







Languages
Pages
Legal


Instituto de Pesquisas Jardim Botânico do Rio de Janeiro
Escola Nacional de Botânica Tropical
Programa de Pós-Graduação Strictu Sensu
Dissertação de Mestrado
Caracterização de núcleos de vegetação
arbustivo/arbóreos como subsídio à restauração passiva
em pastagens abandonadas
Pollyanna Rodrigues de Oliveira dos Santos
Rio de Janeiro
2014

II
Instituto de Pesquisas Jardim Botânico do Rio de Janeiro
Escola Nacional de Botânica Tropical
Programa de Pós-Graduação Strictu Sensu
Caracterização de núcleos de vegetação
arbustivo/arbóreos como subsídio à restauração passiva
em pastagens abandonadas
Pollyanna Rodrigues de Oliveira dos Santos
Dissertação apresentada ao Programa de Pós-
Graduação em Botânica, Escola Nacional de
Botânica Tropical, do Instituto de Pesquisas
Jardim Botânico do Rio de Janeiro, como
parte dos requisitos necessários para a
obtenção do título de Mestre em Botânica.
Orientador: Dr. João Marcelo
Alvarenga Braga.
Coorientador: Dr. Ricardo Valcarcel.
Rio de Janeiro
2014

III
Caracterização de núcleos de vegetação arbustivo/arbóreos como
subsídio à restauração passiva em pastagens abandonadas
Pollyanna Rodrigues de Oliveira dos Santos
Dissertação submetida ao Programa de Pós-Graduação em Botânica
da Escola Nacional de Botânica Tropical, Instituto de Pesquisas Jardim
Botânico do Rio de Janeiro - JBRJ, como parte dos requisitos necessários
para a obtenção do grau de Mestre.
Aprovada por:
Prof. Dr. João Marcelo A. Braga (Orientador)
Profª. Dra. Marcia Cristina Mendes Marques
Prof. Dr. Luiz Fernando Duarte de Moraes
em 25/02/2014
Rio de Janeiro
2014

IV
Santos, Pollyanna Rodrigues de Oliveira dos.
S237c Caracterização de núcleos de vegetação arbustivo/arbóreos como subsídio à restauração passiva em pastagens abandonadas/ Pollyanna Rodrigues de Oliveira dos Santos. – Rio de Janeiro, 2014.
xiii, 64f. : il. ; 28 cm.
Dissertação (mestrado) – Instituto de Pesquisas Jardim Botânico do Rio de Janeiro / Escola Nacional de Botânica Tropical, 2014.
Orientadores: João Marcelo Alvarenga Braga; Ricardo Valcarcel.
Bibliografia.
1. Regeneração natural. 2. Sucessão ecológica. 3. Resiliência. 4. Mata Atlântica. 5.Reserva Biológica de Poço das Antas (RJ). 6. São João, Rio (RJ). I. Título. II. Escola Nacional de Botânica Tropical.
CDD 577.35

V
Dedico este trabalho
a João e Cristina,
Luana,
e Eluã,
com todo meu amor!

VI
AGRADECIMENTOS
Primeiramente a Deus, o grande responsável por tudo!
Agradeço ao João Marcelo Alvarenga Braga, antes de mais nada, pelo voto de
confiança quando aceitou me orientar. Por me ajudar com as muitas barreiras as quais
me deparei nesta travessia, pelas conversas que tanto contribuíram para que este
trabalho chegasse ao final e pela amizade!
Ao Ricardo Valcarcel, por aceitar ser meu coorientador, e assim seguir
contribuindo com a minha formação. Obrigada pelas diversas e ricas discussões, pela
amizade e confiança!
À Marcia Marques, Luiz Fernando Moraes e Jerônimo Sansevero por aceitarem
compor minha banca avaliadora e contribuírem com o trabalho.
À Escola Nacional de Botânica Tropical por esses anos de muito aprendizado,
em especial à Hevelise pela disposição em ajudar da melhor forma sempre que precisei.
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) pela
bolsa concedida. Ao PPBio Mata Atlântica pelo auxílio nas excursões, em especial ao
Pablo José Pena Rodrigues e Mariela Figueiredo por me ajudarem na logística. Ao
Gustavo Luna Peixoto pelo apoio, e à Reserva Biológica de Poço das Antas por ter
oferecido a infraestrutura necessária para o trabalho de campo.
A todos os que me ajudaram nos trabalhos de campo, em especial ao Adilson
Pintor, pelo empenho que foi essencial para concluir a dissertação, além da amizade!
Ao Pablo pela sua importante colaboração em todas as fases da dissertação, ao
Jerônimo pelas críticas e sugestões feitas na qualificação, ao Richieri Sartori pela ajuda
com a identificação botânica e ao Rodolfo Abreu pelo auxílio nas análises estatísticas.
As contribuições de vocês foram muito importantes!
Aos amigos do Grupo TER (Taxonomia, Ecologia e Restauração), Ana Elena,
Rafael, Luana, Leandro, Pablinho, Augusto, Janilson, Jerônimo, Richieri, Rodolfo,
Andrea, Daniele, enfim, a todos do grupo agradeço pelas conversas que trouxeram tanto
aprendizado, pelas diversas ajudas e discussões científicas, pelas risadas, e pelos ótimos
momentos.
Aos amigos do Laboratório de Manejo de Bacias Hidrográficas da Universidade
Federal Rural do Rio de Janeiro (LMBH/UFRRJ), foram muito bons os momentos que
estive aí com vocês. E ainda na Rural, agradeço aos meus queridos amigos de

VII
graduação, que se fizeram tão presentes no mestrado: Letícia, Dally, Carlão e Wanessa.
Amigos pra toda vida!
Agradeço àqueles que foram a grande força e razão para que eu conseguisse dar
mais este importante passo em minha vida. Aos meus pais João e Cristina, meus
maiores motivadores, obrigada pelo carinho e por estarem comigo em todos os
momentos. A minha querida irmã e amiga Luana, que foi essencial na minha formação,
obrigada por me ajudar tanto, e pelos muitos momentos maravilhosos. E ao Eluã,
obrigada por todo amor, compreensão, por estar sempre ao meu lado, e por juntos
estarmos na busca dos nossos sonhos.
Por fim, tantos foram os que estiveram presentes e me apoiaram nesses dois anos
de mestrado, que sem dúvida, este agradecimento é pouco perto da importância que
cada um teve. Com a contribuição de todos vocês pude dar mais este passo.
Muito obrigada!

VIII
RESUMO
Na Mata Atlântica, as pastagens abandonadas são ambientes comuns que, dependendo
do nível de resiliência, apresentam potenciais distintos de restauração passiva, através
da regeneração natural. O objetivo do trabalho foi caracterizar a dinâmica sucessional
de núcleos de vegetação arbustivo/arbóreos para discutir seu papel ecológico na
restauração passiva na Reserva Biológica de Poço das Antas. Foram selecionados 1,08
ha de pastagem abandonada para a amostragem dos indivíduos arbustivo/arbóreos
isolados ou em núcleos pequenos, médios e grandes. Foram levantados o estrato
regenerante, dominância de gramíneas, aporte de serrapilheira, abertura do dossel e
características do solo a fim de analisar fatores que possam ser modificados pelos
núcleos. Moquiniastrum polymorphum foi a espécie mais abundante tanto isolada como
em núcleos de vegetação. Com o aumento da área dos núcleos, houve um aumento de
espécies zoocóricas e não-pioneiras, tanto para o estrato superior como para o estrato
regenerante. A cobertura de gramíneas e abertura do dossel influenciaram a abundância
de regenerantes, ficando evidente que há poucos indivíduos juvenis em locais com
pouco sombreamento, como na matriz graminóide. Variáveis do solo como o índice de
saturação de bases se mostraram diferentes entre classes de núcleos, e entre estes e a
matriz graminóide, inferindo que onde há a presença de núcleos encontra-se um solo de
maior fertilidade. Concluiu-se que os núcleos de vegetação arbustivo/arbóreos
amenizam as características adversas do ambiente, favorecendo o aumento da
diversidade em pastagens. Entretanto, apesar de efetiva, a restauração passiva gerada a
partir dos núcleos é lenta.
Palavras-chave: pastagens abandonadas, regeneração natural, variáveis ambientais.

IX
ABSTRACT
In the Atlantic Forest, the abandoned pastures are common environments that,
depending on the level of resilience, have distinct potential of passive restoration
through natural regeneration. The aim of this work was to characterize the successional
dynamics of shrub/tree nuclei of vegetation to discuss their ecological role in the passive
restoration in the Poço das Antas Biological Reserve. An area of 1.08 ha of abandoned
pasture was selected for the sampling of isolated shrub/tree individuals or in small,
medium and large nuclei. The data of regeneration, dominance of grasses, supply of
litter, canopy openness, and soil characteristics were collected to examine factors that
may be modified by the nuclei. Moquiniastrum polymorphum was the most abundant
species, found isolated or in nuclei of vegetation. The number of zoochorous and non-
pioneer species increased with the nuclei area, in both upper and regenerating layers.
The grass cover and canopy openness influenced in the abundance of saplings,
evidencing the existence of a few juveniles in places with little shading, as in pasture.
Variables of soil like basis saturation index were different between nuclei classes, and
between nuclei and pasture. This implies in soils with high fertility when in presence of
the nuclei. It was concluded that the shrub/tree nuclei of vegetation alleviate the adverse
characteristics of the environment, favoring the increasing of the diversity in pastures.
However, although effective, passive restoration generated from the nuclei is slow.
Key-words: abandoned pastures, natural regeneration, environmental variables.

X
SUMÁRIO
1. INTRODUÇÃO .................................................................................................................... 1
2. MATERIAIS E MÉTODOS ................................................................................................. 5
2.1. Área de Estudo .............................................................................................................. 5
2.2. Amostragem dos núcleos de vegetação arbustivo/arbóreos .......................................... 8
2.3. Fatores influenciados pelos núcleos de vegetação arbustivo/arbóreos ........................ 11
2.3.1. Regeneração natural em núcleos de vegetação e em matriz graminóide ............ 11
2.3.2. Abertura de dossel ............................................................................................... 12
2.3.3. Dominância de gramíneas ................................................................................... 12
2.3.4. Aporte de serrapilheira ........................................................................................ 12
2.3.5. Análise de Solo.................................................................................................... 13
2.4. Análises dos Dados ..................................................................................................... 14
3. RESULTADOS ................................................................................................................... 15
3.1. Sucessão ecológica em pastagem abandonada ............................................................ 15
3.1.1. Estrato superior ................................................................................................... 15
3.1.2. Estrato regenerante .............................................................................................. 21
3.2. Fatores influenciados pelos núcleos de vegetação ...................................................... 26
4. DISCUSSÃO ....................................................................................................................... 36
5. CONCLUSÃO .................................................................................................................... 42
6. RECOMENDAÇÕES FINAIS ........................................................................................... 43
APÊNDICES ............................................................................................................................... 57
ANEXOS..................................................................................................................................... 62

XI
LISTA DE FIGURAS
Figura 1: Localização da Reserva Biológica de Poço das Antas em relação à cadeia de montanhas da serra do Mar e a planície de inundação. .............................................................. 6
Figura 2: Mapa da vegetação da Reserva Biológica de Poço das Antas, Silva Jardim – RJ. ........ 7
Figura 3: Diferentes padrões de vegetação na Reserva Biológica de Poço das Antas, Silva Jardim – RJ. ................................................................................................................................ 7
Figura 4: Esquema de uma parcela de 30 x 60 metros, com as classificações em árvores isoladas e árvores formadoras de núcleos de vegetação arbustivo/arbóreos, e os respectivos exemplos de cada classificação, Reserva Biológica de Poço das Antas, RJ. ............................................ 10
Figura 5: Parcela de 60 x 30 metros com esquema de alocação de parcelas 1 x 2 metros nos núcleos de vegetação arbustivo/arbóreos, Reserva Biológica de Poço das Antas, RJ. ............. 11
Figura 6: Esquema das coletas de serrapilheira nas parcelas 1 x 2 metros no interior dos núcleos e em matriz graminóide. ........................................................................................................... 13
Figura 7: Esquema das coletas de solo nas parcelas 1 x 2 metros no interior dos núcleos e em matriz graminóide. .................................................................................................................... 14
Figura 8: Diferentes riquezas e espécies mais abundantes encontradas formando núcleos de vegetação arbustivo/arbóreos e de forma isolada . ................................................................... 18
Figura 9: Curva de diversidade e dominância para indivíduos isolados e núcleos de vegetação arbustivo/arbóreos de diferentes tamanhos. .............................................................................. 19
Figura 10: Síndrome de dispersão do estrato superior apresentado em porcentagem, para os indivíduos isolados e diferentes classes de núcleos. ................................................................. 20
Figura 11: Grupo sucessional do estrato superior apresentado em porcentagem, para os indivíduos isolados e diferentes classes de núcleos. ................................................................. 20
Figura 12: Síndrome de dispersão do estrato regenerante apresentado em porcentagem, para as diferentes classes de núcleos. ................................................................................................... 23
Figura 13: Grupo sucessional do estrato regenerante apresentado em porcentagem, para as diferentes classes de núcleos.. .................................................................................................. 24
Figura 14: Trajetória sucessional de pastagens abandonadas na Reserva Biológica de Poço das Antas, Silva Jardim – RJ. .......................................................................................................... 25
Figura 15: Curva de diversidade e dominância para indivíduos em núcleos de vegetação arbustivo/arbóreos e matriz graminóide em diferentes posições na pastagem. ........................ 27
Figura 16: Gráficos gerados com testes de regressão linear para responder o comportamento da regeneração natural à diferentes fatores ambientais. ................................................................ 30

XII
Figura 17: Análise de Componentes Principais (PCA) que infere a relação entre o estrato regenerante e variáveis, em núcleos de vegetação arbustivo/arbóreos e matriz graminóide. ... 31
Figura 18: Análise de Componentes Principais (PCA) referentes às variáveis do solo em núcleos de vegetação arbustivo/arbóreos e matriz graminóide.. ............................................................ 33
Figura 19: Análise de redundância (RDA) das espécies do estrato regenerante relacionadas às variáveis de solo significativas. ................................................................................................ 34
Figura 20: Análise de redundância (RDA) das espécies do estrato superior relacionadas às variáveis de solo significativas. ................................................................................................ 35
Figura 21: Caracterização das influências positivas trazidas pelos núcleos de vegetação arbustivo/arbóreos de acordo com seu distanciamento, em áreas de pastagem abandonada.. .. 39

XIII
LISTA DE TABELAS
Tabela 1: Composição de espécies arbustivo/arbóreas e suas abundâncias, grupo sucessional, síndrome de dispersão e forma de ocorrência em pastagem abandonada, Reserva Biológica de Poço das Antas, Silva Jardim – RJ. .......................................................................................... 15
Tabela 2: Análises de espécies indicadora das diferentes classes de núcleos de vegetação arbustivo/arbóreo. ..................................................................................................................... 21
Tabela 3: Composição de espécies do estrato regenerante e suas abundâncias comparadas ao do estrato superior, grupo sucessional e síndrome de dispersão em pastagem abandonada, Reserva Biológica de Poço das Antas, Silva Jardim – RJ. ..................................................................... 22
Tabela 4: Médias dos fatores e número de indivíduos na matriz graminóide e em núcleos de vegetação arbustivo/arbóreos de pastagens abandonadas na Reserva Biológica de Poço das Antas, Silva Jardim – RJ. .......................................................................................................... 28
Tabela 5: Resultados da ANOVA para as variáveis de solo relacionadas ao estrato regenerante de núcleos de vegetação arbustivo/arbóreos. ............................................................................ 34
Tabela 6: Resultados da ANOVA para as variáveis de solo relacionadas ao estrato superior de núcleos de vegetação arbustivo/arbóreos. ................................................................................. 35

1
1. INTRODUÇÃO
A Mata Atlântica está em extremo estado de degradação, com seus
remanescentes compostos, em grande parte, por fragmentos menores que 50 ha e que
são insuficientes para suportar em longo prazo a sobrevivência da floresta (Ribeiro et
al., 2009). Alterações ambientais são constantemente observadas ao longo do tempo e
em diferentes tipos de paisagens. Segundo a Sociedade de Restauração Ecológica (SER,
2004), um ecossistema sofre transformação quando há a conversão da paisagem e,
consequentemente, dos processos ecológicos. A modificação de ecossistemas gera
principalmente a fragmentação, que interrompe a continuidade espacial e funcional de
habitats (Aronson et al., 2011).
A resiliência, que é considerada a forma e o grau de recuperação da estrutura e
funções iniciais de um ecossistema após perturbação (Westman, 1978), é um fator de
extrema importância para a recuperação natural de áreas perturbadas, e que pode ser
observada em áreas degradadas do bioma Mata Atlântica. As pastagens abandonadas
são um exemplo de ambientes perturbados que compõem este domínio e que,
dependendo do nível de resiliência, pode apresentar formas diferenciadas de
recuperação.
Neste contexto de degradação está presente a restauração ecológica, que é
definida como o processo que auxilia o restabelecimento de um ecossistema degradado,
danificado ou destruído (SER, 2004), podendo ser dividida em restauração passiva e
restauração ativa. A restauração passiva é definida como o retorno espontâneo de um
ecossistema degradado para um estado desejável pré-existente, através da resiliência e
em decorrência da interrupção intencional da perturbação antrópica (Aronson et al.,
2011; Scowcroft & Yeh, 2013). Acontece quando há a ocorrência da sucessão
secundária em áreas onde houve, por exemplo, a remoção de gado e abandono da
agricultura (Morrison & Lindell, 2010).
Já a restauração ativa se diferencia pela introdução intencional de diásporos e
plantio direto, especialmente de sementes e mudas. Ela pode acelerar o processo de
restauração (DeSimone, 2011), por ser uma técnica onde há geralmente escolha das
espécies de rápido crescimento, do espaçamento e do preparo do terreno. Entretanto, os
custos para a implementação de plantios são elevados, e por isso muitas vezes limitam
as opções de restauração (Holl & Aide, 2011).

