
Critical review Sequence skill learning in persons who ... · Implications for...
28
Journal of Fluency Disorders 32 (2007) 251–278 Critical review Sequence skill learning in persons who stutter: Implications for cortico-striato-thalamo-cortical dysfunction Sarah Smits-Bandstra a,∗ , Luc F. De Nil a,b,c a Department of Speech-Language Pathology, University of Toronto, Toronto, Canada b Toronto Western Research Institute, University Health Network, Toronto, Canada c The Hospital for Sick Children Research Institute, Toronto, Toronto, Canada Received 19 March 2007; received in revised form 29 May 2007; accepted 7 June 2007 Abstract The basal ganglia and cortico-striato-thalamo-cortical connections are known to play a critical role in sequence skill learning and increasing automaticity over practice. The current paper reviews four studies comparing the sequence skill learning and the transition to automaticity of persons who stutter (PWS) and fluent speakers (PNS) over practice. Studies One and Two found PWS to have poor finger tap sequencing skill and nonsense syllable sequencing skill after practice, and on retention and transfer tests relative to PNS. Studies Three and Four found PWS to be significantly less accurate and/or significantly slower after practice on dual tasks requiring concurrent sequencing and colour recognition over practice relative to PNS. Evidence of PWS’ deficits in sequence skill learning and automaticity development support the hypothesis that dysfunction in cortico-striato-thalamo-cortical connections may be one etiological component in the development and maintenance of stuttering. Educational objectives: As a result of this activity, the reader will: (1) be able to articulate the research regarding the basal ganglia system relating to sequence skill learning; (2) be able to summarize the research on stuttering with indications of sequence skill learning deficits; and (3) be able to discuss basal ganglia mechanisms with relevance for theory of stuttering. © 2007 Elsevier Inc. All rights reserved. Keywords: Stuttering; Sequencing; Skill learning; Automaticity; Dual task; Basal ganglia; Finger tapping; Nonsense syllables; Practice ∗ Corresponding author at: 2037 Kilbride Street, P.O. Box 679, Kilbride, ON, Canada L0P 1G0. Tel.: +1 905 335 9127; fax: +1 416 978 1596. E-mail address: [email protected] (S. Smits-Bandstra). 0094-730X/$ – see front matter © 2007 Elsevier Inc. All rights reserved. doi:10.1016/j.jfludis.2007.06.001
-
Upload
nguyennguyet -
Category
Documents
-
view
220 -
download
0
Transcript of Critical review Sequence skill learning in persons who ... · Implications for...

Journal of Fluency Disorders 32 (2007) 251–278
Critical review
Sequence skill learning in persons who stutter:Implications for cortico-striato-thalamo-cortical
dysfunction
Sarah Smits-Bandstra a,∗, Luc F. De Nil a,b,c
a Department of Speech-Language Pathology, University of Toronto, Toronto, Canadab Toronto Western Research Institute, University Health Network, Toronto, Canada
c The Hospital for Sick Children Research Institute, Toronto, Toronto, Canada
Received 19 March 2007; received in revised form 29 May 2007; accepted 7 June 2007
Abstract
The basal ganglia and cortico-striato-thalamo-cortical connections are known to play a critical role insequence skill learning and increasing automaticity over practice. The current paper reviews four studiescomparing the sequence skill learning and the transition to automaticity of persons who stutter (PWS) andfluent speakers (PNS) over practice. Studies One and Two found PWS to have poor finger tap sequencingskill and nonsense syllable sequencing skill after practice, and on retention and transfer tests relative toPNS. Studies Three and Four found PWS to be significantly less accurate and/or significantly slower afterpractice on dual tasks requiring concurrent sequencing and colour recognition over practice relative to PNS.Evidence of PWS’ deficits in sequence skill learning and automaticity development support the hypothesisthat dysfunction in cortico-striato-thalamo-cortical connections may be one etiological component in thedevelopment and maintenance of stuttering.
Educational objectives: As a result of this activity, the reader will: (1) be able to articulate the researchregarding the basal ganglia system relating to sequence skill learning; (2) be able to summarize the researchon stuttering with indications of sequence skill learning deficits; and (3) be able to discuss basal gangliamechanisms with relevance for theory of stuttering.© 2007 Elsevier Inc. All rights reserved.
Keywords: Stuttering; Sequencing; Skill learning; Automaticity; Dual task; Basal ganglia; Finger tapping; Nonsensesyllables; Practice
∗ Corresponding author at: 2037 Kilbride Street, P.O. Box 679, Kilbride, ON, Canada L0P 1G0. Tel.: +1 905 335 9127;fax: +1 416 978 1596.
E-mail address: [email protected] (S. Smits-Bandstra).
0094-730X/$ – see front matter © 2007 Elsevier Inc. All rights reserved.doi:10.1016/j.jfludis.2007.06.001

252 S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2522. The importance of the BG for skill learning and automatization . . . . . . . . . . . . . . . . . . . . . . . . . . . 254
2.1. The role of the BG in sequencing skill . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2542.2. The role of the BG in early stages of learning . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2562.3. The role of the BG in complex/ambiguous sequence skill learning. . . . . . . . . . . . . . . . . . . 2562.4. The role of the BG in retention and transfer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2582.5. The role of the BG in automatization . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 258
3. Sequence skill learning and automatization in PWS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2593.1. Sequence skill learning in PWS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2593.2. Retention in PWS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2593.3. Automatization in PWS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 259
3.3.1. Nonspeech dual tasks . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2593.3.2. Cognitive and linguistic dual tasks . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 260
4. Primary objectives . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2605. Study One: syllable sequence reading (Smits-Bandstra & De Nil, 2006) . . . . . . . . . . . . . . . . . . . . 2616. Study Two: finger-tap sequence learning (Smits-Bandstra, De Nil, & Saint-Cyr, 2006) . . . . . . . 2627. Study Three: dual task syllable sequence reading (Smits-Bandstra & De Nil, 2006) . . . . . . . . . 2638. Study Four: finger tapping dual task (Smits-Bandstra, De Nil, & Rochon, 2006) . . . . . . . . . . . . 2649. General discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 265
9.1. Sequence skill learning . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2659.2. Dual task performance . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2659.3. The nature of the proposed deficit . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2669.4. Possible neurological correlates of the proposed deficit . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2679.5. Explaining stuttering symptoms and characteristics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 267
9.5.1. Onset of developmental stuttering . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2689.5.2. Spontaneous recovery in stuttering . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2689.5.3. Fluency enhancing conditions (FEC) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2689.5.4. Treatment effects . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 269
9.6. Integrations of the proposed deficit with stuttering etiology theories . . . . . . . . . . . . . . . . . 26910. Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 270
Acknowledgments . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 272References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 272
1. Introduction
It has been postulated that dysfunction of the basal ganglia (BG) and its associated cortico-striato-thalamo-cortical (CSTC) connections may be an important etiological component ofstuttering (Jones, White, Lawson, & Anderson, 2002; Maguire, Gottschalk, Riley, & Franklin,2000). This proposal is based on evidence from a comparison of the symptomatology and neu-rophysiological characteristics reported for patients with basal ganglia dysfunction (PBG: suchas patients with Parkinson’s disease, Huntington’s disease, Tourette’s syndrome, and dystonia)and persons who stutter (PWS). Some of the similarities noted between PBG and PWS includea developmental onset (Pauls, Leckman, & Cohen, 1993; Yairi, Ambrose, & Cox, 1996), envi-ronmental/situational exacerbation of symptoms (Alsobrook & Pauls, 1998; Bloodstein, 1995),facilitory effects of external sensory cues for movement (Liu, Tubbesing, Aziz, Miall, & Stein,1999; Loucks & De Nil, 2001), facilitory effects of focused attention/de-automatization of move-

S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278 253
ments (Kandel, Schwartz, & Jessel, 2000; Venkatagiri, 2004), and sensitivity to fluctuatingdopamine levels or dopamine-moderating medication (Kandel et al., 2000; Wu et al., 1995).
A review of brain imaging studies reveals some compelling similarities between the brainactivity patterns shown by PBG and PWS. For example, a similar pattern of hypoactivation ofcircumscribed left motor planning areas and hyperactivation of right hemisphere homologues andbilateral cerebellar activity was found for both PBG (Berardelli, Rothwell, Thompson, & Hallet,2001; Catalan, Ishii, Honda, Samii, & Hallet, 1999; Mentis et al., 2003) and PWS (Fox et al.,1996; Fox et al., 2000). In particular, PBG show deactivation in the left primary motor cortexand left supplementary motor cortex (SMA) (Catalan et al., 1999). PBG also show compensatoryhyperactivation left prefrontal cortex, left insula, and bilateral cerebellum (Mentis et al., 2003).In addition, PBG show compensatory hyperactivation in areas responsible for attention to actionincluding recruitment of the right hemisphere, specifically the right prefrontal cortex, SMA, andanterior cingulate cortex (Catalan et al., 1999).
PWS also consistently demonstrate hyperactivation of the cerebellum and right frontal cerebralhemisphere including the right frontal operculum (right homologue of Broca’s area), the rightSMA, the right primary motor cortex, the right insula, and the right superior premotor cortexcorresponding roughly to Brodman’s area 6 (BA 6; Ingham, Fox, Costello Ingham, & Zamarripa,2000). Some left cerebral areas are also hyperactivated by PWS including the left insula, leftpremotor cortex (BA 46), left prefrontal cortex, and the left anterior cingulate cortex (De Nil, Kroll,Kapur, & Houle, 2000). Activation of these areas appears to correspond to increased attention toaction or de-automatization of speech movement (De Nil & Bosshardt, 2001).
Similar to PBG, PWS have also demonstrated specific deactivations, areas less active in PWS vs.fluent speakers (PNS), which have been reported in neuroimaging studies. PWS have demonstrateddeactivations in Wernicke’s area, the left superior and posterior temporal lobes, and hypoactivationin circumscribed areas of the left frontal cortex including the left dorsolateral prefrontal cortices,the left superior frontal operculum (BA 44/45; Braun et al., 1997), the left inferior frontal cortex(BA 47; Fox et al., 1996; Ingham et al., 2000), the left primary motor cortex (Neumann et al.,2003, 2005; Preibisch et al., 2003), although not specifically the left SMA. Neumann et al. (2003,2005) reported that hypoactivation of the left inferior frontal cortex and left primary motor cortex,in particular, in PWS across tasks and across pre- and post-therapy measures over a two-yearperiod suggesting a potential locus of dysfunction.
It is well known that the neurophysiological locus of pathology in Parkinson’s disease origi-nates in the BG complex (see Saint-Cyr, 2003 for detail). A pattern of potential loci of pathologyassociated with acquired stuttering was detected only recently when researchers came to considerboth cortical and subcortical structures as well as their interconnections. For example, acquiredstuttering cases have been reported to have resulted from damage to CSTC connections. Forinstance, Ludlow, Rosenberg, Salazar, Grafman, and Smutok (1987) reported that 8 of the 10patients they examined who had acquired stuttering incurred symptoms after injury to the stria-tum or cortico-striatal connections. Many other authors have also reported acquired stutteringassociated with damage to the BG or other structures which play a critical function in CSTCconnections (Ciabarra, Elkind, Roberts, & Marshall, 2000). For example, stuttering was foundto occur after damage to areas of the thalamus that project to speech control regions of the cor-tex (Andy & Bhatnager, 1992; Heuer, Thayer Sataloff, Mandel, & Travers, 1996; Van Borsel,Van Der Made, & Santens, 2003), the SMA (Ackermann, Hertrich, Zieglers, Bitzer, & Bien,1996; Alexander, Naeser, & Palumbo, 1987; Van Borsel, Van Lierde, Van Cauwenberge, & VanOrshoven, 1998), or during electrical stimulation of either the thalamus or SMA (Penfield &Welch, 1951).

