
An Estimation of Above Ground Tree Biomass of a Mangrove ...
15
TROPICS Vol. r'(4), pp. 243-257. Issued May 1992 An Estimation of Above Ground Tree Biomass of a Mangrove Forest in East Sumatra, Indonesia Cecep KUSMANA Faculty of Agriculture, Kyoto University, Kyoto 606, Japan. Supiandi SABIHAM Faculty of Agriculture, Bogor Agricultural University, Bogor, Indonesia. Kenichi ABE The Center for Southeast Asian Studies, Kyoto University, Kyoto 606, Japan. Hiroyuki WATANABE Faculty of Agriculture, Kyoto University, Kyoto 606, Japan. Abstract Above ground tree biomass was estimated in Bruguiera parviflora, B. sexangula, and Rhizophora apiculata communities of a mangrove forest in Talidendang Besar, East Sumatra, Indonesia. The research was carried out from December 1990 to January 1991, and allometric relation method was used to estimate the above ground biomass of tree species having a diameter of 10 cm and over. The above ground biomass in this forest ranged from 42.94 to 89.68 t d.wt/ha in a B. parviflora community; 75.99 to 279.03 t d.wt/ha in a B. sexangula community; and 40.70 t d.wt/ha in aR. apiculata community. Key words: above !,'Tound biomass / allometric relation method / Bruguiera parviflora / Bruguiera sexan- gula / EastSumatra /Indonesia / mangrove forest / Rhizophora apiculata / Talidendang Besar. Cintron and Novelli (1984) stated that above ground biomass is the amount of standing organic matter per unit area at a given time, which is related to a function of productivity system, stand age and organic allocation, and exportation strategies. The estimation of above ground biomass not only provides increasingly valuable means for making comparisons among ecosystems as well as for evaluating world wide productivity patterns (Rodin and Bazilevich, 1967), but is also very important for the study of the functional aspects of forests such as primary productivity, nutrient cycling and energy flow (Hasse et al., 1985). Conse- quently, biomass data is important in order to understand forest ecosystem characteristics to establish the proper management system based upon the sustainable yield principle. Variability in biomass data may be attributable to stand age, stand history, or structural differences (Lugo and Snedaker, 1974), and climatic factors such as temperature and precipi- tation (Komiyama et al., 1988). The world literature on forest biomass and production expanded rapidly in the 1960s and 1970s. Cannell (1982) reviewed the world forest biomass data from 600 papers which were written by scientists all over the world. However, the papers on the biomass of mangrove forests were scarce. Some researchers who worked on standing biomass of mangrove forests were Golley et al. (1962) in Puerto Rico, Golley et al. (1969) in Panama, Lugo and Snedaker (1974) in South Florida, Briggs (1977) in Australia, Christensen (1978) in Phuket Island - Southern Thailand, Nakasuga (1979) in Ryukyu Islands - Southern Japan, Suzuki and Tagawa (1983) in Ishigaki Island - Southern Japan, Ong et al. (1984) in Malaysia, Tarnai et al. (1986) in Ranong - Southern Thailand, and Komiyama et al. (1988) in Maluku Island - East Indonesia. In Indonesia, while the mangrove forests are varied from place to place in structure and species composition (Soemodihardjo and Soerianegara, 1989) and are believed to be the
Transcript of An Estimation of Above Ground Tree Biomass of a Mangrove ...

TROPICS Vol. r'(4), pp. 243-257. Issued May 1992
An Estimation of Above Ground Tree Biomass of a Mangrove Forest in East Sumatra, Indonesia
Cecep KUSMANA Faculty of Agriculture, Kyoto University, Kyoto 606, Japan.
Supiandi SABIHAM Faculty of Agriculture, Bogor Agricultural University, Bogor, Indonesia.
Kenichi ABE The Center for Southeast Asian Studies, Kyoto University, Kyoto 606, Japan.
Hiroyuki WATANABE Faculty of Agriculture, Kyoto University, Kyoto 606, Japan.
Abstract Above ground tree biomass was estimated in Bruguiera parviflora, B. sexangula, and Rhizophora apiculata communities of a mangrove forest in Talidendang Besar, East Sumatra, Indonesia. The research was carried out from December 1990 to January 1991, and allometric relation method was used to estimate the above ground biomass of tree species having a diameter of 10 cm and over. The above ground biomass in this forest ranged from 42.94 to 89.68 t d.wt/ha in a B. parviflora community; 75.99 to 279.03 t d.wt/ha in a B. sexangula community; and 40.70 t d.wt/ha in aR. apiculata community.
Key words: above !,'Tound biomass / allometric relation method / Bruguiera parviflora / Bruguiera sexangula / EastSumatra /Indonesia / mangrove forest / Rhizophora apiculata / Talidendang Besar.
Cintron and Novelli (1984) stated that above ground biomass is the amount of standing organic matter per unit area at a given time, which is related to a function of productivity system, stand age and organic allocation, and exportation strategies. The estimation of above ground biomass not only provides increasingly valuable means for making comparisons
among ecosystems as well as for evaluating world wide productivity patterns (Rodin and Bazilevich, 1967), but is also very important for the study of the functional aspects of forests
such as primary productivity, nutrient cycling and energy flow (Hasse et al., 1985). Conse
quently, biomass data is important in order to understand forest ecosystem characteristics to
establish the proper management system based upon the sustainable yield principle. Variability in biomass data may be attributable to stand age, stand history, or structural
differences (Lugo and Snedaker, 1974), and climatic factors such as temperature and precipitation (Komiyama et al., 1988). The world literature on forest biomass and production
expanded rapidly in the 1960s and 1970s. Cannell (1982) reviewed the world forest biomass data from 600 papers which were written by scientists all over the world. However, the
papers on the biomass of mangrove forests were scarce. Some researchers who worked on standing biomass of mangrove forests were Golley et al. (1962) in Puerto Rico, Golley et al. (1969) in Panama, Lugo and Snedaker (1974) in South Florida, Briggs (1977) in Australia, Christensen (1978) in Phuket Island - Southern Thailand, Nakasuga (1979) in Ryukyu Islands - Southern Japan, Suzuki and Tagawa (1983) in Ishigaki Island - Southern Japan, Ong et al. (1984) in Malaysia, Tarnai et al. (1986) in Ranong - Southern Thailand, and Komiyama
et al. (1988) in Maluku Island - East Indonesia. In Indonesia, while the mangrove forests are varied from place to place in structure and
species composition (Soemodihardjo and Soerianegara, 1989) and are believed to be the

244 C. KusuANA, S. SaSTHAM, K. AeE & H, WnTnXABE
largest (in terms of total coverage) in the world (Christensen, 1982), there was only one paper
published on the biomass of mangrove, i.e. the paper on the biomass of a natural mangrove
forest in Maluku Island (Komiyama et al., 1988). Therefore, some mangrove forests inIndonesia had been damaged due to limited scientific knowledge of their productivity inmanaging and utilizing them (Soegiarto, 1984; Soemodihardjo and Soerianegara, 1989) and
haphazard felling had led to severe exploitation during Dutch colonization in the 16th century
(Sukardjo, 1987).
