
1 структура ак цен ферментов герман
29
Ферменты. Структура активного центра в ферментах (Часть 2-ая) К.Э. Герман Зав.кафедрой естественнонаучных дисциплин МИ РЕАВИЗ
-
Upload
konstantin-german -
Category
Documents
-
view
98 -
download
1
Transcript of 1 структура ак цен ферментов герман

Ферменты.Структура активного центра
в ферментах
(Часть 2-ая)
К.Э. ГерманЗав.кафедрой естественнонаучных дисциплин
МИ РЕАВИЗ
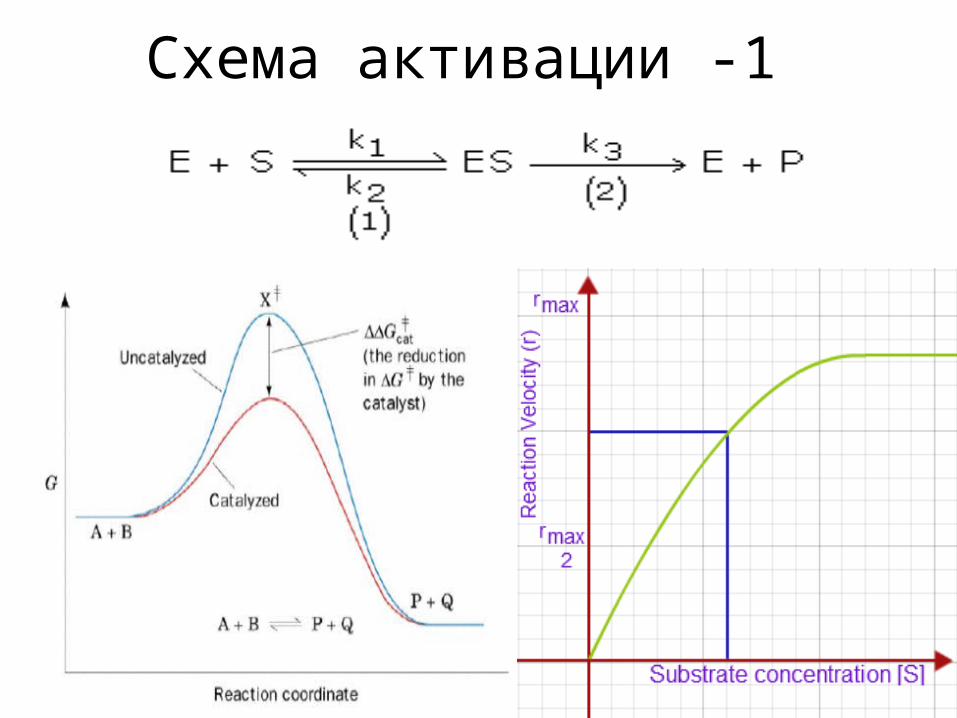
Схема активации -1
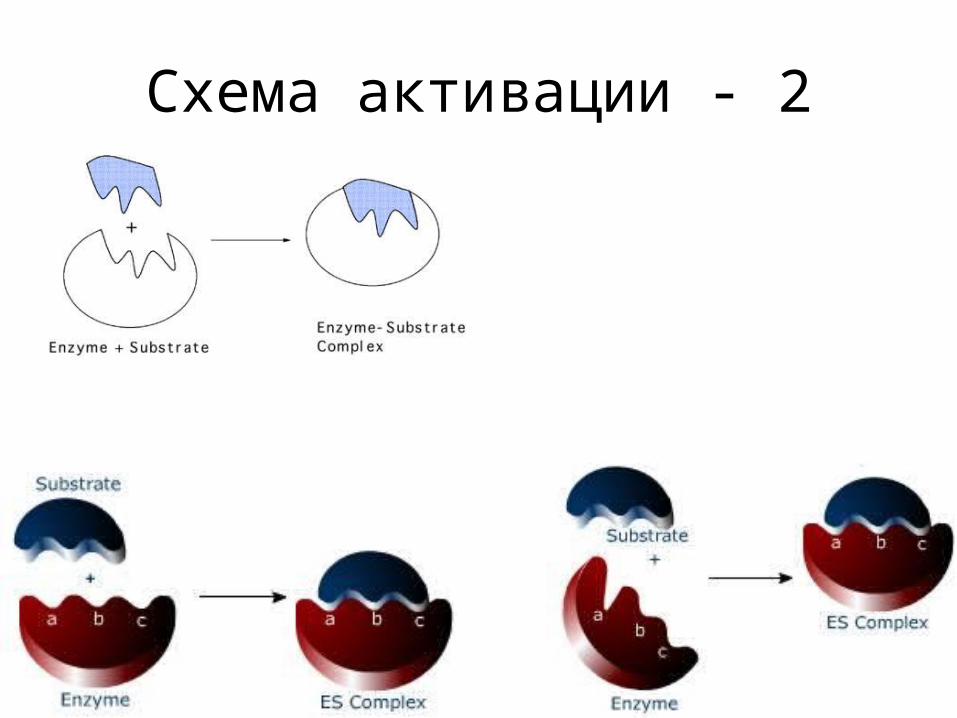
Схема активации - 2

Структуры ферментов
• http://www.rcsb.org/pdb/explore/explore.do?structureId=1GOF PDB
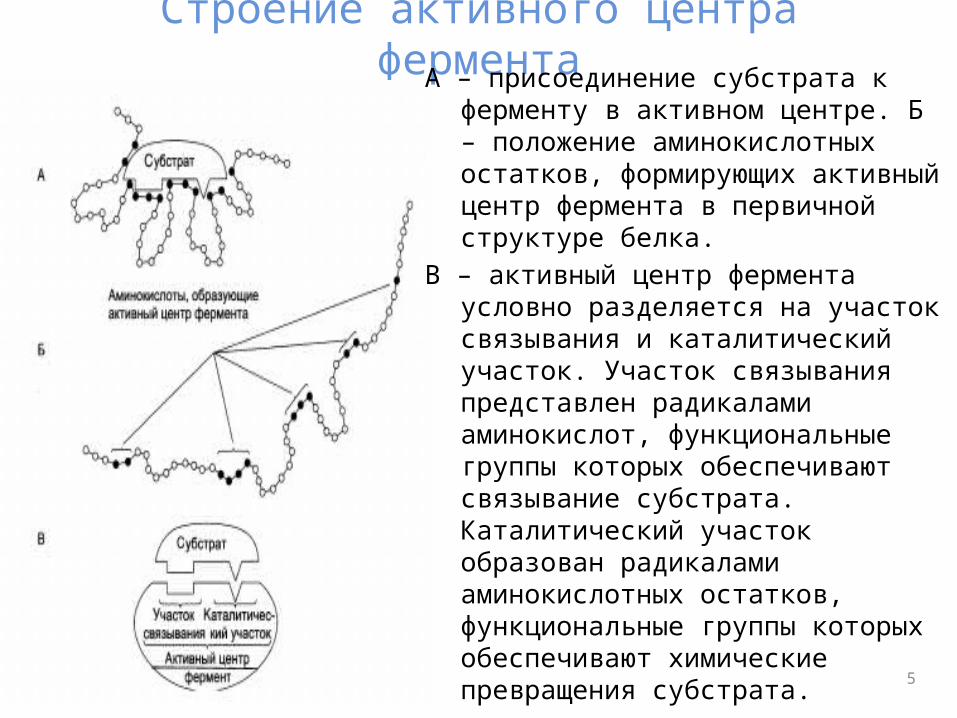
Строение активного центра ферментаА – присоединение субстрата к ферменту в
активном центре. Б – положение аминокислотных остатков, формирующих активный центр фермента в первичной структуре белка.
