
in the haemolymph of the protochordate Botrylloides leachii
10
Immunology 1981 42 241 Identification of two carbohydrate-specific erythrocyte agglutinins in the haemolymph of the protochordate Botrylloides leachii S. F. SCHLUTER, P. L. EY, DEIRDRE R. KEOUGH & C. R. JENKIN Department of Microbiology and Immunology and Department of Zoology, The University of Adelaide, Adelaide, South Australia Acceptedfor publication 7 September 1980 Summary. The haemolymph of the colonial ascidian Botrylloides leachii contains two haemagglutinins which have been termed HA- I and HA-2 respectively. The HA- I agglutinin is specific for guinea-pig erythro- cytes, requires Ca2+ ions for its activity and has an apparent mol.wt. of about 200,000. It can be adsorbed by guinea-pig but not human, sheep, mouse or pigeon erythrocytes. The agglutination of guinea-pig cells is reversibly inhibited by various mono- and oligosac- charides, lactose and D-galactose being among the most effective of those tested. The HA-2 agglutinin is smaller (mol.wt. approximately 63,000) and agglu- tinates a variety of erythrocyte types, including those from the human, guinea-pig, mouse, sheep, pigeon and chicken. The agglutination of all these cells is inhibited by lactose. Data from sugar-inhibition and adsorption experiments indicate that the HA-2 agglu- tinins are homogeneous with respect to binding site specificity. Divalent cations are not required for activity. INTRODUCTION Sufficient evidence has now accumulated to indicate that the phagocytic cells of many invertebrates recog- Correspondence: Dr P. L. Ey, Department of Micro- biology and Immunology, University of Adelaide, GPO Box 498, Adelaide, South Australia 5001. 0019-2805/81/0200-0241S02.00 ©) 1981 Blackwell Scientific Publications nize foreign materials by a system analogous to that observed in the vertebrates, i.e. by the interaction of specific humoral proteins (immunoglobulins, in the vertebrates) with the foreign material. However, the factors present in the haemolymph of invertebrates which are considered responsible for recognition of 'non-self' are structurally unrelated to the immunoglo- bulins of vertebrates (reviewed by Jenkin, 1976). During an investigation into the mechanisms re- sponsible for recognition of foreign material by Botrylloides leachii (Urochordata, Ascidiacea), amoe- bocytes from these animals have been observed to bind and subsequently ingest mammalian erythrocytes both in vivo and in vitro. The binding seems to be mediated by factors present on the surface of the amoebocytes (Keough, Ey, Jenkin & Schluter, unpub- lished observations). The haemolymph of this particu- lar animal, like that of other invertebrates and proto- chordates (Fuke & Sugai, 1972; Gold & Balding, 1975; Parrinello & Patricolo, 1975; Anderson & Good, 1975), contains erythrocyte agglutinins which, in view of biological data from other invertebrates (Jenkin, 1976; Renwrantz & Mohr, 1978), may function as opsonins. With one exception (Renwrantz & Mohr, 1978), we are not aware of any study in which a purified invertebrate agglutinin has been used to assess its importance in the recognition and clearance of foreign particles in the homologous organism. The present paper describes the resolution of two hae- magglutinins from the haemolymph of B. leachii as a preliminary step in achieving this aim. 241
Transcript of in the haemolymph of the protochordate Botrylloides leachii

Immunology 1981 42 241
Identification of two carbohydrate-specific erythrocyte agglutininsin the haemolymph of the protochordate Botrylloides leachii
S. F. SCHLUTER, P. L. EY, DEIRDRE R. KEOUGH & C. R. JENKIN Department ofMicrobiology andImmunology and Department ofZoology, The University of Adelaide, Adelaide, South Australia
Acceptedfor publication 7 September 1980
Summary. The haemolymph of the colonial ascidianBotrylloides leachii contains two haemagglutininswhich have been termed HA- I and HA-2 respectively.The HA- I agglutinin is specific for guinea-pig erythro-cytes, requires Ca2+ ions for its activity and has anapparent mol.wt. of about 200,000. It can be adsorbedby guinea-pig but not human, sheep, mouse or pigeonerythrocytes. The agglutination of guinea-pig cells isreversibly inhibited by various mono- and oligosac-charides, lactose and D-galactose being among themost effective of those tested. The HA-2 agglutinin issmaller (mol.wt. approximately 63,000) and agglu-tinates a variety of erythrocyte types, including thosefrom the human, guinea-pig, mouse, sheep, pigeonand chicken. The agglutination of all these cells isinhibited by lactose. Data from sugar-inhibition andadsorption experiments indicate that the HA-2 agglu-tinins are homogeneous with respect to binding sitespecificity. Divalent cations are not required foractivity.
INTRODUCTION
Sufficient evidence has now accumulated to indicatethat the phagocytic cells of many invertebrates recog-
Correspondence: Dr P. L. Ey, Department of Micro-biology and Immunology, University of Adelaide, GPO Box498, Adelaide, South Australia 5001.0019-2805/81/0200-0241S02.00©) 1981 Blackwell Scientific Publications
nize foreign materials by a system analogous to thatobserved in the vertebrates, i.e. by the interaction ofspecific humoral proteins (immunoglobulins, in thevertebrates) with the foreign material. However, thefactors present in the haemolymph of invertebrateswhich are considered responsible for recognition of'non-self' are structurally unrelated to the immunoglo-bulins of vertebrates (reviewed by Jenkin, 1976).
