
Competitor Rule Priming: Evidence for priming of task rules in task switching
17
ORIGINAL ARTICLE Competitor Rule Priming: Evidence for priming of task rules in task switching Maayan Katzir • Bnaya Ori • Shulan Hsieh • Nachshon Meiran Received: 12 February 2014 / Accepted: 2 June 2014 Ó Springer-Verlag Berlin Heidelberg 2014 Abstract In task-switching experiments, participants switch between task rules, and each task rule describes how responses are mapped to stimulus information. Importantly, task rules do not pertain to any specific response but to all possible responses. This work examined the hypothesis that task rules, as wholes, rather than (just) specific responses are primed by their execution, such that, in the following trial, response conflicts are exacerbated when the compet- ing responses are generated by these recently primed rules, and performance becomes relatively poor. This hypothesis was supported in two task-switching experiments and re- analyses of additional three published experiments, thus indicating Competitor Rule Priming. Importantly, the Competitor Rule-Priming effect was independent of response repetition vs. switch, suggesting that it reflects the priming of the entire task rule rather than the priming (or suppression) of specific responses. Moreover, this effect was obtained regardless of Backward Inhibition, suggest- ing these effects are unrelated. Introduction Consider the following situation: you are on your way to have lunch with your friends at the cafeteria, which is located near the post office. When you arrive to the cafe- teria and see the post office, you recall that you also have to mail a letter. In such a situation, the action plan of mailing the letter may compete with the action plan of having lunch with your friends. However, action plans are derived from abstract goals (e.g., the letter is your grant proposal, and thus serves the goal of obtaining this grant; see Kruglanski et al., 2002 for a similar idea). Thus, it is reasonable that resolving the competition between action plans may involve the goals that these actions serve. In this work, we provide behavioral evidence indicating that the degree of competition between action plans (i.e., responses) is determined in part by the abstract goals that govern these actions. To this end, we used a task-switching paradigm in which actions (key-presses) are governed by goals (to execute a specific task rule). Recent evidence suggests that the choice between con- flicting action plans takes place not only at the level of the plans themselves, but also at the level of the abstract rules that have generated these plans. Moreover, the argument is that such conflict resolution is performed by determining the relative strength (or activation) of competing action plans as well as between the competing task rules that generate them (Meiran, Hsieh, & Dimov, 2010; Astle, Jackson, & Swainson, 2012; Tipper, Weaver, & Houghton, 1994). For example, Meiran, Hsieh and colleagues showed recently that when an action plan generates a response conflict, the entire task rule that has generated the conflict is being suppressed. They called this rule ‘‘competitor rule’’. In the present work, we addressed the question what determines the strength of competition between action plans? And in specific, we asked how this strength is influenced by the priming of task rules? The methodology we used is somewhat unique. Prior research that employed two task setups (e.g., Allport, Styles, & Hsieh, 1994; Allport & Wylie, 2000; Waszak, Hommel, & Allport, 2003) could not distinguish between the priming of what became the relevant rule in Trial N vs. M. Katzir (&) Á B. Ori Á N. Meiran Ben Gurion University of the Negev, Beer-Sheva, Israel e-mail: [email protected] S. Hsieh National Cheng Kung University, Tainan, Taiwan 123 Psychological Research DOI 10.1007/s00426-014-0583-3
Transcript of Competitor Rule Priming: Evidence for priming of task rules in task switching

ORIGINAL ARTICLE
Competitor Rule Priming: Evidence for priming of task rulesin task switching
Maayan Katzir • Bnaya Ori • Shulan Hsieh •
Nachshon Meiran
Received: 12 February 2014 / Accepted: 2 June 2014
� Springer-Verlag Berlin Heidelberg 2014
Abstract In task-switching experiments, participants
switch between task rules, and each task rule describes how
responses are mapped to stimulus information. Importantly,
task rules do not pertain to any specific response but to all
possible responses. This work examined the hypothesis that
task rules, as wholes, rather than (just) specific responses
are primed by their execution, such that, in the following
trial, response conflicts are exacerbated when the compet-
ing responses are generated by these recently primed rules,
and performance becomes relatively poor. This hypothesis
was supported in two task-switching experiments and re-
analyses of additional three published experiments, thus
indicating Competitor Rule Priming. Importantly, the
Competitor Rule-Priming effect was independent of
response repetition vs. switch, suggesting that it reflects the
priming of the entire task rule rather than the priming (or
suppression) of specific responses. Moreover, this effect
was obtained regardless of Backward Inhibition, suggest-
ing these effects are unrelated.
Introduction
Consider the following situation: you are on your way to
have lunch with your friends at the cafeteria, which is
located near the post office. When you arrive to the cafe-
teria and see the post office, you recall that you also have to
mail a letter. In such a situation, the action plan of mailing
the letter may compete with the action plan of having lunch
with your friends. However, action plans are derived from
abstract goals (e.g., the letter is your grant proposal, and
thus serves the goal of obtaining this grant; see Kruglanski
et al., 2002 for a similar idea). Thus, it is reasonable that
resolving the competition between action plans may
involve the goals that these actions serve. In this work, we
provide behavioral evidence indicating that the degree of
competition between action plans (i.e., responses) is
determined in part by the abstract goals that govern these
actions. To this end, we used a task-switching paradigm in
which actions (key-presses) are governed by goals (to
execute a specific task rule).
Recent evidence suggests that the choice between con-
flicting action plans takes place not only at the level of the
plans themselves, but also at the level of the abstract rules
that have generated these plans. Moreover, the argument is
that such conflict resolution is performed by determining
the relative strength (or activation) of competing action
plans as well as between the competing task rules that
generate them (Meiran, Hsieh, & Dimov, 2010; Astle,
Jackson, & Swainson, 2012; Tipper, Weaver, & Houghton,
1994). For example, Meiran, Hsieh and colleagues showed
recently that when an action plan generates a response
conflict, the entire task rule that has generated the conflict
is being suppressed. They called this rule ‘‘competitor
rule’’. In the present work, we addressed the question what
determines the strength of competition between action
plans? And in specific, we asked how this strength is
influenced by the priming of task rules?
The methodology we used is somewhat unique. Prior
research that employed two task setups (e.g., Allport,
Styles, & Hsieh, 1994; Allport & Wylie, 2000; Waszak,
Hommel, & Allport, 2003) could not distinguish between
the priming of what became the relevant rule in Trial N vs.
M. Katzir (&) � B. Ori � N. Meiran
Ben Gurion University of the Negev, Beer-Sheva, Israel
e-mail: [email protected]
S. Hsieh
National Cheng Kung University, Tainan, Taiwan
123
Psychological Research
DOI 10.1007/s00426-014-0583-3

priming of what became the irrelevant rule in Trial N (see
more on that issue below). To distinguish between these
two options, studies must employ three or more rules.
Previous studies that used such setups mainly examined
what influences the priming of the relevant rule in Trial N
(e.g., Mayr & Keele, 2000; Meiran et al., 2010; Yeung &
Monsell, 2003a). In this study, we used a four-task setup
and focused on the priming of what became the irrelevant
rule in Trial N. In particular, we examined whether such
priming would influence the degree of interference posed
by the irrelevant rule when it becomes a competitor rule.
To this end, we capitalized on the Task-Rule Congruency
Effect (see Sudevan & Taylor, 1987, for the first demon-
stration and Meiran & Kessler, 2008, for review). In the
Task-Rule Congruency Effect, trials with competing
responses (i.e., incongruent trials) lead to performance cost
both in reaction time (RT) and proportion of errors (PE)
relative to trials with no competing responses (congruent
trials). Note that the interference in incongruent trials is
generated by responses. Nevertheless, considering that
responses are generated by task rules, we examined whe-
ther task rules have a unique contribution to the strength of
the interference in incongruent trials, above and beyond
that of responses. Our main assertion is that the strength of
the interference of the competitor rule (i.e., a rule that
generates a competing response) in Trial N is being
determined, in part, in Trial N–1 by either positive or
negative priming of this entire rule (for a similar idea see
Allport et al., 1994, and the ‘‘General discussion’’). In what
follows, we show that a rule which generates a competing
response (i.e., competitor rule) in Trial N poses enhanced
interference when this competitor rule was primed
beforehand, namely it was the relevant rule in Trial N–1.
We label this phenomenon Competitor Rule Priming.
Defining task rules
Prior to explaining the rationale for our assertion and
analysis, we explain in detail what we mean by ‘‘task rule’’,
and emphasize the difference between task rules and
response rules. Response rules associate keys (i.e.,
responses) with meanings. For example, a response rule
may associate key 1 with the meaning ‘red’ (e.g., IF red
PRESS key 1). In many task-switching paradigms, a few
meanings are attached to each key, and therefore key 1
could mean ‘green’ and also ‘circle’. In choice-RT tasks,
task rules involve several response rules. Each task rule
refers to one stimulus dimension (e.g., color, shape, etc.,).
Task rules therefore specify which stimulus dimension
determines the response, and also specify the correct
response to a given stimulus. As such, ‘task rule’ is a
broader concept than ‘response rule’.
The Task-Rule Congruency Effect
Task-switching experiments typically involve several task
rules that pertain to the same set of stimuli (e.g., colored
shapes, when switching between color classification and
shape classification). Such stimuli are labeled ‘‘multi-
valent’’. Moreover, the responses (typically, key-presses)
also overlap between the tasks (‘‘multivalent responses’’).
To illustrate, ‘‘IF red PRESS key 1; IF green PRESS key
2’’ is an example for a task rule that should be implemented
when the task is to classify the stimuli according to color.
On a given trial, a stimulus is presented and participants are
required to respond according to a specified task rule (e.g.,
color, henceforth ‘‘relevant rule’’). In such setups, the
correct response according to the relevant task rule (e.g.,
key 1) could be either congruent or incongruent with the
correct response generated by an irrelevant rule(s).
Whereas in congruent trials, the currently irrelevant task
rules activate the correct response (e.g., when classifying
the color of a red circle, both red and circle are mapped to
key 1), in incongruent trials, they activate a competing
response (e.g., when classifying the color of a green circle,
green is mapped to key 2 but circle is mapped to key 1).
