
First record of bluish ˛ ˘! L ˚- # ˆ˘! · thism (i.e., lacking yellow pigment) is used to...
6
227-256. LOuRENçO-dE-MORAES, R. & BATiSTA, V. G. & FERREiRA, R. B. (2014): defensive behaviors of Lep - todactylus chaquensis (Anura: Leptodactylidae).- Herpetology Notes, Braunschweig; 7: 391-392. MAR - cHiSiN, A. & ANdERSON, J. d. (1978): Strategies em- ployed by frogs and toads (Amphibia, Anura) to avoid predation by snakes (Reptilia, Serpentes).- Journal of Herpetology, Houston; 12 (2): 151-155. MiyATAkE, T. & NAkAyANA, S. & NiSHi, y. & NAkAJiMA, S. (2009): Tonically immobilized selfish prey can survive by sac- rificing others.- Proceedings of the Royal Society, London; (B) 276: 2762-2767. RiBEiRO, S. c. & ROBER - TO, i. J. & SALES, d. L. & ÁViLA, R. W. & ALMEidA, W. O. (2012): Amphibians and reptiles from the Araripe bioregion, northeastern Brazil.- Salamandra, Mann- heim; 48 (3): 133-146. SANTANA, d. J. & MâNGiA, S. & SiLVEiRA-FiLHO, R. R. & BARROS, L. c. S. & ANdRAdE, i. & NAPOLi, M. F. & JuNcÁ, F. & GARdA, A. A. (2015): Anurans from the middle Jaguaribe River region, ceará state, northeastern Brazil.- Biota Neo- tropica, campinas; 15 (3): 1-8. STEBBiNS, R. c. & cOHEN, N. W. (1995): A natural history of amphibians. Princeton (Princeton university Press), pp. 316. TOLEdO, R. c. & JAREd, c. (1995): cutaneous granular glands and amphibian venoms.- comparative Biochemistry and Physiology, London; 111A (1): 1-29. TOLEdO, L. F. & SAZiMA, i. & HAddAd, c. F. B. (2011): Behavioral defences of anurans: an overview.- Ethology Ecology & Evolution, Abingdon; 23: 1-25. TOLEdO, L. F. & TOZETTi, A. M. & ZiNA, J. (2005): Leptodactylus labyrinthicus (Pepper frog): Repertoire of defensive behavior.- Herpetological Bulletin, London; 91: 30-31. TOLEdO, L. F. & SAZiMA, i. & HAddAd, c. F. B. (2010): is it all death feigning? case in anurans.- Journal of Natural History, London; 44 (31-32): 1979-1988. V AMOSi, S. M. (2005): On the role of enemies in divergence and diversification of prey: a review and synthesis.- canadian Journal of Zoology, Ottawa; 83: 894-910. WiLLiAMS, c. R. & BROdiE Jr. E. d. & TyLER, M. J. & WALkER, S. J. (2000): Anti pre- dator mechanisms of Australian frogs.- Journal of Her- petology, Houston; 34 (3): 431-443. ZAMPROGNO, c. & ZAMPROGNO, M. G. F. & TEixEiRA, R. L. (1998): Bufo paracnemis (Sapo-boi). death feigning.- Herpetologi- cal Review; New york; 29 (2): 96-97. kEy WORdS: Amphibia: Anura: Leptodactyli- dae; Leptodactylus vastus; ethology, antipredator be- havior, body-raising, thanatosis; Brazil SuBMiTTEd: May 30, 2016. AuTHORS: déborah P. dE cASTRO 1, 2) (corre- sponding author, < [email protected] >), diva M. BORGES-NOJOSA 1, 2) , John A. A. OLiVEiRA 2) , Maria J. BORGES-LEiTE 1, 2) , Margarida M. x. dA SiLVA 2) , Tiago A. dE SOuSA 1, 2) & david James HARRiS 3) 1) Programa de Pós-Graduação em Ecologia e Recursos Naturais, universidade Federal do ceará. campus do Pici, Bloco 906, cEP 60.455-760, Forta- leza - cE, Brazil; 2) Núcleo Regional de Ofiologia, universidade Federal do ceará (NuROF-uFc) campus do Pici, Bloco 905, cEP 60.455-760, Fortaleza - cE, Brazil; 3) centro de investigação da Biodiversidade e Recursos Genéticos (ciBiO). universidade do Porto, campus Agrário de Vairão, Rua Padre Armando Quintas, cEP 4485-661, Vairão, Portugal. First record of bluish Podarcis muralis (LAuRENTi, 1768) Lizard coloration depends on the com- bined action of three classes of pigment or light-reflecting cells (i.e., chromatophores) located in the dermal layer of the skin. The