







Languages
Pages
Legal


The Effects of Cacophony Mutations
on Sleep in Drosophila melanogaster
By: Derek Willson
Schuylerville High School
Mentored By: Professor Christopher Vecsey
Skidmore College

Willson-2
Acknowledgement
I would like to acknowledge and thank Professor Christopher Vecsey, for mentoring me,
guiding me, and always being there when I needed him throughout the course of this research
project. I would also like to thank Dr. Andrew Frank of the University of Iowa, for provided the
mutant fly lines used in the project. I would lastly like to thank Mr. David Conneally, for
allowing me the opportunity to take part in our schools wonderful science research project and
for listening to me being overly excited about flies sleeping.

Willson-3
Abstract
Migraines, characterized by throbbing headaches, are often accompanied by symptoms
including nausea and dizziness. Familial Hemiplegic Migraine (FHM), a subtype of migraine
that runs in families, can be caused by a missense mutation in the CACNA1A gene, which is
involved in the production of CAv2.1 calcium channels. Although migraine sufferers often report
sleep problems, little is known about the relationship between FHM mutations and sleep. This
study examines the mechanisms between two CACNA1A mutations and sleep in Drosophila.
Lines containing the UAS-CAC-GFPS/L(3-8m) human transgene and lines containing the UAS-
CAC-GFP786C transgene were crossed with the ELAV-GAL4 driver, which expresses these
mutations across all neurons in their offspring. Using Drosophila Activity Monitors, the sleep of
the offspring was monitored. Males expressing either transgene and females expressing the
GFPS/L(3-8m) transgene saw increased sleep frequency during LD and DD cycles. Both male
and female Drosophila populations expressing the GFPS/L(3-8m) transgene were classified as
arrhythmic. The altered circadian rhythms in Drosophila expressing the S218L mutation may
help in understanding the mechanism linking sleep issues to migraines. This is because the
altered rhythms may cause the sleep issues and this should be pursued in future studies. Data
from all controls varied, which in future studies may be avoided by backcrossing the flies to a
common wild-type strain. Many males expressing the GFPS/L(3-8m) transgene died during the
experiment, which may have been caused by the expression across all neurons. Future studies
should address this by using drivers that express the transgene in a more localized area.

Table of Contents:
Acknowledgement- Page 2
Abstract- Page 3
Table of Contents/List of Figures- Page 4
Introduction- Page 5
Methods and Materials- Page 5
Drosophila Lines- Page 6
Crosses- Page 6
Sleeping Monitoring- Page 7
Results- Page 8
Light/ Dark Cycle Males- Page 8
Dark/Dark Cycle Males- Page 10
Light/Dark Cycle Females- Page 14
Dark/Dark Cycle Females- Page 14
Discussion and Conclusion- Page 17
References- Page 19
List of Figures:
Figure 1- Page 9
Figure 2- Page 9
Figure 3- Page 11
Figure 4- Page 11
Figure 5- Page 12
Figure 6- Page 13
Figure 7- Page 13
Figure 8- Page 15
Figure 9- Page 15
Figure 10- Page 16

Willson-5
Introduction
Characterized by some degree of hemiparesis during aura, Familial hemiplegic migraine
(FHM) is a rare, monogenic subtype of migraine with aura (1). In addition to this motor
weakness, diagnoses of FHM, requires a first or second degree relative to have migraine with
motor weakness aura (1). There are four variations of FHM, with each being categorized by the
specific gene mutated (1).
In this study, a heavy focus is laid upon Familial Hemiplegic Migraine Type 1(FHM-1).
FHM-1, is caused by missense mutations in the CACNA1A gene, which encodes the CAv2.1
subunit of calcium channels on the human chromosome 19 (1,2,3,4,5). In Drosophila, the CAC
gene, has been known to be involved in male courtship behavior as well as the encoding of the
voltage-gated calcium channel (6,7).
The specific CACNA1A mutation targeted in this study is the S218L mutation, one of the
more common FHM-1 mutations found in mammals (4).
Many migraine sufferers report many sleep problems with the most prominent being, trouble
falling asleep, waking up during the night, and lack of refreshing sleep (8,9,10,11). However,
little is known about the mechanisms that link FHM to sleep and circadian rhythms. The purpose
of this study is to gain a better understanding of how FHM mutations affect sleep and circadian
rhythms and to establish a baseline for future studies.
Prior to experimentation, it was hypothesized that abnormal sleep data will be obtained from
the Drosophila expressing either of the transgenes and that this effect will take place across both
genders.
Methods and Materials