During an investigation into the mechanisms re-sponsible for recognition of foreign material byBotrylloides leachii (Urochordata, Ascidiacea), amoe-bocytes from these animals have been observed to bindand subsequently ingest mammalian erythrocytesboth in vivo and in vitro. The binding seems to bemediated by factors present on the surface of theamoebocytes (Keough, Ey, Jenkin & Schluter, unpub-lished observations). The haemolymph of this particu-lar animal, like that of other invertebrates and proto-chordates (Fuke & Sugai, 1972; Gold & Balding, 1975;Parrinello & Patricolo, 1975; Anderson & Good,1975), contains erythrocyte agglutinins which, in viewof biological data from other invertebrates (Jenkin,1976; Renwrantz & Mohr, 1978), may function asopsonins. With one exception (Renwrantz & Mohr,1978), we are not aware of any study in which apurified invertebrate agglutinin has been used to assessits importance in the recognition and clearance offoreign particles in the homologous organism. Thepresent paper describes the resolution of two hae-magglutinins from the haemolymph of B. leachii as apreliminary step in achieving this aim.
241

S. F. Schluter et al.
MATERIAiLS AND METHODS
Collection ofhaemolymphColonies of B. leachii were collected from jetty piles atRapid Bay, South Australia using Scuba and main-tained at 12-14° in holding tanks which were flushedcontinuously with aerated sea water. After removingsurface water with a paper towel, incisions were madein the tunic matrix beneath the zooids and the haemo-lymph was collected on ice. As much as 10- 15 ml couldbe obtained from a colony of approximately 15 cmdiameter. After removing the cells by centrifugation(900g, 5 min), the haemolymph, which does not clot,was pooled and stored at - 20°. When required, it wasthawed and clarified by centrifuging at 30,000g for 30min (4'). The resulting fluid had an absorbance at 280nm of approximately 1 2 and a protein content, usingthe Folin-phenol reagent (Schacterle & Pollack, 1973)with bovine serum albumin as a standard, of 0 4-0 5mg ml-'.
ErythrocytesSheep and human blood was drawn aseptically intoAlsever's solution and stored at 4° for no longer than 3weeks. Blood collected by cardiac puncture fromguinea-pigs, rats and pigeons was also mixed withAlsever's solution and discarded after several days.The erythrocytes were centrifuged (900g, 5 min),washed three times in 0 15 M NaCI and stored at 4° as a10% v/v stock suspension for no longer than 3 days.Blood collected from (BALB/c x C57BI)FI mice wasmixed with 0 013 M trisodium citrate-0 168 M NaCl.The erythrocytes were washed in 0168 M NaCI andused immediately.
SugarsExcept where indicated, sugars were purchased fromBDH Chemicals Ltd. Laboratory reagent grade:D-arabinose, L-arabinose, L-fructose, N-acetyl-D-galactosamine (B grade, Calbiochem.), maltose,D-xylose. Analytical grade: D-fucose, L-fucose (Agrade, Calbiochem.), D-galactose, D-glucose, lactose,mannose, melibiose (A grade, Calbiochem.), sta-chyose and sucrose. The D-fucose and stachyose(Sigma Chem. Co.) were kindly donated by Dr J.Redmond, Chemistry Department, MacquarieUniversity, New South Wales.
Haemagglutination assaysHaemagglutinating activity was usually titrated bymixing two-fold serial dilutions of the sample with an
equal volume of a 0 5% erythrocyte suspension in thewells of a microtitre tray. The diluent was 10 mmphosphate-buffered saline, pH 7-4 (PBS). Agglutin-ation was evaluated after the cells had settled at roomtemperature. Titres are expressed as the greatest dilu-tion of the sample which caused complete agglutin-ation.
For titrating agglutinating activity in adsorptionexperiments (e.g. Table 6), greater endpoint accuracywas required. Thus, after determining the approxi-mate agglutination titre (as above), each sample wasretitrated using fractional dilutions around the end-point dilution. The trays were then photographed andthe cell pellet diameters measured from enlargedprints. A plot of (diameter)2 v. dilution allowed thedilution corresponding to 50% agglutination to beaccurately determined. This was chosen as the end-point.
Haemagglainahion: inhibition by sugarsTo assess the capacity of a given sugar to inhibithaemagglutination, the agglutinin sample was titratedin the presence of various concentrations of the sugar(Table 2). The latter was usually dissolved in water togive a solution iso-osmotic with saline. This wasdiluted in PBS, when necessary, for addition to thewells. The concentration of sugar which halved theagglutinating titre was thus determined (Tables 2& 3).
Sephadex G-200 chromatographyA column of Sephadex G-200 (Pharmacia; 2 5 x 98 5cm) was equilibrated at 40 with PBS condtaining 50 mMlactose, 100 mm galactose and 8 iM-NaN3 (eluantbuffer). The column was calibrated withr blieAdextran2000 (Pharmacia), 251-labelled moust IgqG (Ey,Prowse & Jenkin, 1978), Na3251 (IMS.3Og,ie Radio-chemical Centre, Amersham), ferritin (Ty i, Sigma),bovine serum albumin (Calbiochem.), catalase (Nutri-tional Biochemicals Corp.), and horse skeletal musclemyoglobin (Calbiochem.). Various combinations ofthese markers were applied to the column in 5 ml ofbuffer and the column was washed at a flow rate of 16ml hr-', 4 9 ml fractions being collected. The elutionpositions of the various markers, measured by absor-bance (280 nm) or radioactivity (Packard Autogammacounter), were used to calculate values of Ka,[ =(Ve- Vo)/(Vt - Vo), where Ve= elution volume ofsolute, Vo= void volume and V, = gel bed volume](Laurent & Killander, 1964). Blue dextran 2000 and1251- were used to determine Vo and V,, respectively.With the exception of ferritin (Siegel & Monty, 1966),
242
Haemagglutinins of Botrylloides leachii
the Stoke's radii of the marker proteins were calcu-lated from their diffusion constants (Sober, 1970)using the equation a= k T/6ir M D where a is theStoke's radius, k the Boltzman constant, T the abso-lute temperature, M the viscosity of the medium and Dthe diffusion coefficient.