The resultant interference in incongruent trials relative to
congruent trials is labeled ‘‘Task Rule Congruency Effect’’
(Sudevan & Taylor, 1987; Meiran & Kessler, 2008; see
Table 1). Additionally, the rule that generates a competing
response is labeled ‘‘competitor rule’’. Arguably, the Task-
Rule Congruency Effect seen in reaction time (RT-TRCE)
mostly reflects activated overlearned response category
codes in long-term memory (Kiesel, Wendt, & Peters,
2007; Meiran & Kessler, 2008; Wendt & Kiesel, 2008),
whereas the Task-Rule Congruency Effect observed in the
proportion of errors (PE-TRCE) mostly reflects the appli-
cation of the irrelevant task rule (Meiran & Daichman,
2005; cf. Steinhauser & Hubner, 2006).
The Task-Rule Congruency Effect is reflected in per-
formance costs in incongruent trials. This cost is enhanced
when the number of competitors increases (Meiran et al.,
2010). In this work, we additionally show that the inter-
ference from a single competitor rule is defined by the
priming of this rule in Trial N–1. Specifically, we show that
the execution of a rule primes this rule, and this priming
results in enhanced competition from that rule on the
subsequent trial.
Relevant sequential effects in task switching
Given the fact that the effect we demonstrate, namely,
Competitor Rule Priming, represents a sequential effect, in
the following section, we briefly review other sequential
effects in task switching, which are relevant to the current
Psychological Research
123

work. One such effect is ‘‘Backward Inhibition’’ (Mayr &
Keele, 2000; lag-2 repetition costs, Koch, Gade, Schuch, &
Philipp, 2010, for review). According to Backward Inhibi-
tion, in order to switch from task A to task B (in Trial N–1),
task A is being inhibited, presumably to facilitate the exe-
cution of task B. This inhibition incurs performance cost
when returning to task A (in Trial N). Thus, A–B–A
sequences are compared with C–B–A sequences, in which
rule C was inhibited in Trial N–1, and not rule A (see Table 1).
Competitor Rule Suppression is another relevant effect
that has been shown to be independent of Backward Inhi-
bition both in terms of its formal definition, and empirically
(Hsieh, Chang, & Meiran, 2012; Meiran et al., 2010;
Meiran, Hsieh, & Chang, 2011; see also Astle et al., 2012).
Competitor Rule Suppression presumably reflects the
operation of a mechanism that is responsible for resolving
response conflicts. Specifically, Meiran, Hsieh and col-
leagues showed that performance in Trial N was hampered
Table 1 Behavioral effects in task switching
Trial Effect Slow Moderately Slow
Moderately Quick
Quick
Task Rule Congruency Effect (competitor rules)
3 2 1 0
Trial N a1b2c2d2 a1b2c2d1 a1b2c1d1 a1b1c1d1
Backward Inhibition BI+ BI –
Trial N–2 a*b*c*d* a*b*c*d*
Trial N–1 a*b*c*d* a*b*c*d*
Trial N a*b*c*d* a*b*c*d*
Competitor RuleSuppression
CRS+ CRS –
Trial N–1 a1b2c*d* a1b1c*d*
Trial N a*b*c*d* a*b*c*d*
Competitor RulePriming
CRP+ CRP –
Trial N–1 a*b*c*d* a*b*c*d*
Trial N a2b1c1d1 a2b1c1d1
Competitor RuleRepetition
No Competitor
Rule Repetition
Competitor Rule
Repetition
Trial N–1 a1b*c1d2 a2b*c1d1
Trial N a2b1c*d1 a2b1c*d1
Lowercase letters (e.g., a, b) represent stimulus dimensions (e.g., color, shape) and the numbers on the right to each letter represent the dimension
values (e.g., red and green for the dimension color) that go with a specific response key (key 1, key 2) according to the task rules. The numbers
are replaced by an asterisk to denote that the key’s identity is unimportant. The relevant dimension is underscored and also emphasized by
boldface and italics. Thus, a1b1 represents a bivalent stimulus such as a circle that is also red in which both dimensions are associated with key 1
(namely, the correct response to circle and to red is key 1) and dimension A is the relevant dimension. a1b2 represents a bivalent stimulus such as
a circle that is also green in which the shape dimension is associated with key 1, but the color dimension is associated with key 2 and dimension
A is the relevant dimension. This logic is applied to four dimensions (a, b, c, d)
The critical comparison that defines a given phenomenon is always indicated by frame and a double-headed arrow /?. The condition (predicted
to be) associated with poorer performance (slow) is presented left to the arrow, whereas the quicker condition is presented on its right (with the
exception of the Task-Rule Congruency Effect, which does not have a double-headed arrow, because it is calculated as a linear trend)
Psychological Research
123

if the relevant task rule in this trial was a competitor rule in
Trial N–1. They termed this inhibitory effect Competitor
Rule Suppression, as it indicates that the competitor rule in
Trial N–1 was suppressed (see Table 1).
Backward Inhibition and Competitor Rule Suppression
are considered to reflect the carryover of inhibition/sup-
pression from from Trail N-1 to Trail N. Other sequential
effects are attributed to the carryover of activation, like the
switch cost asymmetry effect and the reversed Stroop
effect. Switch cost asymmetry is the increased task-
switching cost when switching from a difficult task (e.g.,
color naming) to an easy task (e.g., word reading) than
when switching in the opposite direction (cf. Allport &
Wylie, 2000; Barutchu, Becker, Carter, Hester, & Levy,
2013; Yeung & Monsell, 2003a, 2003b). Additionally,
when switching from (the difficult) color-naming task to
(the easy) word-reading task, an atypical interference effect
(known as ‘‘reversed Stroop effect’’, see Macleod, 1991,
for review) emerges. Specifically, a congruency effect in
the easy task (word reading) is observed, indicating inter-
ference from the (more difficult) color-naming task. Both
phenomena—the switch cost asymmetry and the reversed
Stroop effect—are attributed to the activation carryover of
the difficult task, which impairs performance when swit-
ched to the easy task (Allport & Wylie, 2000; Yeung &
Monsell, 2003a; but see Bryck & Mayr, 2008).
Whereas sequential effects in task switching usually
indicate influences on the relevant rule, in this work we
examine the sequential influence that priming has on a
competitor rule.
The current study
In the current study, we present direct behavioral evidence
to the influence that priming of a competitor rule has on the
degree of interference in incongruent trials. Our rationale
was that priming of the relevant rule in Trial N–1 is carried
over into Trial N, resulting in performance cost when the
previously relevant rule in Trial N–1 becomes the com-
petitor rule in Trial N. Thus, a competitor rule will incur a
high-performance cost when this competitor rule was
primed beforehand because it was the relevant rule in Trial
N–1. We compare this condition to trials in which the
competitor rule was not previously primed (as strongly),
because this rule was irrelevant in Trial N–1. The increased
competition from the previously primed competitor rule
should be seen in a performance cost in Trial N, thus
indicating Competitor Rule Priming.
We define ‘‘competitor rule’’ as an irrelevant rule that
generates response conflict. Note that we argue that it is the
entire rule, and not merely the response, that influences the
strength of the competition. If competition is determined at
the response level, then priming of the interfering response
will determine the amount of interference of a competing
response. However, if competition is determined at the rule
level, priming of the entire rule that generates this response
will determine the amount of interference of a competing
response, even if the specific response was not previously
primed. Our design enables us to examine the unique
contribution of task rules to the strength of the interference
in incongruent trials (as materializes in Competitor Rule
Priming). We will examine whether this phenomenon is
independent of the contribution of the response, by entering
the response variable (response repetition vs. response
switch) to the analysis.
Competitor Rule Priming will be demonstrated by
comparing trials in which the competitor rule in Trial N
was the relevant rule in Trial N–1 (CRP? trials) to trials in
which the competitor rule in Trial N was irrelevant in Trial
N–1 (CRP- trials). To illustrate, without loss of generality,
assume that rule B is the relevant rule in Trial N, and that
rule A is the only competitor rule in Trial N. In CRP?
trials, rule A was the relevant rule in Trial N–1. In CRP-
trials, rule C was the relevant rule in Trial N–1 (see
Table 1). The defining feature of Competitor Rule Priming
is that the competitor rule in Trial N was the relevant rule
in Trial N–1. The reason is that priming of the relevant rule
in Trial N–1 was carried into Trial N, resulting in a
stronger response competition in Trial N.
Note that, like with Competitor Rule Suppression and
Backward Inhibition, comparing CRP? trials to CRP-
trials cannot be performed in paradigms that incorporate
only two tasks. With regard to Competitor Rule Priming,
what distinguishes between CRP? and CRP- trials when
executing rule A (using the example from the previous
paragraph) is whether the relevant rule in Trial N–1 was
rule B or C, making it necessary to use at least three rules
(rules A, B, and C). For reasons that are clarified in the
Appendix, even three rules are insufficient to rule out
potential alternative accounts and a minimum of four rules
must be used.
To overcome this methodological problem, we used a
switching task that incorporated four tasks that were per-
formed on shapes (Experiment 1A) or digits (Experiment
1B). In both experiments, two tasks involved object clas-
sification (classifying the shape or the color of the object in
Experiment 1A; classifying the parity or magnitude of the
digit in Experiment 1B) and two tasks involved spatial
classification (classifying the shapes according to their
horizontal location or their vertical location). The motiva-
tion for running two experiments was not derived solely
from the need for replication and extension. In Experiment
1A, the decisions are perceptual and therefore stimulus
dimensions (i.e., color and shape) and the task rules are
indistinguishable. Competitor Rule Priming, if obtained,
Psychological Research
123

may indicate interference coming from a perceptual
dimension, not from a semantic task rule. In Experiment
1B, the object tasks involved semantic task rules, which
allowed us to examine whether a semantic task rule, that is
independent of a perceptual dimension, is primed in Trial
N–1 and then influences the strength of a competitor rule.
To maximize the need for control processes, we fol-
lowed Meiran et al. (2010, 2011) and used a relatively short
cue–target interval (Druey & Hubner, 2007; but see Grange
& Houghton, 2009) and avoided task repetitions (Philipp &
Koch, 2006). We predicted a performance cost in trials in
which the competitor rule was the relevant rule in Trial N–
1 (CRP?) relative to trials in which the competitor rule
was not the relevant rule in Trial N–1 (CRP-).
