







Languages
Pages
Legal


HAL Id: hal-00895791https://hal.archives-ouvertes.fr/hal-00895791
Submitted on 1 Jan 2008
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come fromteaching and research institutions in France orabroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, estdestinée au dépôt et à la diffusion de documentsscientifiques de niveau recherche, publiés ou non,émanant des établissements d’enseignement et derecherche français ou étrangers, des laboratoirespublics ou privés.
Parmigiano Reggiano cheese: evolution of cultivable andtotal lactic microflora and peptidase activities during
manufacture and ripeningJuliano De Dea Lindner, Valentina Bernini, Angela De Lorentiis, Alberto
Pecorari, Erasmo Neviani, Monica Gatti
To cite this version:Juliano De Dea Lindner, Valentina Bernini, Angela De Lorentiis, Alberto Pecorari, Erasmo Neviani,et al.. Parmigiano Reggiano cheese: evolution of cultivable and total lactic microflora and peptidaseactivities during manufacture and ripening. Dairy Science & Technology, EDP sciences/Springer,2008, 88 (4-5), pp.511-523. <hal-00895791>

Dairy Sci. Technol. 88 (2008) 511–523 Available online at:c© INRA, EDP Sciences, 2008 www.dairy-journal.orgDOI: 10.1051/dst:2008019
Original article
Parmigiano Reggiano cheese: evolutionof cultivable and total lactic microflora
and peptidase activities during manufactureand ripening
Juliano De Dea Lindner1, Valentina Bernini1, Angela De Lorentiis1,Alberto Pecorari2, Erasmo Neviani1, Monica Gatti1*
1 Department of Genetics, Biology of Microorganisms, Anthropology, Evolution,University of Parma, Parma, Italy
2 Consorzio del Formaggio Parmigiano-Reggiano, Reggio Emilia, Italy
Abstract – Parmigiano Reggiano is a Protected Designation of Origin, long-ripened cheese, madefrom cow’s milk supplemented with natural whey starter, which thus contains a large microbialbiodiversity. The aim of this study was to understand the population dynamics of the total lacticmicroflora throughout the manufacture and ripening of this cheese. Several approaches were com-bined to determine the quantitative changes in the different bacterial populations during 20 monthsof ripening of Parmigiano Reggiano cheeses from the same cheesemaking. Total and viable cellswere enumerated after fluorescent labeling. Culturable bacteria were enumerated on different platecount agar media, including original media prepared from curd and ripened cheese. Six peptidaseactivities were quantified in curd and cheese samples free from cells. While the total bacterial cul-tivable population remained high and similar for the first six months, a decrease in viable starterlactic acid bacteria was observed during the first 48 h. The non-starter lactic acid bacteria popula-tions, initially present in low numbers, began to grow after the brining and remained at high levels(about 107 CFU·g−1) for at least 10 months. During ripening, a strong decrease in the total bacterialpopulation and a marked increase in 4 out of 6 peptidase activities were observed. In the externaland internal zones of Parmigiano Reggiano cheese different trends in microbial growth, cell autol-ysis and peptidase activity were observed. This study gives for the first time a global view of thepossible contribution of total, viable, cultivable and lysed bacterial cells throughout the ripening ofParmigiano Reggiano cheese.
Parmigiano Reggiano cheese / ripening / lactic acid microflora / bacterial cell lysis / cellviability / peptidase activity
摘摘摘要要要 – 干干干酪酪酪制制制造造造和和和成成成熟熟熟期期期间间间乳乳乳酸酸酸菌菌菌菌菌菌群群群的的的可可可培培培养养养性性性和和和肽肽肽酶酶酶活活活性性性的的的评评评价价价。。。ParmigianoReggiano 是一种受原产地名号保护 (PDO) 的产品, 是牛乳添加自然发酵形成的乳清发酵剂而制成的长成熟期干酪, 因此所含的微生物种类繁多。本研究目的是了解该干酪在整个制造和成熟期间乳酸菌菌群的动态变化。采用了不同的方法对来自同一厂家的成熟期为 20 个月的 Parmigiano Reggiano 干酪中不同的细菌菌数进行了定量分析。用荧光标记法进行了总细胞和活细胞计数。在不同的琼脂平板培养基上对最初凝乳和成熟干酪中的可培养细菌进行了计数。定量测定了凝乳和干酪中的六种肽酶活性。研究发现总的可培养细菌数始终较高水平,
* Corresponding author (通讯作者): [email protected]
Article published by EDP Sciences

512 J. De Dea Lindner et al.
并且在前六个月细菌总数基本上相同,在最初的 48小时发酵期内,乳酸菌发酵剂的数量明显降低。发酵初期,非发酵剂乳酸菌数量较低,但加盐渍后非发酵剂乳酸菌的数量持续上升,至少在 10个月内持续增长并保持在较高水平 (约 107 CFU·g−1)。成熟过程中,细菌总数显著下降, 6种肽酶中有 4种活性显着增加。同时,对 Parmigiano Reggiano干酪的外部和内部区域微生物生长的趋势,细胞自溶和肽酶的活性也进行了研究。本文首次研究了总乳酸菌、活菌,可培养乳酸菌和溶解的菌体细胞对 Parmigiano Reggiano干酪整个成熟期间的作用。
Parmigiano Reggiano干干干酪酪酪 /成成成熟熟熟 /乳乳乳酸酸酸菌菌菌菌菌菌群群群 /细细细菌菌菌细细细胞胞胞溶溶溶解解解 /细细细胞胞胞活活活性性性 /肽肽肽酶酶酶活活活性性性
Résumé – Le Parmigiano Reggiano : évolution de la microflore lactique cultivable et totale etdes activités peptidasiques pendant la fabrication et l’affinage. Le Parmigiano Reggiano est unfromage d’Appellation d’Origine Protégée à affinage long, fabriqué à partir de lait cru supplémentéd’un levain naturel issu du lactosérum. Ce fromage contient, en conséquence, une large biodiversitémicrobienne. L’objectif de cette étude était de comprendre la dynamique de population de la micro-flore lactique pendant la fabrication et l’affinage du Parmigiano Reggiano. Plusieurs approches ontété combinées pour déterminer l’évolution des différentes populations bactériennes dans ce fromageau cours de 20 mois d’affinage de fromages issus de la même série de fabrications. Le nombre debactéries totales et viables a été mesuré après marquage fluorescent. Les bactéries cultivables ontété déterminées en utilisant différents milieux gélosés, incluant des milieux originaux préparés àpartir de caillé et de fromage affiné. Six activités peptidasiques ont été quantifiées dans des échan-tillons de caillé et de fromages exempts de cellules bactériennes. La population de bactéries viablesdiminuait dans le caillé pendant les 48 premières heures, alors que la population totale de bactériescultivables restait élevée et constante sur les six premiers mois d’affinage. Les populations de bac-téries lactiques non levains, présentes initialement en faible nombre, commençaient à se développerdès le saumurage puis restaient à un niveau élevé (environ 107 UFC·g−1) pendant plus de 10 mois.Une forte chute de la population bactérienne totale était observée peandant l’affinage, accompagnéed’une hausse marquée de 4 activités peptidasiques. Dans les parties externes et internes du Parmi-giano Reggiano, différentes tendances de croissance, d’autolyse et d’activité peptidasique étaientobservées. Cette étude donne pour la première fois un aperçu global de la contribution possible despopulations totales, viables, cultivables et lysées tout au long de l’affinage du Parmigiano Reggiano.