2
Para que ocorra a restauração passiva é importante que a área esteja em estado
de abandono, sem nenhum tipo de uso (e.g. pecuária, agricultura) ou de grandes
distúrbios (e.g. fogo, enchente). A recuperação natural dessas áreas se torna mais lenta
por estar submetida a intensivo efeito humano, como degradação do solo e perturbações
recorrentes (Shono et al., 2007). Além disso, com um menor uso da área, há menor
interferência no processo de sucessão ecológica.
A partir de uma perturbação é que ocorre a regeneração natural através das
etapas da sucessão ecológica, ocasionando a mudança espontânea na composição de
espécies e na estrutura da comunidade em função do tempo e do espaço (Smith, 1996).
Connell & Slatyer (1977) sugeriram três modelos de sucessão, sendo denominados de
facilitação, inibição e tolerância. O modelo de facilitação ocorre quando espécies em
estágios iniciais modificam e preparam o ambiente para as espécies nas fases finais. Já o
modelo de inibição é competitivo, onde as espécies que se estabeleceram primeiro
dificultam ou impossibilitam que o local seja adequado para a recolonização de outras
espécies. O modelo de tolerância é intermediário entre os dois primeiros, onde espécies
invadem um local, se estabelecem e crescem até a maturidade juntamente com os
indivíduos antecessores, tendo uma maior tolerância a poucos recursos.
Das interações mais frequentes observadas na sucessão, em especial em área de
pastagem abandonada, tem-se a relação entre competição e facilitação. Entre as plantas
que crescem próximas umas das outras podem prevalecer efeitos negativos, havendo
competição que limita os recursos como água, nutrientes, luz e espaço (Padilla &
Pugnaire, 2006). Já a facilitação pode exercer influências positivas, aumentando a
sobrevivência de indivíduos e a diversidade local, modificando as condições bióticas
causadas por toda e qualquer presença de espécies (Stachowicz, 2001; Padilla &
Pugnaire, 2006; McIntire & Fajardo, 2014). As espécies facilitadoras colonizam uma
determinada área num estágio inicial, capacitando o ambiente para a invasão e
desenvolvimento dos estágios posteriores (Ricklefs, 1996).
A competição principal nas áreas de pastagem é devido à dominância de
gramíneas invasoras. Segundo Holl (2002), comparando grandes e pequenas manchas
de arbustos com área de pastagem, a sobrevivência de plântulas e juvenis é maior nas
grandes manchas e menor em pastagem, devido à competição de gramíneas. Uma maior
cobertura do solo causada pelo aumento de biomassa das espécies herbáceas em

3
pastagens dificulta a germinação de sementes e o estabelecimento de espécies arbóreas
(Cheung et al., 2009).
Entender como se dá o processo de sucessão ecológica em áreas perturbadas é
importante para aperfeiçoar técnicas de recuperação da cobertura florestal.
Compreender a dinâmica da vegetação possibilitará ampliar teorias de sucessão e os
esforços na restauração de florestas tropicais (Holl, 2002). Para a aplicação da
restauração, o desafio é identificar quais são as interações positivas, e como
potencializá-las para aumentar a biodiversidade (McIntire & Fajardo, 2014).
Os indivíduos arbóreos remanescentes em áreas abandonadas em pastagens
podem possuir capacidade nucleadora (Reis et al., 2003b), podendo ser primordial para
a posterior chegada e estabelecimento de outros indivíduos (Yarranton & Morrison,
1974). Entretanto, para o estabelecimento de grupos de indivíduos arbustivo/arbóreos
ou núcleos de vegetação em meio às pastagens, além do estabelecimento do primeiro
indivíduo, é necessário que o mesmo acarrete condições para o desenvolvimento de
outros indivíduos. As interações positivas são importantes para a comunidade por
modificar o habitat, onde um indivíduo altera as condições ambientais fazendo ele se
tornar menos estressante para o estabelecimento de outros indivíduos, podendo
aumentar a diversidade de espécies (Stachowicz, 2001, Yarranton & Morrison, 1974).
Quando o projeto de restauração tem como base os conceitos da nucleação, ele
estará facilitando a sucessão, e quanto mais numerosos e diversificados forem os
núcleos formados, a restauração se tornará mais efetiva (Reis et al., 2003a). O modelo
agregado de sucessão e o efeito da facilitação fazem com que a nucleação seja um meio
de simplificar os processos na restauração (Rodrigues, 2013).
A regeneração natural dos núcleos de vegetação arbustivo/arbóreos pode
contribuir consideravelmente para a restauração passiva de áreas de pastagens. Além de
poder facilitar a regeneração em pastos degradados, os núcleos ainda podem exercer o
mecanismo de eliminar remanescentes dominados por gramíneas (Kolb, 1993) e gerar
sombra pelas copas que atua protegendo as mudas e as plantas menores de altas
temperaturas, reduzindo a perda de água e a foto-inibição (Callaway, 1995). Os núcleos
também servem de abrigo e fonte de alimentação para os animais, em especial pássaros
e morcegos (Kolb, 1993; Reis et al., 2003b), além de atrair polinizadores e proteger
contra herbívoros (Callaway, 1995).

4
O aumento da diversidade ao redor de uma árvore depende mais se ela está
isolada ou em grupo do que de sua espécie (Rodrigues, 2013), principalmente devido à
estrutura formada pelo núcleo que gera benefícios para indivíduos menos tolerantes a
situações extremas. Entretanto, não é importante para a restauração da paisagem se o
surgimento de um grupo de novas árvores for apenas bem sucedida, pois existe ainda a
necessidade da expansão florestal para áreas adjacentes, diminuindo assim o efeito da
fragmentação (Scowcroft & Yeh, 2013).
Alguns indivíduos geram melhorias das características ambientais, como a
alteração de substrato ou aumento da disponibilidade de um recurso, ou até através de
outros benefícios, como a introdução de micorrizas no solo (Callaway, 1995) e o
aumento da disponibilidade de nutrientes. Existem diversas maneiras de deposição dos
nutrientes no sistema, entre elas a serrapilheira, que depois de acumulada, intercepta luz,
reduz a amplitude térmica do solo e gera sombra para sementes e plântulas (Barbosa &
Faria, 2006). O pH do solo também pode ser alterado com essas estruturas (Jirová et al.,
2012).
Ao mesmo tempo em que a vegetação pode contribuir com as condições do solo,
este também influi diretamente na vegetação que junto se desenvolve. Segundo Pueyo et
al. (2009), uma boa preservação da vegetação favorece a relação entre planta e solo, que
acaba mantendo as boas condições do solo e este, por sua vez, permite o
estabelecimento e sobrevivência da vegetação.
Em áreas de Mata Atlântica do estado do Rio de Janeiro, um local potencial para
entender a dinâmica dos núcleos de vegetação arbustivo/arbóreos é a Reserva Biológica
de Poço das Antas, por se tratar de uma Unidade de Conservação de Proteção Integral.
As Reservas Biológicas se destacam por serem áreas onde não há interferência humana
intensa ou modificações ambientais, em exceção as medidas de recuperação e ações de
manejo (SNUC, 2000). Como Poço das Antas, antes de se tornar uma Unidade de
Conservação a cerca de 40 anos, era uma fazenda de criação de gado com culturas
agrícolas, hoje ainda são encontradas pastagens abandonadas.
O objetivo deste capítulo foi caracterizar a dinâmica sucessional de núcleos de
vegetação arbustivo/arbóreos em pastagens abandonadas, para analisar seu papel
ecológico na restauração passiva. Para isso, o trabalho buscou responder os seguintes
objetivos específicos:

5
• Em pastagens abandonadas, é possível observar dinamismo sucessional e
evolução dos núcleos de vegetação arbustivo/arbóreos?
• Os núcleos de vegetação arbustivo/arbóreos são estruturas que favorecem o
ambiente para impulsionar a recuperação dessas áreas?
• Quais são os principais fatores que são favorecidos por estas estruturas?
• Os núcleos de vegetação arbustivo/arbóreos são capazes de restaurar
efetivamente áreas de pastagens abandonadas?
2. MATERIAIS E MÉTODOS
1.1. Área de Estudo
O estudo foi realizado na Reserva Biológica de Poço das Antas, que em 1974 foi
criada por meio do Decreto Federal nº 73.791 de 11/03/1974, e no ano seguinte foi
redelimitada pelo Decreto nº 76.534 de 03/11/1975, totalizando 40 anos de sua
formação.
A Reserva está localizada no município de Silva Jardim, Estado do Rio de
Janeiro, inserida na região da baixada litorânea do nordeste fluminense entre as
coordenadas 22°30’ e 22°33’S; 42°15’ e 42°19’w (Oliveira, 2012). A área apresenta
aproximadamente 5000 ha, sendo limítrofe com o rio São João e a Represa de
Juturnaíba, se estendendo ao norte até a rodovia BR-101.
A Reserva ocupa a área central da bacia hidrográfica do Rio São João, abaixo da
cadeia de montanhas da Serra do Mar, com parte inserida na planície de inundação
(Figura 1). Devido esta configuração, a área possui um vínculo com essas duas zonas,
por receber influência tanto da planície de inundação como da cadeia de serras.

6
Figura 1: Localização da Reserva Biológica de Poço das Antas em relação à cadeia de montanhas da
serra do Mar e a planície de inundação. Fonte: imagem Google Earth.
Nas áreas de planície com a presença de morrotes o processo inicial de
colonização foi dado principalmente com a cultura do café e da cana-de-açúcar, e após o
declínio dessas atividades, as áreas passaram a ser ocupadas pela criação extensiva de
gado e o cultivo de cítricos (Cunha e Freitas, 2004).
Além do histórico de exploração e mudanças no uso do solo ocorridos na bacia,
principalmente nas áreas de baixada, houve também alterações físicas com a retificação
do rio São João e a construção da represa de Juturnaíba. Isso gerou assoreamento do rio,
alagamento de trechos a montante da barragem, submersão das formas de relevo,
alteração das taxas de infiltração que favoreceu a ocupação agrícola de cultivo irrigados
da planície a jusante da barragem, além da modificação da fauna e flora aquática,
ciliares e da planície (Cunha, 1995).
A Reserva está inserida num trecho onde, originalmente, era coberta por Floresta
Ombrófila Densa, e atualmente é dividida em seis unidades fisionômicas: floresta
submontana (34,3%), floresta aluvial (17,9%), formação pioneira com influência fluvial
(20,0%), capoeira submontana (4,3%), capoeira aluvial (11,3%) e campo antrópico
(pastagem) (12,3%). (Lima et al., 2006) (Figuras 2 e 3).
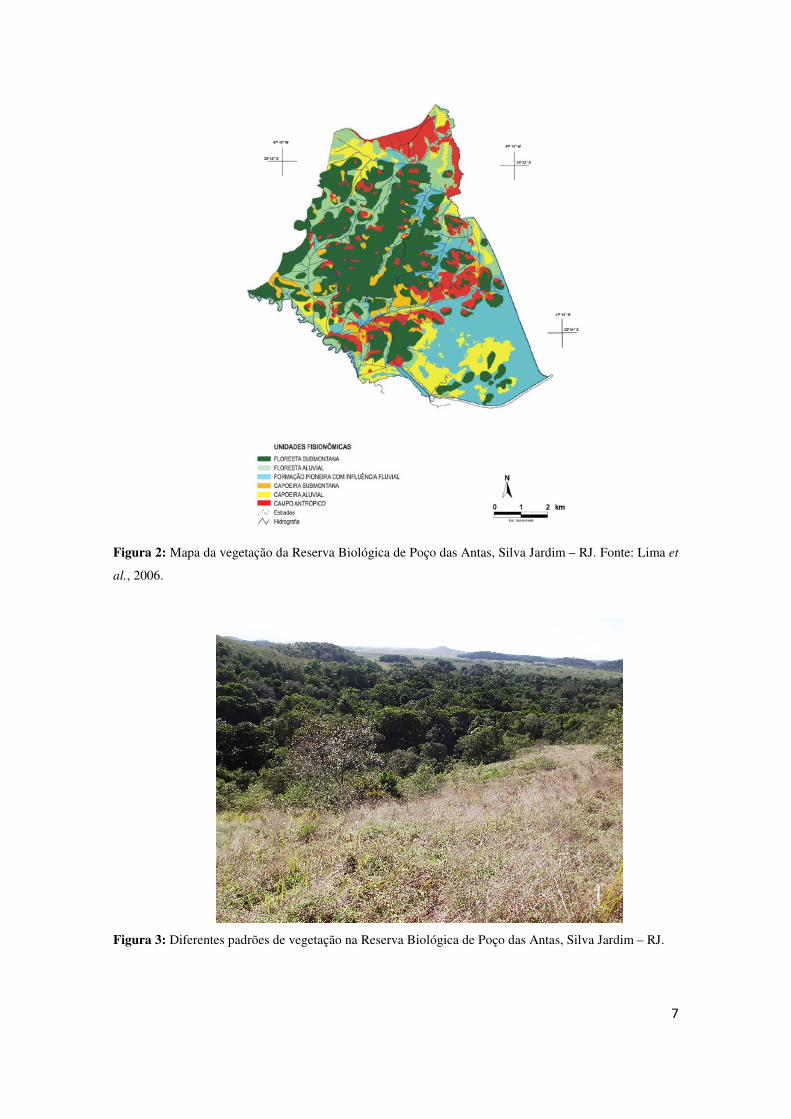
7
Figura 2: Mapa da vegetação da Reserva Biológica de Poço das Antas, Silva Jardim – RJ. Fonte: Lima et
al., 2006.
Figura 3: Diferentes padrões de vegetação na Reserva Biológica de Poço das Antas, Silva Jardim – RJ.

8
Anteriormente, as áreas de pastagens foram utilizadas para criação de gado, e
hoje são dominadas por espécies invasoras como sapê (Imperata brasiliensis), capim-
gordura (Melinis minutiflora), capim-colonião (Panicum maximum) e samambaia
(Pteridium aquilinum) (Oliveira, 2012). As antigas pastagens foram abandonadas após a
criação da Unidade de Conservação, dando início ao processo de sucessão secundária e
estabelecimento da vegetação lenhosa. Entretanto, mesmo existindo fragmentos
florestais, potenciais fontes de propágulo, próximos às áreas estudadas, Vieira & Pessoa
(2001) concluíram que o domínio da vegetação densa formada por espécies herbáceas,
afeta a composição florística e dificulta o estabelecimento de espécies arbóreas, além de
históricos recentes de incêndios (Oliveira, 2012).
A Reserva Biológica de Poço das Antas propiciou em 40 anos a suspensão de
atividades agropastoris, permitindo o restabelecimento da vegetação em locais com
maior capacidade de resiliência. É provável que, em áreas onde os usos foram mais
intensos, a regeneração tenha sido mais lenta, sendo a recíproca verdadeira, o que
permite confirmar que nas áreas com vegetação melhor estruturada tenha havido usos
menos impactantes, e/ou as áreas reúnam atributos ambientais que confiram maior
resiliência.
Na Reserva, são encontradas atualmente áreas de várzeas temporária ou
permanentemente inundadas, onde predominam Neossolos flúvicos, e morrotes com
altitude de até 200 metros, onde predominam Argissolos (Moraes et al., 2006), sendo
encontrados unidades geomorfológicas colinas e maciços costeiros caracterizados por
topografia deprimida com reduzidos valores altimétricos (Neves & Peixoto, 2008).
As temperaturas médias são elevadas durante quase todo o ano, apresentando
máxima de 32°C e mínima de 18°C, não apresentando inverno pronunciado (Vieira &
Pessoa, 2001). O clima é considerado quente e úmido, que, segundo a classificação de
Koppen se enquadra no tipo Aw, com precipitação média anual de aproximadamente
1900 mm (Moraes et al., 2006).
1.2. Amostragem dos núcleos de vegetação arbustivo/arbóreos
Foram selecionadas ao longo da Reserva seis áreas de pastagem abandonada a
40 anos, em diferentes encostas de morrotes mamelonares, onde alocou-se uma parcela
de 30 x 60 metros por área. Estas parcelas delimitaram áreas de encosta onde foram
amostrados os núcleos de vegetação arbustivo/arbóreos.

9
Ao escolher as áreas, foi necessário que as mesmas apresentassem no mínimo
um núcleo de vegetação arbustivo/arbóreos. Foi considerado um núcleo quando no
mínimo duas árvores apresentassem copas sobrepostas, ou quando apresentassem
contato entre as mesmas, ou ainda quando as copas se distanciavam em no máximo 0,5
metros.
Numa área total de 1,08 ha, composta pelas seis áreas delimitadas, foram
levantados 50 núcleos de vegetação encontrados no interior das parcelas, além de
indivíduos isolados dispersos. Para melhor representação, os indivíduos
arbustivo/arbóreos foram então classificados em isolados, que são aqueles indivíduos
que não estão agrupados com outros indivíduos, e árvores formadoras de núcleos de
vegetação, que se encontram agrupadas com outros indivíduos (Figura 4).
Todos os núcleos de vegetação encontrados tiveram seus diâmetros medidos
(m), obtendo-se os dois diâmetros cruzados (Da e Db) com auxílio de uma trena. A área
dos núcleos foram obtidas através da fórmula da área da elipse (π*Da/2*Db/2), por esta
ser a forma geométrica que mais se assemelha aos núcleos (Kolb, 1993; Maza-
Villalobos et al., 2011). Ao todo, os núcleos apresentaram cerca de 20% da área total de
pastagem amostrada (1,08 ha). Foi feita a classificação dos núcleos em pequenos (< 23
m²), médios (23 – 50 m²) e grandes (> 50 m²), objetivando comparar a estrutura dos
núcleos em três diferentes momentos. As classes foram compostas por 16 núcleos
pequenos, 17 médios e 17 grandes, sendo que para as posteriores análises foram
excluídos dois núcleos, um pequeno e um grande, por possuírem características
peculiares discrepantes dos demais.

10
Figura 4: Esquema de uma parcela de 30 x 60 metros, com as classificações em árvores isoladas e
árvores formadoras de núcleos de vegetação arbustivo/arbóreos, e os respectivos exemplos de cada
classificação, Reserva Biológica de Poço das Antas, RJ.
Os indivíduos tiveram medidos o diâmetro à altura do peito (DAP, cm) e altura
(m), apresentando limite de inclusão DAP > 2,5 cm e altura mínima de 1,5 m. Todos os
indivíduos foram identificados no campo e, havendo necessidade, foram coletados para
identificação no herbário do Jardim Botânico do Rio de Janeiro (RB) ou com auxílio de
consultas à literatura e especialistas.
Com os indivíduos classificados e mensurados, foi realizada a avaliação
estrutural da vegetação para os indivíduos isolados e para os formadores de núcleos de
vegetação, para se chegar à espécie com maior valor de importância (VI) de cada
formação. Para isso, foi encontrada a densidade relativa (DR), frequência relativa (FR) e
dominância relativa (DoR) de cada espécie (Moro & Martins, 2011).
Foi confeccionada também uma listagem do grupo sucessional ao qual cada
espécie pertence, classificada em pioneiras e não pioneiras, e a síndrome de dispersão.
Para elaborar esta classificação, foi feito uso de bibliografias especializadas (Silva et al.,
2003; Nappo et al., 2004; Lorenzi, 2008; Nóbrega et al., 2008; Pereira et al., 2008;
Árvores isoladas
60 metros
30 metros Árvores formadoras de ilhas de vegetação Árvores formadoras de núcleos de vegetação
Árvores isoladas

11
Lorenzi, 2009; Lorenzi et al., 2010; Valentini et al., 2010; Sansevero et al., 2011; Braz
et al., 2012; Santos & Alves, 2012; Souza, 2013).
O levantamento do estrato regenerante que compõe os núcleos de vegetação foi
feito por censo da regeneração natural nos núcleos. Foram levantados todos os
indivíduos de 0,10 a 1,50 metros de altura, e com diâmetro a altura do solo (DAS) < 2,5
cm, incluindo plântulas e indivíduos juvenis, para os quais foram medidos DAS, altura e
determinada a espécie. As espécies deste estrato também foram classificadas quanto ao
seu grupo sucessional e dispersão, fazendo uso das mesmas bibliografias utilizadas para
classificar as espécies do estrato superior.
1.3. Fatores influenciados pelos núcleos de vegetação arbustivo/arbóreos
1.1.1. Regeneração natural em núcleos de vegetação e em matriz
graminóide
Para a comparação do estrato regenerante formado no interior dos núcleos de
vegetação e em matriz graminóide, foram alocadas quatro parcelas de 1 x 2 metros em
cada núcleo de vegetação, em linha voltada para o norte, sendo duas localizadas no
interior e duas fora do núcleo (Figura 5).
Figura 5: Parcela de 60 x 30 metros com esquema de alocação de parcelas 1 x 2 metros nos núcleos de
vegetação arbustivo/arbóreos, Reserva Biológica de Poço das Antas, RJ.

