
유전성 유방암·난소암에서 BRCA1/BRCA2 돌연변이 검출을 위한 ... · 2019. 11. 14. · brca2 유전자는 27개 엑손으로 이루어져 있으며 크기는 10.3
Upload
luca-pellegriniCategory
view
227download
8
Emerging functions of BRCA2 in DNArecombinationLuca Pellegrini1 and Ashok Venkitaraman2
1Department of Biochemistry, University of Cambridge, Tennis Court Road, Cambridge CB2 1GA, UK2CR UK Department of Oncology & The Medical Research Council Cancer Cell Unit, University of Cambridge, Hills Road,
Cambridge CB2 2XZ, UK
Mutations in the breast cancer susceptibility protein
BRCA2 cause inherited susceptibility to breast, ovarian
and other cancers. There is now compelling experi-
mental evidence that a major biological function of
BRCA2 is the maintenance of chromosome structure
stability in dividing cells by regulation of steps in
recombination between homologous DNA strands.
Recent experimental findings shape the current models
for BRCA2 function, and structural and biochemical
advances shed new light on the interactions between
BRCA2, the RAD51 recombinase and single-stranded
DNA during DNA recombination.
Breast cancer will affect about one in every eight womenat some point in their lives. Approximately 10% of cases ofthe disease exhibit a familial pattern of inheritance [1].30–50% of the familial cases involve mutations in one oftwo breast cancer susceptibility genes BRCA1 or BRCA2,which were identified in 1994–1995 using reverse geneticapproaches [2,3]. These women (and in particular, thosecarrying BRCA1 mutations [4,5]) are also highly suscep-tible to ovarian cancer [6]. Moreover, in populations inwhich founder mutations affecting the BRCA genes occur,such as in Iceland or amongst Ashkenazi Jews, mutationcarriers are at significantly elevated risk of developingseveral different epithelial cancers, including carcinomasof the prostate, pancreas and male breast [7,8]. Recently,particular mutations affecting BRCA2 have been linked toFanconi anaemia, a rare inherited disease characterizedby haematological abnormalities, defective developmentand susceptibility to acute myeloid leukaemia [9,10].BRCA2 mutations in Fanconi anaemia give rise to acharacteristic syndrome associated with tumours of thebreast and brain [11].
BRCA1 and BRCA2 encode large proteins that lacksignificant resemblance to proteins of known function andlocalize to the nucleus in dividing cells from severaldifferent tissues [12,13]. Despite their shared associationwith an inherited breast and ovarian cancer syndrome,BRCA1 and BRCA2 are not homologous to one another.However, there is now compelling evidence that bothproteins work in pathways that are required for themaintenance of chromosome structure in dividing cells.Thus, a remarkable characteristic of murine or humancells that lack either of the BRCA proteins is the
accumulation of structurally aberrant or brokenchromosomes [14–16], a feature first recognized instudies on murine cells homozygous for a targetedtruncation in Brca2 [14].
BRCA proteins work in the biological response to DNA
damage
Why disruption of BRCA1 or BRCA2 should give rise tochromosomal instability and cancer predisposition has notyet been fully elucidated. Emerging evidence indicates,however, that both proteins are indispensable for thecellular response to DNA damage caused by exogenousagents, as well as endogenous damage that arises duringprocesses such as DNA replication or transcription(reviewed in Refs [12,17,18]). It is also clear that BRCA1and BRCA2 work in distinct molecular mechanismswithin a complex network of pathways that constitutethe DNA-damage response.
In particular, disruption of BRCA2 prevents theefficient repair of DNA double-strand breaks (DSBs),which are introduced into chromosomes by mechanismsthat involve recombination between homologous DNAstrands [19–21]. BRCA2 binds directly to RAD51 [22,23] –an enzyme that is central to recombination reactions – viashort, evolutionarily conserved motifs found within itslarge exon 11, which codes for the central portion ofBRCA2 [24]. These motifs of ,30 amino acid – termedBRC repeats – were first identified by computationalmethods [25]. When expressed in vitro, each BRC repeatcan interact with RAD51, although this capacity variesconsiderably between the different repeats [22,26].
RAD51 – the eukaryotic orthologue of bacterial RecA –is essential for DNA recombination. In vitro, it polymerizeson single-stranded DNA (ssDNA) to form a helicalnucleoprotein filament that mediates the strand-pairingand exchange reactions between homologous DNA thatunderlie recombination (reviewed in Ref. [27]). Biochemi-cal studies in which incubation of a BRC-repeat peptidewith RAD51 and DNA substrates led to disruption of theRAD51–DNA interaction and reversion of RAD51 to amonomeric form suggested that BRCA2 could regulate theability of RAD51 to form nucleoprotein filaments [28].
RAD51 recognition by the BRC repeats of BRCA2
The recent crystallographic determination of the structureof the RecA-homology region of human RAD51 bound to aBRC4-repeat polypeptide represents a step towards the
Corresponding author: Luca Pellegrini ([email protected]).
