
Form-deprivation myopia induces activation of scleral matrix ...
16
Form-Deprivation Myopia Induces Activation of Scleral Matrix Metalloproteinase-2 in Tree Shrew Jeremy A. Guggenheim and Neville A. McBrien Purpose. To investigate whether structural changes to the sclera during form-deprivation myo- pia are caused by active tissue remodeling, the gelatinase activity of tree shrew scleras was studied in normal animals, form-vision deprived animals, and animals recovering from myopia. Methods. Infant tree shrews were monocularly deprived (MD) of form vision with translucent occluders for 5 days. Recovery animals were allowed 3 days of binocularly unoccluded vision after the period of form deprivation. Eyes were removed and dissected to provide scleral samples corresponding to equatorial and posterior regions. Gelatinase activity was assessed by quantitative sodium dodecyl sulfate-polyacrylamide gel electrophoresis or SDS-PAGE, gelatin, zymography of scleral matrix metalloproteinase (MMP) extracts. Results. The major gelatinolytic species present in tree shrew sclera was found to be MMP-2 (gela- tinase A). In normal (nondeprived) animals, most of the MMP-2 was found to be in the latent form (the ratio of active-to-latent MMP-2 was 0.23 ± 0.05 and 0.34 ± 0.06 in equatorial and posterior samples, respectively; n = 10 eyes from five animals). After 5 days of MD, there was a threefold increase in the amount of active scleral MMP-2 in myopic eyes compared to contralateral control eyes, whereas latent MMP-2 activity levels were not altered significantly. This increase in active MMP- 2 was seen in both the equatorial and posterior sclera of myopic eyes (active-to-latent MMP-2 ratios were 0.53 ± 0.10 and 0.81 ± 0.09 in equatorial and posterior regions, respectively; n = 6 animals). Contralateral control eyes had levels of both active and latent MMP-2 not significantly different from normal eyes. After only 3 days of unoccluded vision, previously deprived eyes that were now recovering from myopia had a fivefold lower level of active MMP-2 than that seen in deprived eyes after 5 days of MD. In fact, active (and latent) MMP-2 levels were reduced in recovering eyes even below the levels found in their contralateral control eyes. Active-to-latent ratios for recovering eyes were 0.11 ± 0.03 and 0.15 ± 0.03 in equatorial and posterior sclera, respectively (rc = 5 animals). Conclusions. These results demonstrate that form-deprivation myopia and recovery from myo- pia alter scleral catabolism and provide further support for the theory that changes in eye size during mammalian refractive development are the result of active tissue remodeling rather than passive scleral stretching alone. Invest Ophthalmol Vis Sci. 1996; 37:1380-1395. JL he prevalence of myopia in the adult populations of western Europe and the United States is approxi- mately 25% to 30%. l>2 In a significant minority of per- sons with myopia, the degree of myopia is sufficiently high to result in visual loss from ocular pathology asso- From the Department of Optomelry and Vision Sciences, University of Wales, Cardiff, United Kingdoin. Presented in part at the 1995 meeting of the Association for Research in Vision and Ophthalmology, Port iMiiderdale, Florida. Supported by Wellcome Trust grant 037531 (NAM) Submitted for publication August 24, 1995; revised February 28, 1996; accepted February 29, 1996. Proprietary interest category: N. Reprint requests: Neville A. McBrien, Department of Optometry and Vision Sciences, University of Wales, Cardiff, P.O. Box 905, Cardiff CF1 3XF, Wales, United Kingdom. ciated with the excessive elongation of the eye. Patho- logic consequences of high myopia, such as retinal detachment and Fuch's spot, mean that myopia is the fourth or fifth leading cause of blindness in developed countries. 3 ' 4 High myopia in humans is associated with enlargement of the globe in all meridians, but princi- pally in the axial dimension. Commonly associated with this axial elongation of the myopic human eye is a marked thinning of the posterior sclera 5 and a reduction in scleral collagen fibril diameter. 6 ' 7 Based on these findings, it has been proposed that the ocular enlargement found in high myopia could be the result of scleral stretching. 8 Further support for this essen- tially "passive scleral stretch" hypothesis of myopic 1380 Investigative Ophthalmology & Visual Science, June 1996, Vol. 37, No. 7 Copyright © Association for Research in Vision and Ophthalmology Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933417/ on 03/24/2018
-
Upload
truongthien -
Category
Documents
-
view
223 -
download
1
Transcript of Form-deprivation myopia induces activation of scleral matrix ...

Form-Deprivation Myopia Induces Activation of ScleralMatrix Metalloproteinase-2 in Tree Shrew
Jeremy A. Guggenheim and Neville A. McBrien
Purpose. To investigate whether structural changes to the sclera during form-deprivation myo-pia are caused by active tissue remodeling, the gelatinase activity of tree shrew scleras wasstudied in normal animals, form-vision deprived animals, and animals recovering from myopia.
Methods. Infant tree shrews were monocularly deprived (MD) of form vision with translucentoccluders for 5 days. Recovery animals were allowed 3 days of binocularly unoccluded visionafter the period of form deprivation. Eyes were removed and dissected to provide scleralsamples corresponding to equatorial and posterior regions. Gelatinase activity was assessedby quantitative sodium dodecyl sulfate-polyacrylamide gel electrophoresis or SDS-PAGE,gelatin, zymography of scleral matrix metalloproteinase (MMP) extracts.
Results. The major gelatinolytic species present in tree shrew sclera was found to be MMP-2 (gela-tinase A). In normal (nondeprived) animals, most of the MMP-2 was found to be in the latentform (the ratio of active-to-latent MMP-2 was 0.23 ± 0.05 and 0.34 ± 0.06 in equatorial and posteriorsamples, respectively; n = 10 eyes from five animals). After 5 days of MD, there was a threefoldincrease in the amount of active scleral MMP-2 in myopic eyes compared to contralateral controleyes, whereas latent MMP-2 activity levels were not altered significantly. This increase in active MMP-2 was seen in both the equatorial and posterior sclera of myopic eyes (active-to-latent MMP-2 ratioswere 0.53 ± 0.10 and 0.81 ± 0.09 in equatorial and posterior regions, respectively; n = 6 animals).Contralateral control eyes had levels of both active and latent MMP-2 not significantly differentfrom normal eyes. After only 3 days of unoccluded vision, previously deprived eyes that were nowrecovering from myopia had a fivefold lower level of active MMP-2 than that seen in deprived eyesafter 5 days of MD. In fact, active (and latent) MMP-2 levels were reduced in recovering eyes evenbelow the levels found in their contralateral control eyes. Active-to-latent ratios for recovering eyeswere 0.11 ± 0.03 and 0.15 ± 0.03 in equatorial and posterior sclera, respectively (rc = 5 animals).
Conclusions. These results demonstrate that form-deprivation myopia and recovery from myo-pia alter scleral catabolism and provide further support for the theory that changes in eyesize during mammalian refractive development are the result of active tissue remodelingrather than passive scleral stretching alone. Invest Ophthalmol Vis Sci. 1996; 37:1380-1395.
JL he prevalence of myopia in the adult populationsof western Europe and the United States is approxi-mately 25% to 30%.l>2 In a significant minority of per-sons with myopia, the degree of myopia is sufficientlyhigh to result in visual loss from ocular pathology asso-
From the Department of Optomelry and Vision Sciences, University of Wales,Cardiff, United Kingdoin.Presented in part at the 1995 meeting of the Association for Research in Vision andOphthalmology, Port iMiiderdale, Florida.Supported by Wellcome Trust grant 037531 (NAM)Submitted for publication August 24, 1995; revised February 28, 1996; acceptedFebruary 29, 1996.Proprietary interest category: N.Reprint requests: Neville A. McBrien, Department of Optometry and Vision Sciences,University of Wales, Cardiff, P.O. Box 905, Cardiff CF1 3XF, Wales, UnitedKingdom.
ciated with the excessive elongation of the eye. Patho-logic consequences of high myopia, such as retinaldetachment and Fuch's spot, mean that myopia is thefourth or fifth leading cause of blindness in developedcountries.3'4 High myopia in humans is associated withenlargement of the globe in all meridians, but princi-pally in the axial dimension. Commonly associatedwith this axial elongation of the myopic human eyeis a marked thinning of the posterior sclera5 and areduction in scleral collagen fibril diameter.6'7 Basedon these findings, it has been proposed that the ocularenlargement found in high myopia could be the resultof scleral stretching.8 Further support for this essen-tially "passive scleral stretch" hypothesis of myopic
1380Investigative Ophthalmology & Visual Science, June 1996, Vol. 37, No. 7Copyright © Association for Research in Vision and Ophthalmology
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933417/ on 03/24/2018

Form-Deprivation Myopia Induces Activation of MMP-2 1381
progression comes from observations of embryonicand neonatal eye growth under extremes of intraocu-lar pressure. If the intraocular pressure of the embry-onic chick eye is reduced artificially, normal eye en-largement does not occur.910 Conversely, in humanneonates suffering from buphthalmos, elevated intra-ocular pressure typically leads to ocular enlarge-ment."
In a small minority of human subjects, high myo-pia may be found as part of a syndrome that resultsfrom a known single-gene defect.12"15 In the defectsreported to date, the disease locus has been mappedto a gene encoding a structural component of theextracellular matrix (ECM), such as collagen or elas-tin, or to an enzyme involved in the biosynthesis ofsuch structural components. These defects may predis-pose affected persons to the development of myopiaby changing the biomechanical properties of thesclera, choroid, or vitreous. Perhaps the simplest the-ory to explain the development of myopia in suchconditions is that each gene defect produces a weak-ened sclera, which then stretches excessively underphysiological intraocular pressure. This theory is sup-ported by evidence that collagen gene defects can af-fect profoundly the biomechanical properties of thetissues in which they are expressed"5 and that the bio-mechanical properties of the sclera are affected inhuman high myopia.17
The excessive axial elongation, posterior scleralthinning, and reduction in collagen fibril diameternoted in human high myopia are also evident in mam-malian models of axial myopia (monkey1819 and treeshrew20'21). However, recent findings from animalmodels of myopia question a passive scleral stretchhypothesis and give support to the idea of active scleraltissue remodeling. It has been found that the thinningof the posterior sclera in highly myopic tree shreweyes is not simply caused by a passive stretching of thesclera to cover the enlarged eye because there is asignificant loss of scleral tissue (dry weight) in myopiceyes, particularly at the posterior pole.22 In the chickmodel of myopia, increases in the synthesis of scleralprotein, proteoglycan, and DNA have been reportedin eyes developing myopia.23'24 In both chick and treeshrew, it has been found that eyes are able to recoverfrom induced axial myopia when form deprivationceases.25'26 These eyes are able to slow or stop their rateof elongation, relative to contralateral control eyes,during the recovery period. This recovery effect can-not be accounted for solely by changes in choroidalthickness27 and does not appear to be related tochanges in intraocular pressure.28 In addition, inchick29'30 and tree shrew,31 when myopia is inducedby producing a defocused retinal image with negativelenses, once the eye has compensated for the defocus
(by reaching a functional emmetropia), it arrests itsincreased rate of elongation to maintain a clear image.Findings from recovery experiments and lens-induceddefocus experiments demonstrate that eye enlarge-ment in these animal models of axial myopia can bemodulated. This argues against a model in which pas-sive scleral stretch is the sole causative mechanism ofmyopia development because passive scleral stretchwould predict gradual and continuous axial elonga-tion that is incapable of such fine control.
