
1 摂食・嚥下の生理・解剖 · 2015-07-23 · 1 1 摂食・嚥下の生理・解剖 (1)解剖 (2)生理 「摂食・嚥下の流れ」 図Aを挿入(別添資料)

淡水・沿岸域におけるアミ類の摂餌生態
誌名誌名 日本プランクトン学会報
ISSNISSN 03878961
巻/号巻/号 511
掲載ページ掲載ページ p. 46-72
発行年月発行年月 2004年2月
農林水産省 農林水産技術会議事務局筑波産学連携支援センターTsukuba Business-Academia Cooperation Support Center, Agriculture, Forestry and Fisheries Research CouncilSecretariat

Bull. Plankton Soc. Japan 51(1): 46-72, 2004
淡水・沿岸域におけるアミ類の摂餌生態(総説)
高橋一生
水産総合研究センター東北区水産研究所 干985-0001宮城県塩竃市新浜町 327-5
日本プランクトン学会報
⑥ The Plankton Society of Japan 2004
Feeding ecology of mysids in freshwater and coastal marine habitats: A review
KAZUT AKA TAKAHASHI
Tohoku Nαtional Fisheries Research Institute, 3 27 5 Shinhαma, Shiogama, Miyagi 985 0001
E-mail: [email protected]》
Abstract This paper reviews the feeding ecology of mysids in freshwater and marine coastal
environments, focusing particularly feeding mechanisms, stomach contents, feeding rate, food
selectivity, and its impact on ecosystems. Mysids exhibit two distinct feeding modes; a suspen-
sion feeding mode in which small particles such as detritus, phytoplankton and small zooplankton
are acquired; a raptorial feeding mode in which large preys are actively seized with the thoracic
appendages. In addition, some species seem to be able to obtain benthic diets. Generally, most
mysids exhibit omnivorous feeding habits consuming a wide range of prey items, including
detritus, algae, diatoms, rotifers, copepods, cladocerans, amphipods, carrion, fragments of terres-
trial plants’leaves and of macroalgae, seeds, etc. Diel and seasonal variations in stomach contents
are noticed in many species. Most mysid species feed actively at night in association with vertical
or horizontal migration. Seasonal change in diet is most probably related to food availability and
ontogenetical change in diets. Small sized food items such as phytoplankton, rotifers, detritus are
more frequently eaten by smaller individuals, whereas larger individuals exhibit a higher degree
of carnivory. The tendency for carnivory increases with growth (life-history omnivory) reflect
their increased success in prey capture. Laboratory and field measurements of predation rate
indicate that mysids have a high potential to consume mesozooplankton. Predation rates range from 0.4 to 42.6 μgC ind-1 h一lfor copepods (3 30% of body carbon per day), and from 1.3 to 102
μgC ind 1 h 1 for cladocerans (24-301 % of body carbon per day). Diatoms seem to be preferable
food items compared to other planktonic algae such as green algae and blue-green algae. The
significance of detritus as a diet is variable and depends on its origin and availability of other food
items. Mysids can feed selectively on different species and size groups of zooplankton, depending
on prey availability, predator-prey encounter frequency, prey escape response and mysid’s ability
to capture and handling. In some circumstances, mysids can optimize their feeding behavior by
switching their feeding mode in relation to food abundance. Flexible omnivory in mysids is
advantageous, providing effective feeding throughout the year. This feeding behavior also
enables shallow water-dwelling mysids to utilize the organic matter from various sources, i.e. water column, benthic and terrestrial ecosystems.
Key words: Mysida, feeding, omnivory, zooplanktivory, detritus
2003年 5月7日受付, 2004年 1月10日受理

高橋:淡水・沿岸域におけるアミ類の摂餌生態 47
はじめに
太陽光による光合成産物に依存する生態系は,生きた
植物体を起点とする「生食連鎖」と,動植物の遺骸,排
池物ならびにその分解物などのデトライタスを起点とす
る「腐食連鎖」の二つに大別される.一般的に水界生態
系では陸上生態系に比べ生食連鎖の比重が高いが,陸域
からデトライタスが流入する陸水域や汽水・沿岸域では
腐食連鎖の占める割合が高くなる(Odum& Heald
1972, Begon et al. 1996).このような水域ではデトライ
タスから植物,動物などに至る多様な餌を摂る「雑食者」
が食物網の重要な構成員となる.これらの水域における
雑食者の代表的動物プランクトンとしてアミ類が挙げら
れる.
アミ類はエビ類に似た,通常体長が 1-2cm程度の甲
殻類である.外見はエビ類,あるいはオキアミ類に類似
してはいるが,分類学上はエビ類,オキアミ類の属する
ホンエビ上目(Eucarida)ではなく,ヨコエビ類や等脚
類などと同じフクロエビ上目(Peracarida)に分類され
る.フクロエビの名が示すとおり,すべてのアミ類成熟
雌は腹部に保育嚢を有し,ここに受精卵を瞬化するまで
保持する.原則として海産であり,沿岸から外洋,深海,
洞窟まで世界中の海洋から出現が報告されている.また
一部の種は海跡種として淡水域にも適応している.多く
の種では海底,あるいは何らかの構造物と密接な関係を
保ちつつ生活する「近底層性動物プランクトン(demer-
sal zooplankton)」であり通常のプランクトンネットで
採集される機会は少ないが,実際には汽水域,沿岸域な
どでは高い生物量を示し,多くの魚類や大型甲殻類の重
要な餌料となっている(Mauchline1980).
アミ類は周囲の餌環境にあわせて様々な餌料を摂食・
利用する能力をもち,その胃中からはデトライタス,底
生,浮遊性の動植物,唾上起源の動植物など多様な餌料
が普遍的に見出される(Mauchline 1980).多様な餌項
目を含むアミ類の食性は餌料の定量化を阻み,本動物群
の食物網内における位置づけを困難なものとしてきた.
この結果,アミ類が雑食性であるという見方に異論はな
いものの,食物網内で、果たす役割については様々な見解
が示されてきた.採集された試料個体の消化管を解剖し
顕微鏡で直接観察した初期の研究結果は,アミ類の多く
がデトライタス,動植物プランクトン,底生生物,陸上
植物由来物質など様々な物質を利用できる雑食者である
ことを明らかにしてきた(Mauchline1980).とりわけ,
ほとんどの種の消化管内に卓越した不定形の同定不能物
質(unidentifiedmaterial)の存在からアミ類は海底に沈
殿したデトライタスやこれに付随する微小生物などと魚
類とを直接結び、つける役割を担い,物質循環を短縮・能
率化に貢献しているという考え方が強調された(村野
1974, Mauchline 1980).一方,北米,北欧の淡水域で
は1950年頃から, サケ科魚類の餌料環境を底上げする
目的で,海跡種である Mysisrelictaを,本種が元来生息
していない場所へ移植する事業が盛んに行われてきた
(Lasenby et al. 1986).移植当初,研究者はM.relictaが
雑食性であることは認識していたが,動物プランクトン
に対する捕食の影響は小さいと考えた. しかし移植後,
湖に本来生息していた動物プランクトン群集組成の変
化,とくに Dゅhniaなどの枝角類が激減・消滅する現
象が報告されるようになり(Zyblut1970, Richards et
al. 197 5, Goldman et al. 1979), M. relictaの捕食がその
原因であることが次第に明らかになってきた(Morgan
1982, Nesler & Bergersen 1991).このような経緯から
1980年頃からは次第にアミ類の肉食の能力が注目され
るようになり,動物プランクトンに対する捕食速度,摂
餌選択性に関する研究が増えた.近年はこれらの知見を
ふまえ飼育実験や安定同位体比の分析から季節や生活史
を通して各々の餌料項目の相対的な重要性を評価し,食
物網内での位置を明らかにしようとする研究例が増えつ
つある(例えば Viherluotoet al. 2000, Johannsson et
al. 2001, Mulkins et al. 2002, Kibrige et al. 2003な
ど).
アミ類の生物学全般については Mauchline(1980)の
優れた総説が発表されているが,アミ類の摂餌生態につ
いての記述はわずかである.この総説の出版と前後し
て, Mysisrelictaの移植に関連したアミ類の摂餌生態に
対する関心が高まり,加えて近年の観察・分析技術の発
達によってアミ類の食性に関する知見は大幅に増えた.
その結果,徐々にではあるが本動物群が多様な食性を通
じて食物網内においてどのような役割を担っているのか
という点についても明らかになりつつある.本論文では
近年の研究結果を中心に,アミ類の食性と摂食過程に関
する知見を整理,解説し,アミ類が摂餌活動を通じて水
圏生態系に果たす役割について考える.
アミ類は分類学上,主に外洋深海域に生息するロフォ
ガスター目(Lophogastrida)と,ほとんどの沿岸種,淡
水種を含むアミ日(Mysida)に大別される.両者間では
付属肢の形態の差異から摂餌方法,摂食生態が異なる可
能性がある(Hessler1985).本稿ではアミ類の多くが属
するアミ目に関する知見について解説する.
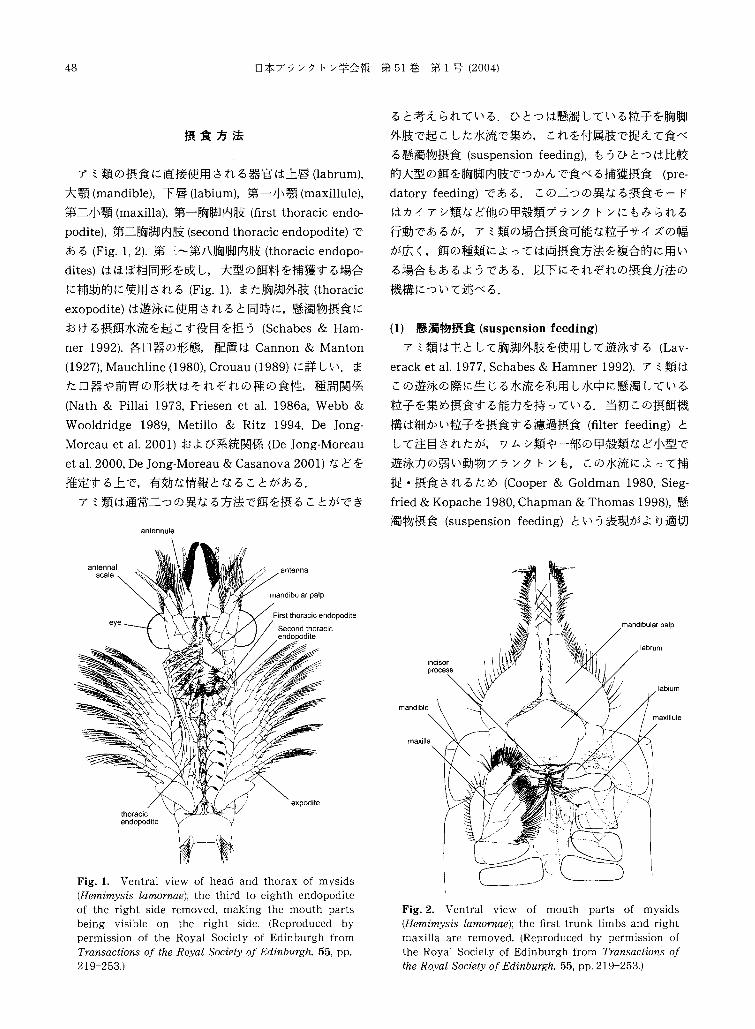
48 日本プランクトン学会報第 51巻第 l号(2004)
ると考えられている.ひとつは懸濁している粒子を胸脚
摂食方法 外肢で起こした水流で集め,これを付属肢で捉えて食べ
アミ類の摂食に直接使用される器官は上唇(labrum),
大顎(mandible),下唇(labium),第一小顎(maxillule),
第二小顎(maxilla),第一胸脚内肢(白rstthoracic endo-
podite),第二胸脚内肢(secondthoracic endopodite)で
ある(Fig.1, 2).第三~第八胸脚内肢(thoracicendopo-
dites)はほぼ相同形を成し,大型の餌料を捕獲する場合
に補助的に使用される(Fig.1).また胸脚外肢(thoracic
exopodite)は遊泳に使用されると同時に,懸濁物摂食に
おける摂餌水流を起こす役目を担う(Schabes& Ham-
ner 1992).各口器の形態,配置は Cannon& Manton
(1927), Mauchline (1980), Crouau (1989)に詳しい.ま
た口器や前胃の形状はそれぞれの種の食性,種間関係
(Nath & Pillai 1973, Friesen et al. 1986a, Webb &
Wooldridge 1989, Metillo & Ritz 1994, De Jong-
Moreau et al. 2001)およひ、系統関係(DeJong
et al. 2000, De Jong-Moreau & Casanova 2001)なと、を
推定する上で、,有効な情報となることがある.
アミ類は通常二つの異なる方法で餌を摂ることができ
antennule
thoracic endopodite
antenna
expodite
Fig. 1. Ventral view of head and thorax of mysids (Hemimysis lamornae); the third to eighth巴ndopoditeof the right side removed, making the mouth parts b巴ing visible on the right side. (Reproduced by permission of the Royal Society of Edinburgh from Transactions of the Royal Socie砂 ofEdinburgh, 55, pp. 219 253.)
る懸濁物摂食(suspensionfeeding),もうひとつは比較
的大型の餌を胸脚内肢で、つかんで・食べる捕獲摂食(pre-
datory feeding)である. この二つの異なる摂食モード
はカイアシ類など他の甲殻類プランクトンにもみられる
行動であるが,アミ類の場合摂食可能な粒子サイズの幅
が広く,餌の種類によっては両摂食方法を複合的に用い
る場合もあるようである.以下にそれぞれの摂食方法の
機構について述べる.
(1) 懸濁物摂食(suspensionfeeding)
アミ類は主として胸脚外肢を使用して遊泳する(Lav-
erack et al. 1977, Schabes & Hamner 1992).アミ類は
この遊泳の際に生じる水流を利用し水中に懸濁している
粒子を集め摂食する能力を持っている.当初この摂餌機
構は細かい粒子を摂食する議過摂食(filterfeeding)と
して注目されたが, ワムシ類や一部の甲殻類など小型で
遊泳力の弱い動物プランクトンも,この水流によって捕
捉・摂食されるため(Cooper& Goldman 1980, Sieg-
fried & Kopache 1980, Chapman & Thomas 1998),懸
濁物摂食(suspensionfeeding)という表現がより適切
labium
mandible
maxilla
Fig. 2. Ventral view of mouth parts of mysids (Hemimysis lamornae); the first trunk limbs and right maxilla are removed. (Reproduced by permission of the Royal Society of Edinburgh from Transactions of the Royal Society of Edinburgh, 55, pp. 219-253.)

高橋・淡水・沿岸域におけるアミ類の摂餌生態 49
であると思われる(Viitasalo& Rautio 1998).ただし以
下に述べるとおり,餌粒子捕捉から摂食に至る過程は粒
子サイズ、の大小によって異なるようだ.
細かい粒子を摂食対象とした, いわゆる「漣過摂食」
の機構については古くから相反する意見が出され混乱が
続いてきた.アミ類が「濃過摂食」によって非常に細か
い粒子を摂餌する能力をもつことを初めて明らかにした
のは Depdolla(1923)である.彼はライススターチ(直
径 39μm)を使って Praunusflexuosusが細かい粒子
を摂餌できることを実験的に証明した.彼の観察によれ
ば胸脚外肢によって体の両側に対称形に生じる水流は体
軸に対し前方約 30° の角度で第二触角鱗片(antennal
scale)方向に近づき合流した後,胸脚内肢の聞を後ろ向
きに流れ,この水流によって水中内に懸濁している粒子
が口器付近に集められる.また集められた粒子は第一,
第二胸脚内肢先端の刺毛と,第二小顎の羽状剛毛によっ
て捉えられ,これらの付属肢によって口部まで運ばれ
る.これに対し Cannon& Manton (1927)は Hemi-
mysis lamornaeを用いた観察結果から,胸脚外肢に
よって生じる水流は,前方に加え上下および左右から胸
部に向かつて流れるとした.ただし前方向からの水はア
ミの胸脚内肢の聞には,一切流れ込まないとした上で,
これに変わる摂餌水流として,それぞれの胸脚外肢の回
転運動によって体の側方から胸脚基部の聞に吸い込まれ
る流れを提唱し,これを「foodcurrentJとよんだ.彼ら
はこの摂餌水流は胸脚基部の聞を通った後,胸部下側中
央に体軸に沿って存在する「foodgroove」と呼ばれる
溝状の空間で合流した後,胸部後方から前方に向かう流
れとなって口部に達すると考え,摂餌水流が前方から流
れ込むとした Depdollaの観察結果を否定した.このの
ちCannon& Mantonの説は広く受け入れられ(Tatter-
sall & Tattersall 1951, Brown & Talbot 1972, Mauch-
line 1980),オキアミ類の漉過摂餌機構の説明にまで取
り入れられた(Mauchline& Fisher 1969).しかしなが
らAttramadal(1981)は, Depdollaおよび Cannon&
Mantonと同じ条件で観察を行い,アミ類胸部下側の
「foodgrooveJ内に見られる水流は,観察に十分な深さ
を持たない時計皿を用いた場合に限って生じる人為的現
象であり,自由に遊泳する個体においては前方向,後方
向のいずれの水流も観察されなかったと報告した.さら
にSchabes& Hamner (1992)は, ビデオ画像によって
A ntarctomysis sp., Acαnthomysis sculpta, Neomysis
rayii 3種の胸脚外肢が引き起こす水流の方向を観察し,
全ての種類において Cannon& Mantonが主張した側
方から胸部下側に流れ込む「foodcurren tJは一切認め
られず,胸部下側には前方からのみ水流が流れ込むこと
を報告した.またオキアミ類についても現在では瀦過摂
食水流は前方から後方に向かつて流れるとされ, Can-
non & Mantonの観察結果は支持されていない(Ham-
ner 1988).アミ類の「液過摂食」は,普遍的かっ主要な
摂餌方法(Tattersall& Tattersall 19 51)と目され,植
物プランクトンやデトライタスを効率よく摂食できるこ
とが実験的に証明されているが(後述),その機構につい
ては現在のところ,これ以上の知見はない.外見上アミ
類と類似しているオキアミ類では,その漉過摂食機構に
ついて詳細な研究例があるが(Hamner1988),両者間で
は遊泳方法ならび、に漉過水流の起こし方が異なっている
上に,胸脚の形態にも差異が認められることから,アミ
類は独自の漉過摂食機構をもっ可能性が考えられる.と
くに胸部下側に流れ込んだ水および餌粒子の動き,餌粒
子の捕捉・運搬方法と各付属肢の機能,餌粒子サイズが
これらに及ぼす影響などに関しては研究の余地がある.
アミ類の起こす摂餌水流は一部の動物性餌料を捕捉す
ることができる.ただし感覚・運動能力に優れたカイア
シ類成体などでは,アミ類の接近をいち早く検知し逃避
するため(Viitasaloet al. 1998),この水流によって捕捉
されるのはワムシ類,カイアシ類幼生,一部の枝角類な
ど,小型で遊泳力の弱い動物プランクトンに限られる.
水流によってアミ類の近くまで引き寄せられた餌生物は
胸脚内肢によって捕獲され摂食される(Metillo& Ritz
1993;捕獲摂食の項参照).また胸脚が起こす水流に対
して餌生物が示す逃避反応は,アミ類が餌生物を検知し
捕獲摂食行動に移るシグナルとなっている可能性を示す
観察結果もあることから(Viitasaloet al. 1998),摂餌水
流は様々なタイプの餌料を摂るアミ類にとって餌捕捉手
段であると同時に,次項に述べる捕獲摂食において餌を
探索する一種のソナーのような役目をもっているのかも
しれない.
(2) 捕獲摂食(predatoryfeeding, raptorial feeding)
摂餌水流によって粒子を捕らえる懸濁物摂食のほか
に,アミ類は比較的大型の餌を直接認知しこれを捕獲す
ることができる.アミ類の捕食対象となるのは主として
枝角類,カイアシ類などの動物プランクトンである.ア
ミ類はこれらの餌生物を物理刺激,視覚,化学物質など
によって知覚すると考えられる.
物理刺激はアミ類が餌生物を知覚する際に最も重要だ
と考えられている(Cooper& Goldman 1980, Mur-
taugh 198la, Ramcharan et al. 1985, Viitasalo et al.
1998, Viherluoto & Viitasalo 2001 b).アミ類は体の前

