
28 August 2018 ISSN 1996-0808 DOI: 10.5897/AJMR www ...
41
28 August 2018 ISSN 1996-0808 DOI: 10.5897/AJMR www.academicjournals.org OPEN ACCESS African Journal of Microbiology Research
Transcript of 28 August 2018 ISSN 1996-0808 DOI: 10.5897/AJMR www ...

28 August 2018ISSN 1996-0808 DOI: 10.5897/AJMRwww.academicjournals.org
OPEN AC C ESS
African Journal of
Microbiology Research

The African Journal of Microbiology Research (AJMR) is published weekly (one volume per year) by Academic Journals.
provides rapid publication (weekly) of articles in all areas of Microbiology such as: Environmental Microbiology, Clinical Microbiology, Immunology, Virology, Bacteriology, Phycology, Mycology and Parasitology, Protozoology, Microbial Ecology, Probiotics and Prebiotics, Molecular Microbiology, Biotechnology, Food Microbiology, Industrial Microbiology, Cell Physiology, Environmental Biotechnology, Genetics, Enzymology, Molecular and Cellular Biology, Plant Pathology, Entomology, Biomedical Sciences, Botany and Plant Sciences, Soil and Environmental Sciences, Zoology, Endocrinology, Toxicology. The Journal welcomes the submission of manuscripts that meet the general criteria of significance and scientific excellence. Papers will be published shortly after acceptance. All articles are peer-reviewed.
Contact Us
Editorial Office: [email protected]
Help Desk: [email protected]
Website: http://www.academicjournals.org/journal/AJMR
Submit manuscript online http://ms.academicjournals.me/

Editors
Prof. Adriano Gomes da Cruz University of Campinas (UNICAMP), Brazil. Prof. Ashok Kumar School of Biotechnology Banaras Hindu UniversityUttar Pradesh, India. Dr. Mohd Fuat Abd Razak Infectious Disease Research Centre, Institute for Medical Research, Jalan Pahang, Malaysia. Dr. Adibe Maxwell Ogochukwu Department of Clinical Pharmacy and Pharmacy Management, University of Nigeria Nsukka, Nigeria. Dr. Mehdi Azami Parasitology & Mycology Department Baghaeei Lab. Isfahan, Iran. Dr. Franco Mutinelli Istituto Zooprofilattico Sperimentale delle Venezie Italy. Prof. Ebiamadon Andi Brisibe University of Calabar, Calabar, Nigeria.
Prof. Nazime Mercan Dogan Department of Biology Faculty of Science and Arts University Denizli Turkey. Prof. Long-Liu Lin Department of Applied Chemistry National Chiayi University Chiayi County Taiwan. Prof. Natasha Potgieter University of Venda South Africa. Dr. Tamer Edirne Department of Family Medicine University of Pamukkale Turkey. Dr Kwabena Ofori-Kwakye Department of Pharmaceutics Kwame Nkrumah University of Science & Technology Kumasi, Ghana. Dr. Tülin Askun Department of Biology Faculty of Sciences & Arts Balikesir University Turkey. Dr. Mahmoud A. M. Mohammed Department of Food Hygiene and Control Faculty of Veterinary Medicine Mansoura University Egypt.

Dr. James Stefan Rokem Department of Microbiology & Molecular Genetics Institute of Medical Research Israel – Canada The Hebrew University – Hadassah Medical School Jerusalem, Israel. Dr. Afework Kassu University of Gondar Ethiopia. Dr. Wael Elnaggar Faculty of Pharmacy Northern Border University Rafha Saudi Arabia. Dr. Maulin Shah Industrial Waste Water Research Laboratory Division of Applied & Environmental Microbiology, Enviro Technology Limited Gujarat, India. Dr. Ahmed Mohammed Pathological Analysis Department Thi-Qar University College of Science Iraq. Prof. Naziha Hassanein Department of Microbiology Faculty of Science Ain Shams University Egypt. Dr. Shikha Thakur Department of Microbiology Sai Institute of Paramedical and Allied Sciences India.
Dr. Samuel K Ameyaw Civista Medical Center USA. Dr. Anubrata Ghosal Department of Biology MIT - Massachusetts Institute of Technology USA. Dr. Bellamkonda Ramesh Department of Food Technology Vikrama Simhapuri University India. Dr. Sabiha Yusuf Essack Department of Pharmaceutical Sciences University of KwaZulu-Natal South Africa. Dr. Navneet Rai Genome Center University of California Davis USA. Dr. Iheanyi Omezuruike Okonko Department of Virology Faculty of Basic Medical Sciences University of Ibadan Ibadan, Nigeria. Dr. Mike Agenbag Municipal Health Services, Joe Gqabi, South Africa. Dr. Abdel-Hady El-Gilany Department of Public Health & Community Medicine, Faculty of Medicine Mansoura University Egypt.

African Journal of Microbiology Research
Table of Content: Volume 12 Number 32 28 August, 2018
ARTICLES
Molecular detection and characterization of Escherichia coli, Salmonella spp. and Campylobacter spp. isolated from broiler meat in Jamalpur, Tangail, Netrokona and Kishoreganj districts of Bangladesh Md. Kamrul Islam, S. M. Lutful Kabir, A. K. M. Ziaul Haque, Y. A. Sarker and M. H. Sikder Screening of human diarrhoeal samples in Mymensingh city of Bangladesh for the isolation, identification and antimicrobial resistance profiles of Campylobacter spp. Sudarsan Karmaker, S. M. Lutful Kabir, A. K. M. Ziaul Haque, Mohammad Ferdousur Rahman Khan and Yousuf Ali Sarker Assessment of dominant bacterial strains isolated from Ntoba mbodi, an indigenous African alkaline-fermented food, and their potential enzyme activities Didine Priscilla Moutou-Tchitoula, Etienne Nguimbi, Stéphanie Giusti-Miller, Philippe Mora, Simon Charles Kobawila, Edouard Miambi Isolation and characterization of starch degrading rhizobacteria from soil of Jimma University Main Campus, Ethiopia Gudeta Dida

Vol. 12(32), pp. 761-770, 28 August, 2018
DOI: 10.5897/AJMR2018.8945
Article Number: 4BE680558399
ISSN: 1996-0808
Copyright ©2018
Author(s) retain the copyright of this article
http://www.academicjournals.org/AJMR
African Journal of Microbiology Research
Full Length Research Paper
Molecular detection and characterization of Escherichia coli, Salmonella spp. and Campylobacter spp. isolated from broiler meat in Jamalpur, Tangail, Netrokona and
Kishoreganj districts of Bangladesh
Md. Kamrul Islam1, S. M. Lutful Kabir1*, A. K. M. Ziaul Haque1, Y. A. Sarker2 and M. H. Sikder2
1Department of Microbiology and Hygiene, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh.
2Department of Pharmacology, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh.
Received 23 July, 2018; Accepted 13 August, 2018
The study was conducted to isolate, identify and characterize bacterial samples from broiler meat collected from 20 different upazilla markets of Jamalpur, Tangail, Netrokona and Kishoreganj districts of Bangladesh. A total of 20 samples were subjected to bacteriological isolation and identification, and the isolated bacteria were subjected to antimicrobial susceptibility testing using disc diffusion method. Among the samples, 75% (n=15) were contaminated with Campylobacter spp., 70% (n=14) were with Salmonella species and 85% (n=17) were contaminated with Escherichia coli. The Campylobacter spp., Salmonella spp. and E. coli were isolated and identified by culturing on Blood agar, Xylose-Lysine Deoxycholate (XLD) agar, and MacConky and eosin methylene blue (EMB) agar respectively. Isolates of Campylobacter spp., Salmonella spp. and E. coli were biochemically analyzed. Campylobacter specific 16S rRNA genes were amplified from the isolates. Campylobacter spp. and E. coli isolates were positive to 16S rRNA gene based polymerase chain reaction (PCR). Almost all isolates of Campylobacter spp., Salmonella spp. and E. coli showed highest susceptibility to ciprofloxacin, norfloxacin and gentamicin. However, most isolates were resistant to amoxicillin and erythromycin. Some isolates showed susceptibility to tetracycline, streptomycin and azithromycin. The findings of this study revealed that there is presence of multidrug resistant isolates of Campylobacter spp., Salmonella spp. and E. coli in broiler meat. Results of this research project demonstrated the high level of microbial contamination and occurrence of pathogenic bacteria in broiler meat sold in markets of Bangladesh. Key words: Broiler meat, Escherichia coli, Salmonella, Campylobacter, molecular detection, characterization.
INTRODUCTION Poultry industry which has started during 1980s is an excellent agribusiness (Haque, 2001) now in Bangladesh.
Over the last decades surprising development has occurred in this sector (Rahman, 2003). It has become a
*Corresponding author. E-mail: [email protected], [email protected]. Tel: +8801754987218.
Author(s) agree that this article remain permanently open access under the terms of the Creative Commons Attribution
License 4.0 International License

762 Afr. J. Microbiol. Res. vital sector for generating employment, creating additional income and improving the nutritional level of the country. Broiler meat is more popular to the consumers because of its easy digestibility and acceptance by the majority of people, although it could be contaminated with various potential food borne pathogens such as Salmonella, Campylobacter, Escherichia coli (Mulder et al., 1999). Broiler entering slaughter processing is highly contaminated by microorganisms such as Salmonella and Campylobacter and tends to be disseminated during processing (Mead et al., 1994).
Particular concern for human health is the inappropriate use of antibiotics in poultry production and the development of antibiotic resistant strains of bacteria (Sarker et al., 2018). Effective control systems are critical in ensuring product safety, and considerable information is available on how to minimize the risks (FAO, 2013). Various pathogenic microbes, such as E. coli, Salmonella spp. and Staphylococcus have been implicated to reduce the growth of poultry including duck.
Campylobacter is one of the most important pathogen and is regarded major bacterial cause of human gastroenteritis worldwide. Food animals, mainly poultry, cattle, sheep and pigs, may act as asymptomatic intestinal carriers of Campylobacter and animal food products can become contaminated by this pathogen during slaughter and carcass dressing
(Berndtson et al.,
1996). The abusive use of antimicrobials in food animals has resulted in the emergence and dissemination of antimicrobial resistant bacteria, including antimicrobial resistant Campylobacter, which has potentially serious impact on human health. Moreover, Campylobacter infections pose a serious public health problem for which many countries have monitored their infection and antimicrobial resistance patterns (Kabir et al., 2014). E. coli frequently cause bacterial infections including urinary tract infection, cholangitis, bacterimia and traveler’s diarrhea. Enteropathogenic E. coli (EPEC) are an important cause of diarrhea in humans (Savkovic et al., 2005). Haemolytic uramic syndrome caused by Shiga toxin producing E. coli (STEC) is dependent on release of Shiga toxin during intestinal infection and subsequent absorption into the blood stream. Poultry meat can be contaminated with E. coli during processing. Any food that has been in contact with raw meat can also be contaminated. The bacteria also spread from person to person, usually when infected person does not wash his hands well after using a toilet. E. coli cause different types of public health hazards including cholangitis, bacteremia, traveler’s diarrhea, Shiga toxicity etc. (Savkovic et al., 2005).
Salmonella spp. is potentially responsible for various pathogenic processes in man and animal including poultry (Freeman, 1985). It can cause diarrhea, vomition, fever, abdominal cramps in human. Sometimes severe diarrhea requires medical interventions such as
intravenous fluid therapy. In cases, where bacteria enter into the bloodstream, symptoms include high fever, malaise, pain in the thorax and abdomen, chills and anorexia (Bell, 2002).
Antibiotics are extensively used in poultry industry either as growth promoters or to control infectious diseases (Sarker et al., 2018). Concern about antibiotic resistance and its transmission to human is important because these resistant bacteria may colonize the human gastrointestinal tract and may contribute to the development of resistance genes to human through R-factor, conjugative plasmids or chromosomal elements as reviewed by Kabir (2010). Therefore, the disease causing microbes that have become resistant to antibiotic drug therapy are increasing public health importance.
Undoubtedly, the poultry slaughtered and dressed under Bangladesh conditions carry extremely high initial contamination loading from the point of slaughtering process to the point at which the consumers are offered the product. There occurs biomagnifications at all levels of handling, poor transport and retailing conditions. Therefore, considering the present perspective, the present research project was designed to detect and characterize the bacteria specifically Campylobacter spp. Salmonella spp. and E. coli and their antimicrobial resistance patterns in broiler meat.
MATERIALS AND METHODS Collection and transportation of samples A total of 20 apparently healthy dressed broilers were collected randomly from 20 different upazilla live bird markets of Jamalpur, Tangail, Kishoreganj and Netrokona district of Bangladesh. Five upazilla were selected randomly from each district. After collection, samples were immediately brought to Bacteriology Laboratory of the Department of Microbiology and Hygiene, Bangladesh Agricultural University, Mymensingh through maintaining cool chain using ice box.
Bacterial culture media Solid and liquid culture media were used to isolate the bacteria. Blood agar (BA), MacConkey (MC), Salmonella-Shigella (SS), Eosin Methylene Blue (EMB), Xylose-Lysine Deoxycholate (XLD), Mueller Hinton agars were used as solid culture media for this study. The liquid media used in the study were nutrient broth (NB), peptone broth, methyl-red and voges-proskauer broth (MR-VP), and sugar media, 1% hippurate solution, 3.5% ninhydrin solution, oxidase solution and sugar media.
Isolation and identification of bacteria
Pure culture of E. coli and Salmonella spp. were obtained as per the methods described by Krieg et al. (1994). Briefly; 10 g of samples were homogenized with 90 ml of 0.1% peptone water and 50 µl of homogenized sample was poured on to selective agar media and spread with glass spreader and incubated at 37°C for 24 h. Isolation of Campylobacter spp. was carried out by filtration method (0.45 μm filter) according to Shiramaru et al. (2012). The

Islam et al. 763 Table 1. List of primers used in this study.
Primer Sequence (5'-3') Target Amplicon
size (bp) References
16S9F GAGTTTGATCCTGGCTC Campylobacter
16S rRNA gene 1530
Samosornsuk
et al. (2007) 16S154OR AAGGAGGTGATCCAGCC
Upper strand ACTGGCGTTATCCCTCTGGTG Histidine transport
Operon gene 496 Cohen et al. (1993)
Lower strand ATGTTGTCCTGCCCCTGGTAAGAGA
ECO-1 GACCATCGGTTTAGTTCACAGA E. coli 16S rRNA gene 585 Schippa et al. (2010)
ECO-2 CACACGCTGACGCTGACCA
collected samples were allowed to prepare meat homogenates and then 50 μl of meat homogenates were spread on the filter papers that were placed on the surface of Blood base agar no.2 and allowed to stand for 30 min at room temperature, after 30 min filter papers were removed from the BA and the plates were incubated at 37°C for 48 h in microaerobic condition (5% O2, 10% CO2 and 85% N2). The colonies of primary cultures were repeatedly sub-cultured by streak plate method (Cheesbrough, 1985) until the pure cultures with homogenous colonies appeared. The representative bacterial colonies were characterized morphologically using Gram's stain according to the method describe by Merchant and Packer (1967). Biochemical characterizations of the E. coli and Salmonella isolates were performed with Sugar fermentation test, Methyl Red test (MR), Voges-Proskauer test (V-P) and indole test (Cheesbrough, 1985). Differentiation of isolated Campylobacter spp. with supporting growth characteristics were subjected to various biochemical tests such as catalase, oxidase and hippurate hydrolysis test according to the procedures followed by Nachamkin (2003) and Foster et al. (2004). Molecular identification by polymerase chain reaction (PCR) DNA template was prepared by boiling method as described by Rawool et al. (2007). All the samples were examined by two pairs of primers (Table 1) to detect 16S rRNA gene of Campylobacter spp., E. coli and Histidine transport operon gene of Salmonella spp. For Campylobacter spp. the PCR reactions were carried out using a thermocycler (ASTEC, Japan) with the following programme: initial denaturation for 5 minutes at 94°C, followed by 30 cycles of denaturation at 94°C for 30 s, annealing at 47°C for 30 s and extension at 72°C for 1 min and 30 s. The final extension was conducted at 72°C for 10 min. For E. coli, the PCR reactions were carried out using a thermocycler (ASTEC, Japan) with the following programme: Initial denaturation for 5 min at 95°C followed by 30 cycles of denaturation at 94°C for 45 s, annealing at 55°C for 45 s and extension at 72°C for 1 min. The final extension was conducted at 72°C for 5 min. For Histidine transport operon gene identification in Salmonella spp. initial denaturation for 5 min at 94°C, followed by 30 cycles of denaturation at 94°C for 30 s, annealing at 56°C for 30 s and extension at 72°C for 45 s. The final extension was conducted at 72°C for 5 min. 1 and 2% agarose (Invitrogen, USA) gel was used for electrophoresis of the PCR products.
Antibiotic sensitivity test
All isolates randomly selected from the three genera were tested for antimicrobial drug susceptibility against eight commonly used
antibiotics by disc diffusion method according to the guidelines of Clinical and Laboratory Standard Institute (CLSI), 2012. The selected antibiotics used were ciprofloxacin (5 μg/disc), azithromycin (30 μg/disc), amoxicillin (30 μg/disc), gentamicin (10 μg/disc), Norfloxacin (10 μg/disc), erythromycin (30 μg/disc), streptomycin (10 μg/disc), and tetracycline (30 μg/disc). The interpretation on susceptibility was done according to the guidelines of Clinical and Laboratory Standard Institute (CLSI, 2007) (Table 1).
RESULTS Isolation of bacteria from poultry meat A total of 20 samples were collected for isolation of bacteria from broiler meat of different upazilla markets of four districts where three types of bacteria were isolated from the collected meat samples. The isolates were identified as E. coli, Salmonella spp. and Campylobacter spp. on the basis of their morphological, cultural properties, biochemical characteristics with standard reference organisms and molecular methods. Among the isolated bacteria, E. coli were detected in 85% (n=17), Salmonella and Campylobacter in 70% (n=14) and 75% (n=15), respectively of the samples. The highest percentage of E. coli (100%) isolates was observed in Jamalpur and Tangail district. Salmonella isolates were higher (80%) in samples of Jamalpur, Tangail and Kishoreganj. Furthermore, Campylobacter isolates were 100% in samples of Jamalpur District. Molecular detection of bacteria Screening of E. coli isolates from meat sample in this study was performed by genus specific (16S rRNA gene) polymerase chain reaction (PCR). The PCR assay was able to amplify 585 bp fragment of the targeted gene from the genomic DNA of E. coli successfully (Figure 1). Specific 496 bp fragment of targeted histidine transport operon gene of Salmonella was amplified successfully (Figure 2). Genus specific (16S rRNA gene) PCR was performed for Campylobacter spp. 1530 bp fragment of