Active remodeling of connective tissues is a com-plex process. Because extracellular matrices providestructural strength and form to organs, remodelingrequires a delicate balance between degradation andsynthesis. The matrix metalloproteinases (MMPs)have been implicated in the degradation of many con-nective tissues during development and pathology.MMPs are zinc-containing, Ca++-dependent proteo-lytic enzymes capable of degrading most ECM compo-nents. There are at least 10 known members of theMMP family (for reviews, see refs. 32 and 33), eachof which has a separate, though usually overlapping,substrate specificity. All MMPs are secreted as inactiveprecursors (zymogens, pro-enzymes) activated bycleavage of an inhibitory domain, with an accompa-nying reduction in molecular weight.
To search for evidence that eye enlargement dur-ing the development of form-deprivation myopia re-sults from active tissue remodeling, the current studymonitored the levels of scleral gelatinases in the eyesof normal, monocularly deprived (MD), and recov-ering tree shrews.
MATERIALS AND METHODS
Animals
Tree shrews (Tupaia belangeri) were reared in ourbreeding colony on a 15-hour light-9-hour dark cycle.Food and water were available ad libitum. Animalswere assigned randomly to one of three groups: nor-mal, MD, or recovery. On the 15th day after eye open-ing, all animals were removed from the maternal cageand housed in individual cages. Monocularly deprivedand recovery group tree shrews were given anesthesiaand were fitted with a head-mounted goggle con-taining a translucent occluder, essentially as de-scribed.34 The eye to be occluded was assigned on arandom basis.
Optical and Structural Measures
On the 20th day after eye opening, animals from allgroups were anesthetized, and optical and structuralmeasures were taken as previously described (CottriallCL, McBrien NA, p. 1368 this issue). Briefly, anterior
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933417/ on 03/24/2018

1382 Investigative Ophthalmology & Visual Science, June 1996, Vol. 37, No. 7
corneal curvature was measured by a modified kerato-meter in the horizontal and vertical meridians. In 5-day MD and normal animals, cycloplegic (atropine1%) ocular refraction was measured by streak retinos-copy to the nearest 0.5 D, and results were reportedas equivalent spherical correction at the corneal apex.In recovery animals, atropine cycloplegia was not per-formed before ocular measures to avoid long-term ef-fects on recovery. Therefore, in recovery animals, pu-pil dilation was achieved by instillation of two 15-fAdrops of phenylephrine 2%. Axial ocular dimensionswere measured by A-scan ultrasonography (10-MHzfocused transducer; Lecroy [Geneva, Switzerland] dig-ital storage oscilloscope). On completion of the opti-cal measures, MD and normal animals were deeplyanesthetized (80 mg/kg sodium pentobarbital), andthe eyes were enucleated. Recovery animals were per-mitted 3 days of binocularly unoccluded visual experi-ence, optical measures were repeated (phenylephrinemydriasis), and eyes were enucleated.
Enucleated eyes were trimmed of extraneous or-bital tissue and weighed, and equatorial dimensionswere measured with digital calipers. The cornea andan accompanying 1.5-mm rim of anterior sclera wereremoved, as was the lens. A posterior pole button wasexcised with a 7-mm diameter trephine. This buttonand the residual equatorial ring of sclera were cleanedof vitreous and of retinal and choroidal tissue andthen frozen in liquid nitrogen. Tissue was stored at—80°C until use. Zymographic analysis was performed"blind," without knowledge of whether tissue was de-rived from treated or control eyes. All animals weretreated in accordance with the ARVO Statement forthe use of Animals in Ophthalmic and Vision Re-search.
Choice of Extraction Procedure
We found the choice of MMP extraction procedureto be crucial to the success of this investigation. Inpreliminary experiments, with an extraction tech-nique involving repeated rounds of freezing and thaw-ing in the presence of a nonionic detergent,35 wefound a considerable variation in the scleral gelatinaseactivity between individual, and even between the twoeyes of, normal tree shrews.36 A subsequent compari-son with other extraction techniques demonstratedthat the freeze-thaw method was highly inefficient forscleral tissue. We chose an improved extraction proce-dure that used sodium dodecyl sulfate (SDS) in con-junction with a calcium chelating agent (EGTA). Therationale for the choice of this extraction method wasthat SDS would partially denature matrix proteins anddissociate MMP-tissue inhibitor of metalloproteinases(TIMP) complexes.37 Chelating agents were used tosequester extracellular calcium, which may have al-
tered the conformation of the calcium-dependentMMPs or prevented any calcium-dependent attach-ment to their substrate. Although both EDTA andEGTA were found to be equally effective extractionagents (appearing to act synergistically with SDS, atthe concentrations tested; Fig. 1A), EGTA was chosenbecause it chelates zinc ions to a lesser extent thanEDTA (by approximately four orders of magnitude38).Further advantages to the use of SDS and EGTA werethat they both inhibit conversion of latent MMPs totheir active forms (Fig. IB).3'1'40 This ensured that thelevels of latent and active MMPs measured by zymogra-phy represented, as closely as possible, those presentin the tissue.
Extraction of Metalloproteinases
Scleral samples were freeze-dried, and their dryweights were recorded in triplicate (dry weights ofboth posterior and equatorial samples approximately2 mg each; weighing accuracy ± 10 //g). Each samplewas rehydrated for 3 minutes at room temperatureby submersion in a 50-^tl droplet of water (18 MQconductivity). The tissue was then snap frozen in liq-uid nitrogen and powdered in a freezer mill (Mikrodi-smembrator; B. Braun, Melsungen GmbH, Germany).The powder was recovered as a suspension in a totalvolume of 950 /A water (preliminary experiments dem-onstrated that the recovery of tissue was >95%). Analiquot of extraction solution (2% wt/vol SDS; 200 mMEGTA; 100 mM Tris-HCl, pH 7.5) was added to givea final concentration of 0.1% SDS and 10 mM EGTA.The solution was vortexed, and then the extractionwas allowed to proceed for 20 minutes at 37°C. Insolu-ble material was pelleted by centrifugation (13,000rpm X 5 minutes, at room temperature) and the su-pernatant, containing the extracted MMPs, was re-moved and freeze-dried.
MMP-2 Standards
Primary cultures of tree shrew skin fibroblasts weregrown in Dulbecco's modified Eagle's medium with10% fetal calf serum until confluent. The cells werewashed in serum-free medium and then incubated for24 hours with serum-free medium containing 100 ng/ml phorbol 12-myristate 13-acetate to stimulate MMPexpression. The conditioned medium was collected,concentrated 50-fold over a 10 kDa-cut ultrafiltrationmembrane, divided into aliquots, and stored at —80°C.Zymographic analysis showed that MMP-2 accountedfor >95% of the gelatinolytic activity of the condi-tioned medium; the activity of the active and latentforms was approximately equal (as judged by scanningdensitometry). A 1:128,000 dilution of the condi-tioned medium standard was the minimum amountthat could be detected with our zymography tech-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933417/ on 03/24/2018
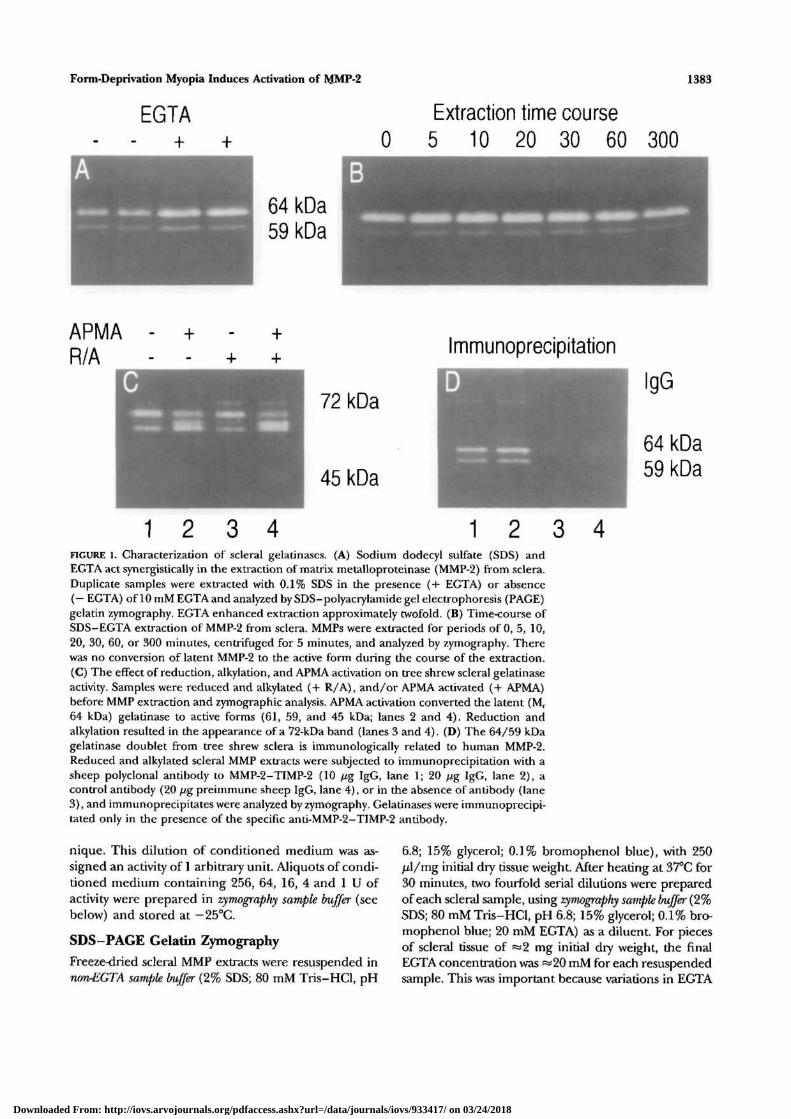
Form-Deprivation Myopia Induces Activation of MMP-2 1383
EGTA0
Extraction time course5 10 20 30 60
64kDa59kDa
APMAR/A Immunoprecipitation
72kDa
45kDa
1 2 3 4FIGURE 1. Characterization of scleral gelatinases. (A) Sodium dodecyl sulfate (SDS) andEGTA act synergistically in the extraction of matrix metalloproteinase (MMP-2) from sclera.Duplicate samples were extracted with 0.1% SDS in the presence (+ EGTA) or absence(— EGTA) of 10 mM EGTA and analyzed by SDS-polyacrylamide gel electrophoresis (PAGE)gelatin zymography. EGTA enhanced extraction approximately twofold. (B) Time-course ofSDS-EGTA extraction of MMP-2 from sclera. MMPs were extracted for periods of 0, 5, 10,20, 30, 60, or 300 minutes, centrifuged for 5 minutes, and analyzed by zymography. Therewas no conversion of latent MMP-2 to the active form during the course of the extraction.(C) The effect of reduction, alkylation, and APMA activation on tree shrew scleral gelatinaseactivity. Samples were reduced and alkylated (+ R/A), and/or APMA activated (+ APMA)before MMP extraction and zymographic analysis. APMA activation converted the latent (Mr
64 kDa) gelatinase to active forms (61, 59, and 45 kDa; lanes 2 and 4). Reduction andalkylation resulted in the appearance of a 72-kDa band (lanes 3 and 4). (D) The 64/59 kDagelatinase doublet from tree shrew sclera is immunologically related to human MMP-2.Reduced and alkylated scleral MMP extracts were subjected to immunoprecipitation with asheep polyclonal antibody to MMP-2-TIMP-2 (10 //g IgG, lane 1; 20 j/g IgG, lane 2), acontrol antibody (20 jug preimmune sheep IgG, lane 4), or in the absence of antibody (lane3), and immunoprecipitates were analyzed by zymography. Gelatinases were immunoprecipi-tated only in the presence of the specific anti-MMP-2-TIMP-2 antibody.