In Indonesia, the mangrove rree species belonging to the genus Rhizophora, Bruguiera,
and Ceriops are considered as commercial rree species (Direktur Jenderal Kehutanan, 1978),
so that the biomass of those free species in the forests is useful information for the mangrove
forest management practices.
In order to contribute to the understanding of the Indonesian mangrove forestproductivity, this research was undertaken as an attempt to estimate the above ground
biomass of various morphological compartments of the trees which have commercial size
(diameter 10 cm and over) in the mangrove forest communities in Taiidendang Besar, Riau
in relation to the mangrove tree species composition.
STUDY SITES
This research was carried out from December 1990 to January 1991 in a mangrove forest
concession area of Bina Lestari Company at Talidendang Besar, Riau (l,ong. 103" 28' to 103"
48' E, Lat. 0o 21' to lo N) which lies at the east coast of Sumatra, Riau Province, Indonesia
(Fig. 1). This concession area is on gentle topography and includes some small rivers. Soil is
organosols and glei humus originating from alluvial parent materials (kmbaga Penelitian
Tanah, 1964).No meteorological data were available for Talidendang Besar. The climatic data of
Mandah, the neare.st area to Talidendang Besar, provides a fairly good approximation ofthe climatic conditions (Fig. l). Average annual rainfall is about 1,335 mm. According to
the Schmidt and Ferguson system (1951), the climate of this area belongs to the minfall type
B (Q-value 28.6Vo) with seven wet months, two dry months, and three humid months.
The mangrove forest of Talidendang Besar is characterized by the existance of R. apiculata
in the transition zone with a fresh water swamp forest and Bruguiera species which spread
from seacoast to landward. The seaward fringe tends to be occupied by B. parviflora, whllethe landward zone tends to be occupied by B. sexangula.In the transition zone which is infre-
quently submerged by sea water, the stand of Rhizophora apiculata which intermingles withNypa fruticans showed a well-developed growth, where the tallest and largest diameter tree
recorded were 34 m and 49.8 cm, respectively.
METHODS
In order to cover all conditions of the research sites, six 50 X 50 m sample plots were set up at
representative areas in this mangrove forest. A sample plot (P1) was set up in a well-devel-oped stand of B. parviflora and another (P2) in an area where many individual trees were
dead in a B. parviflora community. In a,B. sexangula community, a sample plot (P3) was set
up in an area frequently submerged by sea water, another (P4) in an area further inland which
is less submerged by sea water and another (P5) in a well-developed stand of B. sexangula

Above ground tree biomass of a mangrove forest in East Sumatra 245
114" 138"
12"N
12" S
N210
190
170
150
1S
110
90
70
50
30
t0
Fig. 1. Location and climatic condition of the mangrove forest in Talidendang Besar, Riau,East Sumatra.
near to the transition zone which is least submerged by sea water. A sample plot (P6) wasalso set up in a well-developed stand of R. apiculara in the transition zone.
Within each sample plot, all the frees with diameters of 10 cm and over were identifiedand measured in height and diameter. Diameters of the trees were measured at20 cm above
the highest prop-roots for R. apiculata and at the breast-height (1.3 m above the ground level,DBH) for the other species.
Twenty one frees which covered the range of diameter size classes in the sites were fellednear ground level. These sample trees consisted of seven of B . parviflora, nine of B. sexangu-la, and f,rve of R. apiculata which were selected from B. parviflora, B. sexangula, andR.apiculata communities, respectively. The largest diameters of the sample tree of B.parviflora, B. sexangula, and R. apiculata were 24.8 cm, 40.5 cm, and 49.5 cm, respectively.Crown width, height of the extent of prop-roots (for R. apiculata), stem diameter at ground
level, stem diameter at breast-height, stem diameter at 20 cm above the highest prop-roots,stem diameter at the lowest living branch, stem diameter at 0.1 of total height, total tree
height, and the height of the lowest Iiving branch were measured from each sample tree.
Aerial parts of knee roots of B. parviJlora as well as B. sexangula,which sprouted from the
soil, were not harvested, because we could not determine which of them were from thesample tree without excavating. The stem of the sample tree was divided into horizons of I mlong. From each horizon, a fresh weight of each morphological tree compartment, i.e. stems,
branches, leaves, flowers and fruits, and prop-roots were separately weighed and recorded. A
-ccoEEE
6c6E
I Research Area Wffin $j::fiF"*l

246 C. KUSvTANA, S. SaSTHAM, K. AgE & H, WNTNNABE
small amount sample of each tree compartment from each stem horizon was taken and dried
in an oven at 80t for 48 hours to obtain the constant dry weight. lraf area (one-sided) ofsmall amount sample of leaves from each stem horizon was measured with a leaf-area meter
(Type AAM-7, Hayashi Denko).
RESULTS
Table I shows the description of mangrove tree species in the sample plot in each forest
community. Three species of the trees (diameter 10 cm and over) were recorded within the
sample plots in this mangrove forest. Remarkably, all trees recorded in this forest belong to
the genera Rhizophora and Bruguiera; therefore, all of them are regarded as commercial tree
species. B. paniflora was a dominant species in sample plots 1 and 2. Density and basal area
of this species were 300 ind./ha and 5.63 mzlha in the sample plot 1, and 96 ind.lha and2.tImzlha in the sample plot 2. Sample plots 3, 4 and 5 were dominated by B. sexangula. Density
and basal area of this species were 112 ind.lha and 3.70 mzlha in the sample plot 3, 248 ind.
/ha and 14.05 mzlha in the sample plot 4, and296 ind./ha and2o.9l mzlha in the sample plot
5. In the sample plot 6, R. apiculata was a dominant species with a density of 32 ind./ha and
basal area of 2.50 m2lha.
