
Henryk Kozlowski - Home - people.unica.it - Università...
60
Henryk Kozlowski Faculty of Chemistry University of Wroclaw CAGLIARI, SEPTEMBER, 2013
Transcript of Henryk Kozlowski - Home - people.unica.it - Università...

Henryk Kozlowski Faculty of Chemistry
University of Wroclaw
CAGLIARI, SEPTEMBER, 2013
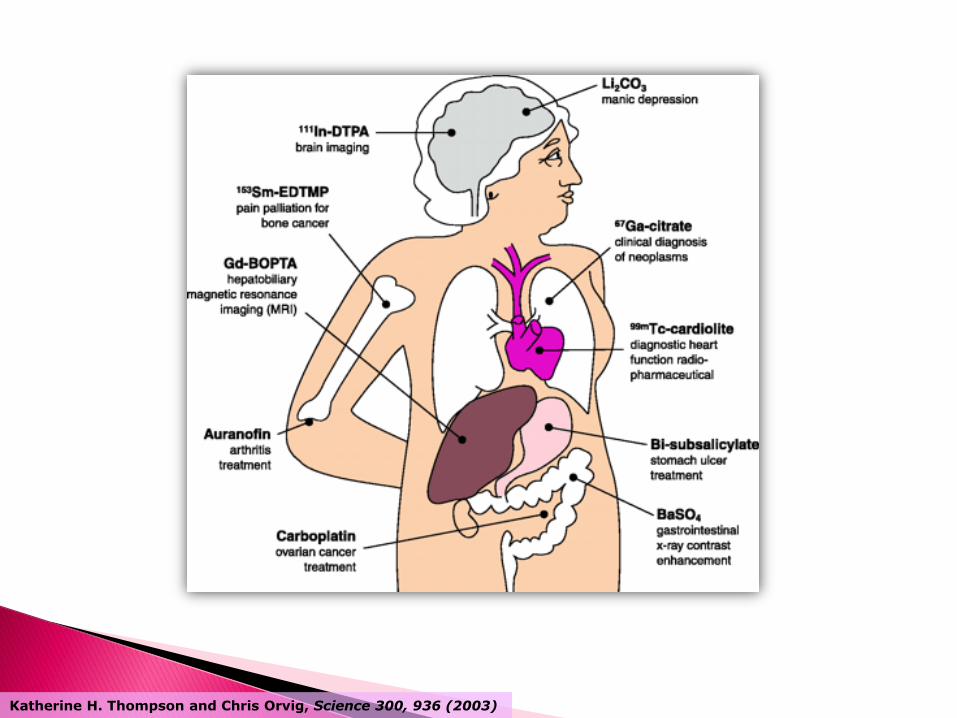
Katherine H. Thompson and Chris Orvig, Science 300, 936 (2003)


Barry J. Marshall
J. Robin Warren

Barry J. Marshall
J. Robin Warren

Urease makes envi-ronment around the bacteria more basic
Hydrogenase plays a vital role in anaerobic metabolism
Urease and hydrogenase accessory proteins, involved in the homeostasis of Ni2+
How can Helicobacter pylori survive in adverse conditions of our stomach?

CO(NH2)2 + H2O→ 2 NH3 + CO2
The active site of urease contains two nickel atoms (green) bridged by a carboxylated lysine and a hydroxyl group.
The urease active site
Eric L. Carter,a Nicholas Flugga, Jodi L. Boer,b Scott B. Mulrooneya and Robert P. Hausinger, Metallomics, 2009, 1, 207-221

Urease maturation
D. Witkowska, M. Rowińska-Żyrek, G. Valensin and H. Kozłowski. Coord.Chem.Rev., 2012, 256, 133-148.

The active site of [NiFe] hydrogenase
W. Zhu, A. C. Marr, Q. Wang, F. Neese, D. J. E. Spencer, A. J. Blake, P. A. Cooke, C. Wilson, and M. Schroder, PNAS, 2005, 18280–18285

Fe2+
HycE
HypC
HypC
HypD
HypE
HypF
HypA
HypB
SlyD
ATP
ADP+Pi
ATPCP
AMP+PPi
HycE
Ni2+GTP
HycEHypC
HypC
GDP+Pi
HycE
HycI
HycI
Fe
Ni
S SFe
Ni
S S
Fe
Ni
S S
Fe
Fe2+
HycE
HypC
HypC
HypD
HypE
HypF
HypA
HypB
SlyD
ATP
ADP+Pi
ATPCP
AMP+PPi
HycE
Ni2+GTP
HycEHypC
HypC
GDP+Pi
HycE
HycI
HycI
Fe
Ni
S SFe
Ni
S S
Fe
Ni
S S
Fe
D. Witkowska, M. Rowińska-Żyrek, G. Valensin and H. Kozłowski. Coord.Chem.Rev., 2012, 256, 133-148.
Hydrogenase maturation