IgG
64 kDa59 kDa
1 2 3 4
nique. This dilution of conditioned medium was as-signed an activity of 1 arbitrary unit. Aliquots of condi-tioned medium containing 256, 64, 16, 4 and 1 U ofactivity were prepared in zymography sample buffer (seebelow) and stored at — 25°C.
SDS-PAGE Gelatin ZymographyFreeze-dried scleral MMP extracts were resuspended innm-EGTA sample buffer (2% SDS; 80 mM Tris-HCl, pH
6.8; 15% glycerol; 0.1% bromophenol blue), with 250/ul/mg initial dry tissue weight. After heating at 37°C for30 minutes, two fourfold serial dilutions were preparedof each scleral sample, using zymography sample buffer (2%SDS; 80 mM Tris-HCl, pH 6.8; 15% glycerol; 0.1% bro-mophenol blue; 20 mM EGTA) as a diluent. For piecesof scleral tissue of « 2 mg initial dry weight, the finalEGTA concentration was «20 mM for each resuspendedsample. This was important because variations in EGTA
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933417/ on 03/24/2018

1384 Investigative Ophthalmology & Visual Science, June 1996, Vol. 37, No. 7
concentration between samples would have producedbands of variable width on the final gel.
Zymography was carried out as described pre-viously3541'42 with slight modifications. Ten-microlitersamples of scleral extract were run on 0.5-mm thick10% SDS-PAGE mini-gels containing 1 mg/ml gela-tin (Sigma G-9382). Conditioned medium MMP-2standards were included on every gel. After electro-phoresis, gels were rinsed at room temperature for 15minutes in 0.03% SDS and then for 45 minutes in2.5% Triton X-100. These steps served to remove resid-ual EGTA from the gel and to renature SDS-denaturedMMPs. Gelatinase activity was initiated by incubationin reaction buffed5 (50 mM Tris-HCl, pH 7.6; 500 mMNaCl; 50 mM CaCl2) and continued for 18 hours at37°C. To visualize regions of gelatin degradation, gelswere stained with Coomassie blue R, destained, in-fused with glycerol, and dried between sheets of cello-phane.
Quantification by Scanning Densitometry
Dried gels were scanned at high resolution (64 (im X64 fim) on a Howtek (Hudson, NH) Scanmaster 3scanner. Images were analyzed with the Quantity Onesoftware package from PDI (Huntington Station, NY)running on a Sun Sparc computer (Sun Microsystems,Mountain View, CA). Zymogram images were "con-trast-inverted" (to give images with dark bands on alight background), and the background was sub-tracted by taking the average pixel density over a 30%area of the gel containing no bands. The pixel densityof individual bands was integrated over a constant areaof gel (3 mm in width and 1 mm in height) analogousto measuring the area under the curve in two dimen-sions. For each of the five MMP-2 standards, this banddensity value was plotted against the known activity oftheir latent band (1 to 256 arbitrary activity units; seeMMP-2 Standards above) to create a standard curve.The MMP-2 activity of samples was calculated from thestandard curve using a point-to-point regime becausestandard curves were only linear from 1 to 64 arbitraryunits. Because both standards and samples were di-luted in fourfold steps, each sample dilution was mea-sured against a separate part of the standard curve,minimizing the effects of gel loading or running arti-facts. The activity of both the latent and the activeMMP-2 bands was measured against the same standardcurve (derived from the latent band of the standards).To account for any slight variations in active-to-latentratios between experimental runs (resulting from vari-ations in the efficiency of the EGTA-removal step),the active MMP-2 activity of samples was normalized.This was achieved by measuring the active-to-latentratio of the MMP-2 standards on each run and nor-malizing the scleral active MMP-2 activities accordingly
(this correction factor averaged 1.007 ± 0.04, mean± SEM).
Characterization of Gelatinases
Normal tree shrew sclera (dry wt « 2 mg) was pow-dered in a freezer mill, as described above in Extrac-tion of Metalloproteinases, and collected as a suspen-sion in 2 ml of water. Five hundred-microliter aliquotsof this suspension were distributed to microcentrifugespin-filter columns (500 fA capacity, 0.22 ^m celluloseacetate membrane; Costar, Cambridge, MA) to facili-tate a series of efficient solution changes. Each samplewas resuspended in 400 /A of 50 mM Tris-HCl (pH7.5), 2 mM CaCl2, and 0.5 mM phenylmethylsulfonylfluoride, and transferred to a 37°C water bath. Twosamples were reduced and alkylated43 (5 mM dithio-threitol, 30 minutes; 20 mM iodoacetamide, 30 min-utes) . One of these samples and one of the untreatedsamples were then activated with p-aminophenyl mer-curic acetate (APMA; 1 mM, 60 minutes). Metallopro-teinases were eluted from the powdered tissue by re-suspension in (200 fA/mg initial dry weight of) EGTA-containing zymography sample buffer, and 10-/zl aliquotswere analyzed by gelatin zymography.
To test whether the 64/59 kDa tree shrew gela-tinase doublet was immunologically related to humanMMP-2, we performed a zymographic analysis of scle-ral MMPs immunoprecipitated by a polyclonal anti-body to human MMP-2-TIMP-244 (generously do-nated by Dr. G. Murphy, Strangeways Research Labo-ratory, Cambridge, UK). This antibody was raised (insheep) against purified MMP-2 and crossreacts withboth MMP-2 and TIMP-2. To prevent immunoprecipi-tation of scleral gelatinase-TIMP complexes by TIMPimmunoreactivity, we reduced and alkylated samples(to remove TIMPs) before immunoprecipitation.Briefly, powdered tree shrew sclera was reduced andalkylated (as described above) and rinsed twice withwater. MMPs were then extracted from the tissue with0.1% SDS + 10 mM EGTA, diluted fourfold in buffer(1% bovine serum albumin, 20 mM Tris-HCl, pH 7.4,150 mM NaCl, 10 mM EGTA, 0.2% Triton X-100),and incubated with protein G-sepharose beads thathad been primed with 20 fig of either sheep anti-MMP-2-TIMP-2 immunoglobulin G (IgG) or control(preimmune) sheep IgG. After a 16-hour incubation,beads were washed extensively, and immunoprecipi-tated proteins were eluted with EGTA-containing zy-mography sample buffer and analyzed by zymography, asdescribed above.
Data Analysis
Latent and active MMP-2 activity data were transferredto a spreadsheet (Excel; Microsoft) for paired samplestatistical analyses (t-test). Analysis of variance (Mini-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933417/ on 03/24/2018

Form-Deprivation Myopia Induces Activation of MMP-2 1385
TABLE l. Optical and Structural Measures
Normals
Age-matched to 5 day MDsRight eyeLeft eyeDifference (right - left)
MDs
After 5 days MDDeprived eyeControl eyeDifference (deprived - control)
Recoveries
After 5 days MDDeprived eyeControl eyeDifference (deprived — control)
5 days MD + 3 days recoveryRecovering eyeControl eyeDifference (recovering — control)
RefractiveStatus (D)
+8.3 ± 0.5+8.2 ± 0.6+0.1 ± 0.2
+ 1.3 ± 0.7+8.7 ± 0.1-7.4 ± 0.7J
+0.8 ± 0.8+8.5 ± 0.1-7.7 ± 0.8J
+5.6 ± 0.7+8.8 ± 0.2-3.1 ± 0.6f
Anterior SegmentDepth (mm)
1.05 ± 0.021.05 ± 0.020.00 ± 0.01
1.09 ± 0.021.06 ± 0.020.03 ± 0.01
1.09 ± 0.011.07 ± 0.010.03 ± 0.02
1.08 ± 0.021.08 ± 0.010.01 ± 0.02
Lens Thickness(mm)
3.19 ± 0.033.18 ± 0.010.01 ± 0.02
3.20 ± 0.013.21 ± 0.02
-0.01 ± 0.01
3.19 ± 0.023.22 ± 0.02
-0.03 ± 0.01
3.23 ± 0.033.24 ± 0.03
-0.01 ± 0.01
Vitreous ChamberDepth (mm)
2.83 ± 0.042.81 ± 0.040.02 ± 0.02
2.94 ± 0.032.79 ± 0.020.15 ± 0.02|
2.91 ± 0.022.76 ± 0.020.15 ± 0.02f
2.84 ± 0.022.77 ± 0.010.07 ± 0.02*
Axial length(mm)
7.07 ± 0.057.04 ± 0.050.03 ± 0.02
7.23 ± 0.067.06 ± 0.050.17 ± 0.02|
7.20 ± 0.057.05 ± 0.030.15 ± 0.02f
7.16 ± 0.057.09 ± 0.040.07 ± 0.03
Values are mean ± SEM; n = 5 for normal and recovery groups; n = 6 for MD group. Significant differences on paired t-test indicatedas *P < 0.05, \P < 0.01, and %P < 0.001.
tab\ Minitab, State College, PA) was used to examinedifferences between groups, with Tukey's HSD testused to assess individual group differences. All resultsare expressed as mean ± SEM. Results were consid-ered statistically significant if P < 0.05.