50 日本プランクトン学会報第 51巻第 1号(2004)
方に良く発達した二対の触角(antennule,antenna)を
もち, 餌生物の発する物理的刺激を受容する. Crouau
(1986, 1989)は複眼の退化している洞窟性のアミ類An-
tromysis juberthieiを用いて,餌の発見過程における感
覚器官,とくに触角上に生えている刺毛(感覚毛)の役
割を,組織学的手法および行動観察に基づき考察し,物
理刺激が餌の発見に重要な役割を果たしていることを示
した.この物理刺激は,遊泳時に強い水流を起こす胸脚
外肢から十分離れている触角鞭節中央および末端部にあ
る感覚毛で主に受容される(Crauau 1989). Neomysis
integer では物理刺激によってカイアシ類 Eurytemora
を検知する距離が 0.92cmであることが報告されてい
る(Viitasaloet al. 1998).
またアミ類の多くは体の前方に良く発達した一対の複
眼を持っている.この複眼は捕食者の発見とならび餌生
物の知覚に一定の役割を果たしている可能性がある.ア
ミ類の複眼は約 600から 5000の個眼より構成され
(Hallberg 1977),離れた距離,とくに前方やや上方にあ
る小型の粒子を高い解像度で探知するのに最適な構造で
あるとされている(Nilsson& Modlin 1994). Ramcha-
ran & Sprules (1986)は光条件を段階的に調節した環境
下で Mysisrelictaに枝角類, カイアシ類を捕食させた
結果,明条件下(0.43Wm 2以下)では暗条件下よりも
捕食速度が高くなることを報告し,視覚が本種の捕獲摂
食に重要であることを示した.完全な暗黒下で摂食速度
が低下する現象は Neomysisamericana, Mysidopsis bi-
gelowiでも報告されており,例えばM.bigelowiがカイ
アシ類 Acartiαtonsaを捕食する際,暗条件下(24D)で
は明暗周期 (12D:12L)と比べ,漏水速度が約 50-90%
低下する(Fulton1982b).一方, M.relict,αを含め Neo-
mysis mercedis, Praunusβexuosus, Tenagomysis chil-
toniなどでは光の有無はアミ類の摂食速度に影響を与え
ないという報告も多い(Siegfried& Kopache 1980,
Cooper & Goldman 1982, Chapman & Thomas 1998,
Viherluoto & Viitasalo 2001 b).また,北部バルト海の
Mysis mixtaのような比較的大規模な日周鉛直移動を行
う種では,光はむしろ捕食速度を低下させることも報告
されていることから(Gorokhova& Hansson 1997, Vi-
herluoto & Viitasalo 2001 b),アミ類の餌生物捕獲にお
ける視覚の果たす役割は環境や種によって異なると考え
られる.
アミ類の触角上には物理刺激を受容するものとはタイ
プの異なる感覚毛があり,化学物質を受容することがで
きると考えられている(Crouau1989, Johansson &
Hallberg 1992).タスマニア沿岸に生息する Anisa-
mysis mixta australis, Paramesopodopsis rufa, Tenaga・
mysis tasmaniaeの3種は,グリシンあるいは塩酸ベタ
インとアンモニアの混合物に反応して摂餌行動を示すこ
とが報告されている(Metillo& Ritz 2003).嘆覚刺激物
質を感知した際の各種の反応パターンは,化学物質の種
類やその混合比に依存した種特異性を示すことから, 3
種が同所的に共存していく上で化学刺激による餌の食い
分けが重要な要素となっている可能性が示唆されている
の1etillo& Ritz 2003).
これらの刺激を受容したアミ類は餌生物に対する接
近・攻撃行動に移る. 肉眼による観察では, Mysisre-
lictaやRhoραloρhthalmusterranatalisが短い距離 (1-
5cm程度)を前方あるいは側方へ飛びついて餌を捕獲
する様子が報告されている(DeGraeve& Reynolds
197 5, Wooldridge & Bailey 1982, Ramcharan et al.
1985).また Neomysisintegerでは執掛に餌を追跡する
一方で, 水槽の底に留まり頭上の餌を捕る「待ち伏せ」
(ambush mode)を示すことも報告されており(Vii-
tasalo et al. 1998),アミ類が動物プランクトン捕食に際
して多様な捕獲法を駆使していることがうかがわれる.
しかしながらアミ類の餌生物探索から攻撃に至る過程に
は依然不明な点が多い.この理由として,多くのアミ類
で餌生物の探索から攻撃までが一連の遊泳行動の中で行
われるため,各捕食過程を詳細に区別することが難しい
点が挙げられる(Ramcharanet al. 1985).今後,実際に
餌生物を検知(発見)している距離(detectiondistance)
の測定や,物理,視覚,化学の各刺激がこの距離に与え
る影響など、について研究の進展が望まれる.
アミ類の動物プランクトンに対する捕獲から摂食に至
る過程と各付属肢の機能については Paramesoρadopsis
ruf aが枝角類を捕食する様子がビデオ撮影によって解
析されている(Metillo& Ritz 1993). P. rufaは遊泳中,
胸脚内肢を胸部下側に押しつけ体を流線型に保っている
が,餌が近づくと第二~八胸脚内肢が下部に向かつて一
斉に伸張する.この急激な動きは餌生物の接近によって
誘発され,胸脚内側の流体密度の低下を引き起すことか
ら,結果として餌が胸脚内肢の聞に吸い寄せられると考
えられている.餌の捕獲に直接関わるのは主に第二,第
三胸脚内肢である.これより後方の胸脚内肢は通常は捕
獲に関与しないが,獲物が小型の場合は後方の胸脚内肢
でも捕獲されることがある.アミ類の胸脚内肢に生えて
いる刺毛は数が少なく柔軟であることから,オキアミ類
でみられる feedingbasket (Hamner 1988)形成のよう
な機械的(力学的)機能より,むしろ感覚器としての機
能を担い(Crouau1989),餌生物の接触や動きを探知す

高橋:淡水・沿岸域におけるアミ類の摂餌生態 51
ることで,捕獲を助けていると考えられる.捕獲された
餌生物は,胸脚内肢によって前方へ受け渡された後,第
一,第二胸脚内肢によってしっかりと捕まえられ,第一
胸脚内肢と口器によって大顎の聞に押し込まれ,かみ砕
かれる.この際に,第三~八胸脚内肢は餌生物の逃避を
防ぐためアミ自身の流線型を保つようにカゴ(cage)型
を形成し(Siegfried& Kopache 1980, Metillo & Ritz
1993),大顎触髭(mandibularpalp)は前方からの餌の
逃亡を防ぐカゴのフタとして機能する(Metillo& Ritz
1993).
Paramesoρadoρsis ruf aは捕らえた枝角類を摂食する
際,頭部から好んで摂食し始めることから,最初に獲物
を動けなくすることで,その後の摂食をより容易にして
いる可能性が示唆されている(Metillo& Ritz 1993).一
方, Neomysismercedisがカイアシ類Eurytemoraを捕
らえた際,頭部から食べ始める確率は 50%であること
が報告されている(Siegfried& Kopache 1980).大顎
鋸歯(incisorprocess)によって引きちぎられた餌の破
砕およびその後の食道に至る過程と各口器の働きについ
ては Cannon& Manton (1927), Crouau (1989)に詳し
(3) その他の摂食方法
上述した二つの主要な摂餌方法はいずれも水柱内に懸
濁あるいは浮遊している粒子を摂食する点で共通してい
るが,アミ類は水柱以外からも餌を摂る能力をもってい
る.Hemimysis lamornae, Neomysis integerでは水槽内
で懸濁物が少なくなってくると底質に対して頭部を下に
垂直に接し,胸脚外肢で水流を起こし,沈殿しているデ
トライタスを上方に巻き上げ,これを漉過摂食する
(Cannon & Manton 1927, Parker & West 1979).また,
底質直上を遊泳しながら第一,第二触角を用いて沈殿し
ている泥やデトライタスなどを掘り起こし,これらの塊
を直接胸脚内肢によってつかみ取り摂食する行動がH.
lamornae, Gastrosaccus psammodytesで知られている
(Cannon & Manton 1927, Brown & Talbot 1972). N
integerでは自らが排池途中の糞粒を, 腹部を折り曲げ
直接摂餌することが報告されている(Parker& West
1979). Neomysis integerでは基質に繁茂した付着珪藻
を直接かじりとる行動も観察されている (Irvineet al.
1993).
餌料組成
現場でのアミ類の餌料を特定するために,採集された
標本の胃内容物の直接観察および生化学的な分析が行わ
れてきた.これまでの研究をまとめると, Mysidoρsis,
Rhopaloρhthalmus属など肉食性の強いグループが一部
に認められるものの(Mauchline1970b, Fulton 1982b,
Wooldridge & Bailey 1982),ほとんどの種類が動物,
植物,デトライタスを利用する雑食性であることを示し
ている(Table1).以下に主要な餌項目について解説す
る.
(1) 動物性餌料
動物性餌料で最も普遍的に胃中から出現するのは浮遊
性の小型甲殻類で,同定不能の甲殻類破片の大部分もこ
れに由来すると考えられる.浮遊性カイアシ類は淡水
域,汽水域,沿岸域を問わず重要な餌料となっており,
淡水域では Epischura,Diαρtomus, Limnocalαnus, Cy-
clopsなどが(Murtaugh1981a, Kinsten & Olsen 1982,
Grossnickle 1982),汽水, 沿岸域では Eurytemora,
Pseudodiaptomus, Acartia, Temora, Pseudocalanus
などが(Siegfried& Kopache 1980, Wooldridge &
Bailey 1982, Hansson et al. 1990a, Rudstam et al.
1992, Irvine et al. 1993, Aaser et al. 1995, Fockeday &
Mees 1999など),よく摂餌されている.カイアシ類
ノープリウス幼生は重要な餌料となっていることが多い
が(Bowers& Vanderploeg 1982, Jansen 1985, Irvine
et al. 1993, Aaser et al. 1995, Jerling & Wooldridge
1995, Takahashi & Kawaguchi 1998),一方で明瞭な
負の選択性を示すという報告もあり(Siegfried& Ko-
pache 1980, Murtaugh 1981a),餌料としての重要性は
アミ類の種や発育段階,および周辺の餌環境に大きく左
右されるようである(後述).枝角類は淡水域,汽水域で
重要な餌料となっており, Bosmina,Daρhnia, Chydorus
などがよく捕食されている(Lasenby& Langford
1973, Murtaugh 1981a, Jansen 1985, Irvine et al.
1993, Viherluoto et al. 2000).また飼育下の Neomysis
integerではフジツボ類ノープリウス幼生に対する捕食
が観察されている(Mauchline1971 b).
汽水域,沿岸域では底生性ハルパクチクス目カイアシ
類も比較的よく胃内容物中に出現する(Mauchline
1971 b. c, Siegfried & Kopache 1980, Johnston & La-
senby 1982, Fenton 1996, Takahashi & Kawaguchi
1998など).ヨコエビ類が胃内容物から見出される頻度
は必ずしも高くないが (Jansen1985, Johannsson et al.
2001),飼育下では頻繁に捕食する様子が観察されてい
る(Nath& Pillai 1973, Parker 1979, Wilhelm et al.
2002).多くのヨコエビ類は比較的大型であるため,捕

52 日本フ。ランクトン学会報第 51巻第 l号(2004)
食され胃内容物中に存在しでも,口器による破砕で原形
を失なっていたり,柔組織だけが摂食されているなどの
理由によって見過ごされている可能性が高く(Parker
1979), Mysis relict,αでは餌料としての重要性を改めて
評価する研究も発表されている (Johannssonet al.
2001).南アフリカ河口域に生息する Rhoραloρhthal-
mus terranαtαlisは,同所的に出現するアミ類 Mesaρ0・
doρsis wooldridgeiの幼体を主な餌料としている(Wool-
dridge & Bailey 1982, Wooldridge & Webb 1988).ま
た飼育環境下では Anisomysis mix似 australis,Piαra-
mesopodoρsis rufa, Tenagomysistasmαniae, Metamysi-
doρsis elongata, M. relicta, R. terranαtalisなど多くの種
で共食い,とくに小型個体を捕食する現象が観察されて
いる(DeGraeve& Reynolds 1975, Clutter 1969, Wool-
dridge & Webb 1988, Johnston & Ritz 2001, Quirt &
Lasenby 2002).さらに多くの種で死亡した個体や傷つ
いた個体は速やかに他個体によって摂食されることが知
られ(Lucas1936,村野 1963,Green 1970, Nath &
Pillai 1973, Parker & West 1979, Yamada et al. 1995
など),雌の保育嚢から落出した匹なども摂餌対象とな
る場合がある(Parker& West 1979,佐藤・村野
1994).
ワムシ類は淡水域から汽水域にかけて Keratellα,
Kellicottia, Notholca, Brachionusなどが広く摂餌され
ており(村野 1963,Johnston & Lasenby 1982, Kin-
sten & Olsen 1982, Jansen 1985, Martinez & Berger-
sen 1991, Jerling & Wooldridge 1995, Fockedey &
Mees 1999など), とくに小型個体の餌料として重要性
が高い.有鐘繊毛虫は汽水域から沿岸域に生息する種に
おいて比較的よく見出されるが,優占することはない
(Jansen 1985, Viherluoto et al. 2000).微小動物プラン
クトンの多くは脆弱で固定液による保存が難しいため,
消化管内容物の直接観察によって検出することは難し
い.現場群集を餌料とした実験では南アフリカ汽水域に
生息する Mesopodoρsiswooldridgei, Rhoρalaρhthαl-
mus terranatalisが直径 10-60μm程度の繊毛虫,鞭毛
虫などを,ある程度摂食できることが示されている (Jer-
ling & Wooldridge 1995, Froneman 2001).
汽水域に生息する Neomysis属の種からは昆虫類, ユ
スリカ幼虫(Lasenby& Langford 1973, Johnston &
Lasenby 1982, Jansen 1985)や半題類幼虫(Fockedey
& Mees 1999)などが見出されている.動物性餌料とし
てはこのほかに,有孔虫類(Fenton1996, Takahashi &
Kawaguchi 1998),線虫類 (Jansen 1985),環形動物
(Johnston & Lasenby 1982, Irvine et al. 1993),軟体動
物の幼生(Takahashi & Kawaguchi 1998),介形類
(Friesen et al. l 986b),ダニ類(Fockedey& Mees
1999),魚類仔魚(Wooldridge& Bailey 1982)などの出
現が報告されているが,頻度,量ともに重要性は低い.
形態が特徴的なカイメン類の骨片は消化管中から比較的
よく見出されるが量的な寄与は極めて低い.おそらくは
底泥やデトライタスなどを摂食する際に一緒に摂食され
たものと考えられる(Mauchline 1968).また大陸棚域
では複数の種がスケソウダラの卵を捕食していることが
免疫学的手法により確かめられている(Baileyet al.
1993).
(2) 植物性餌料
珪藻類は殻が消化されにくいこともあって,淡水域か
ら沿岸域までほとんどの種の消化管内から普遍的に見出
される.一度の調査で 40種以上の珪藻が同定されるこ
ともあるが(Lasenby& Langford 1973, Baldo Kost
&Knight 1975),よく摂食される種類は比較的限られて
おり, 淡水域・汽水域では Melosira, Coscinodiscus,
Synedra, Cyclotellα,Naviculaなど(Lasenby& Lang-
ford 1973, Baldo Kost & Knight 1975, Siegfried &
Kopache 1980),沿岸域では Navicula, Cocconeis,
Chaetoceros, Coscinodiscus やその他中心日の種など
(Mauchline 1967, 1968, 1971a, b, Yamada et al. 1995,
高橋未発表)が頻繁に見出される.また南アフリカ砂浜
域では砕波帯周辺に特異的に生息する珪藻 Anaulusbi-
rostratusがMesopodopsiswooldridgeiや Gastrosaccus
ρsammodytesの主要な餌料となっている(Webbet al.
1987, 1988, Wooldridge 1989, Webb & Wooldridge
1989).珪藻類に多く含まれる n-3系高度不飽和脂肪酸
はとくにアミ類の再生産に重要な働きをすることが知ら
れており(Kreegeret al. 1991, Yamada et al. 1995),た
とえ少量でも餌料としての重要性は高いと考えられる.
渦鞭毛藻は汽水域から沿岸域に生息するアミ類におい
て見出されるが,優占することは希である. Ceratium,
Peridinium, Prorocentrum, Dinoρhysis, Phαlacromaな
どが比較的よく摂食されている(Mauchline1967, Fen-
ton 1996, Takahashi & Kawaguchi 1998, Viherluoto
et al. 2000).緑藻類は淡水域から汽水域にかけて生息す
るアミ類より見出され, Scenedesmus,Planctonema, Pe-
diastrum, Manoraρhidium, Oocyctisなどが同定されて
いる(村野 1963,Baldo Kost & Knight 1975, Bremer
& Vijverberg 1982 , Irvine et al. 1993, Fockedey &
恥1ees1999, Viherluoto et al. 2000).藍藻類は汽水域に
生息する種で報告されている(Bremer& Vijverberg

