







Languages
Pages
Legal


[CANCER RESEARCH 50. 103-107. January I. I990|
Insulin-like Growth Factor I and Transforming Growth Factor a as AutocrineGrowth Factors in Human Pancreatic Cancer Cell Growth1
Eiji <Minium, Masaki Okada, INoritaka Onoda, Y'oshinobu Kamiya, Hitomi Murakami, Toshio Tsushima, and
Kazuo ShizumeDepartment of Internal Medicine, Institute of Clinical Endocrinology, Tokyo Women's Medical College, 8-1 Kawada-cho ¡E.O., N. O., Y. A'., H. M., T. T., K. S.J, and
Research Institute for (irotrth Sciences ¡M.O./, Shinjuku-ku, Tokyo, 162 Japan
ABSTRACT
The roles of insulin-like growth factor I (K.I -I) and transforminggrowth factor a (TCF-a) as autocrine factors in the proliferation of \11 VPaCa 2 cells (human pancreatic cancer cells, PC cells) were investigated.Furthermore, the mechanism(s) of inhibition of PC cell growth by aphorbol ester in relation to these two kinds of growth factor was alsostudied.
PC cells grew autonomously when Dulbecco's modified essential me
dium supplemented with 4% fetal calf serum was changed to serum-freemedium (0.3% bovine serum albumin-Dulbecco's modified essential me
dium). In addition, serum-free conditioned medium from PC cells dialyzedagainst fresh Dulbecco's modified essential medium had a stimulatory
action on the growth of the same kind of cells when compared with thatinduced by nonconditioncd medium. These observations suggest that afactor(s) produced and released by PC cells stimulates their own growth.Analysis of conditioned medium from PC cells revealed the presence ofimmunoreactive (IR)-IGF-I and IR-TGF-a. The molecular size of IR-IGF-I was similar to that of authentic K.I -I. On the other hand, IR-TGF-a was present as multiple forms when analyzed using gel chroma-tography. Authentic IGF-I and TGF-a added to culture medium stimulated PC cell growth by 1.45- and 1.5-fold above control value, respectively. A monoclonal antibody to IGF-I receptor was able to inhibit PCcell growth.
PC cell proliferation was markedly inhibited by 12-O-tetradecanoyl-13-acetate (>0.16 nisi), whereas cell growth of human fibroblasts wasstimulated by it. 12-0-Tetradecanoyl-phorbol-13-acetate also reducedthe binding of 12*I-TGF-a, but not 12'I-IGF-I, to PC cells. Decrease inTGF-a binding was mainly due to the reduced affinity of receptors to theligand.
These results suggest that IGF-I and TGF-a are involved in PC cellproliferation as autocrinc factors. Further, the inhibition of PC cellgrowth by phorbol ester could be, at least partly, due to the decreasedbinding of TGF-a to the cells.
INTRODUCTION
Because of a diminished, or complete lack of, requirementfor serum or specific growth factors, neoplastic cells in cultureare presumed to have autonomous activating pathways in theirgrowth system, namely: (a) autonomous production of growthfactors; (b) synthesis of an altered receptor for growth factors;and (c) activation of postreceptor pathway (1-3).
Many types of tumor cells grown in culture secrete a varietyof peptide growth factors into their culture medium, and thesame cells usually have functional receptors for such growthfactors (2, 3). For example, a recent study has shown that mostmalignant cells derived from solid tumors synthesize TGF-«2and all such cells express mRNA for EGF/TGF-« receptor (4).
Received 6/13/89; revised 9/26/89; accepted 10/3/89.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1The present work was supported by grants from the Ministry of Education,
the Ministry of Health and Welfare, and the Foundation for Growth Science.2The abbreviations used are: TGF, transforming growth factor; EGF, epider
mal growth factor; IGF, insulin-like growth factor; PC cell. MIA-PaCa 2 humanpancreatic carcinoma cell; DM EM. Dulbecco's modified essential medium; FCS,fetal calf serum; BSA, bovine serum albumin; TPA, 12-O-tetradecanoy Iphorbol-13-acetate: CM, conditioned medium; IR, immunoreactive; PK-C protein kinase C.
In addition, growth factors such as IGF-I, TGF-«, TGF-/3,platelet-derived growth factor, and bombesin have been reported to act on cell growth in an autocrine manner in neoplastic and nonneoplastic cells (2, 3, 5-11). Furthermore, a specificmonoclonal antibody against a receptor for a growth factor cansuppress the cell growth, which is consistent with the autocrinetheory in tumor growth (12, 13).
The role of growth factors in the development of pancreaticcancer has been reported by several investigators (14-17). Chester et al. (14) have shown that the incidence of pancreatic cancerinduced by A'-nitrosobis(2-oxopropyl)amine was increased by
EGF in Syrian hamsters. EGF has been shown to promotephosphorylation in cultured pancreatic cancer cells, MIA PaCa-2 (15). Moreover, over-expression of EGF receptor has beenfound in pancreatic cancer cells ( 16). Recently, Smith etal.(\l)revealed that mRNA of TGF-a, which is thought to act throughthe EGF receptor, is expressed in several kinds of pancreaticcancer cells.