Parmigiano Reggiano / affinage / bactéries lactiques / lyse bactérienne / viabilité cellulaire /activité peptidasique
1. INTRODUCTION
Parmigiano Reggiano (PR) is a Pro-tected Designation of Origin (PDO), hard-textured, cooked and long-ripened cheese.It is made following strict manufacturingprocedures, from cow’s milk supplementedwith natural whey starter only (http://www.parmigiano-reggiano.it). Only unheatedraw milk, obtained by mixing eveningmilk (partially skimmed by natural cream-ing) with full-cream milk of the follow-ing morning can be used. During the nat-ural creaming the reduction of the bacteriaascending with the fat globules and thegrowth of the residual non-starter lacticacid bacteria (NSLAB) occur [28]. Natural
whey starter consists of a culture of ther-mophilic lactic acid bacteria (SLAB) se-lected by cooking of the curd and by in-cubation of the sweet whey which resultsfrom each day’s cheesemaking as it natu-rally cools down.
Other important technological charac-teristics are: the use of calf rennet, thecooking temperature of the curd (54–55 ◦C), slow heat dissipation during lac-tic acid fermentation, whey drainage fromthe curd, and brining with NaCl and slowdiffusion into the cheese, as well as a longripening time of at least 12 months [25].
A very interesting recent study high-lighted the effect of the curd cooking tem-perature on the viability of SLAB and

Microbiology of Parmigiano Reggiano cheese 513
NSLAB, cheese composition and ripen-ing indices of an experimental semi-hardcheese [35]. The cooking temperatureplays a key role in the acidification pro-cess in the first hours after PR cheesemak-ing [32]. Furthermore, the temperature atthe end of cooking has been shown to in-fluence the moisture content of cheese andto affect the acidification curves in the in-ner zone of the cheese, where slight varia-tions in temperature can advance or delaythe beginning of the lactic fermentation.
Microbiological features of PR havebeen studied by Coppola et al. [8, 9], whoisolated a great number of strains froma large amount of samples representativeof the production and the earlier and ad-vanced stages of ripening on traditionalgrowth media. Recently, the diversity ofthe dominant lactic acid bacteria popula-tion in six samples of 12-month ripenedParmigiano Reggiano cheese was also in-vestigated by a culture-independent ap-proach [15]. However, the evolution ofLAB during the ripening of the samecheesemaking, to our knowledge, hasnever been investigated.
During the ripening period, LABcontribute to cheese flavor developmentthrough several basic mechanisms thatinclude carbohydrate fermentation, con-version of milk proteins into peptides andfree amino acids, catabolism of aminoacids into aroma compounds, hydrolysisof milk lipids into free fatty acids, andcitrate catabolism [13]. In particular, forlong-ripened cheeses such as PR, theproteolysis may be considered an essentialmulti-step biochemical event [14, 29].
Over the last few years, some LABcomplex peptidase systems with differ-ent natures, specificities and localizationhave been characterized [6, 21, 26, 33, 38].Residual enzymatic activity in cheese canbe considered as the trace of the mi-crobial ecosystem evolution during ag-ing [17, 40]. In a comprehensive studyof the relationship between starter autol-
ysis and the proteolytic enzyme comple-ment released in Cheddar cheese, Sheehanet al. [34] found that differences in lacto-coccal starter autolysis influenced the pro-teolytic activities of the released enzymesduring ripening. Moreover, in a recentstudy the positive impact of Lactobacillushelveticus autolysis on flavor precursor de-velopment in ripened Cheddar cheese hasbeen demonstrated [19].
For many reasons, the study of lacticmicroflora and peptidase activities is im-portant to understand the essential bio-chemical events during PR manufactureand ripening. In this work, a comprehen-sive sampling design allowed the analysisof this PDO cheese to be performed duringits production and over 20 months of ripen-ing. The SLAB and NSLAB cultivabilityand viability were assessed. Furthermore,in order to stress the role of microbialenzymes released after cellular lysis, theaminopeptidase and dipeptidylaminopepti-dase activities in the extract of cheese, freefrom cells, were evaluated.
2. MATERIALS AND METHODS
2.1. Cheesemaking
With the aim of obtaining a suitablenumber of PR cheeses necessary for thisstudy, a well-equipped factory able to guar-antee the production of twin wheels waschosen. The milk and whey starter wereheld and processed in the same tanks andusing a volume counter pump, the milkwas distributed in equal volumes in thevats. The cheese was produced accordingto the PR production protocol (http://www.parmigiano-reggiano.it).
Natural whey starter was added to eightvats each containing 1040 kg of milk(510 kg of partially skimmed evening milkand 530 kg of whole morning milk) at33 g·L−1, bringing the pH of the mixture to6.20. After the cheesemaking and molding,the wheels were held for three days and

514 J. De Dea Lindner et al.
frequently turned to enable complete wheydrainage. Sixteen wheels were then saltedby immersion in brine at 300 g NaCl·L−1
for 23 days. Ripening was done in ag-ing rooms with 85% relative humidity for20 months.
2.2. Dairy samples
First of all, aliquots of the total massof whey starter, raw milk and curd weresampled. Subsequently, for each of the fol-lowing samples, curd 6, 12 and 48 h af-ter extraction, salted cheese (1 month) andcheese at different stages of ripening (2, 3,4, 6, 8, 10, 12, 16 and 20 months) wereused; the wheels were cut lengthwise alongthe vertical axis and a central cheese sec-tion (1.0 cm thick) was obtained. Internaland external (0.5 cm from the cheese crust)portions were obtained from these cheesesections and grated before the analysis.