Willson-6
Drosophila Lines
In this study, four separate lines of Drosophila were raised. The first of the experimental
groups was the UAS-CAC-GFP786C mutation. This mutation was produced by a transgene
insertion on the third chromosome (12). The second of the two experimental groups consisted of
Drosophila containing the UAS-CAC-GFPS/L(3-8m) transgene mutation. This transgene was
constructed by using PCR to alter the serine 161 codon of the UAS-CAC- GFP786C mutation to
leucine. This substitution corresponds to the S218L mammalian CACNA1A mutation (13,14).
These two Drosophila lines were generously provided by Dr. Andrew Frank of the University of
Iowa. The driver line ELAV-GAL4 was also raised. This driver allows for the expression of
transgenes in all neurons, but no other cells (15). Lastly, a W1118CS line was raised. This line
serves as a white eyed wild type, which has no driver capabilities (16). As lines grew in
population, virgin females and males were collected, and individually stored for later crosses.
Crosses
The GAL4/UAS system was used in this study to select which neurons would express the
desired transgene (17,18). This system works by having the UAS transgenes present in all cells,
but only produce in select cells where GAL4 is active (17,18). Since the ELAV-GAL4 driver is
used, all neurons will express the desired transgenes (15). The gender of the lines crossed was
based upon availability.
Crosses between UAS-CAC-GFPS/L(3-8m) males and ELAV-GAL4 virgin females, and
between UAS-CAC-GFPS/L(3-8m) males and W1118 virgin females were set up. In this case the
GFPS/L x GAL4 resulting offspring will be used as the experimental group, with the control
consisting of the GFPS/L x W1118 offspring. This is because the UAS-GAL4 will cause the
expression of the transgene, while the W1118CS will not (15,16).

Willson-7
Crosses between UAS-CAC- GFP786C virgin females and ELAV-GAL4 males and between
UAS-CAC- GFP786C virgin females and W1118 males were also set up. In this second set of
crosses, again the GFP786C x GAL4 resulting offspring will be used as the experimental group,
with the control consisting of the GFP786C x W1118CS offspring. This is also because the GFP786C
x GAL4 will cause the expression of the transgene, while the GFP786C x W1118CS will not
(15,16).
A final cross between ELAV-GAL4 males and W1118 virgin females was set up. This cross
served as another control for each of the two cases.
Sleep Monitoring
In this study data was obtained through the use of Drosophila Activity Monitors (DAM).
These DAMs monitor the activity of individual Drosophila in sealed tubes. They accomplish this
by making use of an infrared beam in the midpoint of the tubes, which measures fly activity by
how often the fly crosses the beam. It is from this information that data regarding sleep can be
obtained (19). Information obtained from these monitors was collected every minute throughout
the experiment.
Prior to monitoring the sleep of the resultant offspring, all female Drosophila had to have
mated. This is because sleep patterns in females, can be affected by whether or not mating has
occurred (20,21). This variable was eliminated by leaving male Drosophila, with the female
Drosophila, that will be used in the experiment, for approximately 24 hours. This ensured that
mating occurred.
After this time period expired, populations of 32 male and female flies were collected from
each of the crosses. This number was determined based upon the maximum capacity of
Drosophila Activity Monitors (19). These flies were then placed into individual tubes, and then