- Metronidazole 500 mg + Amoxycycline 1 g + PPI
- Klatromycine 500 mg +
Metronidazol 500 mg + PPI - Klatromycine 500 mg +
Amoxycycline 1 g + PPI
PPI: omeprazole, lansoprazole, rabeprazole, esomeprazole, pantoprozole
RBS, rantidine bismuth subcitrate (Tritec, Pylorid) Bismuth(III) compounds CBS, colloidal bismuth subcitrate (De-Nol) BSS, bismuth subsalicate
Getting rid of H. pylori

M. Rowińska-Żyrek, D. Valensin, Ł. Szyrwiel, Z. Grzonka, H. Kozłowski, Dalton Trans., 2009, 9131-9140
Species log β logK
HL
H2L
H3L
H4L
H5L
H6L
10.75 (2)
21.02 (1)
30.13 (2)
38.57 (2)
46.40 (2)
50.13 (3)
10.75 Lys
10.25 Cys
9.11 Cys
8.44 Cys
7.83 Cys
3.75 Asp
BiH2L 52.29 (8)
BiHL 48.51 (4) 3.78
BiL 39.32 (5) 9.19 2 4 6 8 10
0
20
40
60
80
100
BiL
BiHLBiH
2L
Ac-GCASCDNCRAAKK-NH2 : Bi
3+ = 2:1
% fo
rma
tio
n r
ela
tive
to
Bi3
+
pH
free Bi3+
The complicated relationship between bismuth and protein fragments:
• Cysteine thiol sulfurs are the only anchoring sites for bismuth • Extremaly stable complexes are formed already at pH < 1

Competition plot of Ac-CGC-NH2, MDPETCPCP-NH2, Ac-CHCH-NH2 and Ac-ACCHDHKKH-NH2 with Bi3+ (1:1:1:1:2)
• Coordination of Bi3+ to the CXC sequence – much stronger than the coordination to the CC sequence • Involvement of histidines in the binding of Bi3+ enhances the stability of the complex
M. Rowińska-Żyrek, D. Valensin, L. Szyrwiel, Z. Grzonka and H. Kozlowski, Dalton Trans., 2009, 9131 M. Rowińska-Żyrek, D. Witkowska, D. Valensin, W. Kamysz and H. Kozlowski, Dalton Trans, 2010, 39 , 5814.
2 4 6 8
0
20
40
60
80
100
Bi3+
-[Ac-CGC-NH2]2 Bi
3+-Ac-ACCHDHKKH-NH
2
Bi3+
-MDPETCPCP-NH2
Bi3+
-Ac-CHCH-NH2
% fo
rma
tio
n r
ela
tive
to
Bi3
+
pH
free Bi3+

How strong are Bi3+ - imidazole interactions?
2 4 6 8 10
0
10
20
30
40
50
60
70
80
90
100
Bi3+
Bi3+
-Ac-CHCH-NH2
Bi3+
-MPGCPCPGCG-NH2
Bi3+
: MPGCPCPGCG-NH2 : Ac-CHCH-NH
2 = 1:1:1
% fo
rma
tio
n r
ela
tive
to
[B
i3+]
pH
Potocki S., Rowińska-Żyrek M., Valensin D., Krzywoszyńska K, Witkowska D., Łuczkowski M, Kozłowski H., Inorg. Chem, 2011

