
第十章 蛋白质的生物合成
46
第第第 第第第第第第第第
description
第十章 蛋白质的生物合成. 蛋白质合成体系的组分. 第一节 蛋白质合成体系的重要组分. mRNA ---- 模板 rRNA ---- 构成核糖体作为蛋白质合成场所 tRNA ---- 搬运工具 氨基酸 ---- 原料 蛋白质因子 ( 起始因子 IF ,延伸因子 EF ,释放因子 RF ,等等 ) 酶 ---- 氨酰 -tRNA 合成酶,转肽酶,转位酶等 无机离子 ----Mg ++. 一、 mRNA 和遗传密码. mRNA 特点:短命 原核:半衰期几秒 — 几分 真核:半衰期数小时。 - PowerPoint PPT Presentation
Transcript of 第十章 蛋白质的生物合成

第十章蛋白质的生物合成

mRNA---- 模板rRNA---- 构成核糖体作为蛋白质合成场所tRNA---- 搬运工具氨基酸 ---- 原料蛋白质因子 ( 起始因子 IF ,延伸因子 EF ,释放因子 R
F ,等等 )酶 ---- 氨酰 -tRNA 合成酶,转肽酶,转位酶等无机离子 ----Mg++
第一节 蛋白质合成体系的重要组分

mRNA 特点:短命 原核:半衰期几秒—几分 真核:半衰期数小时。
mRNA 功能是 : 蛋白质合成的模板,多肽链 AA排列顺序就取决于 mRNA 的核苷酸的排列顺序。
mRNA 为模板的蛋白质合成过程被称为翻译或转译。
一、一、 mRNAmRNA 和遗传密码和遗传密码

mRNA 分子中四种不同碱基( A 、 G 、 C 和U )构成特定顺序决定蛋白质分子中 20 种 AA所构成的序列。
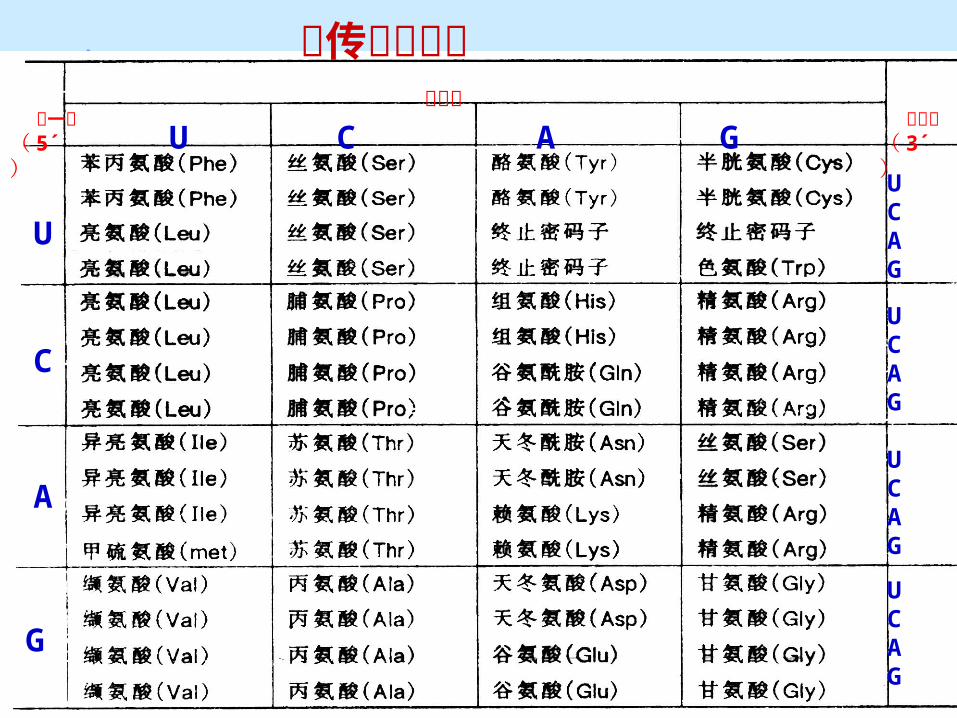
mRNA 上每 3 个相邻的核苷酸编码蛋白质多肽链中的一个氨基酸,这三个核苷酸就称为一个密码子或三联体密码。
四种核苷酸,能有 43 =64 组密码子。 20 种基本氨基酸对应 61 个密码子 , 一个密码子作为起始密码子 , 三个终止密码子。
(二)遗传密码(二)遗传密码
U
A
C
G
UCAG
U C A G第二位
第一位( 5ˊ )
第三位( 3ˊ )
UCAG
UCAG
UCAG
遗传密码字典

