
Biodiversität des Makrozoobenthos der österreichischen ... · Biodiversität des Makrozoobenthos...
20
Biodiversität des Makrozoobenthos der österreichischen Donau unter Berücksichtigung quantitativer Befunde der Freien Fließstrecke unterhalb Wiens C. FESL, U.H. HUMPESCH & E.R. Wöss Abstract: Biodiversity of the macrozoobenthos of the Austrian Danube, including quantitative stud- ies in a free-flowing stretch below Vienna. The Austrian part of the Danube is alpine in character, but only two unimpeded free-flowing stretches of the river remain - the Wachau region above Vienna, and the section below Vienna down to the Austrian border at river-km 1874. The latter section contains 310 species of macroinvertebrates, of which 58 species have not been found in other parts of the river upstream. For conservation of the rarer species of macroinvertebrates, the preservation of their natural habitat is vital; it is therefore important to preserve the unimpeded nature of the river below Vienna. Because the river is directly connected to its riverside forests and backwaters, an exchange of fauna can occur, thereby promoting a potential increase in biodiversity. In the main channel of the free-flowing section downstream of Vienna, the macroinvertebrate commu- nity was investigated in terms of biodiversity, spatial resource utilisation and persistence over a period of one year. Oligochaetes and chironomids accounted for 95 % of the total numbers of macroinverte- brates, the remaining five percent belonged to the Crustacea, Trichoptera, Bivalvia, Gastropoda, Bryo- zoa, Ephemeroptera and Hirudinea. Three species of bryozoans were recorded based on colony findings, and Fredericella sultana was most abundant. An analysis of the relationship between nine attributes of the communities and six environmental factors indicates that habitat stability expressed as sediment turnover was the major factor. Lower sediment turnover near the riverbanks coincided with increased densities and decreased species turnover. Highest values for species richness and total abundance occurred at a riverbank site of intermediate stability and greatest sediment heterogeneity, suggesting that the communities were structured by a combination of several factors. More studies are needed on the ecological relationship between native and recent invasive species, and the manner in which the latter may or will affect the current structures of communities within the riv- er fauna. Key words: Large river, sampling devices, invaders, biodiversity, bryozoan settlement, habitat stability. Einleitung diese Zahlen unterstreichen die ungeheure internationale Bedeutung dieses Flusses. Die Donau ist mit ihrer Länge von ca. 2.850 km der zweitgrößte Fluss Europas. Sie Die österreichische Donau hat eine Län- entspringt im Schwarzwald (Deutschland) ge von ungefähr 351 km und ein durch- und mündet in das Schwarze Meer (Rumä- schnittliches Gefälle von 40 cm pro Kilo- nien, Ukraine), in ihrem Einzugsgebiet lie- meter. Wie alle Gebirgsflüsse, so hat auch gen noch 12 weitere Länder: die Schweiz, die Donau in der österreichischen Strom- Österreich, Tschechische Republik, Slowa- strecke im Verlauf der Geschichte mannig- kei, Ungarn, Slowenien, Kroatien, Bos- faltige Änderungen des Längs- und Quer- nien/Herzegowina, Serbien/Montenegro, profils erfahren. Die systematische Regulie- Albanien, Bulgarien und Moldawien (Abb. rungstätigkeit begann im vorletzten Jahr- 1). In dem Einzugsgebiet von ca. 805.200 hundert und hatte einen schrittweisen Denisia 16, zugleich Kataloge km 2 leben rund 90 Millionen Menschen, Übergang vom Naturstrom zum gebändig- Neue Serie 28(2005), 139-158 © Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
-
Upload
duongduong -
Category
Documents
-
view
219 -
download
0
Transcript of Biodiversität des Makrozoobenthos der österreichischen ... · Biodiversität des Makrozoobenthos...

Biodiversität des Makrozoobenthos derösterreichischen Donau unter
Berücksichtigung quantitativer Befundeder Freien Fließstrecke unterhalb Wiens
C . F E S L , U . H . H U M P E S C H & E . R . W ö s s
Abstract: Biodiversity of the macrozoobenthos of the Austrian Danube, including quantitative stud-ies in a free-flowing stretch below Vienna. The Austrian part of the Danube is alpine in character, but
only two unimpeded free-flowing stretches of the river remain - the Wachau region above Vienna, and
the section below Vienna down to the Austrian border at river-km 1874. The latter section contains
310 species of macroinvertebrates, of which 58 species have not been found in other parts of the river
upstream. For conservation of the rarer species of macroinvertebrates, the preservation of their natural
habitat is vital; it is therefore important to preserve the unimpeded nature of the river below Vienna.
Because the river is directly connected to its riverside forests and backwaters, an exchange of fauna can
occur, thereby promoting a potential increase in biodiversity.
In the main channel of the free-flowing section downstream of Vienna, the macroinvertebrate commu-
nity was investigated in terms of biodiversity, spatial resource utilisation and persistence over a period
of one year. Oligochaetes and chironomids accounted for 95 % of the total numbers of macroinverte-
brates, the remaining five percent belonged to the Crustacea, Trichoptera, Bivalvia, Gastropoda, Bryo-
zoa, Ephemeroptera and Hirudinea. Three species of bryozoans were recorded based on colony findings,
and Fredericella sultana was most abundant. An analysis of the relationship between nine attributes of
the communities and six environmental factors indicates that habitat stability expressed as sediment
turnover was the major factor. Lower sediment turnover near the riverbanks coincided with increased
densities and decreased species turnover. Highest values for species richness and total abundance
occurred at a riverbank site of intermediate stability and greatest sediment heterogeneity, suggesting
that the communities were structured by a combination of several factors.
More studies are needed on the ecological relationship between native and recent invasive species, and
the manner in which the latter may or will affect the current structures of communities within the riv-
er fauna.
Key words: Large river, sampling devices, invaders, biodiversity, bryozoan settlement, habitat stability.
Einleitung diese Zahlen unterstreichen die ungeheure
internationale Bedeutung dieses Flusses.Die Donau ist mit ihrer Länge von ca.
2.850 km der zweitgrößte Fluss Europas. Sie Die österreichische Donau hat eine Län-
entspringt im Schwarzwald (Deutschland) ge von ungefähr 351 km und ein durch-
und mündet in das Schwarze Meer (Rumä- schnittliches Gefälle von 40 cm pro Kilo-
nien, Ukraine), in ihrem Einzugsgebiet lie- meter. Wie alle Gebirgsflüsse, so hat auch
gen noch 12 weitere Länder: die Schweiz, die Donau in der österreichischen Strom-
Österreich, Tschechische Republik, Slowa- strecke im Verlauf der Geschichte mannig-
kei, Ungarn, Slowenien, Kroatien, Bos- faltige Änderungen des Längs- und Quer-
nien/Herzegowina, Serbien/Montenegro, profils erfahren. Die systematische Regulie-
Albanien, Bulgarien und Moldawien (Abb. rungstätigkeit begann im vorletzten Jahr-
1). In dem Einzugsgebiet von ca. 805.200 hundert und hatte einen schrittweisen Denisia 16, zugleich Kataloge
km2 leben rund 90 Millionen Menschen, Übergang vom Naturstrom zum gebändig- Neue Serie 28(2005), 139-158
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
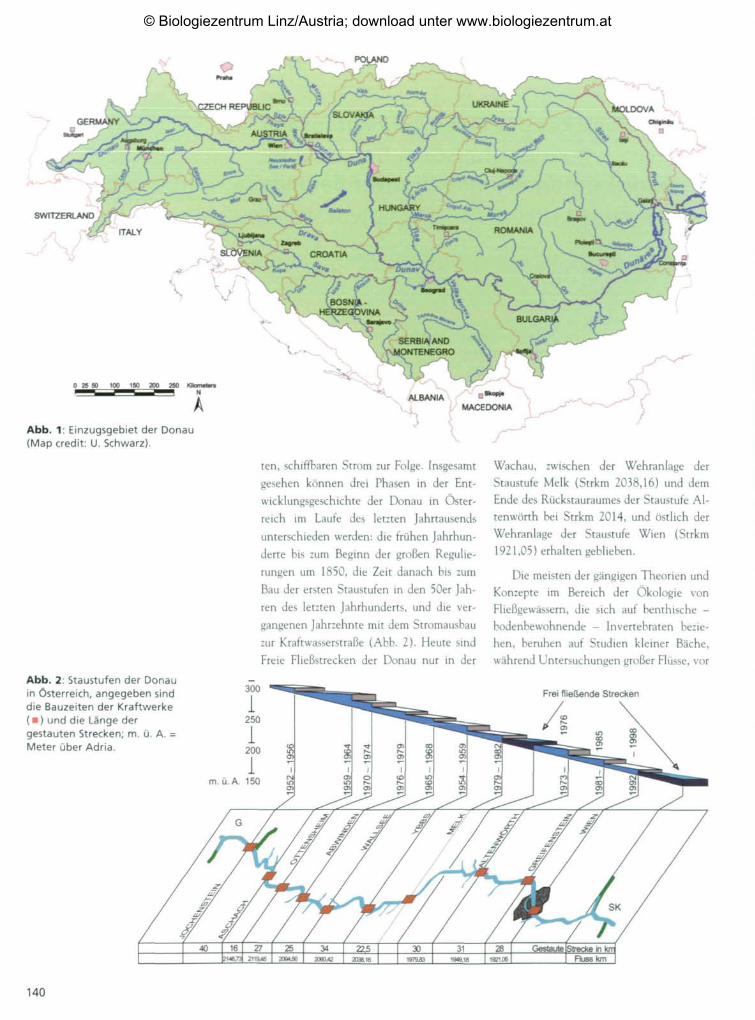
Abb. 1: Einzugsgebiet der Donau(Map credit: U. Schwarz).
ten, schiffbaren Strom zur Folge. Insgesamt
gesehen können drei Phasen in der Ent-
wicklungsgeschichte der Donau in Öster-
reich im Laufe des letzten Jahrtausends
unterschieden werden: die frühen Jahrhun-
derte bis zum Beginn der großen Regulie-
rungen um 1850, die Zeit danach bis zum
Bau der ersten Staustufen in den 50er Jah-
ren des letzten Jahrhunderts, und die ver-
gangenen Jahrzehnte mit dem Stromausbau
zur Kraftwasserstraße (Abb. 2). Heute sind
Freie Fließstrecken der Donau nur in der
Abb. 2: Staustufen der Donauin Österreich, angegeben sinddie Bauzeiten der Kraftwerke( • ) und die Länge dergestauten Strecken; m. ü. A. =Meter über Adria.
Wachau, zwischen der Wehranlage der
Staustufe Melk (Strkm 2038,16) und dem
Ende des Rückstauraumes der Staustufe Al-
tenwörth bei Strkm 2014, und östlich der
Wehranlage der Staustufe Wien (Strkm
1921,05) erhalten geblieben.
Die meisten der gängigen Theorien und
Konzepte im Bereich der Ökologie von
Fließgewässern, die sich auf benthische -
bodenbewohnende - Invertebraten bezie-
hen, beruhen auf Studien kleiner Bäche,
während Untersuchungen großer Flüsse, vor
Frei fließende Strecken
40 16 272119«
25Tsmso
342QHQ/C
22320» 16
30197943
311948.18
281921XB
Gestaute Streckein kmHusskm
140
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at