254 S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278
Additional similarities in movement characteristics have been reported for both PBG andPWS. For example, four hallmark characteristics associated with BG dysfunction include; actionand rest tremor (Liu et al., 1999), involuntary movements including chorea, dyskinesia, tics,and dystonia (Abwender et al., 1998; akinesia (Low, Miller, & Vierck, 2002), and bradykinesia(Berardelli et al., 2001). Research studies indicate that many PWS also demonstrate tremor (Denny& Smith, 1992), involuntary movements (Mulligan, Anderson, Jones, Williams, & Donaldson,2001), reaction time slowing (Smits-Bandstra, De Nil, & Rochon, 2006), and slow movementdurations (Max & Yudman, 2003) relative to PNS.
In his review, Alm (2004) briefly mentioned the importance of BG systems and dopamineregulation in reward prediction during learning. What was not discussed in as much detail in hispaper was the role these systems play in sequence skill learning and automaticity. Neuroimagingand primate studies have established that the BG and CSTC connections play a critical rolein sequence skill learning and increasing automaticity over practice (Catalan et al., 1999). Thesequence skill learning deficits of PBG are well established in the literature (Benecke, Rothwell,Dick, Day, & Marsden, 1986; Doyon et al., 1997), as are PBG’s poor performance on transfer andretention tests and on dual tasks designed to assess automaticity (Smiley-Oyen, Worringham, &Cross, 2003).
In this paper, we will first review studies that establish sequence skill learning deficits andpoor automaticity development in PBG. Second, those limited skill learning and dual task stud-ies that do exist in the stuttering literature are reviewed. Third, we will summarize the mainfindings from four studies specifically investigating sequence skill learning and the transitionto increasing automaticity in PWS. Finally tentative conclusions surrounding the etiologicalimplications of sequence skill learning deficits and poor automaticity development in PWS arediscussed.
2. The importance of the BG for skill learning and automatization
2.1. The role of the BG in sequencing skill
A distributed system of cortical and subcortical structures is involved in sequence skill learning,retention, and transfer (see Fig. 1). Recent studies have emphasized the primary role of theBG and CSTC connections for these skills (Laforce & Doyon, 2001; see Saint-Cyr, 2003 fora review). Because evidence of BG involvement in sequence learning is well established in theliterature, this paper will be limited to representative studies imparting findings most relevant to thediscussion.
The basal ganglia is a complex cluster of subcortical nuclei which has extensive direct andindirect neural connections to the lateral cortex and the spinal cord (see Fig. 1). These connectionshave both an inhibitory and excitatory effect on neural activity. As reviewed by Alm (2004), inputfrom the BG to the SMA, and higher order motor cortical areas via the thalamus, appears tosignal the timing of practiced sequential elements (Brotchie, Iansek, & Horne, 1991; Sawaguchi& Goldman-Rakic, 1991). Research has shown that striatal neurons show rapid firing changesin the early trials of motor sequence learning. Pre-movement firing activity begins progressivelyearlier in these neurons during learning (Pasupathy & Miller, 2005). These striatal neurons providea predictive signal for initiation of an increasingly automated, quick, fluent movement sequencein response to a specified stimulus (Pasupathy & Miller, 2005; Shizgal & Arvanitogiannis, 2003).Disruption of the timing of BG signaling via CSTC connections resulted in arrest, perseveration, orhesitations in fluent sequence execution in primates (Pasupathy & Miller, 2005). Weiss, Stelmach,

S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278 255
Fig. 1. Diagram of input and output connections of the basal ganglia-thalamus-cortex circuitry. Major basal ganglia nucleiare represented by shaded boxes. Excitatory connections are shown as full arrows, inhibitory connections as dotted arrows(adapted from Kandel et al., 2000, p. 856).
& Hefter (1997) found that Parkinson’s disease was associated with abnormal hesitancy andintersegmental latencies in the step-by-step performance of a sequence of hand movements. Thisinformation has led several researchers to propose that CSTC connections, particularly those ofthe left hemisphere, serve to integrate sensory input in order to initiate and regulate the executiontiming of sequences (Roy, Saint-Cyr, Taylor, & Lang, 1993; Seitz & Roland, 1992).
Folion, Tremblay, and Bedard (1988) present a physiological explanation for disruption in thetiming between BG and CSTC connections. They reported that experimentally induced dopamineloss in monkeys resulted in a remarkable loss of the specificity of movement encoding of BGneurons. When deprived of dopamine, cells in the BG lost their specificity for encoding move-ment direction. The firing patterns of the cells become slow, defocused, and cells responded tomovements in several directions. In his review of BG function, Saint-Cyr (2003) seconded theproposal that an imbalance in dopamine regulation resulted in a delay in the cells’ ability to selectappropriate responses due to ambiguity of undifferentiated choices. As seen in primates withinduced lesions and PBG this defocused cell activity resulted in slow sequence initiation andhesitations/delays in sequence production during sequence learning (Pasupathy & Miller, 2005;Roy et al., 1993).

256 S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278
2.2. The role of the BG in early stages of learning
More recent primate studies have confirmed the importance of the BG in the earliest stages ofsequence skill learning. Using single cell recording procedures, Pasupathy and Miller (2005) foundthat the BG were the first to encode stimulus response associations in a visuomotor learning task.They found that BG (specifically caudate) activity began earlier and earlier relative to prefrontalcortex activity during practice repetitions (a sort of anticipation) in order to facilitate quickerresponse initiation.
Saint-Cyr (2003) suggested that the BG are critical for finding the “ballpark,” or recognizing andselecting appropriate pre-existing movement control patterns while inhibiting irrelevant ones inearly stages of learning. In support of this interpretation, Frith, Bloxham, and Carpenter (1986) hadpatients with Parkinson’s disease (PPD) and control subjects track a target by moving a joystick.During the first minute of each practice session control subjects showed a marked improvement inperformance while PPD did not. The authors suggested that this rapid but temporary improvementin performance reflected the acquisition of a motor “set” and that PPD had difficulty in maintainingsuch sets.
Korman, Raz, Flash, and Karni (2003), Frith et al. (1986), and Rowe et al. (2002) concurredthat CSTC connections were critical during early stage sequence learning for acquisition of set.Evarts, Shinoda, and Wise (1984) defined “set” as a matching of the parameters of a skill tosuit the context in which it is being performed. Grafton, Hazeltine, and Ivry (2002) and Houk,Buckingham, and Barto (1996) proposed that the integrity of CSTC connections were criticalspecifically for the acquisition of set for learning complex sequential behaviours.
2.3. The role of the BG in complex/ambiguous sequence skill learning
Skill learning impairments associated with BG and CSTC connection dysfunction appear tobe specific to the sequence type. Significant differences between PPD and control subjects werefound predominantly for procedural skill learning. Specifically, PPD were found to have slowerreaction times for implicit sequence learning tasks such as the serial reaction time tasks (SRT;see Siegert, Taylor, Weatherall, & Abernethy, 2006, for a meta-analysis). Significant differencesbetween PPD and control subjects were also found for procedural learning tasks with a cognitiveprocessing component, but not for declarative learning tasks (see Saint-Cyr, Taylor, & Lang, 1988for a review).
Saint-Cyr et al. (1988) compared 24 early-stage PPD, 24 patients in various stages of Hunt-ington’s disease, 24 healthy control subjects and 2 patients with severe declarative memoryimpairments. All subjects performed tests of declarative memory including parts of the Wech-sler Memory Scale, the Buschke Selective Reminding Test, and a standard declarative test ofverbal supraspan learning. Subjects also performed the “Tower of Toronto” test of cognitiveprocedural learning which requires learning of the most efficient sequence of movements to relo-cate four coloured discs onto three pegs (see appendix of Saint-Cyr et al., 1988). Huntington’sdisease patients were somewhat impaired on both procedural and declarative tasks. However,in comparison to matched controls, PPD performed normally on all declarative tests but werespecifically impaired on the Tower of Toronto. In sharp contrast, patients with declarative deficitswere clearly impaired on all declarative memory tests but demonstrated no difficulties with theTower of Toronto.
Saint-Cyr and colleagues pointed out that study methodology must attempt to control, asbest as possible, the extent/stage of disease. Early stage Parkinson’s disease is thought to be

S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278 257
largely restricted to alteration of BG/subcortical structure function while later stages of bothPPD and Huntington’s disease typically involve dementia associated with additional degenerationof cortical areas such as prefrontal areas resulting in more generalized performance deficits. Inaddition, similar studies comparing declarative and procedural learning skills of PPD found that thetype of procedural learning task is important to detect learning deficits in PPD (Dujardin & Laurent,2003; Harrington, Haaland, Yeo, & Marder, 1990). For example, in a study by Helmuth, Mayr,and Daum (2000) PPD and control subjects were administered a manual serial response time task.PPD were unimpaired at learning a sequence of spatial locations, but showed a deficit at learninga stimulus-to-motor-response sequence. The authors suggested that sequencing impairments inParkinson’s disease were not general, but specific to stimulus-to-motor-response sequencing.
Further investigation also indicates significant differences between PPD and control subjectsfor relatively long and complex sequences involving integration of several different movements,but not for simple or repetitive sequences (Shohamy, Myers, Grossman, Sage, & Gluck, 2005).Mentis et al. (2003) compared early stage PPD with matched control subjects on a trial anderror sequence-learning task. Subjects moved a mouse out to, and back from, eight spatial tar-gets to a central location. Sequence learning was performed with timing cues so that sequenceelement reaction times were equivalent, however PPD demonstrated significantly more errors inthe first 10 s than the control group. PPD also demonstrated a significantly slower rate of newelement acquisition over the first 30 s of training relative to control subjects. These investigatorsalso conducted positron emission tomography (PET) scanning during learning and observed thatPPD demonstrated wide spread differences in brain function, specifically increased activation inpreSMA, premotor cortex and insula for both right and left hemispheres relative to control sub-jects despite a similar number of movements in time. Mentis and colleagues suggested that thesebrain recruitment patterns were effective for behavioural compensation when sequences were nottoo long or complex and patients could take their time to learn them. They also concluded that“the dramatic failure in learning that occurs when task difficulty is high or Parkinson’s disease isadvanced may occur when activation regions can not longer compensate to maintain function (p.617)”.
Roy et al. (1993) compared PPD and age-matched control subjects on a task requiring subjectsto learn a sequence of three or four different hand movements to a criterion of five consecutivecorrect trials. The control sequence required simple repetition of the same hand posture. The groupsdid not differ in their accuracy levels, however PPD demonstrated significantly slower reactiontime, total movement time, and reaction time for individual movements within the sequence (inter-response time). Significant differences were found only for the different movement sequences andnot for the repetitive movement sequence. Shohamy et al. (2005) found that PPD patients tested“off” L-dopa medication performed as well as matched control subjects on a first order sequencetask (a task requiring learning of simple stimulus-response associations), in which subjects clickedthe mouse on the correct coloured door to access the route toward a treasure. In contrast, PPDwere impaired at learning the task when second order/hierarchical learning was required, wheresubjects clicked the mouse on a sequence of correct coloured doors to access the route toward atreasure. In contrast, PPD when tested “on” L-dopa performed equivalent to control subjects onboth first and second order sequence learning.
Sequence learning deficits associated with BG dysfunction were found for speech as well asnonspeech sequencing tasks (Schulz, Leon, & Sulc, 2001; Smith & McDowall, 2004; Smith,Siegert, McDowall, & Abernethy, 2001; Westwater, McDowall, Siegert, Mossman, & Abernethy,1998). Schulz et al. (2001) measured the accuracy and articulatory patterns of PPD relative tocontrol subjects while learning a novel speech utterance, a nonsense phrase “Thraim po fra mo