12
As parcelas apresentam 1 metro de distância entre si, então, para esta medição,
não foram utilizados os núcleos pequenos (< 23 m²), para ser possível a alocação das
parcelas em seu interior. Foram utilizados 32 núcleos dos 50 levantados inicialmente
(médios e grandes), totalizando 128 parcelas.
Nestas parcelas amostraram-se todos os indivíduos da regeneração (0,10 a 1,50
metros de altura e DAP < 2,5 cm), incluindo plântulas e indivíduos juvenis. Todos os
indivíduos foram medidos e devidamente determinados.
Com os dados levantados, obteve-se o Valor de Importância da Regeneração
Natural (VIRN), que segundo Ribeiro & Felfili (2009), é calculado pela seguinte
fórmula: VIRN= DR + FR, sendo necessário então avaliar os parâmetros
fitossociológicos densidade relativa (DR), frequência absoluta (FA) e frequência
relativa (FR) (Munhoz & Araújo, 2011).
1.1.1. Abertura de dossel
A abertura do dossel nos núcleos de vegetação e na matriz graminóide foi obtida
seguindo metodologia de Abreu et al. (2011), com fotografias utilizando lente
hemisférica do tipo fisheye (Nikon FC-08) acoplada à câmera Nikon modelo Coolpix
4500. Foram feitas fotografias ao lado de cada parcela 1 x 2 m (N1, N2, P1, P2), a 1
metro de altura, em condições de céu nublado, ou antes do nascer e depois do pôr do
sol, evitando-se que a luz incidisse diretamente na lente e alterasse os resultados. As
fotos foram preparadas no programa SideLook 1.0, com “threshold” em modo
automático e o máximo contraste de azul, e após a conversão da imagem em preto e
branco foi feita a análise no programa Gap Light Analyzer 2.0.
1.1.2. Dominância de gramíneas
Para classificar as áreas de interior de núcleos e matriz graminóide quanto à
dominância de gramíneas, primeiramente foi obtida visualmente a cobertura de
gramínea (%) que ocupou o interior das parcelas 1 x 2 m (N1, N2, P1, P2). Em seguida,
no centro das mesmas parcelas, obteve-se a altura máxima da gramínea dominante com
o uso de uma vara graduada.
1.1.3. Aporte de serrapilheira
Para a caracterização do aporte de serrapilheira foi obtida primeiramente a taxa
de acúmulo de serrapilheira (Kg/m²). Para isso, foi utilizado gabarito de 25 x 25 cm para
a coleta do material, que para não afetar os estudos de regeneração, foi realizada ao lado

13
das parcelas 1 x 2 m (N1, N2, P1, P2), sendo coletadas seis amostras para formar uma
composta por parcela (Figura 6). Em seguida, as amostras foram acondicionadas em
sacos de papel para, em laboratório, serem secos em estufa a 70°C até atingir peso
constante (Miranda, 2012). Os trabalhos de laboratório foram realizados no Laboratório
de Manejo de Bacias Hidrográficas da Universidade Federal Rural do Rio de Janeiro
(LMBH/UFRRJ).
Neste mesmo processo, foi feita a pesagem da serrapilheira antes de ser levada à
estufa (peso úmido), e também pesada após estufa (peso seco), tendo-se assim a
umidade retida na serrapilheira.
Figura 6: Esquema das coletas de serrapilheira nas parcelas 1 x 2 metros no interior dos núcleos e em
matriz graminóide.
1.1.4. Análise de Solo
Foram coletadas quatro amostras simples para formar uma composta, em duas
das quatro parcelas alocadas (N1 e P2), em cada núcleo amostrado (Figura 12). Para não
afetar os estudos de regeneração, as coletas foram realizadas ao lado das parcelas 1 x 2
m (N1e P2).
A coleta do solo foi realizada segundo as instruções de Embrapa (2012),
retirando-se com um trado as amostras da camada superficial do solo, numa
profundidade de até 20 centímetros, limpando anteriormente a superfície e removendo
as folhas e outros detritos.
Foram retiradas amostras com cerca de 500 gramas, que foram devidamente
identificadas e enviadas para a análise no Departamento de Ciência do Solo da
Universidade Federal de Lavras. Foram realizadas análises em relação aos parâmetros
usuais químicos e de textura.

14
Figura 7: Esquema das coletas de solo nas parcelas 1 x 2 metros no interior dos núcleos e em matriz
graminóide.
1.1. Análises dos Dados
Para o estudo das relações e influências presentes na sucessão de pastagens
abandonadas, foram feitos testes de correlação de Pearson (r) para comparar os
seguintes parâmetros: área do núcleo de vegetação e abundância do estrato superior,
área do núcleo de vegetação e riqueza do estrato superior, riqueza do estrato superior e
riqueza do estrato regenerante, área do núcleo de vegetação e abundância do estrato
regenerante, abundância do estrato superior e abundância do estrato regenerante.
Também foi realizado o teste “análise de espécies indicadoras”, objetivando
encontrar a espécie indicadora de cada classe de tamanho de núcleo analisada. Para a
realização deste teste, foi necessário que as três classes de núcleo (pequeno, médio e
grande) tivessem a mesma quantidade de núcleos. Para isso, foi realizado sorteio para a
escolha aleatória dos núcleos que deveriam ser excluídos da análise. Com isso, as três
classes apresentaram 15 núcleos.
Para relacionar os fatores influenciados pelos núcleos de vegetação
arbustivo/arbóreos foram feitos testes de regressão linear para os seguintes parâmetros:
altura de gramíneas e abundância de regenerantes, cobertura de gramíneas e abundância
de regenerantes, serrapilheira acumulada e abundância de regenerantes, umidade retida
na serrapilheira e abundância de regenerantes, abertura do dossel e abundância de
regenerantes. Para todas as análises de regressão linear realizadas, foi feito
anteriormente teste de Shapiro-Wilk (Shapiro & Wilk, 1965) para avaliar a normalidade
dos dados, e quando necessário, o grupo de dados foi normalizado através de logaritmo.

15
Para avaliar a relação dos fatores com o estrato regenerante, foi realizada uma
ordenação através da Análise de Componentes Principais (PCA). Desta forma, foi
possível observar o nível de relação entre os fatores e a regeneração natural.
Para os dados de solo, primeiramente foi realizada uma PCA para testar a
diferença das variáveis de solo entre núcleo de vegetação e matriz graminóide.
Posteriormente, foi realizada análise de redundância (RDA) para relacionar a
abundância das espécies encontradas nos estratos arbustivo/arbóreo e regenerante
(variáveis dependentes) com as variáveis do solo (variáveis independentes). Para a
realização da RDA, foi feita a normalização dos dados de abundância (função
“normalize”) e das variáveis (função “standardize”), e encontrado-se quais fatores de
solo são significativos através de ANOVA (p < 0,05), onde foi feita a escolha das
variáveis para a realização da RDA.
Todas as análises estatísticas foram feitas no software R 2.13.2 (R Development
Core Team, 2013), com a utilização dos pacotes labdsv (Roberts, 2013) para realização
do teste de análise de espécies indicadoras, e vegan 2.0-9 (Oksanen et al., 2013) para os
demais testes.
3. RESULTADOS
1.1. Sucessão ecológica em pastagem abandonada
1.1.1. Estrato superior
Foram amostrados ao todo 468 indivíduos, pertencentes a 33 espécies e 23
famílias. Dos 50 núcleos de vegetação levantados, 300 indivíduos foram encontrados
compondo esses núcleos e 168 indivíduos foram encontrados de forma isolada pela
pastagem. Na Tabela 1 são apresentadas as espécies encontradas em ambas as
formações.
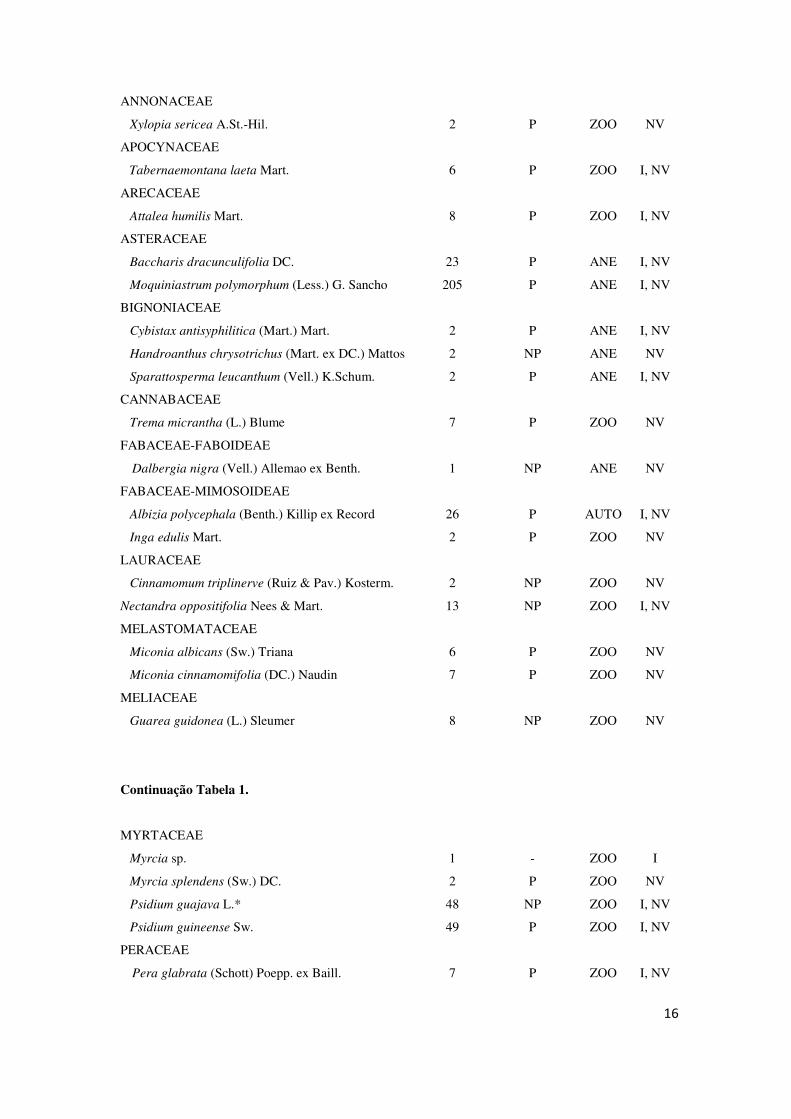
Tabela 1: Composição de espécies arbustivo/arbóreas e suas abundâncias (ABUND), grupo sucessional
(GS), síndrome de dispersão (SD) e forma de ocorrência (FO) em pastagem abandonada, Reserva
Biológica de Poço das Antas, Silva Jardim – RJ. Legenda: GS: P – pioneira; NP – não pioneira. SD: ZOO
– zoocoria; ANE – anemocoria; AUTO – autocoria. FO: I – isoladas; NV – núcleos de vegetação.
*espécie exótica.
FAMÍLIA/ESPÉCIE ABUND GS SD FO
ANACARDIACEAE
Schinus terebinthifolius Raddi
Tapirira guianensis Aubl.
9
1
P
P
ZOO
ZOO
NV
NV
16
ANNONACEAE
Xylopia sericea A.St.-Hil.
2
P
ZOO
NV
APOCYNACEAE
Tabernaemontana laeta Mart.
6 P ZOO I, NV
ARECACEAE
Attalea humilis Mart.
8
P
ZOO
I, NV
ASTERACEAE
Baccharis dracunculifolia DC.
Moquiniastrum polymorphum (Less.) G. Sancho
23
205
P
P
ANE
ANE
I, NV
I, NV
BIGNONIACEAE
Cybistax antisyphilitica (Mart.) Mart.
Handroanthus chrysotrichus (Mart. ex DC.) Mattos
Sparattosperma leucanthum (Vell.) K.Schum.
2
2
2
P
NP
P
ANE
ANE
ANE
I, NV
NV
I, NV
CANNABACEAE
Trema micrantha (L.) Blume
7 P ZOO
NV
FABACEAE-FABOIDEAE
Dalbergia nigra (Vell.) Allemao ex Benth.
1 NP ANE NV
FABACEAE-MIMOSOIDEAE
Albizia polycephala (Benth.) Killip ex Record
Inga edulis Mart.
26
2
P
P
AUTO
ZOO
I, NV
NV
LAURACEAE
Cinnamomum triplinerve (Ruiz & Pav.) Kosterm.
Nectandra oppositifolia Nees & Mart.
2
13
NP
NP
ZOO
ZOO
NV
I, NV
MELASTOMATACEAE
Miconia albicans (Sw.) Triana
Miconia cinnamomifolia (DC.) Naudin
6
7
P
P
ZOO
ZOO
NV
NV
MELIACEAE
Guarea guidonea (L.) Sleumer
8 NP ZOO NV
Continuação Tabela 1.
MYRTACEAE
Myrcia sp.
Myrcia splendens (Sw.) DC.
Psidium guajava L.*
Psidium guineense Sw.
1
2
48
49
-
P
NP
P
ZOO
ZOO
ZOO
ZOO
I
NV
I, NV
I, NV
PERACEAE
Pera glabrata (Schott) Poepp. ex Baill.
7 P ZOO I, NV

17
PRIMULACEAE
Myrsine coriacea (Sw.) R.Br. ex Roem. & Schult.
4 P ZOO NV
RUBIACEAE
Tocoyena bullata (Vell.) Mart.
1
NP
ZOO
NV
RUTACEAE
Zanthoxylum rhoifolium Lam.
1 P ZOO NV
SALICACEAE
Casearia sylvestris Sw.
4 P ZOO NV
SAPINDACEAE
Cupania racemosa (Vell.) Radlk.
3 P ZOO NV
SIPARUNACEAE
Siparuna guianensis Aubl.
9
NP
ZOO
NV
SOLANACEAE
Solanum inaequilaterum Domin.
1 - ZOO NV
STYRACACEAE
Styrax glabratum Spreng.
2 - ZOO NV
URTICACEAE
Cecropia pachystachya Trécul
4 P ZOO I, NV
A única espécie encontrada exclusivamente de forma isolada foi Myrcia sp.,
enquanto Schinus terebinthifolius, Tapirira guianensis, Xylopia sericea,
Handroanthus chrysotrichus, Trema micrantha, Dalbergia nigra, Inga edulis,
Cinnamomum triplinerve, Miconia albicans, Miconia cinnamomifolia, Guarea
guidonea, Myrcia splendens, Myrsine coriacea, Tocoyena bullata,
Zanthoxylum rhoifolium, Casearia sylvestris, Cupania racemosa, Siparuna guianensis,
Solanum inaequilaterum e Styrax glabratum foram encontradas exclusivamente em
núcleos de vegetação.
A espécie mais abundante, tanto ocorrendo de forma isolada como em núcleos
de vegetação arbustivo/arbóreos foi Moquiniastrum polymorphum, com 72 indivíduos
isolados e 133 indivíduos em núcleos, fazendo com que Asteraceae se tornasse a família
mais representativa. A seguir observa-se a riqueza de espécies nas duas circunstâncias,
com destaque para a espécie de maior abundância (Figura 8).

18
Figura 8: Diferentes riquezas e espécies mais abundantes encontradas formando núcleos de vegetação
arbustivo/arbóreos (A) e de forma isolada (B).
Na figura anterior se observa que, embora a espécie dominante tenha sido
Moquiniastrum polymorphum, foram nos núcleos de vegetação que se encontrou o
maior número de espécies.
A contribuição de cada espécie para as áreas de pastagem, sendo indivíduos
isolados ou formadores de núcleos, é apresentada nos resultados de fitossociologia
(Apêndices 1 e 2). A espécie Attalea humilis, por ser acaule, foi excluída desta análise.
Estes resultados confirmam que, tanto nas formações isoladas (40,59%) como
em núcleos de vegetação (38,59%), a espécie de maior importância é Moquiniastrum
polymorphum, sendo esta uma espécie pioneira e de dispersão anemocórica. Entretanto,
deve-se levar em consideração que o valor de VI desta espécie foi maior de forma
isolada do que em núcleo, e foi entre as isoladas que se encontrou um número menor de
espécies. A diferença de VI entre Moquiniastrum polymorphum e Psidium guajava (2º
maior VI) em espécies de núcleo foi elevada, inferindo a importância da espécie para a
formação, e também sugerindo que isso pode ter acontecido pelo grande número de
espécies encontradas.
A riqueza também apresenta diferença quando analisada nas diferentes classes
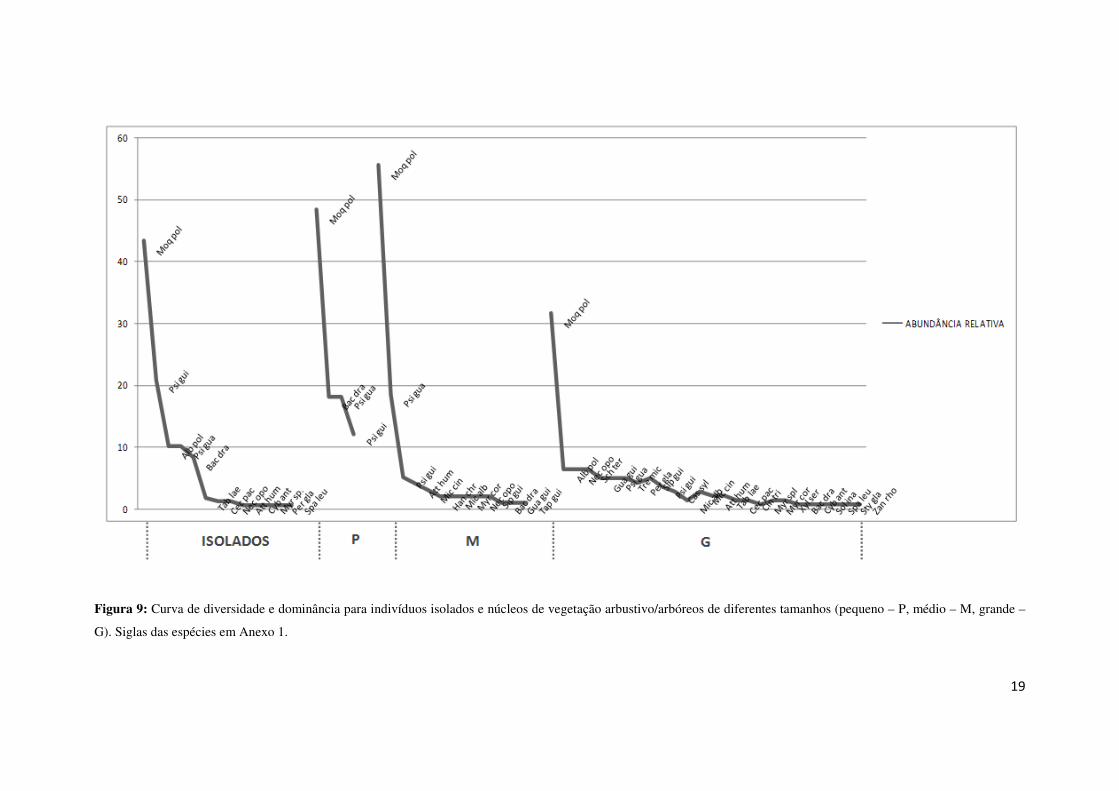
de núcleos. Na Figura 9 observa-se a variação no padrão de diversidade e dominância
de cada tipo de formação.
A B
Psidium guajava (31) Psidium guineense (35)
Moquiniastrum polymorphum (133) Moquiniastrum polymorphum (72)
19
Figura 9: Curva de diversidade e dominância para indivíduos isolados e núcleos de vegetação arbustivo/arbóreos de diferentes tamanhos (pequeno – P, médio – M, grande –
G). Siglas das espécies em Anexo 1.