⑴ 密码子的方向性 阅读方向为 5-3’ ,与 mRNA 链合成方向相同。
⑵ 密码子的简并性 由一种以上密码子编码同一个氨基酸的现象称为简
并性( dogeneracy ) . 64-3=61 个代表示 20 种氨基酸,仅甲硫氨酸、色氨
酸只有一个密码子。 编码同一个氨基酸的一组密码子称为同义密码子。 密码的简并性可以减少有害突变 。
遗传密码的特点

⑶ 密码子的连续性(读码) 从正确起点开始至终止信号,密码子的排列是连续的。既不存在间隔(无标点),也无重叠。 在 mRNA 分子上插入或删去一个碱基,会使该点以后的读码发生改变,称为移码,由这种情况引起的突变称为移码突变。
3’
起始密码子
5’

⑷ 密码子的基本通用性(近于完全通用)
对于高等、低等生物都适用。
只有一个例外:真核生物线粒体 DNA 。
另一些原核生物中利用终止密码翻译 AA(如 UGA-Trp\ 硒代半胱氨酸)

人线粒体中变异的密码子
UGA 终止信号 Trp
AUA Ile Met
AGA Arg 终止信号
AGG Arg 终止信号
密码子 正常情况下编码 线粒体 DNA 编码

⑸ 起始密码子和终止密码子
起始密码子: AUG (少数 GUG )→原核:甲酰蛋氨酸 真核:蛋氨酸
终止密码子: UAA 、 UAG 、 UGA→ 终止或无意义密码子,不编码任何氨基酸。

⑹ 密码子的摆动性(变偶性)
如丙氨酸: GCU , GCC , GCA , GCG ,只第三位不同 ,显然密码子的专一性基本取决于前两位碱基,第三位碱基有较大灵活性。
tRNA 上的反密码子与 mRNA 上的密码子配对时,密码子的第一位、第二位碱基配对是严格的,第三位碱基可以有一定变动,这种现象称为密码的摆动性或变偶性。

反密码子与密码子之间的碱基配对
A U
C G
反密码子第一位碱基 密码子第三位碱基
GU
C
UA
G
I
U
C
A

• 为每个三联体密码子译成氨基酸提供接合体,准确无误地将活化的氨基酸运送到核糖体中 mRNA 模板上。
• 同功受体 tRNA: 一种氨基酸可以有多种 tRNA 作为运载工具。把携带相同氨基酸而反密码子不同的一组 tRNA 称为同功受体 tRNA
反密码子 : tRNA 分子上三个特定的碱基组成一个反密码子,位于反密码子环上。
二、二、 tRNAtRNA

tRNA 的接头 (adaptor)作用 3´-端上的氨基酸接受位点 :3’端 CCA, 接
受氨基酸,形成氨酰 -tRNA 。需 ATP提供活化氨基酸所需的能量
识别氨酰 - tRNA 合成酶的位点 :倒 L中部的 DHU臂和反密码子环,氨基酸臂参与这一作用
核糖体识别位点 :倒 L中部的 TψC环 反密码子位点 : 与 mRNA结合部位

反密码子
反密码子环
合成酶
aa 位点
可变环
D 臂
TψC 环
TψC 臂

3’
5’
I
CCA-OH
5’ 3’
CCA-OH
G G C
C C G
密码子与反密码子的阅读方向均为 5‘ 3’,两者反向平行配对。

•tRNA 的表示方法:•tRNACys : 右上角标上所转运的氨基酸
•虽然蛋氨酸仅一组密码子( AUG ),却至少有两种 tRNA
•原核 tRNAMet— 将Met 运到肽链中间 tRNAffMet—携带甲酰蛋氨酰参于蛋白质
合成的起始。•真核 tRNAMet— 将Met 运到肽链中间 tRNAiMet—携带Met参于蛋白质合成的
起始。

