
41 How Plants Grow in Response to Their Environment
30
807 41 How Plants Grow in Response to Their Environment Concept Outline 41.1 Plant growth is often guided by environmental cues. Tropisms. Plant growth is often influenced by light, gravity, and contact with other plants and animals. Dormancy. The ability to cease growth allows plants to wait out the bad times. 41.2 The hormones that guide growth are keyed to the environment. Plant Hormones. Hormones are grouped into seven classes. Auxin. Auxin is involved in the elongation of stems. Cytokinins. Cytokinins stimulate cell division. Gibberellins. Gibberellins control stem elongation. Brassinosteroids and Oligosaccharins. There are several recent additions to the plant hormone family. Ethylene. Ethylene controls leaves and flower abscision. Abscisic Acid. Abscisic acid suppresses growth of buds and promotes leaf senescence. 41.3 The environment influences flowering. Plants Undergo Metamorphosis. The transition of a shoot meristem from vegetative to adult is called phase change. Pathways Leading to Flower Production. Photoperiod is regulated in complex ways. Identity Genes and the Formation of Floral Meristems and Floral Organs. Floral meristem identity genes activate floral organ identity genes. 41.4 Many short-term responses to the environment do not require growth. Turgor Movement. Changes in the water pressure within plant cells result in quick and reversible plant movements. Plant Defense Responses. In addition to generalized defense mechanisms, some plants have highly evolved recognition mechanisms for specific pathogens. A ll organisms sense and interact with their environ- ment. This is particularly true of plants. Plant survival and growth is critically influenced by abiotic factors includ- ing water, wind, and light. In this chapter, we will explore how a plant senses such factors, and transduces these sig- nals to elicit an optimal physiological, growth, or develop- mental response. Hormones play an important role in the internal signaling that brings about environmental re- sponses, and is keyed in many ways to the environment. The effect of the local environment on plant growth also accounts for much of the variation in adult form within a species (figure 41.1). Precisely regulated responses to the environment not only allow a plant to survive from day to day but also determine when a flowering plant will produce a flower. The entire process of constructing a flower in turn sets the stage for intricate reproductive strategies that will be discussed in the next chapter. FIGURE 41.1 Plant growth is affected by environmental cues. The branches of this fallen tree are growing straight up in response to gravity and light.
Transcript of 41 How Plants Grow in Response to Their Environment

807
41How Plants Grow in
Response to TheirEnvironment
Concept Outline
41.1 Plant growth is often guided by environmentalcues.
Tropisms. Plant growth is often influenced by light,gravity, and contact with other plants and animals.Dormancy. The ability to cease growth allows plants towait out the bad times.
41.2 The hormones that guide growth are keyed tothe environment.
Plant Hormones. Hormones are grouped into sevenclasses.Auxin. Auxin is involved in the elongation of stems.Cytokinins. Cytokinins stimulate cell division.Gibberellins. Gibberellins control stem elongation.Brassinosteroids and Oligosaccharins. There areseveral recent additions to the plant hormone family.Ethylene. Ethylene controls leaves and flower abscision.Abscisic Acid. Abscisic acid suppresses growth of budsand promotes leaf senescence.
41.3 The environment influences flowering.
Plants Undergo Metamorphosis. The transition of ashoot meristem from vegetative to adult is called phasechange.Pathways Leading to Flower Production. Photoperiodis regulated in complex ways.Identity Genes and the Formation of Floral Meristemsand Floral Organs. Floral meristem identity genesactivate floral organ identity genes.
41.4 Many short-term responses to the environmentdo not require growth.
Turgor Movement. Changes in the water pressurewithin plant cells result in quick and reversible plantmovements. Plant Defense Responses. In addition to generalizeddefense mechanisms, some plants have highly evolvedrecognition mechanisms for specific pathogens.
All organisms sense and interact with their environ-ment. This is particularly true of plants. Plant survival
and growth is critically influenced by abiotic factors includ-ing water, wind, and light. In this chapter, we will explorehow a plant senses such factors, and transduces these sig-nals to elicit an optimal physiological, growth, or develop-mental response. Hormones play an important role in theinternal signaling that brings about environmental re-sponses, and is keyed in many ways to the environment.The effect of the local environment on plant growth alsoaccounts for much of the variation in adult form within aspecies (figure 41.1). Precisely regulated responses to theenvironment not only allow a plant to survive from day today but also determine when a flowering plant will producea flower. The entire process of constructing a flower inturn sets the stage for intricate reproductive strategies thatwill be discussed in the next chapter.
FIGURE 41.1Plant growth is affected by environmental cues. The branchesof this fallen tree are growing straight up in response to gravityand light.

given biological reaction that is affected by phytochromewill occur. When most of the Pfr has been replaced by Pr,the reaction will not occur (figure 41.3). While we referto phytochrome as a single molecule here, it is importantto note that several different phytochromes have nowbeen identified that appear to have specific biologicalfunctions.
Phytochrome is a light receptor, but it does not act di-rectly to bring about reactions to light. The existence ofphytochrome was conclusively demonstrated in 1959 byHarry A. Borthwick and his collaborators at the U.S. De-partment of Agriculture Research Center at Beltsville,Maryland. It has since been shown that the molecule con-sists of two parts: a smaller one that is sensitive to lightand a larger portion that is a protein. The protein compo-nent initiates a signal transduction leading to a particulartropism. The phytochrome pigment is blue, and its light-sensitive portion is similar in structure to the phycobilinsthat occur in cyanobacteria and red algae. Phytochrome ispresent in all groups of plants and in a few genera ofgreen algae, but not in bacteria, fungi, or protists (otherthan the few green algae). It is likely that phytochromesystems for measuring light evolved among the greenalgae and were present in the common ancestor of theplants.
Phytochrome is involved in many plant growth re-sponses. For example, seed germination is inhibited by far-red light and stimulated by red light in many plants. Be-cause chlorophyll absorbs red light strongly but does notabsorb far-red light, light passing through green leaves in-hibits seed germination. Consequently, seeds on the
808 Part XI Plant Growth and Reproduction
TropismsGrowth patterns in plants are often guided by environ-mental signals. Tropisms (from trope, the Greek word for“turn”) are positive or negative growth responses of plantsto external stimuli that usually come from one direction.Some responses occur independently of the direction ofthe stimuli and are referred to as nastic movements. Forexample, a tendril of a pea plant will always coil in one di-rection when touched. Tropisms, on the other hand, aredirectional and offer significant compensation for theplant’s inability to get up and walk away from unfavorableenvironmental conditions. Tropisms contribute the vari-ety of branching patterns we see within a species. Here wewill consider three major classes of plant tropisms: pho-totropism, gravitropism, and thigmotropism. Tropismsare particularly intriguing because they challenge us toconnect environmental signals with cellular perception ofthe signal, transduction into biochemical pathways, andultimately an altered growth response.
Phototropism
Phototropic responses involve the bending of growingstems and other plant parts toward sources of light (figure41.2). In general, stems are positively phototropic, growingtoward a light source, while most roots do not respond tolight or, in exceptional cases, exhibit only a weak negativephototropic response. The phototropic reactions of stemsare clearly of adaptive value, giving plants greater exposureto available light. They are also important in determiningthe development of plant organs and, therefore, the ap-pearance of the plant. Individual leaves may display pho-totropic responses. The position of leaves is important tothe photosynthetic efficiency of the plant. A plant hormonecalled auxin (discussed later in this chapter) is probably in-volved in most, if not all, of the phototropic growth re-sponses of plants.
The first step in a phototropic response is perceivingthe light. Photoreceptors perceive different wavelengthsof light with blue and red being the most common. Bluelight receptors are being characterized and we are begin-ning to understand how plants “see blue.” Much more isknown about “seeing red” and translating that perceptioninto a signal transduction pathway leading to an alteredgrowth response. Plants contain a pigment, phy-tochrome, which exists in two interconvertible forms, Prand Pfr. In the first form, phytochrome absorbs red light;in the second, it absorbs far-red light. When a moleculeof Pr absorbs a photon of red light (660 nm), it is instantlyconverted into a molecule of Pfr, and when a molecule ofPfr absorbs a photon of far-red light (730 nm), it is in-stantly converted to Pr. Pfr is biologically active and Pr isbiologically inactive. In other words, when Pfr is present, a
41.1 Plant growth is often guided by environmental cues.
FIGURE 41.2Phototropism. Impatiens plant growing toward light.

ground under deciduous plants that lose their leaves inwinter are more apt to germinate in the spring after theleaves have decomposed and the seedlings are exposed todirect sunlight. This greatly improves the chances theseedlings will become established.
A second example of these relationships is the elonga-tion of the shoot in an etiolated seedling (one that is paleand slender from having been kept in the dark). Such plantsbecome normal when exposed to light, especially red light,but the effects of such exposure are canceled by far-redlight. This indicates a relationship similar to that observedin seed germination. There appears to be a link betweenphytochrome light perception and brassinosteroids in theetiolation response. Etiolation is an energy conservationstrategy to help plants growing in the dark reach the lightbefore they die. They don't green-up until there is light,and they divert energy to growing as tall as possiblethrough internode elongation. The de-etiolated (det2) Ara-bidopsis mutant has a poor etiolation response. It does nothave elongated internodes and greens up a bit in the dark.It turns out that det2 mutants are defective in an enzymenecessary for brassinosteroid biosynthesis. Researchers sus-pect that brassinosteroids play a role in how plants respondto light through phytochrome. Thus, because det2 mutantslack brassinosteroids, they do not respond to light, or lackof light, as normal plants do, and the det2 mutants grownormally in the dark.
Red and far-red light also are used as signals for plantspacing. The closer plants are together, the more likelythey are to grow tall and try to outcompete others for thesunshine. Plants somehow measure the amount of far-redlight being bounced back to them from neighboring trees.If their perception is messed up by putting a collar aroundthe stem with a solution that blocks light absorption, theelongation response is no longer seen.
Gravitropism
When a potted plant is tipped over, the shoot bends andgrows upward (figure 41.4). The same thing happens whena storm pushes over plants in a field. These are examples ofgravitropism, the response of a plant to the gravitationalfield of the earth. We saw in chapter 40 that brown algaeorient their first cell division so the rhizoid grows down-ward. Rhizoids also develop away from a unilateral lightsource. Separating out phototropic effects is important inthe study of gravitropisms.
Gravitropic responses are present at germination whenthe root grows down and the shoot grows up. Why does ashoot have a negative gravitropic response (growth awayfrom gravity), while a root has a positive gravitropic re-sponse? The opportunity to experiment on the spaceshuttle in a gravity-free environment has accelerated re-search in this area. Auxins play a primary role in gravit-ropic responses, but they may not be the only way gravita-tional information is sent through the plant. When John
Glenn made his second trip into space, he was accompa-nied by an experiment designed to test the role of gravityand electrical signaling in root bending. Analysis of grav-itropic mutants is also adding to our understanding ofgravitropism. There are four steps that lead to a gravit-ropic response:
1. Gravity is perceived by the cell2. Signals form in the cell that perceives gravity3. The signal is transduced intra- and intercellularly4. Differential cell elongation occurs between cells in
the “up” and “down” sides of the root or shoot.
Chapter 41 How Plants Grow in Response to Their Environment 809
Far-red light(730 nm)
Biological responseis blocked
Pd
Red light(660 nm)
PhytochromePr
Long periodof darkness
Destruction
Synthesis
PhytochromePfr
PrecursorPp
FIGURE 41.3How phytochrome works. Phytochrome is synthesized in the Prform from amino acids, designated Pp for phytochrome precursor.When exposed to red light, Pr changes to Pfr, which is the activeform that elicits a response in plants. Pfr is converted to Pr whenexposed to far-red light, and it also converts to Pr or is destroyedin darkness. The destruction product is designated Pd.
FIGURE 41.4Plant response togravity. This plant(Zebrina pendula) wasplaced horizontally andallowed to grow for 7 days. Note thenegative gravitationalresponse of the shoot.

