
Infezione toxoplasmica nel cane: indagine sierologica ... · la reale importanza di questa...
97
UNIVERSITÀ DI PISA Dipartimento di Scienze Veterinarie Corso di Laurea Specialistica in Medicina Veterinaria Infezione toxoplasmica nel cane: indagine sierologica retrospettiva su cani viventi in Toscana e Liguria Candidato: Tempori Chiara Relatore: Prof.ssa Francesca Mancianti Correlatore: Dott.ssa Simona Nardoni ANNO ACCADEMICO 2013-2014
Transcript of Infezione toxoplasmica nel cane: indagine sierologica ... · la reale importanza di questa...

UNIVERSITÀ DI PISA
Dipartimento di Scienze Veterinarie
Corso di Laurea Specialistica in Medicina Veterinaria
Infezione toxoplasmica nel cane: indagine
sierologica retrospettiva su cani viventi in
Toscana e Liguria
Candidato: Tempori Chiara Relatore: Prof.ssa Francesca Mancianti
Correlatore: Dott.ssa Simona Nardoni
ANNO ACCADEMICO 2013-2014

2
indice
RIASSUNTO……………………………………………………...pag. 4
ABSTRACT……………………………………………………….pag. 5
Capitolo 1 Definizione, riferimenti storici e importanza….pag. 6
1.1 Definizione……………………………………….pag. 6
1.2 Riferimenti Storici………………………………..pag. 7
1.3 Diffusione nel mondo…………………………….pag. 9
1.4 Importanza dell’infezione umana……………….pag. 12
Capitolo 2 Ciclo biologico e morfologia…………………..pag. 15
2.1 Ciclo biologico……………..
2.2 Morfologia……………………………………...pag. 18
2.3 Genotipi………………………………………...pag. 26
Capitolo 3 Modalità di trasmissione………………………pag. 27
3.1 Ingestione delle oocisti sporulate……………….pag. 27
3.2 Ingestione dei bradizoiti………………………...pag. 27
3.3 Ingestione dei tachizoiti………………………...pag. 28
3.4 Trasmissione transplacentare…………………...pag. 28
3.5 Modalità di trasmissione in ambiente acquatico..pag. 29
Capitolo 4 Immunità anti-toxoplasma e patogenesi……...pag. 31
4.1 Immunità anti T.gondii………………………….pag. 31
4.2 Patogenesi……………………………………....pag. 32

3
Capitolo 5 Sintomi clinici…………………………………..pag. 33
5.1 Sintomi clinici nell’infezione congenita………..pag. 33
5.2 Sintomi clinici nell’infezione post-natale……….pag. 33
5.3 Reperti anatomopatologici e lesioni istologiche..pag. 35
Capitolo 6 Diagnosi………………………………………...pag. 36
6.1 Diagnosi diretta……………………………...….pag. 36
6.2 Diagnosi indiretta……………………………….pag. 37
6.2.1 Tecnica IFAT………………………..pag. 39
Capitolo 7 Terapia e prevenzione…………………………pag. 43
7.1 Trattamento farmacologico……………………..pag. 44
7.2 Profilassi vaccinale…………………………...…pag. 44
Capitolo 8 Scopo del lavoro………………………………..pag. 45
Capitolo 9 Materiali e metodi……………………………...pag. 46
9.1 Campionamento………………………………...pag. 46
9.2 Immunofluorescenza indiretta…………………..pag. 47
9.3 Leishmania Infantum e Neospora Caninum….....pag. 48
Capitolo 10 Risultati……………………………………..….pag. 50
Capitolo 11 Discussione e conclusioni……………………...pag. 59
Bibliografia………………………………………………………pag. 66
Ringraziamenti…………………………………………………..pag. 96

4
RIASSUNTO
Parole chiave: Toxoplasma gondii, cane, IFAT, sierologia, Toscana.
La toxoplasmosi è una zoonosi cosmopolita, diffusa in tutto il mondo e con forte impatto sulla
salute pubblica. L’agente eziologico, Toxoplasma gondii, è un protozoo ,parassita obbligato
delle cellule nucleate degli animali omeotermi, circa 200 specie di mammiferi (uomo compreso)
e 62 specie di uccelli. L’epidemiologia della toxoplasmosi del gatto è ampiamente conosciuta
sono invece pochi i dati sul cane e gli articoli presenti in bibliografia riguardano solamente paesi
esteri. Gli studi sulla toxoplasmosi canina in Italia sono pochi e non recenti, assenti quelli sulla
prevalenza del parassita in Toscana.
Lo scopo del lavoro è stato quello valutare la sieroprevalenza di T.gondii in uno studio
retrospettivo (gennaio 2012 a dicembre 2013), condotto su 1562 sieri di cane, pervenuti presso il
laboratorio di sierologia parassitaria del Dipartimento di Scienze veterinarie dell’Università di
Pisa, per il controllo sierologico della leishmaniosi, sono stati sottoposti alla tecnica IFAT per la
ricerca di anticorpi anti-toxoplasma; 9 degli animali esaminati sono risultati affetti da segni
clinici compatibili. Sui positivi è stata eseguita anche l’IFAT per Neospora caninum. 13 erano
positivi e l’analisi dei titoli anticorpali sembrava indicare una cross reazione con toxoplasma. I
dati ottenuti sono stati analizzati e confrontati con i dati riguardanti altri paesi, indicando una
buona corrispondenza. In Toscana T.gondii, così come N.caninum, è presente ma scarsamente
riconosciuto e spesso non vengono effettuati test diagnostici per attestarne la presenza.

5
ABSTRACT
Key-words : Toxoplasma gondii, dog, IFAT, serology.
Toxoplasmosis is a cosmopolitan zoonosis, spread all over the world, which has a strong impact
on public health. The etiological agent, Toxoplasma gondii, is a protozoan, an obligate parasite
of the nucleated cells of homeothermic animals, around 200 species of mammals (including
humans) and 62 species of birds. The epidemiology of toxoplasmosis in cats is widely known,
there is little information concerning the toxoplasmosis in dogs instead, and the articles in the
bibliography only refer to foreign countries. Studies on canine toxoplasmosis in Italy are few and
are not recent, and the ones relating to the prevalence of the parasite are absent in Tuscany.
The purpose of this paper was to evaluate the seroprevalence of T. gondii in a retrospective study
(January 2012 to December 2013). The study was conducted on 1,562 dog sera, which were sent
to the parasitic serology laboratory of the Department of Veterinary Sciences of the University of
Pisa, for the serological test of leishmaniasis. The IFAT technique was performed for the
detection of anti-Toxoplasma antibodies. Nine of the examined animals were found to be
suffering from compatible clinical signs. On the positive samples was also performed IFAT for
the detection of Neospora caninum. Thirteen samples were positive and the analysis of antibody
titers seemed to indicate a cross-reaction with toxoplasma. The obtained data was analyzed and
compared with data concerning other countries, showing a good match. T. gondii, as well as N.
caninum, is present in Tuscany but is inadequately detected and often no diagnostic tests are
carried out in order to verify its presence.

6
Capitolo 1 Definizione, riferimenti storici e importanza
1.1 Definizione
I protozoi (dal greco “protos”= primo e “zoon”= animale) rappresentano i progenitori di tutti gli
animali. L’organizzazione unicellulare in questi microrganismi ha raggiunto una struttura
citologica estremamente complessa.
Figura 1. Classificazione dei protozoi di interesse parassitologico (da Urquart et al., 1988).
Toxoplasma gondii è un parassita appartenente al Phylum “Apicomplexa” (Levine, 1970), classe
“Sporozoasida” (Leukart, 1879), subclasse “Coccidiasina” (Leukart, 1879), ordine
“Eimeriorina” (Leger, 1911) famiglia “Toxoplasmatidae” (Biocca, 1956),genere “Protozoa”, del
quale è l’unica specie.
T. gondii è un protozoo ubiquitario, parassita obbligato delle cellule nucleate degli animali
omeotermi, circa 200 specie di mammiferi (uomo compreso) e 62 specie di uccelli; l’infezione
sembra essere molto frequente tra gli animali selvatici, soprattutto canidi (Jakubek et al., 2007).
Gli ospiti definitivi sono il gatto e i felidi selvatici, considerati comunque “ospiti
completi”perché possono presentare la fase di sviluppo asessuata extraintestinale del
parassita,tipica dell’ospite intermedio. T.gondii ha una vasta gamma di ospiti secondari (Frenkel
et al., 1970): carnivori selvatici, suini d’allevamento, orsi, felidi, volpi, procioni, moffette fino

7
addirittura ai cervi selvatici, ungulati, marsupiali , scimmie e cetacei, lontra di mare colpita in
modo massivo nel 2005 (Hill, 2005).
1.2 Riferimenti Storici
Il nome toxoplasma deriva dal greco “toxòn”e “plasma” cioè forma ad arco, che descrive la
forma a mezzaluna assunta dal parassita nella fase del tachizoite; ”gondii” deriva invece dal
nome di un roditore nord-africano (Ctenodactylus gondii) da cui fu isolato per la prima volta.
Nel 1908 Nicolle e Manceaux isolarono per caso Toxoplasma, durante delle ricerche sulla
presenza di Leishmania nei roditori del Nord Africa. Nello stesso anno,fu l’italiano Alfonso
Splendore a isolarlo nei conigli. Nel 1917 Chatton e Blanc ritennero Toxoplasma un parassita
ematico, trasmesso da un artropode, perché isolato nel sangue degli ospiti; studi svolti negli anni
successivi scoprirono le caratteristiche biologiche e morfologiche di Toxoplasma e fu compresa
la reale importanza di questa infezione sulla sanità pubblica (Dubey, 2009). Nel 1923 è stato
riportato il primo caso di infezione oculare da Toxoplasma nell’uomo (Janku, 1923). Nel 1938
Wolf e Cowen studiarono la forma congenita della toxoplasmosi in una neonata e questo aprì le
porte a ulteriori approfondimenti sulla trasmissione, che venne dimostrata in diverse specie
animali (Hartley e Marshall, 1957; Dubey, 2009a).
Nel 1948 si raggiunse uno dei più importanti step nel percorso di conoscenza del toxoplasma, la
realizzazione del “Dye Test”da parte di Sabin e Feldmann. Questo test era utilizzato per la
diagnosi nell’uomo mediante l’analisi di campioni di siero ed ha permesso di mettere in
evidenza la massiva diffusione del parassita nella popolazione umana. Tutto ciò ha portato i
ricercatori a valutare l’esistenza di altre vie di trasmissione, poiché la sola congenita, non poteva
essere responsabile di tale prevalenza (Innes, 2010). Nel 1960 Jacobs, insieme ad altri autori,
dimostrarono la trasmissione orizzontale del parassita, mettendo in evidenza come la parete della
ciste fosse distrutta dagli enzimi digestivi dell’ospite, mentre i bradizoiti all’interno venivano
rilasciati (Jacobs et al., 1960).
L’evento determinante nella “storia del toxoplasma” fu la scoperta delle oocisti nelle feci di
gatto, nel 1970,che permise anche la classificazione del parassita tra i Coccidi, vista la grande
similarità con la già nota ”Isospora bigemina” (Hutchison et al. 1969, 1970, 1971; Sheffield e
Melton, 1969; Frenkel et al., 1970; Weiland e Kuhn, 1970).
Hutchinson nel 1965 eseguì un esperimento con il quale verificò che l’infettività di T.gondii era
legata alle feci di gatto, cadendo però in errore poiché riteneva che la trasmissione avvenisse
attraverso le uova di T. cati. Il ricercatore infettò con cisti tissutali di toxoplasma un gatto già
colpito da T.cati, raccogliendo feci contenenti uova di nematodi; flottò, poi, le feci in soluzione

8
al 33% di solfato di zinco e le conservò in acqua per 12 mesi, in un recipiente senza tappo per
farle embrionare. Hutchinson infettò poi dei topolini, i quali svilupparono la toxoplasmosi.
(Hutchison, 1965). L’oocisti risultò essere la forma resistente all’acqua e nell’ambiente esterno,
mentre le forme note fino a quel momento erano sensibili all’acqua. L’esame microscopico delle
feci, però, rivelò apparentemente la presenza soltanto di uova di T. cati e di oocisti di Isospora.
Successivamente Hutchison ripetè l’esperimento su due gatti infettati con T. cati e due esenti
dall’infezione e Toxoplasma fu trasmesso solo in associazione con l’infezione con il nematode;
proprio per tali risultati il ricercatore ipotizzò che Toxoplasma fosse trasmesso con le uova di
Toxocara (Hutchison, 1965). Nel 1969 furono isolate oocisti di toxoplasma in feci di gatto non
infettate da Toxocara, cadde così la teoria della coniugazione nematode-toxoplasma (Frenkel et
al., 1969; Sheffield and Melton, 1969). Nel 1970 le oocisti furono caratterizzate dal punto di
vista morfologico e biologico (Dubey et al., 1970). Numerose specie animali furono infettate con
toxoplasma, ma solo i felidi emisero le cisti poiché esclusivamente questi mammiferi sono ospiti
definitivi. Successivamente furono eseguiti studi su alcune isole nell’Oceano Pacifico (Wallace,
1969), in Australia (Dubey et al., 1997) e negli USA (Munday, 1972) nelle quali non viveva il
gatto e non vi venne dimostrata neanche la presenza del Toxoplasma. I felidi hanno un ruolo
essenziale nella trasmissione del parassita infatti, a sostegno di tale ipotesi, fu riscontrato che
marsupiali australiani erano molto più sensibili alla toxoplasmosi clinica rispetto a altri
esemplari nati in America o nel vecchio mondo; questi ultimi, infatti, entrarono in contatto con il
gatto solo dopo la colonizzazione europea. Anche le scimmie mostrarono una forte sensibilità
verso il toxoplasma, poiché vivendo sugli alberi, venivano raramente a contatto con le feci di
gatto parassitate (Dubey e Beattie, 1988).
Gli ultimi studi sulla trasmissione di toxoplasma hanno rivelato che i felidi possono infettarsi con
qualsiasi stadio del Toxoplasma, smentendo la precedente ipotesi secondo cui gli sporozoiti
contenuti nelle oocisti non infettavano i gatti, mentre anche una sola trasmetteva l’infezione
nell’uomo, nel suino e nei topi (Dubey et al., 1996).
Le ricerche effettuate su Toxoplasma erano rivolte alla ricerca di tachizoiti dal liquido ascitico di
topi infettati sperimentalmente attraverso l’inoculazione intraperitoneale di diversi stadi del
parassita, ma nuovi studi hanno evidenziato che nel liquido peritoneale sono presenti anche
bradizoiti e fasi di transizione (Dubey, 2005).
Nel 2007 Wainwright evidenziò la presenza di una membrana impermeabile intorno delle oocisti
che le rende resistenti a molti disinfettanti (Wainwright et al.,2007).Tra gli animali colpiti da
toxoplasma vengono attualmente considerati anche i mammiferi marini,morti di quest’infezione
acquisita dall’ingestione di acque dolci contaminate da oocisti,probabilmente defluite da terreni
con feci di gatto. (Miller et al., 2002; Dubey et al., 2003b; Conrad et al., 2005; Dubey e Jones,

9
2008). Toxoplasma non infetta animali a sangue freddo (Dubey e Beattie, 1988),tranne i
molluschi lamellibranchi che possono essere trasportatori delle oocisti (Miller et al.,
2002;Arkush et al., 2003; Lindsay et al., 2004).
1.3 Diffusione nel mondo
Figura 2. Sieroprevalenzo globale di T.gondii. Rosso scuro:prevalenza: 60%. Rosso chiaro: 40-60%.
Giallo: 20-40%. Blue:10-20%. Bianco: assenza (Toxoplasmosis snapshots: Global status of Toxoplasma
condii seroprevalence and implications for pregnancy and congenital toxoplasmosis.
2009 International journal parasitology: 1385-1394)
L’infezione da T. gondii è una delle zoonosi più importanti e ubiquitaria del mondo, è stata
evidenziata dall’Alaska all’Australia. L’uomo acquisisce l’infezione attraverso l’ingestione di
carne cruda o poco cotta, il contatto con feci di gatto e molto più raramente bevendo acqua
contaminata o attraverso trapianti con organi inquinati (Hill and Dubey, 2002). Nell’uomo
l’infezione è spesso subclinica, può talvolta essere confusa con una malattia virale poco virulenta
(Montoya e Liesenfeld, 2004). Si stima che oltre mezzo miliardo di persone nel mondo
presentino nel siero anticorpi specifici (Dubey, 1997) e quasi un terzo dell’umanità è stato
esposto al parassita. La prevalenza nell’uomo è massima nei paesi tropicali, forse a causa di
scarse condizioni igienico-sanitarie (70%). Numerosi studi hanno evidenziato un’ elevata

10
presenza del parassita in tutta l’America latina, in particolare in Brasile, dove la presenza di
valenza supera il 60% (Cabrale et al., 2008). Studi sulla sieroprevalenza realizzati nelle
popolazioni native americane, gli amerindi, hanno evidenziato lo stretto legame tra l’elevata
presenza del parassita con le abitudini alimentari e le pratiche sanitarie di questo popolo
(alimentazione a base di scimmie e topi) (Boia et al., 2008); le condizioni climatiche delle foreste
pluviali giocano un ruolo essenziale nella sopravvivenza ambientale del parassita (De la rosa et
al., 1999). Guatemala e Chile seguono l’andamento degli altri paesi dell’America Latina, con una
prevalenza maggiore per il primo (37,9%) (Conteras et al.,1996). Il Messico è invece
caratterizzato da una elevata prevalenza a sud, mentre il nord è poco interessato (Valasco
Castrejon et al., 1992). Tuttavia indagini più recenti hanno evidenziato una riduzione della siero-
prevalenza nello stato del Durango, nel Messico centro-settentrionale (Pappas et al., 2009).
Negli USA la sieroprevalenza è notevolmente ridotta (Jones et al., 2001), è più alta negli
afroamericani e nelle persone di origine messicana, soprattutto nei soggetti non nati negli USA.
La correlazione tra la presenza del toxoplasma e lo stato socio-economico, ha portato gli esperti
a proporre uno screening di massa per la trasmissione congenita del toxoplasma (Hotez, 2008).
In Canada la prevalenza si attesta intorno al 18,6% (Shuhaiber et al., 2003), mentre nella
popolazione “Inuit” la prevalenza sale al 59,8%, per il massivo consumo di carni di foca e di
caribù (Messier et al.,2009).
I dati riguardanti l’Europa sono molto variegati ma allo stesso tempo discontinui. Recentemente
è stato dimostrato come in Europa la sorveglianza epidemiologica per il toxoplasma congenito
sia limitato (Bernard et al., 2008); in realtà solo la Germania e la Francia hanno avviato
programmi di sorveglianza nazionale, mentre l’Austria sta intraprendendo questa strada,
abbandonata recentemente dalla Danimarca. Questi programmi sono organizzati da ”Eurotoxo”,
un progetto che riunisce scienziati che cooperano per analizzare tutti gli aspetti della
Toxoplasmosi congenita. Un ulteriore ostacolo a un’ interpretazione efficiente dei dati è
evidenziabile nelle continue variazioni che interessano la popolazione campionata, soprattutto a
causa dell’immigrazione dall’est-Europa, che interessa la Grecia e la Spagna, dove i tassi sono
elevati. Il paese con la più altra prevalenza in Europa è la Francia, con un tasso del 59% nel 1995
(Barile et al., 1999), sceso al 40%nel 2005, secondo studi dell’EFSA (EFSA, 2007) per il
consumo di carne cruda o poco cotta. L’est Europa è caratterizzata da una elevata siero
prevalenza, soprattutto Lituania, Estonia, Slovacchia e Repubblica Cieca (50%). Nei paesi del
nord, Svezia, Islanda, Danimarca i tassi restano sotto il 20%, mentre si alzano fino al 60% in
Ungheria (Szenasi et al.,1997).

11
Il Portogallo è caratterizzato da una prevalenza medio alta al nord, mentre il sud del paese
sembra essere quasi indenne, questo è causato da una minore consapevolezza della presenza
della malattia.
In Africa sono l’Egitto e la Tunisia ad avere la prevalenza maggiore, con il 59% ( Hussein et al.,
2001; Bouratbine et al., 2001); la Nigeria ha una prevalenza che sfiora il 20 % (Uneke et al.,
2005), mentre studi eseguiti sui donatori di sangue del Mali hanno evidenziato una prevalenza
del 21% (Maiga et al., 2001). In questi paesi la diffusione di T. gondii è collegato al consumo di
carne di ratto e al contatto con il suolo. Studi non recenti e sommari evidenziano una prevalenza
del 71,2% in Gabon (Nabias et al., 1998), 40,2%in Senegal (Faye et al., 1998), e dell’83% in
Madagascar (Lelong et al., 1995).
L’Asia è caratterizzata da un’elevata prevalenza in Turchia, Iran, Iraq e Kuwait, mentre l’Arabia
Saudita, Bahrain e Qatar sono interessata per un 25% (Al-Qurashi et al,2001; Al-Qurashi et
al.,2001; Tabbara and Saleh, 2005; Abu-Madi et al., 2008).In studi successivi, alcuni autori,
prospettarono il timore di un incremento della siero prevalenza in relazione alla presenza di quasi
2 milioni di gatti nella città di Doha, in Qatar; in realtà la prevalenza è molto maggiore nelle città
vicine, rispetto a quella zona che mantiene comunque un tasso esiguo. Il sud est asiatico è
interessato da un’elevata prevalenza, soprattutto Indonesia, Malesia,mentre il Vietnam e la
Tailandia hanno tassi meno elevati; questi dati variano molto in base all’etnia delle persone, in
Tailandia, per esempio, la prevalenza nell’etnia di origine cinese è molto minore rispetto alle
popolazioni aborigene, che sfiora il 40% (Fan et al., 2003). In Malesia, inoltre, il tasso più alto
interessa gli immigrati di origine Nepalese e i malesiani stessi, noti per essere amanti dei gatti e
cani, mentre altre etnie di immigrati sono meno interessate. In uno studio sulla Toxoplasmosi
congenita nel Massachusetts, USA (Jara et al., 2001) ha dimostrato che è molto più comune nei
bambini nati da madri Cambogia e del Laos.
La Cina ha una delle minori prevalenze del mondo, 1%, nella provincia dello Shangdong (Chen
et al., 2005). L’India ha dati molto vari, 9,4% in Rajasthan 48.2% in Kerala (Dhumne et al.,
2007).In Oceania e Nuova Zelanda la prevalenza è diminuita notevolmente rispetto a studi
passati (Cursons et al., 1982).
Uno studio del 2002 ha dimostrato che il 16-40% della popolazione degli Stati Uniti e del Regno
Unito è infetta, mentre in Centro e sud Africa e in Europa questa percentuale arriva fino all’80%
(Hill and Dubey, 2002).

12
1.4 Importanza dell’infezione umana
Figura 3. Ciclo di T.gondii nell'uomo (da http://wwwcentroveterinariofioranese.blogspot.com)
Nel corso dell’evoluzione, T. gondii ha sviluppato una grande varietà di possibili vie di
trasmissione; nella maggior parte dei casi l’infezione umana è acquisita nella fase postnatale e
non è ancora chiara quale sia la più importante fonte di contagio dal punto di vista della salute
pubblica. I tachizoiti , che giocano un ruolo significativo nella trasmissione verticale, sono molto
sensibili alle condizioni ambientali e sono frequentemente uccisi fuori dall’ospite, percui questa
via non è epidemiologicamente rilevante. I tachizoiti sono stati isolati in numerosi fluidi corporei
come saliva, urina, lacrime, seme e latte della maggior parte degli ospiti intermedi (pecore,
capre, bovini e cammelli) ( Tenter et al., 2000; Dekhordi et al., 2013). Studi più recenti
dimostrano come il tachizoita sia stato isolato da uovo crudo di gallina, sperimentalmente
infettata (Jacobs e Melton,1966).
I tachizoiti sono comunque inattivati dagli enzimi proteolitici del tratto intestinale, che sono però
ridotti nei bambini e questo sembra spiegare l’infezione in quelli allattati al seno da madri che
hanno acquisito un’infezione primaria (Bonametti et al., 1997). In realtà l’infezione da tachizoiti
può interessare anche gli adulti, poiché alcuni cibi solidi alzano il pH dello stomaco fino a 5 per
alcune ore e il parassita può arrivare al piccolo intestino, penetrare la mucosa e arrivare al circolo
sanguigno e linfatico (Riemann et al.1975, Sacks et al. 1982, Johnson 1997). I tachizoiti sono

13
comunque in grado, almeno nel gatto, di attraversare le mucose. Fino ad oggi, la maggior parte
di casi clinici negli umani è stato causato dall’assunzione di latte di capra non pastorizzato
(Riemann et al., 1975, Sacks et al., 1982, De Andrade et al., 1984, Skinner et al., 1990), mentre il
rischio d’infezione con quella di vacca è molto inferiore (Jackson e
Hutchison, 1989, Dubey, 1991). Tuttavia non è possibile escludere il rischio d’infezione con il
latte di vacca: in Polonia, uno studio condotto su donne in età fertile, ha rivelato che bere latte
può comportare l’infezione per la trasmissione orizzontale (Paul, 1998). Le popolazioni nomadi
dei pastori del Sudan hanno una sieroprevalenza alta (67%), causata dall’assunzione di latte di
cammello crudo (Elamin et al., 1992).
Uno studio effettuato su campioni di latte di diverse specie, quali bovini, ovini, caprini, cammelli
e bufali in Iran, mediante test in vitro, ELISA e PCR, ha evidenziato come T.gondii fosse
presente nel latte di tutte le specie, con prevalenza nei caprini (10%) con picchi nella stagione
estiva (76%) ( Dehkordi FS,2014).
Non è possibile, perciò,escludere la trasmissione umana attraverso i tachizoiti; l’unico modo per
interrompere questa trasmissione è non consumare il latte crudo, ma bollito o pastorizzato e
cuocere le uova .
La trasmissione animali-uomo è prevalentemente causata dall’ingestione di cisti presenti nelle
carni e nelle frattaglie di animali infetti o per assunzione di acqua o alimenti contaminati da
oocisti sporulate, presenti nell’ambiente o, raramente, direttamente dalle feci di gatto. Le
categorie professionali maggiormente a rischio sono i macellai, i cacciatori e tutti coloro che
lavorano le carni. A differenza dei tachizoiti, i bradizoiti sono maggiormente resistenti agli
enzimi digestivi (Jacobs et al., 1960, Dubey, 1998), quali tripsina e pepsina e alle variazioni di
temperatura, rimanendo infettanti fino a 3 settimane nella carne tritata, mantenuta a temperatura
di refrigerazione; la maggior parte delle cisti tissutali è eliminata alla temperatura di -12, ma
alcuni ceppi di T. gondii resistono alla surgelazione e al congelamento (Grosklaus e
Baumgarten,1967, Kotula et al., 1991, Kuticic e Wikerhauser
1996, Dubey 2000). Alcuni studi hanno dimostrato come le cisti siano inattivate dalle consuete
procedure industriali di conservazione degli alimenti, come la salagione, dall’aggiunta di
saccarosio o l’affumicatura. Le oocisti sporulate, invece, sono molto resistenti nell’ambiente,
sopravvivono poco tempo alle basse temperature e alla disidratazione,ma resistono più di un
anno nel terreno o nella sabbia umida. Le oocisti, inoltre, sono impermeabili e resistono ai
disinfettanti comuni (Kuticic &e Wikerhauser, 1996, Frenkel, 2000). Nell’ambiente le oocisti
sono distribuite dal vento, dalla pioggia, dalle acque di superficie vertebrati coprofagi, ma si
possono comportare da vettori anche i vermi di terra, invertebrati coprofagi. L’importanza di
questa via di trasmissione per la sanità pubblica è evidenziato dall’alto tasso di sieroprevalenza

