
ELEMENTOS DEL PAISAJE SOBRE LA DIVERSIDAD DE … · PALMA DE ACEITE (Elaeis guineensis ......
116
EFECTO DE LA ESTRUCTURA DEL CULTIVO DE PALMA DE ACEITE ( Elaeis guineensis ), Y OTROS ELEMENTOS DEL PAISAJE SOBRE LA DIVERSIDAD DE AVES EN DOS MUNICIPIOS DE META (COLOMBIA) DIANA PATRICIA TAMARIS TURIZO, MSc. UNIVERSIDAD NACIONAL DE COLOMBIA Facultad de Ciencias, Departamento de Biología Bogotá, DC., Colombia 2017
Transcript of ELEMENTOS DEL PAISAJE SOBRE LA DIVERSIDAD DE … · PALMA DE ACEITE (Elaeis guineensis ......

EFECTO DE LA ESTRUCTURA DEL CULTIVO DE
PALMA DE ACEITE (Elaeis guineensis), Y OTROS
ELEMENTOS DEL PAISAJE SOBRE LA DIVERSIDAD DE
AVES EN DOS MUNICIPIOS DE META (COLOMBIA)
DIANA PATRICIA TAMARIS TURIZO, MSc.
UNIVERSIDAD NACIONAL DE COLOMBIA
Facultad de Ciencias, Departamento de Biología
Bogotá, DC., Colombia
2017

EFECTO DE LA ESTRUCTURA DEL CULTIVO DE
PALMA DE ACEITE (Elaeis guineensis), Y OTROS
ELEMENTOS DEL PAISAJE SOBRE LA DIVERSIDAD DE
AVES EN DOS MUNICIPIOS DE META (COLOMBIA)
DIANA PATRICIA TAMARIS TURIZO, MSc.
Tesis presentada como requisito paricial para optar al título de:
Doctora en Ciencias Biología
Director:
D.C., M.Sc. Hugo Fernando López Arévalo
UNIVERSIDAD NACIONAL DE COLOMBIA
Facultad de Ciencias, Departamento de Biología
Bogotá, DC., Colombia
2017
(Dedicatoria o lema)

VII
“Hay grandeza en esta concepción de que la vida, con sus
diferentes facultades, fue originariamente alentada por el
Creador en unas cuantas formas o en una sola, y que,
mientras este planeta ha ido girando según la constante ley de
la gravitación, se han desarrollado y se están desarrollando, a
partir de un comienzo tan sencillo, infinidad de formas cada
vez más bellas y maravillosas”.
-Charles Darwin.


I
Agradecimientos
A la Unversidad Nacional de Colombia y al Institudo de Ciencias Naturales por ayudarme a crecer
como profesional.
Un especial agraedecimiento a mi director de tesis Dr. Hugo F. López Arévalo, porque todos sus
consejos, orientaciones y enseñanzas fueron fundamentales para mi formación y para el desarrollo
y conclusión de esta investigación.
A Colciencias por otórgame el crédito educativo a través del programa Estudios de Doctorado
Nacional y a la Universidad del Magdalena por la comisión de estudios que me brindaron para este
fin.
Idea Wild por la donación de equipos e insumos que facilitaron el trabajo de campo.
Al grupo de Investigación en Conservación y Manejo de Vida Silvestre de la Universidad Nacional
de Colombia y los profesores Olga Lucía Montenegro y Pedro Sánchez Palomino, quienes me
aconsejaron, asesoraron y guiaron en el momento oportuno.
En el INECOL (Xalapa, Veracruz, México), expreso un especial agradecimiento a la Dra. Fabiola
López, Roberto Monroy, Roberto Monroy Rosario Landgrave y al grupo de Investigación de
Ecología Funcional, por recibirme durante mi estancia y por el valioso acompañamiento que me
brindaron.
A mi familia de mamíferos Cata, Darwin, Karen, Catherin, Arlensiu, Jorge y Cindy, quienes
adoptaron a una ornitóloga y me ayudaron en todo lo que necesité de ustedes.
A mi compañera de clases Zulima Isabel y a mi compañero de investigación y de trabajo de campo
Nabhi Romero, porque en cada momento encontré en ustedes un valioso apoyo para el desarrollo
de esta investigación.
El Grupo de Ornitología de la Universidad Nacional (GOUN) también fue un pilar de apoyo en
esta investigación, de manera especial les agradezco a Nicolás Téllez, Camilo Alfonso, Gina Díaz,
German Gutiérrez, Ángela Mayorga y Miguel Ángel Muñoz, quienes trabajaron de manera
comprometida en el trabajo de campo.

II
A los propietarios y administradores de Inversiones Tierra Viva e Inversiones La Mejorana por la
autorización y todo el apoyo logístico que nos brindaros para el desarrollo de la investigación, de
manera particular al Ing. Jairo Jeréz Jaramillo, Birman Correal, Juan Manuel Correal y al Sr.
Freddy Capera.
Expreso todo mi agradecimiento y dedicación de esta obra a mi querida familia: mi hijo Daniel
Alejandro, la más hermosa bendición de Dios, el motor de mi vida y fuente de alegría en mi diario
vivir. Al cómplice de mis sueños y compañero de mi vida, Daniel José, por estar a siempre a mi
lado y brindarme todo su amor, apoyo y comprensión en todo momento. A mi querida madre: la
Sra. Berta, mami, cualquier palabra que exprese no es suficiente para darte las agracias por el
apoyo que me has brindado para alcanzar esta meta. A mi papá y mis queridos hermanos, y resto
de familia. También agradezco con todo mi corazón a mi familia de Bogotá: Merita, Tina, la Sra.
Edna y demás miembros de esta hermosa familia quienes me abrieron su corazón y me han
ayudado enormemente durante mi estancia en esta ciudad.
Mis amistades en Santa Marta siempre estuvieron muy pendientes de mi, dándome fuerzas y ánimo
en todo momento: Luza, Judi y Jorge, muchas gracias mis queridos amigos.
Por último, a todas aquellas personas que de una u otra forma han hecho de este proyecto una
realidad y un logro alcanzado.

VII
Contenido
Pág.
Lista de Figuras ............................................................................................................... V Lista de Tablas ............................................................................................................... VI
Resumen .......................................................................................................................... 7
Abstract............................................................................................................................ 8
PRESENTACIÓN .............................................................................................................. 9
INTRODUCCIÓN GENERAL .......................................................................................... 10 Ecología de las comunidades de aves ............................................................................... 10 Avifauna de la Región Oriental de Colombia .................................................................... 11 Cultivos de Palma de Aceite ........................................................................................... 12 Problema ...................................................................................................................... 15 Los cultivos de palma de aceite en Colombia .................................................................... 17 Aves en Sistemas Agrícolas: Diversidad e implicaciones del paisaje .................................... 18 OBJETIVO GENERAL ................................................................................................. 20 Preguntas de Investigación ............................................................................................. 20 Métodos Generales ........................................................................................................ 21 Área de Estudio ............................................................................................................. 21 Referencias ................................................................................................................... 24
CAPÍTULO 1. EFECTO DE LA ESTRUCTURA DEL CULTIVO DE PALMA DE ACEITE Elaeis guineensis SOBRE LA DIVERSIDAD DE AVES EN UN PAISAJE TROPICAL
DE COLOMBIA ............................................................................................................... 30 Resumen ...................................................................................................................... 31 Introducción ................................................................................................................. 32 Métodos ....................................................................................................................... 34 Resultados .................................................................................................................... 37 Discusión ..................................................................................................................... 38 Referencias ................................................................................................................... 47
CAPÍTULO 2. USO DE RECURSOS DE AVES EN PAISAJES DOMINADOS POR CULTIVOS DE PALMA DE ACEITE (Elaeis guineensis) .............................................. 51
Resumen ...................................................................................................................... 52 Introducción ................................................................................................................. 53 Métodos ....................................................................................................................... 54 Resultados .................................................................................................................... 58 Discusión ..................................................................................................................... 59 Referencias ................................................................................................................... 64

IV
CAPÍTULO 3. CARACTERIZACIÓN DE GREMIOS Y RASGOS FUNCIONALES DE AVES EN PAISAJES DOMINADOS POR CULTIVOS DE PALMA DE ACEITE (Elaeis
guineensis) .....................................................................................................................68 Resumen ....................................................................................................................... 69 Introducción .................................................................................................................. 70 Métodos ........................................................................................................................ 72 Resultados .................................................................................................................... 74 Discusión ...................................................................................................................... 75 Referencias ................................................................................................................... 80
CAPÍTULO 4. RESPUESTA DE UNA COMUNIDAD DE AVES ANTE EL ESTABLECIMIENTO DEL CULTIVO DE PALMA DE ACEITE: UN ENFOQUE A
ESCALA DE PAISAJE ....................................................................................................84 Resumen ..................................................................................................................... 85 4.1 Introducción ....................................................................................................... 86 4.2 Métodos ............................................................................................................. 88 4.3 Resultados .......................................................................................................... 93 4.4 Discusión ........................................................................................................... 95 1.1 Referencias ....................................................................................................... 103
CAPÍTULO 5. CONCLUSIONES Y RECOMENDACIONES .......................................... 108 Conclusiones ............................................................................................................... 108 Recomendaciones ........................................................................................................ 110
SÍNTESIS GENERAL .................................................................................................... 112

V
Lista de Figuras
Figura 1 Ubicación de las fincas Tierra Viva (TIV) y La Mejorana (MEJ) en San Martín de los
Llanos y Acacias, Meta ...................................................................................................... 21
Figura 2.3 Índice de selectividad de recursos para gremios de alimentación de aves en el cultivo de
palma de aceite. a) hojas de palma grande, b) troncos de palma grande, c) hojas de palma mediana,
d) hojas de palma pequeña. ................................................................................................ 63
Figura 1. 1 Distribución de la abundancia de especies en cada clase de altura del cultivo de palma
de aceite Elaeis guineensis ................................................................................................. 43
Figura 1. 2 Distribución de la abundancia promedio de especies en cada clase de altura del cultivo
de palma de aceite Elaeis guineensis ................................................................................... 43
Figura 1. 3 Curva de acumulación de especies de aves: A) asociadas a todo el cultivo de palma de
aceite y B) a cada clase de altura ......................................................................................... 44
Figura 1. 4 Modelo de distribución de abundancia de especies en aves asociadas a cultivos de
palma de aceite Elaeis guineensis. G: grande, M: mediano, P: pequeño, Total: todas las clases de
altura ............................................................................................................................... 45
Figura 1. 5 Análisis de correspondencia rectificado (DCA) de los gremios de alimentación de aves
en las clases de altura del cultivo de palma de aceite Elaeis guineensis .................................... 45
Figura 2. 1 Frecuencias de actividades realizadas por las aves en el cultivo de palma de aceite. P:
descanso en percha, C: canto, F: forrajeo, A: alimentación ..................................................... 61
Figura 2. 2 Proyección de variables del cultivo y uso proporcional de recursos, en el cultivo en
palma de aceite ................................................................................................................. 61
Figura 3. 1 Coeficiente de la distancia la Euclidiana A) de la abundancia proporcional de
individuos en cada gremio de alimentación y B) de los gremios de alimentación en relación al
bosque y las clases de altura del cultivo de palma de aceite .................................................... 78
Figura 3. 2 Comparación de rasgos funcionales (talla-peso) en aves asociadas a bosque y cultivo de
palma .............................................................................................................................. 78
Figura 3. 3 Comparación de rasgos funcionales (medidas del pico) en aves asociadas a bosque y
cultivo de palma. B: bosque, P: cultivo de palma .................................................................. 79
Figura 4. 1 Mapa elementos (A) y coberturas (B) y del paisaje en el predio Tierra Viva ............. 99
Figura 4. 2 Mapa elementos (A) y coberturas (B) y del paisaje en el predio Mejorana ..............100
Figura 4. 3 Distribución de gremios de alimentación de aves en función a coberturas y métricas del
paisaje MEJ_Bga: ............................................................................................................101
Figura 4. 4. Análisis de correspondencia rectificado (DCA) de la diversidad de aves y rasgos
morfológicos con relación a los tipos de cobertura en los predios Mejorana y Tierra Viva.........102

VI Título de la tesis o trabajo de investigación
Lista de Tablas
Pág. Tabla 1.1. Cálculos de diversidad de aves asociadas a cultivos de palmas grandes, medianas y
pequeñas del cultivo de palma de aceite. ............................................................................... 46
Tabla 1 2. Gremios de alimentación de aves asociadas a cultivos de palma de aceite. ................. 46
Tabla 2 1. Variables del cultivo y proporción de uso del cultivo de palma de aceite ................... 57
Tabla 3 1. Composición, riqueza y abundancia de individuos de cada gremio de alimentación
asociado a bosque y cultivo de palma de aceite. ..................................................................... 77
Tabla 4. 1 Elementos del paisaje y coberturas asociadas en las fincas Tierra Viva y Mejorana .... 90
Tabla 4. 2 Definición de las coberturas del paisaje ................................................................. 91
Tabla 4. 3 Descripción de las métricas de paisaje a nivel de clase ............................................ 93
Tabla 4. 4 Comparación de métricas de paisaje en los predios Mejorana y Tierra Viva ............... 98

7
Resumen
La palma de aceite es uno de los sistemas agrícolas con mayor área de siembra en
Colombia (466.185 ha) cuyo establecimiento implica remoción y/o transformación de la
vegetación y uso del suelo, desconociéndose en gran medida sus efectos sobre la
biodiversidad. Este trabajo se llevó a cabo con el objetivo de conocer el efecto del cultivo
de la palma de aceite y la altura de estas sobre la diversidad de aves en dos fincas con
paisajes dominados por plantaciones comerciales en el Departamento del Meta
(Colombia). Para ello se realizaron medidas de diversidad ecológica de las aves a partir de
censos visuales y de captura, así como análisis de uso de recursos, gremios de
alimentación y atributos del paisaje. A partir de las técnicas de observación encontramos
44 especies de aves asociadas al cultivo de palma de aceite. La altura de la palma no tuvo
una incidencia significativa en la diversidad de aves ni en el uso de recursos, siendo el
descanso en percha la actividad más recurrente dentro de la plantación. Usando las
técnicas de captura encontramos que el bosque tuvo mayor asociación de especies (88 vs
33 en el cultivo), y mayor representatividad de aves de cada gremio de alimentación. En el
análisis a escala de paisaje identificamos que las aves respondieron de manera diferencial
ante los atributos del paisaje en tanto que algunas especies se pueden beneficiar de la
cantidad de parches y de borde por ser cazadoras aéreas (ej. Tyrannus, Myozetetes,
Myiodinastes), mientras que aves insectívoras de interior de bosque (Myrmeciza) podrían
ser más sensibles a la fragmentación cuando se reduce el área del parche y aumenta la
cantidad de borde. También encontramos que la comunidad tuvo mayor afinidad por los
matorrales de ambas fincas, pero la abundancia de las especies puede variar con el área y
el tipo de cobertura. En conclusión, los cultivos comerciales de palma de aceite mostraron
bajos valores de diversidad de aves independientemente de la altura de las palmas. La
escasa complejidad estructural del cultivo limitó la diversidad de gremios de alimentación
y uso de recursos, afectando con ello el papel funcional de las aves. Esta situación
trasciende a otras áreas del país afectando aves residentes y migratorias para las cuales este
tipo de monocultivo, representa un entorno con recursos limitados a lo largo de su ciclo de
vida, del cual algunas especies se pueden beneficiar, pero otras de mayor valor de
conservación podrían estar reduciendo sus poblaciones.

8
Palabras clave: comunidad de aves, gremios de alimentación, palma de aceite, Orinoquía
colombiana, estructura de la vegetación
Abstract
Palm oil crops it´s one of agricultural system more extensive in Colombia (466.185 ha), in
which imply to remove and/or change the vegetation and land use, unknowing about the
effects on the biodiversity. Our study had the objetive of to know the effect of the palm oil
crop and her size, on birds diversity in two farms with landscape dominated by comercial
plantations, in Meta (Colombia). We measured ecological diversity from both, visual and
capture techniques, resource use, feeding guilds and landscape characteristics. From visual
techniques we recorded 44 bird species into palm oil crop. Size of the palms had no
significant incidence in the bird diversity, neither resource use, and perch was the most
frequently activity into plantation. Using capture techniques we found the forest had most
bird species number (88 vs. 33 in the crop), and representativeness in each feeding guild.
Analysis at landscape scale the birds showed a differential response to landscape
attributes, for example, some species could be benefit the patch and edge amount because
they are hunting-air (ej. Tyrannus, Myozetetes, Myiodinastes), while the insectivorous
birds of core forest (ej. Myrmeciza) could show more sensitivity to fragmentation when to
reduce the patch area and increase edge amount. Bird community showed relationship to
scrub in both farm, but abundance could change due type of cover and area. We conclude
that commercial palm oil crops showed low values of bird diversity regardless size of
palms. Low structural complexity of the crop limited the feeding guild diversity and
resource use, affecting the functional role of the birds, too. Current situation transcend in
another areas of country affecting both, resident and migratory birds that visit a
monoculture with lacking resource, along your life cycle in which some species could
benefit, but that ones of importance of conservation could be reducing their populations.
Key words: bird community, feeding guilds, palm oil, Orinoquia colombiana, structure of
vegetation

9
PRESENTACIÓN
A los lectores:
Aprovecho este espacio para introducirlo de manera resumida a los capítulos que se
describen en esta investigación, los cuales tienen varios enfoques relacionados a la
ecología de las comunidades de aves: en el primer objetivo de investigación hice un
análisis de la diversidad y estructura trófica de aves asociadas al cultivos de palma,
seguidamente me refiero a un análisis sobre uso de hábitat de las aves dentro de la
plantación, con relación al tercer objetivo se hizo una comparación de la estructura de
gremios de alimentación entre el cultivo y las formaciones vegetales originales, y por
último estudié la respuesta de la comunidad ante la configuración del paisaje analizado a
partir de los gremios de alimentación.
Aunque mucho se ha documentado sobre los diversos factores que afectan la
biodiversidad, las acciones de mitigación, contingencia y reparación son escasas y lentas.
Como factores que afectan la biodiversidad me refiero a proyectos agrícolas, mineros,
industriales y toda aquella actividad humana que causa una perturbación a gran escala
sobre los ecosistemas. Las aves de Colombia son un componente sumamente importante y
valioso de nuestra biodiversidad, el país es reconocido por tener la mayor riqueza de
especies del mundo, sin embargo, este patrimonio heredado de la evolución y de los
procesos de espacialización en el uso y explotación de recursos específicos, en un país
tropical, puede estar amenazado por la desarticulación de los proyectos de desarrollo con
acciones que garanticen el sostén de la biodiversidad a largo plazo.
Los resultados de esta investigación nos ayudan a entender cómo son los procesos
funcionales de las aves dentro del monocultivo de palma de aceite, en qué medida las
especies se benefician o perjudican y nos permite identificar qué aspecto del entorno se
deben mantener o modificar para favorecer la biodiversidad y el sostenimiento de las
comunidades de aves a largo plazo en paisajes donde predominan los cultivos de palma de
aceite.

10
INTRODUCCIÓN GENERAL
Ecología de las comunidades de aves
Las comunidades ecológicas son estructuras que se derivan de eventos del pasado (Drake,
1991), se definen como entidades complejas y dinámicas, compuestas de poblaciones que
interactúan en diversas direcciones: competencia, depredación, herbivoría, dispersión,
polinización, parasitismo, entre otras, causando cambios continuos en el flujo de energía
en espacio y tiempo (Carnicer et al., 2009). Esta definición abarca todos los niveles
tróficos e incluye especies y procesos que operan en distintas escalas y niveles (Allen &
Starr 1982; ONeill et al., 1986).
La ecología de comunidades aplicada al estudio de las aves tuvo mayor fuerza a partir de
los años 40 y 50, cuando las publicaciones de David Lack (Lack & Southern 1949) y
Robert MacArthur (1958) enfatizaron en la importancia de la competencia interespecífica
en la estructura de las comunidades a través de sus efectos sobre el desarrollo evolutivo de
las diferencias morfológicas y ecológicas entre especies que co-existen (Wiens 1989). Los
estudios de Lack y MacArthur incorporaron formulaciones matemáticas basadas en
ecología de poblaciones y tuvieron un efecto profundo en la ecología de comunidades
porque fue el momento en el que la ecología pasa de ser una ciencia descriptiva a ser una
ciencia predictiva acompañada de teorías cuantitativas que reconocen ampliamente los
patrones de la naturaleza (Wiens, 1989).
La ecología de las comunidades de aves entonces tiene como principal objetivo conocer y
determinar el papel que cumplen las aves en la estructura y función de las comunidades
ecológicas (MacMahon et al., 1981; Morrison et al., 1988) y cómo la distribución y
abundancia de los recursos influyen en la dinámica de las poblaciones e interacciones entre
especies (Morrison et al., 1988; Wiens, 1984). Tales interacciones son procesos básicos

11
elementales que pueden influir en la abundancia de las especies y la composición de una
comunidad (Morin, 2011).
La relación entre las aves y la vegetación ha sido un punto de partida para comprender las
interacciones en la distribución de las especies (Jankowski et al., 2013), y cómo estas
relaciones inciden sobre la diversidad y estructura de una comunidad, la oferta y
disponibilidad de recursos, el establecimiento de poblaciones y segregación de gremios
(Cody, 1981; Cousin & Phillips, 2008). Las aves ocupan hábitats que pueden ser
estructuralmente complejos u homogéneos y la selección dependerá de las adaptaciones en
términos de explotación de recursos, la competencia interespecífica y como responden de
manera específica a la estructura del hábitat (Alatalo, 1981; Cousin & Phillips, 2008).
Avifauna de la Región Oriental de Colombia
Los Llanos Orientales de Colombia, también conocidos como Orinoquía colombiana,
cuenta con una extensión de 158.855 km2, presenta diversos paisajes como el piedemonte,
las terrazas, llanura aluvial de desborde, llanura eólica, la altillanura y aluviones, también
se destaca la dominancia de sabanas tropicales con una matriz herbácea (Mora et al.,
2011).
Esta es una de las regiones con mayor diversidad de aves del país, en la cual se han
registrado 761 especies que incluye 14 amenazadas, 6 casi endémicas, 15 con rango
restringido, 150 están incluidas en el listado CITES, 60 congregatorias y 1 introducida
(Acevedo-Charry et al., 2014; Murillo-Pacheco 2005), las cuales se distribuyen entre
sabanas inundables con lámina de agua, sabanas con estrato herbáceo, bosques de galería,
esta última es la formación reconocida por albergar la mayor cantidad de especies (Lau-
Pérez, 2008).
Los ecosistemas terrestres de la Orinoquía varían en composición, estructura y
complejidad de la vegetación (Blydestein, 1967; Rangel-Ch. & Minorta-Cely, 2014), y así

12
mismo la distribución y asociación de la avifauna (McNish, 2007). Para la región
Orinoquense se han decrito diversas formaciones de vegetación relacionadas al
piedemonte, terrazas aluviales y altillanura, así como 10 tipos de ecosistemas de sabana
(Blydestein, 1967; Rangel-Ch & Minorta-Cely, 2014;), a partir de los cuales se puede
predecir la presencia de ciertas especies. Por ejemplo, especies como Athene cunicularia,
Burhinus bistriatus, Anthus lutescens son comunes en áreas abiertas cubiertas de
pastizales. En esteros y pastos inundables frecuentan Eudocimus ruber, Theristicus
caudatus, Jabiru mycteria, Ciconia maguari, Mycteria americana, Ardea cocoi. Entre los
morichales se observa la guacamaya Orthopsittaca manilata, Amazona amazonica,
Amazona ochrocephala. Cuando la vegetación tiene mayor grado de obstrucción como en
las matas de monte o en los bosques de galería se pueden encontrar especies como
Momotus subrufecens, Galbula gálbula, Ramphastos tucanus, Myrmeciza longipes,
Myrmeciza athrotorax, Manacus manacus, entre otros ejemplares (McNish, 2007).
Los cambios en el uso del suelo en las áreas de bosque y sabanas a monocultivos de palma
ya han generado extinciones locales y/o sustitución de especies (Olivares, 1982; Acevedo-
Charry et. al., 2014) Varias especies que se alimentan en sabanas, nidifican en matorrales
riparios vecinos a estas, por los que las alteraciones drásticas en las sabanas con fines
agropecuarios pueden afectar el éxito reproductivo de las especies (Marín et al., 2007,
Ramo & Busto 1984). En estos ecosistemas se han observado procesos de dispersión en
función a la estacionalidad (Lau-Pérez 2008), también se ha destacado la importancia de
conservar corredores amplios y bosques de galería para garantizar la permanencia de
especies que viven en el bosque, pero se alimentan en los pastizales (Marín et al., 2007,
Tubelis 2002).
Cultivos de Palma de Aceite
La palma de aceite o palma africana (Elaeis guineensis) es considerada la especie más
productiva y de mayor rendimiento en toneladas métricas de aceite por hectárea en
comparación con otros cultivos oleaginosos como la soya, girasol, maní, algodón y
ajonjolí (Corley & Tinker, 2003; Raygada-Zambrano, 2005).