allem auf Grund methodischer Schwierig-keiten, lange Zeit eine untergeordnete Rol-le spielten. In dieser Hinsicht bietet die Do-nau in Osterreich eine hervorragende Mög-lichkeit wissenschaftliche Hypothesen aneinem großen Fließgewässer zu überprüfenoder neu zu formulieren.
Das Hauptinteresse der vorliegendenBearbeitung liegt in der Untersuchung derBiodiversität, d.h. der Anzahl der Arten undderen relative Anteile an der Lebensge-meinschaft, sowie ihrer steuernden Umwelt-faktoren. Die Biodiversität selbst ist in derÖkologie von fundamentaler Bedeutung, dasie der Frage nachgeht, warum manche Ar-ten häufig, manche selten sind, und wel-chen Einfluss deren Anteile an der Strukturund Funktion der gesamten Lebensgemein-schaft darstellen.
Allgemeine Sammelmethoden
Die meisten der in der vorliegenden Ar-beit verwendeten Geräte wurden bereits vonanderen Autoren beschrieben (siehe HUM-PESCH & ELLIOTT 1990). Für die biologische
Besammlung der Donau, für die ein entspre-chend ausgerüstetes Schiff vorhanden seinmuss (Abb. 3), wurden diese Geräte aller-dings den Gegebenheiten im Fluss - Tiefe,Bodenbeschaffenheit, Strömung - angepasstund daher verändert. Die Sammelgeräte sindin Abbildung 4 exemplarisch zusammenge-fasst. Die Abbildung zeigt auch in welchenFlussabschnitten - Freie Fließstrecke oderStau - die Geräte verwendet wurden.
Für qualitative Aufsammlungen, Auf-sammlungen also, deren Ergebnis auf eineArtenliste, auf die Schätzung relativer Ab-undanzen und die Berechnung einiger„Community" oder biotischer Indices, dieauf einer Rangordnung oder der Feststellungder Diversität basieren, beschränkt ist, ka-men die folgenden Geräte zum Einsatz(Abb. 4, 5):
• Fast Dredge (Abb. 4d, 5a, b),• Air-Liftsammelgerät (Abb. 5c, d),
• Bodensauger für die Bodenfauna (Abb.4c),
• Besiedlungssammelgerät für die Fauna desBodens und der Ufer (Abb. 4g) und
• ein automatisches, kontinuierlich mes-sendes Driftgerät (Abb. 4a) für im freienWasser treibende wirbellose Tiere.
Für quantitative Aufsammlungen, Aut-sammlungen also, deren Ergebnis die Schät-zung der Anzahl oder der Biomasse derPflanzen und Tiere pro Flächeneinheit ist,um die räumliche und zeitliche Verteilungder Lebewesen zu erfassen und darüber hin-aus über parametrische Tests auch geringfü-gige Veränderungen der Wasserqualität fest-zustellen und Wachstums-, Reproduktions-und Sterberaten berechnen zu können, eig-neten sich die folgenden Geräte:
für die Freiwasserzone
• Deck-Inkubator für das Phytoplankton,• Schindler-Planktonschöpfer (Abb. 4b)
für das Phyto- und Zooplankton.
für die Bodenzone - grobe Sedimente
• Stickstoff-Gefriercorer (Abb. 4f, 5i-m) fürdie Bodenfauna der seichten und tiefenStellen,
• Petersen-Greifer (Abb. 4e, 5e-h) für dieBodenfauna der tiefen Stellen,
• Besiedlungssammelgerät für die Boden-fauna der seichten und tiefen Stellen.
für die Bodenfauna - feine Sedimente
• „RINIMUCO"-Multicorer (Abb. 4h) fürdie Bodenfauna der seichten und tiefenStellen.
für die Ufer• Echomessgerät für Makrophyten.
• „Box"-Aufsammelgerät für die epiphy-tisch und
• Besiedlungssammelgerät (Abb. 4i) für dieepilithisch lebenden wirbellosen Tiere.
Die Grundlage für diese Empfehlungenist die Erfahrung, die während der langjähri-gen Aufsammlungstätigkeit in der Donaugemacht wurde.
Abb. 3: Untersuchungsschiff auf der Donau(v. r. Messschiff, Zugschiff,Untersuchungsschiff).
141
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at

schnellgrob
Abb. 4: Schematische Übersicht überProbennahmegeräte für die Flora undFauna der Donau, a: Driftsammeigerät; b:Schindler-Planktonschöpfer; c:Bodensauger; d: Fast Dredge; e: Petersen-Greifer; f: Stickstoff-Gefriercorer; g:Kolonisationssammelgerät; h: RINIMUCO-Multicorer; i: „Box"-Sammelgerät. Der Pfeilgibt die Strömungsrichtung an.
Strömu ng sgeschwi nd ig keitSubstratbeschaffenheit
Spezielle Sammelmethodenfür das Makrozoobenthos
Qualitative Aufsammlungen
• Fast - Bodendredge (Abb. 4d, 5a, b): Die
Dredge ist 41 kg schwer und wird von ei-
nem Schiff aus gegen die Strömung über
den Flussboden gezogen. Gitter verschie-
dener Maschenweiten verhindern, dass
größere Steine in das Netz gelangen, in
dem die Bodentiere gefangen werden, die
auf der Flusssohle leben.
• Air-Lift - Sammelgerät (Abb. 5c, d): Die
Funktionsweise basiert auf dem Prinzip,
dass ein vertikales Rohr im Wasser auf
den Flussboden aufgesetzt wird und Luft
in das Rohr gepumpt wird. Das Gemisch
aus Luft und Flusswasser, inklusive der
Bodenteile mit den darauf lebenden Bo-
denorganismen, steigt deshalb auf, weil
dieses spezifische Gewicht geringer ist als
das Wasser, das das Rohr umgibt.
Der Vorteil der beiden Geräte ist, dass inrelativ kurzer Zeit große Flächen des Fluss-
langsamfein
bodens mit seinen unterschiedlichen Habi-
taten besammelt werden können. Das Er-
gebnis ist dann eine relativ vollständige
Faunenliste.
Quantitative Aufsammlungen (Anzahl der
Tiere pro Flächeneinheit)
• Petersen-Greifer (in der modifizierten
Form von R. Niederreiter; Abb. 4e, 5e-h):
Der Greifer wiegt 45 kg, zusätzliche Ge-
wichte können angebracht werden, und
besammelt eine Fläche von 0.062 m2 bis
in eine Bodentiefe von ca. 7 cm. Der
Greifer besteht im Wesentlichen aus ei-
nem offen gehaltenen Schaufelpaar, de-
ren Fixierung sich beim Aufsetzen auf den
Flussboden über einen Mechanismus löst
und die sich beim Herausziehen des Grei-
fers schließen. So wird das von den
Schaufeln eingeschlossene Material,
Schottersedimente inkl. Organismen, aus
dem Wasser an Bord gebracht. Dabei ver-
hindert eine zweite Schaufel, dass, wenn
sich das Schaufelpaar infolge eines klem-
142
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at

4cm u- 123cm
a
Abb. 5: Einzeldarstellung der Geräte für (a-d) qualitative, (e-m) quantitative Beprobung des Makrozoobenthos der Freien Fließstreckeder Donau, a: Fast Dredge, schematische Übersicht; b: Fast Dredge vor einer Beprobung; c: Air-Lift, schematische Übersicht (aus: PEHOFER1998, verändert); d: Air-Lift vor einer Beprobung; e: Petersen-Greifer, schematische Übersicht; f: Petersen-Greifer offen, vor demHerablassen auf den Flussboden; g: Petersen-Greifer geschlossen, nach dem Herausholen aus dem Wasser; h: Petersen-Greifer geöffnetzum Erhalten der Probe.
143
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at

Abb. 5: Einzeldarstellung der Geräte für (a-d) qualitative, (e-m) quantitative Beprobung des Makrozoobenthos der Freien Fließstreckeder Donau, i: Stickstoff-Gefriercorer, schematische Übersicht (aus HUMPESCH & NIEDERREITER 1993): 1a Gefriercorer, 1b Corerrohr, 2Einschlaghammer, 3 Plattform, 4 Stickstoffbehälter, 5 Winde, 6 Untersuchungsschiff; Pfeil gibt die Strömungsrichtung an; j : Corerrohrwird in das Wasser gelassen (links der N2-Behälter); k: Einfüllen des flüssigen N2 aus N2-Behälter; I: Corer, nachdem er aus dem Wassergezogen wurde; m: Corer wird in 10 cm Teilstücke geschnitten.
144
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at