20
A riqueza nos núcleos grandes é maior que nos núcleos pequenos, mostrando
que há uma evolução com o aumento da área do núcleo. Entretanto, também encontrou-
se mais espécies de indivíduos isolados que em núcleos pequenos, sugerindo que esta
classe de núcleos também está sujeita a desaparecer e gerar indivíduos isolados. Ainda
assim, nas quatro situações, Moquiniastrum polymorphum é a espécie mais abundante,
se destacando por apresentar uma abundância bastante superior às demais espécies.
Os núcleos grandes apresentam maior número de indivíduos zoocóricos,
enquanto que os demais núcleos e os indivíduos isolados são representados por
indivíduos anemocóricos (Figura 10), principalmente devido à abundância de
Moquiniastrum polymorphum. Também devido a esta espécie, a maioria dos indivíduos
encontrados são pioneiros, em ambas as classes (Figura 11).
0
10
20
30
40
50
60
70
Isoladas Pequeno Médio Grande
Nú
me
ro d
e i
nd
ivíd
uo
s (%
)
ANE
AUTO
ZOO
Figura 10: Síndrome de dispersão do estrato superior apresentado em porcentagem, para os indivíduos
isolados e diferentes classes de núcleos. ANE: anemocórica; AUTO: autocórica; ZOO: zoocórica.
0
10
20
30
40
50
60
70
80
90
100
Isoladas Pequeno Médio Grande
Nú
me
ro d
e in
div
ídu
os
(%)
P
NP
Figura 11: Grupo sucessional do estrato superior apresentado em porcentagem, para os indivíduos
isolados e diferentes classes de núcleos. P: pioneiras; NP: não pioneiras.

21
A relação entre a área do núcleo de vegetação arbustivo/arbóreo com a
abundância e riqueza do estrato superior mostra que a área do núcleo apresentou
correlação maior com a abundância de árvores (r = 0,67; DF = 46; P < 0,0001) do que
com a riqueza (r = 0,49; DF = 46; P = 0,00039). Em ambas as variáveis a correlação é
positiva, mostrando que quanto maior a área, maior também será a abundância e a
riqueza. Entretanto, a correlação com a riqueza não apresentou significância (P >
0,0001).
A análise de espécies indicadoras mostrou que algumas espécies são indicadoras
de diferentes classes de núcleos de vegetação, relacionando a presença das mesmas com
os diferentes estágios dos núcleos. A Tabela 2 apresenta os resultados significativos da
análise (p = 0,05).
Tabela 2: Análises de espécies indicadora das diferentes classes de núcleos de vegetação
arbustivo/arbóreo (pequeno – P, médio – M, grande – G).
Espécie P (%)
M (%)
G (%)
Maior % indicador
Classe indicada
P valor
Siparuna guianensis 0,000 0,000 0,333 0,333 G 0,004
Baccharis dracunculifolia 0,259 0,007 0,007 0,259 P 0,052
Nectandra opositifolia 0,000 0,007 0,240 0,240 G 0,058
O teste apresentou significância das espécies Siparuna guianensis e Nectandra
opositifolia como sendo indicadoras dos núcleos grandes, e Baccharis dracunculifolia
indicando os núcleos pequenos. Como as demais espécies não foram significativas, não
foram apresentadas na tabela.
1.1.2. Estrato regenerante
A maioria das espécies encontradas no estrato regenerante são as mesmas que
compõe o estrato superior, que são as fontes de propágulos mais próximas. A diferença
é encontrada principalmente no número de indivíduos de algumas espécies que compõe
tanto o estrato regenerante como o superior. Na Tabela 3 estão apresentados estes
valores de abundância, assim como o grupo sucessional e síndrome de dispersão.
22
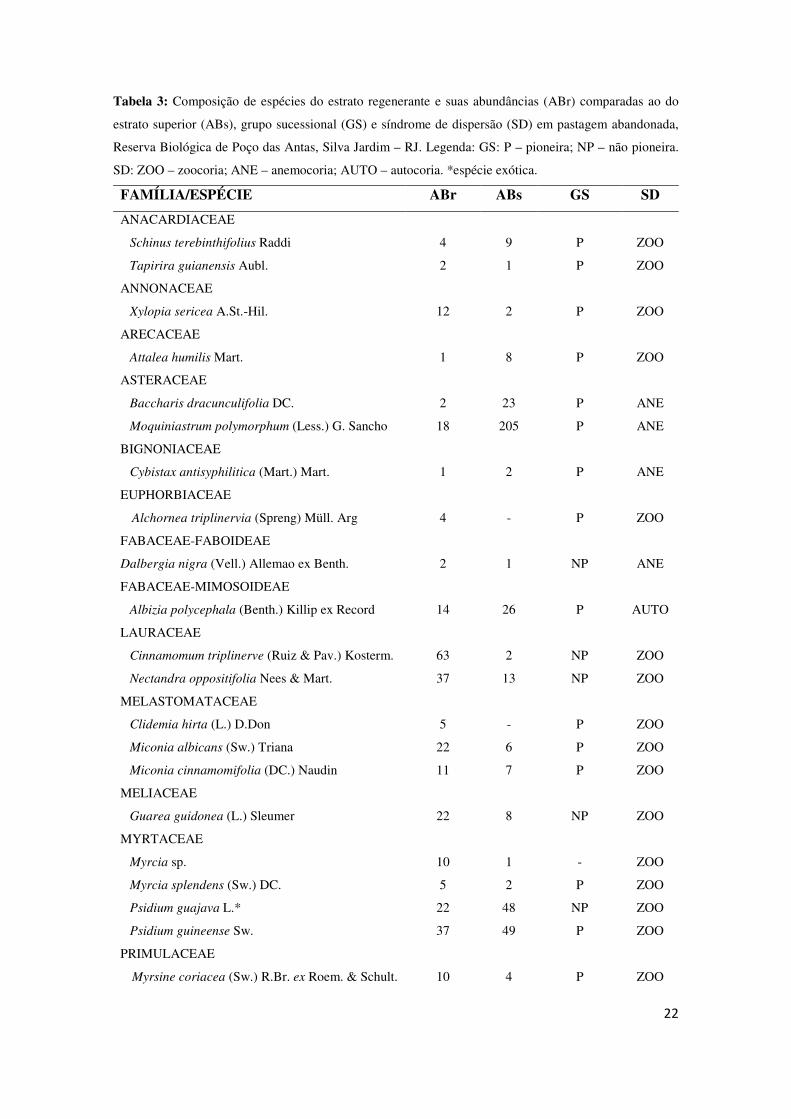
Tabela 3: Composição de espécies do estrato regenerante e suas abundâncias (ABr) comparadas ao do
estrato superior (ABs), grupo sucessional (GS) e síndrome de dispersão (SD) em pastagem abandonada,
Reserva Biológica de Poço das Antas, Silva Jardim – RJ. Legenda: GS: P – pioneira; NP – não pioneira.
SD: ZOO – zoocoria; ANE – anemocoria; AUTO – autocoria. *espécie exótica.
FAMÍLIA/ESPÉCIE ABr ABs GS SD
ANACARDIACEAE
Schinus terebinthifolius Raddi
Tapirira guianensis Aubl.
4
2
9
1
P
P
ZOO
ZOO
ANNONACEAE
Xylopia sericea A.St.-Hil.
12
2 P ZOO
ARECACEAE
Attalea humilis Mart.
1
8 P ZOO
ASTERACEAE
Baccharis dracunculifolia DC.
Moquiniastrum polymorphum (Less.) G. Sancho
2
18
23
205
P
P
ANE
ANE
BIGNONIACEAE
Cybistax antisyphilitica (Mart.) Mart.
1
2 P ANE
EUPHORBIACEAE
Alchornea triplinervia (Spreng) Müll. Arg
4
- P ZOO
FABACEAE-FABOIDEAE
Dalbergia nigra (Vell.) Allemao ex Benth.
2
1
NP ANE
FABACEAE-MIMOSOIDEAE
Albizia polycephala (Benth.) Killip ex Record
14
26 P AUTO
LAURACEAE
Cinnamomum triplinerve (Ruiz & Pav.) Kosterm.
Nectandra oppositifolia Nees & Mart.
63
37
2
13
NP
NP
ZOO
ZOO
MELASTOMATACEAE
Clidemia hirta (L.) D.Don
Miconia albicans (Sw.) Triana
Miconia cinnamomifolia (DC.) Naudin
5
22
11
-
6
7
P
P
P
ZOO
ZOO
ZOO
MELIACEAE
Guarea guidonea (L.) Sleumer
22
8 NP ZOO
MYRTACEAE
Myrcia sp.
Myrcia splendens (Sw.) DC.
Psidium guajava L.*
Psidium guineense Sw.
10
5
22
37
1
2
48
49
-
P
NP
P
ZOO
ZOO
ZOO
ZOO
PRIMULACEAE
Myrsine coriacea (Sw.) R.Br. ex Roem. & Schult.
10
4 P ZOO

23
Continuação Tabela 3.
SIPARUNACEAE
Siparuna guianensis Aubl.
78
9 NP ZOO
As espécies Xylopia sericea, Cinnamomum triplinerve, Nectandra oppositifolia,
Miconia albicans, Guarea guidonea, Myrcia sp., e Siparuna guianensis se destacaram
por ocorrerem pouco no estrato superior, mas que aumentaram consideravelmente o
número de indivíduos no estrato regenerante. Já com as espécies Baccharis
dracunculifolia, Moquiniastrum polymorphum e Psidium guajava ocorreu o contrário,
foram espécies de grande abundância no estrato superior, mas pouco encontrada na
regeneração. A espécie Moquiniastrum polymorphum é a que mais se destaca, por ser a
espécie mais abundante entre as arbóreas adultas (205 indivíduos), encontrando-se
somente 18 indivíduos no estrato regenerante.
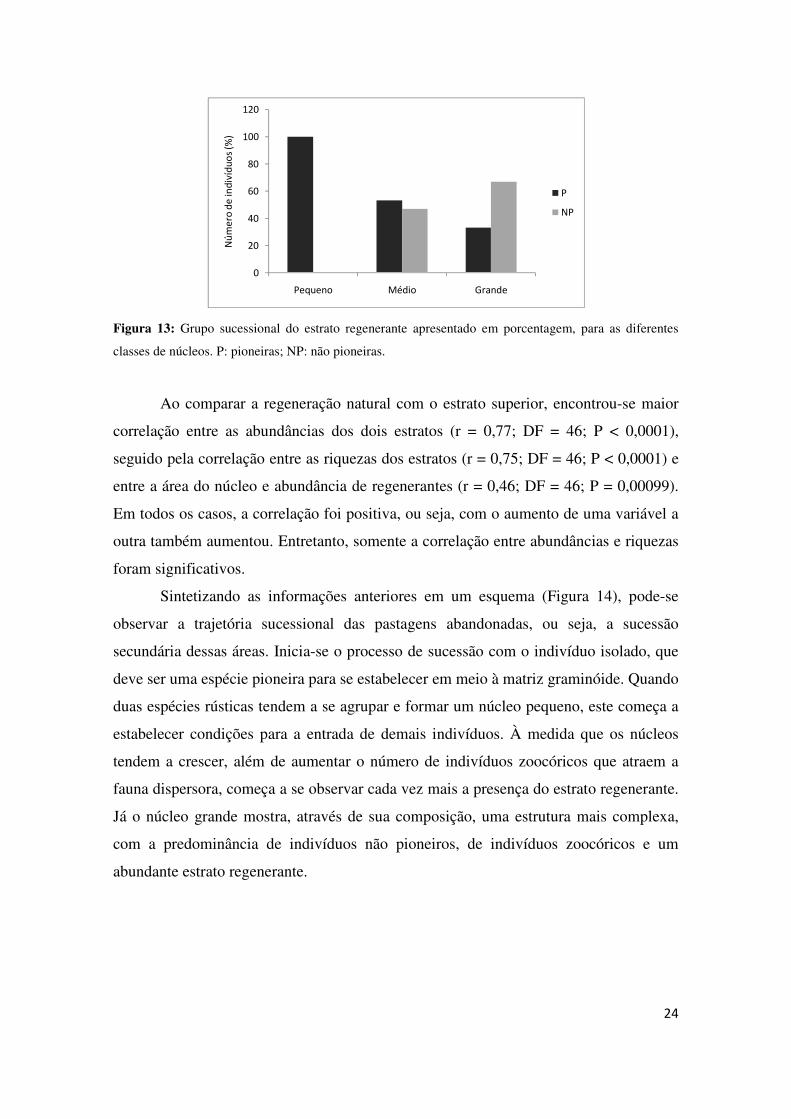
Analisando a síndrome de dispersão e o grupo sucessional (Figuras 12 e 13), é
possível verificar que à medida que os núcleos aumentam sua área, há um aumento de
indivíduos zoocóricos e não pioneiros e, consequentemente, a diminuição dos
indivíduos pioneiros e anemocóricos. Nos gráficos também se observa que os núcleos
grandes apresentam maior número de indivíduos regenerantes que as demais classes.
0
10
20
30
40
50
60
70
80
90
100
Pequeno Médio Grande
Nú
me
ro d
e i
nd
ivíd
uo
s (%
)
ANE
AUTO
ZOO
Figura 12: Síndrome de dispersão do estrato regenerante apresentado em porcentagem, para as diferentes
classes de núcleos. ANE: anemocórica; AUTO: autocórica; ZOO: zoocórica.
24
0
20
40
60
80
100
120
Pequeno Médio Grande
Nú
me
ro d
e in
div
ídu
os
(%)
P
NP
Figura 13: Grupo sucessional do estrato regenerante apresentado em porcentagem, para as diferentes
classes de núcleos. P: pioneiras; NP: não pioneiras.
Ao comparar a regeneração natural com o estrato superior, encontrou-se maior
correlação entre as abundâncias dos dois estratos (r = 0,77; DF = 46; P < 0,0001),
seguido pela correlação entre as riquezas dos estratos (r = 0,75; DF = 46; P < 0,0001) e
entre a área do núcleo e abundância de regenerantes (r = 0,46; DF = 46; P = 0,00099).
Em todos os casos, a correlação foi positiva, ou seja, com o aumento de uma variável a
outra também aumentou. Entretanto, somente a correlação entre abundâncias e riquezas
foram significativos.
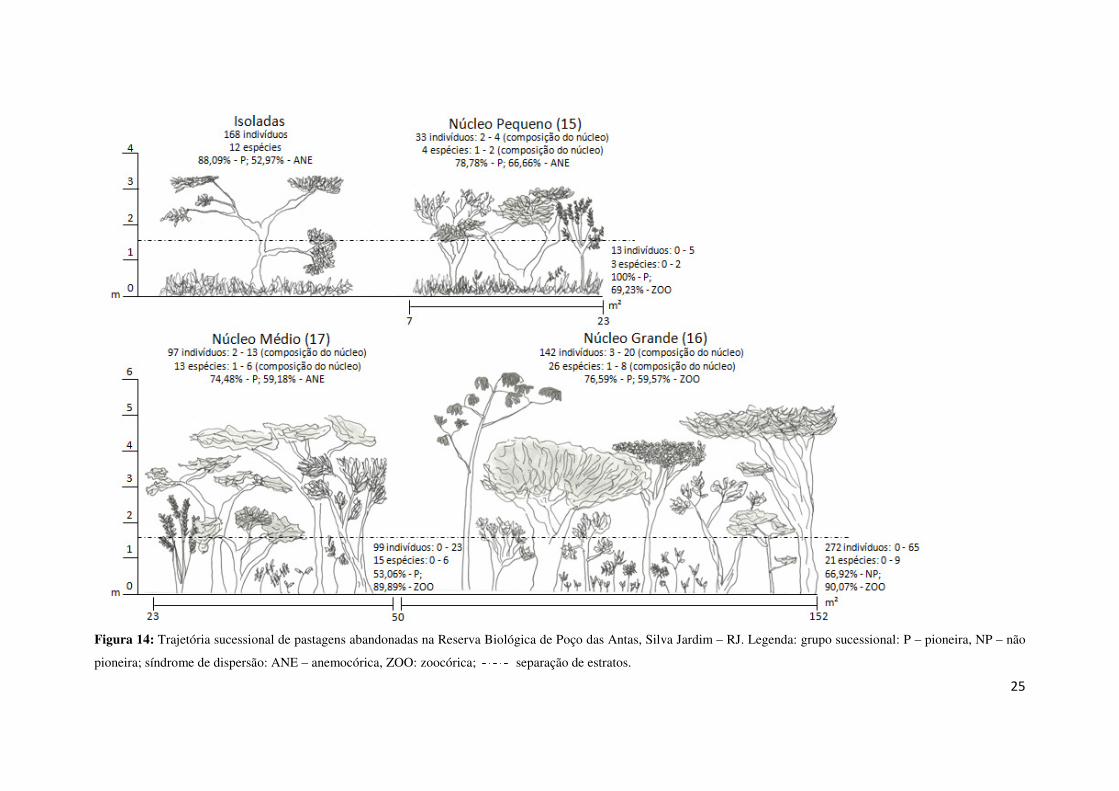
Sintetizando as informações anteriores em um esquema (Figura 14), pode-se
observar a trajetória sucessional das pastagens abandonadas, ou seja, a sucessão
secundária dessas áreas. Inicia-se o processo de sucessão com o indivíduo isolado, que
deve ser uma espécie pioneira para se estabelecer em meio à matriz graminóide. Quando
duas espécies rústicas tendem a se agrupar e formar um núcleo pequeno, este começa a
estabelecer condições para a entrada de demais indivíduos. À medida que os núcleos
tendem a crescer, além de aumentar o número de indivíduos zoocóricos que atraem a
fauna dispersora, começa a se observar cada vez mais a presença do estrato regenerante.
Já o núcleo grande mostra, através de sua composição, uma estrutura mais complexa,
com a predominância de indivíduos não pioneiros, de indivíduos zoocóricos e um
abundante estrato regenerante.
25
Figura 14: Trajetória sucessional de pastagens abandonadas na Reserva Biológica de Poço das Antas, Silva Jardim – RJ. Legenda: grupo sucessional: P – pioneira, NP – não
pioneira; síndrome de dispersão: ANE – anemocórica, ZOO: zoocórica; separação de estratos.