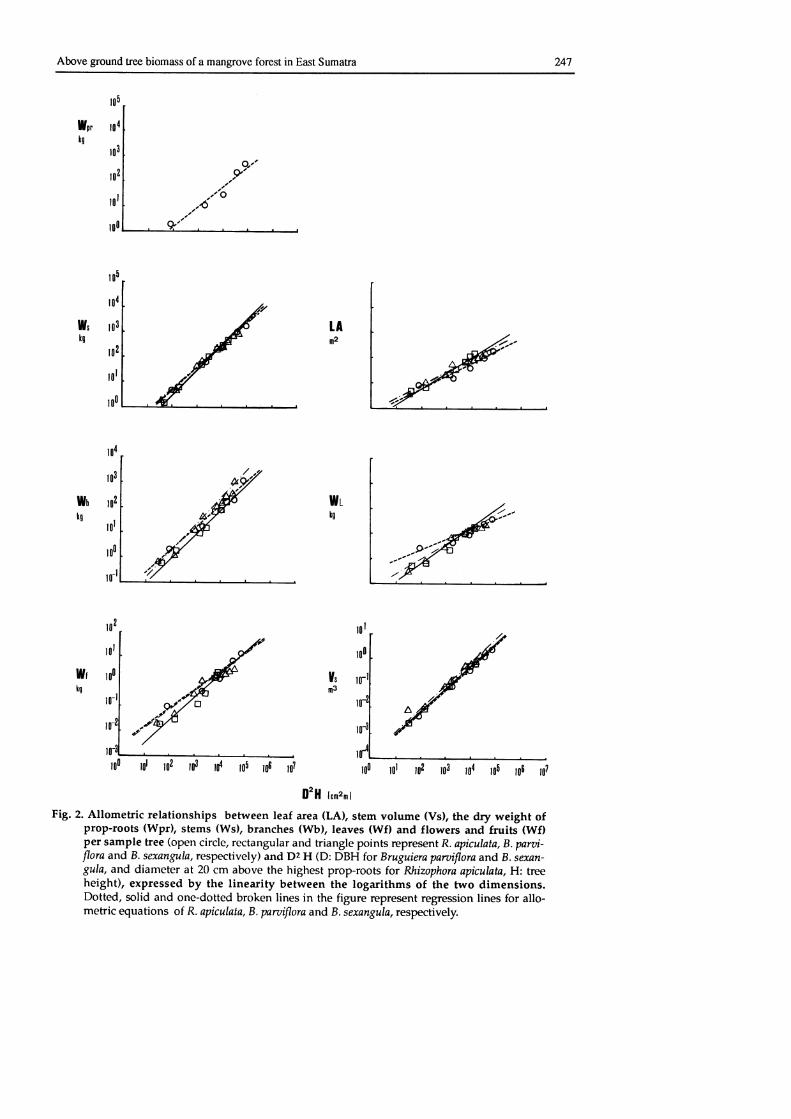
Fig. 2 shows the relationships between the dry weight of each above ground morphologi-
cal compartment, leaf area and stem volume of sample trees, and the prcduct of the square ofdiameter and rree height (D2H). These relationships can be expressed by the following simple
allometric equation:
Wx = b (D2H)a
where Wx is the dry weight of the compartment x (x is s for stem, b for branches, I for leaves,
f for flowers and fruits, and pr for prop-roots) while Wx indicates leaf area when Wx is LAand stem volume when Wx is Vs; D is the diameter at 20 cm above the highest prop-roots for
R. apiculata and diameter at breast-height (DBH) for B. parviflora and B. sexangula; H istotal length of the tree stem above ground level; and a and b are constants. The stem volume
of an individual tree is the total volume of each stem horizon (log) excluding the butress or
stem in which prop-roots exist. Smalian's formula (cited from Loetsch et aI., 1973) was
adopted to estimate the volume of each stem horizon of a sample tree as given byi
r. (Gt + G2)\/c
= f
2 L-t
where Vs is the volume of the log (stem horizon), G 1
ends of the log, and L is the log length.
The coefficients of allometric equations, a and b,
method and the results are shown in Table 2.
and G2 are cross-sectional area at both
were determined by the least square
According to t-statistical test, the regression coefficients "a" of stems, branches and stem
volume of each tree species as well as that of flowers and fruits of. B. parviflorc and prop-
roots of R. apiculata were significantly close to I at the level of P < 0.01. This means the dry
weights of these parts proportionately increased with DzH. However, the regression coeffi-
cients "a" of leaves and leaf areas of each tree species as well as that of flowers and fruits ofB. sexangulc and R. apiculata were significantly less than 1 at the level of P < 0.05;
Above ground tree biomass of a mangrove forest in East Sumatra 247
att'ttt Q
.'6
Wr
ls
t03 ro5 to8
D2ll lcmzml
Fig. 2. Allometric relationships between leaf area (LA), stem volume (Vs), the dry weight ofProP'roots (Wpr), stems (Ws), branches (Wb), leaves (W0 and flowers and fruits (wf)Per sample tree (open circle, rectangular and triangle points represent R. aViculata, B. part i-flora and B. semngula, respectively) and Dr H (D: DBH for Bruguiera paruiflora and B. semn-gula, and diameter at 20 cm above the highest proproots for Rhinphora apicuhta, H: treeheight), expressed by the linearity between the logarithms of the two dimensions.Dotted, solid and one'dotted broken lines in the figure rcpresent regression lines for allo-mehic equations of R. apiculata, B. pantiflora and B. semngul4 respectively.
t05
Wpr toilg
t03
t02
l0t
t00
U lo3kg
t02
t0l
100
105
t04
tAm2
t04
t03
Wrt0
Wr tozks ror
100
| 0-l
Ut63
t08t05104t03n2t0lt00t02t00

248 C. KusvANA, S. SasrHAM, K. Aee & H, WnrnxABE
Table l. The description of mangrove free species having diameter of 10 cm and over at eachsample plot in the mangrove forest communities in Talidendang Besar, Riau.
Forest community Sample Tree species Density Basal area Aug. Aug.Diam. (cm) H (m)plot (ind. /ha) (m2 /ha)
Bruguiera palvifloracommunitv
Bruguiera sexangulacommunity
Rhizophora apiculatacommunity
B. paruifloraB. sexangula
B. paruifloraB. sexangula
B. pantifloraB. sexangula
B. paruifloraB. sexangula
B. paruifloraB. sexangula
R. apiculata
300100
9660
76712
60248
52296
32
5.633.58
2.171.87
1.313.70
1.1514.05
1.2020.97
2.50
15.727.5
16.519.9
74.820.5
75.426.7
16.929.7
31.8
20.117.5
20.818.2
15.518.6
17.223.7
17.825.7
29.5
therefore, the increasing rate of the dry weights of these parts decreased with increases ofDzH. The growth rate of the trees seems to decrease with age or increasing tree size because
the biomass ofphotosynthetic parts (leaves) does not increase as fast as that ofnon-photosyn-thetic parts. Conversely, the increasing rate of the dry weight of flowers and fruits of B. parv-
iflora which proportionally increased with increasing tree size means that the bigger trees
have proportionally more flowers and fruits than smaller trees, and more or less B. parviflora
is regarded as a vigorous tree species to produce flowers and fruits more than the other two
species.
