







Languages
Pages
Legal


UNIVERSITE CHEIKH ANTA DIOP
FACULTE DES SCIENCES ET TECHNIQUES
Département de Biologie Animale
Année : 2011 numéro : 103
Mémoire de Diplôme de Master II en Biologie Animale
Spécialité : PARASITOLOGIE
Présenté et soutenu en Février 2012
Par
Masse SAMBOU
Né le 15 décembre 1985 à Cadjinolle Kagnao (Sénégal)
JURY
Président : Professeur Bhen Sikina TOGUEBAYE (FST/UCAD)
Membres : Docteur Franselyse CLOTILDE BA (FST/UCAD)
Docteur Cheikhna DIEBAKATE (FST/UCAD)
Professeur Ngor FAYE (FST/UCAD)
Docteur Oleg MEDIANNIKOV (IRD/Marseille)
IDENTIFICATION DE RICKETTSIES PATHOGENES
CHEZ LES TIQUES IXODIDES DE LA ZONE DE KEUR
MOMAR SARR (LOUGA), SENEGAL

REMERCIEMENTS
A Monsieur le représentant de l’IRD au Sénégal Georges De NONI, pour avoir accepté de nous
accueillir au sein de votre institut et de mettre à notre disposition le matériel nécessaire à notre
travail. Nous vous en remercions.
Au Docteur Cheikh SOKHNA, vous avez bien voulu nous accueillir dans votre laboratoire. Nous
vous exprimons notre profonde gratitude.
Au Docteur Oleg MEDIANNIKOV, vous avez proposé et accepté de diriger ce travail en vous
mettant entièrement à notre disposition malgré vos multiples occupations. Vous êtes pour nous un
modèle scientifique dans le travail. Ce travail est le vôtre et nous vous serons toujours
reconnaissant.
Au Professeur Ngor FAYE, pour l’intérêt que vous avez porté à notre travail et pour avoir accepté
de suivre sa réalisation et de veiller à sa rédaction.
A Monsieur Bhen Sikina TOGUEBAYE, pour avoir accepté de nous faire l’honneur d’être le
président du jury.
Au Docteur Franselyse CLOTILDE BA, pour avoir accepté de juger notre travail.
Au Docteur Cheikhna DIEBAKATE, pour avoir accepté de juger notre travail.
Au Docteur Georges DIATTA, pour la rigueur, la franchise, l’amour, la qualité du travail et la
disponibilité avec lesquelles vous avez accompagné toutes mes études depuis les premières heures
de recherche à la rédaction de ce mémoire. Ce travail est également le votre.
A Monsieur Hubert BASSENE, vous avez été toujours disponible et compréhensible. Nous vous
remercions pour n’avoir aménagé aucun effort à la réalisation de ce travail.
A l’ensemble du personnel de l’UMR 198 URMITE de l’IRD de Dakar. Soyez en remerciés.
A l’ensemble des éleveurs des villages de collecte, votre compréhension a permis de réaliser ce
travail. Soyez en remerciés.
A mes professeurs et camarades de promotion.
DEDICACES
A ma mère Kouloumy et mon père Elisse, pour avoir mis vos moyens à notre éducation. Votre
affection et votre soutien sont le fruit de ce travail. Que le tout puissant soit toujours avec vous.
A ma grande sœur Diminga Ango, tu as été toujours un modèle pour la famille et une confidente
pour moi. Merci pour ton soutien et amour. Et à mon grand frère Jonas Silonor, pour ta gentillesse
et l’intérêt que tu accordes à mes études.
A mes cousins Thomas N. MANGA, Jeannot SAMBOU pour votre soutien.
A Flora D. SAMBOU, pour ta compréhension et ton amour. Que le tout puissant soit avec toi.
A mes oncles et tantes.
A César BASSENE, Paulin SAMBOU, Boubacar KEITA, Hyacinth SAMBOU, Abdoulaye
DIALLO, Basil Z. SAMBOU, Simon Jr SAMBOU, Ampa D. DIATTA, Jacques S. SAMBOU.

SOMMAIRE INTRODUCTION ................................................................................................................................... 1
CHAPITRE I : SYNTHESE BIBLIOGRAPHIQUE .............................................................................. 3
I.1. Généralités sur les rickettsioses .................................................................................................... 3
I.2. Les fièvres boutonneuses à tiques ................................................................................................. 5
I.2.1. Eléments d’épidémiologie ......................................................................................... 6
I.2.1.1. Vecteurs et réservoirs des Rickettsia du groupe boutonneux ................................ 6
I.2.1.2. Importance .............................................................................................................. 7
I.2.1.3. Répartition géographique des rickettsioses à tiques ............................................... 7
I.2.2. Signes cliniques ......................................................................................................... 8
I.3. Généralités sur les tiques ixodides vectrices des rickettsies ........................................................ 9
I.3.1. Bio-écologie ............................................................................................................. 10
I.3.2. Position systématique .............................................................................................. 10
I.3.3. Morphologie générale ............................................................................................. 12
I.3.4. Cycles des tiques ixodides ....................................................................................... 14
I.3.4.1. Cycle de développement ...................................................................................... 14
I.3.4.2. Cycles parasitaires ................................................................................................ 15
CHAPITRE II : MATERIEL ET METHODES .................................................................................... 16
II.1. Zone d’étude .............................................................................................................................. 16
II.2. Matériel biologique .................................................................................................................... 18
II.3. Matériel de collecte .................................................................................................................... 18
II.4. Matériel de laboratoire ............................................................................................................... 18
II.5. Méthodes de collecte des tiques ixodides .................................................................................. 20
II.6. Méthodes d’étude au laboratoire................................................................................................ 21
II.6.1. Lavage des tiques ixodides ..................................................................................... 21
II.6.2. Détermination morphologique ............................................................................... 21
II.6.3. Analyses moléculaires ............................................................................................ 21
II.6.3.1. Extraction de l’ADN de tiques ............................................................................ 21
II.6.3.2. Amplification par PCR ........................................................................................ 22
II.6.3.3. Migration et révélation des produits PCR amplifiés ........................................... 23
II.6.3.4. Le séquençage ..................................................................................................... 23
CHAPITRE III : RESULTATS ET DISCUSSION .............................................................................. 24
III.1. Résultats ................................................................................................................................... 24

III.1.1. Echantillonnage et détermination morphologique des tiques ............................... 24
III.1.2. Détection des Rickettsia par PCR ........................................................................ 26
III.1.3. Caractérisation génétique des Rickettsia .............................................................. 26
III.2. Discussion ................................................................................................................................ 27
III.2.1. Prévalences de l’infection chez des tiques ixodides ............................................. 27
III.2.2. Circulation des rickettsies dans la zone d’étude ................................................... 28
CONCLUSION ET PERSPECTIVES .................................................................................................. 30
BIBLIOGRAPHIE ................................................................................................................................ 31

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
1 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
INTRODUCTION
Les rickettsies sont des bactéries intracellulaires obligatoires, gram négatif, répandues
dans le monde entier (Raoult & Parola, 2008). Celles du genre Rickettsia sont en général de
très petits bacilles (500 nm) associés à des arthropodes comme les tiques qui jouent le rôle de
vecteurs et/ou de réservoirs invertébrés (Renvoisé & Raoult, 2009). Ces Rickettsia sont des
agents pathogènes de rickettsioses à tiques du groupe boutonneux (Lagier et al., 2009). Leur
diagnostic est difficile à cause de leur croissance strictement intracellulaire. Beaucoup de
rickettsies du genre Rickettsia sont transmises par des tiques ixodides et seulement quelques
unes d’entre elles étaient connues en médecine humaine avant l’utilisation des techniques de
biologie moléculaire. Avec l’avènement de la biologie moléculaire et grâce à la mise au point
d’une nouvelle technique de culture cellulaire dérivée des cultures virales, le diagnostic des
rickettsioses a connu des progrès décisifs au cours des 20 dernières années (Ngwamidiba et
al., 2006). La technique de séquençage basée sur la comparaison des séquences des gènes
ompA, ompB et sca4 a permis d’identifier des rickettsies du genre Rickettsia et de déterminer
leur appartenance phylogénétique. De nouvelles rickettsies pathogènes qui étaient encore mal
connues des cliniciens au Sénégal ont été identifiées par cette technique, parmi lesquelles on
peut citer Rickettsia africae, l’agent de la fièvre à tiques africaine, Rickettsia conorii,
Rickettsia aeschlimannii, agents de la fièvre boutonneuse méditerranéenne, Rickettsia
massiliae et Rickettsia sibirica mongolitimonae (Mediannikov et al., 2010)
Pour ces raisons, l’Unité de Recherche sur les Maladies Infectieuses et Tropicales
Émergentes (URMITE) de l’Institut de Recherche pour le Développement (IRD) de Dakar a
initié une étude pour «déterminer les agents pathogènes responsables de fièvres non palustres
au Sénégal». Cette étude est réalisée dans cinq zones représentatives des différents faciès éco-
épidémiologique du pays et vise à diagnostiquer et identifier par des techniques de biologie
moléculaire les causes de fièvre chez des malades consultant dans des structures sanitaires. La
région de Louga et notamment la zone de Keur Momar Sarr est l’une des cinq zones
d’intervention de URMITE Dakar où les infections à Rickettsia et leur fréquence n’avaient
pas été explorées ni connues tant chez les tiques ixodides vectrices que chez l’homme.

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
2 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
Nos investigations sur les rickettsies pathogènes chez les tiques ixodides de la zone de
Keur Momar Sarr s’inscrit donc dans le cadre des objectifs sus-évoqués visant à identifier les
causes de fièvre chez des sujets fébriles.
Dans ce travail, nous présenterons dans le premier chapitre une synthèse
bibliographique, décrirons dans le deuxième chapitre le matériel et les méthodes avant de
présenter et discuter nos résultats dans le troisième chapitre.

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
3 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
CHAPITRE I : SYNTHESE BIBLIOGRAPHIQUE
I.1. Généralités sur les rickettsioses
Les rickettsioses représentent des maladies infectieuses causées par des rickettsies.
Elles sont associées aux arthropodes, essentiellement les tiques, mais aussi les poux, les puces
(Aubry, 2009 et 2010).
D’après Aubry (2009 et 2010), on distingue trois grands groupes :
Le groupe Typhus comprend Rickettsia prowazekii et Rickettsia typhi ;
Le groupe Orientia qui comprend des rickettsies proches de la bactérie Orientia
tsutsugamushi ;
Le groupe Boutonneux regroupant des pathogènes du genre Rickettsia, responsables
de la fièvre boutonneuse méditerranéenne, de la fièvre pourprée des montagnes rocheuses, de
la fièvre à tiques africaine, de la fièvre vésiculeuse ou rickettsialpox et de la fièvre
boutonneuse à puces.
La taxonomie des rickettsies a été modifiée par le développement des techniques de
biologie moléculaire (Aubry, 2009 et 2010). En 1989, il a été montré que le genre Rickettsia
appartient la classe alpha des protéobactéries (Weisburg et al., 1989) et à la famille des
Rickettsiaceae (Renvoisé & Raoult, 2009) (Tableau 1). Sur la base des caractères
phénotypiques, les rickettsies pathogènes du genre Rickettsia sont classiquement séparées en
deux groupes (Halos, 2005) :
- les bactéries du groupe typhus qui ont une localisation exclusivement intra-
cytoplasmique,
- celles du groupe boutonneux localisées dans le cytoplasme et le noyau des
cellules infectées.
Des études génomiques ont montré que les rickettsies du groupe boutonneux possèdent
le gène ompA qui est absent ou non exprimé chez celles du groupe typhus (Halos, 2005).
Du fait de leur développement strictement intracellulaire (Figure 1), la culture des Rickettsia
sur milieu inerte n’est pas possible, mais la plupart des espèces poussent en cultures
cellulaires (Raoult & Roux, 1997).