Willson-8
into the DAMs. The DAMs were subsequently placed into incubators with stable environmental
conditions.
For the first 7 days, all Drosophila were exposed to consistent cycle of 12 hours of light
followed by 12 hours of darkness (LD cycle). The following 5 days, all Drosophila were kept in
a constant 24 hours of Darkness (DD cycle). The LD cycle, was used to obtain data pertaining to
sleep when in the presence of an external stimulus, in this case light. While the DD cycle was
used to provide information on the natural circadian rhythms of the Drosophila, since there will
be no external stimulus, such as light (22,23).
Results
Usable data from 17 GFPS/L x GAL4 males, 30 GFPS/L x W1118 males, 30 W1118x GAL4 males,
31 GFP786C x W1118 males, 31 GFP786C x GAL4 males, 32 GFPS/L x GAL4 females, 31 GFPS/L x
W1118 females, 29 W1118x GAL4 females, 32 GFP786C x W1118 females and 31 GFP786C x GAL4
females was obtained. Data were considered usable if Drosophila lived throughout the entire
duration of the experiment.
Light/Dark Cycle Males
The GFPS/L x GAL males in a LD were noted sleeping on average many more times a day
than GFPS/L x W1118 males or W1118CS x GAL4 males (Figure 2B). GFPS/L x GAL males also
saw slightly shorter daily average episodic sleep duration than either GFPS/L x W1118 males or
W1118CS x GAL4 males (Figure 2C). However, GFPS/L x GAL males had a much shorter
episodic sleep duration than either of its controls during the 12 hour dark period (Figure 2C). On
average GFPS/L x GAL males showed small increase over its controls in total sleep duration
(Figure 2A), and tended to sleep slightly more towards the end of the 12 light period than either
controls (Figure 1B).

Willson-9
Figure 1. Male LD cycle Average Sleep per 30 minutes
A) Graph A depicts the average sleep per 30 minutes for all male flies for each cross during the LD cycle period.
B) Graph B depicts the average sleep per 30 minutes all crosses pertaining to the UAS-CAC-GFPS/L(3-8m) transgene
and its controls during the LD cycle period.
C) Graph C depicts the average sleep per 30 minutes all crosses pertaining to the UAS-CAC- GFP786C transgene and its
controls during the LD cycle period.
A
B C
B
A
C
Figure 2. Male LD Cycle Sleep Graphs
A) Graph A depicts the average total sleep duration across all male flies for each cross during the LD cycle period.
B) Graph B depicts the average number of sleep episodes across all male flies for each cross during the LD cycle period.
C) Graph C depicts the average episodic sleep duration across all male flies for each cross during the LD cycle period.