One of the first steps in perceiving gravity is that amy-loplasts, plastids that contain starch, sink toward the gravi-tational field. These may interact with the cytoskeleton,but the net effect is that auxin becomes more concentratedon the lower side of the stem axis than on the upper side.The increased auxin concentration on the lower side instems causes the cells in that area to grow more than thecells on the upper side. The result is a bending upward ofthe stem against the force of gravity—in other words thereis a negative gravitropic response. Such differences in hor-mone concentration have not been as well documented inroots. Nevertheless, the upper sides of roots oriented hori-zontally grow more rapidly than the lower sides, causingthe root ultimately to grow downward; this phenomenonis known as positive gravitropism. In shoots, the gravity-sensing cells are in the endoderm. Mutants like scarecrowand short root in Arabidopsis that lack normal endodermaldevelopment fail to have a normal gravitropic response.These endodermal cells are the sites of the amyloplasts inthe stems.
In roots, the gravity-sensing cells are located in theroot cap and the cells that actually do the asymmetricgrowth are in the distal elongation zone which is closestto the root cap. How the information gets transferredover this distance is an intriguing problem. Auxin may beinvolved, but when auxin transport is suppressed, there isstill a gravitropic response in the distal elongation zone.Some type of electrical signaling involving membrane po-larization has been hypothesized and this was testedaboard the space shuttle. So far the verdict is still not inon the exact mechanism.
It may surprise you to learn that in tropical rain forests,roots of some plants may grow up the stems of neighboringplants, instead of exhibiting the normal positive gravitropicresponses typical of other roots. The rainwater dissolvesnutrients, both while passing through the lush uppercanopy of the forest, and also subsequently as it tricklesdown tree trunks. Such water functions as a more reliablesource of nutrients for the roots than the nutrient-poorsoils in which the plants are anchored. Explaining this interms of current hypotheses is a challenge. It has been pro-posed that roots are more sensitive to auxin than shootsand that auxin may actually inhibit growth on the lowerside of a root, resulting in a positive gravitropic response.Perhaps in these tropical plants, the sensitivity to auxin inroots is reduced.
Thigmotropism
Thigmotropism is a name derived from the Greek rootthigma, meaning “touch.” A thigmotropism is a response ofa plant or plant part to contact with the touch of an object,animal, plant, or even the wind. (figure 41.5). When a ten-dril makes contact with an object, specialized epidermalcells, whose action is not clearly understood, perceive thecontact and promote uneven growth, causing the tendril to
curl around the object, sometimes within as little as 3 to 10minutes. Both auxin and ethylene appear to be involved intendril movements, and they can induce coiling in the ab-sence of any contact stimulus. In other plants, such asclematis, bindweed, and dodder, leaf petioles or unmodifiedstems twine around other stems or solid objects.
Again, Arabidopsis is proving valuable as a model system.A gene has been identified that is expressed in 100-foldhigher levels 10 to 30 minutes after touch. Given the valueof a molecular genetics approach in dissecting the pathwaysleading from an environmental signal to a growth response,this gene provides a promising first step in understandinghow plants respond to touch.
Other Tropisms
The tropisms just discussed are among the best known, butothers have been recognized. They include electrotropism(responses to electricity); chemotropism (response to chemi-cals); traumotropism (response to wounding which we dis-cuss on page 834); thermotropism (response to temperature);aerotropism (response to oxygen); skototropism (response todark); and geomagnetotropism (response to magnetic fields).Roots will often follow a diffusion gradient of water com-ing from a cracked pipe and enter the crack. Some call suchgrowth movement hydrotropism, but there is disagreementwhether responses to water and several other “stimuli” aretrue tropisms.
While plants can’t move away or toward optimalconditions, they can grow. Phototropisms are growthresponses of plants to a unidirectional source of light.Gravitropism, the response of a plant to gravity,generally causes shoots to grow up (negativegravitropism) and roots to grow down (positivegravitropism). Thigmotropisms are growth responses ofplants to contact.
810 Part XI Plant Growth and Reproduction
FIGURE 41.5Thigmotropism.The thigmotropicresponse of thesetwining stems causesthem to coil aroundthe object with whichthey have come incontact.

DormancySometimes modifying the direction of growth is notenough to protect a plant from harsh conditions. The abil-ity to cease growth and go into a dormant stage provides asurvival advantage. The extreme example is seed dormancy,but there are intermediate approaches to waiting out thebad times as well. Environmental signals both initiate andend dormant phases in the life of a plant.
In temperate regions, we generally associate dormancywith winter, when freezing temperatures and the accompa-nying unavailability of water make it impossible for plantsto grow. During this season, buds of deciduous trees andshrubs remain dormant, and apical meristems remain wellprotected inside enfolding scales. Perennial herbs spend thewinter underground as stout stems or roots packed withstored food. Many other kinds of plants, including most an-nuals, pass the winter as seeds.
In some seasonally dry climates, seed dormancy occursprimarily during the dry season, often the summer. Rain-falls trigger germination when conditions for survival aremore favorable. Annual plants occur frequently in areas ofseasonal drought. Seeds are ideal for allowing annual plantsto bypass the dry season, when there is insufficient waterfor growth. When it rains, they can germinate and theplants can grow rapidly, having adapted to the relativelyshort periods when water is available. Chapter 40 coveredsome of the mechanisms involved in breaking seed dor-mancy and allowing germination under favorable circum-stances. These include the leaching from the seed coats ofchemicals that inhibit germination, or mechanically crack-ing the seed coats, a procedure that is particularly suitablefor promoting growth in seasonally dry areas. Wheneverrains occur, they will leach out the chemicals from the seedcoats, and the hard coats of other seeds may be crackedwhen they are being washed down along temporarilyflooded arroyos (figure 41.6).
Seeds may remain dormant for a surprisingly long time.Many legumes (plants of the pea and bean family,Fabaceae) have tough seeds that are virtually impermeableto water and oxygen. These seeds often last decades andeven longer without special care; they will eventually ger-minate when their seed coats have been cracked and wateris available. Seeds that are thousands of years old have beensuccessfully germinated!
A period of cold is necessary before some kinds ofseeds will germinate, as we mentioned in chapter 40. Theseeds of other plants will germinate only when adequatewater is available and the temperatures are relatively high.For this reason, certain weeds germinate and grow in thecooler part of the year and others in the warmer part ofthe year. Similarly, a period of cold is needed before thebuds of some trees and shrubs will break dormancy anddevelop normally. For this reason, many plants that nor-mally grow in temperate regions do not thrive in warmerregions near the equator, because even at high elevations
in the tropics it still does not get cold enough, and theday-length relationships are different from those of tem-perate regions.
Mature plants may become dormant in dry or coldseasons that are unfavorable for growth. Dormantplants usually lose their leaves and drought-resistantwinter buds are produced. Long unfavorable periodsmay be bypassed through the production of seeds,which themselves can remain dormant for longperiods.
Chapter 41 How Plants Grow in Response to Their Environment 811
FIGURE 41.6Palo verde (Cercidium floridum). This desert tree (a) has toughseeds (b) that germinate only after they are cracked.
(a)
(b)

Plant HormonesWhile initial responses of plants to environmental signalsmay rely primarily on electrical signaling, longer-term re-sponses that alter morphology rely on complex physiologi-cal networks. Many internal signaling pathways involveplant hormones, which are the focus of this section. Hor-mones are involved in responses to the environment, aswell as internally regulated development (examples ofwhich you saw in chapter 40).
Hormones are chemical substances produced in small,often minute, quantities in one part of an organism andthen transported to another part, where they bring aboutphysiological or developmental responses. The activity ofhormones results from their capacity to stimulate certainphysiological processes and to inhibit others (figure41.7). How they act in a particular instance is influencedboth by the hormone and the tissue that receives themessage.
In animals, hormones are usually produced at definitesites, usually organs. In plants, hormones are not producedin specialized tissues but, instead, in tissues that also carry
out other, usually more obvious, functions. There are sevenmajor kinds of plant hormones: auxin, cytokinins, gib-berellins, brassinosteroids, oligosaccharins, ethylene, andabscisic acid (table 41.1). Current research is focused on thebiosynthesis of hormones and on characterizing the hor-mone receptors that trigger signal transduction pathways.Much of the molecular basis of hormone function remainsenigmatic.
Because hormones are involved in so many aspects ofplant function and development, we have chosen to inte-grate examples of hormone activity with specific aspects ofplant biology throughout the text. In this section, our goalis to give you a brief overview of these hormones. Use thissection as a reference when specific hormones are discussedin the next few chapters.
There are seven major kinds of plant hormones: auxin,cytokinins, gibberellins, brassinosteroids,oligosaccharins, ethylene, and abscisic acid.
812 Part XI Plant Growth and Reproduction
41.2 The hormones that guide growth are keyed to the environment.
FIGURE 41.7Effects of plant hormones. Plant hormones, oftenacting together, influence many aspects of plantgrowth and development, including (a) leafabscission and (b) the formation of mature fruit.
(a)
(b)
Chapter 41 How Plants Grow in Response to Their Environment 813
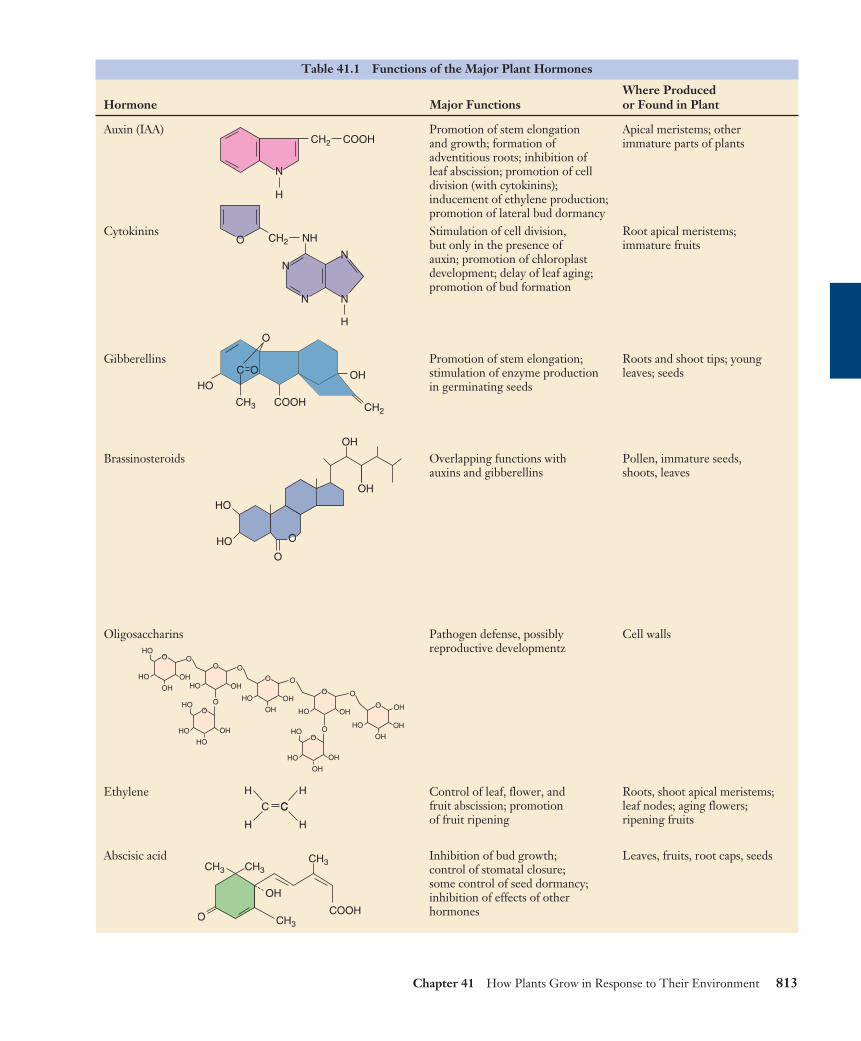
Table 41.1 Functions of the Major Plant Hormones
Where ProducedHormone Major Functions or Found in Plant
Auxin (IAA) Promotion of stem elongation Apical meristems; otherand growth; formation of immature parts of plantsadventitious roots; inhibition of leaf abscission; promotion of cell division (with cytokinins); inducement of ethylene production; promotion of lateral bud dormancy
Cytokinins Stimulation of cell division, Root apical meristems;but only in the presence of immature fruitsauxin; promotion of chloroplast development; delay of leaf aging; promotion of bud formation
Gibberellins Promotion of stem elongation; Roots and shoot tips; young stimulation of enzyme production leaves; seedsin germinating seeds
Brassinosteroids Overlapping functions with Pollen, immature seeds,auxins and gibberellins shoots, leaves
Oligosaccharins Pathogen defense, possibly Cell wallsreproductive developmentz
Ethylene Control of leaf, flower, and Roots, shoot apical meristems; fruit abscission; promotion leaf nodes; aging flowers; of fruit ripening ripening fruits
Abscisic acid Inhibition of bud growth; Leaves, fruits, root caps, seedscontrol of stomatal closure; some control of seed dormancy; inhibition of effects of other hormones
CH2 NH
NN
N
H
N
CH2 COOH
OH
CH2COOHCH3
O
HO
C O
H
O
HO
HO
O
O
OH
OH
N
O
OH
OHHO
HOO
O
OHHO
O
OH
OHHO
O
O
OHHO
O
OH
OHHO
OH
OH
HO OH
HOOH
OH HO
OH
O
O
O
O
O
O
CH3CH3
OH
CH3
CH3
COOHO
C CC
H
H
H
H

