
Effect of salmon plasma protein on Pacific whiting surimi ...
26
Effect of salmon plasma protein on Pacific whiting surimi gelation under various ohmic heating conditions Fowler, M. R., & Park, J. W. (2015). Effect of salmon plasma protein on Pacific whiting surimi gelation under various ohmic heating conditions. LWT-Food Science and Technology, 61(2), 309-315. doi:10.1016/j.lwt.2014.12.049 10.1016/j.lwt.2014.12.049 Elsevier Accepted Manuscript http://cdss.library.oregonstate.edu/sa-termsofuse
Transcript of Effect of salmon plasma protein on Pacific whiting surimi ...

Effect of salmon plasma protein on Pacific whiting surimi gelation under various ohmic heating conditions
Fowler, M. R., & Park, J. W. (2015). Effect of salmon plasma protein on Pacific whiting surimi gelation under various ohmic heating conditions. LWT-Food Science and Technology, 61(2), 309-315. doi:10.1016/j.lwt.2014.12.049
10.1016/j.lwt.2014.12.049
Elsevier
Accepted Manuscript
http://cdss.library.oregonstate.edu/sa-termsofuse

1
Effect of Salmon Plasma Protein on Pacific Whiting Surimi Gelation under Various 1
Ohmic Heating Conditions 2
3
Matthew R Fowler, Jae W Park 4
5
6
Oregon State University Seafood Research and Education Center 7
2001 Marine Dr Rm 253, Astoria, OR 97103, USA 8
9
Corresponding Author: 10
Jae W Park 11
(503) 325-4531 Ext #3 12
14
15
To be submitted to LWT- Food Sci and Tech 16
17

2
ABSTRACT: 18
The effect of salmon blood plasma (SPP) on the gelation of Pacific whiting surimi under 19
different ohmic heating conditions was investigated. SPP was found to significantly increase gel 20
strength in gels heated ohmically to and held at 60° for 30 min followed by heating ohmically to 21
90°C. SPP at a level of 1 g/100g was also found to increase gel strength in gels held at 25°C for 2 22
h prior to ohmic heating. This increase was not seen in gels where EDTA was added to inhibit 23
the activity of endogenous transglutaminase (ETG). SPP also created a more pronounced setting 24
effect as measured by dynamic rheology and SDS-PAGE. SPP was found to effectively inhibit 25
protease activity through TCA-soluble peptide analysis. Scanning electron microcopy revealed a 26
loosely arranged gel network caused by protease enzymes. It was reversed by the addition of 27
SPP as well as setting at 25oC due to ETG. 28
29
Keywords: salmon plasma, surimi gelation, ohmic heating, transglutaminase, protease 30
31

3
Highlights: 32
• Various ohmic heating rates effectively isolated enzyme activity in surimi 33
• Salmon plasma protein effectively inhibited protease in Pacific whiting surimi 34
• Salmon plasma protein increased gel strength during setting 35
• Salmon plasma protein is an effective inhibitor at low levels (0.5 g/100g or less) 36
37

4
Introduction 38
In the United States, surimi is made from two types of fish: Alaska pollock (AP) and Pacific 39
whiting (PW). Unlike AP, PW contains a high amount of protease enzymes that degrade the 40
quality of the surimi gel when heated slowly (Klesk, Yongsawatdigul, Park, Viratchakul, & 41
Virulhakul, 2000). The major protease enzymes found in PW are cathepsins B, H, and L. After 42
PW has gone through the rinsing step of surimi manufacturing, most of cathepsin B and almost 43
all of cathepsin H proteases are removed. However, cathepsin L, a protease associated with 44
myofibrillar proteins, is not removed during the washing process and was found to be the main 45
protease responsible for degradation of the surimi gel (An, Weerasinghe, Seymour, & 46
Morrissey, 1994). Cathepsin L is a heat activated cysteine protease, having an optimum 47
temperature of around 55-60°C (Seymour, Morrissey, Peters, & An, 1994; Visessanguan, 48
Benjakul, & An, 2003). Incubating PW surimi around this temperature range for 30 min before 49
heating to 90oC will result in a complete disappearance of the myosin heavy chain as well as an 50
inability to form a gel network (Morrissey, Wu, Lin, & An, 1993; Rawdkuen, Benjakul, 51
Visessanguan, & Lanier, 2007a). Surimi that is heated at a slow rate (such as in a water bath) 52
also suffers protoeolytic degradation (Yongsawatdigul, Park, Kolbe, Dagga, & Morrissey, 1995). 53
In the past, bovine blood plasma (BPP) was added to PW surimi as a protease inhibitor. This 54
practice was discontinued however due to public fear of Bovine Spongiform Encephalopathy 55
(BSE). Since then, BPP has been replaced by dried egg white (DEW), which contains mainly 56
serine protease inhibitors (Yongswatdigul, Hemung, & Choi, 2014). Since DEW is not as effective 57
as BPP at inhibiting cysteine proteases, such as cathepsin L (Yongswatdigul et al., 2014), finding 58
an alternative inhibitor that can be used at small concentrations would be beneficial. Blood 59