Using the allometric equations with the coefficients as shown in Table 2, the dry weight ofeach compartment of all individual trees having commercial size (diameter l0 cm and over)
in the sample plots were calculated. The above ground biomass of the trees having commer-
cial size was estimated by summing up the dry weight of their above ground compartments
in the sample plots (Table 3). The mean above ground biomass of the trees having commer-
cial size were estimated to be 66.31 t d.rt/ha ranging from 42.94 to 89.68 t d.wtlha in a.B.
parviflora community; 177.94 t d.wt/ha ranging from 75.99 , 178.81 to 279.03 t d.wt/ha in a
B. sexangula community; and 40.70 t d.wt/ha in a R. apiculata community. The estimated
leaf area index (LAI) and stem volume of those tree species ranged from 1.07 to 2.14 halha
and 12.82 to 26.81 63/ha in a B. parviflora community, l.41to 3.61 ha/ha and 21.99 to 8O.62
m3lha in a B. sexangula community, and 0.23 ha,/ha and 8.98 m:/ha in a R. apiculata commu-
nity, respectively.
The above ground tree biomass in the sample plots tended to increase with increases ofbasal area and density (Fig. 3). The above gtound tree biomass in the sample plots in this
mangrove forest linearly increased with increases of basal area, but it did not significantlyincrease proportionately with increases of density (Fig. 3).
The stand in R. apiculatc community had a taller canopy, but its density and basal area
were much smaller than the stands in the other communities. It resulted in a smaller above
ground biomass of the rrees in a R. apiculata community.

Above ground tree biomass of a mangrove forest in East Sumatra 249
Table 2. Coefficients "a" and "b" of allometric equations between Dz H and the dry weight ofthe above ground morphological tree compartmcnts, leaf arca and stem volume.
B. paruiflora B. sexangula R. apiculata
No. Trees
compartment
1. Stem
2. Branch
3. Flower and fruit
4. Leaf
5. Prop-root
6. Leaf area
7. Stem volume
0.98710 0.03715
0.96790 0.01093
0.90224 0.00041
0.58504 0.02s22
0.52765 0.30562
0.96592 0.00007
0.99** o.g9o23
0.98** t.o2g25
o.g4** o.73o2g
0.96** o.60l(>4
o.g'** o.sog24
0.99** o.gMtg
0.07957 0.gg**
0.07257 0.95**
0.00211 0.gg**
0.05247 0.97**
0.77 672 0.g5**
0.00009 0.gg**
0.90292 0.0g070 0.gg**
0.903g0 0.02940 0.97**
0.7 477 4 0.0020g 0.96**
0.40M7 0.33853 0.92*
0.87637 0.03594 0.95*
0.47357 0.82118 0.89*
0.92397 0.00009 0.gg**
r means coefficient correlation.* and ** indicate significance at the levels of P < 0.05 and P < 0.01, respectively.
Table 3. Estimated above ground biomass, leaf area index (LAI), and stem volume of the treeswith a diameter of l0 cm and over at the sample plots in the mangrove forest commu-nities in Talidendang Besar, Riau.
No. Vegetationcharacteristics
B. paraifloracommunity
P1 P2
B. sexangulacommunity
R. apiculatacommunity
P6P3 P5P4
1. Above ground biomass( t a. wt/ ha)
1.1. Stem
7.2. Branch
1.3. Flower and fruit
7.4. Prop-root
Total non-pho tosyn theticcomponent
1.5. Leaf
Total above ground biomass
6.LAI(ha/ha)
7. Stem volume (me /ha)
69.46(77.4s)16.42
(18.31)
0.37( 0.41)
86.25(96.17)
3.43( 3.83)
89.68
2.14
26.87
33.35(77.67)
7.84(18.26)
0.20( 0.46)
41.39
(e6.3e)
1.55
( 3.61)
42.94
7.07
12.82 21.99 52.77
765.96(se.48)706.44(38.1s)
0.91( 0.32)
273.37
(97.es)
5.72( 2.0s)
279.03
3.61,
80.62
25.55
62.78)9.46
(23.24)
0.12( 0.2e)
5.72(12.58)
40.25
(e8.8e)
0.45( 1 .11)
40.70
0.23
8.98
46.87 707.99(61.68) (60.3e)
26.72 66.03(3s.16) (36.e3)0.28 0.62
( 0.37) ( 0.3s)
73.87 774.&(e7.27) (97.67)
2.12 4.1,7
( 2.7q ( 2.33)75.99 779.91,
7.41 2.69
Pl to P6 indicate sample plots 1 to 6.
Values in the parentheses are percentage of the biomass of the tree morphological compartment inthe total above ground biomass in the sample plot.
Within each sample plot in each forest community, non-photosynthetic componentcontributed a large proportion (above 96Vo) to the above ground biomass matters, where less
than l%o of which corresponded to flowers and fruits. However, leaves as photosyntheticcomponent showed very little portion, i.e. averaged 3.72Vo, 2.39Vo, and l.llVo to the aboveground biomass in a B. parviflora, B. sexangula. and R. apiculata communities, respectively.

250 C. KUSTT,TANA, S. SNSTHAM, K. Agg & H, WNTNNABE
I80
:oa.40I@
ozBr*E,ouJ
o1 24o
o 11o toirnr,r, ( r'o./u )
Fig.3. The relationship between above gtound biomass and basal area (A) and density G)of the trees at the sample plot in the study area.
1 to 5 indicate sample plot 1 to 6. Asterisk (**), significant at the level of P < 0.01.
In a R. apiculata community prop-roots shared about l2.58Vo to the above ground biomass
matters. Consequently, prop-roots biomass seems to enlarge non-photosynthetic component
of the above ground biomass of the trees in this community.

Above ground tree biomass of a mangrove forest in East Sumatra 25r
The mean specific leaf area (SLA) of B. parviflora, B. sexangula, and R. apiculata areestimated to be 98.66, 90.47, and 57.28 cm2lg d.wt, respectively. It indicates that while theleaves of R. apiculata were the thickest, the leaves of B. sexangula were thicker than those ofB. parviflora.