Methods
Participants
In each experiment, 24 undergraduate students from Ben-
Gurion University of the Negev participated in return for
partial course credit or for 35 NIS (approximately 9 US$).
The participants reported having normal or corrected-to-
normal vision.
Stimuli and procedure
The software was programmed in E-Prime 1.0 (Experiment
1A) and E-Prime 2.0 (Experiment 1B) (Schneider, Esch-
man & Zuccolotto, 2002).
Experiment 1A
The stimuli consisted of an object presented inside a 2 9 2
grid subtending a visual angle of approximately 13.10
(width) 9 13.10 (height) degrees. The object was a colored
shape, the stimuli were two triangles (2.38� 9 2.86�) and
two circles (diameter 2.38�), two in red and two in green,
forming all the four combinations of shape and color. The
task cues were task-related icons that were presented in the
center of the grid (Fig. 1a), while the target-object was
presented in the grid’s quadrants (Fig. 1b). We asked
participants to classify the stimuli according to the task
cue. In the object tasks, they classified the stimuli either
according to their color or according to their shape. In the
spatial classification tasks, they classified the stimuli either
according to their horizontal (left–right) location or
according to their vertical (up–down) location.
The experiment began with verbal explanation and
illustration of the tasks and the stimuli. Afterwards, the
participants were required to execute one practice block
and 18 experimental blocks. Each block consisted of 64
trials, rendering a total of 1,052 trials for each participant,
excluding the practice block. We gave the participants a
2-min rest between the blocks in an effort to keep them
alert. The total session took approximately 1 h and 10 min.
The participants were asked to be as accurate and quick as
possible.
A trial started with a response–cue interval of 500 ms in
which a black screen was presented. This was followed by
the task–cue presentation in the center of the empty grid
array for 500 ms. Then the object was added to the display,
which was kept on the screen until the response was given
(Fig. 1b). A beep sound of 400 MHz was heard after the
response if an error was made. The task always switched
from trial to trial.
The response key arrangement was counterbalanced
between the participants. Half of the participants used the
upper left (7) and lower right (3) keys on the keypad to
indicate up/left or down/right, while the other half used
upper right (9) and lower left (1) keys to indicate up/right
or down/left (Fig. 2). The assignment of object values
(such as circle, green, etc.,) to the same response keys used
for the spatial tasks was counterbalanced as well, yielding a
total of eight counterbalancing combinations. The keypad
was aligned with the center of the screen, and the partici-
pants were instructed to respond with their two index
fingers.
Experiment 1B
The procedure was identical to that of Experiment 1A with
a few minor changes. Instead of colored shapes, we used
8 9 6 mm digits (1–9, excluding 5). The task cues were
the Hebrew words for parity, magnitude, and task-related
icons for the spatial tasks. Therefore, in the object tasks,
participants classified the stimuli either according to their
magnitude (smaller or larger than 5) or according to their
parity (Schuch & Koch, 2003).
Results
We followed Meiran et al. (2010, 2011) and excluded trials
that followed an error either immediately or after two trials.
We analyzed RT only for correct trials and excluded from
the analysis RTs \100 ms (anticipatory errors) or longer
than 3,000 ms (outliers). For each analysis, we computed
the cell means separately for each participant.
Sequential phenomena in task switching
First, we wanted to make sure that our paradigm produces
well-documented effects in task switching (i.e., Backward
Inhibition, Competitor Rule Suppression). Table 2 shows
Psychological Research
123

that Backward Inhibition and Competitor Rule Suppression
were reliable.
Task-Rule Congruency Effect analysis
Because Competitor Rule Priming is a phenomenon relat-
ing to the interference coming from competitor rules,
before demonstrating Competitor Rule Priming, we wished
to demonstrate that competitors in fact yielded interference
in this paradigm. To do that, we analyzed the Task-Rule
Congruency Effect. Because there were four tasks, there
could be 0, 1, 2, or 3 competitor rules on a given trial (see
Table 1). The Task-Rule Congruency Effect was expected
to manifest as a linear trend showing that interference is
increased when the number of competitors increases
(Meiran et al., 2010). We included the relevant task in Trial
N as a variable in the analysis to see if the effect may be
task specific. The design of this analysis, therefore,
employed two independent variables: Task (i.e., shape,
color, horizontal, or vertical), and Number of Competitor
Rules (0, 1, 2, and 3). To estimate the Task-Rule Con-
gruency Effect as a linear trend, we used a linear contrast
(Pinhas, Tzelgov, & Ganor-Stren, 2012).
Experiment 1A
Response time (RT) A repeated measures ANOVA of
Task (shape, color, horizontal, vertical) 9 Number of
Fig. 1 a Stimuli and task cues. b Example of a trial and of cue–target pairs and response key arrangements in Experiment 1A
Fig. 2 The two possible mapping conditions for the spatial tasks.
a Participants respond with key 1 to down and left, and with key 9 to
up and right. b Participants respond with key 7 to up and left, and with
key 3 to down and right. Note that the object task’s responses were
also mapped to the same keys (1 and 9 in scenario a, 3 and 7 in
scenario b), in a counterbalanced order, resulting in eight mapping
conditions (4 in scenario a and 4 in scenario b). Figure 2 presents
only one possible mapping of the object tasks
Psychological Research
123

Competitors (0, 1, 2, 3) on RT yielded a main effect of
Task, F(3, 69) = 54.69, MSE = 84,807, p \ .001,
gp2 = .70, representing a difference between mean RT of
935, 898, 508, and 571 ms for shape, color, horizontal, and
vertical, respectively. More importantly, there was a sig-
nificant linear trend for Number of Competitors (i.e.,
the Task-Rule Congruency Effect), F(1, 23) = 30.86, MSE
= 8,250, p \ .001, gp2 = .57, representing a difference
between mean RT of 690, 721, 741, and 760 ms for 0, 1, 2,
and 3 competing rules, respectively. The interaction
between Task and the linear trend of Number of Compet-
itors was non-significant.
Proportion of errors (PE) A repeated measures ANOVA
with the same design yielded a main effect of Task, F(3,
69) = 11.82, MSE = .008, p \ .001, gp2 = .34, representing
a difference between mean PE of .07, .08, .01, and .04 for
shape, color, horizontal, and vertical, respectively. More
importantly, there was a significant linear trend for Number
of Competitors (i.e., the Task-Rule Congruency Effect),
F(1, 23) = 26.51, MSE = .009, p \ .001, gp2 = .54, repre-
senting a difference between mean PE of .02, .03, .05, and
.09 for 0, 1, 2, and 3 competing rules, respectively. The
interaction between Task and the linear trend of Number of
Competitors was also significant, F(3, 69) = 8.78, MSE =
.003, p \ .001, gp2 = .28, but the linear trend of Number of
Competitors was significant in all the tasks, F’s(1,
23) = 19.41, 17.48, 5.73, 17.74, MSE’s = .008, .007,
.0004, .002, p’s \ .05, gp2’s = .46, .43, .20, .44, for shape,
color, horizontal, and vertical, respectively.
Experiment 1B
RT The ANOVA yielded a main effect of Task, F(3,
69) = 37.49, MSE = 46,346, p \ .001, gp2 = .62, repre-
senting a difference between mean RT of 733, 828, 533,
and 588 ms for parity, magnitude, horizontal, and vertical,
respectively. More importantly, there was a significant
linear trend for Number of Competitors (i.e., the Task-Rule
Congruency Effect), F(1, 23) = 23.35, MSE = 7,710,
p \ .001, gp2 = .50, representing a difference between
mean RT of 640, 665, 674, and 702 ms for 0, 1, 2, and 3
competing rules, respectively. The interaction between
Task and the linear trend of Number of Competitors was
also significant, F(9, 207) = 5.54, MSE = 3,761,
p = .002, gp2 = .19, but the linear trend of Number of
Competitors was significant or marginally significant in all
the tasks, F’s = 3.04, 3.62, 6.62, 36.92, MSE’s 4,688,
5,345, 4,023, 4,938, p’s = .094, .070, .017, \.001,
gp2 = .12, .14, .22, .62, in the parity, magnitude, horizontal,
and vertical, respectively.
PE The ANOVA yielded a main effect of Task, F(3,
69) = 12.15, MSE = .006, p \ .001, gp2 = .35, representing
a difference between mean PE of .08, .08, .02, and .05 for
parity, magnitude, horizontal, and vertical, respectively.
More importantly, the linear trend for Number of Com-
petitors was also significant, F(1, 23) = 8.52, MSE = .01,
p = .008, gp2 = .27, representing a difference between
mean PE of .04, .04, .06, and .09 for 0, 1, 2, and 3 com-
peting rules, respectively. The interaction between Task
and the linear trend of Number of Competitors was non-
significant.
Competitor Rule-Priming analysis
To demonstrate the basic Competitor Rule-Priming effect,
we compared trials in which the competitor rule was the
relevant rule in Trial N–1 (CRP?) with trials in which the
competitor rule was irrelevant in Trial N–1 (CRP-). As
explained above, the response variable was included in the
analysis, to assure that Competitor Rule Priming has a
unique contribution above and beyond that of the response.
Task (i.e., shape, color, horizontal, or vertical) was also
included as independent variable in the analysis (to be
explained in the ‘‘Selection criteria’’ section below). The
design of this analysis, therefore, employed three inde-
pendent variables: Task (e.g., shape, color, horizontal, or
vertical), Response (repetition, switch) and Competitor
Rule Priming (CRP? vs. CRP-).
Selection criteria
The straightforward Competitor Rule-Priming analysis
included many confounds. Elaborated description of these
confounds and how we controlled for them can be found in
the ‘‘Appendix’’. Briefly, we equated CRP? trials and
CRP- trials on the degree of interference in Trial N
(namely, the number of competitor rules), the degree of
interference in Trial N–1 (Botvinick, Braver, Barch, Carter,
& Cohen, 2001), Competitor Rule Suppression, and
Table 2 Analysis of sequential phenomena in task switching
Size-RT (ms) gp2 Size-PE gp
2
Experiment 1A
BI 31* .42 \.001 \.001
CRS 26* .29 .007* .07
Experiment 1B
BI 18* .26 -.006* .13
CRS 15* .13 .014* .46
Effect size in RT (in ms) and PE of well-documented effects in task
switching: Backward Inhibition and Competitor Rule Suppression
BI Backward Inhibition, CRS Competitor Rule Suppression, RT
response time, PE proportion of errors
* One-sided significance, p \ .05
Psychological Research
123

competitor rule repetition (whether the competitor rule was
the same in Trials N–1 and N; Meiran et al., 2011).