高橋 淡水・沿岸域におけるアミ類の摂餌生態 53
1982, Viherluoto et al. 2000).バルト海では Mysis
mixta, Mysis relicta の消化管から Aphanizomenon,
Merismoρedia, Nodular切などが見出されているが(Vi-
herluoto et al. 2000),これらの藍藻類はしばしば有毒
であるうえに,繊維状の藻体が口器に絡まり摂餌行動を
妨けるため好適な餌料となることは少ないと考えられる
(Bremer & Vijverberg 1982, Engstrom et al. 2001 ).
藻類以外の植物性餌料として花粉,胞子類などが淡水
から沿岸域に生息するアミ類において頻繁に見出される
が量的に卓越することはほとんどない(村野 1963,
恥1auchline1967, 1970a, 197lb, c, Fenton 1996, Fock-
edey & Mees 1999).
(3) “デトライタス”または同定不能物質
ほとんどのアミ類の消化管中には同定できない不定形
の物質が見出され,これらは総じてデトライタスとして
扱われることが多い(Table1).この同定不能物質(un-
identified material)は,しばしばアミ類消化管内容物の
大部分を占め,餌料源として重要であると考えられる
が,おそらくその起源は多様であり,栄養的価値もまた
その由来によって異なると考えられる.
アマモに代表される海草類,亜熱帯域のマングローブ
や塩生沼沢周辺に生育する維管束植物はデトライタスの
重要な供給源である(Mann1988).汽水域から沿岸域の
アミ類の消化管内容物中には同定不能物質中に混じって
繊維質の形状を残した維管束植物の破片が広く認められ
る(Mauchline1967, 1969, 1970a, 1971a, c, Odum &
Heald 1972, Johnston & Lasenby 1982).また Neo-
mysis americanαでは消化管中に優占するデトライタス
が塩生沼沢に生育するイネ科植物勾artinaalternijlora
に由来することが免疫学的手法により確認されている
(Zagursky & Feller 1985).維管束植物に多く含まれる
セルロース系の構造炭水化物は難消化性であるが,
Mysis stenolepisはこのセルロースを 3050%の同化効
率で摂取できることが知られている(Foulds& Mann
1978).この同化効率は微生物が付着したセルロースよ
り滅菌したセルロースで高いことから,本種は基質上に
生育するバクテリアではなくセルロースそのものを栄養
源としていると考えられている(Foulds& Mann 1978).
Weinright & Mann (1982)は抗生物質を添加した海水
中では M.stenoleρisのセルロース同化能力が失われる
ことを示し,セルラーゼ分泌に消化管内細菌が関与して
いる可能性を示唆した.しかし Friesenet al. (1986a, b)
は,同種の消化管内に認められる細菌数は,餌料である
デトライタスに付着しているものとほぼ同数であり,さ
らには消化管の形態そのものも腸内細菌を共生させるに
は不適であることを示し,アミ自身がセルラーゼを分泌
する以外,本種のセルロース分解能力を説明することは
できないと結論した.
大型藻類の破片は沿岸域に生息する種で高い頻度で見
出される(Hobson& Chess 1976, Fenton 1996).大型
海藻類や浮滋・底生性の微細藻類に由来するデトライタ
スは,維管束植物由来のデトライタスに比べ繊維が少な
く窒素含有量が高いため,デトライタス食者にとって栄
養価値が高い(Mann1988).地中海の海草藻場周辺に生
息する Lゆtomysis属数種では,炭素安定同位体比分析
結果から,海草由来炭素よりもプランクトン,あるいは
底生藻類由来の炭素が餌料源としてより重要であること
が示されている(Dauby1995).
熱帯域ではデトライタスの供給源としてサンゴ類が細
胞外に分泌する粘液物質が重要であり,それに対する摂
食速度や同化効率がアミ類でも調べられている(Gott・
fried & Roman 1983, Mullin & Roman 1986).米国フ
ロリダに生息する Mysidiumintegrumはサンゴの分泌
した粘液とこれに付着したバクテリアを摂食,同化する
能力をもち,この餌料のみで 2ヶ月以上飼育可能である
ことが報告されている(Gottfried& Roman 1983).
消化管中の査定不能物質中には, しばしば砂やシルト
などが混じって認められるため底土も餌料として摂取さ
れていると考えられる.Mysis relict,αやNeomysisinte-
gerなどでは飼育下で実際に底土を摂食することが報告
されている(ValKlump et al. 1991, Roast et al. 2000).
Fockedey & Mees (1999)は汽水域に生息する N.inte-
gerの消化管中に卓越する綿状の不定形物質の組成を元
素分析装置(EDAX)で分析した結果,この物質が水柱に
懸濁する珪素,アルミニウムを主体とする粘土(clay
minerals)であることを明らかにし, エネルギー源とし
ての餌料価値は低いと結論している.おそらく底土摂食
の利点は,底土中に生息する生物を摂取できる点にあ
る.実際に M.relictaやNeomysismercedisなどは,表
在性の動植物が混在する底土から有機物に富んだ餌料だ
けを選択的に摂餌できるようだ(Lasenby& Langford
197 3, Johnston & Lasenby 1982).また現場の底土を飼
育下のアミ類に与えることでアミ類の生残,成長に好影
響を与える例もしばしば報告されている(村野 1963,
Cripe et al. 2000). 村野(1963)は Neomysisawat-
schensisの飼育に際し諏訪湖の底土を敷いた水槽では
無給餌にもかかわらず,高い生残率を示すことを報告
し,原生動物や藻類を含んだ底土が本種の餌料として重
要であると結論している.

第 1号(2004)第 51巻日本プランクトン学会報54
Habitat
Species
J<'reshwater
My sis阿 佐ta
M1人口Srel町ta
M戸日開lieta
My sis問 l町ta
Mys is問l町加
Nee mysis intermedza
Neomysis mercedis
Brackish & Estuarine water
Afysidop幻saim}叩
Mysis mixta
Mysis mi叫α
凡f}sismix乙α
M,sis問 lieta
Neomysis integer
Neomysis integer
Neomysi.s integer
Neomysis integer
N即 mysisinteger
Neomysis m開 cedzs
Neom戸時間ercedis
Necmysis mercedis
RhopalophthαImus terranatal!s
Taphromys日bο叩mani
Coastal water
Acanthοmysis mit.suku門Z
Acαnthomysis sculpta
Ani.scmysi.s m目的 australis
A町haenmys日jap仁nica
Archaeamysis kokuboi
t:rythれap.sele島即日
Eηthrops se庁ata
Gastrosacces psammodytes
Leptomysis E杭αcilis
Mesopodopsis uooldridgei
MysidοPsis bigelowi
Mysidopsis dide砂hys
MysidoPsis gibbc sa
Mys日stenalep日
Necmysi.s ame円cana
Neomysis ame門cana
Neorrり.sisinteger
f勺ramesopadopsis叩 fαPara mys日α開 ncsa
Praunus fiexuοSUS
Praunus neglectu.s
Schistomysis kernillez
Schistomysis ornata
SchistDmysis spritus
Siriella Pacifica
Tenagcmysi.s tosmaniae
* Other crustaceans contain unidentified crustacean fragments, ostracods, amphipods, and mysids. 料 Otheraquatic animals contain forminiferans, sponge spinule, nematods, mollusc larvae, annelids, and fish larvae.
Diets of mysids from freshwater, brackish, and coastal marine habitats. Relative importance Table 1.
Tintinnids Rotifers
十件
+付
um汁
+
Pollen
十十
++
Vascular
plant detritus
Macrophyto
fragments
Dmo
flagellates
Blue green
algae
Green
algae
十+
+件
++
Diatoms
川廿刊附
U竹川廿掛川市
Algae
十+
U廿
件
付
朴
+
+
川
市
川
村
山
市
川
竹
川
竹
+
件
什
+
十
+
+
+
+
++
+十
+
+
+
十件
十+
制
+
榊
掛
川
怖
+十
+
十件
#
++
件
十件
+朴
+件
品市川市川市
+件
十+
+什
仲山市川竹
++
汁
判+
+
十件
IH
+仲
+件
什
U廿川村山市
川廿汁川計什廿+
什
什
++
+件
榊
竹
川
竹
十
+
+!
+
十
十
十
U廿H廿+川市川竹川汁
+
十
++
++
掛
川
市
+
掛
川
竹
掛
川
市
川
廿
件
+
H汁
十
川市川計+
+朴
+
+
+
以
内
十
川
市
+特
十+
+併
+
+
+
+
十件
十朴
++
十件
+
++
Mysis relictaにおいて,デトライタスが存在しない餌料
環境下で消化管中に同定不能物質が検出されることを認
め,この同定不能物質が他餌料の消化物であると結論し
ている.消化管中の同定不能物質の由来を推定する場合
にはその形状や色,同時に出現する餌料などに注意する
必要があるだろう(Fockedey& Mees 1999参照).
アミ類に摂餌された餌料は大顎や胃内歯によって細か
くすり潰されるため,消化管中に見られる同定不能物質
のうち,少なくともその一部はデトライタスではなく甲
殻類や藻類の消化物や内容物であるという可能性も指摘
されている(Rybock1978, Siegfried & Kopache 1980,
Jansen 1985, Rudstam et al. 1989, Fockedey & Mees
1999).飼育環境下で珪藻を摂食させた Mesoρodopsis
wooldridgeiの消化管内容物は, その大半が同定不能の
物質となる(Webbet al. 1987).また Rybock(1978)は

55 高橋: j炎水・沿岸域におけるアミ類の摂餌生態
of food item is expressed by frequency of occurrence;州: Common,*: Occasional, +: Rare.
Unidentified
material
Other aquatIC
animals料
Other
crustaceans*
Harpacticoid
cope pods References
榊
川
廿
川
計
川
竹
川
廿
Lasenby & Langford 1973
Lasenby & Langford 1973
Johann58nn et al. 2001
Tattersall & Tattersall 1951
Rybcck 1978
Murano 1963
Murtaugh 1981a
Sediments
十い
+糾
++
川
村
山
材
料
Insects
++
川
廿
川
廿
川
廿
Cladocerans
十件
掛
川
廿
川
村
十
川
和
Copepcds
Odum & Heald 1972
Hansson et al. l 990a
Rudstam et al. 1989
Viherluoto et al. 2000
Viherluoto et al. 2000
Aaser et al. 1995
Irvine et al. 1993
Fockedey & Mees 1999
Bremer & Vijverberg 1982
Jansen 1985
Baldo Kost & Knight 1975
Siegfried & Kopache 1980
Johnston & Lasenby 1982
Wooldridge & Bailey ! 982
Odum & Heald 1972
++
十+
+件
十+
川
市
川
市
川
市
以
情
川
叶
川
情
川
市
+
川
市
川
市
川
竹
+
十
+
+
+
+十
+
+
十
十+
十件
+
紛
糾
十
十
+
川
市
掛
川
市
十
+
+十
+仲
+件
+
+
+
川
竹
川
市
川
情
川
市
川
前
Mm山
村
山
村
山
村
山
村
山
材
料
Yamada et al. 1995
Hobson & Chess 1976
Fenton 1996
Takahashi & Kawaguchi 1998
Takahashi & Kawaguchi 1998
Mauchline 1968
Mauchline 1968
Webb eta! 1988
Mauchline 1969
Webb & Wooldridge 1989
Fulton 1982b
Mauchline l 970b
Mauchline l 970b
Friesen et al. l 986b
Fulton l 982b
Zagursky & Filler 1985
Mauchline 197lb
Fenton 1996
Mauchline 197la
Mauchline 197lc
Mauchline 197lc
Mauchlinc 197ld
Mauchline l 970a
Mauchline 1967
Hobson & CheS> 1976
Fenton 1996
++
+仲
+仲
+仲
掛
川
汁
川
村
tH
+件
++
十件
um掛
川
市
柵
川
市
川
市
川
市
川
愉
+
川
市
川
市
川
市
川
計
川
市
川
情
山
市
川
市
山
前
+
川
市
十十
+
+
+
+
件
掛
け
竹
川
市
川
市
H件
州
州
川
村
山
村
十+
+件
+什
+
+糾
+軒
tit
IH
十什
++
十件
+仲
榊
以
川
以
川
以
山
村十
十+
十件
+計
十軒
+ 十件
+件
十+
* +件
十件
べ,夜間に上層に浮上し水柱内で枝角類やワムシなどの
動物プランクトンを捕食する(Lasenby& Langford
1973).一方,米国ミシガン湖のM.relictaは夏季夜間,
亜表層クロロフィル極大層に浮上し,植物プランク卜ン
を摂食する(Bowers& Grossnickle 1978, Grossnickle
1979).バルト海に生息する Mysismixt,αは昼間,海底
でデトライタスを食べ,夜間は水住内で枝角類やカイア
シ類を捕食している(Rudstamet al. 1989).また三陸海
岸砂浜域砕波帯に生息する Archaeomysiskokuboiは,
摂餌活動の変化
(1) 摂餌日周性
アミ類では, しばしば昼夜間で摂餌活性や餌料内容が
変化する例が報告されている.これまでの報告では夜間
に摂餌活動が活発になる例が多く,その大部分は鉛直・
水平的な移動を伴う(Table2).米国ス卜ニー湖に生息す
る Mysisrelictaは昼間深層に留まりデトライタスを食
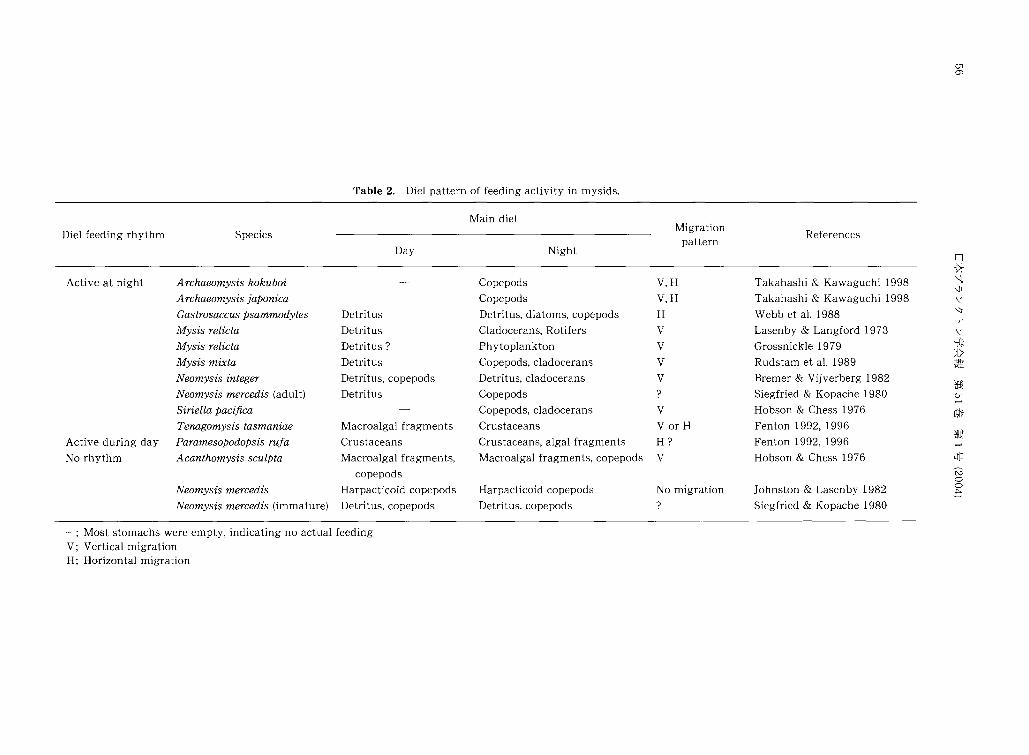
01 σコ
Diel pattern of feeding activity in mysids.
Main diet
Table 2.
Referenc巴sMigration
pattern Species Diel feeding rhythm
司、サリ判。AWV\守
7
ぃ、二村山町世間
制巴山附
制州
Hd
(
N
。。品)
Night
Active at night
Active during day
No rhythm
Takahashi & Kawaguchi 1998
Takahashi & Kawaguchi 1998
Webb et al. 1988
Lasenby & Langford 1973
Grossnickle 1979
Rudstam et al. 1989
Bremer & Vijverberg 1982
Siegfried & Kopache 1980
Hobson & Chess 1976
F巴nton1992 1996
Fenton 1992 1996
Hobson & Chess 1976
V,H
VH
H
v v v v つ
v Vor H
H?
v
Cope pods
Copepods
Detritus, diatoms, copepods
C!adocerans, Rotifers
Phytoplankton
Copepods, cladocerans
Detritus, cladocerans
Cope pods
Copepods, cladocerans
Crustaceans
Crustaceans, algal fragments
Macroalgal fragments, copepods
Day
Detritus
Detritus
Detritus?
Detritus
Detritus, copepods
Detritus
Macroalgal fragments
Crustaceans
Macroalgal fragments,
copepods
Harpacticoid copepods
Detritus, copepods
Archaeomysis kokuboi
Archaeomysis japonica
Gastrosaccus psammodytes
Mysis relicta
Mysis relicta
Mysis mixtα
Neomysis integer
Neomysis mercedis (adult)
Siriellaρacificα Tenagomysis tasmaniae
Paramesopodoρsis rufa Acanthomysis sculpta
Johnston & Lasenby 1982
Siegfried & Kopache 1980
No migration
つ
Harpacticoid copepods
Detritus, copepods
Neomysis mercedis
Neomysis mercedis (immature)
←; Most stomachs were empty, indicating no actual feeding
V; Vertical migration H; Horizontal migration

