
Detection of genomic alterations in carcinogen-induced mouse liver tumors by DNA fingerprint...
5
MOLECULAR CARCINOGENESIS 3:330-334 (1990) Detection of Genomic Alterations in Carcinogen-Induced Mouse Liver Tumors by DNA Fingerprint Analysis Oliver Muller, Angel Alonso, Albrecht Buchrnann, Richard Bauer-Hofrnann, Karl-Walter Bock, and Michael Schwarz' German Cancer Research Center; Institute of Experimental Pathology, im Neuenheimer Feld 280, 6900 Heidelberg, Germany (OM, AA, AB, RBH, MS) and Institute of Toxicology, University of Tubingen, Wilhelmstr 56, 7400 Tubingen, Germany (KWB) DNA fingerprint analysis was used to study structural abnormalities in the genome of mouse liver tumor cells. Liver tumors were induced in three strains of mice, namely C57BU6J, C3H/He and B6C3F1, by a single injection of 20 pg/g body wt. diethylnitrosamine on day 15 after birth. DNA from liver tumors was digested with Hinfl restriction enzyme and hybridized on Southern blots with wild-type bacteriophage M13 DNA as probe. The resulting fingerprints of tumor DNA were compared with those of DNA from normal liver tissue. Genomic aberrations were detected in two out of 68 tumors analyzed, one stemming from a C57BU6J and the other from a C3H/He mouse. Key words: Hepatocarcinogenesis, liver tumors, DNA fingerprinting INTRODUCTION According to the present understanding of the carcino- genic process, the transition of a normal cell to a tumor cell is mediated by sequential genetic alterations in critical cell regulatory genes such as proto-oncogenes or tumor suppressor genes that may lead to either the activation or the inactivation of these genes [reviewed in 11. Thus, point mutations in proto-oncogenes resulting in a gene product with differing activity have been reported to occur in a large variety of human and experimental tumors [reviewed in 21. Activation of proto-oncogenes by gene amplifica- tion or translocation is also frequently seen in cancerous tissues (31, as is the loss of function of tumor suppressor genes in hereditary and sporadic cases of human cancers [I]. This latter phenomenon often finds its molecular cor- relate in structural changes of the genome ranging from specific inactivating point mutations to loss of entire chro- mosomes [4-71. Evidence for structural alterations in the genome of tu- mor cells has classically been accumulated from cytoge- netic studies [5,8,9]. More recently, tumor-associated chromosomal aberrations have been described that were detected by restriction fragment length polymorphism (RFLP) analyses. In these experiments, single-copy DNA se- quences of known chromosomal location were used as "informative" probes for Southern blot hybridization of tu- mor DNA digested with different restriction enzymes [ 10- 121. The power of this procedure lies in its ability to allow the direct assignment of a genomic abnormality to a cer- tain chromosomal location. The disadvantage, however, is that a large number of different "informative" DNA probes have to be applied to score for unknown genomic alter- ations within tumor cells. Alternatively, oligonucleotide probes specific for nonecotropic endogenous murine leu- kemia viruses present in inbred strains of mice [I31 could be used for these analyses. With these probes, approxi- mately 50 such endogenous murine viruses were recently identified in different inbred mouse strains and were for the most part assigned to distinct chromosomal loca- tions [ 141. In the present study we used DNA fingerprinting in which DNA from the bacteriophage MI 3 was employed as a specific hybridization probe to analyze genomic ab- normalities in rodent liver tumors. The recently developed technique of DNA fingerprinting essentially makes use of the fact that genomic DNA harbors repetitive sequences that are dispersed over the entire genome that, upon di- gestion of DNA with appropriate restriction enzymes, in turn give rise to well-defined banding patterns of specific probes hybridized on Southern blots. So far, DNA finger- print analyses have been used predominantly in paternity testing, in forensic analyses, and in studies on the charac- terization of cell line contamination [I 5-1 71. The advan- tage of using M I 3 minisatellite DNA as probe for DNA fingerprinting is that the recognition sequence is highly repetitive in the mouse genome, thus enabling the screen- ing for structural alterations in a comparatively large frac- tion of the genome. The obvious disadvantage of this and similar probes, however, lies in the lack of information on the chromosomal location of their recognition sequences. A few studies have been reported in which genomic ab- normalities in tumor cells have been analyzed by DNA fin- gerprinting using human minisatellites [ 151 as hybridization probes [18-201. Wild-type MI3 phage DNA has been widely used for fingerprinting of mammalian DNA [21-241 'Corresponding author: German Cancer Research Center, Institute of Experimental Pathology, Im Neuenheimer Feld 280, 6900 Heidel- berg, Germany. Abbreviations: DEN, diethylnitrosamine; RFLP, restriction fragment length polymorphism, SDS, sodium dodecyl sulfate; SSC, standard saline citrate. CC) 1990 WILEY-LISS, INC
-
Upload
oliver-mueller -
Category
Documents
-
view
212 -
download
0
Transcript of Detection of genomic alterations in carcinogen-induced mouse liver tumors by DNA fingerprint...