DISCUSSION
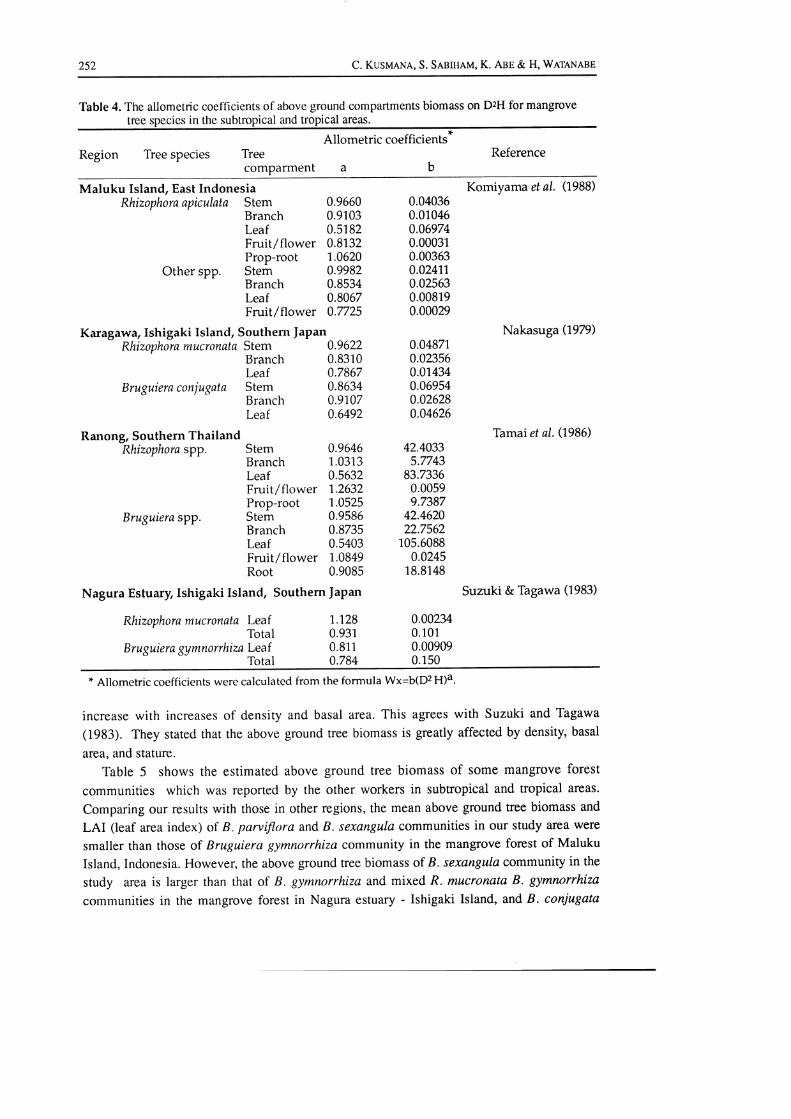
Table 4 shows the allometric coefficients which were calculated from the allometric relationsbetween the biomass of various tree compartments on DzH for mangrove tree species in thesubtropical and tropical areas. Comparison of our allometric coefficients obtained with thosereported for mangrove tree species in the other mangrove forests shows that the allometriccoefficient "a" of the stem, branch, and leaf biomass for R. apiculata, B. parviflora, and B.sexangula in the study area was comparable to that of Rhizophora and.Bruguiera species inthe mangrove forest of Maluku Island - East Indonesia, Karagawa - Ishigaki Island - SouthernJapan, Ranong - Southern Thailand, and Puerto Rico. While the allometric coefficient "a" ofthe fruit and flower biomass of B. paniflora in the study area was comparable to that ofBruguiera spp. in the mangrove forest of Ranong - Southern Thailand, the allometric coeffi-cient "a" of the prop-root biomass of R. apiculara in the study area was comparable (close to1) to that of Rhizophora species in the mangrove forest of Maluku Island, Ranong, and puer-
to Rico. In conrrast to the Rhizophora and Bruguiera species in the other mangrove forests, R.mucronata and B. gymnorrhiza in the mangrove forest of Nagura estuary, Ishigaki Island -Southern Japan seem to grow vigorously to produce leaves as reflected by the allometriccoefficient "a" which was close to I of the leaf biomass of those species. Besides, theconsiderably close to 1 of the allometric coefficient "a" of the fruit and flower biomass ofRhizophora spp. and Bruguiera spp. in the Ranong mangrove forest as well as that of B.parviflora in our study area means that these tree species in these forests vigorously grow toproduce fruits and flowers as tree size increases as compared with the Rhizophora andBruguiera species in the other mangrove forests.
The stand of R. apiculdta community in this mangrove forest had about l2.58Vo of thetotal above ground biomass as prop-roots and is recognized as comparable to that of theRhizophora community in the mangrove forest of Maluku Island in which the prop-rootsbiomass is as large as 13.5 t/ha (Komiyama et a1.,1988). This foresr community must be clas-sified as "estuarine" forest according to the description of Florida mangroves given by Lugoand Snedaker (1974). They found a lower proportion ofprop-roots in estuarine (2Vo, l5vo)than in finge (23o/o,26Vo, 47Vo), overwash (35Vo, 4OVo) and scrub (53Vo) forests. In this case,Christensen (1978) stated that the relatively small number of prop-roots in Rhizophora forestsgrowing under estuarine conditions might be due to reduced salinity or a better nutrientsupply.
As indicated by specific leaf area, the greater thickness of the leaves of R. apiculata thanthoseof B.parvifloraandB. sexangulaindicatesthatR. apiculataisashade-intoleranttreespecies as reported by Macnae (1968). It is also reported thatR. apiculata in the mangroveforest of Maluku lsland had sparser foliage of thicker leaves than B. gymnorrhiza (Komiyamaet al.,1988), and similar results were also reported by Ninomiya et al. (1989) for R. stylosaand B. gymnorrhiza in the mangrove forest of Funaura Bay, Okinawa, Japan.
In this mangrove forest, the above gtound tree biomass in the sample plots tended to
252 C. KUSUANA, S. SEETHAM, K. AEE & H, WNTNNABE
Table 4. The allometric coefficients of above ground compartments biomass on D2H for mangrovetree species in the subtropical and tropical areas.
Allometric coefficients*
abRegion Tree species Tree
comparmentReference
Maluku Island, East IndonesiaRhizophora apiculata Stem
BranchLeafFruit/flowerProp-root
Other spp. StemBranchLeafFruit/flower
Karag awa,Ishigaki Island, Southern |apanRhizophora mucronata Stem
BranchLeaf
Bruguiera conjugata StemBranchLeaf
Ranong, Southern ThailandRhizophora spp. Stem
BranchLeaf
Bruguiera spp.