xantophores are the most superficial and contain pigments (i.e., pteridines and/or caro- tenoids) that absorb short-wavelength light and reflect long wavelengths. iridophores contain intracellular guanine platelets that scatter the incident light. Melanophores oc- cupy a basal position in the dermis and con- tain eumelanin that absorbs all light trans- mitted by the xantophores and the irido - phores (cOOPER & GREENBERG 2002; GRE - THER et al. 2004). Variation in the abundance and in the spatial arrangement of these cell types can produce the great array of skin colors found in lizards (e.g., SAENkO et al. 2013). Blue coloration has long attracted the attention of researchers and herpetocultur- ists due, among other reasons, to the fact that blue pigments are almost absent in nature (BAGNARA 2007; uMBERS 2013; but see GOdA & FuJi 1995). in vertebrates, blue is generally thought to be a structural color that results from selective light scattering by nanoscale elements that differ in refractive index (BAGNARA 2007). in particular, short- wavelength colors (blue and ultraviolet- blue) in lizards are structural colors pro- duced by light scattering in the iridophores, although these colors also depend on inter- actions with xanthophores and the underly- ing layer of melanophores (MENTER et al. 1979; kuRiyAMA et al. 2006; BAGNARA et al. 2007). in Anolis carolinensis VOiGT, 1832, the isolated iridophores appear blue-green under reflected light, and the color intensi- fies if a layer of melanophores is added under the iridophores (presumably due to absorption of longer wavelengths, ROHR - LicH & PORTER 1972; ROHRLicH 1974). The addition of xanthophores containing yellow pigments results in the normal brown-green skin color of the species. These results sug- gest that blue colors are produced when xanthophores contain few or no pigments (allowing almost all wavelengths of the incident light to interact directly with the iri- dophores). consequently, the term axan- 218 SHORT NOTE HERPETOZOA 29 (3/4) Wien, 30. Jänner 2017 SHORT NOTE
Transcript of First record of bluish ˛ ˘! L ˚- # ˆ˘! · thism (i.e., lacking yellow pigment) is used to...

227-256. LOuRENçO-dE-MORAES, R. & BATiSTA, V. G.& FERREiRA, R. B. (2014): defensive behaviors of Lep-todactylus chaquensis (Anura: Leptodactylidae).-Herpetology Notes, Braunschweig; 7: 391-392. MAR -cHiSiN, A. & ANdERSON, J. d. (1978): Strategies em -ployed by frogs and toads (Amphibia, Anura) to avoidpredation by snakes (Reptilia, Serpentes).- Journal ofHerpetology, Houston; 12 (2): 151-155. MiyATAkE, T.& NAkAyANA, S. & NiSHi, y. & NAkAJiMA, S. (2009):Tonically immobilized selfish prey can survive by sac-rificing others.- Proceedings of the Royal Society,London; (B) 276: 2762-2767. RiBEiRO, S. c. & ROBER -TO, i. J. & SALES, d. L. & ÁViLA, R. W. & ALMEidA, W.O. (2012): Amphibians and reptiles from the Araripebioregion, northeastern Brazil.- Salamandra, Mann -heim; 48 (3): 133-146. SANTANA, d. J. & MâNGiA, S.& SiLVEiRA-FiLHO, R. R. & BARROS, L. c. S. &ANdRAdE, i. & NAPOLi, M. F. & JuNcÁ, F. & GARdA, A.A. (2015): Anurans from the middle Jaguaribe Riverregion, ceará state, northeastern Brazil.- Biota Neo -tropica, campinas; 15 (3): 1-8. STEBBiNS, R. c. &cOHEN, N. W. (1995): A natural history of amphibians.Princeton (Princeton university Press), pp. 316.TOLEdO, R. c. & JAREd, c. (1995): cutaneous granularglands and amphibian venoms.- comparativeBiochemistry and Physiology, London; 111A (1): 1-29.TOLEdO, L. F. & SAZiMA, i. & HAddAd, c. F. B.