In the present study, we demonstrate that IGF-I and TGF-«stimulate pancreatic cancer cell growth, presumably in an autocrine manner. In addition, we show that phorbol ester, aprotein kinase C activator (18), has a potent inhibitory activityon the cell growth. The mechanism(s) of this inhibitory actionin terms of the above two growth factors will be discussed.
MATERIALS AND METHODS
Cell Culture. MIA-PaCa 2 human pancreatic carcinoma cells wereobtained from the American Type Culture Collection (Rockville, MD)and maintained by once weekly passage in DM EM (Gibco, GrandIsland, NY) containing 4% FCS (Filtron, Australia) and antibiotics(Gibco). Human fibroblast cultures were established from the forearmskin of normal adults and were grown in RPMI (Gibco) supplementedwith 10% FCS and antibiotics. The cells were cultured at 37°Cin 5%
CO2/95% humidified air atmosphere.Proliferation Study. PC cells or human fibroblasts were seeded into
12-well plates (Costar, MA) at the density of 1.5-2.0 x IO4cells/well.
Cultured cells were usually used after growing for 3 days in 4% FCS/DMEM. The cells were washed with serum-free DMEM and proliferation studies were performed in 0.3% BSA/DMEM. The test materials,dissolved with 0.3% BSA/DMEM, were added at the initiation of theexperiment. Media and materials were changed every 2-3 days. At theend of the experiments, cells isolated by trypsinizing were countedusing a Coulter Counter (Coulter Electronics, Inc.) Recombinant IGF-I and TCP-« were kindly supplied by Fujisawa Pharmaceutical Co.(Osaka, Japan) and Earth Chemical Co. (Hyogo, Japan). TPA wasobtained from Sigma Chemical Co. (St. Louis, MO).
The mean ±SD was calculated for each group and significance wasdetermined by the Student's / test.
Binding Studies. Confluent cells cultured in nmlii\\cll (12 wells)plates were used for the binding studies. The cells were washed oncewith ice-cold 0.3% BSA/DMEM supplemented with 20 mM 4-(2-hydroxyethyl)-l-piperazineethanesulfonicacid (pH 7.4) and then incubated at 4°Cfor 24 h in the same medium with or without peptides.
Labeled ligands (40,000-50,000 cpm/well) were added and incubatedat 4°Cfor 20 h. Reaction was stopped by removing the medium and
washing the cells twice with the ice-cold fresh medium. PC cells were
103
Research. on February 5, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from
GROWTH FACTORS AND HUMAN PANCREATIC CANCER CELL GROWTH
0 2 4 6 8 10 12DAYS AFTER PLATING
10
5
10
05
01-
Lo 5 7 9 n 13
DAYS AFTER PLATING
Fig. I. A, PC cell growth in two different concentrations (2% and 10%) ofFCS. B, PC cell growth in serum-free medium. Initially, cells were cultured in4% FCS/DMEM; then medium was changed to 0.1% or 0.5% BSA/DMEM.Medium was changed every 2 days. Values are means ±SD (bars) of threedeterminations.
then solubili/ed with 0.5 ml l N NaOH and transferred to tubes forcounting radioactivity.
Collection and Extraction of Conditioned Medium. PC cells weregrown in flasks to confluence in 4% FCS/DMEM. Cells were washedonce with serum-free DMEM. Following 48 h of culture of these cellsin 0.3% BSA/DMEM, CM was collected and centrifuged at 500 x gfor 5 min. The supernatant was dialyzed (molecular weight cutoff,3500) against distilled water for 48 h at 4°C.Freeze-dried CM prepa
rations were treated with I Macetic acid for 20 h at 4 ( and centrifugedat 2000 x g for IO min. The supernatant was neutralized when necessaryand analyzed for the growth factors of interest.
Radioimmunoassay for IGF-I or IGF-a. Antibodies against synthetichuman TGF-« (Sumitomo Pharmaceutical Co., Osaka, Japan) andrecombinant IGF-I (Fujisawa) were obtained by immunizing rabbitswith peptides conjugated with thyroglobulin. Antibodies to IGF-I andTGF-a were used at final dilution of 1:50,000. Human IGF-I and ratTGF-a (Peninsula, CA) labeled with Na'"I (NEN, Boston, MA) by thechloramine-T method were used as tracers. The sensitivity in the IGF-I assay system was 20 pg/tube and the 50% inhibitory dose was almost0.2 ng/tube. IGF-II cross-reacted with this antibody at approximately0.1%. In the TGF-a radioimmunoassay system, the sensitivity was 0.3ng/tube, and 50% inhibitory dose was about 3 ng/tube. Cross-reactionof EGF in this assay system was less than 0.01%.
RESULTS
Growth of PC Cells in Medium Supplemented with FCS orBSA. Fig. \A shows the growth of PC cells in 2% and 10%FCS/DMEM. The doubling time was about 24 h at the twoconcentrations. When FCS/DMEM was changed to 0.1% or0.3% BSA/DMEM on day 5, the growth rate of PC cells becameslower, but the final cell density on day 13 was comparable withthat obtained in the presence of 5% FCS (Fig. \B).