Willson-10
GFP786C x GAL4 males were seen sleeping on average a few more times daily than either the
GFP786C x W1118 males or the W1118x GAL4 males. The largest difference occurred during the 12
dark period (Figure 2B). GFP786C x GAL4 males also showed a shorter average daily episodic
sleep duration than either of the controls, with the largest discrepancy occurring during the 12
hour dark period (Figure 2C). GFP786C x GAL4 males were seen sleeping on average more than
the W1118x GAL4 males during the 12 hour light cycle, but less than GFP786C x W1118 males
during both the light and dark cycles (Figure 1C, Figure 2A).
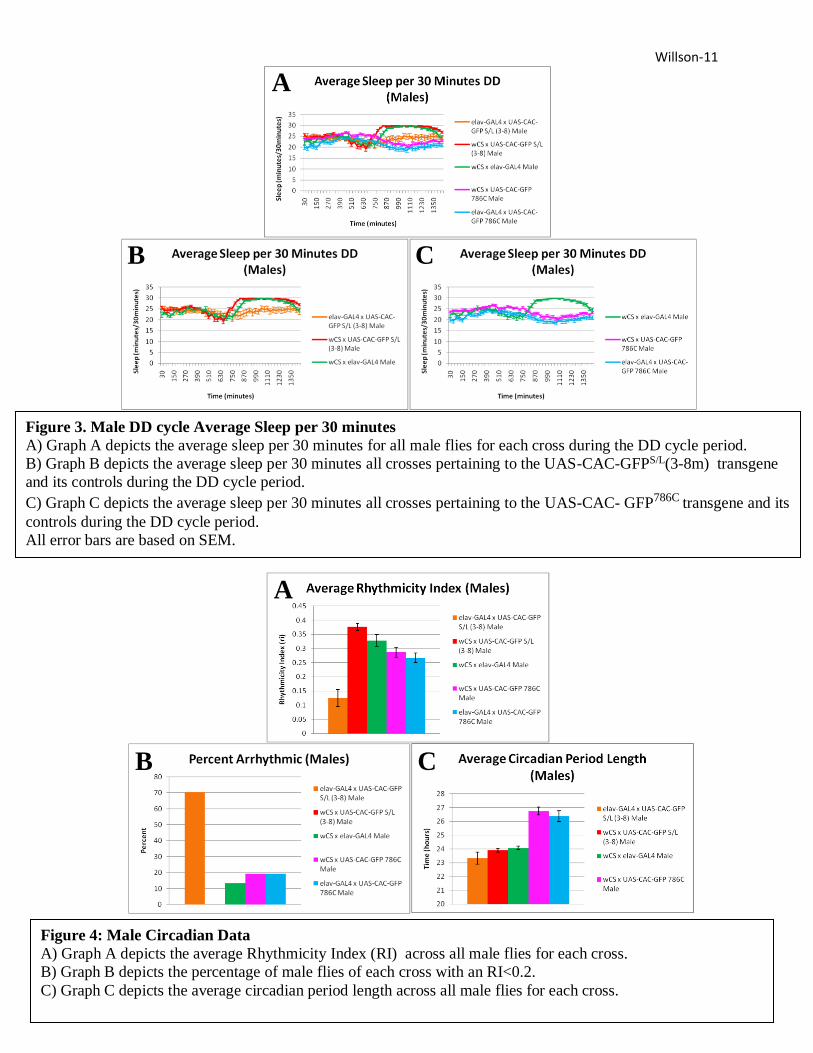
Dark/Dark Cycle Males
The GFPS/L x GAL4 males in DD had on average a much lower rhythmicity index (RI) than
either GFPS/L x W1118 males or W1118CS x GAL4 males (Figure 4A). In fact, 70.6% of the GFPS/L
x GAL4 males were classified as arrhythmic (RI<0.2), meaning the population studied was
arrhythmic. Neither of its controls was classified as arrhythmic and each had a much lower
percentage of arrhythmic flies (Figure 4B). The GFPS/L x GAL4 males are noted as having a
much flatter line in the average sleep per 30 minutes graph than either of their controls, showing
their weaker rhythmicity (Figure 3B). These flies also had a slightly shorter circadian period
length than their controls (Figure 4C). GFPS/L x GAL4 males were noted sleeping many times
more per day than GFPS/L x W1118 males or W1118CS x GAL4 males (Figure 5A) and having a
much shorter average episodic sleep duration, especially during the second period of darkness,
than these controls (Figure 5C). The average total sleep duration of GFPS/L x GAL4 males, was
slightly less than their controls (Figure 5A).
Willson-11
Figure 3. Male DD cycle Average Sleep per 30 minutes
A) Graph A depicts the average sleep per 30 minutes for all male flies for each cross during the DD cycle period.
B) Graph B depicts the average sleep per 30 minutes all crosses pertaining to the UAS-CAC-GFPS/L(3-8m) transgene
and its controls during the DD cycle period.
C) Graph C depicts the average sleep per 30 minutes all crosses pertaining to the UAS-CAC- GFP786C transgene and its
controls during the DD cycle period.
All error bars are based on SEM.
A
A
B C
Figure 4: Male Circadian Data
A) Graph A depicts the average Rhythmicity Index (RI) across all male flies for each cross.
B) Graph B depicts the percentage of male flies of each cross with an RI<0.2.
C) Graph C depicts the average circadian period length across all male flies for each cross.
A
B C