13
Es una especie nativa del occidente de África, pero actualmente la palma de aceite se
encuentra ampliamente distribuida en países de Asia, África, Centro y Sur América
(Corley & Tinker, 2003; Raygada-Zambrano, 2005). La palma crece hasta los 500
m.s.n.m., con pendientes menores a 15º, bajo una temperatura ambiental que varía de 21 a
33oC, el brillo solar anual debe ser superior a 1500 horas, con un rango de precipitación
promedio anual de 1800 a 2200 mm y a una humedad relativa del 80% (Mignorance et al.,
2004). El diseño del cultivo se hace a tresbolillo (formando un triángulo equilátero con
lados de 9 m) para obtener 143 plantas por ha (Raygada-Zambrano, 2005), las plantas se
llevan al campo de siembra definitiva después de los 24 meses de su germinación (USDA,
2007).
El manejo del cultivo de palma de aceite es una labor permanente que consiste en labores
culturales y de fertilización que consiste en la limpieza y remoción de herbáceas y arbustos
que le resta complejidad estructura al sistema (Raygada-Zambrano, 2005), ambas buscan
garantizar la sanidad y optima producción de la plantación (IICA, 2006). Las labores
culturales son las actividades agronómicas para reducir las poblaciones de plagas por
debajo de los niveles que puedan causar pérdidas económicas (Lizarazo & Alfonso, 2011)
mediante el control de malezas con métodos mecánicos y químicos, poda sanitaria y
coyoleo (recolección de frutos caídos para evitar el nacimiento de plántulas),
mantenimiento de círculos e interlineas, poda sanitaria, polinización asistida y entomófila,
cosecha y transporte de racimos, poda pos cosecha, control de plagas y enfermedades,
mantenimiento de drenaje y mantenimiento de vías de comunicación (IICA, 2006;
Raygada-Zambrano, 2005). En los primeros cuatro años en campo definitivo se hace
control de maleza dos o tres veces al año mediante control químico o mecánico (Raygada-
Zambrano, 2005).
Las labores de fertilización están orientadas a suministrar nutrientes para promover el
crecimiento vegetativo, la resistencia a plagas y enfermedades y remplazar los nutrientes
exportados por los racimos en la cosecha (IICA, 2006; Raygada-Zambrano, 2005), la

14
dosificación se programa según la edad, los requerimientos del cultivo, la fertilidad del
suelo, los resultados del análisis químico foliar y los niveles de rendimiento (IICA, 2006).
La producción inicia entre los 24 y 36 meses después de la siembra definitiva en campo
(IICA, 2006; Raygada-Zambrano, 2005; USDA, 2007). El producto de la plantación se usa
con fines comestibles y no comestibles. Como producto comestible se utiliza como aceite
de cocina, se emplea para elaborar productos de panadería, pastelería, confitería, sopas
instantáneas, salsas, productos congelados y deshidratados. Dentro de los usos no
comestibles es útil para la elaboración de detergente líquido, jabón, champú, lápiz labial,
ceras, grasas lubricantes, también lo emplean para reducir la fricción durante la fabricación
del acero y es la materia prima del biodiesel (Danielsen et al., 2008; Clay, 2004).
En América Latina estas plantaciones se han establecido en México, Costa Rica,
Nicaragua, Ecuador, Perú, Brasil, Venezuela, Panamá y Colombia, en este último el área
sembrada y la producción se ha incrementado en los últimos años en cuatro zonas del
territorio nacional (norte, oriente, centro y occidente). Los reportes indican que el área de
siembra pasó de 306.961 ha en el 2007 a 427.368 ha en el año 2011, así mismo la
producción pasó de 733. 000 a 941.000 ton en el mismo periodo de tiempo (Fedepalma,
2012). Parte de la producción del aceite de palma que se origina en Colombia se distribuye
al Reino Unido, Alemania, España, Holanda, Venezuela, República Dominicana, entre
otros, en el año 2007 se exportaron 341.875 ton representando un crecimiento promedio
del 43% con respecto a los años anteriores (USAID, 2009).
El desarrollo del sector palmero ha sido rápido y continuo en las diferentes zonas de
producción del territorio nacional, especialmente por los incentivos que el gobierno viene
otorgando desde el año 2000 entre los que se destacan la exención del impuesto de renta
para nuevos cultivos, el certificado de reembolso tributario (CERT) el cual promueve las
exportaciones mediante la devolución total o parcial de impuestos y tasas de exportación,
el Incentivo de Capitalización Rural (ICR), de igual forma también se han otorgado
exenciones tributarias para la producción de biocombustible en el sector palmicultor

15
(Álvarez-Aristizabal 2009, Ministerio del Ambiente, Vivienda y Desarrollo Territorial
2010). Por su parte las autoridades ambientales como el Ministerio de Ambiente, Vivienda
y Desarrollo Territorial y Corporaciones Autónomas Regionales no exigen licencias
ambientales a estos proyectos de producción agrícola aun cuando es claro el impacto
ambiental que se presenta a nivel biótico, abiótico y paisajístico (Seeboldt y Salinas
Abdala, 2010) y muchas veces con una escasa regulación de las medidas de mitigación y
prevención establecidas en el Decreto 1220 del 21 de abril de 2005. No obstante,
Fedepalma como órgano representante del gremio palmicultor en Colombia, se ha
vinculado a la iniciativa internacional de la RSPO (Roundtable on Sustainable Palm Oil)
con el fin de aumentar la competitividad y producción de forma económica,
ambientalmente viable y socialmente sostenible, que bajo el cumplimiento de un conjunto
de principios y criterios busca certificar a las empresas productoras que cumplan con leyes
y regulaciones locales, nacionales e internacionales, que implementen el uso de prácticas
apropiadas en los procesos operativos para plantaciones y plantas extractoras que tengan
responsabilidad con el medio ambiente y conservación de los recursos naturales y la
biodiversidad, entre otros compromisos (RSPO, 2007).
Problema
La mayor biodiversidad de los ecosistemas terrestres está localizada en los trópicos, donde
hay óptimas condiciones para el cultivo de la palma de aceite (Elaeis guineensis) y por
consiguiente donde está la mayor cobertura de esta plantación que representa una amenaza
para las comunidades de aves y otros representantes de la biota (BirdLife International,
2008; Persey & Anhar 2010). Los bajos costos, la alta producción y las óptimas
condiciones climáticas y edáficas en zonas tropicales ha generado un aumento en las áreas
de cultivo debido al alto rendimiento de la cosecha durante todo el año (Casson, 2000).
¿Cómo han impactado los cultivos de palma de aceite en la biodiversidad?
El cultivo de palma de aceite es uno de los principales causantes de la pérdida de la
biodiversidad, debido a las extensas áreas cultivadas que se han establecido en las regiones

16
de mayor biodiversidad del planeta (FAO, 2011; Turner et al., 2011). El aumento en la
demanda del aceite de palma para productos alimenticios y no consumibles conduce a
cambios ambientales, que a pesar de su importancia han sido poco estudiados y que aún es
necesario entender cómo los paisajes modificados por el hombre pueden ser manejados
para permitir una producción sostenible de este cultivo, que amerita una importancia
global, así como el mantenimiento de la biodiversidad (Turner et al., 2011).
Las investigaciones a cerca de los impactos de la conversión de bosques a cultivos de
palma de palma sobre la biodiversidad son muy escasos (Foster et al., 2011; Turner et al.,
2008, Turner et al., 2011), a pesar de esto existe evidencia contundente que la conversión
de habitats naturales a cultivos de palma ha generado impactos negativos sobre la
biodiversidad (Fitzherbert et al., 2008; Foster et al., 2011, Turner et al., 2011). Estudios
realizados con diferentes grupos taxonómicos: insectos (hormigas, escarabajos, abejas,
mariposas), mamíferos (primates, musarañas, ardillas murciélagos), aves y lagartos, han
mostrado una reducción en la riqueza de especies cuando hubo transformación de otros
ecosistemas a cultivos de palma, los estudios destacaron un alto nivel de la pérdida de la
biodiversidad como resultado de la expansión global de este monocultivo (Turner et al.,
2011). Inversamente también han encontrado que algunas especes de murciélagos
(Danielsen & Heegaard, 1995), coleópteros coprófagos (Davis & Philips, 2005), y
lagartijas (Glor et al., 2001) incrementaron su abundancia como resultado de la conversión
del hábitat y por ser poblaciones tolerantes a las perturbacioes (Turner et al., 2011).
El establecimiento de este monocultivo también reduce la complejidad estructural de la
vegetación (Nájera & Simonetti, 2010). La estructura de la plantación varía de acuerdo al
crecimiento de la palma principalmente en cuanto a la altura, cobertura de dosel,
crecimiento de plantas epífitas sobre troncos y crecimiento de plantas en el suelo (Cagod
& Nuñeza, 2012; Luskin & Potts, 2011). Aunque la asociación de aves con respecto a los
cambios en la altura de las palmas y la complejidad estructural de los cultivos de palma es
un aspecto poco estudiado, se considera un factor determinante en los procesos ecológicos
que sustenta la diversidad funcional de las aves en cultivos de palma de aceite así como en

17
la composición, riqueza y abundancia de aves que frecuentan este tipo de cobertura (Azhar
et al., 2013; Cagod & Nuñeza, 2012; Nájera & Simonetti, 2012b).
Se ha encontrado que los cultivos de palma de aceite pueden interrumpir, reducir o
eliminar el desplazamiento de individuos debido a que la fragmentación y el aislamiento
entre los parches disminuye la conectividad del paisaje, en este contexto la persistencia de
las especies depende de cómo estas responden a la nueva configuración, particularmente a
la capacidad de moverse a través de los elementos de la matriz (Awade & Metzger, 2008).
Los cultivos de palma de aceite en Colombia
Colombia se ubica como el primer país productor de palma de aceite en América Latina y
el cuarto a nivel mundial después de Tailandia Indonesia y Filipinas (Clay, 2004; Corley &
Tinker, 2003). Los cultivos de palma del país se concentran en el piedemonte llanero,
Cesar, Magdalena, Sucre, Córdoba, Bolívar y la Costa Pacífica. La superficie cultivada ha
crecido a una tasa anual del 10,5% desde 1999 hasta el año 2006 con un área de siembra
de 326.033 ha en el territorio nacional (Cenipalma-Fedepalma, 2009; Clay, 2004). La
actividad agrícola en la llanura oriental del país ha transformado y reducido bosques y
sabanas inundables del piedemonte andino generando cambios en la composición de aves
y pérdida de la estructura vegetal nativa (Blydenstein, 1967; McKay, 1980; Romero Ruíz,
2011). Estudios han revelado que las extensas áreas de esta plantación pueden ser una
matriz inhóspita para las aves, si se considera que en sistemas mayores de 15 años se ha
reportado una reducción del 80% de las especies de bosque (Foster et al., 2011, Peh et al.,
2006) ya que las condiciones microclimáticas, los recursos alimenticios y las plantas
epífitas que usan algunas aves para anidar, varían de acuerdo a la edad del cultivo (Luskin
& Potts, 2011, Foster et al., 2011, Hassan et al., 2011).
El sector palmero de Colombia se ha fortalecido en los últimos 60 años expandiendo el
cultivo en más de 124 municipios con un área de siembra superior a 450.132 hectáreas
(Fedepalma, 2015; Leech, 2009). Se estima que los pequeños productores suman el 25%
del cultivo de palma a nivel nacional (Leech, 2009). Aunado a esto, el gobierno nacional, a

18
través del Ministerio de Agricultura y Desarrollo Rural, viene otorgando el Incentivo de
Capitalización Rural (ICR), como un beneficio económico que le otorgan a pequeños,
medianos y grandes palmicultores (entre otros gremios del sector agrícola) para
incrementar el área de siembra y mejorar las condiciones de productividad (IAvH, 2000),
aunque esto implique la destrucción de ecosistemas naturales.
Aves en Sistemas Agrícolas: Diversidad e implicaciones del paisaje
Existen plantaciones que pueden favorecer la asociación de especies, como por ejemplo
los cultivos de cacao, café con sombra y de arroz que se consideran amigables porque
funcionan como hábitat y brindan recursos de alimento, sitios de reproducción y refugio
para la subsistencia y permanencia de muchas especies, inclusive en aves migratorias
(Díaz-Bohórquez et al., 2014; Wunderle & Latta, 1998). La cercanía con la vegetación
nativa también es un factor que puede favorecer la asociación de especies dentro de un
cultivo (De Chenon & Susanto, 2006), sin embargo la carencia de estas características en
los sistemas agrícolas productivos a gran escala propicia un entorno inhóspito para muchas
especies de aves, resultando en la reducción de la riqueza de especies, cambios en la
estructura trófica, se hospedan pocas especies de importancia de conservación y algunas
especies también se ven afectadas por el efecto de borde (Aratrakorn et al., 2006;
Fitzherbert et al., 2008), como por ejemplo los casos de depredación de nidos y la
permeabilidad o el paso de individuos entre ecosistemas (Andrén, 1994).
Los patrones en el uso de la tierra pueden alterar la dirección de los procesos naturales en
el entorno donde los organismos deben vivir, reproducirse y dispersarse (Turner et al.,
2001). La ampliación de la frontera agrícola modifica los ecosistemas y crea paisajes
fragmentados, la vegetación remanente queda como parches pequeños aislados en una
matriz agrícola (Fuentes-Montemayor et al., 2013, Saunders et al., 1991), que varían en el
grado de conectividad y albergan poca diversidad con especies de bajo valor de
conservación (Collard et al., 2009; McIntyre & Hobbs, 1999; Saunders et al., 1991), la
estructura de la vegetación y la configuración espacial se ve alterada, afectando
comunidades biológicas, por ejemplo, creando barreras de dispersión y efecto de borde

19
(Fuentes-Montemayor et al., 2013). En las aves, los procesos de fragmentación también
generan cambios en la diversidad y composición de la comunidad, aunque los parches con
relictos de vegetación remanente pueden proporcionar un hábitat importante para las aves
nativas (Collard et al., 2009).

20
OBJETIVO GENERAL
Conocer el efecto del cultivo de palma de aceite sobre la diversidad de aves, y evaluar la
influencia de los tipos de vegetación sobre los atributos de la comunidad.
Preguntas de Investigación
Para identificar y comprender si la comunidad de aves podría estar afectada de forma
directa o indirecta, por factores relacionados a la estructura vertical del cultivo de palma de
aceite, se consideró pertinente contestar las siguientes preguntas:
1. ¿Cuál es el efecto de la altura de la palma sobre diversidad de aves?
2. ¿Qué recursos utilizan las aves dentro del cultivo de palma de acite y cuáles son los
gremios de alimentación más favorecidos?
3. ¿Cuáles son las principales diferencias de la composición y estructura de los gremios de
alimentación de aves, entre el cultivo de palma de aceite y el bosque?
4. ¿Cómo inciden los tipos de vegetación y los atributos del paisaje sobre en la diversidad
de aves?

21
Métodos Generales
Área de Estudio
La presente investigación se hizo en las fincas con cultivos de palma de aceite Tierra Viva
y La Mejorana, ubicadas en las veredas Barbascal y Montelibano delos municipios San
Martín de los Llanos y Acacias, respectivamente, departamento del Meta.
Tierra Viva (TIV) tiene un área total de 1400 ha, se encuentran a los 03°37'51.8" N,
073°20'59.7" W y a 420 m.s.n.m, por su parte, la finca Mejorana (MEJ) tiene un área total
de 1480 ha, se encuentra ubicada a los 03°57'37.18"N, 73°37'32.93W y a 498 m.s.n.m.
(Figura 1.1). Cada plantación tiene lotes con distintas edades, que varían desde plantas
recién sembradas hasta aquellas que tienen cerca de los 22 años.
Figura 1 Ubicación de las fincas Tierra Viva (TIV) y La Mejorana (MEJ) en San Martín de
los Llanos y Acacias, Meta

22
San Martín de los Llanos se encuentra en el centro occidente del Meta, según la Alcaldía
de San Martín de los Llanos (2009) la región se caracteriza por presentar cuatro zonas
definidas en cuanto al uso del suelo. La zona aluvial tiene suelos fértiles y en esta se
concentran la mayoría de los cultivos comerciales así como áreas de ganadería, la zona de
terrazas tiene suelos con pendientes leves (menores del 3%) destinadas principalmente a la
ganadería, la zona de piedemonte tiene pendientes más pronunciadas (de 3 a mayores de
50%) donde predominan cultivos de cítricos, y la zona de altillanura, ubicada en el oriente
del municipio con pendientes del 3 al 5%, presenta erosión del suelo por quemas y el
pastoreo del ganado.
El paisaje de San Martín de los Llanos incluye abanicos aluviales, llanuras de inundación,
llanura aluvial de desborde y la altillanura, estas formaciones tienen una dinámica
relacionada con el clima, la agricultura, la ganadería y los sistemas fluviales. La
temperatura varía entre los 25 y 28oC, el régimen de lluvias es monomodal con
precipitaciones de marzo a noviembre y un periodo seco de diciembre a febrero, siendo
mayo y diciembre los picos máximos de cada periodo (Alcaldía de San Martín de los
Llanos 2009). El componente vegetal está representado por bosques de galería, bosque
húmedo tropical, sabanas con gramíneas naturales e introducidas y grandes áreas de
explotación agrícola comercial y de subsistencia (Alcaldía de San Martín de los Llanos,
2009).
El municipio de Acacias se encuentra en el noroccidente del departamento del Meta, los
valores promedios de temperatura y precipitación son de 24oC y 5000 mm,
respectivamente. El sector rural del municipio presenta piedemonte andino, reservas
forestales, pastoreo del ganado, cultivos permanentes de palma de aceite, y agricultura
intensiva con cultivos de arroz, maíz, soya, algodón y frutales. Los registros de fauna
incluyen especies de aves acuáticas, cerca de 20 especies de mamíferos, no obstante este
componente ha estado presionado por la caza y comercialización ilegal de aves, mamíferos
y reptiles, principalmente (Cormacarena, 2006).

23
Denominaciones: Bosque y Tipos de Coberturas
En el capítulo 4 se compara la estructura de los gremios de alimentación entre bosque y
cultivo de palma. El término Bosque se usó para unificar la comparación entre el sistema
agrícola y las formaciones de vegetación natural. No obstante, en el capítulo 5 se analizan
3 tipos de coberturas (bosque de galería, fragmento de bosque y matorral) que hicieron
parte de la comparación de los atributos del paisaje con la diversidad de aves, y dichas
coberturas corresponden a lo denominado como bosque en el capítulo 4.

24
Referencias
Acevedo-Charry, O., A. Pinto-Gómez & J. O Rangel-Ch. 2014. Las aves de la Orinoquía
colombiana: una revisión de sus registros. En Rangel-Ch, J. O. (Ed.) 2014.
Colombia Diversidad Biótica XIV. La región de la Orinoquía de Colombia.
Universidad Nacional de Colombia – Instituto de Ciencias Naturales. 895pp.
Bogotá, D.C.
Alatalo, R. V. 1981. Habitat selection of forest birds in the seasonal environment of
Finland. Annales Zoologici Fennici, 18: 103-114.
Alcaldía de San Martín de los Llanos. 2009. Agenda Ambiental San Martín de los Llanos,
Meta. Ministerio de Ambiente, Vivienda y Desarrollo Territorial- Cormacarena.
Allen, T. F. H. & T. B. Starr 1982. Hierarchy: Perspectives for Ecological Complexity.
Chicago: University of Chicago Press, 1982, 310 pp.
Álvarez Aristizábal, A. 2009. Efectos del monocultivo de palma de aceite en los medios de
vida de las comunidades campesinas. El caso de Simití – Sur de Bolívar. Tesis de
Maestría en Desarrollo Rural. Pontificia Universidad Javeriana. Bogotá, Col.
Andrén, H. 1994. Effects of hábitat fragmentation on birds and mammals with different
proportion of situable hábitat: a review. Oikos, 71: 355-366.
Aratrakorn, S., Thunhikorn, S., & Donald, P. F. (2006). Changes in bird communities
following conversion of lowland forest to oil palm and rubber plantations in
southern Thailand. Bird Conservation International, 16, 71-82. Awade, M. y J. P.
Metzger. 2008. Using gap-crossing capacity to evaluate functional connectivity
of two Atlantic rainforest birds and their response to fragmentation. Austral
Ecology, 33: 863–871.
Azhar, B., D. B. Lindenmayer, J. Wood, J. Fischer, A. Manning, C. McElhinny & M.
Zakaria. 2013. The influence of agricultural system, stand structural complexity
and landscape context on foraging birds in oil palm landscapes. Ibis, 155: 297-
312.
BirdLife International, 2008. Many forest birds cannot survive in oil palm and rubber
plantations. Presented as part of the BirdLife State of the world's birds website.
Available from: http://www.birdlife.org/datazone/sowb/casestudy/138. Checked:
04/03/2011
Blydenstein, J. 1967. Tropical savanna vegetation of the Llanos de Colombia. Ecology,
48(1): 1–15.

25
Cagod, B. M. & O. M. Nuñeza. 2012. Avian species diversity in oil palm plantations of
Agusan Del Sur and Compostela Valley, Philippines. AES. Bioflux, 4(2): 85–
105.
Carnicer, J., P. Jordano & C. Melián. 2009. The temporal dynamics of resource use by
frugivorous birds: a network approach. Ecology, 90(7): 1958-1970.
Casson, A. 2000. The Hesitant Boom: Indonesia’s Oil Palm Sub-Sector in an Era of
Economic Crisis and Political Change. CIFOR (Center of International Forest
Research) Occasional Paper No. 29. 50 pp.
Cenipalma-Fedepalma. 2009. Agenda prospectiva de investigación y desarrollo 38
tecnológico para la cadena productiva de palma de aceite en Colombia con
énfasis en oleína roja. Bogotá, Colombia.
Clay, J. 2004. Agriculture and the environment. A commodity by commodity guide to
impacts and practices. Island Press. Washingtong D.C. 570 pp.
Cody, M. L. 1981. Habitat selection in birds: The roles of vegetation structure,
competidors and productivity. BioScience, 31(2): 107-113.
Collard, S., A. Le Brocque & Ch. Zammit- 2009. Bird assemblages in fragmented
agricultural landscapes: the role of small brigalow remnants and adjoining land
uses. Biodiversity Conservatio, 18: 1649-1670.
Corley, R. H. & P. B. Tinker. 2003. The palm oil. 4th Edition. Oxford Blackwell Science.
562pp.
Cormacarena. 2006. Agenda Ambiental. Municipio de Acacias, Departamento del Meta.
Disponible en: http://www.acacias-meta.gov.co
Cousin, J. & R. D. Phillips. 2008. Habitat complexity explains species-specific occupancy
but not species richness in a Western Australian Woodland. Australian Journal
Zoology, 56: 95-102.
Danielsen, F. & M. Heegaard. 1995. Impact of logging and plantation development on
species diversity: a case study from Sumatra, In Management of tropical forests:
towards an integrated perspective, O. Sandbukt, (Ed.), Centre for Development
and the Environment, Oslo, Norway.
Danielsen, F., H. Beukema, N. D. Burgess, F. Parish, C. A. Brühl, P. F. Donald, D.
Murdiyarso, B. Phalan, L. Reijnders, M. Struebig & E. B. Fitzherbert. 2008.
Biofuel Plantations on Forested Lands: Double Jeopardy for Biodiversity and
Climate. Conservation Biology. 23(2): 348-358.
Davis, A. L. V. & T. K. Philips. 2005. Effect of Deforestation on a Southwest Ghana Dung
Beetle Assemblage (Coleoptera: Scarabaeidae) at the Periphery of Ankasa
Conservation Area. Environmental Entomology, 34(5): 1081-1088.

26
De Chenon, R. & A. Susanto. 2006. Ecological observations on diurnal birds in Indonesian
oil palm plantations. Journal of Oil Palm Research (Special Issue April): 122–
143.
Díaz-Bohórquez, A. M., N. J. Bayly, J. E. Botero & C. Gómez. (2014). Aves migratorias
en agroecosistemas del norte de Latinoamérica, con énfasis en Colombia.
Ornitología Colombiana, 14, 3-27.
Drake, J. A. 1991. Community-Assembly mechanics and the structure of an experimental
species ensemble. The American Naturalist, 137(1): 1-26.
FAO. 2011. In: FAOSTAT Online Statistical Service. Food and Agriculture Organization
of the United Nations, Accessed March 2011, Available from:
http://faostat.fao.org
Fedepalma. 2012. Anuario Estadístico 2007 - 2011. La agroindustria de la palma de aceite
en Colombia y el mundo. Fedepalma, Bogotá Colombia
Fedepalma. 2015. Anuario Estadístico 2010 - 2014. La agroindustria de la palma de aceite
en Colombia y el mundo. Fedepalma, Bogotá Colombia
Fitzherbert, E. B., M. J. Struebig, A. Morel, F. Danielsen, C. A. Brühl, P. F. Donald & B.
Phalan. 2008. How will oil palm expansion affect biodiversity?. Trends in
Ecology and Evolution. 23(10): 538-545.
Foster, W. A., J. L. Snaddon, E. C. Turner, T. M. Fayle, T. D. Cockerill, M. D. Farnon
Ellwood, G. R. Broad, A. Y. C. Chung, P. Eggleton, C. V. Khen & K. M. Yusah.
2011. Establishing the evidence base for maintaining biodiversity and ecosystem
function in the oil palm landscapes of South East Asia. Phil. Trans. R. Soc. B.,
366: 3277–3291.
Fuentes-Montemayor, E., D. Goulson, L. Cavin, J. M. Wallace & K. J. Park. 2013.
Fragmented woodlands in agricultural landscapes: The influence of woodland
character and landscape context on bats and their insect prey. Agriculture,
Ecosystems and Environment, 172: 6-15.
Glor, R. E., A. S. Flecker, M. F. Benard & A. J. Power. 2001. Lizard diversity and
agricultural disturbance in a Caribbean forest landscape. Biodiversity and
Conservation, 10(5): 711-723.
IICA (Instituto Interamericano de Cooperación para la Agricultura). 2006. Cultivo de la
palma africana Guía Técnica. Managua, Nicaragua.
Jankowski, J. E., Merkord, C. L., Rios, W. F., Cabrera, K. G., Revilla, N. S. & Silman, M.
R. 2013. The relationship of tropical bird communities to tree species
composition and vegetation structure along an Andean elevational gradient. J.
Biogeogr., 40: 950–962.

27
Lack, D & H. N. Southern. 1949. Birds of Tenerife. The Ibis, 607-626.
Lau-Pérez, P. A. 2008. Patrones de utilización de los hábitats por parte de las aves, en una
región de sabanas bien drenadas de los Llanos Orientales en Venezuela.
Ecotrópicos, 21(1): 13–33.
Lizarazo, I. A. & O. A. Alfonzo. 2011. Aplicaciones de la agricultura de precisión en
palma de aceite “Elaeis Guineensis” e hibrido O x G. Revista de Ingeniería
Universidad de los Andes, 33: 124–130.
Luskin, M. S. & M. D. Potts. 2011. Microclimate and habitat heterogeneity through the oil
palm lifecycle. Basic and Applied Ecology, 12: 540–551.
MacArthur, R. H. 1958. Population ecology of some Warblers of Northeastern coniferous
forest. Ecology, 39(4): 599-619.
MacMahon, J. A., D. J. Schimpf, D. C. Andersen, K. G. Smith & R. L. Bayn Jr.1981. An
organism-centered approach to some community and ecosystem concepts.
Journal of Theoretical Biology, 88(2): 287-307.
Marín, G., L. Bastidas, J. Muñoz, O. Oliveros, R. Navarro & B. Marcano. 2007. Perfil
ecológico de la avifauna de los Llanos Orientales de Venezuela en función de los
impactos antrópicos. Interciencia, 32(6): 391–398.
McIntyre S, & Hobbs RJ (1999). A framework for conceptualizing human effects on
landscapes and its relevance to management and research models. Conserv Biol
13:1282–1292.
McKay, W. D. (1980). The influence of agriculture on avian communities near
Villavicencio, Colombia. The Wilson Bull. 92(3): 381-389
Mignorance, F., F. Minelli & H. Le Du. 2004. El cultivo de la palma africana en el Chocó.
Legalidad Ambiental, Territorial y Derechos Humanos. Human Rights
Everywhere.Colombia. 221p.
Mora Fernández C., C. Castellanos Castro, A. Cardona Cardozo, L. Pinzón Pérez & O.
Vargas. 2011. Historia de transformación del paisaje, de la cuenca baja del río
Pauto, Casanare (Colombia). En: Tomás León Sicard (Ed.) 2011. Mamíferos,
Reptiles y Ecosistemas del Bloque Cubiro (Casanare). Educación Ambiental para
la Conservación. Instituto de Estudios Ambientales, Universidad Nacional de
Colombia, Bogotá.
Morin, P. J. 2011. Community Ecology. 2nd Edition. Wiley Blackwell. USA. 407p.
Morrison, M. L., C. J. Ralph, J. Verner & J. R. Jehl (Eds).1988. Avian foraging: Theory,
Methodology and applications. Studies in Avian Biology No. 13. Cooper
Ornithological Society.