(2011): Behavioral defences of anurans: an overview.-Ethology Ecology & Evolution, Abingdon; 23: 1-25.TOLEdO, L. F. & TOZETTi, A. M. & ZiNA, J. (2005):Leptodactylus labyrinthicus (Pepper frog): Repertoireof defensive behavior.- Herpetological Bulletin,London; 91: 30-31. TOLEdO, L. F. & SAZiMA, i. &HAddAd, c. F. B. (2010): is it all death feigning? casein anurans.- Journal of Natural History, London; 44(31-32): 1979-1988. VAMOSi, S. M. (2005): On the roleof enemies in divergence and diversification of prey: areview and synthesis.- canadian Journal of Zoology,Ottawa; 83: 894-910. WiLLiAMS, c. R. & BROdiE Jr. E.d. & TyLER, M. J. & WALkER, S. J. (2000): Anti pre -dator mechanisms of Australian frogs.- Journal of Her -petology, Houston; 34 (3): 431-443. ZAMPROGNO, c. &ZAMPROGNO, M. G. F. & TEixEiRA, R. L. (1998): Bufoparacnemis (Sapo-boi). death feigning.- Herpetologi -cal Review; New york; 29 (2): 96-97.
kEy WORdS: Amphibia: Anura: Leptodactyli -dae; Leptodactylus vastus; ethology, antipredator be -havior, body-raising, thanatosis; Brazil
SuBMiTTEd: May 30, 2016.
AuTHORS: déborah P. dE cASTRO 1, 2) (corre -sponding author, < [email protected] >),diva M. BORGES-NOJOSA 1, 2), John A. A. OLiVEiRA 2),Maria J. BORGES-LEiTE 1, 2), Margarida M. x. dA SiLVA 2),Tiago A. dE SOuSA 1, 2) & david James HARRiS 3)
1) Programa de Pós-Graduação em Ecologia eRecursos Naturais, universidade Federal do ceará.campus do Pici, Bloco 906, cEP 60.455-760, Forta -leza - cE, Brazil;
2) Núcleo Regional de Ofiologia, universidadeFederal do ceará (NuROF-uFc) campus do Pici, Bloco905, cEP 60.455-760, Fortaleza - cE, Brazil;
3) centro de investigação da Biodiversidade eRecursos Genéticos (ciBiO). universidade do Porto,campus Agrário de Vairão, Rua Padre ArmandoQuintas, cEP 4485-661, Vairão, Portugal.
First record of bluish Podarcismuralis(LAuRENTi, 1768)
Lizard coloration depends on the com-bined action of three classes of pigment orlight-reflecting cells (i.e., chromatophores)located in the dermal layer of the skin. Thexantophores are the most superficial andcontain pigments (i.e., pteridines and/or caro -tenoids) that absorb short-wavelength lightand reflect long wavelengths. iridophorescontain intracellular guanine platelets thatscatter the incident light. Melanophores oc -cupy a basal position in the dermis and con-tain eumelanin that absorbs all light trans-mitted by the xantophores and the irido -phores (cOOPER & GREENBERG 2002; GRE -THER et al. 2004). Variation in the abundanceand in the spatial arrangement of these celltypes can produce the great array of skincolors found in lizards (e.g., SAENkO et al.2013).