14
tra i vegetariani (24-47%) (Hall et al., 1999, Roghmann et al., 1999), che si infettano sia per
ingestione di verdure crude contaminate, sia nell’attività di giardinaggio (Cook et al., 2000). Uno
studio su donne gravide in Norvegia ha evidenziato la stretta connessione tra acquisizione della
parassitosi e l’assunzione di vegetali crudi e frutta non lavata (Kapperud et al., 1996).
L’infezione acquisita nell’uomo ha un decorso asintomatico e autolimitante; solo nel 10-20% dei
casi si manifesta con linfoadenopatia laterocervicale, ascellare e inguinale associata a febbre e
linfocitosi (Piergili Fioretti, 2004).
Nei soggetti immunocompetenti la parassitemia viene facilmente contrastata, ma le cisti possono
sopravvivere nei tessuti e, in seguito a compromissione del sistema immunitario, possono
riattivarsi come nel caso di pazienti affetti da HIV e pazienti sottoposti a trapianti d’organo; la
toxoplasmosi è, quindi, un problema rilevante di sanità pubblica per le categorie più deboli,
quali donne in gravidanza e soggetti immunodepressi colpiti da AIDS (Brandonisio 2006).
Nei pazienti colpiti da AIDS la toxoplasmosi rappresenta un’infezione secondaria , subdola e
mortale fino a qualche anno fa; con le nuove terapie antiretrovirali si è ridotta sia l’incidenza di
encefalite che della mortalità (Brandonisio 2006). Nei paesi con una bassa prevalenza di T.
gondii, la probabilità che una donna in gravidanza s’infetti per la prima volta è molto bassa, ma
se ciò avviene è rilevante il rischio di aborto o di infezione congenita; nei paesi con siero
prevalenza alta,al contrario, è molto probabile che una donna sia venuta a contatto con T. gondii
prima della gravidanza, eliminando ogni rischio di trasmissione al nascituro. Una sorveglianza
sulle donne a inizio gravidanza, con un semplice esame del sangue, soprattutto in queste zona ad
alta prevalenza, permetterebbe a molte donne di assumere comportamenti idonei a evitare la
trasmissione. Nella maggior parte dei paesi “sviluppati” è previsto uno screening prenatale sul
siero delle gestanti nei primi mesi di gravidanza, introdotto in Italia con il decreto ministeriale
del 1998 (G.U. 20/10/98), ma viene effettuato anche in Francia e l'Austria (Hill and Dubey,
2002).In Italia la percentuale di donne sieronegative in gravidanza è del 40%, quindi circa 600 su
1000 sono a rischio di contrarre l’infezione, l’infezione primaria attesa è di 1:10 su 1000
gravidanze (SNLG, 2010).
L’impatto sociale della toxoplasmosi è rilevante in tutto il mondo, sia per le sofferenze causate
direttamente dalla patologia, sia per i costi legati all’assistenza sanitaria delle persone colpite,
soprattutto bambini che presentano spesso i sintomi più importanti (il ritardo mentale e cecità) e i
malati di AIDS, nei quali la toxoplasmosi è l’infezione opportunistica più comune (Dubey et al.,
1998).
15
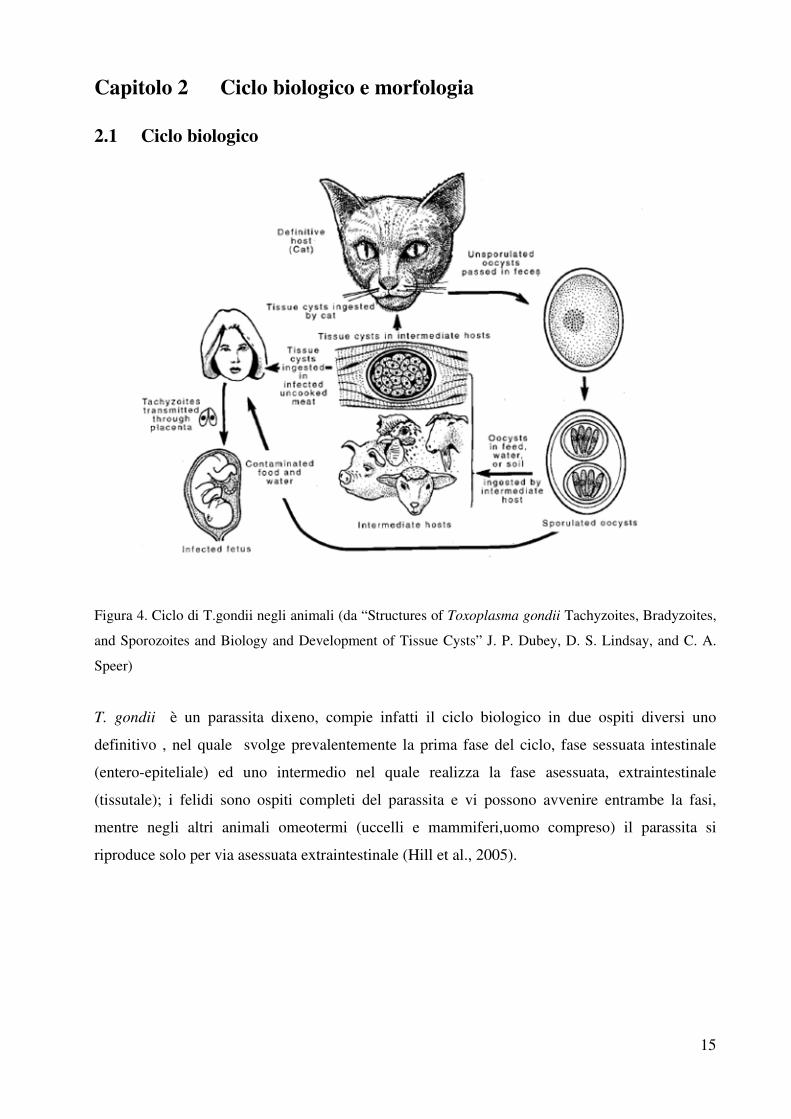
Capitolo 2 Ciclo biologico e morfologia
2.1 Ciclo biologico
Figura 4. Ciclo di T.gondii negli animali (da “Structures of Toxoplasma gondii Tachyzoites, Bradyzoites,
and Sporozoites and Biology and Development of Tissue Cysts” J. P. Dubey, D. S. Lindsay, and C. A.
Speer)
T. gondii è un parassita dixeno, compie infatti il ciclo biologico in due ospiti diversi uno
definitivo , nel quale svolge prevalentemente la prima fase del ciclo, fase sessuata intestinale
(entero-epiteliale) ed uno intermedio nel quale realizza la fase asessuata, extraintestinale
(tissutale); i felidi sono ospiti completi del parassita e vi possono avvenire entrambe la fasi,
mentre negli altri animali omeotermi (uccelli e mammiferi,uomo compreso) il parassita si
riproduce solo per via asessuata extraintestinale (Hill et al., 2005).

16
Ciclo intestinale
Nei felini T.gondii si comporta come un classico coccidio se l’infezione è contratta mediante
l’ingestione di ospiti intermedi contenenti cisti con bradizoiti e pseudocisti con tachizoiti. Altra
via di contagio è costituita dalle oocisti emesse con le feci e sporulate nell’ambiente. Il potere
infettante delle oocisti è molto inferiore alle altre forme (Dubey 2006); la via di trasmissione più
importante, infatti, è l’ingestione dei bradizoiti maturi contenuti nelle cisti,che causa il rilascio di
massive quantità di oocisti, molto più rilevanti rispetto alle altre vie di contagio (Urquhart et al.,
1998). Studi in materia rivelano che l’ingestione di cisti tissutali con bradizoiti da origine nel
97% dei casi a ulteriore emissione di oocisti; questa percentuale si abbassa notevolmente se il
contagio avviene tramite oocisti sporulate o tachizoiti (16-20% ) (Dubey, 1996 ). Dopo
l’ingestione di un ospite intermedio infetto, in genere un roditore, gli enzimi proteolitici dello
stomaco dell’ospite definitivo digeriscono la parete delle cisti tessutali ed i bradizoiti
raggiungono le cellule epiteliali del piccolo intestino. Alcuni bradizoiti penetrano la lamina
propria e si moltiplicano come tachizoiti, disseminandosi per via ematica o linfatica ai tessuti
extraintestinali del gatto; altri penetrano nelle cellule epiteliali e iniziano un ciclo schizogonico,
con varie generazioni di schizogonie fino allo stadio di schizonte (Dubey e Frenkel, 1972). Nello
schizonte si ha la formazione di numerosi merozoiti (4-32), fino alla rottura della cellula
intestinale che lo ospita . I merozoiti rilasciati invadono altri enterociti, dove iniziano nuovi cicli
schizogonici ; secondo un meccanismo non ancora conosciuto questi cicli si interrompono
quando alcuni merozoiti si differenziano in micro e macro gameti. Il microgamete, una volta
maturo, si stacca dal citoplasma residuo e appare come una struttura allungata con nucleo
anteriore,un mitocondrio e i corpi basali dei 2 flagelli ; questo gamete è detto maschile per la
forte similitudine con una cellula spermatica (Ferguson et al., 1974;Dubey et al., 1998). Il
macrogamete ha un’importanza determinante per la formazione dello zigote, perché sintetizza e
stocca gli elementi nutrizionali per la sporulazione nell’ambiente esterno e sostiene la
sopravvivenza degli sporozoiti fino ad un anno; sintetizza , inoltre, componenti necessari per la
formazione della parete. Il microgamete distrugge l’enterocita, entra nell’enterocita parassitato
dal gamete femminile e lo fertilizza, dando origine allo zigote. La nuova cellula, originata
dall’unione del macro e microgamete, ancora all’interno della cellula intestinale, e si circonda di
una parete cistica, diventando oocisti; la cellula che le accoglie si rompe, l’oocisti si riversa nel
lume intestinale ed è eliminata nell’ambiente esterno con le feci per circa 15 giorni. Il periodo di
prepatenza può essere breve (3-10giorni) se l’animale ingerisce cisti tissutali, lungo (18-49
giorni) se ingerisce oocisti sporulate (Dubey, 1996).Un gatto può emettere fino a 100 milioni di
oocisti,raramente però le emette dopo una seconda infezione per lo stabilirsi di una risposta

17
immunitaria; a conferma di ciò è stato osservato che in gatti trattati con trattamenti
immunosoppressori, per esempio alti dosaggi di corticosteroidi o per lungo periodo, si ha una
ripresa dell’emissione di oocisti (Lindsay, 1997). Alcuni studi hanno dimostrato che questo non
succede per malattie infettive come FIV e FeLV (Lappin et al.,1992,Lappin et al.,
1996,Svobodova et all 1998).
Nell’ambiente esterno l’oocisti sporula in 1-5 giorni ed ha all’interno 2 sporocisti con 4
sporozoiti ciascuna ed è infettante.
Nel caso in cui l’ospite definitivo ingerisca le oocisti sporulate si ha la liberazione nell’intestino
di sporozoiti che invadono le cellule della mucosa epiteliale del tenue, dove iniziano la fase
schizogonia asessuata dando origine allo schizonte nel digiuno e nell’ileo (Rinaldi, 2010).Il
ciclo, poi, prosegue come nel caso precedentemente osservato.
Ciclo extraintestinale
Avviene in mammiferi a sangue caldo, gatto compreso, o uccelli, che si infettano per ingestione
di oocisti mature, di tachizoiti o per il consumo di carni di altri animali con cisti terminali
contenenti i bradizoiti. E’ ospite intermedio anche il cane. Una volta entrato nel nuovo ospite,
Toxoplasma, passa la barriera intestinale e per via ematogena, veicolato da macrofagi e altre
cellule della serie bianca, invade le cellule di altri tessuti. Nelle cellule dei tessuti invasi si
formano dei vacuoli parassitofori, nei quali il protozoo si divide per endoduogenia, in due
tachizoiti avvolti dalla pellicola della cellula madre. Dopo circa 3-4 divisioni binarie si sono
formati 8-16 tachizoiti che determinano la rottura della cellula ospite; i tachizoiti riversati in
circolo invadono altre cellule e continuano a riprodursi oppure vengono inglobati da macrofagi e
diffondono a tutto l’organismo. Questa fase acuta dura circa 1-2 settimane, in genere senza una
sintomatologia importante, dopodiche l’ospite acquisisce un’immunità,con la produzione di IgM
che limitano l’invasività e il numero dei tachizoiti localizzati nel tessuto nervoso e muscolare.
In queste sedi i tachizoiti si riproducono per endoduogenia, ma molto più lentamente per sfuggire
all’azione anticorpale, rispetto alla precedente divisione, formando nella cellula madre, qualche
centinaio di bradizoiti (forma simile ai tachizoiti ma di minori dimensioni). I bradizoiti hanno
una bassa capacità di moltiplicazione e restano quiescenti nelle cisti,che hanno sedi preferenziali
di localizzazione , cioè la muscolatura scheletrica, cardiaca, l’ occhio, il sistema nervoso
centrale, raramente il polmone e il fegato (Rinaldi, 2010). I bradizoiti vi rimangono vivi e vitali
per tutta la vita dell’ospite, inattaccabili dalle difese dell’organismo (Dubey et al., 1998). La
formazione della cisti è legata all’equilibrio tra parassita e sistema immunitario dell’ospite ; l
‘infezione decorre senza comparsa dei sintomi clinici. Le cisti tissutali periodicamente si

18
rompono liberando bradizoiti che vengono immediatamente circondati da un involucro cistico.
Un’eventuale caduta delle difese immunitarie riacutizza l’infezione, con attivazione delle forme
quiescenti che si trasformano in tachizoiti e che portano alla formazione di nuove pseudocisti.
Se l’ospite definitivo ingerisce cisti presenti nei tessuti di prede infette i bradizoiti si liberano,
invadono il piccolo intestino, si differenziano in tachizoiti e scatenano la fase acuta in un nuovo
individuo.
2.2 Morfologia
Gli stadi infettanti di T.gondii sono tre:
• Tachizoiti (in gruppo o cloni nei tessuti dell’ospite intermedio)
• Bradizoiti (in cisti tessutali)
• Sporozoiti (in oocisti presenti nell’ambiente)
( Dubey et al., 1998)
Tachizoiti
Il termine tachizoita significa “rapido”, dal greco “tacus”, fu coniato da Frenkel per definire il
veloce moltiplicarsi in ogni cellula dell’ ospite intermedio e nell’epitelio intestinale nell’opite
definitivo. Questa struttura veniva definita precedentemente con il termine “trofozoita”, dal
greco “Trophos” alimentazione; aggregati di più tachizoiti sono definiti cloni, colonie terminali
o gruppi. I tachizoiti presentano una forma a mezzaluna e larga circa 2 micron e lunga 6, con
una parte anteriore conoidale appuntita ed una terminale arrotondata.
Ultrastrutturalmente presenta vari organelli e corpi inclusi rivestiti da una pellicola esterna:
• Anelli apicali
• Anelli polari
• Conoide
• Rostri
• Micronemi
• Micropori
• Mitocondri
• Microtubuli sottopellicolari
• Reticolo endoplasmatico liscio e rugoso
• Apparato del Golgi
• Ribosomi
• Granuli densi+granuli di amilopectina (possono non essere presenti)

19
• Plastide legato alla membrana chiamato “apicoplasto”
(De Melo et al., 1997; De Souza et al., 1978,Kohl et al., 1997; Sheffield et al., 1968,Vivier et al.,
1972)
Figura 5. Schematizzazione di tachizoita (a sx) e bradizoita (a dx) Dubey JP, Lindsay DS, and Speer CA
(1998). Structure of Toxoplasma gondii Tachyzoites, Bradyziotes, and Sporozoites and Biology and
Development of Tissue Cyst. Clinical Microbiology Reviews, Apr. 1998, p. 267 -299).
Il nucleo è generalmente situato nella parte centrale della cellula, contiene ciuffi di cromatina e
un nucleolo ben definito.
Il rivestimento è costituito da tre membrane, il plasmalemma e due membrane strettamente
adese, che formano la membrana interna, caratterizzata da una maglia di vescicole piatte
(Morrissette et al., 1997; Porchet-Hennere et al., 1985). La membrana interna è discontinua sopra
l’anello polare, ai micropori, a cui è posta lateralmente e alla punta posteriore dello zoita.
L’anello polare è un ispessimento denso del complesso della membrana interna nella parte
anteriore del tachizoita, il quale circonda una struttura a tronco di cono, detta “conoide” che
consiste in 6-8 elementi micro tubulari, ripiegati come una molla compressa. 22 microtubuli
sottopellicolari originati dal secondo anello polare, corrono longitudinalmente quasi per tutta la
lunghezza della cellula, proprio sotto il complesso della membrana interna, mentre 2 microtubuli
interni terminano nel conoide.

20
I microtubuli sono come una gabbia costale e sono organizzati a spirale; singoli microtubuli
hanno strie trasverse prominenti. Nello spazio compreso tra la parte anteriore e il nucleolo ci
sono 8-10 organelli a forma di clava(Sulzer et al., 1974), chiamati “rhoptries”, che sono strutture
escretorie con uno stretto collo anteriormente che si estende all’interno del conoide ed una parte
posteriore sacciforme, spesso intricata. I Micronemi sono delle strutture presenti
prevalentemente nella parte anteriore del parassita.
Figura 6. Complesso apicale del tachizoita.(da Dubey JP, Lindsay DS, and Speer CA 1998, Structure of
Toxoplasma gondii Tachyzoites, Bradyziotes, and Sporozoites and Biology and Development of Tissue
Cyst. Clinical Microbiology Reviews, Apr. 1998, p. 267 -299).
Il tachizoita si può muovere per scivolamento, flettendosi, ondulando o ruotando su se stesso ma
non hanno visibili mezzi di locomozione come come ciglia, flagelli o pseudopodi. Le funzioni di
alcune strutture, come i conoidi, i roptrii e i micronemi, non sono ancora conosciute ma si pensa
che abbiano un ruolo nella penetrazione della cellula ospite e nella creazione di un ambiente
intracellulare adatto alla crescita e allo sviluppo del parassita. Il conoide può ruotare, inclinarsi,
allungarsi e retrarsi in modo che il parassita possa esplorare prima della penetrazione il
plasmalemma della cellula ospite (Chiappino,et al., 1984). I roptrii hanno una funzione secretoria
associata alla penetrazione poiché riversano il loro secreto ricco di enzimi proteolitici (Saffer et
al., 1992) attraverso il plasma lemma (Nichols et al., 1983). I micropori sono strutture simili ai
citosomi, formati dall’invaginamento della membrana interna( Nichols,et al., 1994).

21
I tachizoiti penetrano nella cellula sia attivamente, attraverso la membrana oppure attraverso
fagocitosi; subito dopo cambia forma e diventa ovoidale ed è circondato di vacuolo parassito
foro, che sembra derivare sia dal parassita che dall’ospite.
Immediatamente dopo la penetrazione una rete membranosa tubulo vescicolare si sviluppa
all’interno del vacuolo parassito foro e sembra originare dalla terminazione posteriore del
tachizoita (Bonhomme et al., 1982; Dubremetz et al., 1993; Joiner, 1993; Morisaki, 1995;
Nichols, 1981; Sibley, 1995; Silva, 1982; Speer, 1997; Werk, 1995). Il tachozoita comincia a
riprodursi asessualmente nella cellula ospite per endoduogenia, forma specializzata di
replicazione in cui due progenie si formano all’interno della cellula madre parassita. L’apparato
di Golgi si duplica per primo e si formano due complessi alla terminazione anteriore del
nucleolo. Successivamente la porzione anteriore del complesso di membrane, i microtubuli
sottopellicolari delle cellule figlie appaiono a forma di cupola; il nucleolo assume la forma di
ferro di cavallo. Il complesso di membrane interne e i microtubuli sottopellicolari continuano a
estendersi posteriormente e circondano parte del nucleolo che si può dividere in due. Le cellule
figlie continuano a crescere fino a raggiungere la superficie delle cellule parentali, le quali
perdono il complesso di membrane interne e la membrana interna diventa il plasma lemma delle
cellule figlie. Quando la cellula ospite non può più sostenere la crescita si rompe e libera
tachizoiti che a loro volta invadono altre cellule; i tassi di invasione e crescita dipendono dal
ceppo del parassita e dalla cellula ospite (Appleford, 1997; Kaufman, 1962). Dopo la
penetrazione del tachizoita nella cellula ospite c’è un periodo di arresto prima che il parassita
inizi la divisione e questa fase è in parte, dipendente dal parassita.
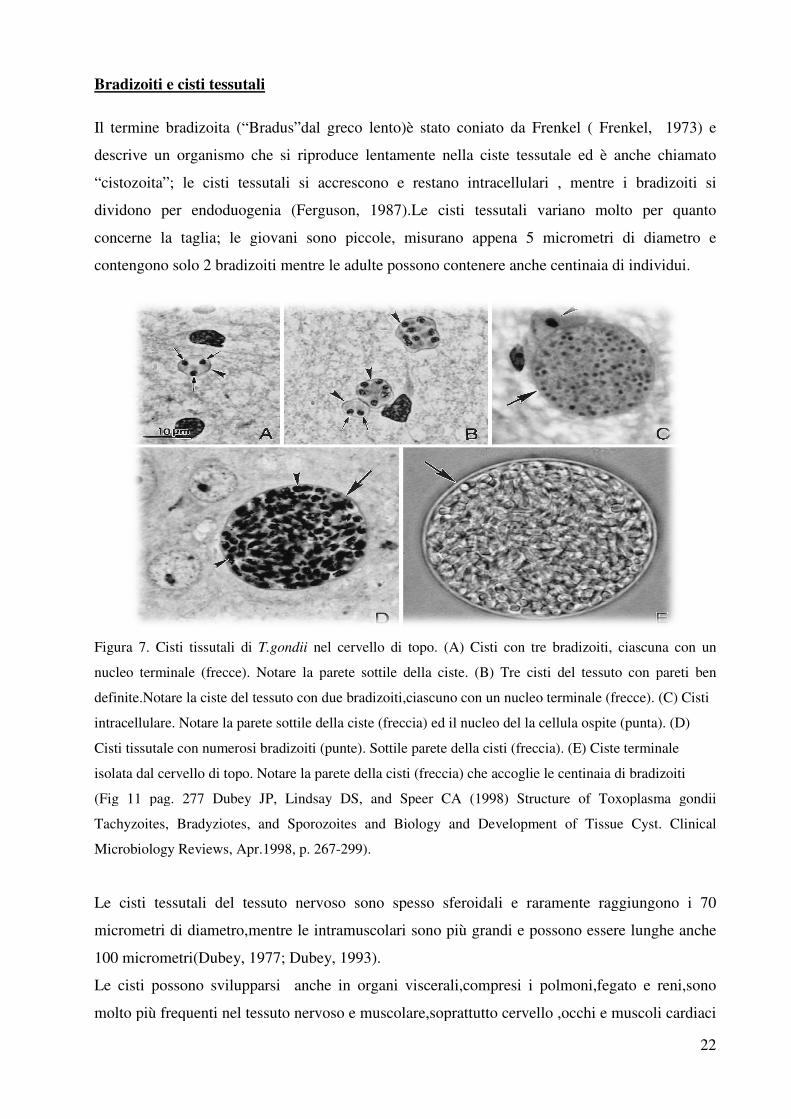
Bradizoiti e cisti tessutali
Il termine bradizoita (“Bradus”dal greco lento)è stato coniato da
descrive un organismo che si riproduce lentamente nella ciste tessutale ed è anche chiamato
“cistozoita”; le cisti tessutali si accrescono e restano intracellulari ,
dividono per endoduogenia (
concerne la taglia; le giovani sono piccole,
contengono solo 2 bradizoiti mentre le adulte possono contenere anche centinaia di individui.
Figura 7. Cisti tissutali di T.gondii
nucleo terminale (frecce). Notare la pa
definite.Notare la ciste del tessuto con
intracellulare. Notare la parete sottile della ciste (freccia) ed il nucleo del la cellula ospite (punta). (D)
Cisti tissutale con numerosi bradizoiti (punte). Sottile parete della cisti (
isolata dal cervello di topo. Notare la parete della cisti (freccia) che accoglie le centinaia di bradizoiti
(Fig 11 pag. 277 Dubey JP, Lindsay DS, and Speer CA (1998) Structure of Toxoplasma gondii
Tachyzoites, Bradyziotes, and Sporozoites and Biology and Development of Tissue Cyst. Clinical
Microbiology Reviews, Apr.1998, p. 267
Le cisti tessutali del tessuto nervoso sono spesso sferoidali e raramente raggiungono i 70
micrometri di diametro,mentre le intramuscolari son
100 micrometri(Dubey, 1977;
Le cisti possono svilupparsi anche in organi viscerali,compresi i polmoni,fegato e reni,sono
molto più frequenti nel tessuto nervoso e muscolare,soprattutto cervello ,occ
Il termine bradizoita (“Bradus”dal greco lento)è stato coniato da Frenkel
descrive un organismo che si riproduce lentamente nella ciste tessutale ed è anche chiamato
le cisti tessutali si accrescono e restano intracellulari , mentre i bradizoiti si
dividono per endoduogenia (Ferguson, 1987).Le cisti tessutali variano molto per quanto
le giovani sono piccole, misurano appena 5 micrometri di diametro
contengono solo 2 bradizoiti mentre le adulte possono contenere anche centinaia di individui.
T.gondii nel cervello di topo. (A) Cisti con tre bradizoiti, ciascuna con un
erminale (frecce). Notare la parete sottile della ciste. (B) Tre cisti del tessuto con pareti ben
definite.Notare la ciste del tessuto con due bradizoiti,ciascuno con un nucleo terminale (frecce). (C) Cisti
intracellulare. Notare la parete sottile della ciste (freccia) ed il nucleo del la cellula ospite (punta). (D)
Cisti tissutale con numerosi bradizoiti (punte). Sottile parete della cisti (freccia). (E) Ciste terminale
isolata dal cervello di topo. Notare la parete della cisti (freccia) che accoglie le centinaia di bradizoiti
(Fig 11 pag. 277 Dubey JP, Lindsay DS, and Speer CA (1998) Structure of Toxoplasma gondii
nd Sporozoites and Biology and Development of Tissue Cyst. Clinical
Microbiology Reviews, Apr.1998, p. 267-299).
Le cisti tessutali del tessuto nervoso sono spesso sferoidali e raramente raggiungono i 70
micrometri di diametro,mentre le intramuscolari sono più grandi e possono essere lunghe anche
Dubey, 1993).
Le cisti possono svilupparsi anche in organi viscerali,compresi i polmoni,fegato e reni,sono
molto più frequenti nel tessuto nervoso e muscolare,soprattutto cervello ,occ
22
Frenkel ( Frenkel, 1973) e
descrive un organismo che si riproduce lentamente nella ciste tessutale ed è anche chiamato
mentre i bradizoiti si
1987).Le cisti tessutali variano molto per quanto
5 micrometri di diametro e
contengono solo 2 bradizoiti mentre le adulte possono contenere anche centinaia di individui.
nel cervello di topo. (A) Cisti con tre bradizoiti, ciascuna con un
rete sottile della ciste. (B) Tre cisti del tessuto con pareti ben
due bradizoiti,ciascuno con un nucleo terminale (frecce). (C) Cisti
intracellulare. Notare la parete sottile della ciste (freccia) ed il nucleo del la cellula ospite (punta). (D)
freccia). (E) Ciste terminale
isolata dal cervello di topo. Notare la parete della cisti (freccia) che accoglie le centinaia di bradizoiti
(Fig 11 pag. 277 Dubey JP, Lindsay DS, and Speer CA (1998) Structure of Toxoplasma gondii
nd Sporozoites and Biology and Development of Tissue Cyst. Clinical
Le cisti tessutali del tessuto nervoso sono spesso sferoidali e raramente raggiungono i 70
o più grandi e possono essere lunghe anche
Le cisti possono svilupparsi anche in organi viscerali,compresi i polmoni,fegato e reni,sono
molto più frequenti nel tessuto nervoso e muscolare,soprattutto cervello ,occhi e muscoli cardiaci

23
e scheletrici. Le cisti intatte non causano danni e persistono per tutta la vita dell’ospite senza
causare alcuna risposta infiammatoria. La parete della ciste è elastica e sottile,circa 0.5 micron
contiene centinaia di bradizoiti falcifomi, ciascuno misura dai 7 a 1,5 micron(Melhorn e
Frenkel, 1980). La mancanza di colorazione con l’impregnazione argentica indica che la parte
della ciste non contiene glicogeno ne altri polisaccaridi: è composta da elementi dell’ospite e del
parassita (Ferguson e Hutchison, 1987; Ferguson e Hutchison, 1987; Sim et al., 1988).
La parete della ciste è rivestita da materiale granulare che riempie anche lo spazio tra i
bradizoiti. Alcuni bradizoiti degenerano, specialmente nelle cisti di vecchia data ( Pavesio et al.,
1992).
I bradizoiti strutturalmente sono molto simili ai tachizoiti, il nucleo situato verso l’estremità
posteriore mentre nei secondi è localizzato più centralmente. Il contenuto dei roptrii è labirintico
nei tachizoiti, mentre nei bradizoiti elettrondenso; in quest’ultimo caso varia molto in base
all’età della cisti tessutale: se è giovane i roptrii sono labirintici, se di vecchia data sono
elettrondensi. Inoltre molti bradizoiti hanno da 1 a 3 roptrii che sono collegati tra loro. I
bradizoiti contengono granuli di amilopectina, che possono essere presenti o assenti nei
tachizoiti. I bradizoiti sono più sottili dei tachizoiti e sono perciò più sensibili agli enzimi
proteolitici (Jacobs, Remington e Melton, 1960); da ciò deriva che il periodo prepatente nel gatto
dopo l’ingestione dei bradizoiti è molto più corto di quello dopo ingestione di tachizoiti ( Dubey
e Frenkel., 1976).