RESULTS
Refractive Effects of Form Deprivation andRecovery
Five days of monocular deprivation induced —7.4 ±0.7 D of myopia in occluded eyes compared to contra-lateral control eyes (Table 1). This difference in re-fraction was accounted for by an elongation of thevitreous chamber in occluded eyes relative to contra-lateral control eyes. There were no significant differ-ences in corneal curvature, anterior segment depth(corneal thickness + anterior chamber depth) or lensthickness in any of the three groups.
Three days of binocular visual experience (recov-ery) after 5 days of MD reduced the amount of myopiain previously occluded eyes, compared to their contra-lateral controls, by 60% (Table 1). This demonstratedthat the 3-day recovery period was sufficient to allowa significant emmetropization response to occur inrecovering eyes. This reduction in myopia was associ-ated with a significant reduction (P < 0.002) in therelative difference in vitreous chamber depth between
the two eyes. An interesting finding was that the reduc-tion in vitreous chamber difference was achieved pre-dominantly by the myopic eye actually becomingsmaller (see Table 1), an effect that did not appearto be caused by choroidal thickening (unpublishedobservations, 1995). Recently, a similar finding wasreported,45 and a more detailed investigation of thiseffect will be presented in a separate communication.
Characterization of Scleral Gelatinases
Zymographic analysis of MMP extracts from tree shrewsclera demonstrated the presence of two major gela-tinases that migrated as a doublet of 64 and 59 kDa(Fig. 1). This doublet accounted for >95% of scleralgelatinase activity. Highly concentrated scleral extractsamples consistently demonstrated the presence ofother minor gelatinases (a »90 kDa form, which mayhave been MMP-9, and a «28 kDa form, which mayhave been the gelatinolytically active cleavage productof a larger MMP). All gelatinase activity could be inhib-ited by replacing the Ca++ in the zymography reactionbuffer with EDTA (not shown), but activity was notinhibited by the irreversible serine protease inhibitorphenylmethylsulfonyl fluoride (Fig. 1C).
Activation of scleral samples with APMA convertedsome of the 64 kDa gelatinase to the 59-kDa form,suggesting that the 64-kDa and 59-kDa bands repre-sented the latent and active forms, respectively, of a
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933417/ on 03/24/2018

1386 Investigative Ophthalmology & Visual Science, June 1996, Vol. 37, No. 7
single metalloproteinase (Fig. 1C, lanes 2, 4). Thisactivation process was accompanied by the appearanceof a 61-kDa band of gelatinase activity, suggestive ofan intermediary product formed as part of the 64/59kDa conversion reaction.
Several lines of evidence suggested that the 64/59 kDa gelatinase doublet represented the latent andactive forms of tree shrew MMP-2 (gelatinase A, 72-kDa type IV collagenase). Most important, the molecu-lar weights corresponded to those expected for latentMMP-2 and its activated form when run under nonre-ducing SDS-PAGE conditions.4241'47 The appearanceof an intermediary gelatinase product after APMA-activation is also characteristic of MMP-2.46 When scle-ral gelatinases were reduced and alkylated before zy-mography, a new band of gelatinase activity with anM, of 72 kDa appeared (Fig. 1C, lanes 3, 4), againconsistent with results expected for MMP-247 (this 72-kDa band corresponds to a reducing agent-induced,conformationally distinct form of latent MMP-2). The64/59 kDa scleral gelatinase doublet comigrated withthe gelatinase doublet secreted by tree shrew skin fi-broblasts when stimulated by the tumor promoter,phorbol 12-myristate 13-acetate (phorbol esters havebeen shown to induce the expression of MMPs, includ-ing MMP-2, in many fibroblast cell lines and also toinduce the activation of a considerable proportion ofthe secreted zymogens40'46'48). The similar electropho-retic migration patterns of the fibroblast-secreted gela-tinase doublet and the scleral gelatinase doublet sug-gest that these proteins are closely related. Finally, thescleral gelatinase doublet could be immunoprecipi-tated by an antibody to human MMP-2-TIMP-2, butnot by a control (sheep IgG) antibody, used at thesame concentration (Fig. ID). In these experiments,immunoprecipitation was conducted on scleral ex-tracts that had undergone reduction and alkylationto remove TIMPs to prevent immunoprecipitation ofgelatinase-TIMP complexes by TIMP immunoreactiv-ity. This result demonstrates that the scleral gelatinasedoublet is irnmunologically related to human MMP-2.In conclusion, we propose that the 64/59 kDa scleralgelatinase doublet represents the latent and activeforms of tree shrew MMP-2.
Active and Latent MMP-2 Are Present inDeveloping Tree Shrew Sclera
Zymograms of the gelatinase activity of normal treeshrew sclera demonstrated the presence of active andlatent MMP-2 (Fig. 2). The absolute amounts of latentand active MMP-2 found in the right and left eyes ofnormal animals (n = 5) are shown in Table 2 andFigure 3A. There were no significant differences inlatent or active MMP-2 levels between right and lefteyes. Although there was no difference in the absolute
amount of latent MMP-2 between equatorial and poste-rior sclera, there was significantly more active MMP-2in posterior, compared to equatorial, sclera (n = 10eyes from five animals; P < 0.01).
In the normal animals, most of the scleral MMP-2 was in the latent form (Fig. 3D). The ratio of active-to-latent MMP-2 was 0.23 ± 0.05 and 0.34 ± 0.06 (n= 10 eyes from five animals) in the equatorial andposterior sclera, respectively. This equatorial-poste-rior difference in active-to-latent ratios was significant(P < 0.01), caused by the higher level of active MMP-2 in posterior sclera noted above. These results suggestthat the posterior sclera undergoes more intensive re-modeling than the equatorial region at this stage ofnormal ocular development (20 days after eye open-ing).
Form-Deprived Myopic Eyes Show IncreasedLevels of Active MMP-2
There were substantial differences in the appearanceof zymograms from animals that had undergone 5days of form deprivation, compared to normals (Fig.4). There appeared to be much greater amounts ofactive MMP-2 in equatorial and posterior sclera ofform-deprived myopic eyes compared to their contra-lateral controls. These differences were confirmed byquantitative densitometric analysis (Table 2, Fig. 3B).
There was a significant increase in the absoluteamount of active MMP-2 in form-deprived myopic eyescompared to their contralateral control eyes (P <0.001 and P < 0.02 in equatorial and posterior sclera,respectively). There were no corresponding differ-ences in latent MMP-2 levels between control and myo-pic eyes in either the equatorial or the posterior sclera(P = 0.12 and P = 0.78, respectively). When data fromthe two eyes of MD animals were pooled (i.e., activityin the equatorial region of both eyes versus activity inthe posterior region of both eyes), the equatorialsclera had significantly higher latent MMP-2 levelsthan posterior sclera (161.8 ± 15.7 versus 119.6 ± 9U, respectively; P< 0.005). This difference is unlikelyto represent a methodological effect because the equa-torial and posterior samples from individual animalswere processed simultaneously.
In the sclera of normal tree shrews, most of theMMP-2 was in the latent form (see previous section).This was also the case in the control eye of MD ani-mals, but in deprived eyes in which myopia was devel-oping, there was a considerable increase in the active-to-latent ratio so that, in some cases, there was almostas much active MMP-2 as latent MMP-2. Differencesin the active-to-latent MMP-2 ratio between deprivedand control eyes were significant (P < 0.01 for bothequatorial and posterior sclera). Additionally, thehigher active-to-latent ratio of the posterior sclera,
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933417/ on 03/24/2018

Form-Deprivation Myopia Induces Activation of MMP-2
Standards Right eye
Equatorial
1387
Left eye
Latent (64 kDa)Active (59 kDa)
Posterior Latent (64 kDa)Active (59 kDa)
1 7 8 9 10 11FIGURE 2. SDS—PAGE gelatin zymography of normal group tree shrew sclera. MMP wereextracted from sclera and analyzed by zymography as described in Materials and Methods.{upper and lower gel panels) Equatorial and posterior scleral extracts, respectively (from asingle animal). Lanes 1 to 5, MMP-2 standards (256 U to 1 U per lane). Lanes 6 to 8, scleralextracts from right eye (fourfold serial dilution). Lanes 9 to 11, scleral extracts from lefteye (fourfold serial dilution). Comparisons between the two eyes were possible directly fromthe gel because sample loadings were normalized on the basis of tissue dry weight.
compared to the equatorial sclera, of deprived eyeswas significant (P < 0.01), as was found for normalanimals. In fact, the active-to-latent ratio for the poste-rior sclera of deprived eyes was significantly higherthan the active-to-latent ratio of all other groups (P <0.01,Tukey's).