2.3. Physico-chemical analyses
The pH of samples was directly mea-sured using a pH meter (350 pH, BeckmanCoulter, Fullerton, CA, USA).
Lactose, galactose, L-lactate andD-lactate were determined according toCareri et al. [4], by enzymatic assays ing·L−1 with Boehringer-Mannheim kits(Boehringer-Mannheim, Germany). Totallactate was obtained through the sum ofL-lactate plus D-lactate.
Citrate, succinate, acetic acid and py-roglutamic acid were determined usinga DAD 540 liquid chromatography sys-tem with a variable UV/visible wavelength(Kontron Instruments, Italy) and refrac-tometric detectors (Refractive Index 475,Kontron Instruments, Italy). Analyses wereperformed using an Aminex HPX 87Hcation exchange column (Bio-Rad Lab-oratories, CA, USA), and filtered anddegassed 0.009 N reagent grade H2SO4
(Carlo Erba, Italy) as the mobile phaseat a flow rate of 0.6 mL·min−1. Eluates
were monitored at 215 nm. The calibra-tion curves were obtained by preparing astandard mix of the organic acids (Sigma,USA). The resulting peaks areas werecalculated for duplicate 25-μL injectionsand plotted against concentration usingthe software Kromasystem 2000 (KontronInstruments, Italy).
Each assay was carried out in du-plicate and average values expressed ing·100 g−1 for the lactose, galactose,L-lactate, D-lactate, citrate, succinate, andacetic and pyroglutamic acid concentra-tion during PR manufacturing and ripen-ing. The estimated error for the enzymaticassays was less than 5% and for the chro-matographic assays it was less than 3%.
2.4. Microbiological analyses
2.4.1. Media and growth conditions
Whey starter and milk were serially di-luted tenfold in 0.05 mol·L−1 sodium cit-rate (Sigma, Italy) buffer, pH 7.5, while10 g of curd and 10 g of the internal orexternal cheese samples were first homog-enized for 3 min in a blender (Stomacher400, Seward, UK) with sodium citrate be-fore being serially diluted. In order to re-cover the cultivable bacterial population,the following media and temperature/timeconditions of incubation were used: MRSagar pH 5.4 (Oxoid, Italy) at 42 ◦C for48 h and 25 ◦C for 72 h to facilitate, re-spectively, the growth of thermophilic andmesophilic lactobacilli; M17-SSW [12] at42 ◦C for 42 h to count streptococci, and25 ◦C for 72 h to count lactococci; andwhey agar medium (WAM) [18] at 42 ◦Cfor 48 h to facilitate the growth of thestrains arising from natural whey starter.Moreover, curd agar medium (CURDAM)[22] and cheese agar medium (CAM) [10,22], which are media mainly composedof grated curd and ripened cheese, re-spectively, were used at 42 ◦C for 72 hto select low nutritional-demanding LAB.

Microbiology of Parmigiano Reggiano cheese 515
The media were all incubated under anaer-obic conditions. The microbial counts werecarried out in duplicate and the standarddeviation of mean values was calculated.The estimated error was less than 10%.
2.4.2. Cell viability count
Cell viability was directly checked us-ing a LIVE/DEAD BacLight Bacterial Vi-ability Kit (Molecular Probes Inc., Eugene,USA) and an epifluorescence microscope(Nikon 80i, Tokyo, Japan) [16]. Curd andcheese samples were diluted as previouslydescribed, centrifuged (8000× g, 10 min,4 ˚C) and pelleted. The resulting pellet wasdiluted in 1 mL of sterile water and treatedwith 0.7 μL of Syto9 stain and 1 μL of pro-pidium iodide stain. Direct counts of total,viable and unviable bacterial populationswere evaluated using a Thoma chamber(Brand, Germany). Each sample was pre-pared and counted in duplicate and averagevalues were calculated. The estimated errorwas about 3% [16].
2.5. Determination of peptidaseactivities
Sample extracts were prepared accord-ing to Gatti et al. [17]. A dialysis step wasincluded to eliminate any low-molecular-weight substances such as salts whichcould interfere with the successive reac-tions. The sample suspension obtained af-ter 24 h of dialysis in tube-shaped cellu-lose (Spectra/por, Spectrum LaboratoriesInc., USA), with a cutoff of 3000 g·mol−1,was centrifuged (10 000× g, 10 min, 4 ◦C),filtered through a cellulose acetate mem-brane with 0.22 μm pore size (Sartorius,Italy) and incubated with 0.650 mmol·L−1
solutions of amino acid β-naphthylamide(βNA) derivates (Bachem FeinchemikalienAG, Switzerland) and 0.05 mol·L−1 phos-phate buffer pH 7.0 at 40 ◦C in differ-ent amounts for different times: Lys-βNA,Arg-βNA, Leu-βNA: 50 μL of dialyzed
suspension, 125 μL of βNA substrate,125 μL of phosphate buffer and 200 μLof bidistilled water for 1 h for the firsttwo substrates and 3 h for the third one;Pro-βNA: 300 μL of dialyzed suspension,100 μL of βNA substrate and 100 μLof phosphate buffer for 24 h; Glu-βNA:350 μL of dialyzed suspension, 75 μL ofβNA substrate and 75 μL of phosphatebuffer for 24 h; Phe-Pro-βNA: 50 μL ofdialyzed suspension, 125 μL of βNA sub-strate, and 325 μL of bidistilled water for1 h. The reaction was stopped by the ad-dition of 250 μL of 2.0 mol·L−1 HCl. Thedegree of hydrolysis was determined bymeasuring the colored product of an azo-copulation reaction by reading spectropho-tometrically at 580 nm (A580nm) accordingto Boquien et al. [1]. Each assay was car-ried out in duplicate and the average val-ues were calculated. The estimated errorwas less than 1.5%. The arbitrary unit ofenzyme activity (EA) was defined as thenumber of micromoles of β-naphthylamidereleased in one hour in one mL of eachsample.
3. RESULTS AND DISCUSSION
3.1. Chemical analyses
Organic acids are important indicatorsof biochemical metabolic processes occur-ring during the ripening of cheese. The re-sults obtained from the evaluation of or-ganic acids can be used as a reflection ofthe bacterial metabolism and fermentationwhich characterize PR cheese [27].