258 S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278
dis” practiced 50 times. They found only control subjects demonstrated increased accuracy withreductions in duration and variability of productions over practice. The authors suggested thatthe PPD appeared to increase accuracy at the expense of reductions in movement durations andconsistency. Smith and McDowall (2004) conducted a serial response time task that required verbalresponses to a 12-item sequence. Twenty PPD and 37 matched controls participated. Following avisual stimulus subjects said aloud “A, B, C, or D” to indicate one of four spatial locations wherethe next anticipated stimulus would appear. Their results indicated that the speech reaction timesof PPD improved significantly less and more gradually than those of the control subjects overpractice, while the error rates for both groups were equivalent.
2.4. The role of the BG in retention and transfer
Studies with PPD have also demonstrated the importance of the BG and CSTC connections forretention (Doyon et al., 1998; Smiley-Oyen et al., 2003), and transfer (Dominey, Ventre-Dominey,Broussolle, & Jeannerod, 1997) of various skills such as finger tap sequencing or linear scalingarm movements.
Smiley-Oyen et al. (2003) compared nine PPD and matched controls on a learned linear armmovement-scaling task over two days. Subjects were required to make arm movements with theirtwo arms proportional to the length of visually presented target bars. Scaling was acquired throughknowledge of results following every second trial. PPD demonstrated poor initial acquisition of thelinear scaling movement as well as significantly poorer retention of scaling skill after a retentionperiod of 24 h. Doyon et al. (1998) compared the performance of 11 PPD and 11 matched controlsubjects on the serial response time task. This test consisted of a visual reaction time task witha fixed embedded sequence of finger movements. The subjects practiced the sequence 40 timesper weekly session over six weeks. Eight to 16 months later subjects were retested revealing thatPPD who changed from Stage 1 to Stage 2 of the disease did not show as much retention of theability to produce the repeating visuomotor sequence. The authors determined that the differencein performance in the rention session was not due merely to a motor deterioration per se.
Based on a comparison of PPD and control subjects on a finger movement sequence learningstudy, Dominey et al. (1997) concluded that sequence learning and transfer appears to be impairedin PPD only when feedback/knowledge of results is not present to guide learning. They proposedthat the learning mechanisms of PPD rely on augmented explicit sensory feedback. To substantiatethis hypothesis, Guadagnoli, Leis, Van Gemmert, & Stelmach (2002) reported PPD required 100%knowledge of results while control subjects required only 20% knowledge of results in order todemonstrate commensurate retention of a simple timing movement.
2.5. The role of the BG in automatization
Dual-tasks are well-established in the literature as a means to investigate the level of autom-atization of a skill. This method assesses the level of attentional resources that a subject canallocate to a second task, as a primary task becomes well practiced and increasingly automated(Doyon et al., 1998). Thomas, Reymann, Lieury, & Allain (1996) found that PPD demonstratedpoor performance relative to control subjects during early stage learning of a maze tracing andan arithmetic task, as well as after continued practice during the “automation” stage. Similarly,Doyon et al. (1997, see above) found PPD showed impairment of sequence learning after contin-ued practice during the automatization stage. Patients with dystonia (Jahanshahi, Rowe, & Fuller,2003), Huntington’s disease (Brown, Redondo-Verge, Chacon, Lucas, & Channon, 2001), and

S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278 259
PPD (Benecke et al., 1986; Doyon et al., 1998) each demonstrate poor performance on dual tasksrelative to control subjects, indicating lower levels of automaticity after practice. These studiesprovide evidence for the continuing role of the BG and CSTC connections in the development ofautomaticity in sequencing skill.
3. Sequence skill learning and automatization in PWS
3.1. Sequence skill learning in PWS
Like PPD, PWS demonstrate poor skill learning relative to fluent speakers (PNS) on nonspeechsequencing tasks after practice. Weinstein, Caruso, Severing, and VerHoeve (1989) found thatPWS, unlike PNS, did not show a significant decrease in the time it took to initiate the firstsaccade (small rapid eye movement) of a multi-saccadic sequence after practice. Webster (1986)evaluated the ability of PWS and PNS to learn 4-element finger tapping sequences and foundthat PWS did not show improved accuracy after practice on sequences with no repeated elements(2-1-3-4 as opposed to 2-1-3-1) compared to PNS.
PWS also demonstrate relatively slower and more inaccurate performance on speech sequenc-ing tasks after practice relative to PNS. Ludlow, Siren, and Zikira (1997) asked PNS and PWSto practice two, 4-syllable nonsense words. Subjects practiced recalling and producing the wordsbetween trials of a listening comprehension secondary task. Ludlow et al. found that PWS wereimpaired in their rate of learning as well as the overall accuracy of their nonsense word produc-tions. Similarly, Cooper and Allen (1977) found that PWS required significantly more repetitionsto demonstrate increased speed of repetition for read-aloud paragraphs and sentences compared toPNS when they used between 16 and 110 trial repetitions. Children who stutter have also demon-strated a diminished ability to learn phonological sequences relative to fluent speaking children.Berman Hakim and Bernstein Ratner (2004) found that children who stutter were significantlyless accurate than fluent speaking children CNS when reproducing learned nonsense syllablesequences that were three or more syllables long.
3.2. Retention in PWS
Only one unpublished study of which we are aware has compared PWS and PNS on retentionof a learned motor speech task. Namasivayam and van Lieshout (2004) measured the speed andcoordination of lip movements of PWS and PNS using electromagnetic articulography. PWS wereasked to repeat the pronounceable nonsense words/bapi/, /bipa/, /bapiter/, and /bipater/for 12 swith a bite block in place. When retention and transfer were assessed two and three days later,PWS demonstrated slower nonsense word performance and more variable lip coordination duringproduction as compared to PNS.
3.3. Automatization in PWS
3.3.1. Nonspeech dual tasksFitzgerald, Cooke, and Greiner (1984) asked PWS and PNS to write the numbers 1–12 in
a vertical line down a sheet of paper without visual feedback, simultaneously with both handswhile performing a concurrent speaking task. PWS demonstrated poorly organized non-dominanthandwriting, more reversed numbers, and greater differences in dominant/non-dominant handscores compared to PNS. The authors suggested that their results could be interpreted as “the

260 S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278
result of an inability to allocate attention freely to processes competing for functional cerebralspace” (p. 63).
Using a similar methodology, in Webster’s (1988) experiment, subjects listened to words andwere asked to repeat them and then write the initial letters of the words as quickly as possiblewith two hands simultaneously. Results indicated that PWS made more reversed letters, showedpoorer letter formation quality, and were significantly slower in responding during the dual taskcompared to PNS. These results were interpreted by Webster as implicating the SMA in stuttering,however he also stated that “it may be more parsimonious at this time to interpret the data in termsof group differences in processing resources” (p. 241).
Webster (1989) compared PWS’ and PNS’ ability to concurrently tap a number sequence withone hand and turn a knob back and forth 30 degrees with the other hand. PWS performed rela-tively poorly on both tasks under dual task conditions. Also using a tapping paradigm, Greiner,Fitzgerald, and Cooke (1986) found that PWS had slower tapping rates than PNS while simultane-ously speaking. PWS demonstrated a larger dual task interference effect evidenced by increasedtapping errors and increased rates of stuttered speech. In a later study Forster and Webster (2001)found PWS performed more poorly and made more on-line corrections on an “etch-a-sketch”bimanual crank task relative to PNS speakers.
3.3.2. Cognitive and linguistic dual tasksUsing a cognitive/linguistic dual task, De Nil and Bosshardt (2001) found that PWS showed
significant impairment of rhyming and category decision-making during the dual task condition.PWS did not differ from PNS in the number of words used for sentence formulation, howevertheir sentences tended to contain fewer syllables and were less grammatically complex. Neu-roimaging revealed increased brain activity in areas associated with early stages of motor learningand attentive processing. These brain activity patterns suggested less automatized processing ofspeech/language tasks in PWS as compared to PNS.
Bosshardt (1999) asked PWS and PNS to repeat random three-word sequences containingthree-syllable nouns (e.g., buttermilk) for the first task. The second task required participants tocalculate the addition of three addends (22 + 12 + 3). Under dual task conditions, the stutteringrate of PWS increased beyond the base level for single task word repetition. In yet another study,Bosshardt (2002) found that the disfluencies of PWS significantly increased when simultaneouslyrepeating words and silent reading or memorizing words.
The consistently poor performance manifested by PWS on these various dual tasks can beinterpreted to suggest that PWS performed the speaking and/or the secondary tasks with relativelylow levels of automaticity compared to PNS. However, this evidence speaks only indirectly to theissue. Further investigative studies which have directly evaluated the process of automatizationover practice bring critical clarification to the issue and are presented below.
4. Primary objectives
Select patients with BG/CSTC connection dysfunction have consistently demonstratedimpaired sequence skill acquisition and an impaired ability to develop automaticity over practice(Laforce & Doyon, 2001). If present, impaired skill acquisition in PWS may indicate possibleetiological factors of stuttering. There is limited evidence in the existing literature, which speaksdirectly to skill learning and automaticity development in the stuttering population. To this end,four recent studies are reviewed for evidence that (1) PWS demonstrate poor speech and nonspeechsequencing skill over practice, and poor performance on retention and transfer tests compared
S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278 261
to PNS, and that (2) PWS demonstrate a transition to automaticity comparable to PNS duringspeech and nonspeech sequencing practice within a dual task paradigm. Some of these studyresults have been published elsewhere and details of subject recruitment and methodology canbe found in the published studies (Smits-Bandstra, De Nil, & Rochon, 2006; Smits-Bandstra,De Nil, & Saint-Cyr, 2006). However, we believe that the implications of the combined studyresults for the involvement of BG/CSTC connection dysfunction as a possible etiological factor instuttering are important for our understanding of the etiology and development of developmentalstuttering.
5. Study One: syllable sequence reading (Smits-Bandstra & De Nil, 2006)
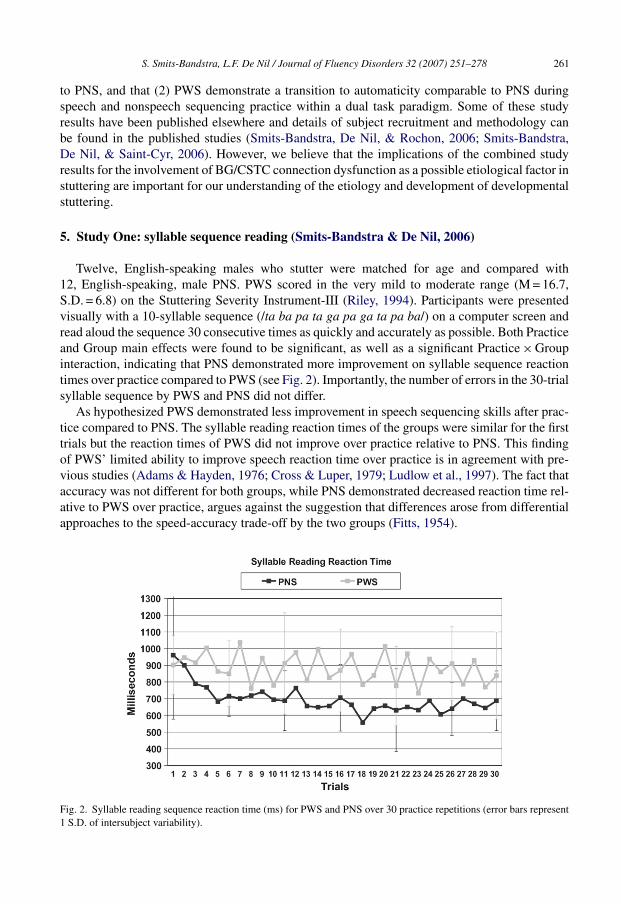
Twelve, English-speaking males who stutter were matched for age and compared with12, English-speaking, male PNS. PWS scored in the very mild to moderate range (M = 16.7,S.D. = 6.8) on the Stuttering Severity Instrument-III (Riley, 1994). Participants were presentedvisually with a 10-syllable sequence (/ta ba pa ta ga pa ga ta pa ba/) on a computer screen andread aloud the sequence 30 consecutive times as quickly and accurately as possible. Both Practiceand Group main effects were found to be significant, as well as a significant Practice × Groupinteraction, indicating that PNS demonstrated more improvement on syllable sequence reactiontimes over practice compared to PWS (see Fig. 2). Importantly, the number of errors in the 30-trialsyllable sequence by PWS and PNS did not differ.
As hypothesized PWS demonstrated less improvement in speech sequencing skills after prac-tice compared to PNS. The syllable reading reaction times of the groups were similar for the firsttrials but the reaction times of PWS did not improve over practice relative to PNS. This findingof PWS’ limited ability to improve speech reaction time over practice is in agreement with pre-vious studies (Adams & Hayden, 1976; Cross & Luper, 1979; Ludlow et al., 1997). The fact thataccuracy was not different for both groups, while PNS demonstrated decreased reaction time rel-ative to PWS over practice, argues against the suggestion that differences arose from differentialapproaches to the speed-accuracy trade-off by the two groups (Fitts, 1954).
Fig. 2. Syllable reading sequence reaction time (ms) for PWS and PNS over 30 practice repetitions (error bars represent1 S.D. of intersubject variability).
262 S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278
6. Study Two: finger-tap sequence learning (Smits-Bandstra, De Nil, & Saint-Cyr, 2006)
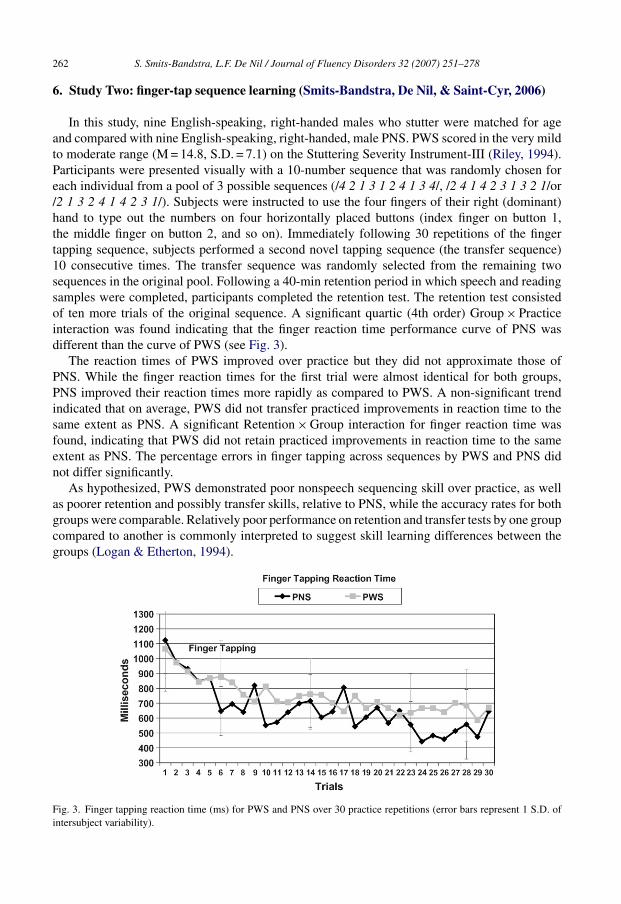
In this study, nine English-speaking, right-handed males who stutter were matched for ageand compared with nine English-speaking, right-handed, male PNS. PWS scored in the very mildto moderate range (M = 14.8, S.D. = 7.1) on the Stuttering Severity Instrument-III (Riley, 1994).Participants were presented visually with a 10-number sequence that was randomly chosen foreach individual from a pool of 3 possible sequences (/4 2 1 3 1 2 4 1 3 4/, /2 4 1 4 2 3 1 3 2 1/or/2 1 3 2 4 1 4 2 3 1/). Subjects were instructed to use the four fingers of their right (dominant)hand to type out the numbers on four horizontally placed buttons (index finger on button 1,the middle finger on button 2, and so on). Immediately following 30 repetitions of the fingertapping sequence, subjects performed a second novel tapping sequence (the transfer sequence)10 consecutive times. The transfer sequence was randomly selected from the remaining twosequences in the original pool. Following a 40-min retention period in which speech and readingsamples were completed, participants completed the retention test. The retention test consistedof ten more trials of the original sequence. A significant quartic (4th order) Group × Practiceinteraction was found indicating that the finger reaction time performance curve of PNS wasdifferent than the curve of PWS (see Fig. 3).
The reaction times of PWS improved over practice but they did not approximate those ofPNS. While the finger reaction times for the first trial were almost identical for both groups,PNS improved their reaction times more rapidly as compared to PWS. A non-significant trendindicated that on average, PWS did not transfer practiced improvements in reaction time to thesame extent as PNS. A significant Retention × Group interaction for finger reaction time wasfound, indicating that PWS did not retain practiced improvements in reaction time to the sameextent as PNS. The percentage errors in finger tapping across sequences by PWS and PNS didnot differ significantly.
As hypothesized, PWS demonstrated poor nonspeech sequencing skill over practice, as wellas poorer retention and possibly transfer skills, relative to PNS, while the accuracy rates for bothgroups were comparable. Relatively poor performance on retention and transfer tests by one groupcompared to another is commonly interpreted to suggest skill learning differences between thegroups (Logan & Etherton, 1994).
Fig. 3. Finger tapping reaction time (ms) for PWS and PNS over 30 practice repetitions (error bars represent 1 S.D. ofintersubject variability).