24
Oocisti
Oocisti non sporulate sono di forma sferica o subsferica, dai 10 ai 12 micrometri di diametro. In
microscopia chiara la parete dell’oocisti è costituita da 2 strati incolori,i granuli polari sono
assenti e lo sporonte riempie tutta l’oociste. La sporulazione avviene all’esterno in 1-5 giorni a
seconda della temperatura e della tensione di ossigeno.
Figura 8. A.oocisti non sporulata ,la massa centrale è lo sporonte . B.oocisti sporulata con 2 sporocisti, nei
quali sono visibili 4 sporozoiti. Oocisti di toxoplasma gondii. (A) Oocisti non sporulata. (B)Oocisti
sporulata con due sporocisti, e 4 sporozoiti visibili solo in una sporociste. (C) Micro scopia elettronica di
una oocisti sporulata. Notare la parete sottile del oocisti (grande freccia), due sporocisti e gli sporozoiti, di
cui uno in sezione trasversale ed uno in sezione longitudinale(piccole frecce).Dubey JP, Lindsay DS, and
Speer CA (1998) Structure of Toxoplasma gondii Tachyzoites, Bradyziotes, and Sporozoites and Biology
and Development of Tissue Cyst. Clinical Microbiology Reviews, Apr. 1998, p. 267 -299).
Le oocisti sporulate sono sub sferiche o ellissoidali, di11-13 micron di diametro ed ognuna
contiene e due sporocisti ellissoidali di 6-8 micron, senza corpi di Stieda; sono presenti residui
delle sporocisti ma non delle oocisti. Ogni sporociste contiene 4 sporozoiti. Il citoplasma delle
oocisti non sporulate (zigote) ha un grosso nucleo con nucleoplasmi amorfi e un nucleolo
distinto. Lo zigote è delimitato da un'unica membrana con pochi micropori, il nucleo si divide 2
volte ,dando origine a 4 nuclei, localizzati alla periferia dello zigote; si forma in questo stadio la
seconda membrana e dopo la divisione citoplasmatica si formano 2 sporoblasti sferici ognuno
con 2 nuclei (Ferguson, 1979).
A questo punto gli sporoblasti si allungano formando così le sporocisti; le due membrane esterne
degli sporoblasti diventano lo strato esterno della parete della sporocisti e il plasmalemma della
massa citoplasmatica si trasforma nello strato interno(Ferguson, 1979). Infine 4 placche arcuate

25
formano lo strato interno della sporociste; le placche sono unite da 4 suture che formano nel
punto di giunzione, delle depressioni sulla superficie della sporociste. La formazione dello
sporozoita inizia quando due placche dense appaiono a entrambe le estremità della sporociste;
ogni nucleo si divide in due ed è incorporato nella nuova struttura che si sta formando,ogni
sporociste contiene 4 sporozoiti. Dopo che lo sporozoita si è formato restano una serie di corpi
residuali all’interno della membrana (Ferguson, 1979).Ultrastrutturalmente la parete dell’oociste
sporulata consiste in 3 strati:
• Strato elettrondenso esterno;
• Strato elettrolucente medio;
• Strato moderatamente elettrodenso interno;
Lo strato mediano è costituito da resti delle membrane rimasti tra le superfici interne ed esterne
durante la formazione della parete dell’oociti (Speer et al., 1997). Il trattamento con sodio
ipoclorito (Clorox) al 1,3% rimuove lo strato interno.
La parete dell’oociste contiene un micropilo, piccolo, di circa 350 nanometri, presente sulla
parete stessa senza una precisa localizzazione; il micropilo è costituito da 3 strati che si
continuano con quelli dell’oociste; tuttavia lo strato esterno del micropilo è sottile e
moderatamente elettrodenso, mentre quello interno elettrodenso, discoidale e leggermente più
spesso rispetto allo strato interno della parete dell’oociste.
La funzione micropilo non è conosciuta, forse può rappresentare un punto di permeabilità della
parete dell’oociste che diventa suscettibile all’azione della CO2 vari enzimi, che consente
l’entrata dei Sali biliari e della tripsina, che provoca la disincistazione dello sporozoita.
Ricapitolando la parete della sporociste consiste in 2 strati ben distinti,uno esterno sottile ed
elettrondenso ed un altro interno, spesso e moderatamente elettrondenso, costituito da 4 placche
arcuate (Christie, 1978; Ferguson, 1979).
Durante la disincistazione, in presenza di sali biliari e tripsina, la sporocisti si rompe
improvvisamente con la fuoriuscita degli sporozoiti. I residui della sporociste sono granuli di
amilopectina e corpi lipidici. Ultrastutturalmente lo sporozoita e il tachizoita sono molto simili,
differiscono solo per il fatto che nel primo ci sono molti micronemi, roptrii e granuli di
amilopectina.
Gli sporozoiti misurano dai 2 ai 6-8 micron con nucleo terminale, non presentano corpi
cristalloidi o rifrangenti.

26
2.3 Genotipi
In passato, studi molecolari e di biologia delle popolazioni, avevano ipotizzato che T.gondii
avesse tre ceppi clonali diversi per virulenza e patogenicità, (Sibley e Boothroyd, 1992; Howe e
Sibley, 1995). Negli ultimi anni sono stati isolati ceppi diversi sia in uomini malati di AIDS che
in soggetti immunocompetenti nella Guyana francese, affetti da grave sintomatologia
(Ajzenberg et al., 2004; Demar et al.,2008); negli animali questi ceppi sono stati isolati nei suini
in Brasile (Belfort-Neto et al., 2007; Ferreira et al., 2008), nelle otarie in California centrale
(Miller et al., 2004; Conrad et al., 2005). Analizzando campioni prelevati sia da ospiti
apparentemente sani sia da quelli malati, i positivi sono stati classificati secondo tre tipi genetici
(I, II e III) in base al polimorfismo di lunghezza dei frammenti di restrizione (RFLP) (Howe e
Sibley, 1995). Grazie a nuovi marcatori per la caratterizzazione genetica è stata rivelata una
variabilità genetica molto più elevata rispetto a quello che si pensava (Ajzenberg et al., 2002;
Ajzenberg et al., 2004; Khan et al., 2006; Lehmann et al., 2006).I tipi clonali I, II e III hanno
rilevanti differenze per quanto riguarda la virulenza: il tipo clonale I sembra causare infezioni
letali nei topi “outbred”, mentre i tipi clonali II e III sono significativamente meno virulenti
(Sibley e Boothroyd, 1992).
In Asia e in Sud-America sembrano essere presenti genotipi atipici (Dubey et al., 2007, 2008;
Lehmann et al., 2006), molto virulenti sia nei topi (Pena et al., 2008)che in uomini adulti
immunocompetenti(Carme et al., 2009).
In California sono stati isolati due nuovi ceppi ( A-XX) nelle otarie (Sundar et al., 2009.), ma
anche in altre specie che vivono sulla costa, come i bivalvi e altri invertebrati filtratori. Studi
genetici hanno evidenziato che alcune proteine dei “rhoptries” che vengono rilasciate al
momento della penetrazione nelle cellule ospite (ad esempio ROP18) sono collegate con la
virulenza nei vari ceppi genetici (Yuan et al., 2010); altre analisi genetiche condotte su nuovi loci
variabili sembrano suggerire un’evoluzione distinta di ceppi sudamericani ed eurasiatici fino a
tempi relativamente recenti. Il trasporto di animali, soprattutto gatti, ratti e topi, sulle navi che
trasportavano gli schiavi avrebbe fornito ulteriori possibilità di migrazione di ceppi distinti da un
continente all’altro ( Lehmann et al., 2006).

27
Capitolo 3 Modalità di trasmissione
La trasmissione di T.gondii può essere sia orizzontale che verticale,ma ancora non si conosce
quale via sia la più frequente; è noto il ruolo fondamentale del gatto e dei felidi selvatici nel
mantenimento del parassita nell’ambiente (Dubey et al., 1997). L’infezione segue quattro vie di
trasmissione:
a) ORIZZONTALE: ingestione di oocisti sporulate dall’ambiente;
b) ORIZZONTALE: ingestione di cisti tessutali contenute in carne cruda o poco
cotta o da frattaglie primarie (visceri) di ospiti intermedi;
c) VERTICALE: trasmissione transplacentare dei tachizoiti (Dubey et al., 1998;
Dubey 1991; dubey 1991; jackson e hutchison 1989; Dubey e Beattie 1988)
d) TRAMITE SECRETI (latte) ed escreti (saliva,urine)
3.1 Ingestione oocisti sporulate
Gli ospiti intermedi ,uccelli compresi, possono infettarsi ingerendo direttamente le oocisti
sporulate presenti nell’ambiente esterno come nei pascoli , nel fieno, nel grano e nel grano
contaminato ( Buxton,1990). Il gatto può infettarsi per assunzione di oocisti rilasciate da altri
felini. Gli insetti, mosche e blatte soprattutto, possono essere vettori di trasporto meccanico delle
oocisti sulle zampe o parti dell’apparato boccale (Hartley e Munday 1974,Wallace 1972-1973-
1971).
La trasmissione mediante oocisti è più adatta per l’infezione di ospiti intermedi, mentre nel gatto,
che è un predatore, è più frequente la trasmissione attraverso bradizoiti (Frenkel et al., 1970;
Dubey, 2001). Meno della metà dei gatti alimentati con oocisti emettono a loro volta oocisti; il
preriodo prepatente dopo l’ingestione di oocisti , inoltre, è molto lungo e non completamente
conosciuto (Dubey, 2005).
3.2 Ingestione di bradizoiti
Il cane,così come tutti i carnivori e gli uccelli predatori,può infettarsi per ingestione di prede che
contengono nei loro tessuti cisti (Dubey, 1986;Rothe et al., 1985). Questa via di trasmissione è
molto importante anche nell’uomo, poiché le cisti tessutali sono frequenti in animali da carne
quali i suini, ovini e caprini, meno in avicoli, cunicoli e equini (Tenter et al., 2000). Non sono
escluse però anche carni di piccioni, lepri, cinghiali, cervi e canguri mentre i bovini hanno un
sistema immunitario tale per cui il pericolo di contrarre la malattia è ridotto (Innes,1997) .Il

28
rischio sussiste se le carni sono crude o poco cotte, poiché una cottura a 70°per 10 minuti
distrugge gli organismi, mentre non è sufficiente la cottura “al sangue”(Dubey, 1986).
3.3 Ingestione di tachizoiti
Evenienze più rare sono l’infezione mediante il trasferimento di tachizoiti dalla madre alla prole
con il latte (Powell et al., 2001; Tender, 2000; Jackson e Hutchison, 1989; Dubey e Beattie,
1988).
Un’altra forma di trasmissione è quella attraverso il trapianto di organi quali il cuore, fegato,
reni, midollo osseo e trasfusioni di sangue e liquidi corporei. L’infezione può insorgere per
impianto di organi da un donatore infetto in un soggetto non immune o immunocompromesso
oppure per induzione di malattia in un soggetto immucompromesso con infezione latente;
sembra infatti che le fonti d’infezione siano sia le cisti tissutali presenti nei tessuti, sia l’infezione
latente nei pazienti che son subiscono l’impianto. In entrambi i casi viene somministrata una
forte terapia immunosoppressiva, per ridurre il rigetto molto frequente in questi casi, ma questo
favorisce la riattivazione dell’infezione e l’insorgenza della malattia (D.Hill et al.,2002) .
3.4 Trasmissione transplacentare
L’infezione si trasmette dalla madre, priva di anticorpi anti-toxoplasma, al feto, prevalentemente
nell’uomo, nella pecora e nella capra (Dubey et al., 1981; Masala et al., 2007). Negli ovini
l’infezione è molto importante poiché causa un’elevata percentuale di aborti nei greggi e quindi
determina importanti perdite economiche (Dubey et Beattie, 1988). L’infezione nelle
popolazioni ovine varia con gradi diversi per i singoli greggi, oscilla tra 5% e 95% (Blewett,
1983). In seguito all’infezione , infatti,i tachizoiti sono attratti dagli ormoni, possono
moltiplicarsi nella placenta e nel feto (Cook et al., 2000; Tender et al.,2000) e causare aborto,
morte neonatale, anomalie fetali quali cecità e ritardo mentale (Dubey e Beattle, 1988). Il 20-
50% delle donne infette trasmetterà l’infezione al feto (Jones et al., 2001), ma il rischio aumenta
con il progredire della gestazione (Hohlfeld et al., 1989; Jenum et al., 1998; Dunn et al., 1999;
Foulon et al., 1999; Remington et al., 2005), mentre i sintomi sono più gravi quanto più
l’infezione è ad uno stadio precoce, che corrisponde alla fase di embriogenesi (Hohlfeld et
al.1989; Dunn et al., 1999; Montoya e Liesenfeld, 2004). Studi recenti hanno evidenziato
l’importanza della trasmissione congenita, che in un gregge di pecore avviene nel 61% delle
gravidanze (Duncanson et al., 2001) e hanno rivalutato l’importanza della trasmissione verticale
nel mantenimento del parassita negli ospiti intermedi (Owen e trees, 1998;de Roever –Bonnet,
1969; Remington et al., 1961; Beverly, 1959). La trasmissione congenita può raramente avvenire

29
anche nei felidi, ma la gravità dell’infezione varia a seconda della fase di gravidanza al momento
dell’infezione: più è precoce più saranno evidenti i sintomi. Gattini nati da madri infettate in
gravidanza possono contrarre l’infezione per via transplacentare o trans lattea (Dubey et
al.,1995; Dubey e Carpenter, 1993; Sato e al., 1993). L’infezione transplacentare è molto più
grave rispetto a quella trans lattea, poiché la prima può portare a feti nati morti, mortalità
presvezzamento, la seconda invece causa toxoplasmosi acuta ma ha un’incidenza di mortalità
molto inferiore (Dubey et al.,1995; Dubey e Carpenter, 1993).
3.5 Modalità di trasmissione delle oocisti in ambiente acquatico
Recenti studi sulla Toxoplasmosi hanno evidenziato l’importanza della trasmissione
dell’infezione attraverso l’ambiente acquatico, sia marino che d’acqua dolce; il campanello
d’allarme è stato il ritrovamento di mammiferi marini positivi o morti per toxoplasma e s ‘è
scoperto che toxoplasma è un importante patogeno dei mammiferi marini delle zone costiere
(Lindsay e Dubey, 2009). Le principali fonti d’infezione sono le fognature e gli scarichi dei
liquami che possono contenere anche la sabbia delle letti eredi gatti domestici e che sono portate
dalle correnti fluviali al mare (Feyer et al.,2004). Negli estuari si concentra la maggior parte
delle oocisti, questo è dimostrato dall’elevato numero di lontre della California decedute in
seguito all’ingestione di queste (Miller et al., 2002). Un fattore importante per la contaminazione
delle acque con oocisti di felidi selvatici è il dilavamento dei terreni ad opera delle piogge
torrenziali (Miller et al., 2008). Anche l’ingestione casuale di acqua dolce di laghi o fiumi o
salata del mare e comunque acqua non trattata può causare infezione nell’uomo ma anche nel
cane e tutti gli animali che si abbeverano. Le oocisti nell’acqua marina artificiale sporulano in 3
giorni, a concentrazione di 15 ppm e 32 ppm di NaCl a 24°C, restano infettanti per 6 mesi a 22-
24°C (Lindsay et al.,2003), mentre a 4°anche 24 mesi (Lindsay e Dubey, 2009). La dose di
acqua necessaria a trasmettere l’infezione non è nota, i delfini, per esempio, assumono solo poca
acqua mentre il resto della componente idrica è assunta con il cibo, prevalentemente cefalopodi
e pesci che comunque non sono noti portatori di Toxoplasma (Dubey et al., 2003; Jones et
Dubey, 2009). Recentemente è stato dimostrato che le oocisti di T.gondii possono sporulare in
acqua marina e restare infettanti per i topi fino a 6 mesi (Lindsay and Dubey, 2009); le ostriche
della specie “Crassostrea virginica” rilasciano oocisti in acqua salata fino a 21 giorni post–
infezione , che rimagono infettanti (Lindsay et al., 2001; Arkush et al., 2003; Lindsay et al.,
2004).
Un altro elemento fondamentale da prendere in considerazione è il ruolo di ospiti paratenici dei
molluschi bivalvi. Secondo prove sperimentali le cozze (“Mytilus galloprovincialis”) e le

30
ostriche (“C. virginica”) possono assumente oocisti dall’acqua marina e concentrarle
nell’appartato digerente e restare infettanti per alcuni mesi (Lindsay et al., 2004); questa ipotesi è
stata confermata dal ritrovamento mitili del genere “Mytilus californianus”, nell’estuario della
baia di Monterey, in Calfornia, positiva per il ceppo X di Toxoplasma (Miller et al., 2008),
proprio quello che infetta le lontre della stessa zona (Miller et al., 2003; Conrad et al., 2005). I
molluschi bivalvi , se consumati crudi sono un pericolo per l’uomo ma anche per gli animali. Le
ostriche sono tra i molluschi più apprezzati nel mondo e tradizionalmente mangiati crudi, poco
cotti o cotti al vapore, quindi a rischio trasmissione per l’uomo ( Putignani et al.,2010).

31
Capitolo 4 Immunità anti Toxoplasma gondii e patogenesi
4.1 Immunità anti T.gondii
Numerosi studi hanno rivelato che il controllo di T.gondii richiede la precoce produzione di
Citochine IL-12, le quali stimolano le Natural Killer (NK) e i CD4+ e CD8+ Cell T a rilasciare
INF gamma. Sebbene la cellula infetta liberi linfociti citotossici per il parassita, il ruolo
principale di queste cellule è la produzione di INF gamma, il quale è determinante per una serie
di meccanismi antimicrobici, sia in cellule emopoietiche che non,che limitano la moltiplicazione
del parassita. In particolare INF gamma altera il metabolismo cellulare che trasforma il
triptofano in fibroblasto e che trasporta il ferro alle cellule intestinali. Queste citochine stimolano
i fagociti a produrre ossido di azoto che danneggia T.gondii e ne impedisce la crescita nei
macrofagi. Alcuni studi effettuati sui topi, hanno rivelato l’esistenza di un meccanismo di
resistenza al INF gamma dipendente, ma iNOS (ossido nitrico sintetasi) indipendente. Questo
ha rivelato che il GPTa indotto dall ‘INF gamma ha un’influenza importante nell’eliminazione
del parassita; il meccanismo non è definito, ma sembra che le GTPasi comportino la rottura del
vacuolo contenente il parassita e la sua digestione nel citosol. Tra le GTPasi è compresa la p67
cioè la GBPs, che è stato recentemente scoperto essere molto importante nel controllo nel topo.
L’infezione delle cellule è associata a una ridotta espressione del complesso maggiore di
istocompatibilità(MHC). Nonostante questo meccanismo, l’infezione con il ceppo 2 porta
all’attivazione e espansione del DCs e ad una risposta cellulare importante(CD8+). L’infezione
con il ceppo 1,invece,induce una debole risposta(Christopher A. Hunter,2012).

32
4.2 Patogenesi
Il tipo e la gravità della malattia clinica dipendono da diversi fattori, sia dell’ospite (età, sesso,
specie) ma anche del parassita (numero di organismi, fase del parassita ingerito, il ceppo). Alcuni
tipi di T.gondii, infatti potrebbero avere affinità tessutale; per esempio alcuni ceppi sembrano
avere una maggior probabilità di essere associati a malattie oculari nel gatto (Powell e Lappin,
2001). Gli elementi più importanti che concorrono alla toxoplasmosi clinica sono
l’immunosoppressione e le malattie concomitanti,poichè se la risposta immunitaria dopo
un’infezione primaria è debole,si verifica la riproduzione massiccia dei tachizoiti con necrosi
tessutale (Dubey e Lappin 2006; Dubey et al , 2009). Questo meccanismo è probabile che si
verifichi anche nei gatti affetti da toxoplasmosi cronica che vanno incontro a
immunosoppressione. Il gatto affetto da FeLV, FIV o FIP è molto più suscettibile sia
all’infezione primaria, che alla riattivazione della toxoplasmosi (Davidson et al., 1993), mentre
nel cane sono determinanti soprattutto il Cimurro e l’Ehrlichiosi. L’immunosoppressione
iatrogena è causata da trattamenti prolungati di glicocorticoidi o altri farmaci immunosoppressivi
come la ciclosporina (Beatty e Barrs, 2003; Bernsteen et al.,1999). La toxoplasmosi acquisita è,
generalmente, meno grave della forma congenita. Dopo l’ingestione di cisti tessutali o delle
oocisti i segni clinici sono localizzati all’intestino e con necrosi dell’epitelio e degli organi
linfoidi associati per la proliferazione dei tachizoiti (Dubey e Lappin, 1998); successivamente
per via linfoematica e all’interno dei monociti, il parassita diffonde in altri organi tra i quali
fegato, polmone, cervello e miocardio con aree di necrosi diffuse (Dubey e Lappin, 1998). Nella
terza settimana post-infezione i tachizoiti scompaiono e compaiono le cisti con i bradizoiti ;
queste possono rompersi e i bradizoiti rilasciti possono causare recidive in soggetti
immunodepressi. La forma cronica interessa prevalentemente il cervello,i muscoli scheletrici, il
fegato, i polmoni e gli occhi. Le cisti tessutali persistono per tutta la vita dell’ospite senza dare
particolari reazioni infiammatorie, possono riattivarsi in seguito a immunosoppressione.

33
Capitolo 5 Sintomi clinici
La Toxoplasmosi clinica nel cane raramente si presenta come malattia primaria, ma è quasi
sempre associata ad uno stato di debilitazione indotto da altre malattie o condizioni di
immunodepressione (Dubey ,2003). Come già detto nel cane T.gondii è spesso associato al virus
del Cimurro, all’Ehrlichiosi o all’uso terapeutico dei glicocorticoidi. L’infezione decorre spesso
in modo asintomatico e la gravità della sintomatologia clinica dipende da vari fattori,
fondamentalmente riconducibili a:
• Età
• Organi coinvolti
• Grado d’infezione
5.1 Sintomi clinici nell’ infezione congenita
La trasmissione congenita nel cane è rara ma possibile se l’infezione è contratta nelle prime fasi
di gestazione,quando i tachizoiti si diffondono per via transplacentare; i sintomi riscontrati sono
prevalentemente aborti e morti neonatali o perinatali. Nei soggetti che non muoio alla nascita
sono presenti addome aumentato di volume,spesso associato a epatomegalia e ascite; le forme
nervose sono prevalentemente caratterizzate da encefaliti con ipertermia e forme polmonari con
polmoniti, dispnea e difficoltà respiratoria (Dubey, 2003).
5.2 Sintomi clinici nell’ infezione post-natale
I sintomi clinici nel cane sono dovuti prevalentemente alla disseminazione extraintestinale dei
tachizoiti e alla successiva formazione di cisti contenenti i bradizoiti. Gli organi colpiti più
frequentemente sono i polmoni, il sistema neuromuscolare , l’oculare e il tratto gastroenterico. I
soggetti più sensibili sono cani sotto all’anno di età e gli anziani ovvero categorie con un sistema
immunitario più debole; i soggetti giovani, nelle fasi iniziali dell’infezione, manifestano sintomi
sistemici non patognomonici, come anoressia, letargia , febbre, diarrea, vomito, difficoltà
respiratorie e tonsillite. Sono frequenti anche polmoniti con difficoltà respiratorie e
interessamento epatico con ittero.
Nei soggetti anziani sono più frequenti forme neuromuscolari sistemiche (Dubey, 2000). In studi
effettuati in Brasile nel 2012 è stato dimostrato come T.gondii sia una delle principali cause di
sintomi neurologici nel cane in associazione al Cimurro (Langoni , 2012). I cani affetti
presentano:

34
• Atassia
• Variazioni comportamentali
• Circuitazione
• tremori
• Paralisi e paresi
• Andatura rigida
• Paraplegia
• Zoppia
• Encefaliti
Meno frequenti nel cane rispetto al gatto, ma comunque evidenziati sono sintomi oculari, quali
corio retinite, uveite con glaucoma, distacco del cristallino o della retina,neurite del nervo ottico,
intorbidamento dell’umor acqueo (Da Silva, 2005).
Recenti studi hanno evidenziato lesioni cutanee associate a T.gondii in cani trattati con terapia
immunosoppressiva per altre patologie; questa diagnosi è stata confermata mediante
immunoistochimica e PCR.
Questo studio considera due cani, un maschio Akita e di 9 anni ed una femmina Rhodesian
ridgeback di 7. Nel primo caso il cane era affetto da malattia emolitica immunomediata e da
infezioni alle basse vie urinarie;dopo un trattamento con Prednisolone ,ciclosporine ,sucralfato e
amoxicillina con acido clavulanico s’è risolto la forma infettiva e si è stabilizzato l’ematocrito,
ma dopo 20 giorni è descritta la comparsa di melena,diarrea trattata con metronidazolo. Dopo 20
giorni il cane è stato portato ad una visita specialistica per la comparsa di pustole e prurito. La
biopsia cutanea ha evidenziato un’infiammazione piogranulomatosa, estesa dallo strato
superficiale del derma fino a sottocute con neutrofili, macrofagi e frammenti necrotici. Nel
citoplasma di cellule endoteliali ,di cheratinociti , macrofagi e fibroblasti sono stati evidenziati
organismi basofilici, allungati con al centro tachizoiti, singoli o aggregati. Nel secondo caso il
cane era stato sottoposto a terapia immunomediata per trombocitopenia immunomediata ,
endocardite valvolare e infezione del tratto urinario (prednisolone, azathioprine, enrofloxacina e
amoxicillina). Dopo 4 settimane è stato osservato un nodulo cutaneo sulla spalla destra e il
citologico evidenziò la presenza del protozoo. Oltre alle lesioni viste nei due casi clinici in cani
e gatti affetti da toxoplasma sono state descritte dermatiti piogranulomatose o granulomatose e
necrotizzanti, pannicoliti con vasculite, iperplasia dell’epidermide e ipercheratosi. T.gondii può
essere organizzato in gruppo o da solo localizzato fuori della cellula o nel citoplasma del
follicolo epidermico, nei macrofagi, nell’epitelio ghiandolare o nei fibroblasti ( Hoffmann,2012).