AuxinMore than a century ago, an organic substance known asauxin became the first plant hormone to be discovered.Auxin increases the plasticity of plant cell walls and is in-volved in elongation of stems. Cells can enlarge in re-sponse to changes in turgor pressure when their cell wallshave enhanced plasticity from auxin action. The discov-ery of auxin and its role in plant growth is an elegant ex-ample of thoughtful experimental design. The historicalstory is recounted here for that reason. Recent effortshave uncovered an auxin receptor. Transport mecha-nisms are also being unraveled. As with all the classes ofhormones, we are just beginning to understand, at a cel-lular and molecular level, how hormones regulate growthand development.
Discovery of Auxin
In his later years, the great evolutionist, Charles Darwin,became increasingly devoted to the study of plants. In1881, he and his son Francis published a book called ThePower of Movement of Plants. In this book, the Darwins re-ported their systematic experiments on the response ofgrowing plants to light—responses that came to beknown as phototropisms. They used germinating oat(Avena sativa) and canary grass (Phalaris canariensis)seedlings in their experiments and made many observa-tions in this field.
Charles and Francis Darwin knew that if light came pri-marily from one direction, the seedlings would bendstrongly toward it. If they covered the tip of the shoot witha thin glass tube, the shoot would bend as if it were notcovered. However, if they used a metal foil cap to excludelight from the plant tip, the shoot would not bend (figure41.8). They also found that using an opaque collar to ex-clude light from the stem below the tip did not keep thearea above the collar from bending.
In explaining these unexpected findings, the Darwinshypothesized that when the shoots were illuminated fromone side, they bent toward the light in response to an “in-fluence” that was transmitted downward from its source atthe tip of the shoot. For some 30 years, the Darwins’ per-ceptive experiments remained the sole source of informa-tion about this interesting phenomenon. Then Danishplant physiologist Peter Boysen-Jensen and the Hungarianplant physiologist Arpad Paal independently demonstratedthat the substance that caused the shoots to bend was achemical. They showed that if the tip of a germinatinggrass seedling was cut off and then replaced with a smallblock of agar separating it from the rest of the seedling, theseedling would grow as if there had been no change. Some-thing evidently was passing from the tip of the seedlingthrough the agar into the region where the bending oc-curred. On the basis of these observations under conditionsof uniform illumination or of darkness, Paal suggested that
814 Part XI Plant Growth and Reproduction
(a)
1
2
3
4
Light
Lightproof cap
Transparent cap
Lightproof collar
(b)
FIGURE 41.8The Darwins’ experiment. (a) Young grass seedlings normallybend toward the light. (b) The bending (1) did not occur when thetip of a seedling was covered with a lightproof cap (2), but didoccur when it was covered with a transparent one (3). When acollar was placed below the tip (4), the characteristic lightresponse took place. From these experiments, the Darwinsconcluded that, in response to light, an “influence” that causedbending was transmitted from the tip of the seedling to the areabelow, where bending normally occurs.
an unknown substance continually moves down from thetips of grass seedlings and promotes growth on all sides.Such a light pattern would not, of course, cause the shootto bend.
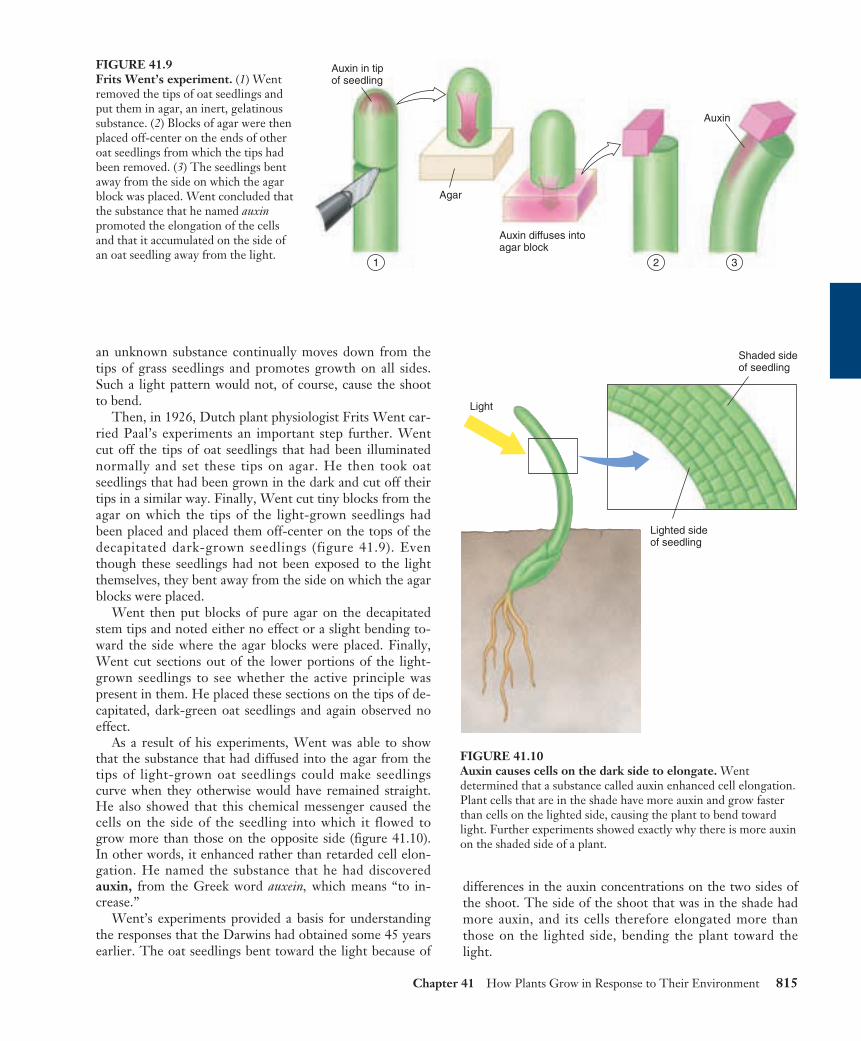
Then, in 1926, Dutch plant physiologist Frits Went car-ried Paal’s experiments an important step further. Wentcut off the tips of oat seedlings that had been illuminatednormally and set these tips on agar. He then took oatseedlings that had been grown in the dark and cut off theirtips in a similar way. Finally, Went cut tiny blocks from theagar on which the tips of the light-grown seedlings hadbeen placed and placed them off-center on the tops of thedecapitated dark-grown seedlings (figure 41.9). Eventhough these seedlings had not been exposed to the lightthemselves, they bent away from the side on which the agarblocks were placed.
Went then put blocks of pure agar on the decapitatedstem tips and noted either no effect or a slight bending to-ward the side where the agar blocks were placed. Finally,Went cut sections out of the lower portions of the light-grown seedlings to see whether the active principle waspresent in them. He placed these sections on the tips of de-capitated, dark-green oat seedlings and again observed noeffect.
As a result of his experiments, Went was able to showthat the substance that had diffused into the agar from thetips of light-grown oat seedlings could make seedlingscurve when they otherwise would have remained straight.He also showed that this chemical messenger caused thecells on the side of the seedling into which it flowed togrow more than those on the opposite side (figure 41.10).In other words, it enhanced rather than retarded cell elon-gation. He named the substance that he had discoveredauxin, from the Greek word auxein, which means “to in-crease.”
Went’s experiments provided a basis for understandingthe responses that the Darwins had obtained some 45 yearsearlier. The oat seedlings bent toward the light because of
differences in the auxin concentrations on the two sides ofthe shoot. The side of the shoot that was in the shade hadmore auxin, and its cells therefore elongated more thanthose on the lighted side, bending the plant toward thelight.
Chapter 41 How Plants Grow in Response to Their Environment 815
Auxin in tipof seedling
Agar
Auxin diffuses intoagar block
Auxin
1 2 3
FIGURE 41.9Frits Went’s experiment. (1) Wentremoved the tips of oat seedlings andput them in agar, an inert, gelatinoussubstance. (2) Blocks of agar were thenplaced off-center on the ends of otheroat seedlings from which the tips hadbeen removed. (3) The seedlings bentaway from the side on which the agarblock was placed. Went concluded thatthe substance that he named auxinpromoted the elongation of the cellsand that it accumulated on the side ofan oat seedling away from the light.
Light
Lighted sideof seedling
Shaded sideof seedling
FIGURE 41.10Auxin causes cells on the dark side to elongate. Wentdetermined that a substance called auxin enhanced cell elongation.Plant cells that are in the shade have more auxin and grow fasterthan cells on the lighted side, causing the plant to bend towardlight. Further experiments showed exactly why there is more auxinon the shaded side of a plant.