В – активный центр фермента условно разделяется на участок связывания и каталитический участок. Участок связывания представлен радикалами аминокислот, функциональные группы которых обеспечивают связывание субстрата. Каталитический участок образован радикалами аминокислотных остатков, функциональные группы которых обеспечивают химические превращения субстрата.
5

6
Активный центр ферментовИзучение механизма химической реакции, катализируемой ферментом наряду с определением промежуточных и конечных продуктов на разных стадиях реакции подразумевает точное знание геометрии третичной структуры фермента, природы функциональных групп его молекулы, обеспечивающих специфичность действия и высокую каталитическую активность на данный субстрат, а также химической природы участка (участков) молекулы фермента, который обеспечивает высокую скорость каталитической реакции. Обычно молекулы субстрата, участвующие в ферментативных реакциях, по сравнению с молекулами ферментов имеют относительно небольшие размеры. Таким образом, при образовании фермент-субстратных комплексов в непосредственное химическое взаимодействие вступают лишь ограниченные фрагменты аминокислотной последовательности полипептидной цепи — «активный центр» — уникальная комбинация остатков аминокислот в молекуле фермента, обеспечивающая непосредственное взаимодействие с молекулой субстрата и прямое участие в акте катализа[10].
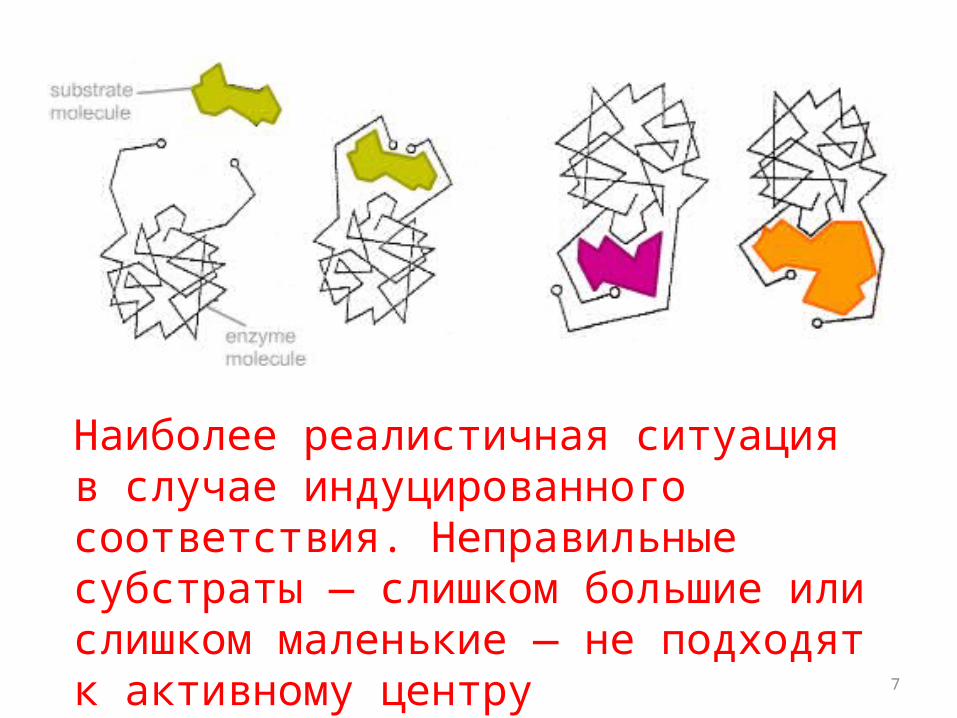
7
Наиболее реалистичная ситуация в случае индуцированного соответствия. Неправильные субстраты — слишком большие или слишком маленькие — не подходят к активному центру

В PDB много других примеров сериновых протеаз, построенные для пищеварения, активизации гормонов,
свертывания крови, активации иммунной системы и многих других функций.
Они входят в необычную коллекцию производных аминокислот, предназначенных для оказания помощи в
реакции резки белков, которые были подобраны природой снова и снова в процессе эволюции

Серин в ферментах
Центром этой «машины» является аминокислота серин, которая активируется с помощью гистидина и аспартата.
Вместе эти три аминокислоты были названы «системой передачи заряда).
Гистидин и аспарат помогают в удалении атома водорода от серина (белого цвета), что делает его более реакционноспособным при атаке на цепь целевого белка.