2 4 6 8 10
0
10
20
30
40
50
60
70
80
90
100
Bi-Ac-CCSTSDSHHQ-NH2
Bi-MDPETCPCP-NH2
Bi-Ac-GCASCDNARAAKK-NH2
Ac-GCASCDNARAAKK-NH2 : MDPETCPCP-NH
2 : Ac-CCSTSDSHHQ-NH
2 : Bi
3+ = 1:1:1
% fo
rma
tio
n r
ela
tive
to
[B
i3+]
pH
What is the impact of the distance between Cys residues in peptide fragments on the stability of the complexes they form with Bi3+ ions?
CXXC
CXC
CC
Rowińska-Żyrek M., Witkowska D., Bielinska S., Kamysz W. and Kozlowski H., Dalton Trans., 2011, 40, 5604-5610.
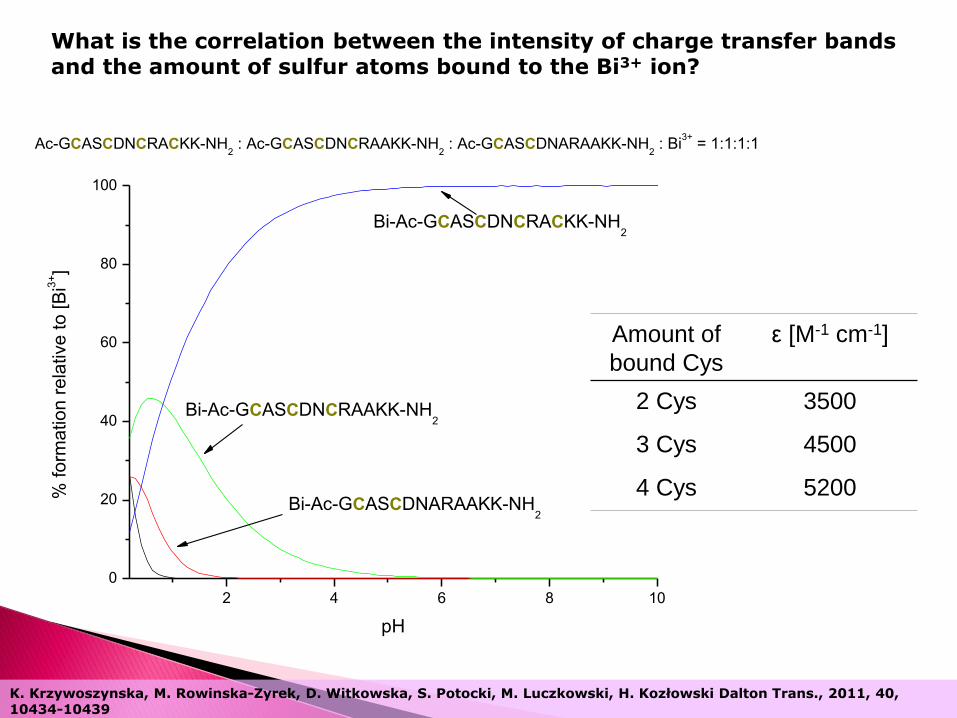
2 4 6 8 10
0
20
40
60
80
100
Bi-Ac-GCASCDNARAAKK-NH2
Bi-Ac-GCASCDNCRAAKK-NH2
Bi-Ac-GCASCDNCRACKK-NH2
Ac-GCASCDNCRACKK-NH2 : Ac-GCASCDNCRAAKK-NH
2 : Ac-GCASCDNARAAKK-NH
2 : Bi
3+ = 1:1:1:1
% fo
rma
tio
n r
ela
tive
to
[B
i3+]
pH
What is the correlation between the intensity of charge transfer bands and the amount of sulfur atoms bound to the Bi3+ ion?
Amount of
bound Cys
ε [M-1 cm-1]
2 Cys 3500
3 Cys 4500
4 Cys 5200
K. Krzywoszynska, M. Rowinska-Zyrek, D. Witkowska, S. Potocki, M. Luczkowski, H. Kozłowski Dalton Trans., 2011, 40, 10434-10439

91GSCCHTGNHDHKHAKEHEACCHDHKKH118
Cun et al., J. Biol. Chem. 2008, 283, 22, p. 15142

300 400 500
-10
0
10
NiLBi0,6
NiLBi0,45
NiLBi0,3
NiLBi0,15
NiL
[nm]
CD spectra of Ni2+ complexes with the B-domain peptide at pH6, titrated with Bi3+ ions in a step of 0.15 molar equivalents. Final Bi3+ : Ni2+ : L ratio = 1 : 1 : 2.
Bi3+ is able to displace Ni2+ from it’s binding sites:
M. Rowińska-Żyrek, D. Witkowska, D. Valensin, H. Kozlowski, Dalton Trans. 2010, 39 , 5814.