Willson-12
The GFP786C x GAL4 males showed only a slightly lower average RI when compared to GFP-
786C x W1118 males or W1118x GAL4 males (Figure 4A). The average circadian period length of
GFP786C x GAL4 males was longer than that of W1118x GAL4 males, but shorter than that of
GFP786C x W1118 males (4C). GFP786C x GAL4 males slept on average more times per day and for
a shorter average episodic duration than either of the controls groups (Figure 5B, Figure 5C),
the larger difference for both cases was between GFP786C x GAL4 males and W1118x GAL4
males.
Figure 5. Male LD Cycle Sleep Graphs
A) Graph A depicts the average total sleep duration across all male flies for each cross during the DD cycle period.
B) Graph B depicts the average number of sleep episodes across all male flies for each cross during the DD cycle period.
C) Graph C depicts the average episodic sleep duration across all male flies for each cross during the DD cycle period.
All error bars are based on SEM.
B A
C

Willson-13
Figure 6. Female LD cycle Average Sleep per 30 minutes
A) Graph A depicts the average sleep per 30 minutes for all female flies for each cross during the LD cycle period.
B) Graph B depicts the average sleep per 30 minutes all crosses pertaining to the UAS-CAC-GFPS/L(3-8m) transgene
and its controls during the LD cycle period.
C) Graph C depicts the average sleep per 30 minutes all crosses pertaining to the UAS-CAC- GFP786C transgene and its
controls during the LD cycle period.
All error bars are based on SEM.
A
A
B C
Figure 7. Female LD Cycle Sleep Graphs
A) Graph A depicts the average total sleep duration across all female flies for each cross during the LD cycle period.
B) Graph B depicts the average number of sleep episodes across all female flies for each cross during the LD cycle
period.
C) Graph C depicts the average episodic sleep duration across all female flies for each cross during the LD cycle period.
All error bars are based on SEM.
B
A
C

Willson-14
GFP786C x GAL4 males were also noted having a slightly lower average total sleep duration than
their controls (Figure 5A).
Light/Dark Cycle Females
In LD the GFPS/L x GAL4 females were seen having a much higher average total sleep then
their controls, which were the GFPS/L x W1118 females and the W1118CS x GAL4 females (Figure
6b, Figure 7A). This is likely caused by the GFPS/L x GAL4 females having a higher number of
average sleep episodes per day (Figure 7B), while also having a slight increase in average daily
episodic sleep duration than either of their controls (7C).
The GFP786C x GAL4 females were noted having an increase in average number of sleep
episodes per day, when compared to the GFP786C x W1118 females or the W1118x GAL4 females
(Figure 7B). The average episodic sleep duration for GFP786C x GAL4 females, was slightly less
than their controls over the course of a day, and much less during the second 12 dark periods
(Figure 7C). Lastly, the average total sleep of the GFP786C x GAL4 females was slightly more
than that of the W1118x GAL4 females and slightly less than that of the GFP786C x W1118 females
(Figure 7A).
Dark/Dark Cycle Females
In DD the GFPS/L x GAL4 females had an average RI value much lower than that of the
GFPS/L x W1118 females or the W1118CS x GAL4 females (Figure 9). This lower average RI
translate to approximately 80.4% of GFPS/L x GAL4 females being classified as arrhythmic
(RI<0.2) (Figure 9B). This percentage is much higher than either of the controls, which is likely
the reason for the GFPS/L x GAL4 females showing a much flatter curve in the average sleep per
30 minutes graph than the two controls (figure 8B). However, the average circadian period

Willson-15
Figure 8. Female DD cycle Average Sleep per 30 minutes
A) Graph A depicts the average sleep per 30 minutes for all female flies for each cross during the DD cycle period.
B) Graph B depicts the average sleep per 30 minutes all crosses pertaining to the UAS-CAC-GFPS/L(3-8m) transgene
and its controls during the DD cycle period.
C) Graph C depicts the average sleep per 30 minutes all crosses pertaining to the UAS-CAC- GFP786C transgene and its
controls during the DD cycle period.
All error bars are based on SEM.
A
A
B C
Figure 9: Female Circadian Data
A) Graph A depicts the average Rhythmicity Index (RI) across all female flies for each cross.
B) Graph B depicts the percentage of female flies of each cross with an RI<0.2.
C) Graph C depicts the average circadian period length across all female flies for each cross.
All error bars are based on SEM
A
B C