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
4 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
Figure 1 : Cellule infectée par des Ricckettsia
Source : http://ifr48.timone.univ-mrs.fr/Fiches/FBM.html
Tableau 1 : Position systématique des rickettsies du genre Rickettsia
Domaine Bacteria
Phylum Proteobacteria
Classe Alpha-proteobacteria
Ordre Rickettsiales
Famille Rickettsiaceae
Genre Rickettsia
Les premiers cas de rickettsioses ont été rapportés en Afrique du Nord par Conor &
Bruch (1910) et en Afrique du Sud par McNaught (1911) puis Sant’Anna (1912). Tous les cas
de fièvre pourprée observés en Afrique sub-saharien étaient considérés comme des cas de
fièvre boutonneuse méditerranéenne encore appelée «Spotted Fever Mediterrenean (SFM) »
due à Rickettsia conorii (Gear, 1938), malgré l’avis contraire de Pijper (1936). Pendant
longtemps, on a pensé que la fièvre boutonneuse méditerranéenne était la seule rickettsiose
connue en Afrique (Socolovschi et al., 2008). Cependant, un nouveau cas de rickettsiose due
à Rickettsia africae affilié au groupe de la fièvre pourprée ou «Spotted Fever Group (SFG) » a
été décrit au Zimbabwe chez une femme de 36 ans (Kelly et al., 1992). Cet agent pathogène
R. africae était identique aux souches de Rickettsia isolées chez les tiques Amblyomma
variegatum collectées en Ethiopie (Philip et al., 1966) et Ambryomma hebraeum, provenant
du Zimbabwe (Beati et al., 1995). R. africae a été, par la suite, identifiée par la technique de
Polymérase Chain Reaction (PCR) dans plusieurs pays d’Afrique sub-saharienne notamment

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
5 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
au Niger, Mali, Burundi et Soudan (Parola et al., 2001), au Tchad et en Ethiopie (Mura et al.,
2008) et dans beaucoup de pays d’Afrique Equatoriale et du Sud (Cazorla et al., 2008).
Rickettsia aeschlimannii isolée chez la tique Hyalomma marginatum marginatum
collectée au Maroc en 1997, a été considérée comme un agent pathogène du groupe de la
fièvre pourprée (Beati et al., 1997 ; Parola et al., 2005). Cet agent pathogène avait été détecté
par PCR au Zimbabwe chez la tique Hyalomma marginatum rufipes, au Portugal chez H.
marginatum marginatum (Béati et al., 1995), en Egypte chez H. impeltatum (Loftis et al.,
2006), au Sénégal chez les tiques H. marginatum rufipes, H. truncatum et Rhipicephalus
evertsi evertsi (Mediannikov et al., 2010). Rickettsia conorii l’agent pathogène de la fièvre
boutonneuse méditerranéenne est un complexe de quatre sous-espèces génétiquement
identiques (Renvoisé & Raoult, 2009). Il s’agit de Rickettsia conorii conorii, R. israelensis, R.
caspia et R. indica (Zhu et al., 2005). Cette rickettsie est transmise par la tique brune du
chien, Rhipicephalus sanguineus. R. israelensis est l’agent étiologique de la fièvre
boutonneuse d’Israël (Zhu et al., 2005), mais de nouveaux cas isolés en Italie et au Portugal
font penser que sa distribution géographique est beaucoup plus large (Renvoisé & Raoult,
2009). La fièvre d’Astrakhan est causée par R. caspia (Zhu et al., 2005). La maladie sévit
autour de la mer caspienne, mais sa distribution pourrait être plus vaste. La bactérie R. indica
est l’agent suspecté de la fièvre à tiques indienne «Indian tick typhus» (Renvoisé & Raoult,
2009). Rickettsia felis est l’agent causal de la fièvre boutonneuse à puces (Desenclos &
Lecollinet, 2009). Il a été identifié pour la première fois en 1918 chez des puces de chat,
Ctenocephalides felis, puis en 1990 (Aubry, 2009 et 2010). R. felis a été détecté chez des
puces dans de nombreuses régions du monde telles que l’Europe (Capelli et al., 2009), l’Asie
(Jiang et al., 2006), l’Afrique du Nord et Sub-saharienne (Bitam et al., 2006 ; Sakal et al.,
2008), l’Amérique du Nord et du Sud (Labruna et al., 2007 ; Reif & Macaluso, 2009), et
plusieurs cas de rickettsiose à R. felis ont d’ailleurs été récemment décrits en Afrique
(Socolovschi et al., 2010).
I.2. Les fièvres boutonneuses à tiques
Elles ont en commun l’apparition d’une fièvre et d’une escarre (lésion cutanée en
plaque) après une piqûre de tiques (Aubry, 2009 et 2010 ; Renvoisé & Raoult, 2009), de
frissons, de myalgies et de céphalées (Socolovschi et al., 2008).
Les tiques vectrices sont des ixodides (ou tiques dures) qui sont les vecteurs des
rickettsies du groupe boutonneux. Ce sont des acariens hématophages. Chacun des 3 stades de
développement de ces tiques (larves, nymphes, adultes mâle et femelle) ne prend qu'un repas

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
6 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
sanguin. L'épidémiologie des rickettsioses à tiques est liée aux caractéristiques écologiques et
comportementales des tiques vectrices.
Les fièvres boutonneuses à tiques comprennent près de 20 espèces de rickettsies qui peuvent
infecter l’homme (Aubry, 2009 et 2010). On distingue :
La fièvre boutonneuse méditerranéenne, encore appelée « mediterranean spotted
fever » (MSF) due à R. conorii (Renvoisé & Raoult, 2009), est une maladie urbaine et péri-
urbaine, endémique dans le pourtour méditerranéen, mais elle peut aussi survenir en Europe
centrale et en Afrique centrale et du Sud. C'est une maladie saisonnière estivale. Les
symptômes durent 12 à 20 jours et l’amélioration clinique survient après 48 heures de
traitement.
La fièvre à tiques africaine, Elle est la principale rickettsiose en Afrique sub-
saharienne (Renvoisé & Raoult, 2009). Le plus souvent les patients atteints sont des hommes
adultes. La fièvre à tiques africaine est donc une des étiologies de fièvre au retour de voyage,
à évoquer systématiquement après un séjour en zone d’endémie (Renvoisé & Raoult, 2009). Il
est important de préciser que l’élévation des titres sérologiques est plus tardive que dans les
autres rickettsioses (Brouqui et al., 2007).
La Fièvre pourprée des montagnes rocheuses, est due à Rickettsia rickettsii. Elle
est décrite dans tous les Etats-Unis et dans certains pays d'Amérique centrale et du sud
(Estripeaut et al., 2007). Les cas surviennent à la fois dans les zones rurales et urbaines,
pendant la période d’avril à septembre. Il n’y a généralement pas d’escarre au niveau de la
zone de piqûre (Aubry, 2009 et 2010). La fièvre pourprée des montagnes rocheuses est la plus
sévère des rickettsioses à tiques et reste potentiellement mortelle (Aubry, 2009 et 2010).
I.2.1. Eléments d’épidémiologie
I.2.1.1. Vecteurs et réservoirs des Rickettsia du groupe boutonneux
Les Rickettsia se maintiennent dans les populations de tiques par la transmission
transtadiale et transovarienne. Ainsi, la tique joue le rôle de vecteur et de réservoir de la
bactérie. Les Rickettsia se multiplient dans pratiquement tous les organes de leur hôte
invertébré y compris les ovaires et les oocytes chez la femelle (Aubry, 2009 et 2010). Le taux
d’œufs infectant d’une femelle de tique infectée par une souche de Rickettsia varie sous
l’influence de facteurs encore inconnus (Raoult & Roux, 1997). Chaque espèce de rickettsie
est étroitement liée à son vecteur et l'identification morphologique des tiques est d’une grande
importance.

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
7 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
La tique du chien, Rhipicephalus sanguineus, est le principal vecteur du complexe
Rickettsia conorii agent pathogène de la fièvre boutonneuse méditerranéenne (R. conorii
subsp. conorii, israelensis, caspia et indica) (Renvoisé & Raoult, 2009). Récemment, les
travaux de Mediannikov et collaborateurs (2010) ont montré la présence de R. conorii chez la
tique Rhipicephalus evertsi evertsi, au Sénégal. La tique Rh. evertsi evertsi a été également
impliquée dans le maintien de certaines rickettsies telles que, Rickettsia africae et Rickettsia
aeschlimannii (Mediannikov et al. 2010). R. aeschlimannii a pour vecteur principal les tiques
Hyalomma marginatum marginatum, H. marginatum rufipes (Renvoisé & Raoult, 2009) et H.
impeltatum (Loftis et al., 2006). Le genre Amblyomma est un vecteur potentiel (A.
variegatum, A. hebraeum et A. lepidum) de R. africae (Socolovschi et al., 2008).
I.2.1.2. Importance
L’importance des rickettsioses, comme celle de la plupart des maladies à tiques, reste
mal connue. Des données ponctuelles provenant de campagnes d’épidémiosurveillance
permettent d’avoir un aperçu de l’importance de ces maladies. Au Sénégal, une étude récente
a montré une prévalence élevée de Rickettsia africae chez la tique A. variegatum
(Mediannikov et al., 2010). Des données en provenance de divers pays d’Afrique (Mali,
Burkina Faso, République Centrafricaine, Zimbabwe) ont montré une forte prévalence
d’anticorps chez le malade (Aubry, 2009 et 2010). De plus, une rickettsie récemment
reconnue pathogène, Rickettsia aeschlimannii a été détectée à Dielmo et Ndiop, chez la tique
Hyalomma marginatum rufipes avec une incidence supérieure à 51,3% (Mediannikov et al.,
2010). Les tiques Rhipicephalus evertsi evertsi et Boophilus annulatus pouvaient être
porteuses de cette nouvelle rickettsie pathogène très peu connue (Mediannikov et al., 2010).
Ces tiques (A. variegatum, H. marginatum rufipes, Rh. evertsi evertsi et B. annulatus) peuvent
piquer accidentellement l’homme (Walker et al., 2003). L’incidence connue de la fièvre
boutonneuse méditerranéenne chez l’homme est de 0,7% des cas de rickettsioses observés
dans les villages de Dielmo et Ndiop (Mediannikov et al., 2010).
I.2.1.3. Répartition géographique des rickettsioses à tiques
Les hypothèses expliquant la dissémination des rickettsioses transmises par les tiques
reposent sur le concept de coévolution entre micro-organismes pathogènes, la tique et de
l’animal hôte (Parola & Raoult, 2001). Les rickettsioses à tiques sévissent dans des zones
optimales pour le développement de la tique vectrice et des hôtes réservoirs. Le maintien des
infections dans de nouvelles régions dépend de la présence de tiques et d’hôtes susceptibles
de porter l’infection et qui peuvent assurer la survie de l’agent pathogène (Halos, 2005).