MOLECULAR CARCINOGENESIS 3:330-334 (1990)
Detection of Genomic Alterations in Carcinogen-Induced Mouse Liver Tumors by DNA Fingerprint Analysis Oliver Muller, Angel Alonso, Albrecht Buchrnann, Richard Bauer-Hofrnann, Karl-Walter Bock, and Michael Schwarz'
German Cancer Research Center; Institute of Experimental Pathology, im Neuenheimer Feld 280, 6900 Heidelberg, Germany (OM, AA, AB, RBH, MS) and Institute of Toxicology, University of Tubingen, Wilhelmstr 56, 7400 Tubingen, Germany (KWB)
DNA fingerprint analysis was used t o study structural abnormalities in the genome o f mouse liver tumor cells. Liver tumors were induced in three strains of mice, namely C57BU6J, C3H/He and B6C3F1, by a single injection of 20 pg/g body wt. diethylnitrosamine o n day 15 after birth. DNA from liver tumors was digested w i th Hinfl restriction enzyme and hybridized on Southern blots with wild-type bacteriophage M13 DNA as probe. The resulting fingerprints of tumor DNA were compared with those of DNA from normal liver tissue. Genomic aberrations were detected in t w o ou t o f 68 tumors analyzed, one stemming from a C57BU6J and the other f rom a C3H/He mouse.
Key words: Hepatocarcinogenesis, liver tumors, DNA fingerprinting
INTRODUCTION
According to the present understanding of the carcino- genic process, the transition of a normal cell to a tumor cell is mediated by sequential genetic alterations in critical cell regulatory genes such as proto-oncogenes or tumor suppressor genes that may lead to either the activation or the inactivation of these genes [reviewed in 11. Thus, point mutations in proto-oncogenes resulting in a gene product with differing activity have been reported to occur in a large variety of human and experimental tumors [reviewed in 21. Activation of proto-oncogenes by gene amplifica- tion or translocation is also frequently seen in cancerous tissues (31, as is the loss of function of tumor suppressor genes in hereditary and sporadic cases of human cancers [I]. This latter phenomenon often finds its molecular cor- relate in structural changes of the genome ranging from specific inactivating point mutations to loss of entire chro- mosomes [4-71.
Evidence for structural alterations in the genome of tu- mor cells has classically been accumulated from cytoge- netic studies [5,8,9]. More recently, tumor-associated chromosomal aberrations have been described that were detected by restriction fragment length polymorphism (RFLP) analyses. In these experiments, single-copy DNA se- quences of known chromosomal location were used as "informative" probes for Southern blot hybridization of tu- mor DNA digested with different restriction enzymes [ 10- 121. The power of this procedure lies in its ability to allow the direct assignment of a genomic abnormality to a cer- tain chromosomal location. The disadvantage, however, is that a large number of different "informative" DNA probes have to be applied to score for unknown genomic alter- ations within tumor cells. Alternatively, oligonucleotide probes specific for nonecotropic endogenous murine leu- kemia viruses present in inbred strains of mice [I31 could be used for these analyses. With these probes, approxi-
mately 50 such endogenous murine viruses were recently identified in different inbred mouse strains and were for the most part assigned to distinct chromosomal loca- tions [ 141.
