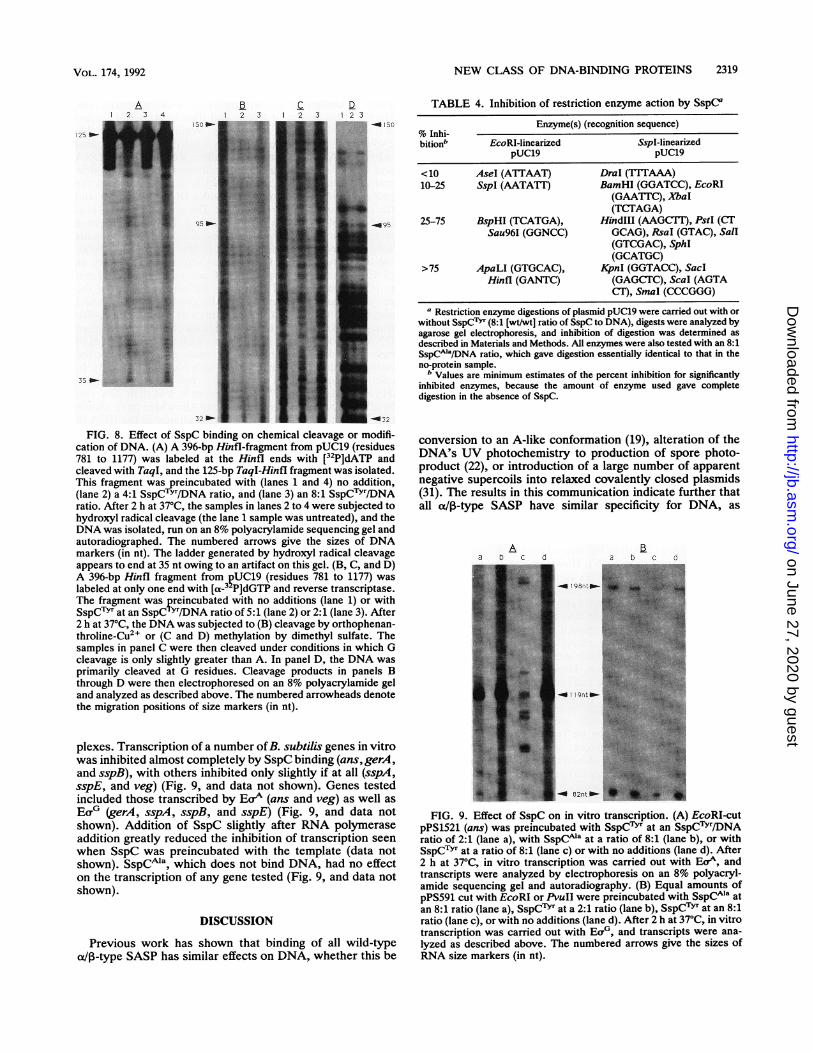
FIG. 8. Effect of SspC binding on chemical cleavage or modifi-cation of DNA. (A) A 396-bp HinfI-fragment from pUC19 (residues781 to 1177) was labeled at the Hinfl ends with [32P]dATP andcleaved with TaqI, and the 125-bp TaqI-Hinfl fragment was isolated.This fragment was reincubated with (lanes 1 and 4) no addition,(lane 2) a 4:1 SspCTYT/DNA ratio, and (lane 3) an 8:1 SspCTYr/DNAratio. After 2 h at 37°C, the samples in lanes 2 to 4 were subjected tohydroxyl radical cleavage (the lane 1 sample was untreated), and theDNA was isolated, run on an 8% polyacrylamide sequencing gel andautoradiographed. The numbered arrows give the sizes of DNAmarkers (in nt). The ladder generated by hydroxyl radical cleavageappears to end at 35 nt owing to an artifact on this gel. (B, C, and D)A 396-bp Hinfl fragment from ?UC19 (residues 781 to 1177) waslabeled at only one end with [a-3 P]dGTP and reverse transcriptase.The fragment was preincubated with no additions (lane 1) or withSspCTYr at an SspCTYr/DNA ratio of 5:1 (lane 2) or 2:1 (lane 3). After2 h at 37°C, the DNA was subjected to (B) cleavage by orthophenan-throline-Cu2, or (C and D) methylation by dimethyl sulfate. Thesamples in panel C were then cleaved under conditions in which Gcleavage is only slightly greater than A. In panel D, the DNA wasprimarily cleaved at G residues. Cleavage products in panels Bthrough D were then electrophoresed on an 8% polyacrylamide geland analyzed as described above. The numbered arrowheads denotethe migration positions of size markers (in nt).