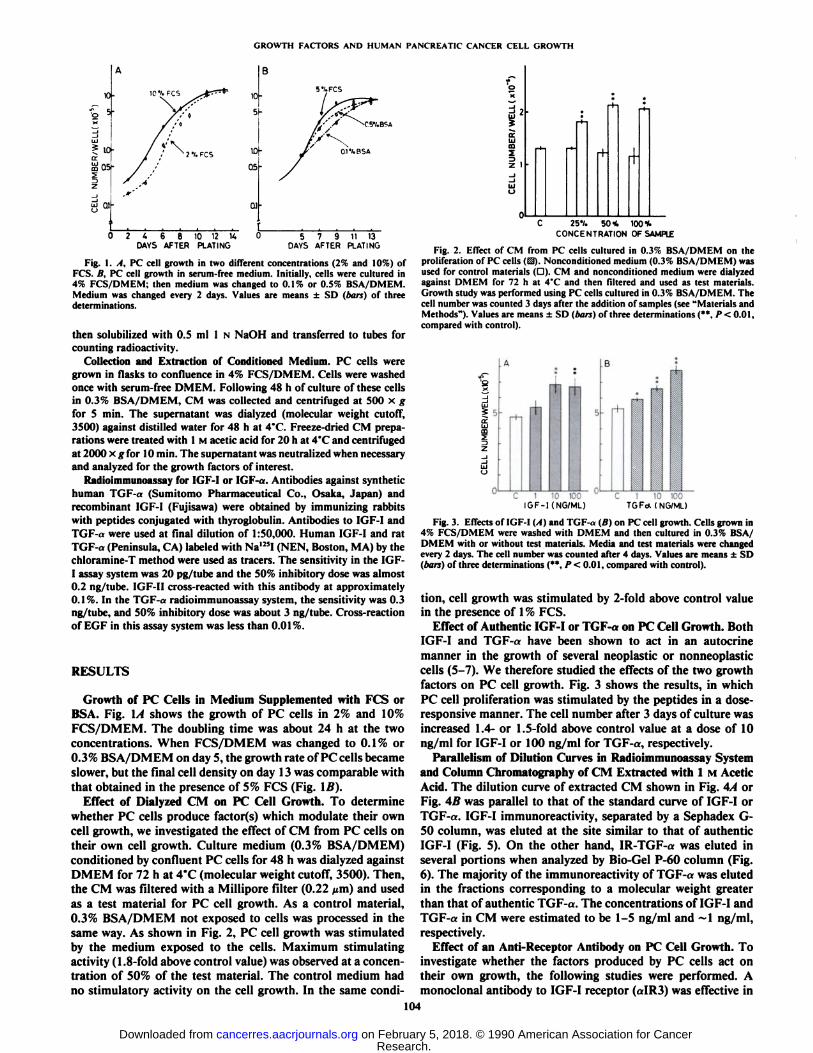
Effect of Dialyzed CM on PC Cell Growth. To determinewhether PC cells produce factor(s) which modulate their owncell growth, we investigated the effect of CM from PC cells ontheir own cell growth. Culture medium (0.3% BSA/DMEM)conditioned by confluent PC cells for 48 h was dialyzed againstDMEM for 72 h at 4°C(molecular weight cutoff, 3500). Then,
the CM was filtered with a Millipore filter (0.22 ^m) and usedas a test material for PC cell growth. As a control material,0.3% BSA/DMEM not exposed to cells was processed in thesame way. As shown in Fig. 2, PC cell growth was stimulatedby the medium exposed to the cells. Maximum stimulatingactivity (1.8-fold above control value) was observed at a concentration of 50% of the test material. The control medium hadno stimulatory activity on the cell growth. In the same condi-
UJce
i. 1+ rf
C 25'/. 50* 100*
CONCENTRATION OF SAMPLE
Fig. 2. Effect of CM from PC cells cultured in 0.3% BSA/DMEM on theproliferation of PC cells (13). Nonconditioned medium (0.3% BSA/DMEM) wasused for control materials (D). CM and noncondilioned medium were dialyzedagainst DMEM for 72 h at 4°Cand then filtered and used as test materials.
Growth study was performed using PC cells cultured in 0.3% BSA/DMEM. Thecell number was counted 3 days after the addition of samples (see "Materials andMethods"). Values are means ±SD (bars) of three determinations (**. P< 0.01.
compared with control).
«r-B
A**1
_LTTC
1 T•T.B5-ñlri•00" C!IGF-KNG/ML)
TGF**„L.111
%10
100(NG/ML
Fig. 3. Effects of IGF-I (A) and TGF-«(B) on PC cell growth. Cells grown in4% FCS/DMEM were washed with DMEM and then cultured in 0.3% BSA/DMEM with or without test materials. Media and test materials were changedevery 2 days. The cell number was counted after 4 days. Values are means ±SD(bars) of three determinations (**, P< 0.01, compared with control).
tion, cell growth was stimulated by 2-fold above control valuein the presence of 1% FCS.