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
8 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
L’homme peut aussi jouer un rôle important dans la dispersion des rickettsioses en
modifiant l’habitat des tiques ou par le transport d’animaux sur de longues distances. Ainsi,
l’introduction de bétail en Guadeloupe et en Inde, en provenance du Sénégal a entrainé la
présence d’A. variegatum, vecteur de R. africae agent de la fièvre à tiques africaine, dans ces
pays où la maladie est devenue endémique dans les années 30 (Hoogstraal, 1956). En Afrique,
la répartition des rickettsioses (Figure 2) est peu connue surtout en Afrique de l’Ouest, plus
particulièrement au Sénégal où des études récentes ont montré la présence de certains
Rickettsia dont Rickettsia aeschlimannii, Rickettsia mongolitimonae, Rickettsia massiliae et
Rickettsia conorii (Mediannikov et al., 2010).
Figure 2: Répartition géographique des différentes espèces de rickettsies du groupe
boutonneux en Afrique (Socolovschi et al., 2008).
I.2.2. Signes cliniques
Les rickettsioses à tiques ont des manifestations cliniques communes (Socolovschi et
al., 2008). Lors de leur transmission à l’Homme ou à l’hôte mammifère, les Rickettsia
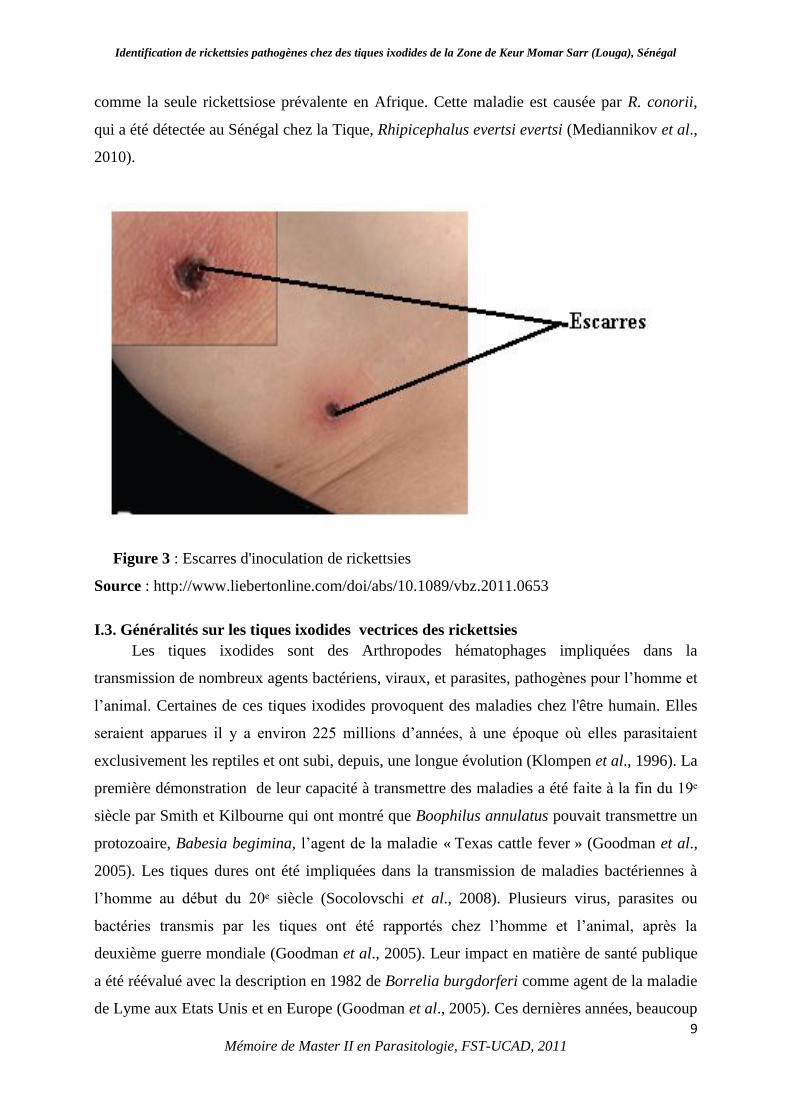
pathogènes se multiplient à leur point d’inoculation pour donner une escarre (Figure 3) suivie
de vascularité, d’inflammation péri-vasculaire et de thrombose et, éventuellement, d’atteintes
viscérales (Brouqui et al., 2004). Les rickettsioses se manifestent aussi par une fièvre
importante (> 39 °C), des myalgies, arthralgies, et céphalées (Halos, 2005). La sévérité
dépend de l’espèce pathogène (Socolovschi et al., 2008). La fièvre à tiques africaine en
Afrique subsaharienne est plutôt bénigne. La rickettsiose historiquement la plus connue en
Afrique est la fièvre boutonneuse méditerranéenne qui, jusqu’à récemment, était considérée
Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
9 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
comme la seule rickettsiose prévalente en Afrique. Cette maladie est causée par R. conorii,
qui a été détectée au Sénégal chez la Tique, Rhipicephalus evertsi evertsi (Mediannikov et al.,
2010).
Figure 3 : Escarres d'inoculation de rickettsies
Source : http://www.liebertonline.com/doi/abs/10.1089/vbz.2011.0653
I.3. Généralités sur les tiques ixodides vectrices des rickettsies
Les tiques ixodides sont des Arthropodes hématophages impliquées dans la
transmission de nombreux agents bactériens, viraux, et parasites, pathogènes pour l’homme et
l’animal. Certaines de ces tiques ixodides provoquent des maladies chez l'être humain. Elles
seraient apparues il y a environ 225 millions d’années, à une époque où elles parasitaient
exclusivement les reptiles et ont subi, depuis, une longue évolution (Klompen et al., 1996). La
première démonstration de leur capacité à transmettre des maladies a été faite à la fin du 19ᵉ
siècle par Smith et Kilbourne qui ont montré que Boophilus annulatus pouvait transmettre un
protozoaire, Babesia begimina, l’agent de la maladie « Texas cattle fever » (Goodman et al.,
2005). Les tiques dures ont été impliquées dans la transmission de maladies bactériennes à
l’homme au début du 20ᵉ siècle (Socolovschi et al., 2008). Plusieurs virus, parasites ou
bactéries transmis par les tiques ont été rapportés chez l’homme et l’animal, après la
deuxième guerre mondiale (Goodman et al., 2005). Leur impact en matière de santé publique
a été réévalué avec la description en 1982 de Borrelia burgdorferi comme agent de la maladie
de Lyme aux Etats Unis et en Europe (Goodman et al., 2005). Ces dernières années, beaucoup

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
10 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
de nouvelles maladies humaines transmises par les tiques ixodides ont été décrites à travers le
monde. Les tiques sont considérées comme le deuxième groupe de vecteurs de maladies
humaines après les moustiques (Goodman et al., 2005). A l'inverse des moustiques, elles
restent longtemps en contact (7 à 10 jours) avec des hôtes diversifiés par lesquels elles vont
assurer la diffusion des agents pathogènes. Ainsi, les germes circulent d’une population
réservoir d’hôtes vertébrés aux tiques qui vont les transmettre à d’autres hôtes vertébrés. La
longévité exceptionnelle de la tique en fait à la fois un bon vecteur et un excellent réservoir.
I.3.1. Bio-écologie
Les tiques dures vivent dans un écosystème particulier où leur vie est influencée par la
végétation, les conditions climatiques et les interactions qu’elles entretiennent avec les autres
êtres vivants. La distribution géographique de ces tiques, leur cycle de vie, la variation
saisonnière de leur activité, la dynamique des populations et leur comportement sont
essentiellement influencés par les facteurs climatiques (Daniel & Dusbabek, 1994).
Cependant, chaque espèce de tique vectrice présente une distribution géographique
particulière (Socolovschi et al., 2008).
I.3.2. Position systématique
Les tiques appartiennent à l’embranchement des arthropodes, à la sous-classe des
acariens et à l’ordre des ixodida (Figure 4). La classification des tiques fait toujours l’objet de
discussions parmi les écoles de systématique, américaine, française et russe. Ainsi, la lecture
des différentes publications concernant les tiques est rendue difficile par le nombre important
de noms qui ont pu être attribués aux différents taxa à travers l’histoire (Socolovschi et al.,
2008). Cependant, Camicas et al. ont publié une synthèse des différents noms et synonymes
utilisés dans la littérature avec 869 espèces ou sous-espèces répertoriées au 1ᵉʳ janvier 1996.
Cette publication constitue la référence majeure francophone en taxonomie (Camicas et al.,
1998 ; Socolovschi et al., 2008). Il existe 223 espèces de tiques en Afrique dont 180 sont des
tiques dures et 43 des tiques molles (Socolovschi et al., 2008).

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
11 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
Figure 4: classification des tiques d'après Camicas et al., 1998 (le nombre d’espèces présentes
dans la région afro tropicale est indiqué en astérisque).