M. Rowińska-Żyrek, D. Witkowska, D. Valensin, H. Kozlowski, Dalton Trans. 2010, 39 , 5814.


W. Xia, H. Li, K. H. Sze and H. Sun, J. Am. Chem. Soc. 131 (2009) 10031–10040.
Metallochaperone HypA

Communication between the zinc and nickel sites in dimeric HypA
Herbst R.W.; Perovic I.; Martin-Diaconescu V.; O’Brien K.; Chivers P.T.; Pochapsky S.S.; Pochapsky T.C.; Maroney M.J.; J. Am. Chem. Soc. 2010, 132 (30), 10338–10351

W. Xia, H. Li, K. H. Sze and H. Sun, J. Am. Chem. Soc. 131 (2009) 10031–10040.
Metallochaperone HypA

Ac-ELECKDCSHVFKPNALDYGVCEKCHS-NH2
HypA loop sequence - residues 71-96

764.3
746.3
749.8
755.6
763.9765.1
766.1 768.8770.9
745 750 755 760 765 770 775 m/z0.0
0.2
0.4
0.6
0.8
1.0
1.2
8x10
Intens.
ZnL4+
L4+
NaL4+
Ac-ELECKDCSHVFKPNALDYGVCEKCHS-NH2 + 1 eq. Zn(II)
763.6
749.8
755.3
745 750 755 760 765 770 775 m/z0
1
2
3
4
5
6
8x10
Intens.
NiL4+
L4+
NaL4+
Ac-ELECKDCSHVFKPNALDYGVCEKCHS-NH2 + 1 eq. Ni(II)
ESI-MS spectra of a system containing Ac-ELECKDCSHVFKPNALDYGVCEKCHS-NH2,
with Zn2+ (A) and Ni2+ ions (B) in a 1 : 1 : 1 stoichiometry
M. Rowinska-Zyrek, S. Potocki, , D. Witkowska, D.Valensin H. Kozlowski, Dalton Trans., 42, 6012-6020, (2013)

4 6 8 10
0
20
40
60
80
100
% fo
rma
tion
re
lativ
e to
lig
an
d
pH
ZnL
NiL
Ac-ELECKDCSHVFKPNALDYGVCEKCHS-NH2 : Zn
2+ : Ni
2+ = 1 : 1 : 1
A competition plot of Ni2+ and Zn2+ complexes with the
Ac-ELECKDCSHVFKPNALDYGVCEKCHS-NH2 fragment
M. Rowinska-Zyrek, S. Potocki, , D. Witkowska, D.Valensin H. Kozlowski, Dalton Trans. 42, 6012-6020, (2013)

conserved –CXXC- motifs
Ac-ELECKDCSHVFKPNALDYGVCEKCHS-NH2
Asp
Cys Ser
His
Val Ni

Communication between the zinc and nickel sites in dimeric HypA
Herbst R.W.; Perovic I.; Martin-Diaconescu V.; O’Brien K.; Chivers P.T.; Pochapsky S.S.; Pochapsky T.C.; Maroney M.J.; J. Am.
Chem. Soc. 2010, 132 (30), 10338–10351

Types of binding modes in Zn2+ complexes with the
Ac-ELECKDCSHVFKPNALDYGVCEKCHS-NH2 fragment
M. Rowinska-Zyrek, D. Witkowska, D.Valensin, S. Potocki, H. Kozlowski, Dalton Trans., 42, 6012-6020, (2013)
4 6 8 10
0
20
40
60
80
100
Zn2+
ZnHL
ZnLZnH
2L
ZnH3L
ZnH4L
ZnH5L
% fo
rma
tio
n r
ela
tive to
[Z
n2
+]
pH
{3S-,Nim} {4S-}


Metal ions + Hpn
R. Ge, Y. Zhang, X. Sun, R. Watt, Q-Y. He, J-D. Huang, D. Wilcox, H. Sun, J. Am. Chem. Soc., 2006, 128 (35), 11330
MAHHEEQHGG HHHHHHHTHH HHYHGGEHHH
HHHSSHHEEG CCSTSDSHHQ EEGCCHGHHE