Recovering Eyes Show Reduced Levels ofMMP-2
The levels of scleral MMP-2 in animals recoveringfrom form-deprivation myopia are shown in Table 2and Figure 3C. A typical zymogram from a recoverygroup animal is shown in Figure 5. After only 3 daysof recovery, there was a fivefold reduction in the level
of active MMP-2 in both the equatorial and posteriorsclera of eyes recovering from myopia, when com-pared to the deprived eyes of MD animals (equatorialP < 0.05; posterior P < 0.01; Tukey's). No significantdifferences in latent MMP-2 levels were found betweeneyes recovering from myopia and the deprived eyesof MD animals. These findings resulted in significantreductions in the active-to-latent MMP-2 ratio of boththe equatorial and posterior regions of recoveringeyes, compared to the deprived eyes of MD animals(equatorial P < 0.05; posterior P < 0.01; Tukey's).
The amount of active MMP-2 was reduced slightlyin recovering eyes, compared to their contralateralcontrols in both equatorial and posterior sclera. How-
TABLE 2. MMP-2 Activity of Tree Shrew Sclera
Equatorial ScleraActiveLatentTotalActive/Latent Ratio
Posterior ScleraActiveLatentTotalActive/Latent Ratio
Normals
Right
35.0 ± 15.9146.3 ± 18.8181.3 ± 33.40.21 ± 0.07
65.1 ± 20.9172.3 ± 21.0237.4 ± 38.50.36 ± 0.09
I
39.6 i155.1 i194.6 t0.26 d
67.6 d183.3 d
t 13.9t 16.9t 25.4t 0.08
t 27.9t 28.8
250.9 ± 54.80.33 ± 0.09
Deprived
86.7 ± 13.8172.6 ± 19.1259.3 ± 23.10.53 ± 0.10
99.3 ± 16.3121.2 ± 15.4220.5 ± 29.80.81 ± 0.09
MDs
Control
25.7 ± 5.0151.0 ± 25.8176.7 ± 26.40.19 ± 0.05
27.1 ± 9.2118.0 ± 11.0145.2 ± 12.30.25 ± 0.10
Recoveries
Recovering
17.1 ± 6.3155.9 ± 34.2173.0 ± 36.80.11 ± 0.03
17.9 ± 6.7115.9 ± 17.4133.8 ± 22.80.15 ± 0.03
Control
37.3 ± 11.3182.5 ± 23.2219.8 ± 31.3
0.20 ± 0.04
38.1 ± 15.8163.2 ± 31.1201.2 ± 42.80.22 ± 0.06
Values are mean i SEM; n = 5 for normal and recovery groups, n = 6 for MD group. Activity measured in arbitrary units.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933417/ on 03/24/2018
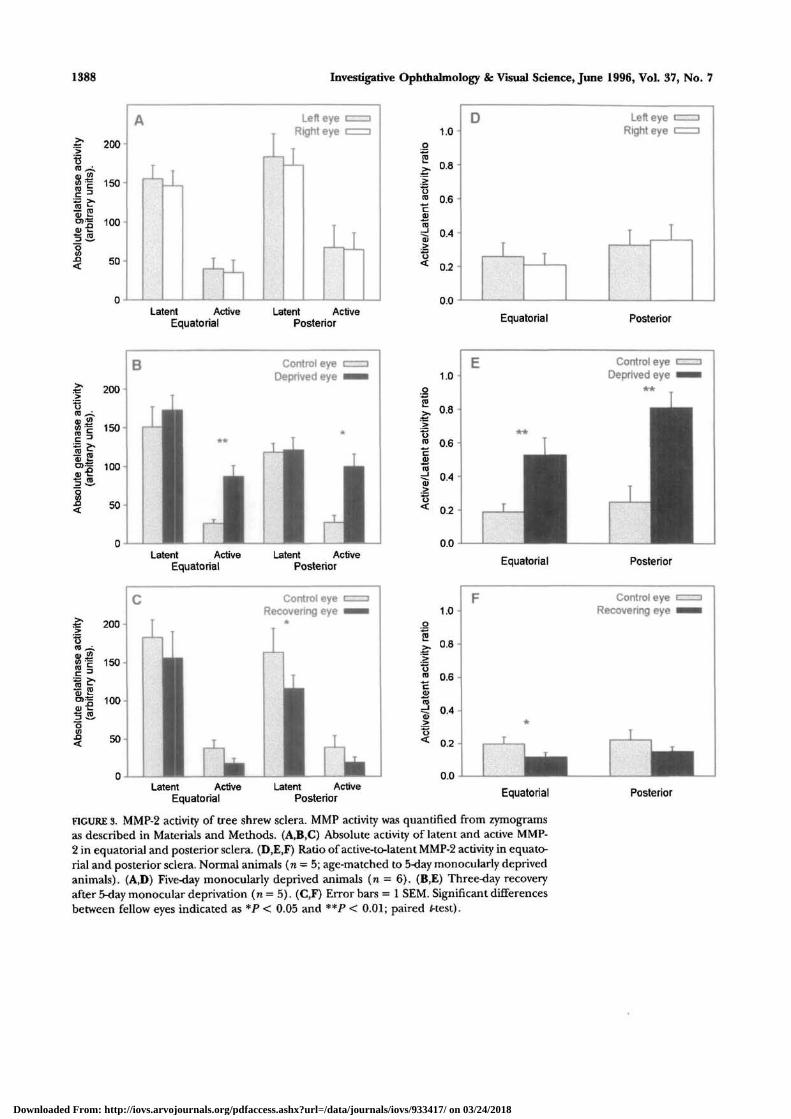
1388 Investigative Ophthalmology & Visual Science, June 1996, Vol. 37, No. 7
.-=* 200
8 |
8JD<
150-
100-
50-
Latent ActiveEquatorial
Latent ActivePosterior Equatorial Posterior
Latent ActiveEquatorial
Latent ActivePosterior
Control eyeRecovering eye
Latent ActiveEquatorial
Latent ActivePosterior
1.0
0.8
0.6-
0.4
Equatorial Posterior
Control eyeRecovering eye
Equatorial Posterior
FIGURE 3. MMP-2 activity of tree shrew sclera. MMP activity was quantified from zymogramsas described in Materials and Methods. (A,B,C) Absolute activity of latent and active MMP-2 in equatorial and posterior sclera. (D,E,F) Ratio of active-to-latent MMP-2 activity in equato-rial and posterior sclera. Normal animals (n = 5; age-matched to 5-day monocularly deprivedanimals). (A,D) Five-day monocularly deprived animals (n = 6). (B,E) Three-day recoveryafter 5-day monocular deprivation ( n = 5). (C,F) Error bars = 1 SEM. Significant differencesbetween fellow eyes indicated as *P < 0.05 and **P < 0.01; paired f-test).
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933417/ on 03/24/2018

Form-Deprivation Myopia Induces Activation of MMP-2 1389
ever, this difference only reached statistical signifi-cance when the equatorial and posterior enzyme activ-ities of each eye were pooled (P < 0.05, n = 10 pairsof measurements). Interestingly, for posterior sclera,the level of latent MMP-2 was also reduced in recov-ering eyes compared to contralateral control eyes (P< 0.05). Monocularly deprived animals showed nosuch difference in latent MMP-2 levels (see previoussection). The reduction in both active and latentMMP-2 levels in the posterior sclera of eyes recoveringfrom axial myopia resulted in an active-to-latent ratiothat was not significantly different to that of contralat-eral control eyes (P = 0.10). Conversely, for the equa-torial sclera, there was a significant reduction in theactive-to-latent ratio of recovering eyes compared tocontrol eyes (P < 0.02) because, equatorially, latentMMP-2 levels were similar in both recovering and con-trol eyes. There were no significant differences in ei-ther active or latent MMP-2 levels, nor in active-to-latent ratios, between the contralateral control eyes ofrecovery group animals and the eyes of normals (itshould be noted, however, that normals were agematched to 5-day MD animals and not to recoverygroup animals).
DISCUSSION
We hypothesized that if ocular enlargement duringthe development of form-deprivation myopia in treeshrews was the result of active tissue remodeling ratherthan just passive scleral stretching, we would find evi-
dence of increased ECM degradative activity in thescleras of myopic eyes. As potential markers of ECMdegradative activity, we chose to study scleral gela-tinases. Results demonstrated that scleral gelatinaseactivity is increased markedly during axial myopia de-velopment and is reduced during the recovery frommyopia.
In normal tree shrew sclera, MMP-2 was found tobe by far the most prominent gelatinase (a «90-kDaenzyme presumed to be latent MMP-9 accounted forless than 5% of all activity on gelatin zymograms).Most of the scleral MMP-2 was found to be in thelatent form. This suggests that in vivo the enzyme isprevented from autocatalytic activation, probably byan association with the inhibitor TIMP-2. The discov-ery of a proportion of MMP-2 in the active form indi-cates that at this stage of normal scleral development,some active tissue remodeling is occurring becauseactivated MMP-2 is catalytically active, even in the pres-ence of TIMP-2.:w 40
In myopic eyes that underwent 5 days of MD, thelevel of active MMP-2 was increased approximatelythreefold over that in contralateral control eyes andtwofold over that in age-matched normal eyes. LatentMMP-2 levels were not significantly different betweenthe deprived and control eyes of MD animals. Theincrease in active MMP-2 content shifted the balanceof active-to-latent MMP-2 in the sclera of deprived,myopic eyes so that, in posterior sclera, there was al-most as much active MMP-2 as latent MMP-2 (see Figs.3B, 4). Although the zymography technique does not
Standards Control eye Deprived eye
EquatorialLatent (64 kDa)Active (59 kDa)
Posterior Latent (64 kDa)Active (59 kDa)
1 8 10 11
FIGURE 4. SDS-PAGE gelatin zymography of monocularly deprived (MD) tree shrew sclera.{upper and loxver gel panels) Equatorial and posterior scleral extracts, respectively (from asingle animal). Lanes 1 to 5, MMP-2 standards (256 U to 1 U per lane panels). Lanes 6 to8, scleral extracts from control eye (fourfold serial dilution). Lanes 9 to 11, scleral extractsfrom deprived eye (fourfold serial dilution). Comparisons between the two eyes are possibledirectly from the gel.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933417/ on 03/24/2018

1390
Equatorial
Investigative Ophthalmology & Visual Science, June 1996, Vol. 37, No. 7
Standards Recoverina eve Control eve
Latent (64 kDa)Active (59 kDa)
Posterior Latent (64 kDa)Active (59 kDa)
1 8 10 11FIGURE 5. SDS-PAGE gelatin zymography of recovery group tree shrew sclera. {upper andbwer gel panels) Equatorial and posterior scleral extracts, respectively (from a single animal).Lanes 1 to 5, MMP-2 standards (256 U to 1 U per lane panels). Lanes 6 to 8, scleral extractsfrom recovering eye (fourfold serial dilution), Lanes 9 to 11, scleral extracts from controleye (fourfold serial dilution). Comparisons between the two eyes are possible direcdy fromthe gel.
measure the in situ level of gelatinase activity, becauseit is independent of scleral TIMP/TIMP-2 concentra-tions, the observed increase in the level of active MMP-2 indicates that scleral gelatinolytic activity is increasedmarkedly in form-deprived myopic eyes.
