
Formation of net-like structures Ho to make long branched filaments? (please switch to the full...
21
Formation of net-like structures Ho to make long branched filaments? (please switch to the full presentation mode to see the animated simulations) In the following a model is described that was already published in 1976: Meinhardt, H., Morphogenesis of lines and nets. Differentiation 6, 117-123; see also: Models of Biological Pattern Formation (1982; Academic Press); three nerve cells of a fly’s brain embryonic blood vessels of a chick trachea of a beetle skeleton of an aspen leaf
-
Upload
samuel-parker -
Category
Documents
-
view
216 -
download
3
Transcript of Formation of net-like structures Ho to make long branched filaments? (please switch to the full...

Formation of net-like structuresHo to make long branched filaments?
(please switch to the full presentation mode to see the animated simulations)
In the following a model is described that was already published in 1976: Meinhardt, H., Morphogenesis of lines and nets. Differentiation 6, 117-123; see also: Models of Biological Pattern Formation (1982; Academic Press); (PDF’s of both are available on our website)
three nerve cells of a fly’s brain embryonic blood vessels of a chick trachea of a beetle skeleton of an aspen leaf

Formation of netlike structures
Differentiation 6, 117-123 (1976)
Net-like Structures are common and essential for all higher organisms. The veins of leaves, blood vessels, tracheae, lung branching or growing axons are examples. In this 1976-proposed model it is assumed that a tip signal is generated by local self-enhancement and long-ranging inhibition. It causes the elongation of the filament and thus its own shift. Long filaments are generated in this way. The elongation occurs towards a region of a higher level of an attractant that can be uniformly produced in the field. For blood vessels, it has turned out that VEGF works as attractor and that Delta-Notch is responsible for specifying the tip region.

Formation of netlike structures:traces behind a shifting signal
Differentiation 6, 117-123 (1976)
The tip-signal can be also used to produce pseudopods that cause an extension of the cell into the correct direction (see the model for the chemotactic orientation of cells) or to orient cell division of the filament-forming cells. Crucial is that the tip signal causes local elongation of the filament and enforces in this way its own shift.

Formation of a netlike structure:a trace behind a shifting signal
Differentiation 6, 117-123 (1976)
Assumed is an activator - inhibitor system. A high activator concentration leads to the differentiation of the exposed cell . Differentiated cells remove a substance such as VEGF or auxin from the surrounding cells. Since the activator production depends on this substrate, the activator maximum becomes quenched in newly differentiated cells and shifts toward a region of higher substrate concentration. Long filaments of differentiated cells are formed behind wandering activator maxima. If the moving tips of the filaments become sufficiently remote from each other and enough space is available, a baseline activator production in the differentiated cells can trigger new maxima – the initiation of branches.
In detail:
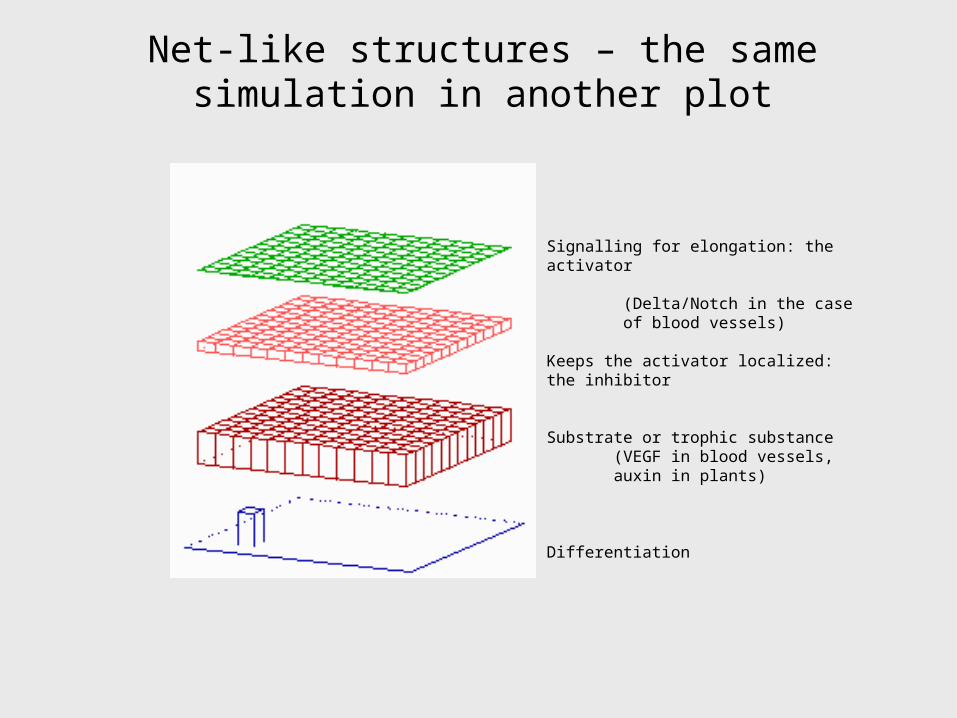
Net-like structures – the same simulation in another plot
Signalling for elongation: the activator
(Delta/Notch in the case of blood vessels)
Keeps the activator localized: the inhibitor
Substrate or trophic substance (VEGF in blood vessels, auxin in plants)
Differentiation

Regeneration of a net-like structure
Differentiation 6, 117-123 (1976)
After removal of some veins, e.g., by an injury, substrate accumulates in the deprived region. This attracts new veins. The resulting pattern is similar but not identical