…a specific sequence of string of six to nine histidine residues
…used in vectors for productions of recombinant proteins Recombinant protein with a six histidine or poly-His tag connected to N- or C-terminus
Expressed His-tagged proteins can be purified and
detected easily because the string of histidine residues binds to several types of immobilized metal ions, including nickel, zinc and copper, under specific buffer conditions.
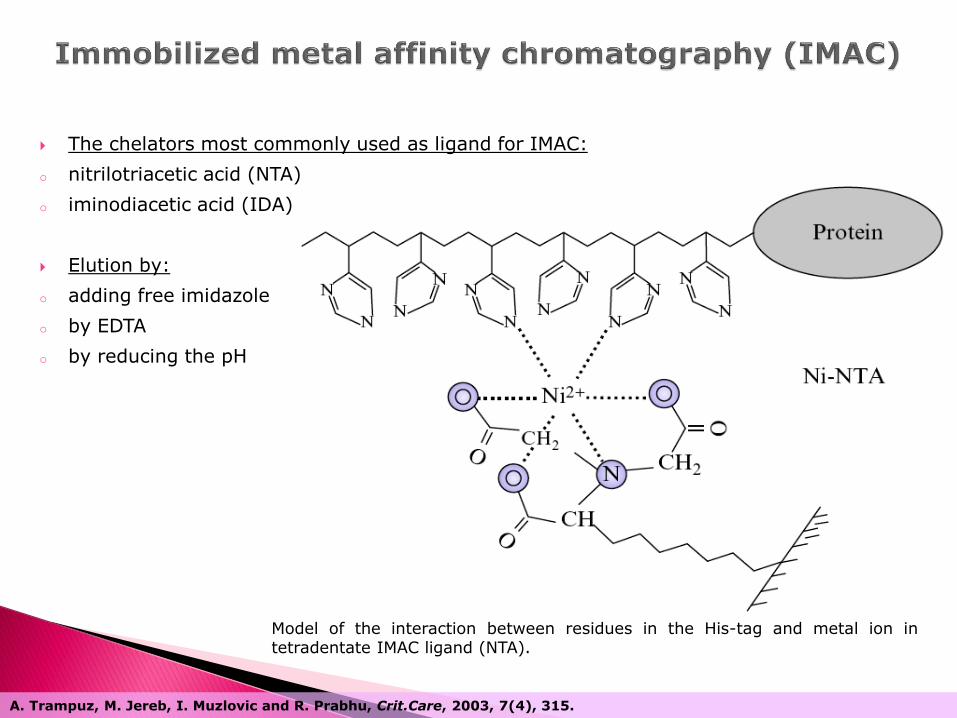
The chelators most commonly used as ligand for IMAC:
o nitrilotriacetic acid (NTA)
o iminodiacetic acid (IDA)
Elution by:
o adding free imidazole
o by EDTA
o by reducing the pH
Model of the interaction between residues in the His-tag and metal ion in tetradentate IMAC ligand (NTA).
A. Trampuz, M. Jereb, I. Muzlovic and R. Prabhu, Crit.Care, 2003, 7(4), 315.

4 6 8 10
0
20
40
60
80
100
free L
Ni2+
-Ac-CCSTSDSHHQ-NH2
Zn2+
-Ac-CCSTSDSHHQ-NH2
% fo
rma
tio
n r
ela
tive
to
L
pH
The Ni2+ complex of Ac-CCSTSDSHHQ-NH2 peptide
from H.pylori Hpn protein is much more stable than that of zinc
M. Rowińska-Żyrek, D. Witkowska, S. Bielińska, W. Kamysz, H. Kozłowski. Dalton Trans., 2011, 40, 5604-5610
MAHHEEQHGG HHHHHHHTHH HHYHGGEHHH
HHHSSHHEEG CCSTSDSHHQ EEGCCHGHHE

4 6 8 10
0
20
40
60
80
100
% fo
rma
tion
re
lativ
e to
L
free L
Zn-Ac-EEGCCHGHHE-NH2
Ni2+
-Ac-EEGCCHGHHE-NH2
pH
The Ni2+ complex of Ac-EEGCCHGHHE-NH2 peptide from H.pylori Hpn protein is much more stable
than that of zinc
M. Rowińska-Żyrek, D. Witkowska, S. Bielińska, W. Kamysz, H. Kozłowski. Dalton Trans., 2011, 40, 5604-5610
MAHHEEQHGG HHHHHHHTHH HHYHGGEHHH
HHHSSHHEEG CCSTSDSHHQ EEGCCHGHHE

300 400 500 600 700 800
-20
-10
0
10
20
(mol
-1cm
-1)
pH4
pH7-pH9
Ni2+
-Ac-EEGCCHGHHE-NH2
pH61
[nm]
pH5
300 400 500 600 700 800
-50
-40
-30
-20
-10
0
10
20
30
40
50
Ni2+
-Ac-CC-NH2
pH7-pH10
pH6
pH5,3/m
olxc
m[nm]
pH4
M. Rowińska-Żyrek, D. Witkowska, S. Bielińska, W. Kamysz, H. Kozłowski. Dalton Trans., 2011, 40, 5604-5610
CD spectra of Ni2+complexes of Ac-CC-NH2 and Ac-EEGCCHGHHE-NH2
peptides
X
NH2CH
CH2
O
S
N
CHCH2
O
S
NH2
Ni