S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278 263
7. Study Three: dual task syllable sequence reading (Smits-Bandstra & De Nil, 2006)
Nine, native-English-speaking males who stutter were matched for age and compared withnine English-speaking male PNS. PWS scored in the very mild to moderate range (M = 17.3,S.D. = 7.6) on the SSI-III (Riley, 1994). A dual task paradigm was used in which subjects wereinstructed to complete two different tasks either separately or simultaneously. The two tasks usedwere a syllable reading sequencing task similar to Study One (see above), and a colour recognitiontask. The single and dual task conditions were repeated in a continuous, interleaved design (e.g.,30 trials of single task, 30 trials of dual task, 30 trials of single task, and 30 trials of dual task).The single condition syllable task was identical to Study One, with the exception that the 10-syllable ambiguous sequence was changed to/ga pa ta ba pa ta pa ga ba ta/. In the single taskcondition of the colour monitoring task, subjects were presented with a string of squares in aspecific colour (e.g., red). These squares changed colours three times before disappearing fromthe screen (e.g., red, blue, green, red). After viewing the stimulus squares, subjects were requiredto indicate as quickly as they could (by saying “yes”, or “no” aloud) whether or not the samecolour was presented twice.
In the dual task condition, the syllable sequence was presented in colour (e.g., red), thenchanged colours three times before disappearing from the screen (e.g., red, blue, green, red). Thesubjects read the sequence aloud as quickly and accurately as possible while at the same timepaying attention to the colours. After completing the sequence, subjects were required to sayaloud “yes” or “no” as quickly as they could in order to indicate whether or not the same colourwas presented twice. A significant Condition x Group interaction was found indicating that thereaction times of PNS were significantly faster under single task vs. dual task conditions while thereaction times of PWS were similarly slow under both single and dual task conditions. Relativelysmall group differences were apparent on the first practice trial under single task conditions,before practice. However this difference between the groups grew (often more than doubled) aspractice continued. No significant differences in group accuracy rates were found, however PWSdemonstrated a non-significant trend toward more errors on the colour monitoring task under dualtask conditions relative to PNS (see Fig. 4).
Fig. 4. Syllable sequence accuracy and colour monitoring accuracy for PWS and PNS under single and dual task conditions(error bars represent 1 S.D. of intersubject variability).

264 S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278
One of the most surprising and interesting findings of this study was that PWS’ reaction times,unlike those of PNS, did not differ across conditions. While PNS demonstrated two distinctperformance patterns, consisting of faster reaction times over practice during the single task, anda switch back to slower reaction times during the dual task condition, the performance of PWS, incontrast, was consistently slow, and essentially similar to the dual task performance of PNS. Thisfinding suggests that the motor planning/preparation system of the PWS was unable to transitionto an increasingly efficient, relatively automated mode during task performance.
8. Study Four: finger tapping dual task (Smits-Bandstra, De Nil, & Rochon, 2006)
The twelve PNS and PWS of this study were the same subjects described in Study One (seeabove). Additionally, subjects were matched for typing experience (hours per day). The fingertap sequence task was identical to Study Two with the exception that the sequence used was /31 3 2 4 1 4 2 3 1/ printed in black. The colour recognition procedure was identical to StudyThree with the exception that instead of saying “yes” or “no” aloud, subjects were required topress a “yes” or “no” button to indicate whether or not the same colour was presented twice. Twoimportant findings of the study are presented here. A significant three-way interaction was found,which revealed that the reaction times of PNS improved more after practice compared to PWSunder single task conditions only (see Fig. 5). PWS also demonstrated significantly more colourrecognition errors under single and dual task conditions compared to PNS.
As hypothesized, PWS demonstrated difficulty in transitioning to increasingly automatizedperformance compared to PNS. PNS demonstrated steeply sloping performance curves undersingle task conditions and shallow, gradually sloping performance under dual task conditions.This finding suggested that PNS’ performance improvement was hampered by limited attention.PWS demonstrated a consistent, shallow performance curve across conditions. One speculativeexplanation for this finding was that performing even a single task for PWS required the attentionalresources that would normally be reserved for dual tasks in PNS. In this study, group differenceswere apparent on the first practice trial under single task conditions. Similar to the results of Study
Fig. 5. Finger tapping sequence reaction time (ms) obtained by PWS and PNS over single and dual task conditions (errorbars represent 1 S.D. of intersubject variability).

S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278 265
Three, while initial group differences were relatively small, the difference between the groupsgrew (often more than doubled) as practice continued.
9. General discussion
9.1. Sequence skill learning
The most significant effect found for Studies One and Two was that PWS did not demonstratereaction time improvements over practice to the same extent as PNS. The deficits in learningefficiency were most apparent in the first 10 trials. Thus while performance was similar betweenthe two groups on the first trial, PNS improved their performance much more quickly in subsequenttrials. This early stage of skill acquisition has been termed “acquisition of set’ and has beendefined as a state of readiness (Evarts et al., 1984), optimization of task parameters (Friston, Frith,Passingham, Liddle, & Frackowiak, 1992), or establishing the “ballpark” of required responseparameters (Saint-Cyr, 2003). Grafton et al. (2002) proposed that attention can be used to optimizeadoption of set, or conversely that acquisition of set is negatively affected by lack of attention.
Acquisition of set can be observed as a steeply sloping curve of reaction time decreases overpractice trials (Newell & Rosenbloom, 1981; Schmidt & Wrisberg, 2004). Venkatagiri (2004)proposed that this reaction time decrease results because movement plans/memories have beenestablished and need only to be retrieved instead of built from ‘scratch’. This process of reactiontime improvement has also been described as a result of chunking (the close binding of previouslyindependent units; Anderson, 1982; Klapp, 2003), or strengthened/automatized stimulus-responseassociations (Schneider, Dumais, & Shiffrin, 1984).
Neuroscientific evidence suggests that all three of these proposals may simply be differentways of describing the same underlying phenomenon. Baddeley et al. (2000) state that memoriesare not inflexible structures but neurochemical traces in plastic neural networks. These traces,which develop after continued exposure to specific stimulus-response (SR) associations, becomeincreasingly resistant to interference as they become increasingly automatized (Baddeley et al.,2000). These movement traces become increasingly efficient, or automatized, over practice.
9.2. Dual task performance
The most significant effect found for Studies Three and Four was that PWS did not demonstrateautomatization of the practiced sequence to the same extent as PNS after practice. As stated earlierin this paper, neuroscientific research suggests that predictive/anticipatory signals from the BG tothe frontal cortex via the thalamus facilitate the shift to increasingly automated performance overpractice (Pasupathy & Miller, 2005; Shizgal & Arvanitogiannis, 2003). In accordance with thisresearch, PNS clearly demonstrated the ability to transition to the quick accurate performancecharacteristic of a relatively high level of automaticity from an attention-demanding, slow, vari-able, and effortful performance, observed over the first 10 trials of the first single task condition.One of the most interesting findings of the current studies was that PWS, on the other hand, demon-strated only one, consistent pattern of sequencing performance regardless of condition. PWS didnot demonstrate the ability to transition to quick accurate performance to the same extent asPNS. Instead, the performance of PWS was relatively slow, inaccurate, and attention-demandingas though system which triggered and facilitated the transition to increasing automaticity wasimpaired.