menden Steines, nicht vollständigschließt, der Inhalt der Schaufel beimHeraufziehen des Greifers ausrinnenkann.
• Stickstoff-Gefriercorer (Abb. 4f, 5i-m):Die Funktionsweise basiert auf dem Prin-zip, dass ein Rohr, das bis zu einer be-stimmten Tiefe, z.B. 1 m, in den Flussbo-den geschlagen wird, mit flüssigem Sick-stoff (-195,8 °C) so versorgt wird, dass dieFlussbodenbestandteile an das Rohr an-frieren. Danach wird das Rohr mit demangefrorenen Substrat und den darin be-findlichen Organismen aus dem Flussbo-den gezogen. Durch das Schneiden dessog. Cores, z.B. in 10 cm Schichten, kannso auch die Verteilung der Organismen indie Tiefe analysiert werden.
Der Vorteil der beiden Geräte ist, dassnicht nur die Bodenorganismen sondernauch das Substrat auf dem sie leben quanti-tativ erfasst werden und so Beziehungen zwi-schen der Lebensgemeinschaft und dem Le-bensraum analysiert werden können.
Artenzusammensetzungder Donau
In den Arbeiten von MOOG et al. (1995,2000) sind alle Arten der bodenbewohnen-den Makroinvertebraten, die aus der öster-reichischen Donau bekannt sind, zusammen-gefasst. Insgesamt wurden 1.289 Arten fest-gestellt, wenn von dieser Zahl die Collembo-len und bestimmte Zweiflügler abgezogenwerden, ergibt sich eine Zahl von 1.122 Ar-ten. Zu letzteren tragen die Dipteren (größ-tenteils Zuckmücken) mit 493 Arten oder44 % bei. Von übrigen 20 Hauptgruppen ge-hören zu den Coleopteren 138 und zu denTrichopteren 106 Arten, alle übrigen Grup-pen weisen eine geringere Artenzahl auf(Tab. 1). Eine Auswahl von Arten, die fürdie österreichische Donau typisch sind, ist inAbbildung 6 wiedergegeben.
Die Makroinvertebraten können imStrom vier Hauptlebensorten zugeordnetwerden (Tab. 1): der Region der FreienFließstrecke des Hauptgerinnes und seinerangeschlossenen Augewässer, der der ge-stauten Strecke, den Stauräumen und dendiesen angeschlossenen Augewässern. Inletzteren wurden 713 Arten aller Haupt-
Tab. 1: Gesamtzahl der benthischen Makroinvertebraten, die in der österreichischenDonau und ihren vier Haupthabitaten gefunden wurden. (Daten aus MOOG et al. 1995,2000).
HauptqruppenNicht-InsektenPoriferaHydrozoaTurbellariaMolluscaPolychaetaOligochaetaHirudineaAcarina
AraneaeCrustaceaBryozoaInsekten (excl.EphemeropteraPlecopteraOdonataHeteropteraMegalopteraPlanipenniaColeopteraTrichopteraLepidoptera
Diptera
Gesamtzahl
Gesamtzahl
22
25682
632423
1238
Diptera)40
66030
22
138106
4
493
1122
FreieHauptgerinne
005
321
261030
112
15012025
32
0163310
FließstreckeAuqewässer
105
460
311601
105
80
3026
22
53622
130430
GestauteStauraum
10
1047
2571330
112
18273107
510
229
464
StreckenAuqewässer
22
16612
1722190
158
334
582222
13192
2
203
713
gruppen gefunden. Die Stauräume enthiel-ten 464 Arten, die Augewässer der FreienFließstrecke 430 Arten jeweils von 17 Grup-pen und die Freie Fließstrecke selbst 310 Ar-ten von 15 Gruppen.
Von der Gesamtzahl der Arten, die in al-len 4 Haupthabitaten vorkommen, dominie-ren die Dipteren, in der Freien Fließstreckedie Trichopteren und Mollusken und in ih-ren Augewässern die Trichopteren, Coleop-teren und Mollusken (Tab. 1). Die Oligo-chaeten, Trichopteren und Mollusken warenin den Stauräumen dominant, in deren Au-gewässer die Coleopteren und Trichopteren.Die Arten der übrigen Gruppen machtenweniger als 10 % der Gesamtzahl der Artenin den einzelnen Hauptlebensräumen aus.
Gegenwärtig enthält die Datenbasis der„Roten Liste Österreichs" (www.roteliste.at)nur Informationen über die Mammalier, Vö-gel, Reptilien, Amphibien, Fische undSchmetterlinge, während der Wissensstandüber den Gefährdungsgrad der Arten desMakrozoobenthos der Donau sehr lücken-haft ist: gefährdet gelten nach BAUERNFEIND&. HUMPESCH (2001) zehn Ephemeropte-ren-, nach MALICKY (1994) 25 Trichopte-
145
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at

146
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at

Abb. 6: Typische Arten des Makrozoobenthos der Freien Fließstrecke der Donau: a: Strudelwurm - Dendrocoelum lacteum (MULLER)(Photo: P. Kölbl); b: Schnecke - Lithoglyphus natkoides, lebend (Photo: S. Linden); c: L. naticoides, Schalen (Photo: 2000 TerraFoundation, www.terra.hu); d: Muschel - Corbicula fluminea/fluminalis (Photo: A. Kureck); e: Krebs - Echinogammarus ischnus(Photo: Kahlert); f: Eintagsfliegen - Ephoron virgo (OLIVIER), Larve in Wohnröhre; g: E. virgo, Larvenkopf; h: E. virgo, er Imaginalhäutung;i: E. virgo, 9 Massenflug (z.B. beobachtet am 24. August 1994 Donaulände Linz) (Photo: A. Kureck); j : Potamanthus luteus PICTET, Larve;k: P. luteus, er Subimago; I: P. luteus, er Imago (Photo: P. Maihöfer); m: Köcherfliegenlarven - Ceraclea dissimilis (STEPHENS);n: Lepidostoma hirtum (FABRICIUS); O: Hydropsyche sp. (Photos: J. Waringer); p: Kriebelmücken - Simulium colombaschense, Larve;q: S. angustipes (EDWARDS), Kopf von oben, mit Haarfächer der ein- und ausklappbaren Fortsätzen der Oberlippe (Photos: W. Lechthaler);r: Simuliiden-Larven auf Substrat (Pfeil gibt die Strömungsrichtung des Wassers an); s: Simuliidae, blutsaugendes 9 (Photo: USDepartment of Agriculture); t : Schwammfliege - Larve von Sisyra dalii MCLACHLAN (Photo: W. Weißmair).
ren-, nach FRANK & REISCHÜTZ (1994) 45
Molluskenarten und nach GEPP (1994) eine
Sisyridenart.
Einwanderung neuer Arten vonMakroinvertebraten
Die Einwanderung von neuen Arten ist
eine weltweite Erscheinung und von großer
ökologischer Bedeutung. Ursprünglich führ-
ten die „Einwanderer" zu einer Zunahme der
Biodiversität in den betroffenen Gebieten,
doch bedeuten die Einwanderer auch eine
mögliche Gefahr für die ursprüngliche Le-
bensgemeinschaft, da sie im Einwande-
rungsgebiet das Verschwinden der dort le-
benden, seltenen, aber auch häufigen Arten
bedingen können. Weiters beeinflussen sie
die dortige ökologische Struktur und Funk-
tionsweise der Lebensgemeinschaft wesent-
lich. Die Donau fließt durch verschiedene
tiergeographische Regionen und ist außer-
dem seit 1992 über den Main mit dem
Rhein künstlich verbunden. Diese Verbin-
dungen ermöglichen die Einwanderung von
Faunenelementen aus diesen Gebieten in
die österreichische Donau und zwar sowohl
vom Westen flussabwärts als auch vom Os-
ten -aufwärts (Tab. 2).
Als Folge der Veränderung des Lebens-
raumes im Flusslauf, wie durch Staustufen,
z.B. hinsichtlich der Strömung und der Bo-
denstruktur (siehe z.B. HUMPESCH 1992)
wurden neue Lebensräume geschaffen, die
für das Überleben der Einwanderer sehr gu-
te Umweltbedingungen bedeuten.
Als Folge dieser Einwanderungsvorgän-
ge wird die ursprüngliche Zusammensetzung
der Lebensgemeinschaft wesentlich verän-
dert. Typische Beispiele dafür sind die von
Osten her aktiv eingewanderten Krebsarten
Corophium curvispinum, Echinogammarus
ischnus, Dikerogammarus haemobaphes, D.
viüosus und )aera isai bzw. die passiv vertra-
genen Arten wie die Muschel Dreissena poly-
morpha und der Süßwasservielborster Hypa-
nia invalida. Alle diese Arten sind ursprüng-
lich Element der pontokaspischen Fauna
mit ihrer ursprünglichen Verbreitung auf
147
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
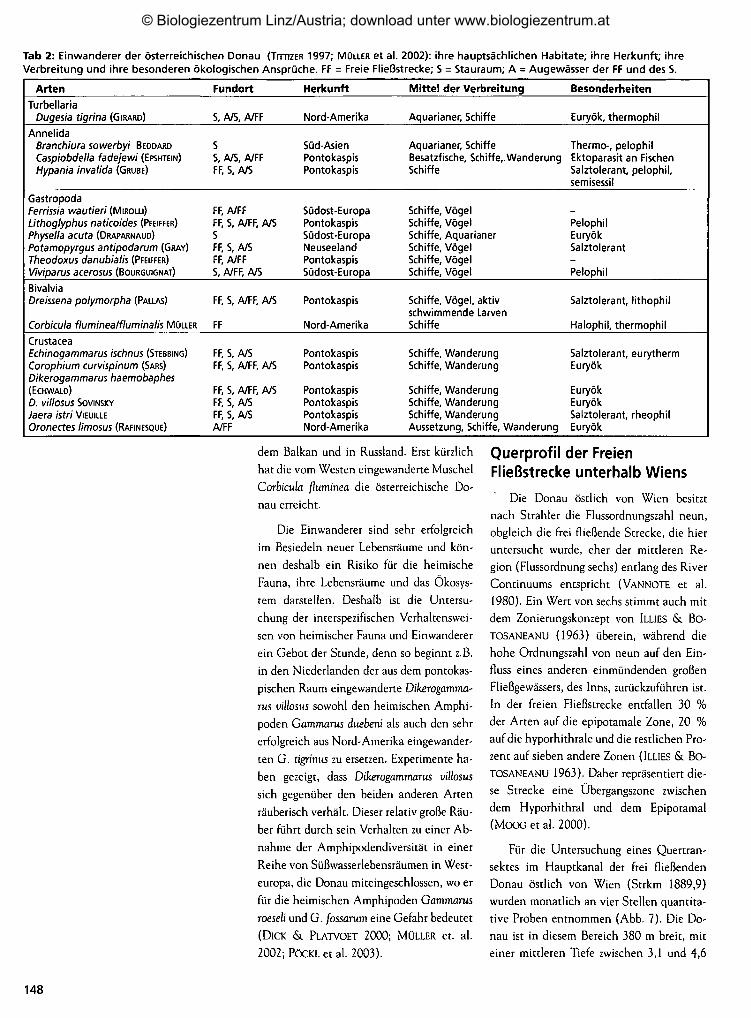
Tab 2: Einwanderer der österreichischen Donau (TrmzER 1997; MOLLER et al. 2002): ihre hauptsächlichen Habitate; ihre Herkunft; ihreVerbreitung und ihre besonderen ökologischen Ansprüche. FF = Freie Fließstrecke; S = Stauraum; A = Augewässer der FF und des S.
Arten
TurbellariaDugesia tigrina (GIRARD)
AnnelidaBranchiura sowerbyi BEODAROCaspiobdella fadejewi (EPSHTEIN)Hypania invalida (GRUBE)
GastropodaFerrissia wautieri (MIROLLI)Lithoglyphus naticoides (PFEIFFER)Physella acuta (DRAPARNAUD)Potamopyrgus antipodarum (GRAY)Theodoxus danubialis (PFEIFFER)Viviparus acerosus (BOURGUIGNAT)
BivalviaDreissena polymorpha (PALLAS)
Corbicula fluminealfluminalis MÜLLER
CrustaceaEchinogammarus ischnus (STEBBING)Corophium curvispinum (SARS)Dikerogammarus haemobaphes(ECHWALD)
D. villosus SOVINSKY
Jaera istri VIEUILLEOronectes limosus (RAFINESQUE)
Fundort
S, A/S, A/FF
SS, A/S, A/FFFF, S, A/S
FF, A/FFFF, S, A/FF, A/SSFF, S, A/SFF, A/FFS, A/FF, A/S
FF, S, A/FF, A/S
FF
FF, S, A/SFF, S, A/FF, A/S
FF, S, A/FF, A/SFF, S, A/SFF, S, A/SA/FF
Herkunft
Nord-Amerika
Süd-AsienPontokaspisPontokaspis
Südost-EuropaPontokaspisSüdost-EuropaNeuseelandPontokaspisSüdost-Europa
Pontokaspis
Nord-Amerika
PontokaspisPontokaspis
PontokaspisPontokaspisPontokaspisNord-Amerika
Mittel der Verbreitung
Aquarianer, Schiffe
Aquarianer, SchiffeBesatzfische, Schiffe,.WanderungSchiffe
Schiffe, VögelSchiffe, VögelSchiffe, AquarianerSchiffe, VögelSchiffe, VögelSchiffe, Vögel
Schiffe, Vögel, aktivschwimmende LarvenSchiffe
Schiffe, WanderungSchiffe, Wanderung
Schiffe, WanderungSchiffe, WanderungSchiffe, WanderungAussetzung, Schiffe, Wanderung
Besonderheiten
Euryök, thermophil
Thermo-, pelophilEktoparasit an FischenSalztolerant, pelophil.semisessil
PelophilEuryökSalztolerant-Pelophil
Salztolerant, lithophil
Halophil, thermophil
Salztolerant, eurythermEuryök
EuryökEuryökSalztolerant, rheophilEuryök
dem Balkan und in Russland. Erst kürzlich
hat die vom Westen eingewanderte Muschel
Corbicula fluminea die österreichische Do-
nau erreicht.
Die Einwanderer sind sehr erfolgreich
im Besiedeln neuer Lebensräume und kön-
nen deshalb ein Risiko für die heimische
Fauna, ihre Lebensräume und das Ökosys-
tem darstellen. Deshalb ist die Untersu-
chung der interspezifischen Verhaltenswei-
sen von heimischer Fauna und Einwanderer
ein Gebot der Stunde, denn so beginnt z.B.
in den Niederlanden der aus dem pontokas-
pischen Raum eingewanderte Dikerogamma-
rus viüosus sowohl den heimischen Amphi-
poden Gammarus duebeni als auch den sehr
erfolgreich aus Nord-Amerika eingewander-
ten G. tigrinus zu ersetzen. Experimente ha-
ben gezeigt, dass Dikerogammarus villosus
sich gegenüber den beiden anderen Arten
räuberisch verhält. Dieser relativ große Räu-
ber führt durch sein Verhalten zu einer Ab-
nahme der Amphipodendiversität in einer
Reihe von Süßwasserlebensräumen in West-
europa, die Donau miteingeschlossen, wo er
für die heimischen Amphipoden Gammarus
roeseli und G. fossarum eine Gefahr bedeutet
(DICK & PLATVOET 2000; MÜLLER et. al.
2002; PöCKL et al. 2003).
Querprofil der FreienFließstrecke unterhalb Wiens
Die Donau östlich von Wien besitzt
nach Strahler die Flussordnungszahl neun,
obgleich die frei fließende Strecke, die hier
untersucht wurde, eher der mittleren Re-
gion (Flussordnung sechs) entlang des River
Continuums entspricht (VANNOTE et al.
1980). Ein Wert von sechs stimmt auch mit
dem Zonierungskonzept von ILLIES & B o
TOSANEANU (1963) überein, während die
hohe Ordnungszahl von neun auf den Ein-
fluss eines anderen einmündenden großen
Fließgewässers, des Inns, zurückzuführen ist.
In der freien Fließstrecke entfallen 30 %
der Arten auf die epipotamale Zone, 20 %
auf die hyporhithrale und die restlichen Pro-
zent auf sieben andere Zonen (ILLIES & Bo-
TOSANEANU 1963). Daher repräsentiert die-
se Strecke eine Übergangszone zwischen
dem Hyporhithral und dem Epipotamal
(MOOG et al. 2000).
Für die Untersuchung eines Quertran-
sektes im Hauptkanal der frei fließenden
Donau östlich von Wien (Strkm 1889,9)
wurden monatlich an vier Stellen quantita-
tive Proben entnommen (Abb. 7). Die Do-
nau ist in diesem Bereich 380 m breit, mit
einer mittleren Tiefe zwischen 3,1 und 4,6
148
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at