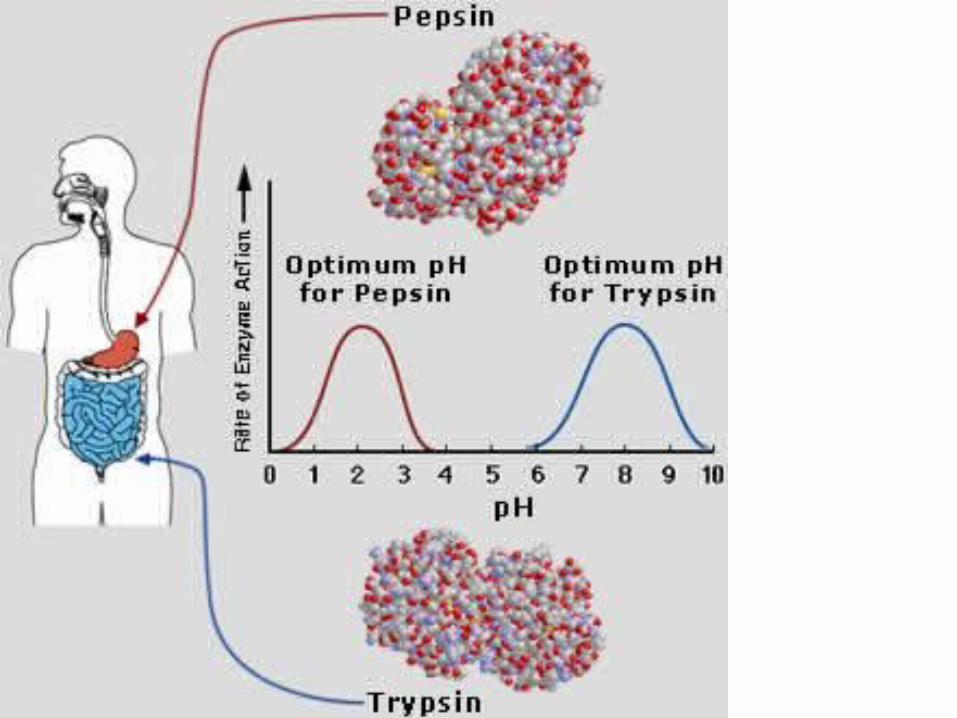
Трипсины ряда животных получены в кристаллическом виде (впервые в 1932). Молекула бычьего трипсина (молекулярная масса около 24 кДа) состоит из 223 аминокислотных остатков, образующих одну полипептидную цепь, и содержит 6 дисульфидных связей. Изоэлектрическая точка трипсина лежит при pH 10,8, а оптимум каталитической активности — при pH 7,8—8,0.
Трипсины относятся к группе сериновых протеаз и содержат в активном центре отатки серина и гистидина.
Пепсин относятся к
Трипсины легко подвергаются самоперевариванию (аутолизу), что приводит к загрязнению препаратов трипсинов неактивными продуктами
Пепсин и Трипсин — ферменты класса гидролаз, расщепляющие пептиды и белки
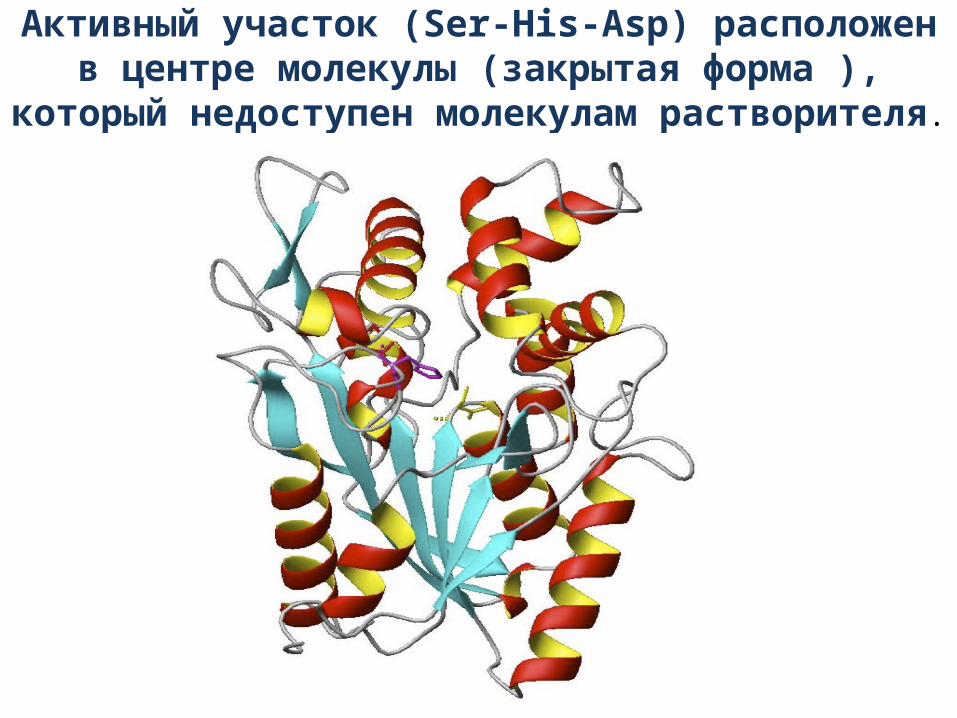
Активный участок (Ser-His-Asp) расположен в центре молекулы (закрытая форма ), который недоступен
молекулам растворителя.