高橋 淡;J<・沿岸域におけるアミ類の摂餌生態 57
昼間岸に近い波打ち際近傍海底に留まり全く摂餌活動を
行わず,夜間に沖側に泳ぎ出しカイアシ類などを捕食す
る(Takahashi& Kawaguchi 1998).同じく砕波帯に生
息する Gastorosaccusρsammodytesは昼間,珪藻やデ
トライタスを主に摂取し,夜間になると肉食性を強め,
水住内に泳ぎ出してカイアシ類を捕食する(Webbet al.
1988).
一方,夜間よりも昼間に摂餌活動が活発化する報告が
一例のみ知られている.タスマニア沿岸に生息する
Pαramesoρodopsis ruf aは比較的肉食性の強い種であ
ると考えられているが(Metillo& Ritz 1994),その消化
管充満度は夜間よりも日中および日没直後に高く,昼間
活発にカイアシ類を捕食していると考えられている
(Fenton 1996).
明瞭な摂餌リズムを示さない場合もある.カナダ西岸
河口域に生息する Neomysismercedisは底生性のハル
パクチクス目カイアシ類を主な餌料としているが,昼夜
間とも摂餌内容に差が見られない (Johnston& Lasen-
by 1982).またカリフォルニアのジャイアントケルプ藻
場に生息する Acanthomysissculρtaの消化管は昼夜と
も,その藻体由来のデトライタスやカイアシ類で満たさ
れている(Hobson& Chess 1976).また南アフリカの
Mesopodopsis wooldridgeiでは,消化管色素の経時変化
に日周性が認められないことが報告されている(Frone-
man 2001).
発育段階によって摂餌リズムが変化する例も知られて
いる. 米国サクラメントの運河に生息する Neomysis
mercedis 成体は夜間になるとより活発にカイアシ類を
捕食するが,未成熟個体ではこのような変化を示さない
(Siegfried & Kopache 1980).一方, Mysisrelicta, Ar-
chaeomysis kokuboi, Archaeomysis jaρonicaなどでは
発育段階に関係なく,全ての個体が夜間に摂餌が活発化
する摂餌リズムを示す(Grossnickle1979, Takahashi
& Kawaguchi 1998).
(2) 季節変化
アミ類は利用できる餌料範囲が幅広いため,餌料の季
節変化は原則として周辺の餌料環境の変化に左右される
と考えられる.植物プランクトンブルームが起こる春季
には珪藻の重要性が高くなるという報告が多い.サクラ
メント運河に生息する Neomysismercedぬでは春に珪
藻 (Melosira)の優占度が高くなる(Siegfried& Ko-
pache 1980).同様に米国オンタリオ湖の Mysisrelict,α
では春季同化した炭素の約半分が珪藻に由来しているこ
とが報告されている (Johannssonet al. 2001).また植
物プランクトンが減少する夏季にはワムシ類,枝角類,
カイアシ類などの動物プランクトンを中心とした動物性
餌料の重要性が高くなる(Siegfried& Kopache 1980,
Arndt & Jansen 1986,恥1artinez& Bergersen 1991,
Viherluoto et al. 2000).動植物の生物量が減少する冬
季にはデトライタスの重要性が高くなるという報告が多
い(BaldoKost & Knight 1975, Arndt & Jansen 1986,
Zagursky & Feller 1985).一方,餌料源の供給が安定し
ている種,個体群では,明瞭な季節性を示さないことも
ある (Johnston& Lasenby 1982, Fenton 1996).カナ
ダ西岸河口域に生息する N mercedisは通年にわたり底
生性ハルパクチクス目カイアシ類を主な餌料としている
(Johnston & Lasenby 1982).また窒素安定同位体比の
解析より,霞ヶ浦のNeomysisawatschensisの中・大型
個体は年聞を通じて植物プランクトンを主体とする粒状
有機物を主な餌料としていると推定されている(Toda
& Wada 1990;ただし動物プランクトン捕食の可能性
について Hanazato1990も参照).
アミ類の世代時聞が数ヶ月に及ぶ場合,餌料の季節変
化は成長に伴って発達する餌料獲得能力に強く影響され
る. Viherluoto et al. (2000)はバルト海北部の Mysis
mixt,αの成長期(6-9月)における餌料組成の変化を,ア
ミ類の発育段階および餌料環境を考慮に入れて詳細に解
析した.M. mixtaの小型個体は 6月頃に出現するが,こ
の時期には主に海底近くで摂餌を行い,海底上に沈殿し
た植物プランクトンや底生性珪藻を餌料として成長す
る.その後, 7-8月になると成長に伴って徐々に肉食性
を強め,海底の餌に加え水柱内でワムシ類や枝角類,カ
イアシ類などの甲殻類プランクトンを捕食するようにな
り, 9月には浮遊性のカイアシ類を主な餌料とするよう
になる.ただし逃避能力の高いカイアシ類を捕食できる
ようになるには体長 8-11mm程度まで成長する必要が
あり,この大きさに達しない時期には水住内にカイアシ
類が卓越するにもかかわらず,その餌料中に占める割合
は低;いレベルにとどまる(Viherluotoet al. 2000).
(3) 発育段階および性による変化
多くのアミ類では成長に伴って餌料の種類やサイズが
変化する.直達発生を行うアミ類では摂餌機構自体は大
きく変化しないことが予想されるため,この餌料変化は
主に個体の餌料要求あるいは餌料獲得能力の発達によっ
て決定されていると考えられる.
小型のアミ類は動物性の餌料を追跡,捕獲するために
十分な能力を備えていないため,生活史の初期には珪藻
類や小型の動物プランクトンが重要な餌料となる.南ア

58 日本プランクトン学会報第 51巻第 l号(2004)
フリカ汽水域に生息する Mesaρodoρsiswooldridgeiで
は,幼体(体長 4mm以下)はカイアシ類ノープリウス
幼生を摂食することができず,一方,成体 (15mm以
上)では lOμm以下の小型藻類を摂食する能力がない
(Froneman 2001 ).サクラメント運河に生息する Neo-
mysis mercedisでは餌料組成が成長に伴って珪藻やデ
トライタスからカイアシ類主体へ変化することが報告さ
れている(Siegfried& Kopache 1980).本種は大型個体
ほどよく甲殻類を捕食しており,体長 3mm以下の個体
では甲殻類の捕食は認められない.成体がカイアシ類を
主食とするバルト海のMysismixtaも,カイアシ類を捕
獲できるサイズ(体長811 mm)に達するまでは植物
プランクトンや小型の動物プランクトンを主餌料として
いる(Viherluotoet al. 2000).ノースカロライナ沿岸の
Neomysis americana, Mysidopsis bigelowiでは,消化
管内におけるカイアシ類の出現頻度が幼体ではそれぞれ
O,および 20%であるのに対し成体では 42,および
72%にそれぞれ増加する(Fulton1982 b).北米の
Mysis relicta,バルト海の M.mixta, Neomysis integer
などでは安定同位体比の分析によって,成長に伴う栄養
段階の上昇が認められている(Gorokhova& Hansson
1999, Branstrator et al. 2000).ドイツ汽水域の N.inte-
gerではワムシ類, カイアシ類ノープリウス幼生が小型
個体の重要な餌料となっている (Jansen1985, Arndt &
ロッパ汽水域の N.integerでは,すべての発育段階がカ
ラヌス目カイアシ類の成体およびコペポダイト期幼生を
主餌料としているが,ここでも小型のアミ類は,より小
型のカイアシ類を捕食する傾向があることが知られてい
る(Fockedey&恥1ees1999).
雌雄間で餌料組成を調査した例は少ないが,これまで
の報告からは雌雄差が認められたという例はない(Ta-
kahashi & Kawaguchi 1998, Fockedey &恥1ees1999).
ただし種によっては雌が雄に比べ大型に成長する場合も
あり,このような状況下では餌料組成に性差が生じる可
能性も考えられる(例えば Wilhelmet al. 2002).また
摂食速度に関しては種によって雌雄間で差が生じる例が
報告されている(後述).
アミ類の成長に伴った餌料種・サイズの変化は,各発
育段階個体聞の種内競争を緩和する上で重要な意昧をも
っと考えられる(Siegfried& Kopache 1980).さらに北
米海跡湖の Mysi冶relictaでは,一生を通じて複数の栄
養段階に所属するという性質が(Life history omni-
vory),餌生物に与える捕食圧を分散させ,結果としてプ
ランクトン食物網の安定化に寄与している可能性が示さ
れているのprules& Bowerman 1988).
同化効率
Jansen 1986).砂浜域に生息する Archaeomysiskoku- 摂食した餌料の総量中,生物活動(成長,再生産,脱
boi小型個体ではカイアシ類ノープリウス幼生が重要で 皮,呼吸)に費やされた割合を示す同化効率は餌料の種
あり,成長すると浮遊性カイアシ類を主な餌料とするよ 類によって変化する. 植物性餌料ではこれまでに Neo-
うになる(Takahashi& Kawaguchi 1998).一方,ヨ- mysis mirabilisが渦鞭毛藻(Pechen'-Finenko& Pav-
Table 3. Assimilation efficiency in mysids on various diets.
Species of mysids Diet Assimilation
References efficiency (%)
Plant Neomysis mir,αbilis Gymnodinium kowalevskii 83-87 Pechen'-Finenko & Pavlovskaya 1973
Neomysis integer Chloγella vulgaris 80-90 Arndt & Jansen 1986 Neomysis integer Phytoplankton 62-77 Arndt & Jansen 1986
Mysidae sp. Phytoplankton 90 Clutter & Theilacker 1971
Animal Archaeomysis kokuboi Artemia nauplii 74 Takahashi (unpublished)
Archaeomysis japonica Artemia nauplii 74 Takahashi (unpublished)
Mysis relicta Daphnia pulex 85 Lasenby & Langford 1973 Mysis relicta Chrinomid larvae 82 Lasenby & Langford 1973
Neomysis mercedis Scottolana canadensis 82 Johnston & Lasenby 1982
Neomysis intermedia Daphnia 15-60 Toda et al. 1987 Others (detritus)
Neomysis integer Sediment (Ash-free fraction) 35 Roast et al. 2000 Mysidium integrurη Coral mucus 39 Gottfried & Roman 1983
Mysis stenolepis Cellulose 30-50 Foulds & Mann 1978
Mysis steno!ゅis Barley hay 20-35 Foulds & Mann 1978

高橋:淡水・沿岸域におけるアミ類の摂餌生態 59
lovskaya 1973), Neomysis integerが緑藻(Arndt&
Jansen 1986),アミ科未同定種が植物プランクトン群集
(Clutter & Theilacker 1971)をそれぞれ摂食した際の
同化効率が測定されており,その値は概ね 80-90%の範
囲内にある(Table3).
動物性餌料では MysisrelictaがDaρhniaおよびユス
リカ幼虫(Lasenby& Langford 1973), Neomysis mer-
cedisがハルパクチクス目カイアシ類 (Johnston& La-
sen by 1982), Archαeo明 ysis属2種がアルテミアノープ
リウス幼生(高橋 未発表)をそれぞれ摂食した場合の
同化効率が測定されており,その値はアミ類の種や餌料
に関係なく概ね 7080 %の同化効率を示している(Ta-
ble 3).一方, Todaet al. (1987)が見積もった Neomysis
awatschensis のDaphniaに対する同化効率(15-60%)
は動物性餌料に対する値としては低く,彼らはこの原因
として捕食時に破砕された餌料の一部が摂取されずに漏
出している可能性を示唆している.
デトライタスを形成する様々な非生体有機物に対する
同化効率も何例か報告がある. Foulds & Mann (1978)
はアマモ場に生息する Mysisstenolゆisに滅菌したセル
ロースおよび大麦の干し草を摂食させ,その同化効率を
それぞれ 30-50%,20 35%と見積もった. さらに彼ら
は,これらの餌料にバクテリアが付着していた場合,同
化効率が低下することを報告している. Neomysis inte-
gerの生物を取り除いた底泥の有機物画分に対する同化
効率は 35%と見積もられている(Roastet al. 2000).ま
たフロリダのサンゴ礁に生息する Mysidiumintegrum
がサンゴ分泌粘液を摂食した際の同化効率は 39%であ
ることが報告されている(Gottfried& Roman 1983).
同化効率は発育段階によっても変化する.Neomysis
integerの緑藻, 植物プランクトンに対する同化効率は
幼体で 77,90%であるのに対し,成体では 62,80%に
それぞれ低下する(Arndt& Jansen 1986).一方, Neo-
mysis awatschensisのDaphniaに対する同化効率は成
長するにつれ高くなり,成熟(雌)すると再度低下するこ
とが報告されている(Todaet al. 1987).
摂餌速度
様々な餌料に対するアミ類の摂食速度が室内,あるい
は野外での実験によって求められている.多くの動物プ
ランクトンの摂食速度と同様に,アミ類の摂食速度も一
般に餌密度の増加に伴って速くなり,ある濃度以上では
一定となる機能的応答(functionalresponse)を示す.
応答の形や速度は餌種のサイズや形態,逃避能力や化学
組成などに加え,アミ類自身が懸濁物摂食,捕獲摂食の
どちらの摂食モードを使用したかなどの点に影響を受け
る.
(1) 動物プランクトン
動物プランク卜ン関係では主に枝角類,カイアシ類お
よび現場で採集されたプランクトン群集を餌料として摂
食速度が見積もられている(Table4).アミ類の動物プ
ランクトンに対する漉水速度は最大で 1日あたり概ね
ト10リットルの範囲にあり,カイアシ類を餌料とした
場合,多くの種で一時間に 0.78個体程度を捕食する
(Table 4). Praunus fiexuosusの大型個体では 1時間に
Acartiaを 15個体捕食する場合もあるが(Viitasalo&
Rautio 1998),これらを含め体炭素量比では 1日に概ね
7-30%程度を摂食する能力があるようだ.カイアシ類
ノープリウス幼生に対する摂食速度は Neomysisinteger
で測定されており, 1時間で Eurytemoraのノープリウ
ス幼生 52個体を摂食する(Aaseret al. 1995).
枝角類に対する摂食速度は Neomysismercedis, Prau-
nusβexuosus, Tenagomysis chiltoniなど‘淡水域から汽
水域に生息する種で測定されている.枝角類に対する摂
食速度は総じてカイアシ類のそれより高い傾向を示す場
合が多く (Table 4),逃避能力の弱L、枝角類はアミ類に
とってより捕食しやすい餌であると考えられている
(Murtaugh 198la, Nero & Sprules 1986b, Viitasalo &
Rautio 1998,餌料選択性の項参照).
現場の動物プランクトン群集(原生生物を除く)を餌
料として摂餌速度を測定した研究は Mysisrelicta,
Mysis mixta, Neomysis integer, Mesaρodopsis wool-
dridgeiなどで知られている(Bowers& Vanderploeg
1982, Uitto et al. 1995, Mohammadian et al. 1997,
Froneman 2001, Viherluoto & Viitasalo 200la). M.
mix taは主にワムシ類および甲殻類プランクトン(枝角
類,カイアシ類)から構成される動物プランクトン群集
に対して最大で 40μgCind 1 h l,体炭素の 63%を一
日に摂食する能力をもっている(Viherluoto& Viita-
salo 200la).一方, M.wooldridgeiでは動物プランクト
ン群集(カイアシ類コペポダイト期が優占)に対する摂
食速度として 0.2μgC ind 1 h一l,一日の摂食量として
体炭素の 4%と,室内実験としては比較的低い値が報告
されているが(Froneman2001),これは餌密度の差に起
因していると考えられる.野外での摂餌速度も報告例が
ある. N integerは野外メソコズムにおいて動物プラン
クトン群集(ワムシ類,カイアシ類など)を 4μgCind 1
h l,体炭素量として 11%d一lを摂餌する(Uittoet al.