m bei einer mittleren Schüttung von 2170m' pro Sekunde (FESL et al. 1999). Die
Stromsohle besteht aus Fels, Stein undSchotter mit gelegentlich Sand undSchlick. Sie wurde mittels modifiziertem Pe-terson-Greifer (HüMPESCH & ELLIOTT1990) bis in eine Tiefe von 0,26 bis 11,1 cmbesammelt.
Wichtige Umweltvariablen imQuertransekt
Eine Hauptkomponentenanalyse auf derBasis von 32 gemessenen oder errechnetenUmweltvariablen resultierte in sechs Haupt-komponenten (Tab. 3): mittlere Strömungs-geschwindigkeit, mittlere Korngröße,Heterogenität des Sediments (Zusammen-setzung des Sediments aus gleichen oderverschieden großen Körnern), Lückenraumim Sediment, der Umlagerung (Instabilität)des Sediments und die Wassertemperatur.Letztere variierte über das Untersuchungs-jahr von 2 bis 19,5 °C, mit einer mittlerenTemperatur von 10 °C.
An der Stelle 4 entlang des Quertran-sekts, nahe dem linken Ufer, ist die Strö-mungsgeschwindigkeit signifikant geringergegenüber den anderen Stellen (Tab. 3). ImZeitverlauf wurde die größte Strömung imMai und Juli während der Schneeschmelzein den Alpen beobachtet.
Auch die Substratzusammensetzung va-riiert entlang des Quertransekts. Das rechteUfer ist durch Blockwurf stabilisiert, und dienahe gelegene Stelle 1 weist signifikant grö-ßere Korngrößen verglichen mit den übri-gen Stellen auf. Die kleinsten mittlerenKorngrößen wurden nahe dem linken Uferhinter einer Buhne beobachtet; die Sedi-mentpartikel nahe der Strommitte an derStelle 3 liegen in ihrer Größe zwischen je-nen der Stellen 1 und 4. Die mittlerenKorngrößen variieren auch im Jahresverlauf.Die Heterogenität der Sedimentzusammen-setzung nimmt vom rechten zum linkenUfer hin leicht zu, während die zeitliche Va-riation nur gering ist.
Die Sedimentumlagerung unterscheidetsich zwischen den Stellen und ihre zeit-lichen Veränderungen entsprechend denSchüttungsverhältnissen mit geringerer Se-dimentbewegung im Winter und höherenUmlagerungen im späten Frühling und
50 100 150 200Entfernung vom rechten Ufer (m)
250 300
Abb. 7: Netzgrafik des Untersuchungs-areals bei Niedrigwasserstand (Durchfluss1182 m ! s ' ) bei Flusskilometer 1889,9 derDonau östlich von Wien. Die Kreise zeigendie möglichen der zufällig gewähltenPositionen von zehn Proben innerhalb dervier Stellen entlang des Quertransektes(Stellen 1, 2, 3 und 4: 30 m, 100 m, 200 mund 250 m vom rechten Ufer). Die Probenwurden monatlich von September 1995 bisAugust 1996 entnommen. Der gelbe Pfeilgibt die Strömungsrichtung an, der rotePfeil die Anbindung an die Au (nach FESL2002).
Sommer. Das Sediment nahe dem linkenUfer ist durch einen geringeren Lückenraumim Vergleich zu den übrigen Stellen ge-kennzeichnet, was auf die geringere Strö-mungsgeschwindigkeit und damit eine stär-kere Sedimentation kleiner Partikel zurück-zuführen ist. Im Zeitverlauf wurde der ge-ringste mittlere Lückenraum im Juli gefun-den.
Biodiversität der Makrozoo-benthos-Gemeinschaft inBeziehung zu Umweltvariablenin einem Quertransekt derDonau flussabwärts von Wien
In der vorliegenden Arbeit wurden ver-schiedene Attribute der Invertebraten-Ge-meinschaft für detaillierte Analysen ver-wendet. Die wichtigsten Ergebnisse werdenim Folgenden angeführt.
Taxonomische Zusammensetzung undIndividuendichten der Arten
Insgesamt wurden 164 Arten an vierStellen entlang des Quertransektes gefun-den (Tab. 4), von denen 81 Arten den Chir-onomiden (v.a. Orthociadiinae) und 33 denOligochaeten (v.a. Naididae) zuzuordnen
Tab. 3: Fünf hydrophysikalische Variablen an vier Probennahmestellen ineinem Quertransekt der Donau bei Strkm 1889,9 (FESL 2002). Die Probenwurden monatlich von September 1995 bis August 1996 entnommen. DiePosition der Stellen ist Abbildung 7 zu entnehmen.
VariableMittlere Strömung (cm pro sek)Median der Korngrößen (mm)SubstratheterogenitätSedimentumlagerungLückenraum (%)Wassertemperatur CC)
Stelle 1
182,059,51.48
1,530,8
Stelle 2 !
206,228,91,583,5
33,3Mitte
»eile 3197,440,71,672,6
31,510 °C
Stelle 4
139,627,91,75
2,125,6
149
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at