Blue coloration has long attracted theattention of researchers and herpetocultur-ists due, among other reasons, to the factthat blue pigments are almost absent innature (BAGNARA 2007; uMBERS 2013; butsee GOdA & FuJi 1995). in vertebrates, blueis generally thought to be a structural colorthat results from selective light scattering bynanoscale elements that differ in refractiveindex (BAGNARA 2007). in particular, short-wavelength colors (blue and ultraviolet-blue) in lizards are structural colors pro-duced by light scattering in the iridophores,although these colors also depend on inter-actions with xanthophores and the underly-ing layer of melanophores (MENTER et al.1979; kuRiyAMA et al. 2006; BAGNARA et al.2007). in Anoliscarolinensis VOiGT, 1832,the isolated iridophores appear blue-greenunder reflected light, and the color intensi-fies if a layer of melanophores is addedunder the iridophores (presumably due toabsorption of longer wavelengths, ROHR -LicH & PORTER 1972; ROHRLicH 1974). Theaddition of xanthophores containing yellowpigments results in the normal brown-greenskin color of the species. These results sug-gest that blue colors are produced whenxanthophores contain few or no pigments(allowing almost all wavelengths of theincident light to interact directly with the iri-dophores). consequently, the term axan-
218 SHORT NOTE HERPETOZOA 29 (3/4) Wien, 30. Jänner 2017 SHORT NOTE
ALL_SHORT_NOTES:SHORT_NOTE.qxd 08.02.2017 16:41 Seite 36

thism (i.e., lacking yellow pigment) is usedto designate individuals exhibiting abnor-mal bluish coloration (BAGNARA et al. 1978,2007). Although the mechanisms of colorproduction are often unclear, blue colorationin lacertids is generally attributed to lack ofpigments in the xantophores (e.g., Ibero-
lacerta martinezricai(ARRiBAS, 1996), AR -RiBAS et al. 2008). Alternatively, it has beensuggested that the blue coloration of someinsular populations of Podarcis siculus(RAFiNESQuE-ScHMALTZ, 1810), may becaused by an increase in the concentrationof dermal melanin and is therefore consid-
SHORT NOTE HERPETOZOA 29 (3/4) Wien, 30. Jänner 2017 SHORT NOTE 219
Fig. 1: dorsal and ventral views of the bluish (B) Podarcismuralis(LAuRENTi, 1768), and of same-sex and same location individuals showing white (W) ventral background coloration for comparison.
Above ‒ bluish female from Porta (Languedoc-Roussillon, département Pyrénées Orientales, France; 42°31’38’’N, 1°49’36’’E); note orange scales on the throat.
Below ‒ bluish male from Latour de carol (Languedoc-Roussillon, département Pyrénées Orientales, France; 42°27’57’’N, 1°53’23’’E).
ALL_SHORT_NOTES:SHORT_NOTE.qxd 08.02.2017 16:41 Seite 37

ered a type of melanism (RAiA et al. 2010;NOVOSOLOV et al. 2013; see also QuiNN &HEWS 2003).
in June 2015, during fieldwork in theeastern Pyrenees (France; for locality detailssee caption of Fig. 1), the authors found twoindividuals of Podarcismuralis (LAuRENTi,1768) with an unusual blue coloration intwo localities ca. nine km apart. One was anadult female (snout-vent length, SVL: 59mm, body mass: 4.3 g) with a light blue col-oration over all her body (especially in theventral surface), and some orange scales inthe throat (Fig. 1). The other was an adultmale (SVL: 61 mm, body mass: 5.5 g)showing a darker, more saturated blue col-oration than the female, also affecting theentire body surface (Fig. 1). Only these two
individuals from a total of 1,118 lizards cap-tured during this field season presented thisabnormal blue coloration (˂ 0.18 %) andnone were observed during previous fieldcampaigns (2005-2014). Both animals werefound basking on stone walls (Fig. 2). Theanimals were captured by noosing, putinside individual cloth bags, and transferredto a dark room for spectrometric measure-ments. Reflectance spectra were obtainedwith a portable uSB-2000 spectrometer anda Px-2 xenon strobe light (Ocean Opticsinc., dunedin, Fl. u.S.A.) using standardspectrophotometric techniques (FONT et al.2009; PéREZ i dE LANuZA & FONT 2011).Measurements extended over the 300-700nm range, which encompasses the entirevisual spectrum of lacertids (PéREZ i dELANuZA & FONT 2014; MARTiN et al. 2015).To further characterize the unusual colora -tion, the reflectance spectra of the bluishlizards were compared to the mean reflec -tance curve of a sample of males (N =14)and females (N= 13) with normal colorationcaptured at the same localities (Fig. 3). Forthis comparison the authors used animalshaving exclusively white ventral colorationas this phenotype is the most common inthis area. Both lizards were released at theircapture sites after measurements. The malewas again sighted in May 2016, at the samelocation where it was originally captured.