MAHHEQQQQQQA-NH2 (Hpn-like) MAHHEEQHG-NH2 (Hpn) Ac-EEGCCHGHHE-NH2 (Hpn) Ac-THHHHYHGG-NH2 (Hpn) Ac-ACCHDHKKH-NH2 (HspA)
Copper and nickel binding to fragments
of Helicobacter pylori proteins

4 6 8 10
0
20
40
60
80
100
DAH-NH2 : Ac-ACCHDHKKH-NH
2 : Ni
2+= 1:1:1
free Ni2+
Ni2+
-Ac-ACCHDHKKH-NH2
% fo
rmat
ion
rela
tive
to N
i2+
pHM. Rowinska-Zyrek, D. Witkowska, D. Valensin, W. Kamysz and H. Kozlowski, Dalton Trans, 2010, 39 , 5814 - 5826 P. Mlynarz, D. Valensin, K. Kociolek, J. Zabrocki, J. Olejnik, H. Kozlowski, New J. Chem., 2002, 26, p.264
The distribution of Ni2+ between DAH-NH2 and Ac-ACCHDHKKH-NH2 peptides
HN
CH
C
H2C
O
S
NCH
C
CH2
O
S
NH2
C
3HC
O Ni
X
Ni
N
N
N
H2N
N
O
OX
Y

NH2
CH
CH2
O
N
NH
NCH
CH2
O
N
NH
NH
CHCH2
O
N
NH
NH
CH
CH2
O
N
NH NH
CH
CH2
O
OH
OH
NiNH
CHCH2
O
S
H3C
N
CH
CH3
O
NCH
CH2
O
N+
NH
OH
Ni
X
NH2CH
CH2
O
S
N
CHCH2
O
S
NH2
Ni
Witkowska D., Politano R., Rowińska-Żyrek M., Guerrini R., Remelli M., Kozłowski H., Chem. Eur. J., 2012, 18, 11088-11099.
Binding modes of nickel to various Hpn sequences
MAHHEEQHGG HHHHHHHTHH HHYHGGEHHH
HHHSSHHEEG CCSTSDSHHQ EEGCCHGHHE

Competition plots of Ac-THHHHHYHGG-NH2 and MAHHEEQHG-NH2 with Cu2+ and Ni2+
His-rich complexes are more stable only at below–physiological pH
Rowińska-Żyrek M., Witkowska D., Potocki S., Remelli M., Kozłowski H., New J. Chem., 2013, 37, 58-70.

Ni
N
N
N
H2N
N
O
OX
Y
ALBUMIN-LIKE BINDING
Xaa-Yaa-His-

MAHHEQQQQQQA-NH2 (Hpn-like) MAHHEEQHG-NH2 (Hpn) Ac-EEGCCHGHHE-NH2 (Hpn) Ac-THHHHYHGG-NH2 (Hpn) Ac-ACCHDHKKH-NH2 (HspA)
Copper and nickel binding to fragments
of Helicobacter pylori proteins

4 6 8 10
0
20
40
60
80
100
free Ni2+
Ni2+
-MAHHEEQHG-NH2
Ni2+
-MAHHEQQQQQQA-NH2
% f
orm
ation r
ela
tive t
o N
i2+
pH
D. Witkowska, S.Bielińska, W.Kamysz and H.Kozlowski, J. Inorg. Biochem., 2011, 105, 2, 208-214
The distribution of Ni2+ between MAHHEEQHG-NH2
and MAHHEQQQQQQA-NH2
(0.5:1:1) The N-terminal fragment of Hpn-like (MAHHEQQQQQQA-NH2) is much more effective in binding both Cu2+ and Ni2+ ions than the N-terminal fragment of Hpn

MAHHE-NH2...M2+
MAHHEEQHG-NH2...M2+
MAHHEQHQA-NH2...M2+
MAHHEQQQQQQA-NH2...M2+
Cu2+
MAHHEQQQQQQA-NH2...Cu2+
Common M2+ binding
pattern
Ni2+
MAHHEEQHG-NH2...Ni2+
Method: full DFT
(M052X/6-31G(d,p)
Full optimization of the
structure of the complex
(M=Cu, Ni)
N.M. Chiera, M. Rowinska-Zyrek, R. Wieczorek, M. Remelli, H. Kozlowski, Metallomics, 5, 214-221, (2013)