Effect of Authentic IGF-I or TGF-a on PC Cell Growth. BothIGF-I and TGF-a have been shown to act in an autocrinemanner in the growth of several neoplastic or nonneoplasticcells (5-7). We therefore studied the effects of the two growthfactors on PC cell growth. Fig. 3 shows the results, in whichPC cell proliferation was stimulated by the peptides in a dose-responsive manner. The cell number after 3 days of culture wasincreased 1.4- or 1.5-fold above control value at a dose of 10ng/ml for IGF-I or 100 ng/ml for TGF-a, respectively.
Parallelism of Dilution Curves in Radioimmunoassay Systemand Column Chromatography of CM Extracted with l M AceticAcid. The dilution curve of extracted CM shown in Fig. 4A orFig. 4B was parallel to that of the standard curve of IGF-I orTGF-a. IGF-I immunoreactivity, separated by a Sephadex G-50 column, was eluted at the site similar to that of authenticIGF-I (Fig. 5). On the other hand, IR-TGF-a was eluted inseveral portions when analyzed by Bio-Gel P-60 column (Fig.6). The majority of the immunoreactivity of TGF-«was elutedin the fractions corresponding to a molecular weight greaterthan that of authentic TGF-a. The concentrations of IGF-I andTGF-a in CM were estimated to be 1-5 ng/ml and ~1 ng/ml,respectively.
Effect of an Anti-Receptor Antibody on PC Cell Growth. Toinvestigate whether the factors produced by PC cells act ontheir own growth, the following studies were performed. Amonoclonal antibody to IGF-I receptor («IR3)was effective in
104
Research. on February 5, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from

GROWTH FACTORS AND HUMAN PANCREATIC CANCER CELL GROWTH
;o5-
0039 0156 0625IGF-1 (NO/TUBE)
25
05-
0312 125 5 20TGF*(NG/TUBE)
Fig. 4. Dilution curves of CM extracted by l M acetic acid in IGF-I (A) andTGF-a (A) radioimmunoassay systems. CM dialyzed against distilled water waslyophilizcd and extracted by I M acetic acid. Neutralized samples were dilutedand used as materials. Medium containing authentic IGF-I (1 ng/ml) or TGF-»
(1 ng/ml) was extracted in the same way and plotted in A (A) or B (O).
100
zo
50u_O
O 0.125 025 0.5 %CONCENTRATION OF ANTIBODY
Fig. 7. Effect of anti-lGF-I receptor antibody on PC cell growth in the presenceof 0.3% BSA (El) and 2% PCS (Q). Cells grown in 4% FCS/DMEM were washedwith DMEM and then cultured in 0.3% BSA/DMEM or 2% FCS/DMEM withanti-receptor antibody. Ascites which does not contain anti-receptor antibody was
usedas control. The cell number was counted 3 days after the addition of antibody(**, P< 0.01, compared with each control). Bars, SD.
56-50 IMAA
10 20 30 ¿,0ELUTION VOLUME(ML)
Fig. 5. Elution profile of acid-extracted CM from PC cells. Material appliedon Sephadex G-SO was eluted with I M acetic acid (A.A.). Sample in each fractionwas lyophilized and dissolved with radioimmunoassay buffer. Then the level ofpeptide was measured. CHY.A, chymotrypsinogen A.
—¿�7
UJ eS6
u 2CL
PC CELL \
0 0016 008 0.16 0¿OB 16T PA(NM)
Fig. 8. Effect of TPA on cell growth. Cultured cells were washed with serum-free media and growth studies were done in 0.3% BSA/media with TPA (0.016-1.6 nMl or without TPA. Media and test materials were changed every 2 days.Cell numbers were counted after 4 days of culture. F.B., fibroblast.
8
7
6
i*I*±3
B.G. P-60 IM AA
V.BSA
U
TGFoi
IV.I
10 20ELUTION
30VOLUME(ML)
60
Fig. 6. Bio-Gel P-60 chromatography of 1 M acetic acid (A.A.(-extracted CMfrom PC cells. Sample in each fraction eluted with 1 M acetic acid were lyophilizedand dissolved with radioimmunoassay buffer. Then peptide level was measured.
inhibiting PC cell growth in both the absence (0.3% BSA) andpresence (2% PCS) of serum (Fig. 7), when compared with theactivity of control ascites. Since anti-EGF/TGF-a receptorantibody, which effectively blocks the binding of ligands, wasnot available, a similar study focused on TGF-«was not carriedout.