5
plasma from other sources, such as pork (Benjakul, Srivilai, & Visessanguan, 2001; 60
Visessanguan, Benjakul, & An, 2000) and chicken (Rawdkuen et al., 2007a; Rawdkuen, Lanier, 61
Visessanguan, & Benjakul, 2004, 2007b), have been found to be effective inhibitors of protease 62
enzymes found in surimi. 63
In addition to protease inhibitors, blood plasma also contains other proteins that may enhance 64
the gelation of surimi. Blood plasma proteins such as fibrinogen exhibit their own gelling ability 65
upon heating (Davila, Pares, Cuvelier, & Relkin, 2007). Also, blood plasma has been shown to 66
contain endogenous transglutaminase (ETG) enzymes (Lorand, 2007). ETG is a naturally 67
occurring enzyme in PW and other species of fish. ETG is a calcium dependent enzyme that 68
mediates covalent cross linking of myofibrillar proteins, resulting in a higher gel strength (An, 69
Peters, & Seymour, 1996). Since endogenous PW and AP ETG has an optimum temperature of 70
around 25°C, leaving surimi paste at room temperature for 1 or 2 hr before heating results in 71
stronger gels. This ETG mediated formation of non-disulfide covalent cross links before heating 72
is known as “setting”. Addition of calcium and calcium containing compounds to surimi has 73
been shown to increase the effect of setting (Lee & Park, 1998) and the addition of calcium 74
chelating compounds, such as EDTA, has been shown to completely inhibit setting (Kumazawa, 75
Numazawa, Seguro, & Motoki, 1995). Since blood plasma contains ETG, it may also contribute 76
to the setting phenomenon when added to surimi in sufficient amounts. 77
The activity of these two different types of enzymes (proteases and ETG), pose a problem when 78
evaluating the quality of PW surimi gel. Traditionally, surimi is heated in a water bath to 90°C 79
before conducting gel texture measurement. This slow heating allows for the activity of both 80
ETG (enhances gel strength) and proteases (lowers gel strength). Surimi crabstick, however, is 81

6
manufactured in a thin sheet under gas and/or steam heating, which quickly deactivates both 82
types of enzymes and does not allow for any activity beyond 75oC. Therefore, these testing 83
methods do not accurately assess the quality of the surimi seafood being produced in a thin 84
sheet under fast heating. Rapid heating methods, such as ohmic heating, allow for a better 85
assessment of surimi containing protease enzymes (Yongsawatdigul et al., 1995). The objective 86
of this study was to isolate the activities of both proteases and ETG at various heating rates 87
under ohmic heating, and evaluate the effect of salmon plasma protein (SPP) on the gelation of 88
PW surimi in combination with these enzymes. 89
90
2. Materials and Methods 91
2.1. Materials 92
Pacific whiting surimi (FA grade) without the addition of egg white, 2 months old, was obtained 93
from American Seafoods (Seattle, WA, USA) and kept at -30°C until used. Protein markers and 94
other electrophoresis chemicals were purchased from Bio-Rad Laboratories (Hercules, CA, 95
USA). All other chemicals used were of reagent grade. 96
97
2.2. Collection of salmon blood and preparation of plasma 98
Whole blood was collected at the Klaskanine Fish Hatchery (Astoria, OR, USA) from female 99
Chinook salmon immediately before roe collection. Blood was collected from bleeding fish 100
into bottles containing 3.8g/100mL sodium citrate (as an anti-coagulant), and gently mixed at a 101
ratio of 9:1 (mL:mL) blood to sodium citrate (Li, Lin, & Kim, 2008; Rawdkuen et al., 2007b). 102
Blood was kept on ice and transported back to the Oregon State Seafood Laboratory (Astoria, 103

7
OR, USA) where it was centrifuged for 15 min at 1,500 × g at 4°C using a Beckman J6-MI 104
centrifuge (Beckman Coulter, Fullerton, CA, USA). The supernatant was then lyophilized in a 105
Labconco freeze drier (Kansas City, MO, USA) and regarded as salmon plasma protein (SPP). SPP 106
was stored at -80°C until used. Samples were not stored longer than 3 months. 107
108
2.3. Surimi gel preparation 109
Paste and gels were prepared according the method of Poowakanjana, Mayer, and Park (2012) 110
with various heating methods. Partially thawed surimi was chopped at 1,800 rpm for 1 min 111
using a silent cutter (UM 5 universal, Stephan Machinery Corp, Columbus, OH, USA). After a 112
2g/100g addition of salt, surimi was chopped for an additional 1 min at 1,800 rpm. Moisture 113
content was then adjusted to 78g/100g using ice. At this time SPP (0, 0.5 or 1 g/100g) as well as 114
EDTA (0 or 0.1 g/100g) was added. A preliminary study conducted in our laboratory indicated 115
that 0.1 g/100g EDTA was sufficient to completely inhibit ETG activity in PW surimi (data not 116
shown). Following the addition of ice and other dry ingredients, surimi was chopped again for 1 117
min at 1,800 rpm. Chopping was then continued at 3,600 rpm under vacuum (40-60 kPa) for an 118
additional 3 min and a total chopping time of 6 min. Care was taken so that the final 119
temperature of the surimi paste was less than 15°C. Paste was packed in a polyethylene bag 120
and subjected to vacuum to remove any air pockets developed during packing. The paste was 121
then stuffed into a 15 cm x 3 cm nylon tube. 3 different heating methods were used: 1. Ohmic 122
(rapid) heating to 90°C at a voltage gradient of 12.62 V/cm with settings of 250 V and 10 kHz to 123
prevent the activity of both ETG and proteases (OH); 2. Ohmic heating to 60°C followed by 124
ohmically holding at 60°C for 30 min before ohmically heating to 90°C to prevent the activity of 125