In the present study we used DNA fingerprinting in which DNA from the bacteriophage M I 3 was employed as a specific hybridization probe to analyze genomic ab- normalities in rodent liver tumors. The recently developed technique of DNA fingerprinting essentially makes use of the fact that genomic DNA harbors repetitive sequences that are dispersed over the entire genome that, upon di- gestion of DNA with appropriate restriction enzymes, in turn give rise to well-defined banding patterns of specific probes hybridized on Southern blots. So far, DNA finger- print analyses have been used predominantly in paternity testing, in forensic analyses, and in studies on the charac- terization of cell line contamination [ I 5-1 71. The advan- tage of using M I 3 minisatellite DNA as probe for DNA fingerprinting is that the recognition sequence is highly repetitive in the mouse genome, thus enabling the screen- ing for structural alterations in a comparatively large frac- tion of the genome. The obvious disadvantage of this and similar probes, however, lies in the lack of information on the chromosomal location of their recognition sequences.
A few studies have been reported in which genomic ab- normalities in tumor cells have been analyzed by DNA fin- gerprinting using human minisatellites [ 151 as hybridization probes [18-201. Wild-type MI3 phage DNA has been widely used for fingerprinting of mammalian DNA [21-241
'Corresponding author: German Cancer Research Center, Institute of Experimental Pathology, Im Neuenheimer Feld 280, 6900 Heidel- berg, Germany.
Abbreviations: DEN, diethylnitrosamine; RFLP, restriction fragment length polymorphism, SDS, sodium dodecyl sulfate; SSC, standard saline citrate.
CC) 1990 WILEY-LISS, INC

CENOMICALTERATIONS IN MOUSE LIVER TUMORS 33 1
and has also been demonstrated to be suitable for detect- ing structural changes in the genome of human tumor cells (25,261.
MATERIALS AND METHODS Source of DNA
C3H/He and C57BU6J mice were obtained from the Zentralinstitut fur Versuchstierkunde (Hannover, Germany) and mated in our laboratory for breeding of C3H/He, C57BU6J, and B6C3F1 (C3H x C57BL) mice. Liver tumors were induced in male offspring by a single i.p. injection of diethylnitrosamine (DEN; 20 pg/g body wt.) on day 15 af- ter birth. Animals were killed sequentially thereafter. In ac- cordance with the literature [27], the onset of liver nodule and tumor formation was characteristically different be- tween the three strains. In C3H/He and B6C3F1 mice, small nodules (diameters <2 mm) started to appear on the sur- face of the livers at 17 to 23 wk after treatment and liver tumors with diameters larger than 5 mm were observed after 29 wk. In contrast, in C57BU6J mice the tumor re- sponse was considerably delayed, with small liver nodules being present a t 36 wk and large liver tumors at 45 wk after DEN treatment. A total of 68 tumors were analyzed in this study, 22 from C3H/He, 24 from C57BU6J. and 22 from B6C3F1 mice. The average tumor size was 6 mm, ranging from 3 to 15 mm. To minimize contamination of tumor material by normal liver tissue, only tumors that were well circumscribed were included. In addition, tis- sue samples from the surrounding normal liver were COIL lected. High molecular weight DNA was extracted as described by Krieg et al. [28].
DNA Fingerprinting
Equivalent amounts (1 0 pg) of DNA from normal liver and from liver tumors were digested by several different restriction enzymes according to the recommendations of the manufacturer (Boehringer Mannheim, Mannheim, Germany). The following enzymes were tested for their suitability in producing "informative" fingerprints: BamHI, Bgll, EcoRI, Haell, Haelll, Hindlll, Hinfl, Hpal, Hpall, Mspl, Ndell, Taql, Sau3A, and Xhol. Digested DNA was separated through a 0.6% agarose gel of 20-cm length. Electropho- resis was performed overnight at 2.4 V/cm until fragments of approximately 0.6 kb each ran off the gel. The DNA was then transferred onto Bio Trace RP nylon membranes (Gelman Sciences, Dreieich, Germany) and baked for 2 h at 80°C. Fifty nanograms of bacteriophage M 13mp8 DNA (Pharmacia, Uppsala, Sweden) were labeled by random priming to a specific activity of 1 O8 to 1 O9 cpm/pg [29]. Completely labeled M I 3 DNA in a solution of 0.26 M Na2HP04, pH 7.2, 7% sodium dodecyl sulfate (SDS), 1 m M EDTA, pH 8.0, and 1 % bovine serum albumin at 55°C was applied to blotted nylon membrane according to the procedure described by Westneat et al. [301. The temperature for stringent washes in 2 x standard saline citrate (SSC), 0.1% SDS was 55°C. The washed filters were exposed at - 70°C to Kodak XAR films with inten- sifying screens.
