
A multiplex PCR assay, targeting a di-guanylate cyclase encoding ...
17
1 1 A multiplex PCR assay, targeting a di-guanylate cyclase encoding gene, cdgc1 to differentiate 2 species within the genus Cronobacter 3 4 L. Carter 1 , L. A. Lindsey 1 , C. J. Grim 1, 2 , V. Sathyamoorthy 1 , K. G. Jarvis 1 , G. Gopinath 1 , C. 5 Lee 1 , J. A. Sadowski 1 , L. Trach 1 , M. Pava-Ripoll 3 , B. A. McCardell 1 , B. D. Tall 1 and L. Hu 1* 6 7 CFSAN, FDA, 1 Laurel, 3 College Park, MD, 2 Oak Ridge Institute for Science and Education, Oak 8 Ridge, TN, 9 10 Running title: Cronobacter species-specific GGDEF multiplex PCR assay 11 Key words: Cronobacter, GGDEF, multiplex PCR assay, cyclic diguanylate 12 13 14 15 *Please send reviewers' comments and galley proofs to: Lan Hu, M.D., Ph.D. Room, 3408 MOD 16 1 Facility, Virulence Mechanisms Branch, (HFS-025), Division of Virulence Assessment, 17 OARSA, Center for Food Safety and Applied Nutrition, U. S. Food and Drug Administration, 18 8301 Muirkirk Rd, Laurel, Maryland 20708, Phone#: 301-210-6279; Fax#: 301-210-7976, Email 19 address: [email protected] 20 21 22 Copyright © 2012, American Society for Microbiology. All Rights Reserved. Appl. Environ. Microbiol. doi:10.1128/AEM.02898-12 AEM Accepts, published online ahead of print on 9 November 2012 on April 8, 2018 by guest http://aem.asm.org/ Downloaded from
Transcript of A multiplex PCR assay, targeting a di-guanylate cyclase encoding ...

1
1
A multiplex PCR assay, targeting a di-guanylate cyclase encoding gene, cdgc1 to differentiate 2
species within the genus Cronobacter 3
4
L. Carter1, L. A. Lindsey1, C. J. Grim1, 2, V. Sathyamoorthy1, K. G. Jarvis1, G. Gopinath1, C. 5
Lee1, J. A. Sadowski1, L. Trach1, M. Pava-Ripoll3, B. A. McCardell1, B. D. Tall1 and L. Hu1* 6
7
CFSAN, FDA, 1Laurel, 3College Park, MD, 2Oak Ridge Institute for Science and Education, Oak 8
Ridge, TN, 9
10
Running title: Cronobacter species-specific GGDEF multiplex PCR assay 11
Key words: Cronobacter, GGDEF, multiplex PCR assay, cyclic diguanylate 12
13
14
15
*Please send reviewers' comments and galley proofs to: Lan Hu, M.D., Ph.D. Room, 3408 MOD 16
1 Facility, Virulence Mechanisms Branch, (HFS-025), Division of Virulence Assessment, 17
OARSA, Center for Food Safety and Applied Nutrition, U. S. Food and Drug Administration, 18
8301 Muirkirk Rd, Laurel, Maryland 20708, Phone#: 301-210-6279; Fax#: 301-210-7976, Email 19
address: [email protected] 20
21
22
Copyright © 2012, American Society for Microbiology. All Rights Reserved.Appl. Environ. Microbiol. doi:10.1128/AEM.02898-12 AEM Accepts, published online ahead of print on 9 November 2012
on April 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

2
Abstract 23
Compared to the widely used Cronobacter rpoB-PCR assay, a highly specific 24
multiplexed PCR assay, based on cdgc1, a di-guanylate cyclase gene was designed that 25
identified all of the targeted six species among 305 Cronobacter isolates. This assay will be a 26
valuable tool for identifying suspected Cronobacter isolates from foodborne investigations.27
on April 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