Table I shows that at the beginning ofthe cheesemaking, lactose and galactosewere totally consumed by thermophilicSLAB. At the same time, an expected in-crease in lactate was observed. Lactateproduced from lactose was in agreementwith other authors, who described it asthe most abundant organic acid in Parmi-giano Reggiano cheese, ranging from 1.30

516 J. De Dea Lindner et al.
Table I. pH and mean values in g·100 g−1 obtained after duplicate assays for the concentrationof lactose, galactose, L-lactate, D-lactate, lactate, citrate, succinate, acetic and pyroglutamic acidduring Parmigiano Reggiano manufacturing and ripening. Total lactate was obtained through thesum of L-lactate plus D-lactate.
pH Lactose Galactose L- D- Lactate Citrate Succinate Acetic PyroglutamicLactate Lactate acid acid
Milk 6.73 4.81 – – – – 0.16 – – –Whey starter 3.25 – – – – – – – – –Curd at vat 6.00 1.36 0.09 0.10 0.09 0.19 – – – –extractionCurd 6 h 5.75 0.40 0.39 0.22 0.20 0.43 – – – –Curd 12 h 5.57 0.15 0.56 0.24 0.24 0.48 – – – –Curd 48 h 4.36 0 0 0.47 0.58 1.05 0.05 0.03 0.04 –Salted cheese 5.35 0 0 0.77 0.61 1.38 – – – –(1 month)Cheese 2 months 5.36 0 0 0.73 0.57 1.30 – – – –Cheese 3 months 5.37 0 0 0.72 0.67 1.39 – – – –Cheese 4 months 5.36 0 0 0.85 0.60 1.45 – – – –Cheese 6 months 5.33 0 0 0.73 0.69 1.42 0.05 0.04 0.09 0.18Cheese 8 months 5.41 0 0 0.72 0.62 1.34 – – – –Cheese 10 months 5.37 0 0 0.74 0.63 1.37 – – – –Cheese 12 months 5.42 0 0 0.73 0.63 1.36 – – – –Cheese 16 months 5.41 0 0 0.69 0.63 1.32 – – – –Cheese 20 months 5.41 0 0 0.75 0.68 1.43 0.05 0.05 0.12 0.51–: Not determined.
to 1.45 g·100 g−1 [4, 28]. The concentra-tions of L- and D-lactic acid were approx-imately the same (L:D ratio approx. 1:1).In fact, both optical isomers are producedfrom lactose by the thermophilic LABcomponent of the natural starter. The val-ues for citrate and succinate are in agree-ment with the results described by Careriet al. [4], but disagree with Coppola et al.[8], who observed a gradual decrease incitrate concentration during aging. Dur-ing our PR cheesemaking, citrate was ap-parently not used as an energy source byfacultative heterofermentative LAB, eventhough many of them have the potentialto metabolize it as well when fermentablesugars are present [7]. Our findings suggestthat the growth of the microorganisms af-ter the start of the ripening was probablynot connected to their ability to utilize cit-rate as a source of energy. The acetic acid
values found are in agreement with previ-ous reports [4]. Pyroglutamic acid is re-lated to the age of PR [27]. The associ-ation of glutamic acid with pyroglutamicacid cyclase activity was demonstrated forthermophilic lactic species. The cycliza-tion of glutamic acid in pyroglutamic acidwas proposed by Mucchetti et al. [24] asa ripening marker in long-ripened cookedcheese. Our results for this organic acidsupport this hypothesis.
3.2. Microbial counts and growthtrends
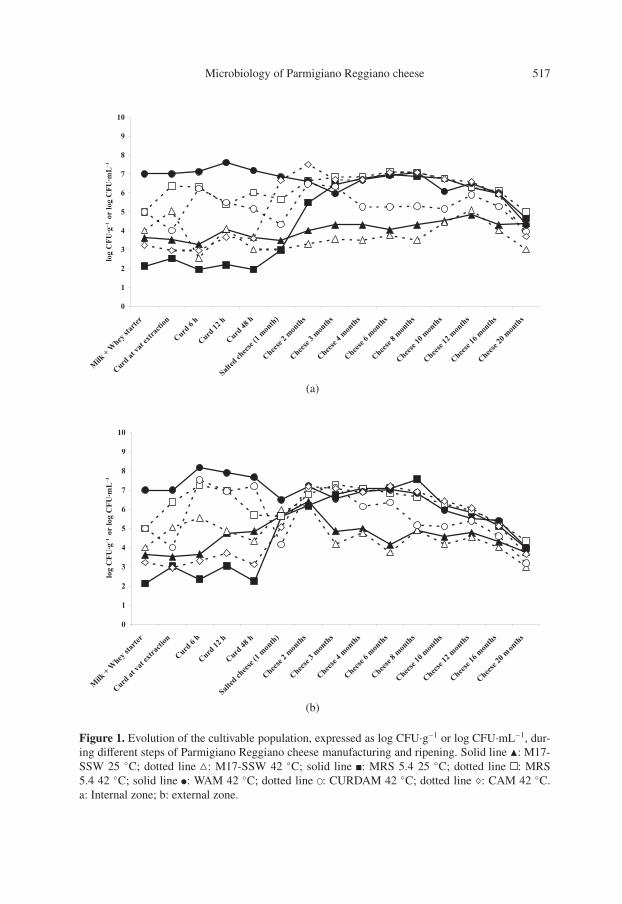
LAB cultivability was evaluated dur-ing PR manufacturing and ripening. Fig-ures 1a and 1b show the growth trends ofcultivable populations in MRS 5.4, M17-SSW, WAM, CURDAM and CAM forboth internal and external zones of the
Microbiology of Parmigiano Reggiano cheese 517
0
1
2
3
4
5
6
7
8
9
10
Milk
+ Whey
starte
r
Curd at
vat ex
tracti
on
Curd6 h
Curd 12
h
Curd 48
h
Salted
chees
e (1 mon
th)
Cheese 2
mon
ths
Cheese 3
months
Cheese 4
months
Cheese 6
months
Cheese 8 mon
ths
Cheese 10
mon
ths
Cheese 12
mon
ths
Cheese 1
6 mon
ths
Cheese 2
0 mon
ths
log
CFU
. g−1
or lo
g C
FU. m
L−1
(a)
0
1
2
3
4
5
6
7
8
9
10
Milk + W
hey st
arter
Curd at
vat ex
tracti
on
Curd6 h
Curd 12
h
Curd 48
h
Salted
chees
e (1 mon
th)
Cheese 2
mon
ths
Cheese 3
mon
ths
Cheese 4
months
Cheese 6
mon
ths
Cheese 8
months
Cheese 1
0 mon
ths
Cheese 12
mon
ths
Cheese 1
6 mon
ths
Cheese 2
0 months
log
CFU
. g−1
or lo
g C
FU. m
L−1
(b)
Figure 1. Evolution of the cultivable population, expressed as log CFU·g−1 or log CFU·mL−1, dur-ing different steps of Parmigiano Reggiano cheese manufacturing and ripening. Solid line �: M17-SSW 25 ◦C; dotted line : M17-SSW 42 ◦C; solid line : MRS 5.4 25 ◦C; dotted line : MRS5.4 42 ◦C; solid line : WAM 42 ◦C; dotted line : CURDAM 42 ◦C; dotted line ♦: CAM 42 ◦C.a: Internal zone; b: external zone.