764 Afr. J. Microbiol. Res.
Figure 1. PCR targeting 16s rRNA gene for identification of E. coli. Lane 1: 100 bp DNA marker, lane 2, 3, 4, 5, 6: DNA of E. coli, lane 7: negative control without DNA.
Figure 2. Detection of Salmonella spp. by histidine transport operon gene based PCR. Lane 1: 100 bp DNA marker, lane 2, 3, 4: DNA of Salmonella.
presented in Figure 3.
Figure 1. PCR targeting 16s rRNA gene for identification of E. coli.
1 2 5 6 3 4 7
585 bp
control without DNA.
Figure 2. Detection of Salmonella spp. by histidine transport operon gene
based PCR. Lane 1: 100 bp DNA marker, lane 2, 3,
1 2 3 4
496 bp

Islam et al. 765
Figure 3. Detection of Campylobacter spp. by 16S rRNA gene based PCR. Lane 1: 100 bp DNA marker, lane 2, 3, 4, 5: DNA of Campylobacter spp. and lane 6: negative control.
targeted gene was amplified successfully. The result of PCR is presented in Figure 3. Antimicrobial susceptibility of isolated E. coli, Salmonella and Campylobacter spp. A total of 17 E. coli isolates were subjected to antimicrobial susceptibility testing. Most of the isolates were resistant to erythromycin (70.58%) and amoxicillin (88.24%) and few isolates were intermediate. Almost all the isolates of E. coli showed their highest sensitivity to gentamicin (76.47%), norfloxacin (82.35%), ciprofloxacin (82.35%) and azithromycin (64.70%). Some isolates of E. coli were resistant and some were sensitive to tetracycline. The result is presented in Table 2. A total of 14 Salmonella isolates were subjected to antimicrobial susceptibility testing against eight selected antibiotics. Almost all isolates of Salmonella spp. were resistant to tetracycline (85.71%) and erythromycin (64.28%) whereas most of the isolates of Salmonella spp. were susceptible to gentamicin (92.85%), norfloxacin (78.57%) and ciprofloxacin (78.57%) respectively. 5 (35.71%) were sensitive to streptomycin and 10 (71.42%) isolates were intermediate to amoxicillin. Results are presented in Table 2.
The result of antimicrobial susceptibility of Campylobacter jejuni and Campylobacter coli are summarized in Table 2. Out of 11 C. jejuni isolates, 10 (90.90%) isolates were susceptible to gentamicin, 9 (81.81%) were sensitive to norfloxacin, 8 (72.72%) were
sensitive to ciprofloxacin and 7 (63.63%) were susceptible to streptomycin. Most of the isolates were resistant to amoxicillin (81.81%) and 9 (81.81%) isolates were intermediate resistant to erythromycin. On the other hand, out of four C. coli isolates, all (100%) were sensitive to both ciprofloxacin and norfloxacin. 3 (75%) were sensitive to gentamicin, and 4 (100%) were resistant to amoxicillin, 2 were resistant to streptomycin. Antimicrobial resistance pattern Antimicrobial resistance pattern of E. coli The results of the antimicrobial resistance pattern by disk diffusion method with 8 chosen antimicrobial agents are presented in Table 3. Out of 17 isolates, 6 (35.29%) were resistant to 1 antimicrobial agent. Each 1 (5.88%) was resistant to each of 2 antibiotics. Moreover, 2 (11.76%) were resistant to 2 antibiotics. Furthermore, each 1 (5.88%) was resistant to each of 3 antibiotics. Antimicrobial resistance pattern of Salmonella spp. The results of the antimicrobial resistance pattern by disk diffusion method with 8 chosen antimicrobial agents are presented in Table 4. Out of 14 isolates no resistance observed in 1 isolate. 1 (7.14%) was resistant to 1 antibiotics. Furthermore, each 2 (14.28%) were resistant to each of 2 antibiotics. Moreover, 1 (7.14%), 2 (14.28%),
Figure 3. Detection of Campylobacter spp. by 16S rRNA gene based PCR.
1530 bp
1 2 3 4 5 6

766 Afr. J. Microbiol. Res.
Table 2. Antimicrobial susceptibility pattern of E. coli, Salmonella spp., C. jejuni and C. coli identified by disk diffusion method.
Antimicrobial agents No. (%) of isolates
S I R
E. coli
Amoxicillin 0 (0.0%) 2 (11.76%) 15 (88.24%)
Azithromycin 11(64.70%) 4 (23.52%) 2 (11.76%)
Ciprofloxacin 14 (82.35%) 3 (17.64%) 0 (0.0%)
Erythromycin 0 (0.0%) 5 (29.41%) 12 (70.58%)
Gentamicin 13 (76.47%) 2 (11.76%) 2 (11.76%)
Norfloxacin 14 (82.35%) 2 (11.76%) 1 (5.88%)
Streptomycin 4 (23.52%) 10 (58.82%) 3 (17.64%)
Tetracycline 9 (52.94%) 5 (29.41%) 3 (17.64%)
Salmonella spp.
Amoxicillin 0 (0.0%) 10 (71.42%) 4 (28.57%)
Azithromycin 3 (21.42%) 7 (50%) 4 (28.57%)
Ciprofloxacin 11 (78.57%) 2 (14.28%) 1 (7.14%)
Erythromycin 0 (0.0%) 5 (35.71%) 9 (64.28%)
Gentamicin 13 (92.85%) 1 (7.14%) 0 (0.0%)
Norfloxacin 11 (78.57%) 3 (21.42%) 0 (0.0%)
Streptomycin 5 (35.71%) 2 (14.28%) 7 (50%)
Tetracycline 0 (0.0%) 2 (14.28%) 12 (85.71%)
C. jejuni
Amoxicillin 0 (0.0%) 2 (18.18%) 9 (81.81%)
Azithromycin 6 (54.54%) 3 (27.27%) 2 (18.18%)
Ciprofloxacin 8 (72.72%) 2 (18.18%) 1 (9.09%)
Erythromycin 0 (0.0%) 9 (81.81%) 2 (18.18%)
Gentamicin 10 (90.90%) 1 (9.09%) 0 (0.0%)
Norfloxacin 9 (81.81%) 1 (9.09%) 1 (9.09%)
Streptomycin 7 (63.63%) 0 (0.00%) 4 (36.36%)
Tetracycline 3 (27.27%) 2 (18.18%) 6 (54.54%)
C. coli
Amoxicillin 0 (0.0%) 0 (0.0%) 4 (100%)
Azithromycin 2 (50.00%) 2 (50.00%) 0 (0.0%)
Ciprofloxacin 4 (100%) 0 (0.0%) 0 (0.0%)
Erythromycin 0 (0.0%) 3 (75.00%) 1 (25.00%)
Gentamicin 3 (75.00%) 1 (25.00%) 0 (0.0%)
Norfloxacin 4 (100%) 0 (0.0%) 0 (0.0%)
Streptomycin 2 (50.00%) 0 (0.00%) 2 (50.00%)
Tetracycline 0 (0.0%) 1 (25.00%) 3 (75.00%)
S= Susceptible; I= Intermediate; R= Resistant.
1 (7.14%) and 1 (7.14) were resistant to each of 3 antibiotics respectively. Furthermore, 1 (7.14%) and 1 (7.14%) were resistant to each of 4 antimicrobial agents. On the other hand, 1 was resistant to 5 antimicrobial agents. Antimicrobial resistance pattern of Campylobacter spp. The result of antimicrobial resistance patterns of C. jejuni and C. coli are summarized in Table 5. Out of 11 C. jejuni isolates, each 1 (9.09%) was resistant to each
1 antimicrobial agent. Furthermore, 3 (27.27%) and 1 (9.09%) were resistant to each of 2 antibiotics respectively. Moreover, 1 (9.09%) and 1 (9.09%) were resistant to each of 3 antibiotics. On the other hand, 1 (9.09%) and 1 (9.09%) were resistant to each of 3 and 4 antibiotics respectively. Out of 4 C. coli isolates, each 1 (25%) were resistant to each of 1 antibiotic. Furthermore, 2 (50%) were resistant to 3 antibiotics. DISCUSSION Meat products are important not only from nutritional

Islam et al. 767
Table 3. Results of antimicrobial resistance pattern of E. coli.
Isolates Resistance profiles No. of isolates (%)
E. coli (n=17)
No resistance demonstrated -
Resistant to 1 agent AMX 6 (35.29%)
Resistant to 2 agent (AMX-NOR) 1 (5.88%)
Resistant to 2 agent (S-TE) 1 (5.88%)
Resistant to 2 agents (AMX- S) 1 (5.88%)
Resistant to 2 agents (AMX-E) 2 (11.76%)
Resistant to 3 agents (AMX-E-S) 1 (5.88%)
Resistant to 3 agents (AMX-E-AZM) 1 (5.88%)
Resistant to 3 agents (E-GEN-TE) 1 (5.88%)
Resistant to 3 agents (AMX-E-TE) 1 (5.88%)
Resistant to 3 agents (E-AMX-GEN) 1 (5.88%)
Resistant to 3 agents (AMX-S-AZM) 1 (5.88%)
Total resistant isolates 17 (100%)
Table 4. Results of antimicrobial resistance pattern of Salmonella spp.
Isolates Resistance profiles No. of isolates (%)
Salmonella spp. (n=14)
No resistance demonstrated 1 (7.14%)
Resistant to 1 agent TE 1 (7.14%)
Resistant to 2 agent (E-TE) 2 (14.28%)
Resistant to 2 agent (S-TE) 2 (14.28%)
Resistant to 3 agents (E- AMX-TE) 1 (7.14%)
Resistant to 3 agents (S-E-TE) 2 (14.28%)
Resistant to 3 agents (AMX-AZM-TE) 1 (7.14%)
Resistant to 3 agents (E-AZM-TE) 1 (7.14%)
Resistant to 4 agents (AMX-E-S-TE) 1 (7.14%)
Resistant to 4 agents (E-AZM-S-TE) 1 (7.14%)
Resistant to 5 agents (AMX-S-E-CIP-AZM) 1 (7.14%)
Total resistant isolates 14 (100%)
Table 5. Results of antimicrobial resistance pattern of Campylobacter spp.
Isolates Resistance profiles No. of isolates (%)
C. jejuni (n=11)
No resistance demonstrated -
Resistant to 1 agent AMX 1 (9.09%)
Resistant to 1 agent (E) 1(9.09%)
Resistant to 1 agent (AZM) 1(9.09%)
Resistant to 2 agents (AMX- TE) 3 (27.27%)
Resistant to 2 agents (AMX-S) 1(9.09%)
Resistant to 3 agents (AMX-S-TE) 1 (9.09%)
Resistant to 3 agents (E-S-CIP) 1 (9.09%)
Resistant to 4 agents (AMX-NOR-AZM-TE) 1 (9.09%)
Resistant to 5 agents (AMX-S-E-AZM-TE) 1 (9.09%)
Total resistant isolates 11 (100%)
C. coli (n=4)
Resistant to 2 agent (AMX-TE) 1 (25%)
Resistant to 2 agent (AMX-E) 1 (25%)
Resistant to 3 agent (AMX-S-TE) 2 (50%)
Total resistant isolates 4 (100%)

768 Afr. J. Microbiol. Res. point of view, but also as an item of international trade and foreign exchange for a number of countries. However, they can also function as carriers of several microbial and other health hazards. The greatest risk to human health is due to the consumption of raw or contaminated meat and meat products.
In this study, E. coli, Salmonella and Campylobacter were found in the poultry meat, which was similar to the reports by Zhao et al. (2001), Ahmed et al. (2009), Awad-Alla et al. (2010), Torok et al. (2011), Voidarou et al. (2011), Sudershan et al. (2012), Malmuthuge et al. (2012) and Hossain et al. (2015). Several selective culture media were used simultaneously in this study to culture the organism. The media used in this study were selected considering the experience of the past researcher worked in various fields relevant to the present study by Kabir et al. (2014). The colony characteristics of Campylobacter spp. were gray color spreading colony which was supported by (Doyle, 1990; Rowe and Madden, 2000). In Gram's staining, the morphology of the isolated Campylobacter spp. was also supported by Doyle (1990). The colony characteristics of E. coli observed in Mac conkey, EMB agar were similar to the findings of Sharada et al. (1999). And in gram staining isolated bacteria showed pink color small rod shaped organism which was reported by others previously Sharada et al. (1999) and Merchant and Packer (1967). The colony morphology of Salmonella spp. was similar to that of Sarker et al. (2009) and Khan et al. (2005). In Gram's staining, the morphology of the isolated bacteria exhibited Gram negative small rod arranged in single or paired and motile which was supported by Sharada et al. (1999).
In catalase test, all the isolates (n = 15) produced bubbles those indicated positive for Campylobacter. In oxidase test, a purple color change was observed within ten seconds in all the isolates (n=15). In hippurate hydrolysis test, some of the isolates (n=4) develop a faint purple to no color change that indicated the isolates were C. coli and some of the test isolates (n=11) developed deep purple color that indicated the isolates were C. jejuni. These findings are similar to the findings of Kabir et al. (2014) and Jamshidi et al. (2008).
The E. coli isolates revealed a complete fermentation of 5 basic sugars by producing both acid and gas which was supported by Thomas (1998) and Beutin et al. (1991). The isolates also revealed positive reaction in MR test and Indole test but negative reaction in VP test (Honda et al., 1982). The antimicrobial susceptibility of most of the isolates was sensitive to ciprofloxacin; gentamicin and all the isolates were resistant to amoxycillin. Some of the isolates were multidrug resistant which was similar to the result of Kabir et al. (2014). In this study, it was observed that the E. coli isolates were sensitive to ciprofloxacin, norfloxacin and gentamicin. The results strengthen the earlier observations of Akond et al. (2009) and Islam et al. (2004). Resistance of E. coli was observed against
erythromycin, amoxycillin. The result was supported by Akond et al. (2009). The cause of such resistance by the E. coli might be for the fact that the organisms might have gained the resistance property due to the indiscriminate use of antibiotics.
The occurrence of isolation of bacterial pathogens from broiler should be considered as hazardous to health and advocate the preventing risk factors. However, in the present study ciprofloxacin were proved to be the best antibiotics to treat E. coli infection since they were highly effective. The results agreed with those reported by several investigations of Islam et al. (2004) and Ozaki et al. (2011) who also obtained similar resistant patterns of E. coli isolated from broiler. It was revealed that Salmonella spp. were sensitive to ciprofloxacin, gentamicin and norfloxacin. This result was supported by Jahan et al. (2013) and Khan et al. (2005) where isolates were sensitive to ciprofloxacin and chloramphenicol. The result is also consistent with Wouafo et al. (2010). The isolates were resistant to erythromycin, and amoxycillin which is similar to report of Hyeon et al. (2011) and Khan et al. (2005).
Conclusion In this study, bacteria were isolated from only 20 samples of 20 different upazila of Bangladesh. The findings of the present study revealed the presence of multidrug resistant bacteria in broiler meat sold in markets of Bangladesh. Most of the isolates showed resistance to amoxycillin but sensitive to ciprofloxacin and gentamicin. Some isolates showed multidrug resistance. CONFLICT OF INTERESTS The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
This study was performed in partial fulfillment of the requirements of a M.S. thesis for Md. Kamrul Islam from the Department of Microbiology and Hygiene, Bangladesh Agricultural University, Mymensingh, Bangladesh. We would also like to thank the Food and Agriculture Organization of the United Nations (FAO) for giving financial and logistic support to the present research project and management of this research. REFERENCES
Ahmed AM, Shimabukuro H, Shimamoto T (2009). Isolation and
molecular characterization of multidrug-resistant strains of E. coli and Salmonella from retail chicken meat in Japan. Journal of Food Science 74:405-410.