OliChirCrustTrichBivaGastBryoEphHiru
2 3 4Stelle
Abb. 8: Relative Individuendichten (log.Skala) von neun taxonomischen Gruppen,die bis zum Artniveau bestimmt wurden.Oli, Oligochaeta; Chir, Chironomidae; Crust,Crustacea; Trich, Trichoptera; Biva, Bivalvia;Gast, Gastropoda; Bryo, Bryozoa; Eph,Ephemeroptera; Hiru, Hirudinea.
waren. Die Stelle 4, nahe dem linken Ufer,weist 142 Arten mit 68 Chironomiden-Ar-ten (29 davon kommen nur an dieser Stellevor) und 30 Arten an Oligochaeta (davon 2ausschließlich an der Stelle 4) auf. Weiteresieben Großgruppen beinhalten zusammen50 Arten. An der Stelle 4 wurden die höch-sten Artenzahlen an Trichopteren (10 voninsgesamt 11 Arten), Ephemeropteren(9/11), Bivalven (8/8), Gastropoden (7/8),Crustaceen (6/7) und Hirudineen (2/2) ge-funden; 2 Bryozoen-Arten kommen an dreider vier Stellen vor (Tab. 4). Insgesamtweist die Stelle 4 die weitaus größte Anzahlan Arten (47) auf, die an keiner anderenStelle gefunden wurde.
Die Stelle 4 ist neben der höchsten Ar-tenzahl (142) durch die geringsten mittlerenStrömungsgeschwindigkeiten und Korngrö-ßen gekennzeichnet. Im Vergleich dazu, istdie Stelle 1, nahe dem rechten Ufer, durchdie größten Korngrößen und die geringsteSedimentumlagerung (Tab. 3) charakteri-siert. Darüber hinaus weist sie geringere Ar-tenzahlen (93) als die Stelle 4, aber, mit
Ausnahme der Ephemeropteren, leicht hö-here als die Stellen 2 und 3, in der Strom-mitte auf (Tab. 4). Sieben von 93 Artenkommen nur an der Stelle 1 vor, 11 von 82nur an der Stelle 2 und 4 von 71 Arten nuran der Stelle 3. Von diesen 22 Arten miteingeschränkter Verteilung sind 16 Chi-ronomiden und Oligochaeten und zweiEphemeropteren.
Bezüglich der Gesamtindividuendichtenpro rrr Stromsohle sind die Oligochaeta amzahlreichsten (Chaetogaster diastrophusheto-
sus (SVETLOV) und Nais elinguis MÜLLER amhäufigsten), gefolgt von den Chironomiden(Cladotanytarsus cf. vanderwulpi (EDWARDS)und C. cf. daviesi BlLYlj & DAV1ES am häu-figsten). Auf diese beiden Gruppen entfal-len mehr als 95 % der Gesamtindividuen-zahlen an allen Stellen (Abb. 8). Die mitt-leren Gesamtabundanzen aller Arten zusam-men sind an der Stelle 4 am höchsten(11.146 Individuen pro nr), doppelt so vielals die mittleren Abundanzen an der Stelle1 und das Zehnfache jener an der Stelle 3;an der Stelle 2 beträgt die durchschnittlicheIndividuenzahl nur 623 Individuen pro m2
(Tab. 4).
Unter den restlichen identifizierten 50Makroinvertebraten-Arten, sind die Crusta-ceen vergleichsweise abundant, v.a. ]aera is-
tri (35 %) und Corophium curvispinum (32%), gefolgt von einer Bryozoen-Art, Frederi-
cella sultana (4 %), und Trichopteren, imwesentlichen Hydropsyche contubemalis andPsychomya pusilla (3 %). Mit mittleren rela-tiven Abundanzen von weniger als 1 % tre-ten die Bivalven, die Gastropoden, dieEphemeropteren und Hirudineen auf, was
Tab. 4: Gesamtartenzahlen (S), Artenzahlen an den einzelnen Stellen und mittlere Individuendichtenentlang des Quertransektes flussabwärts von Wien; fettgedruckte Zahlen in Klammern entsprechen denArtenzahlen, die ausschließlich an der jeweiligen Stelle gefunden wurden. Die Daten für Chironomidenwurden FESL (2002), für Oligochaeten FESL & HUMPESCH (2003) und für alle anderen Gruppen FESL &HUMPESCH (2005) entnommen. Die Lage der Untersuchungsstellen sind der Abbildung 7 zu entnehmen.
Großgruppe
ChironomidaeOligochaetaEphemeropteraTrichopteraBivalviaGastropodaCrustaceaBryozoaHirudinea
Gesamtzahl der ArtenMittlere Ind.-Dichten (per m2
S
8133111188732
164) —
Stelle 1
40282754421
935.314
(3)(1)(1)(1)
(1)
(7)
Stelle 2
37245421621
82623
(8)(1)(1)
(1)
(11)
Stelle 3
27 (2)25 (1)343241 (1)1
70 (4)1.195
Stelle 4
68309
1087622
14211.146
(29)(2)(5)(3)(2)(4)(1)
(1)(47)
150
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at

(a)
collectors
microbes - * < h r e d d e r s
predators
auf eine große Anzahl seltener Arten hin-
deutet. Die Gesamtindividuendichten aller
Arten zeigen signifikante Unterschiede zwi-
schen den Stellen und eine große zeitliche
Variation, mit Spitzen in November und
März und den geringsten Werten im Juli
nach durch die Schneeschmelze in den Al-
pen verursachten Hochwässern.
Ein Vergleich der Anteile an den funk-
tionellen Ernährungstypen - shredders (Zer-
kleinerer), collectors (Sammler), grazers
(Weidegänger) und predators (Räuber) -
weist auf eine Dominanz der collectors hin
(Abb. 9).
Aspekte der Bryozoenbesiedlung
Bereits LlEPOLT (1967) gibt in seinerMonographie zur Limnologie der Donau einVorkommen von insgesamt acht Bryozoen-Arten an, deren Verbreitungsgebiet imStrom als unregelmäßig klassifiziert wird.Allerdings führt er auch mit Victorella pavida
KENT eine Brackwasserform, die sich auf dasDonaudelta beschränkt. Für den österreichi-schen Abschnitt des Donaustroms gilt beidiesem Autor einzig Plumatella repens (LlN-NAEUS) als belegt. In MOOG et al. (1995)werden aus früheren Untersuchungen für dieFließstrecke unterhalb Wiens die beiden Ar-ten Fredericeüa sultana (BLUMENBACH) undCristatella mucedo CuviER genannt (Abb.10a, e). PfiCSI & ERDELICS (1970) beschrei-ben zudem im angrenzenden ungarischenDonauabschnitt das Auftreten von Paludicel-
la articulata (EHRENBERG). In der vorliegen-den Arbeit beinhalten die 480 Proben, dieinnerhalb eines Jahres zu zwölf Terminen an
den vier Stellen des Quertransekts bei Strkm1889 genommen wurden, Kolonien von Fre-dericeüa sultana, PaludiceUa articulata und Plu-
mateüa emarginata ALLMAN (Abb. 10a-d, h).Fredericeüa sultana tritt im Durchschnittrund 20 x so häufig auf als Pcdudiceüa articu-
lata, während der Anteil von Plumateüa
emargiruita verschwindend gering bleibt(Tab. 5), so finden sich nur in einer einzigenProbe des Monats August Kolonien der letzt-genannten Art. In den folgenden beidenAbbildungen 11 und 12 werden ausschließ-lich die Koloniefunde von Fredericeüa sultana
und PaludiceUa articulata berücksichtigt.
Die Darstellung des jahreszeitlichenVerlaufs (Abb. 11) zeigt, dass für diese bei-den Arten in den Wintermonaten und imFrühjahr die Abundanzen geringer sind, be-ziehungsweise, dass Funde vollkommen feh-len. Dies entspräche dem natürlichen Rück-gang der Kolonien bis hin zum völligen Aus-fall zu Beginn der kälteren Jahreszeit, ob-wohl gerade im Falle von Fredericeüa sultana
und PaludiceUa articulata eine Überwinte-rungsfähigkeit der Kolonien angenommenwerden kann (WlEBACH 1960). Die geringeHäufigkeit von P. articulata, die nur bei dreiTerminen (Mai, August und September)vertreten ist, lässt allerdings nur beschränktRückschlüsse zu. Fredericeüa sultana fehltnur im März und April; besonders auffälligist die hohe Dichte an Kolonien im Juni, sosind 52 % des Jahresanteils diesem Monatzuzurechnen, gefolgt vom Juli (15 %) undAugust (13 %).
Vergleicht man die vier Probenstellen
des Quertransekts (Abb. 12), so ist die mitt-
active,passivefilterercollectors
shredders
predators
Abb. 9: a: Erwartete relative Anteile derfunktionellen Ernährungstypen gemäß demRiver Continuum Concept (adaptiert von:Oxbow River & Stream Restoration 2003).b: Mittlere Zusammensetzung derfunktionellen Ernährungstypen in derFreien Fließstrecke der Donau flussabwärtsvon Wien. Collectors = Sammler, grazers =Weidegänger, filterer collectors = Filtrierer,shredders = Zerkleinerer, predators =Räuber, microbes = Mikroorganismen.
151
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at