518 J. De Dea Lindner et al.
cheeses, respectively. The microbiologi-cal counts for the whey starter were esti-mated as 7.7 × 104 colony-forming units(CFU)·mL−1 in M17-SSW incubated at42 ◦C, 1.0 × 107 CFU·mL−1 in MRS 5.442 ◦C, 3.4 × 108 CFU·mL−1 in WAM,1.0 × 107 CFU·mL−1 in CURDAM and10 CFU·mL−1 in CAM. The microbio-logical counts for milk were estimated as4.1 × 103 CFU·mL−1 in M17-SSW incu-bated at 25 ◦C, 7.8 × 103 CFU·mL−1 inM17-SSW 25 ◦C, 1.4 × 102 CFU·mL−1 inMRS 5.4 25 ◦C, 90 CFU·mL−1 in MRS5.4 42 ◦C, 2.7 × 102 CFU·mL−1 in WAM,1.5 × 103 CFU·mL−1 in CURDAM and1.7 × 103 CFU·mL−1 in CAM.
Figures 1a and 1b show that the totalmicrobial cultivable population remainedhigh and similar for the first six months.After six months, a progressive decreasewas observed. The evolution of the micro-bial counts in the five growth media wasdifferent for the first 48 h, and split intotwo groups after four months: the highestcounts were in WAM, MRS 5.4 42 ◦C andCAM; the lowest in M17-SSW 25 ◦C and42 ◦C. In particular, when three monthsof ripening were reached in the internaland external zones, the values of microbialcounts on MRS 5.4, WAM and CAM resultoverlapped and remained comparable un-til the end of ripening. After 9–10 monthsof ripening a decrease in the number ofcultivable bacteria was observed. Countsin CURDAM were low (in the order of10−1 to 10−2), and the lowest counts wereobserved for the streptococcal and lacto-coccal populations, evaluated in M17-SSWagar, respectively, at 42 and 25 ◦C. We hy-pothesize that the majority of the micro-bial populations found in cheese after fourmonths of ripening were not able to me-tabolize lactose, present in M17-SSW andCURDAM media as a carbon source.
Microbial counts in WAM confirmedthat after the addition of whey starter, be-cause of the temperatures used during pro-duction in the first 48 h, the thermophilic
microflora (SLAB) became dominant in re-spect to the mesophilic microflora (Figs. 1aand 1b). In this phase, the nutritionally de-manding microorganisms found in WAMhad the best conditions for growth. WAMattempts to reproduce the natural compo-sition of the whey; it supports the origi-nal population from the whey starter andcan be considered the growth medium thatbest suits their growth. Different microbialcounts were detected using WAM for theinternal and external 6-h curd, suggestingthat the early decrease in temperature in theexternal zone of the wheel could have fa-vored the growth of SLAB.
Microorganisms cultivated at 25 ◦C inMRS 5.4 and at 42 ◦C in CAM were un-able to grow in the first 48 h. However, inthe advanced phase of ripening (Figs. 1aand 1b), they demonstrated the ability touse alternative potential energy sourcesand also to metabolize carbon sourcesother than lactose and citrate. These mi-croflora were probably NSLAB originat-ing from milk, as observed in other hardcooked cheeses such as Swiss Gruyère[5]. Generally, only a few strains survivethe processing conditions and grow duringripening [5].
The number of SLAB commonly ex-ceeds 109 CFU·g−1 in many young semi-hard and hard ripened cheeses [2]. At thesame time, NSLAB populations occur innumbers of about 102 CFU·g−1 and be-gin to grow and eventually plateau at celldensities of 107–109 CFU·g−1 after sev-eral months of aging [2]. Using the platecount agar technique, we found similar val-ues only for NSLAB, whereas the value ofSLAB, at the beginning of ripening, wasmore than 1 log less than the populationgenerally found [2]. This difference maybe due to the intrinsic limits of traditionalagar-based and culture-dependent methodsapplied to estimating the SLAB arisingfrom a natural whey starter [16]. More-over, we found that in advanced phases ofPR ripening (over 12 months), presumptive

Microbiology of Parmigiano Reggiano cheese 519
0
1
2
3
4
5
6
7
8
9
10
Curd at
vat ex
tracti
on
Curd 6
h
Curd 12
h
Curd 48
h
Salted
chees
e (1 m
onth)
Cheese 2
mon
ths
Cheese 3 mon
ths
Cheese 4 mon
ths
Cheese 6 m
onths
Cheese 8
mon
ths
Cheese 1
0 months
Cheese 1
2 mon
ths
Cheese 1
6 mon
ths
Cheese 2
0 months
log
cells
. g-1
Figure 2. Evolution of the viable �, not viable � and total population , expressed as log cells·g−1, ininternal (dotted line) and external (solid line) zones, during different steps of Parmigiano Reggianocheese manufacturing and ripening.
mesophilic lactobacilli counts were higherthan presumptive lactococci in both cheeseportions.
3.3. Cell viability counts
Cell viability counts using a BacterialViability Kit allowed the determinationof the viable (undamaged cell membrane)and the unviable (damaged cell membrane)bacteria. The sum of viable and unviablecounts gives an indication of the directtotal population. Total microbial countsusing the cell viability method gives ad-ditional information compared with classi-cal plate counts, where there is the influ-ence of adaptability, choice of nutritionalsubstrate and incubation parameters. Fig-ures 1 and 2 show that cultivable popu-lations were always lower than total di-rect population counts, especially at thebeginning of cheese production. Duringripening, the culture medium CAM sup-ported the recovery of the total cultivable
population, as demonstrated by an overlapof both plate and direct count results. In thefirst 12 h of manufacture, viable, unviableand total populations were higher in the ex-ternal portion of cheese than in the internalportion (Fig. 2). In particular, viable cellsin the external portion of the curd were onelogarithm higher than those in the internalportion.