Available online 6 May 2004
Review TRENDS in Biochemical Sciences Vol.29 No.6 June 2004
www.sciencedirect.com 0968-0004/$ - see front matter q 2004 Elsevier Ltd. All rights reserved. doi:10.1016/j.tibs.2004.04.009
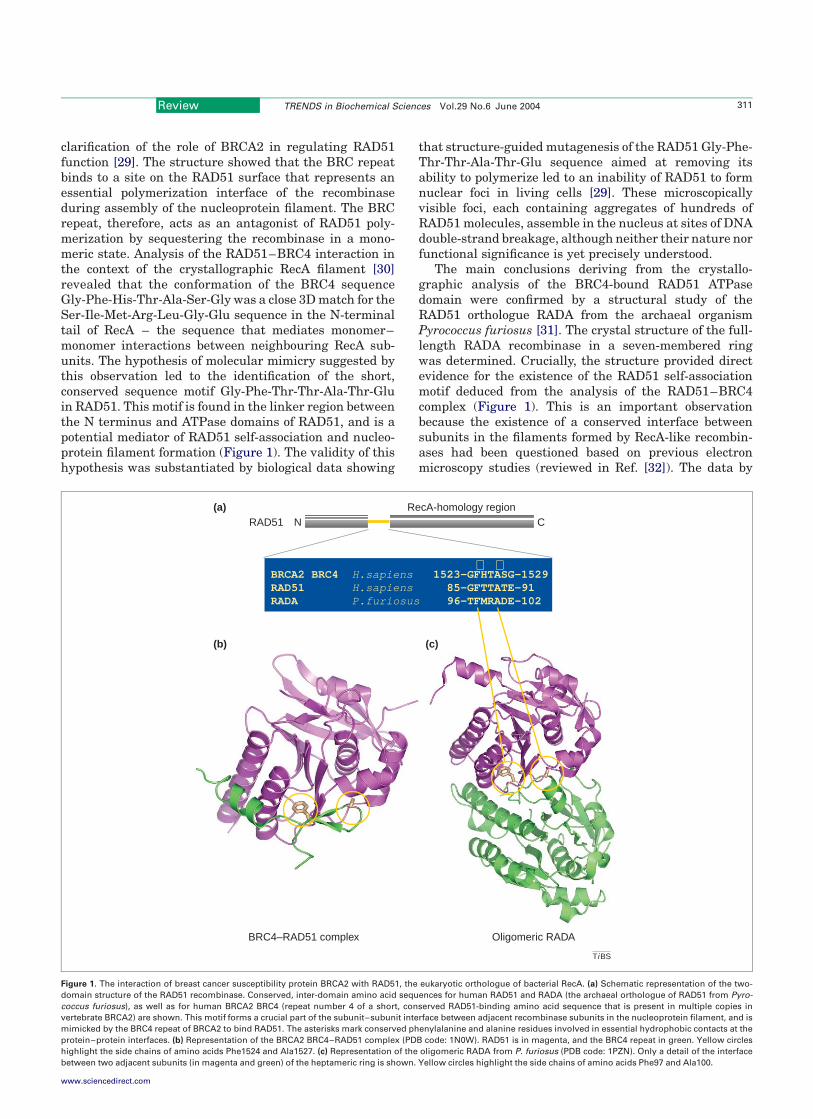
clarification of the role of BRCA2 in regulating RAD51function [29]. The structure showed that the BRC repeatbinds to a site on the RAD51 surface that represents anessential polymerization interface of the recombinaseduring assembly of the nucleoprotein filament. The BRCrepeat, therefore, acts as an antagonist of RAD51 poly-merization by sequestering the recombinase in a mono-meric state. Analysis of the RAD51–BRC4 interaction inthe context of the crystallographic RecA filament [30]revealed that the conformation of the BRC4 sequenceGly-Phe-His-Thr-Ala-Ser-Gly was a close 3D match for theSer-Ile-Met-Arg-Leu-Gly-Glu sequence in the N-terminaltail of RecA – the sequence that mediates monomer–monomer interactions between neighbouring RecA sub-units. The hypothesis of molecular mimicry suggested bythis observation led to the identification of the short,conserved sequence motif Gly-Phe-Thr-Thr-Ala-Thr-Gluin RAD51. This motif is found in the linker region betweenthe N terminus and ATPase domains of RAD51, and is apotential mediator of RAD51 self-association and nucleo-protein filament formation (Figure 1). The validity of thishypothesis was substantiated by biological data showing
that structure-guided mutagenesis of the RAD51 Gly-Phe-Thr-Thr-Ala-Thr-Glu sequence aimed at removing itsability to polymerize led to an inability of RAD51 to formnuclear foci in living cells [29]. These microscopicallyvisible foci, each containing aggregates of hundreds ofRAD51 molecules, assemble in the nucleus at sites of DNAdouble-strand breakage, although neither their nature norfunctional significance is yet precisely understood.