RESULTS
Agglutination of erythrocytesThe data presented in Table 1 show that B. leachiihaemolymph agglutinated erythrocytes from variousvertebrate species. The titre was similar (1/16-1/64)for most of the erythrocytes tested, but exceptionallyhigh for those from the guinea-pig and pigeon. Nodifference in titre was observed when human type A, Bor 0 erythrocytes were used. Little variation was
observed between different batches of haemolymph.
Table 1. Haemagglutinating activity of B. leachiihaemolymph
Source of erythrocytes Agglutination titre-
Guinea-pig 2000-4000*Chicken 8-16Pigeon 1600Mouse 64Rat 64Rabbit 32Sheep 64-128Human 64
* Usually enhanced two to four-fold if CaC12 wasincluded in the diluent.
Inhibition by sugarsMany agglutinins isolated from both plants and inver-tebrates have binding specificities for simple sugars(Gold & Balding, 1975). To determine whether the B.leachii haemagglutinins were also sugar-specific, anumber of sugars were tested for their capacity toinhibit the agglutination of erythrocytes from severalvertebrate species. Such data are useful not only forthe purification of an agglutinin by affinity chromato-graphy, but also in assessing whether or not the agglu-tinins in a sample are homogeneous. The results of thesugar inhibition tests (Tables 2 and 3) suggested thatmore than one haemagglutinin occurred in B. keachiihaemolymph. For example, whereas the agglutinationof guinea-pig erythrocytes was inhibited by a number
Table 2. The effect of several saccharides on the agglutination ofguinea-pig erythrocytes by B. leachii haemolymph
Inhibitor concentration (mM)
Inhibitor 0 0 5 1 2 4 8 16
Agglutination titre- I
Lactose 3200 1600 800 400 200 < 200 < 200D-Galactose 3200 1600 1600 400 200 < 200 < 200D-Glucose 3200 3200 3200 3200 3200 1600 800D-Arabinose 3200 3200 3200 3200 3200 3200 3200
Replicate titrations (serial two-fold dilutions; 25 p1) of a 1: 100dilution of haemolymph were made across the rows of a micro-titre tray. An equal volume of PBS was then added to each wellof the first two rows (duplicate controls), whilst the next tworows received PBS containing a fixed amount of the test sugar.This procedure was repeated using a range of sugar concentra-tions. Guinea-pig erythrocytes (50 p1, 055%) were then addedand agglutination was evaluated after the cells had settled.
Table 3. The capacity of sugars to inhibit the agglutinationof erythrocytes by haemolymph*
Concentration of sugar (mM)required to halve theagglutination titre for
Guinea-pig Sheep PigeonSugar erythrocytes erythrocytes erythrocytes
Melibioset 0-5 > 67 4Lactose: 0 5 8 0.1D-Fucose I > 32 > 32D-Galactose I > 67 2Stachyose§ 2 > 67 4N-Acetyl-D-
galactosamine 2 > 4 NDL-Arabinose 2 ND > 67D-Glucose 8 > 67 > 67D-Xylose 8 > 67 > 67L-Fucose 16 > 67 > 67D-Arabinose 32 > 67 > 67Maltose 32 > 67 > 67Mannose > 67 > 67 > 67Sucrose > 67 > 67 > 67D-Fructose > 67 > 67 > 67
* Haemolymph was dialysed against 100 vol. of PBS + 8mM NaN3 (40, overnight). Experimental details are de-scribed in the Materials and Methods and Table 2.
t D-Gal-oa I 6 D-Glu.: D-Gal-# I 4- D-Glu.§ D-Gal-al -+6-D-Gal-ae-I-16-D-Glu-fIl - 2-D-Fru.ND = not done.
243
S. F. Schluter et al.
of sugars (melibiose lactose > D-galactose _ D-fucose) and that ofpigeon erythrocytes by lactose > D-galactose> melibiose stachyose, the only sugar toinhibit the agglutination of sheep erythrocytes waslactose (Table 3). Furthermore, although lactose wasin each case the most effective inhibitor, the concentra-tion required to halve the titre was different for eachtype of cell.
Fractionation of haemolymphAs a preliminary step in the characterization of theagglutinins, a sample of haemolymph was chromato-graphed on Sephadex G-200 using PBS as the eluant.The passage of the agglutinating activity for bothsheep and guinea-pig erythrocytes was significantlyretarded ( > I bed volume), presumably due to adsorp-tion to the Sephadex (a polymer of glucose). To mini-mize this interaction, lactose and galactose were in-cluded in the eluant at concentrations of 50 mm and100 mm respectively (see Materials and Methods).Fractions collected under these conditions were dia-lysed individually to remove the sugars, before being
12
1 1
0
- 9Lu
z
27
4
a
4.4
3
2
Vo Ferritin IgG Albumin Myoglobin
II I I I
HA-2
40 SO 60 70 80
FRACTION NUMBER
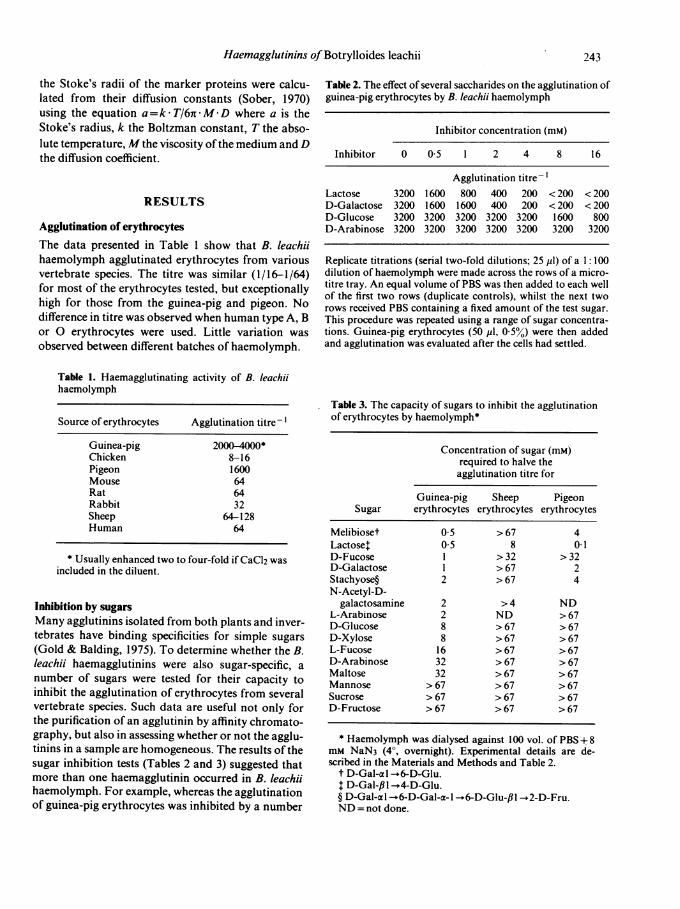
Figure 1. Separation of B. leachii haemagglutinins by chro-matography ofhaemolymph on Sephadex G-200. The eluantbuffer contained 50 mm lactose and 100mm D-galactose. Theagglutinating activities for guinea-pig (a), sheep (-) andpigeon (4) erythrocytes, as well as the elution positions ofthe marker proteins, are shown.