Additionally, the other exclusion criteria led to the exclu-
sion of stimulus repetitions between Trials N–1 and N (see
‘‘Appendix’’). Eventually, we only analyzed trials with one
or two competing responses (because CRP- cannot be
materialized in trials with three competing rules, and
CRP? cannot be materialized in trials with zero competing
rules).
Controlling for all the possible confounds when con-
sidering Trial N–1 to Trial N transitions, reduces immen-
sely the amount of valid trials. Note that because tasks
were selected randomly, and so did the order of stimuli
selection, the number of trials in each condition varied for
each participant. In the switching task, which consisted of
1,052 trials, after excluding outlier trials and trials fol-
lowing errors, there were 203 trials (126–243), on average,
for each participant in the CRP- condition, and 206 trials
(129–256), on average in the CRP? condition, in Experi-
ment 1A. In Experiment 1B there were 196 trials
(133–228), on average, for each participant in the CRP-
condition, and 203 trials (132–248), on average in the
CRP? condition.
Given that performance on the spatial tasks was better
than performance on the object tasks, and given that tasks
were selected randomly, it could be that the random
selection would randomly yield more object trials in the
CRP? condition than in the CRP- condition. In such sit-
uations, the Competitor Rule-Priming effect would be a
result of noise coming from the task variable, and not the
real Competitor Rule-Priming effect. To control for this
possibility, we entered the task variable to the analysis.
When the task and the response variables were consid-
ered, there were 26 trials (8–44 in the CRP- condition and
8–41 in the CRP? condition), on average, in each cell in
the design in Experiment 1A. In Experiment 1B, there were
25 trials (10–36 in the CRP- condition and 12–41 in the
CRP? condition), on average, in each cell in the design.
Experiment 1A
RT A repeated measures ANOVA of Task (shape, color,
horizontal, vertical) 9 Response (repetition, switch) 9
Competitor Rule Priming (CRP?, CRP-) on RT yielded a
main effect of Task, F(3, 69) = 53.09, MSE = 83,896,
p \ .001, gp2 = .70, representing a difference between
mean RT of 924, 897, 511, and 569 ms for shape, color,
horizontal, and vertical, respectively. The analysis also
yielded a main effect of Response, F(1, 23) = 10.77, MSE
= 9,607, p = .003, gp2 = .32, indicating that responses were
slower when the response in Trial N–1 was repeated in trial
N (M = 742 ms) than when it switched (M = 709 ms). No
other effects were significant.
PE A similar ANOVA on PE yielded a main effect of
Task, F(3, 69) = 7.36, MSE = .01, p \ .001, gp2 = .24,
representing a difference between mean PE of .05, .07, .01,
and .04 for shape, color, horizontal, and vertical, respec-
tively. There was also a main effect for Response, F(1,
23) = 8.24, MSE = .004, p = .008, gp2 = .26, indicating
that participants performed more errors following response
repetition trials (M = .05), relative to response switch tri-
als (M = .03). The interaction Task 9 Response was also
significant, F(3, 69) = 4.13, MSE = .003, p = .009,
gp2 = .15, indicating that the response effect was present
only in the object tasks, F(1, 23) = 8.10, MSE = .008,
p = .009, gp2 = .26, but not in the spatial tasks, F \ 1.
More importantly, we found the predicted main effect of
Competitor Rule Priming, F(1, 23) = 4.76, MSE = .002,
p = .040, gp2 = .17, indicating that PE was higher in
CRP? trials (.05) than in CRP- trials (.04, demonstrating
a Competitor Rule-Priming effect of .01). Note that Com-
petitor Rule Priming did not interact with Task, Response,
or Task 9 Response interaction, Fs \ 1.
Experiment 1B
RT The ANOVA yielded a main effect of Task, F(3,
69) = 37.12, MSE = 47,308, p \ .001, gp2 = .62, repre-
senting a difference between mean RT of 736, 829, 538,
and 581 ms for parity, magnitude, horizontal, and vertical.
The main effects for Response, which indicated that
responses were slower when the response in Trial N–1
repeated in Trial N (M = 678 ms), than when it switched
(M = 664 ms), and the main effect for Competitor Rule
Priming, which indicated that as predicted, participants
were slower in CRP? trials (M = 677 ms) relative to
CRP- trials (M = 665 ms), failed to reach significance,
Fs(1, 23) = 2.67 and 2.69, MSE’s = 6,889 and 4,751,
ps = .116 and .114, gp2 = .10 and .10 for Response and
Competitor Rule Priming, respectively. Also, the interac-
tion, Task 9 Competitor Rule Priming was marginally
significant, F(3, 69) = 2.27, MSE = 4,838, p = .088,
gp2 = .09, indicating that the Competitor Rule-Priming
effect was significant in the spatial tasks (M’s = 573 and
546 ms, in the CRP? and CRP- trials, respectively), F(1,
23) = 7.94, MSE = 4,181, p = .010, gp2 = .26, but not in
the semantic tasks (Ms = 784 and 780 ms, in the CRP?
and CRP- trials, respectively), F \ 1. No other effects
were significant.
PE A similar ANOVA on PE yielded a significant main
effect of Task, F(3, 69) = 11.95, MSE = .005, p \ .001,
gp2 = .34, representing a difference between mean PE of
.08, .06, .02, and .05 for parity, magnitude, horizontal, and
vertical, respectively. The main effect for Response was
marginally significant, F(1, 23) = 2.97, MSE = .003,
Psychological Research
123
p = .099, gp2 = .11, indicating that participants performed
more errors following response repetition trials (M = .06),
relative to response switch trials (M = .05).
More importantly, we found the predicted main effect of
Competitor Rule Priming, F(1, 23) = 4.87, MSE = .001,
p = .038, gp2 = .17, indicating that PE was higher in
CRP? trials (.06) than in CRP- trials (.05, demonstrating
a Competitor Rule-Priming effect of .01). Note that Com-
petitor Rule Priming interacted with task and response,
F(3, 69) = 4.16, MSE = .001, p = .009, gp2 = .15, indi-
cating that the Competitor Rule-Priming effect was found
in the parity and the horizontal tasks when the response
switched, but in the magnitude task it was found when the
response repeated. At the very least, we can conclude that
the Competitor Rule-Priming effect is not systematically
related to response switch or repetition.
Discussion
Experiments 1A and 1B revealed all the standard effects.
More importantly, in both experiments, we found the
predicted main effect for Competitor Rule Priming in PE
(although in Experiment 1B it was present in some, but not
all the conditions), indicating that if the competitor rule in
Trial N was also the relevant rule in Trial N–1, it poses
higher interference than if it was not the relevant rule in
Trial N–1. Note that the Competitor Rule-Priming effect
was independent of response repetition (or at least, not
consistently related to it), suggesting that rule priming
influences the degree of interference posed by a competitor
rule, above and beyond that of responses. Given the small
Competitor Rule-Priming effect, which manifested only in
PE, we wished to examine the robustness of this effect
before drawing conclusions.
Is Competitor Rule Priming a robust phenomenon?
Given the small Competitor Rule-Priming effect, and
since it was consistently found only in PE, we wished to
examine the robustness of this effect. To do that, we
analyzed previously published experiments. Overall, we
analyzed the results of three experiments that were
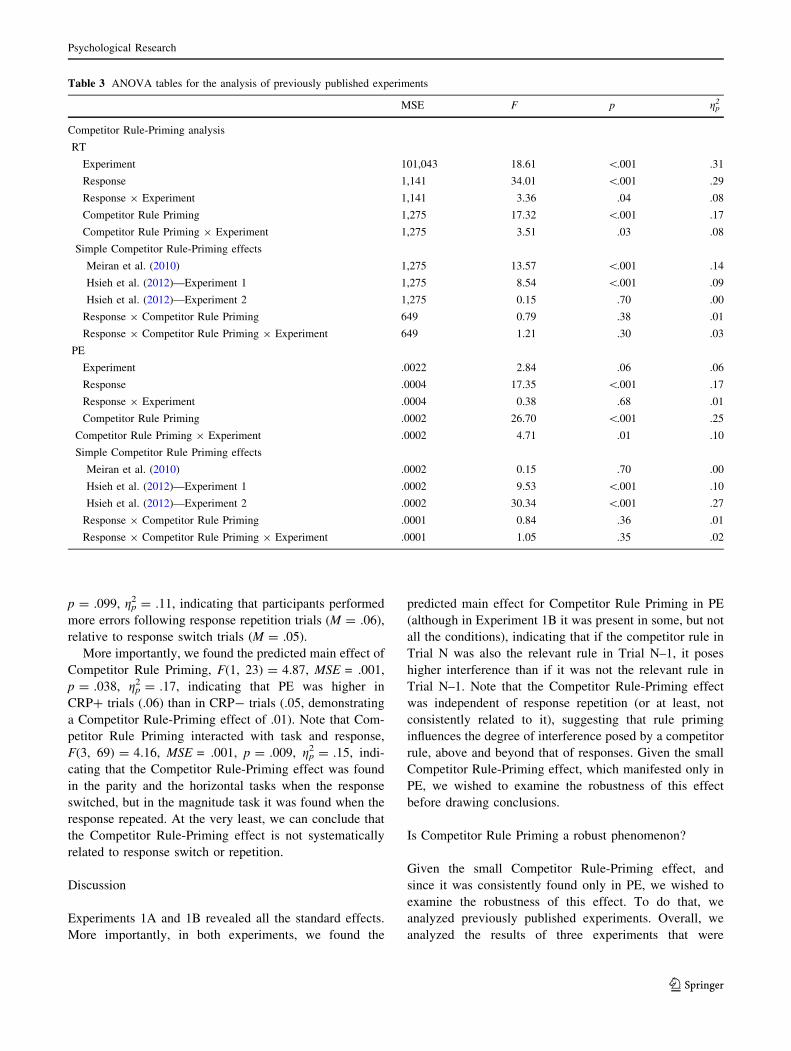
Table 3 ANOVA tables for the analysis of previously published experiments
MSE F p gp2
Competitor Rule-Priming analysis
RT
Experiment 101,043 18.61 \.001 .31
Response 1,141 34.01 \.001 .29
Response 9 Experiment 1,141 3.36 .04 .08
Competitor Rule Priming 1,275 17.32 \.001 .17
Competitor Rule Priming 9 Experiment 1,275 3.51 .03 .08
Simple Competitor Rule-Priming effects
Meiran et al. (2010) 1,275 13.57 \.001 .14
Hsieh et al. (2012)—Experiment 1 1,275 8.54 \.001 .09
Hsieh et al. (2012)—Experiment 2 1,275 0.15 .70 .00
Response 9 Competitor Rule Priming 649 0.79 .38 .01
Response 9 Competitor Rule Priming 9 Experiment 649 1.21 .30 .03
PE
Experiment .0022 2.84 .06 .06
Response .0004 17.35 \.001 .17
Response 9 Experiment .0004 0.38 .68 .01
Competitor Rule Priming .0002 26.70 \.001 .25
Competitor Rule Priming 9 Experiment .0002 4.71 .01 .10
Simple Competitor Rule Priming effects
Meiran et al. (2010) .0002 0.15 .70 .00
Hsieh et al. (2012)—Experiment 1 .0002 9.53 \.001 .10
Hsieh et al. (2012)—Experiment 2 .0002 30.34 \.001 .27
Response 9 Competitor Rule Priming .0001 0.84 .36 .01
Response 9 Competitor Rule Priming 9 Experiment .0001 1.05 .35 .02
Psychological Research
123

published elsewhere (Hsieh et al., 2012; Meiran et al.,
2010). The experiments used a similar design in which
there were four tasks, two involving location-based clas-
sifications and two involving object-based classifications.