The main conclusions deriving from the crystallo-graphic analysis of the BRC4-bound RAD51 ATPasedomain were confirmed by a structural study of theRAD51 orthologue RADA from the archaeal organismPyrococcus furiosus [31]. The crystal structure of the full-length RADA recombinase in a seven-membered ringwas determined. Crucially, the structure provided directevidence for the existence of the RAD51 self-associationmotif deduced from the analysis of the RAD51–BRC4complex (Figure 1). This is an important observationbecause the existence of a conserved interface betweensubunits in the filaments formed by RecA-like recombin-ases had been questioned based on previous electronmicroscopy studies (reviewed in Ref. [32]). The data by
Figure 1. The interaction of breast cancer susceptibility protein BRCA2 with RAD51, the eukaryotic orthologue of bacterial RecA. (a) Schematic representation of the two-
domain structure of the RAD51 recombinase. Conserved, inter-domain amino acid sequences for human RAD51 and RADA (the archaeal orthologue of RAD51 from Pyro-
coccus furiosus), as well as for human BRCA2 BRC4 (repeat number 4 of a short, conserved RAD51-binding amino acid sequence that is present in multiple copies in
vertebrate BRCA2) are shown. This motif forms a crucial part of the subunit–subunit interface between adjacent recombinase subunits in the nucleoprotein filament, and is
mimicked by the BRC4 repeat of BRCA2 to bind RAD51. The asterisks mark conserved phenylalanine and alanine residues involved in essential hydrophobic contacts at the
protein–protein interfaces. (b) Representation of the BRCA2 BRC4–RAD51 complex (PDB code: 1N0W). RAD51 is in magenta, and the BRC4 repeat in green. Yellow circles
highlight the side chains of amino acids Phe1524 and Ala1527. (c) Representation of the oligomeric RADA from P. furiosus (PDB code: 1PZN). Only a detail of the interface
between two adjacent subunits (in magenta and green) of the heptameric ring is shown. Yellow circles highlight the side chains of amino acids Phe97 and Ala100.
Ti BS
N CRAD51
BRCA2 BRC4 H.sapiens 1523-GFHTASG-1529RAD51 H.sapiens 85-GFTTATE-91RADA P.furiosus 96-TFMRADE-102
∗ ∗
(b) (c)
BRC4–RAD51 complex Oligomeric RADA
RecA-homology region(a)
Review TRENDS in Biochemical Sciences Vol.29 No.6 June 2004 311
www.sciencedirect.com

Shin et al. [31] provide vital evidence in support of thehypothesis that RAD51 orthologues do indeed share acommon mechanism of self-assembly within the nucleo-protein filament. Further experimental support for theexistence of the proposed RAD51–RAD51 interface comesfrom fluorescent measurements of residues at the pro-posed RAD51 subunit interface in the presence of DNAand ATP [33]. Consequently, the available data suggestthat the proposed model for RAD51 self-association in thefilament is correct, but high-resolution crystallographic orelectron microscopic studies of the RAD51–DNA filamentwill be required to confirm this.
A significant advance in our understanding of BRCA2function was achieved through the determination of thecrystal structure of a large, evolutionarily conserved,carboxyl-terminal domain (CTD; ,800 amino acids) ofmurine BRCA2 bound to the smaller DSS1 (deleted in splithand/split foot) protein [34] (Figure 2). The study revealedthat the BRCA2 CTD folds into a multi-domain structurecomprising an N-terminal helical domain followed by threesuccessive oligonucleotide-binding (OB) domains, whichare widespread protein modules that can bind ssDNA in asequence-specific or non-specific manner. The helical andOB domains are tightly associated with each other, withsignificant surface area buried at their interfaces. In thestructure, DSS1 does not fold into a globular domain, andits extended conformation is intimately associated withthe helical and OB1 domains through a series ofhydrophobic and buried, charged interactions. Therefore,it seems likely that DSS1 has evolved to become anintegral part of the BRCA2 architecture, as demonstratedindirectly by the observation that DSS1 is needed toproduce correctly folded recombinant BRCA2 CTD [34].However, a further functional role for DSS1 in DNArepair that is independent of BRCA2 cannot be ruled out atthis stage.
In addition, the authors presented the structure of aBRCA2–DSS1 complex bound to a ssDNA oligonucleotide[34] (Figure 2). A continuous span of six deoxythymidinenucleotides binds in an extended conformation to CTD
domains OB2 and OB3. The phosphate groups are exposedto solvent, whereas the bases point towards the protein.The four bases at the 50 end of the oligonucleotide areinserted into a deep groove lined with aromatic side-chainsthat mediate the most significant protein–DNA inter-actions. Importantly, the authors presented biochemicalevidence for the ability of BRCA2 to bind ssDNA, and forthe stimulation of RAD51-dependent recombination bythe CTD. The stimulatory effect of the BRCA2 CTD in thein vitro assay depended on the presence of replicationprotein A (RPA), an abundant nuclear protein that alsorelies on OB domains to bind ssDNA in a non-specificmanner. Thus, BRCA2 appears to facilitate RAD51 fila-ment formation through a ‘mediator’ mechanism reminis-cent of that proposed for RAD52, which involves multipleinteractions with RAD51, RPA and ssDNA [35–37]. TheDNA-binding OB domains of BRCA2 could help thedisplacement of RPA from the ssDNA at the site ofdamage in preparation for DNA binding by RAD51. Aspecific, DNA-independent interaction of BRCA2 withRPA has also been reported, which, however, involves theN-terminal region of BRCA2 and, thus, lies outside theCTD [38].