28
Murillo-Pacheco, J. 2005. Evaluación de la distribución y estado actual de los registros
ornitológicos de los Llanos Orientales de Colombia. Tesis de pregrado,
Universidad de Nariño. Pasto, Col. 141p.
Nájera A. & J. A. Simonetti. 2010. Can oil palm plantations become bird friendly?.
Agroforestry Systems, 80: 203-209
Olivares, A. 1982. Aves de la Orinoquía colombiana. 2da Edición Instituto de Ciencias
Naturales Uiversidad Nacional de Colombia. Bogotá.
O´Neill, R. V., R. H. Gardner & M. G. Turner. 1986. A hierarchical neutral model for
landscape analysis’. Landscape Ecology,7(1): 55-61.
Peh, K. S.-H., Sodhi, N. S., De Jong, J., Sekercioglu, C. H., Yap, C. A.-M. & Lim, S. L.-H.
.2006. Conservation value of degraded habitats for forest birds in southern
Peninsular Malaysia. Diversity and Distributions, 12: 572–581.
Persey, S. & S. Anhar. 2010. Biodiversity information for Oil Palm. International
Conference on Oil Palm and Environment. Bali, Indonesia.
Ramo C. & B. Busto. 1984. Nidificación de los Passeriformes en los llanos de Apure
(Venezuela). Biotropica 16: 59–68. Ramo C. y B. Busto. 1984. Nidificación de
los Passeriformes en los llanos de Apure (Venezuela). Biotropica 16: 59–68.
Raygada-Zambrano, R. 2005. Manual técnico para el cultivo de la palma aceite. Comisión
Nacional para el Desarrollo y Vida sin Drogas – DEVIDA, Proyecto de
Desarrollo Alternativo Tocache-Uchiza – PRODATU. Perú. 109 pp.
RSPO (Roundtable on Sustainable Palm Oil). 2007. RSPO Principles and Criteria for
Sustainable Palm Oil Production. Including Indicators and Guidance. Disponible
44 en [http://www.rspo.org] 8 May 2012.
Saunders, D. A., Hobbs, R. J. & Margules, C. R.1991. Biological Consequences of
Ecosystem Fragmentation: A Review. Conservation Biology, 5: 18–32
Seeboldt, S. & Y. Salinas Abdala (2010). Responsabilidad y sostenibilidad de la industria
de la palma ¿Son factibles los principios y criterios de la RSPO en Colombia?
INDEPAZ. Disponible en
Tubelis D.2004. Species composition and seasonal occurrence of mixed-species flocksof
forest birds in savannas in central Cerrado, Brazil. Ararajuba, 12: 105–111.
Turner, M. G., R. H. Gardner & R. V. O´Neill. 2001. Landscape ecology in theory and
practice. Patter ans Process. Springer. NY.
Turner E. C., J. L. Snaddon., T. M. Fayle, & W. A. Foster. 2008. Oil Palm Research in
Context: Identifying the Need for Biodiversity Assessment. PlosOne, 3(2): 1-4.

29
Turner, E. C., J. L Snaddon, R. M Ewers, T. M Fayle & W. A Foster. 2011. The Impact of
Oil Palm Expansion on Environmental Change: Putting Conservation Research in
Context. En: Environmental Impact of Biofuels. Marco Aurélio dos Santos
Bernardes (Ed.) InTech. 280p.
USAID (United State Agency International Development), 2009. Situación actual y
perspectivas del mercado del aceite de palma colombiano. Sostenibilidad de los
proyectos de palma apoyados por USAID 81–99.
USDA (United State Department of Agriculture). 2007. Indonesia: Palm Oil Production
Prospects Continue to Grow. Foreing Agriculture Service. Disponible en
[http://www.pecad.fas.usda.gov].
Wiens, J. A. 1984. Resource systems, populations, and communities, p. 397-436. En: W.
Price, C. N Slobodchikoff, y W. S. Gaud (Eds.). A new ecology: novel
approaches to interactive systems. John Wiley and Sons, New York.
Wiens, J. A. 1989. The ecology of bird comminities. Vol1. Foundations and patterns.
Cambirfge University Press. NY. 539p.
Wunderle, J. M. & S. C. Latta. (1998). Avian resource in Dominican shade coffe
plantations. Wilson Bulletin, 110, 271-281.

30
CAPÍTULO 1. EFECTO DE LA ESTRUCTURA DEL CULTIVO DE PALMA DE
ACEITE Elaeis guineensis SOBRE LA DIVERSIDAD DE AVES EN UN PAISAJE
TROPICAL DE COLOMBIA
Diana Patricia Tamaris-Turizo, Hugo Fernando López-Arévalo y Nabhi Romero Rodríguez

31
Resumen
La expansión del cultivo de palma de aceite ha ocasionado cambios drásticos en los
hábitats naturales y modificado la estructura de la vegetación, reduciendo el hábitat
disponible para las especies de aves nativas. Se estudiaron las aves asociadas a lotes de
palma de aceite de diferentes clases de alturas en el piedemonte de la Orinoquia
colombiana, entre agosto de 2013 y febrero de 2014. Las clases de altura fueron tres:
pequeña hasta seis metros, mediana hasta 10 metros y grande hasta15 metros, en cada lote
se censaron las aves mediante transectos lineales y se comparó la composición, riqueza,
abundancia y describió la estructura de los gremios de alimentación. Registramos 468
individuos de 44 especies y 23 familias. Gymnomystax mexicanus, Crotophaga ani y
Milvago chimachima fueron las especies más abundantes, las dos primeras representan el
4.5% de las especies abundantes, el 25% tuvo abundancias intermedias y el 70.4% fueron
especies raras en el área del cultivo. En términos de la abundancia proporcional de
especies de aves, las palmas grandes y medianas fueron más parecidas. La comunidad de
aves encontrada en parcelas de palma grande tuvo mayor diversidad y uniformidad (2.63 y
0.8907, respectivamente), mientras que en las palmas pequeñas hubo mayor dominancia y
abundancia de individuos (0.1825 and 252, cada cual). En todo el cultivo predominaron
aves insectívoras y carnívoras, las cuales tuvieron mayor afinidad por palmas pequeñas y
grandes. La palma de aceite puede funcionar como un hábitat favorable para un bajo
número de especies de amplia distribución y la altura no parece ser un variable que
beneficie de manera significativa la composición, riqueza y abundancia de las especies de
aves asociadas a la plantación.

32
Introducción
La complejidad estructural es un factor que regula la riqueza y diversidad de aves
terrestres (Cousin & Phillips, 2008; Hilden, 1965; MacArthur & MacArthur, 1962). La
asociación y organización de las aves en un ecosistema, también dependen de los recursos
disponibles para el forrajeo, la reproducción, el refugio frente a las adversidades del clima,
la presencia de parásitos y depredadores (Cousin & Phillips, 2008; Hilden, 1965). Por lo
tanto, las actividades humanas que conducen a los cambios en la composición y estructura
de la vegetación, pueden afectar la persistencia de aves por la insuficiencia de recursos,
aislamiento entre hábitats o niveles elevados de parasitismo y depredación (Miller & Cale,
2000).
La Orinoquía colombiana se ha destacado como una de las regiones que alberga
mayor riqueza de aves en Colombia, los registros incluyen 761 especies de aves que
representan aproximadamente el 7.6% de las especies a nivel mundial y el 40% de las
especies registradas para el país (McNish, 2007; Rangel-Ch & Minorta-Cely, 2014). Sin
embargo, esta diversidad de aves está en riesgo debido al extensivo y acelerado remplazo
de la vegetación nativa transformada en un monocultivo de palma de aceite, que viene
generando cambios en la composición y estructura de la vegetación (Umaña-Villaveces et
al., 2009). El sector palmero considera la Orinoquía como una región con gran potencial
para la expansión del cultivo de palma de aceite (Andrade et al., 2009). Estas plantaciones
se han establecido en la mencionada región desde los años 70´s y aportan cerca del 30% de
la producción nacional de aceite de palma (Andrade et al., 2009; Fedepalma, 2007). Su
producción y expansión es una respuesta de la alta demanda de biodiesel, productos
comestibles y otros productos no comestibles, lo cual resulta en acelerados cambios
ambientales (Koh & Ghazoul, 2008). Actualmente, el gobierno colombiano fomenta la
ampliación de la frontera agrícola a través del programa Desarrollo Rural con Equidad
(DRE) y del Incentivo a la Capitalización Rural (ICR) para pequeños y medianos
productores.

33
Los cultivos de palma de aceite son sistemas uniformes, el establecimiento de una
plantación implica la remoción de la vegetación nativa (Fitzherbert et al., 2008), las
variaciones estructurales del cultivo se reducen a la vegetación epífita que crece sobre
troncos, la vegetación que crece en el suelo y el sotobosque, cuando forma parte del
manejo del cultivo (Luskin & Potts 2011; Nájera & Simonetti 2010). La siembra de palma
de aceite a escala industrial, ha generado impactos negativos en diversos componentes de
la biodiversidad (Aratrakorn et al., 2006; Edwards et. al., 2010; Fitzherbert et al., 2008;
García-Ulloa et al., 2012), a través de la degradación de hábitat, destrucción y
reemplazamiento de la vegetación nativa, que en el caso de la Orinoquía afecta a bosques
húmedos, bosques secos, bosques de galería, pastizales y humedales (Blydenstein, 1967;
Bolwing et al., 2006; Pinzón Pinzón et al., 2009;), lo que genera cambios en la
composición y estructura de la vegetación (Correa & Stevenson, 2010).
Estos cultivos son dominados por un reducido número de aves comunes que tienen
amplio rango de distribución, mientras que todas aquellas especies asociadas a bosque
quedan confinadas a los relictos de vegetación nativa (Achondo et al., 2011; Aratrakorn et
al., 2006; Bolwing et al., 2006). Las aves asociadas a estos monocultivos habitan
diferentes espacios de la plantación y de acuerdo a la edad de la palma se han encontrado
especies distintas. Así, por ejemplo, se han encontrado especies de importancia de
conservación (endémicas y amenazadas) en palmares jóvenes cercanos a bosques nativos
(De Chenon & Susanto, 2006). La riqueza de especies de aves puede aumentar cuando hay
mayor complejidad estructural dentro del cultivo, p.e. epífitas, plantas presentes en el
suelo, sotobosque y árboles nativos dentro de la plantación (Cagod & Nuñeza, 2012;
Fitzherbert et al., 2008), pero también por el efecto de borde, cuando hay disponibilidad de
fragmentos de bosque, y por la complejidad estructural de la vegetación cercana al cultivo
(Cagod & Nuñeza, 2012).
El gremio palmicultor adelantan el proyecto Conservación de la Biodiversidad en
las Zonas de Cultivo de Palma de Aceite como una iniciativa orientada a la sostenibilidad
ambiental y que busca promover el principio de la responsabilidad con el medio ambiente
y conservación de recursos naturales y la biodiversidad (RSPO, 2014), sin embargo, en

34
Colombia hace falta rigor para que las prácticas de agronegocios se realicen de forma
sostenible. Con el fin de aportar información a la discusión sobre el impacto ambiental de
los agronegocios, se estudió la composición, riqueza y abundancia de aves, así como la
estructura de los gremios de alimentación, y se comparó con tres clases de altura del
cultivo de palma de aceite, en la región oriental de Colombia.
Métodos
Área de estudio: El estudio se realizó en dos fincas con cultivos de palma de aceite Tierra
Viva (TIV) y La Mejorana (MEJ). TIV pertenece al municipio de San Martín de los
Llanos, tiene un área de 1 400 ha y está ubicada a los 03°37'51" N & 73°20'59" W, y a 420
m.s.n.m. MEJ se encuentra en el municipio de Acacias, la finca tiene un área de 1 480 ha y
se encuentra ubicada a los 03°57'37.18" N & 73°37'32.93" W, y a 498 msnm. Estos
municipios pertenecen al departamento del Meta (Llanos Orientales de Colombia).
El área de estudio forma parte de la unidad fisiográfica piedemonte depositacional,
derivado de rocas terciarias y cretáceas (Casanare y Meta), con un área total de 2.532.526
ha, que corresponde al 6.7% del área total de la cuenca del Orinoco, dentro del zonobioma
húmedo tropical conformado por 67 ecosistemas que se localizan entre los 50 a 1 100
m.s.n.m. (Correa et al., 2005). Las formaciones vegetales más representativas son bosques
de estrato arbóreo con elementos de 18 y 30 m de altura, herbazales con elementos
leñosos, pastizales con resto de bosque, pastizales con elementos de 20 cm de altura donde
domina Paspalum carinatum y palmares con predominio de Socratea exorrhiza e Iriartea
deltoidea (Blydenstein, 1967; Rangel-Ch & Minorta-Cely, 2014). Este tipo de vegetación
se ha modificado desde el siglo XX por la actividad ganadera y por el establecimiento y
consolidación de la agricultura comercial basada en cultivos de palma de aceite, arroz,
maíz, sorgo, soya, frutales, entre otros (Mora Fernández et al., 2011; Rippstein et al.,
2001).
Censos de aves: Se hicieron observaciones de aves durante cuatro días consecutivos en
junio, agosto y noviembre de 2013, y en febrero y julio de 2014, en 18 lotes (parcelas):

35
seis de palmas pequeñas (de 2 a 6 m), seis de palmas medianas (7-10 m) y seis de palmas
grandes (11-15 m). Se definieron estas clases de altura teniendo en cuenta que la altura
máxima promedio de una palma en una plantación es de 15 a 18 m y que a partir de este
rango de altura puede haber reemplazo de las palmas, por la dificultad para la cosecha de
frutos (ASD, 2006). Los meses de muestreo coincidieron con eventos de sequía (febrero) y
lluvia (julio, agosto y noviembre), correspondientes al régimen unimodal biestacional
característico de la región (Minorta-Cely & Rangel-Ch). El área de los lotes fue variable
(23.3 a 48.7 ha), incluyendo 160 palmas/ha. La altura de las palmas se consideró desde la
base del tronco hasta el extremo superior del dosel (Luskin & Potts, 2011). En general los
troncos tuvieron plantas epífitas, y en ambas localidades se implementaron técnicas
manuales y mecánicas para el control de hierbas y prevenir el crecimiento de vegetación
arbustiva.
Las aves se contaron con el método de transecto lineal de ancho variable (Buckland et al.,
1993). Para ello, en cada lote, se estableció una línea de 500 m, y se censaron todas las
aves dentro del lote, en cada caso se tuvo en cuenta la distancia del ave con respecto al
transecto, lo cual se usó para calcular la densidad de individuos por km2 para todo el
cultivo y por clase de altura. Los transectos se ubicaron teniendo en cuenta el tamaño del
lote y la accesibilidad a éste, manteniendo una distancia mínima de 800 m con respecto al
transecto más cercano, con el fin de garantizar la independencia de las observaciones. Este
método de conteo es recomendable en hábitats de baja densidad con una estructura
uniforme (Bibby et al., 2000). Cada transecto se recorrió seis veces durante el periodo de
estudio, completando 108 recorridos en total, pero para el análisis de datos se tuvieron en
cuenta los recorridos donde hubo registro de especies (81 recorridos). El orden para
recorrer los transectos se escogió aleatoriamente en cada visita, para evitar sesgos en los
conteos según la hora del día (Bibby et al., 2000), y se dedicó 40 minutos en cada
recorrido. En el registro se incluyeron las especies observadas y escuchadas (Bibby et al.,
2000). Un observador tomó los registros en la mañana (06:00 a 10:30) y en la tarde (15:00
a 17:30), y se invirtió un esfuerzo de muestreo mínimo diario de 2.5 km/día; se utilizó
binoculares Nikon Action de alcance 8 × 40 mm. Durante las observaciones no se tuvieron

36
en cuenta los individuos de paso o que sobrevolaron el sitio de estudio por encima de los
20 m (Codesido & Bilenca, 2004). Todas las especies observadas fueron asignadas a una
de las siguientes categorías de grupo trófico: carnívoro (CAR), carnívoro correñoero
(CAR-CRR), carreñero (CRR), frugívoro (FRU), granívoro (GRA), insectívoro (INS),
nectarívoro (NEC) y omnívoro (OMN). Estas se basaron tanto en las observaciones de
campo como en la literatura disponible (Hilty & Brown 2001; Restall et al., 2006).
Análisis de datos
Se construyó una curva de acumulación de especies utilizando los registros de aves (global
para el cultivo y discriminado por clases de altura) para identificar si el esfuerzo de
muestreo fue lo suficientemente representativo. También se estimó el número de especies
posibles con los estimadores de riqueza Chao1 y ACE. Se graficaron los valores de
abundancia de cada especie (n), registradas en cada clase de altura, para visualizar la
proporción de las aves dentro de la plantación.
La diversidad de la comunidad de aves fue analizada a partir de índices de diversidad
(Magurran, 1988; Moreno, 2001. La diversidad α fue el número total de especies obtenido
en palmas de cada clase de altura. El índice de diversidad de Brillouin (H), el índice de
equitatividad Pielou (J) y el índice de dominancia de Simpson (λ), mientras que la
diversidad β (recambio de especies entre palmas grandes: G, medianas: M y pequeñas: P)
se calculó, a partir de los valores de riqueza (S) y de abundancia (N) en cada clase de
altura, con el Coeficiente de Similaridad, el índice de Whittaker (βW) y el índice de
Complementareidad según lo describe Magurran (1988). También se generó el modelo de
distribución de abundancias (Magurran, 1988) para describir la estructura de la comunidad
en términos de la abundancia proporcional de las especies.
A partir de un coeficiente de correlación (Krebs, 1999) se analizó el grado de similitud
teniendo en cuenta la abundancia de las especies, entre cultivos de palmas grandes,
medianas y pequeñas, utilizando el programa PAST 3. Un análisis de correspondencia
rectificado (DCA) se usó para identificar si los gremios de alimentación tuvieron una
segregación con respecto a las clases de altura de la plantación, asimismo, se aplicó la

37
prueba Chi2 para identificar posibles diferencias significativas de la abundancia de aves de
cada grupo trófico, con respecto a las clases de altura de la palma. Finalmente, se utilizó la
prueba no paramétrica Friedman para identificar si existían diferencias significativas de la
abundancia de individuos entre las clases de altura.
Resultados
Durante el estudio se registraron 468 individuos asociados al cultivo de palma, agrupados
en 44 especies, 23 familias y 14 órdenes. A partir de las abundancias absolutas, las
especies más numerosas dentro del cultivo de palma fueron Gymnomystax mexicanus,
Crotophaga ani y Milvago chimachima; mientras que Orthopsittaca, Crotophaga ani y
Pteroglosus castanotis fueron predominantes al analizar la abundancia promedio. Las dos
primeras fueron las especies más dominantes en palma pequeña, mientras que M.
chimachima fue la especie más dominante en lotes de palma mediana y grande (Fig. 1.1 y
1.2).
La curva de acumulación de especies total (Fig. 2.3A) no se estabilizó, lo cual indica que
el censo que se hizo en campo no fue suficiente para proporcionar una caracterización
completa de las aves presentes en el cultivo de palma de aceite, aunque la riqueza
observada (44 especies) corresponde al 88% de la riqueza calculada por el estimador
Chao1y ACE (48 y 50 especies, respectivamente). La rareza de las especies generó un
efecto ascendente en las curvas de acumulación. Un comportamiento parecido al anterior
se observó en la curva de acumulación de especies para cada clase de altura (Fig. 1.3 B).
La riqueza observada en cada clase de altura correspondió al 96 (n=27), 80 (n=20) y 93%
(n=30) con respecto a la riqueza calculada por el estimador de riqueza Chao 1 en palmas
grandes, medianas y pequeñas, respectivamente.
La similitud a partir del coeficiente de correlación entre pares de sitios indicó que las
palmas grandes y medianas fueron más similares entre sí (0.816) en términos de la
abundancia proporcional de las especies (Cuadro 1.1). La densidad de individuos fue
mayor en las palmas pequeñas (6.9 ind./km2, SD± 2.65), reduciéndose a medida que

38
aumentó la altura (4.0 ind./ km2, SD± 8.5 y 2.8 ind./ km2, SD± 6.6, en palmas medianas y
grandes, respectivamente). Los modelos de la distribución de abundancia de las especies
fueron semejantes en las tres clases de altura del cultivo de palma y no hubo diferencia
significativa entre clases de altura (Fig. 1.4, Friedman= 1,67, p=0.3670). Encontramos que
el 70% de la comunidad fueron especies raras (especies con 10 o menos individuos), el
25% tuvieron abundancias intermedias (especies con 11 a 49 individuos) y el 4.5% de las
especies fueron abundantes dentro de la plantación (especies con 50 o más individuos), lo
cual se ajusta al modelo Serie Logarítmica. Los índices de diversidad demostraron que la
palma grande tuvo mayor equidad (H) y uniformidad (J´) en la abundancia proporcional de
las especies, mientras que en las palmas pequeñas tuvieron mayor riqueza (S) y
dominancia (λ) de especies (Tabla 1.1). Al comparar entre pares de biotas, la mayor
diferencia en la riqueza de especies fue entre palmas grandes y pequeñas (βW), siendo a la
vez más complementarias (CA,B) (Tabla 1.1).
La estructura trófica de las aves asociadas al cultivo de palma de aceite estuvo conformada
por ocho grupos y predominaron las especies insectívoras, seguido de las carnívoras, las
cuales estuvieron principalmente presentes en palmas pequeñas y medianas,
respectivamente (Chi2= 342.4, valor de p < 0.001) (Tabla 1.2). Las aves insectívoras más
comunes estuvieron representadas por Gymnomystax mexicanus, Crotophaga ani,
Tyrannus melancholicus y Cyanocorax violaceus, y las carnívoras por Milvago
chimachima, Rupornis magnirostris y Tyto alba. Finalmente, con el análisis de
ordenación, se pudo identificar que las asociaciones más importantes fueron de aves
nectarívoras en palmas pequeñas y medianas; por su parte las aves insectívoras mostraron
mayor afinidad a palmas pequeñas y grandes, y las aves carnívoras y carroñeras tuvieron
mayor asociación con palmas grandes y medianas (Fig. 1.5).
Discusión
La abundancia de aves no fue significativamente diferente entre las clases de altura de las
palmas. La riqueza total observada (44 especies) refleja que el monocultivo estudiado
soportó un bajo número de especies si se tiene en cuenta que representan el 5.78% de las

39
especies reportadas para la región, el 6.44% de las especies reportadas para el
departamento del Meta, y corresponde al 10.6% de las especies reportadas para el
piedemonte de la Orinoquía colombiana (Acevedo-Charry et al., 2014; McNish, 2007). El
estudio como el de Enríquez Bernal (2006) también encontró un reducido número de
especies de aves asociadas al cultivo de palma de aceite (27 y 29 en palmas pequeñas y
grandes), en comparación con otros ecosistemas como bosque de galería y pastizales
donde registraron 41 y 26 especies, respectivamente. Nuestros resultados también
demostraron que puede presentarse una baja diversidad de aves dentro del monocultivo
indistintamente de la clase de altura, al considerar que en los recorridos donde no hubo
registro de aves (n=27), ocurrieron en igual proporción en sectores de palmas grandes,
medianas y pequeñas.
Las curvas de rarefacción estuvieron influenciadas por las especies únicas y duplicadas, lo
que genera la mayoría de las veces que los muestreos se consideren incompletos. Sin
embargo, esto no invalida el trabajo, más bien demuestra que son pocos individuos, de las
diferentes especies reportadas, que hacen uso de las plantaciones de palmas. La ausencia
de un porcentaje considerable de especies de aves propias del piedemonte, puede estar
relacionada con la pérdida de bosque, la expansión del monocultivo y los cambios en el
uso del suelo en los municipios de Acacias y San Martín de los Llanos, en los cuales se ha
reportado un crecimiento en el área cultivada del 10.23 y 5.67%, respectivamente, entre
los años 1988 y 2005 (IDEAM, 2016; Pinzón Pinzón et al., 2009). De igual forma ha
ocurrido en Villanueva (Casanare), donde encontraron que el cultivo de palma de aceite ha
sido la cobertura de mayor expansión entre el 2002 y el 2012 con respecto a otros usos de
suelo, periodo en el cual presentó una ganancia de 1 923.15 ha, y que a su vez ha
sustituido pastos, bosques de galería y vegetación secundaria, considerados ecosistemas
estratégicos para la conservación de la biodiversidad (González & Romero, 2013).
La siembra a escala industrial ha promovido la remoción de formaciones vegetales
originales: bosques altos, palmares de Socratea exorrhiza, vegetación acuática, matorrales
y pastizales con restos de bosque (Rangel-Ch & Minorta-Cely, 2014), a su vez que se han
generado eventos de extinción local de las aves asociadas al piedemonte: Anhima cornuta,

40
Neochen jubata, Amazonetta brasilensis, Aburria aburri, Mitu tomentosum, Crax
daubentoni, Harpia harpia, Opisthocomus hoazín, Eurypyga helias, Geotrigon linearis,
Orthopsittaca manilata, Atene cunicularia, Nyctibius grandis, Eutoxeres aquila,
Phaetornis malaris, Pachyramphus cinnamomeus, P. polychopterus, Anthu lutescens, entre
otras especies (Acevedo-Charry et al., 2014, McNish, 2007, Olivares, 1982).
El reemplazo de la vegetación original que incluye bosques con un estrato arbóreo de 30 m
(Cassia moschata, como uno de los árboles dominantes), por un sistema que a gran escala
es uniforme en edad y estructura, de bajo dosel (13 m), escaso sotobosque y un microclima
más severo (Fitzherbert et al., 2008; Luskin & Potts 2011), reduce la asociación de
especies en comparación con las formaciones vegetales originales (Acevedo-Charry et al.,
2014). Tendencia que también documentó Romero (2014) al reportar 59 especies de aves
en vegetación arbustiva y arbórea de Barbascal (Meta), de las cuales 49 no estuvieron
presentes en una plantación comercial de palma de aceite adyacentes la vegetación
analizada. Prescott et al. (2016) también sostienen que la riqueza y la diversidad funcional
de las aves es menor en palmares de aceite que fueron comparados con remanentes de
bosque.
La reducción de la riqueza de aves dentro de este monocultivo es mayor que en otros
cultivos, donde se ha reportado mayor número de especies. Por ejemplo, en cultivos de
café con sombra se han registrado 141 especies (Bayly & Gómez, 2010), en arrozales 150
(Cifuentes-Sarmiento & Castillo Cortés 2016), mientras el presente estudio reporta 44 aves
asociadas al cultivo de palma de aceite, y estudios previos han registrado 34 y 29 especies
en plantaciones localizadas en el piedemonte amazónico y acacias, respectivamente
(Enríquez Bernal 2006; Velásquez Valencia, 2009).
La abundancia de individuos fue notablemente mayor en palmas pequeñas. Esta tendencia
difiere con otros estudios donde han encontrado mayor abundancia de aves en plantaciones
maduras que en palmas jóvenes como en palmares de Indonesia y Filipinas (Cagod &
Nuñeza, 2012; De Chenon & Susanto, 2006). Se argumenta que las palmas maduras
soportan mayor biodiversidad que las palmas recién sembradas porque tienen mayor
complejidad estructural, mayor desarrollo de sotobosque y un microclima más conveniente

41
(Azhar et al., 2015; Luskin & Potts, 2011). En nuestro caso la remoción de plantas
herbáceas y arbustivas del suelo le resta complejidad al cultivo, la asociación de
invertebrados y artrópodos y con ello recursos potenciales de forrajeo y alimentación para
las aves (Cagod & Nuñeza, 2012).
La estructura trófica de la comunidad estuvo conformada por ocho grupos y las aves
insectívoras fueron predominantes. Esta tendencia se ha encontrado en estudios previos y
se ha considerado que estos conjuntos de especies actúan sobre el control de insectos que
representan riesgos para el cultivo (Achondo et al., 2011, Azhar et al., 2013, Enríquez
Bernal, 2006). Sin embargo, también se ha encontrado que las aves generalistas y
frugívoras son abundantes. Azhar et al. (2013) encontraron que las aves frugívoras fueron
abundantes en palmas pequeñas, posiblemente porque los lotes donde fueron registradas
tuvieron mayor proximidad con árboles nativos, y que la mayoría de los gremios de
alimentación fueron dominados por pocas especies. Las aves nectarívoras pueden ser uno
de los grupos con mayor sensibilidad al establecimiento de la plantación si se tiene en
cuenta la escasa abundancia proporcional de los individuos (Amazilia versicolor). Estudios
previos no relacionan la presencia de este grupo trófico con el cultivo de palma, lo que
concuerda con la reducción la diversidad funcional de las aves en paisajes dominados por
cultivos de palma de aceite (Azhar et al., 2013, Prescot et al., 2016).
Esta investigación destaca que los cultivos de palma de aceite pueden funcionar como un
hábitat favorable para un bajo número de especies, en este caso Gymnomystax mexicanus,
Crotophaga ani, Milvago chimachima y Rupornis magnirostris, si nos basamos en su
abundancia proporcional, pero también pueden funcionar como un hábitat poco amigable o
como una barrera de movimiento para el 75% de las especies que fueron raras dentro de la
comunidad, debido a la pérdida y fragmentación del bosque, y porque el aislamiento entre
parches de bosque, impide el movimiento de aves, aunque rutinariamente muchas cruzan
terrenos inhóspitos (Bélisle et al., 2001). Este conjunto de especies (el 75%) frecuenta
principalmente ecosistemas con vegetación arbustiva y arbórea (Hilty & Brown, 2001),
pero la escasa complejidad del cultivo reduce la explotación de recursos y el bajo grado de
asociación de las aves dentro de este sistema.