Akond MA, Hassan SMR, Alam S, Shirin M (2009). Antibiotic resistance
of E. coli isolated from poultry and poultry environment of Bangladesh. American Journal of Environmental Science 5:47-52.
Awad-Alla ME, Abdien HM, Dessouki AA (2010). Prevalence of bacteria and parasites in White Ibis in Egypt. Veterinaria Italians 46: 277-86.
Bell C (2002). Foodborne pathogens: Hazards, risk analysis and control. Woodhead Publishing and CRC Press pp. 307-335.
Berndtson E, Emanuelson U, Engvall A, Danielsson-Tham ML (1996). A 1-year epidemiological study of Campylobacters in 18 Swedish chicken farms. Preventive Veterinary Medicine 26:167-185.
Beutin L, Geier D, Zimmeronann S, Aleksic S, Gillespie HA, Whittarn TS (1991). Epidemiological relatednes and clonal types of natural populations of E. coli strains producing shiga toxin between stx genotype and Stx2 expression level in Shiga 96 toxin-producing E. coli 0157 strain in separate population of cattle and sheep. Applied and Environmental Microbiology 63:2175-2180.
Cheesbrough M (1985). Medical laboratory manual for tropical countries. Microbiology 2:400-480.
Clinical and Laboratory Standards Insitute (CLSI) (2007). M100-S17. Performance standards for antimicrobial susceptibility testing; 16th informational supplement. Clinical and Laboratory Standards Institute, Wayne, PA.
Clinical and Laboratory Standards Insitute (CLSI) (2012). Performance Standards for Antimicrobial Susceptebility Testing: Twenty-second Informational Supplement M100-S22. CLSI, Wayne, PA, USA.
Cohen ND, Neibergs HL, McGruder ED, Whitford H, Behle RW, Ray PM, Hargis BM (1993). Genus-specific detection of salmonellae using the polymerase chain reaction (PCR). Journal of Veterinary Diagnostic Investigation 5:368-371.
Doyle MP (1990). C. jejuni, p. 217-222. In D. O. Cliver (ed.), Foodborne diseases. Academic Press, Inc., Boston, MA.
FAO (2013). Food and Agricultural Organization of the United Nation report on Poultry and Human Health. Publication on Veterinary Public Health.
Foster G, Holmes B, Steigerwalt AG, Lawson PA, Thorne P, Byrer DE, Ross HM, Xerry J, Thompson PM, Collins MD (2004). Campylobacter insulaenigrae sp. nov., isolated from marine mammals. International Journal of Systematic and Evolutionary Microbiology 54: 2369-2373.
Freeman BA (1985). Burrows Textbook of Microbiology. In: W. B. Saunders company, philadephia, London, Toronto, Mexico city, Rio de Janerio, Sydney, Tokyo. 22: 464-475.
Haque QMF (2001). Poultry industry in Bangladesh and strategies for its improvement. In Proceedings of 2
nd International Poultry Show and
Seminar, February 2001, held in IDB Bhaban, Dhaka, Bangladesh. pp. 34-39.
Honda T, Arita M, Takela Y, Miwatani T (1982). Further evaluation of the Biken Test (Modified Eleck Test) for deletion of enterotoxigenic E. coli producing heat stable enterotoxin and application of the test to sampling of heat stable enterotoxin. Journal of Clinical Microbiology 16:60-62
Hossain M, Hoda N, Hossen MJ, Hassan MM, Rahman SME, Kabir SML (2015). Assessment of bacterial load of poultry meat used at dining hall of Bangladesh Agricultural University campus. Asian Journal of Medical and Biological Research 1:9-16.
Hyeon JY, Chon JW, Hwang IG, Kwak HS, Kim MS, Kim SK, Choi IS, Song CS, Park C, Seo KH (2011). Prevalence, antibiotic resistance, and molecular characterization of Salmonella serovars in retail meat products. Journal of Food Protection 74:161-6.
Islam MT, Islam MA, Samad MA, Kabir SML (2004). Charachterization and antibiogram of Eshcherechia coli associated with mortality in broilers and duckling in Bangladesh. Bangladesh Journal of Veterinary Medicine 2:9-14.
Jahan F, Kabir SML, Amin MM (2013). Identification and antimicrobial resistance profiles of Salmonellae isolated from the broiler dressing plants associated with their environments. Advanced Research Journal of Microbiology 1:001-009.
Jamshidi A, Bassami MR, Farkhondeh L (2008). Isolation and identification of Campylobacter spp. and C. coli from poultry carcasses by conventional culture method and multiplex PCR in Mashhad, Iran. Iranian Journal of Veterinary Research 9:132-137.
Kabir SML (2010). Avian Colibacillosis and Salmonellosis: A closer look at Epidemiology, pathogenesis, Diagnosis, Control and Public Health
Islam et al. 769
Concerns. International Journal of Environmental Research and Public Health 7:89-114.
Kabir SML, Sumon MH, Amin MM, Yamasaki S (2014). Isolation, Identification and Antimicrobial Resistance Patterns of Campylobacter Species from Broiler Meat Sold at KR Market of Bangladesh Agricultural University Campus, MymensinghJournal of Agriculture and Food Technology 4:15-21.
Khan MFR, Rahman MB, Sarker SK (2005). Seroprevalence of Mycoplasma gallisepticum infection in poultry in some selected farms of Mymensingh. Journal of Bangladesh Society for Agricultural Science and Technology 2:1-4.
Krieg NR, Holt JG, Sneath PHA, Staley JT, Williams ST (1994). Bergey’s Manual of Determinative Bacteriology, Williams & Wilkins, Baltimore, Md, USA, 9th edition.
Malmuthuge N, Li M, Chen Y, Fries P, Griebel PJ, Baurhoo B, Zhao X, Guan LL (2012). Distinct commensal bacteria associated with ingests and mucosal epithelium in the gastrointestinal tracts of calves and chickens. FEMS Microbiology Ecology 79:337-47.
Mead GC, Hudson WR and Hiton MH (1994). Use of a marker organism in poultry processing to identify sites of cross-contamination and evaluate possible measures. British Poultry Science 35:345-354.
Merchant IA, Packer RA (1967). Veterinary Bacteriology and Virology. Lowa State University Press, Ames. Lowa USA. 7:286- 306.
Mulder RWAW, Schlundt J (1999). Safety of poultry meat: from farm to table. In: International Consultative Group on Food Irradiation (ICGFI), FAO/IAEA/WHO.
Nachamkin I (2003). Campylobacter and Arcobacter, pp. 902-914. In: P. R. Murray, E. J. Baron, J. H. Jorgensen, M. A. Pfaller, and R. H. Yolken (ed.), Manual of Clinical Microbiology, ASM Press, Washington, D.C.
Ozaki H, Esaki H, Takemoto K, Ikeda A, Nakatani Y, Someya A, Hirayama N, Murase T (2011). Antimicrobial resistance in fecal E. coli isolated from growing chickens on commercial broiler farms. Journal of Veterinary Microbiology 150:13-29.
Rahman MM (2003). Growth of poultry industry in Bangladesh, poverty alleviation and employment opportunity. Proceeding of 3rd international poultry show and seminar, Bangladesh china friendship conference centre, Dhaka, Bangladesh, 28th February - 2
nd March
2003. Rawool DB, Malik SVS, Barbuddhe SB, Shakuntala I, Aurora R (2007).
A multiplex PCR for detection of virulence associated genes in Listeria monocytogenes. International Journal of Food Safety 9:56-62.
Rowe MT, Madden RH (2000). Campylobacter: Introduction, In: Robinson, R. K.; Batt, C. A. and Patel, P. D. (eds.), Encyclopedia of food microbiology. Academic Press, San Diego, CA. pp. 335-341.
Samosornsuk W, Asakura M, Yoshida E (2007). Evaluation of a cytolethal distending toxin (cdt) gene-based species-specific multiplex PCR assay for the identification of Campylobacter strains isolated from poultry in Thailand. Microbiology and Immunology 51:909-917.
Sarker SK, Rahman MB, Rahman M, Khan MFR, Akand MSI, Rahman MS (2009). Seroprevalence study, isolation and identification of Salmonella in chickens selected model breeder poultry farms of Bangladesh. International Journal of Bio Research 2:25-29.
Sarker YA, Hasan MM, Paul TK, Rashid SZ, Alam MN, Sikder MH (2018). Screening of antibiotic residues in chicken meat in Bangladesh by thin layer chromatography. Journal of Advanced Veterinary and Animal Research 5(2):140-145.
Savkovic SD, Villanueva J, Turner JR, Matkowskyj KA, Hecht G (2005). Mouse model ofbenteropathogenic E. coli infection. Infectious and Immunity 73:1161-1170.
Schippa S, Iebba V, Barbato M, Nardo GD, Totino V, Checchi MP, Longhi C, Maiella G, Cucchiara S, Conte MP (2010). A distinctive 'microbial signature' in celiac pediatric patients. BMC Microbiology 10:175.
Sharada R, Krishnappa G, Raghavan R, Sreevinas G, Upandra HA (1999). Isolation and serotyping of E. coli from different pathological conditions in poultry. Indian Journal of Poultry Science 34:366-369.
Shiramaru S, Asakura M, Inoue H, Nagita A, Matsuhisa A, Yamasaki S (2012). A cytolethal distending toxin gene-based multiplex PCR assay for detection of Campylobacter spp. in stool specimens and

770 Afr. J. Microbiol. Res.
comparison with culture method. The Journal of Veterinary Medical Science 74:857-862.
Sudershan RV, Naveen Kumar R, Kashinath L, Bhaskar V, Polasa K (2012). Microbiological hazard identification and exposure assessment of poultry products sold in various localities of Hyderabad, India. Scientific World Journal 12:736-40.
Thomas CGA (1998). Gram negative bacilli. Medical Microbiology 6:273-274.
Torok VA, Hughes RJ, Mikkelsen LL, Perez-Maldonado R, Balding K, MacAlpine R, Percy NJ, Ophel-Keller K (2011). Identification and characterization of potential performance-related gut microbiotas in broiler chickens across various feeding trials. Applied and Environmental Microbiology 77:5868-5878.
Voidarou C, Vassos D, Rozos G, Alexopoulos A, Plessas S, Tsinas A, Skoufou M, Stavropoulou E, Bezirtzoglou E (2011). Microbial challenges of poultry meat production. Anarobe 17:341-343.
Wouafo M, Nzouankeu A, Kinfack JA, Fonkoua MC, Ejenguele G, Njine
T, Ngandjio A (2010). Prevalence and antimicrobial resistance of Salmonella serotypes in chickens from retail markets in Yaounde (Cameroon). Microbial Drug Resistance 16:171-176.
Zhao C, Ge B, De Villena J, Sudler R, Yeh E, Zhao S, Meng J (2001). Prevalence of Campylobacter spp., E. coli, and Salmonella serovars in retail chicken, turkey, pork, and beef from the Greater Washington, DC, area. Applied and Environmental Microbiology 67:5431-5436.

Vol. 12(32), pp. 771-778, 28 August, 2018
DOI: 10.5897/AJMR2018.8946
Article Number: 863B6F758401
ISSN: 1996-0808
Copyright ©2018
Author(s) retain the copyright of this article
http://www.academicjournals.org/AJMR
African Journal of Microbiology Research
Full Length Research Paper
Screening of human diarrhoeal samples in Mymensingh city of Bangladesh for the isolation, identification and
antimicrobial resistance profiles of Campylobacter spp.
Sudarsan Karmaker1, S. M. Lutful Kabir1*, A. K. M. Ziaul Haque1, Mohammad Ferdousur Rahman Khan1 and Yousuf Ali Sarker2
1Department of Microbiology and Hygiene, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh.
2Department of Pharmacology, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh.
Received 23 July, 2018; Accepted 13 August, 2018
Campylobacter spp. (Campylobacter jejuni and Campylobacter coli) are one of the major cause of food-borne bacterial diarrhoea in human worldwide. This study was conducted for the isolation, identification and antimicrobial resistance profiling of Campylobacter spp. from diarrhoeal samples of human collected from Surya Kanta (SK) hospital, Mymensingh Medical College, Mymensingh during the period of August 2016 to October 2017. Using cultural and biochemical techniques, a total number of 150 samples were subjected to Campylobacter isolation and identification. The isolated Campylobacter species (C. jejuni and C. coli) were characterized by antimicrobial susceptibility testing. Among 40 positive Campylobacter isolates, 23 (57.50%) were C. jejuni and the rest 17 (42.50%) isolates were C. coli. Furthermore, out of 40 Campylobacter like organisms, 22 Campylobacter isolates were found in male patient and 18 Campylobacter isolates were found in female. 13 (16.04%) C. jejuni and 9 (11.11%) C. coli were found in male and 10 (14.49%) C. jejuni and 8 (11.59%) C. coli were found in female. Considering the different age groups, 5 (33.33%), 12 (27.91%), 19 (29.68%) and 4 (14.28%) Campylobacter isolates were found in 1 to 15, 16 to 30, 31 to 50 and above 50 years respectively during the period of August 2016 to October 2017. Majority of the Campylobacter jejuni were resistant to ampicillin, nalidixic acid, tetracycline and norfloxacin. However, majority of the Campylobacter jejuni were susceptible to gentamycin, chloramphenicol, ciprofloxacin, azithromycin and streptomycin. Furthermore, C. coli were resistant to ampicillin, tetracycline, erythromycin, nalidixic acid, norfloxacin and susceptible to streptomycin, chloramphenicol, gentamycin and ciprofloxacin. Out of 40 Campylobacter isolates 65.21% C. jejuni and 52.94% Campylobacter coli were detected as multidrug resistant. The findings of the study revealed the presence of multidrug resistant Campylobacter species in human diarrhoeal samples in Mymensingh. Key words: Human diarrhoeal samples, Campylobacter spp., isolation, identification, antimicrobial resistance profiles.
INTRODUCTION Campylobacter is one of the most commonly identified bacterial infections and is considered as the vital bacterial cause of acute human gastroenteritis worldwide (Allos
and Taylor, 1998). Campylobacteriosis is the major public health hazard and a common cause of gastro-enteritis of human in the industrialized world (Friedman

772 Afr. J. Microbiol. Res. et al., 2000). FoodNet surveillance identified 13.4 diagnosed Campylobacter infections per 100,000 persons (2002). Approximately 95% of diagnosed Campylobacter infections are due to C. jejuni (Altekruse et al., 1999). Campylobacter species are commonly inhibited in the intestinal tracts of poultry, livestock and food products of animal origin which are mostly associated with diarrhoea (de Boer et al., 2000). Campylobacter spp. is highly pathogenic causing acute diarrhoea and sometimes reactive arthritis and Guillain –Barre syndrome (Zia et al., 2003). Campylobacter spp. has a zoonotic impact (Anonymous, 2008) and causes food-borne infection resulting to human gastro-enteritis (Butzler et al., 1991; White et al., 1997; Kabir, 2011).
The primary sources of infection are considered to be the poultry and poultry products. Therefore, food animals such as cattle, sheep and pigs may be considered as asymptomatic carriers of Campylobacter species and during slaughter and carcass dressing, these animal food products can be contaminated by this pathogen (Berndtson et al., 1996). It is also reported that illness was associated with the consumption of raw milk and participation in hand milking of cows (Black et al., 1988).
Consumption of contaminated food stuffs are the major source of Campylobacteriosis in human which is predominantly acquired through it (Humphrey et al., 2007). About 2.5 million cases of Campylobacteriosis. The Centers for Disease Control and Prevention (CDC) estimates that over 200 deaths occur annually in the United States. Although, Campylobacter out-breaks are relatively rare, waterborne out-breaks have occurred worldwide in many developed countries (Karagiannis et al., 2010).
In campylobacterial diarrhoea, most of the patients had diarrhoea with vomiting and fever and the onset was mostly sudden which usually lasted for less than a week (Goossens et al., 1990). In human and veterinary practices, microbial resistance to antibiotic agents is a matter of concern all over the world (Sarker et al., 2018). Resistance of Campylobacter species to antimicrobial agents have been reported worldwide (Isenbarger et al., 2002). This condition seems to exaggerate rapidly in developing countries where widespread and uncontrolled use of antibiotics is practiced (Englen et al., 2003). Increasing level of antimicrobial resistant Campylobacter species is due to the excessive use of antimicrobial agents in food animals resulting to the emergence and dissemination of antimicrobial resistant bacteria (Hassan et al., 2014; Engberg et al., 2001), which has a serious impact on both Veterinary and Human health with regard to food safety. In veterinary industry antibiotics are commonly used as a therapeutic, prophylactic and growth promoting agents for livestock and poultry production and
its causes accumulation in antibiotic residue in animal food products and development of antibiotic resistance (Donoghue, 2003).
As a microaerophilic bacteria, Campylobacter can be detrimental to their survival within certain environmental stresses like exposure to air, drying, low pH and prolonged storage (Oh et al., 2015). Campylobacter is frequently isolated from stools of infants with diarrhoea in developing countries which is resulted from the consumption of contaminated food or water (Coker et al., 2002). Only few studies in Bangladesh have been reported for the isolation of Campylobacter species from patients having diarrhoea (Blaser et al., 1997; Alam et al., 2006). Furthermore, no documented reports exist yet on the prevalence and antimicrobial resistance of Campylobacter species in diarrhoeal patients in Mymensingh where patients are available in that case.
Therefore, the aim of the present study was to isolate, identify and analyze the antimicrobial resistance patterns of Campylobacter species from diarrhoeal patients in Surya Kanta (SK) hospital, Mymensingh Medical College, Mymensingh, Bangladesh. MATERIALS AND METHODS Collection and transportation of samples A total of 150 diarrhoeal samples were collected and immediately taken to the Bacteriology Laboratory of the Department of Microbiology and Hygiene, Bangladesh Agricultural University, Mymensingh through cool chain maintaining. Thereafter, the samples were processed immediately for the isolation and identification of Campylobacter spp. Isolation and identification of bacteria Isolation of Campylobacter spp. were carried out by filtration method (0.45 μm filter) as described by Shiramaru et al. (2012). Each of the stool samples was suspended in 500 μl of sterile saline. 100 μl of the samples was spread on the filters that were placed on the surface of Blood agar base no. 2 and allowed to stand for 30 min at room temperature. After 30 min, the filter was removed from the BBA and then the plates were incubated at 37°C for 48 h in microaerophilic condition (5% O2, 10% CO2 and 85% N2). After 48 h, the incubated media were then examined for growth of bacteria. Grey, flat and irregularly spreading colonies were observed on BBA. The colony was then subjected to Gram’s Method of staining and observed under microscope for Gram negative curve. The organisms from the agar media were then sub-cultured into Blood agar base no. 2 with the help of an inoculating loop in the case of Gram negative curve in the smears. In the case of Blood agar grey, flat and irregularly spreading colony were observed. Thus, single pure colony was obtained. These pure isolates obtained were used for further study.
For differentiation of isolation of Campylobacter, isolated organisms with supporting growth characteristics of Campylobacter
*Corresponding author. E-mail: [email protected], [email protected]. Tel +8801754987218.
Author(s) agree that this article remain permanently open access under the terms of the Creative Commons Attribution
License 4.0 International License

Karmaker et al. 773 Table 1. List of primer used for Campylobacter spp.
Primer Sequence (5’-3’) Target Amplicon size (bp)
Reference
16S9F
16S1540R
GAGTTTGATCCTGGCTC
AAGGAGGTGATCCAGCC Campylobacter spp.16S rRNA gene 1530 Samosornsuk et al.( 2007)
Table 2. Isolation of Campylobacter spp. according to sex and age of patients.
Parameter Number of
patients Number (%) of Campylobacter
positive case
Number (%) of Campylobacter spp.
C. jejuni (%) C. coli (%)
Sex
Male 81 22(27.16) 13 (16.04) 9 (11.11)
Female 69 18(26.09) 10 (14.49) 8 (11.59)
Total 150 40 (26.67) 23 (15.33) 17 (11.33)
Age (years)
1-15 15 5 (33.33) 3 (20.00) 2 (13.33)
16-30 43 12 (27.91) 7 (16.27) 5 (11.62)
31-50 64 19 (29.68) 11 (17.18) 8 (12.50)
>50 28 4 (14.28) 2 (7.14) 2 (7.14)
Total 150 40 (26.67) 23 (15.33) 17 (11.33)
were subjected to various tests according to the procedures previously described by Nachamkin (2003) and Foster et al. (2004).
Molecular identification by PCR
DNA templates were prepared with the boiling method as described by Hoshino et al. (1998). All the samples were examined by two pairs of primers (Table 1) to detect the 16S rRNA gene of Campylobacter spp.. The PCR reactions were carried out using a thermocycler (ASTEC, Japan) with the following programme: initial denaturation for 5 min at 94°C, followed by 30 cycles of denaturation at 94°C for 30 s, annealing at 47°C for 30 sand extension at 72°C for 1 min and 30 s. The final extension was conducted at 72°C for 10 min. The holding temperature was 4°C until the thermo cycler was removed. 1.5% agarose (Invitrogen, USA) gel was used for electrophoresis of the PCR products.
Antimicrobial sensitivity test
All Campylobacter strains were tested against ampicillin (10 μg), tetracycline (30 μg), chloramphenicol (30 μg), streptomycin (10 μg), gentamicin (10 μg), erythromycin (15 μg), azithromycin (15 μg), nalidixic acid (30 μg), ciprofloxacin (5 μg) and norfloxacin (10 μg) by disk diffusion method as described by Luangtongkum et al. (2007) with few modifications. All antimicrobial disks were obtained from Hi Media Laboratories Pvt. Ltd, India. Briefly, within 15 min after adjusting the turbidity of the inoculum suspension (equivalent to 0.5 McFarland turbidity), a sterile cotton swab was dipped into the adjusted suspension and then, the swab was rotated several times followed by pressed firmly on the inside wall of the tube above the fluid level to remove excess inoculum from the swab. Thereafter, the dried surface of a Muller-Hinton agar supplemented with 5% defibrinated sheep blood was inoculated by streaking the swab over the entire sterile agar surface and this procedure was repeated two more times, and the plate was rotated at 60° each time to ensure a confluent lawn of bacterial growth. After the inoculates were dry, five antimicrobial disks were applied per plate and incubated in the
inverted position at 37°C for 48 h under microaerophilic conditions (5% O2, 10% CO2 and 85% N2). The zone diameter breakpoints of each antimicrobial agent were determined according to the breakpoints used by the National Antimicrobial Resistance Monitoring System (NARMS) and the CLSI-established guideline for bacteria isolated from animals (CDC, 2003; National Committee for Clinical Laboratory Standards 2002a, 2002b).
RESULTS
Cultural examination and biochemical tests
Campylobacter spp. produced grey color spreading colonies on blood agar base no. 2 media after 48 h of incubation at 37°C. In Gram’s staining under microscope, the organism revealed Gram negative, pink color, small curved shape arranged as single or pair. The organisms were checked and confirmed by their purity using special blood agar media (Blood agar base no. 2). All the isolates of Campylobacter spp. were found positive for catalase test and oxidase test. Only C. jejuni were found positive in hippurate hydrolysis test but C. coli were found negative for hippurate hydrolysis test.
Isolation of Campylobacter spp. from patients according to sex and age
A total of 150 diarrhoeal samples (male=81 and female=69) were subjected to isolation of Campylobacter strains by filtration method. A total of 40 (26.67%) Campylobacter organisms were found in the studied samples where 22 (27.16%) were male and 18 (26.09%) were female as shown in Table 2.