Two other studies, both in chick, have found dif-ferences in scleral gelatinase activity during the devel-opment of form-deprivation myopia.49'50 Using zymog-raphy, Jones et al'19 found increased latent and activegelatinase activity in chick sclera from myopic form-deprived eyes compared to controls, coupled with anincreased prevalence of low molecular weight cleavageproducts. The gelatinase activity was attributed to anenzyme of appropriate molecular weight to be MMP-2. A more detailed study of chick scleral gelatinaseactivity has been reported recently by Rada andBrenza.50 Using a [HH] gelatin degradation assay ratherthan the zymography technique, this group reportedan increase in latent, but not active, gelatinase activityin the posterior sclera of eyes developing form-depri-vation myopia compared to their contralateral controleyes. This contrasts with our results on tree shrewsclera showing that increased active gelatinase levelswere apparent in deprived eyes. As in the currentstudy, however, the metalloproteinase responsible forgelatinolytic activity was demonstrated to be MMP-2.In chick sclera, major differences in MMP-2 activitybetween anterior and posterior regions were apparentin deprived and contralateral control eyes, but therewere no significant differences in equatorial scleralgelatinase activity between die two eyes.r>0 This againcontrasts with our results from tree shrew sclera show-
ing similar levels of gelatinase activity in equatorialand posterior sclera and demonstrating that the ef-fects of form-deprivation on MMP-2 levels were quanti-tatively similar in the two regions. A surprising resultfrom the study of Rada and Brenza50 was that, in thechick, the only difference found in gelatinase, colla-genase, or proteoglycanase activity between deprivedand control eyes was for the latent (inactive) form ofgelatinase in posterior sclera. Because marked differ-ences in ECM turnover are apparent in chick scleraduring form deprivation,2324'51 this result suggests ei-ther that the turnover of ECM components is regu-lated primarily by synthesis rates alone (not by a com-bination of synthesis and degradation rates) or thatat the time tested (11 days of MD), scleral ECM degra-datory activity had returned to basal levels. An argu-ment in support of the latter explanation is that ifMMP-2 synthesis and activation were regulated inde-pendendy, MMP-2 activation in chick sclera may havebeen downregulated while enzyme synthesis was stillin an upregulated state. This would have led to anincreased abundance of MMP-2 in the latent form,consistent with the findings of Rada and Brenza. How-ever, the above hypothesis infers that by 11 days ofdeprivation, the rate of chick scleral ECM turnover indeprived eyes has slowed to levels comparable withcontrol eyes—a suggestion that seems inconsistentwith reports of turnover rates of, for example, chickscleral proteoglycans.51 V2 Therefore, it seems likelythat differences in scleral MMP-2 activity betweenchick and tree shrew eyes undergoing myopia develop-ment are mainly caused by species differences. Unlike
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933417/ on 03/24/2018
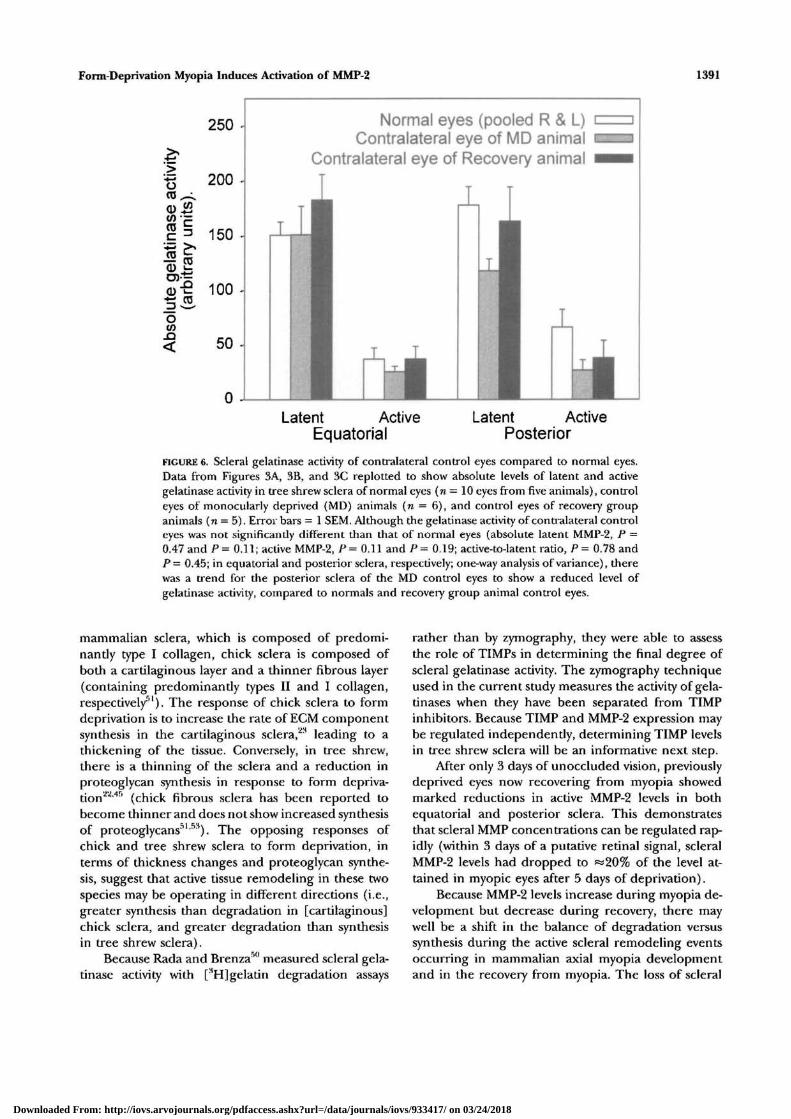
Form-Deprivation Myopia Induces Activation of MMP-2 1391
250
« 200o(0<D</)(Cc
elat
i
o(0
(0
'c
:rary
Normal eyes (pooled R & L)Contralateral eye of MD animal
Contralateral eye of Recovery animal
Latent ActiveEquatorial
Latent ActivePosterior
FIGURE 6. Scleral gelatinase activity of contralateral control eyes compared to normal eyes.Data from Figures 3A, 3B, and 3C replotted to show absolute levels of latent and activegelatinase activity in tree shrew sclera of normal eyes (n = 10 eyes from five animals), controleyes of monocularly deprived (MD) animals (re = 6), and control eyes of recovery groupanimals (n = 5). Error bars = 1 SEM. Although the gelatinase activity of contralateral controleyes was not significantly different than that of normal eyes (absolute latent MMP-2, P =0.47 and P= 0.11; active MMP-2, P= 0.11 and/> = 0.19; active-to-latent ratio, P = 0.78 andP — 0.45; in equatorial and posterior sclera, respectively; one-way analysis of variance), therewas a trend for the posterior sclera of the MD control eyes to show a reduced level ofgelatinase activity, compared to normals and recovery group animal control eyes.
mammalian sclera, which is composed of predomi-nantly type I collagen, chick sclera is composed ofboth a cartilaginous layer and a thinner fibrous layer(containing predominantly types II and I collagen,respectively51). The response of chick sclera to formdeprivation is to increase the rate of ECM componentsynthesis in the cartilaginous sclera,23 leading to athickening of the tissue. Conversely, in tree shrew,there is a thinning of the sclera and a reduction inproteoglycan synthesis in response to form depriva-tion2''''5 (chick fibrous sclera has been reported tobecome thinner and does not show increased synthesisof proteoglycans51"53). The opposing responses ofchick and tree shrew sclera to form deprivation, interms of thickness changes and proteoglycan synthe-sis, suggest that active tissue remodeling in these twospecies may be operating in different directions (i.e.,greater synthesis than degradation in [cartilaginous]chick sclera, and greater degradation than synthesisin tree shrew sclera).
Because Rada and Brenza50 measured scleral gela-tinase activity with [3H]gelatin degradation assays
rather than by zymography, they were able to assessthe role of TIMPs in determining the final degree ofscleral gelatinase activity. The zymography techniqueused in the current study measures the activity of gela-tinases when they have been separated from TIMPinhibitors. Because TIMP and MMP-2 expression maybe regulated independently, determining TIMP levelsin tree shrew sclera will be an informative next step.
After only 3 days of unoccluded vision, previouslydeprived eyes now recovering from myopia showedmarked reductions in active MMP-2 levels in bothequatorial and posterior sclera. This demonstratesthat scleral MMP concentrations can be regulated rap-idly (within 3 days of a putative retinal signal, scleralMMP-2 levels had dropped to «20% of the level at-tained in myopic eyes after 5 days of deprivation).