titrated for agglutinating activity against guinea-pig,sheep, pigeon, human and mouse erythrocytes.The results of one fractionation are depicted in Fig.
1. Two peaks of agglutinating activity were resolved.The first (HA-1) peak contained agglutinins specificfor guinea-pig erythrocytes and accounted for essen-tially all (>50%) of the guinea-pig-specific activityapplied to the column. In contrast, the material in thesecond (HA-2) peak represented < 0-5% ofthe appliedactivity against guinea-pig erythrocytes, but agglu-tinated erythrocytes from all the species tested. Therecovery ofthe latter activities was usually > 50%. Theagglutinating titre of fraction 69 for all the differenttypes of erythrocytes listed in Table 1 was 1/8-1/16,except for pigeon erythrocytes (1/64).
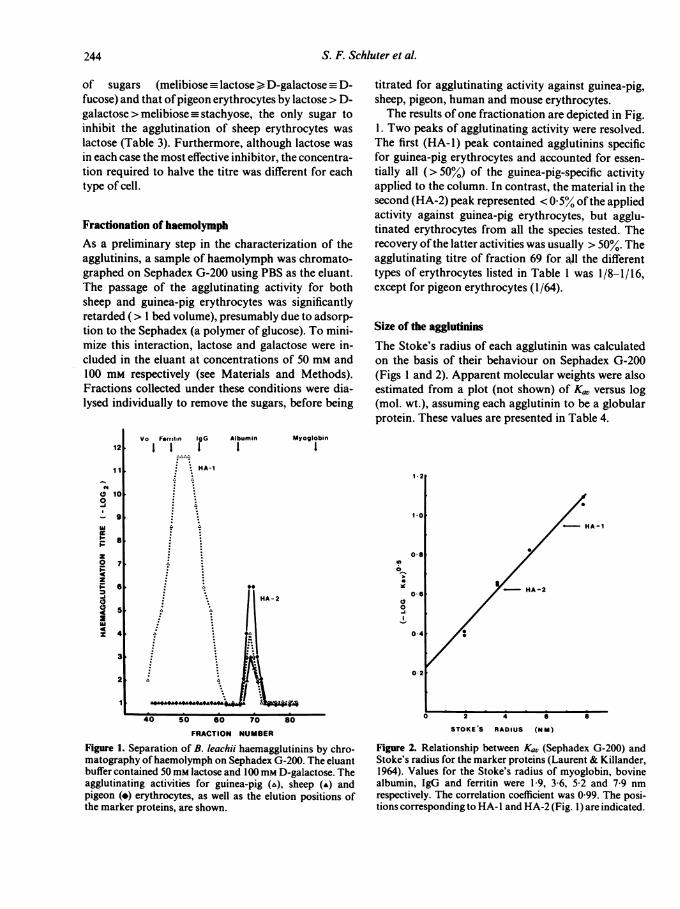
Size of the agglutininsThe Stoke's radius of each agglutinin was calculatedon the basis of their behaviour on Sephadex G-200(Figs I and 2). Apparent molecular weights were alsoestimated from a plot (not shown) of Ku,, versus log(mol. wt.), assuming each agglutinin to be a globularprotein. These values are presented in Table 4.
1-2
1-0HA-1
0-8
O 6 < HA-2
04
0-2
0 2 4 6 S
STOKE'S RADIUS (NbM)
Figure 2. Relationship between Ka, (Sephadex G-200) andStoke's radius for the marker proteins (Laurent & Killander,1964). Values for the Stoke's radius of myoglobin, bovinealbumin, IgG and ferritin were 1-9, 3-6, 5 2 and 7 9 nmrespectively. The correlation coefficient was 0 99. The posi-tions corresponding to HA- I and HA-2 (Fig. 1) are indicated.
244
7
5
5
4
Haemagglutinins ofBotrylloides leachii
Table 4. Molecular size of the HA-1 and HA-2agglutinins
Stoke's radius* ApparentAgglutinin (nm) mol. wt.t
HA-1 6-5 200,000HA-2 3-6 63,000
* Estimated by gel filtration (Figs 1 and 2).t Estimated by plotting K0, (Fig. 1) versus log
(mol. wt.) using IgG (150,000), albumin (67,000)and myoglobin (17,000) as the calibration stan-dards (Pharmacia gel filtration handbook, 1979).