Meiran et al. (2010) used the same spatial tasks as
reported in Experiments 1A and 1B, but the stimuli they
used were faces. Hsieh et al. (2012) used four boxes
arranged in a column and participants indicated whether
the stimulus’ location was up versus down or inner versus
outer. The stimuli were colored shapes. We employed the
same Competitor Rule-Priming analysis we performed in
Experiments 1A and 1B, with a few minor changes. Since
the tasks varied across experiments, we did not enter the
Task variable to the analysis. However, to prevent noise
from the Task variable, we created the cells in the ana-
lysis by averaging across task (i.e., giving equal weight to
each task). We also included the Experiment variable in
the analysis.
RT
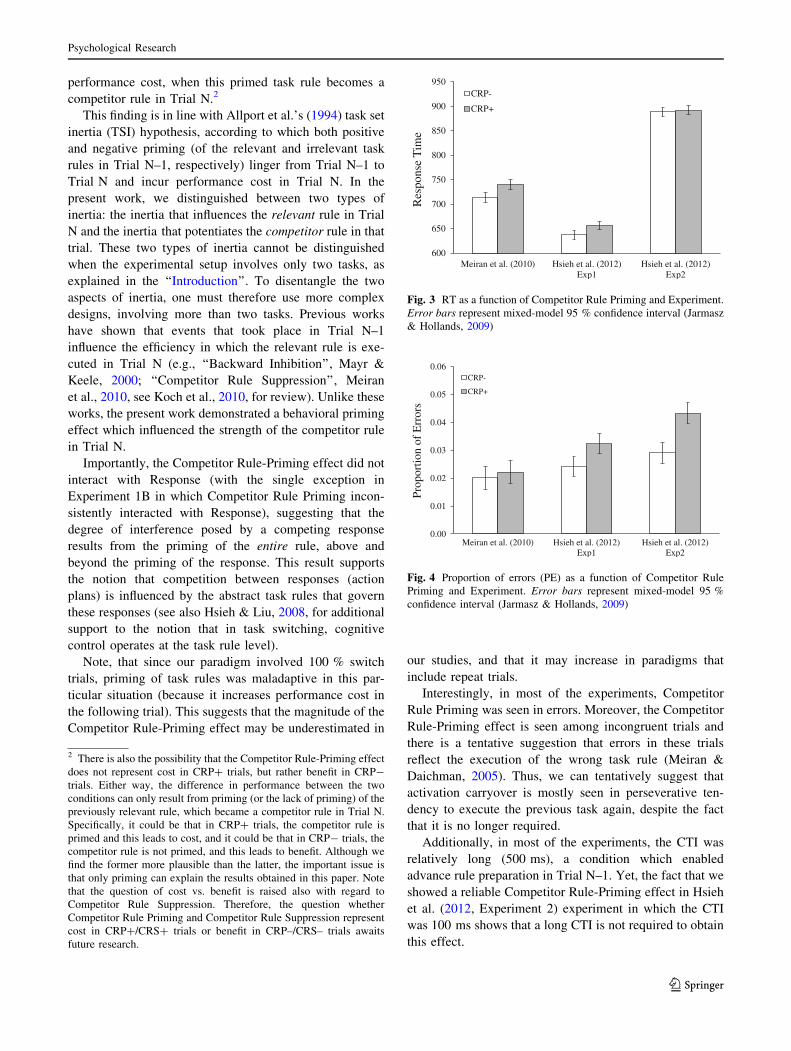
As Table 3 shows, a repeated measures ANOVA of
Response (repetition, switch) 9 Competitor Rule Priming
(CRP?, CRP-) with Experiment as a between-participants
variable showed a reliable main effect (gp2 = .17) for
competitor rule priming, indicating that as predicted,
responses were slower in CRP? trials (M = 769 ms) rel-
ative to CRP- trials (M = 754 ms, demonstrating a
Competitor Rule-Priming effect of 15 ms). The interaction
Competitor Rule Priming 9 Experiment was significant,
indicating that the Competitor Rule-Priming effect was
reliable in Meiran et al. (2010) and Hsieh et al. (2012)—
Experiment 1, but not in Hsieh et al. (2012)—Experiment
2, demonstrating a Competitor Rule-Priming effect of 27,
19 and 2 ms in Meiran et al. (2010), Hsieh et al. (2012)—
Experiment 1 and Hsieh et al. (2012)—Experiment 2,
respectively (see Fig. 3). Importantly, Competitor Rule
Priming did not interact with Response.
PE
As Table 3 shows, a similar ANOVA on PE also yielded a
reliable Competitor Rule-Priming main effect (gp2 = .25),
indicating that PE was higher in CRP? trials (.03) than in
CRP- trials (.02, demonstrating a Competitor Rule-Prim-
ing effect of .01). The interaction Competitor Rule Prim-
ing 9 Experiment was significant, indicating that the
Competitor Rule-Priming effect was reliable in Hsieh et al.
(2012)—Experiment 1 and Hsieh et al. (2012)—Experi-
ment 2, but not in Meiran et al. (2010), demonstrating a
Competitor Rule-Priming effect of .01, .01 and \.01 in
Hsieh et al. (2012)—Experiment 1, Hsieh et al. (2012)—
Experiment 2, and Meiran et al. (2010), respectively (see
Fig. 4). Importantly, Competitor Rule Priming did not
interact with response.
To conclude, Competitor Rule Priming is a robust
phenomenon, which was manifested in every experiment
either in RT or in PE,1 with no indication for speed accu-
racy tradeoff. Importantly, Competitor Rule Priming did
not interact with response, suggesting that the priming of
the entire rule contributes to the degree of the interference
posed by a competing response, above and beyond the
priming of the response.
General discussion
The present study asked the question what determines the
strength of competition between action plans. We spe-
cifically asked whether it is influenced by priming of
abstract task rules. Unlike previous studies, which tackled
similar questions by focusing on the strength of the rel-
evant rule in Trial N, we focused on the irrelevant rule in
Trial N and examined whether priming of the relevant
rule in Trial N–1 influences the degree of interference this
rule poses when it becomes the irrelevant competitor rule
in Trial N.
In two experiments using a task-switching paradigm that
incorporated four tasks and involved 100 % switch trials,
we found performance cost (mainly in PE) in trials in
which the competitor rule in Trial N was the relevant rule
in Trial N–1 (CRP? trials) relative to trials in which the
competitor rule in Trial N was not the relevant rule in Trial
N–1 (CRP– trials). Re-analyses of three additional pub-
lished experiments showed that Competitor Rule Priming,
despite being a small effect, is a robust effect that is usually
found in errors. This finding renders support to the idea that
rule execution (in Trial N–1) primes this task rule. This
priming then carries over to Trial N and incurs
1 It is important to note that the analyses reported in this paper
include a confound relating to the number of competitors in Trial N.
Specifically, due to the selection criteria of the Competitor Rule-
Priming effect, CRP? included more trials with two competitor rules
and fewer trials with one competitor rule than CRP– trials. This
confound works in favor of the Competitor Rule-Priming effect. To
assure that the Competitor Rule-Priming effect is not solely a result of
this confound, we ran another repeated measures ANOVA with
Congruency in Trial N, and Competitor Rule Priming as within
participants independent variables, and Experiment as between-
participants variable (cells were created by averaging across tasks).
The Competitor Rule-Priming effect remained significant both in RT,
F(1, 128) = 6.18, p = .014, gp2 = .05, and in PE, F(1, 128) = 19.51,
p \ .001, gp2 = .13. Importantly, the Competitor Rule-Priming effect
did not interact with Congruency in Trial N or with the interaction
Congruency in Trial N 9 Experiment, in RT and in PE, all Fs \ 1.
This analysis therefore rules out the aforementioned potential
confound.