35
5.3 Reperti anatomopatologici e lesioni istologiche
La principale lesione evidenziabile nelle infezioni da T.gondii nel cane, ma anche negli altri
animali colpiti è la necrosi e gli organi maggiormente interessati sono:
• il sistema nervoso centrale :
� aree atrofiche e pallide nel cervelletto;
� meningoencefalite multifocale non suppurativa con
necrosi,gliosi e vasculite;
� necrosi neuronale ,malacia,astrocitosi;
� infiltrato multifocale a livello delle meningi consistente in
macrofagi cellule plasmatiche, linfociti e neutrofili;
• il polmone:
� noduli bianco-grigi di 5 mm di diametro localizzati sotto la pleura;
� linfonodi bronchiali necrotici e aumentati di volume;
� essudato fibrinoso, necrosi delle pareti alveolari, dei vasi e dei
bronchioli;
� il lume alveolare con fibrina, linfociti, neutrofili ed eosinofili;
• tratto gastrointestinale :
� ulcere di più di 10 mm di diametro;
• pancreas,reni e milza:
� foci necrotici diffusi;
• Muscoli:
� miosite diffusa(soprattutto agli arti);
� necrosi delle miofibrille e fibrosi dei vasi e del connettivo;
� Atrofia muscolare;
� atrofia muscolare con sostituzione del tessuto muscolare con connettivo;
(Dubey e Lappin, 2006)

36
Capitolo 6 Diagnosi
La sintomatologia clinica e le analisi emato-biochimiche non sono patognomiche per la diagnosi
di T.gondii, perciò è necessaria una ricerca diretta e indiretta del parassita. Il controllo della
malattia nell’uomo e imprescindibilmente negli animali e l’individuazione delle fonti d’infezione
principali sono al centro della politica sanitaria europea, che ha infatti incaricato l’EFSA,
European Food Safety Authority, di fornire linee guida sui metodi diagnostici che permettano
una migliore identificazione del parassita negli animali (The EFSA Journal, 2007).
6.1 Diagnosi Diretta
Permette di identificare il parassita mediante ricerca microscopica, isolamento in vivo ed in vitro
e amplificazione del DNA con metodi molecolari.
� Esame coprologico
Eseguito mediante la tecnica di flottazione con soluzione satura di Nacl e ZnSO4 al 33% o la
tecnica di Telemann, eseguita con la sedimentazione con acido acetico ed etere. Permette di
evidenziare le oocisti eliminate con le feci dall’ospite definitivo ed è, quindi, esclusivo per il
gatto o altri felidi, e al di là della problematica della diagnosi differenziale con le oocisti di
Isospora (I.felis, I.rivolta), Sarcocystis (S.bovifelis, S.ovifelis) e soprattutto Hammondia
hammondi, vengono ritenuti di scarso significato diagnostico dato il breve periodo di patenza
della parassitosi (fino a 15 giorni) (Piergili Fioretti, 2010).
� Isolamento in vivo o in vitro
Il parassita può essere coltivato in vitro in colture di fibroblasti umani, questo metodo è rapido
(7gg) ma poco sensibile per perdite di vitalità del parassita a seguito di trattamenti
farmacologici; in vivo viene inoculato nel peritoneo di topi da laboratorio. Si tratta di un test
lento (4-6 settimane) ma specifico e sensibile (Piergili Fioretti, 2010).
� Istologia
Eseguita su campioni bioptici di organi target (linfonodi, milza , fegato, polmone, pancreas in
caso di infezione acuta, SNC in quella cronica) utilizzando le procedure routinarie di fissazione,
disidratazione, inclusione, colorazione elettiva delle sezioni con ematossilina-eosina. Permette di
evidenziare tachizoiti e cisti tessutali. Infezioni non massive possono però risultare in falsi
negativi per la scarsa quantità di materiale bioptico (the EFSA Journal , 2007).L’identificazione

37
dell’organismo da lesioni cutanee con l’esame istologico può essere una vera e propria sfida: in
soggetti sierologicamente positivi per T.gondii con lesioni cutanee suggestive di questa
infezione, solo nel 50% è possibile isolarlo dai tessuti (Hoffmann,2012).
� Immunofluorescenza e immunoistochimica
Vengono eseguite sui campioni bioptici per evidenziare i tachizoiti e le pseudocisti,
nell’immunoistochimica sono utilizzati anticorpi policlonali che si legano agli antigeni del
parassita (Piergili Fioretti, 2010).
� PCR
La PCR permette di amplificare il DNA del parassita dal siero, sangue, liquor e umor acqueo ed
è una delle tecniche più sensibili;tuttavia la piccola quantità di materiale necessario ne limita la
sensibilità, poiché la distribuzione delle cisti tessutali nel campione è casuale e la densità del
parassita può essere bassa (Hill et al.,2006). Questa tecnica è utilizzata per evidenziare il DNA di
T.gondii nelle feci dei gatti e per la diagnosi differenziale da altri protozoi (Mancianti et al.,2014;
Salant et al., 2010).
� Sierologia
I test sierologici evidenziano la presenza di antigeni circolanti.
6.2 Diagnosi indiretta
Permette di mettere in evidenza la risposta anticorpale che l’organismo mette in atto in seguito
all’infezione; in corso di Toxoplasmosi si ha un primo movimento anticorpale di IgM nei primi
5 giorni dall’infezione, con un picco a 2-3 settimane e scomparsa a 3 -6 mesi; le IgG compaiono
a 2 settimane post infezione, hanno un picco a 2-3 mesi e diminuiscono a 1-2 anni, permangono
però per tutta la vita dell’animale. Le IgA hanno un’insorgenza incostante tra IgM e IgG e
sembrano scomparire in un anno.

38
Figura 9. Grafico che rispecchia i risultati dei diversi studi condotti sulle quantità di oocisti di T. gondii
emesse da gatti infettati sperimentalmente o naturalmente nutriti con tachizoiti o bradizoiti contenuti nei
tessuti suini o murini, o essudati peritoneali di topo.(Dal review di Dabritz e Conrad, 2010)
In bibliografia, sia recente che datata, sono riportati studi solo sui felidi, mentre sul cane non è
presente nessun dato; su gatti affetti da toxoplasmosi clinica sono stati rilevati titoli di IgM
trovati nel 93,3% dei sieri e IgG nel 60%, pertanto si ritiene che le IgM abbiano un valore
predittivo della positività alla forma clinica rispetto alle IgG (Lappin et al., 1989; Lappin, 1996).
Nell’80% dei gatti sani infettati sperimentalmente è possibile rilevare IgM T.gondii-specifiche,
nelle 2-4 settimane post inoculazione (Lappin,2010); questo titolo diventa negativo a 16
settimane circa (Lappi net al.,1989). In gatti con T.gondii oculare e con FIV sono stati descritti
titoli di IgM sopra le 16 settimane (Lappin et al.,1926). Nella maggior parte dei casi il periodo
dell’emissione delle oocisti da parte dell’ospite è terminato quando si rilevano le IgG nel siero,
percui gatti sieropositivi per queste immunoglobuline non dovrebbero rappresentare un rischio
d’infezione. É, però, doveroso usare il congiuntivo, poiché studi recenti hanno riportano casi di
gatti positivi alla ricerca del DNA di T.gondii nelle feci e con IgG nel siero (Mancianti et al.,
2010; Lappin et al., 1989; Lappin, 1996). Titoli anticorpali IgG sono stati rilevati nel siero
anche 6 anni post-infezione, ma si ritiene che persistano tutta la vita del soggetto, visto che il
protozoo può resistere nei tessuti per molti anni (Dubey et al., 1995). Titoli elevati di IgG non
indicano, quindi, necessariamente un’infezione recente o attiva. Il periodo che intercorre tra il
primo rilievo di titoli elevati di IgG e il picco, è di circa 2-3 settimane, per questo motivo al
momento del test sierologico le IgG potrebbero già essere diminuite (Lappin,2010). Infine nel
gatto le IgG aumentano raramente in seguito alla riattivazione cronica (Lappin , 2010).Alla luce
di questi dati è inutile ripetere la titolazione anticorpale dopo la risoluzione della malattia clinica,
né una terapia in soggetti che non presentano sintomi di Toxoplasmosi ( Lappin, 2010).
Le principali tecniche che utilizzano tachizoiti interi sono il Dye Test, MAT, IFAT, mentre
ELISA, IHA usano antigeni endocitoplasmatici . MAT è un test di agglutinazione ,utilizzato per

39
determinare sia le IgG sia le IgM: ELISA e IFAT sono considerati test di riferimento per la
diagnosi perché saggiano tutte le classi anticorpali.
6.2.1 Tecnica IFAT ( Immunofluorescence antibody test)
L’immunofluorescenza è una tecnica di laboratorio usata in quasi tutte le branche della biologia
per ricerche per la diagnostica clinica; questa tecnica si basa sul lavoro pioneristico di Coons e
Kaplan (Coons , et al. , 1941; Coons e Kaplan, 1950) e successivamente di Mary Osborne
(Weber et al., 1975). Con questa tecnica è possibile rendere visibile una reazione antigene -
anticorpo, marcando uno dei due con fluorocromi quali istiocianato di fluoresceina (FITC) o
tetrametil rodamina istiocianato (TRITC). Il FITC è un composto giallo che può essere legato ad
anticorpi senza alterare la loro reattività; quando irradiato da luce ultravioletta o celeste riemette
una luce visibile verde chiaro. Il TRiTC emette una fluorescenza rossa. Questa tecnica è
utilizzata sia per individuare un microrganismo patogeno da tessuti infetti sia complessi
antigene-anticorpo . Viene utilizzata anche per rilevare anticorpi specifici dal siero di sangue,ma
in questo caso l’antigene verso cui si cercano gli anticorpi viene fissato su vetrino e si procede
con la tecnica indiretta. L’immunofluorescenza può essere diretta o indiretta;la prima è utilizzata
per identificare la presenza degli antigeni, mentre l’altra è usata per la titolazione degli anticorpi
o per la ricerca degli antigeni.
Figura 10. Immunofluorescenza diretta e indiretta (da Robinson ,immunohistochemical staining methods
2009, http://www.dako.com/08002_03aug09_ihc_guidebook_5th_edition_chapter_10.pdf ).

Immunofluorescenza diretta
Gli anticorpi diretti verso un antigene specifico sono marcati con un
fissato il tessuto da analizzare su un vetrino è incubato con l’antisiero marcato;
il vetrino viene lavato in PBS per eliminare gli anticorpi non legati. Il vetrino sarà osservato ad n
microscopio a luce ultraviolet
antigene anticorpo ci sarà una fluorescenza brillante.
Figura 11. Immunofluorescenza diretta
UTET, pag. 685)
Immunofluorescenza indiretta
È la tecnica utilizzata in questo lavoro,
incubato con un siero sospetto d’avere anticorpi per quell’antigene;
lasciando solo anticorpi specifici legati agli antigeni.
solo dopo incubazione con antiglobul
non legate. La quantità di anticorpi presenti è valutata mediante una diluizione cres
Questa tecnica, rispetto all’indiretta
può legare più antiglobuline marcate
antiglobuline specifiche per ogni classe di antico
dell’anticorpo (Tizard, 2006).
munofluorescenza diretta
Gli anticorpi diretti verso un antigene specifico sono marcati con un fluorocromo e dopo aver
fissato il tessuto da analizzare su un vetrino è incubato con l’antisiero marcato;
il vetrino viene lavato in PBS per eliminare gli anticorpi non legati. Il vetrino sarà osservato ad n
microscopio a luce ultravioletta e dove c’è una reazione positiva,quindi c’è un legame tra
antigene anticorpo ci sarà una fluorescenza brillante.
mmunofluorescenza diretta (da “Microbiologia e immunologia veterinaria
Immunofluorescenza indiretta
la tecnica utilizzata in questo lavoro, in cui un antigene presente in un campione viene
incubato con un siero sospetto d’avere anticorpi per quell’antigene; il siero viene poi lavat
specifici legati agli antigeni. Il legame anticorpo-antigene è visualizzato
solo dopo incubazione con antiglobuline marcate e dopo aver rimosso, mediante lavaggio
La quantità di anticorpi presenti è valutata mediante una diluizione cres
rispetto all’indiretta, ha due vantaggi cioè che ogni complesso antigene anticorpo
egare più antiglobuline marcate e la brillantezza è molto più marcata.
antiglobuline specifiche per ogni classe di anticorpi si può determinare la classe specifica
40
fluorocromo e dopo aver
fissato il tessuto da analizzare su un vetrino è incubato con l’antisiero marcato; successivamente
il vetrino viene lavato in PBS per eliminare gli anticorpi non legati. Il vetrino sarà osservato ad n
ta e dove c’è una reazione positiva,quindi c’è un legame tra
Microbiologia e immunologia veterinaria”, Giorgio Poli,
in cui un antigene presente in un campione viene
il siero viene poi lavat o,
antigene è visualizzato
mediante lavaggio, quelle
La quantità di anticorpi presenti è valutata mediante una diluizione crescente di siero.
ha due vantaggi cioè che ogni complesso antigene anticorpo
e la brillantezza è molto più marcata. Inoltre usando
rpi si può determinare la classe specifica
41
Figura 12. Immunofluorescenza indiretta (da “Microbiologia e immunologia veterinaria”,Giorgio
Poli,UTET, pag. 685)
Principi di fluorescenza
Fosforescenza e fluorescenza sono 2 tipi di luminescenza. Quando molecole con proprietà
luminescenti assorbono luce, emettono raggi luminosi a diverse lunghezze d’onda; nella
fluorescenza l’emissione di luce è estremamente rapida mentre nella fosforescenza l’emissione
continua per millisecondi o minuti, dopo che la fonte di energia è stata rimossa. Il materiale
fluorescente emette luce per la struttura atomica; gli elettroni sono organizzati in diversi livelli
intorno al nucleo dell’atomo ed ogni livello ha una predeterminata quantità d’energia. Quando
un elettrone assorbe l’energia da un fotone di luce, diventa “eccitato” e salta al livello di energia
superiore e meno stabile e tale stato non dura più di 10 secondi. L’energia è persa dall’elettrone
sottoforma di calore ,mentre la restante come fotone. La fluorescenza emessa ha minor energia
rispetto alla luce assorbita, così la lunghezza d’onda della luce emessa e maggiore rispetto alla
luce che ha eccitato.
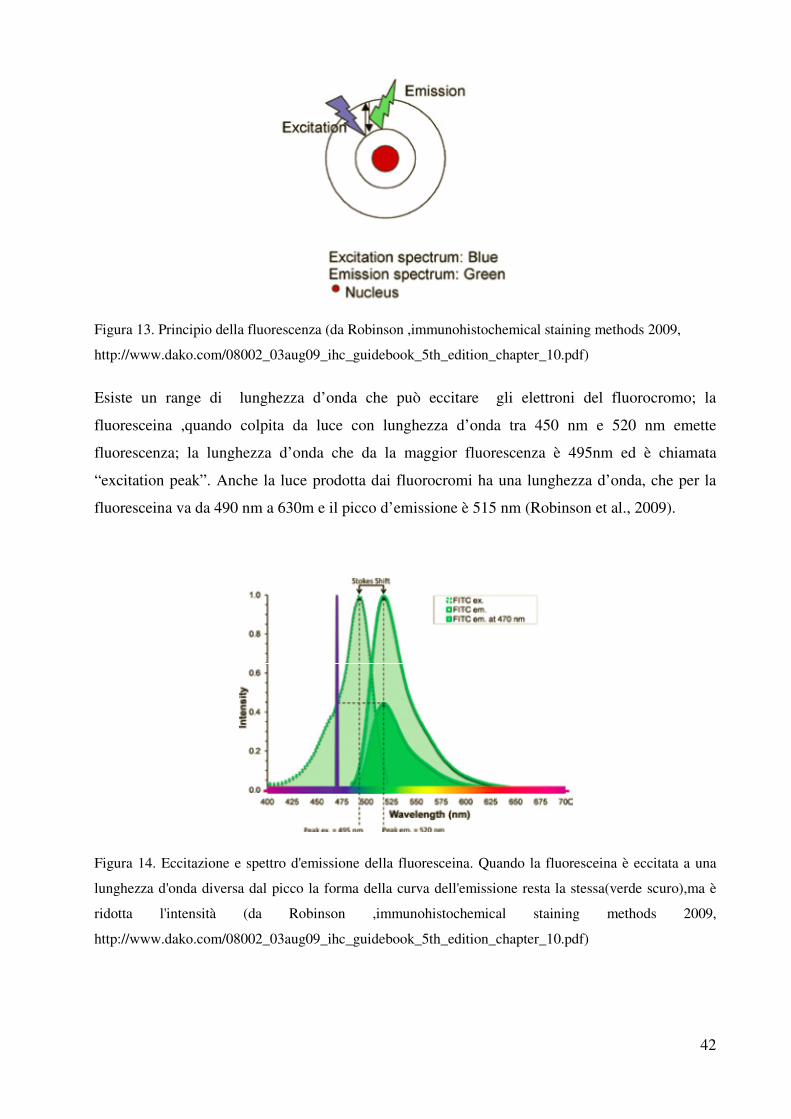
42
Figura 13. Principio della fluorescenza (da Robinson ,immunohistochemical staining methods 2009,
http://www.dako.com/08002_03aug09_ihc_guidebook_5th_edition_chapter_10.pdf)
Esiste un range di lunghezza d’onda che può eccitare gli elettroni del fluorocromo; la
fluoresceina ,quando colpita da luce con lunghezza d’onda tra 450 nm e 520 nm emette
fluorescenza; la lunghezza d’onda che da la maggior fluorescenza è 495nm ed è chiamata
“excitation peak”. Anche la luce prodotta dai fluorocromi ha una lunghezza d’onda, che per la
fluoresceina va da 490 nm a 630m e il picco d’emissione è 515 nm (Robinson et al., 2009).
Figura 14. Eccitazione e spettro d'emissione della fluoresceina. Quando la fluoresceina è eccitata a una
lunghezza d'onda diversa dal picco la forma della curva dell'emissione resta la stessa(verde scuro),ma è
ridotta l'intensità (da Robinson ,immunohistochemical staining methods 2009,
http://www.dako.com/08002_03aug09_ihc_guidebook_5th_edition_chapter_10.pdf)

43
Capitolo 7 Terapia e prevenzione
7.1 Trattamento farmacologico
Studi sul trattamento della Toxoplasmosi con farmaci specifici son iniziati negli anni quaranta
con la descrizione degli effetti terapeutici dei sulfonamidici (Sabine et al., 1941) e proseguiti poi
negli anni 50, con lo studio di una terapia combinata di sulfadiazina e pirimetamina (Eyles et al.,
1953) e spiramicina (Beverly, 1958). Questi farmaci agiscono soprattutto nelle infezioni acute,
poiché si rivolgono contro i tachizoiti ed hanno una scarsa efficacia contro le cisti tessutali
(Innes, 2010). Nel cane la soppressione del midollo osseo per uso di atifolati è meno evidente
che nel gatto, dove sono necessari controlli ematologici per escludere trombocitopenia,
leucopenia e anemia se il trattamento dura più di due settimane. Trimethoprim sulfamidico
oltrepassa la barriera ematoencefalica, ma risulta essere inefficace nel cane per il trattamento di
uveiti gravi e neuriti ottiche (Dubey et al., 1990).
Il farmaco d’elezione è la Clindamicina, un antibiotico facente della classe dei Lincosamidi. La
Clindamicina riduce i sintomi della forma clinica della toxoplasmosi,quali l’ipertermia,la
miosite,l’uveite . Questo farmaco trattamento ha un buon assorbimento intestinale,quindi i
dosaggi parenterali e morali sono simili (25 mg /kg bid ) e i segni clinici, non determinati da
danni al SNC, cominciano a ridursi dopo 24-48 ore dall’inizio della terapia. I sintomi
neurologici, quali la polimiosite , il deficit ai motoneuroni inferiori e l’atrofia muscolare,
richiedono, invece, un tempo maggiore per ridursi (alcune settimane), anche se possono non
risolversi mai a causa dei danni al SNC causati dall’infezione. La Clindamicina somministrata
“per os” può causare anoressia, vomito e diarrea in cani trattati a dosaggi elevati, poiché
comporta un’ irritazione della mucosa gastro-intestinale; questi effetti non si verificano nella
somministrazione parenterale. L’uso dei corticosteroidi non è consigliato per l’effetto
immunosoppressivo che ha, se non per uso locale nel il trattamento di infiammazioni del
segmento anteriore oculare, anche perchè sembra che a dosi cliniche non peggiorino il quadro
sistemico (Lappi net al., 1992). Non esiste, al momento, un farmaco che liberi completamento un
organismo da T.gondii e la prognosi per cani e gatti affetti da forma disseminata è quasi sempre
infausta.

44
7.2 Profilassi Vaccinale
Nel 1967 Frenkel ha dimostrato l’importanza della risposta cellulo-mediata mediante
trasferimento di cellule linfoidi ed ha provato la mediazione dell’immunità da parte di cellule
immunitarie; il trasferimento del siero non ha invece protetto i riceventi. Attualmente l’unico
vaccino contro T.gondii in commercio è utilizzabile negli ovini; non può essere utilizzato
nell’uomo o in altre specie poiché, contenendo tachizoiti vivi, rappresenta un rischio d’infezione
(Innes et al.,2006). I vaccini inattivati non sono in grado di stimolare una risposta cellulo medita
abbastanza importante da proteggere dall’infezione (Innes e Vermeulen, 2006). Questo vaccino,
sviluppato in Nuova Zelanda (o’Connel et al.,1988) e contenente tachizoiti del ceppo S48,
previene la trasmissione congenita nelle pecore poichè stimola una risposta cellulo -
mediata,prima della gestazione (Innes et al.,2009).

45
Capitolo 8 Scopo del lavoro La toxoplasmosi è una delle principali zoonosi conosciute, temuta soprattutto dalle donne in
gravidanza e con un importante impatto sulla sanità pubblica . E’ molto importante, perciò,
conoscere questa patologia in tutti i suoi aspetti, sia per garantire la massima protezione e
sicurezza in gravidanza sia per smentire quelle teorie che considerano gli animali domestici
pericolosi portatori di T.gondii .
La toxoplasmosi del gatto è ampiamente conosciuta, sia per quanto riguarda il ciclo e quindi il
ruolo dei felini nella trasmissione e la sintomatologia, sia per quanto concerne la presenza e la
diffusione del parassita tra le colonie feline; sono invece pochi i dati sul cane e gli articoli
presenti in bibliografia riguardano paesi esteri, soprattutto Brasile e Cina. Gli studi sulla
toxoplasmosi canina in Italia sono pochi e non recenti, praticamente assenti quelli sulla
prevalenza del parassita in Toscana. Partendo da questo vuoto bibliografico, lo scopo del mio
lavoro è stato quello di valutare la sieroprevalenza di T.gondii in uno studio retrospettivo
condotto su sieri di animali viventi sia in Toscana che in Liguria, Genova in particolare e, allo
stesso tempo, quando possibile, valutare i fattori di rischio ambientali associati. Analizzando poi
i dati anamnestici fornitimi dalle strutture veterinarie ho potuto evidenziare quali cani siano stati
maggiormente colpiti in base alle categorie d’ appartenenza (età,razza,habitat…).