26
1.1. Fatores influenciados pelos núcleos de vegetação
Foram amostrados nas 128 parcelas um total de 97 indivíduos. Nas 64 parcelas
do interior dos núcleos encontrou-se 84 indivíduos, sendo a espécie mais abundante
Siparuna guianensis, com 16 indivíduos. Já nas 64 parcelas da matriz graminóide foram
amostrados 13 indivíduos compondo o estrato regenerante, sendo a espécie mais
abundante Miconia albicans com 5 indivíduos. Os valores podem ser reforçados com a
análise fitossociológica apresentadas nos Apêndices 3 e 4.
Nas parcelas do interior do núcleo, as espécies de maior VIRN foram
Siparuna guianensis (30,81%), Cinnamomum triplinerve (22,55%), e Guarea guidonia
(17,79%), enquanto que nas parcelas da matriz graminóide foram Miconia albicans
(71,79%), Moquiniastrum polymorphum (26,50%), e Psidium guineense (26,50%).
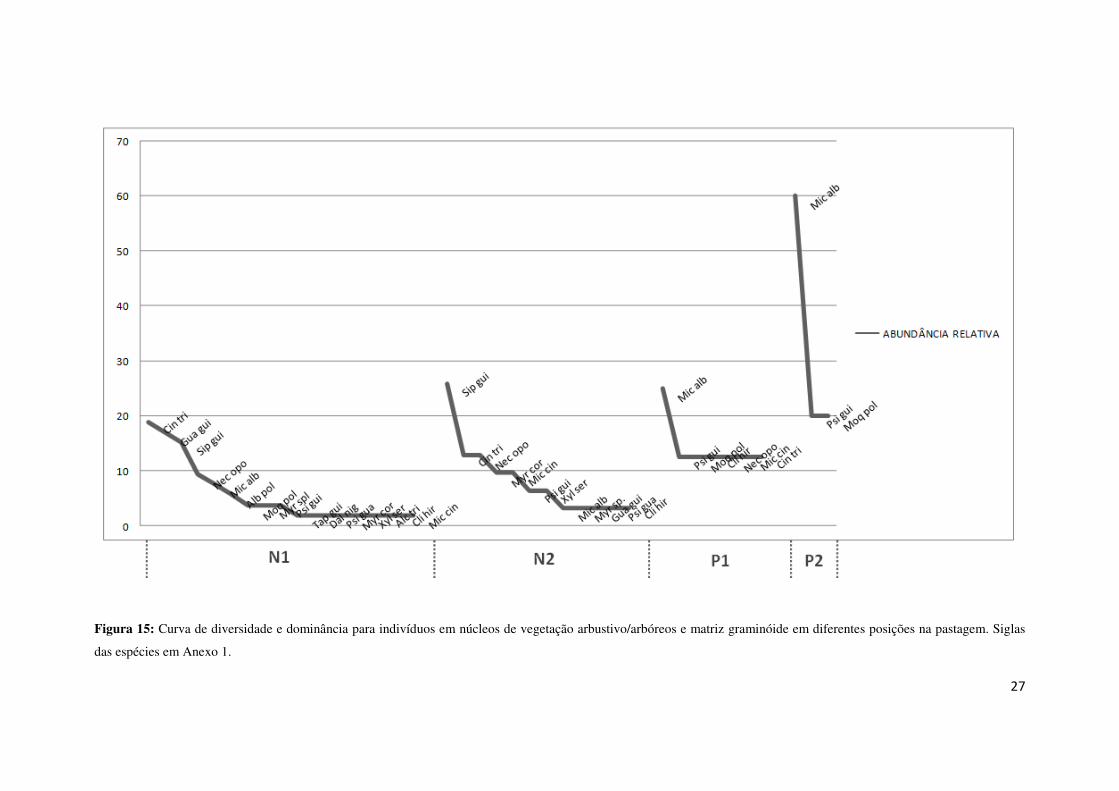
Diferenças também são observadas ao comparar o estrato regenerante entre as
quatro parcelas alocadas (N1, N2, P1, P2). A composição e abundância da regeneração
são mostradas a seguir (Figura 15).
27
Figura 15: Curva de diversidade e dominância para indivíduos em núcleos de vegetação arbustivo/arbóreos e matriz graminóide em diferentes posições na pastagem. Siglas
das espécies em Anexo 1.

28
A figura anterior mostra que a maior riqueza é encontrada no interior dos
núcleos de vegetação, e à medida que vai se afastando do mesmo, a riqueza de espécies
vai diminuindo. Na N2 vê a presença de espécies não pioneiras, mas ainda há a
dominância de uma espécie, sendo esta Siparuna guianensis. Na N1 não se vê uma
dominância muito acentuada, apesar de Cinnamomum triplinerve ter ocorrido mais na
área. A mesma também é composta por indivíduos não pioneiros, mostrando que no
interior dos núcleos o processo de sucessão secundária é mais dinâmico que nos outros
locais.
Na P2, a dominância de Miconia albicans é muito superior que as das demais
espécies. Além da baixa diversidade, todas as espécies que compões esta parcela são
pioneiras. Já entre P1 e P2 infere que, áreas mais próximas aos núcleos de vegetação são
áreas com maior riqueza.
Estes resultados mostram que os núcleos de vegetação afetam de alguma forma
o desenvolvimento da regeneração natural. Na Tabela 4 estão representados os
diferentes fatores que podem explicar a variação encontrada anteriormente.
Tabela 4: Médias dos fatores e número de indivíduos na matriz graminóide e em núcleos de vegetação
arbustivo/arbóreos de pastagens abandonadas na Reserva Biológica de Poço das Antas, Silva Jardim – RJ.
Núcleo de vegetação Matriz graminóide
Número de indivíduos 84 13
Cobertura gramínea (%) 57,28 83,17
Altura gramínea (m) 0,86 0,97
Serrapilheira acumulada (Kg/m²) 7,58 6,49
Umidade retida na serrapilheira (g) 143,13 129,69
Abertura do dossel (%) 36,14 55,76
A tabela mostra que valores como o de cobertura e altura de gramíneas, e
abertura do dossel são maiores na matriz graminóide do que em núcleos de vegetação.
O conjunto de menor presença de gramíneas, menor abertura do dossel, maior umidade
e serrapilheira acumulada reflete no número de indivíduos do estrato regenerante, que
foi maior no interior dos núcleos de vegetação.

29
Foram encontrados baixos coeficientes de regressão (R²) para todos seguintes
parâmetros (Figura 16): cobertura de gramíneas e abundância do estrato regenerante (R²
= 18%; F = 30,35; DF = 126; P < 0,0001), altura de gramíneas e abundância do estrato
regenerante (R² = 11%; F = 16; DF = 126; P = 0,0001075), serrapilheira acumulada e
abundância do estrato regenerante (R² = -0,8%; F = 0,001035; DF = 126; P = 0,9744),
umidade retida na serrapilheira e abundância do estrato regenerante (R² = 3%; F =
4,196; DF = 126; P = 0,0426), abertura do dossel e abundância do estrato regenerante
(R² = 20%; F = 31,16; DF = 126; P < 0,0001).

30
Figura 16: Gráficos gerados com testes de regressão linear para responder o comportamento da
regeneração natural à diferentes fatores ambientais.
R² = 18% p < 0,0001 R² = 11% p = 0,0001075
R² = 0,8% p = 0,9744 R² = 3% p = 0,0426
R² = 20% p < 0,0001

31
Na regressão, somente os valores de cobertura de gramíneas e abertura do dossel
apresentaram significância (P < 0,0001).Foi possível observar, através do gráfico de
abertura, que há uma faixa de núcleos, entre 20 e 80%, onde o número de regenerantes é
nulo.
A Análise de Componentes Principais (PCA) mostrou uma distribuição
sobreposta das parcelas (Figura 17).
Figura 17: Análise de Componentes Principais (PCA) que infere a relação entre o estrato regenerante e
variáveis, em núcleos de vegetação arbustivo/arbóreos e matriz graminóide. Legenda: hcap - altura do
capim; cobcap - cobertura de capim; abert - abertura do dossel; seracum - serrapilheira acumulada;
umidser - umidade retida na serrapilheira.
O teste apresentou 37% de significância do primeiro eixo e 28% do segundo
eixo, que mostraram o agrupamento das parcelas dos núcleos e da matriz graminóide em
relação às variáveis. O gráfico não apresentou uma distribuição clara, com a formação
de grupos. Pode-se destacar que, observando o primeiro eixo (PC1), a maioria das

32
parcelas de matriz graminóide (P1 e P2) estão mais próximas dos fatores de abertura do
dossel e cobertura de gramíneas do que as parcelas de núcleo (N1 e N2), por serem os
locais onde estas variáveis possuem valores mais elevados.
As variáveis do solo também mostraram comportamentos diferentes entre os
núcleos de vegetação e matriz graminóide (Apêndice 6). Foi possível constatar que nos
núcleos de vegetação foram encontrados mais nutrientes que na matriz graminóide. Isto
foi confirmado com a soma de bases trocáveis (SB), onde o valor foi mais alto nos
núcleos, apresentando então maior fertilidade no solo do interior dessas estruturas.
Entretanto, o valor de alumínio trocável (Al) também foi mais alto, sendo
maiores as chances de causar dano às plantas. Contrapondo este valor, o índice de
saturação de alumínio (m) foi maior na matriz graminóide, ou seja, maior é o efeito da
toxidez. Os solos da matriz graminóide pode inclusive ser classificados como álicos, por
ter apresentado um valor de m maior que 50%.
Apesar de ter sido encontrado maior fertilidade no interior dos núcleos, no geral
a fertilidade foi baixa, ficando evidente com os valores de índice de saturação de bases
(V). Os solos são considerados distróficos, devido ao baixo valor de V, que evidencia
que são solos de fertilidade média a baixa. Ainda assim, o valor de V foi maior no
interior dos núcleos de vegetação.
Os valores de matéria orgânica também foram mais elevados no interior dos
núcleos de vegetação, inferindo que nesses locais há maior oferta e deposição de
material orgânico, podendo ter também maior presença de organismos decompositores.
A análise também trouxe a informação de que solo da área, de uma forma geral, varia
entre textura média e argilosa, não variando entre núcleo e matriz graminóide.
Os resultados da PCA não mostraram um agrupamento claro dos núcleos de
vegetação e matriz graminóide, apesar da proporção cumulativa dos dois eixos
principais serem de 67% (Figura 18).
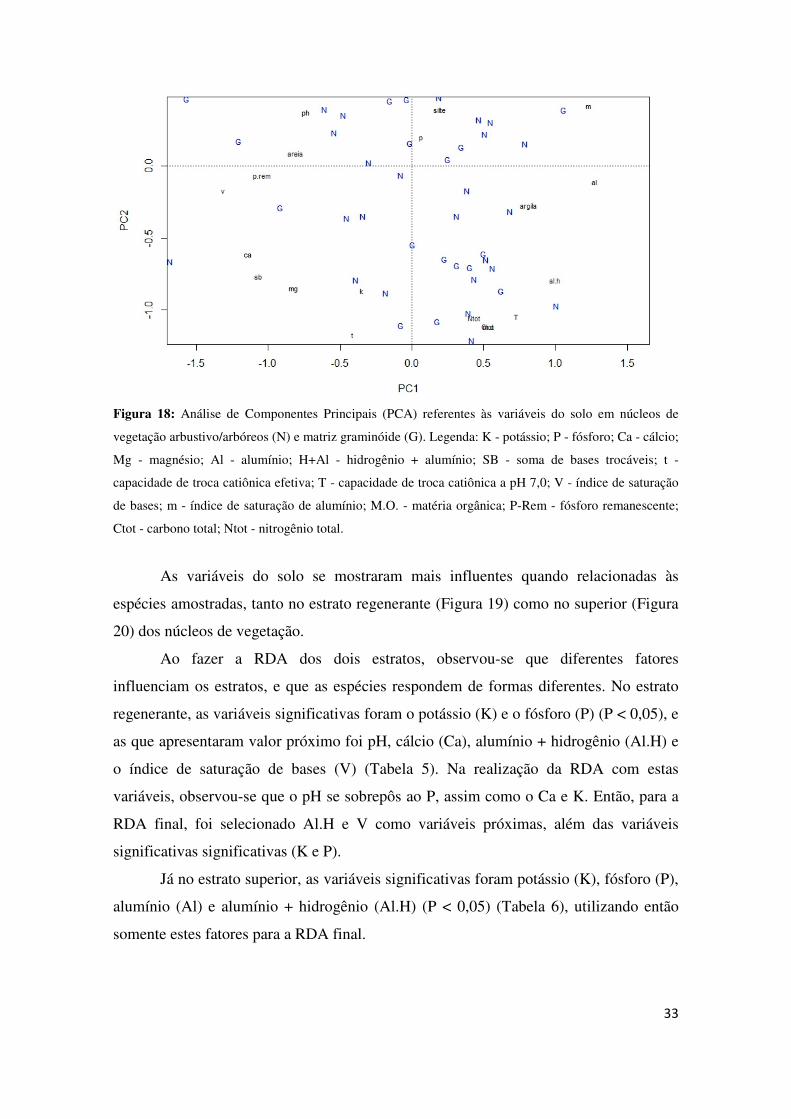
33
Figura 18: Análise de Componentes Principais (PCA) referentes às variáveis do solo em núcleos de
vegetação arbustivo/arbóreos (N) e matriz graminóide (G). Legenda: K - potássio; P - fósforo; Ca - cálcio;
Mg - magnésio; Al - alumínio; H+Al - hidrogênio + alumínio; SB - soma de bases trocáveis; t -
capacidade de troca catiônica efetiva; T - capacidade de troca catiônica a pH 7,0; V - índice de saturação
de bases; m - índice de saturação de alumínio; M.O. - matéria orgânica; P-Rem - fósforo remanescente;
Ctot - carbono total; Ntot - nitrogênio total.
As variáveis do solo se mostraram mais influentes quando relacionadas às
espécies amostradas, tanto no estrato regenerante (Figura 19) como no superior (Figura
20) dos núcleos de vegetação.
Ao fazer a RDA dos dois estratos, observou-se que diferentes fatores
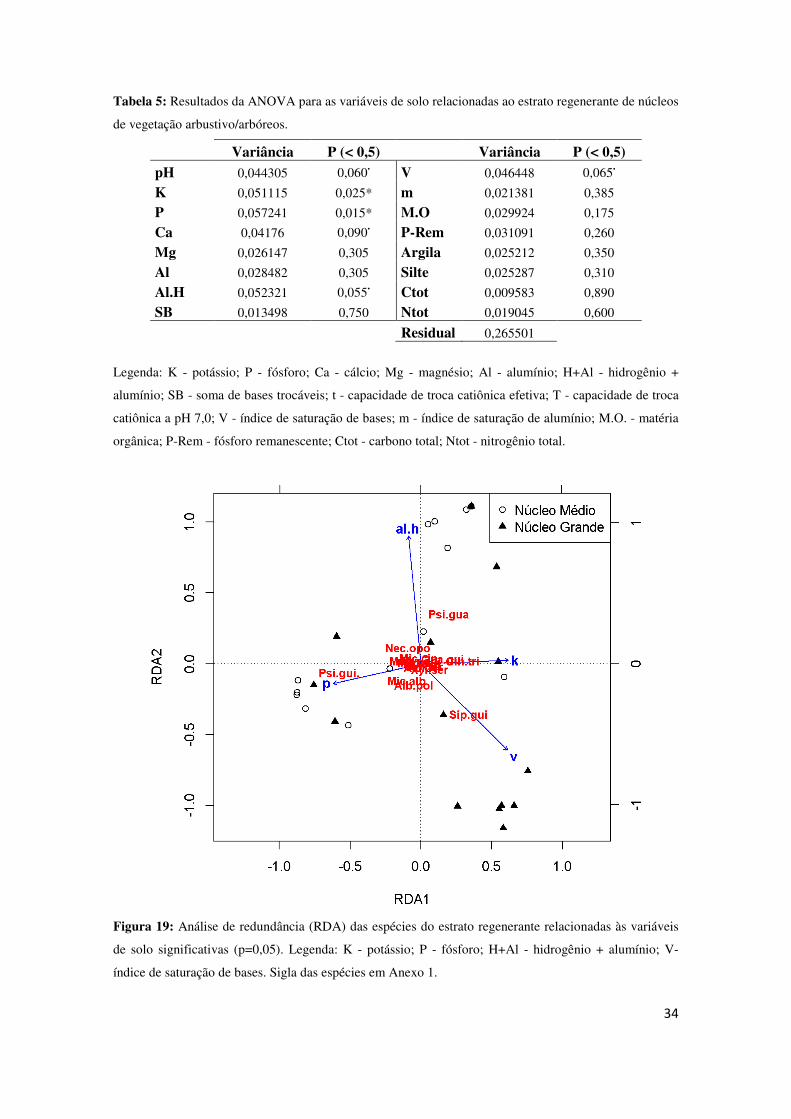
influenciam os estratos, e que as espécies respondem de formas diferentes. No estrato
regenerante, as variáveis significativas foram o potássio (K) e o fósforo (P) (P < 0,05), e
as que apresentaram valor próximo foi pH, cálcio (Ca), alumínio + hidrogênio (Al.H) e
o índice de saturação de bases (V) (Tabela 5). Na realização da RDA com estas
variáveis, observou-se que o pH se sobrepôs ao P, assim como o Ca e K. Então, para a
RDA final, foi selecionado Al.H e V como variáveis próximas, além das variáveis
significativas significativas (K e P).
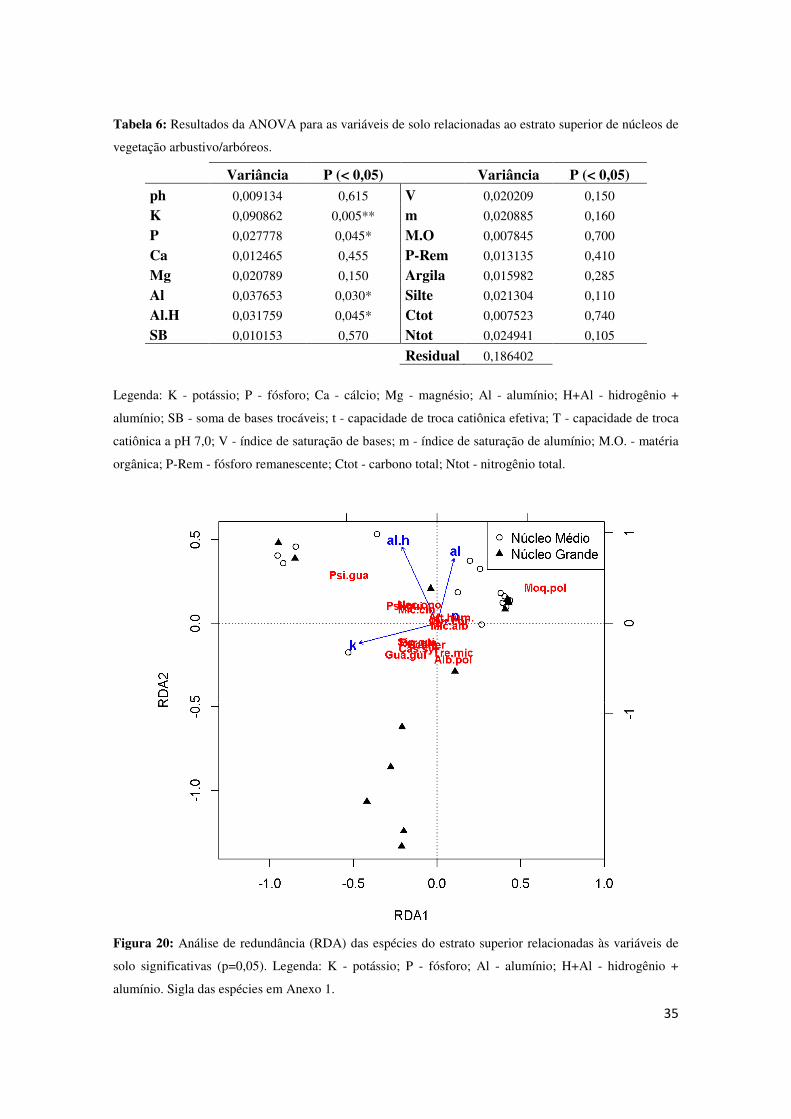
Já no estrato superior, as variáveis significativas foram potássio (K), fósforo (P),
alumínio (Al) e alumínio + hidrogênio (Al.H) (P < 0,05) (Tabela 6), utilizando então
somente estes fatores para a RDA final.
34
Tabela 5: Resultados da ANOVA para as variáveis de solo relacionadas ao estrato regenerante de núcleos
de vegetação arbustivo/arbóreos.
Variância P (< 0,5) Variância P (< 0,5) pH 0,044305 0,060
. V 0,046448 0,065
.
K 0,051115 0,025* m 0,021381 0,385
P 0,057241 0,015* M.O 0,029924 0,175
Ca 0,04176 0,090. P-Rem 0,031091 0,260
Mg 0,026147 0,305 Argila 0,025212 0,350
Al 0,028482 0,305 Silte 0,025287 0,310
Al.H 0,052321 0,055. Ctot 0,009583 0,890
SB 0,013498 0,750 Ntot 0,019045 0,600
Residual 0,265501
Legenda: K - potássio; P - fósforo; Ca - cálcio; Mg - magnésio; Al - alumínio; H+Al - hidrogênio +
alumínio; SB - soma de bases trocáveis; t - capacidade de troca catiônica efetiva; T - capacidade de troca
catiônica a pH 7,0; V - índice de saturação de bases; m - índice de saturação de alumínio; M.O. - matéria
orgânica; P-Rem - fósforo remanescente; Ctot - carbono total; Ntot - nitrogênio total.
Figura 19: Análise de redundância (RDA) das espécies do estrato regenerante relacionadas às variáveis
de solo significativas (p=0,05). Legenda: K - potássio; P - fósforo; H+Al - hidrogênio + alumínio; V-
índice de saturação de bases. Sigla das espécies em Anexo 1.
35
Tabela 6: Resultados da ANOVA para as variáveis de solo relacionadas ao estrato superior de núcleos de
vegetação arbustivo/arbóreos.
Variância P (< 0,05) Variância P (< 0,05) ph 0,009134 0,615 V 0,020209 0,150
K 0,090862 0,005** m 0,020885 0,160
P 0,027778 0,045* M.O 0,007845 0,700
Ca 0,012465 0,455 P-Rem 0,013135 0,410
Mg 0,020789 0,150 Argila 0,015982 0,285
Al 0,037653 0,030* Silte 0,021304 0,110
Al.H 0,031759 0,045* Ctot 0,007523 0,740
SB 0,010153 0,570 Ntot 0,024941 0,105
Residual 0,186402
Legenda: K - potássio; P - fósforo; Ca - cálcio; Mg - magnésio; Al - alumínio; H+Al - hidrogênio +
alumínio; SB - soma de bases trocáveis; t - capacidade de troca catiônica efetiva; T - capacidade de troca
catiônica a pH 7,0; V - índice de saturação de bases; m - índice de saturação de alumínio; M.O. - matéria
orgânica; P-Rem - fósforo remanescente; Ctot - carbono total; Ntot - nitrogênio total.
Figura 20: Análise de redundância (RDA) das espécies do estrato superior relacionadas às variáveis de
solo significativas (p=0,05). Legenda: K - potássio; P - fósforo; Al - alumínio; H+Al - hidrogênio +
alumínio. Sigla das espécies em Anexo 1.