8
ETG and maximize the activity of proteases (60/OH); and 3. Holding in a 25°C water bath for 2 h 126
(to maximize the activity of ETG) followed by ohmic heating to 90°C to prevent the activity of 127
proteases (25/OH). Two sausages were made per heating method. Following heating, gels were 128
placed in a plastic bag, submerged in ice water for 15 min, and stored overnight at 4°C. 129
130
2.4. Oscillatory dynamic measurement 131
Surimi paste was subjected to a temperature sweep using a CVO rheometer (Malvern 132
Instruments Ltd., Worcestershire, UK) using a cone (4° and 40 mm diameter) and plate 133
geometry with a gap of 150 μm. Surimi gels prepared as described above were thinly cut to a 134
thickness of 2 mm and subjected to a frequency sweep using parallel plate geometry (20 mm 135
diameter) and a gap of 1 mm. Surimi gel (3 cm diameter) was trimmed to 2 cm using a small 136
knife and moisture trap containing a moistened sponge was used to minimize drying of sample. 137
Temperature sweeps were conducted from 10 °C-90 °C at a heating rate of 2 °C/min at a fixed 138
frequency of 0.1 Hz. Frequency sweeps were conducted from 0.1 to 10 Hz at a fixed 139
temperature of 25 °C. A shear stress of 50 Pa, determined by stress sweep to be in the linear 140
viscoelastic region, was used. 141
142
2.5. Fracture gel evaluation 143
The day after heating, gels were removed from refrigerated storage and held at room 144
temperature for 2 h prior to testing. Gel samples were cut into 30 mm lengths and the breaking 145
force (g) and penetration distance (mm) were determined using a texture analyzer (TA-XT plus, 146

9
Texture Technologies Corp, NY, USA). Gels were punctured with a spherical probe (5 mm 147
diameter) at 1 mm/sec. 148
149
2.6. Color analysis 150
L*, a*, and b* values of surimi gels were determined from 30 mm samples using a Minolta 151
colorimeter (CR-310; Minolta Camera Co. Ltd., Osaka, Japan). The instrument was standardized 152
using a Minolta calibration plate and a Hunter Lab standard hitching file according to the 153
method of Park (1994). Whiteness was calculated using the equation L*-3b*. 154
155
2.7. Determination of TCA soluble peptides 156
Eighteen mL of 5 g/100mL trichloroacetic acid (TCA) was added to 2 g of sample followed by 157
homogenization for 2 min at 15,000 rpm using a Tissue Tearor homogenizer (Biospec Products 158
Inc., Bartlesville, OK, USA). Homogenate was then held at 4 °C for 1 h before centrifugation at 159
8,000 × g for 5 min using a Sorvall RC-5B centrifuge (DuPont Instruments, Newton, CT, USA). 160
The TCA-soluble peptide content of the supernatant was measured by the method of Lowry et 161
al. (1951) using tyrosine as a standard and expressed as μmol tyrosine/ g sample. Samples were 162
measured in triplicate. 163
164
2.8. SDS PAGE 165
Surimi gels were examined for protein patterns based on their molecular weight according to 166
the method of Laemmli (1970). Gels were solubilized in 5g/100mL sodium dodecyl sulfate at 90 167
°C according to the method of Morrissey et al. (1993). A 4g/100mL acrylamide stacking gel and 168

10
10g/100mL acrylamide separating gel were used. Gels were fixed and stained in 0.125g/100mL 169
Coomassie R-250 (Bio-Rad, Richmond, CA, USA), and de-stained in a 50mL/100mL methanol, 170
10mL/100mL acetic acid solution. Molecular weights of bands were determined by comparison 171
to a molecular weight standard (Protein Plus All Blue, Bio-Rad Laboratories, Hercules, CA, USA). 172
173
2.9. Scanning electron microscopy 174
Gels were cut into 2 mm x 2 mm sections and rinsed two times in distilled water for 30 min 175
prior to fixing for 2 hr in a 0.1 mol/L cacodylate buffer containing 2.5g/100mL glutaraldehyde 176
and 1g/100mL paraformaldehyde. Samples were then dried through serial acetone dilutions 177
(10, 30, 50, 70, 90 and twice in 100%) followed by critical point drying. Samples were then 178
coated with gold and palladium (40:60 ratio) and examined in a Quanta 600 FEG field emission 179
scanning electron microscope (FEI Inc., Hillsboro, OR). This microscopy work was done at the 180
Oregon State University Electron Microscope Facility (Corvallis, OR, USA). 181
182
2.10. Statistical analysis 183
Data were subjected to analysis of variance (ANOVA). Comparison of means was carried out by 184
Tukey test (Ramsey & Schafer, 2012). Statistical analysis was done by Sigma Plot software 185
package (Sigma Plot 12.5, Systat Software Inc, San Jose, CA, USA). Two batches were made for 186
each treatment and all experiments were repeated. 187
188
189
190