Binding site specificitiesBecause the preceding results demonstrated the pre-sence of at least two agglutinins in B. leachii haemo-lymph, it was considered important to show whetheror not they were structurally related. It seemed poss-ible, for instance, that HA-2 molecules could besubunits or a degraded form of the HA-I agglutinin.Apparent differences in agglutinating specificityand/or titre might in such cases result from differencesin the nature and density of receptor sites on various
cells and from differences in the avidity and agglutinat-ing efficiency of the different forms of the agglutinin(Ehrlich, 1979). The binding specificities of both theHA- I and HA-2 agglutinins were therefore examinedby testing the capacity of various sugars to inhibithaemagglutination, as was done previously for unfrac-tionated haemolymph (Table 2). The results, pre-
sented in Table 5, revealed marked differences in thebinding specificity of the two agglutinins. Moreover,when these results were considered in conjunction withthe overall agglutinating activity recovered in the twopeaks (Fig. 1), it was clear that these two factorsaccounted for the overall activity of unfractionatedhaemolymph. The HA-I and HA-2 agglutinins there-fore appeared to be discrete haemagglutinins presentin the haemolymph of B. leachii.
Adsorption experimentsThe binding specificities of the agglutinins were inves-tigated further by adsorbing haemolymph and prepa-
rations ofHA- I and HA-2 with increasing numbers oferythrocytes ofone or another type. The supernatantscollected after centrifugation were titrated for residual
Table 5. The capacity of various sugars to inhibit the agglutination of erythrocytesby HA-I and HA-2 agglutinins*
Concentration of sugar (mM) required to halve thehaemagglutinating titret
HA- I HA-2
Pigeon Guinea-pig Sheep Human MouseGuinea-pig erythro- erythro- erythro- erythro- erythro-
Sugar erythrocytest cytes cytes cytes cytes cytes
Lactose 0 5 0.1 4 8 8 8
D-Galactose 1 2 > 67 > 67 > 67 > 67
Melibiose 0 5 4! > 67 > 67 > 67 > 67
L-Arabinose 4 > 67 > 67 > 67 > 67 > 67D-Glucose 8 > 67 > 67 > 67 > 67 >67
D-Xylose 16 > 67 > 67 > 67 > 67 > 67L-Fucose 16 > 67 > 67 > 67 > 67 > 67D-Arabinose 32 > 67 >67 > 67 > 67 > 67Maltose 67 > 67 > 67 > 67 > 67 > 67
* Each haemagglutinin was isolated by fractionation of haemolymph onSephadex G-200 as per Fig. 1. Appropriate fractions were pooled and dialysedagainst PBS+azide.
t As for Tables 2 and 3.t Agglutinating activity for the other types of erythrocytes was undetectable.
245
S. F. Schluter et al.
Table 6. Effect of lactose on the capacity of sheep erythro-cytes to adsorb the HA-2 agglutinins for sheep and pigeonerythrocytes
Agglutinating activityrecovered in the
Concentration of dialysed supernatant (%)t forlactose present
during adsorption* Sheep Pigeon(mM) erythrocytes erythrocytes
03 <6 607 6 614 6 122-8 25 255-6 50 501 1 100 10022 100 100
* Samples of dialysed haemolymph (0-5 ml) were mixedwith I ml aliquots of a 5% suspension of sheep erythrocytesin PBS containing an appropriate concentration of lactose.After a 1 h incubation (220), each sample was centrifuged.The supernatants were dialysed against PBS overnightbefore being titrated against sheep and pigeon erythrocytes.
t Unadsorbed (100%) titres were 1/32 (sheep erythro-cytes) and 1/256 (pigeon erythrocytes).
agglutinating activity against the same and other typesof erythrocytes. HA-I activity was adsorbed com-pletely by guinea-pig erythrocytes, but not at all bysheep cells (results not shown). Since guinea-pig cellswere also the only type to be agglutinated by the HA- Iagglutinin, the receptor sites recognized by these mole-cules are apparently unique (among the cell typestested) to guinea-pig erythrocytes.
It seemed, from the sugar-inhibition experiments
(Table 5), that the HA-2 agglutinins might be hetero-geneous, one group of molecules agglutinatinghuman, mouse, sheep and guinea-pig erythrocytes anda second group agglutinating pigeon erythrocytes.However, when the different cell types were tested fortheir capacity to adsorb HA-2 activity, the agglutininsfor every other cell type were in each case removed tothe same extent as that for the type oferythrocyte usedfor the adsorption (results not shown). Significantly,sheep erythrocytes adsorbed the agglutinins for pigeonerythrocytes and vice versa. The results were thereforecompatible with the existence of a single HA-2 agglu-tinin binding with different affinity to the variousreceptor sites on the different erythrocytes, or, alterna-tively, with the existence of two or more agglutinins,both ofwhich bind to receptor sites on all the differentcells but one agglutinating pigeon erythrocytes and thesecond agglutinating other cell types (Lis & Sharon,1977). Since the agglutination of pigeon erythrocytesappeared to be more easily inhibited by lactose thandid the agglutination of sheep erythrocytes (Table 5),it should have been possible to separate the agglu-tinins, if they were in fact distinct, by differentialadsorption to sheep erythrocytes in the presence oflowconcentrations (e.g. 1 mM) of lactose. As is evidentfrom Table 6, this could not be done. Using differentconcentrations of lactose to inhibit adsorption of theagglutinin(s) to sheep erythrocytes (Table 6) and inanother experiment, to pigeon erythrocytes (resultsnot shown), the activity for sheep and pigeon erythro-cytes was adsorbed in parallel. However, the concen-
tration of lactose needed to inhibit adsorption by 50%was different for the two adsorbents, 5 mm being
Table 7. Divalent cation requirements of the HA-I and HA-2 agglutinins
Diluent used for titrationDialysed
Agglutinin against Citrate Ca2+ Mg2+ Ca2+ Mg2+
Agglutination titre-iHA-1 Ca2+/Mg2+ < 200 16,000
EDTA <200 16,000 16,000 <200HA-2 Ca2+/Mg2+ 160 160
EDTA 160 160 -
HA- 1 activity was measured using guinea-pig erythrocytes and HA-2 activityusing sheep and human erythrocytes, after haemolymph had been dialysedexhaustively against 25 mm tris-buffered saline (pH 8) containing either 1 mMCaCl2 plus 1 mM MgCl2 or 1 mM EDTA. The diluents were 0-13 M trisodiumcitrate, tris-buffered saline containing 1 mM CaC12 plus 1 mM MgCI2, ortris-buffered saline containing 0-05mM EDTA and either 0-1 mM CaCl2 or 0 1mM MgC12.