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
12 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
I.3.3. Morphologie générale
Les tiques dures sont des acariens de grande taille, au corps globuleux. Le dimorphisme
sexuel est plus ou moins accentué, le mâle étant plus petit que la femelle. Les adultes et les
nymphes ont 4 paires de pattes alors que les larves en ont 3. Les adultes se distinguent des
nymphes par l’ouverture de leur appareil génital. Contrairement aux insectes, les tiques n’ont
pas d’antennes. Leur corps (Figure 5) n’est pas divisé en tête, thorax et abdomen. Mais, il se
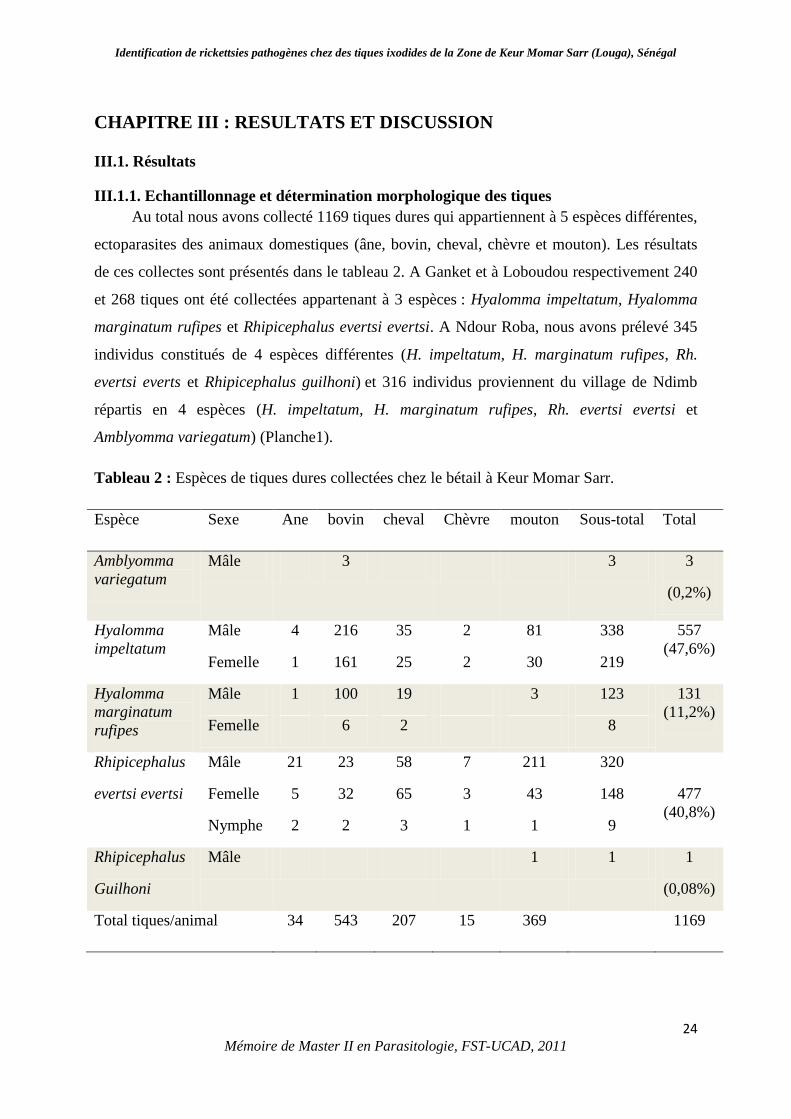
compose d’un gnathosoma et d’un idiosoma. Le gnathosoma est composé d’un capitulum de
forme rectangulaire ou hexagonale (Figure 7) constituant la zone de liaison au corps, et d’un
rostre qui porte les pièces buccales comprenant des organes sensoriels (les pédipalpes), des
organes perforateurs (les chélicères) et un organe médian immobile (hypostome) avec de
nombreuses dents qui ancrent les tiques dans la peau de l’hôte. L’idiosoma (corps) porte les
organes locomoteurs (Halos, 2005 ; Pérez-Eid, 2007). Les tiques ixodides sont caractérisées
par la présence d’une plaque dorsale très dure, le scutum. Le reste du corps est recouvert d’un
tégument extensible qui se distend lors du repas sanguin (Halos, 2005). Chez le mâle, le
scutum recouvre l’intégralité de la surface dorsale, alors qu’il recouvre seulement la partie
antérieure chez la femelle de même que chez les formes immatures, ce qui rend aisée la
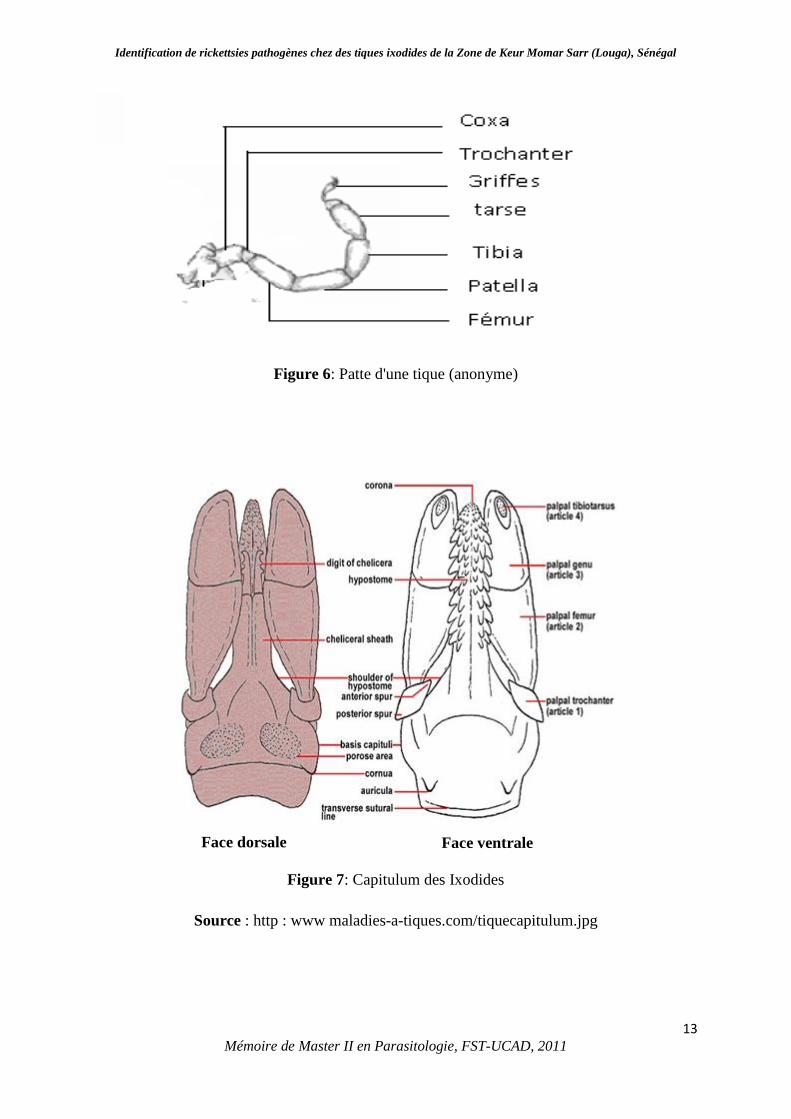
différentiation du sexe (Socolovschi et al., 2008). Les pattes (Figure 6) sont formées de 6
segments : coxa, trochanter, fémur, patelle, tibia, tarse terminé par une ventouse (pulville) et 2
griffes.
Figure 5: Morphologie d'une tique ixodide (Hyalomma impeltatum femelle, vue dorsale)
Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
13 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
Figure 6: Patte d'une tique (anonyme)
Figure 7: Capitulum des Ixodides
Source : http : www maladies-a-tiques.com/tiquecapitulum.jpg
Face dorsale Face ventrale

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
14 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
I.3.4. Cycles des tiques ixodides
I.3.4.1. Cycle de développement
Les tiques ixodides ont 3 stades de développement actifs (Figure 8) : larvaire, nymphal
et adulte. Les stades sont séparés par un repas sanguin qui peut durer plusieurs jours et qui est
suivi d’une mue. Après copulation, la femelle pond environ 1 000 à 20 000 œufs dans un abri.
La durée de l’incubation est de 20 à 50 jours selon les espèces, la température et l’humidité.
La larve éclot 2 à 4 semaines après la ponte. Son repas sanguin dure 3 à 5 jours. Quand
la larve est bien gorgée, elle se laisse tomber au sol, y cherche un abri pour effectuer la mue
qui a une durée de 2 à 8 semaines. Le même comportement est répété par la nymphe (c’est-à-
dire les déplacements, la quête de l’hôte, le repas sanguin de 3 à 5 jours). La métamorphose
en adulte est en général plus longue, jusqu’à 20 à 25 semaines dans les conditions les plus
défavorables (Socolovschi et al., 2008). L’accouplement des adultes se déroule au sol ou sur
l’hôte selon les espèces. La durée du repas sanguin est de 5 à 10 jours. Certaines femelles
commencent leur repas avant l’accouplement et le terminent après la fécondation. La femelle
se détache de l’hôte après son repas de sang, se laisse tomber au sol et recherche une zone
ombragée où elle reste pendant 3 à 4 semaines avant d’entamer la ponte qui dure 10 à 30
jours. La durée d’un cycle est en moyenne de 2 à 4 ans, pouvant aller jusqu’à 7 ans si les
conditions climatiques ne sont pas favorables. Les tiques dures sont très sensibles à la
dessiccation. Si les conditions climatiques ne sont pas favorables, la tique entre en diapause,
un état caractérisé par une chute du métabolisme et un développement retardé.
Figure 8 : Cycle de développement des ixodides
Source: http : //upload.wikimedia.org/wikipedia/commons/a/a5/

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
15 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
I.3.4.2. Cycles parasitaires
Les cycles parasitaires des ixodides (Figure 9) sont de trois types :
Le type monophasique dont les stades se succèdent sur un même et unique hôte
vertébré.
Le type diphasique pour lequel les trois stades se déroulent sur deux hôtes : un premier
hôte où se développent la larve et la nymphe, et un second hôte parasité par l’adulte.
Le type triphasique où les trois stades parasitent trois hôtes différents. La plupart des
espèces de tiques d’intérêt médical et vétérinaire ont un cycle parasitaire triphasique.
Le type de cycle parasitaire est caractéristique de l’espèce de tique (Bussieras & Chermette,
1991).
On distingue des cycles triphasiques monotropes dans lesquels les trois hôtes font partie
du même groupe zoologique. D’autres espèces de tiques ont des préférences de nutrition
propres à chaque stade évolutif et la spécificité d’hôte varie entre les différents stades dans la
même espèce. Dans les cycles triphasiques ditropes, les stades immatures parasitent des
micromammifères, les adultes, des carnivores ou des ongulés.
Enfin, certaines espèces ont une faible spécificité d’hôte. On parle alors de tiques
télotropes. Ainsi les différents stades d’Ixodes ricinus peuvent se nourrir sur plus de 300
espèces d’hôtes qui vont des oiseaux aux grands mammifères. Si les larves se nourrissent de
préférence (90 %) sur les micromammifères et les oiseaux de petite taille, les nymphes sont
plus ubiquistes et se nourrissent indifféremment sur les petits et grands mammifères
(ruminants sauvages ou domestiques) et ce sont elles qui sont les principaux vecteurs de
maladies humaines à tiques. Les adultes, eux, se nourrissent de préférence sur des animaux de
grande taille (Parola & Raoult, 2001 ; Estrada-Peña et al., 2004).
Figure 9 : Cycle parasitaire des Ixodides

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
16 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
CHAPITRE II : MATERIEL ET METHODES
II.1. Zone d’étude
Le village de Keur Momar Sarr (15°55’N, 15°57’W), chef lieu d’arrondissement, est
dans la communauté rurale de Keur Momar Sarr. La communauté rurale de Keur Momar Sarr
est située au Nord-Ouest du Sénégal, à 52 kilomètres de la région de Louga. Elle est limitée
au Nord par la communauté rurale de Ross Béthio, au Sud par celle de Nger Malal, à l'Est par
celle de Syer et à l'Ouest par celle de Sakal (Figures10). Keur Momar Sarr couvre une
superficie d’environ 760 kilomètres carrés, pour 84 villages. Cette communauté rurale
appartient à la région écologique sahélienne caractérisée par une alternance de deux saisons
annuellement contrastées (hivernage et saison sèche) (Plan d’Action Environnemental
Régional (PAER) de Louga, 2007).
Les températures moyennes annuelles sont très élevées avec des variations
interannuelles très importantes (PAER Louga, 2007). Par ailleurs, le climat est fortement
influencé par le microclimat relativement doux du complexe lac de Guiers-Bas Ferlo. La
végétation est dominée par un tapis herbacé bien fourni après les années de bonne
pluviométrie et une strate arbustive, arborée peu dense (PAER, Louga, 2007).
Selon le Programme d'eau potable et d'assainissement du Millénaire (PEPAM), Keur
Momar Sarr compte 1 774 habitants et 171 ménages. Cette population est constituée en
majorité de 54 % de wolofs, de 43 % de peuls et de 3 % de maures et autres ethnies.
Entre autres aspects du développement local, on peut signaler l’existence d’un Collège
d’Enseignement Moyen, d’un marché hebdomadaire influent, d’un foyer des jeunes et
récemment d’un détachement de la gendarmerie nationale du fait de la dynamique actuelle des
populations rurales. On note aussi l’existence d’une station de traitement et de pompage d'eau
potable pour l’approvisionnement de l'agglomération dakaroise, d’un poste de santé et d’une
pharmacie.
L’élevage est l’une des principales activités des populations, avec un cheptel composé
de bovins, caprins, équins et camelins. La transmission de pathogènes à l’homme par les
tiques dures est facilitée par la cohabitation avec le bétail. Notre étude a été réalisée dans
quatre villages de la communauté rurale de Keur Momar Sarr : Ganket, Loboudou, Ndour
Roba et Ndimb (Figure 11).
Le village de Ganket est séparé de Keur Momar Sarr village par le lac de Guier. Il est
situé au Nord Est (15°58’ N ; 15°55’ W) de Keur Momar Sarr et compte 872 habitants et 84