11
3. Results and Discussion 191
3.1. Oscillatory rheology 192
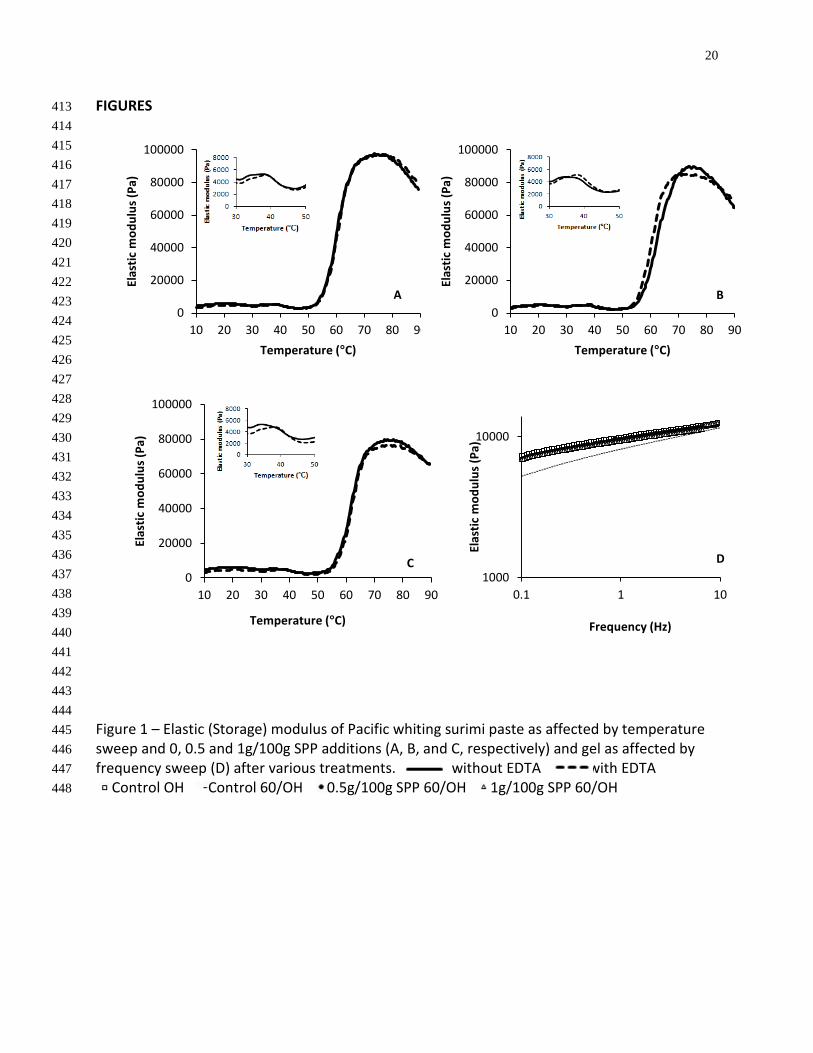
The maximum elastic modulus (G’) of surimi heated at 2°C/min decreased as more SPP was 193
added (Fig 1A-1C). Visessanguan et al. (2000) found that the addition of pork plasma protein to 194
Pacific whiting actomyosin also decreased G’. It was postulated that this was due to plasma 195
proteins having different thermal stabilities and gelation properties than fish muscle proteins. 196
The formation of two different gel networks (plasma protein and fish protein) with different 197
properties may lead to the observed decrease in G’. However, for pastes with 0.5g/100g SPP 198
(Fig 1B) and 1g/100g SPP (Fig 1C), the maximum G’ was higher for samples containing no EDTA 199
than samples containing EDTA. This difference was not observed in the control containing no 200
SPP. The difference between samples with and without EDTA is attributed to the setting effect 201
due to the action of ETG. This activity, however, may be offset by proteases active in the control 202
paste that are inhibited in the samples containing SPP. Therefore, the influence of setting 203
during heating at 2°C/min is more pronounced when SPP is added. In addition, ETG and calcium 204
present in SPP may serve to enhance the setting effect. Yin & Park (2014) found that adding 205
calcium containing nano scale fish bone to surimi also increased gel strength. 206
Rheological properties of the final gel were also evaluated by frequency sweep (Fig 1D). The G’ 207
of the control sample held at 60°C before ohmic heating showed a significantly higher 208
frequency dependence than the control sample ohmically heated directly to 90°C. Samples 209
containing both 0.5g/100g SPP and 1g/100g SPP and held at 60°C before ohmic heating showed 210
similar frequency dependence as the control heated to 90°C. G’ values of less cohesive gels 211
exhibit higher frequency dependency. Therefore, G’ increases at a higher rate as frequency 212