由
亡コFeeding rate of mysids on various types of food under experimental conditions. Values show maximum rates of adult animals in the literature. Table 4.
Carbon ingestion
rate
Predation/Grazing
rate Daily ration Clearance rate
References
% of body Cd 1 μgC mysid I h I Prey (cell) mysid 1
h I mlmysid 1 h 1
Species of mysids Diet/Prey Diet type
Webb et al. 1988
Webbetal.1987
Webbetal.1987
Lucas 1936
Engstrom et al. 2001
Engstrom et al. 2001
249
1082
626
217
5
5
253
253
146
165
3.3
3.2
4.0×106
4.0×106
2.3×106
6.5×106
Gastrosaccus psammodytes15
Mesopodopsis wooldrid耳ei15
Mesopodopsis wooldridgef5
Neomysis integer'
Mysis mixta:8
んfysismαta!B
ENす叫中て山V
7
い\
thw鶏
Engstrom et al. 200 I
Engstrom et al. 2001
Arashkevich & Dirts 1981
Mullin & Roman 1986
I
5 (26) ~·
く0.24
0.9
3.3
1.2
く0.01
21
43
15
10
Mysis mixta:8
Myszs mz.χta!B
Siriella jaltenszs
A nisomysis mullini
A naulus birostratusホl
A naulus birostratus*1
Asterionella glacilis*2
Nitzschia closterium:1
Aphαηizomenon fios-αquae Nodularia sphaerocαゆα
(Non-toxic)
Nodularia spmigena (Toxic)
Brachiomonas submarina
Tetraselmzs suecicα Phytoplankton
Plant
Diatoms
Blue・green algae
Green algae
皿山印戸川断
浦-
d(
NCE)
Viherluoto & Viitasalo 2001 b
Engstrom et al. 200 I
Viherluoto & Viitasalo 2001 b
Fulton 1982b
Fulton l 982a. b
Viitasalo & Rautio 1998
Chapman & Thomas 1998
Fulton 1982b
Fulton l 982a. b
Kinsten & Ols岳n1982
Cooper & Goldman 1980
Viitasalo & Rautio 1998
Aaser et al. 1995
Aaser et al. 1995
QUAせ円
。
1
1
30
11
3
0
6
7
つ臼
1414
12.0
14.7
10.0
6.1
7.7
46.2
1.4
11.0
5.9
1.5
19.8
30.8
10.4
13.9
2.4×105
5.6
4.9
4.7
1.2
1.5
15.0
1.5
0.7
1.1
0.7
3.3
6.0
52.0
8.0
く0.6
138
344
458
300
54
195
167
167
100
380
110
Mysis mixta
Mysis mixta:8
Praunus fiexuosus
Mysidops臼 bigelozιi
Neomysis americana!9 P 庁(8raunus nexuosus
Teηagomysis chiltoni
凡fysidopsisbigelowi
Neomysis ame門'cana:9
Mysis relicta110
Mysis relictal10
Praunusβexuosus18
Neomysis integer
Neomysis integer
A cartia bifilosa
Acα何日 bi)匝losa
A cartia bifilosa
A cartia tons a:'
Acαrtia tonsα:2
A cartia spp.
Calamoecia lucasi:3
Centropages spp. :z Centropages spp. !2
Cyclops scutifer4
Epischura nevαden.,is§
Eurytemoraα<!fin臼
Eurytemora affinis (nauplii)1
Eurytemora affinis
( copepodid)1
Eurytemora hirundoids II
Pseudodiaptomus hessei:5
Others
Animal
Cope pods
Siegfried & Kopache 1980
Wooldridge & Webb 1988
25
7
6.7
0.8
1.3
0.1
Neomysis mercedis:11
Rhopalophthalmus terranαtalzs ’戸
(1mmature)'0
Mesopodopsi.<叩oold門:dgei15
Rhopalophthalmus teγranatalis15
Praunus fiexuosus18
Jerling & Wooldridge 1995
Jerling & Wooldridge 1995
3
5
0.4
4.8
Copepod assemblage
Copepod assemblage
Viitasalo & Rautio 1998
Chapman & Thomas 1998
Chapman & Thomas 1998
Chapman & Thomas 1998
Chigbu & Sibley 1994
Murtaugh 1983
Toda et al. 1987
19
301
82
1.3
9.1
192
45
10
3.3
0.8
50
417
333
42
567
416
Tenagomysis chiltoni
T四時gomysischiltoni
Tenagomysis chiltoni
Neomysis mercedisl11
Neomyszs mercedis
Neomysis integer
Bosm口ialongispina
maritimα Bosmina meridianαlzs Ceriodaphnia dubi.α!3
Daphnia carinata16
Daphnia magnα?
Daphnia spp.
Daphnia sp.
Cladocerans
50 33

Diet type
Mysids
Amphipods Aγ"tern姐
Others
Others
Diet/Prey
Pleopsis polyphemoides
凡1esopodopsiswoold門'dgei:5
凡1ysisγ・efictaH0
Rhopalophthalmus te庁αnatalis15
Paracalliopeβuiiatilzs
A rtemia nauplii
A rtemia naupl口
Zooplankton assemblage
Zooplankton assemblage Zooplankton assemblage Microzooplankton (<200μm)
Microzooplankton (く90μm)
Microzooplankton (く90μm)
Bacteria Algal detritus Coral mucus
Coral mucus Spαrtina detritus
Species of mysids
Praunus flexuosusl8
Rhopalophthalmus
terr anαtalis15 Mysis relicta110
Rhopalophthalmus
ternαnatalis15
Tenagomysis chiltoni
Anisomysis mullini
Siriella jaltensis
Neomysis integer' Mysis mixta:8
凡1esopodopsiswoold門'dgeil5Mesopodopsis woold門'dgef5
M田 opodοpsis問。oldγidgei:5
Rhopalophthalmus terrana tal is:5
Anisomysis mulltnt
Anisomysis mullini
Anisomysis mullini
Mysid口lmintegrum
Neomysis americanal9
Table 4. (continued)
Clearance rate
mlmysid 1 h 1
46
41
108
41.2
く0.6
16 4
42
Predation/Grazing rate
Prey (cell) mysidペh 1
80
0.1
0.1 0.04
0.4 6 4.2
Carbon ingestion Daily ration References
rate
μgC mysid 1 h 1 %。fbody Cd 1
102 24 Viitasalo & Rautio 1998 1.1 2 Wooldridge & Webb 1988
3.5 3 Quirt & Lasenby 2002
1.6 3 Wooldridge & Webb 1988
Wilhelm et al. 2002 4.8 72 Mullin & Roman 1986
(40)' Arashkevich & Dirts 1981
4.4 11 Uitto et al. 1995
40 63 Viherluoto & Viitasalo 2001a 0.2 4 Froneman 2001 0.1 2 Froneman 2001
0.3 2 Jerling & Wooldridge 1995
0.1 0.1 Jerling & Wooldridge 1995
く0.01 <0.24 Mullin & Roman 1986 く0.6 く13.5 Mullin & Roman 1986
<0.6 く1.4 Mullin & Roman 1986 1 7日 Gottfried & Roman 1983
36 233 Zagursky & Feller 1985
* Carbon contents of the diatom are estimated based on Menden-Deuer & Lessard (2000) with length of cell in the following literature; 1. Talbot & Bate (1986), 2.
Burkhardt et al. (1999).
t Dry weight was converted to carbon contents with a factor of 0.4.
: Carbon contents of diets and mysids are estimated based on the following literature (Dry weight was converted to carbon contents with a factor of 0.4); 1. Roman
(1977), 2. Dagg (1977), 3. Forsyth & James (1984), 4. Langeland et al. (1991), 5. Wooldridge & Bailey (1982), 6. Mitchell & Williams (1982), 7. Arndt & Jansen (1986),
8. Viherluoto & Viitasalo (200lb), 9. Mauchline (1980), 10. Sell (1982), 11. Johnston & Northcote (1989).
持 Dryweight value of Epischura lacustris of the same size was used (Langeland et al. 1991 ).
1 Carbon weight value of Eurytemora affinis of the same size was used (Viitasalo & Rautio 1998).
『 Thevalues were estimated based on caloric content.
Z時菰
5喜き号. E『、/,
J主日主.t (;
古十、十
む込
、4,,,
童話fうj萄l!il'l
~諜干
σ3 トー

62 日本プランクトン学会報第 51巻第 l号(2004)
1995).ミシガン湖では M.relictaの摂食速度が現場の
動物プランクトン群集を閉じこめたチャンパー内で測定
され, 1日に体炭素の 27%を摂食していることが報告
されている(Bowers& Vanderploeg 1982, Vander-
ploeg et al. 1982).
微小動物プランクトンに対する摂食は南アフリカの
Mesopodoρsis wooldridgei, Rhopaloρhthalmus terrana-
talis などで測定されており (Jerling & Wooldridge
1995, Froneman 2001), O.l-0.3μg C ind 1 h-1, 1日に
体炭素の 2%以下という値が報告されている(Table4).
(2)植物プランクトン
これまでの報告によると,アミ類の植物プランクトン
に対する摂食速度は分類群によって大きく変動するよう
である.珪藻類に対するアミ類の摂食速度は Neomysis
integer, Castrosαccusρsammodytes, Mesopodopsis wool-
dridgeiなどで報告されている(Lucas1936, Webb et al.
1987, 1988).これらの報告は,いずれもアミ類 1個体あ
たり 1時間に 1-8×106cell程度,炭素量で 160~260
μgの珪藻を摂食できることを示している.これを体炭
素量あたりの比に換算すると, 1日に体炭素の 200%以
上を摂食できる計算になり (Table 4),珪藻の濃密な
パッチは効率の良い餌料源となる可能性を示唆してい
る.
バルト海の Mysismixtaで藍藻類,緑藻類に対する摂
食速度が測定されている(Engstromet al. 2001 ). M.
mix taのこれら藻類に対する摂餌率は,炭素量にして 3
μg程度, 1日に体炭素量の 5%程度にとどまる.また有
毒な藍藻類に対しては,さらに摂食速度が低下する
(Engstrom et al. 2001 ).また Siriellajaltensぬでは緑藻
類 Tetr,αselmis suesicaに対する一個体あたりの摂食速
度が 2.2×105cells h-1. l.2μgC h lであることが報告
されている(Arashkevich& Dirts 1981 ).
野外では消化管蛍光法によって南アフリカ汽水域にお
いて MesopodopsiswoodridgeiがO.lμgCind1 h 1, 1
日に体炭素の約 2%の植物プランクトンを摂餌している
ことが報告されている(Froneman2001).
(3) デトライタスおよびその他
非生体有機物に対する摂食速度がいくつかの種で測定
されている. Neomysis americanαはイネ科植物のデト
ライタスに対して 36μg C ind 1 h一1,1日に体炭素の
230%を摂食する(Zagursky& Feller 1985).サンゴの
分泌する粘液に対する摂食速度は Mysidiumintegrum,
Anisomysis mulliniで測定されており,それぞれ 1日あ
たり体炭素量のト70%,1.4%以下と変動幅が大きいこ
とが示されている(Gottfried& Roman 1983, Mullin &
Roman 1986).サンゴ礁に生息する A.mulliniでは藻
類起源のデトライタスおよびバクテリアに対する摂食率
が測定されており,それぞれ 1日あたり体炭素量の
13.5%以下, 0.24%以下を摂餌する(Mullin& Roman
1986).
底泥に対する摂食率もいくつかの種で報告されてい
る.Johnston & Lasenby (1982)はNeomysismercedis
にメイオベントスを含んだ底泥を与え,その排i世速度か
ら, 1日あたり乾燥重量にして 1.22.4 mg,体重の 13-
27%を摂食すると見積もった. また Mysisrelictaはl
日あたり乾燥重量にして約 1mgの底泥を摂食する
(Val Klump et al. 1991).
摂餌活動における生物・環境要因と人為的影響
アミ類の摂餌速度は様々な環境要因によって変動す
る.摂食速度は一般に水温と正の関係を示し,最適水温
より高くなると摂食速度を低下させる (Johnston&
Lasenby 1982, Toda et al. 1987など). Toda et al.
(1987)は 3℃から 30°Cの範囲で Neomysisawαtschen-
sisの摂食速度を測定した.本種の摂食速度は 3°Cから
20°Cにかけて増加し, 2025°Cで最大値に達する.30°C
では大型個体の摂食速度は変化しないが,小型個体では
やや低下する.一方,水温の変化がアミよりも餌生物に
より大きな影響を与える場合,水温の低下がアミ類の摂
食活動に有利に働くことがある. Cooper & Goldman
(1982)はタホ湖に生息する Mysisrelictaのカイアシ類
2種に対する漉水速度を 5,10,15°Cで測定した.彼らは
M. relictaの漉水速度は逃避能力の劣る Eρischuraを餌
料とした場合は水温の影響を受けないが,逃避能力に優
れた Diaρtomusを餌料とした場合, 5°Cで有意に高くな
ることを報告し原因として低水温がDiaptomusの逃
避能力を低下させた可能性を挙げている.汽水域に生息
する Neomysisintegerでは塩分の上昇に伴って摂餌活
動が活発になることが知られている(Roastet al. 2000).
アミ類の複眼が受容する光の波長域は,種や生息域に
よって異なり(Lindstrom2000),実験の光条件も摂食速
度に影響を与えると考えられる.これまでの報告は光が
摂餌速度に対して正に働く場合 (Neomysisamericana,
Mysidoρsis bigelowi; Fulton l 982b, M. relict,α;Ram-
charan & Sprules 1986),負に働く場合 (Mysismixta;
Gorokhova & Hansson 1997, Viherluoto & Viitasalo
ZOO lb),影響しない場合(M.relicta; Cooper & Gold-

高橋:淡水・沿岸域におけるアミ類の摂餌生態 63
man 1982, Praunus fiexuosus; Viherluoto & Viitasalo
200lb)と様々であり, 実験の際は個々の種の生息環境
や摂餌リズムを考慮に入れる必要がある.
Mesaρadoρsis wooldridgeiの珪藻 Anaulusbirostra-
tusに対する摂食速度は, 同体長の雌雄で比較すると雌
でより速いことが知られている(Webbet al. 1987).一
方, Rhoραloρhthalmusterranatalぬでは摂食速度に性
別は影響しないことが報告されている(Wooldridge&
Webb 1988).このほか,実験時間,絶食期間,飼育水槽
容量や(Cooper& Goldman 1980, 1982, Jerling &
Wooldridge 1995, Gorokhova & Hansson 1997),アミ
類と餌生物密度の相対比(Hanssonet al. 2001)などが
実験結果に影響を与える要素となることが知られてい
る.
餌料選択性と摂餌モードの切り替え(switching)
アミ類は多様な餌料を利用することができるが,実際
には環境中から特定のサイズや種類の餌を選択的に摂食
していることが,多くの実験や現場試料から明らかに
なっている(Table5).
植物プランクトンや微小動物プランクトンなどでは,
原則として個々の餌料の大きさが重要な選択要因となっ
ており,比較的大型の種が好まれ,一定サイズ以下の種
は摂食されないという報告が多い.サクラメン卜の運河
に生息する Neomysismercedisは直径 50μmを超える
Coscinodiscusや群体を作る Melosiraを選択的に摂食
し, lOμm以下の Skeletonemaや5μm以下の Meris-
moρediaは環境中に優占するにもかかわらずほとんど
摂食しない(Siegfried& Kopache 1980).ミシガン湖で
はMysisrelictαが植物プランクトン中, 53μm以上の
群体を形成する珪藻類を選択的に摂食している (Bow-
ers & Grossnickle 1978).また Webbet al. (1987)も
Mesaρadoρsis wooldridgeiが群体を形成する珪藻 Aste-
rionella gracilisを,細胞数では卓越する小型種よりも
好んで摂餌することを報告している. M. wooldridgei,
Rhopaloρhthalmus terranatalisでは植物プランクトン,
微小プランクトン群集中 (90μm以下),動物を中心と
した大型の餌を好み(30-50μm程度),小型(<29μm)
の植物プランクトンに対する選択性は低いことが示され
ている (Jerling& Wooldridge 1995).
動物プランクトンに対するアミ類の捕食選択性は変化
に富み,アミ類の種はもちろんのこと,餌生物のサイズ,
種,組成,密度などに影響される.餌のサイズは,餌生
物の逃避能力および餌の捕獲から摂食終了に至るハンド
リングの過程と関連し,摂餌選択性を左右する重要な要
素となっているが,その影響はアミ類の成長段階によっ
て異なるようだ.Murtaugh (198lb)はDαphniaを餌と
して Neomysismercedisのサイズ選択性を調べた結果,
小型個体は小型の餌を好んで捕食する傾向を示す一方
で,成体は最大クラス(3.0mm)を除く, 全てのサイズ
レンジの Daphniaをほぼ等しく捕食することを明らか
にした.同様の傾向は Mysisrelicta, Tenagomysis chil-
toniなどでも報告されている(Cooper& Goldman
1980, Wilhelm et al. 2002).また,大きすぎる餌は捕獲
しでもうまくハンドリングすることができず破棄された
り(Murtaugh198lb, c, Chapman & Thomas 1998),
たとえ破棄されなくても,小型の餌に比べハンドリング
の時間が長くかかるため,結果として摂食速度や選択性
をある程度低下させる場合がある(Ramcharan et al.
1985, Chigbu & Sibley 1994). N. mercedis成体では,
得られるエネルギーが摂食時間あたり最も高くなる中型
のDゅhniaよりも,ややエネルギー摂取効率の劣る小
型の個体を好んで捕食することから,エネルギー摂取効
率よりも餌の捕まえやすさが選択性決定の上で重要であ
る可能性が示唆されているにhigbu& Sibley 1994).
餌生物種それぞれのもつ行動特’性もアミ類の摂餌選択
性を決定する重要な要因となっている(Cooper& Gold-
man 1980, Murtaugh 198la, 198lb, Chapman & Tho-
mas 1998).アミ類が特定の種,動物群を選択的に捕食
する例として最も有名なのは,本稿冒頭でも述べた淡水
性アミ類 Mysisrelict,αの移植に伴う枝角類の減少・消
滅であろう.多くの研究は M.relictaが比較的大型で逃
避能力に劣る枝角類を選択的に捕食することを証明して
いる(Cooper& Goldman 1980, Bowers & Vander-
ploeg 1982, Grossnickle 1982, Nero & Sprules 1986b,
Spencer et al. 1999). M. relictaに限らず淡水性アミ類
がカイアシ類よりも枝角類を好んで捕食する傾向は
Neomysis属数種や Tenagomysischiltoniなどでも報告
されている(Murtaugh198la, Hanazato 1990, Irvine
et al. 1993, Chapman & Thomas 1998). Ramcharan et
al. (1985)は M.relicta (体長 19mm)のカイアシ類5
種,枝角類 1種に対する捕食選択性が小型種ほど高くな
る傾向があることを示した上で,それぞれの種に対する
捕食過程(攻撃・捕獲・捕食)を観察した(Gerritsen&
Strickler 1977).この実験は暗条件下で行われており
M. relictaは主に物理刺激を頼りに餌を探索したと考え
てよいだろう.観察の結果, M relictaは,遊泳に伴って
大きな物理的擾乱を生じる大型餌種ほど頻繁に攻撃する
傾向を示したが,実際に捕食できる確率(最終攻撃成功