774 Afr. J. Microbiol. Res.
Figure 1. PCR product of 16S rRNA gene. A 1530 bp gene product in the picture confirmed the 16S rRNA gene of Campylobacter spp. Lane, 1 was 100 bp DNA ladder (Promega, USA); lane 2 was negative control and lanes 3 to 7 were positive samples.
The age groups of experimented samples were divided as; 1 to 15 years (n=15), 16 to 30 years (n=43), 31 to 50 years (n=64) and above 50 years (n=28) and were subjected to isolation of Campylobacter strains. Out of 40 Campylobacter organisms; 5 (33.33%) were in 1 to 15 years, 12 (27.91%) were in 16 to 30 years, 19 (29.68%) were in 31 to 50 years and above 50 years, 4 (14.28%) were found (Table 2).
Molecular detection by PCR
Genus specific (16S rRNA gene) polymerase chain reaction (PCR) was performed. 1530 bp fragment of targeted gene was amplified successfully. The results of PCR are shown in Figure 1.
Antibiogram study for Campylobacter spp.
Antimicrobial susceptibility of C. jejuni and C. coli
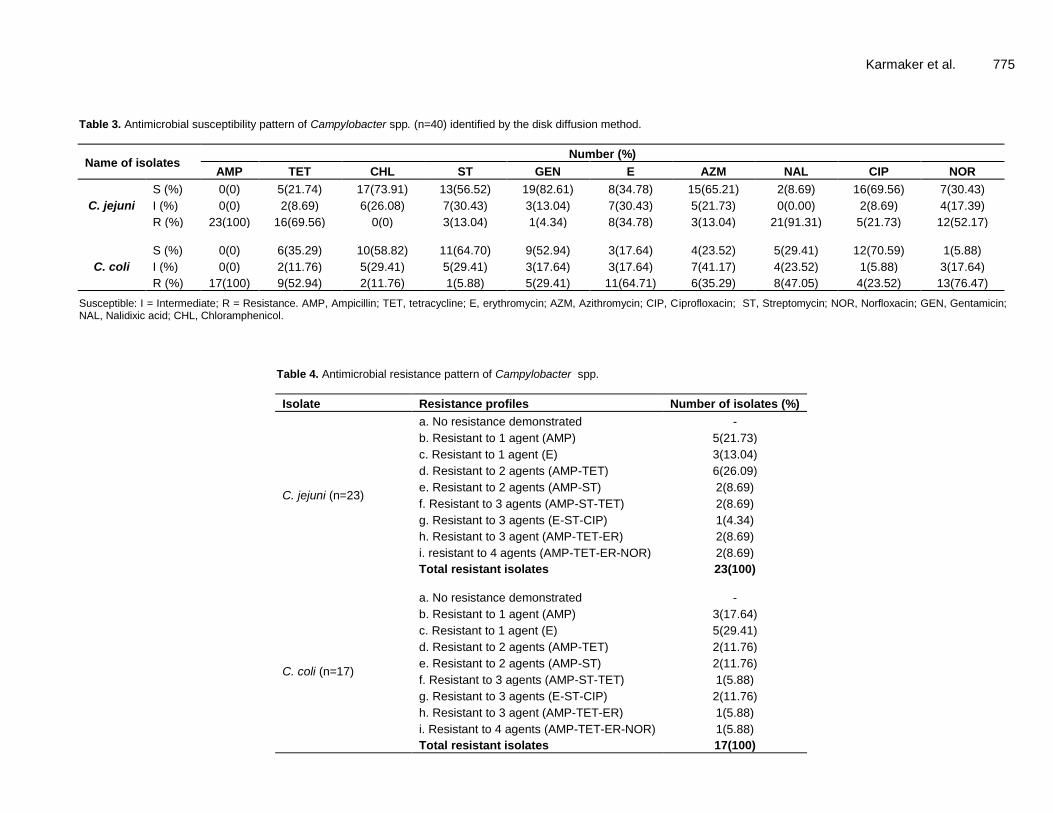
Details results of antimicrobial susceptibility testing by disc diffusion method with 10 chosen antimicrobial agents are presented in Table 3. Out of 23 C. jejuni isolates, 23 (100%) were resistant to ampicillin, 7 (30.43%) were intermediately resistant to streptomycin, and 19 (82.61%) were susceptible to gentamicin.
Out of 17 Campylobacter coli isolates, 17 (100%) were resistant to ampicillin, 7 (41.17%) were intermediately resistant to azithromycin and 12 (70.59%) were susceptible to ciprofloxacin
Antimicrobial resistant pattern of C. jejuni and C. coli
The results of antimicrobial resistance patterns of C.
jejuni and C. coli are summarized in Table 4. Out of 23 C. jejuni isolates, 5 (21.73%) and 3 (13.04%) were resistant to each of 1 antibiotic (AMP) and (E) respectively. Moreover, 6 (26.09%) and 2 (8.69%) were resistant to each of 2 antibiotic agents (AMP-TET) and (AMP-ST) respectively. Furthermore, 2 (8.69%), 1(4.34) and 2 (8.69%) were resistant to each of 3 antibiotic agents: (AMP-ST-TET), (E-ST-CIP) and (AMP-TET-ER) respectively and 2 (8.69%) were resistant to 4 antibiotic agents (AMP-TET-ER-NOR).
Out of 17 C. coli isolates, 3 (17.64%) and 5 (29.41%) were resistant to 1 antibiotic (AMP) and (E) respectively. Furthermore, 2 (11.76%) and 2 (11.76%) were resistant to 2 antibiotics (AMP-TET) and (AMP-ST) respectively. 1 (5.88%), 2 (11.76%) and 1 (5.88%) were resistant to each of 3 antibiotic agents: (AMP-ST-TET), (E-ST-CIP) and (AMP-TET-ER) respectively. 1 (5.88%) were resistant to 4 antibiotic agents (AMP-TET-ER-NOR).
Multidrug resistant Campylobacter spp. was identified by considering resistant to 2 or more drugs. A total of 15 (65.21%) C. jejuni (out of 23) and 9 (52.94%) C. coli (out of 17) were identified as multidrug resistant. DISCUSSION For the characterization of the Campylobacter spp. isolation, cultural characteristics based examination, staining characteristics, biochemical testing and finally PCR were very crucial which were performed from the isolated Campylobacter spp. Blood agar base no. 2 as a selective agar media was used to culture the organism (Campylobacter spp.) under required microaerophilic environment (5% O2, 10 % CO2 and 85% N2) as the experiment was conducted by several researchers (Haseena, 2017; Kabir et al., 2014a, 2014b) and
1500 bp 1530 bp
7 6 5 4 1 2 3
Karmaker et al. 775
Table 3. Antimicrobial susceptibility pattern of Campylobacter spp. (n=40) identified by the disk diffusion method.
Name of isolates Number (%)
AMP TET CHL ST GEN E AZM NAL CIP NOR
C. jejuni
S (%) 0(0) 5(21.74) 17(73.91) 13(56.52) 19(82.61) 8(34.78) 15(65.21) 2(8.69) 16(69.56) 7(30.43)
I (%) 0(0) 2(8.69) 6(26.08) 7(30.43) 3(13.04) 7(30.43) 5(21.73) 0(0.00) 2(8.69) 4(17.39)
R (%) 23(100) 16(69.56) 0(0) 3(13.04) 1(4.34) 8(34.78) 3(13.04) 21(91.31) 5(21.73) 12(52.17)
C. coli
S (%) 0(0) 6(35.29) 10(58.82) 11(64.70) 9(52.94) 3(17.64) 4(23.52) 5(29.41) 12(70.59) 1(5.88)
I (%) 0(0) 2(11.76) 5(29.41) 5(29.41) 3(17.64) 3(17.64) 7(41.17) 4(23.52) 1(5.88) 3(17.64)
R (%) 17(100) 9(52.94) 2(11.76) 1(5.88) 5(29.41) 11(64.71) 6(35.29) 8(47.05) 4(23.52) 13(76.47)
Susceptible: I = Intermediate; R = Resistance. AMP, Ampicillin; TET, tetracycline; E, erythromycin; AZM, Azithromycin; CIP, Ciprofloxacin; ST, Streptomycin; NOR, Norfloxacin; GEN, Gentamicin; NAL, Nalidixic acid; CHL, Chloramphenicol.
Table 4. Antimicrobial resistance pattern of Campylobacter spp.
Isolate Resistance profiles Number of isolates (%)
C. jejuni (n=23)
a. No resistance demonstrated -
b. Resistant to 1 agent (AMP) 5(21.73)
c. Resistant to 1 agent (E) 3(13.04)
d. Resistant to 2 agents (AMP-TET) 6(26.09)
e. Resistant to 2 agents (AMP-ST) 2(8.69)
f. Resistant to 3 agents (AMP-ST-TET) 2(8.69)
g. Resistant to 3 agents (E-ST-CIP) 1(4.34)
h. Resistant to 3 agent (AMP-TET-ER) 2(8.69)
i. resistant to 4 agents (AMP-TET-ER-NOR) 2(8.69)
Total resistant isolates 23(100)
C. coli (n=17)
a. No resistance demonstrated -
b. Resistant to 1 agent (AMP) 3(17.64)
c. Resistant to 1 agent (E) 5(29.41)
d. Resistant to 2 agents (AMP-TET) 2(11.76)
e. Resistant to 2 agents (AMP-ST) 2(11.76)
f. Resistant to 3 agents (AMP-ST-TET) 1(5.88)
g. Resistant to 3 agents (E-ST-CIP) 2(11.76)
h. Resistant to 3 agent (AMP-TET-ER) 1(5.88)
i. Resistant to 4 agents (AMP-TET-ER-NOR) 1(5.88)
Total resistant isolates 17(100)

776 Afr. J. Microbiol. Res. filtration method (0.45 μm filter paper) was used for selection of the Campylobacter spp. (Shiramaru et al., 2012). The colony characteristics of Campylobacter spp. exhibited pink or light pink color, gram negative and slightly curved shaped that is similar to the results of several researchers (Doyle, 1990; Rowe and Madden, 2000; Jamshidi et al., 2008; Kabir, 2011).
This study recorded 40 Campylobacter spp. by biochemical tests. Out of 40 isolates, 23 (57.50%) C. jejuni and 17 (42.50%) C. coli were identified. In this study, for the identification of Campylobacter spp. biochemical tests were performed which were also similar to the studies of a number of researchers (Shiramaru et al., 2012; Gblossi et al., 2012; Kabir et al., 2014a, 2014b).
PCR primers targeting 16S rRNA gene of Campylobacter spp. were amplified. 1530 bp fragments of DNA confirmed the identity of Campylobacter spp. Similar results were also observed by several researchers (Foster et al., 2004; Samosornsuk et al., 2007; Kabir, 2011). Campylobacter spp. were found in the collected samples of diarrhoeal patients in SK Hospital, Mymensingh Medical College, Mymensingh. Similar findings were reported in other studies by Awad et al. (2010); Malmuthuge et al. (2012) and Hossain et al. (2015).
Reliable and reproducible laboratory techniques are required for monitoring drug resistance among the Campylobacter isolates. Campylobacter present difficulties in antimicrobial susceptibility testing like other fastidious bacteria because of their unique growth requirements and test conditions. In this study, susceptibility profiles of Campylobacter isolates were determined through agar disk diffusion method. The methodology has proven to be a relatively accurate method to test antimicrobial susceptibilities of fastidious organisms including Campylobacter spp.
The current study was conducted to detect the antimicrobial resistance patterns by disk diffusion methods of Campylobacter spp. that is used by several researchers (Kabir, 2011; Gblossi et al., 2012; Wieczorek et al., 2013; Kabir et al., 2014a, 2014b). The high level of ciprofloxacin resistance (>80%) and tetracycline had been found by Senok et al. (2007) in human patients that suffered from diarrhoea. Hakanen et al. (2003) found 46% resistance to ciprofloxacin, tetracycline (46%) and ampicillin (17%) respectively from the diarrhoeal patients.
Higher frequency of erythromycin resistance in C. coli than in C. jejuni (0 to 11% in C. jejuni and 0 to 68.4% in C. coli) had been reported by Sjögren et al. (1997) for human that suffered from Campylobacterial diarrhoea. Resistance to erythromycin was low in Japan, Canada, and Finland but recent development of resistance in Thailand and Sweden has been reported by Hoge et al. (1998). The concomitant resistance rates among nalidixic acid resistant C. jejuni isolated from patients (exclusively children) were 2, 12, 12, 97 and 66% for gentamicin, erythromycin, clindamycin, tetracycline, ciprofloxacin
respectively and 90% of the C. coli isolates were concomitantly resistant to clindamycin. The rates of resistance of 51 to 72 human strains of C. jejuni isolated annually from 1998 to 2001 in Montréal, Québec, Canada, varied from 1 to 12% for erythromycin, 43 to 68% for tetracycline and 10 to 47% for ciprofloxacin (Gaudreau, 2003).
For Campylobacter infections, Hoge et al. (1998) found 100% co-resistance between Campylobacter spp. isolates resistant to azithromycin and ciprofloxacin. Fluoroquinolone resistance in C. jejuni, C. coli or combination of C. jejuni and C. coli were 56.9%, 84%, and 75 to 88%, respectively which is supported by the findings of the present study.
This study recorded highest resistance rates to ampicillin, erythromycin, nalidixic acid and norfloxacin against C. jejuni and C. coli. Findings of this study suggest that multidrug resistant Campylobacter spp. isolated from human diarrhoeal samples might be an important concern for human health. This study has generated the first report on the antimicrobial resistance pathogens of Campylobacter spp. isolated from human diarrhoeal samples in Bangladesh. Conclusion
The research project revealed that Campylobacter spp. infection is a common phenomenon in the study area. The findings of the study also revealed the presence of multidrug resistant Campylobacter species in human diarrhoeal samples in Mymensingh. Strictly hygienic measures with personal hygiene, food safety especially during consumption of food stuffs contaminated with Campylobacter spp. organism should strictly be prohibited. Antibiotics could be used after sensitivity test for the treatment of clinical cases of Campylobacter spp. infection in human. It is also necessary to avoid excessive use of antibiotics in food animals due to their antimicrobial resistance properties.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The authors like to sincerely thank the University Grants Commission (UGC) of Bangladesh (Project No. 2015/274/UGC) for giving financial and logistic support for this study. REFERENCES
Alam J, Lastovica AJ, Roux EI, Hossain M.A, Islam MN, Sen SK, Sur GC, Nair GB, Sack DA (2006). Clinical characteristics and serotype