12
increases than is observed in a more cohesive gel (Picout & Ross-Murphy, 2003). These results 213
indicate that adding SPP to surimi to inhibit proteases lead to a less frequency dependent, more 214
cohesive gel network in the finished product. 215
216
3.2. Fracture gel evaluation 217
Both breaking force (Fig 2A) and penetration distance (Fig 2B) of 60/OH gels were greatly 218
increased by the addition of 0.5g/100g and 1g/100g SPP with or without the addition of EDTA 219
(P<0.05). There was no difference for this heating method between 0.5g/100g and 1g/100g 220
SPP. This heating condition maximized the action of endogenous proteases while minimizing 221
the activity of ETG to less than 10% (Park, Ooizumi, & Hunt, 2014). This indicates SPP at a level 222
of 0.5g/100g is sufficient for inhibiting protease activity and no added benefit in this regard is 223
seen from increasing concentration. 224
Compared to OH gels, 25/OH gels without EDTA showed a greater breaking force and 225
penetration distance (P<0.05). This heating method was favored by ETG and the activity of 226
protease enzymes was not significantly noted. Therefore, this increase of gel strength is due to 227
the setting phenomenon. In addition 25/OH gels without EDTA and containing 1g/100g SPP 228
showed the highest breaking force and penetration distance of all samples (P<0.05). It may be 229
postulated that transglutaminase present in SPP contributed additionally to enzyme-mediated 230
covalent cross linking during settings. Both OH and 25/OH gels containing no SPP decreased 231
moderately and significantly, respectively, in breaking force and penetration distance when 232
EDTA was added (P<0.05). EDTA chelates calcium, which in addition to being a cofactor for ETG 233
may also play other roles in gelation (Lee & Park, 1998). However, both OH and 25/OH gels 234

13
containing EDTA showed a higher breaking force and penetration distance with the addition of 235
SPP as compared to the control (P<0.05), indicating a significant role of SPP as a gel enhancer. 236
In addition to transglutaminase being present in SPP, it may also be a source of additional 237
calcium (Heaton & Pomare, 1974; Maye, Keaton, Hurst, & Habener, 1979), leading to greater 238
gel strength. 239
240
3.3 Color gel analysis 241
Whiteness decreased markedly as SPP concentration increased (P<0.05) (Fig 3D). Increase in 242
SPP was also associated with a lower L* value (less lightness) (Fig 3A), more negative a* value 243
(more redness) (Fig 3B) and a higher b* value (more yellowness) (Fig 3C). The decrease in color 244
quality is due to hemolysis in the blood plasma. When blood is collected and plasma is 245
processed, damage to the red blood cells can result in plasma containing a pinkish hue as 246
opposed to a straw yellow color (Field, Elvehjem, & Juday, 1943; Li et al., 2008). In addition, as 247
salmon is a cold water species, blood plasma may exhibit lower thermal stability and therefore 248
be more susceptible to hemolysis than blood plasma from mammals. This issue may be partly 249
intervened by the fact that SPP is needed only at very low levels (maximum of 0.5g/100g as 250
shown by the results in Fig 2) in order to prevent proteolysis in PW surimi. It is suggested 251
however that further studies be conducted to determine collection and processing methods 252
that will reduce hemolysis. In addition, 60/OH gels showed greater L* values, less negative a* 253
values, and higher b* values than other gels (P<0.05). However, overall whiteness was not 254
affected by heating conditions. 255
256

14
257
3.4 TCA soluble peptide content 258
At a TCA concentration of 5g/100mL, all proteins except for small oligopeptides are insoluble 259
(Yvon, Chabanet, & Pelissier, 1989). Since small peptides are the result of the action of 260
endogenous proteases on PW muscle, the protein content in the 5g/100mL TCA supernatant 261
after centrifugation is related to total protease activity in the surimi. Control 60/OH gels with or 262
without EDTA had significantly higher TCA soluble peptide content compared to other gels 263
(P<0.05) (Fig 4). The addition of SPP to these gels showed a significant decrease in TCA soluble 264
peptide content (P<0.05). There was no difference between 0.5g/100g and 1g/100g SPP. 265
Besides 60/OH gels, no difference was seen in TCA soluble peptide content between other 266
treatments and 60/OH gels containing SPP. This confirms the fact that an SPP concentration of 267
0.5g/100g is sufficient to inhibit protease activity and no additional inhibition is seen from 268
increasing concentrations. In addition, this confirms that the fast heating treatments (OH and 269
25/OH) effectively eliminate protease activity. 270
271
3.5 SDS PAGE 272
For OH gels, there was no discernible difference in protein pattern between gels prepared 273
without EDTA (Fig 5A) and gels with EDTA (Fig 5B). Among this group, SPP also had no 274
noticeable effect on protein pattern. This is because the OH treatment eliminated the activity of 275
both protease enzymes and ETG, therefore there was no setting effect or degradation of the 276
myosin heavy chain. For the 25/OH samples, however, a protein band is visible in the high 277
molecular weight range (>250 kDa) for gels treated without EDTA that is not present for gels 278