42
Este estudio encontró que los cultivos de palma de aceite tuvieron una baja asociación de
aves, y las especies que estuvieron presentes son de amplia distribución y baja importancia
de conservación. La variación de altura de las palmas aparentemente no refleja un
beneficio para las aves, al observarse que la abundancia de individuos no presentó
diferencias significativas entre clases de altura. La expansión acelerada de este
monocultivo podría representar un detrimento en la disponibilidad y calidad de hábitat
para las aves, si tenemos en cuenta que el 75% de los miembros de la comunidad fueron
raros dentro de la plantación. Con el panorama actual es recomendable que, si se decide
expandir este monocultivo, se mantenga la vegetación nativa existente, para que estos
relictos puedan proporcionar recursos de alimento, refugio, sitios de reproducción y
movimiento, a las especies que fueron escasas y raras dentro del cultivo y aquellas que
usan este tipo de cobertura.

43
Especie
Gym
nom
ysta
x m
exic
anus
Cro
toph
aga
ani
Milv
ago
chim
achi
ma
Rup
orni
s m
agni
rost
ris
Phi
mos
us in
fusc
atus
Tyr
annu
s m
elan
chol
icus
Ortho
psitt
aca
man
ilata
Cya
noco
rax
viol
aceu
s
Syr
igm
a si
bila
trix
Thr
aupi
s pa
lmar
um
Col
umbi
na ta
lpac
oti
Mim
us g
ilvus
Pat
agio
enas
cay
enne
nsis
Pte
rogl
osus
cas
tano
tis
Van
ellu
s ch
ilens
is
Col
inus
cris
tatu
s
Am
azon
a oc
roce
phal
a
Dry
ocop
us li
neat
us
Pita
ngus
lict
or
Núm
ero
de indiv
iduos
0
20
40
60
80
100
120
Grande
Mediano
Pequeño
Figura 1. 1 Distribución de la abundancia de especies en cada clase de altura del cultivo de
palma de aceite Elaeis guineensis
Especie
Orth
opsitta
ca m
anila
taCro
toph
aga
ani
Pte
rogl
osus
cas
tano
tis
Gym
nom
ysta
x m
exic
anus
Phi
mos
us in
fusc
atus
Cya
noco
rax
viol
aceu
s
Milv
ago
chim
achi
ma
Mim
us g
ilvus
Am
azon
a am
azon
ica
Col
inus
cris
tatu
s
Thra
upis
epi
scop
usSyr
igm
a sibi
latri
x
Pro
med
io d
e la
s a
bund
anci
as (
# de
Indi
vidu
os)
0
2
4
6
8
10
12
14
16
Grande
Mediano
Pequeño
Figura 1. 2 Distribución de la abundancia promedio de especies en cada clase de altura del
cultivo de palma de aceite Elaeis guineensis

44
Número de muestras
0 10 20 30 40 50 60 70 80 90
Núm
ero
de
esp
ecie
s a
cu
mu
lad
o
0
10
20
30
40
50
60
S
ACE
Chao1
Número de muestras
0 5 10 15 20 25 30 35
Núm
ero
de e
specie
s
0
5
10
15
20
25
30
35
S Grande
Chao1 Grande
S Mediano
Chao1 Mediano
S Pequeño
Chao1 Pequeño
Figura 1. 3 Curva de acumulación de especies de aves: A) asociadas a todo el cultivo de
palma de aceite y B) a cada clase de altura
A
B

45
Rango de especies
0 20 40 60 80 100 120 140
Abundancia
(Log10)
-3.0
-2.5
-2.0
-1.5
-1.0
-0.5
0.0 Grande
Mediano
Pequeño
Total
Figura 1. 4 Modelo de distribución de abundancia de especies en aves asociadas a cultivos de
palma de aceite Elaeis guineensis. G: grande, M: mediano, P: pequeño, Total: todas las clases
de altura
Figura 1. 5 Análisis de correspondencia rectificado (DCA) de los gremios de alimentación de
aves en las clases de altura del cultivo de palma de aceite Elaeis guineensis
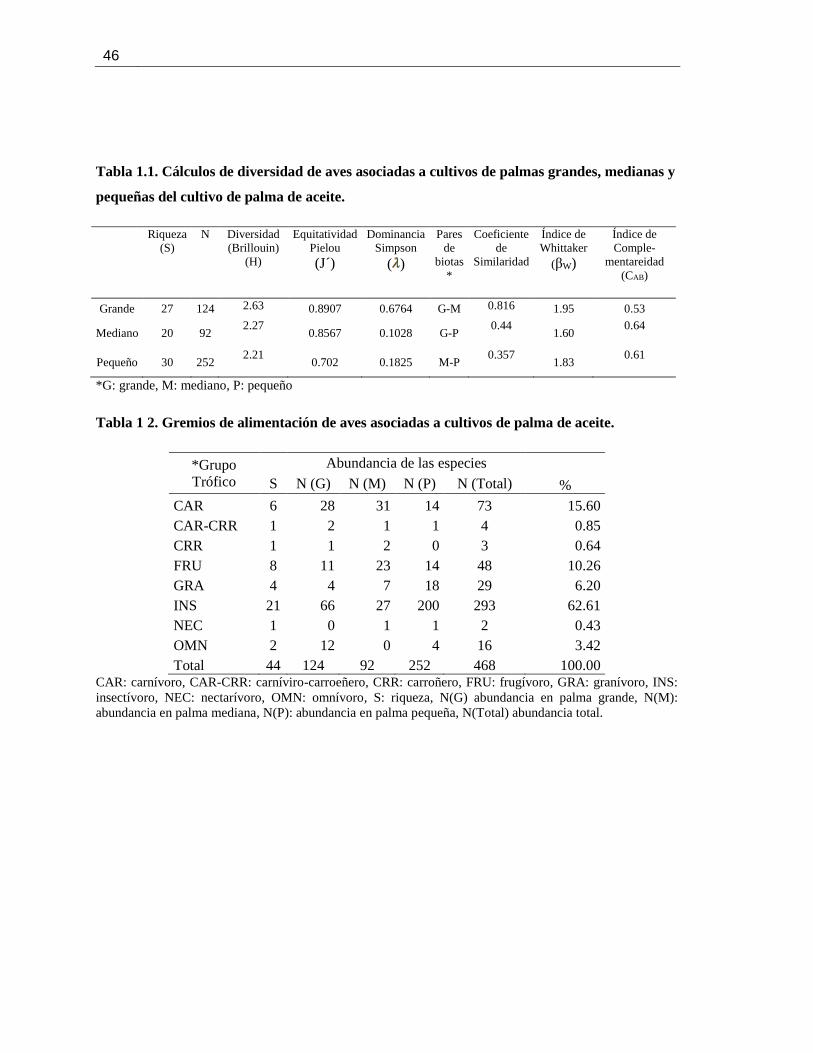
46
Tabla 1.1. Cálculos de diversidad de aves asociadas a cultivos de palmas grandes, medianas y
pequeñas del cultivo de palma de aceite.
Riqueza
(S)
N Diversidad
(Brillouin)
(H)
Equitatividad
Pielou
(J´)
Dominancia
Simpson
( )
Pares
de
biotas
*
Coeficiente
de
Similaridad
Índice de
Whittaker
(βW)
Índice de
Comple-
mentareidad
(CAB)
Grande 27 124 2.63 0.8907 0.6764 G-M 0.816 1.95 0.53
Mediano 20 92 2.27
0.8567 0.1028 G-P 0.44
1.60 0.64
Pequeño 30 252 2.21
0.702 0.1825 M-P 0.357
1.83 0.61
*G: grande, M: mediano, P: pequeño
Tabla 1 2. Gremios de alimentación de aves asociadas a cultivos de palma de aceite.
*Grupo
Trófico
Abundancia de las especies
S N (G) N (M) N (P) N (Total) %
CAR 6 28 31 14 73 15.60
CAR-CRR 1 2 1 1 4 0.85
CRR 1 1 2 0 3 0.64
FRU 8 11 23 14 48 10.26
GRA 4 4 7 18 29 6.20
INS 21 66 27 200 293 62.61
NEC 1 0 1 1 2 0.43
OMN 2 12 0 4 16 3.42
Total 44 124 92 252 468 100.00 CAR: carnívoro, CAR-CRR: carníviro-carroeñero, CRR: carroñero, FRU: frugívoro, GRA: granívoro, INS:
insectívoro, NEC: nectarívoro, OMN: omnívoro, S: riqueza, N(G) abundancia en palma grande, N(M):
abundancia en palma mediana, N(P): abundancia en palma pequeña, N(Total) abundancia total.

47
Referencias
Acevedo-Charry, O., Pinto-Gómez, A., & Rangel-Ch, J. O. (2014). Las aves de la Orinoquía
colombiana: una revisión de sus registros. En Rangel-Ch, J. O. (Ed.), Colombia Diversidad
Biótica XIV. La región de la Orinoquía de Colombia (pp. 691-750). Universidad Nacional de
Colombia – Instituto de Ciencias Naturales. Bogotá, D.C., Colombia.
Achondo, M. J. M., Casin, L. F., Bello, V. P., Tanalgo, K. C., Agduma, A. R., Bretaña, B. L. P.,
Mancao, L. S., Salem, J. G. S., & Supremo, J. P. (2011). Rapid assessment and feeding
guilds of birds in selected Ruber and Oil Palm plantations in North Cotabato. Asian Journal
of Biodiversity, 2, 103-120.
Andrade Pérez, G. I., Castro Gutiérrez, L. G., Durán Durán, A., Rodríguez Becerra, M., Rudas
Lleras, G., Uribe Botero, E., & Wills Herrera, E. (2009). La mejor Orinoquía que podemos
construir Elementos para la sostenibilidad ambiental del desarrollo. Universidad de Los
Andes y Corporinoquía. Bogotá, Colombia.
Aratrakorn, S., Thunhikorn, S., & Donald, P. F. (2006). Changes in bird communities following
conversion of lowland forest to oil palm and rubber plantations in southern Thailand. Bird
Conservation International, 16, 71-82.
ASD (Agricultural Services and Development). (2006). Aspectos Generales del cultivo de la palma
aceitera (Elaeis guineensis Jacq.). San José, Costa Rica: Agricultural Services and
Development.
Azhar, B., Lindenmayer, D. B., Wood, J., Fischer, J., Manning, A., McElhinny, C., & Zakaria, M.
(2013). The influence of agricultural system, stand structural complexity and landscape
context on foraging birds in oil palm landscapes. Ibis, 155, 297-312.
Azhar, B., Saadun, N. Leong Puana, Ch., Kamarudin, N., Aziz, N., Nurhidayu, S., & Fischer, J.
(2015). Promoting landscape heterogeneity to improve the biodiversity benefits of certified
palm oil production: Evidence from Peninsular Malaysia. Global Ecology and Conservation,
3, 553-561.
Bayly, N. J., & Gómez, C. (2010). Las aves de la Hacienda La Victoria. (Informe técnico No.
CEC01). Bogotá, Col. SELVA: Investigación para la conservación en el Neotrópico,
Bogotá.
Bélisle, M., Desrochers, A., & Fortin, M. J. (2001). Influence of forest cover on the movements of
forest birds: a homing experiment. Ecology, 82(7), 1893-1904.
Bibby C. J., Jones, M., & Marsden, S. (2000). Expedition field techniques birds survey. 2nd
Edition Academic Press. London, UK.
Blydenstein, J. (1967). Tropical savanna vegetation of the Llanos de Colombia. Ecology, 48(1), 1-
15.

48
Bolwing, S., Pomeroy, D., Tushabe, H. & Mushabe, D. (2006). Crops, trees, and birds:
Biodiversity change under agricultural intensification in Unganda´s farmed landscape.
Danish Journal of Geography, 106(2), 115-130.
Buckland, S.T., Anderson, D. R., Burnham, K. P., & Laake, J. L. (1993). Distance Sampling:
Estimating Abundance of Biological Populations. Chapman and Hall, London, UK.
Cagod, B. M., & Nuñeza, O. M. (2012). Avian species diversity in oil palm plantations of Agusan
Del Sur and Compostela Valley, Philippines. AES-Bioflux, 4(2), 85-105.
Cifuentes-Sarmiento, Y., & Cantillo Cortés, L. F. (2016). Las alas del arroz: aves acuáticas en
cultivos ecoamigables del Valle del Cauca. Asociación Calidris & Arrocera La Esmeralda
S.A.S. Cali. Colombia.
Codesido, M., & Bilenca, D. (2004). Variación estacional de un ensamble de aves en un bosque
subtropical semiárido del Chaco Argentino. Biotropica, 36(4), 544-554.
Corley, R. H., & Tinker, P. B. (2003). The Palm Oil. 4th Edition. Oxford Blackwell Science. USA.
Correa, H. D, Ruiz, S. L., & Arévalo, L. M. (Eds.) (2005). Plan de acción en biodiversidad de la
cuenca del Orinoco – Colombia / 2005 - 2015 - Propuesta Técnica. Bogotá D.C.
Correa, D. F., & Stevenson, P. R. (2010). Estructura y diversidad de los bosques de galería en una
sabana estacional de los llanos orientales colombianos (Reserva Tomo Grande, Vichada).
Orinoquía, 14 (1), 31-48.
Cousin, J., & Phillips, R. D. (2008). Habitat complexity explains species-specific occupancy but
not species richness in a Western Australian Woodland. Australian Journal Zoology, 56, 95-
102.
De Chenon, R., & Susanto, A. (2006). Ecological observations on diurnal birds in Indonesian oil palm
plantations. Journal of Oil Palm Research (Special Issue April), 122-143.
Edwards, D. P., Hodgson, J. A., Hamer, K. C., Mitchell1, S. L., Ahmad, A. H., Cornell, S. J., &
Wilcove, D. S. (2010). Wildlife-friendly oil palm plantations fail to protect biodiversity
effectively. Conservation Letter, 3, 236-242.
Enríquez Bernal, L. M. (2006). Estructura y composición de la comunidad de aves asociadas a
plantaciones de palma de aceite (Elaeis guineensis Jacq.), en Agropecuaria La Loma
(Acacias - Meta). (Tesis de Pregrado), Pontificia Universidad Javeriana. Bogotá, Colombia.
Fedepalma (Federación Nacional de Cultivadores de Aceite de Palma). (2007). Localización
geográfica de cultivos y planta de beneficio. Sistema de Información Estadística del Sector
Palmero. Recuperado de www.fedepalma.org.
Fitzherbert, E. B., Struebig, M. J., Morel, A., Danielsen, F., Brühl, C. A., Donald, P. F., & Phalan,
B. (2008). How will oil palm expansion affect biodiversity?. Trends in Ecology and
Evolution, 23(10), 538-545.
García-Ulloa, J., Sloan, S., Pacheco, P., Ghazoul, J., & Pin Koh, L. (2012). Lowering
environmental cost of oil-palm expansion in Colombia. Conservation Letters, 5, 366-375.

49
González, L. M., & Romero, A. P. (2013). Análisis multitemporal de los cambios de cobertura de
la tierra e incidencia del cultivo de palma de aceite en el territorio del municipio de
Villanueva Casanare. (Tesis de Maestría). Pontificia Universidad Javeriana. Bogotá,
Colombia.
Hilden, O. (1965). Habitat selection in birds: A review. Annales Zoologici Fennici, 2(1), 53-75.
Hilty, S., & Brown, W. L. (2001). Guía de las aves de Colombia. American Bird Conservancy-
ABC. Imprelibros SA. Cali, Colombia.
Koh, L. P., & Ghazoul, J. (2008). Biofuels, biodiversity, and people: understanding the conflicts
and finding opportunities. Biological Conservation, 141(10), 2450-2460.
Krebs, Ch. J. (1999). Ecological Methodology. 2nd Ed. Addison Wesley Longman. CA. E.U.
Luskin, M. S., & Potts, M. D. (2011). Microclimate and habitat heterogeneity through the oil palm
lifecycle. Basic and Applied Ecology, 12, 540-551.
MacArthur, R. H., & MacArthur, J. W. (1962). On bird species diversity. The American Naturalist,
96 (888), 167-174.
Magurran, A. (1988). Ecological diversity and its measurement. New Jersey, USA.
McNish, T. (2007). Las aves de los Llanos de la Orinoquía. M&B Ltda. Bogotá, Colombia.
Miller, J. R., & Cale, P. (2000). Behavioral mechanism and habitat use by birds in a fragmented
agri cultural landscape. Ecological Applications, 10(6), 1732-1748.
Minorta-Cely, V., & Rangel-Ch. J. O. (2014). Clima de la Orinoquía colombiana. En Rangel-Ch, J.
O. (Ed.), Colombia Diversidad Biótica XIV. La región de la Orinoquía de Colombia (pp.
153-206). Universidad Nacional de Colombia – Instituto de Ciencias Naturales. Bogotá,
D.C., Colombia.
Mora Fernández, C., Castellanos Castro, C., Cardona Cardozo, A., Pinzón Pérez, L., & Vargas, O.
(2011). Historia de transformación del paisaje, de la cuenca baja del río Pauto, Casanare
(Colombia). En: Tomás León Sicard (Ed.). Mamíferos, Reptiles y Ecosistemas del Bloque
Cubiro (Casanare). Educación Ambiental para la Conservación. Instituto de Estudios
Ambientales, Universidad Nacional de Colombia, Bogotá, Colombia.
Moreno, C. (2001). Métodos para medir la biodiversidad. M&T-Manuales y Tesis SEA. Vol. 1.
Zaragoza, España.
Nájera A., & Simonetti, J. A. (2010). Enhancing Avifauna in Commercial Plantations.
Conservation Biology, 24(1), 319-324.
Olivares, A. (1982). Aves de la Orinoquía colombiana. 2da Edición Instituto de Ciencias Naturales
Universidad Nacional de Colombia. Bogotá, Colombia.
Pinzón Pinzón, F., Pardi Lacruz, M. S., Trejos Serrato, J., & Midori Saito, S. (Abril, 2009). Uso de
técnicas de percepción remota para la estimación del incremento de áreas cultivadas de
palma aceitosa y su relación con el conflicto armado en el Departamento del Meta-
Colombia. Anais En. XIV Simpósio Brasileiro de Sensoramiento Remoto. Natal, Brasil.

50
Prescott, G. W., Gilory, J. J., Haugaasen, T., & Medina Uribe, C. A. (2016). Reducing the impacts
of Neotropical oil palm development on functional diversity. Biological Conservation, 197,
139-145.
Rangel-Ch, O., & Minorta-Cely, V. (2014). Tipos de vegetación de la Orinoquía colombiana. En
Rangel-Ch, J. O. (Ed.), Colombia Diversidad Biótica XIV. La región de la Orinoquía de
Colombia. Universidad Nacional de Colombia – Instituto de Ciencias Naturales. Bogotá,
Colombia.
Restall, R., Rodner, C., & Lentino, R. (2006). Birds of northern South America. Christopher Helm.
London.
Rippstein, G., Escobar, G., & Motta, F. (Eds.). (2001). Agroecología y biodiversidad de los Llanos
Orientales de Colombia. Centro Internacional de Agricultura Tropical (CIAT) # 322. Cali,
Colombia.
Romero N. (2014). Efecto de la estructura de la cobertura natural sobre la composición de aves de
sotobosque en áreas dominadas por cultivos de palma africana en San Martin, Meta. (Tesis
de pregrado). Universidad Nacional de Colombia. Bogotá, Colombia.
Roundtable on Sustainable Palm Oil (RSPO) (2014). Principios y Criterios para la producción de
aceite de palma sostenible. Documento Borrador de la Interpretación Nacional (IN) para
Colombia del estándar RSPO 2013 para consulta pública. Fedepalma.
Umaña-Villaveces, A. M., Murillo, J. I., Restrepo-Calle, S. & Álvarez-Rebolledo, M. (2009).
Indicadores de seguimiento de la biodiversidad en el piedemonte orinoquense, sabanas y
bosques asociados al norte del río Guaviare: Aves. En: Romero, M. H., Maldonado-Ocampo,
J. A., Gregory, J. D., Usma, J. S., Umaña-Villaveces, A. M., Murillo, J. I., Restrepo-Calle,
S., Álvarez, M., Palacios-Lozano, M. T., Valbuena, M. S., Mejía, S. L. Aldana-Domínguez,
J & Payán, E. Informe sobre el estado de la biodiversidad en Colombia 2007-2008:
Piedemonte orinoquense, sabanas y bosques asociados al norte del río Guaviare (pp 48-78).
Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Bogotá, D.C.
Velásquez Valencia, A. (2009). Estructura de la comunidad de aves en sistemas de producción del
piedemonte amazónico. (Tesis de Maestría) Universidad Nacional de Colombia.

51
CAPÍTULO 2. USO DE RECURSOS DE AVES EN PAISAJES DOMINADOS POR
CULTIVOS DE PALMA DE ACEITE (Elaeis guineensis)
Diana Patricia Tamaris-Turizo y Hugo Fernando López-Arévalo

52
Resumen
La conversión de bosques tropicales a sistemas agrícolas ha conllevado a la pérdida de hábitat que
usan las aves para el refugio, alimentación y forrajeo. Muchas aves son tolerantes a los cambios en
el hábitat, pero es importate examinar cómo responde una comunidad frente a los entornos
agrícolas y los ecosistemas remanentes. Para conocer la selección diferencial de los recursos que
usa la comunidad de aves, y determinar si estos cambian con la altura de la plantación se hicieron
censos dentro de 18 lotes de palmas grandes, medianas y pequeñas dentro de los cuales se
identificaron como categorías de recursos tronco (T) y hojas (H), y estos se relacionaron con las
actividades de las aves (descanso en percha, canto, forraleo y alimentación). A partir de los
gremios de alimentación se hicieron medidas de selectividad de recursos y amplitud de nicho para
cuantificar la selección de los recursos y el grado de especialización de las aves. Los resultados
demostraron que la mayoría de las aves usaron la plantación para descanso en percha,
indistintamente de la altura de la palma, con una baja frecuencia las aves usaron la plantación para
la alimentación y en ninguna ocasión para la reproducción. Los gremios de alimentación no
mostraron una medida significativa en la especializaciòn del uso de los recursos del cultivo y la
selectividad por troncos y hoja fue negativa o azarosa en todas las clases de altura.
La interpretación de los resultados nos indica que el cultivo de palma de aceite no fue una
cobertura importante para las aves en cuanto a selectividad y uso de recursos, los cambios en la
altura no representaron un beneficio significativo en cuanto a actividades y uso de recursos y
posiblemente este tipo de sistema podría reducir oportunidades para el establecimiento de
poblaciones de especies propias del piedemonte.

53
Introducción
Las aves están presentes en varios tipos de hábitats y juegan un papel importante en la
estructura y función de los ecosistemas (Tanalgo, 2015), este grupo selecciona sitios al
cual estén bien adaptadas en términos de explotación de recursos (Hilden 1965; Alatalo,
1981).
Los recursos en candidades adecuadas son necesarios para mantener las poblaciones de
animales (Rosenzweig, 1981; Manly et al., 2004). Un recurso se caracteriza por ser usado,
aunque no necesariamente consumido, esto abarca el alimento (identidades taxonómicas
de consumo de un organismo); el espacio también puede ser un recurso para expresar una
conducta territorial o social, también son considerados recursos los componentes del
hábitat como sitios para anidar, sitios de perchas o sitios de refugio contra depredadores
(Wiens 1989, Krebs 1999).
La selección diferencial de los recursos es un de los principios que explica la coexistencia
de las especies (Rosenzweig, 1981; Manly et al., 2004), esta es una conducta que obedece
a la selección de elementos particulares (por ejemplo ítems de alimento) dentro de las
características generales del hábitat (sitios de alimentación), y se define como la cantidad
de recurso que es utilizado por un animal (o poblaciones de animales) en un perido de
tiempo establecido y la manera más comúnmente empleada de cuantificar el uso de
recursos es calcular el porcentaje del recurso usado por el porcentaje del recurso
disponible (Manly et al., 2004). Para las aves de hábitat terrestres el alimento es
considerado el recurso más importante y las medidas de forrajeo usualmente se emplean
para indicar cómo, dónde y qué tipo de alimento obtienen (Holmes y Recher, 1986;
Dhindsa y Saini, 1994).
Las aves que habitan los sistemas agrícolas pueden permanecer como poblaciones
interactuantes en cuanto ofrezcan recursos adecuados de hábitat para las especies,
disponibilidad y acceso al alimento, así como sitios de refugio (Johnson et al., 2011,
Cifuentes-Sarmiento y Cantillo Cortés 2016; Bayly y Gómez, 2010), sin embargo la

54
carencia de estas características en los sistemas agrícolas a gran escala propicia un entorno
inhóspito para muchas especies de aves, especialmente para aquellas que dependen de
características estructurales específicas de la vegetación (Cody, 1981; Ramachandran y
Ganesh, 2012).
La abundancia, variedad de recursos y el grado en el que esto son compartidos, son
factores que influyen en la diversidad, distribución y abundancia de especies y el
desarrollo de una comunidad (Block y Bernnan 1993, MacArthur 1972, Finch 1989), pero
ante los acelerados procesos de transformación causados por la agricultura a gran escala es
importante investigar qué aspectos de las comunidades cambian. En los cultivos de palma
de aceite se ha documentado que las palmas maduras pueden soportar mayor diversidad de
aves porque tienen mayor heterogeneidad estructural que las palmas jóvenes (Azhar et al.,
2015), y tal heterogeneidad se entiende como recursos potenciales para las aves. El
objetivo de este estudio fue conocer la selección diferencial de recursos como: tipo de
alimento, sitios para percha, forrajeo y alimentación que usa la comunidad de aves de un
cultivo de palma de aceite y determinar si estos cambian con la altura del cultivo.
Métodos
Área de Estudio
El estudio se hizo en dos fincas con cultivos de palma de aceite llamadas Tierra Viva y
Mejorana, las cuales se encuentran en los municipios de San Martín (03°37'51.8" N,
073°20'59.7" W) de los Llanos y Acacias (03°57'37.18"N, 73°37'32.93W), departamento
del Meta. Los predios tienen un área de 1400 y 1480 ha respectivamente, dentro de los
cuales 600 ha son lotes de palma de diferente grado de crecimiento, el terreno también
incluye remanentes de vegetación nativa de crecimiento secundario y bosques de galería
asociados a cursos de agua de cada sector: caño Barbascal y río Acacias.