plexes. Transcription of a number ofB. subtilis genes in vitrowas inhibited almost completely by SspC binding (ans,gerA,and sspB), with others inhibited only slightly if at all (sspA,sspE, and veg) (Fig. 9, and data not shown). Genes testedincluded those transcribed by Eor (ans and veg) as well asEo0 (gerA, sspA, sspB, and sspE) (Fig. 9, and data notshown). Addition of SspC slightly after RNA polymeraseaddition greatly reduced the inhibition of transcription seenwhen SspC was preincubated with the template (data notshown). SSpCMa, which does not bind DNA, had no effecton the transcription of any gene tested (Fig. 9, and data notshown).
DISCUSSION
Previous work has shown that binding of all wild-typeoa/1-type SASP has similar effects on DNA, whether this be
TABLE 4. Inhibition of restriction enzyme action by SspC"Enzyme(s) (recognition sequence)
% Inhi-bitionb EcoRI-linearized SspI-linearized
pUC19 pUC19
<10 AseI (ATTAAT) DraI (TTTAAA)10-25 SspI (AATATT) BamHI (GGATCC), EcoRI
(GAATTC), XbaI(TCTAGA)
25-75 BspHI (TCATGA), HindIII (AAGCTT), PstI (CTSau96I (GGNCC) GCAG), RsaI (GTAC), SalI
(GTCGAC), SphI(GCATGC)
>75 ApaLI (GTGCAC), KpnI (GGTACC), SaclHinfl (GANTC) (GAGCTC), Scal (AGTA
CT), SmaI (CCCGGG)a Restriction enzyme digestions of plasmid pUC19 were carried out with or
without SspC`YT (8:1 [wt/wt] ratio of SspC to DNA), digests were analyzed byagarose gel electrophoresis, and inhibition of digestion was determined asdescribed in Materials and Methods. All enzymes were also tested with an 8:1SsPCAIa/DNA ratio, which gave digestion essentially identical to that in theno-protein sample.
b Values are minimum estimates of the percent inhibition for significantlyinhibited enzymes, because the amount of enzyme used gave completedigestion in the absence of SspC.
conversion to an A-like conformation (19), alteration of theDNA's UV photochemistry to production of spore photo-product (22), or introduction of a large number of apparentnegative supercoils into relaxed covalently closed plasmids(31). The results in this communication indicate further thatall ot/P-type SASP have similar specificity for DNA, as
Abaa b c d a b c
| _ 198nt_-
9nt -
- 82nt _
FIG. 9. Effect of SspC on in vitro transcription. (A) EcoRI-cutpPS1521 (ans) was preincubated with SspCTYr at an SspC'Y'r/DNAratio of 2:1 (lane a), with SspCMa at a ratio of 8:1 (lane b), or withSspC"Y' at a ratio of 8:1 (lane c) or with no additions (lane d). After2 h at 37°C, in vitro transcription was carried out with EaA, andtranscripts were analyzed by electrophoresis on an 8% polyacryl-amide sequencing gel and autoradiography. (B) Equal amounts ofpPS591 cut with EcoRI or PvuII were preincubated with SspC"' atan 8:1 ratio (lane a), SspC'Yr at a 2:1 ratio (lane b), SspC"T' at an 8:1ratio (lane c), or with no additions (lane d). After 2 h at 37°C, in vitrotranscription was carried out with Ecr, and transcripts were ana-lyzed as described above. The numbered arrows give the sizes ofRNA size markers (in nt).