15
treated with EDTA (dotted line in Fig 5A). Yin and Park (2014) found that ETG-mediated cross 279
linking in surimi led to the appearance of high molecular weight bands. This band is due to the 280
crosslinking of myosin heavy chain proteins, leading to higher molecular weight proteins 281
(Kamath, Lanier, Foegeding, & Hamann, 1992). The disappearance of this band in the EDTA 282
samples confirms the inhibition of ETG by EDTA. The high molecular weight band is also darker 283
for 1g/100g SPP than the control, indicating that SPP might have played a role in the setting 284
phenomenon as transglutaminase is one of various proteins present in blood plasma (Folk, 285
1980). 286
The 60/OH group showed no difference between samples with and without EDTA. In this 287
treatment, only protease enzymes are active and the action of ETG was eliminated. This heating 288
treatment completely destroyed the myosin heavy chain band in the control samples due to the 289
activity of proteases. When SPP was added at 0.5g/100g however, the myosin heavy chain band 290
remained intact. Increasing SPP concentration did not affect the intensity of the myosin heavy 291
chain band. This confirms that 0.5g/100g SPP is sufficient to inhibit proteases and prevent 292
myosin heavy chain degradation. 293
294
3.6 Scanning electron microscopy 295
The 60/OH gel with no SPP added showed the greatest number of voids and the least compact 296
structure among the samples tested (Fig 6B). This is consistent with the results from the 297
puncture test as well as the TCA-soluble peptides. This confirms that protease enzymes active 298
at this temperature serve to break up and weaken the gel structure. 60/OH gel with 1g/100g 299
SPP (Fig 6E) showed a significantly more orderly and continuous gel structure with less voids 300

16
than the 60/OH gel with no SPP present, indicating effective inhibition of protease enzymes. 301
However, there were a greater number of voids present in this gel than in the OH gels (Fig 6A 302
and 6D), indicating residual protease activity. 25/OH gels without and with 1g/100g SPP (Fig 6C 303
and 6F, respectively) had a more compact and continuous structure compared to OH gels 304
without and with 1g/100g SPP (Fig 6A and 6D, respectively). This is due to the addition of extra 305
covalent cross linking in the gel structure due to the action of ETG. In addition, 25/OH gel with 306
1g/100g SPP had a more compact structure with less voids than the 25/OH gel without SPP. 307
This may be due to transglutaminase present in SPP in addition to ETG of surimi, leading to 308
additional cross linking during setting. 25/OH sample containing no SPP and 0.1g/100g EDTA 309
(Fig 6G), had a greater number of voids than the OH samples. The purpose of EDTA addition 310
was to chelate calcium in order to prevent the activity of ETG. However, these results indicate 311
that calcium may play other roles in gelation in addition to being a cofactor for ETG. 312
313
4. Conclusion 314
SPP was found to effectively inhibit endogenous proteases in PW surimi at levels as low as 315
0.5g/100g. Higher concentrations of SPP may also aid in transglutaminase-mediated gel setting, 316
leading to an increase in gel strength. However, increasing SPP concentrations also led to a 317
decrease in the elastic modulus as well as a decrease in whiteness. This balance must be kept in 318
mind when formulating surimi with SPP. Holding at 25°C before ohmic heating was found to 319
optimize ETG activity while controlling protease activity. Rapidly heating to and holding at 60°C 320
before ohmic heating to 90°C was found to optimize protease activity while minimizing ETG 321
activity. 322

17
323
5. Acknowledgment 324
This research was supported by a scholarship (2013 NPRD Graduate Research Award) from the 325
North Pacific Research Board (Anchorage, AK). We are thankful for Teresa Sawyer of the 326
Oregon State University Electron Microscope Facility (Corvallis, OR) for her assistance with the 327
scanning electron microscopy. 328
329
REFERENCES 330
An, H., Peters, M. Y., & Seymour, T. A. (1996). Roles of endogenous enzymes in surimi gelation. 331 Trends in Food Science & Technology, 7(10), 321-327. 332
An, H., Weerasinghe, V., Seymour, T. A., & Morrissey, M. T. (1994). Cathepsin Degradation of 333 Pacific Whiting Surimi Proteins. Journal of Food Science, 59(5), 1013-1017. 334
Benjakul, S., Srivilai, C., & Visessanguan, W. (2001). Porcine plasma protein as proteinase 335 inhibitor in bigeye snapper (Priacanthus tayenus) muscle and surimi. Journal of the 336 Science of Food and Agriculture, 81(10), 1039-1046. 337
Davila, E., Pares, D., Cuvelier, G. r., & Relkin, P. (2007). Heat-induced gelation of porcine blood 338 plasma proteins as affected by pH. Meat Science, 76(2), 216-225. 339
Field, J. B., Elvehjem, C. A., & Juday, C. (1943). A study of the blood constituents of carp and 340 trout. Journal of Biological Chemistry, 148(2), 261-269. 341
Folk, J. E. (1980). Transglutaminases. Annual Review of Biochemistry, 49(1), 517-531. 342 Heaton, K. W., & Pomare, E. W. (1974). Effect of bran on blood lipids and calcium. The Lancet, 343
303(7846), 49-50. 344 Kamath, G. G., Lanier, T. C., Foegeding, E. A., & Hamann, D. D. (1992). Nondisulfide covalent 345
cross linking of myosin heavy chain in setting of Alaska Pollock and Atlantic Croaker 346 surimi. Journal of Food Biochemistry, 16(3), 151-172. 347
Klesk, K., Yongsawatdigul, J., Park, J. W., Viratchakul, S., & Virulhakul, P. (2000). Gel Forming 348 Ability of Tropical Tilapia Surimi as Compared with Alaska Pollock and Pacific Whiting 349 Surimi. Journal of Aquatic Food Product Technology, 9(3), 91-104. 350
Kumazawa, Y., Numazawa, T., Seguro, K., & Motoki, M. (1995). Suppression of surimi gel setting 351 by transglutaminase inhibitors. Journal of Food Science, 60(4), 715-717. 352
Laemmli, U. K. (1970). Cleavage of Structural Proteins during the Assembly of the Head of 353 Bacteriophage T4. Nature, 227(5259), 680-685. 354
Lee, N., & Park, J. W. (1998). Calcium compounds to improve gel functionality of Pacific whiting 355 and Alaska pollock surimi. Journal of Food Science, 63(6), 969-974. 356