MAHHEEQHG-NH2...Ni2+
Ni2+
MAHHEQQQQQQA-NH2...Ni2+
Ni2+
Method: DFT (M052X/6-31G(d,p) Gln-rich
fragment influence on peptide folding
N.M. Chiera, M. Rowinska-Zyrek, R. Wieczorek, M. Remelli, H. Kozlowski, Metallomics, 5, 214-221, (2013)

CONCLUSIONS
Cys-Xaa-Yaa----Cys sites are much more efficient binding sites for Zn(II) when compared to Ni(II)
Poly-Gln sequences may stabilize the metal ion binding to peptide by organizing specific structure around the binding site with hydrogen bond network of side-chain amide groups.
Cys-Cys is a basic and very specific binding site for Ni(II) ions as that of albumin but much more efficient
His-rich complexes are more stable only at below–physiological pH
The thermodynamic stability of Bi(III) complexes is several orders of magnitude higher than those of Ni(II). Bi(III) is able to displace Ni(II) from its binding sites.

Overall binding process of hexahistidine to Ni2+-NTA at moderate concentration. Ni2+
is immobilized to the surface of the sensor chip via NTA. One imidazole group of the hexahis-tag (a) makes first contact with Ni2+ (b). After the monovalent interaction is established the high local concentratiom of his ligand facilitates the interaction with a second imidazole forming a divalent complex with histidine from the i and i+2 position (c). By consecutive dissociation and reassociation (d, e) the molecule ‘slides’ over the chip surface, which explains the observed rebinding effect in the SPR measurments. Finally, the hexahis dissociates from the Ni2+ (f, g).
S. Knecht, D. Ricklin, A. N. Eberle and B. Ernst, J. Mol. Recognit., 2009, 22(4), 270-279.

The image of the His-tag motifs after energy minimization calculations. (a) His-tag2; (b) His-tag3; (c) His-tag4; (d) His-tag5; and (e) His-tag6. Hsuan-Liang Liu, Yih Ho, Chia-Ming Hsu, J. Biomol. Struct. Dynam., 2003, 21(1), 31-40.

Important issues:
There is a relationship between the number of residues in the His-tag and their properties of binding
A bivalent ligand formed by two His at positions n+2 and n and n+5 in hexapeptide are better than the other possible divalent ligands (n+1, n+3, n+4)
The elongation His-tag for example to seven or ten histidines in not profitable to metal ion binding.
Binding affinity KA (1/KD) of different oligohistidines to Ni2+-NTA chips in surface
plasmon resonance experiment.
S. Knecht, D. Ricklin, A. N. Eberle and B. Ernst, J. Mol. Recognit., 2009, 22(4), 270-279

Ac-HHHHHH-NH2 – His-Tag uses in ITC
Ac-EDDHHHHHHHHHG-NH2 – sequence
occurs in nature (from venom glands of the snake)

Ac-HHHHHH-NH2 – log b: 11,61
Ac-EDDHHHHHHHHHG-NH2 – log b: 16,82
We do not know why ???

2 3 4 5 6 7 8 9 10 11
0
20
40
60
80
100
Cu2+
- Ac-HHHHHH-NH2
free Cu2+
Cu2+
- Ac-EDDHHHHHHHHHG-NH2
% fo
rma
tion
re
lativ
e to
[C
u2
+]
pH

2 3 4 5 6 7 8 9 10 11 12
0
20
40
60
80
100free Cu
2+
Cu2+
- Ac-THHHHAHGG-NH2
Cu2+
- Ac-HHHHHH-NH2
% fo
rma
tion
re
lativ
e to
[C
u2
+]
pH

2 3 4 5 6 7 8 9 10 11 12
0
20
40
60
80
100free Cu
2+
Cu2+
- Ac-PVHTGHMGHIGHTGHTGHTGSSGHG-NH2
Cu2+
- Ac-HHHHHH-NH2
% fo
rma
tio
n r
ela
tive
to
[C
u2
+]
pH

2 3 4 5 6 7 8 9 10 11
0
20
40
60
80
100
free Cu2+
Cu2+
- EDDHHHHHHHHHGVGGGGGGGGGG-NH2
Cu2+
- Ac-EDDHHHHHHHHHG-NH2
% fo
rma
tio
n r
ela
tive to
[C
u2
+]
pH