The assessment of total counts high-lighted the induction of bacterial lysis.During aging, the conditions prevailing inthe cheese matrix limit the preservation ofcell viability [2]. The absence of residuallactose and galactose, high levels of NaCl,low pH and the drop in temperature con-tribute to cell autolysis, which results in therelease of intracellular enzymes and othercomponents into the cheese matrix. In theone-month cheese sample, a reduction inthe number of total cells was observed bothin the external and internal portions. Inter-estingly, between the first and the secondmonths an increase in the total population,
520 J. De Dea Lindner et al.
0
500
1000
1500
2000
2500
Curd 6
h
Curd 12
h
Curd 48
h
Salted
chees
e (1mon
th)
Cheese
2 mon
ths
Cheese
3 mon
ths
Cheese
4 mon
ths
Cheese
6 mon
ths
Cheese
8 mon
ths
Cheese
10 m
onths
Cheese
12mon
ths
Cheese
16 m
onths
Cheese
20mon
ths
EA
012345678910
log
cells
. mL
-1
(a)
0
500
1000
1500
2000
2500
Curd 6
h
Curd 12
h
Curd 48
h
Salte
d chees
e (1 m
onth)
Cheese
2 mon
ths
Cheese
3 mon
ths
Cheese
4 mon
ths
Cheese
6 mon
ths
Cheese
8 mon
ths
Cheese
10 m
onths
Cheese
12 m
onths
Cheese
16 m
onths
Cheese
20 m
onths
EA
012345678910
log
cells
. mL
-1
(b)
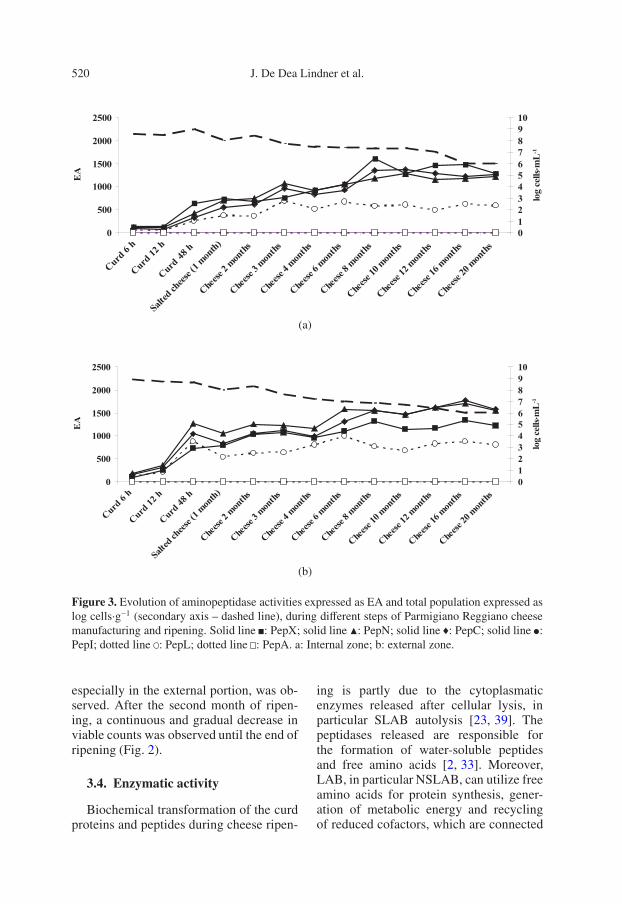
Figure 3. Evolution of aminopeptidase activities expressed as EA and total population expressed aslog cells·g−1 (secondary axis – dashed line), during different steps of Parmigiano Reggiano cheesemanufacturing and ripening. Solid line : PepX; solid line �: PepN; solid line : PepC; solid line :PepI; dotted line : PepL; dotted line : PepA. a: Internal zone; b: external zone.
especially in the external portion, was ob-served. After the second month of ripen-ing, a continuous and gradual decrease inviable counts was observed until the end ofripening (Fig. 2).
3.4. Enzymatic activity
Biochemical transformation of the curdproteins and peptides during cheese ripen-
ing is partly due to the cytoplasmaticenzymes released after cellular lysis, inparticular SLAB autolysis [23, 39]. Thepeptidases released are responsible forthe formation of water-soluble peptidesand free amino acids [2, 33]. Moreover,LAB, in particular NSLAB, can utilize freeamino acids for protein synthesis, gener-ation of metabolic energy and recyclingof reduced cofactors, which are connected

Microbiology of Parmigiano Reggiano cheese 521
with cheese flavor formation [6, 36, 41].Amino acid catabolism involves a widerange of both general and specific reg-ulators and shows significant differencesamong LAB [11].
The aminopeptidase activities tested inthis work were those frequently foundin LAB and capable of hydrolyzing thefollowing six different substrates: broad-specificity aminopeptidase N (PepN) andC (PepC) activities were determined us-ing Lys-βNA and Arg-βNA, respectively,proline aminopeptidase (PepI) activity us-ing Pro-βNA, glutamyl aminopeptidase A(PepA) activity using Glu-βNA, pepti-dase with high specificity for leucine andalanine (PepL) using Leu-βNA, and activ-ity of the X-prolyl dipeptidyl aminopep-tidase (PepX) was determined using Phe-Pro-βNA [6, 33]. The enzymatic activityvalues detected in the cheese extract freefrom cells are shown in Figures 3a and 3bfor the internal and external zones of thecheese. These results reflect an enzymaticactivity due only to cytoplasmatic enzymesreleased after bacterial lysis.