VOL. 174, 1992
on June 27, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

2320 SETLOW ET AL.
a/13-type SASP from three different Bacillus species gavesimilar patterns of DNase-resistant bands in pUC19. Wenote, however, that these patterns were not identical. Thus,it is possible that different a/03-type SASP from the samespecies might have slightly different DNA specificities, andthis could be the reason for multiple ae/n-type SASP in onespecies. Indeed, the at/3-type SASP from C. bifermentansgave a different pattern of DNase-resistant bands in pUC19than that given by proteins from Bacillus species. However,all a/IB-type SASP had similar affinities for various syntheticDNAs, with poly(dG) poly(dC) bound most strongly. Sincethe approximate Kd for an SspC-pUC19 complex was calcu-lated to be <5 x 10-' and >10 M (21), it is likely that theKd for an SspC-poly(dG) poly(dC) complex is significantlyless than 10-6 M. Previous work has shown that the SspCvariant SspCGIn, in which a glutamine residue has replacedan invariant lysine at position 57, no longer binds poly(dA-dT) poly(dA-dT), pUC19, or poly(dG-dC) poly(dG-dC)but still binds, albeit weakly, to poly(dG) poly(dC) (42).The fact that a/n-type SASP bind most strongly to
poly(dG) poly(dC) and not to poly(dA) poly(dT) and dl-containing DNAs is notable in view of the conversion ofDNA from the B conformation to an A-like conformationupon a/,8-type SASP binding (19). GC-rich double-strandedoligonucleotides invariably are found to adopt an A-likestructure in crystals and in solution may be on the borderbetween the transition from a B- to an A-like structure (9, 15,27). Poly(dG) poly(dC) is in an A-like structure, or largelyso, in solution (2, 25, 29). In contrast, poly(dA) poly(dT)and dI-containing polymers have never been seen to adoptan A-like conformation under any condition tested, includingdry films and fibers (1, 27). The situation with the copoly-mers poly(dG-dC) poly(dG-dC) and poly(dA-dT) poly(dA-dT) is intermediate, as both are normally in the Bconformation in solution but can adopt the A conformationat reduced humidities (9, 27). However, the A-like structureof poly(dA-dT) poly(dA-dT) is metastable, while that ofpoly(dG-dC) poly(dG-dC) is stable (27).The lack of binding of a/,3-type SASP to poly(dA) poly
(dT) as well as a stretch of 58 dA residues in a plasmid mightseem at odds with the proposed role of these proteins inbinding spore DNA in vivo, changing the DNA conformationto an A-like structure which somehow results in no TTformation on UV irradiation. However, while we have notdetermined the upper size limit of an oligo(dA) oligo(dT)tract in DNA to which oa/1-type SASP can bind, theseproteins appear to bind regions at least 7 to 8 residues inlength, as regions of this size are present in pUB110 andpUC19, both of which can be completely protected fromDNase by SspC. Furthermore, several circular dichroismspectroscopy studies have shown that double-stranded poly-nucleotides containing oligo(dA) oligo(dT) tracts of variouslengths only adopt the unusual circular dichroism spectrumof poly(dA). poly(dT) (indicative of this polynucleotide'sunusual structure) slowly as a function of tract length (8, 10).Indeed, more than 24 continuous dA residues are required togive a circular dichroism spectrum similar to that of poly(dA) poly(dT) (10). Thus, when present as oligo(dA) oligo(dT) tracts in DNA, regions of moderate length, perhaps of20 or less, may well bind o/13-type SASP (albeit more weaklythan GC-rich regions) and change to an A-like conformation.The agreement between the affinity of aet/-type SASP for
various synthetic DNAs and the relative stability of theA-like conformation of these DNAs is consistent with o/13-type SASP binding converting DNA into an A-like confor-mation. However, these proteins may only bind small re-