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
17 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
ménages selon le programme d’eau potable et d’assainissement du millénaire (PEPAM). Le
village abrite un poste de santé et une mosquée.
Le village de Loboudou se trouve à l’Est (15°57’ N ; 15°55’ W) de Keur Momar Sarr, et
il est séparé de ce dernier par le lac de Guier. L’élevage traditionnel est une des activités les
plus pratiquées par les habitants majoritairement composés de peuls. Il possède aussi un poste
de santé.
Ndour Roba village est situé au Nord Ouest (15°54’ N ; 15°59’ W) de Keur Momar Sarr
village. Il est en majorité peuplé de peuls, qui vivent dans des baraques en carton et/ou en
paille. L’élevage et la culture de l’arachide sont les activités principales qui y sont pratiquées.
Le village de Ndimb est situé au Nord (16°02’ N ; 16°00’ W) de Keur Momar Sarr. Il
est majoritairement habité par des Wolofs. On y trouve également un poste de santé.
L’élevage pratiqué est de type traditionnel.
Figure 10: Carte du Sénégal montrant la zone d'étude

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
18 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
Figure 11: Vue aérienne de Keur Momar Sarr montrant les sites d'étude.
Source : GoogleEarth
II.2. Matériel biologique
Le matériel biologique est représenté par des tiques ixodides collectées à partir des
animaux domestiques (ânes, bovins, chevaux, chèvres et moutons).
II.3. Matériel de collecte
Pour la collecte de tiques dures nous avons utilisé des :
Pots à urine ;
Autocollants ;
Marqueurs ;
Désinfectant ;
Sachets plastique.
II.4. Matériel de laboratoire
Le matériel utilisé pour les études de laboratoire est composé de matériel de lavage,
d’identification et de biologie moléculaire.
Matériel de lavage (Figure 12) :
- Eau de javel ;
- Savon ;

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
19 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
- Bacs ;
- Tamis ;
- Pinces souples ;
- Papier serviette.
Figure 12: Matériel de lavage des tiques dures collectées
Matériel d’identification :
- Aiguille à disséquer ;
- Boites de pétri ;
- Loupe binoculaire ;
- Clé d’identification ;
- Tubes eppendorf (1,5ml) ;
- Boites de rangement.
Matériel destiné à la biologie moléculaire
- Solutions (cetyl trimethyl ammonium bromide à 2%, chloroforme, isopronol, éthanol70°) ;
- Bain marie ;
- Centrifugeuse ;
- Speed bac ;
- Portoirs tubes eppendorf (1,5ml et 0,2ml) ;
- Tubes eppendorf (1,5ml et 0,2ml) ;

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
20 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
- Pipettes (P10µl, P200µl, P1000µl) ;
- Lames de bistouri ;
- Réfrigérateur (T : +2 à +8°c) ;
- Congélateur (T : -20°c) ;
- Gants ;
- Poubelles ;
- Hotte.
Le port de la blouse est obligatoire.
II.5. Méthodes de collecte des tiques ixodides
La technique de collecte des tiques (Figure 13) consiste à maitriser l’animal, puis à
examiner les parties sensibles susceptibles d’être parasitées par les tiques comme la région
anale, les mamelles, les oreilles. Les tiques collectées sont placées dans un pot à urine sur
lequel les indications suivantes sont mentionnées : type d’animal (cheval, bovin etc.), lieu ou
zone de collecte et la date. Pour maintenir les tiques vivantes, le bouchon de chaque pot est
percé (d’un trou d’environ 1,5 cm de diamètre) et fermé avec un tissu en soie. Les tubes sont
mis dans un sachet plastique dans lequel nous introduisons un papier serviette imbibé d’eau
pour y maintenir l’humidité.
Figure 13: Collecte de tiques ixodides

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
21 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
II.6. Méthodes d’étude au laboratoire
II.6.1. Lavage des tiques ixodides
Afin de limiter les contaminations de l’ADN par des micro-organismes présents à la
surface des tiques, toutes les tiques collectées sont lavées dans un bac contenant une solution
de savon, d’eau de javel diluée et un peu d’alcool.
La méthode consiste à vider le contenu du pot à urine dans un tamis, puis les tiques sont
immergées dans la solution contenue dans le bac. Avec une pince souple nous prenons chaque
tique que nous nettoyons soigneusement avec du papier serviette. Après nettoyage, la tique est
conservée dans un tube falcon contenant de l’alcool 70°. Sur le tube falcon nous mentionnons
les mêmes indications figurant sur le pot à urine correspondant. Les tiques sont ainsi
conservées dans l’éthanol jusqu’à leur détermination morphologique.
II.6.2. Détermination morphologique
Chaque tique a été identifiée et le sexe a été déterminé à la loupe binoculaire grâce à la
clé d’identification taxonomique standard des tiques adultes (Hoogstraal, 1956 ; Mathysse et
al., 1987 ; Walker et al., 2000 ; Walker et al., 2003 ).
Le contenu de chaque tube falcon est vidé dans une boite de pétri. Avec des pinces
souples, nous prélevons chaque tique puis nous l’étalons dans le couvercle de la boite de pétri
pour l’examiner à la loupe. Nous procédons à la détermination du sexe, du genre, de l’espèce
et l’âge selon les instructions de la clé d’identification utilisée (Hoogstraal, 1956 ; Mathysse et
al., 1987 ; Walker et al., 2000 ; Walker et al., 2003 ). Après identification, les tiques sont
conservées à sec dans des tubes eppendorfs. Pour chaque espèce identifiée, est attribué un
code mentionné sur son tube. Le code est une référence de base de données où sont notés :
code, nom de l’espèce, sexe, lieu de collecte, animal, âge. Les tubes eppendorfs sont rangés
dans des boites de rangement à température ambiante jusqu’à l’extraction de l acide
désoxyribonucléique(ADN).
II.6.3. Analyses moléculaires
L’objectif principal des études moléculaires vise à détecter la présence de rickettsies
par la technique de Polymerase chain reaction (PCR), après avoir broyé les tiques afin d’en
extraire leur ADN et de déterminer les espèces de Rickettsia des tiques infectées par le
séquençage.
II.6.3.1. Extraction de l’ADN de tiques
L’extraction de l’ADN des tiques ixodides semble être difficile du fait de la présence
d’une cuticule rigide qui rend difficile l’accès aux tissus internes colonisés par les pathogènes.

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
22 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
La technique d’extraction consiste à broyer finement les tissus de la tique entière dans
du Cetyl Trimethyl Ammonium Bromide (CTAB) à 2%, avec une lame de bistouri. Une lame
de bistouri est utilisée par tique. Le broyat a été digéré par chauffage au bain-marie à 65°C
pendant 5 minutes, pour débarrasser les échantillons d’un maximum de protéines pouvant
interférer dans les étapes ultérieurs. Nous avons ajouté au broyat un volume de 200
microlitres (µl) de chloroforme et avons récupéré le surnageant après centrifugation à 12000
tours par minute (trs/minute) pendant 5 minutes. Les acides nucléiques ont été précipités par
200µl d’isopropanol après 15 minutes de centrifugation à 12000 trs/minute. Le culot a été
séché au speed bac pendant 3 à 4 minutes et a été repris par élution avec 200µl d’eau pure. La
solution d’ADN est conservée au réfrigérateur à une température comprise entre 2° et 8°C.
II.6.3.2. Amplification par PCR
Le principe de la PCR consiste à amplifier de façon spécifique une séquence d’acide
nucléique ADN ou ARN dans un échantillon biologique défini par des amorces. Nous
produisons ainsi en grande quantité cet acide nucléique pour le rendre plus facilement
détectable même lorsque l’échantillon d’origine est pauvre en matériel biologique.
Nous avons réalisé toutes les réactions PCR dans un thermocycleur mastercycler.
Chaque réaction a été conduite dans un volume final de 25µl, contenant 2,5µl de tampon PCR
10X (buffer), 2,5µl de désoxyribonucléotides (dNTP), 0,75µl de chlorure de magnésium
(MgCl2), 0,5µl de chaque amorce, 0,1µl de Taq polymérase, 15,15µl d’eau pure et 3,0µl
d’ADN. Deux témoins, positif Rickettsia conorii et négatif eau stérile, ont été inclus à chaque
expérience.
La présence de l’ADN de Rickettsia sp. a été testée en utilisant un couple d’amorces
permettant d’amplifier spécifiquement un fragment d’environ 330 paires de bases du gène de
l’ARNr 16S de Rickettsia sp. (Regnery et al., 1991). Dans nos premiers tests nous avons
amplifié le gène gltA, avec le couple d’amorces (Rp877 - Rp1258), mais sa sensibilité à nos
échantillons était faible. Nous avons repris nos tests avec le gène ompA (couple d’amorces
190.70 - 190.701) qui est un gène rencontré chez toutes les rickettsies du groupe boutonneux.
Ce gène a une longueur d’environ 632 paires de bases. Le témoin positif était un extrait
d’ADN de Rickettsia conorii aimablement fourni par le Docteur Mediannikov de l’Unité des
Rickettsies de Marseille en France.

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
23 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
II.6.3.3. Migration et révélation des produits PCR amplifiés
Les fragments d’ADN amplifiés ont été séparés en fonction de leur taille sur un gel
d’agarose à 1,5% dans du tampon 1mole par litre (X). Le Biothium, utilisé comme révélateur
des acides nucléiques émet une fluorescence lorsqu’il est exposé à une lumière ultra-violette.
Un prélèvement de 10µl de chaque échantillon amplifié est mélangé avec une goutte de
tampon de charge (10X), dont le rôle est d’augmenter la densité du mélange, donc de faciliter
le dépôt de l’échantillon dans les puits, et de pouvoir suivre la migration grâce au bleu de
bromophénol.
Un marqueur de taille, choisi pour donner des bandes de 100 à 1000 paires de bases, est
déposé dans un puits à raison de 2,5µl dans chaque gel. La migration est effectuée sous 100
volts pendant 25 minutes, suivi d’une photographie du gel en lumière ultraviolette.
II.6.3.4. Le séquençage
Le séquençage des amplicons a été réalisé à Marseille, en utilisant le BigDye Teminator
Cycle Sequencing Kit (Perkin Elmer Applied Biosystems) avec séquenceur automatisé ABI
(Applied Biosstems). Les séquences obtenues ont été assemblées (ChromasPro 1.49 beta,
technelysium Pty Ltd, Tewantin, Australia), puis éditées par BioEdit Sequence alignement
editor v. 7.0.9.0 (Hall, 1999) et comparées avec celles disponibles dans GenBank par NCBI
BLAST (http://blast.ncb.nlm.nih.gov/Blast.cgi). Les séquences des gènes gltA et ompA des
Rickettsia choisies dans GenBank pour comparaison ont été concaténées et alignées avec le
programme ClustalW.
Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
24 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
CHAPITRE III : RESULTATS ET DISCUSSION
III.1. Résultats
III.1.1. Echantillonnage et détermination morphologique des tiques
Au total nous avons collecté 1169 tiques dures qui appartiennent à 5 espèces différentes,
ectoparasites des animaux domestiques (âne, bovin, cheval, chèvre et mouton). Les résultats
de ces collectes sont présentés dans le tableau 2. A Ganket et à Loboudou respectivement 240
et 268 tiques ont été collectées appartenant à 3 espèces : Hyalomma impeltatum, Hyalomma
marginatum rufipes et Rhipicephalus evertsi evertsi. A Ndour Roba, nous avons prélevé 345
individus constitués de 4 espèces différentes (H. impeltatum, H. marginatum rufipes, Rh.
evertsi everts et Rhipicephalus guilhoni) et 316 individus proviennent du village de Ndimb
répartis en 4 espèces (H. impeltatum, H. marginatum rufipes, Rh. evertsi evertsi et
Amblyomma variegatum) (Planche1).
Tableau 2 : Espèces de tiques dures collectées chez le bétail à Keur Momar Sarr.
Espèce Sexe Ane bovin cheval Chèvre mouton Sous-total Total
Amblyomma
variegatum
Mâle 3 3 3
(0,2%)
Hyalomma
impeltatum
Mâle 4 216 35 2 81 338 557
(47,6%) Femelle 1 161 25 2 30 219
Hyalomma
marginatum
rufipes
Mâle 1 100 19 3 123 131
(11,2%) Femelle 6 2 8
Rhipicephalus
evertsi evertsi
Mâle 21 23 58 7 211 320
477
(40,8%)
Femelle 5 32 65 3 43 148
Nymphe 2 2 3 1 1 9
Rhipicephalus
Guilhoni
Mâle 1 1 1
(0,08%)
Total tiques/animal 34 543 207 15 369 1169