3
Cronobacter spp. are Gram-negative, opportunistic pathogens that cause meningitis, 28
necrotizing enterocolitis, and septicemia in neonates and elderly individuals (3, 14, 20). 29
Cronobacter are ubiquitous in nature, and have been isolated from clinical, environmental and 30
from food sources, most notably powdered infant formula and other dried foods (14, 20) and 31
more recently, from surfaces and intestinal tracts of wild filth flies (15). The Cronobacter genus 32
consists of seven species: C. condimenti, C. dublinensis, C. malonaticus, C. muytjensii, C. 33
sakazakii, C. turicensis, and C. universalis (6, 8). Although all species, except C. condimenti 34
have been associated with clinical infections, C. sakazakii and C. malonaticus isolates are 35
responsible for causing the majority of infantile illnesses (9). It is important to identify 36
Cronobacter quickly and precisely. Current Cronobacter identification and subtyping methods 37
include 16S rDNA gene sequencing, ribotyping, DNA-DNA hybridization, rpoB PCR, Pulsed-38
field gel electrophoresis, plasmidotyping, and molecular serogrouping assays (1, 2, 6, 7, 13, 18, 39
20). These methods have detected considerable diversity among Cronobacter, however many of 40
these are not rapid or require multiple PCR reactions to identify or characterize isolates. Also, 41
16S rDNA sequence analysis has limitations for discriminating between very closely related 42
organisms, such as C. malonaticus and C. sakazakii, because of minimal sequence diversity or 43
the presence of multiple copies of 16S rDNA loci. It is necessary to ensure that reliable and 44
robust identification methods are used so that the control of contamination by these organisms 45
during the food manufacturing process and the reduction of exposure to susceptible high risk 46
individuals are achieved. 47
Cyclic di-guanylate (c-di-GMP) is a bacterial signal-transduction second messenger 48
recognized for its involvement in the regulation of a number of complex physiological processes, 49
including bacterial virulence, biofilm formation, and persistence (long-term survival) (16). Di-50
on April 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