36
No RDA do estrato regenerante, que apresentou 19% de proporção cumulativa
entre os dois eixos principais (13% no primeiro eixo e 6% no segundo), observa-se que
a espécie Siparuna guianensis está diretamente relacionada aos núcleos de vegetação
grandes, comprovando outras análises já realizadas. Estes núcleos estão relacionados ao
índice de saturação de bases (V), assim como os núcleos médios estão relacionados ao
fósforo (P) e a espécie Psidium guineensis.
Já a RDA do estrato superior (29% de proporção cumulativa entre os eixos
principais, 21% pertencente ao primeiro eixo e 8% ao segundo) mostra o agrupamento
de algumas espécies, sendo algumas Guarea guidonea, Albizia polycephala e Trema
micrantha, relacionadas a núcleos grandes e ao elemento potássio (K). Também se
observa o alumínio (Al), causador da acidez do solo, relacionado à núcleos médios, e
estes núcleos relacionados a espécie Moquiniastrum polymorphum.
4. DISCUSSÃO
Os núcleos de vegetação arbustivo/arbóreos partem de um ponto inicial, que
seriam os indivíduos isolados. A grande maioria desses indivíduos foram encontrados
no interior dos núcleos, mostrando que eles permanecem presentes em estágios mais
avançados da sucessão. Os indivíduos isolados trazem então benefícios à área (García-
Orth & Martínez-Ramos, 2011) principalmente pelo processo de facilitação que
favorece a formação de núcleos de vegetação arbustivo/arbóreos.
O início da sucessão se dá com a presença de espécies pioneiras, em especial
Moquiniastrum polymorphum (= Gochnatia polymorpha), que dominou as pastagens
tanto de forma isolada como em núcleos de vegetação. A importância de M.
polymorphum é principalmente na reabilitação de áreas. A espécie é altamente resistente
ao fogo, dominando áreas com recente histórico de queimada (Sánchez-Tapia, 2011).
Além disso, é encontrada em bordas e capoeiras da região, que coloniza e domina o
ambiente, criando condições para o estabelecimento de outras espécies que necessitam
de algum tipo de sombreamento para germinarem (Neves & Peixoto, 2008).
Entretanto, em pastagens, observou-se que esta espécie quando isolada ou
formando núcleos compostos pela mesma espécie, não apresentou associação com
indivíduos regenerantes, e sim com a presença de gramíneas. Isto infere que a espécie
não traz condições para o recrutamento de outros indivíduos. Isto sugere que M.

37
polymorphum de forma isolada não tem a capacidade de formar um núcleo, exceto em
condição de monodominância, principalmente por esta espécie apresentar uma
conformação de copa que não impede a entrada de luz e, consequentemente, nem a
presença dominante de gramíneas. M. polymorphum em áreas de pastagem é diferente
de Clusia hilariana em restinga (Garbin et al., 2012), que acarreta melhores condições
em áreas de extrema luminosidade e temperatura, facilitando o estabelecimento de
outros indivíduos.
Porém, quando Moquiniastrum polymorphum forma núcleos com outras
espécies, por exemplo Psidium guajava, que foi a segunda maior espécie encontrada em
núcleos, já é possível verificar o estabelecimento diversificado de plântulas. Segundo
Zahawi & Augspurger (1999), a riqueza de espécies, assim como o número de
indivíduos de plântulas, foi consideravelmente maior em pastos compostos por
cobertura de Psidium guajava do que em pastos abertos, mostrando que esta espécie se
comportou como facilitadora, formando ilhas que favoreceram a sucessão da pastagem.
Nos núcleos pequenos, observou-se a presença dominante de espécies pioneiras
(Figura 9). Isto mostra que, a partir deste ponto de co-existência de pioneiras, os núcleos
de vegetação arbustivo/arbóreos podem seguir evoluindo no processo de sucessão.
Entretanto, assim como os núcleos tendem a crescer, eles também podem se tornar
indivíduos isolados novamente, isto porque a riqueza de espécies encontradas isoladas
foi maior que nos núcleos pequenos (Figura 9).
Como as espécies encontradas exclusivamente nos núcleos de vegetação
arbustivo/arbóreos são pouco abundantes na área, mostra que estas são espécies que
provavelmente chegaram após a formação do núcleo, contribuindo com a riqueza. Com
o aumento da área dos núcleos também há o aumento de espécies não pioneiras e
zoocóricas, mostrando a importância de indivíduos atrativos de fauna que acarretam a
dispersão de espécies não pioneiras. Com o estabelecimento deste balanço, é menos
provável que haja o retorno dos núcleos para a forma isolada.
Isto fica claro ao observar que a espécie indicadora de núcleos pequenos é
pioneira, enquanto que as espécies indicadoras de núcleos grandes são não pioneiras
(Tabela 2). Pode-se inferir que a espécie Baccharis dracunculifolia atua no auxílio da
formação dos núcleos, enquanto que Siparuna guianensis e Nectandra opositifolia
chegam quando o núcleo já está desenvolvido, contribuindo para o desenvolvimento do
mesmo.

38
O desenvolvimento do núcleo se dá de uma forma em que, à medida que há o
crescimento dos mesmos, também há o aumento da riqueza. Isto infere que eles tendem
a crescer não somente pela dominância das próprias espécies que os compõem, mas
também pela chegada de novas espécies vindas de outra fonte de propágulo. É provável
que núcleos grandes atraem os dispersores de semente, podendo também criar uma
condição mais favorável no estabelecimento de plântulas (Zahawi & Augspurger, 2006).
Duarte et al. (2006) também constataram que grandes manchas florestadas há um maior
número de espécies exclusivas, ao contrário das pequenas manchas e das facilitadoras.
A área do núcleo está associada então com uma maior tendência sucessional,
tendo em vista que núcleos maiores tende a ter uma dinâmica mais acelerada que os
menores (Duarte et al., 2006). Esta dinâmica, impulsionada pela chegada de diferentes
propágulos, se deve principalmente à fauna dispersora. Este aumento também foi
constatado em outras áreas perturbadas (Duarte et al., 2010; Zahawi & Augspurger,
2006)
Com relação ao estrato regenerante, o presente estudo não constatou elevados
números de indivíduos de Moquiniastrum polymorphum, contrapondo-se ao estrato
superior. Ao avaliar a chuva de sementes na mesma área de estudo, Kolb (1993)
amostrou sementes dessa espécie como sendo as mais abundantes. Isto mostra que o
elevado número de propágulos produzido por esta espécie não é suficiente para manter
sua dominância no estrato regenerante. Já espécies como Cinnamomum triplinerve,
Nectandra oppositifolia, Guarea guidonea e Siparuna guianensis possuem mais
indivíduos neste estrato do que no estrato superior, onde são pouco encontradas. A
abundância de indivíduos não pioneiros na regeneração mostra que, além de haver
evolução da comunidade, infere que os núcleos oferecem subsídios para a germinação
de sementes e posterior estabelecimento das plântulas.
Assim, o padrão encontrado neste trabalho foi semelhante aos resultados de
Cheung et al. (2010), onde a sucessão em pastagem abandonada foi caracterizada pelo
aumento de árvores, de espécies zoocóricas e não pioneiras.
Os núcleos de vegetação arbustivo/arbóreos se mostraram importantes na
paisagem principalmente ao olhar a composição e abundância do estrato regenerante. A
riqueza e o número de regenerantes são superiores no interior do núcleo (Figura 15). Na
borda do mesmo ainda possui uma elevada riqueza, por ainda serem favorecidos pelas
suas características. Na área de matriz graminóide ainda próximo aos núcleos, a

39
abundância de regenerantes foi menor por esta área já não sofrer tanta influência dos
núcleos, e com o aumento dessa distância, a influência é praticamente nula (Figura 21).
Na matriz graminóide, além dos poucos indivíduos regenerantes encontrados, a espécie
mais abundante foi Miconia albicans. Esta se trata de uma espécie pioneira, devendo
obrigatoriamente ter características que as possibilitem estar em ambientes adversos,
como falta de sombreamento e alta competição com gramíneas.
A dificuldade de estabelecimento de indivíduos em meio às gramíneas também
foram observados por Holl (2002), que encontrou maior número de sementes de
espécies não pioneiras em manchas de arbustos em comparação à matriz graminóide, e
por Oliveira (2012), que levantou uma menor riqueza de espécies de plântulas em
campo antrópico (pastagem) em comparação à capoeiras e florestas.
Figura 21: Caracterização das influências positivas trazidas pelos núcleos de vegetação
arbustivo/arbóreos de acordo com seu distanciamento, em áreas de pastagem abandonada. Legenda: R -
presença de regeneração; G - presença de gramínea; P - presença de indivíduos pioneiros; NP - presença
de indivíduos não pioneiros.
Segundo Holl et al. (2000), os processos que influenciam o estabelecimento de
plântulas, limitando a cobertura em pastagem, são a chuva de sementes, germinação e
predação de sementes, e crescimento e sobrevivência de plântulas, e alguns fatores que
afetam a magnitude destes processos são estrutura da vegetação, cobertura de
gramíneas, microclima e stress hídrico, e parâmetros físicos e químicos do solo.
No presente trabalho, as influências trazidas pelos núcleos que mais favoreceram
a fase inicial de estabelecimento da regeneração natural foram o sombreamento e a
baixa cobertura de gramíneas. Observou-se que a abundância de regenerantes diminuía

40
à medida que havia um aumento da abertura do dossel e da cobertura de gramíneas, e
esses fatores foram menores no interior dos núcleos (Figura 16, Tabela 4).
Holl (2002) encontrou este padrão em seu estudo, com 99%, 78% e 49% de
cobertura de herbáceas em matriz graminóide, pequenos arbustos e grandes manchas de
arbustos, respectivamente. A temperatura, amenizada pelo sombreamento, também é
maior em locais abertos do que sob indivíduos isolados (García-Orth & Martínez-
Ramos, 2011), mostrando que este fator propicia melhores condições a partir do
estabelecimento de um primeiro indivíduo.
O efeito do sombreamento pode ser o fator chave para a sobrevivência de
plântulas em núcleos de vegetação, já em áreas de pastagem o microclima pode inibir a
sobrevivência destes indivíduos (Kolb, 1993). A competição com as gramíneas que
compõe a pastagem é o fator primário que impede a sobrevivência de plântulas (Holl et
al., 2000), acarretando desta forma a diminuição da biodiversidade (Martins et al.,
2004).
A cobertura vegetal promovida principalmente pelas herbáceas exóticas
influencia negativamente na regeneração natural, evidenciando o efeito limitante da
cobertura de gramíneas (Cheung et al., 2009; Denslow et al., 2006; Cabin et al., 2002).
Para Eliason & Allen (1997), as gramíneas podem, num primeiro momento, reduzir o
crescimento e a sobrevivência de mudas, entretanto, com o tempo o efeito das
gramíneas diminui, não sendo mais significativo. Isto pôde ser observado com a
presença de núcleos mais evoluídos e muitas vezes sem a presença de gramíneas. Logo,
é na fase inicial de formação dos núcleos que fatores físicos podem afetar mais no
estabelecimento da regeneração.
Características do solo também se mostraram diferentes com a presença dos
núcleos de vegetação arbustivo/arbóreos. As plantas podem se alterarem no ambiente
onde vivem, principalmente por conta das suas próprias características e pela interação
delas com o ambiente. Um exemplo disso é dado por Maharning et al. (2009), que
afirmaram que as espécies de plantas variam sua química e fisiologia, e assim variam a
qualidade e quantidade de serrapilheira produzida.
O interior dos núcleos apresentou solo com maior fertilidade e menor acidez
quando comparado à matriz graminóide (Apêndice 5). Jirová et al. (2012) também
encontraram resultado semelhande ao constatar que em solos sob arbustos de pastagens
o pH é maior (7,22) que em solos de bosque (6,59). Já ao comparar os resultados do

41
presente trabalho com os de Moraes et al.(2008), encontrou-se valores semelhantes de
pH tanto em pastagens abandonadas de várzea (5,37) como de morrote (5,00),
mostrando que este foi um fator pouco afetado. Já K e P obtiveram valores diferentes,
tanto para várzea (82,43 e 2,57 respectivamente) como para pastagens de morrote
(20,30 e 1,33 respectivamente). Estes nutrientes também apareceram de forma
diferenciada na matriz graminóide e nos núcleos de vegetação, influenciando de
diferentes formas os tipos de núcleos encontrados (Figuras 19 e 20).
Os dois estratos mostraram relação de conteúdo de P no solo com a abundância
de espécies pioneiras, e do K com espécies não pioneiras. O P é um elemento estável no
solo, e isto resulta numa baixa solubilidade que pode causar deficiência em sua
disponibilidade (Fassbender, 1980).
Já os valores de Al foram maiores no presente estudo (1,03 em núcleos de
vegetação e 0,98 em matriz graminóide) do que em pastagens de morrote da mesma
área (0,47) (Moraes et al., 2008). Entretanto, os autores encontraram valores mais altos
de Al em solos sob florestas, o que condiz com os resultados de maior teor de Al em
núcleos de vegetação. Estes resultados, de certa forma, não são positivos devido a
elevados fatores de Al gerar toxidez, que danificam as plantas.
Comparando áreas de pastagem com floresta, Holl (1999) encontrou valores
maiores de Ca, Mg, K, M.O. e Ntot na floresta, e níveis de P baixos em ambos os
habitats. No presente trabalho, todos os elementos (K, P, Ca, Mg) foram maiores no
interior dos núcleos de vegetação, podendo ser explicado por este ser um sistema mais
complexo, com maior diversidade florística, imobilizando mais os elementos que são
componentes estruturais dos vegetais, além da alta taxa de renovação de biomassa aérea
e de raízes (Moraes et al., 2008). Martins et al. (2004) também encontraram em área
dominada por capim-gordura elevada acidez, alta saturação de Al, baixa disponibilidade
de P, e baixos níveis de bases trocáveis.
No geral, poucas variáveis do solo foram influentes no processo de sucessão,
concordando com trabalho de Zanini et al. (2006), que afirma que a variação dos
nutrientes não tem um papel expressivo no estágio inicial de sucessão. Segundo Ganade
& Brown (2002), a luminosidade é o principal recurso para o início da sucessão,
enquanto que as taxas de elementos como P e K, podem contribuir com o crescimento
de plântulas quando a luz não for mais um recurso limitante.

42
Com o estudo foi possível observar a importância dos núcleos de vegetação
arbustivo/arbóreos e os fatores que mais influenciam na sucessão secundária de
pastagens abandonadas. Desta forma, avaliar a mudança na estrutura florestal dessas
áreas é importante para verificar a resiliência do sistema (Cheung et al., 2010), além de
entender os processos que ali se desenvolvem.
Com relação às pastagens da Reserva Biológica de Poço das Antas, pode-se
inferir que os núcleos são estruturas que trazem benefícios, principalmente ao comparar
fatores como dominância de gramíneas e sombreamento com a matriz graminóide, além
de observar um aumento na dinâmica sucessional e da diversidade de espécies com o
aumento da área dos núcleos. Entretanto, vendo de uma forma geral, os núcleos de
vegetação arbustivo/arbóreos se desenvolveram a ponto de ocupar 20% de uma área de
1, 08 ha, em 40 anos de abandono das pastagens. Isto mostra que o desenvolvimento
desses locais ocorre, mas de uma forma gradual, inferindo que há diversos fatores que
dificultam o processo de restauração passiva.
5. CONCLUSÃO
Os núcleos de vegetação arbustivo/arbóreo, que ocupam quase 20% das encostas
onde predomina matriz graminóide, se caracterizam quanto a diversidade e tamanho,
pela presença de espécies indicadoras de evolução florística e alteração na composição
da regeneração com o tempo.
A espécie Moquiniastrum polymorphum ocorre desde a forma isolada até os
núcleos grandes, mostrando resistir às condições ambientais inóspitas de ambientes
extremos, embora não apresente indícios de facilitação.
Os núcleos de vegetação acarretam melhores condições ao ecossistema, tanto a
partir das modificações no sombreamento, como em função das características químicas
do solo (capacidade de saturação).
A nucleação se mostrou eficiente com relação à avanços na dinâmica sucessional
em pastagens abandonadas. Entretanto, em 40 anos de abandono, 1,08 hectares de áreas
ocupadas por pastagem apresentaram cerca de 20% ocupados por núcleos de vegetação
arbustivo/arbóreos e 80% por matriz graminóide, inferindo que a restauração passiva de
pastagens abandonadas é um processo lento.