266 S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278
9.3. The nature of the proposed deficit
Based on a series of dual task experiments, Beilock, Carr, MacMahon, and Starkes (2002)proposed that continued conscious effort and attention focused on task performance preventsautomatization (or the expression of automatization) of SR associations over practice. Neurosci-entific evidence suggests that the optimization/automatization of SR binding can also be impededwhen there is insufficient sensory feedback for processing of SR associations (Graybiel, 1998), orwhen the timing of SR associations is not consistent/predictable (Kelso, 1995). Any/all of the threeexplanations of poor automaticity development presented above may serve as possible explana-tions for the poor dual task performance and poor reaction time improvements demonstrated byPWS relative to PNS over practice.
In concordance with Beilock et al.’s (2002) proposal of conscious effort/attention impedingautomatization, van Lieshout, Hulstijn, & Peters (1996) proposed that PWS choose to use a“controlled” strategy to avoid relying on an unstable motor control system. This hypothesis is basedon the idea that PWS are aware that their speech system is unstable, and in reaction to this instabilityhave developed a strategy of increased attention and effort to stabilize their speech system (Kelso,1995). However, the current studies found PWS to demonstrate impaired performance on thefinger-tapping sequencing task as well as the speech task. None of the PWS reported any awarenessor concern about instability of nonspeech motor control suggesting that conscious strategy usewas not likely the reason for performance differences on this task. In addition, PWS and PNSdid not differ on the first practice trial when instability, anxiety levels, and the motivation to usea controlled strategy was likely the highest. Instead performance differences between the twogroups grew over practice, a finding which also suggests that strategy use was not the primaryreason for performance differences.
As proposed by Graybiel (1998), automatization development can be impeded by insufficientsensory feedback for processing SR associations. This second proposal is a more likely explanationfor the sequence learning/automatization deficit demonstrated by PWS. This hypothesis is basedon the idea that performance improvements in early stages of skill learning are thought be relianton accurate sensory feedback (Schmidt & Wrisberg, 2004). Early in practice movements areguided by sensory feedback. Movement is performed more slowly and the demands on attentionalresources are high while sensorimotor processing is performed on-line to optimize movementparameters or “find the ballpark” (Schmidt & Wrisberg, 2004).
Both the cerebellum and the BG complex are thought to influence sensorimotor integrationearly in learning. Patients with cerebellar damage show poor error monitoring and self-correctionduring learning tasks (Fiez, Petersen, Cheney, & Raichle, 1992; Leiner, Leiner, & Dow, 1989). PPDdemonstrate an incomplete automatization of the sensorimotor integration process, forcing thesystem to continue to guide movement by sensory input (Graybiel, 1998). Over-reliance on visualfeedback is characteristic of novices, whereas experts rely predominantly on kinesthetic feedback(Beilock et al., 2002; Kelso, 1995). The automatization of sensorimotor integration builds theefficiency of kinesthetic-feedback-guided movement. The incomplete automatization of sensori-motor integration skills has been associated with impaired use of kinesthetic feedback and/or anover-reliance on visual feedback, such as is seen in the performance of novices (Smiley-Oyen etal., 2003). Both PPD (Smiley-Oyen et al., 2003) and PWS perform poorly relative to PNS on tasksrequiring sensorimotor integration without visual feedback (Archibald & De Nil, 1999; De Nil &Abbs, 1991; Forster & Webster, 2001; Howell, Sackin, & Rustin, 1995; Loucks & De Nil, 2001).
As proposed by Kelso (1995), inconsistent timing of SR associations may result in decreasedautomatization of SR binding. This third proposal is also a likely explanation for the sequence skill

S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278 267
learning/automatization deficit demonstrated by PWS is that stuttering is associated with deficientoptimization/automatization of SR binding due to unpredictable timing of SR associations duringpractice. This explanation is highly compatible with the proposal of a sensorimotor integrationdeficit discussed above, as the two explanations are not mutually exclusive. Instead, deficits insensorimotor integration may result in unpredictable timing of SR associations during practice.In addition, deficits in sensorimotor integration and SR automatization (Berardelli et al., 2001;Graybiel, 1998) are both known outcomes of the dysfunction in CSTC connections associatedwith Parkinson’s disease.
CSTC connections are thought to play a critical role in the timing of SR associations andunit-to-unit associations in sequencing, especially in the visuomotor sequence learning tasks suchas that used in the present studies. Berardelli and colleagues (2001) proposed that the role of theBG system is to reinforce and automatize SR association learning. Previous research has shownthat striatal neurons show rapid firing changes in the early trials of motor sequence learning. Pre-movement firing activity begins progressively earlier in these neurons during learning (Pasupathy& Miller, 2005). These striatal neurons provide a predictive signal for increased speed of responseto a specified stimulus (Shizgal & Arvanitogiannis, 2003). The interconnections between the BGcomplex and the frontal cortex are equally important for reinforcing associations between sub-sequent responses in a movement sequence as has been demonstrated by studies comparing PPDand matched control subjects on sequencing tasks (Benecke et al., 1986; Dominey et al., 1997).
9.4. Possible neurological correlates of the proposed deficit
Motor control deficits associated with PD are known to arise from deficiencies in BG functionand CSTC connections. A review of neuroimaging studies of PPD reveals decreased input fromthe BG to the SMA and primary motor cortex via the thalamus in these patients (Berardelli et al.,2001). These brain activity patterns coincided with sequence skill learning deficits as measuredby reaction time, movement duration, and accuracy (Berardelli et al., 2001). In particular, PPDdemonstrated impaired acquisition of set (Frith et al., 1986).
Neuroimaging studies contrasting the brain activity patterns of PWS and PNS reveal that PWStypically demonstrate structural abnormalities (Beal, Gracco, Lafaille, De Nil, in press; Sommer,Koch, Paulus, Weiller, & Buchel 2002), aberrant timing (Walla, Mayer, Deecke, & Thurner,2004), hypoactivation (Fox et al., 1996, 2000) of the BG (Wu, Riley, Maguire, Najafi, & Tang,1997), and circumscribed areas of the left prefrontal and premotor cortex which form part of theCSTC connections critical for acquisition of set and sequence skill learning. Similar to PPD, thesequence learning difficulties (e.g., poor acquisition of set) evidenced by PWS may have evolvedfrom aberrancies in these motor control structures/connections. These aberrancies disrupt theoptimization/automatization of SR binding. Neumann et al. (2003) proposed that hyperactivationof the right frontal operculum specifically may compensate for deficient CSTC transmissionsbetween Broca’s area and left-sided articulatory motor representations. Preibisch et al. (2003)concurred that hyperactivation of the right frontal operculum likely compensated for deficits notspecific to Broca’s area but areas “downstream” from Broca’s, in particular an anomaly of whitematter below the motor representation of the tongue and larynx identified by Sommer et al. (2002).
9.5. Explaining stuttering symptoms and characteristics
Any hypothesis of stuttering etiology must demonstrate the potential to further explain keydefining characteristics of developmental stuttering as well as a century of literature finding

268 S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278
differences between PWS and PNS on tasks ranging from foot tapping to eye movements tospeaking with delayed auditory feedback (Smith & Kelly, 1997).
9.5.1. Onset of developmental stutteringChildren who stutter are able to encode correctly and retrieve simple movements. They typically
do not stutter when babbling, producing single words, or producing early word combinations(Bloodstein, 1995). Perhaps it is because children who stutter have difficulty retrieving sequencedmovements and acquiring skill in complex sequence production that they typically start stutteringwhen producing words in a syntactically governed sequence (Bernstein Ratner, 1997). It is at thisage when speech sequencing skills must be automatized to ensure fluent adult-like speech rateand intonation with limited susceptibility to interference from competing task demands such aslanguage formulation.
Alm (2004) proposed that neurophysiological changes underly the ability to learn and autom-atize speech sequencing skills. Neuroscientific evidence suggests that learning of complexsequential skills by preschool/early school-age children likely reflects the development of thedopaminergic-dependent basal ganglia complex, CSTC connections, and the frontal/prefrontalcortex (Saint-Cyr & Taylor, 1992). Alm reviewed several studies reporting that dopamine recep-tor sites and receptor site availability (largely governed by the BG complex) change dramaticallybetween the ages of 3 and 6 when children most often manifest and recover from stuttering(Seeman et al., 1987).
9.5.2. Spontaneous recovery in stutteringMaturation of neurological structures of skill learning and automatization provides a tentative
explanation not only for recovery from stuttering but also for the disproportionate spontaneousrecovery of female children who stutter. The ratio of male to female adults who stutter is 4:1 whilethe ratio in children is approximately 1.5:1, suggesting that more girls recover from stutteringthan boys (Yairi & Ambrose, 1992). Rauch and Savage’s (1997) review of anatomical studiesindicated that women have larger caudate volumes than men, but this difference is the result of agradual shift during development. Using neuroimaging, Foundas (2003) found that by age five,children have already established typically left-lateralized activation of the middle and superiortemporal gyrus for auditory processing of language. It is possible that changes in lateralizationand growth within the BG complex, the caudate in particular, during development may result inspontaneous recovery in some children. In support of this hypothesis are the findings of relativelydecreased left-laterality of language processing areas and large caudate volumes of females ingeneral (Rauch & Savage, 1997), and the high rate of spontaneous recovery of female childrenwho stutter (Yairi & Ambrose, 1992).
9.5.3. Fluency enhancing conditions (FEC)Neuroimaging studies have revealed that skill learning and complex skill production is
redundantly represented in the brain with striato-thalamo-cortical circuits and cerebello-thalamo-cortical circuits (Jueptner & Weiller, 1998). These authors reported that the dominant use of thecerebellar circuit is largely determined when the skill to be learned required continuous, on-lineerror monitoring and reliance on sensory feedback (especially visual feedback). In contrast theBG circuit tended to be dominant for automatized tasks that are already proceduralized, inter-nalized (reliant on kinesthetic feedback), and chunked into sequences. When the BG circuit iscompromised, as in Parkinson’s disease, sequencing performance suffers (Benecke et al., 1986).In addition, neuroimaging studies have shown that PPD demonstrated increased activity in the

S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278 269
cerebellar circuit and performance reflecting dependence on the cerebellar circuit (i.e., controlled,monitored, and reliant on external sensory feedback). Alm (2004) suggested that perhaps fluencyenhancing conditions such as metronomic speech, paced speech, or chorus speech reduce stut-tering in PWS because they provide PWS with external sensory feedback, which cues cerebellarcircuits to “kick in” and compensate for a possibly deficient BG circuit. This proposal is supportedby neuroimaging results indicating hyper cerebellar activation in PWS relative to PNS (De Nil,Kroll, & Houle, 2001).
9.5.4. Treatment effectsCompensation for a dysfunctional BG circuit can also be attained by focusing conscious atten-
tion to task performance. Neuroimaging studies show that the simple act of focusing attentionto motor performance and sensory feedback from motor performance significantly enhances theneural activity in the sensorimotor integration areas (e.g., the primary motor cortex, primary sen-sory cortex, and superior temporal gyrus for speech), and prefrontal areas in the left, and less so theright cortex during sequencing (Johansen-Berg & Matthews, 2002). This effect is also observedfor PPD, and partially normalizes the decreased left frontal cortical activity in those areas due topoor input/facilitation by BG signaling.
Enhanced activity in prefrontal and sensorimotor integration areas is also seen when automa-tized tasks are “de-automatized” or segmented. For example Dogil et al. (2002) found that small,isolated tongue movements created more activation bilaterally in the motor and prefrontal corticesthen when the tongue movements were part of an automated speech movement.
Neuroimaging of PWS reveals hyperactivation of right hemisphere SMA, primary motor cortexM1, and premotor areas (BA 6, 46), as well as hypoactivation of left inferior frontal and precentralregions and temporal regions before treatment and more normalized patterns of decreased rightSMA and premotor areas and left temporal regions after treatment (De Nil, Kroll, Lafaille, &Houle, 2003). Saltuklaroglu, Kalinowski, and Guntupalli (2004) proposed that stuttering treatmentand delayed auditory feedback may cause PWS to “de-automatize” or focus conscious attention tocertain aspects of their speech and facilitate fluency through enhanced neural activity restoring theexpected pattern of left hemisphere dominance in motor planning and sensorimotor integrationareas.
9.6. Integrations of the proposed deficit with stuttering etiology theories
A deficit in the optimization/automatization of SR binding fits well with several stutteringetiology theories. The proposal that this SR binding deficit arises from the CSTC connections isan elaboration of the hypothesis of several researchers that stuttering is associated with dysfunctionof BG circuits (Alm, 2004; Jones et al., 2002), including the SMA (Forster & Webster, 2001),and the dopaminergic neurotransmitter system (Maguire et al., 2000).
This proposed deficit is also highly congruent with Max’s (2004) proposal that stuttering isassociated with an impaired “feedforward” mode of speech production. As reported by Weber-Fox(2001), studies comparing the electroencephalographic (EEG) records of PWS and PNS indicatedthat the process of stimulus perception is relatively unimpaired. Max defined stuttering as a deficitin the ability to retrieve the correct movement in response to stimuli quickly and in an increasinglyautomated (feedforward) manner due to an over-reliance on a slow, sensory feedback-dependentmode.
A deficit in the optimization/automatization of SR binding fits with Venkatagiri (2004), andHuinck, van Lieshout, Peters, and Hulstijn’s (2004) proposals. These authors proposed that stut-