4
guanylate cyclase, which synthesizes cyclic-di-guanylate, possesses a conserved active domain 51
of five amino acids, Gly-Gly-Asp-Glu-Phe, or GGDEF (16). Several foodborne pathogens such 52
as Vibrio cholerae, Salmonella, and E. coli encode variable numbers of GGDEF domain proteins 53
(16). 54
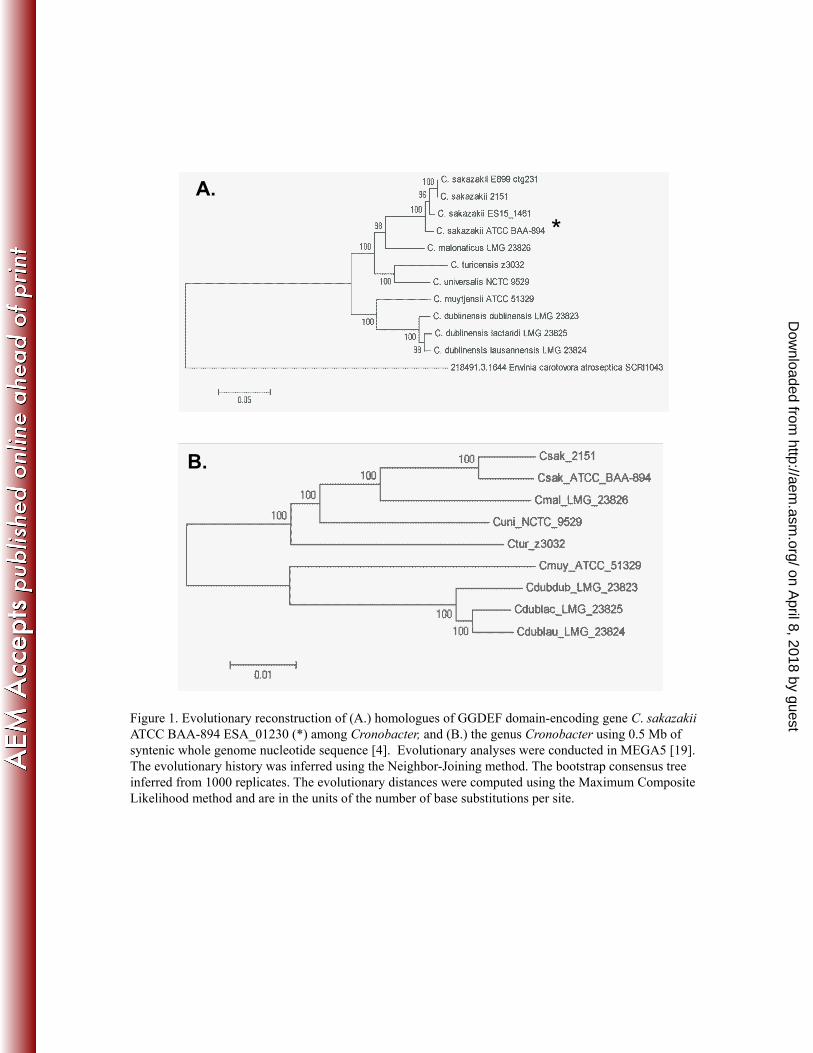
Analysis of 12 Cronobacter genomes revealed seven GGDEF-domain encoding genes, 55
which were conserved among all Cronobacter (5, 12, 17). Phylogenetic analysis of each set of 56
homologous genes from the available genomes revealed that homologs of two of the seven 57
genes, (ESA_04212 and ESA_03399 of C. sakazakii ATCC BAA-894) were highly similar 58
within the genus, and, one of these, within the Enterobacteriaceae (data not shown). In contrast, 59
the other five GGDEF domain-encoding genes (homologs of ESA_01230, ESA_01822, 60
ESA_03401, ESA_03491, and ESA_ 04315 from C. sakazakii ATCC BAA-894) showed 61
species-specific allelic divergence (Fig. 1A), some of which recapitulated the species-specific 62
phylogenetic relationships within the genus (Fig. 1B), previously described by Iversen et al. (6), 63
and as determined through MLST (10), rpoB sequencing analyses (18), and whole genome 64
phylogenetic reconstruction (5). 65
In particular, multiple sequence alignment of homologues of ESA_01230 from C. 66
sakazakii ATCC BAA-894 (Fig. 1A), annotated as a putative Cronobacter di-guanylate cyclase 67
(containing a GGDEF domain) and designated here as cdgc1, yielded a phylogeny in which all 68
six species formed discrete, distinct lineages, with relationships in agreement with whole genome 69
analysis (Fig. 1B). Further, we hypothesized that this gene would be amenable to species-70
specific primer design, based on length of the coding sequences and the depth of branching 71
exhibited between species, which, taken together, would provide a significant number of variable 72
sites (SNPs) for species-specific primer design (Supplemental Figure 1). 73
on April 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

5
These observations led us to design a set of species-specific multiplex PCR primers, 74
which could be used to identify strains of Cronobacter in a single multiplex PCR assay (Table 75
1). Primer sequences were chosen by multiple alignment analysis of the respective cgdg1 76
sequences using MEGA5 (19). The primer sequences were evaluated for their ability to form 77
homo- and heterodimers as well as hairpins by using oligo-analyzer 3.1 (Integrated DNA 78
Technologies, Coralville, IA). All primers were synthesized by (Integrated DNA Technologies). 79
Empirical primer concentration and PCR optimization studies were performed by changing 80
primer and template concentrations and PCR assay parameter conditions so that optimal kinetics 81
were achieved. 82
PCR mixtures were prepared using the GoTaq Green master mix (Promega Corp., 83
Madison, WI) according to the manufacturer’s instructions using boiled genomic DNA 84
preparations (approximately 50 ng DNA/25-μl reaction mixture) as DNA template (11). 85
GoTaq® Hot Start Green Master Mix (a–c) is a premixed, proprietary, ready-to-use solution 86
containing GoTaq® Hot Start Polymerase, dNTPs, MgCl2 and reaction buffers at optimal 87
concentrations for efficient amplification of DNA templates by PCR. GoTaq® Hot Start 88
Polymerase is supplied in 2X Green GoTaq® Reaction Buffer (pH 8.5), 400 μM dATP, 400 μM 89
dGTP, 400 μM dCTP, 400 μM dTTP and 4mM MgCl2. In all PCR reactions, the polymerase 90
was activated by using a 3-min predenaturation step at 94°C, followed by 25 cycles of 91
denaturation at 94°C for 30 s, annealing of 62°C for 30 s, and one min extension at 72°C, 92
following by a final extension step of 72°C for five min. PCR amplicons were subjected to 93
agarose gel electrophoresis using 1.3% Tris-borate-EDTA (TBE, Invitrogen, Carlsbad, CA) 94
agarose gels in a RunOne (Embi Tec, San Diego, CA) horizontal electrophoresis unit and were 95
on April 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