246

Haemagglutinins of Botrylloides leachiii
required when sheep cells were used (Table 6) and0-1-1-0 mm when pigeon cells were used (data notshown).
Divalent cation requirementsTo determine whether divalent cations were necessaryfor the activity of either agglutinin, samples were dia-lysed against tris-buffered saline containing eitherCa2+ and Mg2+ ions or EDTA. Each was then titratedfor agglutinating activity using as diluent, isotonicsodium citrate, or tris-buffered saline containingCaC12, MgCl2 or CaCl2 plus MgCl2 (Table 7). It isclear from the results that the HA-1 agglutinin was
reversibly inactivated in the presence of chelatingagents and that full activity was restored upon theaddition ofCa2+ but not Mg2+ ions. HA-2 activity, onthe other hand, showed no dependency on divalentcations.
DISCUSSION
The existence of haemagglutinating factors in inverte-brates is well documented, although few agglutininshave been purified and characterized physicochemi-cally. Of those for which such information is available(from molluscs, arthropods and sponges), all are
multivalent, polymeric glycoproteins with binding sitespecificities for small saccharide units (Marchalonis &Edelman, 1968; Acton, Bennett, Evans & Schrohen-loher, 1969; Hammarstrom & Kabat, 1969; Hall &Rowlands, 1974; Bretting & Kabat, 1976; Baldo,Sawyer, Stick & Uhlenbruck, 1978). Some of thesefactors may function in vivo as opsonins (Jenkin, 1976;Renwrantz & Mohr, 1978). Much less is known aboutthe protochordates, despite their close affinity to thevertebrates. The haemolymph of a number ofascidianspecies has been reported to contain agglutinatingfactors, but none has yet been purified (Fuke & Sugai,1972; Bretting & Renwrantz, 1973; Wright, 1974;Anderson & Good, 1975; Parrinello & Patricolo,1975).The results of this study show that B. leachii haemo-
lymph contains agglutinins for a wide variety of verte-brate erythrocytes. The titre was similar for most ofthe erythrocytes examined, although higher titres wereobserved using pigeon and guinea-pig cells. The agglu-tination of all cell types was reversibly inhibited bylactose. However, in the case ofpigeon and guinea-pigerythrocytes, several other related sugars were also
inhibitory. The agglutinins, like those from anotherascidian Halocynthia pyriformis (Anderson & Good,1975), therefore recognized carbohydrate residues onthe erythrocytes. This was substantiated when the agg-lutinating factors were found to be adsorbed toSephadex G-200. When haemolymph was applied to acolumn of Sephadex equilibrated with a buffer con-taining high concentrations of galactose and lactose,however, the passage of the agglutinins did not appearto be retarded. Under these conditions, two peaks ofactivity were resolved. The first (HA- 1) peak con-tained agglutinins specific for guinea-pig erythrocytesand accounted for essentially all ofthe original activityfor these cells, whilst the second (HA-2) peak con-tained agglutinins for every type of erythrocyte tested.On the basis of their chromatographic behaviour, themolecular weights of these agglutinins were estimatedto be approximately 200,000 and 63,000 respectively.
It was initially considered that HA- 1 and HA-2 maybe different forms of a single agglutinin. This possibi-lity was rejected for the following reasons. Firstly, theHA-I factors agglutinated only guinea-pig erythro-cytes, whilst the HA-2 molecules agglutinated everyspecies of erythrocyte examined, including those fromthe guinea-pig. Secondly, although guinea-pig eryth-rocytes could adsorb out both HA- I and HA-2 activi-ties, other cell types were able to adsorb only HA-2agglutinins. Thirdly, the agglutination of guinea-pigerythrocytes by the HA- I and HA-2 factors exhibiteddifferent sensitivities to inhibition by various sugars.The agglutinating titre of HA-I was halved, for exam-ple, in the presence of 0-5 mm lactose or I mm D-galac-tose, whereas that of HA-2 was halved in the presenceof 4 mm lactose but was not affected by even 67 mmgalactose (Table 5). Since the inhibitory capacity of asugar is related to its intrinsic association constant forthe binding sites of an agglutinin (Lootiens, vanWauwe & de Bruyne, 1975; see Appendix), theseresults showed that the individual binding sites of thetwo agglutinins were different and that the agglutininswere therefore unrelated. This has been subsequentlyconfirmed by a lack of antigenic cross-reactivity usingspecific antisera raised in rabbits against each agglu-tinin (Schluter, Ey, Keough & Jenkin, manuscript inpreparation).