55
Censos de aves
Las aves se registraron censos mediante transectos lineales en 18 lotes de palmas: seis de
palmas grandes (11 a 15 m), seis de palmas medianas (7 a 10 m) y seis de palmas
pequeñas (2 a 6 m). Cada transecto tuvo una distancia de 500 m, fueron de ancho variable
(Buckland et al., 1993) y mantuvieron una distancia mínima de 800 m para garantizar la
independencia de los datos (Bibby et al., 2000). Cada uno de los 18 transectos se recorrió
seis veces entre junio, agosto y noviembre de 2013, y en febrero y julio de 2014,
totalizando 108 recorridos durante el periodo de estudio. Para efectos del análsisi de datos
se tuvieron en cuenta los recorridos donde hubo registro de aves (81 recorridos).
Las aves observadas fueron asignadas a una de las siguientes categorías de grupo trófico:
carnívoro (CAR), carnívoro correñoero (CAR-CRR), carroñero (CRR), frugívoro (FRU),
granívoro (GRA), insectívoro (INS), nectarívoro (NEC) y omnívoro (OMN), basado tanto
tanto en las observaciones de campo como en la literatura disponible (Hilty & Brown
2001; Restall et al., 2006).
Análisis de los datos
El punto de partida de este análisis fue identificar categorías de recursos y posteriormente
cuantificar en qué proporción las aves usaron las categorías de recursos disponibles dentro
de la plantación, todo esto analizado a partir de los gremios de alimentación.
Teniendo en cuenta la definición de recurso descrita por Wiens (1989), quien expresa que
un recurso puede ser identidades taxonómicas de consumo de un organismo, el espacio o
componentes del hábitat, a partir de las observaciones se identificaron como categorías de
recursos aquellos sustratos o elementos del cultivo que usaron las aves y que se puedieron
cuantificar por unidad de área. En este sentido las categorías de recursos fueron tronco (T),
y hojas (H) y las actividades que realizaron sobre estas categorías de recursos fueron
alimentación, forrajeo, canto y descanso en percha.

56
Se calculó el índice de selectividad de Ivlev (Ivlev, 1961) dado por la fórmula:
Donde corresponde a la proporción de la muestra del recurso usado de la
categoría i. Y corresponde a la proporción de recurso disponible de la
categoría i.
Este es uno de los índices de selección de recursos más usados (Manly et al., 2002) y por
lo tanto más comparable. Los valores del índice varían de -1 (máxima selección negativa)
a + 1(máxima selección positiva) (Atienza, 1994) y se basa en la relación de recursos
usados y la proporción de recursos disponibles de cada categoría analizada (Manly et al.,
2002).
Los recursos disponibles fueron tronco (T) y hojas (H), la disponibilidad de estos
elementos se obtuvo a partir del número de palmas presentes en el área de los lotes con
transectos. El diseño del cultivo conocido como tresbolillo, permite obtener una densidad
de 143 palmas/ha (Raygada-Zambrano, 2005). Durante el estudio hubo regisros de aves
alimentándose del fruto de la palma, pero esta categoría de recurso no se tuvo en cuenta
porque los racimos siempre están sujetos a las labores de cosecha. El índice se calculó con
los gremios de alimentación para usarlos como indicadores del tipo de recursos que las
aves pueden obtener dentro del sistema.
Para probar si las actividades de las aves de cada grupo trófico mostraban una relación
significativa con los recursos del cultivo se hizo una prueba Friedman usando las
frecuencias de las actividades en cada clase de altura de la palma.
Un análisis de ampliud de nicho (Levins 1968) se hizo para cuantificar si hubo algún grado
de especialización en cuanto al uso de los recursos. Aunque el concepto de nicho es
especie específico, el índice también se ha aplicado a estudio de comunidades para
observar la distribución de los organismos dentro de los estados de un recurso dentro de un
hábitat (Krebs, 1999; Colwell y Futuyma, 1971)
Levins (1968) propuso que la amplitud de nicho se puede estimar midiendo la uniformidad
de la distribución de individuos entre los estados de recursos (Krebs 1999), y está dado por
la expresión:

57
Donde B= Amplitu de nicho propuesto por Levins
pj= proporción de individuos que usaron el recurso j
El índice fue estandarizado por Hulbert (1979) para expresar el resultado en una escala de
0 a 1, donde 0 representa el grado de máxima especialización (Krebs 1999).
Con el fin de identificar si las especies se relacionaron con características estructurales del
cultivo o de acuerdo a los recursos disponibles, se hizo un análisis de componentes
principales teniendo en cuenta siete variables del cultivo en cada clase de altura y cinco
variables de la proporción de uso de cada categoría de sustrato (Tabla 2.1).
Las variables del cultivo se obtuvieron a partir de datos tomados en campo e información
secundaria (Corley y Tinker, 2003, Ratanawil et al., 2006). Aunque la matriz contenía
variables con diferentes unidades de medida, los datos no se transformaron para evitar
tendencias erróneas en la estructura de la correlación y sesgos en los resultados (Koròan y
Adamík, 2007; Jackson 1997).
Tabla 2 1. Variables del cultivo y proporción de uso del cultivo de palma de aceite
Variable Grande Mediano Pequeño
Diámetro de tronco (m) ** 0.5 0.6 0.7
Altura promedio de palma (m) 14 12 6
Número de Palmas 186 171 186
Área (ha) 1.3 1.2 1.3
Altura tronco (m) ͌* * 10 4 1.3
Peso racimo (Kg)* 25 15 5
Diámetro dosel (m) 17 12 6
Proporción uso suelo 0.3142857 0.1551724 0.1393443
Proporción uso tronco 0.1 0 0
Proporción uso hojas 0.4714286 0.7413793 0.7786885
Proporción uso racimo 0.0285714 0.0344828 0.0081967
Proporción uso Otros Elementos 0.0857143 0.0689655 0.0737705 * Corley & Tinker (2003), * * Ratanawil et al., 2006, ͌ Luskin & Potts, 2011
Sin asterisco: datos tomados en campo
𝐵 =1
𝑝𝑗2

58
Resultados
La actividad más frecuente de las aves dentro del cultivo de palma fue descanso en percha
(n= 200), sin importar la clase de altura y sin encontrar diferencias significativas de las
actividades realizadas entre clase de altura de las palmas (Friedman Chi2= 6.12, > 0.05),
mientras que la alimentación fue la actividad menos frecuente (n= 45) (Figura 2.1).
A partir de los cálculos de selectividad de recursos se pudo probar que la palma es un
sistema de pobremente seleccionado por las aves. En general los gremios de alimentación
presentaron una selección negativa (valores negativos) o azarosa (valores cercanos a 0) por
seleccionar las categorías de recurso H y T, indistintamente de la clase de altura de la
plantación (Figura 2.3). En la palma grande se detectó que el grupo INS tuvo selección
positiva (iE=0.82) por los troncos, sin embargo, este valor pudo estar sesgado porque hubo
una relación más proporcional entre las unidades de recurso usado y el número total de
unidades de recurso usado (Oi=14/16).
La comunidad tuvo un bajo grado de especialización de las actividades que hicieron dentro
del cultivo (B=0.68) y uso de recursos (B=0.55). Valores intermedios en la escala de
medida de 0 a 1 refeja que las aves no tuvieron un grado de especialización significativo
por usar los recursos de la plantación.
En el análisis de componentes principales (Figua 2.2) identificamos que de izquierda a
derecha se ordenaron las variables de menor a mayor correlación con la abundancia
absoluta y proporcional de individuos. El componente 1 (valor de eje= 7617.56, 98% de
varianza) proyectó la variación de acuerdo al tamaño de la palma y el componente 2 (valor
de eje= 153.22., 1.9% de varianza) de acuerdo a las variables del cultivo y la proporción de
uso del mismo, así palmas grandes y medianas quedaron situadas del lado negativo y
representando una correlación negativa con la abundancia de individuos, las variables del
cultivo tuvieron mayor relación con la riqueza de especies.

59
Las variables de uso proporcional del cultivo se proyectaron cerca del origen e indican que
no hubo una correlación fuerte con los componentes 1 y 2 que resumen la varianza de los
datos.
Discusión
Las aves utilizaron el cultivo de palma principalmente para percha de descanso y los
cambios en la altura no influyeron de manera significativa en las actividades y el uso de
recursos. La alimentación fue una actividad de baja frecuencia (5 a 25%) y este sistema
parece ser una cobertura pobre para proveer sitios de reproducción para las aves. Este
panorama no siempre es igual, pues existe evidencia de otros tipos de cultivos que ofrecen
mayor grado de recursos y asociación a las aves. Por ejemplo, en los cultivos de café con
sombra se ha observado que las plantas de café proveen alimento como insectos e
invertebrados que se posan en las hojas, el fruto y las flores también son recursos
atractivos para diferentes especies (Wunderle & Latta, 1998). Las plantas epífitas que
crecen sobre los árboles que conforman el sotobosque también son un elemento que
favorece a las aves (Cruz-Angón y Greenberg, 2005). Los cultivos de arroz son
reconocidos por la convención RAMSAR porque se comportan como un humedal
temporal que cubre cerca de 193.000ha en la región de la Orinoquía (Cifuentes- Sarmiento
y Castillo-Cortés, 2016). En localidades del Valle del Cauca En ellos se han identificado
más de 150 especies de aves que en algún momento lo usan como sitio de descanso,
alimentación o de paso (Cifuentes- Sarmiento y Castillo-Cortés, 2016). En los cultivos de
arroz del Ornico también se ha documentado que estos entornos agrícolas son sitios
importantes para especies acuáticas, migratorias, congregatorias (Ocampo-Peñuela, 2010).
En relación a los aspectos reproductivos, también se ha encontrado que diversas especies
de aves usan campos agrícolas durante la época de reproducción, incluso, algunas especies
se han especializado para anidar y forrajear en estos entornos (Hiron, 2013). En nuestro
estudio esta actidad fue poco frecuente, considerando que fuera de periodos de censo se
observó el apareamiento de una pareja de Caracara plancus y hubo registros anecdóticos
de nidos de Dendrosygna viduata, Amazona amazonica y Milvago chimachima, en estos

60
casos los nidos se vieron afectados (destruidos) por la cosecha de recimos de la palma.
Muchas especies pueden tener limitaciones para encontrar sitios de reproducción frente a
los cambios de vegetación (Hilden 1965) e inclusio las crías son más vulnerables ante los
depredadores por efecto de borde (Andren, 1994), o por las actividades humanas como en
este caso. La reducción de recursos para las aves por la transformación de la vegetación a
tierras agrícolas se ha documentado ampliamente (Wilson et al., 2005, Aratrakorn et al.,
2006; Danielsen et al., 2008), coincidiendo en que la uniformidad del sistema a gran escala
reduce notoriamente la oferta de recursos como el alimento y los sitios de anidación.
El análisis de selectividad de recursos mostró que las categorías de recursos tronco (T) y
hojas (H) fueron de escasamente seleccionados para los miembros de la comunidad. En la
actualidad hay mucha desinformación sobre estudios que calculen la selectividad de
recursos para las aves en ambientes agrícolas, sin embargo, con base en nuestros
resultados se puede inferir que el tronco y las hojas de las palmas, aunque se comportan
como un sustrato de forrajeo, especialmente para aves insectívoras y carnívoras, esta oferta
es pobremente explotada por los miembros de la comunidad (13 especies insectívoras, 2
carnívoras), indistintamente de la clase de altura.
La diversidad de especies aumenta cuando los recursos son heterogéneos y ampliamente
disponibles (Finch, 1989), nuestros resultados demostraron que la homogenidad estructural
del cultivo de palma de aceite hace que sea un sistema de pobremente seleccionado por las
aves y pobre para ofrecer sitios de reproducción y alimento a la mayoría de los miembros
de la comunidad, sin embargo puede ser más favorable para aves insectívoras de amplia
distribución que de manera frecuente forrajearon dentro de la plantación y para aves
carívoras que perchan en las hojas de las plamas para cazar pequeños vertebrados
(Sánchez, 2000). A pesar de la extención, este sistema no brinda suficiente variedad de
recursos que sean ampliamente seleccionados por los gremios de alimentación de la
comunidad de aves y tampoco se evidenció una relación positva de las actividades que
realizan dentro del cultivo con la clase de altura.

61
Figura 2. 1 Frecuencias de actividades realizadas por las aves en el cultivo de palma de
aceite. P: descanso en percha, C: canto, F: forrajeo, A: alimentación
Figura 2. 2 Proyección de variables del cultivo y uso proporcional de recursos, en el cultivo
en palma de aceite
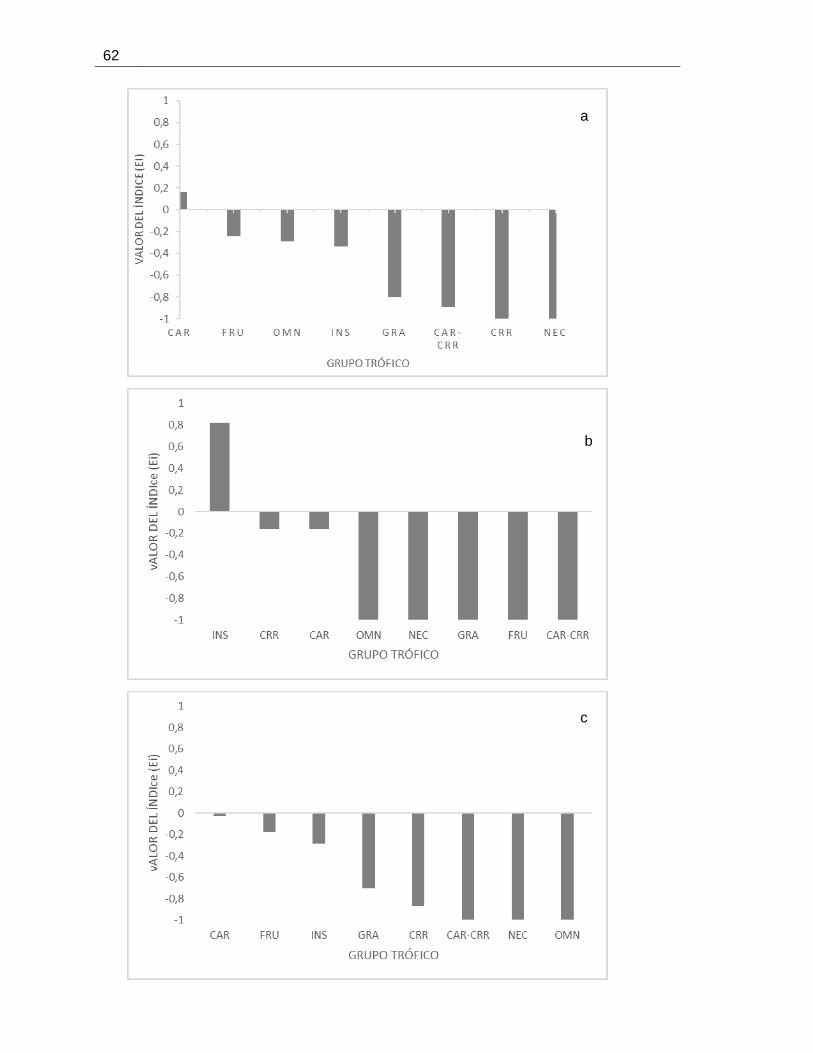
62
a
b
c

63
Figura 2.3 Índice de selectividad de recursos para gremios de alimentación de aves en el
cultivo de palma de aceite. a) hojas de palma grande, b) troncos de palma grande, c) hojas de
palma mediana, d) hojas de palma pequeña.
d

64
Referencias
Alatalo, R. V. 1981. Habitat selection of forest birds in the seasonal environment of
Finland. Annales Zoologici Fennici, 18: 103-114.
Andren, H. 1994. Effects of habitat fragmentation on birds and mammals in landscapes
with different proportions of suitable habitat: a review. Oikos 71: 355-366.
Aratrakorn, S., Thunhikorn, S., & Donald, P. F. (2006). Changes in bird communities
following conversion of lowland forest to oil palm and rubber plantations in
southern Thailand. Bird Conservation International, 16, 71-82.
Atienza, J. C. 1994. La utilización de índices en el estudio de la selección de recursos.
Ardeola, 41(2): 173-175.
Azhar, B., N. Saadun, Ch. Leong Puana, N. Kamarudin, N. Aziz, S. Nurhidayu & J.
Fischer. 2015. Promoting landscape heterogeneity to improve the biodiversity
benefits of certified palm oil production: Evidence from Peninsular Malaysia.
Global Ecology and Conservation, 3: 553-561.
Bayly, N. J. & Gómez, C. 2010. Las aves de la Hacienda La Victoria. SELVA:
Investigación para la conservación en el Neotrópico, Bogotá. Informe técnico No.
CEC01.
Bibby C. J., M. Jones & S. Marsden. 2000. Expedition field techniques birds survey. 2nd
Edition Academic Press. London, UK.
Block, W. M. & L. A. Bernnan. 1993. The habitat concept in ornithology Theory and
applications. En: Current Ornithology, Vol. 11 Dennis M. Power (Ed.). Plenum
Press, New York.
Buckland, S.T., Anderson, D.R., Burnham, K.P. & Laake, J.L. 1993. Distance Sampling:
Estimating Abundance of Biological Populations. Chapman and Hall, London.
446pp Cañas Ch., H. Garrido y J. Calderón. 2004. Codificación de marcas de
lectura a distancia. Anuari Ornitologic De Les Balears A.O.B, 19: 51–60.
Colwel, R. K. & D. J. Futuyma. 1971. On the measurment of niche and overlap. Ecology,
52(4): 567-576.
Corley, R. H. & P. B. Tinker. 2003. The palm oil. 4th Edition. Oxford Blackwell Science.
562pp.
Cruz-Angón, A. & R. Greenberg. 2005. Are epiphytes important for birds in coffee
plantations? An experimental assessment. Journal of Applied Ecology, 42, 150-
159.

65
Danielsen, F., H. Beukema, N. D. Burgess, F. Parish, C. A. Brühl, P. F. Donald, D.
Murdiyarso, B. Phalan, L. Reijnders, M. Struebig & E. B. Fitzherbert. 2008.
Biofuel Plantations on Forested Lands: Double Jeopardy for Biodiversity and
Climate. Conservation Biology. 23(2): 348-358.
Dirzo, R. & M. C. García. 1992. Rates of deforestation in Los Tuxtlas, a Neotropical area
in southeast Mexico. Conserv. Biol. 6: 84-90.
Dhindsa, M. S & H. K SAINI. 1994. Agricultural ornithology: An Indian perspective.
Journal of Bioscience, 9(4): 391-402.
Cifuentes-Sarmiento, Y. & L. F. Cantillo Cortés. 2016. Las alas del arroz: aves acuáticas
en cultivos ecoamigables del Valle del Cauca. Asociación Calidris & Arrocera La
Esmeralda S.A.S. Cali. Colombia.
Cody, M. L. 1981. Habitat selection in birds: The roles of vegetation structure,
competidors and productivity. BioScience, 31(2): 107-113.
Finch, D. 1989. Habitat use and habitat overlap of riparian birds in three elevational zones.
Ecology, 70(4): 866-880.
Hilden, O. 1965. Habitat selection in birds: A review. Annales Zoologici Fennici, 2(1): 53-
75.
Hiron, M. 2013. From Fields to Landscapes: Effects of Agricultural Land Use and
Landscape Heterogeneity on Farmland Birds. Doctoral Thesis. Swedish
University of Agricultural Sciences
Holmes, R. T. & H. F Recher. 1986. Determinants of guild structure in forest bird
communities: an intercontinental comparison. The Condor, 88: 427-439.
Hulbert, S. H. 1978. The measurement of niche overlap and some relatives. Ecology, 59:
67-77.
Ivlev, V.S. 1961. Experimental ecology of the feeding of fishes. Yale University Press.
Johnson, R. J., J. A. Jedlicka, J. E. Quinn & J. R. Brandle. 2011. Global perspective on
Birds in Agricultural Landscapes. En: Campbell, W. B. y S. L. Ortíz (Eds.).
Integrating agriculture, conservation and ecotourism: examples from the field.
Vol. 1. Springer-Verlag Berlin. Alemania
Krebs, Ch. J. 1999. Ecologcical Methodology. 2nd Edition. Addison Wesley
Longman.581p.

66
Koròan, M. & P. Adamík. 2007. Foraging guild structure within a primaeval mixed forest
bird assemblage: a comparison of two concepts. Community Ecology, 8(2): 133-
149.
Levins, R. 1968. Evolution in changing environment: Some Theoretical Explorations.
Princeton University Press, Princeton, New Jersey.
Luskin, M. S. y M. D. Potts. 2011. Microclimate and habitat heterogeneity through the oil
palm lifecycle. Basic and Applied Ecology, 12: 540–551.
MacArthur 1972. Geographical ecology. Patterns in the distribution of species. Harper and
Row, New York, New York, USA.
Manly, B. F. J., L. L. McDonald, D. L. Thomas. T. L. McDonald & W. P. Erickson. 2002.
Resource selection by animals 2nd Edition. Kluwer Academic Publisher.
Netherlands.
Ocampo-Peñuela, N. 2010. El fenómeno de la migración en aves: una Mirada desde la
Orinoquía. Orinoquía, 14(2): 188-200.
Pianka, E. R. 1974. Niche overlap and diffuse competition. Proc. Nat. Acad. Sci, 71(5):
2141-2145.
Ramachandran, V. & T. Ganesh. 2012. Habitat structure and its efects on birds
assemblages in the Kalakad-Mundanthurai Tiger Reserve (KMTR), India. Jornal
of the Bombay Natural History Society, 109(1&2): 87-95.
Ratanawil, T., T. Chumthong & S. Kirdkong. 2006. Investigación sobre las propiedades
mecánicas de los troncos de palma de aceite para la industria del mueble. Journal
of the Oil Plam Research, Special Issue, 114-121.
Raygada-Zambrano, R. 2005. Manual técnico para el cultivo de la palma aceite. Comisión
Nacional para el Desarrollo y Vida sin Drogas – DEVIDA, Proyecto de
Desarrollo Alternativo Tocache-Uchiza – PRODATU. Perú. 109 pp.
Rosenzweig, M. L. (1981). A theory of habitat selection. Ecology 62: 327-335.
Sánchez, S. 2000. Vertegrados silvestres registrados en una parcela de palma aceitera en
Tabasco, México. ASD. Oil Palm Papers, No. 20, 17-18.
Tanalgo, K. C., J. A. Fuentes Pineda, M. E. Agravante & Z. Mamalo Amerol. 2015. Bird
Diversity and Structure in Different Land-use Types in Lowland SouthCentral
Mindanao, Philippines. Tropical Life Sciences Research, 26(2), 85–103.
Wiens, J. A. 1989. The ecology of bird comminities. Vol1. Foundations and patterns.
Cambirfge University Press. NY. 539p.

67
Wilson, J.D., M. J. Whittingham & R. B. Bradbury. 2005. The management of crop
structure: a general approach to reversing the impacts of agricultural
intensification on birds? Ibis, 147: 453-463.
Wunderle, J. M. & S. C. Latta. 1998. Avian resource in Dominican shade coffe
plantations. Wilson Bulletin, 110(2): 271-281.

68
CAPÍTULO 3. CARACTERIZACIÓN DE GREMIOS Y RASGOS FUNCIONALES
DE AVES EN PAISAJES DOMINADOS POR CULTIVOS DE PALMA DE
ACEITE (Elaeis guineensis)
Diana Patricia Tamaris-Turizo y Hugo Fernando López-Arévalo

69
Resumen
La conversión de boque a cultivo de palma genera cambios en el tipo de vegetación y la
escasa complejidad estructural del cultivo reduce la diversidad de especies y genera
cambios en la estrucutra de los gremios de alimentación. Para comparar la estructura
trófica y rasgos funcionales de aves entre el bosque y lotes con cultivo de palma de tres
alturas (grandes, medianas y pequeñas), así como para explicar los cambios de la
comunidad generados por la transformación de la vegetación se hicieron capturas de aves
con redes de niebla. Estas se instalaron dentro de seis lotes de palma y de bosques
adyacentes a dichos lotes. El registro de aves incluye 271 individuos pertenecientes a 88
especies, 74 de las cuales se capturaron dentro del bosque y 33 dentro de la palma. La
composición de los gremios de alimentación fue muy similar entre el bosque y el cultivo
de palma, pero la composición de especies, riqueza y abundancia varió entre los sistemas
analizados. Aves piscívoras fueron exclusivas en el bosque y carnívoras fueron exclusivas
dentro del monocultivo. Aves innsectívoras fueron las más representativas en ambas
coberturas (44% y 20% en bosque y palma respectivamente). La abundancia de aves en el
bosque fue más proporcional con el ensamble registrado en palma pequeña. Las aves
asociadas al cultivo de palma tuvieron el pico más grande que aquellas asociadas al
bosque. Los resultados permitieron concluir que el cultivo de palma de aceite indujo a
cambios en la comunidad de aves y estos cambios se reflejaron en la composición, riqueza
y abundancia de gremios de alimentación y esto podría tener consecuencias en las
interacciones ecológicas y el papel funcional que cumplen las especies dentro de los
ecosistemas.