RESULTS
In the initial phases of our study, the conditions for DNA fingerprinting of mouse liver DNA were optimized. Initial analyses with human L1, a member of the Lines family (long interspersed sequences), as probe for hybridization were unsuccessful since discrete bands of digested geno- mic mouse DNA could not be resolved. In subsequent anal- yses we employed M I 3 DNA as a probe hybridized to genomic mouse liver DNA digested with various restric- tion enzymes. Out of these, Hinfl was found to be most suitable for DNA fingerprint analysis, since it resulted in well-defined patterns in Southern blots (Figure 1). Under the gel separation conditions employed, DNA cut with Hinfl and probed with M13 DNA showed approximately 20 bands in the range of 2 to 25 kb whereby clearcut dif- ferences between different strains of mice were evident. Representative examples obtained with normal liver DNA from C57BU6J. C3H/He, and B6C3F1 mice are shown in Figure 2. As expected, certain bands in DNA digests from livers of the B6C3F1 mouse (the F1 cross between C3H/He and C57BU6J) could be nicely allocated to the parental alleles (Figure 2). In this context it is noteworthy that band- ing within strains did not significantly differ, indicating that the DNA fingerprints observed are specific to each strain.
We found it peculiar that Hindlll fragments of bacterio- phage lambda used as size markers also hybridized with the M13 probe, which made the positions of the marker fragments clearly visible on the autoradiogram (see Fig- ures 1 and 3).
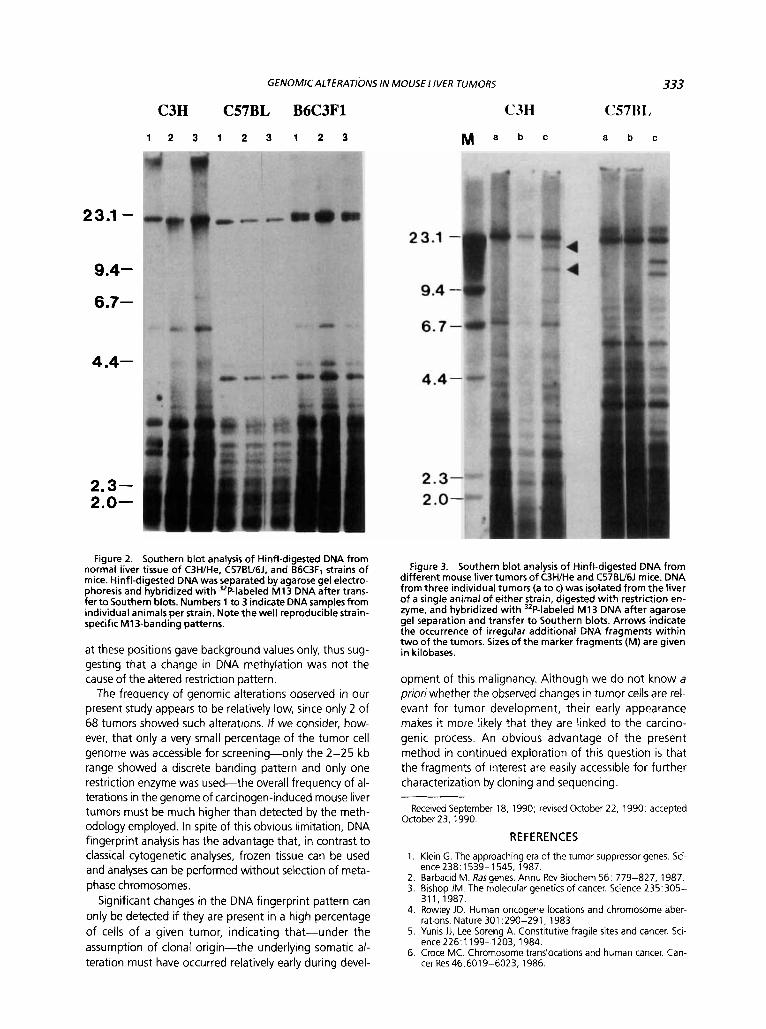