PepC, PepN, PepX and PepL activitieswere higher in the external zone of cheese(Fig. 3b) than in the internal zone (Fig. 3a),especially during the first six months ofaging. In both zones, PepA and PepI ac-tivities were very low, in contrast to thehigh quantity of proline and glutamic acidusually found in free form at the end ofripening [4, 31]. In general, glutamic acidis the main amino acid detected in cheesesproduced with a thermophilic starter andcooked with a temperature gradient suchas PR [4], Swiss cheese [30], Gruyère [3]and Emmental and Bergkäse [20]. This ap-parent contradiction might be explainedin different ways: (i) Glu can be liber-ated by broad-specificity aminopeptidaselike PepC [6]; (ii) the hydrolytic activ-ity towards Glu and Pro is low but al-ways present during ripening; (iii) Glu andPro are highly present in milk proteins[37]; and (iv) Pro is not further converted
into degraded molecules and gradually in-creases in cheese [11].
A significant increase in EA was de-tected in the first 48 h of manufacture.These results are in agreement with the de-crease in total counts observed, in partic-ular in the cheese internal zone. A secondstage of EA increase was observed for theinternal zone during the 6th and 8th months(Fig. 3a) and for the external zone duringthe 4th and 6th months (Fig. 3b). Over the20 months of ripening, similar quantitativeascending trends in the hydrolysis of spe-cific substrates were observed for the inter-nal and external zones of the cheese.
4. CONCLUSION
The Parmigiano Reggiano cheese ripen-ing process is a very complex biochemicalevent. Technological parameters determinethe specific microbial selection in the dif-ferent stages of cheesemaking and ripen-ing and even in the different cheese zones.Our results show that the microbial popula-tions present in curd and in cheese can playdifferent roles in the subsequent phases ofripening and can interact by overlappingtheir metabolic pathways. For example, theSLAB are involved not only in lactose de-pletion and curd acidification, but also inthe production of a considerable amount ofintracellular aminopeptidases released as aconsequence of bacterial lysis, which areactive during the whole ripening.
Regarding NSLAB, they remain viablefor a long time during cheese ripening.They seem to adapt better to cheese thanSLAB and are able to survive and groweven in the absence of lactose. The tech-nological role of this secondary microflorashould be investigated in greater depth.
The microbial dynamics, the iden-tification of all microbial species, theinfraspecies biodiversity and their interac-tions during PR cheesemaking and ripen-ing are not still completely clarified.

522 J. De Dea Lindner et al.
This gap should be filled to understand bet-ter and protect the origin, the tradition andthe quality of this PDO cheese.
Acknowledgements: This work was partiallysupported by the Emilia Romagna Region(LR28/98), Bologna, Italy, Project “SMEPRStudio e modellazione degli aspetti enzi-matici legati alla stagionatura del formag-gio Parmigiano-Reggiano”. The authors thankthe Consorzio Parmigiano Reggiano, ReggioEmilia, Italy and CRPA Spa, Reggio Emilia,Italy, for their organizational and technical con-tributions. The authors are grateful to Dr. R.Ghiglietti (CRA-FLC Lodi, Italy) for the or-ganic acid determination.
REFERENCES
[1] Boquien C.Y., Corrieu G., Desmazeaud M.J.,Enzymatic methods for determining popu-lations of Streptococcus cremoris AM2 andLeuconostoc lactis CNRZ 1091 in pure andmixed cultures, Appl. Microbiol. Biotech. 30(1989) 402–407.
[2] Broadbent J.R., Steele J.L., Cheese flavorand the genomics of lactic acid bacteria,ASM News 71 (2005) 121–125.
[3] Bütikofer U., Fuchs D., Development of freeamino acids in Appenzeller, Emmentaler,Gruyère, Raclette, Sbrinz and Tilsiter cheese,Lait 77 (1997) 91–100.
[4] Careri M., Spagnoli S., Panari G., ZannoniM., Barbieri G., Chemical parameters of thenon-volatile fraction of ripened Parmigiano-Reggiano cheese, Int. Dairy J. 6 (1996) 147–155.
[5] Casey M.G., Häni J.P., Gruskovnjak J.,Schaeren W., Wechsler D., Characterisationof the non-starter lactic acid bacteria(NSLAB) of Gruyère PDO cheese, Lait 86(2006) 407–414.
[6] Christensen J.E., Dudley E.G., PedersonJ.A., Steel J.L., Peptidases and amino acidcatabolism in lactic acid bacteria, Antonievan Leeuwenhoek 76 (1999) 217–249.
[7] Cogan T.M., Beresford T.P., Steele J.,Broadbent J., Shah N.P., Ustunol Z., Invitedreview: Advances in starter cultures and cul-tured foods, J. Dairy Sci. 90 (2007) 4005–4021.
[8] Coppola R., Nanni M., Iorizzo M.,Sorrentino A., Sorrentino E., Chiavari C.,Grazia L., Microbiological characteristicsof Parmigiano Reggiano cheese during the
cheesemaking and the first months of theripening, Lait 80 (2000) 479–490.
[9] Coppola R., Nanni M., Iorizzo M.,Sorrentino A., Sorrentino E., Grazia L.,Survey of lactic acid bacteria isolatedduring the advance stages of the ripening ofParmigiano Reggiano cheese, J. Dairy Res.64 (1997) 305–310.
[10] De Dea Lindner J., Traditional and innova-tive approaches to evaluate microbial contri-bution in long ripened fermented foods: thecase of Parmigiano Reggiano cheese, Ph.D.thesis, University of Parma, Italy, 2008.
[11] Fernández M., Zúñiga M., Amino acidcatabolic pathways of lactic acid bacteria,Crit. Rev. Microbiol. 32 (2006) 155–183.
[12] Fornasari M.E., Rossetti L., Carminati D.,Giraffa G., Cultivability of Streptococcusthermophilus in Grana Padano cheese wheystarters, FEMS Microbiol. Lett. 257 (2006)139–144.
[13] Fox P.F., Wallace J.M., Formation of flavourcompounds in cheese, Adv. Appl. Microbiol.45 (1997) 17–85.
[14] Fox P.F., Wallace J.M., Morgan S., LynchC.M., Niland E.J., Tobin J., Acceleration ofcheese ripening, Antonie van Leeuwenhoek70 (1996) 271–297.
[15] Gala E., Landi S., Solieri L., Nocetti M.,Pulvirenti A., Giudici P., Diversity of lac-tic acid bacteria population in ripenedParmigiano Reggiano cheese, Int. J. FoodMicrobiol. 125 (2008) 347–351.
[16] Gatti M., Bernini V., Lazzi C., Neviani E.,Fluorescence microscopy for studying the vi-ability of micro-organisms in natural wheystarters, Lett. Appl. Microbiol 42 (2006)338–343.