gions of DNA that are already in an A-like conformation,stabilize this region of DNA, and then bind to and furtherstabilize A-like DNA as it forms. While this is a possibleexplanation of oa3-type SASP-DNA binding, we believe thatat/-type SASP interact initially (albeit weakly) with B-DNAand that this interaction then facilitates the B to A confor-mational change in the DNA, which in turn stabilizes theprotein-DNA interaction and provides DNase resistance andthe change in UV photochemistry. The evidence for this is atleast fourfold. (i) With poly(dA-dT) poly(dA-dT), there isno evidence for the presence of any A-like DNA in aqueoussolution, yet a/13-type SASP bind to this polymer. (ii) Bind-ing of oalp-type SASP to a random sequence DNA such aspUC19 is not instantaneous at 37°C and requires up to 10 minto reach completion. This may be the time required for theprotein-dependent alteration in the conformation of shortoligo(dA). oligo(dT) stretches from a B form to an A-likeform. It is known that different regions of random-sequenceDNA can adopt the A conformation with different degrees ofease, with oligo(dA) oligo(dT) regions generally being theones adopting an A-like structure with most difficulty (3). Incontrast, SspC binding to poly(dG) poly(dC) and poly(dA-dT) poly(dA-dT) at 37°C is complete in less than 20 s. (iii)Studies to be reported in more detail elsewhere have shownthat in addition to the conformational change in the DNA inan a/13-type SASP-DNA complex, there is also a conforma-tional change in the protein (18). A very small amount of thislatter change may take place with a/,B-type SASP which nolonger give DNase protection to pUC19 (i.e., SSpCAIa) (18),suggesting that these proteins can associate weakly withDNA but cannot facilitate the B to A conformational change.Presumably this weak interaction is either easily disruptedby DNase or does not hinder DNase attack. (iv) Binding tosome DNAs [i.e., poly(dA-dT) poly(dA-dT)] is extremelycooperative, as oligo(dA-dT) oligo(dA-dT) oligomers ofless than 70 bp are not protected from DNase by SspC. Thissuggests that protein-protein interactions are required tostabilize the A form of this polymer. This is not the case foroligo(dG). oligo(dC), in which the A form is much morestable.While o/ia-type SASP clearly bind best to DNAs which
most easily assume the A conformation, the A conformationalone is not sufficient for nucleic acid binding. Double-stranded RNAs, including poly(G) poly(C), are in the Aconformation in solution (27) yet exhibit no detectable SASPbinding, nor does an RNA- DNA hybrid. Consequently, thepresence of a hydrogen on the 2' residue of the sugar moietyof the nucleic acid is an important determinant of nucleicacid binding by aot3-type SASP. This observation, as well asthe protection of the DNA backbone from attack by bothchemical and enzymatic cleavage, suggests that a/l-typeSASP-DNA binding must involve the nucleic acid backbone.While the protein may contact the bases to some degree, thismay only be at the edges projecting towards the outside ofthe helix, as methylation of purines in DNA was not affectedby SspC binding. This finding could be taken to mean thatoa13-type SASP bind preferentially in DNA's minor grooverather than the major groove, where the N-7 atom of guanineresides. However, since the exact parameters of the A-likeDNA structure in the SASP-DNA complex are not known, itis premature to draw any definitive conclusions on theprecise nature of SASP-DNA binding. In any event, the lackof sequence specificity in t/13-type SASP-DNA binding isconsistent with the protein's not making significant contactwith the bases. As suggested previously, many of the char-acteristics of oa1B-type SASP-DNA binding outlined above
