
Activation of Adenosine Monophosphate–activated...
16
Copyright © 2015, the American Society of Anesthesiologists, Inc. Wolters Kluwer Health, Inc. Unauthorized reproduction of this article is prohibited. Anesthesiology, V 123 • No 5 1170 November 2015 P AIN is a major clinical problem for cancer patients, experienced by more than 64% of those with advanced stage or metastatic cancer. 1 e most frequent pain in patients with metastatic breast, prostate, or lung cancer is bone can- cer pain, which is particularly severe, difficult to treat, and insufficiently relieved. Cancer cells metastasized to the bone generate pain by releasing algogenic substances, which can induce sensitization and activation of nerve fibers that inner- vate the bone. 2 Tumor growth in the bone can also generate a neuropathic pain by directly injuring nerve fibers and induc- ing an active and highly pathologic sprouting of both sensory and sympathetic nerve fibers, which seem to contribute to What We Already Know about This Topic • Robust activation of microglia and astrocytes in the spinal cord plays a pivotal role in the pathology of cancer-related bone pain • The suppression of glial activation may be a viable option for cancer pain management What This Article Tells Us That Is New • Activation of adenosine monophosphate–activated kinase by resveratrol significantly inhibited spinal neuroinflammation in a rat model of bone cancer pain • Resveratrol also reduced pain-related behaviors in these animals Copyright © 2015, the American Society of Anesthesiologists, Inc. Wolters Kluwer Health, Inc. All Rights Reserved. Anesthesiology 2015; 123:1170-85 ABSTRACT Background: Activation of adenosine monophosphate–activated kinase (AMPK) has been associated with the inhibition of inflammatory nociception and the attenuation of morphine antinociceptive tolerance. In this study, the authors investigated the impact of AMPK activation through resveratrol treatment on bone cancer pain. Methods: e nociception was assessed by measuring the incidence of foot withdrawal in response to mechanical indentation in rats (n = 8). Cytokine expression was measured using quantitative polymerase chain reaction (n = 8). Cell signalings were assayed by western blot (n = 4) and immunohistochemistry (n = 5). e microglial cell line BV-2, primary astrocytes, and neuron-like SH-SY5Y cells were cultured to investigate the in vitro effects. Results: Resveratrol and 5-amino-1-β-D-ribofuranosyl-imidazole-4-carboxamide, the AMPK activators, significantly attenu- ated bone cancer pain in rats with tumor cell implantation (TCI; threshold of mechanical withdrawal, resveratrol vs. vehicle: 10.1 ± 0.56 vs. 4.1 ± 0.37; 5-amino-1-β-D-ribofuranosyl-imidazole-4-carboxamide vs. vehicle: 8.2 ± 0.17 vs. 4.1 ± 0.37, mean ± SEM); these effects were reversed by the AMPK inhibitor compound C (compound C vs. resveratrol: 6.2 ± 1.35 vs. 10.1 ± 0.56, mean ± SEM). Resveratrol has an AMPK-dependent inhibitory effect on TCI-evoked astrocyte and microglial activation. e antinociceptive effects of resveratrol were partially mediated by the reduced phosphorylation of mitogen-activated protein kinases and decreased production of proinflammatory cytokines in an AMPK-dependent manner. Furthermore, resveratrol potently inhibited inflammatory factors–mediated protein kinase B/mammalian target of rapamycin signaling in neurons. Acute pain evoked by proinflammatory cytokines in the spinal cord was significantly attenuated by resveratrol. Conclusions: AMPK activation in the spinal glia by resveratrol may have utility in the treatment of TCI-induced neuroinflam- mation, and our results further implicate AMPK as a novel target for the attenuation of bone cancer pain. (A NESTHESIOLOGY 2015; 123:1170-85) The first three authors contributed equally to this work. Submitted for publication August 5, 2014. Accepted for publication July 14, 2015. From the Department of Pharmacology, Jiangsu Key Laboratory of Neurodegeneration, Nanjing Medical University, Jiangsu, China (H.S., Y.H., C.P., X.D., W.D., L.H., C.J., Y.Y., W.L.); State Key Laboratory of Pharmaceutical Biotechnology, School of Life Sciences, Nanjing University, Jiangsu, China (H.S., X.W.); Jiangsu Province Key Laboratory of Anesthesiology, School of Anesthesiology, Xuzhou Medical College, Xuzhou, Jiangsu, China (Y.H.); Department of Pain Management, Cancer Biotherapy Center, The Second Affiliated Hospital of Nanjing Medical University, Jiangsu, China (Z.C.); Department of Medicinal Chemistry, School of Pharmacy, Nanjing Medical University, Jiangsu, China (F.L.); and Research Division of Pharmacology, China Pharmaceutical University, Nanjing, China (X.D., W.D., G.Z.). Activation of Adenosine Monophosphate–activated Protein Kinase Suppresses Neuroinflammation and Ameliorates Bone Cancer Pain Involvement of Inhibition on Mitogen-activated Protein Kinase Huayuan Song, M.D., Yuan Han, M.D., Ph.D., Cailong Pan, B.S., Xueting Deng, M.S., Wenling Dai, B.S., Liang Hu, Ph.D., Chunyi Jiang, Ph.D., Yanjing Yang, M.D., Zhixiang Cheng, M.D., Ph.D., Fei Li, Ph.D., Guangqin Zhang, Ph.D., Xuefeng Wu, Ph.D., Wentao Liu, Ph.D. Downloaded From: http://anesthesiology.pubs.asahq.org/pdfaccess.ashx?url=/data/journals/jasa/934601/ on 09/16/2018
Transcript of Activation of Adenosine Monophosphate–activated...

Copyright © 2015, the American Society of Anesthesiologists, Inc. Wolters Kluwer Health, Inc. Unauthorized reproduction of this article is prohibited.
Anesthesiology, V 123 • No 5 1170 November 2015
P AIN is a major clinical problem for cancer patients, experienced by more than 64% of those with advanced
stage or metastatic cancer.1 The most frequent pain in patients with metastatic breast, prostate, or lung cancer is bone can-cer pain, which is particularly severe, difficult to treat, and insufficiently relieved. Cancer cells metastasized to the bone generate pain by releasing algogenic substances, which can induce sensitization and activation of nerve fibers that inner-vate the bone.2 Tumor growth in the bone can also generate a neuropathic pain by directly injuring nerve fibers and induc-ing an active and highly pathologic sprouting of both sensory and sympathetic nerve fibers, which seem to contribute to
What We Already Know about This Topic
• Robust activation of microglia and astrocytes in the spinalcord plays a pivotal role in the pathology of cancer-relatedbonepain
• Thesuppressionofglialactivationmaybeaviableoptionforcancerpainmanagement
What This Article Tells Us That Is New
• Activationofadenosinemonophosphate–activatedkinasebyresveratrolsignificantlyinhibitedspinalneuroinflammationinaratmodelofbonecancerpain
• Resveratrolalsoreducedpain-relatedbehaviorsintheseanimals
Copyright © 2015, the American Society of Anesthesiologists, Inc. Wolters Kluwer Health, Inc. All Rights Reserved. Anesthesiology 2015; 123:1170-85
ABSTRACT
Background: Activation of adenosine monophosphate–activated kinase (AMPK) has been associated with the inhibition of inflammatory nociception and the attenuation of morphine antinociceptive tolerance. In this study, the authors investigated the impact of AMPK activation through resveratrol treatment on bone cancer pain.Methods: The nociception was assessed by measuring the incidence of foot withdrawal in response to mechanical indentation in rats (n = 8). Cytokine expression was measured using quantitative polymerase chain reaction (n = 8). Cell signalings were assayed by western blot (n = 4) and immunohistochemistry (n = 5). The microglial cell line BV-2, primary astrocytes, and neuron-like SH-SY5Y cells were cultured to investigate the in vitro effects.Results: Resveratrol and 5-amino-1-β-d-ribofuranosyl-imidazole-4-carboxamide, the AMPK activators, significantly attenu-ated bone cancer pain in rats with tumor cell implantation (TCI; threshold of mechanical withdrawal, resveratrol vs. vehicle: 10.1 ± 0.56 vs. 4.1 ± 0.37; 5-amino-1-β-d-ribofuranosyl-imidazole-4-carboxamide vs. vehicle: 8.2 ± 0.17 vs. 4.1 ± 0.37, mean ± SEM); these effects were reversed by the AMPK inhibitor compound C (compound C vs. resveratrol: 6.2 ± 1.35 vs. 10.1 ± 0.56, mean ± SEM). Resveratrol has an AMPK-dependent inhibitory effect on TCI-evoked astrocyte and microglial activation. The antinociceptive effects of resveratrol were partially mediated by the reduced phosphorylation of mitogen-activated protein kinases and decreased production of proinflammatory cytokines in an AMPK-dependent manner. Furthermore, resveratrol potently inhibited inflammatory factors–mediated protein kinase B/mammalian target of rapamycin signaling in neurons. Acute pain evoked by proinflammatory cytokines in the spinal cord was significantly attenuated by resveratrol.Conclusions: AMPK activation in the spinal glia by resveratrol may have utility in the treatment of TCI-induced neuroinflam-mation, and our results further implicate AMPK as a novel target for the attenuation of bone cancer pain. (A nesthesiology 2015; 123:1170-85)
The first three authors contributed equally to this work.
Submitted for publication August 5, 2014. Accepted for publication July 14, 2015. From the Department of Pharmacology, Jiangsu Key Laboratory of Neurodegeneration, Nanjing Medical University, Jiangsu, China (H.S., Y.H., C.P., X.D., W.D., L.H., C.J., Y.Y., W.L.); State Key Laboratory of Pharmaceutical Biotechnology, School of Life Sciences, Nanjing University, Jiangsu, China (H.S., X.W.); Jiangsu Province Key Laboratory of Anesthesiology, School of Anesthesiology, Xuzhou Medical College, Xuzhou, Jiangsu, China (Y.H.); Department of Pain Management, Cancer Biotherapy Center, The Second Affiliated Hospital of Nanjing Medical University, Jiangsu, China (Z.C.); Department of Medicinal Chemistry, School of Pharmacy, Nanjing Medical University, Jiangsu, China (F.L.); and Research Division of Pharmacology, China Pharmaceutical University, Nanjing, China (X.D., W.D., G.Z.).
Activation of Adenosine Monophosphate–activated Protein Kinase Suppresses Neuroinflammation and Ameliorates Bone Cancer Pain
Involvement of Inhibition on Mitogen-activated Protein Kinase
HuayuanSong,M.D.,YuanHan,M.D.,Ph.D.,CailongPan,B.S.,XuetingDeng,M.S.,WenlingDai,B.S.,LiangHu,Ph.D.,ChunyiJiang,Ph.D.,YanjingYang,M.D.,ZhixiangCheng,M.D.,Ph.D.,FeiLi,Ph.D.,GuangqinZhang,Ph.D.,XuefengWu,Ph.D.,WentaoLiu,Ph.D.
Downloaded From: http://anesthesiology.pubs.asahq.org/pdfaccess.ashx?url=/data/journals/jasa/934601/ on 09/16/2018

Copyright © 2015, the American Society of Anesthesiologists, Inc. Wolters Kluwer Health, Inc. Unauthorized reproduction of this article is prohibited.
Anesthesiology 2015; 123:1170-85 1171 Song et al.
PAIN MEDICINE
the peripheral and central sensitization that is common in advanced bone cancer pain.3 Specifically, mounting evidence has indicated that spinal glial activation is strongly involved in the pathology of central sensitization.4,5 Bone cancer pain is linked to the markedly increased expression of glial fibril-lary acidic protein (GFAP) in astrocytes and ionized calcium-binding adapter molecule 1 (IBA-1) in microglia within the dorsal horn (dH) of the spinal cord.6,7 The glial P2X7 recep-tor and chemokines CCL2, CXCL10, and CXCL12 are up-regulated during the induction of bone cancer pain.8–12 As such, the regulation of glial-mediated inflammation may be a useful strategy for the treatment of bone cancer pain.
Adenosine monophosphate (AMP)–activated kinase (AMPK) is primarily associated with cholesterol bio-synthesis as an important regulator of metabolism.13 It has also been recognized for its antiinflammatory effects through the decreased expression of tumor necrosis factor (TNF)-α, interleukin (IL)-1β, and IL-6 induced by mor-phine or lipopolysaccharide in microglia in the central ner-vous system.14,15 In addition, the AMPK activation through 5-amino-1-β-d-ribofuranosyl-imidazole-4-carboxamide (AICAR) may reduce IL-1β–induced IL-6 and IL-8 levels in the adipose tissue of patients and in the skeletal muscles of rats.16 Taken together, these results suggest that AMPK activation may significantly suppress neuroinflammation in the central nervous system.
A possible function of AMPK in pain-related pathways has been revealed in the model of incision-induced acute and chronic pain after treatment with the AMPK activa-tors resveratrol, metformin, or A769662.17–19 Knockout mice lacking the AMPKα2 subunit shows increased noci-ceptive responses that could not be reversed by AMPK activators.20 Based on these results, it seems likely that AMPK would also be involved in other clinically relevant pain states, such as that caused by bone cancer. Ques-tions remain regarding the specific role of AMPK in can-cer pain pathways and optimal regulation of AMPK in a clinically effective and safe way. Herein, we make the hypothesis that AMPK activation may inhibit glial activa-tion, thereby inhibiting neuroinflammation and suppress-ing cancer pain. In this study, we evaluated the effects of resveratrol, a natural AMPK activator widely found in red grapes and wine, in a model of tibia bone cavity tumor cell implantation (TCI) in rats.