Fruit/flowerProp-rootStemBranchLeafFruit/flowerRoot
0.96600.91030.51820.8132,'t.0620
0.99820.85340.80670.7725
0.96220.83100.78670.86340.91070.6492
0.96461 .03130.56321.26321.05250.95860.87350.54031.08490.908s
Nagura Estu Ny,Ishigaki Islaild, Southern f apan
71280.9310.8110.784
0.040360.010460.069740.000310.003630.02411,0.025630.008190.00029
0.048710.023560.01 4340.069540.026280.04626
42.40335.7743
83.73360.00599.7387
42.462022.7562
105.60880.0245
18.8148
0.002340.1010.009090.1 50
Komiyama et al. (1988)
Nakasuga (1979)
Thmai et al. (1986)
Suzuki & Tagawa (1983)
Rhizophora mucronata LeafTotal
Bru guiera gymnorrhiza LeafTotal
* Allometric coefficients were calculated from the formula Wx=b(OaH)4.
increase with increases of density and basal area. This agrees with Suzuki and Tagawa
(1983). They stated that the above ground tree biomass is greatly affected by density, basal
area, and stature.
Table 5 shows the estimated above ground tree biomass of some mangrove forest
communities which was reported by the other workers in subtropical and tropical areas.
Comparing our results with those in other regions, the mean above ground tree biomass and
LAI (leaf area index) of B. parviflora and B. sexangula communities in our study area were
smaller than those of Bruguiera gymnorrhiza community in the mangrove forest of Maluku
Island, Indonesia. However, the above ground tree biomass of B. sexangula community in the
study area is larger than that of B. gymnorrhiza and mixed R. mucronata B. gymnorrhiza
communities in the mangrove forest in Nagura estuary - Ishigaki Island, and B. conjugata

Above ground tree biomass of a mangrove forest in East Sumatra 253
Table 5. Estimated above ground tree biomass, leaf area index (LAI), and vegetation structure ofmangrove forest communities in subtropical and tropical areas.
No. Location and Aboveground biomass L A I Density Basal area Avg. canopy Referenceforest cornmunity (t d.wtlha) (halha) (ind./ha) (mzlha) H (m)
1. Maluku Island, East Indonesia Komiyama et aL (1988)Mizophora apiculata 327.9 3.76 586 23.97 18.3
(299.1-356.8) (3.71-3.81) (411,-761) (22.82-25.11) (1s.s-21..2)
Rhizophora stylosa 178.2 2.31 206 13.96 22.3Bruguiera gymnorrhiza 421,.5 8.2 350 35.03 24.4
(4O6.6-436.4) (7.76-8.62) (300-400) (35.91-%.15) (22.4-26.4)
2. Nagura estuary, Ishigaki Island, Southern )apan Suzuki & Tapwa O983)Mizophoramuuonata 108.1 3.2 ur 31.00 5Rhizophora muuonata- B. gymnorrhiza 78.6 2.5 ur 22.70 6Bruguiera gymnorrhiza 97.6 3.1 ur 32.90 6
3. Ryukyu Islands, SouthernJapan Nakasuga (1979)I(andelia candel 7'1.8 0.64 9,600 24.50 5.8Bruguiera conjugata 707.1 7.60 7,522 25.45 6.3
(1.02.7-'t"t1.s) (5.5-8.7) (4,644-'.t0,400) (24.2-26.7) (6342)Nizophora mucronata 185.3 5.4 9,6N 48.3 7.2
4. Ranong, Southern Thailand Tamai et al. (1986)Rhizophora apiculata 398.8 6.58 1,246 23.97 ur
5. Puerto Rico Golley et aI. (1962)Rhizophoramangle 629 4.4 13,600 ur 8.0
6. Panama Golley et aI. (1969)Mizophora braistyla 279.2 ur ' 712 13.6 (30 - 40)
7. Phuket Island, Southern Thailand Christensen (1928)14 to 15 years old Mizophora apiculataplantation forest 159.1 (3.7 - 4.2') ur ur (5 - 11)
8. Southern Florida Lugo and Snedaker (1974)Rhizophora mangle 124.61 ur ur ur ur
('t't9.58-129.64)
9. Malaysia Onget aI. (1984)25 years old Rhizophora apiculataplantation forest 300 ur ur ur ur
l0.Talidendang Besar, Riau, East Sumatra Present studyBruguieraparotflora 66.3'1 1,.61 278 6.ffi 79.2
g2.94-8,9.68)('1.07-2.14) (155-400) (3.9&9.21) (18.8-19.5)Bruguiera sexangula 177.94 2.57 281 14.39 l9.z
(75.99-279.03) 0.41-3.61) (188-348) (5.8s-22.11) ('17.1-21,.8)
Rhizophora apiculata 40.70 0.23 32 z.fr 29.5
The values in the parentheses show range of the data; ur; unreported.
community in Ryukyu Islands - Southern Japan. Furtherrnore, the above ground tree biomassand LAI of R. apicul4td community in the study area were much smaller than that ofRhizophora communities in the other mangrove forests. The average above ground treebiomass for three forest communities in the study area (94.98 t/ha) is almost the same as thatof mangrove forest communities in Nagura estuary - Ishigaki Island, Southern Japan (94.8
Vha).