The Effects of Auxins
Auxin acts to adapt the plant to its environment in a highlyadvantageous way. It promotes growth and elongation andfacilitates the plant’s response to its environment. Environ-mental signals directly influence the distribution of auxin inthe plant. How does the environment—specifically, light—exert this influence? Theoretically, it might destroy the auxin, decrease the cells’ sensitivity to auxin, or causethe auxin molecules to migrate away from the light into theshaded portion of the shoot. This last possibility has provedto be the case.
In a simple but effective experiment, Winslow Briggs in-serted a thin sheet of mica vertically between the half of theshoot oriented toward the light and the half of the shootoriented away from it (figure 41.11). He found that lightfrom one side does not cause a shoot with such a barrier tobend. When Briggs examined the illuminated plant, hefound equal auxin levels on both the light and dark sides ofthe barrier. He concluded that a normal plant’s response tolight from one direction involves auxin migrating from thelight side to the dark side, and that the mica barrier pre-vented a response by blocking the migration of auxin.
The effects of auxin are numerous and varied. Auxinpromotes the activity of the vascular cambium and the vas-cular tissues. Also, auxins are present in pollen in large
quantities and play a key role in the development of fruits.Synthetic auxins are used commercially for the same pur-pose. Fruits will normally not develop if fertilization hasnot occurred and seeds are not present, but frequently theywill if auxins are applied. Pollination may trigger auxin re-lease in some species leading to fruit development occur-ring even before fertilization.
How Auxin Works
In spite of this long history of research on auxin, its molec-ular basis of action has been an enigma. The chemicalstructure of IAA resembles that of the amino acid trypto-phan, from which it is probably synthesized by plants (fig-ure 41.12). Unlike animal hormones, a specific signal is notsent to specific cells, eliciting a predictable response. Thereare most likely multiple auxin perception sites. Auxin is alsounique among the plant hormones in that it is transportedtoward the base of the plant. Two families of genes havebeen identified in Arabidopsis that are involved in auxintransport. For example, one protein is involved in the topto bottom transport of auxin; while two other proteinsfunction in the root tip to regulate the growth response togravity. We are still a ways from linking the measurableand observable effects of auxin to events that transpire afterit travels to a site and binds to a receptor.
816 Part XI Plant Growth and Reproduction
Light
Light
Auxin in seedling tip
(a) (b) (c)
24�
31�12�
(d)
(a) IAA (Indoleacetic acid)
CH2 CH
NH2
COOH
(b) Tryptophan
CH2O COOH
CH2 COOH
(c) Dichlorophenoxyacetic acid(2,4-D)
Cl Cl
N
H
N
H
FIGURE 41.12Auxins. (a) Indoleacetic acid (IAA), theprincipal naturally occurring auxin. (b)Tryptophan, the amino acid from whichplants probably synthesize IAA. (c)Dichlorophenoxyacetic acid (2,4-D), asynthetic auxin, is a widely used herbicide.
FIGURE 41.11Phototropism and auxin: the Winslow Briggs experiments. The basic design of theseexperiments was to place the tip of an oat seedling on an agar block, apply light from oneside, and observe the degree of curvature produced when the agar blocks were later placedon the decapitated seedlings. However, Briggs inserted a barrier in various places andnoted how this affected the location of auxin. A comparison of (a) and (b) with similarexperiments performed in the dark showed that auxin production does not depend onlight; all produced approximately 24˚ of curvature. If a barrier was inserted in the agarblock (d), light caused the displacement of the auxin away from the light. Finally,experiment (c) showed that it was displacement that had occurred, and not different ratesof auxin production on the dark and light sides, because when displacement was preventedwith a barrier, both sides of the agar block produced about 24˚ of curvature.

One of the downstream effects of auxin is an increase inplasticity of the plant cell wall. This will only work onyoung cell walls without extensive secondary cell wall for-mation. A more plastic wall will stretch more while its pro-toplast is swelling during active cell growth. The acidgrowth hypothesis provides a model linking auxin to cellwall expansion (figure 41.13). Auxin causes responsive cellsto release hydrogen ions into the cell wall. This decreasesthe pH which activates enzymes that can break bonds be-tween cell wall fibers. Remember that different enzymes op-erate optimally at different pHs. This hypothesis has beenexperimentally supported in several ways. Buffers that pre-vent cell wall acidification block cell expansion. Other com-pounds that release hydrogen ions from the cell can alsocause cell expansion. The movement of hydrogen ions hasbeen observed in response to auxin treatment. This hypoth-esis explains the rapid growth response. There are also de-layed responses which most likely involve auxin-stimulatedgene expression.
Synthetic Auxins. Synthetic auxins such as NAA (naph-thalene acetic acid) and IBA (indolebutyric acid) have manyuses in agriculture and horticulture. One of their most im-portant uses is based on their prevention of abscission, theprocess that causes a leaf or other organ to fall from aplant. Synthetic auxins are used to prevent fruit drop in ap-ples before they are ripe and to hold berries on holly that isbeing prepared for shipping. Synthetic auxins are also usedto promote flowering and fruiting in pineapples and to in-duce the formation of roots in cuttings.
Synthetic auxins are routinely used to control weeds.When used as herbicides, they are applied in higher con-centrations than IAA would normally occur in plants. Oneof the most important synthetic auxin herbicides is 2,4-dichlorophenoxyacetic acid, usually known as 2,4-D (seefigure 41.12c). It kills weeds in grass lawns by selectivelyeliminating broad-leaved dicots. The stems of the dicotweeds cease all axial growth.
The herbicide 2,4,5-trichlorophenoxyacetic acid, betterknown as 2,4,5-T, is closely related to 2,4-D. 2,4,5-T waswidely used as a broad-spectrum herbicide to kill weeds andseedlings of woody plants. It became notorious during theVietnam War as a component of a jungle defoliant knownas Agent Orange and was banned in 1979 for most uses inthe United States. When 2,4,5-T is manufactured, it is un-avoidably contaminated with minute amounts of dioxin.Dioxin, in doses as low as a few parts per billion, has pro-duced liver and lung diseases, leukemia, miscarriages, birthdefects, and even death in laboratory animals. Vietnam vet-erans and children of Vietnam veterans exposed to AgentOrange have been among the victims.
Auxin is synthesized in apical meristems of shoots. Itcauses young stems to bend toward light when itmigrates toward the darker side, where it makes youngcell walls more plastic and thereby promotes cellelongation. By interacting with other hormones, auxincan promote an increase in girth and is involved ingrowth responses to gravity and fruit ripening.
Chapter 41 How Plants Grow in Response to Their Environment 817
H+
H+
H+
H+
H+
H+
Turgor
Cytoplasm
Auxin
Cellulose fiber in cell wallEnzyme (inactive)Cross bridge
Activeenzyme
1. Auxin causes cells to pumphydrogen ions into the cell wall.
2. pH in the cell wall decreases,activating enzymes that breakcross-bridges between cellulosefibers in the cell wall.
3. Cellulose fibers loosen and allow the cell to expand as turgorpressure inside the cell pushesagainst the cell wall.
FIGURE 41.13Acid growthhypothesis. Auxinstimulates the release ofhydrogen ions from thetarget cells which altersthe pH of the cell wall.This optimizes theactivity of enzymeswhich break bonds inthe cell wall, allowingthem to expand.

CytokininsCytokinins comprise another group of naturally occurringgrowth hormones in plants. Studies by Gottlieb Haber-landt of Austria around 1913 demonstrated the existence ofan unknown chemical in various tissues of vascular plantsthat, in cut potato tubers, would cause parenchyma cells tobecome meristematic, and would induce the differentiationof a cork cambium. The role of cytokinins, active compo-nents of coconut milk, in promoting the differentiation oforgans in masses of plant tissue growing in culture later ledto their discovery. Subsequent studies have focused on therole cytokinins play in the differentiation of tissues fromcallus.
A cytokinin is a plant hormone that, in combinationwith auxin, stimulates cell division and differentiation inplants. Most cytokinins are produced in the root apicalmeristems and transported throughout the plant. Develop-ing fruits are also important sites of cytokinin synthesis. Inmosses, cytokinins cause the formation of vegetative budson the gametophyte. In all plants, cytokinins, working withother hormones, seem to regulate growth patterns.
All naturally occurring cytokinins are purines that ap-pear to be derivatives of, or have molecule side chains simi-lar to, those of adenine (figure 41.14). Other chemically di-verse molecules, not known to occur naturally, have effectssimilar to those of cytokinins. Cytokinins promote growthof lateral buds into branches (figure 41.15); though, alongwith auxin and ethylene, they also play a role in apicaldominance (the suppression of lateral bud growth). Con-
818 Part XI Plant Growth and Reproduction
Adenine
Kinetin
CH2
O
6-Benzylamino purine (BAP)
CH2 NH
NN
N
H
N
NH2
NN
N
H
N
NH
NN
N
H
N
FIGURE 41.14Some cytokinins. Two commonly used synthetic cytokinins:kinetin and 6-benzylamino purine. Note their resemblance to thepurine base adenine.
Lateral buds
Lateral branches
Apical budremoved
Apical budremoved
Auxin
Lateralbuds
FIGURE 41.15Cytokinins stimulate lateralbud growth. (a) When theapical meristem of a plant isintact, auxin from the apicalbud will inhibit the growth oflateral buds. (b) When theapical bud is removed,cytokinins are able to producethe growth of lateral budsinto branches. (c) When theapical bud is removed andauxin is added to the cutsurface, axillary budoutgrowth is suppressed.
(a) (b) (c)

versely, cytokinins inhibit formation of lateral roots, whileauxins promote their formation. As a consequence of theserelationships, the balance between cytokinins and auxin,along with other factors, determines the appearance of amature plant. In addition, the application of cytokinins toleaves detached from a plant retards their yellowing. Theyfunction as anti-aging hormones.
The action of cytokinins, like that of other hormones,has been studied in terms of its effects on growth and dif-ferentiation of masses of tissue growing in defined media.Plant tissue can form shoots, roots, or an undifferentiatedmass of tissues, depending on the relative amounts of auxinand cytokinin (figure 41.16). In the early cell-growth exper-iments coconut “milk” was an essential factor. Eventually,it was discovered that coconut “milk” is not only rich inamino acids and other reduced nitrogen compounds re-quired for growth, but it also contains cytokinins. Cy-tokinins seem to be essential for mitosis and cell division.They apparently promote the synthesis or activation ofproteins that are specifically required for mitosis.
Cytokinins have also been used against plants bypathogens. The bacteria Agrobacterium, for example, intro-
duces genes into the plant genome that increase the rate ofcytokinin, as well as auxin, production. This causes massivecell division and the formation of a tumor called crown gall(figure 41.17). How these hormone biosynthesis genesended up in a bacterium is an intriguing evolutionary ques-tion. Coevolution does not always work to the plant’sadvantage.
Cytokinins are plant hormones that, in combinationwith auxin, stimulate cell division and, along with anumber of other factors, determine the course ofdifferentiation. In contrast to auxins, cytokinins arepurines that are related to or derived from adenine.
Chapter 41 How Plants Grow in Response to Their Environment 819
Auxin:
Cytokinin:
High
HighLow
Low Intermediate
Intermediate
FIGURE 41.16Relative amounts of cytokinins and auxin affect organregeneration in culture. In the case of tobacco, (a) high auxin tocytokinin ratios favor root development; (b) high cytokinin toauxin ratios favor shoot development; and (c) intermediateconcentrations result in the formation of undifferentiated cells.These developmental responses to cytokinin/auxin ratios inculture are species specific. FIGURE 41.17
Crown gall tumor. Sometimes cytokinins can be used against theplant by a pathogen. In this case Agrobacterium tumefaciens (abacteria) has incorporated a piece of its DNA into the plantgenome. This DNA contains genes coding for enzymes necessaryfor cytokinin and auxin biosynthesis. The increased levels of thesehormones in the plant cause massive cell division and theformation of a tumor.

GibberellinsGibberellins are named after the fungus Gibberella fu-jikuroi, which causes rice plants, on which it is parasitic, togrow abnormally tall. Japanese plant pathologist EiichiKurosawa investigated Bakane (“foolish seedling”) diseasein the 1920s. He grew Gibberella in culture and obtained asubstance that, when applied to rice plants, producedbakane. This substance was isolated and the structural for-mula identified by Japanese chemists in 1939. Britishchemists reconfirmed the formula in 1954. Although suchchemicals were first thought to be only a curiosity, theyhave since turned out to belong to a large class of morethan 100 naturally occurring plant hormones called gib-berellins. All are acidic and are usually abbreviated to GA(for gibberellic acid), with a different subscript (GA1, GA2,and so forth) to distinguish each one. While gibberellinsfunction endogenously as hormones, they also function aspheromones in ferns. In ferns gibberellin-like compoundsreleased from one gametophyte can trigger the develop-ment of male reproductive structures on a neighboring ga-metophyte.
Gibberellins, which are synthesized in the apical por-tions of stems and roots, have important effects on stemelongation. The elongation effect is enhanced if auxin isalso present. The application of gibberellins to dwarf mu-tants is known to restore the normal growth and develop-ment in many plants (figure 41.18). Some dwarf mutantsproduce insufficient amounts of gibberellin; while otherslack the ability to perceive gibberellin. The large numberof gibberellins are all part of a complex biosynthetic path-way that has been unraveled using gibberellin-deficientmutants in maize (corn). While many of these gibberellinsare intermediate forms in the production of GA1, recentwork shows that different forms may have specific biologi-cal roles.
In chapter 41, we noted the role gibberellins stimulatethe production of �-amylase and other hydrolytic en-zymes needed for utilization of food resources during ger-mination and establishment of cereal seedlings. How arethe genes encoding these enzymes transcribed? Experi-mental studies in the aleurone layer surrounding the en-dosperms of cereal grains have shown that transcriptionoccurs when the gibberellins initiate a burst of messengerRNA (mRNA) and protein synthesis. GA somehow en-hances DNA binding proteins, which in turn allow DNAtranscription of a gene. Synthesis of DNA does not seemto occur during the early stages of seed germination butbecomes important when the radicle has grown throughthe seed coats.
Gibberellins also affect a number of other aspects ofplant growth and development. These hormones also has-ten seed germination, apparently because they can substi-tute for the effects of cold or light requirements in thisprocess. Gibberellins are used commercially to space grape
flowers by extending internode length so the fruits havemore room to grow (figure 41.19).
Gibberellins are an important class of plant hormonesthat are produced in the apical regions of shoots androots. They play the major role in controlling stemelongation for most plants, acting in concert with auxinand other hormones.
820 Part XI Plant Growth and Reproduction
FIGURE 41.18Effects of gibberellins. This rapid cycling member of themustard family plant (Brassica rapa) will “bolt” and flower becauseof increased gibberellin levels. Mutants such as the rosette mutantshown here (left) are defective in producing gibberellins. They canbe rescued by applying gibberellins. Other mutants have beenidentified that are defective in perceiving gibberellins and theywill not respond to gibberellin applications.
FIGURE 41.19Applications of gibberellins increase the space betweengrapes. Larger grapes develop because there is more roombetween individual grapes.