70
Introducción
El concepto de gremio representa una de las maneras como se puede analizar la
organización de una comunidad (Tokeshi, 2009). El término fue definido como un grupo
de especies que explotan la misma clase de recursos ambientales de una manera similar
(Root 1967) y se enfoca en la competencia de las especies simpátricas,
independientemente de la relación taxonómica (Simberloff y Dayan, 1991, Hawkins y
MacMahon, 1989).
La identificación y análisis de gremios ecológicos ha sido fundamental para entender los
procesos que determinan la diversidad funcioal, estructura y organización de las
comunidades (González-Salazar et al., 2014). Los miembros de cada gremio juegan un
papel importante dentro de los ecosistemas y se puede usar una clasificación funcional
para analizar el flujo de energía y biomasa dentro de las redes alimenticias (Hawkins y
MacMahon 1989). Las especies asociadas a cada grupo realizan procesos claves dentro de
este tipo de sistemas: dispersión de semillas (frugívoras), consumo de semillas
(granívoras), controlan la herbivoría y regulan la abundancia de los insectos (insectívoras),
facilitan la polinización (nectarívoras) (Pin Koh 2008, Karp et al. 2011), por ello es
importante entender los patrones de explotación de alimento, conocer la estructura trófica,
especialmente presencia o ausencia de gremios específicos.
La expansión de la frontera agrícola y la sustitución ecosistemas naturales a gran escala
reduce la riqueza de especies y la diversidad funcional, trae efectos en la composición y
estructura de gremios, la calidad y disponibilidad del hábitat y de los recursos del cual
dependen las aves (Azhar et al. 2013, Ramírez-Albores, 2010; Dirzo y García, 1992), se ve
afectado el papel ecológico que realizan las aves dentro de los ecosistemas con
implicaciones importantes en la provisión de servicios ecosistémicos (Prescott et al. 2016).
El efecto puede ser más severo con el establecimiento de cultivos perennes (Fitzherbert et.
al., 2006) puesto que muchas especies reducen sus poblaciones por las drásticas
modificaciones del entorno y por la reducción en la disponibilidad, de recursos (Beissinger
y Osborne 1982).

71
En los monocultivos de palma de aceite se ha encontrado que las aves insectívoras son las
más dominanestes dentro del sistema (Azhar et al., 2013, Achondo et al., 2011) estas
actúan como control de insectos y la presenca de este y otros gremios depende de la
cercanía que conserve el cultivo con relictos de bosque (Azhar et al. 2013). Por otro lado
también se ha documentado que la diversidad funcional se reduce con el paso de
remanentes de bosque a un cultivo comercial de palma de aceite (Prescott at al. 2016).
Conocer la diversidad biológica de un ecosistema es clave para entender los mecanismos
de ensamblaje de las comunidades (Salgado Negret y Paz, 2015). En este estudio se quizo
probar como hipótesis: 1) que la conversión de bosque a cultivo de palma de aceite reduce
la diversidad de los gremios de alimentación. 2) los cambios de altura de la palma de
aceite no inducen a cambios significativos en la estrutura y composición de los gremios de
alimentación dentro de la plantación y 3) la diversidad funcional de las aves se reduce
cuando el bosque pasa a ser un monocultivo de palma de aceite. A partir de estas
consideraciones se planteó como objetivo comparar la estructura de gremios de
alimentación y los rasgos funcionales entre aves de bosques y cultivos de palma de aceite.

72
Métodos
Área de estudio
El estudio se hizo en dos fincas del Oriente de Colombia, con cultivos de palma de aceite
llamadas Tierra Vivia y Mejorana, ubicadas en los municipios San Martín de los Llanos
(03°37'51.8" N, 073°20'59.7" W) y Acacias (03°57'37.18" N, 73°37'32.93” W),
respectivamente, departamento del Meta. Cada predio cuenta con un área de
aproximadamente 1.400 y 1480 ha.
El área geográfica de ambas localidades hace parte del piedemonte cordillerano, en la que
se destacan bosques, matorrales, pastizales y áreas con actividades agrícola y ganadera
(Rangel-Ch y Minorta-Cely, 2014; Pinzón Pinzón et al., 2009, Mora Fernández et al.,
2011; Rippstein et al., 2001), lo cual ha generado cambios estructurales en la vegetación,
procesos de fragmentación y alteraciones microclimàticas de los fragmentos (González-
Borrero, 2013). El régimen biestacional hace que la temperatura sea elevada en el los
primeros y últimos meses del año, mietras que a mitad de año se registran las temperaturas
más bajas (Minorta-Cely y Rangel-Ch, 2014). Por su parte, las precipitacines tienen un
margen de variación anual de 2877 a 4000 mm (Minorta-Cely y Rangel-Ch, 2014).
Censos de aves
Para conocer y comparar la estructura trófica de las aves asociadas al cultivo de palma de
aceite y fragmentos de bosque se hicieron censos con redes de niebla. Las redes se
ubicaron en el interior de seis lotes de palmas grandes, mediandas y pequeñas (dos lotes
por cada clase de altura), así como en el interior de seis fragmentos de bosque, los cuales
estuvieron cercanos a lo lotes de palma: distancia entre los bordes más cercanos entre
dichas coberturas no mayor a 1km.

73
En cada unidad de muestreo se instalaron de 7 a 10 redes (12× 2.5 m y ojo de malla de 36
mm) que se abrieron de 6:00 a 10:00 am y de 3:30 a 5:30 pm dedicando 360 horas/red
durante el estudio. Las aves capturadas fueron identificadas (especie, sexo, edad), medidas,
pesadas y liberadas después de tomar registro fotográfico.
Las especies capturadas en palma y bosque fueron asignadas a uno de los siguientes
gremios de alimentación teniendo en cuenta la principal fuente de alimento de cada
especie: 1) carnívoro, 2) frugívoro, 3) frugívoro-insectívoro, 4) granívoro, 5) granivoro-
insectívoro 6) insectívoro, 7) nectarívoro y 8) piscívoro. La asignación de cada grupo se
basó en observaciones directas y revisión de literatura especializada (Restall et al. 2006,
Universidad Icesi 2014).
El análisis de gremios de alimentación se hizo a partir de los individuos capturados. Para
comparar la estructura de los gremios de alimentación entre el bosque y el cultivo de
palma de aceite se hizo un análisis de asociación en modo Q y R con el índice de
similaridad Bray Curtis. A partir de este análisis se obtiene un dendrograma en dos
dimensiones en el cual se organizan los grupos según la disimilaridad en el eje de las
ordenadas (Ramírez González, 2005).
Se emplearon cinco medidas de rasgos funcionales para identificar la relación del tamaño
de las aves con relación al tipo de vegetación. Los rasgos que se utilizaron fueron talla,
peso, longitud del culmen total (LCT), alto del pico (AP), y ancho del pico a la altura de
las narinas (AnP). Estos se tuvieron en cuenta porque están asociados con la locomoción,
movimientos locales, uso de recursos y la capacidad de responder ante cambios en el
ambiete, además la forma y tamaño del pico se relaciona con el tipo de alimento y con el
papel ecológico dentro del ecosistema (López-Ordoñez et al., 2015). Esta comparación se
complementó con una prueba Kruskal Wallis para identificar diferencias significativas
entre las variables (rasgos funcionales de cada gremio de alimentación) y los factores
(bosque y palma).

74
Resultados
Se registraron 271 individuos pertenecientes a 88 especies. En el bosque se registraron 74
especies (84%) y 56 fueron exclusivas. En la plantación se registraron 33 especies (37.5%)
de las cuales 15 fueron capturas únicas de este agrosistema. Las aves registradas dentro del
cultivo de palma fueron residentes, en el bosque se encontraron 5 especies migratorias:
(Elaenia parvirostris, Contopus virens Parkesia novevoracensis, Sporophila lineola y
Setophaga ruticilla), otras migrantes también se observaron fuera de los censos en el área
de estudio (Piranga rubra, Tringa solitaria y Catharus ustulatus).
Identificamos siete gremios de alimentación asociados a ambos tipos de cobertura, las aves
piscívoras y carnívoras fueron exclusivas de bosque y cultivo de palma, respectivamente,
mientras que las aves insectívoras fueron predominantes en ambas coberturas, no obstante,
la proporción de riqueza y abundancia de este últimogrupo fue mayor en el bosque. (Tabla
3.1).
Al comparar la similitud de los gremios de alimentación (descriptores) con el bosque y la
palma (objeto) se observó una segregación estructurada por la abundancia de individuos
(coef. corr. 0.8335). El agrupamiento entre FRU y GRA tuvo la mayor medida de similitud
(90%), NEC e INS también tuvieron una similaridad importante (60%). Estas
agrupaciones se basaron en los promedios de la abundancia proporcional de cada grupo
trófico con respecto a los sitios analizados (Fig. 3.1A).
Al analizar la relación de los sitios (objeto) con respecto a los gremios de alimentación
(descriptores) se detectó que el bosque y la palma pequeña se agruparon por con mayor
grado de similitud (80%), mientras que la palma mediana fue diferente de los demás sitios
por la escasa asociación de individuos asociados a lodentrode cada grupo trófico (coef.
cor. 0.99002), (Fig 3.1B). El análisis entonces demostró que el empaquetamiento de los
grupos se formó por el grado de similitud de la abundancia proporcional de individuos
entre los gremios de alimentación y entre las clases de altura del cultivo de palma.

75
Al comparar el peso entre tipos de coberura se observó que esta medida tuvo mayor
dispersión de los datos en aves asociadas a la palma por la captura de aves pequeñas y
grandes como Amazilia fimbriata y Falco femoralis, que fueron la especies más livianas y
pesadas capturada durante el estudio (5 y 490 gm) (Figura 3.2).
La comparación de rasgos funcionales (Talla, Peso, LCT, AP y AnP) mostró una clara
tendencia: aves de tamaño y picos más grandes estuvieron asociadas al cultivo de palma y
aquellas de tamaño y picos más pequeños fueron frecuentes en el bosque (Figuras 3.2 y
3.3), pero dicha diferencia no fue significativa (KW,> α= 0.05).
Discusión
Nuestros resultados mostraron que los gremios de alimentación presentes entre el bosque y
el cultivo de palma de aceite fueron similares, pero las diferencias estén determinadas por
la composición de cada grupo entre las coberturas. La palma pequeña tuvo mayor
representatividad de individuos y el análisis de rasgos funcionales demostró que la
plantación alberga aves de mayor altura que la palma.
La estructura trófica de la comunidad de aves se caracterizó por la predominancia de
especies insectívoras en ambos tipos de cobertura las cuales estuvieron representadas por
48 espeies en el bosque y 13 dentro de la palma. La mayor proporción de aves insectívoras
en el bosque pudo estar relaciona a la presencia de árboles nativos (Azhar et a., 2013), o
con la complejidad estructural de la vegetación (Alatalo, 1981; Cousin & Phillips, 2008).
Estas formaciones albergan mayor diversidad y abundancia de aves en comparación con
los sistemas agrícolas, y también son fuente de especies polonizadoras que probablemente
se desplazan hacia los cultivos favoreciendo procesos de polinización (Espejo González, et
al. 20014). Otros estudios también han encontrado que este es el grupo más predominante
en otros paisajes agrícolas (Dhindsa y Saini, 1994), así como dentro de monocultivos de
palma (Azhar et al., 2013, Achondo et al., 2011; Cagod y Nuñeza, 2012), dentro de los
cuales se destaca su papel en el control de insectos y en la reducción en la tasa de
herbivoría por parte de los insectos (Johnson et al. 2011, Pin Koh 2008). Sin embargo se
argumenta que las aves insectívoras de bosque son más especializadas que en los entornos

76
agrícolas porque forrajean sobre troncos de sotobosque (Toledo-Lima et al., 2014),
característica estuvo ausente en nuestro sitio de estudio.
Otra diferencia importante en la estructura gremial se basó en que las aves piscívoras
fueron exclusivas del bosque y aves carnívoras fueron exclusivas de la palma. Las especies
carnívoras dentro de laplantación fueron Falco femoralis y Rupornis magnirostris, fuera
de censos también se observó Tyto alba, Buteogallus meridionalis y Caracara palncus.
Aunque no fue un grupo abundante, la presencia se explica porque este conjunto de
especies forrajea dentro del cultivo de palma para cazar vertebrados como aves más
pequeñas, pequeños mamíferos, reptiles y anfibios (Sánchez, 2000). En el caso de las aves
piscívoras, para este estudio conformado por Chloroceryle amazona, Chloroceryle
americana y Chloroceryle inda, estas se observaron en la vegetación aledaña a ríos y
caños donde percharon para cazar peces que hacen parte de su dieta.
La similaridad en la composición de gremios de alimentación entre el bosque y el cultivo
de palma también tiene implicaciones en el papel ecológico que cumplen las aves dentro
de los ecosistemas. Las aves asociadas a sistemas agrícolas se adaptan cuando adquieren
hábitos en la explotación de los recursos y contribuyen en los procesos de polinización y
control de insectos y roedores (Dhindsa y Saini, 1994, Sekercioglu, 2006), en el bosque,
además de los anteriores, las aves también contribuyen en la arquitectura de la vegetación,
propagación de semillas y regeneración de bosques y ciclaje de nutrientes (Sekercioglu,
2006; Sekercioglu, 2012).
Los patrones morfológicos parecen estar relacionados con la partición de nicho entre los
gremios (Simberloff y Dayan, 1991), y podría existir una tendencia entre el tipo de
cobertura con el tamaño corporal y el tamaño del pico de las aves. Pero, ¿qué significa que
aves con picos más grandes estén asociadas al cultivo de palma? Básicamente esto nos
informa que el establecimiento del cultivo induce a un cambio en la composición de
especies y aquellas asociadas a la plantación posiblemente tienen mayor grado de
adaptación en términos de competencia y explotación de recursos en un ambiente
intervenido (Simberloff y Dayan, 1991).

77
A partir de los resultados obtenidos se concluye que las plantaciones comerciales de palma
de aceite a escala comercial generan un cambio en la comunidad de aves, afecta la riqueza
de especies dentro de cada gremio de alimentación, la abundancia de aves en palmas
pequeñas podría sugerir una clase de altura con mayor oportunidades de recursos para las
aves y, que la diversidad funcional y el papel ecológico que cumplen las aves dentro de los
ecosistemas podrían verse reducido o afectado en entornos con cultivos de palma de aceite
donde observamos menor grado de asociación de aves y menor variedad de rasgos
funcionales, al compararse con el bosque.
Tabla 3 1. Composición, riqueza y abundancia de individuos de cada gremio de
alimentación asociado a bosque y cultivo de palma de aceite.
S n % S n G n M n P N (G+M+P) %
CAR 0 0 0 2 1 0 1 2 3,4
FRU 6 36 17 4 1 0 8 9 15,3
FRU-INS 3 20 9 1 0 0 3 3 5,1
GRA 5 18 9 7 1 0 9 10 16,9
GRA-INS 1 6 3 1 0 0 1 1 1,7
INS 48 93 44 13 4 3 13 20 33,9
NEC 8 33 16 5 1 6 7 14 23,7
PIC 3 6 3 0 0 0 0 0 0,0
Total 74 212 100 33 8 9 42 59 100
Total PalmaBosque PalmaGrupo
Trófico

78
Figura 3. 1 Coeficiente de la distancia la Euclidiana A) de la abundancia proporcional de
individuos en cada gremio de alimentación y B) de los gremios de alimentación en relación al
bosque y las clases de altura del cultivo de palma de aceite
Figura 3. 2 Comparación de rasgos funcionales (talla-peso) en aves asociadas a bosque y
cultivo de palma
B A

79
Figura 3. 3 Comparación de rasgos funcionales (medidas del pico) en aves asociadas a bosque
y cultivo de palma. B: bosque, P: cultivo de palma

80
Referencias
Achondo, M. J. M., Casin, L. F., Bello, V. P., Tanalgo, K. C., Agduma, A. R., Bretaña, B.
L. P., Mancao, L. S., Salem, J. G. S., & Supremo, J. P. (2011). Rapid assessment
and feeding guilds of birds in selected Ruber and Oil Palm plantations in North
Cotabato. Asian Journal of Biodiversity, 2, 103-120.Alatalo, R. V. 1981. Habitat
selection of forest birds in the seasonal environment of Finland. Annales
Zoologici Fennici, 18: 103-114.
Alatalo, R. V. 1981. Habitat selection of forest birds in the seasonal environment of
Finland. Annales Zoologici Fennici, 18: 103-114.
Aratrakorn, S., Thunhikorn, S., & Donald, P. F. (2006). Changes in bird communities
following conversion of lowland forest to oil palm and rubber plantations in
southern Thailand. Bird Conservation International, 16, 71-82.Awade, M. y J. P.
Metzger. 2008. Using gap-crossing capacity to evaluate functional connectivity
of two Atlantic rainforest birds and their response to fragmentation. Austral
Ecology, 33: 863–871.
Azhar, B., D. B. Lindenmayer, J. Wood, J. Fischer, A. Manning, C. McElhinny y M.
Zakaria. 2013. The influence of agricultural system, stand structural complexity
and landscape context on foraging birds in oil palm landscapes. Ibis, 155: 297-
312.
Azman, N.M, N. S. Latip, S. A. Sah, M. A. Akil, N. J. Shafie y N. L. Khairuddin NL. 2011
Avian Diversity and Feeding Guilds in a Secondary Forest, an Oil Palm
Plantation and a Paddy Field in Riparian Areas of the Kerian River Basin, Perak,
Malaysia Tropical life science research, 22(29): 45–64.
Cagod, B. M. y O. M. Nuñeza. 2012. Avian species diversity in oil palm plantations of
Agusan Del Sur and Compostela Valley, Philippines. AES. Bioflux, 4(2): 85–
105.
Cousin, J. y R. D. Phillips. 2008. Habitat complexity explains species-specific occupancy
but not species richness in a Western Australian Woodland. Australian Journal
Zoology, 56: 95-102.
Dhindsa, M. S y H. K SAINI. 1994. Agricultural ornithology: an Indian perspective.
Journal of Bioscience, 9(4): 391-402.
Dirzo, R. & M. C. García. 1992. Rates of deforestation in Los Tuxtlas, a neotropical area
in southeast Mexico. Conserv. Biol. 6: 84-90.
Espejo González, D., J. Hidalgo Martin, M. Santmaria Galindo y J. Fernández. 20014.
Insectos asociados entre un cultivo de curuba y un fragmento de bosque
altoandino de la sabana de Bogotá. Inventum, 16: 9-16.

81
Fitzherbert, E. B., M. J. Struebig, A. Morel, F. Danielsen, C. A. Brühl, P. F. Donald y B.
Phalan. 2008. How will oil palm expansion affect biodiversity?. Trends in
Ecology and Evolution. 23(10): 538-545.
González-Borrero, L. H. 2013. Anàlisis multitemporal de los cambios de cobertura de la
tierra e incidencia del cultivo de plama en el territorio del municipio
deVillanueva Casanare. Tesis de Maestría, Pontificia Universidad Javeriana
Gonzáles-Salazar, C., E. Martínez-Meyer y G. López-Santiago. 2014. A hierarchical
classification of trophic guilds for North American birds and mammals. Revista
Mexicana de Biodiversidad, 85: 931-941.
Hawkins C. P. y J. A. MacMahon. 1989. Guilds: the multiple meanings of a concept.
Annual Rev Entomology 34:423–451.
Johnson, R. J., J. A. Jedlicka, J. E. Quinn y J. R. Brandle. 2011. Global perspective on
Birds in Agricultural Landscapes. En: Campbell, W. B. y S. L. Ortíz (Eds.).
Integrating agriculture, conservation and ecotourism: examples from the field.
Vol. 1. Springer-Verlag Berlin. AlemaniaKarp, D. S, G. Ziv, J. Zook, P. R.
Ehrlicha y G. C. Daily. 2011. Resilience and stability in bird guilds across
tropical countryside. PNAS, 18(52): 21134–21139.
López-Ordoñez, L. P., F. G. Styles y J. L. Parra Vergara. 2015. Protocolo para medición
de rasgos funcionales en aves. En: Salgado-Negret, B. (ed). La ecología funcional
como aproximación al estudio, manejo y conservación de la biodiversidad:
protocolos y aplicaciones. Instituto de Investigación de Recursos Biológicos
Alexander von Humboldt. Bogotá, D. C. Colombia. 236 pp.MacFadyen, A. 1957.
Animal ecology: Aims and Methods. Pitman and Sons, London.for conservation.
Fauna & Flora International, Oryx, 45(2), 234–242.
Marinoni, R. 2001. Os grupos tróficos em Coleoptera. Revista Brasileira de Zoologia, 18
(1): 205-224.
Minorta-Cely, V., & Rangel-Ch. J. O. (2014). Clima de la Orinoquía colombiana. En
Rangel-Ch, J. O. (Ed.), Colombia Diversidad Biótica XIV. La región de la
Orinoquía de Colombia (pp. 153-206). Universidad Nacional de Colombia –
Instituto de Ciencias Naturales. Bogotá, D.C., Colombia.
Mora Fernández C., C. Castellanos Castro, A. Cardona Cardozo, L. Pinzón Pérez & O.
Vargas. 2011. Historia de transformación del paisaje, de la cuenca baja del río
Pauto, Casanare (Colombia). En: Tomás León Sicard (Ed.) 2011. Mamíferos,
Reptiles y Ecosistemas del Bloque Cubiro (Casanare). Educación Ambiental para
la Conservación. Instituto de Estudios Ambientales, Universidad Nacional de
Colombia, Bogotá.

82
Pin Koh, L. 2008. Birds defend oil palms form herbivorous insects. Ecological
Applications, 18(4): 821–825.
Pinzón Pinzón, F., Pardi Lacruz, M. S., Trejos Serrato, J., & Midori Saito, S. (Abril, 2009).
Uso de técnicas de percepción remota para la estimación del incremento de áreas
cultivadas de palma aceitosa y su relación con el conflicto armado en el
Departamento del Meta-Colombia. Anais En. XIV Simpósio Brasileiro de
Sensoramiento Remoto. Natal, Brasil.
Prescott, G. W., J. J. Gilory, T. Haugassen, C. A. Medina Uribe, W. A. Foster y D. P.
Edards. 2016. Reducing the impacts of Neotropical oil palm development on
functional diversity. Biological Conservation, 197: 139-145.
Ramírez-Albores, Jorge E. (2010). Diversidad de aves de hábitats naturales y modificados
en un paisaje de la Depresión Central de Chiapas, México. Revista de Biología
Tropical,58(1), 511-528.
Ramírez González, A. 2005.Ecología Aplicada diseño y análisis. Universidad de Bogotá
Jorge Tadeo Lozano. Bogotá, Col.
Rangel-Ch, O., y Minorta-Cely, V. 2014. Tipos de vegetación de la Orinoquía colombiana.
En Rangel-Ch, J. O. (Ed.), Colombia Diversidad Biótica XIV. La región de la
Orinoquía de Colombia. Universidad Nacional de Colombia – Instituto de
Ciencias Naturales. Bogotá, ColombiaRestall, R., Rodner, C., & Lentino, R.
2006. Birds of northern South America. Christopher Helm. London.
Rippstein, G., Escobar, G., & Motta, F. (Eds.). (2001). Agroecología y biodiversidad de
los Llanos Orientales de Colombia. Centro Internacional de Agricultura Tropical
(CIAT) # 322. Cali, Colombia. Roa, M. 1998. Ecología de la avifauna en cercas
vivas en un ecosistema fragmentado del piedemonte llanero. Importancia para su
conservación. Tesis Biología. Pontificia Universidad Javeriana.
Root, R. B. 1967 The niche exploitation pattern of the blue-gray gnatcatcher. Ecology
Monography 37:317–350.
Salgado Negret, B. y H. Paz. 2015. Escalando de los rasgos funcionales a procesos
poblacionales, comunitarios y ecosistémicos. En: Salgado-Negret, B. (ed). La
ecología funcional como aproximación al estudio, manejo y conservación de la
biodiversidad: protocolos y aplicaciones. Instituto de Investigación de Recursos
Biológicos Alexander von Humboldt. Bogotá, D. C. Colombia. 236 pp.
Sánchez, S. 2000. Vertebrdos silvestres registrados en una parcela aceitera en Tabasco,
México. ASD Oil Palm Papers, 20: 17-18.
Sekercioglu, C. H. 2006. Increasing awareness of avian ecological function. Trends in
Ecology and Evolution, 21(8): 464-471.

83
Sekercioglu, C. H. 2012. Bird functional diversity and ecosystem services in tropical
forests, agroforests and agricultural areas. Journal Ornithology. Published online.
Simberloff, D. y T. Dayan. 1991. The guild concept and the structure of ecological
communities. Annual Review of Ecology and Systematics, 22: 115-43.
Tokeshi, M. 1999. Species coexistence ecological and evolutionary perspectives. Wiley-
Blackwell. UK. 468p.
Toledo-Lima, G. S., P. Macario, R. M. De Lyra-Neves, B. P. Teixeira, L. A. F. De Lima,
G. O.S. Sugliano y W. R. 2014 Telino-Júnior. Richness, composition and trophic
groups of an avian community in the Pernambuco Endemism Centre, Alagoas,
Brazil. Annals of the Brazilian Academy of Sciences. 86 (3): 1207-1219.
Universidad Icesi 2014.Wiki Aves Colombia. Disponible: en
http://www.icesi.edu.co/wiki_aves_colombia. [20 Oct 2016].

84
CAPÍTULO 4. RESPUESTA DE UNA COMUNIDAD DE AVES ANTE EL
ESTABLECIMIENTO DEL CULTIVO DE PALMA DE ACEITE: UN ENFOQUE
A ESCALA DE PAISAJE
Diana P. Tamaris Turizo, Hugo F. López Arévalo y María Claudia Franco Rozo

85
Resumen
Las aves exhiben diferentes respuestas ante las características del paisaje así, los elementos
y tipos de cobertura que se encuentren en el terreno tendrán incidencia en la composición
de especies y la estructura de una comunidad. La respuesta de las aves ante la
configuración del paisaje donde predominan cultivos comerciales de palma de aceite se
analizó desde tres aspectos: valores de diversidad, gremios de alimentación y tamaño
corporal de las aves. Para ello se hicieron censos de aves con redes de niebla en 12 tipos de
coberturas, tales como lotes de palma, matorrales, fragmentos de bosque y bosque de
galería. En las unidades de muestreo se tomaron datos corporales de las aves capturadas y
de cada cobertura analizada se obtuvieron métricas de paisaje que se relacionaron con los
gremios de alimentación de las aves. En términos de diversidad, las aves fueron más
numerosas y abundantes en los matorrales del paisaje. El análisis de los gremios de
alimentación mostró que las picívoras estuvieron asociadas a bosque de galería, aves
carnívoras se relacionaron más con la palma y aves granívoras con los matorrales. La
dimensión fractal fue la métrica que tuvo mayor incidencia con el bosque de galería. En
cuanto al tamaño corporal se observó que las aves de mayor peso se asociaron a plamas
grandes y medianas y aves de mayor talla se asociaron a matorrales y bosque de galería. Se
pudo concluir que la comunidad de aves repondió de manera diferente ante los
componentes del paisaje, se evidenció que las coberturas del paisaje con vegetación nativa
son elementos que ayudan al sostén de la mayoría de las aves de la comunidad.