σ3 品
Table 5. Various patterns of feeding selectivity of mysids in the fields and under experimental conditions. “Factor a汀ectingthe selectivity" shows the most
influential element in the process of prey selection. “>” indicates the t巴ndencyof preference in prey selection.
Factor affecting
the selectivity
(pattern of preference) Referenc巴sL巴sspreferred prey Preferred prey Species of mysids Diet type
回新刈。JW
て山可寸て品川ゆ惑
Webb et al. 1987
Siegfried & Kopache 1980
Jerling & Wooldridge 1995
Rybock 1978
Cooper & Goldman 1980
Cooper & Goldman 1980
Skeletonema, Merismopedia
Acartia and Pseudodiaptomus copepodi tes
Diaptomus
Cope pods
Diaρtom us
Anaulus birostr,αtus
泊四戸川附
泌-
4(
NCC品)
Bowers & Vanderploeg 1982
Folt et al. 1982
Ramcharan et al. 1985
Nero & Sprules 1986b
Irvine et al. 1993
Jerling & Wooldridg巴 1995
Adult diaptomids & cyclopoids
Diaρtom us Large copepods
Large copepods, Diaptomus
Eurytem口ra
Pseudodiaptomus copepodites
Coscinodiscus, Melosira
Acartia and Pseudodiaptomus nauplii
Epischura, Kellicottia, Bosmina
Daphnia
Epischura, cladocerans, cyclopoids
Cladocerans
Epischura
Daphnia, small copepods
Small cladocerans, Epischura
Daphnia
Pseudodiaρtomus nauplii
Asterionella gracilis Mesopodoρsis wooldridgei
Neomysis mercedis
Mesopodopsis wooldridgei
Size (large> small) Phytoplankton
My sisγelicta
Mvsisγelicta
Mvsis relicta
Escape response of prey
(vigour less> vigour)
Zooplankton
Mysis relicta
Mysis relicta
Mysis relicta
Mysis relicta
Neomysis integγ
Rhoρalophthalmus terranatalis
Mysis mixta Viitasalo et al. 2001 Bosmina, Pseudocalanus, Acartia
Eurytemora, Temora Frequency of encounter
(high>low)
Wooldridge & Webb 1988
Cooper & Goldman 1980
Wilhelm et al. 2002
Fulton 1982a, b
Viitasalo & Rautio 1998
Pseudodiapto押ius
Diaptomus female
Am phi pods
Centropages
Acartia
Mesopodopsis wooldridgei
Diaptomus male
Daphnia
Acartia
Pleopsis
Rhopalophthalmus
terranatalis
Mysis relicta
Tenagomysis chiltoni
Mysidopsis bigelowi
Praunus fiexuosus
Prey availability
(efficient> less efficient)

高橋・淡水・沿岸域におけるアミ類の摂餌生態 65
率: ultimateattack success)は大型餌種で低く,逆に
小型餌種で高くなった.また枝角類 (Daρhniaρulex)に
対する攻撃頻度は同サイズのカイアシ類 (Diacyclops
thomasi)と比べ半分程度に留まるが,最終攻撃成功率は
4倍以上も高く,本種が M.relictaに容易に捕獲される
ことを示した. Ramcharan et al. (1985)はこの最終攻
撃成功率と捕食選択性が高い相闘を示すことを認め,餌
生物の逃避能力がM.relict,αの捕食選択性を決定する重
要な要因であると結論した.この結果は M.relict,αの枝
角類や小型のカイアシ類に対する選択性が,自ら好んで
捕食した「真の」選択性ではなく,餌生物の逃避能力の
差によって生じた「見かけの」選択性であることを示し
ている.
一方バルト海の Mysismixt,αでは,枝角類よりもカイ
アシ類を好むという淡水性アミ類とは逆の傾向が報告さ
れている(Mohammadianet al. 1997, Viherluoto &
Viitasalo 2001a). Viitasalo et al. (2001)はカイアシ類
4種 (Temoralongicornis, Euηtemora affinis, Pseudo-
calanus elongatus, Acartia sp.),枝角類 1種 (Bosmina
longisρina maritima)に対する M.mixta (体長 319
mm)の選択性と,餌種それぞれの逃避能力を測定し,
M. mixtaの選択性が餌生物の逃避能力よりも, むしろ
サイズに依存する傾向を見出した.枝角類は同サイズの
カイアシ類に比べると,やや好まれる傾向があるもの
の,大型のカイアシ類に対してそれ以上に強い選択性が
認められた.この理由として彼らは物理的刺激を主な餌
探索刺激とする M.mixt,αにとって(Viherluoto& Vii-
tasalo 2001b),遊泳に伴う物理的撹乱の大きい大型種ほ
ど検知されやすいためであると結論している.この実験
で M.mixtaが最も強い選択性を示したEurytemoraは
逃避能力が最も優れた種であり,ゆえに M.mixtaは本
種に対して「真の」選択性を示したと考えられる(Vii-
tasalo et al. 2001). Ramcharan et al. (1985), Viitasalo
et al. (2001)の研究はいずれもアミ類が大型の動物プラ
ンクトンに対して好んで攻撃を仕掛けることを示してい
る.一方で,それぞれの種が示す対照的な選択性は,
個々の餌生物の逃避能力とアミ類の餌生物捕獲能力の差
が捕食選択性を決定する上で重要な要因となっているこ
とを示している.
餌生物の逃避能力とアミ類の捕獲能力の関係は一定で
はなく,餌種の密度や他餌種の存在によって変化する場
合もある.Mysis relictaは2種のカイアシ類Eρischura,
およびDiaρtomusに対して,前種をより好んで捕食す
ることが実験および野外調査から明らかにされているが
(Table 5), Folt et al. (1982)は,このカイアシ類2種に
対する選択性が両種の相対密度(個体数比)よりも,む
しろ合計密度に大きく影響されることを明らかにした.
彼女らの実験結果によると, M.relictaのEρischuraに
対する選択性は Diaρtomusとの合計密度が低いとき
(2.7 ind l 1)に最も強くなり,高密度下(>13 ind J一1)で
は低下(すなわち Diaptomusに対する選択性が相対的
に上昇)する.この原因として高密度条件下での捕食
者・餌生物の行動の変化,例えは、 M.relict,αのDiaρto-
mus捕食能力の向上,あるいは Diaρtomusの逃避能力
の低下などが生じた可能性が指摘されている(Foltet al,
1982, Folt 1987).
アミ類が懸濁物摂食と捕獲摂食の二つの摂餌モード
(摂食機構の項参照)を餌環境によって使い分けること
により摂餌行動を最適化することが最近報告された.
Viitasalo & Rautio (1998)は Praunusfiexuosusにカイ
アシ類Acartiaと枝角類Pleaρsisを単独で,あるいは混
合 (1:2)して様々な密度で摂餌させた結果,高密度下に
おける Acart切に対する摂食速度がPleaρsisと混合す
ることで大幅に低下することを報告した.これは小型で
遊泳力の弱い枝角類Pleaρsisが高密度で存在したこと
により, P.fiexuosusが専ら懸濁物摂食モードを採用し
たためであると考えられ,機能的応答の観点からもこの
摂餌モードのスイッチングが摂餌行動を最適化する上で
有効であることが証明されている(Viitasalo& Rautio
1998).多くのアミ類が好んで摂食するカイアシ類ノー
プリウス幼生 (Irvineet al. 1993, Aaser et al. 1995,
Jerling & Wooldridge 1995など)が,時として全く選
択されない現象は(Siegfried& Kopache 1980, Mur-
taugh 1981a),こういった摂餌モードのスイッチングと
関連していると考えられる.アミ類における摂餌モード
のスイッチングは,本動物群の摂餌生態を考える上で重
要な性質であるが,知見は限られている.とくに一部の
魚類やカイアシ類で,すでに示されているような,ス
イッチングが起こる餌料・環境条件の特定(Leong&
O’Connell 1969, Saiz & Ki0rboe 1995, Ki0rboe et al.
1996)や性質の異なる餌料(動物,植物,デトライタス
など)間でのスイッチングなどは今後の研究課題として
重要である.
アミ類の摂餌活動が食物網に与える影響
これまで述べてきたように,アミ類は環境に応じて
様々な餌を効率よく摂取する能力をもっており,その摂
食活動は時として食物網内の他の生物に大きな影響を与
える.アミ類の摂食活動が生態系に与える影響の最も代

66 日本プランクトン学会報第51巻第 1号(2004)
表的な例は,捕食を通じて動物プランクトン群集の組成
や量を左右するという現象である.淡水域では, Mysis
relic taに関する論文が多く発表されている. M. relict,α
の移植は枝角類や一部の動物プランクトンの減少を招
き, 魚類や基礎生産にまで影響を与えている(Morgan
1982, Nesler & Bergersen 1991 ). M. relic taの移植が生
態系に最も顕著な影響を与えた例は Spenceret al.
(1991)によって報告されている.米国フラットヘッド湖
では M.relictaの捕食による枝角類の減少が,おなじく
枝角類を餌としていたヒメマスの産卵遡上尾数の減少を
招き,この魚を餌としていたワシ,ヒグマなどが減少し
た.またオランダ貯水池ではカスピ海水系からの侵入種
Hemimysis animalaが定着し, その摂餌圧によって植
物プランクトンやワムシ類・枝角類・介形類などが急激
に減少・消滅したことが報告されている(Ketelaarset
al. 1999).
ノースカロライナ沿岸では冬季,カイアシ類群集の優
占種がAcartiaから Centroρages,Sαρhirellaに遷移す
る原因として MysidoρsisbigelowiとNeomysisameri-
canaの捕食が重要であると考えられている(Fulton
1982a, b).この 2種のアミ類はAcartiaを好んで捕食す
るが,その捕食は密度依存的でカイアシ類群集中に
Acartiaの占める割合が低い場合は,カイアシ類群集の
密度を減少させるものの,その組成には影響しないため
カイアシ類群集の多様性の維持に効果があるとした
(Fulton 1982a).この 2種のアミ類は冬季 1日あたりカ
イアシ類Acartiaおよび Centropagesの現存量の 25%
および 9%をそれぞれ捕食する能力を持っている (Ful-
ton 1982b, 1983).イギリスの富栄養汽水湖ではカイア
シ類Eurytemoraの生物量の減少がNeomysisinteger大
型個体の捕食に起因している可能性が示唆されている
(Irvine et al. 1995).
Mysis relictaの動物プランクトンに対する捕食圧は
時としてプランクトン食性魚類をはるかにしのぎ,これ
ら魚類と餌をめぐって競合関係にあると考えられている
(Johannsson et al. 1994, Chipps & Bennett 2000,
Johnson et al. 2002).米国アイダホの湖における枝角類
に対する年間捕食量は,湿重量にしてヒメマスでは 4.5
gm-2であるのに対して M. relicta では 4倍以上の
18.6gm 2であると見積もられている(Chipps& Ben-
nett 2000).バルト海に生息する Mysismixtaはニシン
稚魚と並び動物プランクトン捕食者として重要であるこ
とが報告されている(Hanssonet al. 1990a).本種個体
群の摂食量は夏季に 1日あたり 4-16 mgCm 2にのぼ
り,これは動物プランクトン群集全体の生産量の約 l割
にあたる(Hanssonet al. l 990a).また本種の成長は動
物プランクトン現存量に制限されていることが現場デー
タおよび摂餌実験によって示されており(ただし実験手
法における問題点について Hanssonet al. 2001 も参
照),とくに夏季から秋にかけて本種は餌料において,魚
類なと、の他の動物フ。ランクトン食者と競合している可能
性が示唆されている(Hanssonet al. 1990b, Mohamma-
dian et al. 1997).
特定の動物プランクトンを減少させることで,群集内
の競合関係あるいは捕食関係が変化し,他の種類が増加
することもある.Mysis relict,αが移植された湖では枝角
類が選択的に捕食された結果,枝角類と競合関係にあっ
たカイアシ類が増加することがある(Nero& Sprules
1986a, Spencer et al. 1999).また Neomysisintegerの
捕食によるカイアシ類の減少が,繊毛虫類の増加をもた
らすことが短期間のメソコズム実験で報告されている
(Uitto et al. 1995).デンマークの富栄養汽水湖では, N.
integerが主要植物フ。ランクトン食者であるカイアシ類
Eurytemora個体群を捕食によって減少させ,さらに摂
食活動を通して底質から水柱内へ栄養塩を再循環させる
ことによって植物プランクトン量を増加させ,透明度を
低下させる原因となっている可能性が示唆されている
(Aaser et al. 1995). M. relictaを移植したスウェーデン
の湖でも,アミ類の増加に伴って,植物プランクトンの
ブルームがみられるようになったことが報告されている
(Kinsten & Olsen 1982).
ミシガン湖では Mysisrelictaが珪藻を破砕する過程
で漏出する溶存態有機物が,ノ〈クテリア群集にとって重
要である可能性が示唆されている(Sierszen& Brooks
1982).また Bowers& Grossnickle (1978)はM.relicta
が摂食の際に破砕した大型珪藻細胞が,カイアシ類の餌
料源となっている可能性を示唆した.
底生動物群集に対する影響については研究例が少ない
が,カナダ西岸河口域のNeomysismercedisはメイオベ
ントスを主要な餌料としており, 1日に最大でハルパク
チクス目カイアシ類個体群の 12-13%を摂餌する能力
をもっていることが報告されている (Johnston& La-
senby 1982).またニュージーランドの湖に生息する
Tenagomysis chiltoniは,同じ湖に住むヨコエビ Para-
calliopeβuviatilis個体群に対して,一日あたり春夏季
には個体数の 0.7-5%,秋冬季には 1.5-9.6%を捕食して
いると見積もられている(Wilhelmet al. 2002).
アミ類の日周摂餌リズムは多くの場合,鉛直的に数
メートルから数十メートルの移動伴うため,水柱と海底
聞の物質運搬において重要な働きを担っていると考えら

高橋:淡水・沿岸域におけるアミ類の摂餌生態 67
れる(村野 1974,Mauchline 1980).このような役割は
水平的にもみられる.Mesaρodopsis wooldridgeiは砕波
帯内で夜間摂餌を行い,昼間は砕波帯外へ移動するた
め,砂浜域における岸沖方向の物質輸送に重要な役割を
果たしていると考えられている(Cockcroftet al. 1988,
Webb & Wooldridge 1990).地中海の洞窟性アミ類
Hemimys俗学eluncolaは,夜間洞窟外で摂餌した後,洞
窟内に帰り糞粒を排池するため,洞窟内の有機物供給役
として重要であると考えられている(Comaet al. 1997).
おわりに
以上,近年の研究結果を中心に,アミ類の摂餌生態に
ついて概観した.近年多くのカイアシ類で報告されてい
るように,プランクトン食物網において雑食者は特別な
存在ではなくなりつつある.アミ類が他の雑食者と大き
く異なる特徴として,利用できる餌料の種,大きさが多
様である上に,植食者,肉食者,デトライタス食者のい
ずれとしても優れた能力を発揮するという点が挙げられ
るだろう.Mysis relictaの移植の影響の大きさから,近
年のアミ類の食性研究は肉食者としての側面を強調する
ものが多いが,実際には動物プランクトン以外の餌もま
た同様かそれ以上に重要であるという研究結果が最近に
なって増えてきた(Aaseret al. 1995, Irvine et al. 1993,
Johannsson et al. 1994, 2001).今後は餌料環境の季節
変化やアミ類発育段階毎の餌料獲得能力の変化を考慮に
入れた食性研究が,アミ類の食物網内に果たす役割を知
Terrestrial ecosystems
Bottom
る上で鍵となるだろう.
アミ類の雑食性は餌料環境変化の観点からおそらく二
つの利点を有する.ひとつは植物プランクトンブルーム
や動物プランクトンの繁殖期など生産性の高い時期に豊
富な特定の餌料を効率よく摂食できる点,もうひとつは
冬季など餌料の乏しくなる時期をデトライタスや底生生
物など様々な代替餌料を利用することで,他の甲殻類プ
ランクトンにみられるような休眠期や著しい成長の停滞
を経ずに乗り切ることができる点である.とくに後者の
利点はアミ類の生物量や栄養状態を周年一定のレベルに
保つのに貢献しており(Hanazato1990, Adare & La-
senby 1994),結果としてアミ類が潜在的動物プランク
トン捕食者として食物網内で一定の影響力を保つ原因と
もなっていると考えられる(Fultonl 982b, 1983).多く
のアミ類が生息する淡水から沿岸域は,水柱,海底(湖
底)の双方で生産が行われる上に,陸域からも有機物が
流入する水域である.アミ類の柔軟な雑食性は(Fig.3),
これら様々な有機物を餌料として速やかに利用する上で
有効であり,繁殖戦略(Wittmann1984, Johnson et al.
2001)や捕食者逃避行動(Rademacher& Kils 1996,
Ritz et al. 1997)などとともに,これらの水域でアミ類
が高い生物量を保つ一因となっているといえよう.
謝辞
前本誌編集委員長の北海道大学志賀直信博士には本総
説を執筆する機会を与えて頂きました.東北区水産研究
Water column
. .
Fig. 3. Schematic diagram of flexible omnivory of mysids in freshwater, estuarine, and coastal marine habitats. Open arrows indicate solar radiation, solid arrows indicate feeding of mysids, dotted arrows indicate excretion and decomposition process.