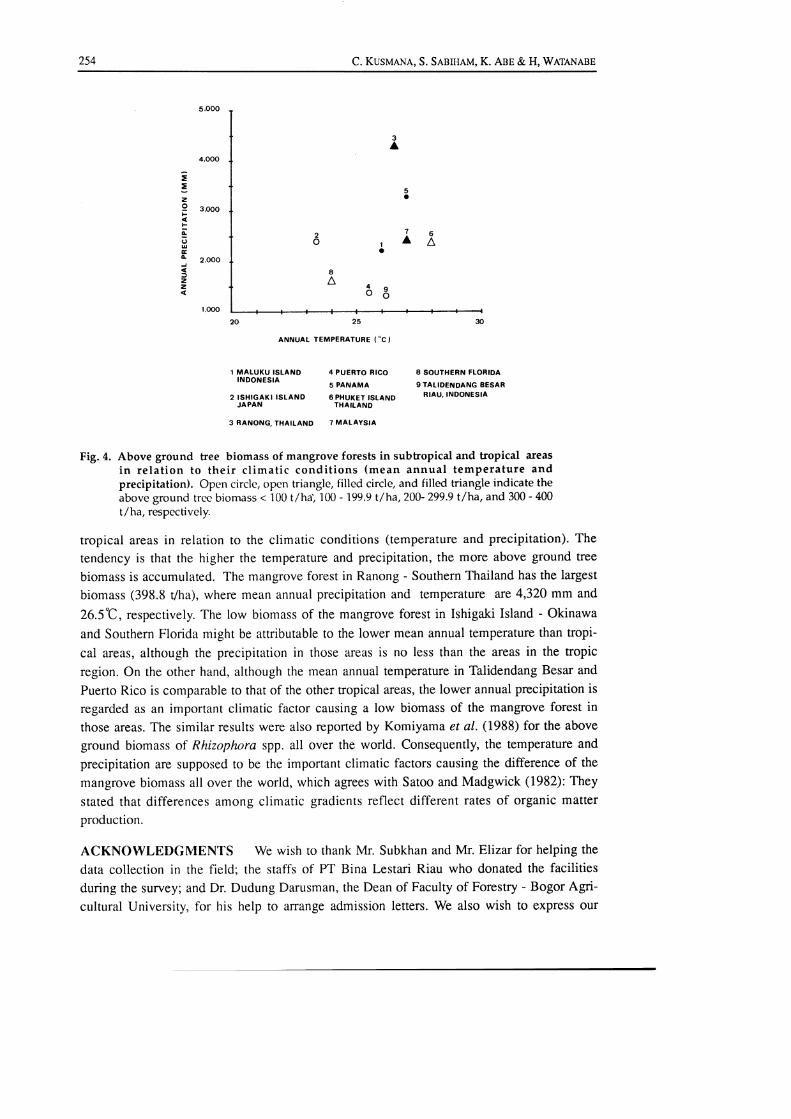
Fig. 4 shows the above ground ffee biomass of the mangrove forests in subtropical and
254 C. KusuANA, S. SaSTHAM, K. Agg & H, WITaNABE
5,ooo
4,OOO
3,OOO
2,OOO
l.OOO
25
ANNUAL TEMPERATURE ( "C )
rNDor{EsrA ,;::[t^"'"" :r].]i::::j:T|1"2lilf,crfK'TSLAND uifiXll.irnl".o"o R|AU'rNDoNEsrA
3 RANOT{G. THAILAND 7 MALAYSIA
Fig. 4. Above ground tree biomass of mangrove forests in subtropical and tropical areas
in relation to their climatic conditions (mean annual temperature andprecipitation). Open circle, open triangle, filled circle, and filled triangle indicate theabove ground trerc biomass < 100 t/ha) 100 - 199.9 t/ha,200-299.9 t/ha, and 300 - 400
t/ha, respcctively.
tropical areas in relation to the climatic conditions (temperature and precipitation). The
tendency is that the higher the temperature and precipitation, the more above ground tree
biomass is accumulated. The mangrove forest in Ranong - Southern Thailand has the largest
biomass (398.3 Vha), where mean annual precipitation and temperature are 4,320 mm and
26.5"C, respectively. The low biomass of the mangrove forest in Ishigaki Island - Okinawa
and Southem Florida might be attributable to the lower mean annual temperature than tropi-
cal areas, although the precipitation in those areas is no less than the areas in the tropic
region. On the other hand, although the mean annual temperature in Talidendang Besar and
Puerto Rico is comparable to that of the other tropical areas, the lower annual precipitation is
regarded as an important climatic factor causing a low biomass of the mangrove forest in
those areas. The similarresults were also reported by Komiyamaet al. (1988) forthe above
ground biomass of Rhizophor4 spp. all over the world. Consequently, the temperature and
precipitation are supposed to be the important climatic factors causing the difference of the
mangrove biomass all over the world, which agrees with Satoo and Madgwick (1982): They
stated that differences among climatic gradients reflect different rates of organic matter
production.
ACKNOWLEDGMENTS We wish to thank Mr. Subkhan and Mr. Elizar for helping the
data collection in the field; the staffs of PT Bina Lestari Riau who donated the facilities
during the survey; and Dr. Dudung Darusman, the Dean of Faculty of Foresury - Bogor Agri-
cultural University, for his help to iurange admission letters. We also wish to express our
=3z9F
Fo-
6UJEGJJzz

Above ground tree biomass of a mangrove forest in East Sumatra 255
thanks to Dr. Akira Komiyama, Faculty of Agriculture - Gifu University for his valuable
comments to prepare the manuscript; Dr. Isamu Yamada, The Center for Southeast Asian
Studies of Kyoto University; Dr. Shinya Takeda, Faculty of Agriculture, Kyoto University;
Mr. Sukristijono Sukardjo, The Center for Oceanological Research of Indonesia; Prof. Ishe
mat Soerianegara and Ir. Evrizal Amzu, MS., Faculty of Forestry, Bogor Agricultural Univer
sity, for their encouragement throughout this research.
REFERENCES
Briggs, S. V. 1977. Estimates of biomass in a temperate mangrove community. 1. Austral.
Ecol., 2: 369-373.
Cannell, M. G. R. 1982. Wor:'d Forest Biomass and Primary Production Data. Academic
Press, London. 391 pp.
Christensen, B. 1978. Biomass and primary production of Rhizophora apiculata B1. in a mangrove in Southern Thailand. Aquatic Botany, 4: 43-52.
--. 1982. Management and Utilization of Mangroves in Asia and the Pacific. FAO Environ
ment Paper 3. FAO, Rome.
Cintron, G. & Novelli, Y. S. 1984. Methods for studying mangrove structure. In: Snedaker,
S. C. and Snedaker, J. G. (eds.), The Mangrove Ecosystem: Research Methods, 91-113.
UNESCO, Paris.
Direktur Jenderal Kehutanan. 1978. Surat keputusan No. 601KptslDjlI/1978 tentang pedoman
sistem silvikultur untuk hutan mangrove. Direktur Jenderal Kehutanan, Departemen Perta
nian Republik Indonesia, Jakarta. 5 pp.
Golley, F. B., Odum, H. T. & Wilson, R. F. 1962. The structure and metabolism of a Puerto
Rican red mangrove forest in May. Ecology, 43: 9-19.
--, McGinnis, J. T., Clements, R. G., Child, G. 1. & Duever, M. J. 1969. The structure of
tropical forests in Panama and Colombia. BioScience, 19 (8): 693-696.
Hase, H., Foelster & Lindheim, M. 1985. On the accuracy of estimating aboveground tree
biomass in an evergreen forest near Manaus, Brazil. A simulation study. Biotropica, 17 (3): 191-195.
Komiyama, A., Moriya, H., Prawiroatmodjo, S., Toma, T. & Ogino, K. 1988. Forest as an
ecosystem, its structure and function: 2. Primary productivity of mangrove forest. In:
Ogino, K. & Chihara, M. (eds.), Biological System of Mangroves: A Report of East
Indonesian Mangrove Expedition 1986,97-106. Ehime University, Japan.