Psychological Research
123
performance cost, when this primed task rule becomes a
competitor rule in Trial N.2
This finding is in line with Allport et al.’s (1994) task set
inertia (TSI) hypothesis, according to which both positive
and negative priming (of the relevant and irrelevant task
rules in Trial N–1, respectively) linger from Trial N–1 to
Trial N and incur performance cost in Trial N. In the
present work, we distinguished between two types of
inertia: the inertia that influences the relevant rule in Trial
N and the inertia that potentiates the competitor rule in that
trial. These two types of inertia cannot be distinguished
when the experimental setup involves only two tasks, as
explained in the ‘‘Introduction’’. To disentangle the two
aspects of inertia, one must therefore use more complex
designs, involving more than two tasks. Previous works
have shown that events that took place in Trial N–1
influence the efficiency in which the relevant rule is exe-
cuted in Trial N (e.g., ‘‘Backward Inhibition’’, Mayr &
Keele, 2000; ‘‘Competitor Rule Suppression’’, Meiran
et al., 2010, see Koch et al., 2010, for review). Unlike these
works, the present work demonstrated a behavioral priming
effect which influenced the strength of the competitor rule
in Trial N.
Importantly, the Competitor Rule-Priming effect did not
interact with Response (with the single exception in
Experiment 1B in which Competitor Rule Priming incon-
sistently interacted with Response), suggesting that the
degree of interference posed by a competing response
results from the priming of the entire rule, above and
beyond the priming of the response. This result supports
the notion that competition between responses (action
plans) is influenced by the abstract task rules that govern
these responses (see also Hsieh & Liu, 2008, for additional
support to the notion that in task switching, cognitive
control operates at the task rule level).
Note, that since our paradigm involved 100 % switch
trials, priming of task rules was maladaptive in this par-
ticular situation (because it increases performance cost in
the following trial). This suggests that the magnitude of the
Competitor Rule-Priming effect may be underestimated in
our studies, and that it may increase in paradigms that
include repeat trials.
Interestingly, in most of the experiments, Competitor
Rule Priming was seen in errors. Moreover, the Competitor
Rule-Priming effect is seen among incongruent trials and
there is a tentative suggestion that errors in these trials
reflect the execution of the wrong task rule (Meiran &
Daichman, 2005). Thus, we can tentatively suggest that
activation carryover is mostly seen in perseverative ten-
dency to execute the previous task again, despite the fact
that it is no longer required.
Additionally, in most of the experiments, the CTI was
relatively long (500 ms), a condition which enabled
advance rule preparation in Trial N–1. Yet, the fact that we
showed a reliable Competitor Rule-Priming effect in Hsieh
et al. (2012, Experiment 2) experiment in which the CTI
was 100 ms shows that a long CTI is not required to obtain
this effect.
600
650
700
750
800
850
900
950
Meiran et al. (2010) Hsieh et al. (2012)Exp1
Hsieh et al. (2012)Exp2
Res
pons
e T
ime
CRP-
CRP+
Fig. 3 RT as a function of Competitor Rule Priming and Experiment.
Error bars represent mixed-model 95 % confidence interval (Jarmasz
& Hollands, 2009)
0.00
0.01
0.02
0.03
0.04
0.05
0.06
Meiran et al. (2010) Hsieh et al. (2012)Exp1
Hsieh et al. (2012)Exp2
Prop
ortio
n of
Err
ors
CRP-
CRP+
Fig. 4 Proportion of errors (PE) as a function of Competitor Rule
Priming and Experiment. Error bars represent mixed-model 95 %
confidence interval (Jarmasz & Hollands, 2009)
2 There is also the possibility that the Competitor Rule-Priming effect
does not represent cost in CRP? trials, but rather benefit in CRP-
trials. Either way, the difference in performance between the two
conditions can only result from priming (or the lack of priming) of the
previously relevant rule, which became a competitor rule in Trial N.
Specifically, it could be that in CRP? trials, the competitor rule is
primed and this leads to cost, and it could be that in CRP- trials, the
competitor rule is not primed, and this leads to benefit. Although we
find the former more plausible than the latter, the important issue is
that only priming can explain the results obtained in this paper. Note
that the question of cost vs. benefit is raised also with regard to
Competitor Rule Suppression. Therefore, the question whether
Competitor Rule Priming and Competitor Rule Suppression represent
cost in CRP?/CRS? trials or benefit in CRP–/CRS– trials awaits
future research.
Psychological Research
123

We now turn to discuss a study which supports the
Competitor Rule-Priming effect. In Hubner et al. (2003)
study, participants switched between three tasks. Each task
was performed on different stimuli (i.e., digits, letters, and
shapes), and participants responded with different keys to
each task. Digits were categorized to odd and even, letters
to consonants and vowels, and shapes to shape with straight
lines only or shapes that also include curved lines. The
authors analyzed only switch trials. In these trials, the
target stimulus always appeared in the middle of the
screen, and it was either flanked by stimuli from the rele-
vant task in Trial N–1 (‘‘preceding’’ condition), or flanked
by stimuli form the third task (‘‘control’’ condition), or not
flanked at all (‘‘none’’ condition). In Experiment 1, half of
the switch trials were pre-cued with a cue that indicated the
relevant task in Trial N, and the other half they were not
cued. Experiment 2 was identical to Experiment 1, with the
exception that the cue only indicated a task switch, but did
not indicate which task is going to be the relevant task in
Trial N. The authors found performance costs on the
‘‘preceding’’ condition relative to the ‘‘control’’ condition
in three out of the four conditions, an effect reminiscent of
Competitor Rule Priming. However, when the pre-cue was
informative about the upcoming task (Experiment 1), they
found performance benefit on the ‘‘preceding’’ condition
(seemingly indicating a reversed Competitor Rule-Priming
effect). Hubner et al. (2003) interpreted this latter result as
expressing Backward Inhibition, suggesting that the for-
merly relevant task is being inhibited only when the to-be-
performed task is known in advance. Note that their latter
result is opposite to the results we obtained in the five
experiments reported here, in which tasks were always pre-
cued. Given the differences in the paradigms, we cannot
tell whether our results represent a failure to replicate
Hubner et al.’s results. However, note that unlike Hubner
et al.’s analyses, in our analyses we ruled out confounds
such as competitor rule repetition, Competitor Rule Sup-
pression, etc. (see ‘‘Appendix’’ for elaboration). In addi-
tion, unlike Hubner et al.’s paradigm, our paradigm
enabled the examination of response repetition effects, a
fact that enabled us to reach the conclusion that Competitor
Rule Priming represents priming of the entire task rule
rather than responses. Another advantage of our paradigm
is that it enables the examination of Backward Inhibition in
addition to Competitor Rule Priming. In this regard, we
find Competitor Rule Priming both in experiments that
show a reliable Backward Inhibition (3 out of the 5
experiments reported in this paper) and in experiments that
did not show a reliable Backward Inhibition (Experiments
1 and 2 in Hsieh et al., 2012), suggesting that Competitor
Rule Priming is unrelated to Backward Inhibition.
Finally, we presented a phenomenon that refers to the
priming of a competitor rule in Trial N if it was the relevant
rule in Trial N–1. Nevertheless, this work is limited in
deciding what is the cognitive process underlying this
phenomenon. The process underlying Competitor Rule
Priming may be the activation of the relevant rule when it
is executed (e.g., Allport & Wylie, 2000; Yeung & Mon-
sell, 2003a). However, the process underlying Competitor
Rule Priming may also be inhibition (e.g., Mayr & Keele,
2000; Koch et al., 2010), and more specifically the
decreased inhibition of that rule relative to other rules.
Namely, if we assume that executing a task is accompanied
by the inhibition of alternative tasks (e.g., Brown et al.,
2007; Mayr & Keele, 2000), task rules that were performed
more recently are inhibited to a lesser degree.3
To conclude, five experiments established the existence
of the Competitor Rule-Priming effect, in which priming of
the relevant rule in Trial N–1 is carried over to Trial N. The
Competitor Rule-Priming effect shows that interference
from competing action plans may be influenced by the
recent execution of the abstract rule that has generated
these plans.
Acknowledgments The research was supported by a Bi-National
Taiwan–Israel research grant to Shulan Hsieh and Nachshon Meiran,
by a research grant from the Israel Science Foundation to Nachshon
Meiran and by a research grant from the Israeli Foundation Trustees
to Maayan Katzir (Fund for Doctoral Students No. 30).
Appendix: Selection criteria
The straightforward Competitor Rule-Priming analysis
included many confounds, which involved the degree of
interference in Trial N (Meiran & Kessler, 2008), the
degree of interference in Trial N–1 (Botvinick et al., 2001),
Competitor Rule Suppression, and competitor repetition
(Meiran et al., 2011). In this section, we briefly explain
how we controlled for these confounds. CRP- can only
occur in trials with zero, one or two competing rules, but
not with three competing rules. This is because if there are
three competing rules, one of them must have been the
relevant rule in Trial N–1 (given that there are four tasks).
Furthermore, CRP? can only occur in trials with one, two
or three competing rules, but not with zero competing
rules, because to obtain Competitor Rule Priming there
must be a competitor rule to be primed. Given these con-
siderations, we excluded trials with zero or three competing
rules to remove this particular confound.
Moreover, we wanted to assure that CRP- and CRP?
trials are equally influenced by conflict in Trial N–1
(Botvinick et al., 2001) and conflict in Trial N (Meiran &
Kessler, 2008). Specifically, according to conflict adapta-
tion theory (Botvinick et al., 2001), control is recruited
3 We thank Andrea Kiesel for suggesting this option.
Psychological Research
123

Table 4 Selection criteria Competitor Rule Priming
Competitor Rule Priming
Competitor
Rule
Suppression
Competitor
Repetition
CRP – CRP +
Number of competitors in Trial N=0
CRP+ cannot be materialized: There cannot be CRP+ if there is no
competitor rule in Trial N.
Number of competitors in Trial N=1
Number of competitors in Trial N–1=0
– – Trial N–1: a1b1c1d1 Trial N–1: a1b1c1d1
Trial N: a1b1c2d1 Trial N: a1b1c2d1
+ There cannot be CRS+ or CRR+ trials if there was no competitor rule
in Trial N–1, because in CRS+ a competitor (in N–1) becomes the
relevant rule (in N) and in CRR+ the competitor (in N–1) remains the
competitor (in N).
+
Number of competitors in Trial N–1=1
– – Trial N–1: a2b1c1d1 Trial N–1: a2b1c1d1
Trial N: a1b1c2d1 Trial N: a1b1c2d1
+ CRP+ cannot be materialized: If the only competitor in Trial N was the
competitor in Trial N–1 (creating CRR+), it could not have been the
relevant rule in Trial N–1 (to create CRP+).