68 日本フ。ランクトン学会報第 51巻第 l号(2004)
所混合域海洋環境部生物環境研究室,斉藤宏明室長には
原稿を校閲して頂きました.桑田 晃,中川至純, Alf
Norkko,太田尚志の各氏には有益な助言を頂きました.
釜石隆,河内直子,小針統,柴田晃,下出信二,
高橋邦夫,新村陽子,花村幸生,贋瀬太郎,松政正俊の
Mar. Ecol. P:γng. Ser. 184: 31 41.
Cannon, H. G. & S. M. Manton 1927. On the feeding mecha-
nism of a mysid crustacean, Hemimysis lamornae. Trans. Roy. Soc. Edinburgh 55: 219 255.
Chapman, M. A. & M. F. Thomas 1998. An experimental
study of feeding in Tenagomysis chiltoni (Crustacea, Mysi-dacea). Arch. Hydrobiol. 143: 197 209.
各氏には文献収集の面でご協力を頂きました. ここに記 Chigbu, P. & T. H. Sibley 1994. Predation by Neomysis
して感謝申し上げます.
引用文献
Aaser, H.F., E. Jepps巴n& M. Sondergaard 1995. Seasonal
dynamics of the mysid Neomysis integer and its predation
on the copepod Eurytemora ajfinis in a shallow hyper-
tropic brackish lake. Mar. Ecol. Prog. Ser. 127:・ 47-56.
Adare, K. I. & D. C. Lasenby 1994. Seasonal changes in the
total lipid content of the opossum shrimp, Mysis relicta
(Malacostraca: Mysidacea). Can. J Fish. Aquat目 Sci.51: 1935-1941.
Arndt, E. A. & W. Jansen 1986. Neomysis integeγ(Leach) in
the chain of boddens south of Drass/Zingst (Western
Baltic) Ecophysiology and population dynamics. Ophe-
lia Suppl. 4: 1 15.
Arashkevich, Y. E.G. & A. V. Dirts 1981. Experimental
study of the feeding habits of the Black Sea mysid Siriella
1αltensis (Czernjavsky). Oceanoloεy 21: 498 502.
A ttramadal, Y. G. 1981. On a non-existent ventral filtration current in Hemimysis lamornae (Couch) and Praunus fiexu-osus (Muller) (Crustacea: Mysidacea). Sarsia 66: 283 286.
Bailey, K. M., R. D. Brodeur, N. Merati & M. M. Yoklavich 1993. Predation on walleye pollock (Ther,α:gra chalco-gramma) eggs and yolk-sac larvae by pelagic crustacean
invertebrates in the western Gulf of Alaska目 Fish.Ocean-ogr. 2: 30 39.
Baldo Kost, A. L. & A. W. Knight 1975. The food of Neo-mysis mercedis Holmes in the Sacramento-San Joaquin
estuary. Calif Fish Game 61: 35 46. Begon,恥1.,J. L. Harper & C.R. Townsend 1996. Ecology.
Individuαls, Populations and Communities (3'ct ed). Black-well Science, 1068 pp.
Bowers, J. A. & N. E. Grossnickle 1978. The herbivorous
habits of Mysis relicta in Lake Michigan. Limnol. Ocea nogr. 23: 767-776.
Bowers, J. A. & H. A. Vanderploeg 1982. In situ predatory
behavior of Mysis relicta in Lake Michigan. Hydrobiologia 93: 121ー131.
Branstrator, D. K., G. Cabana, A. Mazumder & J.B. Ras-
mussen. 2000. Measuring life-history omnivory in the
opossum shrimp, Mysis relicta, with stable nitrogen iso-topes. Limnol. Oceanogr. 45: 463 467.
Bremer, P. & J. Vijverberg 1982. Production, population biology and diet of Neomysis integer (Leach) in a shallow
Frisian Lake (The Netherlands). Hydrobiologia 93:・ 4151.
Brown, A. C. & M. S. Talbot 1972. The ecology of the sandy
beaches of the Cape Peninsula, South Africa. Part 3: a
study of Gastrosaccus psammodytes Tattersall (Crustacea:
Mysidacea). Trans. Roy. Soc. S. Afr. 40: 309 333. Burkhardt, S., U. Riebesell & I. Zondervan. 1999. Stable
carbon isotope fractionation by marine phytoplankton in
r巴sponseto daylength, growth rate, and C02 availability.
mercedis: effects of temperature, Daphnia magna size and
prey density on ingestion rate and size selectivity. Fresh-water Biol. 32: 39 48.
Chipps, S. R. & D. H. Bennett 2000. Zooplanktivory and nu-
trient regeneration by invertebrate (Mysis relicta) and vertebrate (Oncorh::,nchus nerka) planktivores: Implica-tions for trophic interactions in oligotrophic lakes. Trans.
Am. Fish. Soc. 129: 569 583.
Clutter, R. I. 1969. The microdistribution and social be-
habior of some pelagic mysid shrimps. J Exp. Mar. Biol. Ecol. 3: 125-155.
Clutter, R. I. & G. H. Theilacker 1971. Ecological efficiency
of a pelagic mysid shrimp; estimates from growth, energy
budget and mortality studies. Fish. Bull. 69: 93-115.
Cockcroft, A. C., P. Webb & T. Wooldridge 1988. Nitrogen
regeneration by two surf-zone mysids, Mesopodopsis slab beri and Gastrosaccus psammodytes. Mar. Biol. 99:・ 75-82.
Coma, R., M. Carola, T. Riera & M. Zebala 1997. Horizontal
transfer of matter by a cave dwelling mysid. P. S. Z. N.よ
Mar. Ecol. 18: 211 226. Cooper, S. D. & C.R. Goldman 1980. Opossum shrimp
(Mysis relicta) predation on zooplankton. Can. J Aquat. Fish. Sci 37:・ 909-919.
Cooper, S. D. & C.R. Goldman 1982. Environmental factors
a釘ectingpredation rates of Mysis relicta. Can. J Fish. Aquat.目 Sci.39: 203-208.
Cripe, G. M., R. S. Carr, S. S. Foss, P. S. Harris & R. S. Stanley
2000. E紅巳ctsof whole sediments from Corpus Christi
Bay on survival, growth, and reproduction of the mysid,
Americamysis bahia (formerly Mysidopsis bahia). Bull. En-viron. Contam. Toxicol. 64: 426-433.
Crouau, Y. 1986. Antennular m巴chanosensitivityin a cav-
ernicolous mysid crustacean.]. Crustacean Biol. 6: 158
165.
Crouau, Y. 1989. Feeding mechanisms of th巴 Mysidacea,
pp. 153 171. In Functional Morphology of Feeding and Grooming in Crustacea (eds. Felgenhauer, B. E., L. Wal ting, & A. B. Thistle). Balkema, Rotterdam.
Dagg, M. 1977. Some e百ectsof patchy food environments
on copepods. Limnol. Oceanogr. 22:・ 99107.
Dauby, P.A. 1995. Aδ13C study of the feeding habits in
four Mediterranean Lゅtomysisspecies (Crustac巴a:Mysi-
dacea). P. S. Z. N. I. Mar. Ecol. 16: 93 102. DeGraeve, G. M. & J.B. Reynolds 1975. Feeding behavior
and temperature and light tolerance of M_vsis relicta in the laboratory. Trans. Am. Fish. Soc. 103: 394 397.
De Jong-恥1orean,L. & J.-P. Casanova 2001. The foreguts of
the primitive families of the Mysida (Crustacea, Pera-
carida): a transitional link between those of the Lopho
gastrida (Crustacea, Mysidacea) and the most evolved
Mysida. Acta Zool. 82: 137 147.
De Jong-Moreau, L., B. Casanova & J.-P. Casanova 2001.
Detailed comparative morphology of the peri-oral struc-

高橋:淡水・沿岸域におけるアミ類の摂餌生態 69
tures of the Mysidacea and Euphausiacea (Crustacea): an
indication for the food preference. ]. Mar. Biol目 Assoc.u. K. 81: 235-241.
De Jong-Moreau, L., M. Brunet, J.-P. Casanova & ]. Mazza 2000. Comparative structure and ultrastructure of the
midgut and hepatopancreas of five species of Mysidac巴a
(Crustacea): functional implications. Can. ]. Zool. 78: 822-834.
D巴pdolla,P. 1923. Nahrung und Nahrungserwerb bei Prau-nus fiexuosus (Mi.iller). Biol. Zentralbl. 43: 534 546.
Engstrom,]., M. Viherluoto, & M. Viitasalo 2001. E釘ectsof
toxic and non-toxic cyanobacteria on grazing, zooplankti-
vory and survival of the mysid shrimp Mysis mixta. ].
Exp. Mar. Biol. Ecol. 257: 269 280.
Fenton, G. W. 1992. Population dynamics of Tenagomysis tasmaniae Fenton, Anisomysis mixta australis (Zimmer) and Paramesopodoρsis rufa Fenton from south-eastern
Tasmania (Crustacea: Mysidacea). Hydrobiologia 246:
173 193.
Fenton, G. W. 1996. Diet and predators of Tenagomysis tas-maniae Fenton, Anisa押zysismixta australis (Zimmer) and
Paramesopodopsis rufa Fenton from south-eastern Tasma-nia (Crustacea: Mysidacea). Hydrobiologia 323: 31 44.
Fockedey, N. & ]. Mees 1999. Feeding of the hyperbenthic
mysid Neomysis integer in the maximum turbidity zone of
the Elbe, Westerschelde and Gironde estuaries.]. Mar. Sys. 22: 207 228.
Folt, C. L. 1987. An experimental analysis of costs and benefits of zooplankton aggregation, pp. 300 314. In
Predation, Direct and lndirecct Imρacts on Aquatic Commu-nities (eds. Kerfoot, W. C. & A. Sih), University Press of
New England, London.
Folt, C. L.,]. T. Ry bock & C.R. Goldman 1982. The effect of
prey composition and abundance on the predation rate
and selectivity of Mysis relicta. Hydrobめlogiα93:133ー143.
Forsyth, D.]. & M. R. James 1984. Zoopalnkton grazing on
lake bacterioplankton and phytoplankton. ]. Plankton
Res. 6:・ 803-810.
Foulds, ]. B. & K. H. Mann 1978. Cellulose digestion in
Mysis stenolepis and its ecological implications. Limnol. Oceanogγ. 23: 760一766.
Friesen, J. A., K. H. Mann & ]. H. M. Willison 1986a. Gross
anatomy and fine structure of the gut of the marine
mysid shrimp Mysis stenoleρis Smith. Can. ]. Zool. 64:
431 441.
Friesen, ]. A., K. H. Mann & ]. A. Novitsky 1986b. Mysis
digests cellulose in the absence of a gut microflora. Can.]. Zool. 64: 442 446.
Froneman, P. W. 2001. Feeding ecology of the mysid,
Mesopodopsis wooldridgei, in a temperate estuary along
the eastern seaboard of South Africa.]. Plankton Res. 23:
999 1008. Fullon, R. S. III 1982a. Preliminary results of an experi-
mental study of the effects of mysid predation on estuar-
ine zooplankton community structure. Hydrobiologia 93: 79-84.
Fulton, R. S. III 1982b. Predatory feeding of two marine
mysids. Mar. Biol. 72: 183 191.
Fulton, R. S. III 1983. Interactive effects of temperature
and predation on an estuarine zooplankton community.].
ExρMar. Biol. Ecol. 72:・ 6781.
Gerritsen,]. & ]. R. Strickler 1977. Encounter probabiliti巴S
and community structure in zooplankton: a mathematical
model.]. Fish. Res. Ed. Can. 34: 73 82.
Goldman, C. R.. M. D. Morgan, S. T. Threlkeld & N. Angeli 1979. A population dynamics analysis of the cladoceran
disappearance from Lake Tahoe, California-Nevada. Lim-nol. Oceanogr. 24:・ 289ー297.
Gorokhova, E. & S. Hansson 1997. E汀ectsof experimental
conditions on the feeding rate of Mysis mixta (Crustacea, Mysidac巴a).Hydrobiologia. 355: 167 172.
Gorokhova, E. & S. Hansson 1999. An experimental study
on variations in stable carbon and nitrogen isotope frac-
tionation during growth of Mシsismixta and Neomysis
integer. Can.]. Fish. Aquat. Sci. 56: 2203 2210.
Gottfried, M. & M. R. Roman 1983. Ingestion and incorpo-
ration of coral-mucus detritus by reef zooplankton. Mαr. Biol. 72: 211 218.
Green, J.M. 1970. Observations on the behavior and larval development of Acanthomysis sculpta (Tattersall), (Mysi-
dacea). Can. ]. Zool. 48: 289-292.
Grossnickle, N. E. 1979. Nocturnal feeding patterns of
Mysis relicta in Lake Michigan, based on gut content
fluorescence. Limnol. Oceanogr. 24: 777 780.
Grossnickle, N. E. 1982. Feeding habits of Mysis relicta an
overview. Hydrobiologia 93: 101 107.
Hallberg, E. 1977. The fine structure of the compound eyes
of mysids (Crustacea: Mysidacea)目 CellTissue Res. 184:
45 65.
Hamner, W. M. 1988. Biomechanics of filter feeding in the
Antarctic krill Euρhausia superba: review of past work and new observations.よCrustaceanBiol. 8: 149 163.
Hanazato, T. 1990. A comparison between effects on zoo-
plankton communities by Neomysis and Chaoborus. Hy-drobiologia. 198:・ 3340.
Hansson, S., B. T. De Stasio, E. Gorokhova & M.A. Moham-
madian 2001. Ratio-dependent functional responses-
tests with the zooplanktivore Mysis mixta. Mar. Ecol.
Prag. Ser. 216: 181 189.
Hansson, S., U. Larsson & S. Johansson 1990a. Selective predation by herring and mysids, and zooplankton com-
munity structure in a Baltic Sea coastal area.]. Plankton. Res. 12:・ 1099← 1116.
Hansson, S., L. G. Rudstam & S. Johansson 1990b. Are ma-
rin巴planktonicinvertebrates food limited? The case of
Mysis mixta (Crustacea, Mysidacea) in the Baltic Sea. Oe-
cologia 84: 430 432. Hessler, R.R. 1985. Swimming in Crustacea. Trans. Roy.
Soc. Edinburgh, 76: 115 122. Hobson, E. S. & ]. R. Chess 1976. Trophic interactions
among fishes and zooplankters near shore at Santa Cata-
lina Island, California. Fish. Bull。74:567 598.
Irvine, K., B. Moss, M. Bales & D. Snook 1993. The chang-
ing ecosystem of a shallow, brackish lake, Hickling Broad,
Norfolk, U. K. I. Trophic relationships with special refer-
ence to the role of Neomysis integer. Freshwater Biol. 29:
119ー139.
Irvine, K., D目 Snook& B. Moss 1995. Life histories of Neo-
mysis integer, and its copepod prey, Eurytemora affinis, in a
eutrophic and brackish shallow lake. H》drobiologia304:
59 76.
Jansen, W. 1985. Stellung von Neomysis integer Leach (Cru-

70 日本プランクトン学会報第 51巻第 1号(2004)
stacea, Mysidacea) als Konsument im Nahrungsgefilge
der Darβ-Zingster Boddenkette (siidliche Ostee). Fisch-erei Faγschung 23: 55-59.
Jerling H. L. & T. H. Wooldridge 1995. Feeding of two mysid species on plankton in a temperate South African
estuary.]. Exp. Maγ. Biol. Ecol. 188:・ 243259.
Johannsson, 0. E., L. G. Rudstum & D. C. Lasenby 1994.
Mysis relicta: Assessment of metalimnetic feeding and implications for competition with fish in lakes Ontario
and Michigan. Can.]. Fish. Aquat. Sci. 51: 2591-2602.
Johannsson, 0. E., M. F. Leggett, L. G. Rudstam, M. R. Ser-vos, M.A. Mohammadian, G. Gal, R. M. Dermott & R.H.
Hesslein 2001. Diet of Mysisγelicta in Lake Ontario as revealed by stable isotope and gut content analysis. Can.
]. Fish. Aquat. Sci. 58: 1975 1986.
Johansson, K. U. I. & E. Hallberg 1992. The organization of
the olfactory lobes in Euphausiacea and Mysidacea (Crus-
tacea, Malacostraca). Zoomorphology. 112: 81-89.
Johnson, B. M., P. J. Martinez & J. D. Stockwell 2002.
Tracking trophic interactions in cold water reservoirs us-
ing naturally occurring isotopes. Trans. Am. Fish. Soc. 131: 1-13.
Johnson, W. S., M. Stevens & L. Watling 2001. Reproduc-
tion and development of marine p巴racarids.Adv. Mar. Biol. 39:・ 105-260.
Johnston, N. M., & D. A. Ritz 2001. Synchronous develop-
ment and release of broods by the swarming mysids
Anisomysis mixta australis, Paramesopodopsis rufa and Tenagomysis tasmaniae (Mysidacea: Crustacea). Mar. Ecol.
Prog. Ser. 223: 225 233.
Johnston, N. T. & D. C. Lasenby 1982. Diet and feeding of
Neomysis mercedis Holmes (Crustacea, Mysidacea) from the Fraser River Estuary, British Colombia. Can. ]. Zool.
60: 813 824.
Johnston, N. T. & T. G. Northcote 1989. Life-history in Neo-
mysis mercedis Holms (Crustacea) in the Fraser River
Estuary, British Columbia. Caη. ]. Zool. 67: 363 372.
Ketelaars, H. A. M., F. F. Lambregts-Van de Clundert, C. J. Carpentier, A. J. Wagenvoort & W. Hoogenboezem 1999.
Ecological E百ectsof the mass occurrence of the Ponto-
Caspian invader, Hemimysis anomala G. 0. Sars, 1907
(Crustacea: Mysidacea). in a freshwater storage reservoir
in the Netherlands, with notes on its autecology and new
records. Hydrobiologia 394: 233-248.
Kibirige, I. R. Perissinotto, C. Nozais 2003. Grazing rates
and feeding preferences of the mysid shrimp Gastrosaccus brev併ssurain a temporarily open estuary in South Africa
Mar. Ecol. Prag. Ser. 251: 201 210.
Kinsten, B. & P. Olsen 1982. Impact of Mysis relicta Loven introduction on the plankton of two mountain lakes,
Sweden. Rep. Inst. Freshw. Res. Drottningholm 59: 64ー74.
Ki0rboe, T., E. Saiz & M. Viitasalo 1996. Prey switching behaviour in the planklonic copepod Acartia tonsa. Mar.
Ecol. Prag. Ser. 143: 65 75.
Kreeger, K. E., D. A. Kreeger, C. L. Langdon & R. R. Lowry
1991. The nutritional value of Artemia and Tigriops cali-fornicus (Baker) for two Pacific mysid species, Metamysi-dopsis elongata (Holms) and Mysidopsis intii (Holmquist)目
]. Exp. Mar. Biol. Ecol. 148: 147-158.
Langeland, A., L. M. Carl, F. J. Hicks & B. Monroe 1991.
Impact of predation by Mysis relicta and fish on zooplank-
ton in four oligotrophic, north temperate lakes. Am. Fish.
Soc.めmp.9: 88 97.
Lasenby, D. C. & R.R. Langford 1973. Feeding and assimi-
lation of Mysis relicta. Limnol. Oceanogr. 18: 280-285. Lasenby, D. C., T. G. Northcote & M. Furst. 1986. Theory,
practice, and effects of Mysis relicta introductions to
North American and Scandinavian lakes. C包n.]. Fish. Aquat. Sci. 43: 1277 1284.
Laverack. M. S., D. M. Neil & R. M. Robertson 1977. Me tachronal exopodite beating in the mysid Pγaunus fiexu
osus: a quantitative analysis. Proc. Roy. Soc. London B 198:
139 154.
Leong, R. J. H. & C. P. O’Connell 1969. A laboratory study
of particulate and filter fe巴dingof the northern anchovy
(Engraulis mordax).]. Fish. Res. Bd. Can. 26: 557 582.
Lindstrom, M. 2000. Eye function of Mysidacea (Crusta-
cea) in the northern Baltic Sea.]. Exp. Mar. Biol. Ecol. 246: 85-101.
Lucas, C. E. 1936. On certain inter-relations between phy-
toplankton and zooplankton under experimental condi-
tions.]. Conseil Int. Explor. Mer. 11: 343 362.
Mann, K. H. 1988. Production and use of detritus in vari
ous freshwater, estuarine, and coastal marine ecosystems.
Limnol. Oceanogr. 33 (Part 2): 910 930.
Martinez P. J. & E. P. Bergersen 1991. Interactions of zoo-
plankton, Mysis relicta, and kokanees in Lake Granby, Colorado. Am. Fish. Soc. Symp. 9: 49-64.
Mauchline, J. 196 7. The biology of Schistomysis spritus (Crustacea, Mysidacea). ]. Mar. Biol. Assoc. U.K. 47: 383
396. Mauchline, J. 1968. The biology of Erythrops serrata and E. elegans (Crustacea, Mysidacea). ]. Mar. Biol. Assoc. U.K.
48: 455 464.
Mauchline, J. 1969. The biology of Leptomysis gracilis and L. lingiura (Crustacea, Mysidacea). ]. Mar. Biol. Assoc.
ιK. 49: 379 389. Mauchline, J. l 970a. The biology of Schistomysis ornata
(Crustacea, Mysidacea).]. Mar. Biol. Assoc. U.K. 50:・ 169
175. Mauchline, J. l 970b. The biology of Mysidopsis gibbosa, M.
didelphys and M. angusta (Crustacea, Mysidacea).]. Mar. Biol. Assoc. U. K. 50: 381-396.
Mauchline, J. 1971 a. The biology of Paramysis arenosa (Crustacea, Mysidacea).]. Mar. Biol. Assoc. U.K. 51: 339
345. Mauchline, J. 1971 b. The biology of Neomysis integer (Crus-tacea, Mysidacea).]. Maγ. Biol. Assoc. U.K. 51: 347 354.
Mauchline, J. 197lc. The biology of Praunus fiexuosus and P. neglectus (Crustacea, Mysidacea). ]. Mar. Biol. Assoc.
U.K. 51: 641 652. Mauchline, J. 1971 d. The biology of Schistomysis ken illei (Crustacea, Mysidacea).]. Mar. Biol. Assoc. U.K. 51: 653-
658. Mauchline, J. 1980. The biology of mysids and euphau-
siids. Adv. Mar. Biol. 18:・ 1-681.
Mauchline, J. & L. R. Fisher 1969. The biology of euphau-
siids. Adv. Mar. Biol. 7: 1 454目
Menden Deuer, S., & E. J. Lessard 2000. Carbon to volume
relationships for dinoflagellates, diatoms, and other pro-
tist plankton. Limnol. Oceanogr. 45:・ 569579.
Metillo, E. B. & D. A. Ritz 1993. Predatory feeding behav-