Materials and Methods
Ethics StatementAll procedures were strictly performed in accordance with the regulations of the ethics committee of the International Association for the Study of Pain and the Guide for the Care and Use of Laboratory Animals (The Ministry of Science and Technology of China, 2006). All animal experiments were approved by Nanjing Medical University Animal Care and Use Committee and were designed to minimize suffering and the number of animals used.
AnimalsSprague-dawley female rats (weight, 150 to 170 g used in TCI operation or 60 to 80 g used in the expansion of Walker 256 ascites tumor cells) were provided by the Experimental Animal Center at Nanjing Medical University, Nanjing, China. Five to six animals were housed per cage under pathogen-free con-ditions with soft bedding under controlled temperature (22° ± 2°C) and photoperiods (12:12-h light–dark cycle). They were allowed to acclimate to these conditions for at least 2 days before inclusion in experiments. Animals were randomly divided into groups (n = 8). The sample size was designed based on the previous experience21 and to be limited to the minimal as scientifically justified. For each group of experi-ments, the animals were matched by age and body weight. All surgeries were performed under anesthesia with pentobarbital (Sigma, USA, 50 mg/kg, intraperitoneally [IP]).
Drugs and ReagentsFetal bovine serum (FBS) and other cell culture media and supplements were purchased from Hyclone (USA). AICAR, compound C, and resveratrol were purchased from Sigma. Antibody for glyceraldehyde 3-phosphate dehydrogenase was purchased from Sigma. Antibody for phospho-p38 (Tyr182) was from Epitomics (USA). Antibody for IBA-1 was from Wako Pure Chemical Industries (Japan). Anti-body for GFAP and neuronal nuclear protein (NeuN) was from Millipore (USA). Antibodies for IL-1β, TNF-α were from Santa Cruz Biotechnology (USA). Antibody for Ki67 was from Abcam (UK). Antibodies for phosphorylated N-methyl-d-aspartate receptor 1 (NR1) subunit (Ser896), phosphorylated protein kinase C (PKC)γ; Thr514), phos-phorylated extracellular regulated protein kinase (ERK; Thr202/Tyr204), phosphorylated AMP–activated protein kinase (AMPK; Thr172), phosphorylated c-Jun N-terminal kinase (JNK; Thr183/Tyr185), phosphorylated protein kinase B (Akt; Ser 473), and phosphorylated mammalian target of rapamycin (mTOR; Ser 2448) were from Cell Sig-naling Technology (USA). Secondary antibodies were pur-chased from Chemicon (USA). All other chemicals were purchased from Sigma Chemical Co.
Model of Bone Cancer PainSuspension of Walker 256 ascites tumor cells (5 × 106 cells/ml, 0.5 ml) was inoculated into rats IP. Seven days after inoculation, single-cell suspensions were made from the ascites. Tumor-bearing animals were generated as published previously.22 Briefly, rats were anesthetized with sodium pentobarbital (50 mg/kg, IP), and a left knee arthrotomy was performed. Tumor cells (1 × 105 cells/µl) in 5 μl of ster-ile phosphate-buffered saline (PBS) solution were injected directly into the medullary cavity of the distal femur. Addi-tional controls are naive rats with 5 μl of PBS injected into the medullary cavity. The injection hole was plugged with a dental amalgam that tightly seals the injection hole in the bone and prevents the escape of tumor cells from the bone.
Downloaded From: http://anesthesiology.pubs.asahq.org/pdfaccess.ashx?url=/data/journals/jasa/934601/ on 09/16/2018

Copyright © 2015, the American Society of Anesthesiologists, Inc. Wolters Kluwer Health, Inc. Unauthorized reproduction of this article is prohibited.
Anesthesiology 2015; 123:1170-85 1172 Song et al.
AMPK Activation Ameliorates Bone Cancer Pain
Assessment of Bone Cancer–related Pain BehaviorsMechanical allodynia was determined by measuring the inci-dence of foot withdrawal in response to mechanical indenta-tion of the plantar surface of each hind paw with a sharp, cylindrical probe with a uniform tip diameter of approxi-mately 0.2 mm provided by an Electro Von Frey (ALMEMO 2390–5, Anesthesiometer IITC Inc., USA), using a protocol similar to the one we have described.5 The probe was applied to 10 designated loci distributed over the plantar surface of the foot. The minimal force (in grams) that induced paw withdrawal was read off of the display. Threshold of mechan-ical withdrawal in each rat was calculated by averaging the 10 readings, and the force was converted into millinewtons. For the results expressing mechanical allodynia, the values are mean values of ipsilateral feet. The behavioral tests were carried out blind to the treatment condition. The rats were tested on each of 3 successive days before surgery. To exam-ine the immediate effect of drugs and vehicles on the persis-tence of tumor pain, tests were conducted on postoperative days 14 to 18 (2 h after drugs given), and additional tests were conducted at 0, 2, 4, 6, 8, and 24 h after a single injec-tion on the 14th postoperative day. To examine the effect of drugs on IL-1β/TNF-α–induced pain, tests were conducted on 0 to 8 h after the injection of cytokines. drugs were given just before cytokines injection.
Intrathecal Injection ProcedureTo perform intrathecal (IT) injections, the rat was placed in a prone position and the midpoint between the tips of the iliac crest was located. A Hamilton syringe with 30-gauge needle was inserted into the subarachnoid space of the spi-nal cord between the L5 and L6 spinous processes. Proper IT injection was systemically confirmed by observation of a tail flick. IT injection did not affect baseline responses, com-pared with latencies recorded before injection.
Radiologic Bone ExaminationTo determine the tibial destruction from the inoculated tumor, rats were radiographed at day 18 after tumor cell inoculation. Rats were placed on clear plexiglass and were exposed to an x-ray source under sodium pentobarbital anes-thesia (50 mg/kg, IP). Using a Kodak digital Radiographer System (Kodak, USA), tibial radiographs were taken (40 KVP, 2 ma) from hind limbs (n = 4/group).
Histochemical StainingAt day 18 after tumor cell inoculation, animals were killed. After demineralization in EdTA (10%) for 2 to 3 weeks, the tibiae were embedded in paraffin and 5-μm thick sections were cut using a microtome. The sections were stained with Harris hematoxylin and eosin to verify cancer cell infiltra-tion and bone destruction (n = 4/group). The macroscopic examination was blindly carried out by two independent observers. For standardization, the tibial bones were pre-sented in the same section for analysis. Tumor infiltration
was scored 0 to 4 according to the following criteria: 0 = normal, 1 = minimal infiltration (not readily apparent on low magnification), 2 = mild infiltration, 3 = moderate infil-tration with cell aggregates, and 4 = marked infiltration with large aggregates. Bone destruction was scored 0 to 4 accord-ing to the following criteria: 0 = none, 1 = minimal (1 to 2 sites of cartilage loss, visible only at high magnification), 2 = mild (at least three sites of resorption, visible only at high magnification), 3 = moderate (obvious foci of bone resorp-tion, visible at low power), and 4 = marked (large erosions extending through to the marrow space).
Assessment of Motor FunctionMotor function was measured using a rotarod apparatus for rats (Shandong Science Equipment Co., Ltd., China) under different rotor mode (16, 24, 32 rpm). The inter-val from when the animal mounted the rod to when it fell off was recorded as the performance time. The animals were trained for three trials per day for 3 days before drug administration, and the mean duration on the rod was recorded to obtain stable baseline values. Performance on the rotarod test was measured 30 min after each adminis-tration of resveratrol.
Cell CulturesMicroglial BV-2 cells or human neuron-like SH-SY5Y cells were incubated under humidified 5% CO2 and 95% O2 at 37°C in dulbecco’s modified Eagle’s medium (dMEM, Invitrogen, USA) containing 10% FBS and 1% streptomy-cin and penicillin (Invitrogen). Twenty-four hours before experimentation, the culture media was replaced by 0.5% FBS high-glucose dMEM. Then the cells were stimulated with IL-1β (1 ng/ml) and TNF-α (50 ng/ml) for 30 min with or without resveratrol (125 μM), compound C (20 μM), and AICAR (20 μM).
Glia were isolated from the brains of postnatal day 2 pups as described earlier.23,24 In brief, cerebral cortices were removed from the rat brains for glial cultures. The tissue was dissociated in 0.0025% trypsin/EdTA and passed through a 70-µm pore nylon mesh. After centrifugation, the cell pel-let was resuspended in dMEM containing 10% FBS, 50 U/ml penicillin and 50 mg/ml streptomycin. The cell culture dishes were coated with 10 μg/ml type I collagen (Bd Bio-sciences, USA) for 12 h at 37°C. Excess collagen was removed by PBS solution at 37°C. Then the glial cells in dMEM con-taining 10% FBS were loaded into the dishes. After 7 to 10 days, glial cells typically reached 80 confluence and were ready for the experiment. In general, the cultures consisted of more than 95% astrocytes (as indicated by GFAP-positive staining). Cell proliferation was evaluated with a modified Ki67 assay. The cells were activated with IL-1β (1 ng/ml) and TNF-α (50 ng/ml) for 24 h with resveratrol (125 μM), com-pound C (20 μM), or AICAR (20 μM). As for the measure-ment of proteins in astrocytes, cells were collected 30 min after stimulation.
Downloaded From: http://anesthesiology.pubs.asahq.org/pdfaccess.ashx?url=/data/journals/jasa/934601/ on 09/16/2018

Copyright © 2015, the American Society of Anesthesiologists, Inc. Wolters Kluwer Health, Inc. Unauthorized reproduction of this article is prohibited.
Anesthesiology 2015; 123:1170-85 1173 Song et al.
PAIN MEDICINE
Western BlotTo identify temporal expression or the phosphorylated lev-els of proteins, whole protein samples were analyzed. In brief, samples (cells or spinal cord segments at L1 to L6) were collected and washed with ice-cold PBS before being lysed in radio immunoprecipitation assay lysis buffer. Then whole sample lysates were separated by sodium dodecyl sul-fate polyacrylamide gel electrophoresis and electrophoreti-cally transferred onto polyvinylidene fluoride membranes (Millipore). The membranes were blocked with 5% bovine serum albumin for 1 h at room temperature, probed with antibodies overnight at 4°C with the primary antibodies and then incubated with horseradish peroxidase–coupled secondary antibodies. The primary antibodies used included p-p38 (Tyr182), 1:800; IBA-1, 1:1000; p-NR1 (Ser896), 1:1000; pERK1/2 (Thr202/Tyr204), 1:1000; p-JNK (JNK; Thr183/Tyr185), 1:1000; glyceraldehyde 3-phosphate dehy-drogenase, 1:5000; p-PKC (Thr514), 1:1000; p-Akt (Ser 473), 1:800; p-mTOR (Ser 2448), 1:800; GFAP, 1:2000; IL-1β, 1:1000; and TNF-α, 1:1000. The filters were then developed by enhanced chemiluminescence reagents (Perki-nElmer, Waltham, MA) with secondary antibodies (Chemi-con, USA). data were analyzed with the Molecular Imager (Gel docTM XR, 170–8170) and the associated software Quantity One-4.6.5 (Bio-Rad Laboratories, USA).
Immunohistochemistryduring deep anesthesia, animals were transcardially perfused with PBS followed by 4% paraformaldehyde, and L4 and/or L5 lumbar segment was dissected out and postfixed using the same fixative. The embedded blocks were sectioned as 20-μm thick. Sections from each group (five samples in each group) were incubated overnight at 4°C with primary anti-bodies: anti-GFAP antibody (1:100), anti-IBA-1 (1:100), anti-NeuN (1:100), anti-pAMPK (1:100), anti-pp38 (1:100), anti-pJNK (1:50), or anti-pERK (1:500). Rab-bit IgG (1:200, Vector Laboratories, USA) was used as an isotype control. Then the free-floating sections were washed with PBS and incubated with the secondary antibody for 2 h. After washing out three times with PBS, the samples were studied under a confocal microscope (Olympus FV1000 confocal system, Olympus, Japan) for morphologic details of the immunofluorescence staining. Examination was blindly carried out. Images were randomly coded, and the fluores-cence intensities of IBA-1 or GFAP were analyzed by Image Pro Plus 6.0 software (Media Cybernetics Inc., USA). The average green fluorescence intensity of each pixel was nor-malized to the background intensity in the same image.
Glial cells were fixed with ice-cold methanol and were permeabilized with 0.25% Triton X-100/phosphate-buff-ered saline with Tween 20. After blocking with 1% bovine serum albumin in phosphate-buffered saline with Tween 20 for 120 min, the coverslips were incubated overnight at 4°C with the GFAP and Ki67 antibodies diluted in 1% bovine serum albumin (1:300). The coverslips were then exposed
to the fluorescein isothiocyanate (FITC)-conjugated anti-rabbit IgG (1:100, at room temperature for 60 min) after being washed three times with PBS. After that the coverslips were stained with 1 μg/ml 4′,6-diamidino-2-phenylindole), a fluorescent dNA dye to mark nucleus) for 1 min, and then fluorescent confocal images were acquired using a Olympus FV1000 confocal system (Olympus).25 Ki67-positive astro-cytes were counted in a blind fashion.