18
Li, D. K., Lin, H., & Kim, S. M. (2008). Effect of rainbow trout (Oncorhynchus mykiss) plasma 357 protein on the gelation of Alaska pollock (Theragra chalcogramma) Surimi. Journal of 358 Food Science, 73(4), C227-C234. 359
Lorand, L. (2007). Crosslinks in blood: transglutaminase and beyond. The FASEB Journal, 21(8), 360 1627-1632. 361
Lowry, O. H., Rosebrough, N. J., Farr, A. L., & Randall, R. J. (1951). Protein measurement with 362 the Folin phenol reagent. The Journal Of Biological Chemistry, 193(1), 265-275. 363
Maye, G. P., Keaton, J. A., Hurst, J. G., & Habener, J. F. (1979). Effects of plasma calcium 364 concentration on the relative proportion of hormone and carboxyl fragments in 365 parathyroid venous blood. Endocrinology, 104(6), 1778-1784. 366
Morrissey, M. T., Wu, J. W., Lin, D., & An, H. (1993). Protease Inhibitor Effects on Torsion 367 Measurements and Autolysis of Pacific Whiting Surimi. Journal of Food Science, 58(5), 368 1050-1054. 369
Park, J. W. (1994). Functional protein additives in surimi gels. Journal of Food Science, 59(3), 370 525-527. 371
Park, J. W., Ooizumi, T., & Hunt, A. L. (2014). Ingredient technology for surimi and surimi 372 seafood. In J. W. Park (Ed.), Surimi and Surimi Seafood (3 ed.). Boca Raton, FL: CRC Press. 373
Picout, D. R., & Ross-Murphy, S. B. (2003). Rheology of biopolymer solutions and gels. 374 TheScientificWorld Journal, 3, 105-121. 375
Poowakanjana, S., Mayer, S. G., & Park, J. W. (2012). Optimum chopping conditions for Alaska 376 pollock, Pacific whiting, and threadfin bream surimi paste and gel based on rheological 377 and Raman spectroscopic analysis. Journal of Food Science, 77(4), E88-E97. 378
Ramsey, F., & Schafer, D. (2012). The Statistical Sleuth: A Course in Methods of Data Analysis (3 379 ed.). Stamford: Cengage Learning. 380
Rawdkuen, S., Benjakul, S., Visessanguan, W., & Lanier, T. C. (2007a). Effect of cysteine 381 proteinase inhibitor containing fraction from chicken plasma on autolysis and gelation of 382 Pacific whiting surimi. Food Hydrocolloids, 21(7), 1209-1216. 383
Rawdkuen, S., Lanier, T. C., Visessanguan, W., & Benjakul, S. (2004). Chicken plasma protein: 384 Proteinase inhibitory activity and its effect on surimi gel properties. Food Research 385 International, 37(2), 156-165. 386
Rawdkuen, S., Lanier, T. C., Visessanguan, W., & Benjakul, S. (2007b). Cysteine proteinase 387 inhibitor from chicken plasma: Fractionation, characterization and autolysis inhibition of 388 fish myofibrillar proteins Food Chemistry, 101(4), 1647-1657. 389
Seymour, T. A., Morrissey, M. T., Peters, M. Y., & An, H. (1994). Purification and characterization 390 of Pacific whiting proteases. Journal of Agricultural and Food Chemistry, 42(11), 2421-391 2427. 392
Visessanguan, W., Benjakul, S., & An, H. (2000). Porcine plasma proteins as a surimi protease 393 inhibitor: effects on actomyosin gelation. Journal of Food Science, 65(4), 607-611. 394
Visessanguan, W., Benjakul, S., & An, H. (2003). Purification and characterization of cathepsin L 395 in arrowtooth flounder (Atheresthes stomias) muscle. Comparative Biochemistry and 396 Physiology Part B: Biochemistry and Molecular Biology, 134(3), 477-487. 397
Yin, T., & Park, J. W. (2014). Effects of nano-scaled fish bone on the gelation properties of 398 Alaska pollock surimi. Food Chemistry, 150, 463-468. 399