J. BACTERIOL.
on June 27, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

NEW CLASS OF DNA-BINDING PROTEINS 2321
are those of a new class of DNA-binding protein (19, 21, 22,35). Since these proteins readily bind to some small oligo-nucleotides, it should be possible to determine the structureof an at/-type SASP-DNA cocrystal. Possibly this analysiswill reveal some new features of DNA-protein interaction.One further point about the a/d-type SASP-DNA interac-
tion concerns the effect of this interaction on transcription invitro. While it is not possible at present to predict a priorithose genes whose transcription would be inhibited most bya/o-type SASP binding (unless a gene's regulatory regionhad huge stretches of A's or G's), the fact that in vitrotranscription of different genes exhibited differential sensi-tivity to SspC binding suggests that the same phenomenoncould take place in vivo. Indeed, synthesis of SspC in E. coli(30) or failure to degrade a/p-type SASP during sporegermination (28) results in a large decrease in subsequentrates of RNA and protein synthesis. While most genes mightbe turned off by ot/n-type SASP binding, transcription ofothers could be resistant to, or possibly even stimulated by,this binding. Consequently, it is possible that synthesis ofa/a-type SASP during sporulation plays some regulatoryrole in subsequent gene expression. Unfortunately, only onegene (termed 0.3 kb) is known which is synthesized in theforespore well after a/p-type SASP, and this gene is ex-pressed at a very low level (26). However, recent workindicates that expression of 0.3 kb is ca. threefold higherduring sporulation of B. subtilis strains lacking major ot/I-type SASP (o-13- strains) than in a+j3+ strains and that thisincreased expression in aot-- strains is abolished by intro-duction of a plasmid carrying a highly expressed gene codingfor an ac/p-type SASP (16). Thus, in addition to their struc-tural role in spore DNA, cx/p-type SASP may play a regula-tory role in gene expression during sporulation as well.
ACKNOWLEDGMENTS
This work was supported by grants from the Army ResearchOffice and the National Institutes of Health (GM19698).We are grateful to Heather Fairhead, Tara Grey, and Waltraut
Waterman for assistance with some of the experiments.
REFERENCES1. Arnott, S., and E. Selsing. 1974. Structures for the polynucleo-
tide complexes poly(dA) poly(dT) and poly(dT). poly(dA)poly(dT). J. Mol. Biol. 88:509-521.
2. Arnott, S., and E. Selsing. 1974. The structure of polydeoxygua-nylic acid. polydeoxycytidylic acid. J. Mol. Biol. 88:551-552.
3. Becker, M. M., and Z. Wang. 1989. B--A transitions within a SSribosomal RNA gene are highly sequence specific. J. Biol.Chem. 264:4163-4167.
4. Cabrera-Martinez, R., J. M. Mason, B. Setlow, W. M. Waites,and P. Setlow. 1989. Purification and amino acid sequence oftwo small, acid-soluble proteins from Clostridium bifermentansspores. FEMS Lett. 61:139-144.
5. Cabrera-Martinez, R. M., and P. Setlow. 1991. Cloning andnucleotide sequence of three genes coding for small, acid-soluble proteins of Clostridium perfringens spores. FEMS Mi-crobiol. Lett. 77:127-132.
6. Feavers, I. M., J. Foulkes, B. Setlow, D. Sun, W. Nicholson, P.Setlow, and A. Moir. 1990. The regulation of transcription of thegerA spore germination operon of Bacillus subtilis. Mol. Micro-biol. 4:275-282.
7. Francesconi, S. C., T. J. MacAlister, B. Setlow, and P. Setlow.1988. Immunoelectron microscopic localization of small, acid-soluble spore proteins in sporulating cells of Bacillus subtilis. J.Bacteriol. 170:5963-5967.
8. Gudibande, S. R., S. D. Jayasena, and M. J. Behe. 1988. CDstudies of double-stranded polydeoxynucleotides composed ofrepeating units of contiguous homopurine residues. Biopoly-
mers 27:1905-1915.9. Heinemann, U., C. Alings, and H. Lauble. 1989. Structural
features of G/C rich DNA going A or B, p. 39-53. In R. H.Sarma and M. H. Sarma (ed.), Structure and methods: DNAand RNA, vol. 3. Adenine Press, Guilderland, N.Y.