Because MMP-2 levels increase during myopia de-velopment but decrease during recovery, there maywell be a shift in the balance of degradation versussynthesis during the active scleral remodeling eventsoccurring in mammalian axial myopia developmentand in the recovery from myopia. The loss of scleral
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933417/ on 03/24/2018
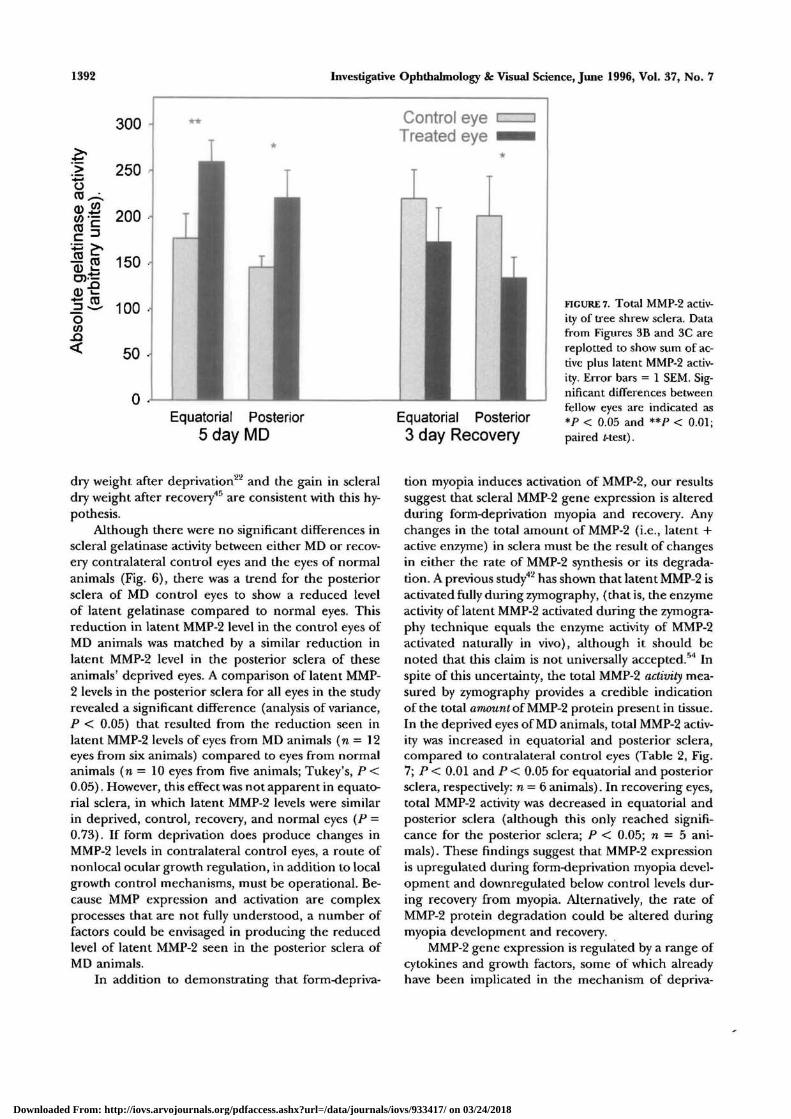
1392 Investigative Ophthalmology & Visual Science, June 1996, Vol. 37, No. 7
s |pD)-g
OCO
300 •
250 •
Control eyeTreated eye
Equatorial Posterior5 day MD
Equatorial Posterior3 day Recovery
FIGURE 7. Total MMP-2 activ-ity of tree shrew sclera. Datafrom Figures 3B and 3C arereplotted to show sum of ac-tive plus latent MMP-2 activ-ity. Error bars = 1 SEM. Sig-nificant differences betweenfellow eyes are indicated as*P < 0.05 and **P < 0.01;paired Mest).
dry weight after deprivation22 and the gain in scleraldry weight after recovery45 are consistent with this hy-pothesis.
Although there were no significant differences inscleral gelatinase activity between either MD or recov-ery contralateral control eyes and the eyes of normalanimals (Fig. 6), there was a trend for the posteriorsclera of MD control eyes to show a reduced levelof latent gelatinase compared to normal eyes. Thisreduction in latent MMP-2 level in the control eyes ofMD animals was matched by a similar reduction inlatent MMP-2 level in the posterior sclera of theseanimals' deprived eyes. A comparison of latent MMP-2 levels in the posterior sclera for all eyes in the studyrevealed a significant difference (analysis of variance,P < 0.05) that resulted from the reduction seen inlatent MMP-2 levels of eyes from MD animals (n = 12eyes from six animals) compared to eyes from normalanimals (n = 10 eyes from five animals; Tukey's, P <0.05). However, this effect was not apparent in equato-rial sclera, in which latent MMP-2 levels were similarin deprived, control, recovery, and normal eyes (P =0.73). If form deprivation does produce changes inMMP-2 levels in contralateral control eyes, a route ofnonlocal ocular growth regulation, in addition to localgrowth control mechanisms, must be operational. Be-cause MMP expression and activation are complexprocesses that are not fully understood, a number offactors could be envisaged in producing the reducedlevel of latent MMP-2 seen in the posterior sclera ofMD animals.
In addition to demonstrating that form-depriva-
tion myopia induces activation of MMP-2, our resultssuggest that scleral MMP-2 gene expression is alteredduring form-deprivation myopia and recovery. Anychanges in the total amount of MMP-2 (i.e., latent +active enzyme) in sclera must be the result of changesin either the rate of MMP-2 synthesis or its degrada-tion. A previous study42 has shown that latent MMP-2 isactivated fully during zymography, (that is, the enzymeactivity of latent MMP-2 activated during the zymogra-phy technique equals the enzyme activity of MMP-2activated naturally in vivo), although it should benoted that this claim is not universally accepted.54 Inspite of this uncertainty, the total MMP-2 activity mea-sured by zymography provides a credible indicationof the total amount of MMP-2 protein present in tissue.In the deprived eyes of MD animals, total MMP-2 activ-ity was increased in equatorial and posterior sclera,compared to contralateral control eyes (Table 2, Fig.7; P < 0.01 and P < 0.05 for equatorial and posteriorsclera, respectively: n = 6 animals). In recovering eyes,total MMP-2 activity was decreased in equatorial andposterior sclera (although this only reached signifi-cance for the posterior sclera; P < 0.05; n = 5 ani-mals) . These findings suggest that MMP-2 expressionis upregulated during form-deprivation myopia devel-opment and downregulated below control levels dur-ing recovery from myopia. Alternatively, the rate ofMMP-2 protein degradation could be altered duringmyopia development and recovery.
MMP-2 gene expression is regulated by a range ofcytokines and growth factors, some of which alreadyhave been implicated in the mechanism of depriva-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933417/ on 03/24/2018

Form-Deprivation Myopia Induces Activation of MMP-2 1393
tion-induced axial myopia. Among the most likely can-didates responsible for induced changes in MMP-2gene expression during form-deprivation myopia areTGF-/5, basic fibroblast growth factor, and the interleu-kins (for example, ILrl and IL-1032'55). However, thecontrol of MMP gene expression is likely to be a com-plex process. Retinoic acid and vasoactive intestinalpolypeptide, for example, are other modulators thatcould play a significant role in determining the degreeof MMP expression.56"58
If scleral MMP-2 is secreted as a complex of latentMMP-2 and TIMP-2, as is the case in most connectivetissues, then the enzyme would be unable to autoacti-vate. Therefore, irrespective of changes in MMP-2 ex-pression, the question arises: How does the enzymebecome activated? Matrix metalloproteinases oftenare expressed and activated coordinately, either byeach other59 or by other proteases.32 The serine prote-ase, plasmin, for example, has been implicated in theactivation of several metalloproteinases and may playa major role in the overall cascade of activation ofMMPs during active tissue remodeling.48'60 At present,the effect of form-deprivation myopia on the plasmin-ogen-plasmin system remains unknown, but it is anarea worthy of investigation.
In conclusion, this study provides further evi-dence that the ocular enlargement that occurs duringthe development of form-deprivation myopia is theresult of active tissue remodeling rather than of pas-sive scleral stretching alone. The involvement of MMP-2 in this response may be particularly relevant to thedegradation of scleral collagen because a recent re-port61 has demonstrated that MMP-2 is capable of di-gesting native, fibrillar type I collagen. This finding isin direct contradiction to the conclusions of earlierstudies suggesting that only MMP-1 and MMP-8 (inter-stitial collagenase and neutrophil collagenase, respec-tively) are capable of degrading fibrillar type I colla-gen. Because type I collagen constitutes 85% to 90%of tree shrew scleral protein,62 MMP-2 may be directlyresponsible for a significant degree of scleral collagendegradation during myopic ocular enlargement. It willnow be important to identify the agent(s) responsiblefor metalloproteinase activation during the develop-ment of myopia as a crucial step in elucidating thepathway by which retinal signals modulate eye growth.
Key Words
gelatinase A, matrix metalloproteinase, myopia, sclera, treeshrew
Acknowledgments
The authors thank Allen Bailey and John Tarlton for adviceand initial assistance with the zymography technique. Theauthors also thank Charles Cottriall, Eleanor Leech, Michele
Crisp, Patrick Lawlor and Shane Wainwright for technicalassistance.
References
1. Fledelius HC. Is myopia getting more frequent? Across-sectional study of 1416 Danes aged 16+. AdaOphthalmol. 1983; 61:545-559.
2. Sperduto RD, Seigel D, Roberts J, Rowland M. Preva-lence of myopia in the United States. Arch Ophthalmol.1983;101:405-407.
3. Curtin BJ. The Myopias: Basic Science and Clinical Man-agement. Philadelphia: Harper & Row; 1985.
4. Grey RH, Burns-Cox CJ, Hughes A. Blind and partialsight registration in Avon. Br J Ophthalmol. 1989;73:88-94.
5. Curtin BJ, Teng CC. Scleral changes in pathological• myopia. Trans Am Acad Ophthalmol Otalaryngol.1958;62:777-790.
6. Garzino A. Modificazione del collagene sclerale nellamiopia maligna. Ross Ital Ottal. 1956:25:241-244.
7. Liu KR, Chen MS, Ko LS. Electron microscopic studiesof the scleral collagen fiber in excessively high myopia.JFormos Med Assoc. 1986;85:1032-1038.
8. Bell GR. The Coleman theory of accommodation andits relevance to myopia. Am J Opt Assoc. 1980;51:582-588.
9. Coulombre AJ. The role of intraocular pressure in thedevelopment of the chick eye. / Exp Zool. 1958;133:211-225.
10. Neath P, Roche SM, Bee JA. Intraocular pressure-de-pendent and independent phases of growth of theembryonic chick eye. Invest Ophthalmol Vis Sri.1991;32:2483-2491.
11. Perkins ES, Phelps CD. Open angle glaucoma, ocularhypertension, low-tension glaucoma, and refraction.Arch Ophthalmol. 1982; 100:1464-1467.
12. Ahmad NN, Ala-Kokko L, Knowlton RG, et al. Stopcodon in the procollagen II gene (COL2A1) in a fam-ily with the Stickler syndrome (arthro-ophthalmopa-thy). ProcNatlAcad Sri USA. 1991; 88:6624-6627.
13. Maumenee IH. The Marfan syndrome is caused by apoint mutation in the fibrillin gene. Arch Ophthalmol.1992:110:472-473.