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
25 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
Planche 1: Différentes espèces de tique ixodides collectées et étudiées (vue dorsale) :
(A) Amblyomma variegatum, mâle ; (B) Rhipicephalus guilhoni, mâle ; (C) & (D) Hyalomma
impeltatum, femelle et mâle ; (E) & (F) Hyalomma marginatum rufipes, femelle et mâle ; (G)
& (H) Rhipicephalus evertsi evertsi, mâle et femelle ;

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
26 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
III.1.2. Détection des Rickettsia par PCR
Nous avons analysé 617 échantillons de tiques. Parmi ces 617 échantillons de tiques
nous avons détecté par PCR 53 (8,5%) individus de tiques infectés par des rickettsies. Les
résultats de l’amplification des gènes gltA et ompA par PCR sont présentés dans le tableau 3.
III.1.3. Caractérisation génétique des Rickettsia
Le séquençage des individus de tiques positifs a conduit à l’identification de 2 espèces
de rickettsies du genre Rickettsia : Rickettsia africae agent de la fièvre à tiques africaine et
Rickettsia aeschlimannii récemment reconnu comme un agent de la fièvre boutonneuse
méditerranéenne (Tableau 3). Chez les H. marginatum rufipes infectés, nous avons trouvé
chez un individu l’espèce Rickettsia africae et chez les autres Rickettsia aeschlimannii. Le
bovin est l’animal domestique où nous avons rencontré le plus de tiques infectées par des
rickettsies (83%).
Tableau 3 : Résultats des analyses PCR et du séquençage
Espèces PCR Séquençage Animal examiné
A. variegatum 2/2 (100%) 1/2 Rickettsia africae BV
H. impeltatum 17/ 364 (4,6%) 14/17 Rickettsia aeschlimannii BV, CV
H. marginatum
rufipes
28/82 (34,1%) 26/28 R. aeschlimannii, R. africae BV, CV
Rh. evertsi evertsi 6/169 (3,5%) 5/6 Rickettsia aeschlimannii CV, MT
Total 53/617 (8,5%) R. africae, R. aeschlimannii 3 (BV, CV et MT)
NB : BV = Bovin ;
CV = Cheval
MT = Mouton

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
27 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
III.2. Discussion
III.2.1. Prévalences de l’infection chez des tiques ixodides
Les études que nous avons réalisées dans le Nord du Sénégal confirment que quatre
espèces de tiques ixodides (tiques dures), impliquées dans la transmission des fièvres
boutonneuses et d’autres maladies infectieuses, sont très répandues chez les animaux
domestiques et en contact permanent avec l’homme.
H. impeltatum, est l’une des tiques les plus abondantes et les plus fréquemment
rencontrées. Elle se nourrit sur une grande variété d’animaux sauvages et domestiques ainsi
que l’homme (Hoogstraal, 1956). Au cours de notre étude, nous avons obtenu une prévalence
élevée de H. impeltatum (47,65%). Cela pourrait s’expliquer par son aire de répartition qui
s’étend de l’Afrique du Nord en passant par le Soudan, l’Erythrée, la Somalie, le Nord du
Kenya, la Tanzanie et les pays de l’Afrique de l’Ouest où le climat est steppique (Walker et
al., 2003). Cette espèce bien que très commune et répandue dans l’Afrique du Nord n’a pas
été souvent étudiée comme un vecteur d’agents pathogènes humain et animal. Elle est plus
connue comme vecteur de maladie humaine dans la transmission du virus de la fièvre
hémorragique de Criminée Congo (VFHCC). Des études faites par Loftis et al. (2006) en
Egypte, ont permis d’identifier chez cette tique Rickettsia aeschlimannii. Au Sénégal, nos
études ont montré pour la première fois la présence de Rickettsia aeschlimannii chez cette
tique. Rickettsia aeschlimannii est récemment connue comme rickettsie pathogène du groupe
de la fièvre boutonneuse méditerranéenne ou « Mediterranean spotted fever » (MSF) (Parola
et al., 2005), montrant ainsi le rôle que H. impeltatum pourrait jouer dans l’épidémiologie des
fièvres boutonneuses. Malgré son faible taux d’infection (lequel est en fait toujours faible
pour tous les vecteurs de MSF) (Rovery et al., 2008), l’abondance de cette tique et son contact
avec l’homme qui vit à proximité du bétail dans les zones rurales, augmentent
considérablement la probabilité de contamination.
La tique Rh. evertsi evertsi se nourrit non seulement sur les animaux domestiques, mais
aussi sur beaucoup d’animaux sauvages en Afrique (Socolovschi et al., 2010). Cette tique est
restreinte à l’Afrique sub-saharienne (Walker et al., 2003). Ce qui explique sa forte
prévalence dans le Nord et le Sud du Sénégal (Mediannikov et al., 2010). Les principaux
pathogènes connus à ce jour et associés à cette tique sont le protozoaire Babesia bigemina
(Hove et al., 1998), le virus de la fièvre hémorragique de Criminée Congo (Zeller et al.,1997)
et Ehrlichia ruminantium en Afrique du Sud ( Allsopp et al., 2007). Des études récentes, de
Mediannikov et collaborateurs (2010) ont montré l’infection de cette tique par des rickettsies

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
28 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
du groupe boutonneux (R. africae, R. aeschlimannii, R. conorii) dans le Sud du Sénégal. Nos
résultats confirment ainsi le portage de R. aeschlimanni au Sénégal, par la tique Rh. evertsi
evertsi avec un pourcentage relativement faible.
Hyalomma marginatum rufipes est largement distribuée dans presque tous les pays de
l’Afrique ; cependant sa répartition est inégale et elle est plus fréquente dans les zones sèches
(Walker et al. 2003). Cette tique est la plus importante en Afrique du Sud dans la transmission
de VFHCC. Elle est également connue dans le portage de R. aeschlimannii en Ethiopie, au
Tchad (Mura et al., 2008) et très récemment au Sud du Sénégal (Mediannikov et al., 2010).
Amblyomma variegatum est une tique connue des régions tropicales et subtropicales
(www.cfsph.iastate.edu/Factsheets/pdfs/amblyomma_variegatum.pdf). La tique se nourrit sur
des animaux domestiques ainsi que sur l’homme. Elle est le principal vecteur de Rickettsia
africae (Mediannikov et al., 2010), agent de la fièvre à tiques africaine ou « African Tick-Bite
Fever » (ATBF) (Kelly et al., 1996 ; Jensenius et al., 2003). Au Sud du Sénégal, Mediannikov
et collaborateurs ont rapporté une prévalence élevée de cette tique en 2010. Au Nord du
Sénégal la prévalence de cette tique est très faible. Nous pensons que cette différence
significative pourrait être due aux facteurs climatiques, car le Nord du Sénégal (Keur Momar
Sarr) est une région appartenant à la zone sahélienne (PAER Louga, 2007). Si nous nous
référons à la littérature classique, la tique A. variegatum exige des conditions climatiques
humides (Mediannikov et al., 2010). Ce qui pourrait expliquer la faible prévalence de la tique
dans le Nord du Sénégal.
III.2.2. Circulation des rickettsies dans la zone d’étude
Rickettsia africae
Nos enquêtes épidémiologiques, nous ont permis de détecter R. africae au Nord du
Sénégal. Cette rickettsie est l’agent de la fièvre à tiques africaine ou « african tick-bite fever »
(ATBF) (Kelly et al., 1996 ; Jensenius et al., 2003). Cet agent pathogène transmis par la tique
A. variegatum, a été décrit dans les années 30 par Pijper en Afrique du Sud, qui distingue
clairement la maladie de la fièvre boutonneuse méditerranéenne due à Rickettsia conorii, tant
sur le plan clinique qu’épidémiologique (Parola et Barre, 2004). Cet agent fut nommé R.
africae après les travaux de Kelly en 1990 au Zimbabwe (Kelly et al., 1996). En 2001, Raoult
et al. ont rapporté la présence d’une rickettsiose à tiques dans le monde, due à Rickettsia
africae chez des touristes européens provenant principalement de l’Afrique (Brouqui et al.,
2007). Très récemment Mediannikov et al. (2010) ont détecté par la technique de biologie
moléculaire (PCR), R. africae chez des tiques A. variegatum comme partout en Afrique et Rh.

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
29 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
evertsi evertsi dans des zones rurales (Dielmo/Ndiop) du Sénégal. La rickettsie a été
auparavant détectée par PCR, dans plusieurs pays d’Afrique dont le Niger, le Mali, le Burundi
et le Soudan (Parola et al., 2001) et aussi dans des pays d’Afrique Equatoriale (Mediannikov
et al., 2010). L’histoire de R. africae dans les différentes régions où elle a été détectée est
comparable aux résultats que nous avons obtenus dans la région Nord du Sénégal. D’après
nos résultats une seule souche de R. africae a été isolée chez deux espèces de tiques
différentes A. variegatum comme partout en Afrique et H. marginatum rufipes.
L’identification de R. africae chez H. marginatum rufipes augmenterait l’aire de répartition de
cette rickettsie dans le nord, car la distribution du pathogène est fonction de l’aire de
répartition de son vecteur et de son hôte.
Rickettsia aeschlimannii
R. aeschlimanni, agent de la fièvre boutonneuse méditerranéenne ou « spotted fever
mediterranean » (SFM) (Parola et al., 2005), a été détectée chez H. impeltatum, H.
marginatum rufipes et Rh. evertsi evertsi. Après sa première détection chez H. impeltatum en
Egypte (Loftis et al., 2006), Afrique Nord-Est, nous rapportons ici sa deuxième détection chez
H. impeltatum en Afrique et la première en Afrique de l’Ouest. La bactérie a été considérée
comme un pathogène appartenant au groupe de la fièvre pourprée « spotted fever group »
(SFG), après son isolement chez la tique H. marginatum marginatum au Maroc (Beati et al.,
1997). Des souches identiques ont été auparavant isolées chez H. marginatum rufipes au
Zimbabwe, et chez H. marginatum marginatum au Portugal (Beati et al., 1995). R.
aeschlimannii a été aussi détectée plus tard au Niger et au Mali (Parola et al., 2001). Au
Sénégal elle a été détectée chez H. marginatum rufipes, Rh. evertsi evertsi et chez H.
truncatum collectées à partir des animaux domestiques à Dielmo / Ndiop (Mediannikov et al.,
2010). D’après nos résultats le nombre élevé d’espèces de tiques infectées par R.
aeschlimannii dans le Nord du Sénégal est comparable à celui du Sud. Avec le comportement
des éleveurs, vivant à proximité de leurs animaux, nous pensons que des cas d’infections
humaines sont possibles, d’autant plus que les travaux de Mokrani et al. (2008) ont confirmé
la pathogénicité de cette rickettsie. Les travaux de Matsumoto et al. (2004) ont montré la
transmision transtadiale et transovarienne de cette rickettsie chez la tique Hyalomma et qui
joue à la fois le rôle de vecteur et réservoir.