Changes in DNA fingerprint patterns resulting from ge- nomic alterations could be demonstrated in 2 of 68 liver tumors analyzed. The first case was a tumor from a C57BL16J mouse. Hinfl-digested DNA from this tumor showed two additional fragments with approximate sizes of 10 and 13 kb (see Figure 3). Four other liver tumors isolated from the same mouse, two of which are shown in Figure 3, did not show this irregular banding pattern, indicating that the appearance of the new bands was not the result of a polymorphism between individual mice. The second case was a tumor from a C3H/He mouse, where again a doublet of additional bands was seen on the Southern blot of Hinfl-digested DNA. The approximate sizes of the two additional fragments were 12 and 18 kb. Five other tumors from the same mouse, two of which are shown in Figure 3, did not show this RFLP
DISCUSSION
Our present investigation was aimed at detecting ab- normalities in the genome of carcinogen-induced mouse liver tumors. With M 13 DNA as probe, no significant dif- ferences existed between DNA fingerprints within each of the three isogenic strains analyzed, although clearcut interstrain differences were evident. These findings con- trast with recent observations of Jeffreys et al. [31], who reported on interindividual and even interorgan differences in DNA banding when digested mouse DNA was hybrid- ized with a sequence of human minisatellite DNA. A pos- sible explanation for this difference could be that the
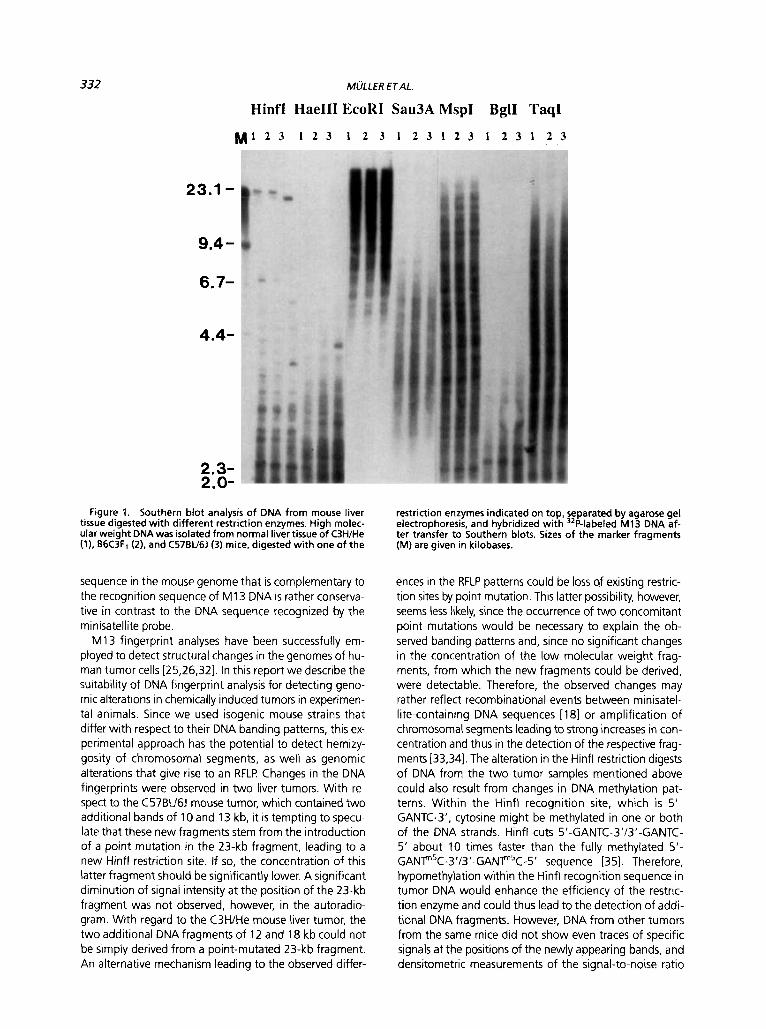
332
23.1 -
9.4 - 6.7-
4.4-
2.3- 2.0-
MULLER ETAL.