152
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at

Tab. 5: Abundanz der Bryozoenkolonien im Quertransekt in der freien Fließstrecke der Donau beiFlusskilometer 1889,9.
Mittlere Koloniezahl per m2
Maximale Koloniezahl per m2
Fredericella sultana
0,53
588,74
Paludicella articulata
0,03
52,84
Plumatella emarginata
0,005
9,81
lere Koloniendichte aller Bryozoenarten an
der linksufrigen Stelle 4 im Durchschnitt et-
wa viermal so hoch als im gesamten Bereich
der übrigen drei Stellen. Allein F. sultana ist
an allen vier Stellen vertreten, ihre mittlere
Gesamtabundanz verteilt sich zu 80 % auf
Stelle 4, gefolgt von 16 % an der rechtsufri-
gen Stelle 1 und vergleichsweise verschwin-
dend kleinen Werten an den Stellen 2 und
3 nahe der Strommitte. Bei Paludicella am-
culata ist die Dichte an Kolonien an Stelle 4
fast doppelt so hoch wie an den gleich stark
besiedelten Stellen 1 und 2.
Moostiere bilden zur Überbrückung un-günstiger Lebensbedingungen Dauerstadienaus, die sich in zahlreichen Benthosprobenin Form von Statoblasten wiederfinden.Diese Art der Dauerknospen werden aus-nahmslos in der Klasse der Phylactolaematagebildet, der alle hier zu besprechendenMoostierarten mit Ausnahme von P. articu-
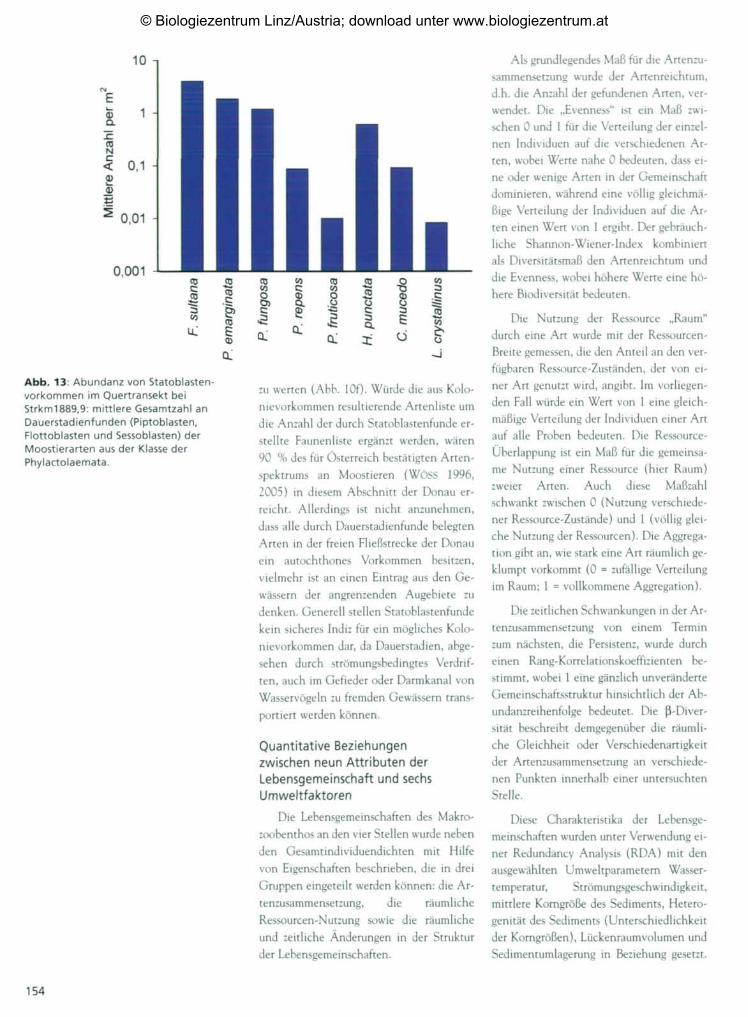
lata angehören. Statoblasten werden jenach Ausbildungsgrad eines Schwimmringsin Flottoblasten, Sessoblasten und Pipto-blasten unterteilt (RYLAND 1970). Diehöchsten Abundanzen an Dauerstadiensind auf die Piptoblastenfunde von Frederi-
cella sultana zurückzuführen (Abb. 13), dieim Mittel fast achtmal so häufig wie Kolo-niefunde anzutreffen sind und pro m2 einenMaximalwert von 14-081 erreichen. Gene-rell übertrifft die mittlere Dauerstadien-dichte von F. sultana (Abb. 10b) jene vonPlumatella emarginata (Abb. 10h) um das
Abb. 10: Bryozoa im Quertransekt beiStrkm 1889,9. a: Fredericella sultana -Kolonie auf Myriophyllum-StengeV, b: F.sultana - Piptoblasten in degenerierterKolonieröhre; c: F. sultana (Bildmitte) undPaludicella articulata (außen; Kolonienzarter und aus konisch verlaufendenRöhren zusammengesetzt); d: Zooide vonFredericella sultana (linke Bildhälfte) undPaludicella articulata (rechte Bildhälfte,Austrittsstelle des Polypids subterminal); e:Cristatella mucedo; f: Lophopuscrystallinus; g: Plumatella fungosa; h: P.emarginata - Flottoblasten einerBenthosprobe.
5,5 n
Fredericella sultanaPaludicella articulata
zweifache bzw. jene von P. fungosa (PALLAS)(Abb. 10g) sogar um das dreifache. Die bei-den letztgenannten Arten bilden sowohlFlottoblasten als auch Sessoblasten aus, wo-bei sich der Flottoblastenanteil in den Pro-ben zwischen 65 % und 75 % bewegt.Nennenswerte Vorkommen an Dauersta-dien sind außerdem von Hyalinella punctata
(HANCOCK) ZU berichten, gefolgt von Cri-
statella mucedo und Plumatella repens. Alsausgesprochene Rarität sind hingegen dieFunde von P. fruticosa ALLMAN und insbe-sonders von Lophopus crystallinus (PALLAS)
2 , 5 -
2,0 •
Abb. 11: Abundanz von Bryozoenkolonienim Jahresverlauf im Quertransekt beiStrkm1889,9: mittlere Anzahl der Kolonienper m2 der Arten Fredericella sultana undPaludicella articulata.
Abb. 12: Abundanz von Bryozoenkolonienan den vier Probenstellen im Quertransektbei Strkm1889,9: mittlere Anzahl derKolonien per m1 der Arten Fredericellasultana und Paludicella articulata.
i.o H
0,5
0,0
Fredericella sultanaPaludicella articulata
1Stelle
153
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
10 -I
n
< 0,1££5
0,01 -I
0,001
& i' e ••§ i I I(0u; g
3 3 pir r> C
Abb. 13: Abundanz von Statoblasten-vorkommen im Quertransekt beiStrkm1889,9: mittlere Gesamtzahl anDauerstadienfunden (Piptoblasten,Flottoblasten und Sessoblasten) derMoostierarten aus der Klasse derPhylactolaemata.
s
zu werten (Abb. 10f)- Würde die aus Kolo-
nievorkommen resultierende Artenliste um
die Anzahl der durch Statoblastenfunde er-
stellte Faunenliste ergänzt werden, wären
90 % des für Osterreich bestätigten Arten-
spektrums an Moostieren (WöSS 1996,
2005) in diesem Abschnitt der Donau er-
reicht. Allerdings ist nicht anzunehmen,
dass alle durch Dauerstadienfunde belegten
Arten in der freien Fließstrecke der Donau
ein autochthones Vorkommen besitzen,
vielmehr ist an einen Eintrag aus den Ge-
wässern der angrenzenden Augebiete zu
denken. Generell stellen Statoblastenfunde
kein sicheres Indiz für ein mögliches Kolo-
nievorkommen dar, da Dauerstadien, abge-
sehen durch strömungsbedingtes Verdrif-
ten, auch im Gefieder oder Darmkanal von
Wasservögeln zu fremden Gewässern trans-
portiert werden können.
Quantitative Beziehungenzwischen neun Attributen derLebensgemeinschaft und sechsUmweltfaktoren
Die Lebensgemeinschaften des Makro-
zoobenthos an den vier Stellen wurde neben
den Gesamtindividuendichten mit Hilfe
von Eigenschaften beschrieben, die in drei
Gruppen eingeteilt werden können: die Ar-
tenzusammensetzung, die räumliche
Ressourcen-Nutzung sowie die räumliche
und zeitliche Änderungen in der Struktur
der Lebensgemeinschaften.
Als grundlegendes Maß für die Artenzu-sammensetrung wurde der Artenreichtum,d.h. die Anzahl der gefundenen Arten, ver-wendet. Die „Evenness" ist ein Maß zwi-schen 0 und 1 für die Verteilung der einzel-nen Individuen auf die verschiedenen Ar-ten, wobei Werte nahe 0 bedeuten, dass ei-ne oder wenige Arten in der Gemeinschaftdominieren, während eine völlig gleichmä-ßige Verteilung der Individuen auf die Ar-ten einen Wert von 1 ergibt. Der gebräuch-liche Shannon-Wiener-Index kombiniertals Diversitätsmaß den Artenreichtum unddie Evenness, wobei höhere Werte eine hö-here Biodiversität bedeuten.
Die Nutzung der Ressource „Raum"durch eine Art wurde mit der Ressourcen-Breite gemessen, die den Anteil an den ver-fügbaren Ressource-Zuständen, der von ei-ner Art genutzt wird, angibt. Im vorliegen-den Fall würde ein Wert von 1 eine gleich-mäßige Verteilung der Individuen einer Artauf alle Proben bedeuten. Die Ressource-Überlappung ist ein Maß für die gemeinsa-me Nutzung einer Ressource (hier Raum)zweier Arten. Auch diese Maßzahlschwankt zwischen 0 (Nutzung verschiede-ner Ressource-Zustände) und 1 (völlig glei-che Nutzung der Ressourcen). Die Aggrega-tion gibt an, wie stark eine Art räumlich ge-klumpt vorkommt (0 = zufällige Verteilungim Raum; 1 = vollkommene Aggregation).
Die zeitlichen Schwankungen in der Ar-tenzusammensetzung von einem Terminzum nächsten, die Persistenz, wurde durcheinen Rang-Korrelationskoeffizienten be-stimmt, wobei 1 eine gänzlich unveränderteGemeinschaftsstruktur hinsichtlich der Ab-undanzreihenfolge bedeutet. Die ß-Diver-sität beschreibt demgegenüber die räumli-che Gleichheit oder Verschiedenartigkeitder Artenzusammensetzung an verschiede-nen Punkten innerhalb einer untersuchtenStelle.
Diese Charakteristika der Lebensge-meinschaften wurden unter Verwendung ei-ner Redundancy Analysis (RDA) mit denausgewählten Umweltparametern Wasser-temperatur, Strömungsgeschwind igkeit,mittlere Korngröße des Sediments, Hetero-genität des Sediments (Unterschiedlichkeitder Korngrößen), Lückenraumvolumen undSedimentumlagerung in Beziehung gesetzt.
154
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
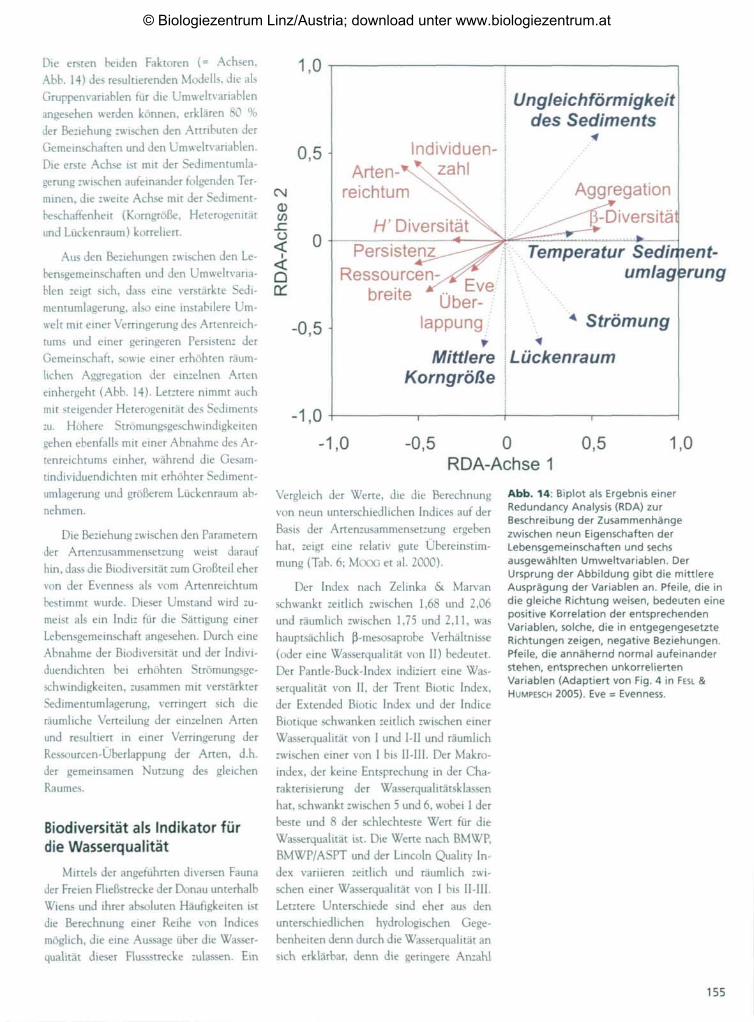
Die ersten beiden Faktoren (= Achsen,
Abb. 14) des resultierenden Modells, die als
Gruppenvariablen für die Umweltvariablen
angesehen werden können, erklären 80 %
der Beziehung zwischen den Attributen der
Gemeinschaften und den Umweltvariablen.
Die erste Achse ist mit der Sedimentumla-
gerung zwischen aufeinander folgenden Ter-
minen, die zweite Achse mit der Sediment-
beschaffenheit (Korngröße, Heterogenität
und Lückenraum) korreliert.
Aus den Beziehungen zwischen den Le-
bensgemeinschaften und den Umweltvaria-
blen zeigt sich, dass eine verstärkte Sedi-
mentumlagerung, also eine instabilere Um-
welt mit einer Verringerung des Artenreich-
tums und einer geringeren Persistenz der
Gemeinschaft, sowie einer erhöhten räum-
lichen Aggregation der einzelnen Arten
einhergeht (Abb. 14)- Letztere nimmt auch
mit steigender Heterogenität des Sediments
zu. Höhere Strömungsgeschwindigkeiten
gehen ebenfalls mit einer Abnahme des Ar-
tenreichtums einher, während die Gesam-
tindividuendichten mit erhöhter Sediment-
umlagerung und größerem Lückenraum ab-
nehmen.
Die Beziehung zwischen den Parameternder Artenzusammensetzung weist daraufhin, dass die Biodiversität zum Großteil ehervon der Evenness als vom Artenreichtumbestimmt wurde. Dieser Umstand wird zu-meist als ein Indiz für die Sättigung einerLebensgemeinschaft angesehen. Durch eineAbnahme der Biodiversität und der Indivi-duendichten bei erhöhten Strömungsge-schwindigkeiten, zusammen mit verstärkterSedimentumlagerung, verringert sich dieräumliche Verteilung der einzelnen Artenund resultiert in einer Verringerung derRessourcen-Überlappung der Arten, d.h.der gemeinsamen Nutzung des gleichenRaumes.
Biodiversität als Indikator fürdie Wasserqualität
Mittels der angeführten diversen Faunader Freien Fließstrecke der Donau unterhalbWiens und ihrer absoluten Häufigkeiten istdie Berechnung einer Reihe von Indicesmöglich, die eine Aussage über die Wasser-qualität dieser Flussstrecke zulassen. Ein
1,0
0,5
CM
CD</)
. C
o<
0
-0,5
-1,0
Individuen-Arten-^V zahl
reichtum
H' DiversitätPersistenz
Ressourcen-breite
lappung
MittlereKorngröße
Ungleichförmigkeitdes Sediments
Aggregation
-Diversitäfl
Temperatur Sedir lentumlag irung
4 Strömung
Lückenraum
-1,0 -0,5 0 0,5RDA-Achse 1
1,0
Vergleich der Werte, die die Berechnungvon neun unterschiedlichen Indices auf derBasis der Artenzusammensetzung ergebenhat, zeigt eine relativ gute Übereinstim-mung (Tab. 6; MOOG et al. 2000).
Der Index nach Zelinka & Marvanschwankt zeitlich zwischen 1,68 und 2,06und räumlich zwischen 1,75 und 2,11, washauptsächlich ß-mesosaprobe Verhältnisse(oder eine Wasserqualität von II) bedeutet.Der Pantle-Buck-lndex indiziert eine Was-serqualität von II, der Trent Biotic Index,der Extended Biotic Index und der IndiceBiotique schwanken zeitlich zwischen einerWasserqualität von I und I-II und räumlichzwischen einer von I bis II-III. Der Makro-index, der keine Entsprechung in der Cha-rakterisierung der Wasserqualitätsklassenhat, schwankt zwischen 5 und 6, wobei 1 derbeste und 8 der schlechteste Wert für dieWasserqualität ist. Die Werte nach BMWP,BMWP/ASPT und der Lincoln Quality In-dex variieren zeitlich und räumlich zwi-schen einer Wasserqualität von I bis II-III.Letztere Unterschiede sind eher aus denunterschiedlichen hydrologischen Gege-benheiten denn durch die Wasserqualität ansich erklärbar, denn die geringere Anzahl
Abb. 14: Biplot als Ergebnis einerRedundancy Analysis (RDA) zurBeschreibung der Zusammenhängezwischen neun Eigenschaften derLebensgemeinschaften und sechsausgewählten Umweltvariablen. DerUrsprung der Abbildung gibt die mittlereAusprägung der Variablen an. Pfeile, die indie gleiche Richtung weisen, bedeuten einepositive Korrelation der entsprechendenVariablen, solche, die in entgegengesetzteRichtungen zeigen, negative Beziehungen.Pfeile, die annähernd normal aufeinanderstehen, entsprechen unkorreliertenVariablen (Adaptiert von Fig. 4 in FESL &HUMPESCH 2005). Eve = Evenness.
155
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at