19
Yongsawatdigul, J., Park, J. W., Kolbe, E., Dagga, Y., & Morrissey, M. T. (1995). Ohmic heating 400 maximizes gel functionality of Pacific whiting surimi. Journal of Food Science, 60(1), 10-401 14. 402
Yongswatdigul, J., Hemung, B. O., & Choi, Y. J. (2014). Proteolytic Enzymes and Control in 403 Surimi. In J. W. Park (Ed.), Surimi and Surimi Seafood (3 ed., pp. 141-167). Baco Raton, 404 FL: Taylor and Francis. 405
Yvon, M., Chabanet, C., & Pelissier, J. P. (1989). Solubility of peptides in trichloroacetic acid 406 (TCA) solutions hypothesis on the precipitation mechanism. International Journal of 407 Peptide and Protein Research, 34(3), 166-176. 408
409 410 411
412
20
1000
10000
0.1 1 10
Elas
tic m
odul
us (P
a)
Frequency (Hz)
0
20000
40000
60000
80000
100000
10 20 30 40 50 60 70 80 90
Elas
tic m
odul
us (P
a)
Temperature (°C)
0
20000
40000
60000
80000
100000
10 20 30 40 50 60 70 80 90
Elas
tic m
odul
us (P
a)
Temperature (°C)
0
20000
40000
60000
80000
100000
10 20 30 40 50 60 70 80 90
Elas
tic m
odul
us (P
a)
Temperature (°C)
FIGURES 413 414 415 416 417 418 419 420 421 422 423 424 425 426 427 428 429 430 431 432 433 434 435 436 437 438 439 440 441 442 443 444 Figure 1 – Elastic (Storage) modulus of Pacific whiting surimi paste as affected by temperature 445 sweep and 0, 0.5 and 1g/100g SPP additions (A, B, and C, respectively) and gel as affected by 446 frequency sweep (D) after various treatments. without EDTA with EDTA 447 Control OH Control 60/OH 0.5g/100g SPP 60/OH 1g/100g SPP 60/OH 448
A B
C D

21
0100200300400500600
Brea
king
For
ce (G
)
02468
101214161820
Pene
trat
ion
Dist
ance
(mm
)
449
450
451
452
453
454
455
456 457 Figure 2 - Breaking force (A) and penetration distance (B) of Pacific whiting surimi gel as 458 affected by SPP concentration, EDTA, and heating method. Error bars represent the standard 459 deviation of 6 determinations. control 0.5g/100g SPP 1g/100g SPP 460 461
462
463
464
465
466
467
468
469
470
471
472
473
B A

22
60
65
70
75
80
85L*
0
2
4
6
8
b*
0
20
40
60
80
Whi
tene
ss (L
*-3b
*)
-4
-3
-2
-1
0
a*
474
475
476
477
478
479 480 481 482
483 484 485 486 487 488 489 490 491 492 493 494 Figure 3 - L* (A), a* (B), b* (C), and whiteness (D) values of Pacific whiting surimi gels as 495 affected by SPP concentration, EDTA, and heating methods. Error bars represent the standard 496 deviation of 6 determinations. 497 498 499 500 501 502 503 504 505 506 507 508 509 510
A B
C D

23
511 Figure 4 - TCA soluble peptide contents in Pacific whiting surimi gel as affected by SPP 512 concentration, EDTA, and heating method. Error bars represent the standard deviation of 3 513 determinations. 514 515 516 517 518 519 520 521 522 523 524 525 526 527 528 529 530 531 532 533 534 535 536 537 538 539 540
0
0.5
1
1.5
2μm
ol ty
rosi
ne/g
sam
ple

24
541 542 543
544 545 Figure 5 - SDS PAGE pattern of surimi gels without (A) and with (B) 0.1g EDTA/100g as affected 546 by SPP concentration and heating methods. MW=molecular weight marker, C=control, 547 0.5=0.5g/100g SPP, 1=1g/100g SPP, MHC=myosin heavy chain, Ac=Actin, 20-250=molecular 548 weight in kDa. 549 550 551 552 553 554 555 556 557 558 559 560 561 562 563 564 565 566 567 568 569 570 571 572 573 574

25
575 576 OH 60/OH 25/OH 577 578 579 580 581 582 Control 583 584 585 586 587 588 589 1g/100g 590 SPP 591 592 593 594 595 596 Control 597 EDTA 598 599 600 601 602 603 Figure 6 – Scanning electron microscope image of surimi gels. A = Ohmic heating with no SPP 604 added; B = 60°C holding for 30 min followed by ohmic heating with no SPP added; C = 25°C 605 holding for 2 hr followed by ohmic heating with no SPP added; D = Ohmic heating with 1g/100g 606 SPP added; E = 60°C holding for 30 min followed by ohmic heating with 1g/100g SPP added; F = 607 25°C holding for 2 hr followed by ohmic heating with 1g/100g SPP added; G = 25°C holding for 2 608 hr followed by ohmic heating with no SPP added and 0.1g EDTA/100g. Magnification = 10,000x. 609 610 611 612 613 614 615 616
A B C
D E F
G


