6
photographed with trans-illuminated UV light using a Bio-Rad Molecular Imager Gel-Chem-Doc 96
XR imaging system (Bio-Rad Laboratories, Hercules, CA). 97
Examples of the cdgc1 PCR assay results for type strains of Cronobacter are shown in 98
Fig. 3. Because C. condimenti is recently described and the taxonomic description is based on a 99
single isolate, this species was not included in these studies. The multiplex PCR assay was 100
evaluated by interrogating a collection of 305 well characterized Cronobacter strains (2, 4-7, 11), 101
which included 15 strains of C. dublinensis, two strains of C. universalis, 12 strains of C. 102
muytjensii, 11 strains of C. turicensis, 231 strains of C. sakazakii, and 34 strains of C. 103
malonaticus. The strain collection included isolates from clinical (69 strains), food (144 strains), 104
environmental (63 strains), flies (18 strains) and unknown (11 strains) sources which were 105
obtained from diverse geographic locations worldwide. These isolates were initially 106
characterized biochemically according to Iversen et al. (6), and later confirmed using the species-107
specific rpoB PCR assays as described by Stoop et al. (18). Additionally, this collection of 108
isolates was subjected to repF1B “plasmidotyping” (2) and molecular serogrouping (7, 13), the 109
results of which further corroborated the rpoB-based PCR species identification for each strain. 110
The multiplex cdgc1 PCR assay correctly identified (100%) the species identity of the 305 111
Cronobacter isolates. These results confirm the species-specificity of the cdgc1 multiplex PCR 112
assay. To test for exclusivity, twenty non-Cronobacter strains, which included Enterobacter 113
amnigenus, E. aerogenes, E. gergoviae, E. asburiae, E. cancerogenus, E. cloacae, E. cloacae 114
subsp. dissolvans, E. helveticus, E. pulveris, and E. turicencis, Citrobacter koseri, Pantoea 115
agglomerans, Erwinia carotovora, Kluyvera intermedia, Listeria monocytogenes, Escherichia 116
coli O157:H7, and Salmonella enterica subsp. enterica serovar Enteritidis, were assayed and all 117
were negative using these PCR primers. 118
on April 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

7
In conclusion this study demonstrates that the Cronobacter multiplex cdgc1 PCR assay 119
can be used to identify Cronobacter strains in a single reaction. The PCR assay described in this 120
report was found to be 100% specific (305/305 correctly identified as Cronobacter spp. strains) 121
and 100% sensitive (did not identify 20/20 non-Cronobacter species). Its main advantage over 122
the currently used rpoB PCR method is that species-identity of C. sakazakii and C. malonaticus 123
can be accomplished in a single reaction as opposed to two separate PCR reactions. The assay 124
reported here will be a valuable tool for identifying suspected Cronobacter isolates from clinical, 125
environmental, and foodborne outbreak and surveillance investigations, quickly and precisely. 126
127
Acknowledgements 128
L. A. Lindsey was a recipient of a 2011 National Association for Equal Opportunity in Higher 129
Education internship program fellowship, Washington, DC and is now a graduate student 130
researcher in the Department of Food and Nutritional Sciences at Tuskegee University in 131
Tuskegee, AL. C. Lee, L. Trach, and J. A. Sadowski were supported by Joint Institute of Food 132
Safety and Applied Nutrition student internship program, University of Maryland College Park. 133
134
on April 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