10. Jayasena, V. K., and M. J. Behe. 1991. Oligopurine oligopyri-midine tracts do not have the same conformation as analogouspolypurine. polypyrimidines. Biopolymers 31:511-518.
11. Magill, N. G., C. A. Loshon, and P. Setlow. 1990. Small,acid-soluble, spore proteins and their genes from two species ofSporosarcina. FEMS Lett. 72:293-298.
12. Mason, J. M., and P. Setlow. 1986. Evidence for an essentialrole for small, acid-soluble, spore proteins in the resistance ofBacillus subtilis spores to UV light. J. Bacteriol. 167:174-178.
13. Mason, J. M., and P. Setlow. 1987. Different small, acid-solubleproteins of the o/1 type have interchangeable roles in the heatand UV radiation resistance of Bacillus subtilis spores. J.Bacteriol. 169:3633-3637.
14. Maxam, A. M., and W. Gilbert. 1980. Sequencing end-labeledDNA with base-specific chemical cleavages. Methods Enzymol.65:499-560.
15. McCall, M., T. Brown, and 0. Kennard. 1985. The crystalstructure of d(GGGGCCCC). A model for poly(dG) poly(dC).J. Mol. Biol. 183:385-396.
16. McGinnis, K., and P. Setlow. Unpublished results.17. Melton, D. A., P. A. Krieg, M. R. Rebagliati, T. Maniatis, K.
Zinn, and M. R. Green. 1984. Efficient in vitro synthesis ofbiologically active RNA and RNA hybridization probes fromplasmids containing a bacteriophage SP6 promoter. NucleicAcids Res. 12:7035-7070.
18. Mohr, S., and P. Setlow. Unpublished results.19. Mohr, S. C., N. V. H. A. Sokolov, C. He, and P. Setlow. 1991.
Binding of small acid-soluble spore proteins from Bacillussubtilis changes the conformation of DNA from B to A. Proc.Natl. Acad. Sci. USA 88:77-81.
20. Moran, C. P., Jr., N. Lang, S. F. J. LeGrice, G. Lee, M.Stephens, A. L. Sonenshein, J. Pero, and R. Losick. 1982.Nucleotide sequences that signal the initiation of transcriptionand translation in Bacillus subtilis. Mol. Gen. Genet. 186:339-346.
21. Nicholson, W. L., B. Setlow, and P. Setlow. 1990. Binding ofDNA in vitro by a small, acid-soluble spore protein and its effecton DNA topology. J. Bacteriol. 172:6900-6906.
22. Nicholson, W. L., B. Setlow, and P. Setlow. 1991. Ultravioletirradiation of DNA complexed with a/6-type small, acid-solubleproteins from spores of Bacillus or Clostridium species makesspore photoproduct but not thymine dimers. Proc. Natl. Acad.Sci. USA 88:8288-8292.
23. Nicholson, W. L., and P. Setlow. 1990. Dramatic increase in thenegative superhelicity of plasmid DNA in the forespore com-partment of sporulating cells of Bacillus subtilis. J. Bacteriol.172:7-14.
24. Nicholson, W. L., D. Sun, B. Setlow, and P. Setlow. 1989.Promoter specificity of sigma-G-containing RNA polymerasefrom sporulating cells of Bacillus subtilis: identification of agroup of forespore-specific promoters. J. Bacteriol. 171:2708-2718.
25. Nishimura, Y., C. Torigoe, and M. Tsuboi. 1985. An A-formpoly(dG) poly(dC) in H20 solution. Biopolymers 24:1841-1844.
26. Panzer, S., R. Losick, D. Sun, and P. Setlow. 1989. Evidence foran additional temporal class of gene expression in the foresporecompartment of sporulating cells of Bacillus subtilis. J. Bacte-riol. 171:561-564.
27. Saenger, W. 1984. Principles of nucleic acid structure. Springer-Verlag, New York.
28. Sanchez-Salas, J.-L., M. L. Santiago-Lara, B. Setlow, M. D.Sussman, and P. Setlow. 1992. Properties Bacillus megateriumand Bacillus subtilis mutants which lack the protease thatdegrades small, acid-soluble proteins during spore germination.J. Bacteriol. 174:807-814.