distribution of Campylobacter jejuni and Campylobacter coli isolated from diarrheic patients in Dhaka, Bangladesh and Cape Town, South Africa. Bangladesh Journal of Microbiology 23:121-124.
Allos BM, Taylor DN (1998). Campylobacter infections.In Bacterial Infections of Humans. pp. 169-190 Springer US.
Altekruse SF, Stern NJ, Fields PI, Swerdlow DL (1999).Campylobacter jejuni-an emerging foodborne pathogen. Emerging infectious diseases 5:28.
Anonymous (2008). Annual New Zealand notifiable disease report 2008.
Awad-Alla ME, Abdien HM, Dessouki AA (2010). Prevalence of bacteria and parasites in White Ibis in Egypt. Veterinaria Italians 46:277-86.
Berndtson E, Emanuelson U, Engvall A, Danielsson-Tham ML (1996). A 1year epidemiological study of Campylobacters in 18 Swedish chicken farms. Preventive Veterinary Medicine 26:167-185.
Black RE, Levine MM, Clements ML, Hughes TP, Blaser MJ (1988). Experimental Campylobacter jejuni infection in humans. Journal of Infectious Diseases 157:472-479.
Blaser MJ (1997). Epidemiologic and clinical features of Campylobacter jejuni infections. Journal of Infectious Diseases 176 (Supplement_2):S103-S105.
Butzler JP, Oosterom J (1991). Campylobacter–pathogenicity and significance in foods. International Journal of Food Microbiology 12:1-8.
Centers for Disease Control and Prevention (CDC) (2003). Preliminary FoodNet data on the incidence of foodborne illnesses--selected sites, United States, (2002). MMWR. Morbidity and mortality weekly report52:340.
Coker AO, Isokpehi RD, Thomas BN, Amisu KO, Obi CL (2002). Human Campylobacteriosis in developing countries. Emerging Infectious Diseases 8:237.
de Boer P, Duim B, Rigter A, van der Plas J, Jacobs-Reitsma WF, Wagenaar JA (2000). Computer-Assisted Analysis and Epidemiological Value of Genotyping Methods for Campylobacter jejuni and Campylobacter coli. Journal of Clinical Microbiology 38:1940-1946.
Donoghue Dan J (2003). Antibiotics residue in poultry tissues and eggs, Human Health Concerns. Poultry Science 82:618-621.
Doyle MP (1990). Campylobacter jejuni, In: D. O. Cliver (ed.), Foodborne diseases. Academic Press, Inc., Boston, MA pp. 217-222
Engberg J, Aarestrup FM, Taylor DE, Gerner-Smidt P, Nachamkin I (2001). Quinolone and macrolide resistance in Campylobacter jejuni and C. coli: resistance mechanisms and trends in human isolates. Emerging Infectious Diseases 7:24.
Englen MD, Ladely SR, Fedorka-Cray PJ (2003). Isolation of Campylobacter and identification by PCR. Methods in Molecular Biology 216:109-121.
Foster G, Holmes B, Steigerwalt AG, Lawson PA, Thorne P, Byrer DE, Ross HM, Xerry J, Thompson PM, Collins MD (2004). Campylobacter insulaenigrae sp. nov., isolated from marine mammals. International Journal of Systematic and Evolutionary Microbiology 54:2369-2373.
Friedman CJ, Neiman J, Wegener HC, Tauxe RV (2000). Epidemiology of Campylobacter jejuniinfection in the United States and other industrialized nations Campylobacter. American society for Microbiology AMS Press pp. 121-138.
Gaudreau C, Gilbert H (2003). Antimicrobial resistance of Campylobacter jejuni subsp. jejuni strains isolated from humans in 1998 to 2001 in Montreal, Canada. Antimicrobial Agents and Chemotherapy 47:2027-2029.
Gblossi GB, Eric A, Solange E, Natalie G, Souleymane B, LamineSébastien N, Mireille D (2012). Prevalence and antimicrobial resistance of thermophilicCampylobacter isolated from chicken in Côte d’Ivoire. International Journal of Microbiology Article ID 150612, 5 pages.
Goossens H, Vlaes L, De Boeck M, Pot B, Kersters K, Levy J, Vandamme P (1990). Is" Campylobacter upsaliensis" an unrecognised cause of human diarrhoea? Lancet (London, England) 335:584-586. Hakanen AJ, Lehtopolku M, Siitonen A, Huovinen P, Kotilainen P (2003). Multidrug resistance in Campylobacter jejuni strains collected from Finnish patients during 1995–2000. Journal of Antimicrobial Chemotherapy 52:1035-1039.
Karmaker et al. 777 Haseena M, Malik MF, Javed A, Arshad S, Asif N, Zulfiqar S, Hanif J
(2017). Water pollution and human health. Environmental Risk Assessment and Remediation 1:16-19
Hassan MM, Amin KB, Ahaduzzaman M Alam M, Faruk MSA, Uddin I (2014). Antimicrobial resis-tance pattern against E. coli and Salmonella in layer poultry. Research Journal for Veterinary Practitioners 2:30-35.
Hoge CW, Gambel JM, Srijan A, Pitarangsi C, Echeverria P (1998). Trends in antibiotic resistance among diarrheal pathogens isolated in Thailand over 15 years. Clinical Infectious Diseases 26:341-345.
Hoshino KS, Yamasaki AK, Mukhopadhyay S, Chakraborti A, Basu SK, Bhattacharya, GB Nair, T Shimada, Takeda Y (1998). Development of evaluation of multiplex PCR assay for rapid detection of Toxicogenic Vibrio cholera O1 and O139. FEMS Immunology and Medical Microbiology 20:201-207.
Hossain M, Hoda N, Hossen MJ, Hasan MM, Rahman SME, Kabir SML (2015) Assessment of bacterial load of poultry meat used at dining hall of Bangladesh Agricultural University campus. Asian Journal of Medical and Biological Research 1:9-16.
Humphrey TJ, Henley A, Lanning DG (2007). The colonization ofbroiler chickens with Campylobacter jejuni: some epidemiological investigations. Epidemiology and Infection 110:601-607.
Isenbarger DW, Hoge CW, Srijan A, Pitarangsi C, Vithayasai N, Bodhidatta L, Hickey KW, Cam PD (2002). Comparative antibiotic resistance of diarrheal pathogens from Vietnam and Thailand, 1996-1999. Emerging Infectious Diseases 8:175-180.
Jamshidi A, Bassami MR, Farkhondeh T (2008). Isolation and identification of Campylobacter spp. and Campylobacter coli from poultry carcasses by conventional culture method and multiplex PCR in Mashhad, Iran. Iranian Journal of Veterinary Research 9:132-137.
Kabir SML (2011). Comparison of molecular methods for the species identification of clinical Campylobacter strains and their antimicrobial resistance. PhD thesis. Osaka Prefecture University, Japan pp. 1-158.
Kabir SML, Islam J, Suman MH, Khan MSR, Yamasaki S (2014a). Isolation, identification and antimicrobial susceptibility profiles of Campylobacter spp. with assessment of the risk factors in broiler flocks of Bangadesh Agricultural University Poultry Farm. Journal of Basic and Applied Science Research 4:160-168.
Kabir SML, Suman MH, Amin MM, Yamasaki S (2014b). Isolation, identification and antimicrobial resistance patterns of Campylobacter spp. from Broiler Meat Sold at KR Market of Bangladesh Agricultural University Campus, Mymensingh.Journal of Agriculture and Food Technology 4:15-21.
Karagiannis I, Sideroglou T, Gkolfinopoulou K, Tsouri A, Lampousaki D, Velonakis EN, Scoulica EV, Mellou K, Panagiotopoulos T, Bonovas S (2010). A waterborne Campylobacter jejuni outbreak on a Greek island. Epidemiology and Infection 138:1726-1734.
Luangtongkum T, Morishita El-Tayeb AB, Ison AJ, Zhang Q (2007). Comparison of antimicrobial susceptibility testing of Campylobacter spp. by the agar dilution and the agar disk diffusion methods. Journal of Clinical Microbiology 45:590-594.
Malmuthuge N, Li M, Chen Y, Fries P, Griebel PJ, Baurhoo B, Zhao X, Guan L L (2012). District commercial Bacteria associated with ingests and mucsal epithelium in the gastrointestinal tracts of calves and chickens. FEMS Microbiology Ecology 79:337-347.
Nachamkin I (2003). Campylobacter and Arcobacter, In Murray PR, Baron EJ, Jorgensen JH, Pfaller MA, Yolken RH (ed.), Manual of Clinical Microbiology, ASM Press, Washington, D.C. pp. 902-914.
National Committee for Clinical Laboratory Standards (2002a). Performance standards for antimicrobial disk and dilution susceptibility tests for bacteria isolated from animals. Approved standard M31-A2, 2nd ed. National Committee for Clinical Laboratory Standards, Wayne, PA.
National Committee for Clinical Laboratory Standards (2002b). Development of in vitro susceptibility testing criteria and quality control parameters for veterinary antimicrobial agents; approved guideline, 2nd ed. M37-A2. National Committee for Clinical Laboratory Standards, Wayne, PA.
Oh E, McMullen L, Jeon B (2015). Impact of oxidative stress defense on bacterial survival and morphological change in Campylobacter jejuni under aerobic conditions. Frontiers in Microbiology 6:295.

778 Afr. J. Microbiol. Res. Rowe MT, Madden RH (2000). Campylobacter: Introduction, In
Robinson RK, Batt CA, Patel PD (eds.), Encyclopedia of food microbiology. Academic Press, San Diego, CA. pp. 335- 341.
Samosornsuk W, Asakura M, Yoshida E, Taguchi T, Nishimura K, Eampokalap B, Yamasaki S (2007). Evaluation of a cytolethal
distending toxin (cdt) Gene‐Based Species‐Specific Multiplex PCR assay for the identification of Campylobacter strains isolated from poultry in Thailand. Microbiology and Immunology 51:909-917.
Sarker YA, Hasan MM, Paul TK, Rashid SZ, Alam MN, Sikder MH (2018). Screening of antibiotic residues in chicken meat in Bangladesh by thin layer chromatography. Journal of Advanced Veterinary and Animal Research 5(2):140-145.
Senok A, Yousif A, Mazi W, Sharaf E, Bindayna K, Elnima E, Botta G (2007). Pattern of antibiotic susceptibility in Campylobacter jejuni isolates of human and poultry origin. Japanese Journal of Infectious Diseases 60:1-4.
Shiramaru S, Asakura M, Inoue H, Nagita A, Matsuhisa A, Yamasaki S (2012). A cytolethal distending toxin gene-based multiplex PCR assay for detection of Campylobacter spp. in stool specimens and comparison with culture method. Journal of Veterinary Medical Science 74:857-862.
Sjögren E, Lindblom GB, Kaijser B (1997). Norfloxacin resistance in
Campylobacter jejuni and Campylobacter coli isolates from Swedish patients. The Journal of Antimicrobial Chemotherapy 40:257-261.
White PL, Baker AR, James WO (1997). Strategies to control Salmonella and Campylobacter in raw poultry products. In contamination of animal products: prevention and risks for public health. Revescientifiqueet technique (International Office of Epizootics) 16: 525-541.
Wieczorek K, Osek J (2013). Antimicrobial resistance mechanisms among Campylobacter. BioMed Research International 2013:340605.
Zia S, Wareing D, Sutton C, Bolton E, Mitchell D, Goodacre JA (2003). Health problems following Campylobacter jejuni enteritis in a Lancashire population. Rheumatology 42:1083-1088.

Vol. 12(32), pp. 779-787, 28 August, 2018
DOI: 10.5897/AJMR2018.8875
Article Number: 87F397558403
ISSN: 1996-0808
Copyright ©2018
Author(s) retain the copyright of this article
http://www.academicjournals.org/AJMR
African Journal of Microbiology Research
Full Length Research Paper
Assessment of dominant bacterial strains isolated from Ntoba mbodi, an indigenous African alkaline-fermented
food, and their potential enzyme activities
Didine Priscilla Moutou-Tchitoula1, Etienne Nguimbi1, Stéphanie Giusti-Miller2, Philippe Mora2, Simon Charles Kobawila1, Edouard Miambi2*
1Laboratoire de Biologie Cellulaire et Moléculaire, Faculté des Sciences et Techniques, Université Marien NGOUABI,
BP 69 – Brazzaville, République du Congo. 2Faculté des Sciences – IEES Paris, Université Paris Est Créteil; 61 Avenue du Général de Gaulle, 94010 Créteil cedex,
France.
Received 26 April, 2018; Accepted 19 July, 2018
Ntoba mbodi is a stable and long-life alkaline-fermented food and the domestic-scale production depends upon microorganisms from the local environment. Previous studies on bacteria in Ntoba mbodi have reported the presence of Bacillus related species and other bacterial taxa. But the abundance of these bacteria in Ntoba mbodi still needs to be determined to assess their ecological importance in this particular environment. In the present study, we used an extinction dilution approach for isolation and taxonomic affiliation of the dominant isolates obtained from the highest positive dilution of Ntoba mbodi. We also tested the hypothesis that there is a convergence in the bacterial community profile in Ntoba mbodi, irrespective of the origins of the food, by assessing the set of dominant bacteria that was found in all the samples studied. The density of the bacteria ranged from 1.67×10
10 to 2.8×10
10 CFU g
-1. Eleven distinct morphotypes were isolated at the highest positive dilution
of Ntoba mbodi and identified using 16S rRNA gene sequencing followed by phylogenetic analyses. Eight isolates were classified as Bacillus related species. A set of five strains was shared between all samples of Ntoba mbodi which were closest to Bacillus altitudinis, Bacillus sp. C-32, Staphylococcus sciuri and Bacterium NLAE-ʫ-H375. Of the eleven predominant isolates, eight showed cellulase activity and five showed xylanase and β-glucosidase activity. Key words: Bacillus, cassava leaves, alkaline-fermentation, Ntoba mbodi, indigenous fermented foods.
INTRODUCTION Fermentation is one of the most ancient forms of food preservation technologies in the world. It uses microorganisms to convert perishable and sometimes inedible raw materials into safe, shelf-stable and
palatable foods or beverages (Ray and Montet, 2017). Indigenous food fermentation represents an extremely
valuable cultural heritage in most regions, and harbors a
huge genetic potential of valuable, but hitherto unidentified,
*Corresponding author. E-mail: [email protected].
Author(s) agree that this article remain permanently open access under the terms of the Creative Commons Attribution
License 4.0 International License

780 Afr. J. Microbiol. Res.
microbial strains (Tamang et al., 2016). Steinkraus (1996) classified fermentation processes
into various categories including lactic acid, alcoholic, acetic acid and alkaline fermentations. Unlike lactic acid and alcoholic fermentations, alkaline fermentation is unusual in that it causes an increase in the pH of the food as a result of the release of ammonia from the breakdown of proteins in the food (Sarkar and Nout, 2014; Ray and Montet, 2017). Alkaline fermented foods are a group of lesser-known food products that are widely consumed almost exclusively in Asian and Africa countries (Streinkraus, 2002); Sarkar and Nout, 2014).
Ntoba mbodi is a fermented food from a household-scale alkaline-fermentation process which depends upon the microorganisms in the local environment (Louembe et al., 2003; Kobawila et al., 2005; Mokemiabeka et al., 2011). A pioneering study of the microflora of Ntoba mbodi, using phenotypic characterization of isolates, reported the presence of Micrococcus varians, Bacillus macerans, Bacillus subtilis, Staphylococcus sciuri and Staphylococcus xylosus (Louembe et al., 2003). Kobawila et al. (2005) using the same bacterial characterization approach reported on the presence of Bacillus subtilis, Bacillus amyloliquefaciens, Bacillus megaterium, Bacillus macerans, Bacillus cereus, Bacillus polymixa, Bacillus brevis and Bacillus pumilus. Recently, Vouidibio-Mbozo et al. (2017), by combining both phenotypic and genotypic methods identified B. safensis, B. siamensis and Lysinibacillus spp. in Ntoba mbodi. A novel species of the genus Lysinibacillus has even been isolated from Ntoba mbodi (Ouoba et al., 2015). Surprisingly, lactic acid bacteria (LAB), which are the microorganisms responsible for acidic indigenous food fermentations (Brauman et al., 1996; Lei and Jakobsen, 2004; Kostinek et al., 2007; Sawadogo-Lingani et al., 2007), were also found in Ntoba mbodi (Ouoba et al., 2010). The LAB isolates were closest to Weissella confusa, Weissella cibaria, Lactobacillus plantarum, Pediococcus pentosaceus, Enterococcus casseliflavus, Enterococcus faecium, Enterococcus faecalis, Enterococcus avium and Enterococcus hirae (Ouoba et al., 2010).
However, it is still unclear whether the isolates previously described were the predominant bacterial taxa in Ntoba mbodi. Indeed, isolation of colonies was based on phenotypic characteristics of Bacillus after cultivation of total mesophilic bacteria (Vouidibio-Mbozo et al., 2017) or LAB (Ouoba et al., 2010). Therefore the bacterial predominance in previous studies represents the fraction of each distinct species complete set of isolates or probably faster growing bacteria but does not reflect the abundance of each cultivable species in the initial samples of Ntoba mbodi. This raises the question of the ecological relevance of isolates obtained under these conditions. Are these bacteria species involved in the fermentation of Ntoba mbodi or simply transient or inactive forms? Knowing the abundance of different species can provide insight into how a community
functions (Verberk, 2011). Likewise, understanding the bacterial diversity and determining the taxonomic classification of dominant species are necessary for determining the bacteria forming the “functional starter” (Leroy and De Vuyst, 2004).
Furthermore, the softening of the cassava leaves which is the main indicator for the end of the fermentation process in Ntoba mbodi has been ascribed to bacterial enzyme activity in Ntoba mbodi (Mokemiabeka et al., 2011). However, this conclusion was based on the results obtained by screening for enzyme activities in homogenates of fermented cassava leaves. This experimental approach could not establish whether the enzyme activities observed came from bacterial activity or from enzymes secreted by cassava leaves themselves. In addition, xylanase activity was not investigated while xylan is the second most abundant polysaccharide found in the plant kingdom after cellulose and is the major hemicellulose in plant cell walls (He et al., 2014).
The present study set out to assess the numerically dominant bacterial species in Ntoba mbodi using a dilution to extinction approach (Garland and Lehman, 1999; Gomez et al., 2004), focusing on colonies obtained from the highest positive dilution of the Ntoba mbodi. We also investigated the hypothesis that there was a convergence of bacterial community profiles in Ntoba mbodi irrespective of the origin of samples in Brazzaville (Congo) as has been demonstrated in other ecosystems (Jia et al., 2016). This hypothesis was tested by assessing the set of bacteria shared across samples of Ntoba mbodi purchased from different producers. Finally, we determined whether the numerically dominant bacteria taxa isolated had the enzyme activities necessary for breaking down the main polysaccharides that make up the plant material.
MATERIALS AND METHODS
Sample collection, culture and bacterial isolation
Samples of Ntoba mbodi were purchased from various markets in Brazzaville (Congo). They were produced in four different workshops. These samples were aseptically collected in containers and transported to the laboratory within 2 h using cooler boxes. The samples were immediately processed in the laboratory under sterile conditions for subsequent analyses. Samples from the same workshop were pooled to make a composite sample. Then a 10 g sub-sample was put into a sterile mortar with 90 mL of sterile 0.9% NaCl solution and homogenized using a pestle. A 1.5 mL-aliquot was centrifuged at 800 x g for 1 min to remove suspended solids. The supernatant was decimally diluted in 0.9% NaCl solution down to 10-8 for use as inocula. The pH was adjusted to 6.8 ± 0.2 and a 30 µL-aliquot of each dilution was plated onto sterile plate count agar (Merck, Germany). The plates were incubated at 37°C for 24 h. The colonies formed on the agar plates were then used to estimate the numbers of bacteria in the original sample. Colonies developing on plates from the last positive dilution were first grouped by their macroscopic and microscopic morphotypes. Then colonies representative of the most abundant morphotypes were