Система фиксации инородного пептида в якорном участке фермента (по Д. Дефо )
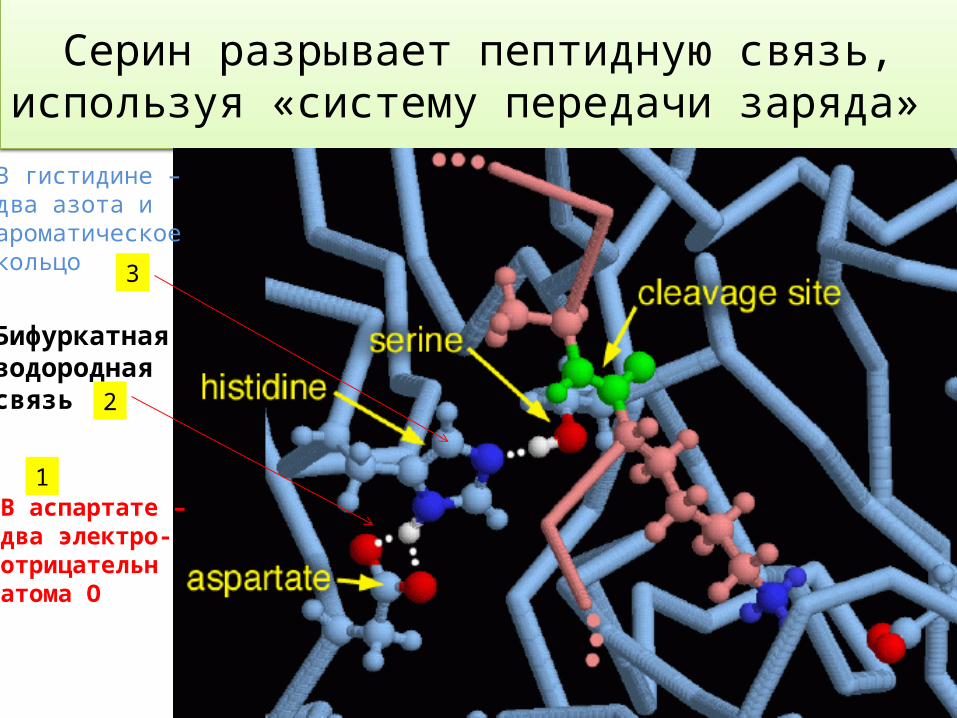
Серин разрывает пептидную связь, используя «систему передачи заряда»
Бифуркатная водороднаясвязь
В гистидине –два азота иароматическоекольцо
В аспартате – два электро-отрицательнатома О
1
2
3
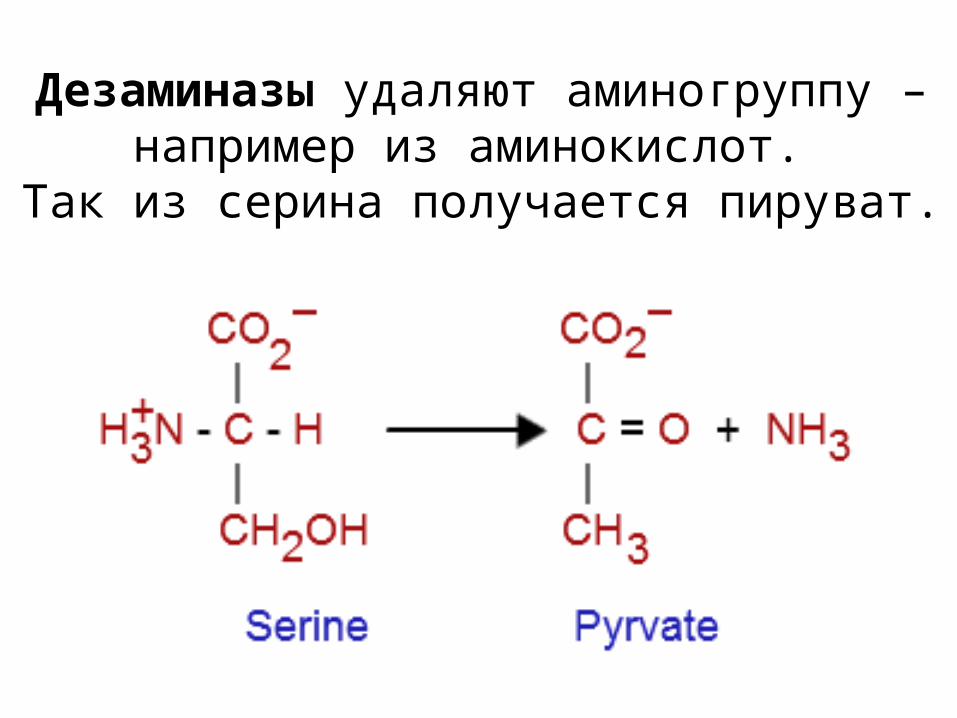
Дезаминазы удаляют аминогруппу – например из аминокислот.
Так из серина получается пируват.
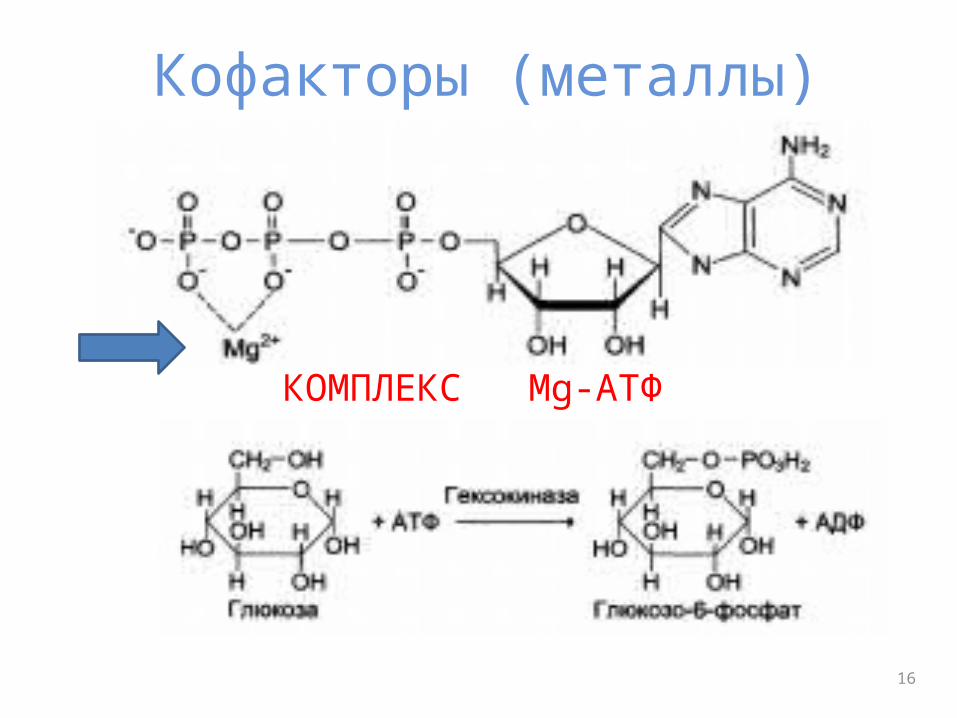
Кофакторы (металлы)
16
КОМПЛЕКС Mg-АТФ
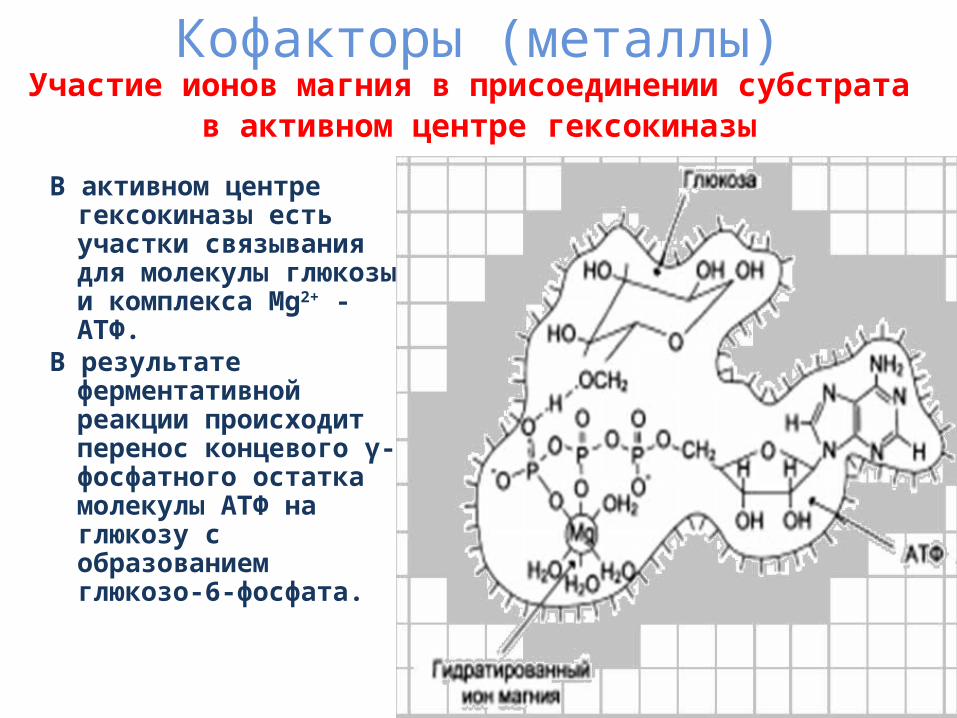
Кофакторы (металлы)Участие ионов магния в присоединении субстрата
в активном центре гексокиназы
17
В активном центре гексокиназы есть участки связывания для молекулы глюкозы и комплекса Mg2+ -АТФ.
В результате ферментативной реакции происходит перенос концевого γ-фосфатного остатка молекулы АТФ на глюкозу с образованием глюкозо-6-фосфата.