+ – Trial N–1:a1b2c1d1 Trial N–1: a1b2c1d1
Trial N: a1b1c2d1 Trial N: a1b1c2d1
+ If there was only one competitor in Trial N–1, it cannot both be the
competitor in Trial N (creating CRR+) and, at the same time, be the
relevant rule (creating CRS+).
Psychological Research
123

Table 4 continued
Number of competitors in Trial N–1=2
– – CRP– cannot be materialized: If the two competitors from Trial N–1
are neither the competitor in Trial N (creating CRR–) nor the relevant
in Trial N (creating CRS–), the relevant rule in Trial N–1 had to be the
competitor in Trial N (creating only CRP+ trials).
+ CRP+ cannot be materialized: if the only competitor in Trial N was the
competitor in Trial N–1 (creating CRR+), it could not have been the
relevant rule in Trial N–1 (to create CRP+).
+ – Trial N–1:a2b2c1d1 Trial N–1: a2b2c1d1
Trial N: a1b1c2d1 Trial N: a1b1c2d1
+ CRP+ cannot be materialized: if the only competitor in Trial N was the
competitor in Trial N–1 (creating CRR+), it could not have been the
relevant rule in Trial N–1 (to create CRP+).
Number of competitors in Trial N–1=3
_ There cannot be CRS– trials because if there were 3 competitors in
Trial N–1, one of them will become the relevant rule.
+ – CRP– cannot be materialized: If the competitor in Trial N was not a
competitor in Trial N–1 (CRR–) then it had to be the relevant (creating
only CRP+).
+ CRP+ cannot be materialized: If the only competitor in Trial N was the
competitor in Trial N–1 (creating CRR+), it could not have been the
relevant rule in Trial N–1 (to create CRP+) .
Number of competitors in Trial N=2
Number of competitors in Trial N–1=0
– – Trial N–1:a1b1c1d1 Trial N–1: a1b1c1d1
Trial N: a2b1c2d1 Trial N: a2b1c2d1
+ There cannot be CRS+ or CRR+ trials if there was no competitor rule
in Trial N–1, because in CRS+ a competitor (in N–1) becomes the
relevant rule (in N) and in CRR+ the competitor (in N–1) remains the
competitor (in N).
+
Psychological Research
123
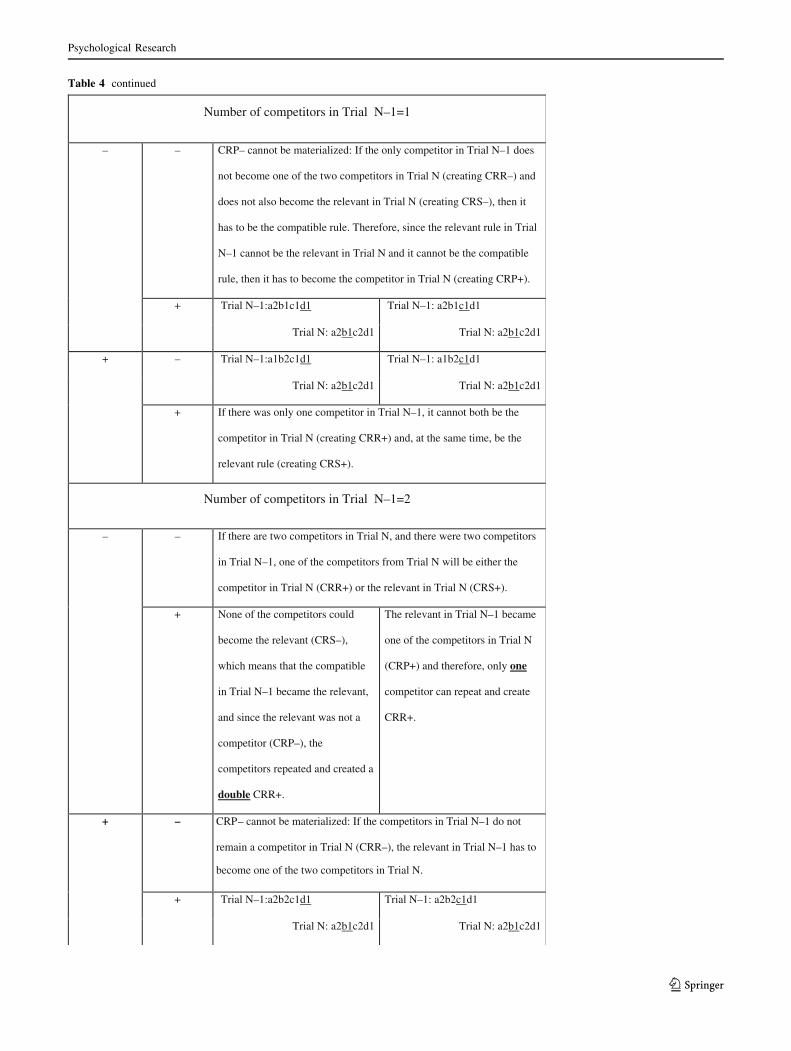
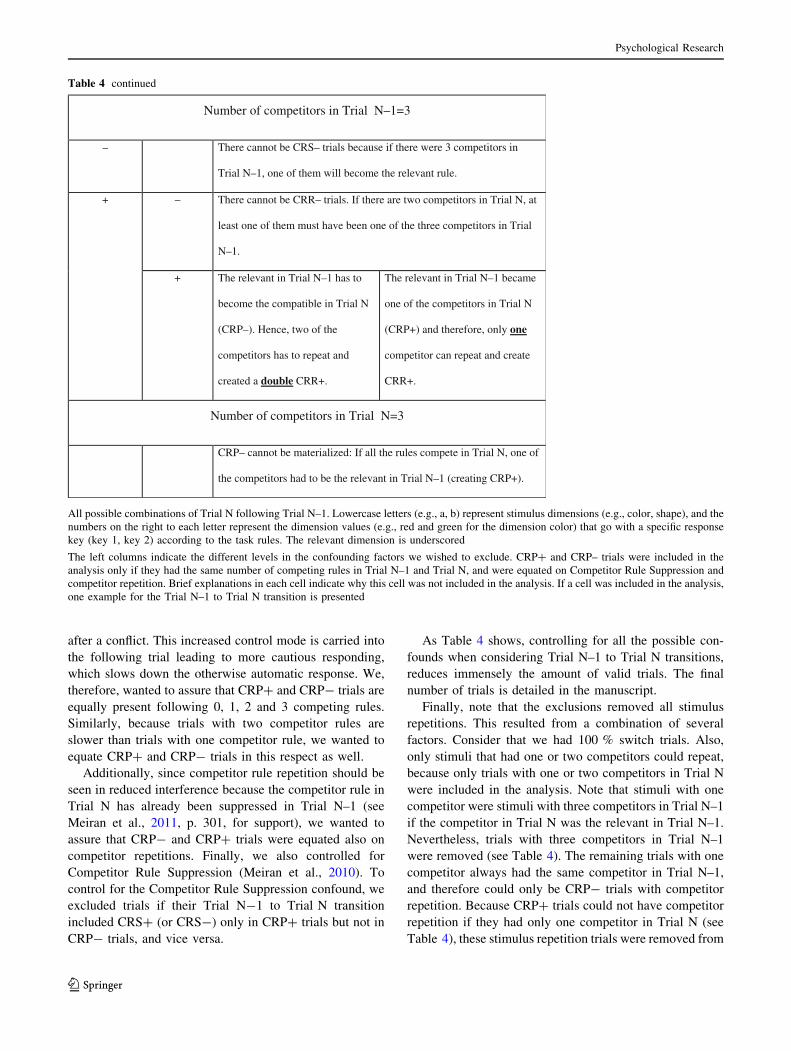
Table 4 continued
Number of competitors in Trial N–1=1
– – CRP– cannot be materialized: If the only competitor in Trial N–1 does
not become one of the two competitors in Trial N (creating CRR–) and
does not also become the relevant in Trial N (creating CRS–), then it
has to be the compatible rule. Therefore, since the relevant rule in Trial
N–1 cannot be the relevant in Trial N and it cannot be the compatible
rule, then it has to become the competitor in Trial N (creating CRP+).
+ Trial N–1:a2b1c1d1 Trial N–1: a2b1c1d1
Trial N: a2b1c2d1 Trial N: a2b1c2d1
+ – Trial N–1:a1b2c1d1 Trial N–1: a1b2c1d1
Trial N: a2b1c2d1 Trial N: a2b1c2d1
+ If there was only one competitor in Trial N–1, it cannot both be the
competitor in Trial N (creating CRR+) and, at the same time, be the
relevant rule (creating CRS+).
Number of competitors in Trial N–1=2
– – If there are two competitors in Trial N, and there were two competitors
in Trial N–1, one of the competitors from Trial N will be either the
competitor in Trial N (CRR+) or the relevant in Trial N (CRS+).
+ None of the competitors could
become the relevant (CRS–),
which means that the compatible
in Trial N–1 became the relevant,
and since the relevant was not a
competitor (CRP–), the
competitors repeated and created a
double CRR+.
The relevant in Trial N–1 became
one of the competitors in Trial N
(CRP+) and therefore, only one
competitor can repeat and create
CRR+.
+ – CRP– cannot be materialized: If the competitors in Trial N–1 do not
remain a competitor in Trial N (CRR–), the relevant in Trial N–1 has to
become one of the two competitors in Trial N.
+ Trial N–1:a2b2c1d1 Trial N–1: a2b2c1d1
Trial N: a2b1c2d1 Trial N: a2b1c2d1
Psychological Research
123
after a conflict. This increased control mode is carried into
the following trial leading to more cautious responding,
which slows down the otherwise automatic response. We,
therefore, wanted to assure that CRP? and CRP- trials are
equally present following 0, 1, 2 and 3 competing rules.
Similarly, because trials with two competitor rules are
slower than trials with one competitor rule, we wanted to
equate CRP? and CRP- trials in this respect as well.