The functional similarities between BRCA2 and RAD52as mediators of RAD51 loading on ssDNA are intriguing. Afunctional overlap between BRCA2 and RAD52 is sup-ported by the observation that the severe phenotypeassociated with mutations ofRAD52 in yeast [39], in whichBRCA2 is absent, is contrasted by the much weakerphenotype observed in vertebrate cells [40,41].
The second OB domain in the BRCA2 CTD includes alarge helical protrusion that culminates in a three-helixbundle containing a helix–turn–helix motif, which iswidely employed for binding double-stranded DNA(dsDNA). This raises the possibility that BRCA2 couldrecognize structures that involve a juxtaposition betweenssDNA and dsDNA. Indeed, there is evidence that DSBrepair by DNA recombination requires the processing offlush-ended DNA into a duplex with a 30-single-strandedtail through the action of an exonuclease that has not yet
Figure 2. Multi-domain structure of breast cancer susceptibility protein BRCA2. (a) Comparison of the domain structure of BRCA2 proteins from Mus musculus and Ustilago
maydis. The regions responsible for RAD51, DSS1 and DNA binding are indicated. (b) Representation of the BRCA2 (blue) CTD bound to DSS1 (yellow) and DNA (shown as
a space-fill model; PDB code: 1MJE). The region of homology between vertebrate and fungal BRCA2 sequences is highlighted in green. Abbreviations: aa, amino acids;
CTD, carboxyl-terminal domain; DSS1, deleted in split hand/split foot) protein; OB, oligonucleotide binding; ssDNA, single-stranded DNA.
Ti BS
(a)
ssDNA
DSS1
AlphaOB1
OB2
OB3
Tower(b)
BRH2 Ustilago maydis (1075 aa)
1N C
BRCA2 Mus musculus (3329 aa)
1 3α 2N C
RAD51 DSS1 ssDNA
BRC repeat Alpha domain OB domain Tower domain
Key:
Review TRENDS in Biochemical Sciences Vol.29 No.6 June 2004312
www.sciencedirect.com

been precisely identified, but might involve the MRE11–RAD50–Nbs1 complex.
Primitive BRCA2
The unexpected identification of the BRCA2-like geneBRH2 from the fungus Ustilago maydis has opened up anew avenue of BRCA2 research [42]. BRH2 is onlyapproximately a third of the size of vertebrate BRCAproteins, and contains a single BRC repeat sequence thatis linked to a slimmed-down version of the CTD. Thefungal orthologue of DSS1 has also been identified [43].Surprisingly, a comparison of fungal BRH2 with verte-brate BRCA2 sequences reveals that its most evolution-arily conserved segment spans OB1 and portions of the a
and OB2 domains adjacent to OB1, and does not thereforecoincide with the DNA-binding site identified in theBRCA2–DSS1–DNA co-crystals (Figure 2). It is possiblethat the ancestral BRCA2 protein had only a single OBdomain, corresponding to OB1 in vertebrate BRCA2, andthat its DNA-binding site underwent a profound trans-formation as BRCA2 evolved to acquire two further OBdomains. In support of this hypothesis, a CTD spanningonly the helical and OB1 domains displayed a weak affinityfor ssDNA [34], which might be considered a remnant of anancestral DNA-binding role. Alternatively, an improveddefinition of the CTD DNA-binding site might have to beobtained by structural studies employing nucleic acidmolecules that more accurately reflect the natural DNAsubstrates of BRCA2.
Why multiple BRC repeats?
Despite the considerable insight into BRCA2 and RAD51function provided by these studies, they probably repre-sent only an initial, partial answer to the question ofhow BRCA2 regulates RAD51 activity. In particular, theyhave highlighted an apparent functional contradiction: ifBRCA2 promotes homologous recombination in livingcells, why do BRCA2 peptides coding for single BRCrepeats inhibit RAD51 filament formation [28,29]? Theanswer to this question is not yet clear. The remarkabledifference in the competency of different BRC repeats todisrupt RAD51–DNA complexes further contributes to thepuzzle. [28] Important questions concerning the BRCA2–RAD51 interaction, such as its stoichiometric ratio or theaffinity of individual repeats for RAD51, are also stillunanswered. Collectively, these observations emphasizethe need to analyse the BRCA2–RAD51 interaction in thecontext of the multiple binding sites found in BRCA2.