Willson-16
length of the GFPS/L x GAL4 females was slightly shorter than that of the GFPS/L x W1118
females but slightly longer than that of W1118CS x GAL4 females (Figure 9C). The GFPS/L x
GAL4 females also had a much higher average number of sleep episodes per day and a slightly
higher average daily total sleep duration over the two control groups (Figure 10B, Figure 10A).
The average episodic sleep duration of the GFPS/L x GAL4 females, was slightly higher than that
of the GFPS/L x W1118 females, but slightly lower than that of the W1118CS x GAL4 females.
Figure 10. Female DD Cycle Sleep Graphs
A) Graph A depicts the average total sleep duration across all female flies for each cross during the DD cycle period.
B) Graph B depicts the average number of sleep episodes across all female flies for each cross during the DD cycle
period.
C) Graph C depicts the average episodic sleep duration across all female flies for each cross during the DD cycle period.
All error bars are based on SEM.
B A
C

Willson-17
The GFP786C x GAL4 females had an average RI that was slightly lower than that of GFP786C
x W1118 females but slightly higher than that of W1118CS x GAL4 females (Figure 9A). The GFP-
786C x GAL4 females also had an average circadian period length that was slightly shorter than
that of GFP786C x W1118 females but slightly longer than that of W1118CS x GAL4 females (Figure
9C). The GFP786C x GAL4 females had a higher average number of sleep episodes per day
(Figure 10B), but a shorter average episodic sleep duration than their two controls (Figure 10C).
This ultimately leads to the GFP786C x GAL4 females having a lower average total sleep duration
than their controls (10A).
Discussion/Conclusion
The main purpose of this study was to gain a better understanding of the mechanism linking
FHM-1 to sleep and circadian rhythms. The sleep patterns of Drosophila with a common FHM-1
mutation were monitored and analyzed, and compared to that of Drosophila with a different
cacophony mutation that doesn’t result in FHM-1. Perhaps the most promising data pertains to
circadian rhythms. The GFPS/L x GAL4 cross produced both males and female populations that
were classified as arrhythmic, having RI of less than 0.2. This finding may prove to be
significant, because mutations in cacophony may lead to altered circadian rhythms. Also, nearly
50% of males expressing the S218L mutation died over the course of the experiment. This may
be a result of broadly expressing this gene in all neurons. Future studies may want to potentially
use drivers that do not express the transgene across all neurons to look for any significance in
this increase mortality rate. Overall the data supported the original hypothesis. This is because
lines expressing either of the transgenes tended to show different sleep data, across the three
major sleep categories analyzed, while abnormal circadian rhythms were observed in both male
and female flies expressing the S218L. In the future, an ANOVA test will be conducted on this

Willson-18
data to gain a better understanding of the significance of the data. From the data the most
pertinent problem faced, was the discrepancies between the control groups. The three control
groups GFPS/L x W1118, W1118x GAL4, GFP786C x W1118 , should have produced similar results.
This is because none of the controls had both a UAS and a GAL4 group which would cause the
expression of the mutated transgene. Despite this the controls produced varying results, which
different greatly in many portions of the data. Potential explanation for the cause of the control
discrepancies could have been the lack of backcrossing, prior to sleep examination (24). This
backcrossing would help to reduce the effects of the flies’ genetic background on the study, and
help to block out some background genes which could alter sleep patterns in Drosophila (24).
This may be a potential avenue for further research, and using the same experiment but
implementing backcrossing into the procedure. A second cause of potential issues, particularly
for the males, would be the inheritance of the desired gene. Being able to only inherit one X
chromosomes, male flies may not inherit the proper genes to express the desired phenotype,
which could lead to varying data. Again, this should be addressed by future studies, in order to
increase the reliability of the controls. Another interesting future area of study would be locating
the specific neurons which may connect these cacophony mutations to sleep and circadian
rhythms would be to implement different drivers. Different drivers, will express these mutations
in different cells in the Drosophila, and by observing the effects of different drivers the
mechanism linking FHM to sleep and circadian rhythms could be further localized and a better
understanding of it be had. The expression of the S218L mutation, the most common mammalian
FHM-1 mutation, resulted in altered circadian rhythms in Drosophila. The fact that a mutation in
the same gene as the S218L, which does not result in FHM, did not alter circadian rhythms
strengthened the case that FHM alter circadian rhythms. This is because only cacophony