46
Capitolo 9 Materiali e metodi 9.1 Campionamento
Per questo studio sono stati utilizzati campioni di siero di cane, pervenuti presso il laboratorio di
sierologia parassitaria del Dipartimento di scienze veterinarie dell’Università di Pisa, per il
controllo sierologico annuale della leishmaniosi. Il periodo di tempo in cui sono stati fatti questi
campionamento è di circa due anni, da gennaio 2012 a dicembre 2013.
Il prelievo di sangue è stato effettuato nelle varie strutture, prevalentemente dalla vena Cefalica,
raccolto in apposite provette, lasciato riposare per circa 2 ore, ad una temperatura che oscilla tra
10°C e i 20°C; dopo la formazione del coagulo questo è stato“scollato”dalle pareti della provetta.
Una volta recuperato il siero, è stato centrifugato per allontanare eventuali globuli rossi rimasti.
La provetta chiusa, a circa 4°C, è stata alloggiata in contenitori termici da trasporto, che lo hanno
protetto sia da sbalzi di temperatura che da insulti meccanici e sono stati così inviati; arrivati in
laboratorio sono stati effettuati i test della Leishmania e poi congelati fino all’esecuzione
dell’immunofluorescenza per T.gondii. Una volta evidenziati i sieri positivi li ho esaminati a
diluizioni maggiori al fine di stabilire il titolo anticorpale .
I 1562 sieri da me analizzati, provengono da strutture presenti in una zona compresa tra Toscana
e Liguria e la popolazione di cani analizzata è mista per quanto riguarda la razza, l’età e
l’ambiente di vita. Questa eterogeneità è fondamentale per il lavoro, poiché non solo studia la
presenza di T.gondii nella popolazione canina in esame ma evidenzia anche quali categorie
siano maggiormente colpite. Sui dati riguardanti la siero prevalenza nelle varie zone è stato poi
fatto un test statistico (chi-quadro). Per questo lavoro è stata essenziale la collaborazione e la disponibilità delle strutture veterinarie
che mi hanno prontamente fornito i dati riguardanti i pazienti ; ho preso in esame fattori che
possono influire sia sul contagio di T.gondii che sulla manifestazione clinica della malattia. In
particolare ho analizzato:
• Razza
• Genere (maschile o femminile)
• Età
• Anamnesi ambientale e contatto con altre specie animali
• Dieta
• Anamnesi remota
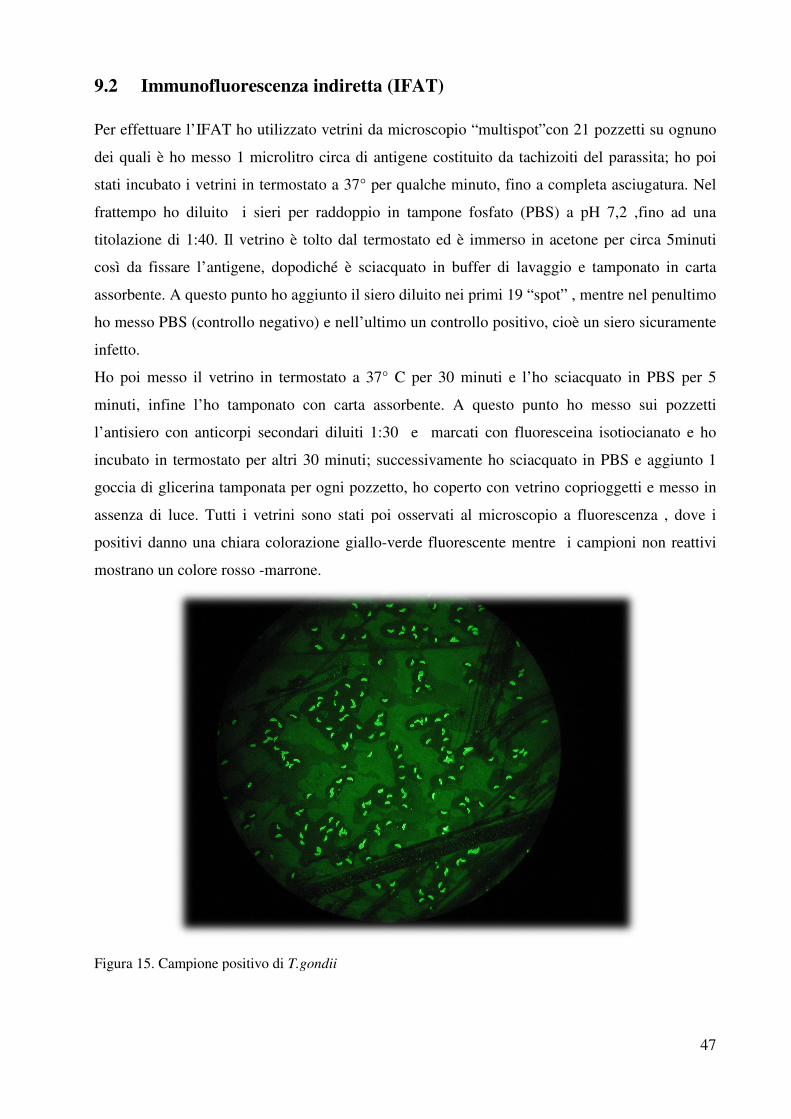
9.2 Immunofluorescenza indiretta (IFAT)
Per effettuare l’IFAT ho utilizzato vetrini da microscopio “multispot”con 21 pozzetti su ognuno
dei quali è ho messo 1 microlitro circa di antigene costituito da
stati incubato i vetrini in termostato a 37° per qualche minuto,
frattempo ho diluito i sieri per raddoppio in tampone fosfato (PBS) a pH 7,2
titolazione di 1:40. Il vetrino è
così da fissare l’antigene, dopodiché è sciacquato in buffer di lavaggio e tamponato in carta
assorbente. A questo punto ho aggiunto il siero diluito nei primi 19 “spot” ,
ho messo PBS (controllo negativo) e n
infetto.
Ho poi messo il vetrino in termostato a 37° C per 30 minuti e l’ho sciacquato in PBS per
minuti, infine l’ho tamponato con carta assorbente.
l’antisiero con anticorpi secondari diluiti 1:
incubato in termostato per altri 30 minuti;
goccia di glicerina tamponata
assenza di luce. Tutti i vetrini sono stati poi osservati al microscopio a fluorescenza ,
positivi danno una chiara colorazione giallo
mostrano un colore rosso -marrone.
Figura 15. Campione positivo di
Immunofluorescenza indiretta (IFAT)
Per effettuare l’IFAT ho utilizzato vetrini da microscopio “multispot”con 21 pozzetti su ognuno
dei quali è ho messo 1 microlitro circa di antigene costituito da tachizoiti del parassita; ho poi
stati incubato i vetrini in termostato a 37° per qualche minuto, fino a completa asciugatura. Nel
frattempo ho diluito i sieri per raddoppio in tampone fosfato (PBS) a pH 7,2
Il vetrino è tolto dal termostato ed è immerso in acetone per circa 5minuti
dopodiché è sciacquato in buffer di lavaggio e tamponato in carta
assorbente. A questo punto ho aggiunto il siero diluito nei primi 19 “spot” ,
(controllo negativo) e nell’ultimo un controllo positivo, cioè un siero sicuramente
Ho poi messo il vetrino in termostato a 37° C per 30 minuti e l’ho sciacquato in PBS per
infine l’ho tamponato con carta assorbente. A questo punto ho messo sui pozzetti
l’antisiero con anticorpi secondari diluiti 1:30 e marcati con fluoresceina isotiocianato e ho
incubato in termostato per altri 30 minuti; successivamente ho sciacquato in PBS
per ogni pozzetto, ho coperto con vetrino coprio
Tutti i vetrini sono stati poi osservati al microscopio a fluorescenza ,
colorazione giallo-verde fluorescente mentre i campioni non reatt
marrone.
ampione positivo di T.gondii
47
Per effettuare l’IFAT ho utilizzato vetrini da microscopio “multispot”con 21 pozzetti su ognuno
tachizoiti del parassita; ho poi
fino a completa asciugatura. Nel
frattempo ho diluito i sieri per raddoppio in tampone fosfato (PBS) a pH 7,2 ,fino ad una
tolto dal termostato ed è immerso in acetone per circa 5minuti
dopodiché è sciacquato in buffer di lavaggio e tamponato in carta
assorbente. A questo punto ho aggiunto il siero diluito nei primi 19 “spot” , mentre nel penultimo
cioè un siero sicuramente
Ho poi messo il vetrino in termostato a 37° C per 30 minuti e l’ho sciacquato in PBS per 5
questo punto ho messo sui pozzetti
e marcati con fluoresceina isotiocianato e ho
cquato in PBS e aggiunto 1
to, ho coperto con vetrino coprioggetti e messo in
Tutti i vetrini sono stati poi osservati al microscopio a fluorescenza , dove i
verde fluorescente mentre i campioni non reattivi

48
9.3 Leishmania e Neospora
Sui sieri è stata eseguita anche l’immunofluorescenza indiretta per Leishmania infantum
(Mancianti e Meciani, 1988) ed i sieri positivi sono stati valutati anche per la presenza di
anticorpi nei confronti di Neospora caninum, protozoo che colpisce i cani causando sintomi che
possono essere confusi con quelli provocati da T.gondii.
L.infantum è un protozoo flagellato, appartenente al phylum “Mastigophora”(Ross ,1903),
parassita obbligato e permanente che ha come ospite preferenziale il cane,ma può colpire anche
la volpe, il gatto e l’uomo. Nell’ospite vertebrato è presente la forma amastigote del parassita, a
livello dei macrofagi cutanei. L’infezione è trasmessa da un insetto vettore appartenente al
genere Phlebotomus che compie il pasto di sangue sull’ospite infetto,ingerisce gli amastigoti
presenti nei macrofagi,che si trasformeranno in o promastigoti e migreranno alle ghiandole
salivari dell’insetto da cui saranno inoculati al prossimo pasto. L.infantum è cosmopolita, si
ritrova nel bacino del Mediterraneo, in Cina, nell’ex URSS, dal nord degli Usa fino
all’Argentina, Brasile compreso (Kuhls et al., 2011).
I sintomi nel cane sono variabili, possono compendere linfoademopatia periferica, lesioni
cutanee, insufficienza renale , uveite e disturbi di deambulazione (Paulan et al., 2012).
N. caninum è un protozoo appartenente al phylum “apicomplexa”, che causa aborto e infertilità
in bovine di tutto il mondo; fu identificato per la prima volta nel cervello di cane (Dubey et al.,
1990), che rappresenta l’ospite definitivo e un fattore importante nella trasmissione agli altri
animali selvatici e domestici (Dubey et al., 2003). Negli ospiti intermedi, bovini , ovini ma anche
canidi, il parassita si trasmette per passaggio transplacentare dei tachizoiti che colonizzano feto
ed invogli, causando nelle prime gravidanze aborti, mentre successivamente nascita di soggetti
infettati; l’ospite definitivo (il cane) si contagia per ingestione di cisti tessutali (Mc Innes et al.,
2006). La differenza sostanziale tra N.caninum e T.gondii è che nella prima il parassita è
particolarmente adattato al passaggio transplacentare, che è parte attiva del contagio, mentre
nella toxoplasmosi il passaggio transplacentare pare essere di minor rilevanza ai fini del ciclo.
Nel cane i sintomi principali sono nervosi, con paresi progressiva ascendente agli arti posteriori
che diventa paralisi, con ipertensione spastica e polimiosite; è presente anche flaccidità e paralisi
della mandibola, insufficienza cardiaca, renale ed epatite (Plugge et al., 2011). Sono presenti
anche lesioni cutanee che possono essere confuse con quelle indotte T.gondii.

49
Capitolo 9 Risultati
La prevalenza totale è risultata 8,9% (140/1562) con variabilità per le diverse zone; la prevalenza
maggiore è stata evidenziata in provincia di Pisa (18,35%), le province di Firenze e Pistoia
hanno mostrato una prevalenza del 7,6%, mentre Genova e Livorno si aggirano intorno all’8,5%;
nelle province di Siena, Lucca e Prato non sono stati rilevati sieri positivi, probabilmente per
l’esiguità del campione.In dettaglio 107 (76.4%) campioni hanno presentato un titolo 1/40, 21
(15%)1/80, 11 (7.9%) 1/20 ed 1 (0.7%) 1/320.
Luogo N°campioni N° campioni
positivi Indice di
prevalenza
Prov.di Pisa
Pisa 69 13 13,2
Pontedera 18 4 4.08
San Miniato 3 1 1,02
Calci 8 0 0
Tot. 98 18 18,3
Prov.di Livorno
Livorno 376 33 8,39
Portoferraio 6 1 0,25
Rosignano Mmo. 5 0 0
Cecina 4 0 0
Tirrenia 1 0 0
Venturina 1 0 0
tot. 393 34 8,65
Prov.di Genova
Tot. 393 38 9,6
Prov.di Firenze
Firenze 293 21 3,59
Sesto F.no 145 13 2,22
Fiesole 86 5 0,85
Scandicci 20 2 0,34
Vinci 19 0 0
Empoli 11 1 0,17
Greve 6 1 0,17
Certaldo 2 0 0
San Giovanni 1 0 0

50
Campi Bisenzio 1 1 0,17
Tot. 584 44 7,51
Prov.di Pistoia
Pistoia 65 3 3,48
Serravalle 17 1 1,16
Lamporecchio 2 1 1,16
Pescia 2 1 1,16
Tot. 86 6 6,96
Prov.di Prato
Tot. 3 0 0
Prov.di Lucca
tot 3 0 0
Prov.di Siena
Tot. 2 0 0
Dall’analisi statistica, eseguita con il test del Chi-quadro, si hanno i seguenti risultati:
� Differenza tra Pisa e Pistoia • Significativa • P=0.0221 • Chi-quadro=5.24 • Odd ratio (95% CI)=3 (1.13-7.95)
� Differenza tra Pisa e Firenze • Altamente significativa • P=0,0006 • Chi-quadro=11.92 • OR (95%CI)=2.76 (1.52-5.01)
� Differenza tra Pisa e Livorno • Altamente significativa • P=0,0052 • Chi-quadro=7.82 • OR (95%CI)=2.38 (1.28-4.42)
� Tra Pisa e Genova • Significativa • P=0,0154 • Chi-quadro=5.87 • OR (95% CI)=2.10 (1.14-3.87)

51
I campioni positivi sono stati suddivisi in 3 categorie d’età, cioè in “giovani”, fino all’anno di
vita, in “adulti” dai 2 ai 7 anni e in “anziani”sopra gli 8 anni. I risultati sono stati i seguenti:
� Giovani: 5% (7 campioni)
� Adulti:44 % (61 campioni)
� Anziani 51% (72 campioni)
Nella prima categoria tutti i campioni hanno titolo anticorpale 1 /40; tra gli adulti 6 hanno un
titolo di 1/80, 42 di 1/40 e 7 di 1/20. Un solo campione di un cane di 15 anni ha titolo 1/320.
Per quanto riguarda il genere,i maschi positivi sono 81,mentre le femmine 59,con una titolazione
così distribuita:
- 1/320=1 maschio;
-1/80= 13 maschi e 7 femmine;
-1/40= 54 maschi e 40 femmine;
-1/20=12 maschi e 8 femmine.
Esiste un sostanziale equilibrio tra i meticci e i cani di razza positivi; tra i 77 meticci la maggior
parte sono di taglia medio grande, così come tra le razze dove prevalgono quelle “da caccia” o
cani con indole molto attiva.
Il titolo 1/320 è evidenziato in un meticcio, 1/80 in 7 di razza e 14 meticci, 1/40 in 48 meticci e
44 di razza, mentre 1 /20 in 9 di razza e 8 meticci.L’anamnesi ambientale dei cani esaminati
riporta che 45 vivono in campagna, 69 in città e 26 in semicampagna. I cani che vivono in
ambiente urbano,soprattutto Genova e Firenze, hanno scarso contatto con quello rurale e con
animali selvatici che ci vivono; da un approfondimento fatto sui cani definiti ”di città” ho potuto
evincere che la maggior parte hanno comunque periodici contatti con zone rurali.
È stata definita la categoria “semicampagna” per quei soggetti che nonostante non vivano in
aperta campagna, sono spesso a contatto con prati, aree verdi non presenza di altre specie, in
particolare il gatto o che vivono in paesi che confinano con aree rurali. I cani di campagna,
infine, vivono a contatto con gatti, topi e altri animali selvatici,potenzialmente infetti.
49 campioni appartengono a cani alimentati esclusivamente con prodotti secchi o umidi
commerciali, mentre 40 con cibo preparato dai proprietari, avanzi o “pastoni”con soprattutto
carboidrati (pasta o riso) e carne. 51 cani sono alimentati con cibo “misto”,con questo si
definisce un ‘alimentazione varia, sia casalinga che commerciale in base alla disponibilità di
risorse e in base alla comodità dei proprietari.
Ho cercato di risalire all’anamnesi sanitaria dei cani per poter valutare se T.gondii fosse presente
in associazione ad altre patologie più evidenti, che hanno causato un indebolimento del sistema

52
immunitario. 41 dei 140 cani sieropositivi sono affetti da patologie più o meno debilitanti del
sistema immunitario o vari sintomi che possono essere così riassunti:
• Patologie neoplastiche;
• Insufficienze d’organo:rene,fegato,cuore
• Disfunzioni metaboliche:ipotiroidismo,Cushing,diabete
• Patologie cutanee quali dermatiti atopiche
• Sindromi autoimmunitarie
Sono stati riferiti 6 casi di cani con sintomi nervosi e cerebrali, con difficoltà alla deambulazione
e con crisi epilettiche, per i quali non era ancora stata fatta una diagnosi precisa; 1 cane presenta
difficoltà visive e cataratta, mentre 2 cani hanno dermatite cronica che non si risolve con nessun
trattamento farmacologico.19 cani sono sieropositivi,oltre che per T.gondii,anche per N.caninum
e per L.infantum:
� 10 sieri positivi per T.gondii e N.caninum;
� 6 sieri positivi per T.gondii e Leishmania;
� 3 sieri per T.gondii,N.caninum e L.infantum;
N°
TIT. ANTICORP
ALE TOXOPLA
SMA
SESSO
ETÀ
RAZZA ALIMENTA
ZIONE HABITA
T ANAMNESI SANITARIA
TITOLO ANTICORPALE
LEISHMANIA
TITOLO ANTICORPA
LE NEOSPORA
1 1/40 ms 5 Meticcio mista semicampag
na n.p. neg. neg.
2 1/40 m 9 Pastore Tedesco
commerciale città mastocitoma neg. neg.
3 1/40 m 12 Meticcio
tg. grande mista
semicampagna
n.p. neg. neg.
4 1/40 f 10 Meticcio tg.media
mista semicampag
na sindrome cerebrale
neg. 1/40
5 1/20 m s 10 Maltese commerciale città dermatite neg. neg.
6 1/40 f 10 Setter Inglese
commerciale città insufficienza
renale neg. neg.
7 1/40 fs 11 Meticcio
taglia media
commerciale città n.p. Neg. neg.
8 1/40 f 12 Setter Inglese
commerciale città ipotiroidismo neg. neg.
9 1/80 m 1 Meticcio tg.piccola
mista campagna n.a. neg. neg.
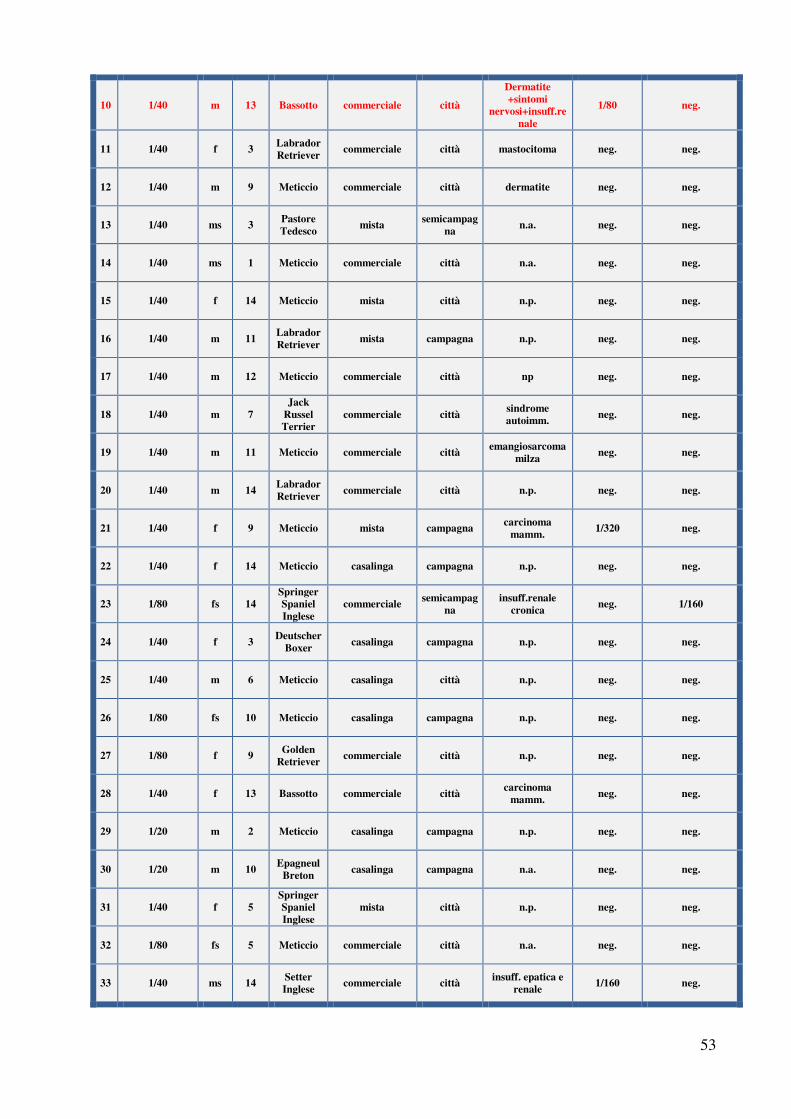
53
10 1/40 m 13 Bassotto commerciale città
Dermatite +sintomi
nervosi+insuff.renale
1/80 neg.
11 1/40 f 3 Labrador Retriever
commerciale città mastocitoma neg. neg.
12 1/40 m 9 Meticcio commerciale città dermatite neg. neg.
13 1/40 ms 3 Pastore Tedesco
mista semicampag
na n.a. neg. neg.
14 1/40 ms 1 Meticcio commerciale città n.a. neg. neg.
15 1/40 f 14 Meticcio mista città n.p. neg. neg.
16 1/40 m 11 Labrador Retriever
mista campagna n.p. neg. neg.
17 1/40 m 12 Meticcio commerciale città np neg. neg.
18 1/40 m 7 Jack
Russel Terrier
commerciale città sindrome autoimm.
neg. neg.
19 1/40 m 11 Meticcio commerciale città emangiosarcoma
milza neg. neg.
20 1/40 m 14 Labrador Retriever
commerciale città n.p. neg. neg.
21 1/40 f 9 Meticcio mista campagna carcinoma
mamm. 1/320 neg.
22 1/40 f 14 Meticcio casalinga campagna n.p. neg. neg.
23 1/80 fs 14 Springer Spaniel Inglese
commerciale semicampag
na insuff.renale
cronica neg. 1/160
24 1/40 f 3 Deutscher
Boxer casalinga campagna n.p. neg. neg.
25 1/40 m 6 Meticcio casalinga città n.p. neg. neg.
26 1/80 fs 10 Meticcio casalinga campagna n.p. neg. neg.
27 1/80 f 9 Golden
Retriever commerciale città n.p. neg. neg.
28 1/40 f 13 Bassotto commerciale città carcinoma
mamm. neg. neg.
29 1/20 m 2 Meticcio casalinga campagna n.p. neg. neg.
30 1/20 m 10 Epagneul
Breton casalinga campagna n.a. neg. neg.
31 1/40 f 5 Springer Spaniel Inglese
mista città n.p. neg. neg.
32 1/80 fs 5 Meticcio commerciale città n.a. neg. neg.
33 1/40 ms 14 Setter Inglese
commerciale città insuff. epatica e
renale 1/160 neg.

54
34 1/40 fs 11 Segugio Italiano
casalinga campagna n.p. neg. neg.
35 1/40 ms 9 Setter Inglese
casalinga campagna n.a. neg. neg.
36 1/40 ms 15 Meticcio mista campagna n.p. neg. neg.
37 1/40 m 10 American Pit Bull Terrier
commerciale semicampag
na mastocitoma neg. neg.
38 1/40 m 9 Deutscher
Boxer commerciale
semicampagna
n.p. neg. neg.
39 1/40 f 6 Meticcio casalinga campagna n.p. neg. neg.
40 1/40 ms 3 Meticcio commerciale città n.p. neg. neg.
41 1/80 f 5 Epagneul
Breton commerciale città n.a. neg. neg.
42 1/40 m 10 Meticcio mista campagna n.a. neg. neg.
43 1/40 ms 3 Golden
Retriever commerciale città n.p. neg. neg.
44 1/40 m s 12 Beagle commerciale città n.p. neg. neg.
45 1/40 m s 8 Weimaran
er commerciale città cataratta neg. 1/40
46 1/20 m 6 American Pit Bull Terrier
commerciale città n.p. neg. neg.
47 1/20 m 4 Weimaran
er mista campagna n.p. neg. neg.
48 1/80 f 8 Meticcio mista campagna n.p. neg. neg.
49 1/40 m 13 Labrador Retriever
commerciale città n.p. neg. neg.
50 1/40 fs 4 Golden
Retriever commerciale città n.p. neg. neg.
51 1/40 m 10 Meticcio commerciale città n.p. neg. neg.
52 1/20 m 4 Meticcio commerciale città n.p. neg. neg.
53 1/40 fs 14 Pastore Tedesco
commerciale città mielopatia
degenerativa neg. neg.
54 1/40 fs 8 Meticcio commerciale città n.a. neg. neg.
55 1/40 m 4 Meticcio mista campagna n.p. neg. 1/40
56 1/40 m 7 Meticcio mista semicampag
na n.p. neg. neg.
57 1/40 fs 8 Pastore
Maremmano
commerciale semicampag
na mastocitoma neg. neg.
58 1/40 fs 15 Bracco Italiano
mista semicampag
na n.p. neg. neg.
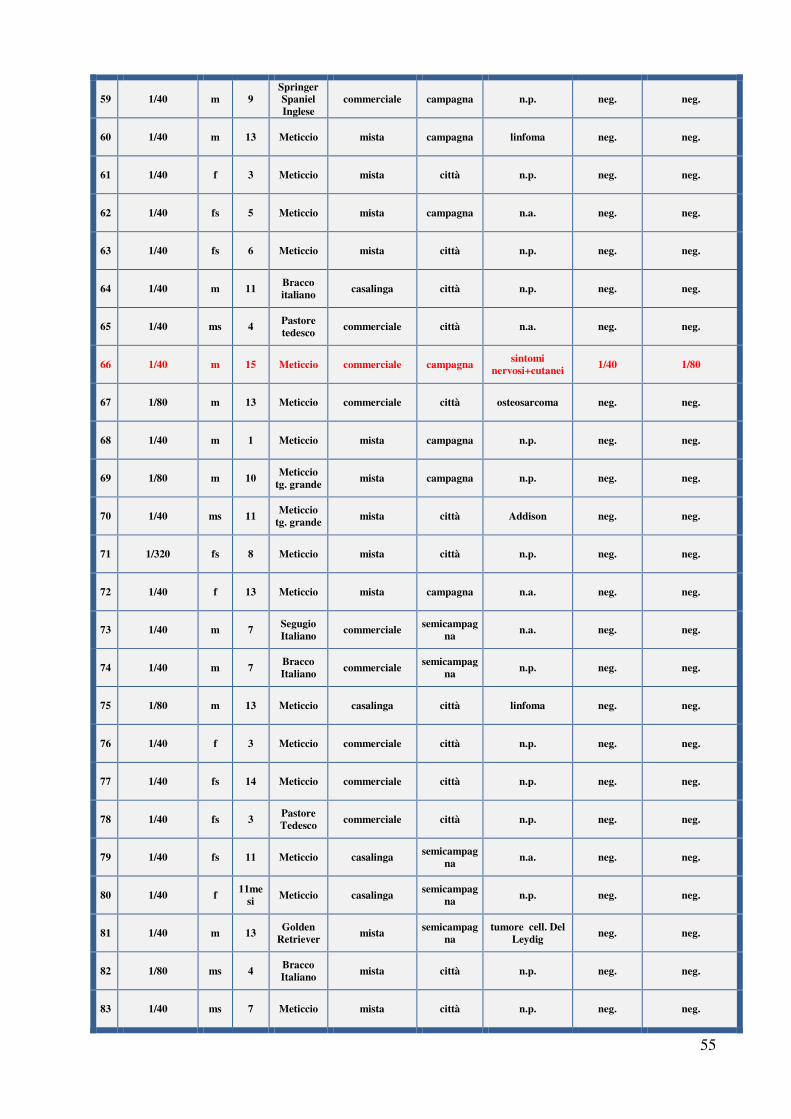
55
59 1/40 m 9 Springer Spaniel Inglese
commerciale campagna n.p. neg. neg.
60 1/40 m 13 Meticcio mista campagna linfoma neg. neg.
61 1/40 f 3 Meticcio mista città n.p. neg. neg.
62 1/40 fs 5 Meticcio mista campagna n.a. neg. neg.
63 1/40 fs 6 Meticcio mista città n.p. neg. neg.
64 1/40 m 11 Bracco italiano
casalinga città n.p. neg. neg.
65 1/40 ms 4 Pastore tedesco
commerciale città n.a. neg. neg.
66 1/40 m 15 Meticcio commerciale campagna sintomi
nervosi+cutanei 1/40 1/80
67 1/80 m 13 Meticcio commerciale città osteosarcoma neg. neg.
68 1/40 m 1 Meticcio mista campagna n.p. neg. neg.
69 1/80 m 10 Meticcio
tg. grande mista campagna n.p. neg. neg.
70 1/40 ms 11 Meticcio
tg. grande mista città Addison neg. neg.
71 1/320 fs 8 Meticcio mista città n.p. neg. neg.
72 1/40 f 13 Meticcio mista campagna n.a. neg. neg.
73 1/40 m 7 Segugio Italiano
commerciale semicampag
na n.a. neg. neg.
74 1/40 m 7 Bracco Italiano
commerciale semicampag
na n.p. neg. neg.
75 1/80 m 13 Meticcio casalinga città linfoma neg. neg.
76 1/40 f 3 Meticcio commerciale città n.p. neg. neg.
77 1/40 fs 14 Meticcio commerciale città n.p. neg. neg.
78 1/40 fs 3 Pastore Tedesco
commerciale città n.p. neg. neg.
79 1/40 fs 11 Meticcio casalinga semicampag
na n.a. neg. neg.
80 1/40 f 11me
si Meticcio casalinga
semicampagna
n.p. neg. neg.
81 1/40 m 13 Golden
Retriever mista
semicampagna
tumore cell. Del Leydig
neg. neg.
82 1/80 ms 4 Bracco Italiano
mista città n.p. neg. neg.
83 1/40 ms 7 Meticcio mista città n.p. neg. neg.