An obvious starting point for the analysis is theobservation that vertebrate BRCA2 proteins have aconserved set of eight BRC-repeat sequences [44] (theonly exception is BRC3, which is missing in chickenBRCA2). One plausible reason for the presence of multiplecopies of the BRC repeat is the necessity to deliver asufficient number of RAD51 molecules onto the ssDNAsimultaneously, to stabilize the nascent nucleoproteinfilament. The mode of RAD51 recognition by the BRCrepeat offers a possible insight into how this might beachieved. The RAD51–BRC repeat interface is, in fact,dominated (not exclusively) by several hydrophobicinteractions that appear to be spatially separate and,
therefore, independent of each other. The presence orabsence of one or a few contact points would, in principle,enable the modulation of the affinity of the BRC repeat forRAD51 without destroying it altogether. Thus, the degreeof sequence divergence visible across BRC repeats mightrepresent an example of evolutionary tuning aimed atproducing binding sites with a range of affinities forRAD51. Under the appropriate conditions, BRC repeatswith weakest affinity could be the first to release RAD51onto the damaged DNA, thus facilitating formation of thenucleoprotein filament.
Why has U. maydis evolved a BRCA2-like protein thatis apparently capable of functioning with a single BRCrepeat? This difference between BRCA2 sequences is evenmore puzzling because the RAD51 nucleoprotein filamentis evolutionarily a highly conserved structure. Perhaps thefungal RAD51 binds ssDNA more avidly than its verte-brate counterparts, thus resembling RecA in its ability toform filaments. But such a difference in behaviour isunlikely given the strong evolutionary conservation ofthe RAD51 sequence (72.6% identity between human andU. maydis RAD51). Alternatively, it is possible that thefungal BRCA2 might be active in an oligomeric state, thuscompensating for the lower BRC-repeat number. Bolder –but more speculative – hypotheses might also be enter-tained. In one such scenario, the difference in number ofRAD51 interaction sites might be deceiving: in realityboth vertebrate and fungal BRCA2 might possess a singleBRC repeat that is competent for recruiting RAD51. Invertebrate BRCA2, the additional BRC repeats might fulfila different role, possibly representing binding sites forother recombination factors.
The BRCA2–RAD51 complex as a dynamic entity
The elucidation of the RAD51 interfaces that are respon-sible for self-association and BRCA2 binding in atomicdetail has enabled the design of mutations that selectivelydisrupt one kind of interaction while leaving the otherintact. This possibility has been exploited to investigatethe dynamic status of RAD51 in the nucleus of a living cell[45]. Fusion of wild-type and mutant versions of RAD51 togreen fluorescent protein followed by in vivo analysis bynucleoplasm photobleaching has enabled the identifi-cation of at least three different pools of nuclear RAD51:one larger, relatively mobile pool, and two smaller oneswherein RAD51 is immobilized through binding to BRCA2or self-association. The existence of two nuclear RAD51pools with distinct kinetic properties was also indepen-dently reported [46]. Importantly, arrest of DNA replica-tion induced by hydroxyurea triggers the transientmobilization of the BRCA2-bound fraction. Thus, itappears that one role of BRCA2 in recombination is totransport RAD51 to the nuclear sites in which its action isrequired, as previously proposed [17], although recentevidence also points to the existence of a BRCA2-independent route of RAD51 transport to the nucleus [47].
The implications of the study by Yu et al. [45], however,go beyond the ‘chaperoning’ function of BRCA2. Toovercome the difficulty of a purely repressive role of theBRCA2 interaction on RAD51 filament formation, onecould postulate that the BRCA2–RAD51 complex might
Review TRENDS in Biochemical Sciences Vol.29 No.6 June 2004 313
www.sciencedirect.com
exist in multiple states. In a putative alternative state,induced by DNA damage, the complex would be competentfor loading RAD51 onto ssDNA in preparation for DNArepair. Yu et al. [45] provide experimental evidencesuggesting that the BRCA2–RAD51 complex might, infact, exist in two states. Investigating the molecularnature of the changes that enable the BRCA2–RAD51complex to alternate between different states will be animportant focus for future research.