Brassinosteroids andOligosaccharinsBrassinosteroids
Although we’ve known about brassinosteroids for 30 years,it is only recently that they have claimed their place as aclass of plant hormones. They were first discovered in Bras-sica pollen, hence the name. Their historical absence in dis-cussions of hormones may be partially due to their func-tional overlap with other plant hormones, especially auxinsand gibberellins. Additive effects among these three classeshave been reported. The application of molecular geneticsto the study of brassinosteroids has led to tremendous ad-vances in our understanding of how they are made and, tosome extent, how they function in signal transduction path-ways. What is particularly intriguing about brassinosteroidsare similarities to animal steroid hormones (figure 41.20).One of the genes coding for an enzyme in the brassinos-teroid biosynthetic pathway has significant similarity to anenzyme used in the synthesis of testosterone and relatedsteroids. Brassinosteroids have been identified in algae andappear to be quite ubiquitous among the plants. It is plausi-ble that their evolutionary origin predated the plant-animalsplit.
Brassinosteroids have a broad spectrum of physiologicaleffects—elongation, cell division, bending of stems, vascu-lar tissue development, delayed senescence, membrane po-larization, and reproductive development. Environmentalsignals can trigger brassinosteroid actions. Mutants havebeen identified that alter the response to brassinosteroid,but signal transduction pathways remain to be uncovered.From an evolutionary perspective, it will be quite interest-ing to see how these pathways compare with animal steroidsignal transduction pathways.
Oligosaccharins
In addition to cellulose, plant cell walls are composed ofnumerous complex carbohydrates called oligosaccharides.There is some evidence that these cell wall componentsfunction as signaling molecules as well as structural wallcomponents. Oligosaccharides that are proposed to have ahormonelike function are called oligosaccharins. Oligosac-charins can be released from the cell wall by enzymes se-creted by pathogens. These carbohydrates are believed tosignal defense responses, such as the hypersensitive re-sponse discussed later in this chapter. Another oligosaccha-rin has been shown to inhibit auxin-stimulated elongationof pea stems. These molecules are active at concentrationsone to two orders of magnitude less than the traditionalplant hormones. You have seen how auxin and cytokininratios can affect organogenesis in culture. Oligosaccharinsalso affect the phenotype of regenerated tobacco tissue, in-hibiting root formation and stimulating flower productionin tissues that are competent to regenerate flowers. How
the culture results translate to in vivo systems is an openquestion. The structural biochemistry of oligosaccharinsmakes them particularly challenging molecules to study.How they interface with cells and initiate signal transduc-tion pathways is an open question.
Brassinosteroids are structurally similar to animalsteroid hormones. They have many effects on plantgrowth and development that parallel those of auxinsand gibberellins. Oligosaccharins are complexcarbohydrates that are released from cell walls andappear to regulate both pathogen responses and growthand development in some plants.
Chapter 41 How Plants Grow in Response to Their Environment 821
HO
HO
O
O
OH
OH
OOH
O
CH2OH
CH3
CH3HO
C
O
OH
CH3
CH3
HO
OHCH3
Estradiol
Testosterone
Cortisol
Brassinolide
FIGURE 41.20Brassinosteroids, such as brassinolide, have structuralsimilarities to animal steroid hormones.
EthyleneLong before its role as a plant hormonewas appreciated, the simple, gaseoushydrocarbon ethylene (H2C�CH2) wasknown to defoliate plants when itleaked from gaslights in streetlamps.Ethylene is, however, a natural productof plant metabolism that, in minuteamounts, interacts with other planthormones. When auxin is transporteddown from the apical meristem of thestem, it stimulates the production ofethylene in the tissues around the lat-eral buds and thus retards their growth.Ethylene also suppresses stem and rootelongation, probably in a similar way.An ethylene receptor has been identi-fied and characterized. It appears tohave evolved early in the evolution ofphotosynthetic organisms, sharing fea-tures with environmental-sensing pro-teins identified in bacteria.
Ethylene plays a major role in fruitripening. At first, auxin, which is pro-duced in significant amounts in polli-nated flowers and developing fruits,stimulates ethylene production; this, inturn, hastens fruit ripening. Complexcarbohydrates are broken down intosimple sugars, chlorophylls are brokendown, cell walls become soft, and thevolatile compounds associated with fla-vor and scent in ripe fruits are produced.
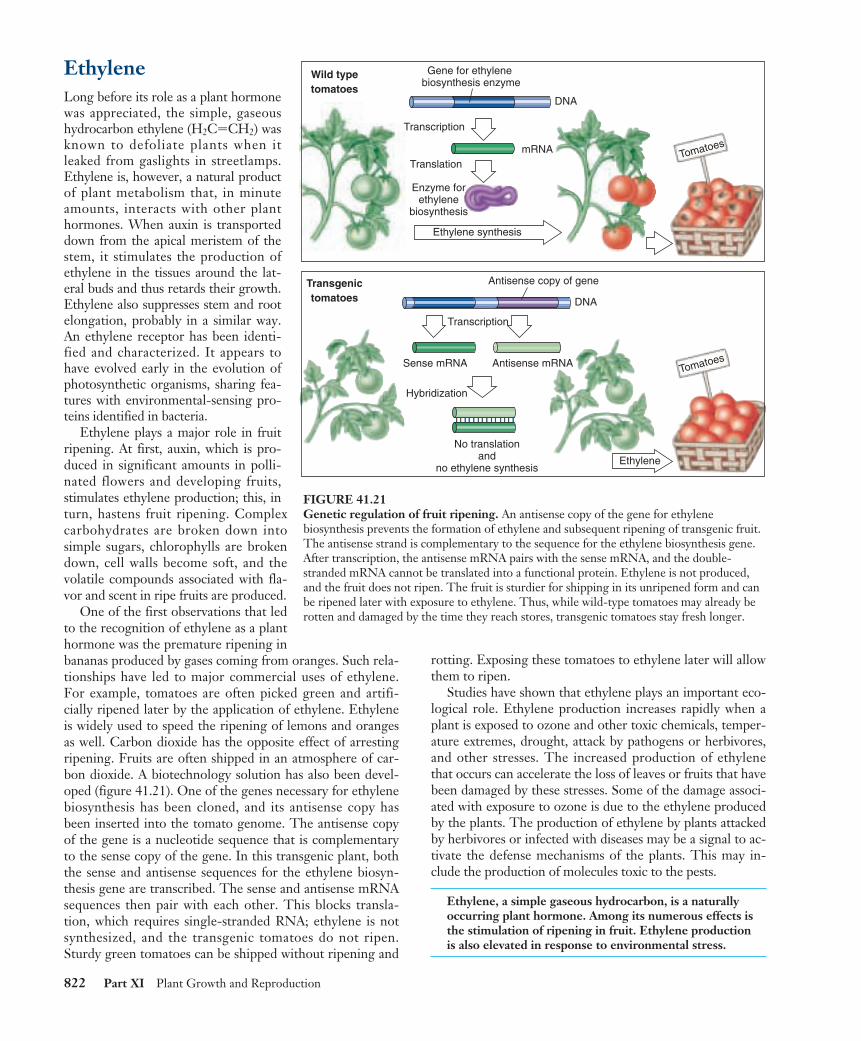
One of the first observations that ledto the recognition of ethylene as a planthormone was the premature ripening inbananas produced by gases coming from oranges. Such rela-tionships have led to major commercial uses of ethylene.For example, tomatoes are often picked green and artifi-cially ripened later by the application of ethylene. Ethyleneis widely used to speed the ripening of lemons and orangesas well. Carbon dioxide has the opposite effect of arrestingripening. Fruits are often shipped in an atmosphere of car-bon dioxide. A biotechnology solution has also been devel-oped (figure 41.21). One of the genes necessary for ethylenebiosynthesis has been cloned, and its antisense copy hasbeen inserted into the tomato genome. The antisense copyof the gene is a nucleotide sequence that is complementaryto the sense copy of the gene. In this transgenic plant, boththe sense and antisense sequences for the ethylene biosyn-thesis gene are transcribed. The sense and antisense mRNAsequences then pair with each other. This blocks transla-tion, which requires single-stranded RNA; ethylene is notsynthesized, and the transgenic tomatoes do not ripen.Sturdy green tomatoes can be shipped without ripening and
rotting. Exposing these tomatoes to ethylene later will allowthem to ripen.
Studies have shown that ethylene plays an important eco-logical role. Ethylene production increases rapidly when aplant is exposed to ozone and other toxic chemicals, temper-ature extremes, drought, attack by pathogens or herbivores,and other stresses. The increased production of ethylenethat occurs can accelerate the loss of leaves or fruits that havebeen damaged by these stresses. Some of the damage associ-ated with exposure to ozone is due to the ethylene producedby the plants. The production of ethylene by plants attackedby herbivores or infected with diseases may be a signal to ac-tivate the defense mechanisms of the plants. This may in-clude the production of molecules toxic to the pests.
Ethylene, a simple gaseous hydrocarbon, is a naturallyoccurring plant hormone. Among its numerous effects isthe stimulation of ripening in fruit. Ethylene productionis also elevated in response to environmental stress.
822 Part XI Plant Growth and Reproduction
Enzyme forethylene
biosynthesis
Gene for ethylene biosynthesis enzyme
Transcription
mRNATranslation
Tomatoes
Tomatoes
Antisense copy of gene
Transcription
Sense mRNA Antisense mRNA
Hybridization
No translationand
no ethylene synthesis
Ethylene synthesis
Ethylene
Wild typetomatoes
Transgenictomatoes
DNA
DNA
FIGURE 41.21Genetic regulation of fruit ripening. An antisense copy of the gene for ethylenebiosynthesis prevents the formation of ethylene and subsequent ripening of transgenic fruit.The antisense strand is complementary to the sequence for the ethylene biosynthesis gene.After transcription, the antisense mRNA pairs with the sense mRNA, and the double-stranded mRNA cannot be translated into a functional protein. Ethylene is not produced,and the fruit does not ripen. The fruit is sturdier for shipping in its unripened form and canbe ripened later with exposure to ethylene. Thus, while wild-type tomatoes may already berotten and damaged by the time they reach stores, transgenic tomatoes stay fresh longer.