14. Wilkin DJ, Bogaert R, Lachman RS, Rimoin DL, EyreDR, Cohn DH. A single amino-acid substitution(G103D) in the type-II collagen triple-helix produceskniest dysplasia. Hum Mol Genet. 1994; 11:1999-2003.
15. Glass IA, Good P, Coleman MP, et al. Genetic map-ping of a cone and rod dysfunction (Aland Island eyedisease) to the proximal short arm of the human Xchromosome./Med Genet. 1993;30:1044-1050.
16. Pereira R, Khillan JS, Helminen HJ, Hume EL, Pro-ckop DJ. Transgenic mice expressing a partially de-leted gene for type-1 procollagen (COL1 Al): A breed-ing line with a phenotype of spontaneous fracturesand decreased bone-collagen and mineral. / Clin In-vest. 1993;91:709-716.
17. Avetisov ES, Savitskaya NF, Vinetskaya MI, IomdinaEN. A study of biochemical and biomechanical quali-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933417/ on 03/24/2018

1394 Investigative Ophthalmology 8c Visual Science, June 1996, Vol. 37, No. 7
ties of normal and myopic eye sclera in humans ofdifferent age groups. Metab Pediatr Syst Ophthalmol.1984;7:183-188.
18. Raviola E, Wiesel TN. An animal model of myopia. iVEnglJMed. 1985;312:1609-1615.
19. Funata M, Tokoro T. Scleral change in experimentallymyopic monkeys. Graefe's Arch Clin Exp Ophthalmol.199O;228:174-179.
20. McBrien NA, Norton TT. Prevention of collagen cross-linking increases form-deprivation myopia in treeshrew. Exp Eye Res. 1994; 59:475-486.
21. Cornell LM, McBrien NA. Alterations in collagen fibrildiameter in the sclera of experimentally myopic treeshrew eyes. ARVO Abstracts. Invest Ophthalmol Vis Sri.1994; 35:2068.
22. Reeder AP, McBrien NA. Biochemical changes in thesclera of tree shrews with high degrees of experimen-tal myopia. Ophthal Physiol Opt. 1993; 13:105. Abstract.
23. Christensen AM, Wallman J. Evidence that increasedscleral growth underlies visual deprivation myopia inchicks. Invest Ophthalmol Vis Sri. 1991; 32:2143-2150.
24. Rada JA, Thoft RA, Hassell AJ. Increased aggrecan(cartilage proteoglycan) production in the sclera ofmyopic chicks. Dev Biol. 1991; 147:303-312.
25. Wallman J, Adams JI. Developmental aspects of exper-imental myopia in chicks: Susceptibility, recovery andrelation to emmetropisation. Vis Res. 1987; 27:1139-1163.
26. Norton TT. Experimental myopia in tree shrews. In:Bock G, Widdows K, eds. Myopia and the Control ofEye Growth. Ciba Foundation Symposium; Chichester:John Wiley & Sons. 1990; 155:178-199.
27. Wallman J, Wildsoet C, Xu A, et al. Moving the retina:Choroidal modulation of refractive state. Vision Res.1995; 35:37-50.
28. Wilkinson JL, Hodos W. Intraocular pressure and eyeenlargement in chicks. CurrEyeRes. 1991;10:163-168.
29. Schaeffel F, Glasser A, Howland HC. Accommodation,refractive error, and eye growth in chickens. VisionRes. 1988;28:639-657.
30. Irving EL, Callender MG, Sivak JG. Inducing myopia,hyperopia, and astigmatism in chicks. Optom Vis Sri.1991;68:364-368.
31. Siegwart JT, Norton TT. Refractive and ocularchanges in tree shrews raised with plus or minuslenses. ARVO Abstracts. Invest Ophthalmol Vis Sri.1993:34:1208.
32. Woessner JF. Matrix metalloproteinases and their in-hibitors in connective tissue remodeling. FASEB J.1991;5:2145-2154.
33. Matrisian LM. The matrix-degrading metalloprotein-ases. Bioessays. 1992; 14:455-463.
34. Siegwart JT, Norton TT. Goggles for controlling thevisual environment of small animals. Lab Animal Sri.1994; 44:213-215.
35. Bailey AJ, Tarlton JF, Van der Stappen J, Sims TJ,Messina A. The continuous elongation technique forsevere Dupuytren's disease. J Hand Surg. 1994;19B(suppl 4):522-527.
36. Guggenheim JA, Crisp MS, Wainwright SD, McBrien
NA. Gelatinase activity in tree shrew sclera. ARVO Ab-stracts. Invest Ophthalmol Vis Sri. 1995;36:S760.
37. Fini ME, Girard MT. Expression of collagenolytic/gelatinolytic metalloproteinases by normal cornea. In-vest Ophthalmol Vis Sri. 199O;31:1779-1788.
38. Bartfai T. Preparation of metal-chelate complexes andthe design of steady-state kinetic experiments involv-ing metal nucleotide complexes. In: Brooker G,Greengard P, Robison GA, eds. Advances in Cyclic Nu-cleotide Research. New York: Raven Press; 1979; 10:219-242.
39. Howard EW, Bullen EC, Banda MJ. Regulation of theautoactivation of human 72-kDa progelatinase by tis-sue inhibitor of metalloproteinases-2. / Biol Chem.1991; 266:13064-13069.
40. Brown PD, Kleiner DE, Unsworth EJ, Stetler-Steven-son WG. Cellular activation of the 72 kDa type IVprocollagenase/TIMP-2 complex. Kidney Int. 1993; 43:163-170.
41. Heussen C, Dowdle EB. Electrophoretic analysis ofplasminogen activators in polyacrylamide gels con-taining sodium dodecyl sulfate. Anal Biochem. 1980;102:196-202.
42. Kleiner DE, Steder-Stevenson WG. Quantitative zy-mography: Detection of picogram quantities of gela-tinases. Anal Biochem. 1994; 218:325-329.
43. Mercer E, Cawston TE, DeSilva M, Hazleman BL. Puri-fication of a metalloproteinase inhibitor from humanrheumatoid synovial fluid. Biochem J. 1985; 231:505-510.
44. Hipps DS, Hembry MR, Docherty AJP, Reynolds JJ,Murphy G. Purification and characterization of hu-man 72-kDa gelatinase (type IV collagenase). BiolChem Hoppe Seyler. 1991; 372:287-296.
45. McBrien NA, Lawlor PL. Increased proteoglycan syn-thesis in the sclera of tree shrew eyes recovering fromform deprivation myopia. ARVO Abstracts. Invest Oph-thalmol Vis Sri. 1995;36:S760.
46. Strongin AY, Marmer BL, Grant GA, Goldberg GI.Plasma membrane-dependent activation of the 72-kDatype IV collagenase is prevented by complex forma-tion with TIMP-2./B?o/ Chem. 1993; 268:14033-14039.
47. Collier IE, Wilhelm SM, Eisen AZ, et al. H-ras onco-gene-transformed human bronchial epithelial cells(TBE-1) secrete a single metalloproteinase capable ofdegrading basement membrane collagen. J Biol Chem.1988:263:6579-6587.
48. Strongin AY, Collier I, Bannikov G, Marmer BL, GrantGA, Goldberg GI. Mechanism of cell surface activationof 72-kDa type IV collagenase. / Biol Chem. 1995;270:5331-5338.
49. Jones BE, Waldbillig RJ, Schoen TJ, Hodos W, ChaderGJ. Extracellular matrix metalloproteinases and insu-lin-like growth factor binding protein (IGFBP) arealtered in experimental myopia. ARVO Abstracts. In-vest Ophthalmol Vis Sri. 1992; 33:1054.
50. Rada JA, Brenza HL. Increased latent gelatinase activ-ity in the sclera of visually deprived chicks. Invest Oph-thalmol Vis Sri. 1995; 36:1555-1565.
51. Rada JA, Mathews AL, Brenza H. Regional proteogly-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933417/ on 03/24/2018

Form-Deprivation Myopia Induces Activation of MMP-2 1395
can synthesis in the sclera of experimentally myopicchicks. ExpEyeRes. 1994;59:747-760.
52. Rada JA, Carlson SR. Proteoglycan turnover in thesclera of normal and experimentally myopic chickeyes. ARVO Abstracts. Invest Ophthalmol Vis Set.1995;36:S760.
53. Gottlieb MD, Joshi HB, Nickla DL. Scleral changesin chicks with form-deprivation myopia. Curr Eye Res.1990;9:1157-1165.
54. Woessner JF Jr. Quantification of matrix metallopro-teinases in tissue samples. Methods Enzymol. 1995;248:510-528.
55. Reitamo S, Remitz A, Tamai K, Uitto J. Interleukin-10 modulates type I collagen and matrix metallopro-teinase gene expression in cultured human skin fi-broblasts. J Clin Invest. 1994;94:2489-2492.
56. Braunhut SJ, Moses MA. Retinoids modulate endothe-lial cell production of matrix-degrading proteases andtissue inhibitors of metalloproteinases (TIMP). J BiolChem. 1994;269:13472-13479.
57. Seko Y, Shimokawa H, Shimizu M, Tokoro T. In vivo &in vitro association of retinoic acid with form-depriva-
tion myopia in chicks. ARVO Abstracts. Invest Ophthal-mol Vis Sri. 1995;36:S758.
58. Goetzl EJ, Xia M, Sreedharan SP, Galardy R, HauserSL, Leppert D. Vasoactive intestinal peptide (VIP)stimulation of matrix metalloproteinase (MMP)-de-pendent chemotaxis (CT) of T cells across a modelbasement membrane (BM). FASEB J. 1995;9(suppl4):A1034
59. Crabbe T, O'Connell JP, Smith BJ, Docherty AJP. Re-ciprocated matrix metalloproteinase activation: A pro-cess performed by interstitial collagenase and progela-tinase A. Biochemistry. 1994; 33:14419-14425.
60. Sato H, Takino T, Okada Y, et al. A matrix metallopro-teinase expressed on the surface of invasive tumourcells. Nature. 1994;370:61-65.
61. Aimes RT, Quigley JP. Matrix metalloproteinase-2 isan interstitial collagenase. JBiol Chem. 1995; 270:5872-5876.
62. Norton TT, Miller EJ. Collagen and protein levels insclera during normal development, induced myopia,and recovery in tree shrews. ARVO Abstracts. InvestOphthalmol Vis Sri. 1995;36:S760.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933417/ on 03/24/2018


