Effect of Phorbol Ester on Cell Growth of PC Cells or HumanFibroblasts. It is reported that phorbol esters (TPA) have aninhibitory activity on cell growth in some kinds of cancer cells(19-21). In Fig. 8, we show that TPA at doses of more than
0.16 nM had a potent inhibitory activity on PC cell growth,although TPA at concentrations lower than 0.08 nM had only
woe
ÃŽ3000
oZ2000
1 IO 100I GF-I (NG/ML)
156 312 625 125 25 50 COTGFcx(NG/ML)
Fig. 9. Effect of TPA on the binding of '"I-IGF-I (A) or '"I-TGF-« (A) to
PC cells. Confluent cells were washed with DMEM and exposured to TPA (16nM) for I h at 37'C. Then the binding studies were performed (see "Materialsand Methods").
a slight stimulatory activity on the cell growth. Conversely, theproliferation of human fibroblasts was enhanced by TPA atdoses greater than 0.8 nM where PC cell growth was markedlyinhibited.
Effect of TPA on Binding of '"I-IGF-I or 125I-TGF-a to PC
Cells. To elucidate the mechanism(s) of the inhibition of PCcells by TPA, we observed the bindings of ligands to the cells.'"I-IGF-I binding to PC cells was not affected by prior treatment with TPA (16 nM) for 1 h (Fig. 9A). However, '"I-TGF-
«binding to the cells was markedly decreased in cells pretreatedwith TPA (Fig. 9fi). Scatchard analysis demonstrated that thisphenomenon was mainly due to a decrease of the affinity of theligand to receptors (Fig. 10).
105
Research. on February 5, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from

GROWTH KACTORS AND HUMAN PANCREATIC CANCER CELL GROWTH
001
KcJ.I.OJnM
10 20TGFd. BOUND
pM
Fig. 10. Scatchard analysis of '"1-TGF-« binding to PC cells. One-h pretreat
ment with TPA (16 n\t) (O) or without TPA (•).Experimental conditions aresame as those in Fig. 9.
DISCUSSION
The regulatory mechanism of malignant cell growth appearsto be complicated. However, autocrine factors are thought toplay an important role in the development of neoplastic cells(1-3). In the present study, we showed that PC cells grew evenin a medium containing a low concentration (0.1 -0.3%) of BSAand finally reached confluence. Furthermore, CM which wasdialyzed against fresh DMEM for supplying the nutrients lostduring the culture periods, had a stimulatory effect on PC cellgrowth. On the other hand, nonconditioned medium did nothave such an effect. These observations indicate the factor(s)produced by PC cells stimulates the proliferation of theseneoplastic cells.
Recently, Smith et al. (17) showed that a human pancreaticcell line, including the PC cells used in the present study,expressed TGF-a mRNA. TGF-a immunoreactivity, however,was not detected in CM from PC cells in their assay system(17). In our study, IR-TGF-a was detectable in CM, althoughthe greater part of such immunoreactive proteins seems to bepresent as larger forms. Large forms of TGF-a activity havealso been reported to be present in other types of human cancer(22, 23). It would appear plausible that larger forms of IR-TGF-a produced by PC cells cannot be detected by a double-sandwich enzyme-linked immunosorbent assay used in theirassay system, because of the molecular variations of the peptidein CM. The presence of the receptor for EGF or TGF-a in PCcells, which has already been reported (17), was confirmed byus (Fig. 9B). Moreover, we showed that authentic TGF-astimulated the proliferation of the cells. The above observationssuggest that TGF-a is one of the autocrine factors whichstimulate PC cell growth.
A similar analysis was done on IGF-I. This peptide is alsoimplicated as an autocrine factor in the proliferation of somekinds of neoplastic cells (5, 24). Our results indicate that IR-IGF-I is present in acid-extracted CM from PC cells, and itssize is similar to that of authentic IGF-I, which stimulated thecell growth. A specific receptor for '"I-IGF-I was also found inPC cells (Fig. 9/1). Moreover, antibody to IGF-I receptor (alR-3) was able to suppress their growth. Like TGF-a, it is conceivable that IGF-I is important in PC cell growth as an autocrinefactor.
It is well known that tumor promoter phorbol esters, such asTPA, activate protein kinase C and evoke various effects on thecells, including a stimulatory or inhibitory action on growth(18, 25). For example, TPA stimulates the growth of myoblasts(26), mammary epithelial cells (27), fibroblastic cells (28), andthyroid cells (29). On the other hand, it inhibits the cell growthof leukemic cells (HL-60) (19) and human breast cancer cells
(MCF-7) (20, 21). Here, we have demonstrated that TPA
strikingly inhibits the proliferation of PC cells. This phenomenon was also observed when another kind of PK-C activator,teleocidin (30), was used in the growth study (data not shown).The data suggest that activation of PK-C is involved in thesuppression of PC cell growth. The precise mechanism of thegrowth arrest after the enzyme activation is not known. According to our present study, the binding of TGF-a to PC cellswas attenuated in the cells treated with TPA, while that of IGF-I was not changed. Since TGF-a produced by PC cells seemsto play an important role in PC cell growth, as mentionedabove, it is conceivable that the growth inhibition of PC cellsby TPA is exerted by decreasing the binding of TGF-a to itsreceptor. The modulations of EGF receptor, which is thoughtto be the same receptor for TGF-a (22), by phorbol ester havebeen reported. In different cell types phorbol ester or other PK-C activators have been shown to decrease the affinity of EGFreceptor to ligands (31, 32) or decrease the number of EGFreceptors (33, 34). Our results indicate that reduced binding ofTGF-a to PC cells treated by TPA appears mainly owing to thedecrease of the affinity of receptors for TGF-a (Fig. 10). Another possible mechanism of the suppressive effect of TPA isthat the disappearance of PK-C following the activation of theenzyme, which occurs in the cells continuously treated withTPA (35, 36), causes this inhibitory effect. However, this possibility would appear to be unlikely, because it is reported thatthe permeant diacylglycerol is effective without affecting thecellular content of the enzyme in inhibiting MCF cell growth(37). Alternatively, since TPA has been shown to inhibit tyro-sine kinase activity in human epidermoid carcinoma cells (38,39), it is possible that the suppression of PC cell growth byTPA is related to the modulation of this kinase activity by PK-C activators. However, we have no evidence to support thishypothesis. In addition, very low concentrations of TPA (lessthan 0.08 nvi) seem to stimulate PC cell growth. The mechanism of this biphasic effect of TPA also remains to be investigated.