Willson-19
mutations that result in FHM resulted in altered circadian rhythms in this study. This finding may
also expand to humans who suffer from FHM-1. Migraines have been known to negatively
impact sleep in humans, but the underline reasons behind this negative impact is not fully
understood. This finding that S2l8L is linked to altered circadian rhythms may help to explain
the negative affects migraines and in particular FHM have on sleep. The worsened sleep of those
suffering from FHM may be attributed to their altered circadian rhythms, and this avenue for
future research should be explored. To do this, mammalian species, such as rats, containing the
S218L mutation should have their sleep monitored, and potential alterations in circadian rhythms
should be looked for. If the same results are duplicated in mammalian species, then a new
explanation for why those with FHM experience worsened sleep may have been discovered. That
discovery would be that FHM does not affect sleep directly; rather it affects circadian rhythms,
which in turn has a negative impact on individuals sleep.
References
(1) The International Classification of Headache Disorders, 3rd edition (beta version).
(2013). Cephalalgia, 33(9), 629-808. doi:10.1177/0333102413485658
(2) Weiss, N., Sandoval, A., Felix, R., Maagdenberg, A. V., & Waard, M. D. (2008, June 26). The
S218L familial hemiplegic migraine mutation promotes deinhibition of Cav2.1 calcium channels
during direct G-protein regulation.
(3) Ophoff, R. A., Terwindt, G. M., Vergouwe, M. N., Eijk, R. V., Oefner, P. J., Hoffman, S. M., . . .
Frants, R. R. (1996). Familial Hemiplegic Migraine and Episodic Ataxia Type-2 Are Caused by
Mutations in the Ca2 Channel Gene CACNL1A4. Cell, 87(3), 543-552. doi:10.1016/s0092-
8674(00)81373-2
(4) Hans, M., Luvisetto, S., Williams, M. E., Spagnolo, M., Urrutia, A., Tottene, A., . . . Pietrobon, D.
(1999, March 01). Functional Consequences of Mutations in the Human α1A Calcium Channel
Subunit Linked to Familial Hemiplegic Migraine.
(5) Schilcher, F. V. (1977). A mutation which changes courtship song inDrosophila
melanogaster. Behavior Genetics, 7(3), 251-259. doi:10.1007/bf01066278
(6) Chan, B., Villella, A., Funes, P., & Hall, J. C. (2002). Courtship and other behaviors affected by a
heat-sensitive, molecularly novel mutation in the cacophony calcium-channel gene of
Drosophila. Genetics, 162(1), 135–153.