Statistical AnalysesSPSS Rel 15 (SPSS Inc., Chicago, IL) was used to conduct all the statistical analyses. Alteration of expression of the pro-teins detected and the behavioral responses were tested with one-way ANOVA, and the differences in latency over time among groups were tested with two-way ANOVA. Bonfer-roni post hoc tests were conducted for all ANOVA models. Results are expressed as mean ± SEM of three independent experiments. Results described as significant are based on a criterion of P < 0.05.
Results
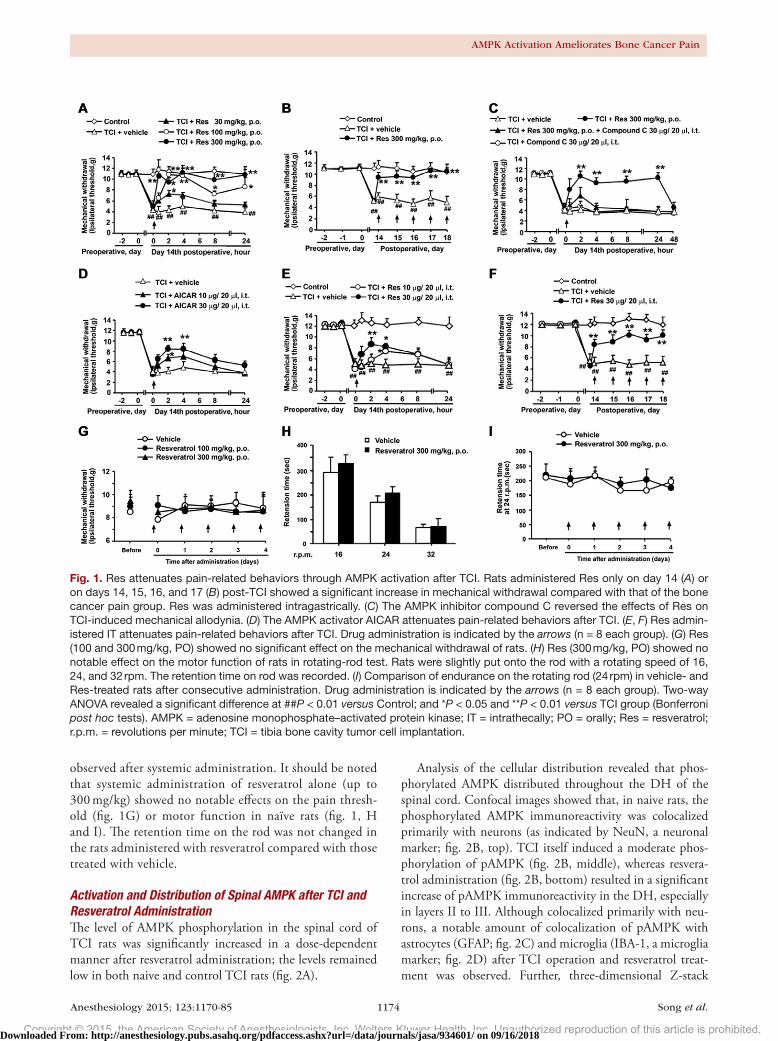
AMPK Activation by Resveratrol Attenuates Mechanical Allodynia in a Rat Model of Bone Cancer PainRats developed tibia tumors after the transplantation of Walker 256 cells and subsequently displayed mechanical allodynia (fig. 1). Pretransplant, there were no significant differences in pain-related behaviors among the sham group and the other groups. A marked decrease in mechanical withdrawal was observed 7 days after TCI followed by a peak on day 14 (4.1 ± 0.37, mean ± SEM). Thus, resveratrol was administered orally (PO) to the TCI rats on day 14. Pain-related behaviors were greatly ameliorated by resveratrol (300 mg/kg, PO, 10.1 ± 0.56), and this effect lasted for more than 24 h (fig. 1A). In addition, consecutive administration of resveratrol at postoperative days 14, 15, 16, 17, and 18 reversed the mechanical allodynia in TCI rats (fig. 1B).
Resveratrol has been reported to inhibit neuroinflam-mation through AMPK activation.15 To explore this puta-tive role of AMPK in resveratrol-mediated inhibition of tumor pain, we questioned whether the AMPK inhibitor compound C could reverse the effects of resveratrol. Our results revealed that compound C (30 μg, IT) significantly inhibited resveratrol-induced alleviation of mechanical allodynia (6.2 ± 1.35), as assessed on postoperative day 14 (fig. 1C). Moreover, the AMPK activator AICAR mim-icked the ameliorative effects of resveratrol on tumor pain (8.2 ± 0.17; fig. 1d).
To provide additional evidence and confirmation, res-veratrol was administered IT to the TCI rats on days 14, 15, 16, 17, and 18. The mechanical allodynia were nota-bly ameliorated by a single dose of resveratrol (fig. 1E) or consecutive administration (fig. 1F). These data par-tially address that local administration targeting spinal cord mechanisms would induce similar effects to those
Downloaded From: http://anesthesiology.pubs.asahq.org/pdfaccess.ashx?url=/data/journals/jasa/934601/ on 09/16/2018
Copyright © 2015, the American Society of Anesthesiologists, Inc. Wolters Kluwer Health, Inc. Unauthorized reproduction of this article is prohibited.
Anesthesiology 2015; 123:1170-85 1174 Song et al.
AMPK Activation Ameliorates Bone Cancer Pain
observed after systemic administration. It should be noted that systemic administration of resveratrol alone (up to 300 mg/kg) showed no notable effects on the pain thresh-old (fig. 1G) or motor function in naïve rats (fig. 1, H and I). The retention time on the rod was not changed in the rats administered with resveratrol compared with those treated with vehicle.
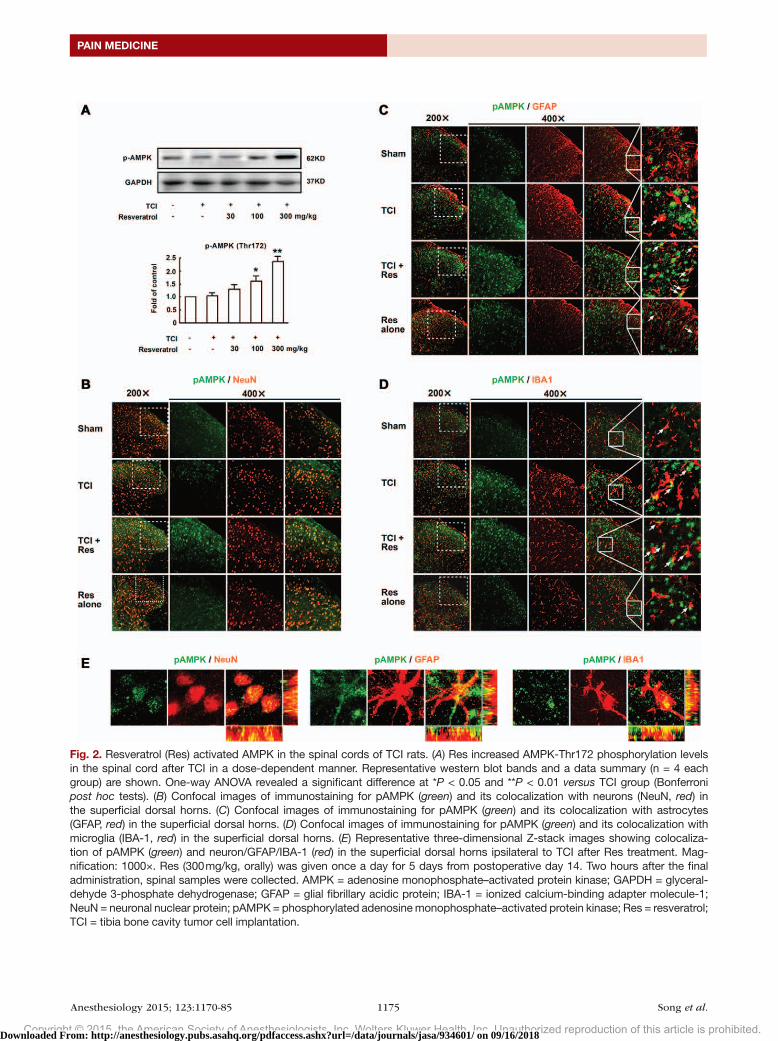
Activation and Distribution of Spinal AMPK after TCI and Resveratrol AdministrationThe level of AMPK phosphorylation in the spinal cord of TCI rats was significantly increased in a dose-dependent manner after resveratrol administration; the levels remained low in both naive and control TCI rats (fig. 2A).
Analysis of the cellular distribution revealed that phos-phorylated AMPK distributed throughout the dH of the spinal cord. Confocal images showed that, in naive rats, the phosphorylated AMPK immunoreactivity was colocalized primarily with neurons (as indicated by NeuN, a neuronal marker; fig. 2B, top). TCI itself induced a moderate phos-phorylation of pAMPK (fig. 2B, middle), whereas resvera-trol administration (fig. 2B, bottom) resulted in a significant increase of pAMPK immunoreactivity in the dH, especially in layers II to III. Although colocalized primarily with neu-rons, a notable amount of colocalization of pAMPK with astrocytes (GFAP; fig. 2C) and microglia (IBA-1, a microglia marker; fig. 2d) after TCI operation and resveratrol treat-ment was observed. Further, three-dimensional Z-stack
Fig. 1. Res attenuates pain-related behaviors through AMPK activation after TCI. Rats administered Res only on day 14 (A) or on days 14, 15, 16, and 17 (B) post-TCI showed a significant increase in mechanical withdrawal compared with that of the bone cancer pain group. Res was administered intragastrically. (C) The AMPK inhibitor compound C reversed the effects of Res on TCI-induced mechanical allodynia. (D) The AMPK activator AICAR attenuates pain-related behaviors after TCI. (E, F) Res admin-istered IT attenuates pain-related behaviors after TCI. Drug administration is indicated by the arrows (n = 8 each group). (G) Res (100 and 300 mg/kg, PO) showed no significant effect on the mechanical withdrawal of rats. (H) Res (300 mg/kg, PO) showed no notable effect on the motor function of rats in rotating-rod test. Rats were slightly put onto the rod with a rotating speed of 16, 24, and 32 rpm. The retention time on rod was recorded. (I) Comparison of endurance on the rotating rod (24 rpm) in vehicle- and Res-treated rats after consecutive administration. Drug administration is indicated by the arrows (n = 8 each group). Two-way ANOVA revealed a significant difference at ##P < 0.01 versus Control; and *P < 0.05 and **P < 0.01 versus TCI group (Bonferroni post hoc tests). AMPK = adenosine monophosphate–activated protein kinase; IT = intrathecally; PO = orally; Res = resveratrol; r.p.m. = revolutions per minute; TCI = tibia bone cavity tumor cell implantation.
Downloaded From: http://anesthesiology.pubs.asahq.org/pdfaccess.ashx?url=/data/journals/jasa/934601/ on 09/16/2018
Copyright © 2015, the American Society of Anesthesiologists, Inc. Wolters Kluwer Health, Inc. Unauthorized reproduction of this article is prohibited.
Anesthesiology 2015; 123:1170-85 1175 Song et al.
PAIN MEDICINE
Fig. 2. Resveratrol (Res) activated AMPK in the spinal cords of TCI rats. (A) Res increased AMPK-Thr172 phosphorylation levels in the spinal cord after TCI in a dose-dependent manner. Representative western blot bands and a data summary (n = 4 each group) are shown. One-way ANOVA revealed a significant difference at *P < 0.05 and **P < 0.01 versus TCI group (Bonferroni post hoc tests). (B) Confocal images of immunostaining for pAMPK (green) and its colocalization with neurons (NeuN, red) in the superficial dorsal horns. (C) Confocal images of immunostaining for pAMPK (green) and its colocalization with astrocytes (GFAP, red) in the superficial dorsal horns. (D) Confocal images of immunostaining for pAMPK (green) and its colocalization with microglia (IBA-1, red) in the superficial dorsal horns. (E) Representative three-dimensional Z-stack images showing colocaliza-tion of pAMPK (green) and neuron/GFAP/IBA-1 (red) in the superficial dorsal horns ipsilateral to TCI after Res treatment. Mag-nification: 1000×. Res (300 mg/kg, orally) was given once a day for 5 days from postoperative day 14. Two hours after the final administration, spinal samples were collected. AMPK = adenosine monophosphate–activated protein kinase; GAPDH = glyceral-dehyde 3-phosphate dehydrogenase; GFAP = glial fibrillary acidic protein; IBA-1 = ionized calcium-binding adapter molecule-1; NeuN = neuronal nuclear protein; pAMPK = phosphorylated adenosine monophosphate–activated protein kinase; Res = resveratrol; TCI = tibia bone cavity tumor cell implantation.
Downloaded From: http://anesthesiology.pubs.asahq.org/pdfaccess.ashx?url=/data/journals/jasa/934601/ on 09/16/2018
Copyright © 2015, the American Society of Anesthesiologists, Inc. Wolters Kluwer Health, Inc. Unauthorized reproduction of this article is prohibited.
Anesthesiology 2015; 123:1170-85 1176 Song et al.