Hinfl HaeIII EcoRI Sau3A MspI BglI TaqI
M I 2 3 1 2 3 1 2 3 1 2 3 1 2 3 1 2 3 1 2 3
Figure 1. Southern blot analysis of DNA from mouse liver tissue digested with different restriction enzymes. High molec- ular weight DNA was isolated from normal liver tissue of C3H/He (1). B6C3F1 (2). and C57BU6J (3) mice, digested with one of the
sequence in the mouse genome that is complementary to the recognition sequence of M13 DNA is rather conserva- tive in contrast to the DNA sequence recognized by the minisatellite probe.
M13 fingerprint analyses have been successfully em- ployed to detect structural changes in the genomes of hu- man tumor cells (25.26.321. In this report we describe the suitability of DNA fingerprint analysis for detecting geno- mic alterations in chemically induced tumors in experimen- tal animals. Since we used isogenic mouse strains that differ with respect to their DNA banding patterns, this ex- perimental approach has the potential to detect hemizy- gosity of chromosomal segments, as well as genomic alterations that give rise to an RFLP Changes in the DNA fingerprints were observed in two liver tumors. With re- spect to the C57BU6J mouse tumor, which contained two additional bands of 10 and 13 kb, it is tempting to specu- late that these new fragments stem from the introduction of a point mutation in the 23-kb fragment, leading to a new Hinfl restriction site. If so, the concentration of this latter fragment should be significantly lower. A significant diminution of signal intensity at the position of the 23-kb fragment was not observed, however, in the autoradio- gram. With regard to the C3H/He mouse liver tumor, the two additional DNA fragments of 12 and 18 kb could not be simply derived from a point-mutated 23-kb fragment. An alternative mechanism leading to the observed differ-
restriction enzymes indicated on top, separated by agarose gel electrophoresis, and hybridized with 32P-labeled M13 DNA af- ter transfer to Southern blots. Sizes of the marker fragments (M) are given in kilobases.
ences in the RFLP patterns could be loss sf existing restric- tion sites by point mutation. This latter possibility, however, seems less likely, since the occurrence of two concomitant point mutations would be necessary to explain the ob- served banding patterns and, since no significant changes in the concentration of the low molecular weight frag- ments, from which the new fragments could be derived, were detectable. Therefore, the observed changes may rather reflect recombinational events between minisatel- lite-containing DNA sequences [ 181 or amplification of chromosomal segments leading to strong increases in con- centration and thus in the detection of the respective frag- ments [33,34]. The alteration in the Hinfl restriction digests of DNA from the two tumor samples mentioned above could also result from changes in DNA methylation pat- terns. Within the Hinfl recognition site, which is 5’- GANTC-3’, cytosine might be methylated in one or both of the DNA strands. Hinfl cuts 5’-GANTC-3‘/3’-GANTC- 5‘ about 10 times faster than the fully methylated 5’- GANF5C-3’/3’-GANF5C-5’ sequence [35]. Therefore, hypomethylation within the Hinfl recognition sequence in tumor DNA would enhance the efficiency of the restric- tion enzyme and could thus lead to the detection of addi- tional DNA fragments. However, DNA from other tumors from the same mice did not show even traces of specific signals at the positions of the newly appearing bands, and densitometric measurements of the signal-to-noise ratio
23.1 -
9.4-
6.7-
GENOMICALTERATIONS IN MOUSE LIVER TUMORS
C3H C57BL B6C3F1 C3H
1 2 3 1 2 3 1 2 3 M a b c
23.1 -
9.4 -
6.7-
4.4- 4.4-
2.3- 2.0-
Fiqure 2. Southern blc analvsis of Hinfl-digested DN from normal liver tissue o f C3H1He. c57BV6J. and 66C3F1 strains of mice. Hinfl-digested DNA was separated by agarose gel electro- phoresis and hybridized with 32P-labeled M13 DNA after trans- fer t o Southern blots. Numbers 1 to 3 indicate DNA samples from individual animals per strain. Note the well reproducible strain- specific M13-banding patterns.
a t these positions gave background values only, thus sug- gesting that a change in DNA methylation was not the cause of the altered restriction pattern.