isolated by streaking three times. The isolates were kept on plate count agar (Merck, Germany) for few days at 4°C before use. Identification of isolates using 16S rRNA gene sequencing and phylogenetic analyses The bacteria were identified using 16S rRNA gene sequencing followed by phylogenetic affiliation analyses. Each isolate was grown in liquid nutrient broth (Merck, Germany) at 37°C for 24 h then a 1.5 mL subsample of the bacterial culture was transferred to a sterile Eppendorf tube and centrifuged at 10,000 g for 10 min. DNA was extracted from the pellets using Mobio kit PowerSoil, following the manufacturer’s instructions. There were three replicates for each sample. The concentration of the extracted DNA was evaluated by UV absorption at 260 nm using a NanoDrop 2000C (Thermo Scientific).The 16S rRNA gene was amplified by Polymerase Chain Reaction (PCR) as described by Miambi et al. (2003) with some modifications. PCR reaction was performed in a 25 µL reaction mixture (total volume) using Taq polymerase Ready-To-Go (Amersham Pharmacia Biotech, USA), 0.25 mM of primer 27F (5’-AGAGTTTGATCCTGGCTCAG-3’), 0.25 mM of primer 1390 R (5’-GAC GGG CGG TGT GTA CAA-3’) and 20 ng of the template DNA. The PCR was carried out using a thermocycler (VWR, France) with an initial denaturation for 8 min at 94°C; 30 cycles of 30 s at 94°C, 1 min at 56°C and 30 s at 72°C, and a final extension for 15 min at 72°C. The PCR products were run on 1.5 % (wt/vol agarose-0.5X TAE) buffer gel stained with SYBR Safe stain (Invitrogen, France) at 80V for 30 min and the bands were recorded using a GelDoc 2000 transilluminator system (BioRad, Richmond, California, USA). The DNA sequence of both strands of the PCR products were determined by Beckman Coulter Genomics (Essex, United Kingdom) using an ABI 3730XL Sanger sequencer (Sanger et al., 1977) with the original primers 27f and 1390R. 16S rRNA gene sequences were aligned using Mega 6 (Tamura et al., 2013) and compared with those available in the NCBI GenBank database using the megaBLAST program (Zhang et al., 2000). Each sequence was aligned with the first 10 database sequences giving the highest scores of sequence similarity and the quality of the database sequences was assessed. Identification at genus level was defined as a 16S rRNA gene sequence similarity of ≥ 97% with that of the closest sequence in GenBank. Enzyme assays The predominant purified isolates obtained after enumeration were used for screening for extracellular enzyme activities. Each of the predominant isolates was transferred into 10 mL of sterile nutrient broth (Merck, Germany) and incubated at 30°C in a stirring incubator for 16 h. Then, the absorbance was measured using a spectrophometer (VWR, France) and adjusted to the same value for all cultures. Aliquots of these cultures were taken and placed in microplates for screening for cellulase, hemicellulase (xylanase), amylase and β-glucosidase activities. The protocol was based on methods described by Popova and Deng (2010); Shao and Lin (2018) with some modifications.
For cellulase, xylanase, amylase and β-glucosidase activities, the activities were assayed using carboxymethyl-cellulose, xylan, starch and p-nitrophenyl-β-D-glucopyranoside as substrates, respectively. All substrates were of analytical grade (from Sigma–Aldrich, St. Louis, MO, USA). For cellulase, hemicellulase and amylase assays, a 100 µL aliquot of culture suspension of each isolate was put into a well in a 96-well microplate with 50 µL of Mac Ilvain buffer (pH 7). Then 50 µL of the substrate was added to each well. The reaction solution was mixed by pipetting up and down several times with multi-channel pipette. Each microplate had a minimum of three replicates for each substrate and three control wells containing
Moutou-Tchitoula1 et al. 781 sterile water, buffer and substrate. The microplate was then incubated at 37°C for 30 min. Aliquots (25 µL) of this reaction solution were transferred to a 96-well microplate with 200 µL of Somogyi’s reagent. The microplates were held at 100°C for 20 min and then cooled on ice. The microplate wells were covered to avoid evaporation. Finally, aliquots (50 µL) of the resulting reaction solution were transferred to a 96-well microplate with 25 µL of Nelson’s reagent. Then, 200 µL of distilled water was added to each well and mixed by pipetting up and down several times using a multi-channel pipette. Absorbance was measured at 630 nm using a microplate reader (Bio-Tek EL800, Bio-Tek Instruments, Winooski, VT). The activities were expressed as pmol glucose min-
1ml-1 for cellulase, xylanase and amylase. The glucose concentrations in the reaction solution were calculated using a calibration curve constructed using glucose solutions at various concentrations.
For -Glucosidase activity, 50 µL of culture suspension from each isolate was put into a well in a 96-well microplate with 25 µL of Mac Ilvain buffer (pH 7). Then 50 µL of the substrate was added to each well and the contents were mixed by pipetting up and down several times. Controls were prepared similarly but without bacterial suspensions. After incubating the microplate at 37 °C for 1 h, they were centrifuged at 14,000 g for 5 min and 50 µL of the supernatant was transferred to a 96-well microplate with 250 µL of 2% Na2CO3 to stop the enzyme reaction. Absorbance was measured at 410 nm using the same microplate reader. The p-Nitrophenol concentrations in the reaction solutions were calculated using a calibration curve that was constructed using pNP standards. There were five replicates for each bacterial suspension and each microplate included a minimum of three replicates three controls. The enzyme activity was expressed as pmol of PNP min-1 mL-1. Statistical analysis The correlations between of isolates obtained from Ntoba mbodi and their potential enzyme activities were tested by principal component analysis (PCA) using ADE 4 (Thioulouse et al., 1997). A permutation test was used to determine whether the differences were large enough to reject the null hypothesis. Significant differences between isolates on the first two axes were determined by one way ANOVA after verifying that the data distribution was normal. The Tukey post hoc test was used after ANOVA. Values were considered different at p < 0.05. Correlations were analyzed using Statgraphics, Centurion XVI (Sigma Plus, France) to establish statistical relationships.
RESULTS
Bacterial enumeration and isolation of predominant morphotypes in Ntoba mbodi
The density of total mesophilic bacteria in Ntoba mbodi varied with the samples (Figure 1). All four samples had a high number of mesophilic bacteria cultivable ranging from 1.67×10
10 to 2.8×10
10 CFU g
-1 (Figure 1). Bacterial
density for samples 1 and 2 were of the same order and lower than samples 3 and 4. Based on macroscopic and microscopic observations of the colonies at the highest dilution, eleven distinct morphotypes were isolated from the four samples of Ntoba mbodi. These isolates are listed in the Table 1. Of the eleven isolates, five (M1, M2, M3, M4 and M7) were found in the all samples and represented the core bacteria of Ntoba mbodi. Isolates

782 Afr. J. Microbiol. Res.
Figure 1. Enumeration of bacterial cells in the different samples of Ntoba-mbodi.
M10 and M11 were specific to samples 1 and 2 while M12 and M8 were only found in samples 4 and 2 respectively (Table 1). Phylogenetic affiliation of predominant isolates Verification of PCR products of the 16S rRNA genes of predominant isolates by electrophoresis in agarose gel indicated that the amplicons were about 1360 pb long (data not shown). The amplicons were nearly full-length and provided enough phylogenetic information for taxonomical identification of 55% of the isolates down to species level (Table 1). Most of the sequences showed 97% identity to the closest sequences already in the databases, and none was found to be apparently chimeric. About 73% of predominant were classified as Bacillus related species including a Bacillus pumilus strain, three strains of Bacillus altitudinis, two strains of Bacillus sp. C-32, two strains of Bacillus sp.01105 and Bacillus aerophilus strain. Non-Bacillus related species account for about 27% of the population and comprised Staphylococcus sciuri, Kurthia sp. B2 and Bacterium NLAE-ʫ-H375. The set of isolates found across all samples of Ntoba mbodi were closest to Bacillus related species (Bacillus altitudinis strain 126YG20, Bacillus sp. C-32) and Staphylococcus sciuri. Screening the isolated bacterial strains for polysaccharide-degrading capacity In the principal component analysis (PCA) of the four enzyme activities and the eleven isolates, axes 1 and 2
accounted for 48.1 and 21.6% of the variance respectively (Figure 2A). Together, these two axes explained about 70% of the variance. Xylanase, cellulase and β-glucosidase activities were opposed to amylase activity on axis 1. One group consisted of the strains M1, M4, M6, M10 and M12. Four of these strains were affiliated with the genus Bacillus and one with the genus Kurthia. They all showed higher xylanase and cellulase activities and, to a lesser extent β-glucosidase activity, than other isolates (Figure 3). A second group had two strains (M8 and M9), one affiliated with the genus Bacillus and one (M2) whose the closest relative was not well determined and classified as a relative of Bacterium NLAE-ʫ-H375 with 96% identity. Unlike the first group, the β-glucosidase, xylanase and cellulase activities of these strains were not significantly different from the other isolates (Figure 3). The strains M3 and M11, affiliated with the genus Bacillus, formed a third group. Their enzyme activities were the same order as those of the first group or second group. There were no significant differences in amylase activities between isolates and the amylase activity was lower than the cellulase, xylanase and β-glucosidase activities (data not shown). DISCUSSION This study set out to assess the variability in the bacterial populations across samples of Ntoba mbodi and the potential enzyme activities associated with the predominant strains isolated from Ntoba mbodi. Consistent with previous studies (Ouoba et al., 2010; Vouidibio-Mbozo et al., 2017), we showed that Ntoba mbodi contains high density of bacteria that varied with
0.00E+00
5.00E+09
1.00E+10
1.50E+10
2.00E+10
2.50E+10
3.00E+10
3.50E+10
Sample 1 Sample 2 Sample 3 Sample 4
Ba
cte
rial
co
un
t (U
FC g
-1)

Moutou-Tchitoula1 et al. 783
Figure 2. Principal component analysis (PCA) based on enzyme activities of the predominant strains. (A) Correlation circle. (B) Ordination of the strains in the plane defined by axes 1 and 2 of the PCA highlighting the profile of enzyme activity of the strains.
1
2
3
B
A

784 Afr. J. Microbiol. Res.
Figure 3. Enzyme activities of the predominant strains. (1) cellulase; (2) xylanase; (3) β-glucosidase. 1 and 2 are expressed as pmol glucose min-1 mL-1 and C as pmol PNP min-1 mL-1. Co: controls. Different letters means that the results are significantly different (p<0.05) according to Fischer's PLSD-test. Mean values (n=5) ± standard error.
the samples from different producers. The variability of the bacterial community underlines that, like most indigenous food fermentation processes, the fermentation of Ntoba mbodi is driven by indigenous microorganisms originating in the raw materials, in autochthonous starter cultures, or in the processing environment itself (Bokulich and Mills, 2013).
The detection of predominant groups within the bacterial assemblages is the basis for determining the “functional starter” (Leroy and De Vuyst, 2004). Therefore we assessed the predominant taxa in samples of Ntoba mbodi from different producers. We found that Ntoba mbodi contain only a few predominant taxa, mainly Bacillus related species (72%), which were closest to
1
2
3
4
Fig.3: Enzyme activities of the predominant strains. (1) cellulase; (2) xylanase; (3) β-glucosidase. 1 5
and 2 are expressed as pmol glucose min-1 mL-1 and C as pmol PNP min-1 mL-1. Co: controls. Different 6
letters means that the results are significantly different (p<0.05) according to Fischer's PLSD-test. 7
Mean values (n=5) ± standard error. 8
9
10
11
(1)
(2)
(3)
Cel
lula
se (
pm
ol g
luco
se m
in-1
mL-1
) X
ylan
ase
(p
mo
l glu
cose
min
-1 m
L-1)
β-g
luco
sid
ase
(p
mo
l PN
P m
in-1
mL-1
)

Bacillus pumilus, Bacillus altitudinis strain 126YG20, Bacillus sp. C-32, Bacillus sp.01105, Bacillus altitudinis strain SH156 and Bacillus aerophilus strain JF11. The presence of Bacillus related species is in agreement with previous studies in Ntoba mbodi (Louembe et al., 2003;
Vouidibio-Mbozo et al., 2017) and in other alkaline food
fermentation products (Steinkraus, 2002; Parkouda et al., 2009; Tamang et al., 2016). Comparison at species level revealed that the Bacillus related species previously reported in Ntoba mbodi, including B. subtilis, B. amyloliquefaciens, B. megaterium, B. macerans, B. cereus, B. polymixa, B. brevis and B. pumilus (Louembe et al., 2003; Kobawila et al., 2005), B. safensis, B. siamensis and Lysinibacillus louembei (Ouoba et al.,
2015; Vouidibio Mbozo et al., 2017), were not found at
the highest dilution of inoculum (Ntoba mbodi). The differences in the composition of bacteria assemblage in African alkaline-fermented foods can be explained by the variability in raw materials (Ouoba et al., 2010; Tamang et al., 2016). But the discrepancy between our results and those reported previously is more likely to be related to the different approaches used for determining the predominant strains using bacterial cultures. In the present study, only colonies obtained from the highest positive dilution of Ntoba mbodi were distinguished, on the basis of differences in macroscopic and microscopic observations, and isolated. These isolates were considered to be the predominant bacterial taxa in Ntoba mbodi. In previous studies, however, the various phenotypes of the Bacillus colonies were selected after
enumeration of mesophilic bacteria (Vouidibio-Mbozo et
al., 2017) and isolated at various dilution factors. The predominance in previous studies represents the percentage of distinct species among the total number of isolates but did not reflect the abundance of each species in initial sample of Ntoba mbodi. Knowing the abundance of different species can provide insight into how a community functions (Verberk, 2011). This raises the question of the relevance of isolates obtained in the present study. Do these bacterial species drive the fermentation of Ntoba mbodi or are they simply transient or inactive forms?
To address this question, we assessed whether there were predominant cultivable mesophilic bacteria found in all samples of Ntoba mbodi, irrespective of the producers. Although the household-scale fermentation process depends upon microorganisms from the local environment, we found a set of five isolates affiliated to Bacillus altitudinis strain 126YG20, Bacillus sp. C-32 (2 isolates), Bacterium NLAE-ʫ-H375 and Staphylococcus sciuri isolate YXY-5 that were found in all samples of Ntoba mbodi. These findings confirm our hypothesis that there is a set of bacteria common to all samples of Ntoba mbodi, irrespective of their origins, in Brazzaville (Congo) and suggest that the colonies isolated in the present study were not simply transient bacterial taxa. Determination of the set of bacteria common to all
Moutou-Tchitoula1 et al. 785
samples of Ntoba mbodi confirmed the convergence in taxonomic profiles of predominant bacteria regardless of the origin of the Ntoba mbodi. The convergence in taxonomic profiles of bacterial community has also been reported for other fermentation processes (Jia et al., 2016). Interestingly, the majority (60%) of the bacteria common to all samples of Ntoba mbodi were Bacillus-related species. Our results confirmed the importance of members of the genus Bacillus in vegetable fermentation processs (Steinkraus, 2002; Parkouda et al., 2009; Tamang et al., 2016). Bacillus spp. are also known for their ability to produce toxins that may cause food poisoning (Stenfors et al., 2008) and potentially pathogenic bacteria such Bacillus cereus have been
reported in Ntoba mbodi (Vouidibio-Mbozo et al., 2017).
Interestingly, these bacteria were not found among predominant isolates obtained in the present study.
Furthermore, the presence of strains affiliated with Staphylococcus sciuri in all samples of Ntoba mbodi in the present study raised the question of role of this bacteria group in fermentation of Ntoba mbodi. Members of this bacterial group are widespread in nature and have been isolated from various food products of animal origin (Garcia et al., 2002; Papamanoli et al., 2002). They are commonly present on skin and mucus membranes of a wide range of pets and farm animals (Kloos, 1980). It is, therefore, possible that their presence in Ntoba mbodi is from contamination during Ntoba mbodi processing due to handling practices. Recently, however, Amao et al. (2018) demonstrated that Staphylococcus spp. played important role by increasing free amino acids in iru (fermented locust beans) and by providing lipolytic enzyme activity during the fermentation. The role of Staphylococcus spp. in the fermentation of Ntoba mbodi still needs to be determined.
The softening of cassava leaves in Ntoba mbodi has been ascribed to bacterial enzyme activities (Mokemiabeka et al., 2011). But, unlike the extensively studied role of bacterial enzyme activities in softening cassava roots (Brauman et al., 1996; Kouhoundé et al., 2014), this has not previously been confirmed. Since it was still unclear whether the enzymes were from the bacteria or from the cassava leaves in Ntoba mbodi, the predominant isolates obtained were screened for some polysaccharide-degrading enzyme activities. Cellulase, xylanase, amylase and β-glucosidase were targeted because plant biomass is made of complex polysaccharides, primarily cellulose and hemicelluloses (Cragg et al., 2015). Out of eleven predominant isolates, eight showed cellulase activity and five showed xylanase and β-glucosidase activities. These findings demonstrate for the first time the bacterial origin of enzyme activities in Ntoba mbodi and confirm that microorganisms change the chemical composition of the raw materials during fermentation (Tamang et al., 2016). It is noteworthy to underline that 18% of predominant isolates including Bacillus related species did not have any of the enzyme