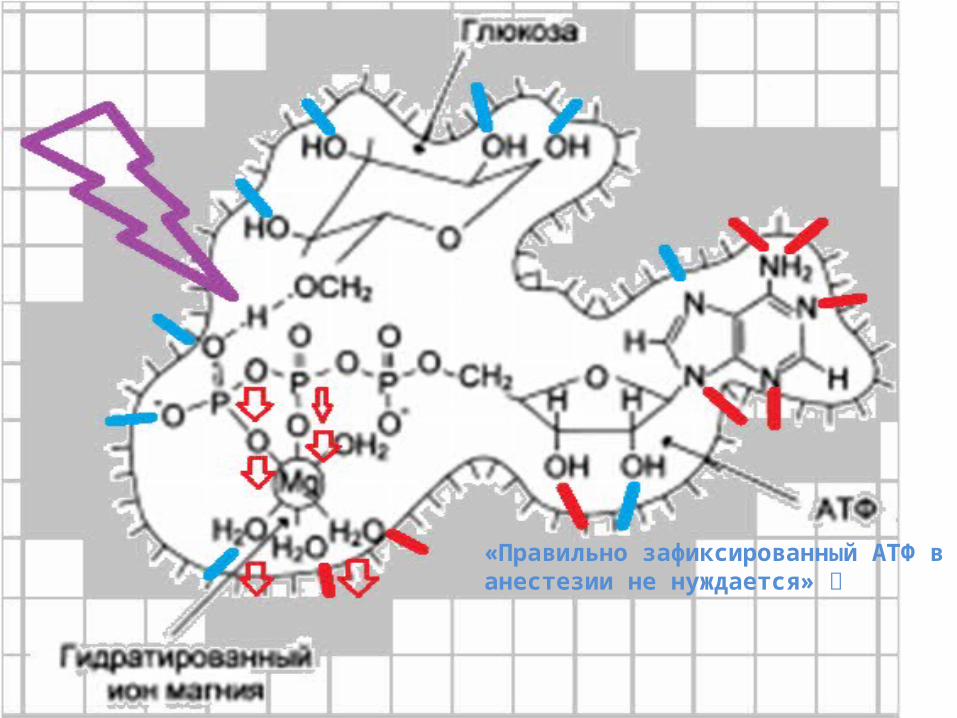
«Правильно зафиксированный АТФ ванестезии не нуждается»
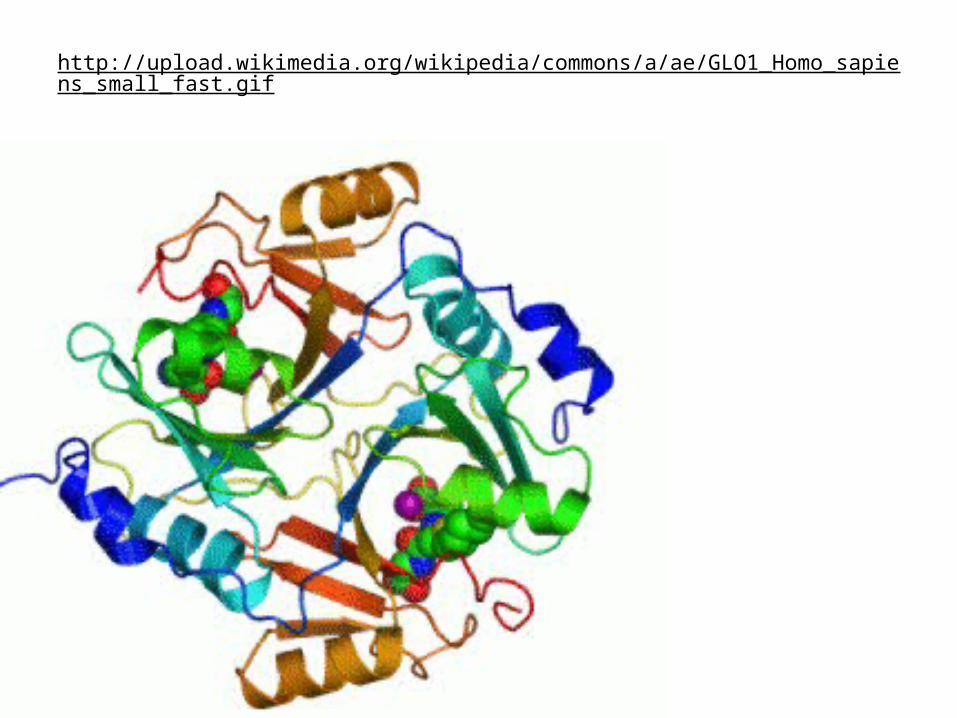
http://upload.wikimedia.org/wikipedia/commons/a/ae/GLO1_Homo_sapiens_small_fast.gif

Фермент активируется после димеризации. Кальций играет важную роль в катализе. Каждый из двух ионов кальция (зеленые шарики) связаны как OMPLA мономеров, таких, что отдельные молекулы OMPLA не может связать кальций эффективно. Два субстрата аналогов приведены расширения вниз от ионов кальция. Эти аналоги зажатой между двумя OMPLA monomers.до сих пор неизвестно, что вызывает образование активного OMPLA димера. Скорее всего мембрана возмущение играет важную роль. Следует отметить, что активный центр ориентирован по направлению к внешней грани наружной мембраны.