AMPK Activation Ameliorates Bone Cancer Pain
Fig. 3. Resveratrol inhibits the activation of spinal astrocytes and microglia after TCI. (A, B) Representative western blot bands and a data summary (n = 4 each group) of expression of GFAP and IBA-1, which are markers of astrocytes and microglia, respectively. (C, D) Confocal images and immunofluorescence analysis data show GFAP expression in the superficial dorsal horns. Quantification of GFAP immunofluorescence is presented as mean fluorescence pixels (n = 5, five images per animal). (E, F) Confocal images and immunofluorescence analysis show IBA-1 expression in the superficial dorsal horns. Quantification
Downloaded From: http://anesthesiology.pubs.asahq.org/pdfaccess.ashx?url=/data/journals/jasa/934601/ on 09/16/2018

Copyright © 2015, the American Society of Anesthesiologists, Inc. Wolters Kluwer Health, Inc. Unauthorized reproduction of this article is prohibited.
Anesthesiology 2015; 123:1170-85 1177 Song et al.
PAIN MEDICINE
images showed that phosphorylated AMPK was expressed in neuron, astrocyte, and microglia in the dH of rats treated with TCI operation and resveratrol administration (fig. 2E).
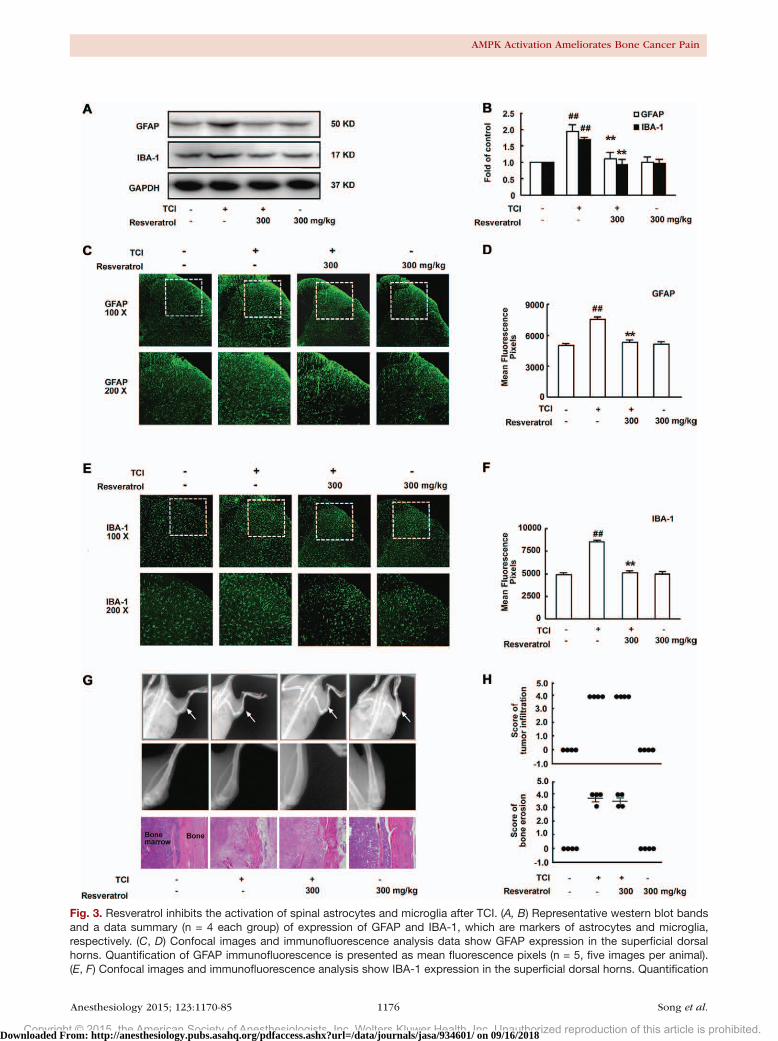
Oral Administration of Resveratrol Suppresses the Activation of Microglia and Astrocytes in Rat Spinal CordsAstrocytes and microglia have been recognized as important contributors to tumor pain pathways. Expression levels of GFAP and IBA-1 have been used as indicators of astrocytic and microglial activity, respectively. Our results showed that the expression of both GFAP and IBA-1 were enhanced by TCI, whereas resveratrol (300 mg/kg, PO, once daily for 5 days from postoperative day 14) down-regulated the expression of both markers (fig. 3, A and B). The increased GFAP and IBA-1 immunoreactivity, which was distributed predominately in the superficial dH ipsilateral to TCI, sug-gested the activation of both astrocytes and microglia. Nota-bly, this activation was alleviated by resveratrol (fig. 3, C–F).
To further verify whether inhibition of bone cancer pain-induced hypersensitivity by resveratrol is because of a decrease in tumor growth and/or spinal sensitization, bone destruction was monitored using radiologic and histologic methods (fig. 3, G and H). Local bone destruction takes place at the proximal epiphysis of the tibia during this time. Histologic examination revealed bone marrow spaces infil-trated with malignant tumor on day 18 after inoculation. However, bone loss and tumor infiltration was not notably changed by resveratrol (300 mg/kg, PO, once daily for 5 days from postoperative day 14). Bone destruction was not observed in the sham or only resveratrol-treated animals.
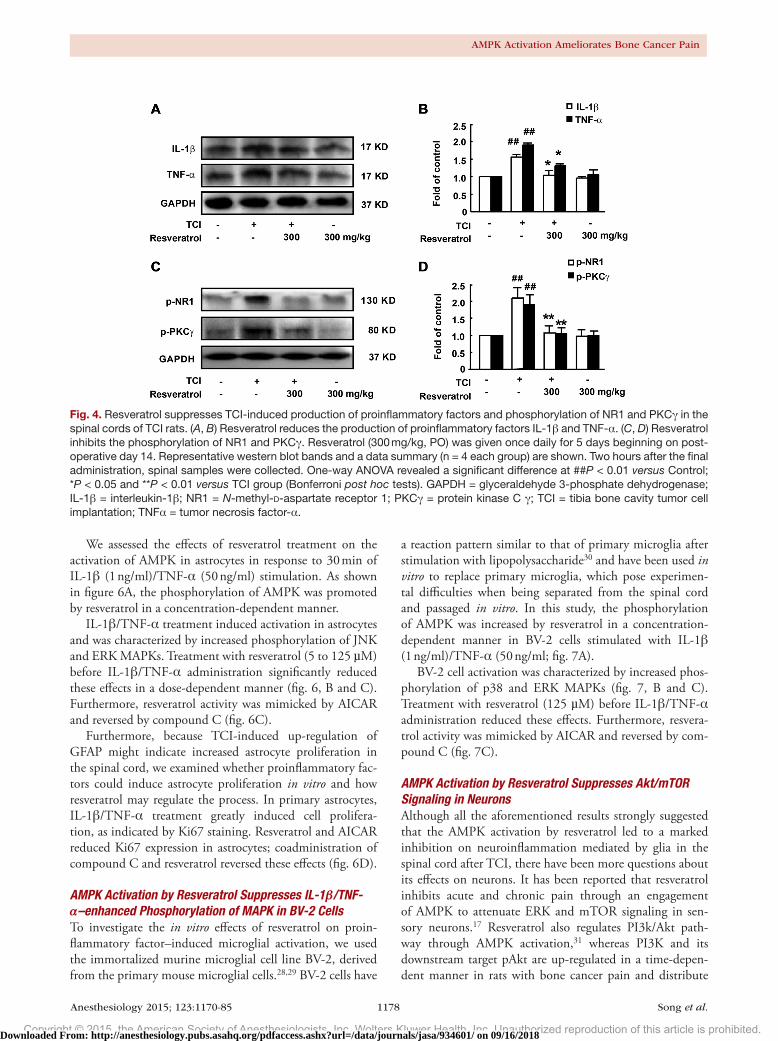
Resveratrol Reduces TCI-induced Production of Proinflammatory Factors and Phosphorylation of NR1, PKCγ, and MAPKs in the Spinal CordThe activation of glia in the context of TCI was followed by an increase in the levels of proinflammatory factors IL-1β and TNF-α. Treatment with resveratrol (300 mg/kg, PO,
once daily for 5 days from postoperative day 14) inhibited the increased production of IL-1β and TNF-α in the spinal cord (fig. 4, A and B). NMdA receptor phosphorylation, which can be activated by proinflammatory factors, has a well-established role in various pain states. PKC kinases can enhance NMdA receptor function. Herein, we showed that TCI treatment significantly increased the levels of phos-phorylated NR1 and PKCγ, whereas repetitive treatment with resveratrol (300 mg/kg, PO, once daily for 5 days from postoperative day 14) inhibited their expression (fig. 4, C and d). Neither the production of proinflammatory factors nor the phosphorylation of NMdA receptors was altered by resveratrol treatment alone in naïve rats.
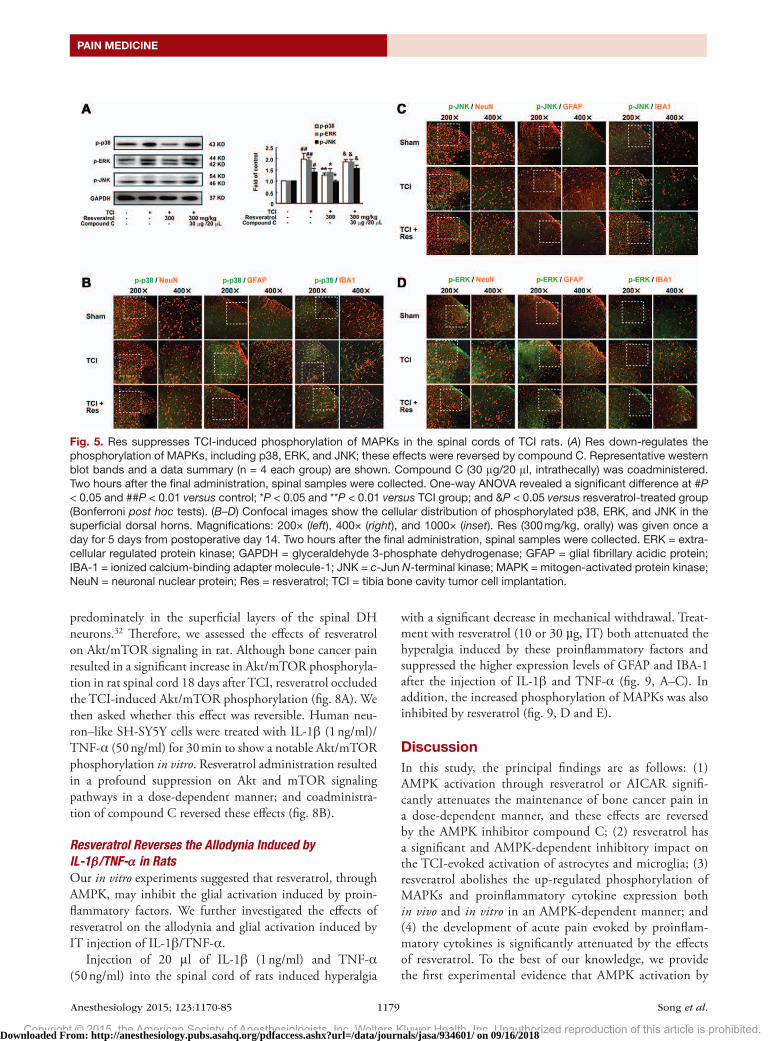
Resveratrol Reduces TCI-induced Phosphorylation of MAPKs in the Spinal CordPrevious studies indicated that the production of both IL-1β and TNF-α might be mediated by mitogen-activated protein kinase (MAPK) signaling pathways in glia.26,27 As shown in figure 5A, administration of resveratrol (300 mg/kg, PO, once daily for 5 days from postoperative day 14) signifi-cantly reduced the levels of phosphorylated MAPKs (includ-ing p38, extracellular signal-regulated protein kinase [ERK] and JNK induced by TCI. These effects were reversed by the coadministration of the AMPK inhibitor compound C (30 μg, IT). These results suggest that resveratrol-mediated sup-pression of MAPKs and subsequent proinflammatory factors and NMdA receptors may be secondary to AMPK signaling activation.
We further determined the cellular distribution and phos-phorylation of MAPKs within the dH after TCI operation and resveratrol treatment. Confocal images showed that, in rats dH, the phosphorylated p38 was colocalized primarily with microglia (IBA-1; fig. 5B), whereas the phosphorylated JNK colocalized with astrocyte (GFAP; fig. 5C). The phos-phorylated ERK could be observed in neuron, astrocyte, and microglia but mainly in neuron (NeuN; fig. 5d). TCI in rats caused a significantly increased phosphorylation of all the three MAPKs in the spinal cords, whereas resveratrol could suppress these effects, in line with the western blot data in figure 5A. Taken together, these results indicated that resve-ratrol inhibited activation of astrocytes and microglia in the spinal cords through MAPKs suppression, thereby reducing the production of proinflammatory cytokines.
AMPK Activation via Resveratrol Treatment Inhibits IL-1β/TNF-α–induced Activation and Proliferation of Primary Astrocytes In VitroOur findings suggest that resveratrol may act through AMPK signaling pathways to attenuate TCI through inhibition of spinal cord glial activation in vivo. We further asked whether the inhibitory effect of AMPK activators on glial activation is direct or indirect. Therefore, we isolated primary rat astro-cytes and cultured them in vitro to investigate the role of resveratrol.