三、核 糖 体1 、核糖体:是由几十种蛋白质和几种 rRNA 组成的亚细胞颗粒 ,其中蛋白质与 rRNA 的重量比约为 1:2 。
蛋白质肽键的合成就是在这种核糖体上进行的。
核糖体的存在形态有三种:单核糖体、核糖体亚基和多核糖体。
真核生物:游离核糖体或与内质网结合 .
原核生物:游离核糖体或与 mRNA 结合成串状的多核糖体
核糖体亚基的聚合与解聚与 Mg2+ 浓度有关

2 、核糖体的组成
34 protein
21 protein
23S RNA 5S RNA
16S RNA
50S subunit
70S ribosome
30S subunit

原核生物核糖体结构示意图
30S subunit
50S subunit

核糖体的大小和 RNA 组成
原核生物
核糖体(S)
亚基( S)
rRNA (S)
真核生物 80
60
40
28
5.8
5
18
5070
30
23
5
16

原核生物核糖体大亚基上有两个与 tRNA结合的位点。各占一个密码子空间: A 位( accepter site ) : 氨酰— tRNA结合部位 P位( peptine site ) : 肽酰— tRNA结合部位
和结合起始的氨酰 -tRNA 大小亚基组成核糖体与 mRNA结合,并按 5’→
3’ 方向沿mRNA 移动,每次移动一个密码子距离,参入一个氨酰— tRNA 。

原核细胞 70S核糖体的 A 位、 P位示意图
30S30S
与 mRNA 结合部位
P位((结合或接受肽基的部位)
A位((结合或接受 AA- tRNA 的部位)
50S50S
5 3mRNA

四、参与蛋白质合成的辅助因子 除 ATP、 GTP、Mg2+外,还有一些蛋白质因子
原核 真核•起始因子 IF1 eIF1-4
IF2 eIF4(A-D)
IF3 eIF5
•延长因子 EF-Tu EF1 (相当于 Tu-Ts) EF-Ts EF2 (相当于 G) EF-G
•终止与释放 RF1 RF2 eRF

IF1: 协助 IF2 、 IF3 起作用IF2:促进氨酰 -tRNA结合在起始密码子上IF3:促进小亚基与 mRNA结合EF-Tu:热不稳定,将氨酰 -tRNA结合在核糖体 A
位点EF-Ts:热稳定,重新生成 EF-Tu-GTPEF-G:依赖于 GTP,又称移位因子(酶)RF1 :识别终止密码子 UAA 和 UAGRF2 :识别终止密码子 UAA 和 UGA

(一)氨基酸的活化在胞液中进行。氨基酸的活化:指各种参加蛋白质合成的 AA 与携带它的相应的 tRNA结合成氨酰 - tRNA 的过程。
活化反应在氨酰 -tRNA 合成酶的催化下进行。消耗两个高能磷酸键。AA 的活化部分是羧基。在氨酰— tRNA 中氨基酸的羧基通过高能酯键连接在 tRNA3ˊ 端 CCA腺苷酸残基 3’-或 2’-羟基上
一、原核生物多肽链的合成过程第二节 蛋白质的合成过程

氨基酸的活化
氨基酸ATP +ATP +
氨酰腺苷酸E-AMPAMP
PPiPPi第一步
AMPAMP
第二步
E
3- 氨酰 -tRNA
AA+ATP+E AA-AMP-E +PPi
Mg 2+
Mn 2+
AA-AMP-E+ tRNA 氨酰-tRNA +AMP+E
P P P
C C A
O C-C-RO H
NH3+
OH

°±»ùËá + tRNA °±»ùõ£-tRNA
°±»ùõ£-tRNAºÏ³Éø
ATP AMP+PPi
氨基酸活化的总反应式:
tRNAIle —— 携带 Ile 的 tRNA Ile- tRNAIle —— 异亮氨酰 -tRNAIle Arg-tRNAarg fMet-tRNAfmet
Met-tRNAimet Met-tRNAmmet
氨基酰 -tRNA 的写法

(二 )肽链合成的起始阶段
1 、起始密码子的识别 小亚基 16s RNA协助辩认起始密码。 SD 序列:距离起始密码子( AUG )上游( 5ˊ-侧)约 10bp处往往有一富含嘌呤的序列,称为 SD 序列。
SD 序列与 16SrRNA 3ˊ端的核苷酸序列形成碱基互补。正是由于这种相互作用,使核糖体能区别起始信号 AUG 与编码肽链中Met 的密码子 AUG ,正确定位于 mRNA 上起始信号的位置。