43
6. RECOMENDAÇÕES FINAIS
Como estratégia de manejo para promover a regeneração natural em áreas de
pastagem, sugere-se a utilização dos núcleos de vegetação para maior enriquecimento
da área. Com a aplicação da restauração passiva, que se trata de um manejo de baixo
custo, basta agir de forma que permita o processo de sucessão ecológica, podendo
cercar a área onde se encontra os núcleos, ou construindo-se aceiros para evitar a
ocorrência de fogo.
Se a opção for uma maior interferência ao restaurar, deve-se atuar de forma que
contribua para o desenvolvimento dos núcleos, com a aplicação de sementes de espécies
não pioneiras em seu interior, ou fazendo a remoção periódica de gramíneas de seu
entorno, para favorecer a expansão do seu diâmetro, e aplicando sementes de espécies
pioneiras de rápido crescimento nessas bordas, para assim aumentar a área de
sombreamento do núcleo.
Por fim, destaca-se que esta é uma técnica mais barata, porém lenta. Se o
objetivo do projeto for restaurar de forma mais rápida, recomenda-se a restauração com
plantio de mudas. Neste contexto, os núcleos de vegetação arbustivo/arbóreos também
podem ser empregados, observando-se se há presente na área a ser restaurada algumas
dessas estruturas, pois estes podem ser fonte de informações de como ocorre a sucessão
secundária, incluindo processos como facilitação e competição.

44
REFERÊNCIAS BIBLIOGRÁFICAS
Abreu, R.C.R.; Assis, G.B.; Frison, S.; Aguirre, A. & Durigan, G. 2011. Can native
vegetation recover after slash pine cultivation in the Brazilian Savanna? Forest
Ecology and Management 262(8): 1452-1459.
Aronson, J.; Durigan, G. & Brancalion, P.H.S. 2011. Conceitos e definições correlatos à
ciência e a prática da restauração ecológica. IF Série de Registros 44:1-38.
Barbosa, J.H.C. & Faria, S.M. 2006. Aporte de serrapilheira ao solo em estágios
sucessionais florestais na Reserva Biológica de Poço das Antas, Rio de Janeiro,
Brasil. Rodriguésia 57(3): 461-476.
Barboza, R.S. 2007. Caracterização das bacias aéreas e avaliação da chuva oculta nos
contrafortes da serra do Mar. Dissertação de Mestrado em Ciências Ambientais e
Florestais. Universidade Federal Rural do Rio de Janeiro, Seropédica. 56p.
Baylão Júnior, H.F. 2010. Espécies com vocação para facilitar processos de
restauração espontânea de ecossistemas perturbados na vertente atlântica da Serra
do Mar, Piraí – RJ. Dissertação de Mestrado em Ciências Ambientais e Florestais.
Universidade Federal Rural do Rio de Janeiro, Seropédica. 73p.
Bidegain, P. 2005. Plano das bacias hidrográficas da região dos Lagos e do rio São
João. Consórcio intermunicipal para gestão das bacias hidrográficas da região dos
Lagos, rio São João e Zona Costeira – CILSJ, Araruama. 153p.
______ & Völker, C.M. 2003. Bacias hidrográficas dos rios São João e das Ostras:
águas, terras e conservação ambiental. Consórcio Intermunicipal Lagos São João,
Rio de Janeiro. 177p.
Braz, D.M.; Santos, P.R.O.; Cysneiros, V.C. & Mendonça Júnior, J.O. 2012. Árvores do
Jardim Botânico da Universidade Federal Rural do Rio de Janeiro: chave para

45
identificação das famílias, dicas para reconhecimento das espécies em campo e
caracterização e uso da madeira. Ed. UFRRJ-EDUR, Seropédica. 300p.
Cabin, R.J.; Weller, S.G.; Lorence, D.H.; Cordell, S.; Hadway, L.J.; Montgomery, R.;
Goo, D. & Urakami, A. 2002. Effects of light, alien grass, and native species
additions on Hawaiian dry forest restoration. Ecological Applications 12(6): 1595-
1610.
Callaway, R.M. 1995. Positive Interactions among Plants. Botanical Review 61(4): 306-
349.
______ & Maron, J.L. 2006. What have exotic plant invasions taught us over the past
20 years? Trends in Ecology and Evolution 21(7): 369-374.
______ & Walker, L.R. 1997. Competition and Facilitation: A Synthetic Approach to
Interactions in Plant Communities. Ecology 78(7): 1958-1965.
Carvalho, F.A.; Nascimento, M.T.; Braga, J.M.A. & Rodrigues, P.J.F.P. 2006. Estrutura
da comunidade arbórea da Floresta Atlântica de Baixada periodicamente inundada
na Reserva Biológica de Poço das Antas, Rio de Janeiro, Brasil. Rodriguésia 57
(3): 503-518.
______; Nascimento, M.T. & Oliveira Filho, A.T. 2008. Composição, riqueza e
heterogeneidade da flora arbórea da bacia do rio São João, RJ, Brasil. Acta
Botanica Brasilica 22(4): 929-940.
Chapman, C.A. & Chapman, L.J. 1999. Forest restoration in abandoned agricultural
land: a case study from east Africa. Conservation Biology 13(6): 1301-1311.
Cheung, K.C.; Liebsch, D. & Marques, M.C.M. 2010. Forest recovery in newly
abandoned pastures in southern Brazil: implications for the Atlantic Rain Forest
resilience. Natureza e Conservação 8(1): 66-70.

46
______; Marques, M.C.M. & Liebsch, D. 2009. Relação entre a presença de vegetação
herbácea e a regeneração natural de espécies lenhosas em pastagens abandonadas
na Floresta Ombrófila Densa do Sul do Brasil. Acta Botanica Brasilica 23(4):
1048-1056.
Christo, A.G.; Guedes-Bruni, R.R. & Fonseca-Kruel, V.S. 2006. Uso de recursos
vegetais em comunidades rurais limítrofes à Reserva Biológica de Poço das Antas,
Silva Jardim, Rio de Janeiro: estudo de caso na gleba Aldeia Velha. Rodriguésia 57
(3): 519-542.
Connell, J.H. & Slatyer, R.O. 1977. Mechanisms of succession in natural communities
and their role in community stability and organization. The American Naturalist
111 (982): 1119-1144.
Consórcio Intermunicipal Lagos São João. Acessado em: junho de 2013.
http://www.lagossaojoao.org.br/
Cortines, E.; Pereira, A.L.; Santos, P.R.O.; Santos, G.L. & Valcarcel, R. 2001.
Vegetação arbórea em vertentes com orientação norte e sul na Floresta Montana,
Nova Friburgo-RJ. Floresta e Ambiente 18(4): 428-437.
Cunha, S.B. 1995. Impactos das obras de engenharia sobre o ambiente biofísico da
Bacia do rio São João (Rio de Janeiro – Brasil). Dissertação de Doutorado em
Geografia Física. Faculdade de Letras da Universidade de Lisboa, Lisboa. 415p.
______ & Freitas, M.W.D. 2004. Geossistemas e gestão ambiental na bacia hidrográfica
do rio São João – RJ. GEOgraphia 6(12): 87-110.
Denslow, J.S.; Uowolo, A.L. & Hughes, R.F. 2006. Limitations to seedling
establishment in a mesic Hawaiian forest. Oecologia 148(1): 118-128.

47
DeSimone, S.A. 2011. Balancing active and passive restoration in a nonchemical,
research-based approach to coastal sage scrub restoration in Southern California.
Ecological Restoration 29(1-2): 45-51.
Duarte, L.S.; Hofmann, G.S.; Santos, M.M.G.; Hartz, S.M. & Pillar, V.S. 2010. Testing
for the influence of niche and neutral factors on sapling community assembly
beneath isolated woody plants in grasslands. Journal of Vegetation Science 21(3):
462-471.
______; Machado, R.E.; Hartz, S.M. & Pillar, V.S. 2006. What saplings can tell us
about forest expansion over natural grasslands. Journal of Vegetation Science
17(6): 799-808.
Egerton, J.J.G.; Banks, J.C.G.; Gibson, A.; Cunningham, R.B. & Ball, M.C. 2000.
Facilitation of seedling establishment: reduction in irradiance enhances winter
growth of Eucalyptus pauciflora. Ecology 81(5): 1437-1449
Eliason, S.A. & Allen, E.B. 1997. Exotic Grass Competition in Suppressing Native
Shrubland Re-establishment. Restoration Ecology 5(3): 245-255.
Embrapa Solos. 2012. Método para coleta de amostras de solos para análise.
Disponível em: www.cnps.embrapa.br/servicos/metodo_coleta.html. Acessado em
15 de abril de 2012.
Fassbender, H.W. 1980. Quimica de suelos con énfasis en suelos de América Latina.
IICA, San José. 398p.
Fundação CEPERJ 2013. Acessado em: julho de 2013.
http://www.ceperj.rj.gov.br/ceep/info_territorios/divis_regional.html
Ganade, G. & Brown, V.K. 2002. Succession in old pastures of Central Amazonia: role
of soil fertility and plant litter. Ecology 83(3): 743-754.

48
Garbin, M.L.; Carrijo, T.T.; Sansevero, J.B.B.; Sánchez-Tapia, A. & Scarano, F.R.
2012. Subordinate, not dominant, woody species promote the diversity of climbing
plants. Perspectives in Plant Ecology, Evolution and Systematics 14(4): 257-265.
García-Orth, X. & Martínez-Ramos, M.M. 2011. Isolated trees and grass removal
improve performance of transplanted Trema micrantha (L.) Blume (Ulmaceae)
saplings in tropical pastures. Restoration Ecology 19(1): 24-34.
Guedes-Bruni, R.R.; Neto, S.J.S.; Morim, M.P. & Mantovani, W. 2006. Composição
florística e estrutura de trecho de Floresta Ombrófila Densa Atlântica Aluvial na
Reserva Biológica de Poço das Antas, Silva Jardim, Rio de Janeiro, Brasil.
Rodriguésia 57(3): 413-428.
Hobbs, R. J. 2009. Woodland restoration in Scotland: Ecology, history, culture,
economics, politics and change. Journal of Environmental Management 90(9):
2857-2865.
______; Higgs, E. & Harris, J.A. 2009. Novel ecosystems: implications for conservation
and restoration. Trends in Ecology and Evolution 24(11): 599-605.
Holl, K. D. 2013. Restoring Tropical Forest. Nature Education Knowledge 4(4): 1-4.
______. 2002. Effect of shrubs on tree seedling establishment in an abandoned tropical
pasture. Journal of Ecology 90(1): 179-187.
______. 1999. Factors Limiting Tropical Rain Forest Regeneration in Abandoned
Pasture: Seed Rain, Seed Germination, Microclimate, and Soil. Biotropica 31(2):
229-242.
______ & Aide, T.M. 2011. When and where to actively restore ecosystems? Forest
Ecology and Management 261(10): 1558-1563.

49
______; Loik, M.E.; Lin, E.H.V. & Samuels, I.A. 2000. Tropical montane forest
restoration in Costa Rica: overcoming barriers to dispersal and establishment.
Restoration Ecology 8(4): 339-349.
______ & Lulow, M.E. 1997. Effects of species, habitat, and distance from edge on
post-dispersal seed predation in a tropical rainforest. Biotropica 29(4): 459-468.
Jirová, A.; Klaudisová, A. & Prach, K. 2012. Spontaneous restoration of target
vegetation in old-fields in a central european landscape: a repeated analysis after
three decades. Applied Vegetation Science 15(2) 245-252.
Kolb, S.R. 1993. Islands of secondary vegetation in degraded pastures of Brazil: their
role in reestablishing Atlantic Coastal Forest. Dissertation submitted to the
Graduate, University of Georgia, 128p.
Lima, H.C.; Pessoa, S.V.A.; Guedes-Bruni, R.R.; Moraes, L.F.D.; Granzotto, S.V.;
Iwamoto, S. & Di Ciero, J. 2006. Caracterização fisionômico-florística e
mapeamento da vegetação da Reserva Biológica de Poço das Antas, Silva Jardim,
Rio de Janeiro, Brasil. Rodriguésia 57(3): 369-389.
Lorenzi, H. 2008. Árvores Brasileiras: Manual de Identificação de Identificação e
Cultivo de Plantas Arbóreas Nativas do Brasil, vol. 1. Instituto Plantarum, Nova
Odessa. 384p.
______. 2009. Árvores Brasileiras: Manual de Identificação de Identificação e Cultivo
de Plantas Arbóreas Nativas do Brasil, vol. 3. Instituto Plantarum, Nova Odessa.
384p.
______; Noblick, L.; Kahn, F. & Ferreira, E. 2010. Flora Brasileira Lorenzi: Arecaceae
(palmeiras). Instituto Plantarum, Nova Odessa. 384p.

50
Maharning, A.R.; Mills, A.A.S. & Adl, S.M. 2009. Soil community changes during
secondary succession to naturalized grasslands. Applied Soil Ecology 41(2): 137-
147.
Martins, C.R.; Leite, L.L. & Haridasan, M. 2004. Capim-gordura (Melinis minutiflora
P. Beauv.), uma gramínea exótica que compromete a recuperação de áreas
degradadas em Unidades de Conservação. Revista Árvore 28(5): 739-747.
Maza-Villalobos, S.; Balvanera, P. & Martínez-Ramos, M. 2011. Early regeneration of
tropical dry forest from abandoned pastures: contrasting chronosequence and
dynamic approaches. Biotropica 43(6): 666-675.
McIntire, E.J.B. & Fajardo, A. 2014. Facilitation as a ubiquitous driver of biodiversity.
New Phytologist 201(2): 403-416.
Meli, P.; Martínez-Ramos, M. & Rey-Benayas, J.M. 2013. Selecting Species for Passive
and Active Riparian Restoration in Southern Mexico. Restoration Ecology
21(2):163–165.
Metzger, J.P. 2001. O que é ecologia de paisagem? Biota Neotropica 1(1/2): 1-9.
Miranda, C.C. 2011. Funcionalidade de núcleos de Clidemia urceolata DC. na
recuperação de ambientes perturbados da Mata Atlântica. Tese de Doutorado em
Ciências Ambientais e Florestais, Universidade Federal Rural do Rio de Janeiro,
Seropédica. 154p.
Moraes, L.F.D.; Assumpção, J.M.; Luchiari, C. & Pereira, T.S. 2006. Plantio de
espécies arbóreas nativas para a restauração ecológica na Reserva Biológica de
Poço das Antas, Rio de Janeiro, Brasil. Rodriguésia 57(3): 477-489.
______; Campello, E.F.C.; Pereira, M.G. & Loss, A. 2008. Características do solo na
restauração de áreas degradadas na Reserva Biológica de Poço das Antas, RJ.
Ciência Florestal 18(2): 193-206.

51
Moro, M. F. & Martins, F. R. 2011. Métodos de levantamento do componente arbóreo –
arbustivo. pp. 174-212. In: Felfili, J.M.; Eisenlohr, P.V.; Melo, M.M.R.F.;
Andrade, L.A. & Neto, J.A.A.M. Fitossociologia no Brasil: métodos e estudo de
casos. v.1. Ed. UVF, Viçosa.
Morrinson, E.B. & Lindell, C.A. 2010. Active or passive Forest restoration? Assessing
restoration alternatives with avian foraging behavior. Restoration Ecology 19(201):
170-177.
Munhoz, C.B.R. & Araújo, G.M. 2011. Métodos de amostragem do estrato herbáceo-
subarbustivo. pp. 213-230. In: Felfili, J.M.; Eisenlohr, P.V.; Melo, M.M.R.F.;
Andrade, L.A. & Neto, J.A.A.M. Fitossociologia no Brasil: métodos e estudo de
casos. v.1. Ed. UVF, Viçosa.
Myers, N.; Mittermeier, R. A.; Mittermeier, C. G.; Fonseca, G. A. B. & Kent, J. 2000.
Biodiversity hotspots for conservation priorities. Nature 403(6772): 853-858.
Nappo, M.E.; Griffith, J.J.; Martins, S.V.; Marco Júnior, P.; Souza, A.L. & Oliveira
Filho, A.T. 2004. Dinâmica da estrutura fitossociológica da regeneração natural em
sub-bosque de Mimosa scabrella Bentham em área minerada, em Poço de Caldas,
MG. Revista Árvore 28(6): 811-829.
Neves, G.M.S. & Peixoto, A.L. 2008. Florística e estrutura da comunidade arbustivo-
arbórea de dois remanescentes em regeneração de Floresta Atlântica secundária na
Reserva Biológica de Poço das Antas, Silva Jardim, Rio de Janeiro. Pesquisas,
Botânica 59: 71-112.
Nóbrega, A.M.F.; Valeri, S.V.; De Paula, R.C. & Silva, S.A. 2008. Regeneração natural
em remanescentes florestais e áreas reflorestadas da várzea do rio Mogi-Guaçu,
Luiz Antônio – SP. Revista Árvore 32(5): 909-920.

52
Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.;
Simpson, G.L.; Solymos, P.; Stevens, M.H.H. & Wagner, H. 2013. vegan:
Community Ecology Package. R package version 2.0-9. http://CRAN.R-
project.org/package=vegan.
Oliveira, F.L. & Mello, E.F. 2007. A mineração de areia e os impactos ambientais na
bacia do rio São João, RJ. Revista Brasileira de Geociências 37(2): 374-389.
Oliveira, G.M.A. 2012. Comunidades de plântulas em campos antropizados e em
plantios para restauração ecológica: estrutura, composição e fatores ambientais.
Dissertação de Mestrado em Botânica, Escola Nacional de Botânica Tropical,
Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, Rio de Janeiro. 94p.
Padilla, F.M. & Pugnaire, F.I. 2006. The role of nurse plants in the restoration of
degraded environments. Frontiers in Ecology and the Environment 4(4): 196-202.
Pereira, T.S.; Costa, M.L.M.N.; Moraes, L.F.D. & Luchiari, C. 2008. Fenologia de
espécies arbóreas em Floresta Atlântica da Reserva Biológica de Poço das Antas,
Rio de Janeiro, Brasil. Iheringia 63(2): 329-339.
Pessoa, S.V.A. & Oliveira, R.R. 2006. Análise estrutural da vegetação arbórea em três
fragmentos florestais na Reserva Biológica de Poço das Antas, Rio de Janeiro,
Brasil. Rodriguésia 57(3): 391-411.
Prach, K.; Jongepierová, I. & Rehounková, K. 2013. Large-scale restoration of dry
grasslands on ex-arable land using a regional seed mixture: establishment of target
species. Restoration Ecology 21(1): 33-39.
Pueyo, Y.; Alados, C.L.; García-Ávila, B.; Kéfi, S.; Maestro, M. & Rietkerk, M. 2009.
Comparing direct abiotic amelioration and facilitation as tools for restoration of
semiarid grasslands. Restoration Ecology 17(6): 908-916.