Fig. 3. (Continued). of IBA-1 immunofluorescence is pre-sented as mean fluorescence pixels (n = 5, five images per animal). (G) Radiologic and histochemical analysis of tumor development in the tibia. To confirm bone destruction of the tibia by tumor, rats were radiographed 18 days after tumor cell inoculation of the ipsilateral hind limb. Arrows indicate the proximal cortical bone. Hematoxylin and eosin staining of the left tibia showed that bone marrow spaces on day 18 after Walker 256 cell inoculation. (H) Pathohistologic grades were evaluated according to the infiltration of tumor cells and bone erosion (n = 4 each group). Resveratrol (300 mg/kg, oral-ly) was given once a day for 5 days from postoperative day 14. Two hours after the final administration, spinal samples were collected. One-way ANOVA revealed a significant differ-ence at ##P < 0.01 versus control and **P < 0.01 versus TCI group (Bonferroni post hoc tests). GAPDH = glyceraldehyde 3-phosphate dehydrogenase; GFAP = glial fibrillary acidic protein; IBA-1 = ionized calcium-binding adapter molecule-1; TCI = tibia bone cavity tumor cell implantation.
Downloaded From: http://anesthesiology.pubs.asahq.org/pdfaccess.ashx?url=/data/journals/jasa/934601/ on 09/16/2018
Copyright © 2015, the American Society of Anesthesiologists, Inc. Wolters Kluwer Health, Inc. Unauthorized reproduction of this article is prohibited.
Anesthesiology 2015; 123:1170-85 1178 Song et al.
AMPK Activation Ameliorates Bone Cancer Pain
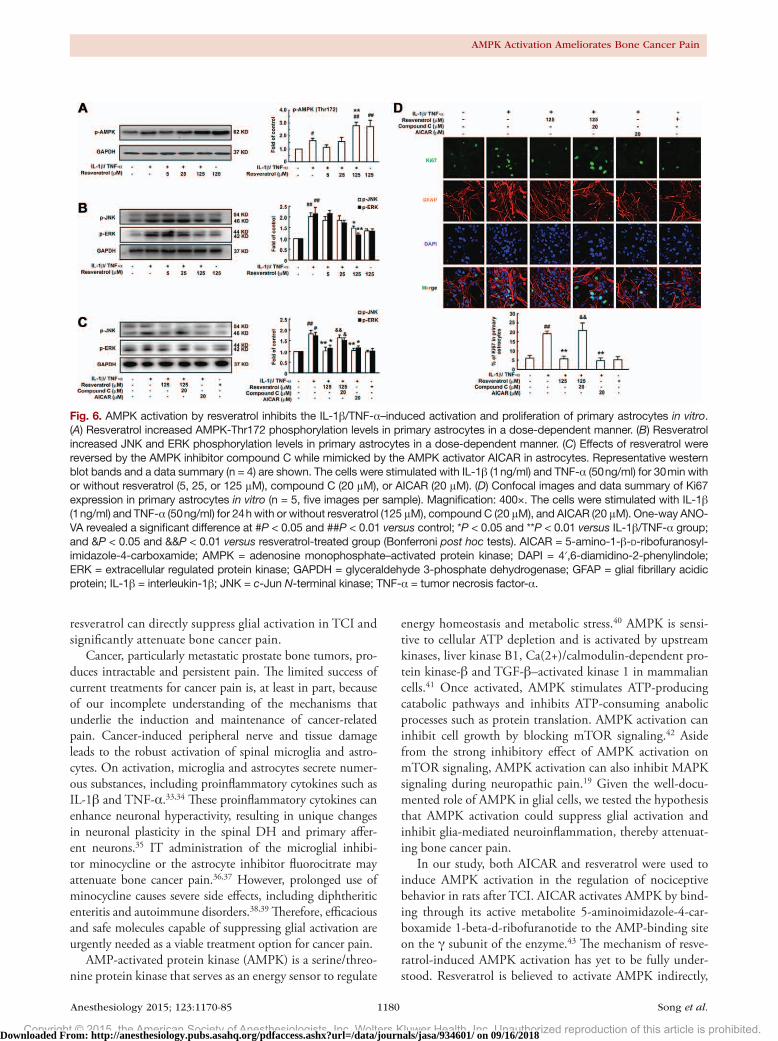
We assessed the effects of resveratrol treatment on the activation of AMPK in astrocytes in response to 30 min of IL-1β (1 ng/ml)/TNF-α (50 ng/ml) stimulation. As shown in figure 6A, the phosphorylation of AMPK was promoted by resveratrol in a concentration-dependent manner.
IL-1β/TNF-α treatment induced activation in astrocytes and was characterized by increased phosphorylation of JNK and ERK MAPKs. Treatment with resveratrol (5 to 125 µM) before IL-1β/TNF-α administration significantly reduced these effects in a dose-dependent manner (fig. 6, B and C). Furthermore, resveratrol activity was mimicked by AICAR and reversed by compound C (fig. 6C).
Furthermore, because TCI-induced up-regulation of GFAP might indicate increased astrocyte proliferation in the spinal cord, we examined whether proinflammatory fac-tors could induce astrocyte proliferation in vitro and how resveratrol may regulate the process. In primary astrocytes, IL-1β/TNF-α treatment greatly induced cell prolifera-tion, as indicated by Ki67 staining. Resveratrol and AICAR reduced Ki67 expression in astrocytes; coadministration of compound C and resveratrol reversed these effects (fig. 6d).
AMPK Activation by Resveratrol Suppresses IL-1β/TNF-α–enhanced Phosphorylation of MAPK in BV-2 CellsTo investigate the in vitro effects of resveratrol on proin-flammatory factor–induced microglial activation, we used the immortalized murine microglial cell line BV-2, derived from the primary mouse microglial cells.28,29 BV-2 cells have
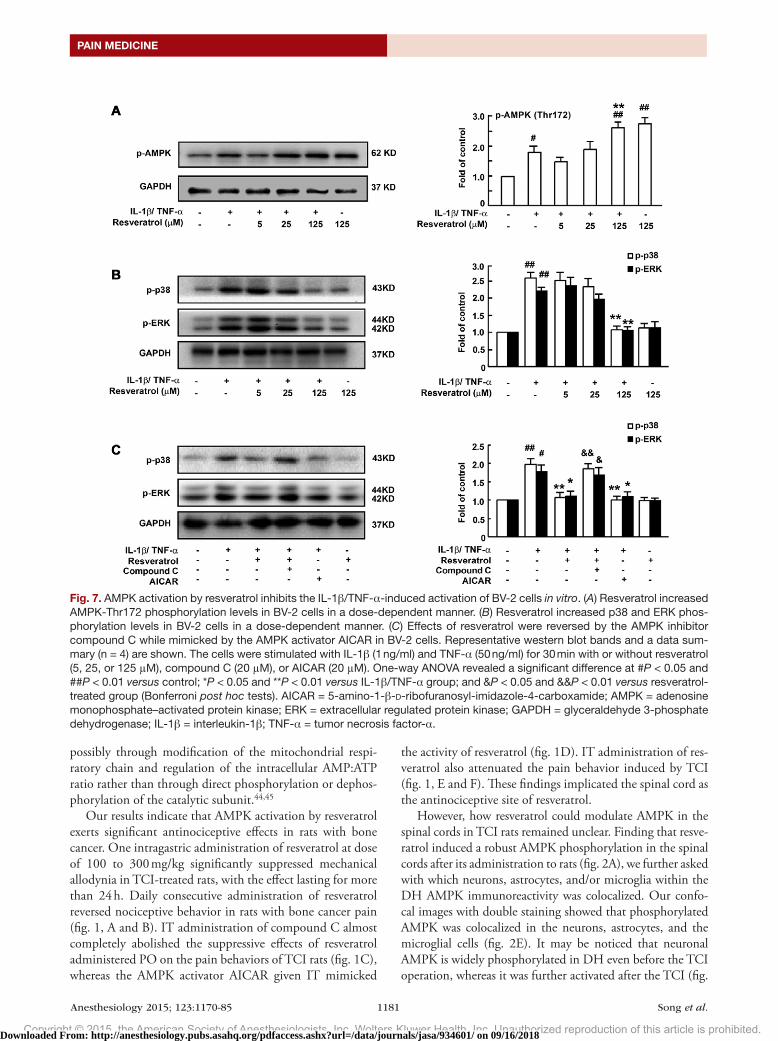
a reaction pattern similar to that of primary microglia after stimulation with lipopolysaccharide30 and have been used in vitro to replace primary microglia, which pose experimen-tal difficulties when being separated from the spinal cord and passaged in vitro. In this study, the phosphorylation of AMPK was increased by resveratrol in a concentration-dependent manner in BV-2 cells stimulated with IL-1β (1 ng/ml)/TNF-α (50 ng/ml; fig. 7A).
BV-2 cell activation was characterized by increased phos-phorylation of p38 and ERK MAPKs (fig. 7, B and C). Treatment with resveratrol (125 µM) before IL-1β/TNF-α administration reduced these effects. Furthermore, resvera-trol activity was mimicked by AICAR and reversed by com-pound C (fig. 7C).
AMPK Activation by Resveratrol Suppresses Akt/mTOR Signaling in NeuronsAlthough all the aforementioned results strongly suggested that the AMPK activation by resveratrol led to a marked inhibition on neuroinflammation mediated by glia in the spinal cord after TCI, there have been more questions about its effects on neurons. It has been reported that resveratrol inhibits acute and chronic pain through an engagement of AMPK to attenuate ERK and mTOR signaling in sen-sory neurons.17 Resveratrol also regulates PI3k/Akt path-way through AMPK activation,31 whereas PI3K and its downstream target pAkt are up-regulated in a time-depen-dent manner in rats with bone cancer pain and distribute
Fig. 4. Resveratrol suppresses TCI-induced production of proinflammatory factors and phosphorylation of NR1 and PKCγ in the spinal cords of TCI rats. (A, B) Resveratrol reduces the production of proinflammatory factors IL-1β and TNF-α. (C, D) Resveratrol inhibits the phosphorylation of NR1 and PKCγ. Resveratrol (300 mg/kg, PO) was given once daily for 5 days beginning on post-operative day 14. Representative western blot bands and a data summary (n = 4 each group) are shown. Two hours after the final administration, spinal samples were collected. One-way ANOVA revealed a significant difference at ##P < 0.01 versus Control; *P < 0.05 and **P < 0.01 versus TCI group (Bonferroni post hoc tests). GAPDH = glyceraldehyde 3-phosphate dehydrogenase; IL-1β = interleukin-1β; NR1 = N-methyl-D-aspartate receptor 1; PKCγ = protein kinase C γ; TCI = tibia bone cavity tumor cell implantation; TNFα = tumor necrosis factor-α.
Downloaded From: http://anesthesiology.pubs.asahq.org/pdfaccess.ashx?url=/data/journals/jasa/934601/ on 09/16/2018
Copyright © 2015, the American Society of Anesthesiologists, Inc. Wolters Kluwer Health, Inc. Unauthorized reproduction of this article is prohibited.
Anesthesiology 2015; 123:1170-85 1179 Song et al.
PAIN MEDICINE
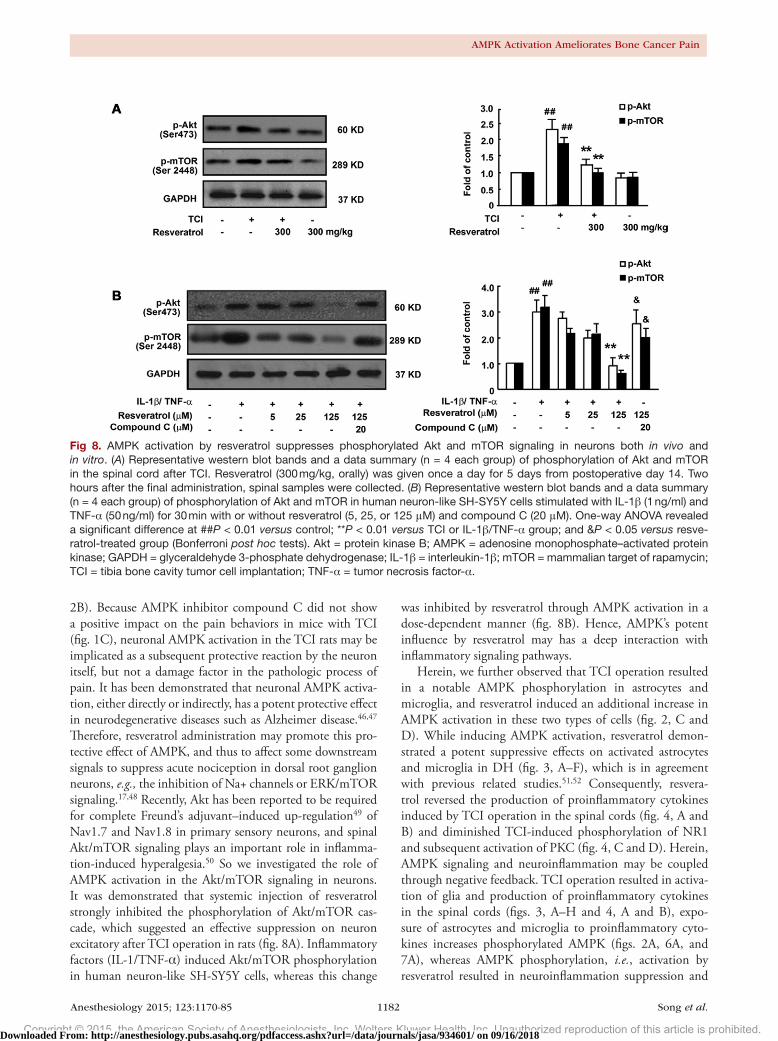
predominately in the superficial layers of the spinal dH neurons.32 Therefore, we assessed the effects of resveratrol on Akt/mTOR signaling in rat. Although bone cancer pain resulted in a significant increase in Akt/mTOR phosphoryla-tion in rat spinal cord 18 days after TCI, resveratrol occluded the TCI-induced Akt/mTOR phosphorylation (fig. 8A). We then asked whether this effect was reversible. Human neu-ron–like SH-SY5Y cells were treated with IL-1β (1 ng/ml)/TNF-α (50 ng/ml) for 30 min to show a notable Akt/mTOR phosphorylation in vitro. Resveratrol administration resulted in a profound suppression on Akt and mTOR signaling pathways in a dose-dependent manner; and coadministra-tion of compound C reversed these effects (fig. 8B).