Reflectance spectra (Fig. 3) under-score the singularity of the blue colorationexhibited by the individuals reported in thisstudy. Spectra from the throat and belly ofnormal-colored white individuals rise steep -ly toward long wavelengths, peaking at~660 nm. in contrast, these spectra arerather flat in the blue male and female, andpeak at lower wavelengths (ca. 420 nm inthe male, ca. 590 nm in the female). Spectraof the two blue individuals differ both inhue (i.e., spectral peak location) and bright-ness, the female having a less chromaticallypure blue than the male. Blue coloration innormal-colored P. muralis, as in other lacer-tids, is restricted to the outer ventral scales(PéREZ i dE LANuZA et al. 2013). Some ofthese scales, particularly in males, displayconspicuous blue patches which, upon spec-trophotometric examination, have theirreflectance peak in the ultraviolet (uV)range (ca. 370 nm). These uV-blue patches
220 SHORT NOTE HERPETOZOA 29 (3/4) Wien, 30. Jänner 2017 SHORT NOTE
Fig. 2: A ‒ Bluish male Podarcismuralis(LAuRENTi,1768), perched in its place of capture.
B ‒ close-up of the head.
ALL_SHORT_NOTES:SHORT_NOTE.qxd 08.02.2017 16:41 Seite 38
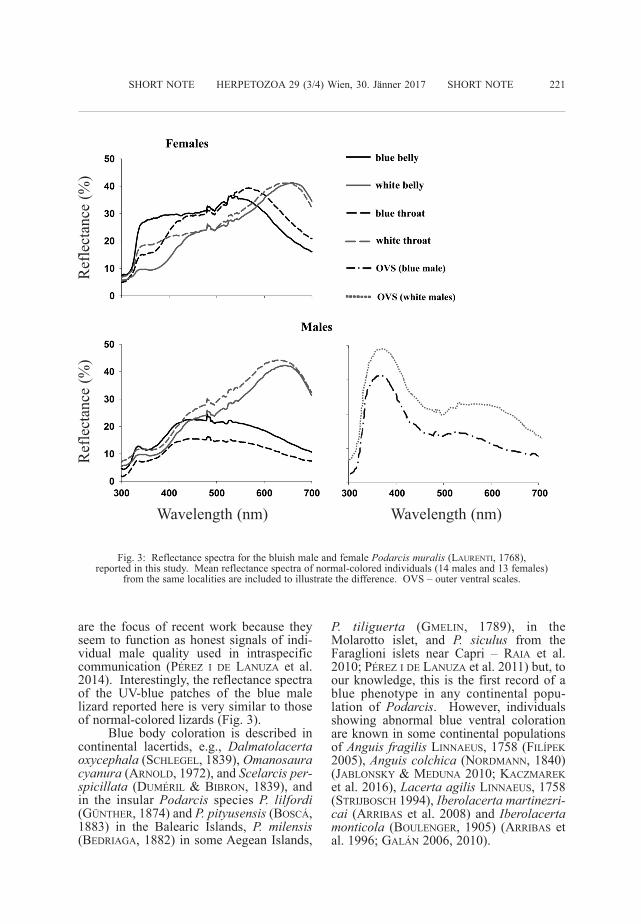
are the focus of recent work because theyseem to function as honest signals of indi-vidual male quality used in intraspecificcommunication (PéREZ i dE LANuZA et al.2014). interestingly, the reflectance spectraof the uV-blue patches of the blue malelizard reported here is very similar to thoseof normal-colored lizards (Fig. 3).