The frequency of genomic alterations observed in our present study appears to be relatively low, since only 2 of 68 tumors showed such alterations. If we consider, how- ever, that only a very small percentage of the tumor cell genome was accessible for screening--only the 2-25 kb range showed a discrete banding pattern and only one restriction enzyme was used-the overall frequency of al- terations in the genome of carcinogen-induced mouse liver tumors must be much higher than detected by the meth- odology employed. In spite of this obvious limitation, DNA fingerprint analysis has the advantage that, in contrast to classical cytogenetic analyses, frozen tissue can be used and analyses can be performed without selection of meta- phase chromosomes.
Significant changes in the DNA fingerprint pattern can only be detected if they are present in a high percentage of cells of a given tumor, indicating that-under the assumption of clonal origin-the underlying somatic al- teration must have occurred relatively early during devel-
2.3- 2 .o-
333
C57H1,
a b c
Figure 3. Southern blot analysis o f Hinfl-digested DNA from different mouse liver tumors of C3H/He and C57BU6J mice. DNA from three individual tumors (a t o c) was isolated from the liver of a single animal of either strain, digested with restriction en- zyme, and hybridized with 32P-labeled M13 DNA after agarose gel separation and transfer t o Southern blots. Arrows indicate the occurrence o f irregular additional DNA fragments within two of the tumors. Sizes of the marker fragments (M) are given in kilobases.
opment of this malignancy. Although we do not know a priori whether the observed changes in tumor cells are rel- evant for tumor development, their early appearance makes it more likely that they are linked to the carcino- genic process. An obvious advantage of the present method in continued exploration of this question is that the fragments of interest are easily accessible for further characterization by cloning and sequencing
Received September 18, 1990; revised October 22, 1990; accepted October 23, 1990.
REFERENCES
1. Klein G. The approaching era of the tumor suppressor genes. Sci-
2. Barbacid M. Rasgenes. Annu Rev Biochem 56: 779-827, 1987. 3. Bishop JM. The molecular genetics of cancer. Science 235:305-
311,1987. 4. Rowley JD. Human oncogene locations and chromosome aber-
rations. Nature 301 :290-291, 1983. 5. Yunis JJ, Lee Soreng A. Constitutive fragile sites and cancer. Sci-
ence226:1199-1203, 1984. 6. Croce MC. Chromosome translocations and human cancer. Can-
cer Res46:6019-6023, 1986.
ence 238: 1539-1 545, 1987.

334 MULLER ETAL.
7 Ponder B. Gene losses in human tumors. Nature 335:400-402, 1988.
8. Sasaki MS, Tsunematsu V. Utsunomiya J, Utsumi J Sitedirected chro- mosome rearrangements in skin fibroblasts from persons carrying genes for hereditary neoplasms. Cancer Res40:4796-4803,1980,
9. Evans HJ. Mutation cytogenetics. Past, present, future. Mutat Res 204:355-363,1987.
10. Koufos A, Hansen MF, Copeland NG, Jenkins NA, Lampkin BC, Cavenee WK. Loss of heterozygosity in three embryonal tumours suggests a common pathogenetic mechanism. Nature 31 6.330- 334,1985.
11. Dracopoli NC, Alhadeff B, Houghton AN, Old U. Loss of hetero- zygosity at autosomal and X-linked loci during tumor progres- sioninapatientwith melanoma. Cancer Res47:3995-4000.1987.
12 Vogelstein B, Fearon EF, Kern SE, et al. Allelotype of colorectal carcinomas. Science 244:207-211, 1989.
13. Stoye JP, Coffin JM. Polymorphism of murine endogenous pro- viruses revealed by using virus class-specific oligonucleotide probes. JVirol62:168-175, 1988.
14 Frankel WN, Stoye JP, Taylor BA, Coffin JM. Genetic identification of endogenous polytropic proviruses by using recombinant in- bred mice. JVirol63:3810-3821, 1989.