[17] Gatti M., Fornasari M.E., Mucchetti G.,Addeo F., Neviani E., Presence of pepti-dase activities in different varieties of cheese,Lett. Appl. Microbiol. 28 (1999) 368–372.
[18] Gatti M., Lazzi C., Rossetti L., Mucchetti G.,Neviani, E., Biodiversity in Lactobacillushelveticus strains present in natural wheystarter used for Parmigiano Reggiano cheese,J. Appl. Microbiol. 95 (2003) 463–470.
[19] Hannon J.A., Kilcawley K.N., WilkinsonM.G., Delahunty C.M., Beresford T.P.,Flavour precursor development in Cheddarcheese due to lactococcal starters and thepresence and lysis of Lactobacillus helveti-cus, Int. Dairy J. 17 (2007) 316–327.
[20] Krause I., Bockhardt A., Klostermeyer H.,Characterization of cheese ripening by freeamino acids and biogenic amines and influ-ence of bactofugation and heat-treatment ofmilk, Lait 77 (1997) 101–108.

Microbiology of Parmigiano Reggiano cheese 523
[21] Kunji E.R.S., Mierau I., Hagting A.,Poolman B., Konings W.N., The proteolyticsystem of lactic acid bacteria, Antonie VanLeeuwenhoek 70 (1996) 187–221.
[22] Lazzi C., Gatti M., Bernini V., De DeaLindner J., Neviani E., Impiego di nuovi ter-reni colturali a base di cagliata e di formag-gio per il recupero e la differenziazione dellamicroflora caratteristica di formaggi a lungastagionatura, Sci. Tecn. Latt.-Cas. 58 (2007)55–69.
[23] Lortal S., Chapot-Chartier M.P., Role, mech-anisms and control of lactic acid bacteria ly-sis in cheese, Int. Dairy J. 15 (2005) 857–871.
[24] Mucchetti G., Locci F., Gatti M., Neviani E.,Addeo F., Dossena A., Marchelli R.,Pyroglutamic acid in cheese: presence,origin, and correlation with ripening timeof Grana Padano cheese, J. Dairy Sci. 83(2000) 659–665.
[25] Mucchetti G., Neviani E., Microbiologiae tecnologia lattiero-casearia. Qualità e si-curezza, Tecniche nuove, Milano, Italy,2006.
[26] O’Cuinn G., Jennigs P.V., Fhaolain I.F.,Booth M., Bacon C.L., McDonnel M.,Wilkinson M., O’Callaghan M.D., FitzgeraldR.J., The contribution of the starter pep-tidases to flavour development in cheese,in: Cogan T.M., Fox P.F., Ross P. (Eds.),Proceedings of the 4th Cheese Symposium,Teagasc, University of Cork, Ireland, 1995,pp. 68–71.
[27] Panari G., L’acido lattico e l’acido piro-glutammico nella maturazione del formaggioParmigiano-Reggiano, Sci. Tecn. Latt.-Cas.36 (1985) 98–109.
[28] Panari G., Reverberi P., Caroli A., NocettiM., Pecorari M., Le variazione del pro-filo microbiologico del latte durantel’affioramento in diverse condizioni op-erative, Sci. Tecn. Latt.-Cas. 58 (2007)83–93.
[29] Pereira C.I., Gomes E.O., Gomes A.M.P.,Malcata F.X., Proteolysis in modelPortuguese cheeses: effects of rennetand starter culture, Food Chem. 108 (2007)862–868.
[30] Preininger M., Warmke R., Grosch W.,Identification of the character impact flavourcompounds of Swiss cheese by sensory stud-ies of models, Z. Lebensm. Unters.-Forsch.202 (1996) 30–34.
[31] Resmini P., Pellegrino L., Pazzaglia C.,Hogenboom J.A., Gli amminoacidi
liberi nella tipizzazione del formaggioParmigiano-Reggiano ed in particolare nelprodotto grattugiato, Sci. Tecn. Latt.-Cas. 37(1985) 557–592.
[32] Sandri S., Tosi F., Fossa E., Scotti C.,Malacarne M., Andamento dei valori di pHmisurati al centro della forma nelle prime oresuccessive alla fabbricazione del ParmigianoReggiano: influenza di alcuni fattori tecno-logici, Sci. Tecn. Latt.-Cas. 58 (2007) 17–28.
[33] Savijoki K., Ingmer H., Varmanen P.,Proteolytic system of lactic acid bacteria,Appl. Microbiol. Biotechnol. 71 (2006) 394–406.
[34] Sheehan A., O’Cuinn G., FitzGerald R.J.,Wilkinson M.G., Proteolytic enzyme ac-tivities in Cheddar cheese juice made us-ing lactococcal starters of differing autolyticproperties, J. Appl. Microbiol. 100 (2006)893–901.
[35] Sheehan J.J., Fenelon M.A., WilkinsonM.G., McSweeney P.L.H., Effect of cooktemperature on starter and non-starter lacticacid bacteria viability, cheese compositionand ripening indices of a semi-hard cheesemanufactured using thermophilic cultures,Int. Dairy J. 17 (2007) 704–716.
[36] Smit G., Smit B.A., Engels W.J.M., Flavourformation by lactic acid bacteria and bio-chemical flavour profiling of cheese prod-ucts, FEMS Microbiol Rev. 29 (2005) 591–610.
[37] Swaisgood H.E., Chemistry of milk proteins,in: Fox P.F. (Ed.), Developments in dairychemistry, Vol. 1: Proteins, Elsevier AppliedScience, London, UK, 1982, pp. 1–59.
[38] Takafuji S., Iwasaki T., Sasaki M.,Tan P.S.T., Proteolytic enzymes of lac-tic acid bacteria, in: Charalambous G.(Ed.), Food Flavors: Generation, Analysisand Process Influence, Elsevier ScienceB.V., Amsterdam, The Netherlands, 1995,pp. 753–767.
[39] Valence F., Deutsch S.M., Richoux R.,Gagnaire V., Lortal S., Autolysis and re-lated proteolysis in Swiss cheese for twoLactobacillus helveticus strains, J. DairyRes. 67 (2000) 261–271.
[40] Wilkinson M.G., Guinee T.P., O’CallaghanD.M., Fox P.F., Autolysis and proteolysisin different strains of starter bacteria duringCheddar cheese ripening, J. Dairy Res. 61(1994) 249–262.
[41] Yvon M., Rijnen L., Cheese flavour forma-tion by amino acid catabolism, Int. Dairy J.11 (2001) 185–201.
Top Related