Blue body coloration is described incontinental lacertids, e.g., Dalmatolacertaoxycephala (ScHLEGEL, 1839), Omanosauracyanura (ARNOLd, 1972), and Scelarcisper-spicillata (duMéRiL & BiBRON, 1839), andin the insular Podarcis species P. lilfordi(GüNTHER, 1874) and P.pityusensis(BOScÁ,1883) in the Balearic islands, P. milensis(BEdRiAGA, 1882) in some Aegean islands,
P. tiliguerta (GMELiN, 1789), in theMolarotto islet, and P. siculus from theFaraglioni islets near capri ‒ RAiA et al.2010; PéREZ i dE LANuZA et al. 2011) but, toour knowledge, this is the first record of ablue phenotype in any continental popu -lation of Podarcis. However, individualsshowing abnormal blue ventral colorationare known in some continental populationsof Anguis fragilisLiNNAEuS, 1758 (FiLíPEk2005), Anguiscolchica (NORdMANN, 1840)(JABLONSky & MEduNA 2010; kAcZMAREket al. 2016), Lacertaagilis LiNNAEuS, 1758(STRiJBOScH 1994), Iberolacertamartinezri-cai (ARRiBAS et al. 2008) and Iberolacertamonticola (BOuLENGER, 1905) (ARRiBAS etal. 1996; GALÁN 2006, 2010).
SHORT NOTE HERPETOZOA 29 (3/4) Wien, 30. Jänner 2017 SHORT NOTE 221
Fig. 3: Reflectance spectra for the bluish male and female Podarcismuralis(LAuRENTi, 1768), reported in this study. Mean reflectance spectra of normal-colored individuals (14 males and 13 females)
from the same localities are included to illustrate the difference. OVS – outer ventral scales.
Wavelength (nm) Wavelength (nm)
Ref
lect
ance
(%
)R
efle
ctan
ce (
%)
ALL_SHORT_NOTES:SHORT_NOTE.qxd 08.02.2017 16:41 Seite 39

The occurrence of sporadic blue spec-imens in mainland populations of a lacertidspecies suggests the transition to blueishphenotypes could be produced by a rathersimple mutation (FROST & MALAciNSki1979; BukOWSki et al. 1990). This mutationwould reduce the concentration of shortwavelength-absorbing pigments in the xan-thophores or cause dermal melanisation,making the lizards appear blue. in conti-nental populations natural selection mayeliminate blue mutants, but in insular popu-lations the selective regime may be different(FuLGiONE et al. 2008; see also MAcEdONiAet al. 2009), which could help the mutationto become fixed. This may explain whyblue individuals are hardly found on thecontinent but are relatively frequent inislands.
AckNOWLEdGMENTS: This work was par-tially supported by an international PostgraduateFellowship at ciBiO-inBiO (universidade do Porto)funded by Fundación Mutua Madrileña. The secondauthor (GPL) was supported by a post-doctoral grant(SFRH/BPd/94582/ 2013) from FcT (Fundação para aciência e a Tecno logia) from the Portuguese Ministérioda Educação e ciência, and a grant from the SpanishMinisterio de Educación y ciencia (cGL2011-23751).Permits were provided by the Préfecture des Pyrénées-Orientales (Arrêté n° 2013095-0001).
REFERENcES: ARRiBAS, O. J. (1996): Taxo -nomic revision of the iberian “Archaeolacertae” i: Anew interpretation of the geographical variation of“Lacerta” monticola BOuLENGER, 1905, and “Lacerta”cyreni MüLLER & HELLMicH, 1937.- Herpetozoa, Wien;9: 31-56. ARRiBAS, O. & cARBONERO, J. & LiZANA, M.(2008): Sobre el dicromatismo ventral verde/azul en lalagartija batueca, Iberolacertamartinezricai(ARRiBAS,1996).- Boletín de la Asociación Herpetológica Españo -la, Madrid; 19: 51-54. BAGNARA, J. T. & FERNANdEZ,P. J. & FuJii, R. (2007): On the blue coloration of ver-tebrates.- Pigment cell Research, Oxford; 20: 14-26.BAGNARA, J. T. & FROST, S. k. & MATSuMOTO, J.