56
84 1/40 f 15 Meticcio mista campagna
sint.nervosi + crisi
epilettiche+sintomi cutanei
neg. 1/80
85 1/40 m 11 Meticcio casalinga città n.p. neg. neg.
86 1/40 m 11 Meticcio casalinga città n.p. neg. neg.
87 1/80 fs 9 Meticcio casalinga città n.a. neg. neg.
88 1/40 ms 5 Meticcio mista campagna n.p. neg. neg.
89 1/40 m 2 Meticcio commerciale semicampag
na sintomi nervosi
+dermatite 1/160 1/80
90 1/40 f 5 Bovaro del
Bernese casalinga città n.p. neg. neg.
91 1/40 m 14 Meticcio mista città trombocitopenia
immunom. neg. neg.
92 1/40 m 8 Meticcio mista semicampag
na n.p. neg. 1/40
93 1/40 m 13 Meticcio mista città n.p. neg. neg.
94 1/40 fs 11me
si Meticcio mista città n.p. neg. neg.
94 1/40 f 8 Pastore Tedesco
mista campagna insufficienza
renale neg. neg.
96 1/40 f 4 Meticcio casalinga campagna n.p. neg. neg.
97 1/80 m 2 Labrador Retriever
mista semicampag
na n.p. neg. neg.
98 1/80 ms 15 Epagneul
Breton casalinga
semicampagna
n.p. neg. neg.
99 1/40 fs 8 Meticcio mista semicampag
na n.a. neg. neg.
100
1/40 fs 11 Meticcio mista semicampag
na anemia emolitica neg. neg.
101
1/40 m 2 Segugio Italiano
casalinga semicampag
na n.p. neg. neg.
102
1/40 f 6 Bracco Italiano
casalinga città n.a. neg. neg.
103
1/80 f 11 Meticcio mista campagna leucemia neg. neg.
104
1/20 f 14 Meticcio casalinga campagna n.p. neg. neg.
105
1/40 ms 4 Springer Spaniel Inglese
commerciale città n.p. neg. neg.
106
1/20 fs 3 Meticcio
tg. grande casalinga campagna
disfunzione poliorgano
1/640 neg.
107
1/20 fs 2 Meticcio
tg. grande mista campagna n.p. neg. 1/640

57
108
1/40 ms 15 Setter Inglese
mista città osteosarcoma neg. neg.
109
1/40 m 12 Springer spaniel inglese
casalinga campagna n.p. neg. neg.
110
1/80 m 5 Setter inglese
casalinga città n.a. neg. neg.
111
1/40 m 8 Meticcio casalinga città n.p. neg. neg.
112
1/80 m 9 Meticcio casalinga campagna n.p. 1/160 neg.
113
1/40 m 8 Meticcio casalinga campagna sintomi nervosi neg. 1/40
114
1/20 m 4 Meticcio casalinga città n.p. neg. neg.
115
1/40 f 3 Labrador Retriever
mista città n.p. neg. neg.
116
1/80 m 7 Epagneul
Breton casalinga campagna tumore epatico neg. neg.
117
1/40 f 8 Golden
Retriever mista città n.p. neg. neg.
118
1/40 m 8 Pastore Tedesco
mista città n.p. neg. neg.
119
1/40 f 11 Meticcio mista città n.a. neg. neg.
120
1/80 f 6 Meticcio mista campagna sindrome autoim. 1/320 neg.
121
1/40 f 1 Bracco Italiano
casalinga semicampag
na n.p. neg. 1/640
122
1/40 m 9 Setter Inglese
mista semicampag
na n.p. neg. neg.
123
1/40 fs 10me
si Meticcio mista campagna neoplasia neg. neg.
124
1/40 f 4 Meticcio
tg. grande casalinga città n.p. neg. neg.
125
1/40 m 6 Meticcio commerciale semicampag
na n.p. neg. neg.
126
1/80 ms 11 Meticcio
tg. grande commerciale
semicampagna
n.p. neg. neg.
127
1/40 ms 5 Meticcio
tg. grande casalinga campagna n.p. neg. neg.
128
1/40 ms 9 Meticcio
tg. grande casalinga campagna anemia emolitica neg. neg.
129
1/40 ms 5 Meticcio casalinga città n.p. neg. neg.
130
1/80 m 7 Meticcio casalinga campagna n.p. neg. neg.
131
1/40 m 9 Labrador Retriever
mista città n.p. neg. neg.
132
1/40 ms 11 Setter Inglese
mista città insuf.mitralica neg. neg.
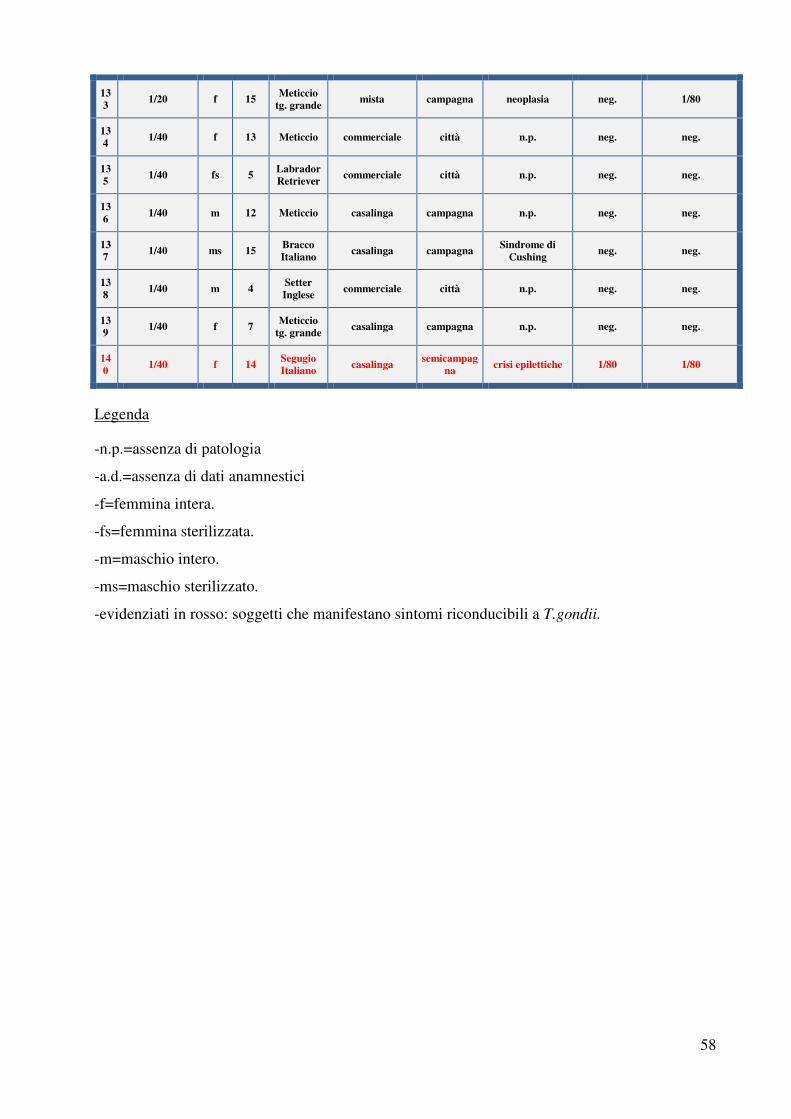
58
133
1/20 f 15 Meticcio
tg. grande mista campagna neoplasia neg. 1/80
134
1/40 f 13 Meticcio commerciale città n.p. neg. neg.
135
1/40 fs 5 Labrador Retriever
commerciale città n.p. neg. neg.
136
1/40 m 12 Meticcio casalinga campagna n.p. neg. neg.
137
1/40 ms 15 Bracco Italiano
casalinga campagna Sindrome di
Cushing neg. neg.
138
1/40 m 4 Setter Inglese
commerciale città n.p. neg. neg.
139
1/40 f 7 Meticcio
tg. grande casalinga campagna n.p. neg. neg.
140
1/40 f 14 Segugio Italiano
casalinga semicampag
na crisi epilettiche 1/80 1/80
Legenda
-n.p.=assenza di patologia
-a.d.=assenza di dati anamnestici
-f=femmina intera.
-fs=femmina sterilizzata.
-m=maschio intero.
-ms=maschio sterilizzato.
-evidenziati in rosso: soggetti che manifestano sintomi riconducibili a T.gondii.

59
Capitolo 11 Discussione e conclusioni
La prevalenza da noi osservata si attesta intorno all’8%. I nostri dati, sono coerenti con parte
della letteratura consultata. Studi sulla sieroprevalenza di T.gondii nel cane riguardano
soprattutto la Cina e il Brasile, paesi dove questa patologia ha un importante impatto sulla salute
pubblica. In Cina è usuale un’alimentazione con carne di cane,questo rappresenta un rischio
elevato per la sanità pubblica e giustifica il numero elevato di indagini fatte in questo paese.
Sono presenti, tuttavia, anche alcuni studi che riguardano la siero prevalenza in Canada, negli
USA (in particolare in America Centrale), in Turchia, in Malesia e Arabia Saudita. Per quanto
concerne l’Europa, i dati provengono soprattutto dalle Baleari (Spagna), dall’Austria, dalle
Repubbliche Ceca e Slovacca e dalla Svezia. Dati italiani sono assenti. La prevalenza evidenziata
in questi lavori è molto variabile soprattutto per quanto riguarda la Cina, dove per ogni area
geografica, sono stati raccolti dati con i seguenti risultati: • nord-est 10%
• est 21,5%
• nord-ovest 11,6%
• ovest 10,42%
• sud-ovest 21,6%
Le titolazioni anticorpali sono medie, con il 50% dei sieri che ha un titolo di 1:20, il 32% 1:40, il
14% 1:80 e a 1:160 il 4% (Wu et al., 2011; Gang Duan et al., 2012; Wang et al., 2012 Yongmei
et al., 2012 ;Yang et al., 2013). Anche in Corea, così come nell’isola di Taiwan la siero
prevalenza è intorno ai 20% ( Nguyen et al., 2012; Tsai, 2008); in quest’ultima i sieri hanno una
titolazione molto alta:
� 122 con 1:32;
� 75 con 1:74;
� 28 con 1:128;
� 36 con 1:256;
� 15 con 1:512;
� 5 con 1 :1024;
� 3 con 1:2048.
In Brasile la sieroprevalenza di T.gondii nei cani si attesta intorno al 52% nell’area
amazzonica,di 21,8% nello stato di San Paolo, a sud del paese e del 22% nella zona sud

60
occidentale (Langoni et al., 2013;Plugge et al., 2011;Minervino et al., 2012); la titolazione
anticorpale in questo paese è alta, 1:200.
Oltre al Brasile, la siero prevalenza in America Centrale, in particolare nello Stato di Grenada, si
attesta intorno a 35,7% anche in un’altra zona, Stato di Trinitad e Tobago,33% (Ali et al.,2003).
La titolazione anticorpale riscontrata è molto alta: 8% ha titolo 1:3200, 10% 1:1600, 13 % 1:100,
10% 1:50 e 8% 1:25.
Per quanto riguarda gli Stati Uniti i dati prevalenti provengono dal Maryland dove la prevalenza
è del 32,2% (Child et Seegar, 1996) e dalla Virginia dove i cani positivi sono il 21% con
titolazione 1:50 (Rosypal et al., 2010); in Canada la prevalenza sfiora il 30% (Salb et al., 2008).
In Messico i cani risultati positivi sono stati 51,5% (Dubey et al., 2007), che con il Brasile sono i
paesi con più alta siero prevalenza di T.gondii nei cani; i titoli anticorpali riportati nello studio
messicano sono medio alti:
• 1:25 in 27 campioni;
• 1:50 in 11;
• 1:100 in 5;
• 1:200 in 4;
• 1:400 in 2;
• 1:800 in 2 ;
• 1:3200 in 1 .
(Dubey et al., 2007)
Studi del 2005 riportano una siero prevalenza nella città di Ankara, in Turchia, del 2,58% (
Aslantas et al., 2005), mentre più recentemente è stata valutata una percentuale del 15% nella
provincia di Kirikkale, nel centro dell’Anatolia con una titolazione di 1:16 (Yildiz et al., 2009). I
dati che provengono dalla Malesia e dall’Arabia Saudita sono molto bassi 9,6%
(Chandrawathani, 2008) e 3% ( Hossain, 1986), anche se la titolazione è alta (1:200).
In Europa la sieroprevalenza si attesta intorno al 59% sull’isola di Maiorca,in Spagna, (Cabezon
et al.,2010), 26% in Austria (Wanha et al., 2005), 20% in Repubblica Ceca e Slovacca (Hejlícek
et al., 1995)e 38 % in Portogallo.
I titoli anticorpali evidenziati in Spagna sono sostanzialmente bassi (1:25) ( Cabezon et al.,
2010), in Austria molto alti, infatti circa il 10% dei sieri hanno titolo 1:6400 (Wanha et
al.,2005). In Portogallo il 45,6 % dei campioni positivi ha titolo alto, infatti in 115 su 252 è di
1:160 (Lopes et al. , 2011).

61
Alla luce dei dati riportati, la sieroprevalenza riscontrata nella provincia di Pisa è pienamente in
accordo con la media delle altre aree analizzate ed anche le percentuali più basse, come quelle
della provincia di Livorno e Genova,non si discostano più di tanto dai dati più bassi riscontrati.
I dati riguardanti i singoli fattori di rischio sono pienamente concordanti; i campioni positivi
sono sia maschi che femmine, senza differenza significativa e per tutti gli autori il genere non è
associato all’insorgere dell’infezione da T.gondii nel cane (Yongmei et al., 2012; Na Yang et
al.,2013; Dubey et al., 2007; Wahna et al., 2005;Aslatas et al., 2005; Lopes et al., 2011).
Questo lavoro è stato possibile grazie alla collaborazione attenta e precisa delle strutture
veterinarie contattate, che mi hanno fornito i dati anamnestici sui cani in esame. La mia
attenzione si è concentrata su alcune informazioni fornitemi, che possono essere considerate o
essere solo apparentemente, i fattori di rischio della toxoplasmosi; avendo esclusivamente l’
anamnesi dei cani positivi, i dati rappresentano solo un’apparente tendenza della sieropositività.
A questo proposito appare interessante notare che l’unico campione con titolo anticorpale
elevato (il N.71 con 1/320) non presentava alcuna sintomatologia. Il fatto che a titoli anticorpali
bassi corrispondano segni clinici compatibili con toxoplasmosi potrebbero indicare una fase
acuta dell’infezione, come peraltro i titoli bassi degli animali asintomatici potrebbero indicare
fase acuta o essere il risultato di un’infezione pregressa.
La maggior parte dei campioni positivi appartengono a cani maschi, tuttavia la percentuale delle
femmine non vi si discosta troppo(42%femmine, 58%maschi); questo non è un dato molto
rilevante,sia perché i sieri inizialmente analizzati erano soprattutto di maschi, sia perché non
valuto il genere un elemento discriminante per l’infezione.
L’età,invece,è un carattere determinante in quest’analisi; il 51% dei positivi sono cani anziani,
che hanno un’età superiore agli 8 anni, mentre solo il 5% sono giovani, fino all’anno di età. L’età
più elevata giustifica la maggior prevalenza, poiché, probabilmente, questi cani hanno avuto
maggiori occasioni per poter venire a contatto con il parassita, rispetto a quelli dai 2 agli 8
anni,che comunque hanno una percentuale elevata (44%); in queste due classi l’infezione è
sicuramente post natale acquisita. Tra i cuccioli solo 7 sono positivi e è alta la probabilità che
siano stati infettati o per via congenita o transcolostrale, ma non è esclusa una nuova infezione.
I meticci e i cani “di razza” positivi sono in sostanziale equilibrio, ma il dato rilevante è che in
entrambe le categorie siano presenti cani di taglia medio grande e comunque di indole attiva;
poiché l’elemento essenziale per il contagio postnatale è il contatto con animali o aree infette è
evidente l’importanza dell’attitudine e del movimento di questi cani. 33 soggetti appartengono a
razze prettamente utilizzati per la caccia, come il “Epagneul Breton ” o il ”Segugio Italiano”che
vengono a contatto con prede selvatiche e con feci di felini, durante la loro attività venatoria. Tra
i positivi si evidenziano,soprattutto, il Pastore Tedesco, Pitt Bull, Rottweiler, Pastore

62
Maremmano,razze dinamiche e vivaci, mentre solo 1-2 soggetti possono essere definiti ”da
appartamento”, quali per esempio la razza maltese.
Le caratteristiche che maggiormente rappresentano un fattori di rischio sono l’habitat di vita e il
tipo di alimentazione , che sono spesso collegati; i cani che vivono in campagna sono quelli
maggiormente presenti( 72), mentre quelli che vivono in aree urbane sono solo 69.27 cani
provengono da zone non definite città, perché in paesi piccoli o comunque a contatto frequente
con aree aperte e animali selvatici. Dalle mie indagini ho potuto evincere che molti soggetti che
vivono in città vengono portati con frequenza in aree rurali e che solo una piccola percentuale ha
effettivamente una vita strettamente urbana. La prevalenza maggiore si evidenzia nella provincia
di Pisa,dove la percentuale nella sola città è del 13,2%,giustificata però dalla geografia di questa
zona, che presenta molti piccoli paesi semirurali al confine con la città, appartenenti però allo
stesso comune. Sono molto più basse le percentuali di Livorno(8,39%), di Genova (8,42%) e
Firenze (3,59%), dove i comuni sono costituiti prevalentemente da sole zone urbane.Un dato
rilevante è quello riguardante l’alimentazione: solo 49 cani vengono abitualmente alimentati con
cibo prettamente commerciale , mentre a 51 soggetti è fornito un cibo misto, cioè sia acquistato,
in preparati umidi o secchi,sia casalingo; in quest’ultima categoria rientrano sia gli avanzi della
tavola sia alimenti preparati per il cane. 40 soggetti sono alimentati solo con preparati
casalinghi. Questi dati sono molto importanti poiché una delle vie di trasmissione più frequente è
proprio alimentazione con carni crude o poco cotte.
L’anamnesi sanitaria dei cani è fondamentale per poter capire quali siano maggiormente colpiti;
purtroppo non ho potuto risalire allo stato di salute di tutti i campionati. Dai risultati ottenuti è
possibile evidenziare come tra i cani esaminati, 44 siano affetti da patologie gravi,croniche e
debilitanti; è stato interessante notare che 9 soggetti presentano sintomi associabili a
toxoplasmosi, quali crisi epilettiche, sindromi cerebrali, alterazione della vista e patologie
dermiche. Per molti di essi non era ancora stata definita una diagnosi certa,vista la variabilità dei
sintomi. Tra i soggetti “sintomatici”i titoli anticorpali non sono alti (8 a 1/40,mentre 1 a 1/20) e
8 sono risultati positivi anche a Neospora e Leishmania.
19 cani presentano possibili coinfezioni con sintomatologia più o meno evidente:
• 3 sono sieropositivi per Toxoplasma, Neospora e Leishmania e hanno sintomi nervosi,
crisi epilettiche e dermatite( 2%); In tutti questi casi il titolo anti toxoplasma è inferiore a
quello anti neospora, pertanto è legittimo sospettare che si tratti di cross reazioni. Per
quanto riguarda leishmania i cani N. 89 e 140 hanno titoli positivi, per quanto riguarda il
N. 66 sarebbero richieste ulteriori indagini per confermare.

63
• 6 cani(4%) mostrano solo anticorpi per Leishmania e Toxoplasma di cui 1 è affetto da
dermatiti,insufficienza renale e sintomi nervosi;
• 10 cani sono positivi per Toxoplasma e Neospora(10%), di cui 4 presentano sintomi
nervosi,cerebrali e cataratta e i restanti 6 sono affetti da patologie degenerative. Anche in
questo caso la metà dei campioni presentano titoli più elevati per Neospora facendo
sospettare una cross reazione nei confronti di Toxoplasma. La prevalenza totale dei
positivi a Toxoplasma è dell’ 8% per Neospora, mentre per Leishmania del 6%.
È importante sottolineare, però, che visti i bassi titoli anticorpali di Neospora nei
campioni analizzati, non è possibile escludere una cross-reazione anticorpale.
La maggior parte degli studi concordano sul fatto che la siero prevalenza si evidenzi soprattutto
in cani sopra ai 6-8 anni per la maggior possibilità di contatto con ambiente infetto; viene,
quindi, considerato un fattore di rischio l’habitat del cane, evidenziando l’importante differenza
tra le positività riscontrate in zone prettamente urbane e quelle rurali o semi rurali (Yongmel et
al., 2012; Na Yang et al., 2013; Dubey et al., 2007; Dubey et al., 2008; Ali et al., 2003; Nguyen
et al., 2012; Lopes et al., 2011; Gang Duan et al., 2012; Wang et al., 2012; Langoni et al., 2013).
I dati raccolti in questo lavoro concordano anche per quanto riguarda l’alimentazione del cane;
sussiste una differenza significativa tra soggetti che ricevono un’alimentazione solo commerciale
e quelli che si cibano di avanzi (soprattutto visceri crudi e poco cotti) o con alimentazione
casalinga ( Lopes et al., 2011;Duan et al., 2012;Ali et al., 2003).
Le coinfezioni tra T.gondii con N.caninum e L.infantum sono molto frequenti e sono numerosi i
lavori sul tema,soprattutto effettuati in Brasile, Corea, Cina e Turchia. T.gondii e N.caninum
sono molto simili, sia nel ciclo, sia per i sintomi sia per la distribuzione geografica e lo sono
anche i fattori di rischio che le favoriscono; questo potrebbe essere il motivo della loro frequente
presenza in associazione (Minervino et al., 2012). Per quanto riguarda L.infantum l’elevata
presenza potrebbe essere collegata all’immunodepressione da essa determinata, che può favorire
l’infezione malattie concomitanti. Tutti gli studi concordano sul fatto che, anche nelle
confezioni, i fattori di rischio sono prevalentemente l’habitat di vita, l’età e il tipo di
alimentazione (Nguyen et al., 2012;Plugge et al., 2011; Minervino et al., 2012); in pochi studi,
tuttavia, è stato evidenziato come N.caninum sia meno influenzata dall’ambiente rispetto a
T.gondii (Yildiz et al., 2009).
Nelle coinfezioni sono frequenti forme nervose e cutanee, come risulta anche da questo studio,
spesso difficili da curare e anche da diagnosticare in modo certo, visto la similarità e la
variabilità dei sintomi (Hoffman et al., 2012).
La maggior parte della bibliografia riporta l’infezione crociata di T.gondii e N.Caninum,con
sintomi nervosi e cutanei similari(Dubey et al., 2007)e con le seguenti percentuali di confezione :

64
• 4,97%Stato di San Paolo (Brasile) (Langoni et al., 2013);
• 2,72% Stato di Paranà (Plugge et al., 2011 );
• 0,72 Corea (Nguyen et al., 2012);
• 1,7% Austria (Wanha et al., 2005)
Le percentuali evidenziate in questo lavoro sono più alte,ma , come detto in precedenza, visti i
bassi livelli anticorpali, non può essere esclusa una cross-reazione.
T.gondii è considerata una delle principali cause di sintomi neurologici del cane e N.caninum
può essere valutata come diagnosi differenziale o come infezione concomitante e la
differenziazione deve essere eseguita con metodi sierologici ( Langoni et al., 2012). Studi
crociati T.gondii e L.infantum sono soprattutto brasiliani e mettono in evidenza la stretta
connessione tra l’immunodepressione sostenuta da Leishmania e l’insorgere d’ infezione
sostenuta Toxoplasma( Paulan et al., 2013). La prevalenza globale di Leishmania, cioè su tutti i
sieri esaminati, è di circa 20%, ciò potrebbe significare che la coinfezione può rappresentare un
campione casuale di sieri di animali viventi in zona endemica.