It is possible that the B. leachii agglutinins have arole analagous to that of vertebrate antibodies, func-tioning as 'recognition' factors for the defence systemof the organism (Wright, 1974; Keough, unpublishedresults). It was therefore of considerable interest todetermine whether the agglutinins present in the HA-2
247

S. F. Schluter et al.
peak were homogeneous or consisted of a populationof related molecules, each with different binding-sitespecificities.The HA-2 agglutinins exhibited similar activity
against every species of erythrocyte tested (Table 1),with the exception ofpigeon cells for which a consider-ably higher titre was apparent. The agglutinating titrewas in each case reduced in the presence oflactose, 4-8mM halving the titre for guinea-pig, sheep, human andmouse erythrocytes but only 0-1 mm being needed withpigeon cells (Table 5). Moreover, D-galactose (2 mM)halved the titre for pigeon erythrocytes whilst 67 mmgalactose had no effect on the agglutination of theother cell types. These results suggested that more thanone type of agglutinin might be present in the HA-2fraction. In a series of adsorption experiments, how-ever, the agglutinins for both pigeon and sheep eryth-rocytes could not be resolved and showed identicalsusceptibility to inhibition by lactose. Additionally,the adsorption of both agglutinins to sheep erythro-cytes was less sensitive to lactose inhibition than wastheir adsorption to pigeon cells. It therefore appearedthat one agglutinin was responsible for the agglutina-tion of both these (and other) cell types. Differences inthe affinity of the agglutinin for the pigeon and sheeperythrocyte receptor(s) and/or in the properties (e.g.size, surface receptor density, surface charge) of thevarious cell types (see Appendix) probably accountedfor these findings.
There are numerous reports in the literature inwhich the activity of invertebrate agglutinins has beenfound to require Ca2+ ions. Examples include thosefrom the arthropods Limulus polyphemus (Marcha-lonis & Edelman, 1968) and Homarus americanus(Hall & Rowlands, 1974), the molluscs Crassostreavirginica (Acton et al., 1969) and Tridacna maxima(Baldo et al., 1978) and the sponge Axinellapolypoides(Bretting & Kabat, 1976). Among the protochordates,haemagglutinins from Halocynthiapyriformis (Ander-son & Good, 1975) and Ciona intestinalis (Wright &Cooper, 1975) have been reported to require Ca2+ions, although the activity of C. intestinalis agglutininshas elsewhere been found to be independent of di-valent cations (Parrinello & Patricolo, 1975). The agg-lutinins from Phallusia mammillata, Ascidia malacaand Styela plicata do not appear to require divalentcations (Fuke & Sugai, 1972; Parrinello & Patricolo,1975). These results may be misleading, however, for ifother ascidians have multiple agglutinins analogous tothe Ca2+-dependent HA-1 and divalent cation-inde-pendent HA-2 agglutinins of B. leachii, any conclu-
sions concerning divalent cation dependence would beinfluenced by whichever agglutinin(s) were detected bythe particular indicator cell(s) used. The haemolymphof a Botryllus sp., an ascidian very closely related to B.leachii, also contains a Ca2+-dependent, galactose/lac-tose-specific guinea-pig erythrocyte agglutinin whichhas a molecular weight similar to that of the B. leachiiHA-I molecule (Schluter, Keough, Ey & Jenkin, un-published data).The agglutinins described in this paper are similar to
those from several other invertebrate species in pos-sessing a specificity for carbohydrate residues (e.g.Anderson & Good, 1975; Bretting & Kabat, 1976;Baldo et al., 1978). The natural function of thesemolecules is unknown. Although they may have rolesunrelated to the defence of the animal, the possibilitythat they act as opsonins cannot yet be excluded, for,despite their restricted binding specificity, they mayrecognize determinants which are common to patho-gens such as marine bacteria.
ACKNOWLEDGMENTS
We thank Ms Carol Heanes for typing the manuscript.This work was supported by the Australian ResearchGrants Committee. P.L.E. holds the position ofSeniorResearch Officer under a grant from the NationalHealth and Medical Research Council of Australia.S.F.S. is the recipient of an education allowance pro-vided by the Commonwealth Department ofVeterans'Affairs.
APPENDIX
A theoretical analysis of the inhibition of haemagglu-tination reactions by small ligands is not available, butthe parameters which must be considered when oneevaluates such data can be identified by a simple exa-mination of the reactions involved. For example, con-sider the interaction of a cell surface-associated recep-tor (R) and a small ligand (L) with a homogeneous,multivalent agglutinin (A). The system can be approxi-mately described by the expression
A+L+RR
The association constants for these reactions may bedefined as
248

Haemagglutinins of Botrylloides leachii 249
[AL][A] [L]
KR [AR] (2)
where [A], [L], [RI, [AL] and [AR] represent the concen-trations of the respective molecular species when thesystem is at equilibrium. For simplicity, the associ-ation constants are considered to include all speciesarising from the multivalency of the agglutinin (e.g.AL, AL2,.-.,AL,). Also, although the concentrationof R (and consequently, also of AR) can only bedescribed by complex mathematical functions(Reynolds, 1979), for the purposes of this analysis itsuffices to recognize that [R] depends on both thenumber of molecules ofR present on each cell and thecell concentration. Dividing equation (2) by equation(1), one obtains
[AR] KR [R] (3)
[AL] KL [L]Providing KR is large (e.g. > 100) and the initial
concentrations [RIo > [A]o, then at equilibrium [A] willbe negligible and [AR] + [AL] - [A]o. These conditionscan be expected to exist when erythrocytes are agglu-tinated by a limiting amount of agglutinin, e.g. near atitration endpoint. Since, in this system, ligand mole-cules compete with the cell receptors for the bindingsites of the agglutinin molecules, one may considerthat, at the concentration of ligand ([L]j) which halvesthe titre of the agglutinin with respect to the uninhi-bited titre (observed in the absence of ligand),[AL]= [AR]. Thus, from equation (3),
[LI KR*[R] (4)KL
It is evident from equation (4) that in any givenexperimental system, the concentration of ligandrequired to halve the titre ofa haemagglutinin dependsnot only on the affinity of the agglutinin for the ligand(KL) (Loontiens et al., 1975), but also on the affinity/-avidity of the agglutinin for the cell receptors (KR) andon the receptor concentration ([R]). Thus, using agiven assay, one agglutinin and one type of cell, theconcentration of a sugar needed to halve the agglu-tinating titre is inversely proportional to the affinity ofthe agglutinin for that sugar. When one compares thecapacity of a particular sugar to inhibit the agglutina-tion of different types of cell, however, the inhibitoryconcentrations depend not only on KL (which is the
same in each case), but also on the affinity/avidity ofthe agglutinin for the receptors on each type of cell aswell as on the effective receptor concentration. Thelatter is a function of cell concentration, cell size andcell surface receptor density (Reynolds, 1979).