270 S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278
tering is not associated with incomplete/incorrect stored movement synergies/memory traces(Venkatagiri, 2004). Rather, stuttering is a deficit in the binding/automatized retrieval of sub-sequent movement synergies in a sequence. For example, Huinck et al. discussed the commonobservation that PWS typically retrieve the initial phoneme of a word, but falter to retrieve thesubsequent vowel (e.g., s-s-s-s-song). Venkatagiri cites clinical observation and reviews severalstudies that confirm that stuttering severity is typically reduced under conditions where a visualor auditory “priming” stimulus is present (e.g., chorus speech, orthographic text, singing).
According to the covert repair hypothesis (Kolk & Postma, 1997), stuttering results fromexcessive monitoring and covert prearticulatory repair of abnormally frequent speech program-ming errors. Neuroscientific findings are compatible with parts of the covert repair hypothesis. Forexample, Preibisch et al. (2003) and Ingham et al. (2000) agreed that the breakdown in sequencingwithin frontal and prefrontal speech control areas (areas shown to be hypoactive in neuroimagingstudies) are compatible with possible deficiencies in phonological planning. Similarly, results ofthe current studies found delays in the reaction times of PWS also suggesting deficiencies in someaspect of output planning.
However, there is less support for Kolk and Postma’s (1997) contention that the wrong itemsare selected. Rather, there is evidence to suggest that PWS have delayed retrieval of syntacticand/or phonological sequence units for production which is partially overcome when primes arepresented (Anderson & Conture, 2004; Wijnen & Boers, 1994).
Intead of the “excessive monitoring” hypothesis (Postma, Kolk, & Povel, 1990), the poordual task performance of PWS in the current studies supports the demands/capacity model(Starkweather, Gottwald, & Halfond, 1990), and more specifically the ‘regression’ or ‘over-load’ hypothesis of stuttering etiology (Kamhi & McOsker, 1982). These models propose thatthe speech skills of PWS are not stable/insufficiently automatized and stuttering results because(1) task demands exceed a limited capacity of resources, and (2) poorly automatized speechskills are susceptible to competing task demands. Bosshardt (2002) elaborated on this hypothesis,stating that the occurrence of stuttering events depended on the amount of attention required forspeech and the amount of interference caused by other concurrently performed tasks that detractedattention from speech.
If stuttering is associated with a deficit in SR binding optimization/automatization, PWS woulddirect additional attentional resources to speech and require additional contributions from otherneural systems to compensate for the deficit. Therefore, a deficit in SR binding optimization mayresult in stuttering depending on whether the amount of attention required for speech and anyother simultaneous tasks surpasses the resource capacity. If SR binding was poorly automated,the SR associations would be relatively weak, unstable, and more susceptible to interference fromongoing activity. Poor SR binding optimization/automatization serves as a viable explanationfor the large variability within and among children who stutter and PWS across time, contexts,audience, and tasks.
10. Conclusions
The present studies were designed to investigate whether or not PWS demonstrated deficitsin sequence skill learning. The results from these speech and nonspeech studies present initialevidence that such deficits exist. Alm (2004) concluded that stuttering may arise from an impairedability of the BG and/or CTSC connections to produce timing cues for the initiation of thenext motor segment in speech, however the studies reviewed in this paper indicate a deficit insequence learning and automatization, rather than a simple deficit in sequencing performance. The

S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278 271
neuroscientific evidence about how stimulus-response connections are developed, optimized, andautomated over practice provides important information about the processes underlying sequenceskill learning and dual task performance. Because the current studies were limited to observingbehavioural differences between PWS and PNS, any proposals specifying underlying deficitswhich are potential causal factors in stuttering are speculative. Although speculative, the proposalthat stuttering may be associated with a deficit in SR binding optimization and automatizationserves as a viable explanation for the deficits observed in the current studies. This proposal alsopresents a plausible ‘common ground’ for several well-established theories of stuttering etiology.
As such, these results raise a plethora of new questions to be answered. One intriguingpossibility is that the neurophysiological correlates of differences between PWS and PNSmay originate in dysfunction of the distributed motor control system, more specifically CSTCconnections. Based on the findings of the current studies, a direct comparison of PWS and PBGon skill learning and dual tasks is called for. Such comparisons have been supported by authorssuch as Ludlow and Loucks (2003) and will likely reveal important neural deficiencies andcompensation that the two groups have in common. Future advances in this area will require bothstructural and functional brain imaging studies in order to investigate whether observed functionaldifferences can be correlated to structural differences at the cortical and/or subcortical level, oralternatively, whether observed structural differences constitute normal intersubject anatomicalvariability or indeed can be associated with atypical functional activation in stuttering speakers.Furthermore, behavioural paradigms done in conjunction with neuroimaging studies with finetemporal (EEG, MEG), spatial (fMRI), and spatiotemporal measurement capabilities (e.g.,electromagnetic midsagittal articulography or EMMA) are especially required to understand thecomplex nature of disorders of connectivity (and the resultant compensations, which stutteringis most likely to be.
CONTINUING EDUCATION
Sequence skill learning in persons who stutter: Implications for cortico-striato-thalamo-cortical dysfunction
QUESTIONS
1. Some of the similarities noted between patients with basal ganglia dysfunction (PBG) andpersons who stutter (PWS) include. . .a. developmental onsetb. environmental exacerbation of symptomsc. sensitivity to fluctuating dopamine levelsd. all of the above
2. A review of brain imaging studies reveals some compelling similarities between the brainactivity patterns shown by PBG and PWS including. . .
a. hypoactivation of the right frontal hemisphereb. hypoactivation of the cerebellumc. hypoactivation of circumscribed left motor areas including the primary motor cortex M1d. none of the above

272 S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278
3. Similar difficulties specific to sequence skill learning reported in the literature for both PBGand PWS include:a. poor acquisition of setb. poor retention but better transfer relative to control subjectsc. poor performance on short repetitive sequences but good performance on long complex
sequencesd. slow perception of sequential information but appropriate motor responses to sequential
stimuli4. Which of the following is a correct description of the review of the literature concerning
the role of the basal ganglia (BG) and its relevant connections for sequence learning andautomatization?a. the SMA contains tonically active neurons which signal the timing of practiced sequential
elementsb. striatal neurons provide a predictive signal for initiation of an increasingly automated,
quick, fluent movement sequencec. the BG and its relevant interconnections serve mainly to monitor feedback and correct
spatial errors of sequence movementd. all of the above
5. Results of recent studies conducted by Smits-Bandstra and colleagues suggest that:a. the speech and nonspeech sequence skill learning of PWS was impaired compared to fluent
speakersb. PWS demonstrated decreased levels of automaticity compared to control subjectsc. control subjects showed a sharp contrast in task performance when transitioning from low
to high levels of automaticity during practice while PWS did notd. all of the above
Acknowledgments
The studies were funded by a grant from the Natural Sciences and Engineering ResearchCouncil of Canada held by Dr. De Nil. The first author deeply appreciates the financial supportthat she has received during her graduate education from the Donalda Jean McGeachy Scholarshipand the Toronto Rehabilitation Institute.
References
Abwender, D. A., Trinidad, K. S., Jones, K. R., Como, P. G., Hymes, E., & Kurlan, R. (1998). Features resemblingTourette’s syndrome in developmental stutterers. Brain and Language, 62, 455–464.
Ackermann, H., Hertrich, I., Zieglers, W., Bitzer, M., & Bien, S. (1996). Acquired disfluencies following infarction of theleft mesiofrontal cortex. Aphasiology, 10, 409–417.
Adams, M. R., & Hayden, P. (1976). The ability of stutterers and nonstutterers to initiate and terminate phonation duringproduction of an isolated vowel. Journal of Speech and Hearing Research, 19, 290–296.
Alexander, M., Naeser, M., & Palumbo, C. (1987). Correlations of subcortical CT lesion sites and aphasia profiles. Brain,110, 961–991.
Alm, P. A. (2004). Stuttering and the basal ganglia circuits: A critical review of possible relations. Journal of Communi-cation Disorders, 37, 325–369.
Alsobrook, J., & Pauls, D. (1998). Molecular approaches to child psychopathology. Human Biology, 70, 413–432.Anderson, J. R. (1982). Acquisition of cognitive skill. Psychological Review, 89, 369–406.Anderson, J. A., & Conture, E. G. (2004). Sentence-structure priming in young children who do and do not stutter. Journal
of Speech Language and Hearing Research, 47, 552–571.

S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278 273
Andy, O., & Bhatnager, S. (1992). Stuttering acquired from subcortical pathologies and its alleviation from thalamicperturbation. Brain and Language, 42, 385–401.
Archibald, L., & De Nil, L. (1999). The relationship between stuttering severity and kinesthetic acuity for jaw movementsin adults who stutter. Journal of Fluency Disorders, 24, 25–42.
Baddeley, A., Bueno, O., Cahill, L., Fuster, J. M., Izquierdo, I., Mcgaugh, J. L., et al. (2000). The brain decade in debate.I: Neurobiology of learning and memory. Brazilian Journal of Medical and Biological Research, 33, 993–1002.
Beal, D., Gracco, V., Lafaille, S. & De Nil, L.F. (in press). Voxel-based morphometry of auditory and speech-relatedcortex of stutterers. NeuroReport.
Beilock, S. L., Carr, T. H., MacMahon, C., & Starkes, J. L. (2002). When paying attention becomes counterproductive:Impact of divided versus skill-focused attention on novice and experienced performance of sensorimotor skills. Journalof Experimental Psychology: Applied, 8, 6–16.
Benecke, R., Rothwell, J. C., Dick, J., Day, B., & Marsden, C. D. (1986). Performance of simultaneous movements inpatients with Parkinson’s disease. Brain, 109, 739–757.
Berardelli, A., Rothwell, J. C., Thompson, P. D., & Hallet, M. (2001). Pathophysiology of bradykinesia in Parkinson’sdisease. Brain, 124, 2131–2146.
Berman Hakim, H., & Bernstein Ratner, N. (2004). Nonword repetition abilities of children who stutter: An exploratorystudy. Journal of Fluency Disorders, 29, 179–199.
Bernstein Ratner, N. (1997). Stuttering: A psycholinguistic perspective. In R. F. Curlee & G. Siegel (Eds.), Nature andtreatment of stuttering: New directions, (2nd ed., pp. 99–127). Boston, MA: Allyn & Bacon.
Bloodstein, O. (1995). A Handbook of Stuttering (4th ed.). Chicago, IL: Easter Seals Society., p. 190.Bosshardt, H. G. (1999). Effects of concurrent mental calculation on stuttering, inhalation, and speech timing. Journal of
Fluency Disorders, 24, 43–72.Bosshardt, H. G. (2002). Effects of concurrent cognitive processing on the fluency of word repetition: Comparison between
persons who do and do not stutter. Journal of Fluency Disorders, 27, 93–114.Braun, A. R., Varga, M., Stager, S. V., Schulz, G. M., Selbie, S., Maisog, J. M., et al. (1997). Altered patterns of cerebral
activity during speech and language production in developmental stuttering: An H215O positron emission tomographystudy. Brain, 120, 761–784.
Brotchie, P., Iansek, R., & Horne, M. K. (1991). Motor function of the monkey globus pallidus. II: Cognitive aspects ofmovement and phasic neural activity. Brain, 114, 1685–1702.
Brown, R. G., Redondo-Verge, L., Chacon, J. R., Lucas, M. L., & Channon, S. (2001). Dissociation between intentionaland incidental sequence learning in Huntington’s disease. Brain, 124, 2188–2202.
Carluer, L., Marie, R. M., Lambert, J., Defer, G. L., Coskun, O., & Rossa, Y. (2000). Acquired and persistent stutteringas the main symptom of striatal infarction. Movement Disorders, 15, 343–346.
Catalan, M. J., Ishii, K., Honda, M., Samii, A., & Hallet, M. (1999). A PET study of sequential finger movements ofvarying length in patients with Parkinson’s disease. Brain, 122, 483–495.
Ciabarra, A. M., Elkind, M. S., Roberts, J. K., & Marshall, R. S. (2000). Subcortical infarction resulting in acquiredstuttering. Journal of Neurology, Neurosurgery, and Psychiatry, 69, 546–549.
Cooper, M. H., & Allen, G. D. (1977). Timing control and accuracy in normal speakers and stutterers. Journal of Speechand Hearing Research, 20, 55–71.
Cross, D. E., & Luper, H. L. (1979). Voice reaction time of stuttering and nonstuttering children and adults. Journal ofFluency Disorders, 4, 59–77.
De Nil, L. F., & Abbs, J. H. (1991). Oral and finger kinesthetic thresholds in stutterers. In H. F. M. Peters, W. Hulstijn, &C. W. Starkweather (Eds.), Speech motor control and stuttering, (pp. 123–130). New York, NY: Elsevier.
De Nil, L. F., & Bosshardt, H. G. (2001). Studying stuttering from a neurological and cognitive information processingperspective. In H. G. Bosshardt, J. S. Yaruss, & H. F. M. Peters (Eds.), Fluency disorders: Theory, research, treatment,and self-help: Proceedings of the third world conference of fluency disorders (pp. 53–58). Nijmegen, The Netherlands:Nijmegen University Press.
De Nil, L. F., Kroll, R. M., & Houle, S. (2001). Functional neuroimaging of cerebellar activation during single wordreading and verb generation in stuttering and nonstuttering adults. Neuroscience Letters, 302, 77–80.
De Nil, L. F., Kroll, R. M., Kapur, S., & Houle, S. (2000). A positron emission tomography study of silent and oralsingle word reading in stuttering and nonstuttering adults. Journal of Speech Language and Hearing Research, 43,1038–1053.
De Nil, L. F., Kroll, R. M., Lafaille, S. J., & Houle, S. (2003). A positron emission tomography study of short- andlong-treatment effects on functional brain activation in adults who stutter. Journal of Fluency Disorders, 28, 357–380.
Denny, M., & Smith, A. (1992). Gradations in a pattern of neuromuscular activity associated with stuttering. Journal ofSpeech and Hearing Research, 35, 1216–1229.