8
References 135 136
1. Brengi SP, O’Brien SB, Pichel M, Iversen C, Arduino M, Binsztein N, Jensen B, 137
Pagotto F, Ribot EM, Stephan R, Cernela N, Cooper K, Fanning S. 2012. 138
Development and validation of a PulseNet standardized protocol for subtyping isolates of 139
Cronobacter species. Foodborne Pathogen Dis. 9:861-867. 140
2. Franco AA, Hu L, Grim CJ, Gopinath G, Sathyamoorthy V, Jarvis KG, Lee C, 141
Sadowski J, Kim J, Kothary MH, McCardell BA, Tall BD. 2011. Characterization of 142
putative virulence genes on the related RepFIB plasmids harbored by Cronobacter spp. 143
Appl. Environ. Microbiol. 77:3255-3267. 144
3. Gosney MA, Martin MV, Wright AE, Gallagher M. 2006. Enterobacter sakazakii in 145
the mouths of stroke patients and its association with aspiration pneumonia. Eur. J. 146
Intern. Med. 17:185-188. 147
4. Grim CJ, Kothary MH, Gopinath G, Jarvis KG, Beaubrun JJ, McClelland M, Tall 148
BD, Franco AA. 2012. Identification and characterization of Cronobacter spp. iron 149
acquisition systems. Appl. Environ. Microbiol. 78:6035-6050. 150
5. Grim CJ, Kotewicz ML, Power KA, Franco AA, Gopinath G, Jarvis KG, 151
Yan QQ, Jackson SA, Hu L, Sathyamoorthy V, Pagotto F, Iversen C, Lehner A, 152
Stephan R, Fanning S, Tall BD. 2012. Genomic analysis of the pan genome of the 153
emerging bacterial foodborne pathogen Cronobacter spp. suggests a species-level 154
bidirectional divergence from a conserved genomic core. BMC Genomics (under review). 155
6. Iversen C, Mullane N, McCardell B, Tall BD, Lehner A, Fanning S, Stephan R, and 156
Joosten H. 2008. Cronobacter gen. nov., a new genus to accommodate the biogroups of 157
on April 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

9
Enterobacter sakazakii, and proposal of Cronobacter sakazakii gen. nov., comb. nov., 158
Cronobacter malonaticus sp. nov., Cronobacter turicensis sp. nov., Cronobacter 159
muytjensii sp. nov., Cronobacter dublinensis sp. nov., Cronobacter genomospecies 1, and 160
of three subspecies, Cronobacter dublinensis subsp. dublinensis subsp. nov., Cronobacter 161
dublinensis subsp. lausannensis subsp. nov. and Cronobacter dublinensis subsp. lactaridi 162
subsp. nov. Int. J. Syst. Evol. Microbiol. 58:1442-1447. 163
7. Jarvis KG, Grim CJ, Franco AA, Gopinath G, Sathyamoorthy V, Hu L, Sadowski 164
JA, Lee CS, Tall BD. 2011. Molecular Characterization of Cronobacter 165
Lipopolysaccharide O-Antigen Gene Clusters and Development of Serotype-Specific 166
PCR Assays. Appl. Environ. Microbiol. 77:4017-4026. 167
8. Joseph S, Cetinkaya E, Drahovska H, Ababneh QO, Levican A, Figueras MJ, 168
Forsythe SJ. 2011. Cronobacter condimenti sp. nov., isolated from spiced meat and 169
Cronobacter universalis sp. nov., a novel species designation for Cronobacter sp. 170
genomospecies 1 recovered from a leg infection water and food ingredients. Int. J. Sys. 171
Evol. Microbiol. 62:1277-1283. 172
9. Joseph S, Forsythe SJ. 2011. Predominance of Cronobacter sakazakii sequence type 4 173
in neonatal infections. Emerg. Infect. Dis. 17:1713-1715. 174
10. Joseph S, Sonbol H, Hariri S, Desai P, McClelland M, Forsythe SJ. 2012. Diversity of 175
the Cronobacter genus as revealed by multi locus sequence typing. J. Clin. Microbiol. 176
50:3031-3039. 177
11. Kothary MH, McCardell BA, Frazer CD, Deer D, and Tall BD. 2007. 178
Characterization of the zinc-containing metalloprotease (zpx) and development of a 179
on April 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