86
4.1 Introducción
Los atributos del paisaje como la composición y configuración de los elementos del
terreno tienen incidencia en la estructura de comunidades dentro de los ecosistemas
(Carrara et al., 2015; Fahrig, 2003; Fahrig et al., 2011). En el caso de las aves se ha
encontrado que los paisajes dominados por bosques primarios sostienen mayor diversidad
α y mayor asociación de aves especialistas que aquellos donde predomina la vegetación
secundaria y dominan especies generalistas (Carrara et al., 2015). Otras características del
paisaje como el área del parche, también puede afectar la diversidad de aves puesto que
generalmente se presenta una relación positiva entre el área y el número de especies: a
mayor área, mayor número de especies (Turner et al., 2001; Diamond y Mayr, 1976).
La distancia de borde causada por la fragmentación de la vegetación también es
características que tiene influencia directa sobre las poblaciones y comunidades de aves
(Fahrig, 2003; Robinson et al., 1995), y tal influencia puede ser perjudicial o no sobre la
composición, distribución y abundancia de las especies (Banks-Leite et al., 2010). Existe
evidencia que el incremento del borde favorece la invasión de fragmentos por especies
generalistas. Estos procesos invasivos pueden afectar tanto la supervivencia de las especies
(por depredación y competencia de recursos), como las funciones ecosistémicas, como
polinización, dispersión de semillas, generando a su vez extinciones locales dentro de los
hábitats remanentes (Santos y Tellería, 2006; Aizen y Feisinger, 1994; Tallmon et al.,
2003).
En países tropicales los paisajes son heterogéneos, pero el aumento poblacional y los
cambios en el uso de la tierra en las últimas décadas ha conducido a la fragmentación de
ecosistemas y reducción del tamaño de las áreas naturales, generando cambios en la
composición y configuración del paisaje (Camacho et al. 2014). Conforme se transforman
los ecosistemas por los cambios en el uso del suelo, resulta necesario entender y predecir

87
los cambios en el comportamiento de los animales (Bevanda et al. 2015) e identificar
cómo se afectan los patrones y procesos a nivel de poblaciones y comunidades.
Con frecuencia las aves son consideradas el grupo menos afectado en los procesos de
cambios y transformación del paisaje, por la facilidad de movimiento, en comparación con
otros grupos taxonómicos de menor movimiento, pero de hecho muchas especies de aves
pueden expresar mayor sensibilidad a la fragmentación, puesto que ya se ha demostrado
que puede afectar severamente la disponibilidad de recursos para la alimentación,
reproducción y la persistencia de los ensambles de aves (Gibson et al. 2011, Suk et al.
2014).
Las aves interactúan con los sistemas agrícolas en diferentes escalas espaciales y
temporales para alimentarse, anidar, forrajear, y cubrir recursos durante los periodos de
migración y fuera de estos (Lima y Zollner 1996, Johnson et al. 2011), pero el
establecimiento de cultivos a gran escala también aumenta la tasa de depredación, influye
en la eficiencia de forrajeo, las poblaciones de depredadores pueden aumentar, al menos a
escala local (Tapper et. al., 2006, Whittingham y Evans, 2004), y en los relictos de
vegetación adyacente algunas especies quedan confinadas a parches que antes eran de
mayor extensión (Wiens 1989, Villard et al. 2001).
Los paisajes con monocultivos de palma de aceite son extensos, homogéneos y se han
considerado como áreas propensas que pueden perjudicar la persistencia y el movimiento
de las especies (Azhar et al 2015). Cuando se establece una plantación surgen drásticas
modificaciones en cuanto a composición y estructura de la vegetación a gran escala, y
precisamente estos eventos de conversión reducen la riqueza de especies y la diversidad
funcional con implicaciones importantes como la provisión de recursos para las especies y
servicios ecosistémicos (Prescott et al., 2016). Estas plantaciones soportan menos gremios
de forrajeo que un bosque, independientemente del manejo que se le de al sistema, la
diversidad y estructura de los gremios también cambia de acuerdo a las nuevas
condiciones ambientales como el microclima, la estructura de la vegetación, y el tipo y
disponibilidad de alimento para las aves (Azhar et al., 2013¸ Bruhl, y Eltz, 2010).

88
Para probar si la diversdiad de aves varía con los atributos del paisaje se analizó la
comunidad desde tres aspectos: la diversidad, los gremios de alimentación y rasgos
morfométricos, para ello se planteó como objetivo identificar los atributos del paisaje que
inciden en la diversidad de aves en paisajes dominados por cultivos de palma de aceite.
4.2 Métodos
Área de estudio
El estudio se hizo en dos fincas del Oriente de Colombia, con cultivos de palma de aceite
llamadas Tierra Vivia y Mejorana, ubicadas en los municipios San Martín de los Llanos
(03°37'51.8" N, 073°20'59.7" W) y Acacias (03°57'37.18" N, 73°37'32.93” W),
respectivamente, departamento del Meta. Cada predio cuenta con un área de
aproximadamente 1.400 y 1480 ha.
El área geográfica de ambas localidades hace parte del piedemonte cordillerano, en la que
se destacan bosques, matorrales, pastizales y áreas con actividades agrícola y ganadera
(Rangel-Ch y Minorta-Cely, 2014; Pinzón Pinzón et al., 2009, Mora Fernández et al.,
2011; Rippstein et al., 2001), lo cual ha generado cambios estructurales en la vegetación,
procesos de fragmentación y alteraciones microclimàticas de los fragmentos (González-
Borrero, 2013). El régimen biestacional hace que la temperatura sea elevada en el los
primeros y últimos meses del año, mietras que a mitad de año se registran las temperaturas
más bajas (Minorta-Cely y Rangel-Ch, 2014). Por su parte, las precipitacines tienen un
margen de variación anual de 2877 a 4000 mm (Minorta-Cely y Rangel-Ch, 2014).
Para conocer la respuesta de la comunidad de aves en paisajes dominados por cultivos de
palma de aceite, se hicieron análisis desde tres enfoques: diversidad de aves entre tipos de
cobertura, se analizaron los gremios de alimentación con respecto a los tipos de cobertura
y a las variables del paisaje de cada predio y se analizaron los tipos de cobertura de cada
predio con variables de riqueza, abundancia y rasgos morfológicos de las aves.

89
Censos de aves
Se hizo un censo de aves mediante capturas con redes de niebla. Las redes se instalaron en
el interior de las coberturas denominadas palma grande, palma mediana, palma pequeña,
matorral y bosque de galería (Tabla 5.2) abarcando 12 unidades de muestreo en total. En
cada unidad de muestreo se instalaron de 7 a 10 redes (12× 2.5 m y ojo de malla de 36
mm) que se abrieron de 6:00 a 10:00 am y de 3:30 a 5:30 pm dedicando 360 horas/red
durante el estudio.
A cada ejemplar capturado se le tomaron medidas morfométricas y fueron asignadas a uno
de los siguientes gremios de alimentación teniendo en cuenta la principal fuente de
alimento: carnívoro (CAR), frugívoro (FRU), frugívoro-insectívoro (FRU-INS), granívoro
(GRA), 5) granivoro-insectívoro (GRA-INS), insectívoro (INS), nectarívoro (NEC) y
piscívoro (PIC). La asignación de cada grupo se basó en observaciones directas y revisión
de literatura especializada (Restall et al. 2006, Universidad Icesi 2014).
Análisis de paisaje
Se utilizó como insumo para la identificación de coberturas las Imágenes satelitales
Landsat 8 registrada en enero 16 de 2016, que captura 11 bandas multiespectrales con una
resolución espacial de 30 metros para las bandas 1-7, 9, de 15 metros para la banda
pancromática 8 y para las bandas TIRS 10 y 11 un remuestreo de 30 metros a una toma
originalmente a 100 metros (NASA, 2016).
Para la interpretación de la imagen se definió un área de trabajo que corresponde al área de
los predios Tierra Viva (1400 ha) y Mejorana (1480 ha) más un buffer de 1,5 km, que
permitió identificar la continuidad de las coberturas más allá de los límites prediales y con
lo cual se obtuvieron dos ventanas de 5539.7 ha y 5894.6 ha.
Para visualizar e identificar las coberturas del terreno se utilizó la herramienta Layer
Stacking de ENVI 5.0 que fusiona las bandas con un orden 5-4-3, a partir de lo cual se
obtuvo una imagen a una resolución de 30 metros. Y con la herramienta Wavelet Fusión

90
del mismo programa, se integra la banda 8, que corresponde a la banda pancromática, para
obtener una imagen con una resolución de 15 metros.
La imagen resultante se trabajó en el programa ArcGIS 10.3, sobre la cual se realizó una
identificación visual de coberturas, teniendo en cuenta las unidades de cobertura definidas
en la tabla 5.1 y 5.2. La identificación de cada unidad del paisaje se verificó con imágenes
Google Earth, World Imagery que incluye el ArcGis 10.3 y con el levantamiento en campo
durante el periodo de estudio.
Tabla 4. 1 Elementos del paisaje y coberturas asociadas en las fincas Tierra Viva y Mejorana
Elemento/Cobertura Asociada
Área Cob
(m2)
Área Elemento
(m2) %
Área Cob
(m2)
Área
Elemento (m2) %
Matriz Palma grande 1405.71 2198.71 37.30 888.60 2877.52 51.94
Palma mediana 394.35 680.88
Palma muerta 0.00 84.49
Palma pequeña 398.66 1223.56
Parche regenrado Matorral 85.32 105.89 1.80 108.59 203.57 3.67
fragmento de bosque 20.58 94.98
Parche introducido Infraestructura 3.95 356.03 6.04 72.40 80.77 1.46
Otros cultivos 352.08 8.37
Parche de disturbio Pastos limpios 1285.84 1301.93 22.09 1680.14 1680.14 30.33
Pastos arbolados 16.09 0.00
Parche de recursos ambientalesHumedal 171.54 171.54 2.91 15.37 15.37 0.28
Corredor Bosque de galería 1756.93 1760.57 29.87 614.96 682.34 12.32
Río 0.00 57.96
Cerca viva 3.641 9.42
Total 5894.67 100 5539.71 100
MejoranaTierra Viva

91
Tabla 4. 2 Definición de las coberturas del paisaje
Elemento Definición*
Palma grande Lotes de palmas con altura igual o mayor a 11 metros
Palma mediana Lotes de palmas con altura de 7 a 10 metros
Palma muerta Lotes de palma inerte con altura igual o mayor a 11 metros
Palma pequeña Lotes de palmas con altura de 2 a 6 metros
Fragmento de
bosque
Es el resultado de la perturbación extensa de la matriz que deja una serie de islas no
afectadas en su estado original, son opuestos a los de perturbación los diferencia la escala
de la perturbación local, que en los parches remanentes es más severa ya que solo deja
islas de la matriz original.
Matorral Formaciones vegetales constituidas principalmente por arbustos; su densidad aunque es
variable, tiende a presentar valores altos
Infraestructura
Son las áreas cubiertas por infraestructura artificial (terrenos cimentados, alquitranados,
asfaltados o estabilizados), las cuales se utilizan también para actividades comerciales o
industriales.Incluye entre otros, instalaciones de ganadería, estanques piscícolas,
invernaderos menores a 5ha, terminales de almacenamiento de petróleo o carbón
(IDEAM, 2010)
Otros cultivos
Son unidades que reúnen dos o más clases de coberturas agrícolas y naturales,
dispuestasen un patrón intrincado de mosaicos geométricos que hace difícil su separación
en coberturas individuales (IDEAM, 2010)
Pastos
arbolados
Cobertura que incluye las tierras cubiertas con pastos, en las cuales se han estructurado
potreros con presencia de árboles de altura superior a cinco metros, distribuidos en forma
dispersa. La cobertura de árboles debe ser mayor a 30% y menor a 50% del área total de la
unidad de pastos (IDEAM, 2010)
Pastos limpios
Esta cobertura comprende las tierras ocupadas por pastos limpios con un porcentaje de
cubrimiento mayor a 70%; la realización de prácticas de manejo (limpieza, encalamiento
y/o fertilización, etc.) y el nivel tecnológico utilizado impiden la presencia o el desarrollo
de otras coberturas. (IDEAM, 2010)
Humedal Superficies o depósitos de agua naturales de carácter abierto o cerrado, dulce o salobre,
que pueden estar conectadas o no con un río o con el mar. (IDEAM, 2010)
Bosque de
galería*
Corresponde a las áreas con vegetación de tipo arbóreo caracterizada por un estrato más o
menos continuo cuya área de cobertura arbórea representa más de 70% del área total de la
unidad, y con altura del dosel entre 5 y 15 metros y que se encuentra localizada en las
franjas adyacentes a los cuerpos de agua (lóticos) (IDEAM, 2010).
Cerca viva
Árboles o arbustos sembrados de forma lineal como soportes para el alambre de púas o
liso, siguiendo los límites de una propiedad o marcando las divisiones entre parcelas
según los diferentes usos del suelo (cultivos anuales o perennes, potreros, bosques, etc.).
Una cerca viva puede estar formada solamente de especies leñosas o de una combinación
de especies leñosas con postes muertos, de especies nativas sembradas o remanentes o por
especies nativas y/o exóticas plantadas artificialmente (Villanueva et al 2005, Roa 1998).

92
Analisis de datos:
La diversidad de aves entre tipos de cobertura se analizó descriptivamente a partir de la
riqueza (S) de esepcies, abundancia de individuos (N) y riqueza de gremios de
alimentación identificados en cada tipo de cobertura.
Para identificar la relación de los gremios de alimentación con los tipos de cobertura y con
las variables del paisaje de cada predio se hizo un Análisis de Correspondencia Canónica y
(ACC), este permite relacionar las comunidades biológicas con variables ambientales. Las
variables biológicas o dependientes fueron los gremios de alimentación de las aves (CAR,
FRU, FRU-INS, GRA, GRA-INS, INS, NEC y PIC). Las variables ambientales fueron
cinco métricas de las coberturas del paisaje donde se instalaron las redes, estas métricas
mostraron mayor relación con las coberturas donde se hicieron censos de aves (Tabla 5.3),
dichas coberturas corresponden al 65.2% y 68.9% del terreno de los predios Mejorana y
Tierra Viva, respectivamente.
Para determinar si los tipos de cobertura de cada predio (TIV, ME) se relacionaban con la
riqueza, abundancia, y los rasgos morfométricos talla y peso, se hizo un Análisis de
Componentes Principales. El análisis se hizo con las coberturas bosque de galería,
fragmento de bosque, matorral, palma grande, palma mediana y palma pequeña, y se
tuvieron en cuenta como variables biológicas la riqueza específica, abundancia, longitud
total máxima (LTmax) y peso máximo (Gmax) de los ejemplares capturados en las
mencionadas coberturas. Finalmente, una prueba Friedman se hizo para identificar
diferencias de la diversidad de aves entre tipos de cobertura de los predios analizados.

93
Tabla 4. 3 Descripción de las métricas de paisaje a nivel de clase
Abreviatura Descripción de la variable*
DFRA Corresponde a la dimensión fractal ponderadaÍndice de forma media ponderada del
área. Es un índice de forma basado en la relación área-perímetro en base a una
transformación logarítmica
MPS Es el tamaño promedio del parche y corresponde al tamaño promedio de los parches
de un tipo de coberturaen el paisaje (m2)
NumP Número de parches, es el número de parches de un tipo de cobertura (número
absoluto)
PA Proporción del área es la proporción de un área de un tipo de cobertura en el paisaje
(%)
TE El borde total es una medida de la cantida de borde que tiene cada tipo de cobertura
(m) *Botequilha et al., 2006.
4.3 Resultados
Caracterización del paisaje:
Se identificaron 14 tipos de coberturas asociados al paisaje MEJ y TIV (Tabla 4.4, Figura
4.1 y 4.2), la palma, los pastos limpios y los pastos arbolados fueron los elementos más
representativos en el terreno (43.98 y 26.27%, respectivamente). Mejorana fue el predio
más fragmentado en el cual se identificaron 252 parches de los cuales el bosque de galería
tuvo la mayor cantidad (52). En esta localidad los lotes de plama (grande, mediana y
pequeña) tuvieron la mayor proporción de área (35.9%). Los pastos limpios también
ocuparon una proporción importante (30%) y en gran medida se encontraron adyacentes a
lotes con cultivos de palma. La palma mediana tuvo un solo parche y fue el de mayor
tamaño promedio (MPS) y el bosque de galería fue la cobertura con mayor medida de
borde (TE) (Tabla 4.4).
Por su parte humedales fue la cobertura con mayor número de parches en el predio Tierra
Viva (20) dentro del cual se identificaron 97 en total. El cultivo de palma totalizó un área
promedio de 37.3%, seguido del bosque de galería (29.8), este último también tuvo la
mayor medida de borde (TE) . La cobertura denominada otros cultivos se destacó por tener
el mayor valor de tamaño promedio de parche (MPS) (Tabla 4.4).

94
Diversidad de aves entre tipos de cobertura
La cobertura matorral tuvo mayor registro de aves (47 especies, 137 individuos), y la
mayor riqueza de gremios de alimentación se registró en la palma pequeña (7), mientras
que la coberuta con menor cantidad de aves y de gremios de alimentación fue la palma
mediana (5 especies y 3 grupos) (Tabla 4.5).
Relación de Gremios de alimentación con variables del paisaje y tipos de cobertura
De acuerdo con el Análisis de Correspondencia Canónica ACC (Fig 4.3) los gremios de
alimentación se relacionaron con ciertos tipos de cobertura, y a su vez se observó que
algunas aves respondieron a los atributos del paisaje (sumatoria de la varianza de los dos
primeros ejes fue del 79%). En el cuadrante superior derecho se observa que las aves PIC
presentaron una relación con el bosque de galería de Tierra Viva, y en menor grado a los
bosques de galería de Mejorana. El grupo de las CAR se asoció a palmas grandes y
pequeñas de Tierra Viva y Mejorana. Las palmas medianas de ambos predios, junto con
los fragmentos de bosque de Mejorana se agruparon en función a la escasa abundancia de
aves. El grupo de las GRA estuvo correlacionado con los matorrales de ambos predios y
con menor fuerza a las palmas pequeñas de Tierra Viva.También se distingue que aves
FRU, FRU-INS e INS estuvieron muy cerca del origen, indicando que fueron grupos
abundantes en la mayoría de las coberturas analizadas. El grupo de las NEC y GRA-INS
tuvieron una relación débil con fragmentos de bosque de Mejorana. El triplot representa la
magnitud que tuvieron de las métricas del paisaje con relación a los grupos tróficos o a los
tipos de cobertura, y con forme se aleja del centro indica mayor grado de asociación. En
este sentido el tamaño medio del parche (MPS) fue una característica que no tuvo una
asociación importante con los grupos trófico ni con las coberturas analizadas. El número
de parches (NumP) fue un atributo principalmente correlacionado con aves INS, las cuales
dominaron en los matorrales de ambos predios.
También se distingue que las aves INS y FRU-INS se estuvieron relacionadas a cuatro
características del paisaje (PA, TE, DFRA y NumP). De forma opuesta CAR y PIC no

95
mostraron una fuerte asociación con las características del paisaje. La dimensión fractal
(DFRA) fue la medida que tuvo mayor relación con la cobertura boque de galería de
Mejorana, sin embargo, no parece ser un atributo importante para las aves.
Relación de los tipos de cobertura con rasgos morfométricos
Apartir del Análisis de Componentes Principales se observó que las aves de mayor peso
(Gmax) se asociaron con palmas grandes y medianas de ambas fincas. Las aves de mayor
tamaño (Ltmax) se asociaron a los matorrales y bosques de galería de ambas fincas. La
riqueza (S) y abundancia (Num Ind) se proyectaron muy cerca del origen indianco que
fueron variables carentes de asociación con los tipos de cobertura (Figura 4.4).
4.4 Discusión
Los resultados mostraron que las aves respondieron de manera diferente ante la
composición del paisaje y los atributos que tuvieron mayor incidencia en la asociación y
diversidad de aves fueron las coberturas con formaciones vegetales naturales y la
dimensión fractal fue el índice más importante.
El análisis del paisaje y la clasificación de coberturas son un reflejo la expansión de la
frontera agrícola con cultivos de palma de aceite en ambos municipios del departamento
del Meta. Una matriz agrícola dominada por estos monocultivos también se presente en 19
mucipios del mismo departamento (Pinzón Pinzón et al, 2009) así como en otros
departamentos como Arauca, Casanare y Vichada que en su conjunto ubican a la zona
Oriental como la de mayor ára sembrada en el país con 181.543 ha para el año 2015
(Fedepalma, 2016). Un caso relevante se presenta en Mapiripan, sur del Meta, donde se ha
establecido 487.385 ha nuevas del cultivo de palma desde el año 2009 en ecosistemas de
sabanas nativas, trayedo modificaciones en la fisionomía el ecosistema y en las
interacciones de los organismos, como ya lo documentaron Olárte González y Escovar
Fadul (2014), quienes reportan por primera vez al mamífero Puma concolor en un sector
de Mapiripan consumiento venado (Mazama americana) dentro del cultivo. Lo anterior es

96
un claro ejemplo de las alteraciones biológicas que trae el establecimiento de plantaciones
comerciales de palma de aceite en los ecosistemas de la Orinoquía colombiana.
Los matorrales tuvieron mayor cantidad de aves en cuanto a número de especies, número
de individuos, y se agruparon en seis grupos trófico, mientras que la palma fue más pobre
en la cantidad de aves registradas mediante técnicas de capturada. En la Orinoquía los
matorrales son formaciones que se caracterizan en parte por tener arbustos de 7 a 8 metros,
Miconia albicans es una de las plantas arbustivas más importantes (Rangel-Ch. y Minorta-
Cely) y llamativa para muchas aves, especialmente del grupo de la tángaras. Por su
arquitectura y composición los matorrales son elementos de mayor complejidad estructural
y éste es un factor fundamental del cual dependen muchas aves para permanecer en un
ecosistema (Cousin y Phillips, 2008, Ramachandran y Ganesh, 2012). Los cultivos de
palma de aceite son sistemas muy uniformes a lo largo de su ciclo de vida, la reducida
complejidad queda expresada en las plantas epífitas que crecen sobre los troncos y en la
vegetación del suelo cuando no es removida (Luskin y Potts 2011), este entorno alberga
pocas especies de aves que en este estudio fueron abundantes principalmente en palmas
pequeñas más que en las demás clases de altura.
La comunidad de aves analizada desde los gremios de alimentación mostró asociaciones
marcadas de aves piscívoras, carnívoras y granívoras en coberturas específicas: bosque de
galería, cultivos de palmas y matorrales, respectivamente. El conjunto de aves piscívoras
estuvo representado por Chloroceryle amazona, Chloroceryle americana y Chloroceryle
inda. Los hábitos de forrajeo explican, en parte, los patrones de movimiento y la selección
de sitios (Morrison et al. 1990). En este sentido los miembros de la familia Alcedinidae
habitan en bosques que tengan cercanía con fuentes de agua (lóticas o lénticas), donde
perchan para cazar peces principalmente (Kirschbaum, 2004). Las aves carnívoras
estuvieron representadas por Falco femoralis y Rupornis magnirostris. Este gremio se
considera parte integral en el control de roedores en los cultivos de palma de aceite
(Quarles, 2011) donde han encontrado un entorno favorable para cazar pequeños
vertebrados como reptiles, anfibios, mamíferos y pequeñas aves (Sánchez, 2000) que
consumen como parte de su dieta y que observan con mayor facilidad dentro de la