Abscisic AcidAbscisic acid, a naturally occurring plant hormone, ap-pears to be synthesized mainly in mature green leaves,fruits, and root caps. The hormone earned its name be-cause applications of it appear to stimulate leaf senescence(aging) and abscission, but there is little evidence that itplays an important role in this process. In fact, it is believedthat abscisic acid may cause ethylene synthesis, and that itis actually the ethylene that promotes senescence and absci-sion. When abscisic acid is applied to a green leaf, the areasof contact turn yellow. Thus, abscisic acid has the exact op-posite effect on a leaf from that of the cytokinins; a yellow-ing leaf will remain green in an area where cytokinins areapplied.
Abscisic acid probably induces the formation of winterbuds—dormant buds that remain through the winter—bysuppressing growth. The conversion of leaf primordia intobud scales follows (figure 41.22a). Like ethylene, it mayalso suppress growth of dormant lateral buds. It appearsthat abscisic acid, by suppressing growth and elongation ofbuds, can counteract some of the effects of gibberellins(which stimulate growth and elongation of buds); it alsopromotes senescence by counteracting auxin (which tendsto retard senescence). Abscisic acid plays a role in seed dor-mancy and is antagonistic to gibberellins during germina-tion. It is also important in controlling the opening andclosing of stomata (figure 41.22b).
Abscisic acid occurs in all groups of plants and appar-ently has been functioning as a growth-regulating sub-stance since early in the evolution of the plant kingdom.Relatively little is known about the exact nature of its phys-iological and biochemical effects. These effects are veryrapid—often taking place within a minute or two—andtherefore they must be at least partly independent of geneexpression. Some longer-term effects of abscisic acid in-volve the regulation of gene expression, but the way thisoccurs is poorly understood. Abscisic acid levels becomegreatly elevated when the plant is subject to stress, espe-cially drought. Like other plant hormones, abscisic acidprobably will prove to have valuable commercial applica-tions when its mode of action is better understood. It is aparticularly strong candidate for understanding desiccationtolerance.
Abscisic acid, produced chiefly in mature green leavesand in fruits, suppresses growth of buds and promotesleaf senescence. It also plays an important role incontrolling the opening and closing of stomata. Abscisicacid may be critical in ensuring survival underenvironmental stress, especially water stresses.
Chapter 41 How Plants Grow in Response to Their Environment 823
FIGURE 41.22Effects of abscisic acid. (a) Abscisic acid plays a role in theformation of these winter buds of an American basswood. Thesebuds will remain dormant for the winter, and bud scales—modified leaves—will protect the buds from desiccation. (b) Abscisic acid also affects the closing of stomata by influencingthe movement of potassium ions out of guard cells.
(a)
(b)

Plants UndergoMetamorphosisOverview of Initiating Flowering
Carefully regulated processes deter-mine when and where flowers willform. Plants must often gain compe-tence to respond to internal or exter-nal signals regulating flowering. Onceplants are competent to reproduce, acombination of factors including light,temperature, and both promotive andinhibitory internal signals determinewhen a flower is produced (figure41.23). These signals turn on genesthat specify where the floral organs,sepals, petals, stamens, and carpels willform. Once cells have instructions to become a specific flo-ral organ, yet another developmental cascade leads to thethree-dimensional construction of flower parts.
Phase Change
Plants go through developmental changes leading to re-productive maturity just like many animals. This shiftfrom juvenile to adult development is seen in the meta-morphosis of a tadpole to an adult frog or caterpillar to abutterfly that can then reproduce. Plants undergo a simi-lar metamorphosis that leads to the production of aflower. Unlike the frog that loses its tail, plants just keepadding on structures to existing structures with theirmeristems. At germination, most plants are incapable of
producing a flower, even if all the environmental cues areoptimal. Internal developmental changes allow plants toobtain competence to respond to external and/or internalsignals that trigger flower formation. This transition is re-ferred to as phase change. Phase change can be morpho-logically obvious or very subtle. Take a look at an oak treein the winter. The lower leaves will still be clinging to thebranches, while the upper ones will be gone (figure41.24a). Those lower branches were initiated by a juvenilemeristem. The fact that they did not respond to environ-mental cues and drop their leaves indicates that they areyoung branches and have not made a phase change. Ivyalso has distinctive juvenile and adult phases of growth(figure 41.24b). Stem tissue produced by a juvenile meris-tem initiates adventitious roots that can cling to walls. If
824 Part XI Plant Growth and Reproduction
41.3 The environment influences flowering.
Phase change
Juvenile Adult
Floral promoters,floral inhibitors
Flowering
Temperature
Light
FIGURE 41.23Factors involved in initiating flowering. This is a model of environmentally cued andinternally processed events that result in a shoot meristem initiating flowers.
FIGURE 41.24Phase change. (a) The lowerbranches of this oak tree representthe juvenile phase of developmentand cling to their leaves in thewinter. The lower leaves are notable to form an abscission layer andbreak off the tree in the fall. Suchvisible changes are marks of phasechange, but the real test is whetheror not the plant is able to flower. (b)Juvenile ivy (left) makes adventitiousroots and has an alternating leafarrangement. Adult ivy (right)cannot make adventitious roots andhas leaves with a differentmorphology that are arranged on anupright stem in a spiral.
(a)
(b)

you look at very old brick buildings that are covered withivy, you will notice the uppermost branches are falling offbecause they have transitioned to the adult phase ofgrowth and have lost the ability to produce adventitiousroots. It is important to remember that even though aplant has reached the adult stage of development, it mayor may not produce reproductive structures. Other factorsmay be necessary to trigger flowering.
Generally it is easier to get a plant to revert from anadult to vegetative state than to induce phase change exper-imentally. Applications of gibberellins and severe pruningcan cause reversion. There is evidence in peas and Ara-bidopsis for genetically controlled repression of flowering.The embryonic flower mutant of Arabidopsis flowers almostimmediately (figure 41.25), which is consistent with the hy-pothesis that the wild-type allele suppresses flowering. It ispossible that flowering is the default state and that mecha-nisms have evolved to delay flowering. This delay allowsthe plant to store more energy to be allocated for repro-duction.
The best example of inducing the juvenile to adulttransition comes from the construction of transgenicplants that overexpress a gene necessary for flowering,that is found in many species. This gene, LEAFY, wascloned in Arabidopsis and its promoter was replaced with aviral promoter that results in constant, high levels ofLEAFY transcription. This gene construct was then in-troduced into cultured aspen cells which were used to re-generate plants. When LEAFY isoverexpressed in aspen, flowering oc-curs in weeks instead of years (figure41.26). Phase change requires bothsufficient signal and the ability to per-ceive the signal. Some plants acquirecompetence in the shoot to perceive asignal of a certain intensity. Othersacquire competence to produce suffi-cient promotive signal(s) and/or de-crease inhibitory signal(s).
Plants become reproductivelycompetent through changes insignaling and perception. Thetransition to the adult stage ofdevelopment where reproductionis possible is called phase change.Plants in the adult phase ofdevelopment may or may notproduce reproductive structures(flowers), depending onenvironmental cues.
Chapter 41 How Plants Grow in Response to Their Environment 825
FIGURE 41.25In Arabidopsis, the embryonic flower gene may repressflowering. The embryonic flower mutant flowers upongermination.
(a) (b)
FIGURE 41.26Overexpression of a flowering gene can accelerate phase change. (a) An aspen treenormally grows for several years before producing flowers. (b) Overexpression of theArabidopsis flowering gene, LEAFY, causes rapid flowering in a transgenic aspen.

Pathways Leading to FlowerProductionThe environment can promote or repress flowering. Insome cases, it can be relatively neutral. Light can be a sig-nal that long, summer days have arrived in a temperate cli-mate and conditions are favorable for reproduction. Inother cases, plants depend on light to accumulate sufficientamounts of sucrose to fuel reproduction, but flower inde-pendently of the length of day. Temperature can also beused as a clue. Gibberellins are important and have beenlinked to the vernalization pathway. Clearly, reproductivesuccess would be unlikely in the middle of a blizzard. As-suming regulation of reproduction first arose in more con-stant tropical environments, many of the day length andtemperature controls would have evolved as plants colo-nized more temperate climates. Plants can rely primarily onone pathway, but all three pathways can be present. Thecomplexity of the flowering pathways has been dissectedphysiologically. Now analysis of flowering mutants is pro-viding insight into the molecular mechanisms of the floralpathways. The redundancy of pathways to flowering en-sures that there will be another generation.
Light-Dependent Pathway
Flowering requires much energy accumulated via photosyn-thesis. Thus, all plants require light for flowering, but this isdistinct from the photoperiodic, or light-dependent, flow-ering pathway. Aspects of growth and development inmost plants are keyed to changes in the proportion oflight to dark in the daily 24-hour cycle (day length). Thisprovides a mechanism for organisms to respond to sea-sonal changes in the relative length of day and night. Daylength changes with the seasons; the farther from theequator, the greater the variation. Flowering responses ofplants to day length fall into several basic categories.When the daylight becomes shorter than a criticallength, flowering is initiated in short-day plants (figure41.27). When the daylight becomes longer than a criticallength, flowering is initiated in long-day plants. Otherplants, such as snapdragons, roses, and many native to thetropics (for example, tomatoes), will flower when matureregardless of day length, as long as they have receivedenough light for normal growth. These are referred to asday-neutral plants. Several grasses (for example, Indiangrass, Sorghastrum nutans), as well as ivy, have two critical
826 Part XI Plant Growth and Reproduction
Long-day plants Short-day plants
Short length of darkrequired for bloom
Iris
Midnight
Noon
6 P.M. 6 A.M.
6 P.M. 6 A.M.
6 P.M. 6 A.M.
Earlysummer
Latefall Midnight
Noon
Flash of light
Noon
(a)
(b)
GoldenrodLong length of darkrequired for bloom
FIGURE 41.27How flowering responds to day length.(a) This iris is a long-day plant that isstimulated by short nights to flower in thespring. The goldenrod is a short-day plantthat, throughout its natural distribution inthe northern hemisphere, is stimulated bylong nights to flower in the fall. (b) If thelong night of winter is artificiallyinterrupted by a flash of light, thegoldenrod will not flower, and the iris will.In each case, it is the duration ofuninterrupted darkness that determineswhen flowering will occur.

photoperiods; they will not flower if the days are toolong, and they also will not flower if the days are tooshort. In some species, there is a sharp distinction be-tween long and short days. In others, flowering occursmore rapidly or slowly depending on the length of day.These plants rely on other flowering pathways as welland are called facultative long- or short-day plants. Thegarden pea is an example of a facultative long-day plant.In all of these plants, it is actually the length of darkness(night), not the length of day, that is physiologically sig-nificant. Using light as a cue permits plants to flowerwhen abiotic environmental conditions are optimal, polli-nators are available, and competition for resources withother plants may be less. For example, the springephemerals flower in the woods before the canopy leafsout, blocking sunlight necessary for photosynthesis.
At middle latitudes, most long-day plants flower in thespring and early summer; examples of such plants includeclover, irises, lettuce, spinach, and hollyhocks. Short-dayplants usually flower in late summer and fall, and includechrysanthemums, goldenrods, poinsettias, soybeans, andmany weeds. Commercial plant growers use these re-sponses to day length to bring plants into flower at spe-cific times. For example, photoperiod is manipulated ingreenhouses so poinsettias flower just in time for thewinter holidays (figure 41.28). The geographic distribu-tion of certain plants may be determined by flowering re-sponses to day length.
Photoperiod is perceived by several different forms ofphytochrome and also a blue-light-sensitive molecule(cryptochrome). The conformational change in a light re-ceptor molecule triggers a cascade of events that leads tothe production of a flower. There is a link between lightand the circadian rhythm regulated by an internal clockthat facilitates or inhibits flowering. At a molecular levelthe gaps between light signaling and production of flowersare rapidly filling in and the control mechanisms are quitecomplex. Here is one example of how day length affects aspecific flowering gene in Arabidopsis, a facultative long-dayplant that flowers in response to both far-red and bluelight. Red light inhibits flowering. The gene CONSTANS(CO) is expressed under long days but not short days. Theloss of CO product does not alter when a plant flowersunder short days, but delays flowering under long days.What happens is that the gene is positively regulated bycryptochrome that perceives blue light under long days.Cryptochrome appears to inhibit the inhibition of flower-ing by phytochrome B exposed to red light. Simply put,flowering is promoted by repressing a gene that repressesflowering! CO is a transcription factor that turns on othergenes which results in the expression of LEAFY. As dis-cussed in the section on phase change, LEAFY is one of thekey genes that “tells” a meristem to switch over to flower-ing. We will see that other pathways also converge on thisimportant gene.
The Flowering Hormone: Does It Exist? The HolyGrail in plant biology has been a flowering hormone,quested unsuccessfully for more than 50 years. A consider-able amount of evidence demonstrates the existence of sub-stances that promote flowering and substances that inhibitit. Grafting experiments have shown that these substancescan move from leaves to shoots. The complexity of theirinteractions, as well as the fact that multiple chemical mes-sengers are evidently involved, has made this scientificallyand commercially interesting search very difficult, and tothis day, the existence of a flowering hormone remainsstrictly hypothetical. We do know that LEAFY can be ex-pressed in the vegetative as well as the reproductive por-tions of plants. Clearly, information about day length gath-ered by leaves is transmitted to shoot apices. Given thatthere are multiple pathways to flowering, several signalsmay be facilitating communication between leaves andshoots. We also know that roots can be a source of floralinhibitors affecting shoot development.
Chapter 41 How Plants Grow in Response to Their Environment 827
FIGURE 41.28Manipulation of photoperiod in greenhouses ensures thatshort-day poinsettias flower in time for the winter holidays.Note that the colorful “petals” are actually sepals. Even afterflowering is induced, there are many developmental events leadingto the production of species-specific flowers.