53
R Development Core Team. 2013. R: A Language and Environment for Statistical
Computing. R Foundation for Statistical Computing, Vienna, Austria. Available
from: http://www.r-project.org/.
Reis, A.; Bechara, F. C.; Espindola, M. B.; Vieira, N. K. & Souza, L.L. 2003a.
Restauração de áreas degradadas: a nucleação como base para incrementar os
processos sucessionais. Natureza & Conservação 1(1): 28-36.
______; Rogalski, J.; Berkenbrock, I.S. & Bourscheid, K. 2003b. A nucleação aplicada
à restauração ambiental. In: Anais Seminário Nacional degradação e recuperação
ambiental, Foz do Iguacú.
http://www.sobrade.com.br/eventos/2003/seminario/index.htm. Acessado em abril
de 2012.
Reis, R.B. 2008. Mapeamento das áreas de preservação permanentes (APP) como
subsídio à elaboração de estratégias de Conservação e recuperação – estudo de
caso na APA do Rio São João/ Mico-Leão-Dourado (RJ). Dissertação de Mestrado
em Geografia, Universidade Federal do Rio de Janeiro, Rio de Janeiro. 120p.
______; Cardoso, P.V.; Cruz, C.B.M. & Vicens, R.S. 2009. Classificação do Uso e
Cobertura do Solo da APA do São João em uma abordagem orientada a objeto. pp.
7087-7094 In: Anais XIV Simpósio Brasileiro de Sensoriamento Remoto, INPE,
Natal.
Ribeiro, G.H.P.M. & Felfili, J.M. 2009. Regeneração natural em diferentes ambientes
da mata de galeria do Capetinga, na Fazenda Água Limpa – DF. Cerne 15(1): 1-9.
Ribeiro, M.C.; Metzger, J.P.; Martensen, A.C.; Ponzoni, F.J. & Hirota, M.M. 2009. The
Brazilian Atlantic Forest: How much is left, and how is the remaining forest
distributed? Implications for conservation. Biological Conservation 142 (6): 1141–
1153.

54
Ricklefs, R.E. 1996. A economia da natureza: um livro-texto em ecologia básica. 3ed.
Guanabara Koogan, Rio de Janeiro. 470p.
Roberts, D.W. 2013. labdsv: Ordination and Multivariate Analysis for Ecology. R
package version 1.6-1. http://CRAN.R-project.org/package=labdsv.
Rodrigues, E. 2013. Ecologia da Restauração. Editora Planta, Londrina, 300p.
Rodrigues R.R.; Brancalion P.H.S. & Isernhagen I. 2009. Pacto pela restauração da
Mata Atlântica: referencial dos conceitos e ações de restauração florestal. Instituto
BioAtlântica, São Paulo. 256p.
Sánchez-Tapia, A. 2011. Regeneração natural e restauração ecológica em capoeiras
antropizadas de Mata Atlântica submetidas a queimadas. Dissertação de Mestrado
em Ecologia, Universidade Federal do Rio de Janeiro, Rio de Janeiro, 95p.
Sansevero, J.B.B. 2008. Processos de regeneração em Mata Atlântica: uma
comparação entre áreas naturais e plantios de restauração ecológica na Reserva
Biológica Poço das Antas, Rio de Janeiro. Dissertação de Mestrado em Botânica,
Escola Nacional de Botânica Tropical, Instituto de Pesquisas Jardim Botânico do
Rio de Janeiro, Rio de Janeiro. 136p.
______; Prieto, P.V.; Moraes, L.F.D. & Rodrigues, P.J.F.P. 2011. Natural Regeneration
in Plantations of Native Trees in Lowland Brazilian Atlantic Forest: Community
Structure, Diversity, and Dispersal Syndromes. Restoration Ecology 19(3): 379-
389.
Santos, S.O. & Alves, M. 2012. Flora da usina São José, Igarassu, Pernambuco:
Lauraceae. Rodriguésia 63(3): 689-703.
Santos, V.E.; Souza, A.F. & Vieira, M.L. 2007. Efeito do Pastejo na Estrutura da
Vegetação de uma Floresta Estacional Ripícola. Revista Brasileira de Biociências
5(1): 171-173.

55
Scarano, F.R. 2006. Plant community structure and function in a swamp forest within
the atlantic rain forest complex: a synthesis. Rodriguésia 57(3): 491-502.
Scowcroft, P.G. & Yeh, J.T. 2013. Passive restoration augments active restoration in
deforested landscapes: The role of root suckering adjacent to planted stands of
Acacia koa. Forest Ecology and Management 305: 138-145.
Seabra , V.S. & Cruz, C.B.M. 2013. Mapeamento da dinâmica da cobertura e uso da
terra na bacia hidrográfica do rio São João, RJ. Sociedade & Natureza 25(2): 411-
426.
SER. 2004. Princípios da SER International sobre a restauração ecológica. Grupo de
Trabalho sobre Ciência e Política - versão 2. Society for Ecological Restoration
International, www.ser.org y Tucson.
Shapiro, S.S. & Wilk, M.B. 1965. An analysis of variance test for normality (complete
samples). Biometrika 52:591-611.
Shono, K.; Cadaweng, E.A. & Durst, P.B. 2007. Application of Assisted Natural
Regeneration to Restore Degraded Tropical Forestlands. Restoration Ecology
15(4): 620-626.
Silva, A.F.; Oliveira, R.V.; Santos, N.R.L. & de Paula, A. 2003. Composição florística e
grupos ecológicos das espécies de um trecho de floresta semidecídua submontana
da fazenda São Geraldo, Viçosa – MG. Revista Árvore 27(3): 311-319.
Souza, R.F.S. 2013. Avaliação da viabilidade de espécies arbóreas nativas da Mata
Atlântica utilizadas em restauração florestal no Sul da Bahia. Trabalho Final de
Mestrado Profissional. Escola Superior de Conservação Ambiental e
Sustentabilidade, Instituto de Pesquisas Ecológicas, Uruçuca. 79p.

56
Smith, R.L. 1996. Ecology and Field Biology. 5ed. Harper Collins College Publishers,
New York. 740p.
SNUC. 2000. Sistema Nacional de Unidades de Conservação. Lei nº 9.985, 18 de julho
de 2000.
Stachowicz, J.J. 2001. Mutualism, facilitation and the structure of ecological
communities. BioScience 51(3): 235-246.
Valentini, C.M.A.; Rodriguez-Ortiz, C.E. & Coelho, M.F.B. 2010. Siparuna guianensis
Aublet (negramina): uma revisão. Revista Brasileira de Plantas Medicinais 12(1):
96-104.
Vieira, C.M. & Pessoa, S.V.A. 2001. Estrutura e composição florística do estrato
herbáceo-subarbustivo de um pasto abandonado na Reserva Biológica de Poço das
Antas, município de Silva Jardim, RJ. Rodriguésia 52(80): 17-30.
Westman, W.E. 1978. Measuring the Inertia and Resilience of Ecosystems. BioScience
28(11): 705-710.
Yarranton, G.A. & Morrison, R.G. 1974. Spatial dynamics of a primary succession:
nucleation. Journal of Ecology 62(2): 417-428.
Zahawi, R.A. & Augspurger, C.K. 2006. Tropical forest restoration: tree islands as
recruitment foci in degraded lands of Honduras. Ecological application 16(2): 464-
478.
______ & Augspurger, C.K. 1999. Early plant succession in abandoned pastures in
Ecuador. Biotropica 31(4): 540-552.
Zanini, L.; Ganade, G. & Hübel, I. 2006. Facilitation and competition influence
succession in a subtropical old field. Plant Ecology 185(2): 179-190.
57
APÊNDICES
Apêndice 1: Parâmetros fitossociológicos das espécies arbustivo/arbóreas amostradas
isoladas em pastagens abandonadas, Reserva Biológica de Poço das Antas, Silva Jardim
– RJ. Legenda: Ni = número de indivíduos; Ab = área basal (m²); DR = densidade
relativa (%); FR = frequência relativa (%); DoR = dominância relativa (%); VI = valor
de importância (%).
Espécies Ni Ab DR FR DoR VI
1 Moquiniastrum polymorphum 72 0,3155 43,11 27,27 51,37 40,59
2 Psidium guineense 35 0,0342 20,96 22,73 5,57 16,42
3 Albizia polycephala 17 0,1930 10,18 4,55 31,42 15,38
4 Baccharis dracunculifolia 15 0,0204 8,98 9,09 3,33 7,13
5 Psidium guajava 17 0,0225 10,18 4,55 3,66 6,13
6 Cecropia pachystachya 2 0,0123 1,20 4,55 2,00 2,58
7 Myrcia sp. 1 0,0095 0,60 4,55 1,55 2,23
8 Tabernaemontana laeta 3 0,0013 1,80 4,55 0,21 2,18
9 Nectandra opositifolia 2 0,0023 1,20 4,55 0,38 2,04
10 Sparattosperma leucanthum 1 0,0022 0,60 4,55 0,36 1,83
11 Cybistax antisyphilitica 1 0,0005 0,60 4,55 0,09 1,74
12 Pera glabrata 1 0,0005 0,60 4,55 0,08 1,74

58
Apêndice 2: Parâmetros fitossociológicos das espécies arbustivo/arbóreas amostradas
formando núcleos de vegetação em pastagens abandonadas, Reserva Biológica de Poço
das Antas, Silva Jardim – RJ. Legenda: Ni = número de indivíduos; Ab = área basal
(m²); DR = densidade relativa (%); FR = frequência relativa (%); DoR = dominância
relativa (%); VI = valor de importância (%).
Espécies Ni Ab DR FR DoR VI
1 Moquiniastrum polymorphum 133 1,2646 45,39 9,09 61,27 38,59
2 Psidium guajava 31 0,0732 10,58 5,45 3,55 6,53
3 Psidium guineense 14 0,0192 4,78 9,09 0,93 4,93
4 Nectandra opositifolia 11 0,0915 3,75 5,45 4,43 4,55
5 Albizia polycephala 9 0,1687 3,07 1,82 8,17 4,35
6 Siparuna guianensis 9 0,0120 3,07 7,27 0,58 3,64
7 Guarea guidonia 8 0,0887 2,73 3,64 4,30 3,55
8 Miconia cinnamomifolia 7 0,0195 2,39 5,45 0,95 2,93
9 Myrsine coriacea 4 0,0163 1,37 5,45 0,79 2,54
10 Baccharis dracunculifolia 8 0,0139 2,73 3,64 0,67 2,35
11 Miconia albicans 6 0,0167 2,05 3,64 0,81 2,16
12 Schinus terebinthifolius 9 0,0330 3,07 1,82 1,60 2,16
13 Inga edulis 2 0,0784 0,68 1,82 3,80 2,10
14 Trema micrantha 7 0,0262 2,39 1,82 1,27 1,83
15 Myrcia splendens 2 0,0101 0,68 3,64 0,49 1,60
16 Pera glabrata 6 0,0146 2,05 1,82 0,71 1,52
17 Cinnamomum triplinerve 2 0,0025 0,68 3,64 0,12 1,48
18 Cupania racemosa 3 0,0319 1,02 1,82 1,55 1,46
19 Casearia sylvestris 4 0,0130 1,37 1,82 0,63 1,27
20 Cecropia pachystachya 2 0,0182 0,68 1,82 0,88 1,13
21 Sparattosperma leucanthum 1 0,0242 0,34 1,82 1,17 1,11
22 Tabernaemontana laeta 3 0,0019 1,02 1,82 0,09 0,98
23 Xylopia sericea 2 0,0039 0,68 1,82 0,19 0,90
24 Styrax glabratum 2 0,0036 0,68 1,82 0,17 0,89
25 Tapirira guianensis 1 0,0101 0,34 1,82 0,49 0,88
26 Handroanthus chrysotrichus 2 0,0027 0,68 1,82 0,13 0,88
27 Solanum inaequilaterum 1 0,0022 0,34 1,82 0,11 0,76
28 Zanthoxylum rhoifolium 1 0,0010 0,34 1,82 0,05 0,74
29 Tocoyena bullata 1 0,0009 0,34 1,82 0,04 0,73
30 Dalbergia nigra 1 0,0009 0,34 1,82 0,04 0,73
31 Cybistax antisyphilitica 1 0,0004 0,34 1,82 0,02 0,73
59
Apêndice 3: Parâmetros fitossociológicos das espécies regenerantes amostradas no
interior de núcleos de vegetação, Reserva Biológica de Poço das Antas, Silva Jardim –
RJ. Legenda: Ni = número de indivíduos; DR = densidade relativa (%); FR = frequência
relativa (%); VIRN = valor de importância da regeneração natural.
Espécies Ni DR FA FR VIRN
1 Siparuna guianensis 16 19,05 6,25 11,76 30,81
2 Cinnamomum triplinerve 14 16,67 3,13 5,88 22,55
3 Guarea guidonia 10 11,90 3,13 5,88 17,79
4 Miconia albicans 5 5,95 6,25 11,76 17,72
5 Nectandra opositifolia 9 10,71 3,13 5,88 16,60
6 Miconia cinnamomifolia 4 4,76 3,13 5,88 10,64
7 Myrsine coriacea 4 4,76 3,13 5,88 10,64
8 Psidium guineense 4 4,76 3,13 5,88 10,64
9 Xylopia sericea 3 3,57 3,13 5,88 9,45
10 Moquiniastrum polymorphum 2 2,38 3,13 5,88 8,26
11 Myrcia splendens 2 2,38 3,13 5,88 8,26
12 Psidium guajava 2 2,38 3,13 5,88 8,26
13 Albizia polycephala 3 3,57 1,56 2,94 6,51
14 Clidemia hirta 2 2,38 1,56 2,94 5,32
15 Alchornea triplinervia 1 1,19 1,56 2,94 4,13
16 Dalbergia nigra 1 1,19 1,56 2,94 4,13
17 Myrcia sp. 1 1,19 1,56 2,94 4,13
18 Tapirira guianensis 1 1,19 1,56 2,94 4,13
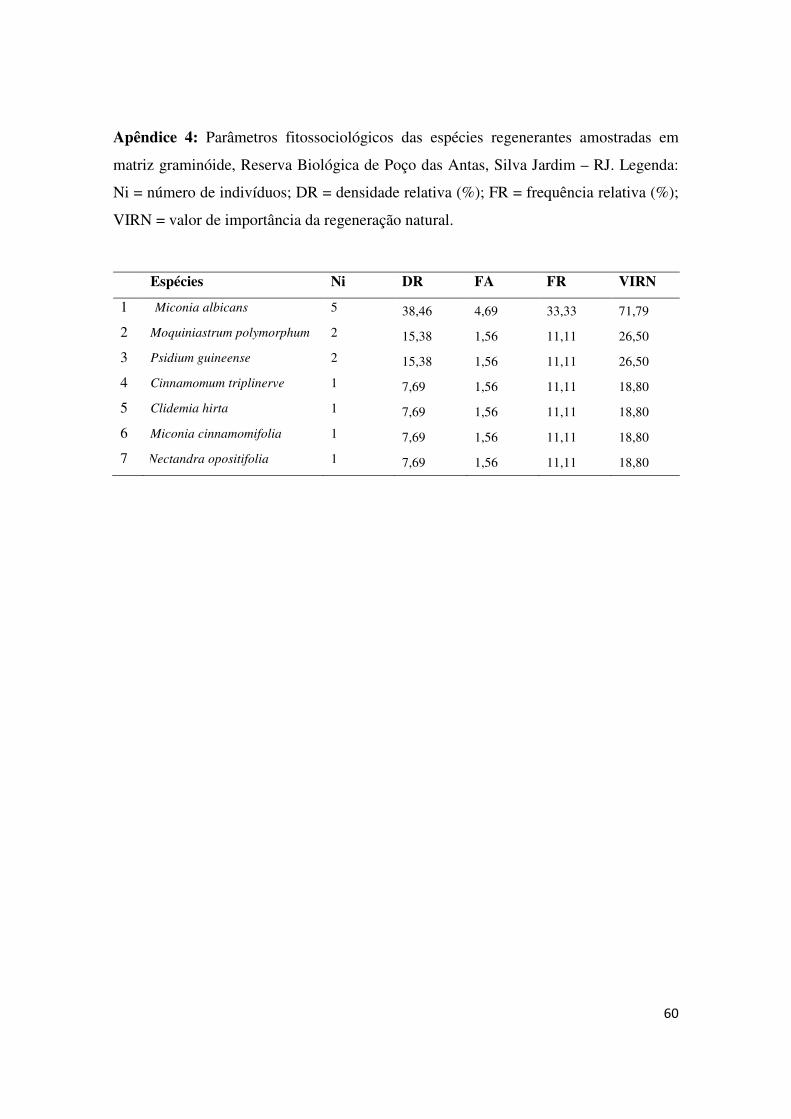
60
Apêndice 4: Parâmetros fitossociológicos das espécies regenerantes amostradas em
matriz graminóide, Reserva Biológica de Poço das Antas, Silva Jardim – RJ. Legenda:
Ni = número de indivíduos; DR = densidade relativa (%); FR = frequência relativa (%);
VIRN = valor de importância da regeneração natural.
Espécies Ni DR FA FR VIRN
1 Miconia albicans
5 38,46 4,69 33,33 71,79
2 Moquiniastrum polymorphum 2 15,38 1,56 11,11 26,50
3 Psidium guineense 2 15,38 1,56 11,11 26,50
4 Cinnamomum triplinerve 1 7,69 1,56 11,11 18,80
5 Clidemia hirta 1 7,69 1,56 11,11 18,80
6 Miconia cinnamomifolia 1 7,69 1,56 11,11 18,80
7 Nectandra opositifolia
1 7,69 1,56 11,11 18,80

61
Apêndice 5: Média dos resultados da análise de solo para as amostras do interior dos núcleos de vegetação e da matriz graminóide de pastagens
abandonadas, Reserva Biológica de Poço das Antas, Silva Jardim – RJ.
pH K P Ca Mg Al H+Al SB t T V m M.O. P-Rem Argila Silte Areia Ctot Ntot
----- mg/dm³ ----- ----- cmol/dm³ ----- ----- cmolc/dm³ ----- ----- % ----- dag/Kg mg/L ----- dag/Kg ----- g/Kg
Núcleo de vegetação
5,0 37,25 0,85 0,68 0,52 1,03 7,43 1,29 2,32 8,71 15,72 46,22 3,13 20,53 38 12 50 1,8 3,3
Matriz graminóide
5,0 30,88 0,77 0,53 0,39 0,98 6,76 1,00 1,98 7,76 13,78 50,54 3,01 20,13 38 11 52 1,7 3,1
Legenda: K - potássio; P - fósforo; Ca - cálcio; Mg - magnésio; Al - alumínio; H+Al - hidrogênio + alumínio; SB - soma de bases trocáveis; t - capacidade de troca catiônica
efetiva; T - capacidade de troca catiônica a pH 7,0; V - índice de saturação de bases; m - índice de saturação de alumínio; M.O. - matéria orgânica; P-Rem - fósforo
remanescente; Ctot - carbono total; Ntot - nitrogênio total.

62
ANEXOS
Anexo 1: Sigla das espécies ocorrentes na área.
Alb pol: Albizia polycephala
Alc tri: Alchornea triplinervia
Att hum : Attalea humilis
Bac dra: Baccharis dracunculifolia
Cas syl: Casearia sylvestris
Cec pac: Cecropia pachystachya
Cin tri: Cinnamomum triplinerve
Cli hir: Clidemia hirta
Cyb ant: Cybistax antisyphilitica
Dal nig: Dalbergia nigra
Gua gui: Guarea Guidonia
Han chr: Handroanthus chrysotrichus
Mic alb: Miconia albicans
Mic cin: Miconia cinnamomifolia
Moq pol: Moquiniastrum polymorphum
Myr sp.: Myrcia sp.
Myr cor: Myrsine coriacea
Myr spl: Myrcia splendens
Nec opo: Nectandra opositifolia
Per gla: Pera glabrata
Psi gua: Psidium guajava
Psi gui : Psidium guineense
Sch ter: Schinus terebinthifolius
Sip gui: Siparuna guianensis
Sol ina: Solanum inaequilaterum
Spa leu: Sparattosperma leucanthum
Sty gla: Styrax glabratum
Tab lae: Tabernaemontana laeta
Tap gui: Tapirira guianensis

63
Tre mic: Trema micrantha
Xyl ser: Xylopia sericea
Zan rho: Zanthoxylum rhoifolium.
Top Related