Regeneration of a net-like structure
Experiments of Wigglesworth, Quart. J. micr. Sci. 95, 115-137 (1954)
Example: after cutting tracheae in one segment, branches from the nearby segment grow over the segment border into the deprived segment to maintain the oxygen supply

Control of vessel density
Regulation of the density of a net. Tumors attract new blood vessels and cause extensive sprouting. A piece of tumor tissue grafted into the cornea of a rabbit cause a massive invasion of blood vessels (redrawn after Folkman, 1976). A piece of cartilage - a tissue that repels blood vessels - largely suppresses such invasion when grafted in front of the tumor. Left: in the model an increase of level of trophic factor (green; e.g., Vegetal Endodermal Growth Factor, VEGF) in the upper half leads to higher rate of branching. The influence of the cartilage is simulated by an increased of a baseline inhibitor production in the center of the field (shaded). Veins preferentially circumvent this area This is somewhat compensated by the increased level of the attractant in the upper central region.
more ‘VEGF’
higher baseline inhibition
Experiments by J. Folkman (1976; this drawing is taken from: Models of Biological Pattern Formation (1982; Academic Press), PDF on our website

Tiling or self-avoidance of growing filaments
Frequently filaments of the same type try to keep distance from each other, making sure that a balanced coverage of an area is achieved. For instance, branches of similar neurons show such a self-avoidance. This is a straightforward property of the model since filaments elongation would not occur towards a region in which the level of the attractant is already lowered due to the drainage by other filament.

Connections of two nets
If two net-forming structures (e.g. veins and arteries) exists, one or both system may attract the moving tips of the other. Assumption in this simulation: each of the two systems have their own attractive factor (green or grey) and each system enhances somewhat production of other attractants. Result: both nets makes connections.
Differentiation 6, 117-123 (1976)Models of Biological Pattern Formation (1982; Academic Press)(available on our website)

Path-finding towards a target region
If the attractant is not uniformly produced but an elevated production occurs in a special source region, a directed extension of a filament towards the source region will occur. Branching occurs preferentially in this target area (as frequently observed with growing axons in the target region)

The components assumed and the equations
2
a a a a
a scar a D b d
t b
2
b b b
bsca r b D b b d
t
c c c c
cb r c c c d D c
t
2
1d
d dd
r ddr d b a
t s d
1. A signal that determines at which position the filament should be elongated or where to initiate a new branch (e.g., an activator-inhibitor
system; Delta/Notch in the case of blood vessels)
2. The elongation of the filaments depends on a trophic factor; it is removed by the filaments (auxin, NGF, VEGF); elongation goes up-hill
3. An irreversible determination that makes the filament cells different from the remaining cells
Differentiation 6, 117-123 (1976)Models of Biological Pattern Formation (1982; Academic Press)(available on our website)

Bifurcations at the tips
In many cases new branches are formed by a split of a growing tip. What mechanism allows peak splitting at the tips? The problem is non-trivial: to localize a peak, lateral inhibition is required. How a peak can split nevertheless?
According to our model, peak splitting can occur in growing systems if the self-enhancement saturates. With increasing distances of the maxima at the tips, the inhibitor can escape into a larger area (or more trophic factor can be obtained from the larger interstices), which leads to an increase in the activation. If the activation saturates, the peak will become broader, not higher. After a certain field size is surpassed, the activation in the center becomes instable since this area has more problems to get rid of the inhibitor or to obtain sufficient trophic factor. The central activation collapses in favor of the flanking regions. The two peaks shift to obtain a greater distance for escaping the inhibitory influence resulting from the other. Saturation facilitates such shifts.

Branch formation by peak splitting at the growing tip

Evolutionary earlier leaves such as the Gingko branching occurs only by splitting, i.e., no lateral
branching of existing veins occur
Even the initial triple branching is reproduced

The problem of closed loops
In contemporary leaves connection between secondary veins are the rule. In terms of the model, this is an open problem. Veins are primarily elongated in directions that are free of veins. Closed loops appear very early in the leaf development. For an updated model, the auxin transport has to be incorporated.

The problem of closed loops
Closed loops are frequent also in other developmental systems.
There are also other mechanism that leads to closed loops …

The formation of stripes
2
2....
(1 )
a a
t h a
The elements of net-like structures have a long extension in one dimension and a short in the other, i.e., these elements are stripes - a frequent pattern developmental biology In terms of the model, stripe formation is possible if the autocatalysis saturates at high activator concentration. Then the inhibitor production is limited too and the mutual competition between neighbouring cells is reduced. Due to the saturation more cells remain activated although at a lower level. Thus, activated cells have to tolerate activated cells in their neighbourhood, independent of the range of inhibition. A further condition is a modest diffusion of the activator: activated regions occur in larger coherent patches since activated cells tend to activate adjacent cells. On the other hand, pattern formation requires that activated cells are close to non-activated cells into which the inhibitor can diffuse. These two seemingly contradictory features, large coherent patches and proximity of non-activated cells, are characteristic for stripe-like patterns. If initiated by random fluctuations, the stripes have random orientations too....

Netlike structures can emerge as stripes around inhibitory centers

Closed loops: stripe formation around inhibitory centers

Closed loops: components and equations
a, b: Activator (blue) - substrate system, marks the center
c, d: An activator (red) - inhibitor system, stripes forms at the highest b levels, i.e., at the largest distance from the centers
Koch, A.J. and Meinhardt, H. (1994). Biological pattern-formation - from basic mechanisms to complex structures. Rev. Modern Physics 66, 1481-1507.


