Temperature-Dependent Pathway
Lysenko solved the problem of winter wheat seed rotting inthe fields in Russia by chilling the seeds and planting themin the spring. Winter wheat would not flower without a pe-riod of chilling, called vernalization. Unfortunately a greatmany problems, including mistreatment of Russian geneti-cists, resulted from this scientifically significant discovery.Lysenko erroneously concluded that he had converted onespecies, winter wheat, to another, spring wheat, by simplyaltering the environment. There was a shift from science topolitics. Genetics and Darwinian evolution were suspect forhalf a century. Social history aside, the valuable lesson hereis that cold temperatures can accelerate or permit floweringin many species. As with light, this ensures that plantsflower at more optimal times.
Vernalization may be necessary for seeds or plants inlater stages of development. Analysis of mutants inArabidopsis and pea indicate that vernalization is a sepa-rate flowering pathway that may be linked to the hor-mone gibberellin. In this pathway, repression may alsolead to flowering. High levels of one of the genes in thepathway may block the promotion of flowering by gib-berellins. When plants are chilled, there is less of thisgene product and gibberellin activity may increase. It isknown that gibberellins enhance the expression ofLEAFY. One proposal is that both the vernalization andautonomous pathways share a common intersection af-fecting gibberellin promotion of flowering. Weigel hasshown that gibberellin actually binds the promoter of theLEAFY gene, so its effect on flowering is direct. The con-nection between gibberellin levels and temperature alsoneeds to be understood.
Autonomous Pathway
The autonomous pathway to flowering is independent ofexternal cues except for basic nutrition. Presumably thiswas the first pathway to evolve. Day-neutral plants oftendepend primarily on the autonomous pathway which allowsplants to “count” and “remember.” A field of day-neutraltobacco will produce a uniform number of nodes beforeflowering. If the shoots of these plants are removed at dif-ferent positions, axillary buds will grow out and producethe same number of nodes as the removed portion of theshoot (figure 41.29). At a certain point in developmentshoots become committed or determined to flower (figure41.30). The upper axillary buds of flowering tobacco willremember their position when rooted or grafted. The ter-minal shoot tip becomes florally determined about fournodes before it initiates a flower. In some other species,this commitment is less stable and/or occurs later.
How do shoots know where they are and at some point“remember” that information? It is clear that there are in-hibitory signals from the roots. If bottomless pots are con-tinuously placed over a growing tobacco plant and filled
with soil, flowering is delayed by the formation of adventi-tious roots (figure 41.31). Control experiments with leaf re-moval show that it is the addition of roots and not the lossof leaves that delays flowering. A balance between floralpromoting and inhibiting signals may regulate when flow-ering occurs in the autonomous pathway and the otherpathways as well.
828 Part XI Plant Growth and Reproduction
Intactplant
Shootremoved
Replacementshoot
Intactplant
Shootremoved
Replacementshoot
(a) Upper axillary bud removed
(b) Lower axillary bud removed
Shootremovedhere
Shootremovedhere
FIGURE 41.29Plants can count. When axillary buds of flowering tobaccoplants are released from apical dominance by removing the mainshoot, they replace the number of nodes that were initiated by themain shoot. (After McDaniel 1996.)

Determination for flowering is tested at the organ orwhole plant level by changing the environment and ascer-taining whether or not the fate has changed. How does flo-ral determination correlate with molecular level changes?In Arabidopsis, floral determination correlates with the in-crease of LEAFY gene expression and has occurred by thetime a second flowering gene, APETALA1, is expressed.Because all three flowering pathways appear to convergewith increased levels of LEAFY, this determination eventshould occur in species with a variety of balances amongthe pathways.
Plants use light receptor molecules to measure thelength of night. This information is then used to signalpathways that promote or inhibit flowering. Lightreceptors in the leaves trigger events that result inchanges in the shoot meristem. Vernalization is therequirement for a period of chilling before a plant canflower. The autonomous pathway leads to floweringindependent of environmental cues. Plants integrateinformation about position in regulating flowering andboth promoters and inhibitors of flowering areimportant.
Chapter 41 How Plants Grow in Response to Their Environment 829
(a) Shoot florally determined (b) Shoot not florally determined
Shootremovedhere
Intactplant
Intactplant
Shootremoved
Shootremoved
Rootedshoot
Floweringrootedshoot
Rootedshoot
Floweringrootedshoot
Shootremovedhere
FIGURE 41.30Plants can remember. At a certain point in the flowering process, shoots become committed to making a flower. This is called floraldetermination. (a) Florally determined axillary buds “remember” their position when rooted in a pot. That is, they produce the same number of nodes that they would have if they had grown out on the plant, and then they flower. (b) Those that are not yet florallydetermined cannot remember how many nodes they have left, so they start counting again. That is, they develop like a seedling and thenflower. (After McDaniel 1996.)
Control plants:no treatment
Experimental plants:pot-on-pot treatment
Control plants:Lower leaves werecontinually removed
FIGURE 41.31Roots can inhibit flowering. Adventitious roots formed asbottomless pots were continuously placed over growing tobaccoplants, delaying flowering. The delay in flowering is caused by theroots, not the loss of the leaves. This was shown by removing leaveson control plants at the same time and in the same position as leaveson experimental plants that became buried as pots were added.

Identity Genes and the Formation ofFloral Meristem and Floral OrgansArabidopsis and snapdragon are valuable model systemsfor identifying flowering genes and understanding theirinteractions. The three pathways, discussed in the previ-ous section, lead to an adult meristem becoming a floralmeristem by either activating or repressing the inhibitionof floral meristem identity genes (figure 41.32). Two ofthe key floral meristem identity genes are LEAFY andAPETALA1. These genes establish the meristem as aflower meristem. They then turn on floral organ identitygenes. The floral organ identity genes define four con-centric whorls moving inward in the floral meristem assepal, petal, stamen, and carpel. Meyerowitz and Coenproposed a model, called the ABC model, to explain howthree classes of floral organ identity genes could specifyfour distinct organ types (figure 41.33). The ABC modelproposes that three classes of organ identity genes (A, B,and C) specify the floral organs in the four floral whorls.By studying mutants the researchers have determined thefollowing:
1. Class A genes alone specify the sepals.2. Class A and class B genes together specify the petals.3. Class B and class C genes together specify the
stamens.4. Class C genes alone specify the carpels.
The beauty of their ABC model is that it is entirely testableby making different combinations of floral organ identity
mutants. Each class of genes is expressed in two whorls,yielding four different combinations of the gene products.When any one class is missing, there are aberrant floral or-gans in predictable positions.
It is important to recognize that this is actually only thebeginning of the making of a flower. These organ identitygenes are transcription factors that turn on many moregenes that will actually give rise to the three-dimensionalflower. There are also genes that “paint” the petals. Com-plex biochemical pathways lead to the accumulation of an-thocyanin pigments in vacuoles. These pigments can be or-ange, red, or purple and the actual color is influenced bypH and by the shape of the petal.
The Formation of Gametes
The ovule within the carpel has origins more ancient thanthe angiosperms. Floral parts are modified leaves, andwithin the ovule is the female gametophyte. This nextgeneration develops from placental tissue in the ovary. Amegaspore mother cell develops and meiotically gives riseto the embryo sac. Usually two layers of integument tissueform around this embryo sac and will become the seedcoat. Genes responsible for the initiation of integumentsand also those responsible for the formation of the integu-ment have been identified. Some also affect leaf structure.This chapter has focused on the complex and elegantprocess that gives rise to the reproductive structure calledthe flower. It is indeed a metamorphosis, but the subtleshift from mitosis to meiosis in the megaspore mother cell
leading to the development of a haploid,gamete-producing gametophyte is per-haps even more critical. The same can besaid for pollen formation in the anther ofthe stamen. As we will see in the nextchapter, the flower houses the haploidgenerations that will produce gametes.The flower also functions to increase theprobability that male and female gametesfrom different (or sometimes the sameplant) will unite.
Floral structures form as a result offloral meristem identity genesturning on floral organ identity geneswhich specify where sepals, petals,stamens, and carpels will form. Thisis followed by organ developmentwhich involves many complexpathways that account for floraldiversity among species.
830 Part XI Plant Growth and Reproduction
Repression of floral inhibitors
Activation of floral meristem identity genes
Phy
toch
rom
e an
d
cryp
toch
rom
e Gibberellin
production
Light Coldtemperature
Light dependentpathway
Autonomouspathway
Temperaturedependentpathway
Adult meristem Floral meristem
FIGURE 41.32Model for flowering. The light-dependent, temperature-dependent, and autonomousflowering pathways promote the formation of floral meristems from adult meristems byrepressing floral inhibitors and activating floral meristem identity genes.

Chapter 41 How Plants Grow in Response to Their Environment 831
Whorl 1Sepal
Whorl 2Petal
Whorl 3Stamen
Whorl 4Carpel
A A
AandB
AandB
BandC
BandC
C
C C
BandC
BandC
BandC
BandC
C
Wild typefloral meristem
Sepals
Petals
Stamens
CarpelsDevelopment
Development
Development
Development
Cross-section of wild type flower
–A mutantfloral meristem:missing geneclass A
–B mutantfloral meristem:missing geneclass B
–C mutantfloral meristem:missing geneclass C
Carpels
Carpels
Stamens
Stamens
Sepals
Sepals
Carpels
Carpels
A A A A
A
A
A
CCC
Cross-section of –A mutant flower
Cross-section of –B mutant flower
Cross-section of –C mutant flower
Sepals
Petals
Petals
Sepals
A and B
A and B
A and B A
and B
FIGURE 41.33ABC model for floral organspecification.Letters labeling whorls indicatewhich gene classes are active.When A function is lost (-A), Cexpands to the first and secondwhorls. When B function is lost(-B), both outer two whorls havejust A function, and both innertwo whorls have just C function;none of the whorls have dualgene function. When C functionis lost (-C), A expands into theinner two whorls. These newcombinations of gene expressionpatterns alter which floralstructures form in each whorl.(Model proposed by Coen andMeyerowitz, 1991.)