65
Bibliografia
ABU-MADI, M.A., AL-MOLAWI, N., BEHNKE, J.M.,
Seroprevalence and epidemiological correlates of Toxoplasma gondii infections among patients
referred for hospital-based serological testing in Doha, Qatar.
2008 Parasitol. Vectors 1, 39.
ALI CN,HARRIS JAwATKINS JD,ADESIYUN AA,
Serepidemiology of Toxoplasma gondii in dogs in Trinidad and Tobago
2003 Vet.Parassitol.113 (3-4):179-87
AL-QURASHI, A.R., GHANDOUR, A.M., OBEID, O.E., AL-MULHIM, A., MAKKI,
S.M.,Seroepidemiological study of Toxoplasma gondii infection in the human
population in the Eastern Region.
2001 Saudi Med. J. 22, 13–18.
ARKUSH K.D., M.A. MILLER, C.M. LEUTENEGGER, I.A. GARDNER, A.E. PACKHAM,
A.R. HECKEROTH, A.M. TENTER, B.C. BARR AND P.A. CONRAD
Molecular and bioassay-based detection of Toxoplasma gondii oocyst uptake by mussels
(Mytilus galloprovincialis),
2003 Int. J. Parasitol. 33, pp. 1087–1097.
AJZENBERG D, COGNÉ N, PARIS L, BESSIÈRES MH, THULLIEZ P, FILISETTI D,
PELLOUX H, MARTY P, DARDÉ ML.,
Genotype of 86 Toxoplasma gondii isolates associated with human congenital toxoplasmosis,
and correlation with clinical findings.
2002 J Infect Dis. 1;186(5):684-9.
AJZENBERG D., A.L. BANULS, C. SU, A. DUMETRE, M. DEMAR, B. CARME AND M.L.
DARDE,
Genetic diversity, clonality and sexuality in Toxoplasma gondii
2004 Int. J. Parasitol. 34, pp. 1185–1196

66
APPLEFORD, P. J., AND J. E. SMITH.,
Toxoplasma gondii: the growth characteristics of three virulent strains.
1997 Acta Trop. 65:97–104.
ASLANTAŞ O1, OZDEMIR V, KILIÇ S, BABÜR C.,
Seroepidemiology of leptospirosis, toxoplasmosis, and leishmaniosis among dogs in Ankara,
Turkey.
2005 Vet Parasitol. ;129(3-4):187-91
BARIL, L., ANCELLE, T., GOULET, V., THULLIEZ, P., TIRARD-FLEURY, V., CARME,
B.,
Risk factors for Toxoplasma infection in pregnancy: a case-control study in France.
1999 Scand. J. Infect. Dis. 31, 305–309.
BEATTY J, BARRS V.,
Acute toxoplasmosis in two cats on cyclosporin therapy
2003 Aust Vet J 81:339
BELFORT-NETO R, NUSSENBLATT V, RIZZO L, MUCCIOLI C, SILVEIRA C,
NUSSENBLATT R, KHAN A, SIBLEY LD, BELFORT R JR.,
high prevalence of unusual genotypes of toxoplasma gondii infection in pork meat samples from
erechim, southern brazil.
2007,An Acad Bras Cienc. 79(1):111-4.
BENARD, A., PETERSEN, E., SALAMON, R., CHENE, G., GILBERT, R., SALMI,
L.R.EUROPEAN TOXO -PREVENTION STUDY GROUP (EUROTOXO)
Survey of European programmes for the epidemiological surveillance of congenital
toxoplasmosis. 2008 Euro Surveill. 13, 257–263.
BERNSTEEN L, GREGORY CR, ARONSON LR, ET AL
Acute toxoplasmosis following renal transplantation in three cats and a dog
1999 J Am Vet Med Assoc 215:1123-1126

67
BEVERLY JKA
A rational approach to the treatment of Toxoplasma uveitis
1958 Trans. Opthalmol. Soc. U.K. 78, 109–121
BEVERLEY J.K.A.
Congenital transmission of toxoplasmosis through successive generations of mice.
1959 Nature. 183: 1348-1349
BIOCCA E.
Some notes on classification of Protozoa and the usefulness of creating a new class of Protozoa
Rev Bras Malariol Doencas Trop. 1956 Jan;8(1):91-102.
BLEWETT DA, MILLER JK, HARDING J.
Simple technique for the direct isolation of toxoplasma tissue cysts from fetal ovine brain.
1983 Vet Rec.112(5):98-100.
BOIA M.N.,CARVALHO-COSTA F.A.,SODRE F.C.,PINTO G.M.,AMENDOEIRA M.R.
Seroprevalence of toxoplasma gondii infection among Indian people living in Lauarete ,Sao
gabriel de Cachoeir ,Amazonas Brazil.
2008 Rev.Ist.Med.Trp.Sao paulo 50,17-20.
BONAMETTI AM, PASSOS J DO N, DA SILVA EMK, BORTOLIERO AL
Surto de toxoplasmose aguda transmitida através da ingestão de carne crua de gado ovino.
1997a Rev Soc Bras Med Trop 30: 21-25
BONHOMME, A., L. PINGRET, AND J. M. PINON
Review: Toxoplasma gondii cellular invasion.
1992 Parassitologia 54:31–43.
BOURATBINE, A., SIALA, E., CHAHED, M.K., AOUN, K., BEN ISMAIL, R.,
Seroepidemiologic profile of toxoplasmosis in northern Tunisia.
2001 Parasite 8, 61–66

68
BRANDONISIO O.
Toxoplasmosi in parassiti e parassitosi umane dalla clinica al laboratorio
2006 Scaglia m.,Gatti s,Rondanelli e.g.,69-88 Ed selecta medica.
BUXTON D. ,
Ovine toxoplasmosis: a review.
1990 J. R. Soc Med, 83: 509-511.
CABRAL A.S.G.,BALBINO F.,ABDALLA L.F.COSTA S.S.S.,VAZ J:A.R.,FONSECA S.F
Seroprevalence of Toxoplasmosis in women of reproductive age in the city of Brasilia ,caoital of
Brazil.
2008 13 th International congress of infectious diseases,Kuala Lumpur Malaysia ,abstract 179
CARME B., M. DEMAR, D. AJZENBERG AND M.L. DARDE ,
Severe acquired toxoplasmosis caused by wild cycle of Toxoplasma gondii
2009 French Guiana, Emerg. Infect. Dis. 15, pp. 656–658 68
CHANDRAWATHANI P1, NURULAINI R, ZANIN CM, PREMAALATHA B, ADNAN
M, JAMNAH O, KHOR SK, KHADIJAH S, LAI SZ, SHAIK MA, SEAH TC, ZATIL SA.
Seroprevalence of Toxoplasma gondii antibodies in pigs,
goats, cattle, dogs and cats in peninsular Malaysia
2008 Tropical Biomedicine 25(3): 257–258
CHEN, X.G., WU, K., LUN, Z.R.,.
Toxoplasmosis researches in China.
2005 Chin. Med. J. (Engl.) 118, 1015–1021.
CHIAPPINO, M. L., B. A. NICHOLS, AND G. R. O’CONNOR.
Scanning electron microscopy of Toxoplasma gondii: parasite torsion and host-cell
responses during invasion.
1984 J. Protozool. 31:288–292.

69
CHRISTIE, E., P. W. PAPPAS, AND J. P. DUBEY.
Ultrastructure of excystment of Toxoplasma gondii oocysts
1978 J. Protozool. 25:438–443.
CONRAD PA, M.A. MILLER, C. KREUDER, E.R. JAMES, J.MAZET, H. DABRITZ, D.A.
JESSUP, F. GULLAND AND M.E. GRIGG
Transmission of Toxoplasma: clues from the study of sea otters as sentinels of Toxoplasma
gondii flow into the marine environment,
2005 Int. J. Parasitol. 35, pp. 1155–1168
DABRITZ HA, CONRAD PA.
Cats and Toxoplasma: implications for public health.
2010 Zoonoses Public Health.;57(1):34-52. Epub 2009 Sep 10. Review.
CONTRERAS M.,SCHENONE H.,SALINAS P.,SANDOVAL L.,ROYAS A,VILLARROEL
F,SOLIS F.,
Seroepidemiology of human toxoplasmosis in chile
1996 Rev.Ist.Med.Trop,san Paulo 38,431-435.
COOK AJ, GILBERT RE, BUFFOLANO W, ZUFFEREY J, PETERSEN E, JENUM PA,
FOULON W, SEMPRINI AE, DUNN DT
Sources of Toxoplasma infection in pregnant women: a European multicentre case-control
study.
2000 Br Med J 15: 142-147.
COONS A H, ET AL.
Immunological properties of an antibody containing a fluorescent group.
1941 Proc Soc e xp Biol Med ;47,200-2
COONS A H AND KAPLAN MH
Localization of antigen in tissue cells. II.Improvements in a method for the detection of antigen
by means of fluorescent antibody.
1950 Journal of e xp Med 1950; 91,1

70
CURSONS, R.M., CEPULIS, S., NAICKER, G., HASLAM, A.,
The prevalence of Toxoplasma antibodies
1982
DA SILVA AC, PEREIRA TDA C.,
Characteristics and current direct costs of hospital admissions due to occupational accidents in
the southwest of Bahia
2005 Rev Bras Epidemiol.;17(2):381-94. English, Portuguese.
DAVIDSON MG, ROTTMAN JB, ENGLISH RV, ET AL
Feline immunodeficiency virus predisposes cats to acute generalized toxoplasmosis
1993 Am J Pathol 143:1486-1497
DEHKORDI FS, BORUJENI MR, RAHIMI E, ABDIZADEH R
Detection of toxoplasma gondii in raw caprine, ovine, buffalo, bovine, and camel milk using cell
cultivation, cat bioassay, capture elisa, and pcr methods in iran.
2012 Foodborne Pathog Dis. 10(2):120-5.
DE LA ROSA M.,BOLIVAR J.,PEREZ H.A.,
Toxoplasma gondii infection in Amerindians of Venezuelan Amazon,
1999 Medicina (B.Aires)59,759-762.
DEMAR M, HOMMEL D, DJOSSOU F, PENEAU C, BOUKHARI R, LOUVEL D,
BOURBIGOT AM, NASSER V, AJZENBERG D, DARDE ML, CARME B.
Acute toxoplasmoses in immunocompetent patients hospitalized in an intensive care unit in
French Guiana.
2012 Clin Microbiol Infect. (7):E221-31. doi: 10.1111/j.1469-0691.2011.03648.x.
DE ROEVER-BONNET, H.
Congenital Toxoplasma infections in mice and hamsters infected with avirulent and virulent
strains.
1979 Tropical and Geographical Medicine . 21: 443-450

71
DE SOUZA W, SOUTO-PADRÓN T.
Ultrastructural localization of basic proteins on the conoid, rhoptries and micronemes of
Toxoplasma gondii.
1978 Z Parasitenkd. 4;56(2):123-9.
DHUMNE, M., SENGUPTA, C., KADIVAL, G., RATHINASWAMY, A., VELUMANI, A.,.
National seroprevalence of Toxoplasma gondii in India
2007. J. Parasitol. 93, 1520–1521.
DUBEY JP, MILLER NL, FRENKEL JF,
The Toxoplasma gondii oocyst from cat feces.
1970 J Exp Med. Oct 1;132(4):636-62.
DUBEY, J. P., AND J. K. FRENKEL
Feline toxoplasmosis from acutely infected mice and the development of Toxoplasma cysts.
1976 J. Protozool. 23:537–546.
DUBEY, J. P.
Toxoplasma, Hammondia, Besnoitia, Sarcocystis, and other tissue cyst-forming coccidia of man
and animals, p. 101–237.
1977 In J. P. Kreier (ed.), Parasitic protozoa, 3rd ed. Academic Press, Inc., New York,
DUBEY JP
A review of toxoplasmosis in pigs.
1986Vet Parasitol,1986, 19:181.
DUBEY JP, AND C. P. BEATTIE
Toxoplasmosis of Animals and Man
1988 CRC Press Boca Raton, Florida
DUBEY J.P.,KOESTERN A.,PIPER RC,
Repeated transplacental trasmission of Neospora Caninum in dogs
1990Mjavma ,194,:857-860

72
DUBEY, J.P.
Toxoplasmosis --an overview.
1991 Southeast Asian Journal of Tropical Medicine and Public Health. 22 Suppl: 88-92.
DUBEY, J. P.
Toxoplasma, Neospora, Sarcocystis, and other tissue cyst-forming coccidia of humans and
animals, p. 1–158. In J. P. Kreier (ed.),
1993 Parasitic protozoa, vol. 6. Academic Press, Inc., New York, N.Y.
DUBEY, J.P. AND CARPENTER, J.L.
Neonatal toxoplasmosis in littermate cats.
1993 Journal of the American Veterinary Medical Assoc iation. 203: 1546-1549
DUBEY, J.P., WEIGEL, R.M., SIEGEL, A.M., THULLIEZ, P., KITRON, U.D.,
MITCHELL, M.A., MANNELLI, A., MATEUS -PINILLA, N.E., SHEN, S.K.,
KWOK, O.C. AND TODD, K.S.,
Sources and reservoirs of Toxoplasma gondii infection on 47 swine farms in Illinois.
1995 Journal of Parasitology. 81: 723-729.
DUBEY J.P. ,
Infectivity and pathogenicity of Toxoplasma gondii oocysts for cats
1996 J. Parasitol. 82 pp. 957–960
DUBEY, J.P., SHEN, S.K., KWOK, O.C. AND THULLIEZ, P. (1997)
Toxoplasmosis in rats (Rattus norvegicus): congenital transmission to
first and second generation offspring an isolation of Toxoplasma gondii
from seronegative rats.
1997 Parasitology. 115: 9-14.
DUBEY, J.P. AND LAPPIN, M.R.
Toxoplasmosis and Neosporosis.
1998 In Infectious Diseases of the Dog and Cat . Greene, C.E. (ed.)
Philadelphia: W. B. Saunders, pp. 493 -509.

73
DUBEY j.p., , D. S. LINDSAY Y2, AND C. A. SPEER3
Structures of Toxoplasma gondiiTachyzoites, Bradyzoites, and Sporozoites and Biology and
Development of Tissue Cysts†
2000 Clin. Microbiol. Rev. April 1998 vol. 11 no. 2 267-299
DUBEY J.P.
Review of Neospora caninum and neosporosis in animals.
2003 Korean Journal of Parasitology,
DUBEY JP, R. ZARNKE, N. J. THOMAS, S. K. WONG, W. VAN BONN, M. BRIGGS, J. W.
DAVIS, R. EWING, M. MENSEA, O. C. H. KWOK, S. ROMAND, AND P. THULLIEZ
Toxoplasma gondii, Neospora caninum, Sarcocystis neurona and Sarcocystis canis-like
infections in marine mammals
2003b Vet. Parasitol. 116, 275–296
DUBEY JP
Unexpected oocyst shedding by cats fed Toxoplasma gondii tachyzoites: in vivo stage
conversion and strain variation
2005 Vet. Parasitol. 133, 289–298
DUBEY JP
Comparative infectivity of oocysts and bradyzoites of Toxoplasma gondii for intermediate
(mice) and definitive (cats) hosts
2006 Vet.Parasitol. 140 , pp. 69
DUBEY JP, X.Q. ZHU, N. SUNDAR, H. ZHANG, O.C. KWOK AND C. SU
Genetic and biologic characterization of Toxoplasma gondii isolates of cats from China
2007 Vet. Parasitol. 145 , pp. 352–35
DUBEY JP, LT. HUONG, BW. LAWSON, D.T. SUBEKTI, P. TASSI, W. CABAJ, N.
SUNDAR, G.V. VELMURUGAN, O.C. KWOK AND C. SU
Seroprevalence and isolation of Toxoplasma gondii from free-range chickens in Ghana,
Indonesia, Italy, Poland, and Vietnam
2008a J. Parasitol. 94, pp. 68–71

74
DUBEY JP, JONES JL
Toxoplasma gondii infection in humans and animals in the United States
2008b Int J Parasitol 38: 1257-1278
DUBEY JP
History of the discovery of the life cycle of Toxoplasma gondii.
2009a
CHOUDHARY S, ZIEGER U, SHARMA RN, CHIKWETO A, TIWARI KP, FERREIRA LR,
OLIVEIRA S, BARKLEY LJ, VERMA SK, KWOK OC, SU C, DUBEY JP
Isolation and RFLP genotyping of Toxoplasma gondii from the mongoose (Herpestes
auropunctatus) in Grenada, West Indies.
2013 J Zoo Wildl Med. ;44(4):1127-30.
DUBREMETZ, J. F., AND J. D. SWARTZMAN
Subcellular organelles of Toxoplasma gondii and host cell invasion.
1993 Res. Immunol. 144:31–33.
DUNCANSON P, TERRY RS, SMITH JE, HIDE G.
High levels of congenital transmission of Toxoplasma gondii in a commercial sheep flock.
2001 Int J Parasitol.;31(14):1699-703.
DUNN DT, LIGHTMAN S, MURRAY PI, PAVESIO CE, GORMLEY PD, MASTERS J,
PARKER SP, STANFORD MR.
Incidence of symptomatic toxoplasma eye disease: aetiology and public health implications.
1999Epidemiol Infect.;123(2):283-9.
ELAMIN EA, ELIAS S, DAUGSCHIES A, ROMMEL M.
Prevalence of Toxoplasma gondii antibodies in pastoral camels (Camelus dromedarius) in the
Butana pla ins, mid-Eastern Sudan
1992 Vet Parasitol.;43(3-4):171-5.
EUROPEAN FOOD SAFETY AUTHORITY,
Surveillance and monitoring of Toxoplasma in humans, food and animals
2007 EFSA J. 583, 1–64.

75
EYLES DE AND N. COLEMAN
Synergistic effect of sulfadiazine and daraprim against toxoplasmosis in mice
1953 Antibiot. Chemother. 3, 483–490
FAN, C.K., LIAO, C.W., WU, M.S., SU, K.E., HAN, B.C.,
Seroepidemiology of Toxoplasma gondii infection among Chinese aboriginal and Han people
residing in mountainous areas of northern Thailand.
J. Parasitol. 89, 1239–1242.
FAYE, O., LEYE, A., DIENG, Y., RICHARD-LENOBLE, D., DIALLO, S., 1998.
TOXOPLASMOSIS IN
DAKAR.,
Seroepidemiologic sampling of 353 women of reproductive age. Bull.
1998 Soc. Pathol. Exot. 91, 249–250.
FERGUSON DJ, HUTCHISON WM, SIIM JC.
Proceedings: Comparison of the methods of asexual multiplication undergone by Toxoplasma
gondii.
1974 Parasitology.;69(2):xiii.
FERGUSON, D. J. P., A. BIRCH-ANDERSEN, J. C. SIIM, AND W. M. HUTCHISON.
An ultrastructural study on the excystation of the sporozoites of Toxoplasma gondii.
1979 Acta Pathol. Microbiol. Scand. Sect. B 87:277–283.
FERGUSON, D. J. P., AND W. M. HUTCHISON
The host-parasite relationship of Toxoplasma gondii in the brains of chronically infected mice.
Virchows
1987 Arch. A 411:39–43.
FERREIRA IM, VIDAL JE, COSTA-SILVA TA, MEIRA CS, HIRAMOTO RM, PENALVA
DE OLIVEIRA AC, PEREIRA-CHIOCCOLA VL.
toxoplasma gondii: genotyping of strains from brazilian aids patients with cerebral
toxoplasmosis by multilocus pcr-rflp markers.
2008 Exp Parasitol. 118(2):221-7.

76
FEYER R., DUBEY J.P., LINDSAY D.S
Zoonotic protozoa: from land to sea.
2004 TRENDS in Parasitology, 20(11): 531-536.
FOULON W, PINON JM, STRAY-PEDERSEN B, POLLAK A, LAPPALAINEN M,
DECOSTER A, VILLENA I, JENUM PA, HAYDE M, NAESSENS A.
Prenatal diagnosis of congenital toxoplasmosis: a multicenter evaluation of different diagnostic
parameters.
1999Am J Obstet Gynecol. ;181(4):843-7.
FRENKEL JK, J. P. DUBEY, AND N. L. MILLER,
Toxoplasma gondii: faecal forms separated from eggs of the nematode Toxocara cati
1969 Science 164, 432–433
FRENKEL JK, DUBEY JP, MILLER NL
Toxoplasma gondii in cats: fecal stages identified as coccidian oocysts
1970 Science. ;167:893–8
FRENKEL JK, DUBEY JP.
Toxoplasmosis and its prevention in cats and man.
1972 J Infect Dis. 126(6):664-73.
FRENKEL, J. K.
Toxoplasma in and around us.
1973 BioScience 23:343–352.
FRENKEL JK, DUBEY JP.
The taxonomic importance of obligate heteroxeny: distinction of Hammondia hammondi from
Toxoplasma gondii--another opinion.
2000 Parasitol Res. ;86(10):783-6.
DUAN G, TIAN YM, LI BF, YANG JF, LIU ZL, YUAN FZ, ZHU XQ, ZOU FC.,
Seroprevalence of Toxoplasma gondii infection in pet dogs in Kunming, Southwest China.
2012 Parasit Vectors.;5:118. doi: 10.1186/1756-3305-5-118.

77
GROSSKLAUS D, BAUMGARTEN HJ
Die überlebensdauer von Toxoplasmen in Schweinefleisch.
1967 Fleischwirtschaft 47: 1372.
HALL SM, PANDIT A, GOLWILKAR A, WILLIAMS TS
How do Jains get Toxoplasma infection?
1999 Lancet 354: 486-487
HARTLEY WJ, AND S. C. MARSHALL
Toxoplasmosis as a cause of ovine perinatal mortality
1957 N. Z. Vet. J. 5, 119–124 76
HARTLEY WJ, MOYLE GG.
Further observations on the epidemiology of ovine Toxoplasma infection.
1974Aust J Exp Biol Med Sci.;52(4):647-53.
HEJLÍCEK K, LITERÁK I, LHOTÁK M.
Prevalence of antibodies to Toxoplasma gondii in army dogs in the Czech Republic and Slovak
Republic.
1995 Vet Med (Praha). ;40(5):137-40.
HILL DOLORES E. AND DUBEY J.P.
Toxoplasma gondii: transmission,
diagnosis and prevention.
2002 Clin Microbiol Infect. 8(10):634-40.
HILL DE, CHIRUKANDOTH S. AND DUBEY J.P.
Biology and epidemiology of Toxoplasma gondii in man and animals.
2005 Animal Health Research Reviews , 6:41 -61.
HILL DE, CHIRUKANDOTH S, DUBEY JP, LUNNEY JK, GAMBLE HR.
Comparison of detection methods for Toxoplasma gondii in naturally and experimentally
infected swine.
2006 Vet Parasitol.;141(1-2):9-17.

78
HOFFMANN AR, CADIEU J, KIUPEL M, LIM A, BOLIN SR, MANSELL J.
Cutaneous toxoplasmosis in two dogs.
2012 J Vet Diagn Invest.):636-40. doi: 10.1177/1040638712440995.
HOHLFELD P, DAFFOS F, THULLIEZ P, AUFRANT C, COUVREUR J, MACALEESE J,
DESCOMBEY D, FORESTIER F.,
Fetal toxoplasmosis: outcome of pregnancy and infant follow-up after in utero treatment.
1989 J Pediatr.;115(5 Pt 1):765-9
HOSSAIN A, BOLBOL AS, BAKIR TM, BASHANDI AM
A serological survey of the prevalence of Toxoplasma gondii antibodies in dogs and cats in
Saudi Arabia
1986 Trop Geogr Med. ;38(3):244-5.
HOTEZ J.P.
Neglected infection of poverty of the united state of america
2008Plso negt trop.dis.2,256
HOWE DK, SIBLEY LD.
Comparison of the major antigens of Neospora caninum and Toxoplasma gondii.
1999 nt J Parasitol.;29(10):1489-96.
HUNTER CA.,DUPONT CD, CHRISTIAN DA.
Semin Immunopathol.;34(6):793-813. doi: 10.1007/s00281-012-0339-3. Epub 2012 Sep 7.
2012 Immune response and immunopathology during toxoplasmosis.
HUSSEIN, A.H., ALI, A.E., SALEH, M.H., NAGATY, I.M., REZK, A.Y.
2001 Prevalence of Toxoplasma infection in Qualyobia governorate, Egypt. J. Egypt. Soc.
Parasitol.
31, 355–363.
HUTCHISON WM, DUNACHIE JF, SIIM J C, WORK K
The life cycle of Toxoplasma gondii
1969 Brit Med J 4: 806

79
HUTCHISON WM, J.F. DUNACHIE, J.C. SIIM AND K. WORK
Coccidian-like nature of Toxoplasma gondii
1970 Br. Med. J. 1, pp. 142–144
HUTCHISON WM, DUNACHIE JF, WORK K, SIIM JC
The life cycle of the coccidian parasite Toxoplasma gondii in the domestic cat.
1971 Trans R Soc Trop Med Hyg 65: 380-399
INNES EA, AND A. N. VERMEULEN
Vaccination as a control strategy against the coccidial parasites Eimeria Toxoplasma and
Neospora
2006 . Parasitology 133, 145–168
INNES EA, BARTLEY PM, MALEY S, KATZER F, BUXTON D.
Veterinary vaccines against Toxoplasma gondii.
2009 Mem Inst Oswaldo Cruz.;104(2):246-51
INNES EA.
A brief history and overview of Toxoplasma gondiI
2010 Zoonoses Public Health.;57(1):1-7.
JACKSON, M.H. AND HUTCHISON, W.M.
The prevalence and source of Toxoplasma infection in the environment.
1989 Advances in Parasitology. 28: 55-105.
JACOBS L., J. S. REMINGTON, AND M. L. MELTON
The resistance of the encysted form of Toxoplasma gondii
1960 J. Parasitol. 46, 11–21
JACOBS L, MELTON ML .
Toxoplasmosis in chickens.
1966 J Parasitol 52: 1158-1162.

80
JAKUBEK E.B.,PALFI V.,MATTSSON J.G.
Prevalence of antibodies against Toxopasma Gondii and Neospora Caninum in Hungarian red
foxes
2007 Veterinary parasitology 144:39-44
JARA, M., HSU, H.W., EATON, R.B., DEMARIA JR., A
Epidemiology of congenital toxoplasmosis identified by population-based newborn screening in
Massachusetts
2001 Pediatr. Infect. Dis. J. 20, 1132–1135.
JANKŮ, J.
Pathogenesis and pathologic anatomy of the “congenital coloboma” of the macula lutea in an eye
of normal size, with microscopic detection of parasites in the rentna
1923 Veterinary parasitology 144:39-44
JENUM PA, HOLBERG-PETERSEN M, MELBY KK, STRAY-PEDERSEN B.
Diagnosis of congenital Toxoplasma gondii infection by polymerase chain reaction (PCR) on
amniotic fluid samples. The Norwegian experience.
1998 APMIS. 106(7):680-6.
JOHNSON AM.
Speculation on possible life cycles for the clonal lineages in the genus toxoplasma.
1997Parasitol Today. 13(10):393-7.
JONES J.L., LOPEZ A., WILSON M., SCHULKIN J., GIBBS R.
Congenital toxoplasmosis: a review.
2001 Obstet Gynecol Surv, 56: 296-305.
JOINER, K.
Cell entry by Toxoplasma gondii: all paths do not lead to success.
1999 Res. Immunol. 144:34–48.
JONES J.L., LOPEZ A., WILSON M., SCHULKIN J., GIBBS R.
Congenital toxoplasmosis: a review.
2001 Obstet Gynecol Surv, 56: 296-305.