Это фосфолипаза OMPLA (белок бета-баррель) встроен в
наружную мембрану, содержащую в основном липополисахариды
(ЛПС) на наружной поверхности и фосфолипиды на внутренней
поверхности.
http://mbb.science.uu.nl/research/researchofegmond.html
Outer membrane phospholipase A1 (OMPLA)
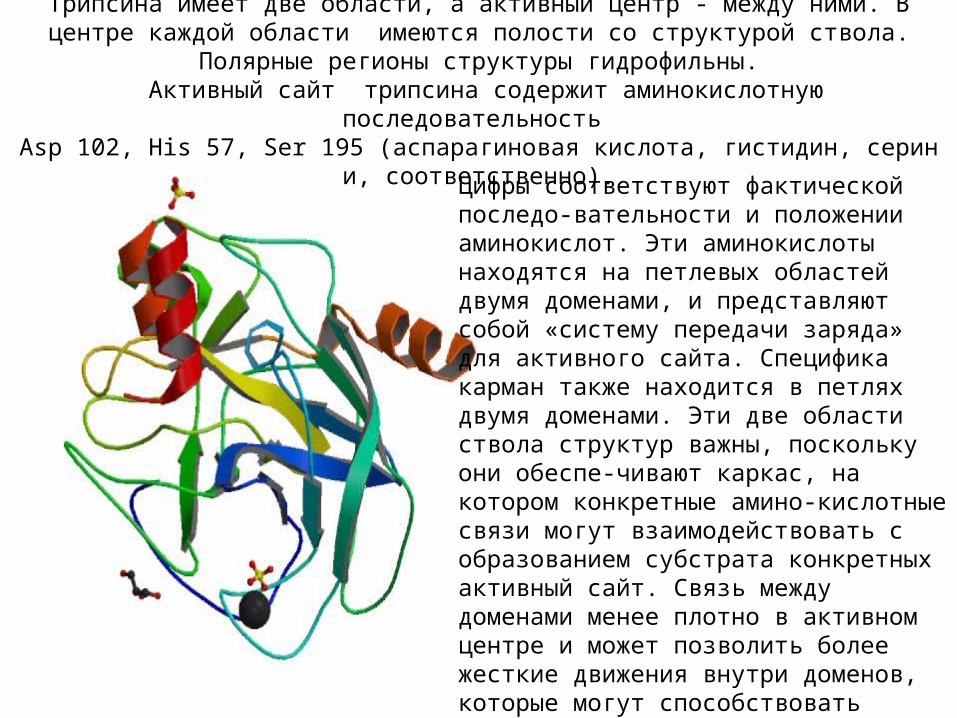
Цифры соответствуют фактической последо-вательности и положении аминокислот. Эти аминокислоты находятся на петлевых областей двумя доменами, и представляют собой «систему передачи заряда» для активного сайта. Специфика карман также находится в петлях двумя доменами. Эти две области ствола структур важны, поскольку они обеспе-чивают каркас, на котором конкретные амино-кислотные связи могут взаимодействовать с образованием субстрата конкретных активный сайт. Связь между доменами менее плотно в активном центре и может позволить более жесткие движения внутри доменов, которые могут способствовать катализа. Эти жесткие движения тела фермента являются основной причиной его каталитического действия.
Фермент : трипсин - в семье серин-протеазы. Активный сайт Трипсина имеет две области, а активный центр - между ними. В центре каждой области имеются полости со структурой
ствола. Полярные регионы структуры гидрофильны. Активный сайт трипсина содержит аминокислотную последовательность
Asp 102, His 57, Ser 195 (аспарагиновая кислота, гистидин, серин и, соответственно).

Охота на вирус СПИДаA Handy EnzymeThis ribbon representation of the RT active domain illustrates its hand-like structure, showing fingers (blue), palm (pink) and thumb (green). The active site (red atoms), where DNA is elongated, is in the palm region. Also shown is an RT-inhibitor drug (yellow) in the pocket where it binds.