Additionally, since competitor rule repetition should be
seen in reduced interference because the competitor rule in
Trial N has already been suppressed in Trial N–1 (see
Meiran et al., 2011, p. 301, for support), we wanted to
assure that CRP- and CRP? trials were equated also on
competitor repetitions. Finally, we also controlled for
Competitor Rule Suppression (Meiran et al., 2010). To
control for the Competitor Rule Suppression confound, we
excluded trials if their Trial N-1 to Trial N transition
included CRS? (or CRS-) only in CRP? trials but not in
CRP- trials, and vice versa.
As Table 4 shows, controlling for all the possible con-
founds when considering Trial N–1 to Trial N transitions,
reduces immensely the amount of valid trials. The final
number of trials is detailed in the manuscript.
Finally, note that the exclusions removed all stimulus
repetitions. This resulted from a combination of several
factors. Consider that we had 100 % switch trials. Also,
only stimuli that had one or two competitors could repeat,
because only trials with one or two competitors in Trial N
were included in the analysis. Note that stimuli with one
competitor were stimuli with three competitors in Trial N–1
if the competitor in Trial N was the relevant in Trial N–1.
Nevertheless, trials with three competitors in Trial N–1
were removed (see Table 4). The remaining trials with one
competitor always had the same competitor in Trial N–1,
and therefore could only be CRP- trials with competitor
repetition. Because CRP? trials could not have competitor
repetition if they had only one competitor in Trial N (see
Table 4), these stimulus repetition trials were removed from
Table 4 continued
Number of competitors in Trial N–1=3
– There cannot be CRS– trials because if there were 3 competitors in
Trial N–1, one of them will become the relevant rule.
+ – There cannot be CRR– trials. If there are two competitors in Trial N, at
least one of them must have been one of the three competitors in Trial
N–1.
+ The relevant in Trial N–1 has to
become the compatible in Trial N
(CRP–). Hence, two of the
competitors has to repeat and
created a double CRR+.
The relevant in Trial N–1 became
one of the competitors in Trial N
(CRP+) and therefore, only one
competitor can repeat and create
CRR+.
Number of competitors in Trial N=3
CRP– cannot be materialized: If all the rules compete in Trial N, one of
the competitors had to be the relevant in Trial N–1 (creating CRP+).
All possible combinations of Trial N following Trial N–1. Lowercase letters (e.g., a, b) represent stimulus dimensions (e.g., color, shape), and the
numbers on the right to each letter represent the dimension values (e.g., red and green for the dimension color) that go with a specific response
key (key 1, key 2) according to the task rules. The relevant dimension is underscored
The left columns indicate the different levels in the confounding factors we wished to exclude. CRP? and CRP– trials were included in the
analysis only if they had the same number of competing rules in Trial N–1 and Trial N, and were equated on Competitor Rule Suppression and
competitor repetition. Brief explanations in each cell indicate why this cell was not included in the analysis. If a cell was included in the analysis,
one example for the Trial N–1 to Trial N transition is presented
Psychological Research
123

the analysis. As for trials with two competitors in Trial N,
these trials can involve stimulus repetition only if there
were also two competitors in Trial N-1. Note that we
included trials with two competitors in Trial N and two
competitors in Trial N-1 only if they were CRS? trials with
competitor repetition (Table 4). In this scenario one of the
competitors necessarily remained the competitor rule and
the other necessarily became the relevant rule. Note that the
correct responses for these two rules had to be equal in Trial
N-1 (because they were both competitors). However, in
Trial N, these two rules were necessarily mapped to dif-
ferent keys (because one rule remained the competitor rule
and the other became the relevant rule), and therefore there
could not be stimulus repetition in this scenario.
References
Allport, A., Styles, E. A., & Hsieh, S. (1994). Shifting intentional set:
exploring the dynamic control of tasks. In C. Umilta & M.
Moscovitch (Eds.), Attention and performance XV: conscious
and nonconsciousinformation processing (pp. 421–452). Cam-
bridge: MIT Press.
Allport, D. A., & Wylie, G. (2000). ‘‘Task–switching’’, stimulus–
response bindings, and negative priming. In S. Monsell & J.
S. Driver (Eds.), Control of cognitive processes: attention and
performance XVIII (pp. 35–70). Cambridge: MIT Press.
Astle, D. E., Jackson, G. M., & Swainson, R. (2012). Two measures
of task-specific inhibition. The Quarterly Journal of Experimen-
tal Psychology, 65(2), 233–251.
Barutchu, A., Becker, S. I., Carter, O., Hester, R., & Levy, N. L.
(2013). The role of task-related learned representations in
explaining asymmetries in task switching. PLoS ONE, 8(4),
e61729. doi:10.1371/journal.pone.0061729.
Botvinick, M., Braver, T., Barch, D. M., Carter, C. S., & Cohen, J. D.
(2001). Conflict monitoring and cognitive control. Psychological
Review, 108(3), 624–652.
Brown, J. W., Reynolds, J. R., & Braver, T. S. (2007). A
computational model of fractionated conflict-control mecha-
nisms in task switching. Cognitive Psychology, 55(1), 37–85.
Bryck, R. L., & Mayr, U. (2008). Task selection cost asymmetry
without task switching. Psychonomic Bulletin and Review, 15(1),
128–134.
Druey, M., & Hubner, R. (2007). The role of temporal cue–target
overlap in backward inhibition under task switching. Psycho-
nomic Bulletin and Review, 14(4), 749–754.
Grange, J. A., & Houghton, G. (2009). Temporal cue–target overlap is
not essential for backward inhibition in task switching. Quarterly
Journal of Experimental Psychology, 62(10), 2068–2079.
Hsieh, S., Chang, C–. C., & Meiran, N. (2012). Episodic retrieval and
decaying inhibition in the competitor-rule suppression phenom-
enon. Acta Psychologica, 41(3), 316–321.
Hsieh, S., & Liu, H. (2008). Electrophysiological correlates of task
conflicts in task-switching. Brain Research, 1203, 116–125.
Hubner, M., Dreisbach, G., Haider, H., & Kluwe, R. H. (2003).
Backward inhibition as a means of sequential task-set control:
evidence for reduction of task competition. Journal of Exper-
imental Psychology. Learning, Memory, and Cognition, 29(2),
289–297.
Jarmasz, J., & Hollands, J. G. (2009). Confidence intervals in
repeated-measures designs: The number of observations
principle. Canadian Journal of Experimental Psychology, 63,
124–138.
Kiesel, A., Wendt, M., & Peters, A. (2007). Task switching: on the
origin of response congruency effects. Psychological Research,
71(2), 117–125.
Koch, I., Gade, M., Schuch, S., & Philipp, A. M. (2010). The role of
inhibition in task switching: a review. Psychonomic Bulletin and
Review, 17(1), 1–14.
Kruglanski, A. W., Shah, J. Y., Fishbach, A., Friedman, R., Chun, W.,
& Sleeth-Keppler, D. (2002). A theory of goal systems. In M.
P. Zanna (Ed.), Advances in experimental social psychology
(Vol. 34, pp. 331–378). San Diego: Academic Press.
MacLeod, C. M. (1991). Half a century of research on the Stroop effect:
an integrative review. Psychological Bulletin, 109(2), 163–203.
Mayr, U., & Keele, S. W. (2000). Changing internal constraints on
action: the role of backward inhibition. Journal of Experimental
Psychology: General, 129(1), 4–26.
Meiran, N., & Daichman, A. (2005). Advance task preparation
reduces task error rate in the cueing-task paradigm. Memory and
Cognition, 33, 1272–1288.
Meiran, N., Hsieh, S., & Chang, C. C. (2011). ‘‘Smart inhibition’’:
electrophysiological evidence for the suppression of conflict-
generating task rules during task switching. Cognitive, Affective
and Behavioral Neuroscience, 11(3), 292–308.
Meiran, N., Hsieh, S., & Dimov, E. (2010). Resolving task rule
incongruence during task switching by competitor rule suppres-
sion. Journal of Experimental Psychology: Learning, Memory,
and Cognition, 36(4), 992–1002.
Meiran, N., & Kessler, Y. (2008). The task rule congruency effect in
task switching reflects activated long term memory. Journal of
Experimental Psychology: Human Perception and Performance,
34(1), 137–157.
Philipp, A. M., & Koch, I. (2006). Task inhibition and task repetition
in task switching. European Journal of Cognitive Psychology,
18(4), 624–639.
Pinhas, M., Tzelgov, J., & Ganor-Stern, D. (2012). Estimating linear
effects in ANOVA designs: the easy way. Behavior Research
Methods, 44(3), 788–794.
Schneider, W., Eschman, A., & Zuccolotto, A. (2002). E-Prime
[Computer software]. Pittsburgh: Psychology Software Tools.
Schuch, S., & Koch, I. (2003). The role of response selection for
inhibition of task sets in task shifting. Journal of Experimental
Psychology: Human Perception and Performance, 29, 92.
Steinhauser, M., & Hubner, R. (2006). Response-based strengthening
in task shifting: evidence from shift effects produced by errors.
Journal of Experimental Psychology: Human Perception and
Performance, 32(3), 517–534.
Sudevan, P., & Taylor, D. A. (1987). The cuing and priming of
cognitive operations. Journal of Experimental Psychology:
Human Perception and Performance, 13, 89–103.
Tipper, S. P., Weaver, B., & Houghton, G. (1994). Behavioural goals
determine inhibitory mechanisms of selective attention. Quar-
terly Journal of Experimental Psychology, 47A, 809–840.
Waszak, F., Hommel, B., & Allport, A. (2003). Task-switching and
long-term priming: Role of episodic stimulus–task bindings in
task-shift costs. Cognitive psychology, 46, 361–413.
Wendt, M., & Kiesel, A. (2008). The impact of stimulus-specific
practice and task instructions on response congruency effects
between tasks. Psychological Research, 72(4), 425–432.
Yeung, N., & Monsell, S. (2003a). Switching between tasks of
unequal familiarity: the role of stimulus-attribute and response-
set selection. Journal of Experimental Psychology: Human
Perception and Performance, 29(2), 455–469.
Yeung, N., & Monsell, S. (2003b). The effects of recent practice on
task switching. Journal of Experimental Psychology: Human
Perception and Performance, 29(5), 919–936.
Psychological Research
123


