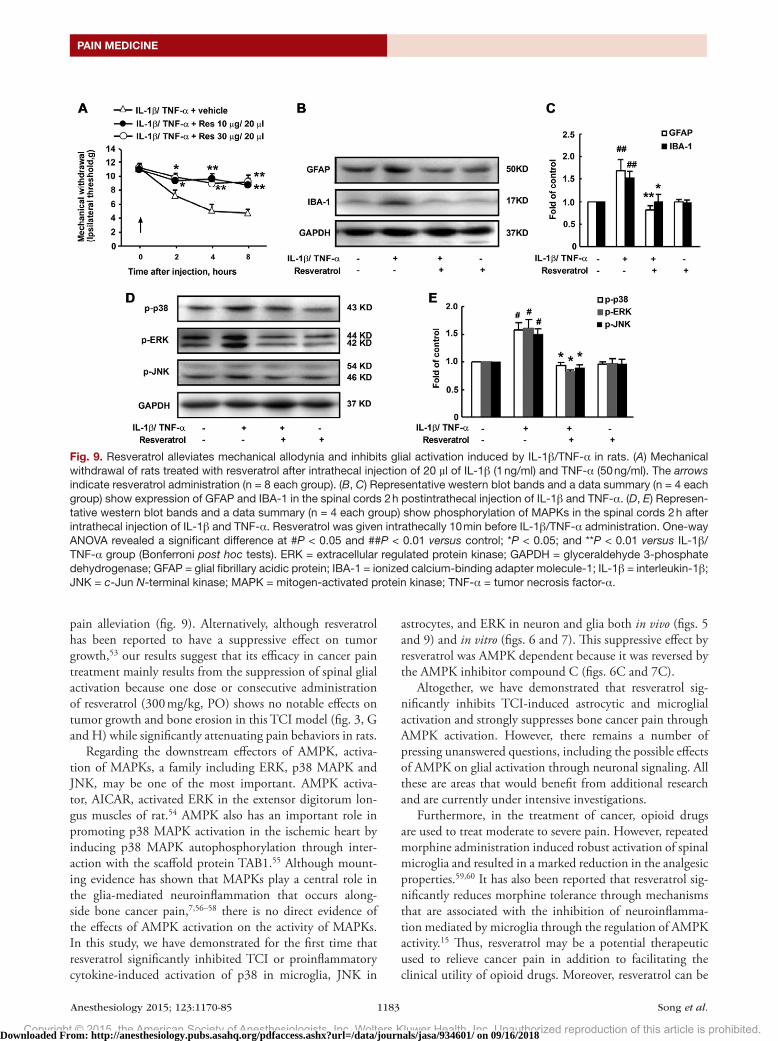
Resveratrol Reverses the Allodynia Induced by IL-1β/TNF-α in RatsOur in vitro experiments suggested that resveratrol, through AMPK, may inhibit the glial activation induced by proin-flammatory factors. We further investigated the effects of resveratrol on the allodynia and glial activation induced by IT injection of IL-1β/TNF-α.
Injection of 20 μl of IL-1β (1 ng/ml) and TNF-α (50 ng/ml) into the spinal cord of rats induced hyperalgia
with a significant decrease in mechanical withdrawal. Treat-ment with resveratrol (10 or 30 µg, IT) both attenuated the hyperalgia induced by these proinflammatory factors and suppressed the higher expression levels of GFAP and IBA-1 after the injection of IL-1β and TNF-α (fig. 9, A–C). In addition, the increased phosphorylation of MAPKs was also inhibited by resveratrol (fig. 9, d and E).
DiscussionIn this study, the principal findings are as follows: (1) AMPK activation through resveratrol or AICAR signifi-cantly attenuates the maintenance of bone cancer pain in a dose-dependent manner, and these effects are reversed by the AMPK inhibitor compound C; (2) resveratrol has a significant and AMPK-dependent inhibitory impact on the TCI-evoked activation of astrocytes and microglia; (3) resveratrol abolishes the up-regulated phosphorylation of MAPKs and proinflammatory cytokine expression both in vivo and in vitro in an AMPK-dependent manner; and (4) the development of acute pain evoked by proinflam-matory cytokines is significantly attenuated by the effects of resveratrol. To the best of our knowledge, we provide the first experimental evidence that AMPK activation by
Fig. 5. Res suppresses TCI-induced phosphorylation of MAPKs in the spinal cords of TCI rats. (A) Res down-regulates the phosphorylation of MAPKs, including p38, ERK, and JNK; these effects were reversed by compound C. Representative western blot bands and a data summary (n = 4 each group) are shown. Compound C (30 μg/20 μl, intrathecally) was coadministered. Two hours after the final administration, spinal samples were collected. One-way ANOVA revealed a significant difference at #P < 0.05 and ##P < 0.01 versus control; *P < 0.05 and **P < 0.01 versus TCI group; and &P < 0.05 versus resveratrol-treated group (Bonferroni post hoc tests). (B–D) Confocal images show the cellular distribution of phosphorylated p38, ERK, and JNK in the superficial dorsal horns. Magnifications: 200× (left), 400× (right), and 1000× (inset). Res (300 mg/kg, orally) was given once a day for 5 days from postoperative day 14. Two hours after the final administration, spinal samples were collected. ERK = extra-cellular regulated protein kinase; GAPDH = glyceraldehyde 3-phosphate dehydrogenase; GFAP = glial fibrillary acidic protein; IBA-1 = ionized calcium-binding adapter molecule-1; JNK = c-Jun N-terminal kinase; MAPK = mitogen-activated protein kinase; NeuN = neuronal nuclear protein; Res = resveratrol; TCI = tibia bone cavity tumor cell implantation.
Downloaded From: http://anesthesiology.pubs.asahq.org/pdfaccess.ashx?url=/data/journals/jasa/934601/ on 09/16/2018
Copyright © 2015, the American Society of Anesthesiologists, Inc. Wolters Kluwer Health, Inc. Unauthorized reproduction of this article is prohibited.
Anesthesiology 2015; 123:1170-85 1180 Song et al.
AMPK Activation Ameliorates Bone Cancer Pain
resveratrol can directly suppress glial activation in TCI and significantly attenuate bone cancer pain.
Cancer, particularly metastatic prostate bone tumors, pro-duces intractable and persistent pain. The limited success of current treatments for cancer pain is, at least in part, because of our incomplete understanding of the mechanisms that underlie the induction and maintenance of cancer-related pain. Cancer-induced peripheral nerve and tissue damage leads to the robust activation of spinal microglia and astro-cytes. On activation, microglia and astrocytes secrete numer-ous substances, including proinflammatory cytokines such as IL-1β and TNF-α.33,34 These proinflammatory cytokines can enhance neuronal hyperactivity, resulting in unique changes in neuronal plasticity in the spinal dH and primary affer-ent neurons.35 IT administration of the microglial inhibi-tor minocycline or the astrocyte inhibitor fluorocitrate may attenuate bone cancer pain.36,37 However, prolonged use of minocycline causes severe side effects, including diphtheritic enteritis and autoimmune disorders.38,39 Therefore, efficacious and safe molecules capable of suppressing glial activation are urgently needed as a viable treatment option for cancer pain.
AMP-activated protein kinase (AMPK) is a serine/threo-nine protein kinase that serves as an energy sensor to regulate
energy homeostasis and metabolic stress.40 AMPK is sensi-tive to cellular ATP depletion and is activated by upstream kinases, liver kinase B1, Ca(2+)/calmodulin-dependent pro-tein kinase-β and TGF-β–activated kinase 1 in mammalian cells.41 Once activated, AMPK stimulates ATP-producing catabolic pathways and inhibits ATP-consuming anabolic processes such as protein translation. AMPK activation can inhibit cell growth by blocking mTOR signaling.42 Aside from the strong inhibitory effect of AMPK activation on mTOR signaling, AMPK activation can also inhibit MAPK signaling during neuropathic pain.19 Given the well-docu-mented role of AMPK in glial cells, we tested the hypothesis that AMPK activation could suppress glial activation and inhibit glia-mediated neuroinflammation, thereby attenuat-ing bone cancer pain.
In our study, both AICAR and resveratrol were used to induce AMPK activation in the regulation of nociceptive behavior in rats after TCI. AICAR activates AMPK by bind-ing through its active metabolite 5-aminoimidazole-4-car-boxamide 1-beta-d-ribofuranotide to the AMP-binding site on the γ subunit of the enzyme.43 The mechanism of resve-ratrol-induced AMPK activation has yet to be fully under-stood. Resveratrol is believed to activate AMPK indirectly,
Fig. 6. AMPK activation by resveratrol inhibits the IL-1β/TNF-α–induced activation and proliferation of primary astrocytes in vitro. (A) Resveratrol increased AMPK-Thr172 phosphorylation levels in primary astrocytes in a dose-dependent manner. (B) Resveratrol increased JNK and ERK phosphorylation levels in primary astrocytes in a dose-dependent manner. (C) Effects of resveratrol were reversed by the AMPK inhibitor compound C while mimicked by the AMPK activator AICAR in astrocytes. Representative western blot bands and a data summary (n = 4) are shown. The cells were stimulated with IL-1β (1 ng/ml) and TNF-α (50 ng/ml) for 30 min with or without resveratrol (5, 25, or 125 μM), compound C (20 μM), or AICAR (20 μM). (D) Confocal images and data summary of Ki67 expression in primary astrocytes in vitro (n = 5, five images per sample). Magnification: 400×. The cells were stimulated with IL-1β (1 ng/ml) and TNF-α (50 ng/ml) for 24 h with or without resveratrol (125 μM), compound C (20 μM), and AICAR (20 μM). One-way ANO-VA revealed a significant difference at #P < 0.05 and ##P < 0.01 versus control; *P < 0.05 and **P < 0.01 versus IL-1β/TNF-α group; and &P < 0.05 and &&P < 0.01 versus resveratrol-treated group (Bonferroni post hoc tests). AICAR = 5-amino-1-β-D-ribofuranosyl-imidazole-4-carboxamide; AMPK = adenosine monophosphate–activated protein kinase; DAPI = 4′,6-diamidino-2-phenylindole; ERK = extracellular regulated protein kinase; GAPDH = glyceraldehyde 3-phosphate dehydrogenase; GFAP = glial fibrillary acidic protein; IL-1β = interleukin-1β; JNK = c-Jun N-terminal kinase; TNF-α = tumor necrosis factor-α.
Downloaded From: http://anesthesiology.pubs.asahq.org/pdfaccess.ashx?url=/data/journals/jasa/934601/ on 09/16/2018
Copyright © 2015, the American Society of Anesthesiologists, Inc. Wolters Kluwer Health, Inc. Unauthorized reproduction of this article is prohibited.
Anesthesiology 2015; 123:1170-85 1181 Song et al.
PAIN MEDICINE
possibly through modification of the mitochondrial respi-ratory chain and regulation of the intracellular AMP:ATP ratio rather than through direct phosphorylation or dephos-phorylation of the catalytic subunit.44,45
Our results indicate that AMPK activation by resveratrol exerts significant antinociceptive effects in rats with bone cancer. One intragastric administration of resveratrol at dose of 100 to 300 mg/kg significantly suppressed mechanical allodynia in TCI-treated rats, with the effect lasting for more than 24 h. daily consecutive administration of resveratrol reversed nociceptive behavior in rats with bone cancer pain (fig. 1, A and B). IT administration of compound C almost completely abolished the suppressive effects of resveratrol administered PO on the pain behaviors of TCI rats (fig. 1C), whereas the AMPK activator AICAR given IT mimicked
the activity of resveratrol (fig. 1d). IT administration of res-veratrol also attenuated the pain behavior induced by TCI (fig. 1, E and F). These findings implicated the spinal cord as the antinociceptive site of resveratrol.
However, how resveratrol could modulate AMPK in the spinal cords in TCI rats remained unclear. Finding that resve-ratrol induced a robust AMPK phosphorylation in the spinal cords after its administration to rats (fig. 2A), we further asked with which neurons, astrocytes, and/or microglia within the dH AMPK immunoreactivity was colocalized. Our confo-cal images with double staining showed that phosphorylated AMPK was colocalized in the neurons, astrocytes, and the microglial cells (fig. 2E). It may be noticed that neuronal AMPK is widely phosphorylated in dH even before the TCI operation, whereas it was further activated after the TCI (fig.