81
KAPPERUD G, JENUM PA, STRAY-PEDERSEN B, MELBY KK, ESKILD A, ENG
J ,
Risk factors for Toxoplasma gondii infection in pregnancy:results of a prospective case-control
study in Norway. Am J
1996 Epidemiol 144: 405-412.
KAUFMAN, H. E., AND E. D. MALONEY.
Multiplication of three strains of Toxoplasma gondii in tissue culture
1962 J. Parasitol. 48:358–361.
CABEZÓN O1, MILLÁN J, GOMIS M, DUBEY JP, FERROGLIO E, ALMERÍA S.,
Kennel dogs as sentinels of Leishmania infantum, Toxoplasma gondii, and Neospora caninum in
Majorca Island, Spain.
2010 Parasitol Res. ;107(6):1505-8. doi: 10.1007/s00436-010-2015-7.
KHAN IA
Toll road for Toxoplasma gondii: the mystery continues.
2007 Trends Parasitol.23(1):1-3.
KOTULA AW, DUBEY JP, SHARAR AK, ANDREWS CD, SHEN SK, LINDSAY
DS .
Effect of freezing on infectivity of Toxoplasma gondii tissue cysts in pork.
1991 J Food Prot 54: 687-690.
KUHLS K, ALAM MZ, CUPOLILLO E, FERREIRA GEM, MAURICIO IL, ODDONE R, ET
AL
Comparative microsatellite typing of new world Leishmania infantum reveals low heterogeneity
among populations and its recent old world origin.
2011 PLoS Negl Trop Dis ; 5(6): e1155. Epud 2011 Jun 7.
KUTICIC V, WIKERHAUSER T
Studies of the effect of various treatments on the viability of Toxoplasma gondii tissue cysts
and oocysts.
1996 In U Gross, Toxoplasma gondii, Springer, Berlin, p. 261-265.

82
LANGONI H, MATTEUCCI G, MEDICI B, CAMOSSI LG, RICHINI-PEREIRA VB, SILVA
RC.
Detection and molecular analysis of Toxoplasma gondii and Neospora caninum from dogs with
neurological disorders.
2012 Rev Soc Bras Med Trop.;45(3):365-
LANGONI H, FORNAZARI F, DA SILVA RC, MONTI ET, VILLA FB.
Prevalence of antibodies against Toxoplasma gondii and Neospora caninum in dogs.
2014 Braz J Microbiol.;44(4):1327-30. eCollection 2013 Dec.
LAPPIN MR, GREENE CE, PRESTWOOD AK, DAWE DL, MARKS A.
Prevalence of Toxoplasma gondii infection in cats in Georgia using enzyme-linked
immunosorbent assays for IgM, IgG, and antigens.
1989 Vet Parasitol.;33(3-4):225-30.
LAPPIN M.R., GASPER P.W., ROSE B.J.
Effect of primary phase feline immunodeficiency virus infection on cats with chronic
toxoplasmosis.
1992Veterinary Immunology and Immunopathology, 35: 121-131.
LAPPIN M.R., GEORGE J.W., PEDERSEN N.C.
Primary and secondary Toxoplasma gondii infections in normal and feline
immunodeficiency virus infected cats.
1996 The Journal of parasitology, 82: 733- 742.
LAPPIN MR, DUBEY JP,GYIMESI ZS,
Application of assays for the diagnosis of toxoplasmosis in a colony of woolly monkeys
(Lagothrix lagotricha).
2006 J Zoo Wildl Med.;37(3):276-80.
LAPPIN MR.
Update on the diagnosis and management of Toxoplasma gondii infection in cats.
2010 Top Companion Anim Med.;25(3):136-41. doi: 10.1053.

83
LEHMANN T., P.L. MARCET, D.H. GRAHAM, E.R. DAHL AND J.P. DUBEY ,
Globalization and the population structure of Toxoplasma gondii
2006 Proc. Natl. Acad. Sci. USA 103 , pp. 11423–11428
LELONG B, RAHELIMINO B, CANDOLFI E, RAVELOJAONA BJ, VILLARD O,
RASAMINDRAKOTROKA AJ, KIEN T.BULL
Prevalence of toxoplasmosis in a population of pregnant women in Antananarivo (Madagascar).
1995,Soc Pathol Exot.;88(1):46-9. French.
LEUCKART, 1879; SLUITER AND SWELLENGREBEL, 1912
Characterization of eimeria species. i. isolation and study of pathogenicity of a pure strain of
eimeria perforans
Z Parasitenkd. 1979 Sep;59(3):227-34.
LEVINE ND
Protozoan parasites of nonhuman primates as zoonotic agents
1970 Lab Anim Care. ;20(2):377-82.
LINDSAY D.S., DUBEY J.P.
Mechanical transmission of Toxoplasma gondii oocysts by dogs.
1997 Veterinary Parasitology 73: 27-33. Parasitol., 28(3): 223-228.
LINDSAY DS.
Toxoplasma gondii research: summary of the seventh international workshops on opportunistic
protists.
J Eukaryot Microbiol. 2001;Suppl:190S.
LINDSAY D.S., DUBEY J.P.
Long term survival of Toxoplasma gondii sporulated oocysts in seawater.
2009 J Parasitol.
LINDSAY DS, M.V. COLLINS, S.M. MITCHELL, C.N. WETCH, A.C. ROSYPAL, G.J.
FLICK, A.M. ZAJAC, A. LINDQUIST AND J.P. DUBEY
Survival of Toxoplasma gondii oocysts in eastern oysters, (Crassostrea virginica)
2004 J. Parasitol. 90, pp. 1054–1057

84
LOPES AP, SARGO R, RODRIGUES M, CARDOSO L.
High seroprevalence of antibodies to Toxoplasma gondii in wild animals from Portugal.
2011 Parasitol Res. ;108(5):1163-9. doi: 10.1007/s00436-010-2158-6. Epub 2010 N
MAIGA, I., KIEMTORE, P., TOUNKARA, A
Prevalence of antitoxoplasma antibodies in patients with acquired immunodeficiency syndrome
and blood donors in Bamako.
2001 Bull. Soc. Pathol. Exot. 94, 268–270.
MANCIANTI F, MECIANI N
Specific serodiagnosis of canine leishmaniasis by indirect immunofluorescence, indirect
hemagglutination, and counterimmunoelectrophoresis.
1988,Am J Vet Res.;49(8):1409-11.
MANCIANTI F, NARDONI S, ARITI G, PARLANTI D, GIULIANI G, PAPINI RA.
Cross-sectional survey of Toxoplasma gondii infection in colony cats from urban Florence
(Italy).
2010 J Feline Med Surg. 12(4):351-4. doi: 10.1016/j.jfms.2009.09.001. Epub 2009 Oct 6.
MANCIANTI F, NARDONI S, MUGNAINI L, ZAMBERNARDI L, GUERRINI
A, GAZZOLA V, PAPINI RA.
A retrospective molecular study of select intestinal protozoa in healthy pet cats from Italy.
J Feline Med Surg. 2014 May 2. [Epub ahead of print]
MASALA G, PORCU R, DAGA C, DENTI S, CANU G, PATTA C, TOLA S
Detection of pathogens in ovine and caprine abortion samples from Sardinia, Italy, by PCR.
2007 J Vet Diagn Invest.;19(1):96-8.
McINNES, L. M. et al.
In vitro isolation and characterization of the first canine Neospora caninum isolate in Australia.
2006 Veterinary Parasitology, v. 137, n. 3-4, p. 355-363, 2006. PMid:16487658

85
MELHORN, H., AND J. K. FRENKEL.
Ultrastructural comparison of cysts and zoites of Toxoplasma gondii, Sarcocystsis muris, and
Hammondia hammondi in skeletal muscle of mice.
1980 J. Parasitol. 66:59–67
MELOT T, GRUEL N, DOUBEIKOVSKI A, SEVENET N, TEILLAUD JL, DELATTRE O.
Production and characterization of mouse monoclonal antibodies to wild-type and oncogenic
FLI-1 proteins.
1997 Hybridoma 16(5):457-64
MESSIER, V., LEVESQUE, B., PROULX, J.F., ROCHETTE, L., LIBMAN, M.D., WARD,
B.J., SERHIR, B., COUILLARD, M., OGDEN, N.H., DEWAILLY, E., HUBERT, B., DERY,
S., BARTHE, C., MURPHY, D., DIXON, B.,
Seroprevalence of Toxoplasma gondii among Nunavik Inuit
(Canada).
2009 Zoonoses Public Health 56, 188–197. MILLER MA, I. A. GARDNER, C. KREUDER, D. M. PARADIES, K. R. WORCESTER, D.
A. JESSUP, E. DODD, M. D. HARRIS, J. A. AMES, A. E. PACKHAM, AND P. A. CONRAD
Coastal freshwater runoff is a risk factor for Toxoplasma gondii infection of southern sea otters
(Enhydra lutris mereis).
2002 Int. J. Parasitol. 32, 997–1006
MILLER MA, M. E. GRIGG, C. KREUDER, E. R. JAMES, A. C. MELLI, P. R. CROSBIE, D.
A. JESSUP, J. C. BOOTHROYD, D. BROWNSTEIN, AND P. A. CONRAD
An unusual genotype of Toxoplasma gondii is common in California sea otters (Enhydra lutris
nereis) and is a cause of mortality
2004 Int. J. Parasitol. 34, 275–284 81

86
MILLER M.A., MILLER W.A., CONRAD P.A., JAMES E.R., MELLI A.C.,
LEUTENEGGER C.M., DABRITZ H.A., PACKHAM A.E., PARADIES D., HARRIS M.,
AMES J., JESSUP D.A., WORCESTER K., GRIGG M.E.
Type X Toxoplasma gondii in a wild mussel and terrestrial carnivores from coastal
California: new linkages between terrestrial mammals, runoff and
toxoplasmosis of sea otters.
2008 Int J Parasitol., 38(11): 1319-28.
MINERVINO AH1, CASSINELLI AB, DE LIMA JT, SOARES HS, MALHEIROS
AF, MARCILI A, GENNARI SM.,
Prevalence of anti- Neospora caninum and anti- Toxoplasma gondii antibodies in dogs from two
different indigenous communities in the Brazilian Amazon Region.
2012 J Parasitol. ;98(6):1276-8. doi: 10.1645/GE-3151.1. Epub 2012 May 2.
MONTOYA E LIESENFELD
toxoplasmosis ,
200 Lancet 3631965-1976)
MORISAKI, J. H., J. E. HEUSER, AND L. D. SIBLEY.
Invasion of Toxoplasma gondii occurs by active penetration of the host cell.
1995 J. Cell Sci. 108:2457–2464.
MORRISSETTE NS1, MURRAY JM, ROOS DS.
Subpellicular microtubules associate with an intramembranous particle lattice in the protozoan
parasite Toxoplasma gondii
1997 J Cell Sci.;110 ( Pt 1):35-42.
MUNDAY BL
Serological evidence of Toxoplasma infection in isolated groups of sheep
1972 Res. Vet. Sci. 13 (1972), pp. 100–102
NABIAS, R., NGOUAMIZOKOU, A., MIGOT-NABIAS, F., MBOU-MOUTSIMBI, R.A.,
LANSOUD-SOUKATE, J
Serological investigation of toxoplasmosis in patients of the M.I.P. center of Franceville
(Gabon). 1998 Bull. Soc. Pathol. Exot. 91, 318–320.

87
NA YANG, MINGYANG MU, HONGKUI LI*, JIANMIN HU, WEIFAN GAO, SHUHUA
YANG, AND JIANBIN HE†
Seroprevalence of Toxoplasma gondii Infection in Pet Dogs in Shenyang,
Northeastern China
2013 J. Parasitol., 99pp. 176–177
NGUYEN THUY THI-DIEU , SE-EUN CHOE1, JAE-WON BYUN1, HONG-BUM KOH3,
Hee-Soo Lee and Seung-Won Kang,
Seroprevalence of Toxoplasma gondii and Neospora caninum in dogs from Korea
2012 Acta Parasitologica, 57(1), 7–12; ISSN 1230-2821
NICHOLS, B. A., AND G. R. O’CONNOR.
Penetration of mouse peritoneal macrophages by the protozoon
1981 Toxoplasma gondii. Lab. Invest. 44:324–335.
NICHOLS, B. A., M. L. CHIAPPINO, AND G. R. O’CONNOR.
Secretion from the rhoptries of Toxoplasma gondii during host-cell invasion.
1983 J. Ultrastruct. Res. 83:85–98.
O’CONNELL, E., WILKINS, M. F. AND TE PUNGA, W. A.
Toxoplasmosis in sheep. II. The ability of a live vaccine to prevent lamb losses after an
intravenous challenge with Toxoplasma gondii.
1988 New Zealand Veterinary Journal 36, 1–4.
OWEN, M.R. AND TREES, A.J.
Vertical transmission of Toxoplasma gondii from chronically infected house ( Mus musculus)
and field (Apodemus sylvaticus) mice determined by polymerase chain reaction.
1998 Parasitology. 116: 299-304
PAPPAS GEORGIOS .,ROUSSOUS NIKOS.,FALAGAS MATTEW.E.
Toxoplasmosis snapshots: Global status of Toxoplasma condii seroprevalence and implications
for pregnancy and congenital toxoplasmosis.
2009 International journal parasitology: 1385-1394)

88
PAUL M.,
Potencjalne źródła zarażenia Toxoplasma gondii w przypadkach badanych w krótkim czasie po
zarażeniu. Przegl
1998 Epidemiol 52: 447-454
PAULAN SC, SILVA HR, LIMA EACF, FLORES EF, TACHIBANA VM, KANDA C, ET AL
.Distribuição espacial da leishmaniose visceral canina em Ilha Solteira, São Paulo, Brasil.
2012 Eng Agríc; 32(4): 765-774.
PAVESIO, C. E. N., M. L. CHIAPPINO, P. Y. SETZER, AND B. A. NICHOLS.
Toxoplasma gondii: differentiation and death of bradyzoites.
1992 Parasitol. Res.78:1–9.
PENA HF, GENNARI SM, DUBEY JP, SU C.
Population structure and mouse-virulence of Toxoplasma gondii in Brazil.
2008 Int J Parasitol.;38(5):561-9.
PIERGILI FIORETTI D.,
Problems and limitations of conventional and innovative methods for the diagnosis of
Toxoplasmosis in humans and animals
2004 Parassitologia 46:177
PLUGGE NF, FERREIRA FM, RICHARTZ RR, DE SIQUEIRA A, DITTRICH RL.,
Occurrence of antibodies against Neospora caninum and/or Toxoplasma gondii in dogs with
neurological signs.
2011 Rev Bras Parasitol Vet. ;20(3):202-6.
PORCHET-HENNERE, E., E. VIVIER, AND G. TORPIER.
Origine des membranes de la paroi chez Toxoplasma.
1985 Ann. Parasitol. Hum. Comp. 60:101–
110.

89
POWELL, C.C., BREWER, M. AND LAPPIN, M.R.
Detection of Toxoplasma gondii in the milk of experimentally infected lactating
cats.
2001 Veterinary Parasitology. 102: 29-33.Published online 2006 July 18. doi:
10.1073/pnas.0601438103
POWELL CC, LAPPIN MR
Clinical ocular toxoplasmosis in neonatal kittens
2001b Vet Ophthalmol 4:87-92
PUTIGNANI L, MENICHELLA D.
Global distribution, public health and clinical impact of the protozoan pathogen cryptosporidium.
2010 Interdiscip Perspect Infect Dis.. pii: 753512. doi: 10.1155/2010/753512. Epub 2010 Jul 14
REMINGTON, J.S., JACOBS, L. AND MELTON, M.L.
Congenital transmission of toxoplasmosis from mother animals with acute and chronic
infections.
1961 Journal of Infectious Diseases. 108: 163-173.
REMINGTON J BOYER KM, HOLFELS E, ROIZEN N, SWISHER C, MACK D, WITHERS
S, MEIER P, MCLEOD R; TOXOPLASMOSIS STUDY GROUP.
Risk factors for Toxoplasma gondii infection in mothers of infants with congenital
toxoplasmosis: Implications for prenatal management and screening.
2005 Am J Obstet Gynecol.;192(2):564-71.
RIEMANN HP, MEYER ME, THEIS JH, KELSO G, BEHYMER DE
Toxoplasmosis in an infant fed unpasteurized goat milk.
1975 J Pediatr 573-576.
RINALDI L.
Toxoplasma.Antropozoonosi parassitarie.
2010 Progetto “Campus Virtuale”dell’Universita degli Studi di Napoli Federico !!

90
J. PAUL ROBINSON PHD, JENNIFER STURGIS BS AND GEORGE L. KUMAR PHD
Immunohistochemical Staining Methods
http://www.dako.com/08002_03aug09_ihc_guidebook_5th_edition_chapter_10.pdf
ROGHMANN MC, FAULKNER CT, LEFKOWITZ A, PATTON S, ZIMMERMAN J,
MORRIS JG JR
decreased seroprevalence for toxoplasma gondii in seventh day adventists in maryland.
1999 Am J Trop Med Hyg. ;60(5):790-2.
ROSYPAL AC,HILL R,LEWIS S,BRAXTON K,ZAJAC AM,LINDSAY DS
Toxoplasma gondii and Trymanosoma cruzi antibodies in dogs from Virginia
2010 Zoonoses Public Health 57(7-8):e 76-80
ROSS R. FURTHER
notes on Leishman’s bodies.
1903 Br Med J 1903; 2(2239): 1401
ROTHE, J., MCDONALD, P. AND JOHNSON, A.M
Detection of Toxoplasma cysts and oocysts in an urban environment in a
developed country.
1985 Pathology. 17: 497-499.
SABIN AB AND J. WARREN
Therapeutic effects of the sulfonamides on infection by an intracellular protozoan (Toxoplasma).
1941 J. Bacteriol. 41, 80–86
SACKS JJ, ROBERTO RR, BROOKS NF.
Toxoplasmosis infection associated with raw goat's milk.
1982 JAMA. 248(14):1728-32
SAFFER, L. D., O. MERCAREAU-PUIJALON, J.-F. DUBREMATZ, AND J. D.
SCHWARTZMAN.
Localization of a Toxoplasma gondii rhoptry protein by immunoelectron
microscopy during and after host cell penetration
1992. J. Protozool. 39:526–530.

91
SALANT H, SPIRA DT, HAMBURGER J.
A comparative analysis of coprologic diagnostic methods for detection of Toxoplama gondii in
cats.
2010 Am J Trop Med Hyg. 82(5):865-70. doi: 10.4269/ajtmh.2010.09-0635.
SATO K, IWAMOTO I, YOSHIKI K.
Experimental toxoplasmosis in pregnant cats.
1993 J Vet Med Sci. ;55(6):1005-9.
SHEFFIELD H G, AND M. L. MELTON
Toxoplasma gondii: transmission through faeces in absence of Toxocara cati eggs
1969 Science 164, 431–432
SHUHAIBER S.,KORENG.,BORKIVIC R.,EINARSONT.R.,SOLDIN O.P.,EINARSON A.,
Seroprevalence of toxoplasma gondii infection among veterinari staff in Ontario Canada(2002),
2008 implication of teratogenic riskBMC infect dis 3,8
SIBLEY LD, LEBLANC AJ, PFEFFERKORN ER, BOOTHROYD JC.
Generation of a restriction fragment length polymorphism linkage map for Toxoplasma gondii.
1992 Genetics. 132(4):1003-15.
SIBLEY, L. D.
Invasion of vertebrate cells by Toxoplasma gondii. Trends
1995 Cell Biol. 5:129–132
SILVA, S. R. L., S. S. MEIRELLES, AND W. DE SOUZA
Mechanism of entry of Toxoplasma gondii into vertebrate cells.
1982 J. Submicrosc. Cytol. 14:471– 482.
SIMS, T. A., J. HAY, AND I. C. TALBOT.
Host-parasite relationship in the brains of mice with congenital toxoplasmosis.
1988 J. Pathol. 156:255–261.

92
SKINNER LJ, TIMPERLEY AC, WIGHTMAN D, CHATTERTON JM, HO-YEN DO
Simultaneous diagnosis of toxoplasmosis in goats and goatowner's family.
Genetics. 132(4):1003-15.
SPEER, C. A., S. CLARKE, AND J. P. DUBEY.
Ultrastructure of the oocysts, sporocysts and sporozoites of the VEG strain of Toxoplasma
gondii. J.Parasitol., in press.
SPEER, C. A., J. P. DUBEY, J. A. BLIXT, AND K. PROKOP.
Time lapse video microscopy and ultrastructure of penetrating sporozoites, types 1 and 2
parasitophorous vacuoles, and the transformation of sporozites to tachyzoites of the VEG strain
of Toxoplasma gondii.
1997 J. Parasitol. 83:565–574.
SULZER AJ, STROBEL PL, SPRINGER EL, ROTH IL, CALLAWAY CS.
A comparative electron microscopic study of the morphology of Toxoplasma gondii by freeze-
etch replication and thin sectioning technic.
1974 J Protozool.;21(5):710-4.
SUNDAR N., R.A. COLE, N.J. THOMAS, D. MAJUMDAR, J.P. DUBEY AND C. SU
Genetic diversity among sea otter isolates of Toxoplama gondii
2003 Vet. Parasitol. 151 , pp. 125–132
SVOBODOVÁ V, LITERÁK I.
Prevalence of IgM and IgG antibodies to Toxoplasma gondii in blood donors in the Czech
Republic.
1998 Eur J Epidemiol.;14(8):803-5.
SZENASI, Z., OZSVAR, Z., NAGY, E., JESZENSZKY, M., SZABO, J., GELLEN, J., VEGH,
M.,VERHOFSTEDE, C.,
Prevention of congenital toxoplasmosis in Szeged,
1997 Hungary. Int. J. Epidemiol. 26, 428–435

93
TABBARA, K.S., SALEH, F.
Serodiagnosis of toxoplasmosis in Bahrain.
2005 Saudi Med. J. 26, 1383–1387.
TENTER AM, HECKEROTH AR, WEISS LM
Toxoplasma gondii: from animals to humans.
2000 Int J Parasitol 30
TIZARD, I.R., FISH, N.A. AND QUINN, J.P.
Some observations on the epidemiology of toxoplasmosis in Canada.
1976 Journal of Hygiene. 77: 11-21.
TSAI YJ, CHUNG WC, FEI AC, HONG CL, TSAI YY, PENG S, WU YL.
Prevalence of Toxoplasma gondii antibodies in stray dogs in Taipei, Taiwan.
2008 J Parasitol.;94(6):1437. doi: 10.1645
UNEKE, C.J., DUHLINSKA, D.D., NJOKU, M.O., NGWU, B.A
Seroprevalence of acquired toxoplasmosis in HIV-infected and apparently healthy individuals in
Jos, Nigeria.
2005 Parassitologia 47, 233–236.
URQUHART M., ARMOUR J., DUNCAN J.L., DUNN A.M., JENNINGS F.W.
(1998). Parassitologia Veterinaria, UTET, Torino
VELASCO-CASTREJÓN O, SALVATIERRA-IZABA B, VALDESPINO JL, SEDANO-LARA
AM, GALINDO-VIRGEN S, MAGOS C, LLAUSÁS A, TAPIA-CONYER R, GUTIÉRREZ G,
SEPÚLVEDA J
Seroepidemiology of toxoplasmosis in Mexico
1992Salud Publica Mex. 34(2):222-9.
VIVI ER E, PETITPREZ A.
Serial sections and three-dimensional reconstitution of the vacuolar system of the hematozoan
Anthemosoma garnhami.
1972 J Ultrastruct Res.;41(3):219-37. French

94
WALLACE GD
Serologic and epidemiologic observations on toxoplasmosis on three Pacific Atolls
1969 Am. J. Epidemiol. 90 , pp. 103–111.
WALLACE GD.
Isolation of Toxoplasma gondii from the feces of naturally infected cats.
1971 J Infect Dis.;124(2):227-8.
WALLACE GD
Experimental transmission of Toxoplasma gondii by cockroaches
1972 J Infect Dis.;126(5):545-7.
WALLACE GD.
Intermediate and transport hosts in the natural history of Toxoplasma gondii.
1973 Am J Trop Med Hyg.;22(4):456-64.
WAINWRIGHT KE, M. A. MILLER, B. C. BARR, I. A. GARDNER, A. C. MELLI, T.
ESSERT, A. E. PACKHAM, T. TRUONG, M. LAGUNAS-SOLAR, AND P. A. CONRAD,
Chemical inactivation of Toxoplasma gondii oocysts in water
2006 J. Parasitol. 93, 925–931
WANG M1, WANG YH, YE Q, MENG P, YIN H, ZHANG DL.
Serological survey of Toxoplasma gondii in Tibetan mastiffs (Canis lupus familiaris) and yaks
(Bos grunniens) in Qinghai, China.
2012 Parasit Vectors.; 5:35. doi: 10.1186/1756-3305-5-35.
WANHA K1, EDELHOFER R, GABLER-EDUARDO C, PROSL H
Prevalence of antibodies against Neospora caninum and Toxoplasma gondii in dogs and foxes in
Austria
2005 Vet Parasitol. ;128(3-4):189-93. Epub 2005 Jan 24.

95
WEBER K, ET AL.
Specific visualization of tubulin-containing structures in
tissue culture cells by immunofluorescence. Cytoplasmic microtubules,
vinblastine-induced paracrystals, and mitotic figures.
1975 E xp Cell Res ;9 5,111-20
WEILAND G. AND D. KÜHN
Experimentelle Toxoplasma-Infektionen bei der Katze. II. Entwicklungsstadien des Parasiten im
Darm
1970 Berl. Münch. Tierärztl. Wochenschr. 83, pp. 128–132
WERK, R.
How does Toxoplasma gondii enter host cells.
1985Rev. Infect. Dis. 7:449–457.
YILDIZ K, YASA DURU S, YAGCI BB, BABUR C, OCAL N, GURCAN S, KARACA S.
Seroprevalence of Neospora caninum and coexistence with Toxoplasma gondii in dogs.
2009 Turkiye Parazitol Derg. (2):116-9.
YONGMEI LI, QUAN LIU`, SHUMIN LI, FENG WEI, HONGTAO JIN, AND MINGHUI
YANG,
Seroprevalence of Toxoplasma gondii Infection in Dogs in Jiangsu ,Eastern China
2012 J.Parassitol.,98 878-879

96
Grazie……
…alla Professoressa Mancianti, per l’attenzione, la dedizione, la pazienza e il
tempo dedicatomi; per avermi aiutato e seguito anche ad agosto, nonostante fosse
in ferie.
Una delle migliori professioniste e persone che ho incontrato in questi anni
universitari.
…alla Dottoressa Nardoni, sempre presente, disponibile ed attenta ad ogni mia
necessità; grazie per la pazienza e la qualità del tempo che m’ha dedicato.
…al Dottor Papini, per l’aiuto, l’attenzione e la professionalità che ha messo a mia
disposizione.
…a Linda e Guido, perché non solo mi hanno seguito con serietà e professionalità,
ma anche guidato, consolato e dato coraggio, come dei veri amici.
…a mamma e babbo, perché hanno sempre creduto in me, più di quanto abbia fatto
io.
…a Alessio, per il suo immenso amore.
…a Barbara e Federica, perchè mi hanno insegnato che nella vita, se vogliamo,
tutto è possibile.
…a nonna, per avermi coccolato, come solo lei sa fare.
…a Enrico, per la sua dolcezza, il suo amore e l’infinita pazienza…
…a Haidi, insostituibile compagna di vita e di tante giornate sui libri…
…a Camilla e a tutti i miei animaletti, per avermi fatto capire quale fosse la mia
strada.
…a Francesca ,una grande amica ed una grande collega….la migliore che potessi
avere….

97
…a Serena, per la sua insostituibile amicizia, pazienza e per tutto il tempo che mi
ha dedicato.
…a Federica ed Erica, le migliori amiche che si possano avere.
…a Alessandro, Federico, Patrizio e a tutti gli altri amici sinceri….per avermi
sopportato e ascoltato, anche nei miei momenti peggiori.
…a me…perché ce l ho fatta!


