29. Sarma, M. H., G. Gupta, and R. H. Sarma. 1986. 500-MHz 'HNMR study of poly(dG). poly(dC) in solution using one-dimen-
VOL. 174, 1992
on June 27, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

2322 SETLOW ET AL.
sional nuclear Overhauser effect. Biochemistry 25:3659-3665.30. Setlow, B., A. R. Hand, and P. Setlow. 1991. Synthesis of a
Bacillus subtilis small, acid-soluble spore protein in Escherichiacoli causes cell DNA to assume some characteristics of sporeDNA. J. Bacteriol. 173:1642-1653.
31. Setlow, B., and P. Setlow. Unpublished results.32. Setlow, B., and P. Setlow. 1979. Localization of low-molecular-
weight basic proteins in Bacillus megaterium spores by irradi-ation with ultraviolet light. J. Bacteriol. 139:486-494.
33. Setlow, B., and P. Setlow. 1987. Thymine-containing dimers aswell as spore photoproducts are found in ultraviolet-irradiatedBacillus subtilis spores that lack small acid-soluble proteins.Proc. Natl. Acad. Sci. USA 84:421-423.
34. Setlow, P. 1975. Purification and properties of some unique lowmolecular weight basic proteins degraded during germination ofBacillus megaterium spores. J. Biol. Chem. 250:8168.
35. Setlow, P. 1988. Small acid-soluble, spore proteins of Bacillusspecies: structure, synthesis, genetics, function and degrada-tion. Annu. Rev. Microbiol. 42:319-338.
36. Setlow, P. 1988. Resistance of bacterial spores to ultravioletlight. Comments Mol. Cell. Biophys. 5:253-264.
37. Setlow, P. 1989. Forespore-specific genes of Bacillus subtilis:function and regulation of expression, p. 211-221. In I. Smith,R. Slepecky, and P. Setlow (ed.), Regulation of procaryoticdevelopment. American Society for Microbiology, Washington,D.C.
38. Sigman, D. S., A. Spassky, S. Rimsky, and H. Buc. 1985.Conformational analysis of lac promoters using the nucleaseactivity of 1,10-phenanthroline-copper ion. Biopolymers 24:183-197.
39. Sun, D., and P. Setlow. 1991. Cloning, nucleotide sequence, andexpression of the Bacillus subtilis ans operon that codes forL-asparaginase and L-aspartase. J. Bacteriol. 173:3831-3845.
40. Sun, D., P. Stragier, and P. Setlow. 1989. Identification of a newa-factor involved in compartmentalized gene expression duringsporulation of Bacillus subtiis. Genes Dev. 3:141-149.
41. Sussman, M. D., and P. Setlow. 1991. Cloning, nucleotidesequence, and regulation of the Bacillus subtilis gpr gene whichcodes for the protease that initiates degradation of small,acid-soluble proteins during spore germination. J. Bacteriol.173:293-300.
42. Tovar-Rojo, F., and P. Setlow. 1991. Analysis of the effects ofmutant small, acid-soluble spore proteins from Bacillus subtilison DNA in vivo and in vitro. J. Bacteriol. 173:4827-4835.
43. Tullius, T. D., and B. A. Dombroski. 1986. Hydroxyl radical"footprinting": high-resolution information about DNA-proteincontacts and application to A repressor and Cro protein. Proc.Natl. Acad. Sci. USA 83:5469-5473.
44. Yuan, K., W. C. Johnson, D. J. Tipper, and P. Setlow. 1981.Comparison of various properties of low-molecular-weight pro-teins from dormant spores of various Bacillus species. J. Bac-teriol. 146:965-971.
J. BACTERIOL.
on June 27, 2020 by guesthttp://jb.asm
.org/D
ownloaded from


