2 、起始复合物的形成
( 1 )起始氨基酸及起始 tRNA原核生物起始氨基酸是甲酰蛋氨酸 (fMet).
一种专一的甲酰化酶催化Met-tRNAf的甲酰化反应 ,这种酶不能催化游离的Met 或Met-tRNAm 的甲酰化。
N- 甲酰甲硫氨酰 -tRNAiMet 的形成
CHO-HN-CH-COO-tRNA
CH2
CH2
S
COO-
+H2N-CH-COO-tRNA
CH2
CH2
S
COO-Met-tRNAf
Met fMet-tRNAtMet
N10-CHO-FH4 FH4
转甲酰酶

( 2 ) 30S复合物形成:
AUGIF3 IF3
AUG
IF3
GTP、 IF1
、IF2 、 fMet-tRNAf
小亚基
AUG
IF1 、 IF2.GTP
fMet
UAC
5

( 3 ) 70S复合物的形成:
AUG
GTP 、 IF1 、 IF2
fMet
UAC
5
+ 50S核糖体AUG
GTP 、 IF1 、 IF2
fMet
UAC
5P位点
A 位点
GDP+Pi、 IF1 、 IF2
起始: fMet-tRNAf 占据 P位,其反密码恰好与起始密码子 AUG结合。空着 A 位准备接受下一个氨酰 -tRNA 。

延伸循环包括 3步反应,每步都是在相应的蛋白质延伸因子催化下完成的,需要 GTP供能。
1 、进位: 在延伸因子 EF-Tu.GTP帮助下,一个新的氨酰— tRN
A进入 A 位。这个氨酰 -tRNA 的反密码子必须与处于A 位的 mRNA 上的密码子相匹配。
EF—Ts 催化 GDP—GTP交换。重新生成 EF-Tu·GTP
(三)、肽链的延伸
EF-Tu-GDP+ EF-Ts EF-Tu-Ts + GDP
EF-Tu-Ts + GTP EF-Tu-GTP + EF-Ts

Tu\Ts 循环
Ts
Ts-GDP

2 、转肽: 在肽酰转移酶作用下,将 P位的 AA (或肽链)转
移至 A 位氨酰 -tRNA 的氨基酸的氨基上形成肽键 .在 A 位上产生肽酰 -tRNA.
把无负载的 tRNA留在 P位 ,A 位上形成肽键。
5’
AA
APfMet
3’ 3’P A
AA-fMet
5’

O
C
CH
NH2
O
R
O
C
CH
NH2
O
H3CSCH2CH2
O
C
CH
NH
C
CH
O
O
H3CSCH2CH2
R
Aλ Pλ AλPλ
תëÄø
NH2
肽链合成方向 N端→ C端

3 、移位: 在 EF-G 移位酶作用下,核糖体沿 5ˊ-3ˊ方向向前移动一
个密码子,结果使原来在 A 位点上的肽酰 -tRNA又回到了P位,空出 A 位。
原 P位上无负载的 tRNA离开核糖体。需 GTP水解供能。
P A5’ 3’
P A5’ 3’P APP AA

肽链的延长:以上三步反应构成一个延伸循环,肽链每掺入一个 AA 就重复一次延伸循环。
22 3322 33
移位
进位
GTPN-端
22 33
55´́ 33´́
C-端
肽键形成
11 2211 22
肽键形成11
55´́ 33´́
( EF-G )
进位
(Tu\Ts)
GTP