Larger predators, microbes, water, and wind often presenta plant with rapid immediate stress. Response, to be effec-tive, must also be immediate. There is little time forgrowth, and plants instead invoke a variety of other kindsof responses. Many environmental cues trigger rapid andreversible localized plant movements, for example. Therapid folding of leaves can startle a potential predator. Leaffolding can also prevent water loss by reducing the surfacearea available for transpiration. Some localized plant move-ments are triggered by unpredictable environmental sig-nals. Other movements are tied into daily internal rhythmsestablished by cyclic environmental signals like light andtemperature. Plants lack a nervous system in the conven-tional sense. Some of the rapid signaling, however, is theresult of electric charge moving through an organ as a waveof membrane ion exchange, not unlike that seen in animals.This is translated into movement by changing the turgorpressure of cells.
Turgor MovementTurgor is pressure within a living cell resulting from diffu-sion of water into it. If water leaves turgid cells (ones with
turgor pressure), the cells may collapse, causing plantmovement; conversely, water entering a limp cell may alsocause movement as the cell once more becomes turgid.
Many other plants, including those of the legume family(Fabaceae), exhibit leaf movements in response to touch orother stimuli. After exposure to a stimulus, the changes inleaf orientation are mostly associated with rapid turgorpressure changes in pulvini (singular: pulvinus), which aremulticellular swellings located at the base of each leaf orleaflet. When leaves with pulvini, such as those of the sen-sitive plant (Mimosa pudica), are stimulated by wind, heat,touch, or, in some instances, intense light, an electrical sig-nal is generated. The electrical signal is translated into achemical signal, with potassium ions, followed by water,migrating from the cells in one half of a pulvinus to the in-tercellular spaces in the other half. The loss of turgor inhalf of the pulvinus causes the leaf to “fold.” The move-ments of the leaves and leaflets of the sensitive plant are es-pecially rapid; the folding occurs within a second or twoafter the leaves are touched (figure 41.34). Over a span ofabout 15 to 30 minutes after the leaves and leaflets havefolded, water usually diffuses back into the same cells fromwhich it left, and the leaf returns to its original position.
832 Part XI Plant Growth and Reproduction
41.4 Many short-term responses to the environment do not require growth.
PulvinusVascular tissue
Cells retainingturgor
Cells losingturgor
(a) (b)
FIGURE 41.34Sensitive plant (Mimosa pudica). (a) Theblades of Mimosa leaves are divided intonumerous leaflets; at the base of each leafletis a swollen structure called a pulvinus. (b) Changes in turgor cause leaflets to foldin response to a stimulus. (c) When leavesare touched (center two leaves above), theyfold due to loss of turgor.

The leaves of some plants with similar mechanisms maytrack the sun, with their blades oriented at right angles toit; how their orientation is directed is, however, poorly un-derstood. Such leaves can move quite rapidly (as much as15° an hour). This movement maximizes photosynthesisand is analogous to solar panels that are designed to trackthe sun.
Some of the most familiar of these reversible changesare seen in leaves and flowers that “open” during the dayand “close” at night. For example, the flowers of fouro’clocks open at 4 P.M. and evening primrose petals open atnight. The blades of plant leaves that exhibit such a dailyshift in position may not actually fold; instead, their orien-tation may be changed as a result of turgor movements.Bean leaves are horizontal during the day when their pul-vini are turgid, but become more or less vertical at night asthe pulvini lose turgor (figure 41.35). These sleep move-ments reduce water loss from transpiration during thenight, but maximize photosynthetic surface area during theday. In these cases, the movement is closely tied to an in-ternal rhythm.
Circadian Clocks
How do leaves know when to “sleep”? They have endoge-nous circadian clocks that set a rhythm with a period ofabout 24 hours (actually it is closer to 22 or 23 hours).While there are shorter and much longer, naturally occur-
ring rhythms, circadian rhythms are particularly commonand widespread because of the day-night cycle on earth.Jean de Mairan, a French astronomer, first identified circa-dian rhythms in 1729. He studied the sensitive plant which,in addition to having a touch response, closes its leafletsand leaves at night like the bean plant described above.When de Mairan put the plants in total darkness, they con-tinued “sleeping” and “waking” just as they did when ex-posed to night and day. This is one of four characteristicsof a circadian rhythm: it must continue to run in the ab-sence of external inputs. It must be about 24 hours in dura-tion and can be reset or entrained. (Perhaps you’ve experi-enced entrainment when traveling to a different time zonein the form of jet-lag recovery.) The fourth characteristic isthat the clock can compensate for differences in tempera-ture. This is quite unique when you consider what youknow about biochemical reactions; most rates of reactionsvary significantly based on temperature. Circadian clocksexist in many organisms and appear to have evolved inde-pendently multiple times. The mechanism behind the clockis not fully understood, but is being actively investigated atthe molecular level.
Turgor movements of plants are reversible and involvechanges in the turgor pressure of specific cells.Circadian clocks are endogenous timekeepers that keepplant movements and other responses synchronizedwith the environment.
Chapter 41 How Plants Grow in Response to Their Environment 833
FIGURE 41.35Sleep movements in bean leaves. Inthe bean plant, leaf blades are orientedhorizontally during the day andvertically at night.
12:00 NOON 3:00 P.M.
10:00 P.M. 12:00 MIDNIGHT

Plant Defense ResponsesInteractions between plants and other organisms can besymbiotic (for example, nitrogen-fixing bacteria and mycorrhizae) or pathogenic. In evolutionary terms, thesetwo types of interactions may simply be opposite sides ofthe same coin. The interactions have many common as-pects and are the result of coevolution between two speciesthat signal and respond to each other. In the case ofpathogens, the microbe or pest is “winning,” at least forthat second in evolutionary time. In chapter 38, we dis-cussed surface barriers the plant constructs to block inva-sion. In this section, we will focus on cellular level re-sponses to attacks by microbes and animals.
Recognizing the Invader
Half a century ago, Flor proposed that there is a plant re-sistance gene (R) whose product interacts with that of apathogen avirulence gene (avr). This is called the gene-for-gene model and several pairs of avr and R genes have beencloned in different species pathogenized by microbes,fungi, and insects, in one case. This has been motivatedpartially by the agronomic benefit of identifying genes thatcan be added to protect other plants from invaders. Muchis now known about the signal transduction pathways thatfollow the recognition of the pathogen by the R gene.These pathways lead to the triggering of the hypersensitiveresponse (HR) which leads to rapid cell death around thesource of the invasion and also a longer-term resistance(figure 41.36). There is not always a gene-for-gene re-sponse, but plants still have defense responses to pathogensand also mechanical wounding. Some of the response path-ways may be similar. Also, oligosaccharins in the cell wallsmay serve as recognition and signaling molecules.
While our focus is on invaders outside the plant king-dom, more is being learned about how parasitic plants in-vade other plants. There are specific molecules releasedfrom the root hairs of the host that the parasitic plant rec-ognizes and responds to with invasive action. Less is knownabout the host response and so far the different defensegenes that are activated appear to be ineffective.
Responding to the Invader
When a plant is attacked and there is gene-for-gene recog-nition, the HR response leads to very rapid cell deatharound the site of attack. This seals off the wounded tissue
to prevent the pathogen or pest from moving into the restof the plant. Hydrogen peroxide and nitric oxide are pro-duced and may signal a cascade of biochemical events re-sulting in the localized death of host cells. They may alsohave negative effects on the pathogen, although antioxidantabilities have coevolved in pathogens. Other antimicrobialagents produced include the phytoalexins which are chemi-cal defense agents. A variety of pathogenesis-related genes(PR genes) are expressed and their proteins can either func-tion as antimicrobial agents or signals for other events thatprotect the plant.
In the case of virulent invaders (no R gene recognition),there are changes in local cell walls that at least partiallyblock the movement of the pathogen or pest farther intothe plant. In this case there is not an HR response and thelocal plant cells are not suicidal.
When an insect takes a mouthful of a leaf, defense responses are also triggered. Mechanical damage causes re-sponses that have some similar components, but the reac-tion may be slower. Biochemically, it is distinct from someof the events triggered by signals in the insect’s mouth.Such responses are collectively called wound responses.Wound responses are a challenge in designing other typesof experiments with plants that involve cutting or otherwisemechanically damaging the tissue. It is important to runcontrol experiments to be sure you are answering yourquestion and not observing a wound response.
Preparing for Future Attacks
In addition to the HR or other local responses, plants arecapable of a systemic response to a pathogen or pest at-tack. This is called a systemic acquired response (SAR).Several pathways lead to broad-ranging resistance thatlasts for a period of days. The signals that induce SAR in-clude salicylic acid and jasmonic acid. Salicylic acid is theactive ingredient in aspirin too! SAR allows the plant torespond more quickly if it is attacked again. However, thisis not the same as the human immune response where an-tibodies (proteins) that recognize specific antigens (for-eign proteins) persist in the body. SAR is neither as spe-cific or long lasting.
Plants defend themselves from invasion in waysreminiscent of the animal immune system. When aninvader is recognized, localized cell death seals off theinfected area.
834 Part XI Plant Growth and Reproduction

Chapter 41 How Plants Grow in Response to Their Environment 835
Plantcell
Plantcells
HRR protein
Microbialprotein
Hypersensitive response (HR):local cell death seals off pathogen
Systemic acquired response (SAR):temporary broad-ranging resistance to pathogen
Pathogenicmicrobial
attack
Signalmolecule
Signalmolecule
SAR
FIGURE 41.36Plant defense responses. In the gene-for-gene response, a cascade of events is triggered leading to local cell death (HR response) andlonger-term resistance in the rest of the plant (SAR).
836 Part XI Plant Growth and Reproduction
Chapter 41Summary Questions Media Resources
41.1 Plant growth is often guided by environmental cues.
• Tropisms in plants are growth responses to externalstimuli, such as light, gravity, or contact.
• Dormancy is a plant adaptation that carries a plantthrough unfavorable seasons or periods of drought.
1. In general, which part of aplant is positively phototropic?What is the adaptive significanceof this reaction?
• Auxin migrates away from light and promotes theelongation of plant cells on the dark side, causingstems to bend in the direction of light.
• Cytokinins are necessary for mitosis and cell divisionin plants. They promote growth of lateral buds andinhibit formation of lateral roots.
• Gibberellins, along with auxin, play a major role instem elongation in most plants. They also tend tohasten the germination of seeds and to breakdormancy in buds.
2. How does auxin affect theplasticity of the plant cell walls? 3. Where are most cytokininsproduced? From whatbiomolecule do cytokininsappear to be derived?4. What plant hormones couldbe lacking in genetically dwarfedplants?
41.2 The hormones that guide growth are keyed to the environment.
• The transition of a shoot meristem from vegetative toadult development is called phase change. Duringphase change, plants gain competence to produce afloral signal(s) and or perceive a signal.
• The light-dependent pathway uses information fromlight receptor molecules integrated with a biologicalclock to determine if the length of night is sufficientfor flowering.
• The autonomous path functions independently ofenvironmental cues. Internal floral inhibitor(s) fromroots and leaves and floral promoter(s) from leavesmove through the plant.
5. A plant has undergone phasechange. Although it is an adult,it does not flower. How mightyou get this plant to flower?6. You have recently movedfrom Canada to Mexico andbrought some seeds from yourfavorite plants. They germinateand produce beautiful leaves, butnever flower. What went wrong?
41.3 The environment influences flowering.
• Changes in turgor pressure reflect responses toenvironmental signals that can protect plants frompredation.
• Other reversible movements in plants are caused bychanges in turgor pressure that are regulated byinternal circadian rhythms.
• Plants have the ability to recognize and respond toinvaders through cellular level recognition andresponse.
7. How are motor cells involvedin the function of the pulvinus?What happens in the motor cellsof the sensitive plant (Mimosapudica) when its leaves aretouched?8. In what ways can a plantprotect itself from pathogenicmicrobes? From animals?
41.4 Many short-term responses to the environment do not require growth.
www.mhhe.com/raven6e www.biocourse.com
• Photoperiod
• Hormones
• Student Research:Plant Growth
• Student Research:Selection in FloweringPlants


