http://www.psc.edu/science/2000/madrid/getting_a_grip_on_aids.html
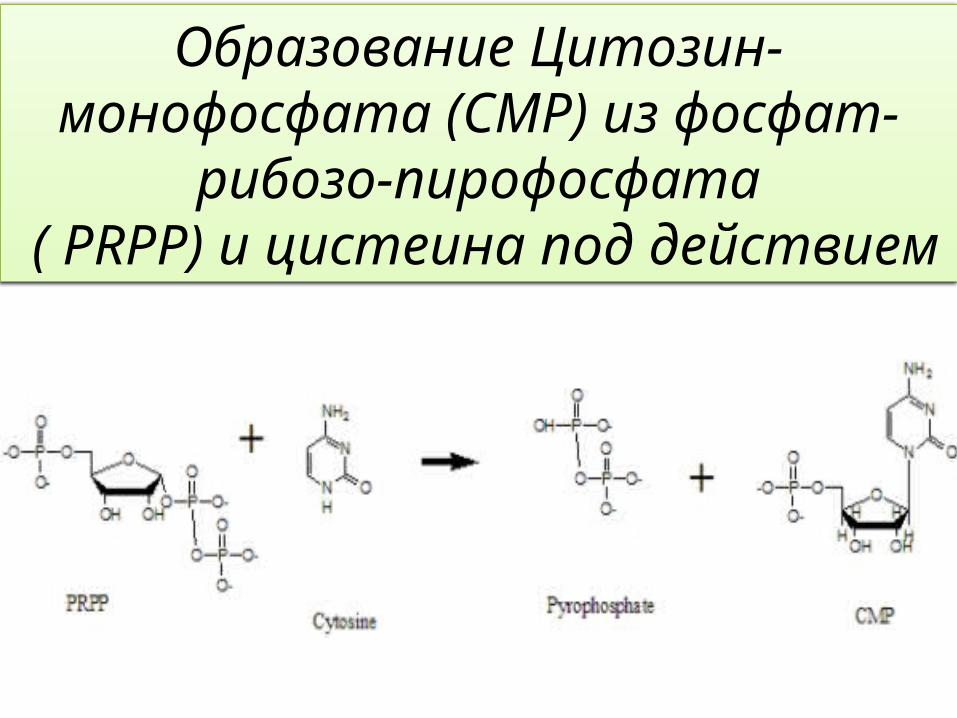
Образование Цитозин-монофосфата (CMP) из фосфат-
рибозо-пирофосфата ( PRPP) и цистеина под действием

Adenine Phosphoribosyltransferase that catalyzes the formation of AMP
from Adenine and PRPP. Это симметричный гомодимер. Крышка-капот (hood)состоит из альфа 1, 2 и бета 1, 2;Ядро состоит из остальных альфа-спиралей и бета-лент. При фиксации каталитического остатка на петле в течение активного кармана на площадке, цикл будет закрыт, когда субстрат связывается с активным сайтом. Этот фермент действует совместно с 9-deazaadenosine (9DA), Mg-ион, ФРПФ
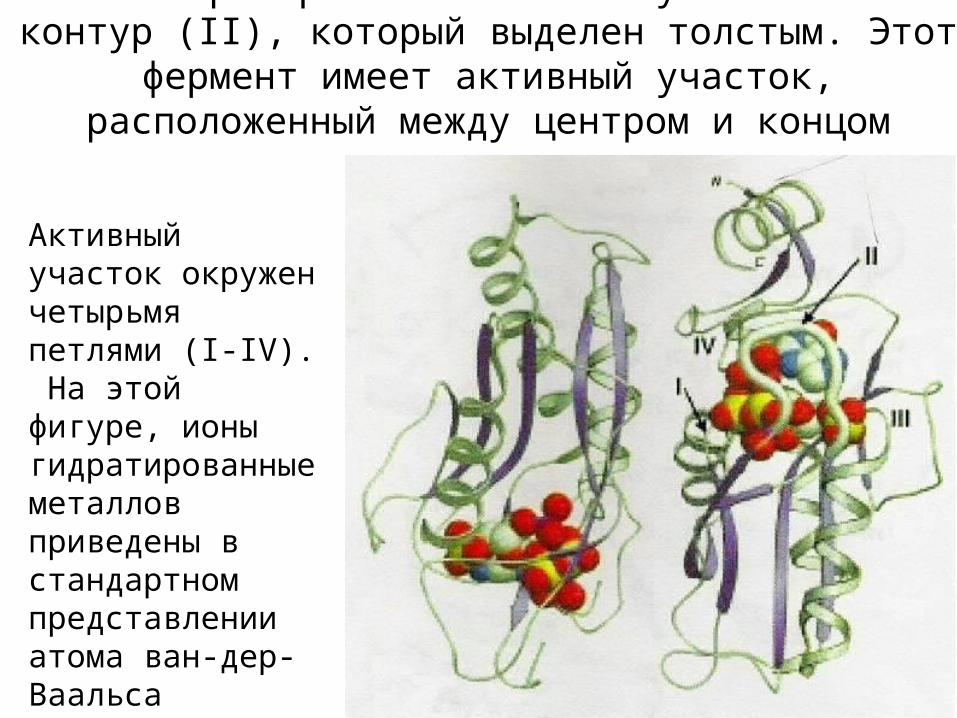
Мономер справа имеет замкнутый гибкий контур (II), который выделен толстым. Этот фермент имеет активный
участок, расположенный между центром и концом домена
Активный участок окружен четырьмя петлями (I-IV). На этой фигуре, ионы гидратированные металлов приведены в стандартном представлении атома ван-дер-Ваальса
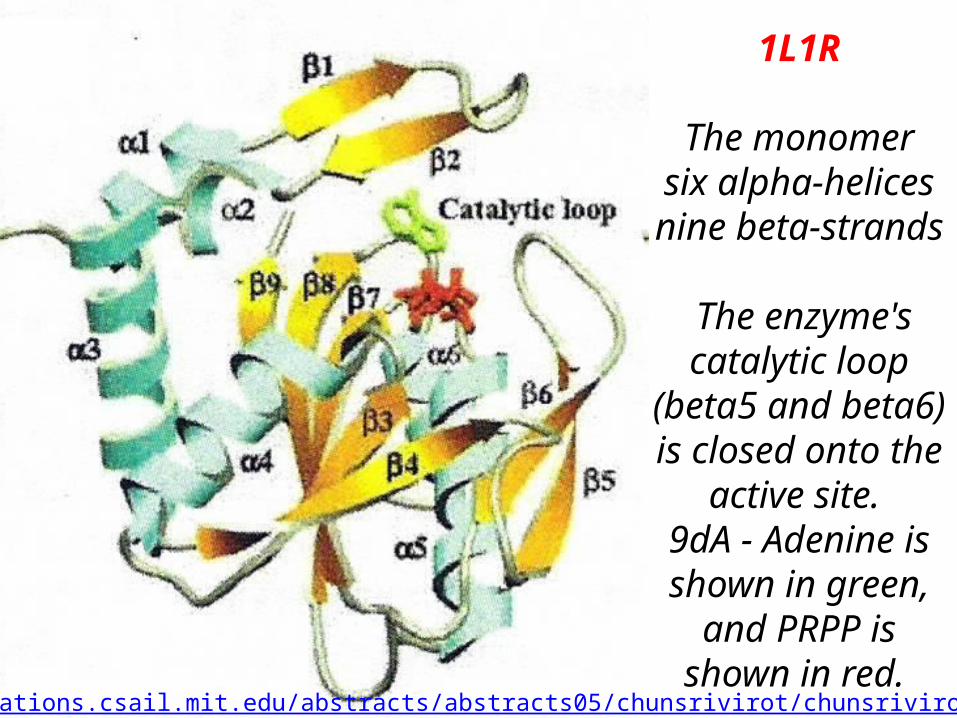
1L1R
The monomer six alpha-helices nine beta-strands
The enzyme's catalytic loop
(beta5 and beta6) is closed onto the
active site. 9dA - Adenine is shown in green,
and PRPP is shown in red.