高橋: i炎水・沿岸域におけるアミ類の摂餌生態 71
iour in Paramesopodopsis rufa Fenton (Crustacea: Mysi-dacea).]. Exp. Mar. Biol. Ecol. 170: 127 141.
Metillo, E. B. & D. A. Ritz 1994. Comparative foregut func-
tional morphology of three co-occurring mysids (Crusta-cea: Mysidacea) from south司easternTasmania. ]. Mar. Biol. Assoc. U.K., 74: 323-336.
Metillo, E. B. & D. A. Ritz 2003目 Differentialchemosensory
feeding behaviour by three co-occurring mysids (Crusta-
cea: Mysidacea) from south-eastern Tasmania. Comp. Bio-
chem. Phisiol. A 134: 399 408. Mitchell, B. D. & W. D. Williams 1982. Population dynam
ics and production of Daphniαcarinata (King) and Simo-ceρhalus exsρinosus (Koch) in waste stabilization ponds. Aust.]. Mar. Freshw. Res. 33: 837-864.
Morgan, M. D. 1982. (editor). Ecology of Mysidacea目 Devel-
opments in Hydrobiology 10 (reprinted from Hydrobiolo gia vol. 93). Junk, Hague, 222 pp.
Mohammadian, M. A., S. Hansson, & B. T. De Stasio 1997.
Are marine planktonic inv巴rtebratesfood limited? The
functional response of Mysis mixta (Crustacea, Mysi-dacea) in the Baltic Sea. Mar. Ecol. Prog. Ser. 150: 113 119.
Mulkins, L. M., D. E. Jelinski, J. D. Karagatzides & A. Carr
2002. Carbon Isotope Composition of Mysids at a Terres-
trial-Marine Ecotone, Clayoquot Sound, British Columbia,
Canada. Est. Coast. Shelf Sci. 54: 669 675.
Mullin, M. M. & M. R目 Roman1986. In situ feeding of a
schooling mysid, Anisomysis sp., on Davies Reef-MECOR
井4.Bull. Mar. Sci. 39: 623 629. 村野正昭 1963.イサザアミ, Neomysisintermediaの漁業生物
学的研究 II.食性について.水産増殖 11:159 165.
村野正昭 1974.“あみ類と近底層プランクトン” pp.111ー128.
海洋プランクトン(丸茂隆三編入東京大学出版会,東京.
Murtaugh, P.A. 1981a. Selective predation by Neomysis 押iercedisin Lake Washington. Limnol. Oceanogr. 26: 445
453.
恥1urtaugh,P.A. 1981 b. Size-selective predation on Daph-
nia by Neomysis mercedis. Ecology 62: 894 900. Murtaugh, P.A. 1981c. Inferring properties of mysid pre
dation from injuries to Daphnia. Limnol. Oceanogr. 26:・
811 821.
Murtaugh, P.A. 1983. Mysid life history and seasonal var-
iation in predation pressure on zooplankton. Can. ]. Fish.
Aquat. Sci. 40: 1968 197 4. Nath, C. N. & N. K. Pillai 1973. The alimentary system of
the littoral mysid Gastrosaccus simulans (Van Beneden).].
Mar. Biol. Assoc. India 15: 577 586. Nero, R. W. & W. G. Sprules 1986a. Zooplankton species
abundance and biomass in relation to occurrence of Mysis γ・eficta (Malacostraca: Mysidacea). Can. ]. Fish. Aquat. Sci.
43: 420 434.
Nero, R. W. & W. G. Sprules 1986b. Predation by three gla-
cial opportunists on natural zooplankton communities.
Can. ]. Zool. 64: 57 64. Nesler, T. P. & E. P. Bergersen 1991. (editors) M》sidsin Fis-
heries: Hard Lessons from Headlong Introductions. Ameri-can. Fisheries. Socciety Symposium 9. American Fisher-
ies Society, Maryland. 199 pp.
Nilsson, D.-E. & R. F. Modlin 1994. A mysid shrimp carry-
ing a pair of binoculars.]. Exp. Biol. 189: 213-236.
Odum, W. E. & E. J. Heald 1972. Trophic analysis of an
estuarine mangrove community. Bull. Mar. Sci. 22: 671
738.
Parker, J. I. 1979. Predation by Mysis relicta on Pontoporeia hoyi: a food chain link of potential importance in the
Great Lakes.]. Great Lakes Res. 6: 164 166. Parker, M. & B. West 1979. The natural history of Neo-mysis integer (Leach) in Lough Furnace, Co. Mayo, a
brackish lough in the west of Ireland. Est. Coast. Mar. Sci. 8・157167.
Pechen'-Finenko, G. A. & T. V. Pavlovskaya 1973. Com-
parative importance of detritus and algae in the food of
Neomysis mirabilis. Hydrobiol.よ11:28-32.
Quirt, ]. & D. Lasenby 2002. Cannibalism and ontogenetic
changes in the response of the freshwater shrimp Mysis relicta to chemical cues from conspecific predators. Can.]. Zool. 80: 1022 1025.
Rademacher, K. & U. Kils 1996目 Predatory/prey dynamics
of fifteen-spined stickleback (Spinachia spinαchiα) and the
mysid (Neomysis integer). Arch. Fish. Mar. Res. 43: 171
181.
Ramcharan, C. W. & W. G. Sprules 1986. Visual predation
in Mysis relicta Loven. Limnol. Oceanogr. 31: 414 420.
Ramcharan, C. W., W. G. Sprules & R. W. Nero 1985. Notes
on tactile feeding behaviour of Mysis relicta Loven (Mala-
costraca: Mysidacea). Verh. Internat. Verein. Limnol. 22: 3215-3219.
Richards R. C. C. R. Goldman T. C. Frants & R. Wickwire
1975. Where have all the Daphnia gone? The decline of a major cladoceran in Lake Tahoe, California-Nevada.
Verh. Intertat. Verein. Limnol. 19: 835 842.
Ritz, D. A.,]. E. Osborn & A. E. ]. Ocken 1997. Influence of
food and predatory attack on mysid swarm dynamics.].
Mar. Biol. Assoc. U.K. 77: 31 42.
Roast, S. D.,]. Widdows & M. B. Jones 2000. Egestion rates of the estuarine mysid Neomysis integer (Peracarida: Mysi dacea) in relation to a variable environment.]. Exp. Mar.
Biol. Ecol. 245: 69 81.
Roman, M. R. 1977目 Feedingof th巴 copepodAcartia tonsa on the diatom Nitzschia closterium and brown algae (Fu-cus vesiculosus) detritus. Mar. Biol. 42: 149 155.
Rudstam, L. G., K. Danielsson, S. Hansson & S. Johansson
1989. Diel vertical migration and feeding patterns of
Mysis mixta (Crustacea, Mysidacea) in the Baltic Sea. Mar. Biol. 101:・ 43-52.
Rudstam, L. G., S. Hansson, S. Johansson & U. Larsson.
1992. Dynamics of planktivory in a coastal area of the
northern Baltic Sea. Mar. Ecol. Prog. Ser. 80: 159-173.
Rybock, J. T. 1978. Mysis relicta Loven. In Lake Tahoe:
Vertical distribution and nocturnal predation. Ph.D. the-
sis, Univ. of California, Davis. 116 pp.
Saiz, E. & T. Ki0rboe 1995. Predatory and suspension feed-
ing of the copepod Acartia tonsa in turbulent environ-ments. Mar Ecol. Prog. Ser. 122: 147-158.
佐藤弘康・村野正昭 1994.アミ類4種に見られた保育嚢から
脱落した幼生の取り込み.La meγ32: 71 74. Schabes, M. & W. Hamner 1992. Mysid locomotion and
feeding: kinematics and water-flow patterns of Antarcto-mysis sp., Acanthomysis scu伊ta,and Neomysis rayii. ].
Crustacean Biol. 12: 1-10. Sell, D. W. 1982. Size-frequency estimates of secondary
production by Mysis relicta in Lakes Michigan and
Huron. Hydrobiologia 93: 69 78.

72 日本フ。ランクトン学会報第 51巻第 l号(2004)
Siegfried, C. A. & M. E. Kopache 1980. Feeding of Neomysis
mercedis (Holmes). Biol. Bull. 159: 193 205. Sierszen, M. E. & A. S. Brooks 1982. The release of dis-
solved organic carbon as a result of diatom fragmenta-tion during feeding by Mysis relicta. Hydrobiologia 93: 155-161.
Spencer, C. N., B. R. McClelland & J. A. Stanford 1991.
Shrimp stocking, salmon collapse, and eagle displace-
ment:. Cascading interactions in the food web of a large
aquatic ecosystem. BioScience 41:・ 1421.
Spencer, C. N., D. S. Potter, R. T. Bukantis & J. A. Stanford
1999. Impact of predation by Mysis relicta on zooplank
ton in Flathead Lake, Montana, USA.]. Plankton Res. 21: 51-64.
Sprules, W. G. & J. E. Bowerman 1988. Omnivory and food
chain length in zooplankton food webs. Ecology 69: 418-
426.
Takahashi, K. & K. Kawaguchi 1998. Diet and feeding rhy-
thm of the sand burrowing mysids, Archaeomysis kokuboi
and A. japonica in Otsuchi Bay, northeastern Japan. Mar. Ecol. Frog. Ser. 162: 191ー199.
Talbot, M. M. B. & G. C. Bate 1986. Diel periodicities in cell characteristics of the surfzone diatom Anaulus birostra-
tus: their role in the dynamics of cell patches. Mar. Ecol. Frog. Ser. 32: 81 89.
Tattersall, W. M. & 0. S. Tattersall 1951. The British Mysi-dacea. Ray Society, London. 468 pp.
Toda, H., T. Arima, M. Takahashi & S. lchimura 1987. Phys
iological evaluation of temperature e百ecton the growth
process of the mysid, Neomysis intermedia Czerniavsky.]. Plankton Res. 9: 51-63.
Toda, H. & E. Wada 1990. Use of 15N/14N ratios to evaluate
the food source of the mysid, Neomysis intermedia Czer-niawsky, in a eutrophic lake in Japan. Hydrobiologia 194:
85 90目
Uitto, A., S. Kaitala, H. Kuosa & R. Pajuniemi 1995. Effect
of nutrient addition and predation of mysid shrimp (Neo-
mysis integer) on a plankton community in a short-term
enclosure experiment in the northern Baltic. Aqua Fen-
nica 25: 23 31. Val Klump, J., J. L. Kaster & M. E. Sierszen 1991. Mysis relicta assimilation of Hexachlorobiphenyl from sedi-
ments. Can. ]. Fish. Aq凶 t.Sci. 48:・ 284289.
Vanderploeg, H. A., J. A. Bowers, 0. Chapelski & H.K. Soo
1982. Measuring in situ predation by Mysis relicta and
observations on underdispers巴d microdistributions of
zooplankton. Hydrobiologia 93:・ 109 119.
Viherluoto, M., H. Kuosa, J. Flinkman & M. Viitasalo 2000.
Food utilisation of pelagic mysids, Mysis mixta and M.
relicta, during their growing season in the northern Baltic Sea Mar. Biol. 136: 553-559.
Viherluoto, M. & M. Viitasalo 200la. Temporal variability
in functional responses and prey selectivity of the pelagic
mysid, Mysis mixta, in natural prey assemblages. Mar. Biol. 138: 575-583.
Viherluoto, M. & Viitasalo, M. 2001 b. Effect of light on the
feeding rates of pelagic and littoral mysid shrimps: a
trade-off between feeding success and predation avoid-
ance.j. Exρ. Mar. Biol. Ecol. 261:・ 237244.
Viitasalo, M., J. Flinkman & M. Viherluoto 2001. Zooplank
tivory in the Baltic Sea: a comparison of prey selectivity
by Clupea harengus and Mysis mixta, with reference to prey escape reactions. Mar. Ecol. Frog. Ser. 216: 191ー200.
Viitasalo, M., T. Ki0rboe, J. Flinkman, L. W. Pedersen & A.
W. Visser 1998. Predation vulnerability of planktonic
copepods: consequences of predator foraging strat巴gies
and prey sensory abilities. Mar. Ecol. Frog. Ser. 175:・ 129
142.
Viitasalo, M. & M. Rautio 1998. Zooplanktivory by Pra-unus fiexuosus (Crustacea: Mysidacea): functional re-
sponse and prey selection in relation to prey escape re-sponses. Mar. Ecol. Frog. Seγ.174:77-87.
Webb, P., R. Perissinotto & T. H. Wooldridge 1987. Feeding
of Mesopodopsis slabberi (Crustacea: Mysidacea) on natu-rally occurring phytoplankton. Mar. Ecol. Frog. Ser. 38:
115 123.
Webb, P., R. Perissinotto & T. H. Wooldridge 1988. Diet
and feeding of Gastrosaccusρsammodytes (Crustacea, Mysidacea) with special reference to the surf diatom Ana-ulus birostratus. Mar. Ecol. Frog. Ser .. 45: 255-261.
Webb, P. & T. H. Wooldridge 1989. Diet elucidation: Sup-
plementary inferences from mysid feeding appendage
morphology. S. Afr.].. Zoo/. 24: 106 109. Webb, P. & T. H. Wooldridge 1990. Diel horizontal migra-
tion of Mesopodopsis slabberi (Crustacea: Mysidacea) in Algoa Bay, southern Africa. Mar .. Ecol. Pγng. Ser. 62: 73-
77. Weinright, P. F. & K. H. Mann 1982. Effect of antimicrobial
substances on the ability of th巴 mysidshrimp Mysis stenolepis to digest c巴llulose.Mar. Ecol. Frog. Ser. 7: 309-
313.
Wilhelm, F. M., J. Hamann & C. W. Burns 2002. Mysid pre-
dation on amphipods and Daphnia in a shallow coastal
lak巴.prey selection and effects of macrophytes. Can. ]. Fish. Aquat. Sci. 59: 1901 1907.
Wittmann, K. J. 1984. Ecophysiology of marsupial devel-
opm巴ntand reproduction in Mysidac巴a(Crustacea). Gee-anogr. Mar. Biol. Ann. Rev. 22: 393 428.
Wooldridge, T. H. 1989. The spatial and temporal distribu-
tion of mysid shrimps and phytoplankton accumulations
in a high energy surfzone. Vie Milieu 39:・ 127133.
Wooldridge T. & C. Bailey 1982. Euryhaline zooplankton
of the Sundays estuary and note on trophic relationships.
S. Afr.]. Zoo/. 17: 151 163. Wooldridge, T. H. & P. Webb 1988. Predator-prey interac-
tions between two species of estuarine mysid shrimps.
Mar .. Ecol. Frog. Ser .. 50: 21 28.
Yamada, H., T. Kawamura, T. Takeuchi & Y. Yamashita
1995. Effects of dietary condition on survival, growth and
reproduction of Acanthomysis mitsukurii (Crustacea, My-sidacea). Bull. Plankton Soc. Jpn. 42: 43-52.
Zagursky, G. & R. J. Feller 1985. Macrophyte detritus in
the winter di巴tof the estuarin巴mysid,Neomysis ameri cana. Estuaries 8: 355 362.
Zyblut, E. R. 1970. Long-term changes in the limnology
and macrozoolankton of a large British Columbia lake.].
βsh. Res. Bd. Can. 27:・ 12391250.


