( 四 ) 、肽链合成的终止与释放1 、终止信号的识别:当终止密码子出现在 A 位时,终
止因子 RF 结合在 A 位,肽链合成终止。 RF1 :识别终止密码子 UAA 和 UAG RF2 :识别终止密码子 UAA 和 UGA
2 、肽链释放:终止因子进入 A 位,使核糖体的肽酰基转移酶变为水解酶,将肽酰基不转移给 A 位 tRNA ,而是转移至水分子上,多肽链从核糖体和 tRNA 上释放出来,核糖体从 mRNA 上释放下来。
3 、核糖体解离:准备去合成另一分子蛋白质, IF3 使核糖体解离为 30S 和 50S非功能性亚基,并与小亚基结合,以防重新聚合。消耗 GTP
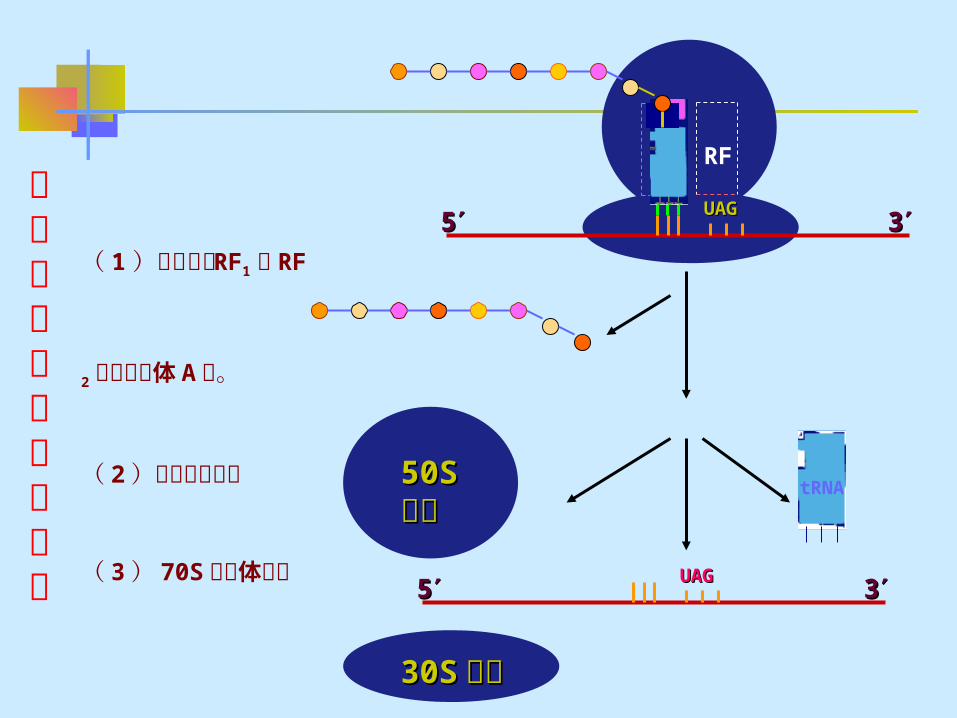
肽链合成的终止及释放
( 1 )释放因子 RF1
或 RF2 进入核糖体 A
位。
( 2 )多肽链的释放
( 3 ) 70S 核糖体解
离
55 33UAGUAG
30S30S 亚基亚基
50S50S亚基亚基
55 33UAGUAG
tRNA
RF


蛋白质的合成是一个高耗能过程
AA 活化 2 个高能磷酸键( ATP) 肽链起始 1 个( 70S复合物形成, GTP) 进位 1 个( GTP) 移位 1 个( GTP) 第一个氨基酸参入需消耗 3 个高能磷酸键(活化 2+起始 1 )
以后每掺入一个 AA需要消耗 4 个高能磷酸键(活化2 +进位 1 个 +移位 1 个)。

三、真核生物蛋白质的生物合成特点核糖体更大, 80S 40S+60S起始 tRNA 和氨基酸:起始密码子为 AUG ,起始氨基酸为甲硫氨酸,起始 tRNA 表示为 tRNAiMet ,起始氨酰 -tRNA 为Met- tRNAiMet
起始:它的上游 5’端无 SD 序列,由帽子结合蛋白促使小亚基与 mRNA 的 5’端 -帽子结合,然后沿mRNA移动寻找 AUG 。真核生物 mRNA 通常只有一个 AUG 密码子,每种 mRNA 只转译出一种多肽。真核中涉及的蛋白因子较多。真核中线粒体、叶绿体核糖体大小、组成及蛋白质合成过程都类似于原核细胞。


本 章 重 点
遗传密码的特点 mRNA 、 tRNA 及核糖体在蛋白质合成中
的作用。 SD 序列概念。 原核生物中蛋白质生物合成的主要过程。 原核细胞与真核细胞的蛋白质生物合成的主要区别


