Willson-20
(7) Yamazaki, S., Ikeno, K., Abe, T., Tohyama, J., & Adachi, Y. (2011). Hemiconvulsion-Hemiplegia-
Epilepsy Syndrome Associated With CACNA1A S218L Mutation. Pediatric Neurology, 45(3),
193-196. doi:10.1016/j.pediatrneurol.2011.04.010
(8) Kelman, L., & Rains, J. C. (2005). Headache and Sleep: Examination of Sleep Patterns and
Complaints in a Large Clinical Sample of Migraineurs. Headache: The Journal of Head and
Face Pain, 45(7), 904-910. doi:10.1111/j.1526-4610.2005.05159.x
(9) Rasmussen, B. K. (1993). Migraine and tension-type headache in a general population:
Precipitating factors, female hormones, sleep pattern and relation to lifestyle. Pain, 53(1), 65-72.
doi:10.1016/0304-3959(93)90057-v
(10) Spierings, E. L., & Hoof, M. (1997). Fatigue and Sleep in Chronic Headache Sufferers: An Age-
and Sex-Controlled Questionnaire Study.Headache: The Journal of Head and Face Pain, 37(9),
549-552. doi:10.1046/j.1526-4610.1997.3709549.x
(11) Vgontzas, A., Cui, L., & Merikangas, K. R. (2008). Are Sleep Difficulties Associated With
Migraine Attributable to Anxiety and Depression?Headache: The Journal of Head and Face
Pain, 48(10), 1451-1459. doi:10.1111/j.1526-4610.2008.01175.x
(12) Kawasaki, F. (2004). Active Zone Localization of Presynaptic Calcium Channels Encoded by the
cacophony Locus of Drosophila. Journal of Neuroscience, 24(1), 282-285.
doi:10.1523/jneurosci.3553-03.2004
(13) Inagaki, A., Frank, C. A., Usachev, Y. M., Benveniste, M., & Lee, A. (2014). Pharmacological
correction of gating defects in the voltage- gated Cav2.1 Ca2+channel due to a familial
hemiplegic migraine mutation. Neuron, 81(1), 91–102.
http://doi.org/10.1016/j.neuron.2013.10.056
(14) Brusich, D. J., Spring, A. M., James, T. D., Helms, T. H., & Frank, C. A. (2017). Drosophila
CaV2 Channels Harboring Human Migraine Mutations Cause Synapse Hyperexcitability That
Can Be Suppressed By Inhibition Of A Ca2+ Store Release Pathway. doi:10.1101/141366
(15) Lin, D. M., & Goodman, C. S. (1994, September). Ectopic and increased expression of Fasciclin II
alters motoneuron growth cone guidance.
(16) Kurkulos, M., Weinberg, J. M., Pepling, M. E., & Mount, S. M. (1991). Polyadenylylation in
copia requires unusually distant upstream sequences. Proceedings of the National Academy of
Sciences, 88(8), 3038-3042. doi:10.1073/pnas.88.8.3038
(17) Osterwalder, T., Yoon, K. S., White, B. H., & Keshishian, H. (2001). A conditional tissue-specific
transgene expression system using inducible GAL4. Proceedings of the National Academy of
Sciences, 98(22), 12596-12601. doi:10.1073/pnas.221303298
(18) AH, Brand & Perrimon, N. (1993). Targeted gene expression as a means of altering cell fates and
generating dominant phenotypes. Development. 118. 401-415.
(19) Pfeiffenberger, C., Lear, B. C., Keegan, K. P., & Allada, R. (2010). Locomotor Activity Level
Monitoring Using the Drosophila Activity Monitoring (DAM) System. Cold Spring Harbor
Protocols, 2010(11). doi:10.1101/pdb.prot5518
(20) Garbe, D. S., Vigderman, A. S., Moscato, E., Dove, A. E., Vecsey, C. G., Kayser, M. S., &
Sehgal, A. (2016). Changes in Female Drosophila Sleep following Mating Are Mediated by
SPSN-SAG Neurons. Journal of Biological Rhythms, 31(6), 551-567.
doi:10.1177/0748730416668048
(21) Dove, A. E., Cook, B. L., Irgebay, Z., & Vecsey, C. G. (2017). Mechanisms of sleep plasticity due
to sexual experience in Drosophila melanogaster.Physiology & Behavior, 180, 146-158.
doi:10.1016/j.physbeh.2017.08.020

Willson-21
(22) Crosthwaite, S. K., Loros, J. J., & Dunlap, J. C. (1995). Light-induced resetting of a circadian
clock is mediated by a rapid increase in frequency transcript. Cell, 81(7), 1003-1012.
doi:10.1016/s0092-8674(05)80005-4
(23) Arpaia, Giuseppina & J. Loros, Jennifer & C. Dunlap, Jay & Morelli, Giorgio & Macho,
Giuseppe. (2017). The lnterplay of Light and the Circadian Clock. .
(24) Chandler, C. H., Chari, S., & Dworkin, I. (2013). Does your gene need a background check? How
genetic background impacts the analysis of mutations, genes, and evolution. Trends in
Genetics, 29(6), 358-366. doi:10.1016/j.tig.2013.01.009
Top Related