Lembaga Penelitian Tanah. 1964. Peta Tanah Eksplorasi Sumatra Bagian Selatan, Skala 1 :
I 000000. Lembaga Penelitian Tanah, Bogor.
Loetsch, E, Zohrer, E & Haller, K. E. 1973. Forest Inventory. BLV Verlagsgesellschaft mbH,
Mtinchen.
Lugo, A. E. & Snedaker, S. C. 1974. The ecology of mangroves. Ann. Rev. Ecol. Syst., 5: 39-
65.
Macnae, W. 1968. A general account of the fauna and flora of mangrove swamps and forests
in the Indowest Pacific region. Adv. Mar. Bioi., 6: 73-270. Nakasuga, T. 1979. Analysis of stand structure of mangroves. Bull. Ryukyu Univ., 26: 413-
519. (in Japanese with English summary) Ninomiya, 1., Komiyama, A., Moriya, H., Tadaki, Y. & Ogino, K. 1989. Production structure

256 C. KUSMANA, S. SABIHAM, K. ABE & H, WATANABE
of Rhizophora stylosa and Bruguiera gymnorrhiza in Funaura, Iriomote. Galaxea, 8: 65-
68.
Ong, J., Wooi-Khoon, G., Chee-Hoong, W. & Dhanarajan, G. 1984. Contribution of aquatic
productivity in managed mangrove ecosystem in Malaysia. In: Soepadmo, Rao, A. N. &
MacIntosh, D. J. (eds.), Proc. Asian Symp. Mangrove Environment: Research and
Management, 209-215. UNESCO, Paris.
Rodin, L. E. & Bazilevich, N. 1. 1967. Production and Mineral Cycling in Terrestrial Vegeta
tion. Oliver & Boyd, London. 288 pp.
Satoo, T. & Madgwick, H. A. I. 1982. Forest Biomass. W. Junk Publishers, The Hague. 152
pp.
Schmidt, F. H. & Ferguson, J. H. A. 1951. Rainfall type based on wet and dry period ratios of
Indonesian with western New Guinea. Kementrian Perhubungan, Djawatan Meteorologi
dan Geojisika, Verhandelingen no. 42. 77 pp.
Soegiarto, A. 1984. The mangrove ecosystem in Indonesia, its problems and management. In:
Teas, H.J. (ed.), Physiology and Management of Mangroves, 69-78. W. Junk Publishers,
The Hague.
Soemodihardjo, S. & Soerianegara, I. 1989. The status of mangrove forests in Indonesia. In : Soerianegara, I., Zamora, P. M., Kartawinata, K., Umaly, R. c., Tjitrosomo, S., Sitompul,
D. M. & Rosalina, U. (eds.), Mangrove Management: Its Ecological and Economics
Considerations. Biotrop Special Publication, 37: 73-114.
Sukardjo, S. 1987. Natural regeneration status of commercial mangrove species (Rhizophora
apiculata and Bruguiera gymnorrhiza) in the mangrove forest of Tanjung Bungin,
Banyuasin District, South Sumatra. Forest Ecology and Management, 20: 233-252.
Suzuki, E. & Tagawa, H. 1983. Biomass of a mangrove forest and a sedge marsh on Ishigaki
island, South Japan. Jpn. 1. Ecol., 33: 231-234.
Tarnai, S., Nakasuga, T., Tabuchi, R. & Ogino, K. 1986. Standing biomass of mangrove
forests in Southern Thailand. 1. Jpn. For. Soc., 68 (9): 384-388.
C. KUSMANA, S. SABIHAM, ~iiJgB~-, )JtilliJJ...iZ - -:;r~*0) J{tt.c gB!~ff;' O)mtE:
Received Jan. 20, 1992
Accepted April 29, 1992
1 /' F ;jv/ 7, -*.1. --.( r '7 . I) 7'/ j'I'1 Talidendang Besar 0)""'7 /':7" 0 - 7·*'±, ?iD:J¥$ip G F9~ ff~~=[f:l]i)'"?-C, Bruguiera parviflora ;!'1::::)v,:f, B. sexangula ;;f;\j-1::::)v,:f, -f-l-C;;f;\j-1::::)v'f'
~ -=t ~= l -C Nipa fruticans ::. 'J /'I.y "/ ~ Rhizophora apiculata 7)' ;\j- 1:::: )v~'O) i t, .Qff~f:H=~
itT .Q 0 :.. 0) --.( /':7' 0 - 7"1f.f0)iJ&I1: Ij:t~;.fll( . flJHJi* ~ 9;0.Q t.: 66, :.. n G 3 ff~O):tt!i...tg:~.lJ.1f:l:O) :f1EJE ~1l' Ij:"? t.:o
50 m X 50 m 0) :J 1·0" '7 - r ~ B. parviflora ,jJ:'F:!.R~= 2.;rp}j·, B. sexangula ff~I= 3 .;rPJT, -t- l-C B.
sexangula - N. fruticans - R. apiculata ~nw:O) R, apiculata O)116T.Q t :.. 0 1= 1 .;rPJTliltlE l, }ffij)~
j][1:£1O(.mJJLO)tO)~1lJ>t~~Jnl1tlt.:iPJt, -tj-1;;:(0)~1j:.Q2Vf~~ft1fUl, -f-n~~, -fx,~,
;fE· *jf, ;Dtli'UIj: t"~=))-f:T, lli:tlijlIJ!E~rj'lj:It" 7 D;!. r 1) - (1§HMffJ:t) ~*66, :tt!i...tg:~.lJ.
H:ill: ~:f1~1E l t.:o

Above ground trl∞ biomass of a mangrove forest in East Sumatra 257
Bruguiera parviflora 群集の立木密度は 156 -400 本lha , B. A (胸高断面積合計)は 4.0 -9.2
m2 /ha , B. sexangurl日 1洋集で 188 -348 4:/ha , 5.0 -22.1 m 2/ha , R. apiculata 群集で 32 本, 2.
5 m2/ha と,かなり疎な林分であった。
推定される地上部現存量はB. parviflora 群集で 42.9 -90.0 t/ha , B. sexangura 群集で 76.0-
279.0 t/ha , R. apiculata 群集ではわずか 40.7 t/ha であった。これらの値は,これまで東南アジ
ア各地のマングローブ林で推定された値と比較して,いずれもかなり低いものであった。これは
年降水量が 1 ,3α)mm と少ないことが影響しているようであった。
東スマトラではまだ広大なマングロープ林が存在するが,その現存量。材責)はかなり低いもの
と考えられ,コンセッションでのマングロープの利用・管理計画の設定に当たっては,この事実
を考慮する必要ーがあろう。


