A mechanism for BRCA2-dependent delivery of RAD51
onto damaged DNA
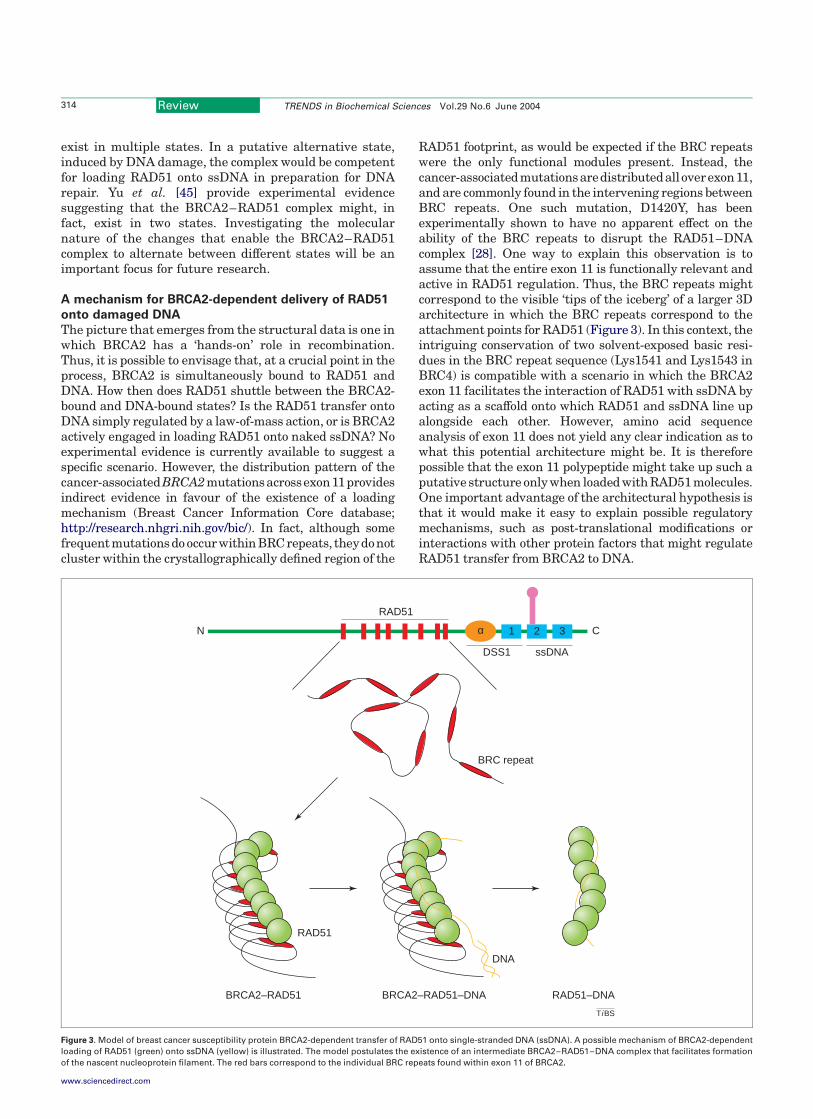
The picture that emerges from the structural data is one inwhich BRCA2 has a ‘hands-on’ role in recombination.Thus, it is possible to envisage that, at a crucial point in theprocess, BRCA2 is simultaneously bound to RAD51 andDNA. How then does RAD51 shuttle between the BRCA2-bound and DNA-bound states? Is the RAD51 transfer ontoDNA simply regulated by a law-of-mass action, or is BRCA2actively engaged in loading RAD51 onto naked ssDNA? Noexperimental evidence is currently available to suggest aspecific scenario. However, the distribution pattern of thecancer-associatedBRCA2mutationsacrossexon11providesindirect evidence in favour of the existence of a loadingmechanism (Breast Cancer Information Core database;http://research.nhgri.nih.gov/bic/). In fact, although somefrequent mutations do occurwithin BRCrepeats, theydonotcluster within the crystallographically defined region of the
RAD51 footprint, as would be expected if the BRC repeatswere the only functional modules present. Instead, thecancer-associatedmutationsaredistributedalloverexon11,and are commonly found in the intervening regions betweenBRC repeats. One such mutation, D1420Y, has beenexperimentally shown to have no apparent effect on theability of the BRC repeats to disrupt the RAD51–DNAcomplex [28]. One way to explain this observation is toassume that the entire exon 11 is functionally relevant andactive in RAD51 regulation. Thus, the BRC repeats mightcorrespond to the visible ‘tips of the iceberg’ of a larger 3Darchitecture in which the BRC repeats correspond to theattachment points for RAD51 (Figure 3). In this context, theintriguing conservation of two solvent-exposed basic resi-dues in the BRC repeat sequence (Lys1541 and Lys1543 inBRC4) is compatible with a scenario in which the BRCA2exon 11 facilitates the interaction of RAD51 with ssDNA byacting as a scaffold onto which RAD51 and ssDNA line upalongside each other. However, amino acid sequenceanalysis of exon 11 does not yield any clear indication as towhat this potential architecture might be. It is thereforepossible that the exon 11 polypeptide might take up such aputative structure only when loaded with RAD51 molecules.One important advantage of the architectural hypothesis isthat it would make it easy to explain possible regulatorymechanisms, such as post-translational modifications orinteractions with other protein factors that might regulateRAD51 transfer from BRCA2 to DNA.
Figure 3. Model of breast cancer susceptibility protein BRCA2-dependent transfer of RAD51 onto single-stranded DNA (ssDNA). A possible mechanism of BRCA2-dependent
loading of RAD51 (green) onto ssDNA (yellow) is illustrated. The model postulates the existence of an intermediate BRCA2–RAD51–DNA complex that facilitates formation
of the nascent nucleoprotein filament. The red bars correspond to the individual BRC repeats found within exon 11 of BRCA2.
Ti BS
RAD51
1 2 3αN C
RAD51
DSS1 ssDNA
BRC repeat
DNA
BRCA2–RAD51 BRCA2–RAD51–DNA RAD51–DNA
Review TRENDS in Biochemical Sciences Vol.29 No.6 June 2004314
www.sciencedirect.com

Future perspectives
Since the first report of its identification in 1994, the geneencoding BRCA2 has become the focus of intense attentionfrom the cancer biology community, in the hope thatunderstanding its function would provide importantinsight into the molecular mechanism underlying thetumourigenic process. Ten years later, we have learnedthat BRCA2 has a vital role in genome maintenancethrough a direct involvement in the process of DNArecombination.