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
30 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
CONCLUSION ET PERSPECTIVES
L’étude des tiques dures dans le Nord du Sénégal comparée à celle de Dielmo/Ndiop,
nous permet de noter une prévalence très élevée de la population de tique Hyalomma
impeltatum, impliquée dans la transmission de nombreux pathogènes comme Rickettsia
aeschlimannii (Loftis et al., 2006). Cette tique est absente dans le sud (Walker et al., 2003).
De plus cette étude nous a permis de noter l’existence de la tique Amblyomma variegatum
dans le nord du Sénégal (région sahélienne) bien que sa prévalence et son abondance soient
faibles par rapport au sud du Sénégal. Les tiques Hyalomma marginatum rufipes et
Rhipicephalus evertsi evertsi ont des prévalences comparables aux résultats obtenus dans le
Sud du Sénégal. La tique Rhipicephalus guilhoni a été collectée bien qu’elle n’était pas
infectée comme c’était le cas à Dielmo/Ndiop. Nos résultats montrent la présence de tiques
ixodides de bétail fréquemment infectées par des rickettsies pathogènes. Ce qui nous amène à
penser qu’il pourrait y avoir des cas d’infections humaines.
Nos travaux nous ont permis de détecter 2 espèces de rickettsies du groupe boutonneux,
Rickettsia africae et R. aeschlimannii. R. africae agent de la fièvre à tiques africaine, a été
identifiée non seulement chez la tique A. variegatum comme partout en Afrique, mais aussi
chez H. marginatum rufipes. Il s’agit d’une première identification de R. africae chez cette
tique H. marginatum rufipes. Ce qui est très intéressante dans la connaissance de la
distribution de la fièvre à tiques africaine. R. aeschlimannii a été identifiée chez la tique H.
impeltatum qui est une première identification en Afrique de l’Ouest et chez H. marginatum
rufipes et Rh. evertsi evertsi comme le cas à Dielmo/Ndiop. Les prévalences élevées des
infections à Rickettsia évoquent un risque majeur de contamination des populations rurales.
Notre étude nous a permis de noter en outre une faible diversité des rickettsies dans le
Nord en comparaison avec les résultats des études réalisées dans le Sud (Dielmo/Ndiop).
Cependant, nos travaux doivent être complétés par une enquête séroépidémiologique
afin d’apprécier le degré d’exposition des populations à R. africae et à R. aeschlimannii. De
plus, la sensibilisation des cliniciens devrait permettre de décrire les premiers cas humains de
rickettsiose dans le Nord du Sénégal.

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
31 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
BIBLIOGRAPHIE
1. ALLSOPP MT, VAN STRIJP MF, FABER E, JOSEMANS AI & ALLSOPP BA, 2007.
Ehrlichia ruminantium variants which do not cause heartwater found in
South Africa. Vet Microbiol, 120: p. 158-166.
2. AUBRY P, Rickettsioses: Actualités 2009. 2009, 2010, Diplôme en
médecine tropicale de l'océan indien.
3. BEATI L, KELLY PJ, MATTHEWMAN LA, MASON P and RAOULT D, 1995.
Prevalence of Rickettsia-like organisms and spotted fever group
Rickettsiae in ticks (Acari : Ixodidae) from Zimbabwe. J Med Entomol, 32:
p. 787–792.
4. BEATI L, MESKINI M, THIERS B & RAOULT D, 1997. Rickettsia aeschlimannii
sp. nov., a new spotted fever group rickettsia associated with Hyalomma
marginatum ticks. Int J Syst Bacteriol., 47(2): p. 548-554.
5. BITAM I, PAROLA P, DE LA CRUZ KD, MATSUMOTO K & AL., 2006.First
molecular detection of Rickettsia felis in fleas from Algeria. Am J Trop Med
Hyg, 74: p. 532-535.
6. BROUQUI P, BACELLAR F, BARANTON G, BIRTLES RJ, BJOERSDORFF A,
BLANCO JR, CARUSO G, RAOULT D & AL., 2004. Guidelines for the diagnosis
of tick-borne bacterial diseases in Europe. Clin Microbiol Infect, 10(10): p.
1108-1132.
7. BROUQUI P, PAROLA P, FOURNIER PE & RAOULT D, 2007. Spotted fever
rickettsioses in southernand easternEurope. FEMS Immunol Med
Microbiol, 49: p. 2–12.
8. BUSSIERAS J and CHERMETTE R, 1991. Entomologie vétérinaire (Abrégé de
parasitologie vétérinaire, fascicule IV). Service de Parasitologie de
l'E.N.V.A :163p.
9. CAMICAS JL, HERVY J, ADAM F & MOREL PC, 1998. Les tiques du Monde
(Acarida, Ixodidea) : nomenclature stades décrits, hôtes, répartition.
Orstom Paris: p. 233 pp.
10. CAPELLI G, MONTARSI F, PORCELLATO E, MAIOLI G & AL., 2009. Occurrence
of Rickettsia felis in dog and cat fleas (Ctenocephalides felis) from Italy.
Parasit Vectors, 2: p. Suppl 1:S8.
11. CAZORLA C, SOCOLOVSCHI C, JENSENIUS M & PAROLA P, 2008. Tick-borne
diseases: tick-borne spotted fever rickettsioses in Africa. Infect Dis Clin
North Am, 22: p. 531–544.
12. CONOR A & BRUCH A, 1910.Une fièvre eruptive observée en Tunisie. Bull
Soc Pathol Exot Filial, 8: p. 492–496.
13. DANIEL M & DUSBABEK F, 1994. Micrometeorological and microhabitats
factors affecting maintenance and dissemination of tick-borne diseases in
the environment. In: Sonenshine DE, Mather TN eds. Ecological dynamics
of tick-borne zoonoses. NewYork: Oxford University Press: p. 91-138.

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
32 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
14. DESENCLOS J-C and LECOLLINET S, 2009.Les zoonozes transmises par les
vecteurs. Lutte antivectorielle en France: p. 92-195.
15. ESTRADA-PEÑA A, BOUATTOUR A, CAMICAS JL & WALKER AR, 2004. Ticks of
domestic animals in the Mediterranean region: a guide to identification of
species. University of Zaragoza, ITG Library, Zaragoza, Espagne: p. 131.
16. ESTRIPEAUT D, ARAMBURÚ MG, SÁEZ-LLORENS X, THOMPSON HA, DASCH GA,
PADDOCK CD & al., 2007. Rocky Mountain spotted fever, Panama. Emerg
Infect Dis, 13: p. 1763-1765.
17. GEAR JHS, 1938.South African typhus. S Afr J Med Sci, 3: p. 134-160.
18. GOODMAN JL, DENNIS DT & SONENSHINE DE, 2005. Tick-borne diseases of
humans. ASM Press,Washington, DC, USA.
19. HALL TA, 1999. BioEdit: a user-friendly biological sequence alignment
editor and analysis program for Windows 95/98/NT. Nucl Acids Symp Ser,
41: p. 95–98.
20. HALOS L, 2005, Détection de bactéries pathogènes dans leur vecteur, les
tiques dures (Acarien : Ixodidae). Paris-Grignon, L’Institut National
Agronomique, 175p.
21. HOOGSTRAAL H, 1956.Ticks of the Sudan (With Special Reference to
Equatoria Province and with Preliminary Reviews of the Genera Boophilus,
Magaropus, and Hyalomma). 3.
22. HOVE T, SITHOLE N, MUNODZANA D & MASAKA S, 1998. Isolation and
characterization of a Babesia species from Rhipicephalus evertsi evertsi
ticks picked off a sable antelope (Hippotragus niger) which died of acute
babesiosis. Onderstepoort J Vet Res, 65: p. 75-80.
23. IBARRA V, OTEO JA, PORTILLO A, SANTIBÁ˜NEZ S, BLANCO JR, METOLA L &
AL. , 2006. Rickettsia slovaca infection: DEBONEL/TIBOLA. Ann N Y Acad
Sci., 1078: p. 206-214.
24. JENSENIUS M, FOURNIER PE, VENE S, HOEL T, HASLE G, HENRIKSEN AZ & AL.,
2003. African tick bite fever in travelers to rural sub-equatorial Africa. Clin
Infect Dis., 36: p. 1411-1417.
25. JIANG J, SOEATMADJI DW, HENRY KM & RATIWAYANTO S, 2006. Rickettsia
felis in Xenopsylla cheopis, Java, Indonesia. Emerg Infect Dis, 12: p. 1281-
1283.
26. KELLY PJ, BEATI L, MASON PR, MATTHEWMAN LA, ROUX V & RAOULT D,
1996. Rickettsia africae sp. nov., the etiological agent of African tick bite
fever. Int J Syst Bacteriol, 46: p. 611-614.
27. KELLY PJ, MATTHEWMAN LA, BEATI L, RAOULT D, MASON P & AL., 1992.
African tick-bite fever: a new spotted fever group rickettsiosis under an old
name. Lancet, 340: p. 982–983.
28. KLOMPEN JHS, BLACK WC, KEIRANS JE & OLIVER JH, 1996. Evolution of
ticks. Annu Rev Entomol, 41: p. 141-161.
29. LABRUNA MB, OGRZEWALSKA M, MORAES-FILHO J, LEPE P & AL.,
2007.Rickettsia felis in Chile. Emerg Infect Dis, 13: p. 1794-1795.