Tab. 6: Gewässergüte der Freien Fließstrecke der Donau unterhalb Wiens, angegeben sinddie Werte der unterschiedlichen Indices; korrespondierende Güteklassen in Klammer;ZaM = Zelinka & Marvan-Saprobienindex; PaB = Pantle-Buck- Index; TBI = Trent BioticIndex; EBI = Extended Biotic Index; IB = Indice Biotique; MI = Makroindex; BMWP =Biological Monitoring Working Party; BMWP/ASPT = Biological Monitoring WorkingParty/ Average Score per Taxon; LQI = Lincoln Quality Index (MOOG et al. 2000).
(a) Zeitliche Unterschiede
Index/Datum
ZaMPaBTBIEBIIBMIBMWPBMWP/ASPTLQI
(b) Räumliche Unterschiede
Index/Entnahmestelle in m vomrechten UferZaMPaBTBIEBIIBMIBMWPBMWP/ASPTLQI
23 Jan
1,68 (l-ll)2,09 (II)9 (l-ll)11(09 (l-ll)570 (Il-Ill)5,38 (Il-Ill)5,5 (l-ll)
302,11(11)2,12(11)9 (l-ll)11-10(1)9 (l-ll)576 (Il-Ill)4,75 (Il-Ill)5(11)
6 Apr
2,06 (II)2,10(11)9 (l-ll)11(1)9 (l-ll)677 (Il-Ill)5,13(11-111)5,5 (l-ll)
1001.91 (II)1,99 (II)8 (II)8 (II)7 (II)638 (III)4,75 (Il-Ill)4,5 (II)
5Jul
1.95 (II)2,11(11)9 (I-M)11(1)8(11)594 (Il-Ill)5,88 (II)6(11)
2001,75 (l-ll)2,07 (II)7 (II)10(1)6 (Il-Ill)531 (III)3,44 (III)3 (Ml)
1 Okt
2,00 (II)2,14(11)9 (l-ll)11(1)9 (l-ll)580 (Il-Ill)5,71 (II)5,5 (l-ll)
2501.85 (II)2,11(11)9 (l-ll)12(1)10(1)5111 (II)6,17(11)6,5 (1)
Gesamt
1.96 (II)2.11(11)9 (l-ll)12(1)10(1)5114(11)6,00 (II)6,5 (1)
Gesamt1,96 (II)2,11(11)9 (l-ll)12(1)10(1)5114(11)6,00 (II)6,5 (1)
der Arten und Individuen an einigen Ent-
nahmestellen, die Grund für eine schlechte-
re Indizierung sind, ergeben sich eher aus
der dort höheren Strömungsgeschwindig-
keit und daher dort größeren Umlagerung
der Stromsohle (siehe dazu das vorherige
Kapitel). Eine zweite Schlussfolgerung die-
ser Ergebnisse ist, dass die Indizes, die auf
der Zusammensetzung von Arten basieren,
ein wirklichkeitsgetreueres Bild der Wasser-
qualität wiedergeben, als die, bei denen für
die Analysen ausschließlich höhere taxono-
mische Niveaus zum Tragen kommen.
Die Fauna deroberflächlichen 0-10 cm unddes tieferen Hyporheons
Die Aussagen des Makrozoobenthos im
angeführten Querprofil bezogen sich auf die
Fauna der ersten 10 Zentimeter der Strom-
sohle. In den meisten Bächen und Flüssen
wird aber auch eine Besiedlung tieferliegen-
der Sedimente, besammelt mittels entspre-
chender Methoden (siehe dazu „Gefrierco-
rer"), festgestellt (z.B. HUMPESCH et al.
2002). Diese Zone wird als Hyporheon be-
zeichnet. Letzteres wird u.a. vom Makro-
zoobenthos dann besiedelt, wenn die Raum-
verhältnisse und andere Umweltbedingun-gen, wie z.B. sauerstoffreiches Wasser, es er-lauben. Frühe Entwicklungsstadien vonsonst an der Oberfläche der Stromsohle le-benden Arten bevorzugen diesen Lebens-raum. Letzteres spiegelt sich in den großensaisonalen Schwankungen in Bezug auf An-zahl, Verbreitung und Vorkommen der Tierewider. Unabhängig davon gibt es Arten, dieausschließlich in diesem Lebensraum vor-kommen, die sog. hyporheische Fauna, unddie auf Grund verschiedener Anpassungdort ihren gesamten Lebenszyklus durchlau-fen können. Zu dieser Fauna gehören u.a.Vertreter der Crustaceen, Oligochaten undHydracarinen.
Einer der größten Vorteile im Hyporheonzu leben ist, dass dort lebensfeindliche Um-weltbedingungen, wie Hochwässer oder Aus-trocknung, überdauert werden können. Nachz.T. katastrophalen Hochwässern in der Do-nau, die die Oberflächenfauna z.T. total eli-minieren, kann von der Fauna, die in tieferenSedimenten diese Zeit verbracht hat, eineWiederbesiedlung der Oberflächensedimenteausgehen. Demnach kann das Hyporheon alsÖkoton zwischen Oberflächen- und Grund-wasser angesehen werden.
Ist Biodiversität vonBedeutung?
Obwohl die Beziehungen zwischen der
Biodiversität und der Funktion bzw. Stabi-
lität von Ökosystemen nach wie vor unklar
sind, sprechen drei Argumente für die Be-
deutung der Biodiversität:
• zunächst das „moralische" Argument, das
besagt „wir sind nur zeitliche Gäste des
Erdplaneten und deshalb haben wir die
Pflicht diesen Lebensraum den nachfol-
genden Generationen in annähernd dem
Zustand zu übergeben, in dem wir ihn vor-
gefunden haben";
• dann das Argument der „Balance in der
Natur": es gibt un- und vorhersehbare
Folgen hervorgerufen durch den Verlust
der Biodiversität, einige dieser Folgen
können sich sehr nachteilig auf den Men-
schen auswirken - die Auslöschung von
Schlüsselarten kann zu einer kompletten
Änderung der Struktur einer Lebensge-
meinschaft führen;
156
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at