REFERENCES
ACTON R.T., BENNETT J.C., EVANS E.E. & SCHROHENLOHERR.E. (1969) Physical and chemical characterization of anoyster hemagglutinin. J. Biol. Chem. 244, 4128.
ANDERSON R.S. & GOOD R.A. (1975) Naturally occurringhaemagglutinin in a tunicate Halocynthia pyriformis.Biol. Bull. 148, 357.
BALDO B.A., SAWYER W.H., STICK R.V. & UHLENBRUCK G.(1978) Purification and characterization of a galactan-reactive agglutinin from the clam Tridacna maxima (Rod-ing) and a study of its combining site. Biochem. J. 175,467.
BRET`TING H. & KABAT E.A. (1976) Purification and charac-terization of the agglutinins from the sponge Axinellapolypoides and a study of their combining sites. Bio-chemistry, 15, 3228.
BRETTING H. & RENWRANTz L. (1973) Investigations of in-vertebrates of the Mediterranean Sea with regard to theircontents ofhaemagglutinins. Z. Immun.-Forsch. 145,242.
EHRLICH P.H. (1979) The effect of multivalency on the speci-ficity of protein and cell interactions. J. Theoret. Biol. 81,123.
Ey P.L., PROWSE S.J. & JENKIN C.R. (1978) Isolation of pureIgG 1, IgG2a and IgG2b immunoglobulins from mouseserum using protein A-Sepharose. Immunochemistry, 15,429.
FUKE M.T. & SUGAI T. (1972) Studies on the naturally occur-ring haemagglutinin in the coelomic fluid of an ascidian.Biol. Bull. 143, 140.
GOLD E.R. & BALDING P. (1975) Receptor-specific proteins.Plant andanimal lectins, Ch. 7. Excerpta Medica, Amster-dam. American Elsevier Publ. Co., Inc. N.Y.
HALL J.L. & ROWLANDs D.T. JR. (1974) Heterogeneity oflobster agglutinins. I. Purification and physicochemicalcharacterization. Biochemistry, 13, 821.
HAMMARSTROM S. & KABAT E.A. (1969) Purification andcharacterization of a blood-group A reactive haemagglu-tinin from the snail Helix pomatia and a study of itscombining site. Biochemistry, 8, 2696.
JENKIN C.R. (1976) Factors involved in the recognition offoreign material by phagocytic cells from invertebrates.Comparative Immunology (Ed. by J. J. Marchalonis), ch.4. Blackwell Scientific Publications, Oxford.
LAURENT T.C. & KILLANDER J. (1964) A theory of gelfiltration and its experimental verification. J. Chroma-togr. 14, 317.
Lis H. & SHARON N. (1977) Lectins: their chemistry andapplications to immunology. The Antigens, vol. 4, p. 429.Academic Press, N.Y.
LOONTIENS F.G., VAN WAUWE J.P. & DE BRUYNE C.K. (1975)Concanavalin A: relationship between hapten inhibitionindexes and association constants for different glycosides.Carbohydr. Res. 44, 150.

250 S. F. Schluter et al.
MARCHALONIS J.J. & EDELMAN G.M. (1968) Isolation andcharacterization of a hemagglutinin from Limulus poly-phemus. J. Mol. Biol. 32,453.
PARRINELLO N. & PATRICOLO E. (1975) Erythrocyte agglu-tinins in the blood of certain ascidians. Experientia, 31,1092.
RENWRANTZ L. & MOHR W. (1978) Opsonizing effect ofserum and albumin gland extracts on the elimination ofhuman erythrocytes from the circulation of Helix poma-tia. J. Invert. Path. 31, 164.
REYNOLDS J.A. (1979) Interaction of divalent antibody withcell surface antigens. Biochemistry, 18, 264.
SCHACTERLE G.R. & POLLACK R.L. (1973) A simplifiedmethod for the quantitative assay of small amounts ofprotein in biologic material. Anal. Biochem. 51, 654.
SIEGEL L.M. & MONTY K.J. (1966) Determination ofmolecu-lar weights and frictional ratios of proteins in impuresystems by use ofgel filtration and density gradient centri-fugation. Application to crude preparations of sulphiteand hydroxylamine reductases. Biochim. Biophys. Acta,112, 346.
SOBER H.A., ed. (1970) Handbook of Biochemistry. Selecteddata for molecular biology. 2nd edn, pp. C3-C24. TheChem. Rubber Co., Cleveland.
WRIGHT R.K. (1974) Protochordate immunity. I. Primaryimmune response of the tunicate Ciona intestinalis tovertebrate erythrocytes. J. Invert. Path. 24, 29.
WRIGHT R.K. & CooPER E.L. (1975) Immunological matu-ration in the tunicate Ciona intestinalis. Amer. Zool. 15,21.















![)JOEBXJ1VCMJTIJOH$PSQPSBUJPO ...con rmedinthemuscles,haemolymph,andreproductive system of A. suum []. TPS was isolated from muscles of A. suum and its properties have been detected.](https://static.fdocuments.net/doc/165x107/60e57c50dafc1611b11f9c61/joebxj1vcmjtijohpsqpsbujpo-con-rmedinthemuscleshaemolymphandreproductive.jpg)