To fully understand the role of BRCA2 in recombina-tion, there are three immediate challenges for the future.First, the initial hypothesis of the inhibitory effect ofRAD51 binding, derived from studies on single BRCrepeats, will have to be confirmed and carried further byanalysing the BRCA2–RAD51 interaction in the context ofmultiple BRC repeats provided by the large central regionof BRCA2. Second, we will need to understand how BRCA2coordinates its RAD51- and ssDNA-binding activities tofacilitate the orderly transfer of RAD51 onto DNA and theestablishment of the nucleoprotein filament. To thispurpose, primitive organisms harbouring a simpler ver-sion of the BRCA2 protein will provide useful modelsystems. Third, it will be necessary to investigate themolecular nature of the signals deployed by the cell tocontrol the mobilization of the BRCA2–RAD51 complex,the delivery of its RAD51 cargo at the site of damage and,possibly, the disassembly of the established filament aftersuccessful completion of the recombination reactions.
References
1 Nathanson, K.L. et al. (2001) Breast cancer genetics: what we knowand what we need. Nat. Med. 7, 552–556
2 Miki, Y. et al. (1994) A strong candidate for the breast and ovariancancer susceptibility gene BRCA1. Science 266, 66–71
3 Wooster, R. et al. (1995) Identification of the breast cancer suscepti-bility gene BRCA2. Nature 378, 789–792
4 Easton, D.F. et al. (1995) Breast and ovarian cancer incidence inBRCA1-mutation carriers. Breast Cancer Linkage Consortium. Am.J. Hum. Genet. 56, 265–271
5 Thompson, D. and Easton, D.F. (2002) Cancer incidence in BRCA1mutation carriers. J. Natl. Cancer Inst. 94, 1358–1365
6 No authors listed (1999) Cancer risks in BRCA2 mutation carriers. TheBreast Cancer Linkage Consortium. J. Natl. Cancer Inst. 91, 1310–1316
7 Struewing, J.P. et al. (1997) The risk of cancer associated with specificmutations of BRCA1 and BRCA2 among Ashkenazi Jews. N. Engl. J.Med. 336, 1401–1408
8 Thorlacius, S. et al. (1996) A single BRCA2 mutation in male andfemale breast cancer families from Iceland with varied cancerphenotypes. Nat. Genet. 13, 117–119
9 Joenje, H. and Patel, K.J. (2001) The emerging genetic and molecularbasis of Fanconi anaemia. Nat. Rev. Genet. 2, 446–457
10 Howlett, N.G. et al. (2002) Biallelic inactivation of BRCA2 in Fanconianemia. Science 297, 606–609
11 Offit, K. et al. (2003) Shared genetic susceptibility to breast cancer,brain tumors, and Fanconi anemia. J. Natl. Cancer Inst. 95,1548–1551
12 Scully, R. and Livingston, D.M. (2000) In search of the tumour-suppressor functions of BRCA1 and BRCA2. Nature 408, 429–432
13 Welcsh, P.L. et al. (2000) Insights into the functions of BRCA1 andBRCA2. Trends Genet. 16, 69–74
14 Patel, K.J. et al. (1998) Involvement of Brca2 in DNA repair. Mol. Cell1, 347–357
15 Xu, X. et al. (1999) Centrosome amplification and a defective G2-M cellcycle checkpoint induce genetic instability in BRCA1 exon 11 isoform-deficient cells. Mol. Cell 3, 389–395
16 Gretarsdottir, S. et al. (1998) BRCA2 and p53 mutations in primarybreast cancer in relation to genetic instability. Cancer Res. 58,859–862
17 Venkitaraman, A.R. (2002) Cancer susceptibility and the functions ofBRCA1 and BRCA2. Cell 108, 171–182
18 Jasin, M. (2002) Homologous repair of DNA damage and tumori-genesis: the BRCA connection. Oncogene 21, 8981–8993
19 Moynahan, M.E. et al. (2001) BRCA2 is required for homology-directedrepair of chromosomal breaks. Mol. Cell 7, 263–272
20 Xia, F. et al. (2001) Deficiency of human BRCA2 leads to impairedhomologous recombination but maintains normal nonhomologous endjoining. Proc. Natl. Acad. Sci. U. S. A. 98, 8644–8649
21 Tutt, A. et al. (2001) Mutation in Brca2 stimulates error-pronehomology-directed repair of DNA double-strand breaks occurringbetween repeated sequences. EMBO J. 20, 4704–4716
22 Wong, A.K. et al. (1997) RAD51 interacts with the evolutionarilyconserved BRC motifs in the human breast cancer susceptibility genebrca2. J. Biol. Chem. 272, 31941–31944
23 Chen, P.L. et al. (1998) The BRC repeats in BRCA2 are critical forRAD51 binding and resistance to methyl methanesulfonate treatment.Proc. Natl. Acad. Sci. U. S. A. 95, 5287–5292