Like TPA, EGF is known to stimulate or inhibit the proliferation of cells. In the latter case, EGF inhibits the growth ofA431 cells (40), rat pituitary tumor cells (41, 42), human breastcancer cells (43), and human squamous cancer cells (44). Sincephosphorylation of EGF receptor by EGF and phorbol esterare similar, but not identical (38,45), it is speculated that somephosphorylated regions of EGF receptor by EGF or phorbolester may be involved in their inhibitory action on the cellgrowth. In PC cells, however, TGF-« stimulated cell growthbut TPA did not. The mechanism of these complicated phenomena in growth inhibition by EGF or TPA remains to be elucidated.
By contrast, the effect of TPA on the growth of fibroblastswas different. At a TPA concentration of more than 1 nM,where PC cell growth was inhibited markedly, the proliferationof human fibroblasts was stimulated. These results emphasizethat the inhibitory effect of TPA on PC cell growth is not anonspecific toxic effect. Although the mechanism of such adiscrepancy in the action of TPA on PC cells and fibroblasts isunclear, it will be interesting to control the growth of malignantand normal cells differently by activating PK-C.
In conclusion, we have shown that the proliferation of pancreatic cancer cells (MIA PaCa-2) is stimulated by humoralfactors, including IGF-I and TGF-a(s) produced by themselves,and PK-C activators suppress PC cell growth presumably, atleast in part, by decreasing the binding of TGF-a to its receptor.
106
Research. on February 5, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from

GROWTH FACTORS AND HUMAN PANCREATIC CANCER CELL GROWTH
ACKNOWLEDGMENTS
We are grateful to Dr. Steven Jacobs who kindly supplied monoclonal anti-IGF-I receptor antibody. We wish to acknowledge SumitomoPharmaceutical Co., Fujisawa Pharmaceutical Co., and Earth ChemicalCo. for providing peptides (IGF-I and TGF-«).
REFERENCES
1. Sporn, M. B., and Todaro, G. J. Autocrine secretion and malignant transformation of cells. N. Engl. J. Med., 303: 878-880, 1980.
2. Sporn, M. B., and Roberts, A. B. Autocrine growth factors and cancer.Nature (Lond.), 313: 745-747, 1985.
3. Goustin. A. S., Leof, E. B., Shipley. G. D.. and Moses, H. Growth factorsand cancer. Cancer Res., 46: 1015-1029, 1986.
4. Derynck, R., Goeddel, D. V., Ullrich. A., Gutterman, J. U., Williams, R. D..Bringman. T. S., and Berger, W. H. Synthesis of messenger RNAs fortransforming growth factors a and tÃand the epidermal growth factor receptorby human tumors. Cancer Res.. 47: 707-712, 1987.
5. Underwood, L. E., D'Ercole. A. J., Clemmons, D. R.. and Van W'yk. J. J.Paracrine functions of somatomedins. Clin. Endocrinol. Metab.. IS: 59-77.1986.
6. Gol-Winkler, R. Paracrine action of transforming growth factors. Clin.Endocrinol. Metab., 15: 99-115, 1986.
7. Van Wyk, J. J., and Trippel, S. B. Endocrine, paracrine. and autocrine effectsof the somatomedins/insulin-like growth factors. In: Growth Hormone—Basic and Clinical Aspects, pp. 337-354. Amsterdam: Elsevier Science Publishers B.V.. 1987.
8. Bronzert, D. A., Pantazis. P., Antoniades, H. N., Kasid. A., Davidson. N.,Dickson. R. B., and Lippman, M. E. Synthesis and secretion of platelet-derived growth factor by human breast cancer cell lines. Proc. Nail. Acad.Sci. USA, 84: 5763-5767. 1987.
9. Hermansson, M.. Nister, M., Betsholtz, C., Heldin, C., Westermark, B.. andFuña.K. Endothelial cell hyperplasia in human glioblastoma: Coexpressionof mRNA for platelet-derived growth factor (PDGF) B chain and PDGFreceptor suggests autocrine growth stimulation. Proc. Nati. Acad. Sci. USA,«5:7748-7752. 1988.