15. Jeffreys AJ, Wilson V, Thein SL. Individual-specific fingerprints of human DNA. Nature 316:67-73, 1985.
16. Gill P, Lygo LE, Fowler SJ, Werrett DJ. An evaluation of DNA fin- gerprinting for forensic purposes. Electrophoresis 8:38-44. 1987
17. Masters JRW, Bedford P, Kearney A, Povey 5, Franks LM. Bladder cancer cell line cross-contamination: Identification using a locus- specific minisatellite probe. Br J Cancer 57:284-286. 1988
18. Thein SL, Jeffreys AJ, Gooi HC, et al. Detection of somatic changes in human cancer DNA by DNA fingerprint analysis Br 1 Cancer 55.353-356, 1987.
19. White JJ, Neuwirth H. Miller CD, Schneider EL. DNA alterations in prostatic adenocarcinoma and benign prostatic hyperplasia. Detection by DNA fingerprinting analyses. Mutat Res 237:37-43, 1990.
20. Uchida T, White JJ, Neuwirth H. DNA minisatellites demonstrate somatic DNA changes in most human bladder cancers Cytogenet Cell Genet 53:61--63, 1990.
21. Vassart G, Georges M, Monsieur R. Brocas H, Lequarre AS, Chris- tophe D. A sequence in M13 phage detects hypervariable minisatellites in human and animal DNA. Science 235:683-684, 1987.
22. Ryskov AP, Jincharadze AG, Prosnyak MI, lvanov PL, Limborska SA. M13 phage DNA as a universal marker for DNA fingerprinting of animals, plants and microorganisms. FEBS Lett 233:388--392, 1988.
23. Georges M, Lequarre A, Castelli M, Hanset R, Vassart G. DNA fingerprinting in domestic animals using four different minisatellite probes Cytogenet Cell Genet47:127-131, 1988.
24. Chen F: Hayward NK, Kidson C, Ellem KAO. Conditions for gen- erating fingerprints using M13 phage DNA. Nucleic Acids Res 18:1065, 1990.
25. Lagoda PJL, Seitz G, Epplen JT, lssinger OG. Increased detectabil- ity of somatic changes in the DNA from human tumours after probing with "synthetic" and "genome-derived" hypervariable multilocus probes. Hum Genet 84:35-40, 1989.
26. Hayward N, Chen P, Nancarrow D, et al. Detection of somatic mutations in tumours of diverse types by DNA fingerprinting with M13 phage DNA. Int J Cancer 45:687-690, 1990.
27. Drinkwater NR, Ginsler JJ. Genetic control of hepatocarcinogene- sis in C57BL16J and C3HIHeJ inbred mice. Carcinogenesis 7: 1701 -1 707, 1986.
28. Krieg P, Amtmann E, Sauer G. The simultaneous extraction of high-molecular-weight DNA and of RNA from solid tumors. Anal Biochem 134:288-294, 1983.
29. Feinberg AP, Vogelstein B. A technique for radiolabeling DNA re- striction endonuclease fragments to high specific activity. Anal Biochem 132:6, 1983.
30 Westneat DF, Noon WA, Reeve HK, Aquadro FA. Improved hy- bridization conditions for DNA "fingerprints" probed with M13 Nucleic AcidsRes 16.4161, 1988.
31. Jeffreys AJ, Wilson V, Kelly R, Taylor BA, Bulfield G. Mouse DNA "fingerprints": analysis of chromosome localization and germ- line stability of hypervariable loci in recombinant inbred strains. Nucleic Acids Res 15:2823-2836, 1987.
32. Pakkala S, Helminen P, Saarinen UM, Alitalo R, Peltonen L. Dif- ferences in DNA-fingerprints between remission and relapse in childhood acute lymphoblastic leukemia. Leukemia Res 12:757- 762,1988.
33. Stark GR, Wahl GM. Gene amplification. Annu Rev Biochem 53:447-457, 1984.
34. Schimke RT. Gene amplification in human cells Cell 37:705-71 1, 1984.
35. Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning. Cold Spring Harbor Laboratory Press Cold Spring Harbor, New York, 1989.


