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
33 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
30. LAGIER JC, DOUDIER B & PAROLA P, 2009. Rickettsioses. Elsevier Masson
SAS, 4-1110: p. résumé.
31. LOFTIS AD, REEVES WA, SZUMLAS DE, ABBASSY MM, HELMY IM, MORIARITY
JR & DASCH GA, 2006. Rickettsial agents in Egyptian ticks collected from
domestic animals. Exp Appl Acarol, 40: p. 67–81.
32. MATSUMOTO K, PAROLA P, BROUQUI P & RAOULT D, 2004. Rickettsia
aeschlimannii in Hyalomma ticks from Corsica. Eur. J. Clin. Microbiol.
Infect. Dis, 23: p. 732-734.
33. MCNAUGHT JG, 1911.A tick-bite in the Union of South Africa. J R Army Med
Corps, 16: p. 505.
34. MEDIANNIKOV O, DIATTA G, FENOLLAR F, SOKHNA C, TRAPE JF & RAOULT D,
2010. Tick-borne rickettsioses, neglected emerging diseases in rural
Senegal. PLoS Negl Trop Dis, 4(9): p. e821.
35. MOKRANI N, PAROLA P, TEBBAL S, DALICHAOUCHE M, AOUATI A & AL.,
2008.Rickettsia aeschlimannii infection, Algeria. Emerg Infect Dis, 14: p.
1814-1815.
36. MURA A, SOCOLOVSCHI C, GINESTA J, LAFRANCE B, MAGNAN SE, ROLAIN JM,
DAVOUST B, RAOULT D & PAROLA P, 2008. Molecular detection of spotted
fever group rickettsiae in ticks from Ethiopia and Chad. sciences direct,
102: p. 945—949.
37. NGWAMIDIBA M, RAOULT D & FOURNIER PE, 2006. Les Rickettsies :
caractères microbiologiques, identification, relations avec les arthropodes,
pathogénie des infections (2e partie). Elsevier Masson SAS, 8(3): p. 166-
174.
38. PAROLA P & BARRE N, 2004. Rickettsia africae, agent de la fièvre à tique
africaine : un pathogène émergent dans les Antilles et l’Ile de La Réunion.
Bull Soc Pathol Exot, 97(3): p. 193-198.
39. PAROLA P, INOKUMA H, CAMICAS JL, BROUQUI P and & RAOULT D, 2001.
Detection and identification of spotted fever group Rickettsiae and
Ehrlichiae in African ticks. . Emerg. Infect. Dis., 7: p. 1014-1017.
40. PAROLA P & RAOULT D, 2001. Ticks and Tickborne Bacterial Diseases in
Humans: An Emerging Infectious Threat. Clinical Infectious Diseases, 32:
p. 897–928.
41. PAROLA P, PADDOCK CD & RAOULT D, 2005. Tick-borne rickettsiosis around
the world: emerging diseases challenging old concepts. Clin Microbiol Rev,
18(4): p. 719-756.
42. PÉREZ-EID C, 2007. Les tiques. Identification, biologie, importance médicale
et vétérinaire. Coll. Monographies de microbiologie: p. 314 p.
43. PHILIP CB, HOOGSTRAAL H, REISS-GUTFREUND R & CLIFFORD CM, 1966.
Evidence of rickettsial disease agents in ticks from Ethiopian cattle. Bull
World Health Organ, 35: p. 127–131.
44. PIJPER A, 1936. Etude expérimentale comparée de la Fièvre boutonneuse
et de la tick-bite-fever. Arch Inst Pasteur Tunis, 25: p. 388-401.

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
34 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
45. RAOULT D & PAROLA P, 2008. Rocky Mountain spotted fever in the USA: a
benign disease or a common diagnostic error? Lancet Infect Dis Clin North
Am, 8(10): p. 587–589.
46. RAOULT D & ROUX V, 1997. Rickettsioses as paradigms of new or emerging
infectious diseases. Clin Microbiol Rev., 10: p. 694-719.
47. REGNERY R, SPRUILL C and PLIKAYTIS B, 1991.Genotypic identification of
Rickettsiae and estimation of intraspecies sequence divergence for
portions of two rickettsial genes. J Bacteriol, 173(5): p. 1576-1589.
48. REIF KE & MACALUSO KR, 2009. Ecology of Rickettsia felis: a review. J Med
Entomol, 46: p. 723-736.
49. RENVOISÉ A & RAOULT D, 2009. An update on rickettsiosis. Medecine et
maladies infectieuses, 39: p. 71-81.
50. ROVERY C & RAOULT D, 2008. Mediterranean spotted fever. Infect Dis Clin
North Am, 22(3): p. 515-530.
51. SACKAL C, LAUDISOIT A, KOSOY M, MASSUNG R & AL., 2008. Bartonella spp.
and Rickettsia felis in fleas, Democratic Republic of Congo. Emerg Infect
Dis, 14: p. 1972-1974.
52. SANT’ANNA JF, 1912. On a disease in man following tick-bites and
occurring in Lourenco Marques. Parasitology, 4: p. 87–88.
53. SOCOLOVSCH C, MEDIANNIKOV O, SOKHNA C, TALL A, DIATTA G, BASSENE H
and AL E, 2010. Rickettsia felis-associated uneruptive fever, Senegal.
Emerg Infect Dis, 16(7): p. 1140-1142.
54. SOCOLOVSCHI C, DOUDIER B, PAGES F & PAROLA P, 2008. Tiques et
maladies transmises à l’homme en Afrique. Med Trop, 68(2): p. 119-133.
55. WALKER A-R, BOUATTOUR A, ESTRADA-PEÑA A, HORAK I-G, LATIF A-A,
PEGRAM R-G and PRESTON P-M, 2003.Ticks of Domestic Animals in Africa:
a Guide to Identification of Species.
56. WALKER J-B, KEIRANS J-E and HORAK I-G, 2000.The Genus Rhipicephalus
( Acari, Ixodidae): a Guide to the Brown Ticks of the World.
57. WEISBURG WG, DOBSON ME, SAMUEL JE, DASCH GA, MALLAVIA LP, BACA O
& AL., 1989. Phylogenetic diversity of the Rickettsiae. J Bacteriol, 171: p.
4202–4206.
58. ZELLER HG, CORNET JP, DIOP A & CAMICAS JL, 1997. Crimean-Congo
hemorrhagic fever in ticks (Acari: Ixodidae) and ruminants: field
observations of an epizootic in Bandia, Senegal (1989-1992). J Med
Entomol, 34: p. 511-516.
59. ZHU Y, FOURNIER PE, EREMEEVA M & RAOULT D, 2005. Proposal to create
subspecies of Rickettsia conorii based on multi-locus sequence typing and
an emended description of Rickettsia conorii. BMC Microbiol, 5: p. 11.

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
35 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
Annexe 1
Protocol d’extraction d’ADN au CTAB, à partir d’une tique entière
Les tiques ou tissus doivent être nettoyés dans l’alcool et à secs.
1- Broyer chaque tique dans 200µl de CTAB à 2%,
2- Mettre au Bain-Marie à 65°c pendant 5 minutes,
3- Ajouter 200µl de chloroforme – Mélanger par vortex/inversion.
4- Centrifuger 5 minutes à 12000tours/munite (trs/mn),
5- Prélever la phase supérieure et la mettre dans un autre tube,
6- Ajouter 200µl d’isopropanol dans le surnageant - Bien mélanger par vortex/inversion
7- Centrifuger 15 minutes à 12000trs/mn,
8- Vider l’isopropanol, bien égoutter et ajouter 200µl d’éthanol 70%,
9- Centrifuger à 5 minutes à 12000trs/mn,
10- Vider l’éthanol,
11- Sécher le culot 5 minutes maximum au speed-vac,
12- Reprendre dans 200µl de tampon d’élution (ou l’eau distillée DNase/RNase free),
13-stocker la solution à T (+2 - +8)

Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
36 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
Annexe 2
Préparation du Gel d’agarose
Préparer un gel à 1,5% d’agarose
1- Peser 1,5 g d’agarose pour 100 ml de tampon TAE.
2- Mélanger puis chauffer à la micro-onde jusqu’à la dissolution complète de l’agarose (le
mélange doit être clair sans bulle) environ 2 minutes.
3- Laisser se refroidir un peu et ajouter 5µl de BIOTIUM pour 100ml environ une goutte.
Préparer la cuve et couler le gel dans le portoir
1- Attendre que le gel soit bien sec avant de retirer les peignes environ 15 minutes.
2- Placer le portoir dans la cuve, remplir de tampon TAE à 1X, ôter les arrêtoirs
3- Charger les puits de 10µl d’amplicons mélangés au tampon de charge (sur PARAFILM)
4- Mettre 2,5 µl de marqueur de taille (small larder) à une extrémité
5- Couvrir la cuve
6- Laisser migrer le gel 25 minutes à 100V
7- A la fin de la migration, éteindre le courant, enlever le couvercle et sortir le portoir, le
placer sur le trans-illuminateur, et visualiser les bandes fluorescentes à l’obscurité.
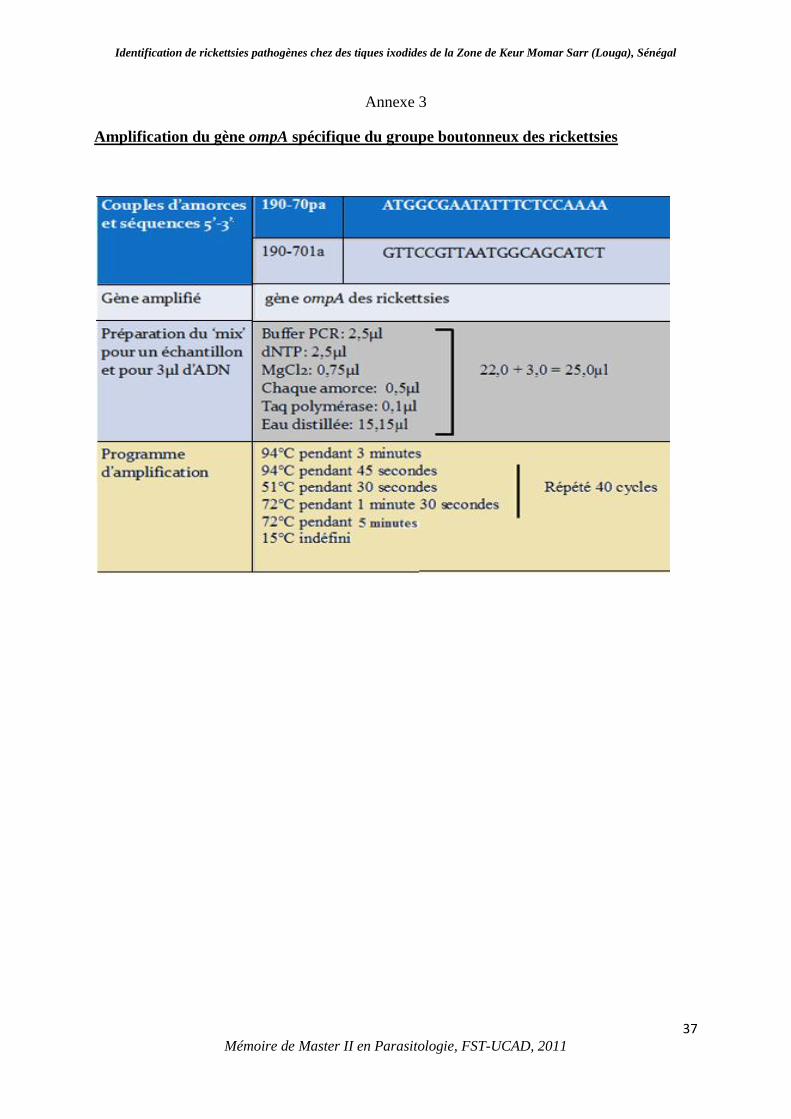
Identification de rickettsies pathogènes chez des tiques ixodides de la Zone de Keur Momar Sarr (Louga), Sénégal
37 Mémoire de Master II en Parasitologie, FST-UCAD, 2011
Annexe 3
Amplification du gène ompA spécifique du groupe boutonneux des rickettsies
Top Related