10
species-specific detection method for Enterobacter sakazakii. Appl. Environ. Microbiol. 180
73:4142–4151. 181
12. Kucerova E, Clifton SW, Xia XQ, Long F, Porwollik S, Fulton L, Fronick C, Minx 182
P, Kyung K, Warren W, Fulton R, Feng D, Wollam A, Shah N, Bhonagiri V, Nash, 183
K. Hallsworth-Pepin WE, Wilson RK, McClelland M, Forsythe SJ 2010. Genome 184
sequence of Cronobacter sakazakii BAA-894 and comparative genomic hybridization 185
analysis with other Cronobacter species. PLoS One. 5:e9556. 186
13. Mullane N, O'Gaora P, Nally JE, Iversen C, Whyte P, Wall PG, Fanning S. 2008. 187
Molecular analysis of the Enterobacter sakazakii O-antigen gene locus. Appl. Environ. 188
Microbiol. 74:3783-3794. 189
14. Nazarowec-White M, Farber JM. 1997. Enterobacter sakazakii: a review. Int. J. Food 190
Microbiol. 34:103-113. 191
15. Pava-Ripoll M, Pearson REG, Miller AK, Ziobro GC. 2012. Prevalence and relative 192
risk of Cronobacter spp., Salmonella spp. and Listeria monocytogenes associated with 193
the body surface and the guts of individual filth flies. Appl. Environ. Microbiol. 78:7891-194
7902. 195
16. Sondermann H, Shikuma NJ, Yildiz FH. 2012. You’ve come a long way: c-di-GMP 196
signaling. Curr. Opin. Microbiol. 15:140-146. 197
17. Stephan R, Lehner A, Tischler P, Rattei T. 2011. Complete genome sequence of 198
Cronobacter turicensis LMG 23827, a food-borne pathogen causing deaths in neonates. 199
J. Bacteriol. 193:309-310. 200
18. Stoop B, Lehner A, Iversen C, Fanning S, Stephan R. 2009. Development and 201
on April 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

11
evaluation of rpoB based PCR systems to differentiate the six proposed species within the 202
genus Cronobacter. Int. J. Food Microbiol. 136:165-168. 203
19. Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. 2011. MEGA5: 204
molecular evolutionary genetics analysis using maximum likelihood, evolutionary 205
distance, and maximum parsimony methods. Mol. Biol. Evol. 28:2731-2739. 206
20. Yan QQ, Condell O, Power KA, Butler F, Tall BD, Fanning S. 2012. Cronobacter 207
species (formerly known as Enterobacter sakazakii) in powdered infant formula: a 208
review of our current understanding of the biology of this bacterium. J. Appl. Microbiol. 209
113:1-15. 210
on April 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

12
Table 1. Cronobacter species-specific cdgc1 PCR primers used in this study 211
212
*The PCR primer, Cdm-469R, used in multiplex reactions, with primers Cdub-40F and Cmuy-213
209F identify C. dublinensis and C. muytjensii strains, respectively. PCR primer Cmstu-825F, 214
used in multiplex reactions, with primers Ctur-1036R, Cuni-1133R, Csak-1317R and Cmal-215
1410R identify C. malonaticus, C. sakazakii, C. turicensis, and C. universalis, strains, 216
respectively. The numbers comprising each primer name represents the 5’ bp location within the 217
aligned nucleotide sequence of cdgc1 (Supplemental Figure 1). 218
219
Primers Sequence Amplicon (bp) Species ID
Cdm-469R* CCACATGGCCGATATGCACGCC
Cdub-40F GATACCTCTCTGGGCCGCAGC 430 C. dublinensis
Cmuy-209F TTCTTCAGGCGGAGCTGACCT 260 C. muytjensii
Cmstu-825F* GGTGGCSGGGTATGACAAAGAC
Ctur-1036R TCGCCATCGAGTGCAGCGTAT 211 C. turicensis
Cuni-1133R GAAACAGGCTGTCCGGTCACG 308 C. universalis
Csak-1317R GGCGGACGAAGCCTCAGAGAGT 492 C. sakazakii
Cmal-1410R GGTGACCACACCTTCAGGCAGA 585 C. malonaticus
on April 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