274 S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278
Dogil, G., Ackermann, H., Grodd, W., Haider, H., Kamp, H., Mayer, J., et al. (2002). The speaking brain: A tutorialintroduction to fMRI experiments in the production of speech, prosody, and syntax. Journal of Neurolinguistics, 15,59–90.
Dominey, P. F., Ventre-Dominey, J., Broussolle, E., & Jeannerod, M. (1997). Analogical transfer is effective in a serialreaction time task in Parkinson’s disease; evidence for a dissociable form of sequence learning. Neuropsychologia,35, 1–9.
Doyon, J., Gaudreau, D., Laforce, R., Jr., Castonguay, M., Bedard, P. J., Bedard, F., et al. (1997). Role of the striatum,cerebellum, and frontal lobes in the learning of a visuomotor sequence. Brain and Cognition, 34, 218–245.
Doyon, J., Laforce, R., Bouchard, G., Gaudreau, D., Roy, J., Poirier, M., et al. (1998). Role of the striatum, cerebellum, andfrontal lobes in the automatization of a repeated visuomotor sequence of movements. Neuropsychologia, 36, 625–641.
Dujardin, K., & Laurent, B. (2003). Dysfunction of the human memory systems: role of the dopaminergic transmission.Current Opinion in Neurology, 16, S11–S16.
Evarts, E., Shinoda, Y., & Wise, S. (1984). Neurophysiology approaches to higher brain function. New York, NY: JohnWiley & Sons., pp. 7–39.
Fattapposta, F., Pierelli, F., Travera, G., My, F., Mostarda, M., Alessio, C., et al. (2000). Preprogramming and controlactivity of bimanual self-paced motor task in Parkinson’s disease. Clinical Neurophysiology, 111, 873–883.
Fiez, J. A., Petersen, S. E., Cheney, M. K., & Raichle, M. E. (1992). Impaired non-motor learning and error detectionassociated with cerebellar damage. Brain, 115, 155–178.
Fitts, P. M. (1954). The information capacity of the human motor system in controlling the amplitude of movement.Journal of Experimental Psychology, 47, 381–391.
Fitzgerald, H. E., Cooke, P. A., & Greiner, J. R. (1984). Speech and bimanual hand organization in adult stutterers andnonstutterers. Journal of Fluency Disorders, 9, 51–65.
Folion, M., Tremblay, L., & Bedard, P. J. (1988). Abnormal influence of passive limb movement on the activity of globuspallidus neurons in Parkinsonian monkeys. Brain Research, 444, 165–176.
Forster, D. C., & Webster, W. G. (2001). Speech-motor control and interhemispheric relations in recovered and persistentstuttering. Developmental Neuropsychology, 19, 125–145.
Foundas, A. L. (2003). Is language laterality established by 5 years of age? Neurology, 60, 1573–1574.Fox, P. T., Ingham, R. J., Ingham, J. C., Hirsch, T. B., Hunter Downs, J., Martin, C., et al. (1996). A PET study of the
neural systems of stuttering. Nature, 382, 158–162.Fox, P. T., Ingham, R. J., Ingham, J. C., Zamarripa, F., Xiong, J. H., & Lancaster, J. L. (2000). Brain correlates of stuttering
and syllable production: A PET performance-correlation analysis. Brain, 123, 1985–2004.Friston, K. J., Frith, C. D., Passingham, R. E., Liddle, P. F., & Frackowiak, R. S. J. (1992). Motor practice and neurophys-
iological adaptation in the cerebellum: A positron tomography study. Proceedings of the Royal Society of London –Series B: Biological Sciences, 248, 223–228.
Frith, C. D., Bloxham, C. A., & Carpenter, K. N. (1986). Impairments in the learning and performance of a new manualskill in patients with Parkinson’s disease. Journal of Neurology, Neurosurgery, and Psychiatry, 49, 661–668.
Grafton, S. T., Hazeltine, E., & Ivry, R. B. (2002). Motor sequence learning with the nondominant left hand: A PETfunctional imaging study. Experimental Brain Research, 146, 369–378.
Graybiel, A. M. (1998). The basal ganglia and chunking of action repertoires. Neurobiology of Learning and Memory,70, 119–136.
Greiner, J. R., Fitzgerald, H. E., & Cooke, P. A. (1986). Speech fluency and hand performance on a sequential tappingtask in left-and right-handed stutterers and nonstutterers. Journal of Fluency Disorders, 11, 55–69.
Guadagnoli, M. A., Leis, B., Van Gemmert, A. W., & Stelmach, G. E. (2002). The relationship between knowledge ofresults and motor learning in Parkinsonian patients. Parkinsonism and Related Disorders, 9, 89–95.
Harrington, D. L., Haaland, K. Y., Yeo, R. A., & Marder, E. (1990). Procedural memory in Parkinson’s disease: Impairedmotor but not visuoperceptual learning. Journal of Clinical and Experimental Neuropsychology, 12, 323–339.
Helmuth, L. L., Mayr, U., & Daum, I. (2000). Sequence learning in Parkinson’s disease: a comparison of spatial-attentionand number-response sequences. Neuropsychologia, 38, 1443–1451.
Heuer, R. J., Thayer Sataloff, R., Mandel, S., & Travers, N. (1996). Neurogenic stuttering: Further corroboration of siteof lesion. Ear, Nose, and Throat Journal, 75, 161–168.
Horska, A., Kaufmann, W., Brant, L., Naidu, S., Harris, J., & Barker, P. (2002). In vivo quantitative proton MRSI studyof brain development from childhood to adolescence. Journal of Magnetic Resonance Imaging, 15, 137–143.
Houk, J. C., Buckingham, J. T., & Barto, A. G. (1996). Models of the cerebellum and motor learning. Behavioral andBrain Sciences, 19, 368–383.
Howell, P., Sackin, S., & Rustin, L. (1995). Comparison of speech motor development in stutterers and fluent speakersbetween 7 and 12 years old. Journal of Fluency Disorders, 20, 243–255.

S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278 275
Huinck, W. J., van Lieshout, P. H. H. M., Peters, H. F. M., & Hulstijn, W. (2004). Gestural overlap in consonant clusters:Effects on the fluent speech of stuttering and non-stuttering subjects. Journal of Fluency Disorders, 29, 3–25.
Ingham, R. J., Fox, P. T., Costello Ingham, J., & Zamarripa, F. (2000). Is overt stuttered speech a prerequisite for theneural activations associated with chronic developmental stuttering? Brain and Language, 75, 163–194.
Jahanshahi, M., Rowe, J., & Fuller, R. (2003). Cognitive executive function in dystonia. Movement Disorders, 18,1470–1481.
Johansen-Berg, H., & Matthews, P. M. (2002). Attention to movement modulates activity in sensorimotor areas, includingprimary motor cortex. Experimental Brain Research, 142, 13–24.
Jonas, S. (1981). The supplementary motor region and speech emission. Journal of Communication Disorders, 14,349–373.
Jones, R. D., White, A. J., Lawson, K. H. C., & Anderson, T. J. (2002). Visuoperceptual and visuomotor deficits indevelopmental stutterers: An exploratory study. Human Movement Science, 21, 603–619.
Jueptner, M., & Weiller, C. (1998). A review of differences between basal ganglia and cerebellar control of movementsas revealed by functional imaging studies. Brain, 121, 1437–1449.
Kamhi, A. G., & McOsker, T. G. (1982). Attention and stuttering: Do stutterers think too much about speech? Journal ofFluency Disorders, 7, 309–321.
Kandel, E., Schwartz, J., & Jessel, T. (2000). Principles of neural science (4th ed.). New York, NY: McGraw-Hill., pp.853–867.
Kelso, S. (1995). Dynamic patterns: The self-organization of brain and behaviour. Boston, MA: MIT Press.Klapp, S. T. (2003). Reaction time analysis of two types of motor preparation for speech articulation: Action as a sequence
of chunks. Journal of Motor Behaviour, 35, 135–150.Kolk, H., & Postma, A. (1997). Stuttering as a covert repair phenomenon. In R. F. Curlee & G. Siegel (Eds.), The nature
and treatment of stuttering (2nd ed., pp. 182–203). Needham Heights, MA: Allyn & Bacon.Korman, M., Raz, N., Flash, T., & Karni, A. (2003). Multiple shifts in the representation of a motor sequence during the
acquisition of skilled performance. Proceeding of the National Academy of Science USA, 100, 12492–12497.Laforce, R., & Doyon, J. (2001). Distinct contributions of the striatum and cerebellum to motor learning. Brain and
Cognition, 45, 189–211.Lehner, M., Taracha, E., Wislowska, A., Zienowicz, M., Maciejak, P., Skorzewska, A., et al. (2003). Neurodevelopmental
theories of schizophrenia—preclinical studies. Psychiatria Polska, 37, 627–639.Leiner, H. C., Leiner, A. L., & Dow, R. S. (1989). Reappraising the cerebellum: What does the hindbrain contribute to
the forebrain? Behavioral Neuroscience, 10, 998–1008.Liu, X., Tubbesing, S., Aziz, T., Miall, R., & Stein, J. (1999). Effects of visual feedback on manual tracking and action
tremor in Parkinson’s disease. Experimental Brain Research, 129, 477–481.Logan, G. D., & Etherton, J. L. (1994). What is learned during automatization? The role of attention in constructing an
instance. Journal of Experimental Psychology: Learning, Memory, and Cognition, 20, 1022–1050.Loucks, T., & De Nil, L. (2001). Oral kinesthetic deficits in stuttering evaluated by movement accuracy and tendon
vibration. In B. Maassen, W. Hulstijn, R. D. Kent, H. F. M. Peters, & P. H. H. M. van Lieshout (Eds.), Speech motorcontrol in normal and disordered speech: Proceedings from the fourth international speech motor conference (pp.307–310). Nijmegen, The Netherlands: Uitgeverij Vantilt.
Low, K., Miller, J., & Vierck, E. (2002). Response slowing in Parkinson’s disease: A psychophysiological analysis ofpremotor and motor processes. Brain, 125, 1980–1994.
Ludlow, C. L., & Loucks, T. (2003). Stuttering: A dynamic motor control disorder. Journal of Fluency Disorders, 28,273–295.
Ludlow, C. L., Rosenberg, J., Salazar, A., Grafman, J., & Smutok, M. (1987). Site of penetrating brain lesions causingchronic acquired stuttering. Annals of Neurology, 22, 60–66.
Ludlow, C. L., Siren, K., & Zikira, M. (1997). Speech production learning in adults with chronic developmental stuttering.In W. Hulstijn, H. F. M. Peters, & P. H. H. M. van Lieshout (Eds.), Speech production: Motor control, brain research,and fluency disorders: Proceedings from the 3rd world conference on speech motor production and fluency disorders(pp. 221–506). Amsterdam, The Netherlands: Elsevier.
Maguire, G. A., Gottschalk, L. A., Riley, G. D., & Franklin, D. L. (2000). Risperidone in the treatment of stuttering.Journal of Clinical Psychopharmacology, 20, 479–482.
Max, L. (2004). Stuttering and internal models for sensorimotor control: A theoretical perspective to generate testablehypotheses. In B. Maassen, R. D. Kent, H. F. M. Peters, P. H. H. M. van Lieshout, & W. Hulstijn (Eds.), Speech motorcontrol in normal and disordered speech (pp. 357–388). Oxford, NY: Oxford University Press.
Max, L., & Yudman, E. M. (2003). Accuracy and variability of isochronous rhythmic timing across motor systems instuttering versus nonstuttering individuals. Journal of Speech, Language, and Hearing Research, 46, 146–163.