24 Bignell, G. et al. (1997) The BRC repeats are conserved in mammalianBRCA2 proteins. Hum. Mol. Genet. 6, 53–58
25 Bork, P. et al. (1996) Internal repeats in the BRCA2 protein sequence.Nat. Genet. 13, 22–23
26 Chen, C.F. et al. (1999) Expression of BRC repeats in breast cancercells disrupts the BRCA2–Rad51 complex and leads to radiationhypersensitivity and loss of G(2)/M checkpoint control. J. Biol. Chem.274, 32931–32935
27 West, S.C. (2003) Molecular views of recombination proteins and theircontrol. Nat. Rev. Mol. Cell Biol. 4, 435–445
28 Davies, A.A. et al. (2001) Role of BRCA2 in control of the RAD51recombination and DNA repair protein. Mol. Cell 7, 273–282
29 Pellegrini, L. et al. (2002) Insights into DNA recombination from thestructure of a RAD51–BRCA2 complex. Nature 420, 287–293
30 Story, R.M. et al. (1992) The structure of the E. coli recA proteinmonomer and polymer. Nature 355, 318–325
31 Shin, D.S. et al. (2003) Full-length archaeal Rad51 structure andmutants: mechanisms for RAD51 assembly and control by BRCA2.EMBO J. 22, 4566–4576
32 Egelman, E.H. (2003) A tale of two polymers: new insights into helicalfilaments. Nat. Rev. Mol. Cell Biol. 4, 621–630
33 Selmane, T. et al. (2004) Identification of the subunit–subunitinterface of Xenopus Rad51.1 protein: similarity to RecA. J. Mol.Biol. 335, 895–904
34 Yang, H. et al. (2002) BRCA2 function in DNA binding andrecombination from a BRCA2–DSS1–ssDNA structure. Science 297,1837–1848
35 Benson, F.E. et al. (1998) Synergistic actions of Rad51 and Rad52 inrecombination and DNA repair. Nature 391, 401–404
36 Shinohara, A. and Ogawa, T. (1998) Stimulation by Rad52 of yeastRad51-mediated recombination. Nature 391, 404–407
37 New, J.H. et al. (1998) Rad52 protein stimulates DNA strand exchangeby Rad51 and replication protein A. Nature 391, 407–410
38 Wong, J.M. et al. (2003) Interaction between BRCA2 and replicationprotein A is compromised by a cancer-predisposing mutation inBRCA2. Oncogene 22, 28–33
39 Game, J.C. (1993) DNA double-strand breaks and the RAD50-RAD57genes in Saccharomyces. Semin. Cancer Biol. 4, 73–83
40 Yamaguchi-Iwai, Y. et al. (1998) Homologous recombination, but notDNA repair, is reduced in vertebrate cells deficient in RAD52. Mol.Cell. Biol. 18, 6430–6435
41 Rijkers, T. et al. (1998) Targeted inactivation of mouse RAD52 reduceshomologous recombination but not resistance to ionizing radiation.Mol. Cell. Biol. 18, 6423–6429
42 Kojic, M. et al. (2002) BRCA2 homolog required for proficiency in DNArepair, recombination, and genome stability in Ustilago maydis. Mol.Cell 10, 683–691
43 Kojic, M. et al. (2003) The BRCA2-interacting protein DSS1 is vital forDNA repair, recombination, and genome stability in Ustilago maydis.Mol. Cell 12, 1043–1049
44 Lo, T. et al. (2003) Sequence fingerprints in BRCA2 and RAD51:
Review TRENDS in Biochemical Sciences Vol.29 No.6 June 2004 315
www.sciencedirect.com

implications for DNA repair and cancer. DNA Repair (Amst.) 2,1015–1028
45 Yu, D.S. et al. (2003) Dynamic control of Rad51 recombinase byself-association and interaction with BRCA2. Mol. Cell 12,1029–1041
46 Essers, J. et al. (2002) Nuclear dynamics of RAD52 group homologousrecombination proteins in response to DNA damage. EMBO J. 21,2030–2037
47 Tarsounas, M. et al. (2003) BRCA2-dependent and independentformation of RAD51 nuclear foci. Oncogene 22, 1115–1123
Getting animated with parasites!
Interested in the molecular cell biology of host–parasite interactions? Then take a look at the online animations produced by Trends
in Parasitology, one of our companion TRENDS journals. The pictures below are snapshots from two of our collection of animations
revealing the latest advances in understanding parasite life cycles. Check them out today!
Microsporidia: how can they invade other cells?By C. Franzen [(2004) Trends Parasitol. 20, 10.1016/j.pt.2004.04.009]
http://archive.bmn.com/supp/part/franzen.html
Interaction of Leishmania with the host macrophageBy E. Handman and D.V.R. Bullen [(2002) Trends Parasitol. 18, 332–334]
http://archive.bmn.com/supp/part/swf012.html
Review TRENDS in Biochemical Sciences Vol.29 No.6 June 2004316
www.sciencedirect.com


