• letztlich das „Ressourcenargument":
Pflanzen und Tiere stellen ein großes ge-
netischen Reservoir dar, das angepasst für
die Landwirtschaft, die Industrie und die
Medizin nutzbar gemacht werden kann.
Die Rolle vieler Arten in einer Lebens-
gemeinschaft, besonders die der seltenen
Arten, ist zum großen Teil nicht bekannt
und hat zu der Theorie geführt, dass es
„überzählige" Arten gibt, die ähnliche Auf-
gaben im Ökosystem haben, wie andere Ar-
ten. Diese ähnlichen Arten erfüllen mögli-
cherweise doch wichtige Aufgaben, je nach-
dem in welchem Zustand sich das Ökosys-
tem befindet. Anstatt sich auf einzelne Ar-
ten und deren Erhalt zu konzentrieren,
scheint der erfolgreichere Ansatz zu sein, die
Lebensräume, in denen diese Arten vor-
kommen, zu erhalten. Letzteres auch des-
halb, weil der Nutzen vieler Arten für den
Menschen oft nicht ersichtlich ist. Die Um-
kehrung dieses Standpunktes könnte sein:
„Welcher Lebensraum ist es nicht Wert er-
halten zu werden"?
Schlussbemerkungen
Der österreichische Teil der Donau, mitseinem alpinen Charakter, enthält nur mehrzwei Freie Fließstrecken - das Gebiet derWachau und der Teil unterhalb des Kraft-werkes Freudenau bis Stromkilometer 1874-Letztere Strecke wird von 310 Arten desMakrozoobenthos besiedelt, von denen nur58 Arten ausschließlich dort gefunden wur-den. Für den Erhalt der selteneren Arten istder ihres Lebensraumes von fundamentalerBedeutung; es ist deshalb notwendig, die un-gestaute Strecke unterhalb Wien als solchezu erhalten. Letzteres auch deshalb, weil derStrom dort direkt mit seinen Augewässernverbunden ist, so dass ein Faunenaustauschzwischen Haupt- und Augewässer stattfin-den und dort zu einer wesentlichen Zunah-me der Biodiversität führen kann.
Die ökologischen Beziehungen zwischen
eingewanderten und ursprünglichen Arten
sind im Wesentlichen nicht bekannt, daher
kann keine Aussage darüber gemacht wer-
den in welcher Form die eingewanderten
Arten die gegenwärtigen Lebensgemein-
schaften des Makrozoobenthos beeinflussen
werden.
Danksagung: Für die finanzielle Unter-
stützung, z.B. die Wissenschafts- und For-
schungsförderung der Gemeinde Wien, Kul-
turabteilung, für diese Arbeiten und die
Mitarbeit einer Reihe von Kolleginnen und
Kollegen ist in den hier zitierten Arbeiten
gedankt worden. Besonders möchten wir
uns bei Herrn Richard Niederreiter/Mond-
see/O.Ö (www.uwitec.at) für die Unterstüt-
zung bei der Entwicklung der Sammelgeräte
bedanken.
LiteraturBAUERNFEIND E. & U.H. HUMPESCH (2001): Die Eintags-
fliegen Zentraleuropas (Insecta: Ephemeropt-era): Bestimmung und Ökologie. — Verlagdes Naturhistorischen Museums Wien: 1-239.
DICK J.T.A. & D. PLATVOET (2000): Invading predato-ry crustacean Dikerogammarus villosus elimi-nates both native and exotic species. — Proc.Roy. Soc, Series B 267: 977-983.
FESL C. (2002): Biodiversity and resource use of lar-val chironomids in relation to environmentalfactor in a large river. — Freshw. Biol. 7:1065-1087.
FESL C. & U.H. HUMPESCH (2003): Community struc-ture and resource use of oligochaetes (Anne-lida) in relation to hydrophysical factors in alarge river. — Arch. Hydrobiol., Suppl. 147,Large Rivers 14: 307-326.
FESL C. & U.H. HUMPESCH (2005): Spatio-temporalvariability of benthic community structureand resource utilisation patterns in relationto environmental factors in a large river. —Arch. Hydrobiol., Suppl., Large Rivers: imDruck.
FESL C, HUMPESCH U.H. S A. ASCHAUER (1999): The re-
lationship between habitat structure and bio-diversity of the macrozoobenthos in the free-flowing section of the Danube in Austria -east of Vienna (preliminary results). — Arch.Hydrobiol., Suppl. 115, Large Rivers 11: 349-374.
FRANK C. & P.L. REISCHUTZ (1994): Rote Liste gefähr-deter Weichtiere Österreichs (Mollusca: Ga-stropoda und Bivalvia). — In: GEPP J. (Hrsg.):Rote Listen gefährdeter Tiere Österreichs. Sty-ria Medienservice, Graz: 283-316.
GEPPJ. (Hrsg.; 1994): Rote Listen gefährdeter TiereÖsterreichs. — Styria Medienservice, Graz: 1-355.
HUMPESCH U.H. (1992): Ecosystem study Alten-wörth: impacts of a hydroelectric power-sta-tion on the River Danube in Austria. — Fresh-water Forum 1: 33-58.
HUMPESCH U.H. S J.M. ELUOTT (Eds.; 1990): Methodsof biological sampling in a large deep river -the Danube in Austria. — Wasser und Abwas-ser, Suppl. 2: 1-83.
157
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at

HUMPESCH U.H. & R. NIEDERREITER (1993): Freeze-coremethod for sampling the vertical distributionof the macrozoobenthos in the main channelof a large deep river. The River Danube at riv-er kilometre 1889. — Arch. Hydrobiol., Suppl.101, Large Rivers 9: 87-90.
HUMPESCH U.H., FESL C. & H. ROGER (2002): The effect
of riverbed management on the habitatstructure and macroinvertebrate communityof a ninth order river, the Danube in Austria.— Arch. Hydrobiol., Suppl. 14, Large Rivers13: 29-46.
ILUES J. & L. BOTOSANEANU (1963): Problemes et me-thodes de la classification et de la zonationecologique des aux courantes, considereessurtout du pont de vue faunistique. — Verh.intern. Verein, theor. angew. Limnol. 12: 1-57.
LIEPOLT R. (1967): Limnologie der Donau. — E.Schweizerbart'sche Verlagsbuchhandlung,Stuttgart V: 4-69.
MAUCKY H. (1994): Rote Liste der gefährdetenKöcherfliegen Österreichs (Trichoptera). In:GEPP J. (Hrsg.): Rote Listen gefährdeter TiereÖsterreichs. Styria Medienservice, Graz: 207-214.
MOOG O., HUMPESCH U.H. & M. KONAR (1995): The
distribution of benthic invertebrates alongthe Austrian stretch of the River Danube andits relevance as an indicator of zoogeograph-ical and water quality patterns - part 1. —Arch. Hydrobiol., Suppl. 101, Large Rivers 9:121-213.
MOOG O., BRUNNER S., HUMPESCH U.H. & A. SCHMIDT-
KLOIBER (2000): The distribution of benthic in-vertebrates along the Austrian stretch of theRiver Danube and its relevance as an indica-tor of zoogeographical and water qualitypatterns - part 2. — Arch. Hydrobiol., Suppl.115, Large Rivers 11: 473-509.
MÜLLER J. C, SCHRAMM S. S A. SErrz (2002): Geneticand morphological differentiation ofDikerogammarus invaders and their invasionhistory in Central Europe. — Freshw. Biol. 47:2039-2048.
Oxbow River & Stream Restoration, Inc. (2003):www.oxbowriver.com / Web_Pages /Stream_Ecology_Pages / Ecology_Riparian /Ecology_RCC.html.
PECSI T. S B. ERDEUCS (1970): PalOdicella articulataEHRENBERG (Bryozoa, Ectoprocta) and Urnatel-la gracilis LEIDY (Kamptozoa), new for theHungarian reach of the Danube. — Arch. Hy-drobiolVSuppl. XXXVI (Donauforschung IV)2/3: 293-298.
PEHOFER H.E. (1998): A new quantitative air-liftsampler for collecting invertebrates designedfor operation in deep, fast-flowing gravelbedrivers. — Arch. Hydrobiol., Suppl. 115, LargeRivers 11: 213-232.
PÖCKL M., WEBB B.W. & D.W. SUTCUFFE (2003): Life
history and reproductive capacity of Oam-marus fossarum and G. roeseli (Crustacea:
Amphipoda) under naturally fluctuating wa-
ter temperatures: a simulation study. —
Freshw. Biol. 48: 53-66.
RYLAND J.S. (1970): Bryozoans. — Hutchinson Univ.
Library. London: 1-175.
TrmzER T. (1997): Ausbreitung aquatischer Neo-
zoen (Makrozoobenthos) in den europäischen
Wasserstraßen, erläutert am Beispiel des
Main-Donau-Kanals. — In: KAVKA G. (Hrsg.):
Güteentwicklung der Donau - Rückblick und
Perspektiven. Schriftenreihe des Bundesamtes
für Wasserwirtschaft 4: 113-134.
VANOTTE R. L., MINSHALL G.W., CUMMINGS K.W., SEDELL
J.R. & C.E. CUSHING (1980): The river continuum
concept. — Canad. J. Fisheries Aquat. Sei. 37:
130-137.
WIEBACH F. (1960): Bryozoa, Moostierchen. — In:
BROHMER P., EHRMANN P. & G. ULMER (Hrsg.): Die
Tierwelt Mitteleuropas, Leipzig 1(8): 1-56.
Wöss E.R. (1996): Life-history variation in freshwa-
ter bryozoans. — In: GORDON D.P., SMITH A.M.
& J.A. GRANT-MACKIE (Eds.): Bryozoans in Space
and Time. National Institute of Water and At-
mospheric Research Ltd. Wellington, New
Zealand: 391-399.
Wöss E.R. (2005): The distribution of freshwater
bryozoans in Austria. — In: MOYANO G.H.I.,
CANCINO J.M. & P.N. WYSE JACKSON (Eds.): Bryo-
zoan Studies 2004. A.A. Balkema Publishers,
Leiden, London, New York, Philadelphia, Sin-
gapore: 369-374.
Anschrift der Autoren:
Mag. Dr. Christian FESL
Department of Freshwater Ecology
University of Vienna
Althanstrasse 14
A-1090 Vienna, Austria
E-Mail: [email protected]
Univ.-Prof. Mag. Dr. Uwe H. HUMPESCH
Institute of Limnology of the Austrian
Academy of Sciences
Mondsees trasse 9
A-5310 Mondsee, Austria
E-Mail: [email protected]
Mag. Dr. Emmy R. WÖSS
Department of Freshwater Ecology
University of Vienna
Althanstrasse 14
A-1090 Wien, Austria
E-Mail: [email protected]
158
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at







![Auswertungen Makrozoobenthos - Bayern · 2020. 8. 11. · Makrozoobenthos 1 Aufgabenstellung Das Bayerische Landesamt für Umwelt [= LfU] betreibt ein Projekt mit dem Titel "25 Jahre](https://static.fdocuments.net/doc/165x107/610d11c683b9f9086c7fde28/auswertungen-makrozoobenthos-bayern-2020-8-11-makrozoobenthos-1-aufgabenstellung.jpg)