We do not know exactly which histidine residues are bound.
WHY POLY-HIS SEQUENCE IS SO EFFECTIVE IN METAL ION BINDING BINDING?
IT IS VERY A GOOD QUESTION

He
Li Be B Ne
Al Ar
Sc Ti Cr Ga Ge As Br Kr
Rb Sr Y Zr Nb Ru Rh Pd Ag Cd In Sb Te Xe
Cs Ba La Hf Ta W Re Os Ir Pt Au Hg Tl Pb Bi
La Ce Pr Nd Sm Eu Gd Tb Dy Ho Er Tm Yb Lu
C
Pt Au
Cr V
Ca Se
Li
99mTc
Fe Mg
Cu
Gd
I
F
Zn
Ag
Al
67Ga
B
N2O
N O
S
Ba
Na
K
A Periodic Table of Medicines
P
Sb
Bi
Mn
Mo
Co Si
Sr Sn
Ti
133Xe 201Tl
As 90Y
188Re 153Sm
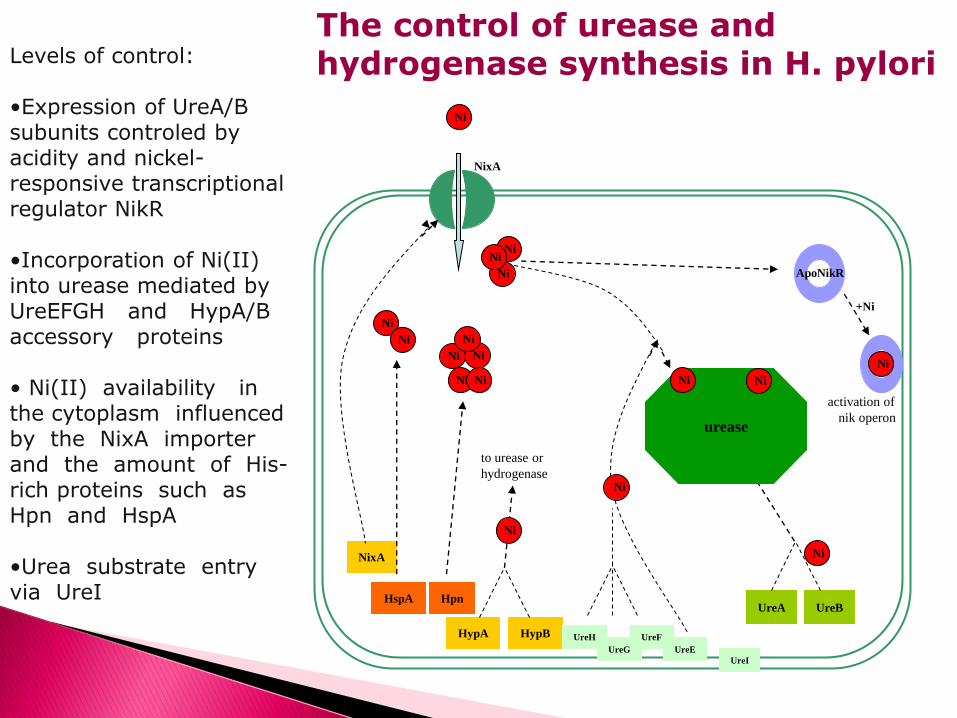
The control of urease and hydrogenase synthesis in H. pylori
Ni
Ni
Ni Ni
NixA
HspA Hpn
HypA HypB UreH
UreG
UreF
UreE
UreI
UreA UreB
Ni
Ni
Ni
Ni
Ni
Ni
Ni
ApoNikR
Ni
urease
Ni
+Ni
Ni
Ni
NixA
to urease or
hydrogenase
Ni
Ni
activation of
nik operon
Levels of control: •Expression of UreA/B subunits controled by acidity and nickel-responsive transcriptional regulator NikR •Incorporation of Ni(II) into urease mediated by UreEFGH and HypA/B accessory proteins • Ni(II) availability in the cytoplasm influenced by the NixA importer and the amount of His-rich proteins such as Hpn and HspA •Urea substrate entry via UreI

4 6 8 10
0
20
40
60
80
100%
fo
rma
tio
n r
ela
tive
to
[C
u2
+]
pH
free Cu2+
DDD
ADSG
ADSGEGDFLAEGGGVR
EDDHHHHHHHHHGVGGGGGGGGGG-NH2


