786 Afr. J. Microbiol. Res. activities targeted. This finding suggests that despite the abundance and the role of Bacillus related species in alkaline fermentation processes (Tamang et al., 2016), not all members of this bacterial group are involved in the degradation of polysaccharides in Ntoba mbodi. It is possible that, in this alkaline fermentation, these bacteria may be involved in the production of bioactive and nutritive compounds during the process (Marco et al., 2017). The absence of amylase activity in all isolates was not expected. These results can be explained by the low starch content in cassava leaves compared to the roots. Conclusions Our study has shown that the density of bacteria in Ntoba mbodi was high and varied with the origin of the samples. Predominant bacteria in Ntoba mbodi comprised a few bacterial taxa dominated by Bacillus related species. Although the fermentation of Ntoba mbodi depends on microorganisms from the environment, there is a core set of bacteria that was found in all final products. More than 50% of predominant strains had enzyme activities for breaking down cellulose and xylan, which are the main components of plant materials. Further studies are needed to investigate the diversity of bacteria in Ntoba mbodi, without using cultures, to gain a better insight into the role played by bacteria in the functioning of this particular ecosystem.
CONFLICT OF INTERESTS The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS Campus France Congo Brazzaville provided funding to Didine Priscilla Moutou-Tchitoula
REFERENCES Amao, JA, Odunfa SA, Mbom CN (2018). The role of Staphylococcus
species in the production of iru during the fermentation of African locust beans (Parkia biglobosa). Food Research 2:187-193.
Bokulich NA, Mills DA (2013). Facility-specific “House” microbiome drives microbial landscapes of artisan cheesemaking Plants. Applied and Environmental Microbiology 79:5214-5223.
Brauman A, Keleke S, Malonga M, Miambi E, Ampe F (1996). Microbiological and Biochemical Characterization of Cassava Retting, a Traditional Lactic Acid Fermentation for Foo-Foo (Cassava Flour) Production. Applied and Environmental Microbiology 62:2854-2858.
Cragg SM, Beckham GT, Bruce NC, Bugg TDH, Distel DL, Dupree P, Etxabe AG, Goodell BS, Jellison J, McGeehan JE, McQueen-Mason SJ, Schnorr K, Walton PH, Watts JEM and Zimmer M (2015). Lignocellulose degradation mechanisms across the Tree of Life. Current Opinion in Chemical Biology 29:108-119.
Garland JL, Lehman RM (1999). Dilution/extinction of community phenotypic characters to estimate relative structural diversity in mixed communities. FEMS Microbiology Ecology 30:333-343.
Gomez EDV, Garland JL, Roberts MS (2004). Microbial structural
diversity estimated by dilution–extinction of phenotypic traits and T-RFLP analysis along a land-use intensification gradient. FEMS Microbiology Ecology 49:253-259.
Sawadogo‐Lingani H, Lei V, Diawara B, Nielsen DS, Møller PL, Traoré AS, Jakobsen M (2007). The biodiversity of predominant lactic acid bacteria in dolo and pito wort for the production of sorghum beer. Journal of Applied Microbiology 103:765-777.
He J, Su L, Sun X, Fu J, Chen J, Wu J (2014). A novel xylanase from Streptomyces sp. FA1: purification, characterization, identification, and heterologous expression. Biotechnology and Bioprocess Engineering 19:8-17.
Jia Y, Wilkins D, Lu H, Cai M, Lee PKH (2016). Long-term enrichment on cellulose or xylan causes functional and taxonomic convergence of microbial communities from anaerobic digesters. Applied and Environmental Microbiology 82:1519-1529.
Kloos WE (1980). Natural populations of the genus Staphylococcus. Annual Review of Microbiology 34:559-592.
Kobawila SC, Louembe D, Keleke S, Hounhouigan J, Gamba C (2005). Reduction of the cyanide content during fermentation of cassava roots and leaves to produce bikedi and ntoba mbodi, two food products from Congo. African Journal of Biotechnology 4:689-696.
Kostinek M, Specht I, Edward VA; Pinto C; Egounlety M, Sossa C; Mbugua S, Dortue C, Thonart P, L.Taljaar L, Mengu M, Franz CMAP, Holzapfel WH (2007). Characterisation and biochemical properties of predominant lactic acid bacteria from fermenting cassava for selection as starter cultures. International Journal of Food Microbiology 114:342-351.
Kouhoundé SHS, Adéoti K, Delvigne F, Savadogo A, Traore AS, Thonart P (2014). The use of microorganisms of cassava retting for the production of pectinolytic enzymes. Journal of Microbiology, Biotechnology and Food Sciences 4:277-281.
Lei V, Jakobsen M (2004). Microbiological characterization and probiotic potential of koko and koko sour water, African spontaneously fermented millet porridge and drink. Journal of Applied Microbiology 96:384-397.
Leroy F, De Vuyst L (2004). Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends in Food Science and Technology 15: 67-78.
Louembe D, Kobawila SC, Bouanga Kalou G, Kéléké S (2003). Etude microbiologique des feuilles fermentées de manioc “Ntoba Mbodi”. Tropicultura 21:106-111.
Marco ML, Heeney D, Binda S, Cifelli CJ, Cotter PD, Foligné B, Gänzle M, Kort R, Pasin G, Pihlanto A, Smid EJ, Hutkins R (2017). Health benefits of fermented foods: microbiota and beyond. Current Opinion in Biotechnology 44:94-102.
Miambi E, Guyot JP, Ampe F (2003). Identification, isolation and quantification of representative bacteria from fermented cassava dough using an integrated approach of culture-dependent and culture-independent methods. International Journal of Food Microbiology 82:111-120.
Mokemiabeka S, Dhellot J, Kobawila SC, Diakabana P, Ntietie RN, Loukombo Nyanga-Koumou AG Louembe D (2011). Softening and mineral content of cassava (Manihot esculenta Crantz) Leaves during the Fermentation to produce Ntoba mbodi. Advance Journal of Food Science and Technology 3:418-423.
Ouoba LII, Nyanga-Koumou CAG, Parkouda C, Sawadogo H, Kobawila SC, Keleke S, Diawara B, Louembe D, Sutherland JP (2010). Genotypic diversity of lactic acid bacteria isolated from African traditional alkaline-fermented foods. Journal of Applied Microbiology 108:2019-2029.
Ouoba LII, Vouidibio Mbozo AB, Thorsen L, Anyogu Amarachukwu, Nielsen DS, Kobawila SC, Sutherland JP (2015). Lysinibacillus louembei sp. nov., a spore-forming bacterium isolated from Ntoba Mbodi, alkaline fermented leaves of cassava from the Republic of the Congo. International Journal of Systematic and Evolutionary Microbiology 65:4256-4262.
Papamanoli E, Kotzekidou P, Tzanetakis N, Litopoulou-Tzanetaki E (2002). Characterization of Micrococcaceae isolated from dry fermented sausage. Food Microbiology 19:441-449.
Parkouda C, Nielsen DS, Azokpota P, Ouoba LII, Amoa-Awua WK, Thorsen L, Hounhouigan JD, Jensen JS, Tano DK, Diawara B,

Jakobsen M (2009). The microbiology of alkaline-fermentation of indigenous seeds used as food condiments in Africa and Asia. Critical Reviews in Microbiology 35:139-156.
Popova IE, Deng S (2010). A high-throughput microplate assay for simultaneous colorimetric quantification of multiple enzyme activities in soil. Applied Soil Ecology 45:315-318.
Ray R, Montet D (2017). Fermented Foods, Part II. Boca Raton: CRC pp. 21-45.
Sanger F, Nicklen S, Coulson AR (1977). DNA sequencing with chain- terminating inhibitors. Proceedings of the National Academy of Sciences 74:5463-5467.
Sarkar P, Nout M (2014). Handbook of indigenous foods Involving alkaline fermentation. Boca Raton: CRC Press. Available at http://library.wur.nl/WebQuery/wurpubs/480686
Shao Y, Lin AHM (2018). Improvement in the quantification of reducing sugars by miniaturizing the Somogyi-Nelson assay using a microtiter plate. Food Chemistry 240:898-903.
Steinkraus KH (1996). Handbook of Indigenous Fermented Foods. Second Edition, Marcel Dekker, Inc., New York. Marcel Dekker Inc., pp. 493-496.
Steinkraus KH (2002). Fermentations in world food processing. Comprehensive reviews in Food Science and Food Safety 1:23-32.
Stenfors ALP, Fagerlund A, Granum PE. (2008). From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiology Reviews 32:579-606.
Tamang JP, Watanabe K and Holzapfel WH (2016) Review: Diversity of Microorganisms in Global Fermented Foods and Beverages. Frontiers in Microbiology 7:1-28.
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013). MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0, Molecular Biology and Evolution 30:2725-2729.
Moutou-Tchitoula1 et al. 787 Thioulouse J, Chessel D, Dolédec S, Olivier JM (1997). ADE-4: a
multivariate analysis and graphical display software. Statistics and Computing 7:75-83.
Verberk, W (2011). Explaining General Patterns in Species Abundance and Distributions. Nature Education Knowledge 3:38.
Vouidibio-Mbozo AB, Kobawila SC, Anyogu A (2017). Investigation of the diversity and safety of the predominant Bacillus pumilus sensu lato and other Bacillus species involved in the alkaline fermentation of cassava leaves for the production of Ntoba Mbodi. Food Control 82:154-162.
Zhang Z, Schwartz S, Wagner L, Miller W (2000). A greedy algorithm for aligning DNA sequences. Journal of Computational Biology 7:203-214.

Vol. 12(32), pp. 788-795, 28 August, 2018
DOI: 10.5897/AJMR2018.8873
Article Number: F59038158405
ISSN: 1996-0808
Copyright ©2018
Author(s) retain the copyright of this article
http://www.academicjournals.org/AJMR
African Journal of Microbiology Research
Full Length Research Paper
Isolation and characterization of starch degrading rhizobacteria from soil of Jimma University Main
Campus, Ethiopia
Gudeta Dida
Microbial Biotechnology Research Program, Agricultural Biotechnology Research Center, Ethiopian Institute of Agricultural Research, Ethiopia.
Received 25 April, 2018; Accepted 10 August, 2018
Starch degrading bacteria are important for different industries such as food, fermentation, textile, and paper. The aim of this study is to isolate and characterize bacteria able to degrade starch from the rhizospheres of various plants at four sites located in Jimma University main campus. Collected soil samples were labeled as kobo (AJUMC), Avocado (BJUMC), Banana (CJUMC), and Cana indica (DJUMC) respectively. Soil samples were serially diluted in sterilized peptone water, poured on sterilized starch agar plates, and incubated at 32ºC for 48 h. The representative colonies showing different morphology was randomly picked up using the streaking method on nutrient agar. A total of 53 bacterial isolates were obtained from the soils rhizospheres. Microscopic characteristics showed that among the 53 isolates, 38 (72%) were Gram-positive rod shaped bacteria, while 15 (28%) were Gram-negative rod shaped bacteria. Based on the biochemical tests, the results revealed that the 38 isolates belonged to the genera Bacillus while the remaining isolates belonged to the genera Pseudomonas. All isolates were catalase positive and only 15 isolates (Pseudomonas) were KOH positive with negative growth at 80°C, while the 38 (Bacillus) isolates have positive growth at 80ºC. The highest values of starch degrading index were the Gram positive bacteria isolates. The amylase activity was also carried out with respect to time, temperature and pH of the media. The maximum activity of amylase at different temperatures from 35 to 45°C was recorded at 35°C (0.94 U/ml) within 24 h, while maximum activity at different pH from 5 to 9 was recorded at pH 7 (1 U/ml). Key words: Rhizobacteria, starch degrading, amylase enzyme, Bacillus, Pseudomonas.
INTRODUCTION The diversity of microbes are associated with plant roots (Berendsen, 2012). The narrow zone of soil directly surrounding the root system is referred to as rhizosphere, while the term „rhizobacteria‟ implies a group of rhizosphere bacteria competent in colonizing the root
environment (Ahmad and Kibret, 2014). The rhizosphere is soil ecological environment for plant-microbe interactions involving colonization of different microorganisms in associative, symbiotic, naturalistic, or parasitic interactions depending upon plant nutrient
E-mail: [email protected].
Author(s) agree that this article remain permanently open access under the terms of the Creative Commons Attribution
License 4.0 International License

status in soil, soil environment, and the type of microorganism proliferating in the rhizosphere zone (Kennedy, 2005). When microbes are close to epidermis, plants secrete signal molecules against invasive microbes in the root zone. Consequently, differentiation takes place between pathogenic, symbiotic, or naturalistic adaptation of microbes with the plant (Hayat et al., 2010).
Rhizosphere bacteria, especially species of Pseudomonas and Bacillus have been identified in the rhizosphere of various leguminous and non-leguminous plants (Halverson and Handelsman, 1991; Parmar, 1999). Different bacterial genera are vital components of soils. They are involved in various biotic activities of the soil ecosystem to make it dynamic for nutrient turnover. Ahmad and Kibret (2014) reported that based on the functional activities, rhizobacteria are classified as (i) bio-fertilizers (increasing the availability of nutrients to plant), (ii) phytostimulators (iii) Rhizoremediators (degrading organic pollutants), (iv) biopesticides (controlling diseases, by production of different metabolites).
There are various starch degrading microorganisms from different sources (Aiba et al., 1983; Tonkova et al, 1993; Kathiresan and Manivannan, 2006). Rhizospere bacteria are one sources of starch degrading microorganism as it contains mostly starch substrate (Alariya et al., 2013). Among the bacterial species, Bacillus spp. Bacillus amyloliquefaciens, Bacillus licheniformis and Pseudomonas are widely used for enzyme production. Other species like Bacillus cereus and Bacillus subtilis also used for production of the amylase enzyme. Amylases produced from B. licheniformis, Bacillus stearothermophilus, and B. amyloliquefaciens show promising potential in a number of industrial applications in processes such as food, fermentation, textiles and paper industries. (Konsoula and Liakopoulou-Kyriakides, 2007; Ha et al., 2001). Thus, the objective of the present study is to isolate and characterize the starch degrading bacteria from different soils rhizopheres of Jimma University. MATERIALS AND METHODS
Description of study area
The study was conducted at Jimma, located 353 km southwest of Addis Ababa, the capital city of Ethiopia. The town‟s geographical coordinates are 7°41‟N latitude, 36°50‟E longitude, and an average altitude of 1, 780 m above sea level. It lies in the climatic zone locally known as “Woyna Daga” (1,500 - 2,400 m above sea level) which is considered to be ideal for agriculture as well as human settlement. The town is generally characterized by warm climate with a mean annual maximum temperature of 30°C and a mean annual minimum temperature of 14°C. The annual rainfall ranges from 1138 - 1690 mm. The maximum precipitation occurs during the three months period from June through August, with minimum rainfall occurring in December and January. From a climatic point of view, abundant rainfall makes this region one of the best watered of Ethiopian highland areas, conducive for agricultural production (Alemu et al., 2011).
Dida 789 Samples collection Ten grams of soil were collected from the rhizopheres of various plants at four sites located in Jimma University. The soil sample were put in the sterilized bag and transferred immediately to the laboratory. Sample preparation and isolation of starch degrading bacteria Each 10 g of soil samples from different sites was mixed with 90 ml of sterile peptone water in different 250 ml beaker and homogenized in a flask for ten minutes using orbital shaker at 110 rpm. Subsequently, 1 ml of each sample was transferred aseptically into 9 ml of sterile peptone water and mixed thoroughly using vortex. The homogenates was serially diluted up to 10-5; thereafter, 0.1 ml aliquot of appropriate dilution was spread properly on starch agar plates. The plates were incubated at 32ºC for 48 h. The bacterial isolates showing different colonies morphology were picked out and purified. A fourth gram of soil samples was collected from four different sites (10 g from each site) of Jimma University plant rooted by streaking on nutrient agar (Oseni and Ekperigin, 2013). Identification of bacterial isolates The bacterial isolates were subjected to identification using morphological characteristics such as Gram reaction and endospore formation according to Silva and Nahas (2002). Biochemical tests including catalase test, KOH test, and the growth at 80°C were also performed (Naik et al., 2008; Williams et al., 1990).
Determination of starch degrading index (SDI) Ability of the bacterial isolates to degrade starch was described by the starch degrading index (SDI): the ratio of the total diameter of clear zone and colony diameter. On the basis of degrading index, potential colonies with the best efficiency were selected as best starch degrading colonies (Nusrat and Rahman, 2007). Screening for amylase activity (Starch Iodine Test) Isolated strains were picked up from each plate containing pure culture and streaked in straight lines in starch agar plates as carbon source. The plates were incubated at 32ºC for 48 h. After incubation, the plates were flooded with (Gram‟s iodine- 250 mg iodine crystals added to 2.5 g potassium iodide solution, and 125 ml of distilled water, to produce a deep blue coloration (Prameela et al., 2016).
Amylase production Amylase was produced by using complex medium containing starch 1.0%, yeast extract 0.04%, (NH4)2HPO4 0.4%, KCl 0.1% and MgSO47H2O 0.05%, and semisynthetic medium containing peptone 0.4%, (NH4)2HPO4 0.4% and KCl 1.0% (Kim et al., 1995). Enzyme assay
A suitable volume of isolated broth culture was incubated for 48 h; thereafter, it was centrifuged at 5000 rpm for 20 min. at 4°C. Supernatant was recovered and amylase determined using