Fig. 7. AMPK activation by resveratrol inhibits the IL-1β/TNF-α-induced activation of BV-2 cells in vitro. (A) Resveratrol increased AMPK-Thr172 phosphorylation levels in BV-2 cells in a dose-dependent manner. (B) Resveratrol increased p38 and ERK phos-phorylation levels in BV-2 cells in a dose-dependent manner. (C) Effects of resveratrol were reversed by the AMPK inhibitor compound C while mimicked by the AMPK activator AICAR in BV-2 cells. Representative western blot bands and a data sum-mary (n = 4) are shown. The cells were stimulated with IL-1β (1 ng/ml) and TNF-α (50 ng/ml) for 30 min with or without resveratrol (5, 25, or 125 μM), compound C (20 μM), or AICAR (20 μM). One-way ANOVA revealed a significant difference at #P < 0.05 and ##P < 0.01 versus control; *P < 0.05 and **P < 0.01 versus IL-1β/TNF-α group; and &P < 0.05 and &&P < 0.01 versus resveratrol-treated group (Bonferroni post hoc tests). AICAR = 5-amino-1-β-D-ribofuranosyl-imidazole-4-carboxamide; AMPK = adenosine monophosphate–activated protein kinase; ERK = extracellular regulated protein kinase; GAPDH = glyceraldehyde 3-phosphate dehydrogenase; IL-1β = interleukin-1β; TNF-α = tumor necrosis factor-α.
Downloaded From: http://anesthesiology.pubs.asahq.org/pdfaccess.ashx?url=/data/journals/jasa/934601/ on 09/16/2018
Copyright © 2015, the American Society of Anesthesiologists, Inc. Wolters Kluwer Health, Inc. Unauthorized reproduction of this article is prohibited.
Anesthesiology 2015; 123:1170-85 1182 Song et al.
AMPK Activation Ameliorates Bone Cancer Pain
2B). Because AMPK inhibitor compound C did not show a positive impact on the pain behaviors in mice with TCI (fig. 1C), neuronal AMPK activation in the TCI rats may be implicated as a subsequent protective reaction by the neuron itself, but not a damage factor in the pathologic process of pain. It has been demonstrated that neuronal AMPK activa-tion, either directly or indirectly, has a potent protective effect in neurodegenerative diseases such as Alzheimer disease.46,47 Therefore, resveratrol administration may promote this pro-tective effect of AMPK, and thus to affect some downstream signals to suppress acute nociception in dorsal root ganglion neurons, e.g., the inhibition of Na+ channels or ERK/mTOR signaling.17,48 Recently, Akt has been reported to be required for complete Freund’s adjuvant–induced up-regulation49 of Nav1.7 and Nav1.8 in primary sensory neurons, and spinal Akt/mTOR signaling plays an important role in inflamma-tion-induced hyperalgesia.50 So we investigated the role of AMPK activation in the Akt/mTOR signaling in neurons. It was demonstrated that systemic injection of resveratrol strongly inhibited the phosphorylation of Akt/mTOR cas-cade, which suggested an effective suppression on neuron excitatory after TCI operation in rats (fig. 8A). Inflammatory factors (IL-1/TNF-α) induced Akt/mTOR phosphorylation in human neuron-like SH-SY5Y cells, whereas this change
was inhibited by resveratrol through AMPK activation in a dose-dependent manner (fig. 8B). Hence, AMPK’s potent influence by resveratrol may has a deep interaction with inflammatory signaling pathways.
Herein, we further observed that TCI operation resulted in a notable AMPK phosphorylation in astrocytes and microglia, and resveratrol induced an additional increase in AMPK activation in these two types of cells (fig. 2, C and d). While inducing AMPK activation, resveratrol demon-strated a potent suppressive effects on activated astrocytes and microglia in dH (fig. 3, A–F), which is in agreement with previous related studies.51,52 Consequently, resvera-trol reversed the production of proinflammatory cytokines induced by TCI operation in the spinal cords (fig. 4, A and B) and diminished TCI-induced phosphorylation of NR1 and subsequent activation of PKC (fig. 4, C and d). Herein, AMPK signaling and neuroinflammation may be coupled through negative feedback. TCI operation resulted in activa-tion of glia and production of proinflammatory cytokines in the spinal cords (figs. 3, A–H and 4, A and B), expo-sure of astrocytes and microglia to proinflammatory cyto-kines increases phosphorylated AMPK (figs. 2A, 6A, and 7A), whereas AMPK phosphorylation, i.e., activation by resveratrol resulted in neuroinflammation suppression and
Fig 8. AMPK activation by resveratrol suppresses phosphorylated Akt and mTOR signaling in neurons both in vivo and in vitro. (A) Representative western blot bands and a data summary (n = 4 each group) of phosphorylation of Akt and mTOR in the spinal cord after TCI. Resveratrol (300 mg/kg, orally) was given once a day for 5 days from postoperative day 14. Two hours after the final administration, spinal samples were collected. (B) Representative western blot bands and a data summary (n = 4 each group) of phosphorylation of Akt and mTOR in human neuron-like SH-SY5Y cells stimulated with IL-1β (1 ng/ml) and TNF-α (50 ng/ml) for 30 min with or without resveratrol (5, 25, or 125 μM) and compound C (20 μM). One-way ANOVA revealed a significant difference at ##P < 0.01 versus control; **P < 0.01 versus TCI or IL-1β/TNF-α group; and &P < 0.05 versus resve-ratrol-treated group (Bonferroni post hoc tests). Akt = protein kinase B; AMPK = adenosine monophosphate–activated protein kinase; GAPDH = glyceraldehyde 3-phosphate dehydrogenase; IL-1β = interleukin-1β; mTOR = mammalian target of rapamycin; TCI = tibia bone cavity tumor cell implantation; TNF-α = tumor necrosis factor-α.
Downloaded From: http://anesthesiology.pubs.asahq.org/pdfaccess.ashx?url=/data/journals/jasa/934601/ on 09/16/2018
Copyright © 2015, the American Society of Anesthesiologists, Inc. Wolters Kluwer Health, Inc. Unauthorized reproduction of this article is prohibited.
Anesthesiology 2015; 123:1170-85 1183 Song et al.
PAIN MEDICINE
pain alleviation (fig. 9). Alternatively, although resveratrol has been reported to have a suppressive effect on tumor growth,53 our results suggest that its efficacy in cancer pain treatment mainly results from the suppression of spinal glial activation because one dose or consecutive administration of resveratrol (300 mg/kg, PO) shows no notable effects on tumor growth and bone erosion in this TCI model (fig. 3, G and H) while significantly attenuating pain behaviors in rats.
Regarding the downstream effectors of AMPK, activa-tion of MAPKs, a family including ERK, p38 MAPK and JNK, may be one of the most important. AMPK activa-tor, AICAR, activated ERK in the extensor digitorum lon-gus muscles of rat.54 AMPK also has an important role in promoting p38 MAPK activation in the ischemic heart by inducing p38 MAPK autophosphorylation through inter-action with the scaffold protein TAB1.55 Although mount-ing evidence has shown that MAPKs play a central role in the glia-mediated neuroinflammation that occurs along-side bone cancer pain,7,56–58 there is no direct evidence of the effects of AMPK activation on the activity of MAPKs. In this study, we have demonstrated for the first time that resveratrol significantly inhibited TCI or proinflammatory cytokine-induced activation of p38 in microglia, JNK in
astrocytes, and ERK in neuron and glia both in vivo (figs. 5 and 9) and in vitro (figs. 6 and 7). This suppressive effect by resveratrol was AMPK dependent because it was reversed by the AMPK inhibitor compound C (figs. 6C and 7C).
Altogether, we have demonstrated that resveratrol sig-nificantly inhibits TCI-induced astrocytic and microglial activation and strongly suppresses bone cancer pain through AMPK activation. However, there remains a number of pressing unanswered questions, including the possible effects of AMPK on glial activation through neuronal signaling. All these are areas that would benefit from additional research and are currently under intensive investigations.
Furthermore, in the treatment of cancer, opioid drugs are used to treat moderate to severe pain. However, repeated morphine administration induced robust activation of spinal microglia and resulted in a marked reduction in the analgesic properties.59,60 It has also been reported that resveratrol sig-nificantly reduces morphine tolerance through mechanisms that are associated with the inhibition of neuroinflamma-tion mediated by microglia through the regulation of AMPK activity.15 Thus, resveratrol may be a potential therapeutic used to relieve cancer pain in addition to facilitating the clinical utility of opioid drugs. Moreover, resveratrol can be
Fig. 9. Resveratrol alleviates mechanical allodynia and inhibits glial activation induced by IL-1β/TNF-α in rats. (A) Mechanical withdrawal of rats treated with resveratrol after intrathecal injection of 20 µl of IL-1β (1 ng/ml) and TNF-α (50 ng/ml). The arrows indicate resveratrol administration (n = 8 each group). (B, C) Representative western blot bands and a data summary (n = 4 each group) show expression of GFAP and IBA-1 in the spinal cords 2 h postintrathecal injection of IL-1β and TNF-α. (D, E) Represen-tative western blot bands and a data summary (n = 4 each group) show phosphorylation of MAPKs in the spinal cords 2 h after intrathecal injection of IL-1β and TNF-α. Resveratrol was given intrathecally 10 min before IL-1β/TNF-α administration. One-way ANOVA revealed a significant difference at #P < 0.05 and ##P < 0.01 versus control; *P < 0.05; and **P < 0.01 versus IL-1β/TNF-α group (Bonferroni post hoc tests). ERK = extracellular regulated protein kinase; GAPDH = glyceraldehyde 3-phosphate dehydrogenase; GFAP = glial fibrillary acidic protein; IBA-1 = ionized calcium-binding adapter molecule-1; IL-1β = interleukin-1β; JNK = c-Jun N-terminal kinase; MAPK = mitogen-activated protein kinase; TNF-α = tumor necrosis factor-α.
Downloaded From: http://anesthesiology.pubs.asahq.org/pdfaccess.ashx?url=/data/journals/jasa/934601/ on 09/16/2018

Copyright © 2015, the American Society of Anesthesiologists, Inc. Wolters Kluwer Health, Inc. Unauthorized reproduction of this article is prohibited.
Anesthesiology 2015; 123:1170-85 1184 Song et al.
AMPK Activation Ameliorates Bone Cancer Pain
administered PO; and no side effects were observed after res-veratrol administration at these dosage levels in either TCI or naïve rats. Therefore, resveratrol may constitute an effective, safe, and convenient treatment option for cancer patients experiencing severe pain.
AcknowledgmentsThis work was supported by National Natural Science Foun-dation of China, Beijing, China (grants 81171044, 81471142, 81200860, 81373466, and 81202513) and Natural Science Foundation of Jiangsu Province, Nanjing, China (grant BK2011053).
Competing InterestsThe authors declare no competing interests.
CorrespondenceAddress correspondence to Dr. Liu: Jiangsu Key Labo-ratory of Neurodegeneration, Nanjing Medical Univer-sity, 140 Hanzhong Road, Nanjing 210029, Jiangsu, China. [email protected]; or Dr. Wu: State Key Laboratory of Pharmaceutical Biotechnology, Nanjing University, Nanjing 210093, Jiangsu, China. [email protected]. Information on pur-chasing reprints may be found at www.anesthesiology.org or on the masthead page at the beginning of this issue. ANeSTHe-
SIoLoGY’s articles are made freely accessible to all readers, for personal use only, 6 months from the cover date of the issue.