10. Bombesin-like peptides can function as autocrine growth factors in humansmall-cell lung cancer. Nature (Lond.), 316: 823-826, 1985.
11. Alexander, R. W., Upp, J. R., Jr., Poston, G. J., Gupta. V., Townsend, C.M., Jr., and Thompson, J. C. Effects of bombesin on growth of small celllung carcinoma in vivo. Cancer Res.. 48: 1439-1441. 1988.
12. Rohlic. Q. T., Adams, D.. Kull. F. C.. Jr.. and Jacobs. S. An antibody to thereceptor for insulin-like growth factor I inhibits the growth of MCF-7 cellsin tissue culture. Biochem. Biophys. Res. Commun., 149: 276-281, 1987.
13. Conover, C. A., Misra, P.. Hintz. R. L., and Rosenfeld. R. G. Effect of ananti-insulin-like growth factor I receptor antibody on insulin-like growthfactor II stimulation of DNA synthesis in human fibroblasts. Biochem.Biophys. Res. Commun., 139: 501-508, 1986.
14. Chester, J. F., Gaissert, H. A., Ross, J. S., and Malt, R. A. Pancreatic cancerin the Syrian hamster induced by A/-nitrosobis(2-oxopropyl)amine: Cocarcin-ogenic effect of epidermal growth factor. Cancer Res., 46: 2954-2957, 1986.
15. Hierowski, M. T., Liebow, C.. Sapin, K., and Serially. A. V. Stimulation bysomatostatin of dephosphorylation of membrane proteins in pancreatic cancer MIA PaCa-2 cell line. FEBS Lett.. 179: 252-256. 1985.
16. Korc, M.. Meltzer, P.. and Trent. J. Enhanced expression of epidermalgrowth factor receptor correlates with alterations of chromosome 7 in humanpancreatic cancer. Proc. Nati. Acad. Sci. USA, 83: 5141-5144, 1986.
17. Smith. J. J., Derynck, R.. and Korc, M. Production of transforming growthfactor «in human pancreatic cancer cells: evidence for a superagonistautocrine cycle. Proc. Nati. Acad. Sci. USA, 84: 7567-7570, 1987.
18. Nishizuka, Y. The role of protein kinase C in cell surface signal transductionand tumour promotion. Nature (Lond.). 308: 693-698, 1984.
19. Huberman, E., and Callaham, M. F. Induction of terminal differentiation inhuman promyelocytic leukemia cells by tumor-promoting agents. Proc. Nati.Acad. Sci. USA. 76: 1293-1297. 1979.
20. Osborne. C. T., Hamilton. B.. Nover, M., and Ziegler, J. Antagonism betweenepidermal growth factor and phorbol ester tumor promoters in human breastcancer cells. J. Clin. Invest., 6 7: 943-951, 1981.
21. Darbon, J., Valette, A., and Bayard, F. Phorbol esters inhibit the proliferationof MCF-7 cells. Biochem. Pharmacol., 35: 2683-2686. 1986.
22. Todaro, G. J., Fryling, C., and De Larco. J. E. Transforming growth factorsproduced by certain human tumor cells: polypeptides that interact withepidermal growth factor receptors. Proc. Nati. Acad. Sci. USA, 77: 5258-5262, 1980.
23. Bates. S. E.. Davidson. N. E., Valverius. E. M.. Fréter,C. E.. Dickson. R.B., Tarn. J. P.. Kudlow. J. E., Lippman, M. E., and Salomon. D. S.Expression of transforming growth factor n and its messenger ribonucleicacid in human breast cancer: its regulation by estrogen and its possiblefunctional significance. Mol. Endocrinol.. 2: 543-555, 1988.
24. Huff, K. K., Knabbe. C.. Lindsey. R., Kaufman, D., Bronzert. D., Lippman,M. E. and Dickson. R. B. Multihormonal regulation of insulin-like growthfactor-I-rclated protein in MCF-7 human breast cancer cells. Mol. Endocrino!.. 2: 200-208. 1988.
25. Diamond. L.. O'Brien, T. G.. and Baird, W. Tumor promoters and themechanism of tumor promotion. Adv. Cancer Res., 32: 1-74, 1980.
26. Cohen. R.. Pacifici. M., Rubinstein. N.. Biehl, J., and Holtzer. H. Effect ofa tumour promoter on myogenesis. Nature (Lond.). 266: 538-540. 1977.
27. Taketani, V'., and Oka, T. Tumor promoter 12-O-tetradecanoyl phorbol 13-
acetate. like epidermal growth factor, stimulates cell proliferation and inhibitsdifferentiation of mouse mammary epithelial cells in culture. Proc. Nati.Acad. Sci. USA. 80: 1646-1649. 1983.
28. Collins, M. K. L.. and Rozengurt. E. Binding of phorbol esters to high-affinity sites on murine fibroblastic cells elicits a mitogenic response. J. Cell.Physiol.. 112: 42-50. 1982.