http://publications.csail.mit.edu/abstracts/abstracts05/chunsrivirot/chunsrivirot.html

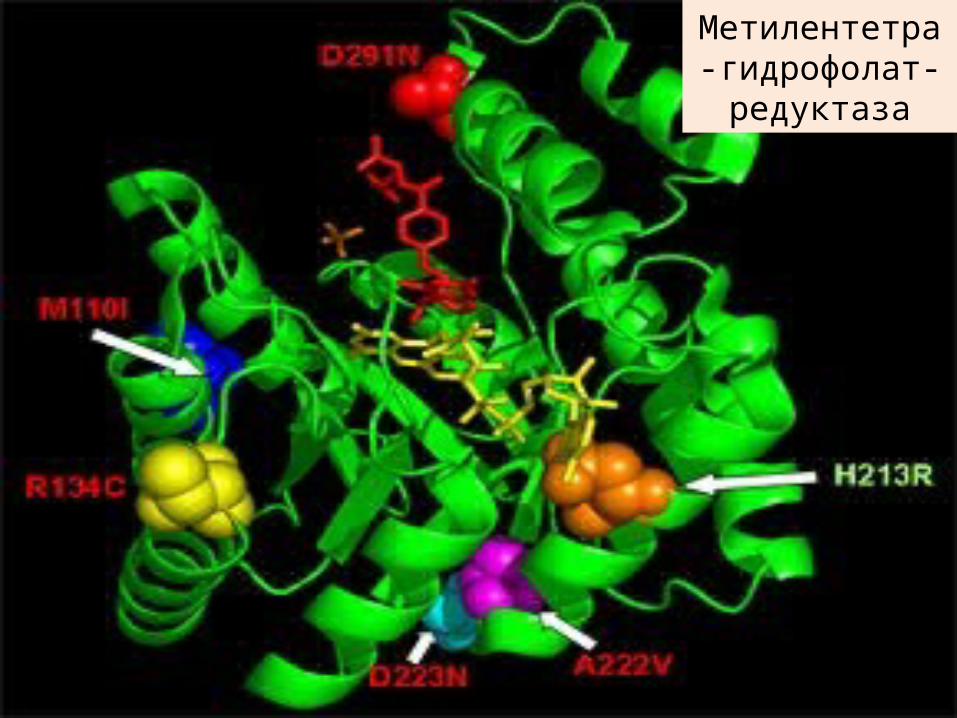
Ферментметилентетрагидрофолатредуктаза
Структура активного сайта фермента метилентетрагидрофолатредуктазы бактерий
(E.coli), который аналогичен по структуре дрожжевого фермента MTHFR.
Пять-сайтов, помеченных красным цветом - места, где общие мутации происходят влияния на
активность фермента; мутации в зеленой меченных сайта не повлиять на
деятельность фермента. Geoff Хорнер
Метилентетра-гидрофолат-
редуктаза
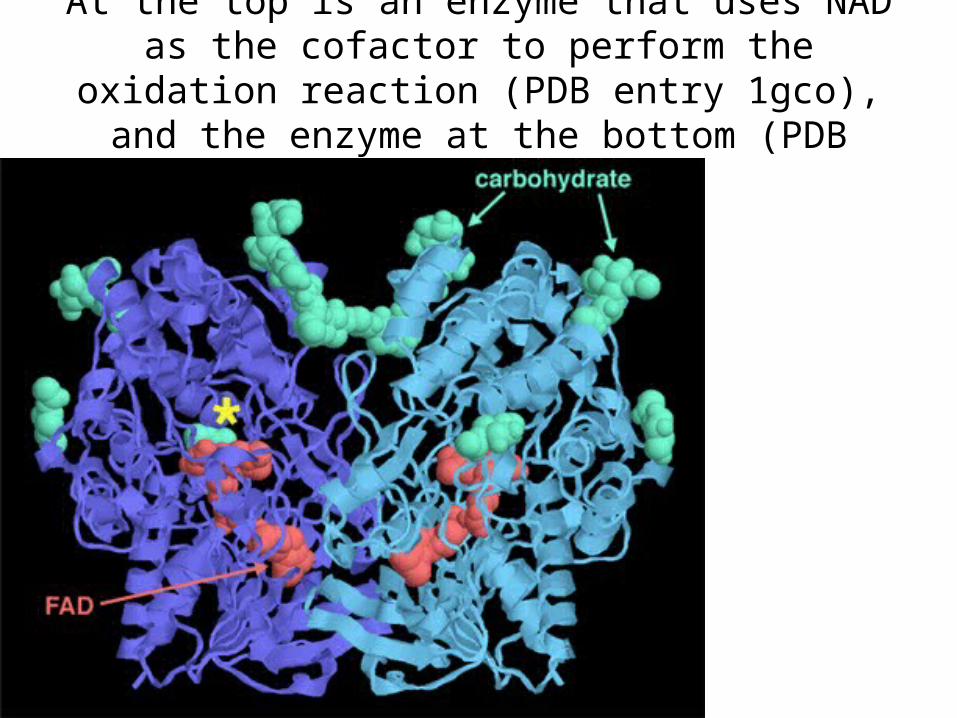
At the top is an enzyme that uses NAD as the cofactor to perform the oxidation reaction (PDB entry 1gco), and
the enzyme at the bottom (PDB entry 1cq1) uses ...


