276 S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278
Mentis, M. J., Dhawan, V., Feigin, A., Delalot, D., Zgaljardic, D., Edwards, C., et al. (2003). Early stage Parkinson’sdisease patients and normal volunteers: Comparative mechanisms of sequence learning. Human Brain Mapping, 20,246–258.
Mulligan, H., Anderson, T., Jones, R., Williams, M., & Donaldson, I. (2001). Dysfluency and involuntary movements: Anew look at developmental stuttering. International Journal of Neuroscience, 109, 23–46.
Namasivayam, A., & van Lieshout, P. (2004). Lip kinematics in people who stutter: Comparisons within and across days.Boston, MA: Poster session presented at the From Sound to Sense conference.
Neumann, K., Euler, H. A., von Gudenberg, W. A., Girard, A. L., Heinrich, L., Lanfermann, H., et al. (2003). Thenature and treatment of stuttering as revealed by fMRI: A within- and between-group comparison. Journal of FluencyDisorders, 28, 381–410.
Neumann, K., Preibisch, C., Euler, H. A., Wolff von Gudenberg, A., Lanfermann, H., Gall, V., et al. (2005). Corticalplasticity associated with stuttering therapy. Journal of Fluency Disorders, 30, 23–39.
Newell, A., & Rosenbloom, P. (1981). Mechanisms of skill acquisition and the law of practice. In J. Anderson (Ed.),Cognitive skills and their acquisition (pp. 1–56). Hillsdale, N.J: Erlbaum.
Pasupathy, A., & Miller, E. (2005). Different time courses of learning-related activity in the prefrontal cortex and striatum.Letters to Nature, 433, 873–876.
Pauls, D., Leckman, J., & Cohen, D. (1993). Familial relationship between Gilles de la Tourette Syndrome, attentiondeficit disorder, learning disability, speech disorders, and stuttering. Journal of the American Academy of Child andAdolescent Psychiatry, 32, 1044–1050.
Penfield, W., & Welch, K. (1951). The supplementary motor area of the cerebral cortex: A clinical and experimental study.Archives of Neurology and Psychiatry, 66, 289–317.
Postma, A., Kolk, H., & Povel, D. J. (1990). Speech planning and execution in stutterers. Journal of Fluency Disorders,15, 49–59.
Preibisch, C., Neumann, K., Raab, P., Euler, H., von Gudenberg, A., Lanfermann, H., et al. (2003). Evidence forcompensation for stuttering by the right frontal operculum. Neuroimage, 20, 1356–1364.
Rauch, S. L., & Savage, C. R. (1997). Neuroimaging and neuropsychology of the striatum: Bridging basic science andclinical practice. Neuropsychiatry of the Basal Ganglia, 20, 741–768.
Riley, G. (1994). A stuttering severity instrument for children and adults (3rd ed.). Austin, TX: Pro-Ed.Rowe, J., Stephan, K., Friston, K. J., Frackowiak, R., Lees, A., & Passingham, R. E. (2002). Attention to
action in Parkinson’s disease: impaired frontal connectivity among frontal cortical regions. Brain, 125, 276–289.
Roy, E. A., Saint-Cyr, J. A., Taylor, A. E., & Lang, A. E. (1993). Motor sequence learning in Parkinson’s disease.International Journal of Neuroscience, 73, 183–194.
Saint-Cyr, J. A. (2003). Frontal-striatal circuit functions: Context, sequence, and consequence. Journal of the InternationalNeuropsychological Society, 9, 103–127.
Saint-Cyr, J. A., & Taylor, A. E. (1992). The mobilization of procedural learning: The “key signature” of the basal ganglia.In N. Butters & L. R. Squire (Eds.), Neuropsychology of memory (pp. 188–202). New York, N.Y: The Guilford Press.
Saint-Cyr, J. A., Taylor, A. E., & Lang, A. E. (1988). Procedural learning and neostriatal dysfunction in man. Brain, 111,941–959.
Saltuklaroglu, T., Kalinowski, J., & Guntupalli, V. K. (2004). Towards a common neural substrate in the immediate andeffective inhibition of stuttering. International Journal of Neuroscience, 114, 435–450.
Sawaguchi, T., & Goldman-Rakic, P. (1991). D1 dopamine receptors in prefrontal cortex: Involvement in working memory.Science, 251, 947–950.
Schmidt, R. A., & Wrisberg, C. A. (2004). Motor learning and performance (3rd ed.). Champaign, IL: Human KineticsPublishers.
Schneider, W., Dumais, S. T., & Shiffrin, R. M. (1984). Automatic and control processing and attention. In R. Parasuraman& D. Davies (Eds.), Varieties of attention (pp. 1–27). Orlando: FL: Academic Press Inc.
Schulz, G. M., Leon, S., & Sulc, S. (2001). Speech motor learning in Parkinson’s disease. In B. Maassen, W. Hulstijn,R. D. Kent, H. F. M. Peters, & P. H. H. M. van Lieshout (Eds.), Speech motor control in normal and disorderedspeech: Proceedings from the fourth international speech motor conference (pp. 332–336). Nijmegen, the Netherlands:Uitgeverij Vantilt.
Seeman, P., Bzowej, N. H., Guan, H. C., Bergeron, C., Becker, L. E., Reynolds, G. P., et al. (1987). Human-braindopamine-receptors in children and aging adults. Synapse, 1, 399–404.
Seitz, R., & Roland, P. E. (1992). Learning of sequential finger movements in man: A combined kinematic and positronemission tomography (PET) study. European Journal of Neuroscience, 4, 154–165.
Shizgal, P., & Arvanitogiannis, A. (2003). Gambling on dopamine. Science, 299, 1856–1861.

S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278 277
Shohamy, D., Myers, C. E., Grossman, S., Sage, J., & Gluck, M. A. (2005). The role of dopamine in cognitive sequencelearning: evidence from Parkinson’s disease. Behavioural Brain Research, 156, 191–199.
Siegert, R. J., Taylor, K. D., Weatherall, M., & Abernethy, D. A. (2006). Is implicit sequence learning impaired inParkinson’s disease? A meta-analysis. Neuropsychology, 20, 490–495.
Smiley-Oyen, A. L., Worringham, C. J., & Cross, C. L. (2003). Motor learning processes in a movement-scaling task inolivopontocerebellar atrophy and Parkinson’s disease. Experimental Brain Research, 152, 453–465.
Smith, A., & Kelly, E. (1997). Stuttering: A dynamic, multifactorial model. In R. F. Curlee & G. Seigal (Eds.), The Natureand Treatment of Stuttering (pp. 204–217). Needham Heights, MA: Allyn and Bacon.
Smith, J. G., & McDowall, J. (2004). Impaired higher order implicit sequence learning on the verbal version of the serialreaction time task in patients with Parkinson’s disease. Neuropsychology, 18, 679–691.
Smith, J. G., Siegert, R., McDowall, J., & Abernethy, D. (2001). Preserved implicit learning on both the serial reactiontime task and artificial grammar in patients with Parkinson’s disease. Brain and Cognition, 45, 378–391.
Smits-Bandstra, S. & De Nil, L. F. (2006). Sequency skill learning and the transition to automaticity in adults who stutter.Dissertation Abstracts International, DAI-B 68/01, 288 (AAT NR22037).
Smits-Bandstra, S., De Nil, L. F., & Rochon, E. (2006). The transition to increased automaticity during finger sequencelearning in adult males who stutter. Journal of Fluency Disorders, 31, 22–42.
Smits-Bandstra, S., De Nil, L. F., & Saint-Cyr, J. A. (2006). Speech and nonspeech sequence skill learning in adults whostutter. Journal of Fluency Disorders, 31, 116–131.
Sommer, M., Koch, M. A., Paulus, W., Weiller, D., & Buchel, C. (2002). Disconnection of speech-relevant brain areas inpersistent developmental stuttering. The Lancet, 360, 380–383.
Starkweather, C. W., Gottwald, S., & Halfond, M. (1990). Stuttering prevention: A clinical method. Englewood Cliffs,N.J: Prentice-Hall.
Thomas, V., Reymann, J. M., Lieury, A., & Allain, H. (1996). Assessment of procedural memory in Parkinson’s disease.Progress in Neuro-Psychopharmacology and Biological Psychiatry, 20, 641–650.
van Lieshout, P. H. H. M., Hulstijn, W., & Peters, H. F. M. (1996). From planning to articulation in speech production:What differentiates a person who stutters from a person who does not stutter? Journal of Speech and Hearing Research,39, 546–564.
Van Borsel, J., Van Der Made, S., & Santens, P. (2003). Thalamic stuttering: A distinct clinical entity? Brain and Language,85, 185–189.
Van Borsel, J., Van Lierde, K., Van Cauwenberge, P., & Van Orshoven, M. (1998). Severe acquired stuttering followinginjury of the left supplementary motor region: A case report. Journal of Fluency Disorders, 23, 49–58.
Venkatagiri, H. S. (2004). Slower and incomplete retrieval of speech motor plans is the proximal source of stuttering:Stutters occur when syllable motor plans stored in memory are concatenated to produce the utterance motor plan.Medical Hypotheses, 62, 401–405.
Walla, P., Mayer, D., Deecke, L., & Thurner, S. (2004). The lack of focused anticipation of verbal information in stutterers:A magneto encephalographic study. Neuroimage, 22, 1321–1327.
Weber-Fox, C. (2001). Neural systems for sentence processing in stuttering. Journal of Speech, Language, and HearingResearch, 44, 814–825.
Webster, W. G. (1986). Neuropsychological models of stuttering-II: Interhemispheric interference. Neuropsychologia, 24,737–741.
Webster, W. G. (1988). Neural mechanisms underlying stuttering: Evidence from bimanual handwriting performance.Brain and Language, 33, 226–244.
Webster, W. G. (1989). Sequence initiation performance by stutterers under conditions of response competition. Brainand Language, 36, 286–300.
Weinstein, J., Caruso, A. J., Severing, K., & VerHoeve, J. (1989). Abnormalities of oculomotor control in stutterers[Abstract]. Investigative Ophthalmology and Visual Science, 30, 78.
Weiss, P., Stelmach, G. E., & Hefter, H. (1997). Programming of a movement sequence in Parkinson’s disease. Brain,120, 91–102.
Westwater, H., McDowall, J., Siegert, R., Mossman, S., & Abernethy, D. (1998). Implicit learning in Parkinson’s disease:Evidence from a verbal version of the serial reaction time task. Journal of Clinical and Experimental Neuropsychology,20, 413–418.
Wijnen, F., & Boers, I. (1994). Phonological priming effects in stutterers. Journal of Fluency Disorders, 19, 1–20.Wu, J. C., Maguire, G., Riley, G., Fallon, J., LaCasse, L., Chin, S., et al. (1995). A positron emission tomography (18F)
deoxyglucose study of developmental stuttering. Neuroreport, 6, 501–505.Wu, J. C., Riley, G., Maguire, G., Najafi, A., & Tang, C. (1997). PET scan evidence of parallel cerebral systems related
to treatment effects: FDG and FDOPA PET scan findings. In W. Hulstijn, H. F. M. Peters, & P. H. H. M. van Lieshout

278 S. Smits-Bandstra, L.F. De Nil / Journal of Fluency Disorders 32 (2007) 251–278
(Eds.), Speech production: Motor control, brain research, and fluency disorders: Proceedings from the third worldconference on speech motor production and fluency disorders (pp. 329–339). Amsterdam, The Netherlands: Elsevier.
Yairi, E., & Ambrose, N. (1992). A longitudinal study of stuttering children: A preliminary report. Journal of Speech andHearing Research, 35, 755–760.
Yairi, E., Ambrose, N., & Cox, N. (1996). Genetics of stuttering: A critical review. Journal of Speech and HearingResearch, 39, 771–784.
Sarah Smits-Bandstra obtained her Ph.D. degree in the Department of Speech-Language Pathology at the Universityof Toronto. She is a registered Speech-Language Pathologist and currently works at Erinoak, Canada. Her post doctoralresearch with Dr. Gracco at McGill University investigates the learning and retention of PWS and Parkinson’s patients.
Luc De Nil is professor and chair of the Department of Speech-Language Pathology at the University of Toronto, anAffiliated Scientist at the Toronto Western Research Institute and an Adjunct Scientist at the Hospital for Sick Children.His research interests include using behavioural and neuroimaging techniques to study the neural bases of developmentaland neurogenic stuttering, and speech disorders in Tourette Syndrome patients. He is Editor-in-Chief of the Journal ofCommunication Disorders.


