29. Bachrach, L. K., Eggo, M. C.. Mak, W. W.. and Burrow, G. N. Phorbolesters stimulate growth and inhibit differentiation in cultured thyroid cells.Endocrinology. /16: 1603-1609. 1985.
30. Fujiki, H.. Tanaka. Y.. Miyake. R.. Kikkawa, U.. Nishizuka. Y., and Sugi-mura, T. Activation of calcium-activated, phospholipid dependent proteinkinase (protein kinase C) by new classes of tumor promoters: telcocidin anddebromoaplysiatoxin. Biochem. Biophys. Res. Commun., 120: 339-343,1984.
31. Shoyab, M., De Larco, J. F... and Todaro, G. J. Biologically active phorbolesters specifically alter affinity of epidermal growth factor membrane receptors. Nature (Lond.). 279: 387-391. 1979.
32. Brown, K. D., Dicker, P., and Rozengurt. E. Inhibition of epidermal growthfactor binding to surface receptors by tumor promoters. Biochem. Biophys.Res. Commun.. 86: 1037-1043, 1979.
33. Lee, L., and Weinstein, I. B. Tumor-promoting phorbol esters inhibit bindingof epidermal growth factor to cellular receptors. Science (Wash. DC), 202:313-315, 1978.
34. Salomon, D. S. Inhibition of epidermal growth factor binding to mouseembryonal carcinoma cells by phorbol esters mediated by specific phorbolester receptors. J. Biol. Chem., 25«:7958-7966, 1981.
35. Rodriguez-Pena. A., and Rozengurt. E. Disappearance of Ca**-sensitive,phospholipid-dependent protein kinase activity in phorbol ester-treated 3T3cells. Biochem. Biophys. Res. Commun., 120: 1053-1059, 1984.
36. Ballester. R., and Rosen. O. M. Fate of immunoprecipitable protein kinaseC in GH3 cells treated with phorbol 12-myristate 13-acetate. J. Biol. Chem.,260: 15194-15199. 1985.
37. Issandou, M., Bayard, F., and Darbon, J. Inhibition of MCF-7 cell growthby 12-O-tetradecanoylphorbol- 13-acetate and 1.2-dioctanoyl-jn-glycerol: distinct effects on protein kinase C activity. Cancer Res.. 48: 6943-6950. 1988.
38. Cochet, C., Gill. G. N., Meisenhelder, J., Cooper. J. A., and Hunter, T. C-kinase phosphorylates the epidermal growth factor receptor and reduces itsepidermal growth factor-stimulated tyrosinc protein kinase activity. J. Biol.Chem., 259: 2553-2558, 1984.
39. Friedman, B.. Frackelton, A. R., Jr., Ross, A. H., Connors, J. M., Fujiki, H.,Sugimura. T.. and Rosner, M. R. Tumor promoters block tyrosine-specificphosphor>lation of the epidermal growth factor receptor. Proc. Nati. Acad.Sci. USA. 81: 3034-3038. 1984.
40. Gill. G. N.. and Lazar. C. S. Increased phosphotyrosine content and inhibition of proliferation in EGF-treated A431 cells. Nature (Lond.), 293: 305-307. 1981.
41. Schonbrunn. A., Krasnoff, M., Weslendorf, J. M., and Tashjian, A. H., Jr.Epidermal growth factor and thyrotropin-releasing hormone act similarly ona clonal pituitary cell strain. J. Cell Biol., 85: 786-797. 1980.
42. Johnson. L. K., Baxter. J. D.. Vlodavsky, I., and Gospodarowicz. D. Epidermal growth factor and expression of specific genes: effects on cultured ratpituitary cells are dissociable from the mitogenic response. Proc. Nati. Acad.Sci. USA, 77: 394-398. 1980.
43. Imai, Y.. Leung. C. K. H., Friesen. H. G.. and Shiu, R. P. C. Epidermalgrowth factor receptors and effect of epidermal growth factor on growth ofhuman breast cancer cells in long-term tissue culture. Cancer Res., 42: 4394-4398, 1982.
44. Kamata, N., Chida. K., Rikimaru, K., Horikoshi. M., Enomoto, S., andKuroki. T. Growth-inhibitory effects of epidermal growth factor and over-expression of its receptors on human squamous cell carcinomas in culture.Cancer Res.. 46: 1648-1653. 1986.
45. Iwashita. S., and Fox, C. F. Epidermal growth factor and potent phorboltumor promoters induce epidermal growth factor receptor phosphorylationin a similar but distinctively different manner in human cpidcrmoid carcinoma A431 cells. J. Biol. Chem.. 259: 2559-2567. 1984.
107
Research. on February 5, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from

1990;50:103-107. Cancer Res Eiji Ohmura, Masaki Okada, Noritaka Onoda, et al. Growthas Autocrine Growth Factors in Human Pancreatic Cancer Cell
αInsulin-like Growth Factor I and Transforming Growth Factor
Updated version
http://cancerres.aacrjournals.org/content/50/1/103
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/50/1/103To request permission to re-use all or part of this article, use this link
Research. on February 5, 2018. © 1990 American Association for Cancercancerres.aacrjournals.org Downloaded from
Top Related