790 Afr. J. Microbiol. Res.
Figure 1. Morphology and biochemical tests.
Table 1. Total isolated colonies and their codes from 4 sites of rhizobacteria (RB).
Isolate code No. of isolates Plant rhizosphere
AJUMC 17 Kobo
BJUMC 10 Avocado
CJUMC 11 Banana
DJUMC 15 Cana indicia
spectrophotometer. One milliliter of crude enzyme was placed into a test tube and 1 ml of 1% soluble starch in sodium phosphate buffer (pH 7) was added into test tube. The test tubes were covered and incubated at 35°C for 10 min (Miller, 1959). Subsequently, 2.0 ml DNS reagent was added in each tube to stop the reaction and kept in boiling water bath for 10 min. After cooling at 25°C, final volume was made to 10 ml with distilled water. The absorbance was measured at 540 nm using spectrophotometer.
Determination of optimum pH
One percent of starch was used as a substrate. Four sets of substrate solution were prepared in sodium phosphate buffer with adjusted pH values 5, 6, 7, 8 and 9. One milliliter of each of crude enzyme solution was added into buffer tubes. Mixture was incubated at 35°C for 10 min; thereafter, 2 ml of DNS reagent was added to terminate the reaction and the mixture was incubated in boiling water for 10 min. After cooling at room temperature, final volume was made to 10 ml with distilled water and the activity of the enzymes was measured based on the absorbance at 540 nm.
Determination of optimum temperature
One ml of the substrate was distributed into six test tubes and 1 ml of phosphate buffer pH 7 was added in each test tubes. Tubes were marked with different temperature (at 30, 35, 40°C). 1 ml of crude enzyme solution was added in each tube; thereafter, tubes were incubated at specific temperature for 10 min. Reactions were terminated by adding 2 ml DNS reagent and the mixture incubated in boiling water for 10 min. After cooling at room temperature, final volume was made to 10 ml with distilled water and the activity of enzymes was determined by taking the absorbance at 540 nm.
Table 2. Morphological features of 53 isolates of rhizobacteria.
Morphology feature Bacteria isolate Number
Cell shape Rod 53
Colony arrangement Chain 41
Single bacillus 12
Endospore + 38
- 15
Gram reaction + 38
- 15
Determination of optimum time The effect of different time duration such as 24, 48 and 72 h on amylase activity was observed. Determination of optimum starch concentration Similarly, to observe the effect of different substrate concentration on amylase activity, the dialyzed aliquot was added to different starch concentrations (0.5, 1.0, 1.5 and 2%) and the activity was observed following the method of Miller (1959).
RESULTS AND DISCUSSION
The morphology and biochemical tests (Figure 1) resulted from four samples of Jimma University main campus plant roots of 53 isolates which were summarized in Tables 1 and 2. In the present study, on the basis of their gram reaction, out of 53 isolates 38(72%) had rod shape Gram positive bacteria while 15(28%) were Gram negative rod shaped bacteria .All the Gram negative were rod shaped, non-spore forming; similarly, Gram positive bacteria were rod shaped and spore former. Biochemically, 53 isolates were catalase
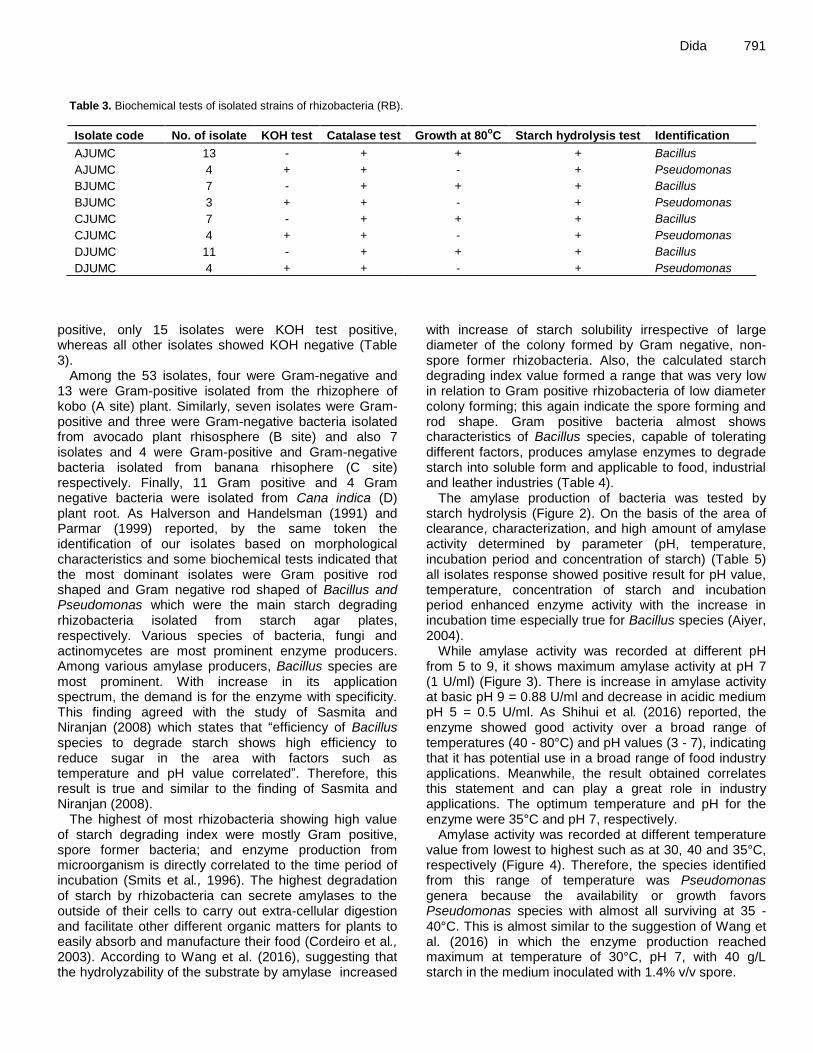
Dida 791
Table 3. Biochemical tests of isolated strains of rhizobacteria (RB).
Isolate code No. of isolate KOH test Catalase test Growth at 80oC Starch hydrolysis test Identification
AJUMC 13 - + + + Bacillus
AJUMC 4 + + - + Pseudomonas
BJUMC 7 - + + + Bacillus
BJUMC 3 + + - + Pseudomonas
CJUMC 7 - + + + Bacillus
CJUMC 4 + + - + Pseudomonas
DJUMC 11 - + + + Bacillus
DJUMC 4 + + - + Pseudomonas
positive, only 15 isolates were KOH test positive, whereas all other isolates showed KOH negative (Table 3).
Among the 53 isolates, four were Gram-negative and 13 were Gram-positive isolated from the rhizophere of kobo (A site) plant. Similarly, seven isolates were Gram-positive and three were Gram-negative bacteria isolated from avocado plant rhisosphere (B site) and also 7 isolates and 4 were Gram-positive and Gram-negative bacteria isolated from banana rhisophere (C site) respectively. Finally, 11 Gram positive and 4 Gram negative bacteria were isolated from Cana indica (D) plant root. As Halverson and Handelsman (1991) and Parmar (1999) reported, by the same token the identification of our isolates based on morphological characteristics and some biochemical tests indicated that the most dominant isolates were Gram positive rod shaped and Gram negative rod shaped of Bacillus and Pseudomonas which were the main starch degrading rhizobacteria isolated from starch agar plates, respectively. Various species of bacteria, fungi and actinomycetes are most prominent enzyme producers. Among various amylase producers, Bacillus species are most prominent. With increase in its application spectrum, the demand is for the enzyme with specificity. This finding agreed with the study of Sasmita and Niranjan (2008) which states that “efficiency of Bacillus species to degrade starch shows high efficiency to reduce sugar in the area with factors such as temperature and pH value correlated”. Therefore, this result is true and similar to the finding of Sasmita and Niranjan (2008).
The highest of most rhizobacteria showing high value of starch degrading index were mostly Gram positive, spore former bacteria; and enzyme production from microorganism is directly correlated to the time period of incubation (Smits et al., 1996). The highest degradation of starch by rhizobacteria can secrete amylases to the outside of their cells to carry out extra-cellular digestion and facilitate other different organic matters for plants to easily absorb and manufacture their food (Cordeiro et al., 2003). According to Wang et al. (2016), suggesting that the hydrolyzability of the substrate by amylase increased
with increase of starch solubility irrespective of large diameter of the colony formed by Gram negative, non-spore former rhizobacteria. Also, the calculated starch degrading index value formed a range that was very low in relation to Gram positive rhizobacteria of low diameter colony forming; this again indicate the spore forming and rod shape. Gram positive bacteria almost shows characteristics of Bacillus species, capable of tolerating different factors, produces amylase enzymes to degrade starch into soluble form and applicable to food, industrial and leather industries (Table 4).
The amylase production of bacteria was tested by starch hydrolysis (Figure 2). On the basis of the area of clearance, characterization, and high amount of amylase activity determined by parameter (pH, temperature, incubation period and concentration of starch) (Table 5) all isolates response showed positive result for pH value, temperature, concentration of starch and incubation period enhanced enzyme activity with the increase in incubation time especially true for Bacillus species (Aiyer, 2004).
While amylase activity was recorded at different pH from 5 to 9, it shows maximum amylase activity at pH 7 (1 U/ml) (Figure 3). There is increase in amylase activity at basic pH 9 = 0.88 U/ml and decrease in acidic medium pH 5 = 0.5 U/ml. As Shihui et al. (2016) reported, the enzyme showed good activity over a broad range of temperatures (40 - 80°C) and pH values (3 - 7), indicating that it has potential use in a broad range of food industry applications. Meanwhile, the result obtained correlates this statement and can play a great role in industry applications. The optimum temperature and pH for the enzyme were 35°C and pH 7, respectively.
Amylase activity was recorded at different temperature value from lowest to highest such as at 30, 40 and 35°C, respectively (Figure 4). Therefore, the species identified from this range of temperature was Pseudomonas genera because the availability or growth favors Pseudomonas species with almost all surviving at 35 - 40°C. This is almost similar to the suggestion of Wang et al. (2016) in which the enzyme production reached maximum at temperature of 30°C, pH 7, with 40 g/L starch in the medium inoculated with 1.4% v/v spore.

792 Afr. J. Microbiol. Res.
Table 4. Starch degrading index of isolated colonies (SDI).
Isolate code Diameter of colony (cm) Diameter of clear zone (cm) SDI of RB
AJUMC1 1.5 2.9 1.933
AJUMC2 1.3 2.7 2.07
AJUMC3 1.6 2.5 1.56
AJUMC4 1.4 3.0 2.14
AJUMC5 1.6 2.8 1.75
AJUMC6 1 1.8 1.8
AJUMC7 1.8 3.0 1.66
AJUMC8 1.3 1.6 1.23
AJUMC9 1.9 3.3 1.74
AJUMC10 1.6 2.8 1.87
AJUMC11 1.5 2.6 1.73
AJUMC12 1.3 2.8 2.15
AJUMC13 2.4 3.6 1.5
AJUMC14 1.6 2.7 1.69
AJUMC15 1.6 2.5 1.56
AJUMC16 1.3 2.8 2.15
AJUMC17 1.6 2.4 1.5
BJUMC1 2.3 3.5 1.52
BJUMC2 1.75 2.4 1.37
BJUMC3 1.2 2.0 1.66
BJUMC4 1.5 2.8 1.86
BJUMC5 1.0 2.1 2.10
BJUMC6 1.2 1.9 1.58
BJUMC7 1.2 2.25 1.875
BJUMC8 1.7 2.8 1.64
BJUMC9 1.2 1.9 1.58
BJUMC10 0.95 1.7 1.79
CJUMC1 1.5 2.6 1.73
CJUMC2 1.4 2.3 1.64
CJUMC3 1.1 1.9 1.72
CJUMC4 1.2 1.8 1.50
CJUMC5 1.8 2.9 1.61
CJUMC6 1.25 2.3 1.84
CJUMC7 2.5 3.6 1.44
CJUMC8 1.8 3 1.67
CJUMC9 1.2 2.1 1.75
CJUMC10 1.3 2.3 1.77
CJUMC11 1.6 2.9 1.81
DJUMC1 2.5 3.5 1.40
DJUMC2 0.8 1.2 1.50
DJUMC3 0.8 1.4 1.75
DJUMC4 1.7 2.7 1.59
DJUMC5 1.2 2.4 2.00
DJUMC6 1.7 2.5 1.47
DJUMC7 1.1 1.9 1.72
DJUMC8 0.9 1.8 2.00
DJUMC9 1.8 2.6 1.44
DJUMC10 0.84 1.3 1.57
DJUMC11 2.1 3.0 1.4
DJUMC12 1.4 2.3 1.64
DJUMC13 1.6 2.6 1.625
DJUMC14 1.2 1.6 1.33
DJUMC15 1.40 2.00 1.43

Dida 793
Figure 2. Amylase activity analysis on Petri-dish.
Table 5. Determination of different parameter.
Isolate code pH (7) Temperature (35°C) Starch con. (1.5%) Incubation period (24 h)
AJUMC + + + +
BJUMC + + + +
CJUMC + + + +
DJUMC + + + +
Figure 3. Amylase activity at different pH.
Conclusion The present study reveals that all four sites soil samples taken from Jimma University plant root were inhabited
with diverse microorganisms mostly of Bacillus and Pseudomonas genera with high potential to degrade starch, thereby degrading starch blended material (polymers) to avoid pollution from the environment. The
0
0.2
0.4
0.6
0.8
1
1.2
PH5 PH6 pH7 pH8 PH9
Am
yla
se
ac
tiv
ity
(U
/ml)
pH
5 6 7 8 9
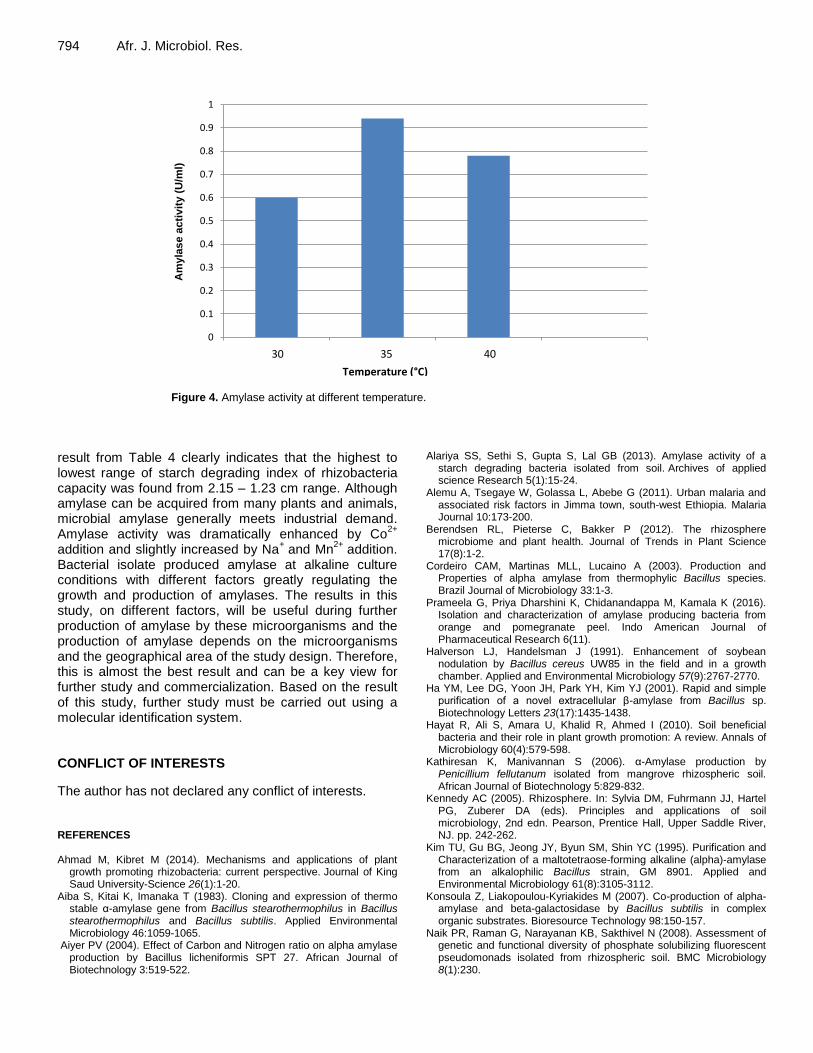
794 Afr. J. Microbiol. Res.
Figure 4. Amylase activity at different temperature.
result from Table 4 clearly indicates that the highest to lowest range of starch degrading index of rhizobacteria capacity was found from 2.15 – 1.23 cm range. Although amylase can be acquired from many plants and animals, microbial amylase generally meets industrial demand. Amylase activity was dramatically enhanced by Co
2+
addition and slightly increased by Na+ and Mn
2+ addition.
Bacterial isolate produced amylase at alkaline culture conditions with different factors greatly regulating the growth and production of amylases. The results in this study, on different factors, will be useful during further production of amylase by these microorganisms and the production of amylase depends on the microorganisms and the geographical area of the study design. Therefore, this is almost the best result and can be a key view for further study and commercialization. Based on the result of this study, further study must be carried out using a molecular identification system. CONFLICT OF INTERESTS
The author has not declared any conflict of interests. REFERENCES Ahmad M, Kibret M (2014). Mechanisms and applications of plant
growth promoting rhizobacteria: current perspective. Journal of King Saud University-Science 26(1):1-20.
Aiba S, Kitai K, Imanaka T (1983). Cloning and expression of thermo stable α-amylase gene from Bacillus stearothermophilus in Bacillus stearothermophilus and Bacillus subtilis. Applied Environmental Microbiology 46:1059-1065.
Aiyer PV (2004). Effect of Carbon and Nitrogen ratio on alpha amylase production by Bacillus licheniformis SPT 27. African Journal of Biotechnology 3:519-522.
Alariya SS, Sethi S, Gupta S, Lal GB (2013). Amylase activity of a starch degrading bacteria isolated from soil. Archives of applied science Research 5(1):15-24.
Alemu A, Tsegaye W, Golassa L, Abebe G (2011). Urban malaria and associated risk factors in Jimma town, south-west Ethiopia. Malaria Journal 10:173-200.
Berendsen RL, Pieterse C, Bakker P (2012). The rhizosphere microbiome and plant health. Journal of Trends in Plant Science 17(8):1-2.
Cordeiro CAM, Martinas MLL, Lucaino A (2003). Production and Properties of alpha amylase from thermophylic Bacillus species. Brazil Journal of Microbiology 33:1-3.
Prameela G, Priya Dharshini K, Chidanandappa M, Kamala K (2016). Isolation and characterization of amylase producing bacteria from orange and pomegranate peel. Indo American Journal of Pharmaceutical Research 6(11).
Halverson LJ, Handelsman J (1991). Enhancement of soybean nodulation by Bacillus cereus UW85 in the field and in a growth chamber. Applied and Environmental Microbiology 57(9):2767-2770.
Ha YM, Lee DG, Yoon JH, Park YH, Kim YJ (2001). Rapid and simple purification of a novel extracellular β-amylase from Bacillus sp. Biotechnology Letters 23(17):1435-1438.
Hayat R, Ali S, Amara U, Khalid R, Ahmed I (2010). Soil beneficial bacteria and their role in plant growth promotion: A review. Annals of Microbiology 60(4):579-598.
Kathiresan K, Manivannan S (2006). α-Amylase production by Penicillium fellutanum isolated from mangrove rhizospheric soil. African Journal of Biotechnology 5:829-832.
Kennedy AC (2005). Rhizosphere. In: Sylvia DM, Fuhrmann JJ, Hartel PG, Zuberer DA (eds). Principles and applications of soil microbiology, 2nd edn. Pearson, Prentice Hall, Upper Saddle River, NJ. pp. 242-262.
Kim TU, Gu BG, Jeong JY, Byun SM, Shin YC (1995). Purification and Characterization of a maltotetraose-forming alkaline (alpha)-amylase from an alkalophilic Bacillus strain, GM 8901. Applied and Environmental Microbiology 61(8):3105-3112.
Konsoula Z, Liakopoulou-Kyriakides M (2007). Co-production of alpha-amylase and beta-galactosidase by Bacillus subtilis in complex organic substrates. Bioresource Technology 98:150-157.
Naik PR, Raman G, Narayanan KB, Sakthivel N (2008). Assessment of genetic and functional diversity of phosphate solubilizing fluorescent pseudomonads isolated from rhizospheric soil. BMC Microbiology 8(1):230.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
temp 30°C temp 35°C temp 40°C
Am
yla
se a
cti
vit
y (
U/m
l)
Temperature (°C)
30 35 40

Nusrat A, Rahman SR (2007). Comparative studies on the production of
extracellular α-amylase by three mesophilic Bacillus isolates. Bangladesh Journal of Microbiology 24(2):129-132.
Oseni O, Ekperigin M (2013). Isolation and activity of alpha amylase from selected bacteria strains in the forest soil. Global Journal of Bioscience and Technology 2(1):17-18.
Parmar N Dadarwal KR (1999). Stimulation of nitrogen fixation and induction of flavonoids like compounds by rhizobacteria. Journal of applied Microbiology 86:3-44.
Sasmita M, Niranjan B (2008). Amylase activity of a starch degrading bacteria isolated from soil receiving kitchen wastes. African Journal of Biotechnology 7(18): 3326-3331.
Silva PD, Nahas E (2002). Bacterial diversity in soil in response to different plans, phosphate fertilizers and liming. Brazilian Journal of Microbiology 33(4):304-310.
Smits JP, Rinzema J, Tramper H, Van M, Knol W (1996). Solid-state fermentation of wheat bran by Trichoderma reesei QM9414: substrate composition changes, C balance, enzyme production, growth and kinetics. Applied Microbial Biotechnology 46:489-496.
Dida 795 Tonkova A, Manolov R, Dobreva E (1993). Thermo stable α-amylase
from derepressed Bacillus lichenform is produced in high yields from glucose. Process Biochemistry 28:539-542.
Wang S, Lin C, Liu Y, Shen Z, Jeyaseelan J and Qin W (2016). Characterization of a starch-hydrolyzing α-amylase produced by Aspergillus niger WLB42 mutated by ethyl methane sulfonate treatment. International journal of biochemistry and molecular biology 7(1):1.
Williams CM, Richter CS, Mackenzie JM, Shih JC (1990). Isolation, identification and characterization of a feather-degrading bacterium. Applied and environmental microbiology 56(6):1509-1515.

Journal of
Microbiology and Antimicrobials
Journal of Microbiology and Antimicrobials
International Journal of
Biotechnology and Molecular
Biology Research
African Journal of
Microbiology Research
African Journal of
Microbiology Research
www.academicjournals.org
African Journal of
Biochemistry Research
African Journal of
Biochemistry Research
Journal of
Bioinformatics and Sequence Analysis
Journal of
Biophysics and Structural Biology


