13
220
221
Figure 1. Evolutionary reconstruction of (A.) homologues of GGDEF domain-encoding gene C. 222
sakazakii ATCC BAA-894 ESA_01230 (*) among Cronobacter, and (B.) the genus Cronobacter 223
using 0.5 Mb of syntenic whole genome nucleotide sequence (4). Evolutionary analyses were 224
conducted in MEGA5 (19). The evolutionary history was inferred using the Neighbor-Joining 225
method. The bootstrap consensus tree inferred from 1000 replicates. The evolutionary distances 226
were computed using the Maximum Composite Likelihood method and are in the units of the 227
number of base substitutions per site. 228
Figure 2. Schematic map of cdgc1 showing locations of PCR primers. Smallest tick of the scale 229
represents 50 bp nucleotide positions with larger ticks representing 100 bp and 1kbp nucleotide 230
positions. See Table 1 for explanation of primer names and nucleotide bp positions. 231
Figure 3. Representative gel image of Cronobacter species-specific GGDEF multiplex PCR. 232
Lanes 1 and 10, TrackIt 100 bp DNA Ladder (Invitrogen); lanes 2, C. dublinensis strain LMG 233
23823T; lane 3, C. universalis strain NCTC 9529; lane 4, C. muytjensii strain ATCC 51329; lane 234
5, C. malonaticus strain LMG 23826T; lane 6, C. sakazakii strains BAA-894; lane 7, C. 235
turicensis strain z3032; lane 8, Enterobacter helveticus strain z513, and lane 9, no DNA template 236
control. Five µl of each PCR reaction (amplicons) was subjected to gel electrophoresis using 237
1.5% agarose gels and were visualized with ethidium bromide (at a final concentration of 0.5 238
µg/ml). 239
240
on April 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

14
Supplemental Figure 1. Multiple alignment of cdcg1 of the different targeted species with 241
marked primer sequence (underlined). 242
243
on April 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from
A.
**
B.
Figure 1. Evolutionary reconstruction of (A.) homologues of GGDEF domain-encoding gene C. sakazakiiATCC BAA-894 ESA_01230 (*) among Cronobacter, and (B.) the genus Cronobacter using 0.5 Mb of syntenic whole genome nucleotide sequence [4]. Evolutionary analyses were conducted in MEGA5 [19]. The evolutionary history was inferred using the Neighbor-Joining method. The bootstrap consensus tree inferred from 1000 replicates. The evolutionary distances were computed using the Maximum Composite Likelihood method and are in the units of the number of base substitutions per site.
on April 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

cdgc1
1 1542
Figure 2. Schematic map of cdgc1showing locations of PCR primers. Smallest tick of the scale represents 50 bp nucleotide positions with larger ticks representing 100 bp p p p g p g pand 1kbp nucleotide positions. See Table 1 for explanation of primer names and nucleotide bp positions.
on April 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from

1 2 3 4 5 6 7 8 109
500 bp500bp
Figure 3. Representative gel image of Cronobacter species-specific GGDEF multiplex PCR. Lanes 1 and 10, TrackIt 100 bp DNA Ladder (Invitrogen); lanes 2, C. dublinensis strain LMG 23823T; lane 3, , p ( g ); , ; ,C. universalis strain NCTC 9529; lane 4, C. muytjensii strain ATCC 51329; lane 5, C. malonaticus strain LMG 23826T; lane 6, C. sakazakii strains BAA-894; lane 7, C. turicensis strain z3032; lane 8, Enterobacter helveticus strain z513, and lane 9, no DNA template control. Five µl of each PCR reaction (amplicons) was subjected to gel electrophoresis using 1.5% agarose gels and were visualized with ethidium bromide (at a final concentration of 0.5 µg/ml).
on April 8, 2018 by guest
http://aem.asm
.org/D
ownloaded from


