(1978): On the development of pigment patterns in am -phibians.- American Zoologist, Lawrence; 18: 301-312. BukOWSki, L. & ERickSON, k. & LyERLA, T. A.(1990): characterization of the yellow pigment in theaxanthic mutant of the Mexican axolotl, Ambystomamexicanum.- Pigment cell Research, Oxford; 3: 123-125. cOOPER Jr., W. E. & GREENBERG, N. (1992):Reptilian coloration and behavior; pp. 298-422. in:GANS, c. & crews, d. (Eds.): Biology of the reptilia.Volume 18. Physiology E : Hormones, brain and be -havior. chicago, London (university of chicagoPress). FiLíPEk, M. (2005): Neobvykle sfarbenýslepúch lámavý.- Živa, Praha; 1: 36. [unusuallycoloured Slow Worm - in czech]. FONT, E. & PéREZ idE LANuZA, G. & SAMPEdRO, c. (2009): ultravioletreflectance and cryptic sexual dichromatism in theocellated lizard, Lacerta (Timon) lepida (Squamata:Lacertidae).- Biological Journal of the Linnean So -
ciety, Oxford; 97: 766-780. FROST, S. k. & MALA -ciNSki, G. M. (1979): The developmental genetics ofpigment mutants in the Mexican axolotl.- develop -mental Genetics, New york; 1: 271-294. FuLGiONE, d.& GuGLiELMi, S. & OdiERNA, G. & RiPPA, d. & cALi -ENdO, M. F. & RASTOGi, R. k. (2008): Morphologicaldifferentiation and genetic structure in island lizardpopulations.- Zoological Science, Tokyo, kawaguchi;25: 465-474. GALÁN, P. (2006). coloración azul atípi-ca en machos de Iberolacertamonticola del extremonorte de Galicia.- Boletín de la Asociación Herpeto -lógica Española, Madrid; 17: 96-99. GALÁN, P. (2010):dicromatismo ventral verde-azul en una población deIberolacertamonticola.- Boletín de la Asociación Her -petológica Española, Madrid; 21: 49-52. GOdA, M. &FuJii, R. (1995): Blue chromatophores in two speciesof callionymid fish.- Zoological Science, Tokyo,kawaguchi; 12: 811-813. GRETHER, G. F. & kOLLuRu,G. R. & NERSiSSiAN, k. (2004): individual colourpatches as multicomponent signals.- BiologicalReviews, Oxford; 79: 583-610. JABLONSki, d. & ME -duNA, P. (2010): Blue colour of the ventral body part ofEastern Slow Worm Anguis colchica (NORdMANN,1840).- Herpetology Notes, Braunschweig; 3: 295-296.kAcZMAREk, P., SkAWińSki, T., & SkóRZEWSki, G.(2016). Blue venter in the slow worm (Anguisfragilis):review and new data.- Herpetological Review, Newyork; 47: 375-377. kuRiyAMA, T. & MiyAJi, k. &SuGiMOTO, M. & HASEGAWA, M. (2006): ultrastructureof the dermal chromatophores in a lizard (Scincidae:Plestiodonlatiscutatus) with conspicuous body and tailcoloration.- Zoological Science, Tokyo, kawaguchi;23: 793-799. MAcEdONiA, J. M. & LAPPiN, A. k. &LOEW, E. R. & McGuiRE, J. A. & HAMiLTON, P. S. &PLASMAN, M. & BRANdT, y. & LEMOS-ESPiNAL, J. A. &kEMP, d. J. (2009): conspicuousness of dickerson’scollared lizard (Crotaphytusdickersonae) through theeyes of conspecifics and predators.- Biological Journalof the Linnean Society, Oxford; 97: 749-765. MARTiN,M. & LE GALLiARd, J. F. & MEyLAN, S. & LOEW, E. R.(2015): The importance of ultraviolet and near-infraredsensitivity for visual discrimination in two species oflacertid lizards.- Journal of Experimental Biology,cambridge; 218: 458-465. MENTER, d. G. & OBikA,M. & TcHEN, T. T. & TAyLOR, J. d. (1979): Leuco -phores and iridophores of Fundulusheteroclitus: bio-physical and ultrastructural properties.- Journal ofMorphology, Hoboken; 160: 103-119. NOVOSOLOV, M.& RAiA, P. & MEiRi, S. (2013): The island syndrome inlizards.- Global Ecology and Biogeography, Oxford;22: 184-191. PéREZ i dE LANuZA, G. & FONT, E.