References 1. Deandrea S, Montanari M, Moja L, Apolone G: Prevalence of
undertreatment in cancer pain. A review of published litera-ture. Ann Oncol 2008; 19:1985–91
2. Joyce JA, Pollard JW: Microenvironmental regulation of metastasis. Nat Rev Cancer 2009; 9:239–52
3. Peters CM, Ghilardi JR, Keyser CP, Kubota K, Lindsay TH, Luger NM, Mach DB, Schwei MJ, Sevcik MA, Mantyh PW: Tumor-induced injury of primary afferent sensory nerve fibers in bone cancer pain. Exp Neurol 2005; 193:85–100
4. Lozano-Ondoua AN, Symons-Liguori AM, Vanderah TW: Cancer-induced bone pain: Mechanisms and models. Neurosci Lett 2013; 557(pt A):52–9
5. Liu S, Liu YP, Song WB, Song XJ: EphrinB-EphB receptor sig-naling contributes to bone cancer pain via Toll-like receptor and proinflammatory cytokines in rat spinal cord. Pain 2013; 154:2823–35
6. Mao-Ying QL, Wang XW, Yang CJ, Li X, Mi WL, Wu GC, Wang YQ: Robust spinal neuroinflammation mediates mechanical allodynia in Walker 256 induced bone cancer rats. Mol Brain 2012; 5:16
7. Wang XW, Li TT, Zhao J, Mao-Ying QL, Zhang H, Hu S, Li Q, Mi WL, Wu GC, Zhang YQ, Wang YQ: Extracellular sig-nal-regulated kinase activation in spinal astrocytes and microglia contributes to cancer-induced bone pain in rats. Neuroscience 2012; 217:172–81
8. Hansen RR, Nielsen CK, Nasser A, Thomsen SI, Eghorn LF, Pham Y, Schulenburg C, Syberg S, Ding M, Stojilkovic SS, Jorgensen NR, Heegaard AM: P2X7 receptor-deficient mice are susceptible to bone cancer pain. Pain 2011; 152:1766–76
9. Pevida M, González-Rodríguez S, Lastra A, García-Suárez O, Hidalgo A, Menéndez L, Baamonde A: Involvement of spi-nal chemokine CCL2 in the hyperalgesia evoked by bone cancer in mice: A role for astroglia and microglia. Cell Mol Neurobiol 2014; 34:143–56
10. Bu H, Shu B, Gao F, Liu C, Guan X, Ke C, Cao F, Hinton AO Jr, Xiang H, Yang H, Tian X, Tian Y: Spinal IFN-γ-induced pro-tein-10 (CXCL10) mediates metastatic breast cancer-induced bone pain by activation of microglia in rat models. Breast Cancer Res Treat 2014; 143:255–63
11. Shen W, Hu XM, Liu YN, Han Y, Chen LP, Wang CC, Song C: CXCL12 in astrocytes contributes to bone cancer pain through CXCR4-mediated neuronal sensitization and glial activation in rat spinal cord. J Neuroinflammation 2014; 11:75
12. Huang ZX, Lu ZJ, Ma WQ, Wu FX, Zhang YQ, Yu WF, Zhao ZQ: Involvement of RVM-expressed P2X7 receptor in bone cancer pain: Mechanism of descending facilitation. Pain 2014; 155:783–91
13. Gruzman A, Babai G, Sasson S: Adenosine monophosphate-activated protein kinase (AMPK) as a new target for anti-diabetic drugs: A review on metabolic, pharmacological and chemical considerations. Rev Diabet Stud 2009; 6:13–36
14. Chen CC, Lin JT, Cheng YF, Kuo CY, Huang CF, Kao SH, Liang YJ, Cheng CY, Chen HM: Amelioration of LPS-induced inflam-mation response in microglia by AMPK activation. Biomed Res Int 2014; 2014:692061
15. Han Y, Jiang C, Tang J, Wang C, Wu P, Zhang G, Liu W, Jamangulova N, Wu X, Song X: Resveratrol reduces morphine tolerance by inhibiting microglial activation via AMPK sig-nalling. Eur J Pain 2014; 18:1458–70
16. Lihn AS, Pedersen SB, Lund S, Richelsen B: The anti-diabetic AMPK activator AICAR reduces IL-6 and IL-8 in human adi-pose tissue and skeletal muscle cells. Mol Cell Endocrinol 2008; 292:36–41
17. Tillu DV, Melemedjian OK, Asiedu MN, Qu N, De Felice M, Dussor G, Price TJ: Resveratrol engages AMPK to attenuate ERK and mTOR signaling in sensory neurons and inhibits incision-induced acute and chronic pain. Mol Pain 2012; 8:5
18. Taylor A, Westveld AH, Szkudlinska M, Guruguri P, Annabi E, Patwardhan A, Price TJ, Yassine HN: The use of metformin is associated with decreased lumbar radiculopathy pain. J Pain Res 2013; 6:755–63
19. Melemedjian OK, Asiedu MN, Tillu DV, Sanoja R, Yan J, Lark A, Khoutorsky A, Johnson J, Peebles KA, Lepow T, Sonenberg N, Dussor G, Price TJ: Targeting adenosine monophosphate-activated protein kinase (AMPK) in preclinical models reveals a potential mechanism for the treatment of neuropathic pain. Mol Pain 2011; 7:70
20. Russe OQ, Möser CV, Kynast KL, King TS, Stephan H, Geisslinger G, Niederberger E: Activation of the AMP-activated protein kinase reduces inflammatory nociception. J Pain 2013; 14:1330–40
21. Liu S, Liu WT, Liu YP, Dong HL, Henkemeyer M, Xiong LZ, Song XJ: Blocking EphB1 receptor forward signaling in spi-nal cord relieves bone cancer pain and rescues analgesic effect of morphine treatment in rodents. Cancer Res 2011; 71:4392–402
22. Medhurst SJ, Walker K, Bowes M, Kidd BL, Glatt M, Muller M, Hattenberger M, Vaxelaire J, O’Reilly T, Wotherspoon G, Winter J, Green J, Urban L: A rat model of bone cancer pain. Pain 2002; 96:129–40
23. Kim HJ, Magrané J: Isolation and culture of neurons and astrocytes from the mouse brain cortex. Methods Mol Biol 2011; 793:63–75
24. Wang CC, Fang KM, Yang CS, Tzeng SF: Reactive oxygen species-induced cell death of rat primary astrocytes through mitochondria-mediated mechanism. J Cell Biochem 2009; 107:933–43
25. Hu LF, Lu M, Tiong CX, Dawe GS, Hu G, Bian JS: Neuroprotective effects of hydrogen sulfide on Parkinson’s disease rat models. Aging Cell 2010; 9:135–46
26. Srinivasan D, Yen JH, Joseph DJ, Friedman W: Cell type-spe-cific interleukin-1beta signaling in the CNS. J Neurosci 2004; 24:6482–8
Downloaded From: http://anesthesiology.pubs.asahq.org/pdfaccess.ashx?url=/data/journals/jasa/934601/ on 09/16/2018

Copyright © 2015, the American Society of Anesthesiologists, Inc. Wolters Kluwer Health, Inc. Unauthorized reproduction of this article is prohibited.
Anesthesiology 2015; 123:1170-85 1185 Song et al.
PAIN MEDICINE
27. Xuan A, Long D, Li J, Ji W, Zhang M, Hong L, Liu J: Hydrogen sulfide attenuates spatial memory impairment and hippocam-pal neuroinflammation in β-amyloid rat model of Alzheimer’s disease. J Neuroinflammation 2012; 9:202
28. Wang X, Loram LC, Ramos K, de Jesus AJ, Thomas J, Cheng K, Reddy A, Somogyi AA, Hutchinson MR, Watkins LR, Yin H: Morphine activates neuroinflammation in a manner parallel to endotoxin. Proc Natl Acad Sci USA 2012; 109:6325–30
29. Blasi E, Barluzzi R, Bocchini V, Mazzolla R, Bistoni F: Immortalization of murine microglial cells by a v-raf/v-myc carrying retrovirus. J Neuroimmunol 1990; 27:229–37
30. Henn A, Lund S, Hedtjärn M, Schrattenholz A, Pörzgen P, Leist M: The suitability of BV2 cells as alternative model sys-tem for primary microglia cultures or for animal experiments examining brain inflammation. ALTEX 2009; 26:83–94
31. Wang G, Amato S, Gilbert J, Man HY: Resveratrol up-regu-lates AMPA receptor expression via AMP-activated protein kinase-mediated protein translation. Neuropharmacology 2015; 95:144–53
32. Guan X, Fu Q, Xiong B, Song Z, Shu B, Bu H, Xu B, Manyande A, Cao F, Tian Y: Activation of PI3Kγ/Akt pathway mediates bone cancer pain in rats. J Neurochem 2015; 134:590–600
33. Taves S, Berta T, Chen G, Ji RR: Microglia and spinal cord synaptic plasticity in persistent pain. Neural Plast 2013; 2013:753656
34. Clark AK, Old EA, Malcangio M: Neuropathic pain and cyto-kines: Current perspectives. J Pain Res 2013; 6:803–14
35. Kuhn TB: Oxygen radicals elicit paralysis and collapse of spinal cord neuron growth cones upon exposure to proin-flammatory cytokines. Biomed Res Int 2014; 2014:191767
36. Wang LN, Yang JP, Zhan Y, Ji FH, Wang XY, Zuo JL, Xu QN: Minocycline-induced reduction of brain-derived neuro-trophic factor expression in relation to cancer-induced bone pain in rats. J Neurosci Res 2012; 90:672–81
37. Liu X, Bu H, Liu C, Gao F, Yang H, Tian X, Xu A, Chen Z, Cao F, Tian Y: Inhibition of glial activation in rostral ventromedial medulla attenuates mechanical allodynia in a rat model of cancer-induced bone pain. J Huazhong Univ Sci Technolog Med Sci 2012; 32:291–8
38. Mongey AB, Hess EV: Drug insight: Autoimmune effects of medications—What’s new? Nat Clin Pract Rheumatol 2008; 4:136–44
39. Ochsendorf F: Minocycline in acne vulgaris: Benefits and risks. Am J Clin Dermatol 2010; 11:327–41
40. Shirwany NA, Zou MH: AMPK: A cellular metabolic and redox sensor. A minireview. Front Biosci (Landmark Ed) 2014; 19:447–74
41. Hardie DG, Ross FA, Hawley SA: AMPK: A nutrient and energy sensor that maintains energy homeostasis. Nat Rev Mol Cell Biol 2012; 13:251–62
42. Vakana E, Altman JK, Glaser H, Donato NJ, Platanias LC: Antileukemic effects of AMPK activators on BCR-ABL-expressing cells. Blood 2011; 118:6399–402
43. Baron SJ, Li J, Russell RR III, Neumann D, Miller EJ, Tuerk R, Wallimann T, Hurley RL, Witters LA, Young LH: Dual mecha-nisms regulating AMPK kinase action in the ischemic heart. Circ Res 2005; 96:337–45
44. Dasgupta B, Milbrandt J: Resveratrol stimulates AMP kinase activity in neurons. Proc Natl Acad Sci USA 2007; 104:7217–22
45. Spasić MR, Callaerts P, Norga KK: AMP-activated protein kinase (AMPK) molecular crossroad for metabolic control and survival of neurons. Neuroscientist 2009; 15:309–16
46. Greco SJ, Sarkar S, Johnston JM, Tezapsidis N: Leptin regu-lates tau phosphorylation and amyloid through AMPK in neu-ronal cells. Biochem Biophys Res Commun 2009; 380:98–104
47. Greco SJ, Hamzelou A, Johnston JM, Smith MA, Ashford JW, Tezapsidis N: Leptin boosts cellular metabolism by activating AMPK and the sirtuins to reduce tau phosphorylation and β-amyloid in neurons. Biochem Biophys Res Commun 2011; 414:170–4
48. Kim HI, Kim TH, Song JH: Resveratrol inhibits Na+ currents in rat dorsal root ganglion neurons. Brain Res 2005; 1045:134–41
49. Liang L, Fan L, Tao B, Yaster M, Tao YX: Protein kinase B/Akt is required for complete Freund’s adjuvant-induced upreg-ulation of Nav1.7 and Nav1.8 in primary sensory neurons. J Pain 2013; 14:638–47
50. Xu Q, Fitzsimmons B, Steinauer J, O’Neill A, Newton AC, Hua XY, Yaksh TL: Spinal phosphinositide 3-kinase-Akt-mamma-lian target of rapamycin signaling cascades in inflammation-induced hyperalgesia. J Neurosci 2011; 31:2113–24
51. Wight RD, Tull CA, Deel MW, Stroope BL, Eubanks AG, Chavis JA, Drew PD, Hensley LL: Resveratrol effects on astrocyte function: Relevance to neurodegenerative diseases. Biochem Biophys Res Commun 2012; 426:112–5
52. Zhang F, Wang H, Wu Q, Lu Y, Nie J, Xie X, Shi J: Resveratrol protects cortical neurons against microglia-mediated neuro-inflammation. Phytother Res 2013; 27:344–9
53. Zhao XY, Yang S, Chen YR, Li PC, Dou MM, Zhang J: Resveratrol and arsenic trioxide act synergistically to kill tumor cells in vitro and in vivo. PLoS One 2014; 9:e98925
54. Chen HC, Bandyopadhyay G, Sajan MP, Kanoh Y, Standaert M, Farese RV Jr, Farese RV: Activation of the ERK pathway and atypical protein kinase C isoforms in exercise- and aminoimidazole-4-carboxamide-1-beta-D-riboside (AICAR)-stimulated glucose transport. J Biol Chem 2002; 277:23554–62
55. Li J, Miller EJ, Ninomiya-Tsuji J, Russell RR III, Young LH: AMP-activated protein kinase activates p38 mitogen-acti-vated protein kinase by increasing recruitment of p38 MAPK to TAB1 in the ischemic heart. Circ Res 2005; 97:872–9
56. Wang LN, Yao M, Yang JP, Peng J, Peng Y, Li CF, Zhang YB, Ji FH, Cheng H, Xu QN, Wang XY, Zuo JL: Cancer-induced bone pain sequentially activates the ERK/MAPK pathway in different cell types in the rat spinal cord. Mol Pain 2011; 7:48
57. Wang XW, Hu S, Mao-Ying QL, Li Q, Yang CJ, Zhang H, Mi WL, Wu GC, Wang YQ: Activation of c-jun N-terminal kinase in spinal cord contributes to breast cancer induced bone pain in rats. Mol Brain 2012; 5:21
58. Hu JH, Yang JP, Liu L, Li CF, Wang LN, Ji FH, Cheng H: Involvement of CX3CR1 in bone cancer pain through the activation of microglia p38 MAPK pathway in the spinal cord. Brain Res 2012; 1465:1–9
59. Cui Y, Liao XX, Liu W, Guo RX, Wu ZZ, Zhao CM, Chen PX, Feng JQ: A novel role of minocycline: Attenuating morphine antino-ciceptive tolerance by inhibition of p38 MAPK in the activated spinal microglia. Brain Behav Immun 2008; 22:114–23
60. Wen YR, Tan PH, Cheng JK, Liu YC, Ji RR: Microglia: A promis-ing target for treating neuropathic and postoperative pain, and morphine tolerance. J Formos Med Assoc 2011; 110:487–94
Downloaded From: http://anesthesiology.pubs.asahq.org/pdfaccess.ashx?url=/data/journals/jasa/934601/ on 09/16/2018


