97
plantación. Un ejemplo de la favorabilidad de aves rapaces en cultivos de palma se
documentó con la lechuza Tyto alba, la cual ha incrementado su población e hizo un
efectivo control de roedores en este tipo de sistemas agrícolas (Quarles, 2011, Duckett,
1991, Chia et al., 1995; Basri et al., 1996). Las granívoras que fueron comunes en
matorrales del área de estudio fueron Ammodramus aurifrons, Columbina minuta,
Columbina talpacoti, Sicalis flaveola y Sporophila lineola, las tres últimas con frecuencia
se perchaN y forrajean en los arbustos del sotobosque buscando frutos o semillas como
parte de su dieta (Morrison et al. 1990, Blake, el al. 1990).
Las aves responden de manera diferencial en paisajes fragmentados y la abundancia de las
especies puede variar por el área del parche, la densidad de borde, entre otros factores
(Crozier y Neimi, 2003). Las métricas de paisaje calculadas a nivel de clase mostraron que
las aves INS y FRU-INS se correlacionaron con el borde (TE), el área (PA) y el número de
parches (NumP). Estas asociaciones nos permitieron corroborar que algunas especies les
resulta conveniente la fragmentación, porque se aumenta el borde y los sitios de percha
para cazar alimento (Anjos, 2006). Sin embargo, otras quedan menos favorecidas,
especialmente en las aves de interior de bosque o aquellas donde el efecto de borde
aumenta la depredación de nidos (Andren, 1994). Especies, como por ejemplo de los
géneros Tyrannus, Myozetetes, Myiodinastes, se benefcian de la cantida de parches y borde
dentro de un pasaje porque son cazadoras aéreas y utilizan los bordes de la vegetación para
cazar y consumir insectos. Mientras que hay aves insectívoras que son de interior de
bosque (como especies del género Myrmeciza), que pueden ser más sensibles a la
fragmentación en cuanto se reduce en tamaño del parche y aumenta la cantidad de borde.
La dimensión fractal (DFRA) se correlacionó con el bosque de galería de MEJ, este índice
refleja el grado de complejidad de un polígono y esta relación se explica porque fue la
cobertura con mayor número de parches en el paisaje de Mejorana.
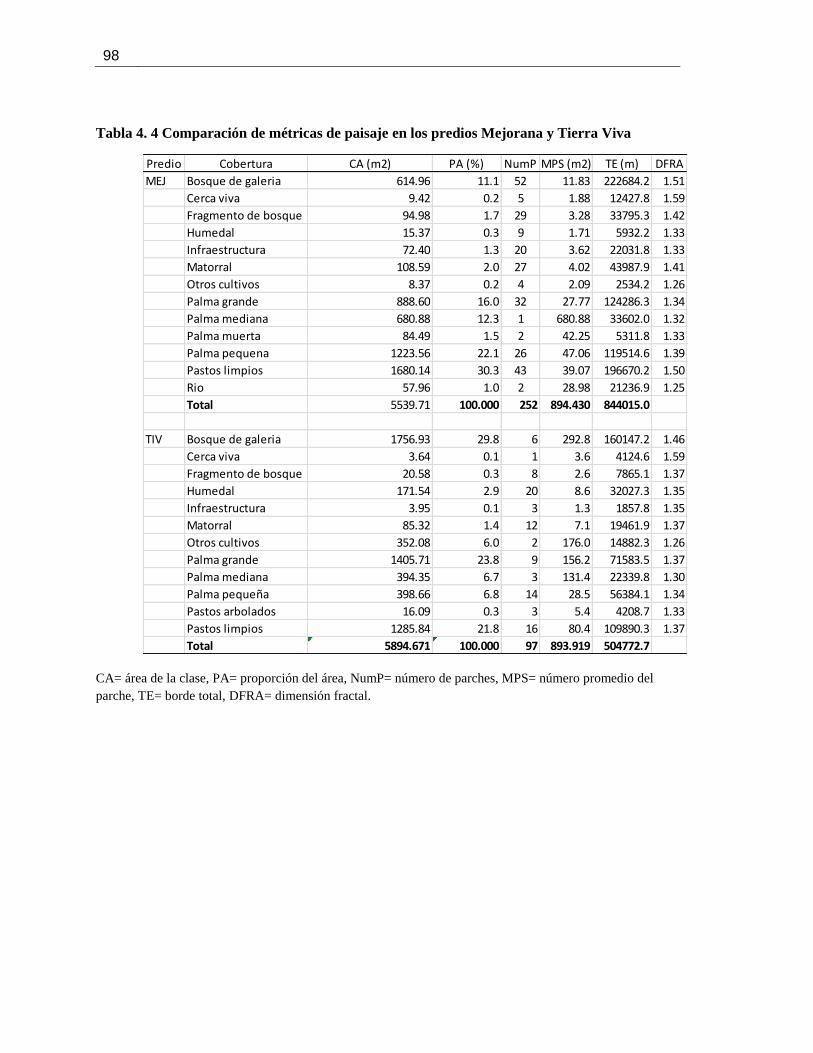
98
Tabla 4. 4 Comparación de métricas de paisaje en los predios Mejorana y Tierra Viva
Predio Cobertura CA (m2) PA (%) NumP MPS (m2) TE (m) DFRA
MEJ Bosque de galeria 614.96 11.1 52 11.83 222684.2 1.51
Cerca viva 9.42 0.2 5 1.88 12427.8 1.59
Fragmento de bosque 94.98 1.7 29 3.28 33795.3 1.42
Humedal 15.37 0.3 9 1.71 5932.2 1.33
Infraestructura 72.40 1.3 20 3.62 22031.8 1.33
Matorral 108.59 2.0 27 4.02 43987.9 1.41
Otros cultivos 8.37 0.2 4 2.09 2534.2 1.26
Palma grande 888.60 16.0 32 27.77 124286.3 1.34
Palma mediana 680.88 12.3 1 680.88 33602.0 1.32
Palma muerta 84.49 1.5 2 42.25 5311.8 1.33
Palma pequena 1223.56 22.1 26 47.06 119514.6 1.39
Pastos limpios 1680.14 30.3 43 39.07 196670.2 1.50
Rio 57.96 1.0 2 28.98 21236.9 1.25
Total 5539.71 100.000 252 894.430 844015.0
TIV Bosque de galeria 1756.93 29.8 6 292.8 160147.2 1.46
Cerca viva 3.64 0.1 1 3.6 4124.6 1.59
Fragmento de bosque 20.58 0.3 8 2.6 7865.1 1.37
Humedal 171.54 2.9 20 8.6 32027.3 1.35
Infraestructura 3.95 0.1 3 1.3 1857.8 1.35
Matorral 85.32 1.4 12 7.1 19461.9 1.37
Otros cultivos 352.08 6.0 2 176.0 14882.3 1.26
Palma grande 1405.71 23.8 9 156.2 71583.5 1.37
Palma mediana 394.35 6.7 3 131.4 22339.8 1.30
Palma pequeña 398.66 6.8 14 28.5 56384.1 1.34
Pastos arbolados 16.09 0.3 3 5.4 4208.7 1.33
Pastos limpios 1285.84 21.8 16 80.4 109890.3 1.37
Total 5894.671 100.000 97 893.919 504772.7
CA= área de la clase, PA= proporción del área, NumP= número de parches, MPS= número promedio del
parche, TE= borde total, DFRA= dimensión fractal.
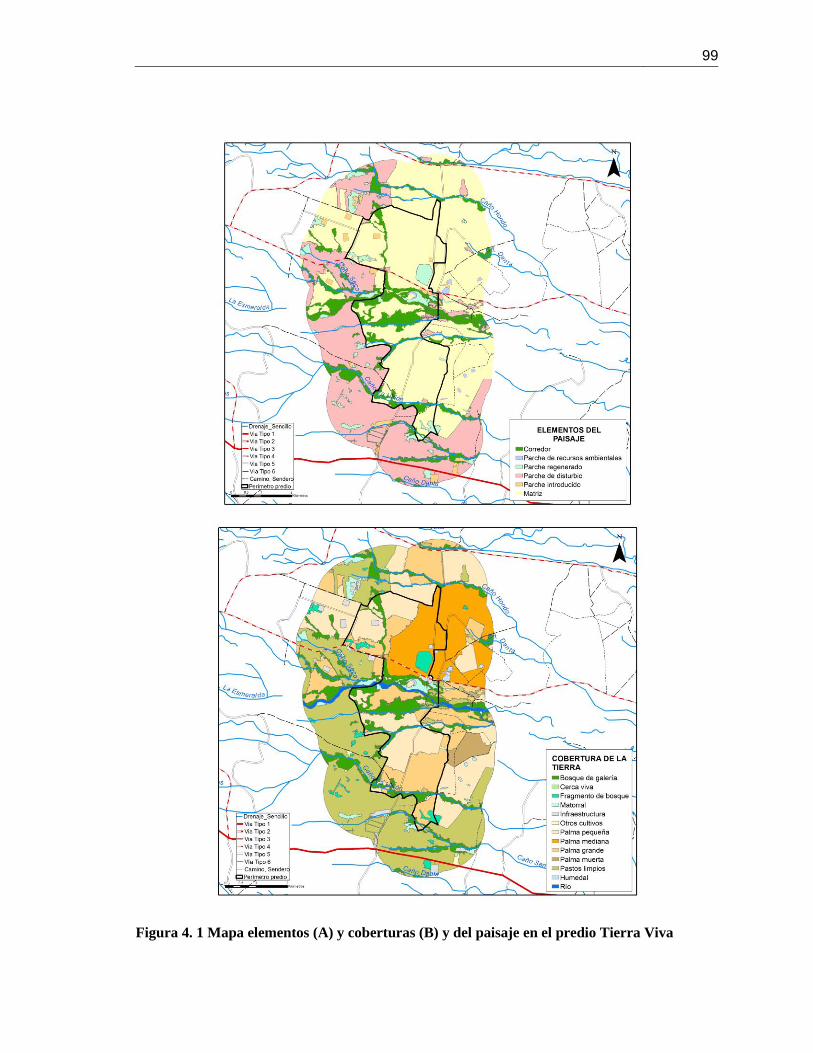
99
Figura 4. 1 Mapa elementos (A) y coberturas (B) y del paisaje en el predio Tierra Viva

100
Figura 4. 2 Mapa elementos (A) y coberturas (B) y del paisaje en el predio Mejorana

101
Tabla 4 5. Diversidad de aves asociadas al cultivo de palma y coberturas del paisaje
Cobertura S N GT
Boque de Galería 20 35 5
Fragmentos de bosque 17 40 5
Matorral 47 137 6
Palma Grande 9 8 5
Palma Mediana 5 9 3
Palma Pequeña 22 42 7 S: riqueza de especies en cada grupo trófico, N: abundancia de individuos en cada grupo trófico, GT: grupo
trófico
Figura 4. 3 Distribución de gremios de alimentación de aves en función a coberturas y
métricas del paisaje MEJ_Bga: bosque de galería Mejorana, TIV_Bga: bosque de galería Tierra viva, MEJ_FrB:
fragmento de bosque Mejorana, TIV _FrB: fragment de bosque Tierra Viva, MEJ_Mat: matorral Mejorana, TIV_Mat:
Matorral Tierra Viva, MEJ_PG: palma grande Mejorana, TIV_PG: palma grande Tierra Viva, MEJ_PM: palma mediana
Mejorana, TIV_PM: palma mediana Tierra Viva, MEJ_PP: palma pequeña Mejorana, TIV_PP: palma pequeña Tierra
Viva
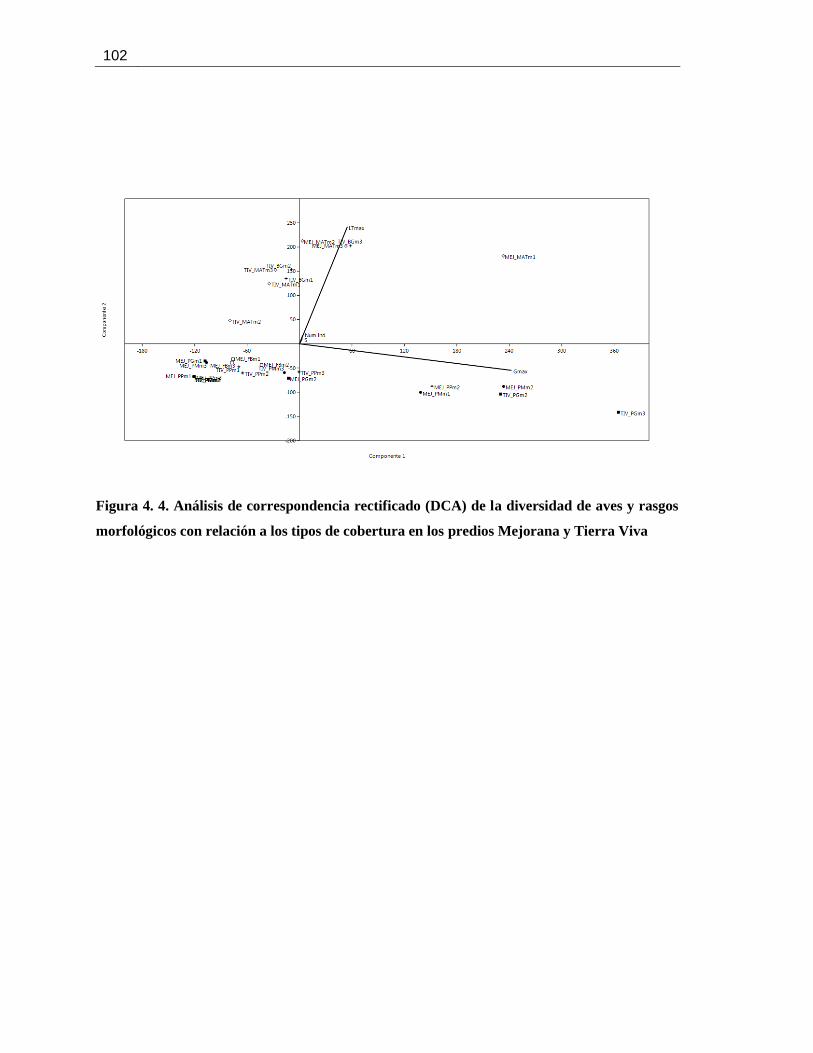
102
Figura 4. 4. Análisis de correspondencia rectificado (DCA) de la diversidad de aves y rasgos
morfológicos con relación a los tipos de cobertura en los predios Mejorana y Tierra Viva

103
1.1 Referencias
Andren, H. 1994. Effects of habitat fragmentation on birds and mammals in landscapes
with different proportions of suitable habitat: a review. Oikos 71: 355-366.
Anjos, L. D. 2006. Bird Species Sensitivity in a Fragmented Landscape of the Atlantic
Forest in Southern Brazil .Biotropica, 38(2): 229-234.
Azhar, B., Lindenmayer, D. B., Wood, J., Fischer, J., Manning, A., McElhinny, C., & Zakaria, M.
(2013). The influence of agricultural system, stand structural complexity and landscape
context on foraging birds in oil palm landscapes. Ibis, 155, 297-312.
Azhar, B., N. Saadun, Ch. Leong Puana, N. Kamarudin, N. Aziz, S. Nurhidayu y J.
Fischer. 2015. Promoting landscape heterogeneity to improve the biodiversity
benefits of certified palm oil production: Evidence from Peninsular Malaysia.
Global Ecology and Conservation, 3: 553-561.
Basri, M. W., S. Ismail y N. Kamarudin.1996. The extend of biological control of rats with
barn owls in Malaysian oil palm plantations. Planter 72 (838): 5-8.
Bevanda, M., E. A. Fronhofer, M. Heurich, J. Mu¨ ller, and B. Reineking. 2015.
Landscape configuration is a majordeterminant of home range size variation.
Ecosphere 6(10):1-12
Blake, J. G., B. A. Loiselle, T. C. Moermond, D. J. Levey y J. S. Denslow. 1990.
Quantifying abundance of fruits for birds in tropical hábitats. Studies in Avia
Bilogy No. 13: 73-79.
Botequilha Leitão A., J. Miller,J. Ahern, y K. McGarigal 2006. Measuring Landscape.A
planner´s handbook. Island Press. USA.
Bruhl, C.A. y Eltz, T. 2010. Fuelling the biodiversity crisis: species loss of ground-welling
forest ants in oil palm plantations in Sabah, Malaysia (Borneo). Biodivers.
Conserv. 19: 519–529
Camacho C, Palacios S, Sa´ez P, Sa´nchez S, Potti J (2014) Human-Induced Changes in
Landscape Configuration Influence Individual Movement Routines: Lessons
from a Versatile, Highly Mobile Species. PLoS ONE 9(8): 1-9
Carrara, E. Víctor Arroyo-Rodríguez, Jorge H. Vega-Rivera, Jorge E. Schondube, Sandra
M. de Freitas y Lenore Fahrig. 2015. Impact of landscape composition and

104
configuration on forest specialist and generalist bird species in the fragmented
Lacandona rainforest, Mexico. Biological Conservation, 184: 117-126.
Chia, T. H., J. L. Limb y B. Buckle. 1995. Barn owl for rat control on oil palm plantations
do the work? Planter71 (828): 109-117.
Cousin, J. y R. D. Phillips. 2008. Habitat complexity explains species-specific occupancy
but not species richness in a Western Australian Woodland. Australian Journal
Zoology, 56: 95-102.
Crozier, G. E y G. J. Niemi. 2003. Using patch and landscape variables to model bird
abundance in a naturally heterogeneous landscape. Canadian Journal Zoology,
81: 441-452.
Diamond, J. M., & E. Mayr. 1976. Species–area relations for birds of the Solomon
archipelago. Proceedings of the National Academy of Sciences, USA 73:262–
266.
Duckett, J. E. 1991. Management of the Barn Owl (Tyto alba javanica) as a predator of
Rats in Oil Palm (Elaeis quineensis) plantations in Malaysia. Birds of Prey
Bulletin, 4: 11-24.
Fahrig. L. 2003. Effects of habitat fragmentation on biodiversity. Annual Review of
Ecology, Evolution, and Systematics, 34: 487-515.
Flohre, A., Fischer, C., Aavik, T., Bengtsson, J., Berendse, F., Bommarco, R., Ceryngier,
P., Clement, L. W., Dennis, C., Eggers, S., Emmerson, M., Geiger, F., Guerrero,
I., Hawro, V., Inchausti, P., Liira, J., Morales, M. B., Oñate, J. J., Pärt, T.,
Weisser, W. W., Winqvist, C., Thies, C. and Tscharntke, T. .2011, Agricultural
intensification and biodiversity partitioning in European landscapes comparing
plants, carabids, and birds. Ecological Applications, 21: 1772–1781.
Fedepalma. 2016. Anuario Estadístico. La Agroindustria de la palma de aceite en
Colombia y en el mundo 2011-2015. Fondo del Fomento Palmero.
Forman, R. T. T.y M. Godron. 1986. Landscape Ecology. Willey, USA,
Gallo-Cajiao, E. y C. J. Idrobo-Medina. 2004. Fragmentos de bosque y conservación de
aves: un estudio de caso en los Andes de Colombia. Memorias: Manejo de Fauna
silvestre en Amazonia y Latinoamérica, 178-185..
Gergel, S. E. y M. G. Turner. 2002. Learing landscape ecology A practical guide to
concepts and techniques. Springer. NY, USE.
Gibson, L., T. Ming Lee, L. Pin Koh, B. W. Brook, T. A. Gardner, J Barlow, C. A. Peres,
C. J. A. Bradshaw W. F. Laurance, T. E. Lovejoy y N. S. Sodhi. 2011. Primary

105
forest are irremplaceable for sustaining tropical biodiversity. Research Letter,
478, 378
González-Borrero, L. H. 2013. Anàlisis multitemporal de los cambios de cobertura de la
tierra e incidencia del cultivo de plama en el territorio del municipio
deVillanueva Casanare. Tesis de Maestría, Pontificia Universidad Javeriana.
Johnson, R. J., J. A. Jedlicka, J. E. Quinn y J. R. Brandle. 2011. Global perspective on
Birds in Agricultural Landscapes. En: Campbell, W. B. y S. L. Ortíz (Eds.).
Integrating agriculture, conservation and ecotourism: examples from the field.
Vol. 1. Springer-Verlag Berlin. Alemania
IDEAM. (2010). Leyenda Nacional de Coberturas de La Tierra: Metodologia Corine Land
Coifver Adaptada para Colombia. Escala 1:100.000. Bogota: IDEAM.
Karp, D.S., Rominger, A.J., Zook, J., Ranganathan, J., Ehrlich, P.R., Daily, G.C., 2012.
Intensive agriculture erodes b-diversity at large scales. Ecology Letters. 15, 963–
970.
Kirschbaum, K. 2004. "Alcedinidae" (On-line), Animal Diversity Web. Accessed April 27,
2013.http://animaldiversity.ummz.umich.edu/site/accounts/information/Alcedinid
ae.html
Lima S.L. y P. A. Zollner.1996. Towards a behavioral ecology of ecological landscapes.
Trends in Ecology and Evolution 11:131–135
Luskin, M. S. y M. D. Potts. 2011. Microclimate and habitat heterogeneity through the oil
palm lifecycle. Basic and Applied Ecology, 12: 540–551.
Minorta-Cely, V., & Rangel-Ch. J. O. (2014). Clima de la Orinoquía colombiana. En
Rangel-Ch, J. O. (Ed.), Colombia Diversidad Biótica XIV. La región de la
Orinoquía de Colombia (pp. 153-206). Universidad Nacional de Colombia –
Instituto de Ciencias Naturales. Bogotá, D.C., Colombia.
Mora Fernández C., C. Castellanos Castro, A. Cardona Cardozo, L. Pinzón Pérez & O.
Vargas. 2011. Historia de transformación del paisaje, de la cuenca baja del río
Pauto, Casanare (Colombia). En: Tomás León Sicard (Ed.) 2011. Mamíferos,
Reptiles y Ecosistemas del Bloque Cubiro (Casanare). Educación Ambiental para
la Conservación. Instituto de Estudios Ambientales, Universidad Nacional de
Colombia, Bogotá.
Morrison, M. L., C. J. Ralph, J. Vernes y J. R. Jehl, Jr. (Eds) 1990. Avian foraging:
theory, methods and applications. Studies in Avia Bilogy No. 13. The Cooper
Ornithological Society.
NASA. (12 de 05 de 2016). http://glovis.usgs.gov/. Obtenido de http://glovis.usgs.gov/: http://glovis.usgs.gov/

106
Olarte González, G. y Escovar Fadul, T. 2014. Presencia de león montuno (Puma
concolor) y las prácticas de conservación de su hábitat en un cultivo de palma de
aceite sostenible en Mapiripán, Meta. Palmas, 35(4), 31-37.
Pinzón Pinzón, F., Pardi Lacruz, M. S., Trejos Serrato, J., & Midori Saito, S. (Abril, 2009).
Uso de técnicas de percepción remota para la estimación del incremento de áreas
cultivadas de palma aceitosa y su relación con el conflicto armado en el
Departamento del Meta-Colombia. Anais En. XIV Simpósio Brasileiro de
Sensoramiento Remoto. Natal, Brasil.
Prescott, G. W., J. J. Gilory, T. Haugassen, C. A. Medina Uribe, W. A. Foster y D. P.
Edards. 2016. Reducing the impacts of Neotropical oil palm development on
functional diversity. Biological Conservation, 197: 139-145.
Quarles, W. 2011. Protecting raptors form rodenticides. Common Sense pest control
quarterly, Vol XXVII, No. 1-4: 3-9.
Ramachandran, V. y T: Ganesh. 2012. Habitat structure and its efects on birds
assemblages in the Kalakad-Mundanthurai Tiger Reserve (KMTR), India. Jornal
of the Bombay Natural History Society, 109(1&2): 87-95.
Rangel-Ch, O., y Minorta-Cely, V. 2014. Tipos de vegetación de la Orinoquía colombiana.
En Rangel-Ch, J. O. (Ed.), Colombia Diversidad Biótica XIV. La región de la
Orinoquía de Colombia. Universidad Nacional de Colombia – Instituto de
Ciencias Naturales. Bogotá, Colombia.
Rippstein, G., Escobar, G., & Motta, F. (Eds.). (2001). Agroecología y biodiversidad de
los Llanos Orientales de Colombia. Centro Internacional de Agricultura Tropical
(CIAT) # 322. Cali, Colombia. Roa, M. 1998. Ecología de la avifauna en cercas
vivas en un ecosistema fragmentado del piedemonte llanero. Importancia para su
conservación. Tesis Biología. Pontificia Universidad Javeriana.
Salgado-Negret, B. (ed). 2015. La ecología funcional como aproximación al estudio,
manejo y conservación de la biodiversidad: protocolos y aplicaciones. Instituto
de Investigación de RecursosBiológicos Alexander von Humboldt. Bogotá, D. C.
Colombia.
Tews J., Brose, U., Grimm, V., Tielbörger, K., .2004. Animal species diversity driven by
habitatheterogeneity/diversity: the importance of keystone structures. Journal of
Biogeography 31, 79–92.
Sánchez, S. 2000. Vertegrados silvestres registrados en una parcela de palma aceitera en
Tabasco, México. ASD. Oil Palm Papers, No. 20, 17-18.

107
Suk, H. Y. O.-S. Chungb, J.-Y. Leec, K.-Ch. Ohd, W.-O. Leee y G.-S, Jang. 2014.
Dynamic influence of patch size on occupancy of woodland birds. Animals Cells
and Systems, 18(1): 68-75.
Villanueva, C; Ibrahim, M; Casasola, F y Arguedas, R. 2005. Las cercas vivas en las
fincas ganaderas. Proyecto GEF y Oxford Forestry Institute.
Wiens, J. A. 1989. The ecology of bird comminities. Vol1. Foundations and patterns.
Cambirfge University Press. NY. 539p.

108
CAPÍTULO 5. CONCLUSIONES Y RECOMENDACIONES
Conclusiones
El cultivo de palma de aceite tuvo una baja diversidad de aves La composición y
riqueza de especies fue similar en palmas grandes, medianas y pequeñas, pero
última podría ofrecer algún tipo de beneficio para las cuatro especies que fueron
abundantes en esta clase de altura.
El cultivo de palma de aceite no representa una cobertura importante para las aves
en cuanto al uso de recursos, considerando que pocas especies del ensamble lo
utilizaron para la alimentación y la reproducción. Los cambios de altura de la
plantación tampoco demostraron un beneficio significativo para las aves del
ensamble y la escasa explotación de los recursos sugiere que este sistema reduce
las oportunidades para el establecimiento de poblaciones.
Los remanentes de vegetación son necesarios para el sostenimiento de las
poblaciones teniendo en cuenta que la riqueza de especies y abundancia de
individuos fue mayor en la primera cobertura.
Aves nectarívoras podrían estar usando la plantación como corredor de movimiento
más que para el consumo de alimento considerando que siempre se observaron
desplazándose.
Los bosques soportaron mayor cantidad de aves que la palma y la diversidad en
cada tipo de cobertura (bosque y palma) demuestra que la siembra comercial de
cultivo de palma modifica la comunidad de aves generando cambios en el tipo y la
abundancia de alimento y favorece especies adaptadas a ambientes intervenidos.

109
Las aves respondieron de manera diferente a las características del paisaje. Algunas
especies se benefician de las modificaciones de la vegetación, pero otro conjunto
de especies, como las que son de interior de bosque, se ven más afectadas
especialmente cuando aumenta el número de parches.
El monocultivo de palma de aceite condujo a la pérdida de diversidad en aves
residentes y migratorias y esto podrían ocasionar extinciones locales de
poblaciones de aves, considerando que es un cultivo permanente y la limitada
oferta de recursos que proporciona.
Analizar la comunidad de aves en entornos agrícolas a diferentes escalas, brinda
una noción más completa sobre la pérdida de la diversidad que se viene
presentando en estos entornos agrícola, lo cual debe ser atendido con la mayor
prontitud.

110
Recomendaciones
¿Podríamos aumentar la diversidad de aves que se pierde por el establecimiento de
cultivos comerciales de palma de aceite?, y cómo se lograría?
La heterogeneidad y la conectividad son aspectos fundamentales para aumetar la
diversidad de aves, esto se lograría implementando cultivos mixtos, permitiendo el
crecimiento de la vegetación en los bordes de los lotes y conformando lotes más pequeños
de diferentes tamaños.
Las formaciones vegetales originales que aún se mantienen son sitios importantes para
sostener la comunidad de aves, tanto para las especies asociadas al bosque como para las
que registramos dentro de la plantación. Es imperativo que estos fragmentos y/o
corredores de vegetación sean intangibles y se propicie la conectividad estructural entre
estos, en para que aporten los recursos necesarios (como alimento, sitios de reproducción y
sitios de refugio) que favorecen el soporte de especies y mantenimiento funcional de los
ecosistemas.
Es altamente recomendable evitar la tala de bosques para expandir el cultivo de palma,
para evitar que se siga reduciendo el hábitat de las aves en cuanto a disponibilidad y
calidad, evitar que siga aumentando el aislamiento de los parches, limitan o impiden los
procesos de movilidad, y aumenta los efectos de borde, conduciendo a la inestabilidad de
las poblaciones, a la extinción local de especies y se alteran los procesos ecológicos de la
comunidad de aves.
Conservar la vegetación de los cursos de agua es necesario. Esta franja de vegetación
reduce la erosión, la evaporación por la radiación directa sobre el espejo de agua, lo cual
conserva por más tiempo el caudal. Las fuentes de agua son un recurso necesario para
muchas aves acuáticas (garzas, ibis, cormoranes), además de que beneficia al palmicultor

111
para mantener el riego en la plantación, especialmente en época de pocas lluvias y se
mantiene el recurso para las demás actividades relacionadas al manejo de la plantación.
El área de estudio cuenta con humedales temporales, que se recargan en la época de lluvia.
Aunque estos no fueron objeto de estudio, las observaciones denotan que son sitos
importantes para las aves de la localidad, en especial del grupo de las acuáticas. Por tanto,
se recomienda que estos sistemas se preserven alejados de la intervención en cuanto a la
siembra de palmas y el paso de tractores que circulan dentro del cultivo.
Finalmente recomendamos que las renovaciones de lotes de palma se realicen en espacios
que ya hayan tenido siembra o ya hayan pasado por un proceso de limpieza del terreno
previamente para evitar la intervención y la remoción de la vegetación que aún se
mantiene.

112
SÍNTESIS GENERAL
En términos generales, el analisis de comunidades de aves en paisajes dominados por
cultivos de palma de aceite se hizo cuantificando la composición, estructura y función, en
dos niveles de organización: ecosistemas-comunidades y paisaje.
En términos de la composición se encontró que la plantación soportó un bajo número de
especies al ser comparadas con las coberturas de vegetación nativa. Sobre la estructura
analizada con los gremios de alimentación, identificamos que el número de gremios fue
muy similar entre el cultivo y el bosque, pero en este último caso los grupos tiene más
especies y en mayor cantidad de individuos. En cuanro a los aspectos funcionales,
encontramos que las aves no mostraron preferencia alguna por los recursos de usan dentro
del cultivo de palma. Desde el punto de vista de gremios, algunos mostraron una relación
más estrecha con el tipo de vegetación. A escala de paisaje pudimos concluir que los
miembros de la comunidad responden de manera diferente ante la composición y
configuración del paisaje, donde las epecies de interior de bosque podrían ser las más
afectadas porque tienen mayor nivel de especialización en los recursos de alimentación y
sitios de forrajeo.
Los monocultivos de palma a gran escala pueden estar funcionanado com una barrera de
movimiento o un espacio que no ofrece un hábitat adecuado para muchas especies de aves.
Este grupo podría tener un futuro incierto en la medida aumente el área de siembra y en
sigan disminuyendo rápidamente las coberturas nativas de la vegetación y los recursos
hídricos. Todo lo anterior nos brinda una visión más integral de los que está ocurriendo
con la diversidad de aves en el país con mayor cantidad de especies, pero con un alto grado
de vulnerabilidad.


