(2011): Lizard blues: Blue body colouration and ultra-violet polychromatism in lacertids.- Revista Españolade Herpetología, Madrid; 24: 67-84. PéREZ i dE LA -NuZA, G. & cARAZO, P. & FONT, E. (2014): colours ofquality: structural (but not pigment) coloration informsabout male quality in a polychromatic lizard.- AnimalBehaviour, Amsterdam, Oxford; 90: 73-81. PéREZ i dELANuZA, G. & FONT, E. (2014): ultraviolet vision inlacertid lizards: evidence from retinal structure, eyetransmittance, SWS1 visual pigment genes and behav-iour.- Journal of Experimental Biology, cambridge;217: 2899-2909. PéREZ i dE LANuZA, G. & FONT, E. &MONTERdE, J. L. (2013): using visual modelling tostudy the evolution of lizard coloration: sexual selec-tion drives the evolution of sexual dichromatism in lac-ertids.- Journal of Evolutionary Biology, Oxford; 26:
222 SHORT NOTE HERPETOZOA 29 (3/4) Wien, 30. Jänner 2017 SHORT NOTE
ALL_SHORT_NOTES:SHORT_NOTE.qxd 08.02.2017 16:41 Seite 40

1826-1835. QuiNN, V. S. & HEWS, d. k. (2003):Positive relationship between abdominal colorationand dermal melanin density in phrynosomatid lizards.-copeia, Washington; 2003: 858-864. RAiA, P. & GuA -RiNO, F. M. & TuRANO, M. & POLESE, G. & RiPPA, d. &cAROTENuTO, F. & MONTi, d. M. & cARdi, M. & FuL -GiONE, d. (2010): The blue lizard spandrel and theisland syndrome.- BMc Evolutionary Biology, London :BioMed central, Berlin, Heidelberg; 10: 289. ROHR -LicH, S. T. (1974): Fine structural demonstration ofordered arrays of cytoplasmic filaments in vertebrateiridophores. A comparative survey.- Journal of cellBiology, New york; 62: 295-304. ROHRLicH, S. T. &PORTER k. R. (1972): Fine structural observations relat-ing to the production of color by the iridophores of alizard, Anolis carolinensis.- Journal of cell Biology,New york; 53: 38-52. SAENkO, S. V. & TEySSiER, J. &VAN dER MAREL, d. & MiLiNkOViTcH, M. c. (2013):Precise colocalization of interacting structural and pig-mentary elements generates extensive color patternvariation in Phelsuma lizards.- BMc EvolutionaryBiology, London : BioMed central, Berlin, Heidelberg;11: 105. STRiJBOScH, H. (1994): Een blauwe Zand -hagedis (Lacerta agilis).- Lacerta, ‘s-Gravenhage; 52(6): 147-148. uMBERS, k. d. (2013): On the percep-tion, production and function of blue colouration inanimals.-Journal of Zoology, London; 289: 229-242.
kEy WORdS: Reptilia: Squamata: Sauria: La -certidae; Podarcismuralis, melanism, blue coloration,axanthism, physiology; eastern Pyrenees, France
SuBMiTTEd: February 16, 2016AuTHORS: Javier ÁBALOS (corresponding
author < [email protected] >) 1, 2), GuillemPéREZ i dE LANuZA 2), Senda REGuERA 3), ArnaudBAdiANE 4, 1), Jindřich BREJcHA 5, 1) & Enrique FONT 1)
1). Ethology Lab, instituto cavanilles deBiodiversidad y Biología Evolutiva, universitat deValència, 46071 Valencia, Spain.
2). centro de investigacão em Biodiversidade eRecursos Genéticos, universidade do Porto, 4485-661Vairão, Vila do conde, Portugal.
3) departamento de Zoología, Facultad deciencias, universidad de Granada, E-18071 Granada,Spain.
4) department of Biological Sciences, Mac-quarie university, Sydney, New South Wales 2109,Australia.
5) department of Philosophy and History ofScience, charles university in Prague, Prague 2 12000, czech Republic.
SHORT NOTE HERPETOZOA 29 (3/4) Wien, 30. Jänner 2017 SHORT NOTE 223
ALL_SHORT_NOTES:SHORT_NOTE.qxd 08.02.2017 16:41 Seite 41


















