







Languages
Pages
Legal

Generación y conducción de potenciales de acción
Fabiola Garrido, Yancarla Maldonado. Medicina USACH.
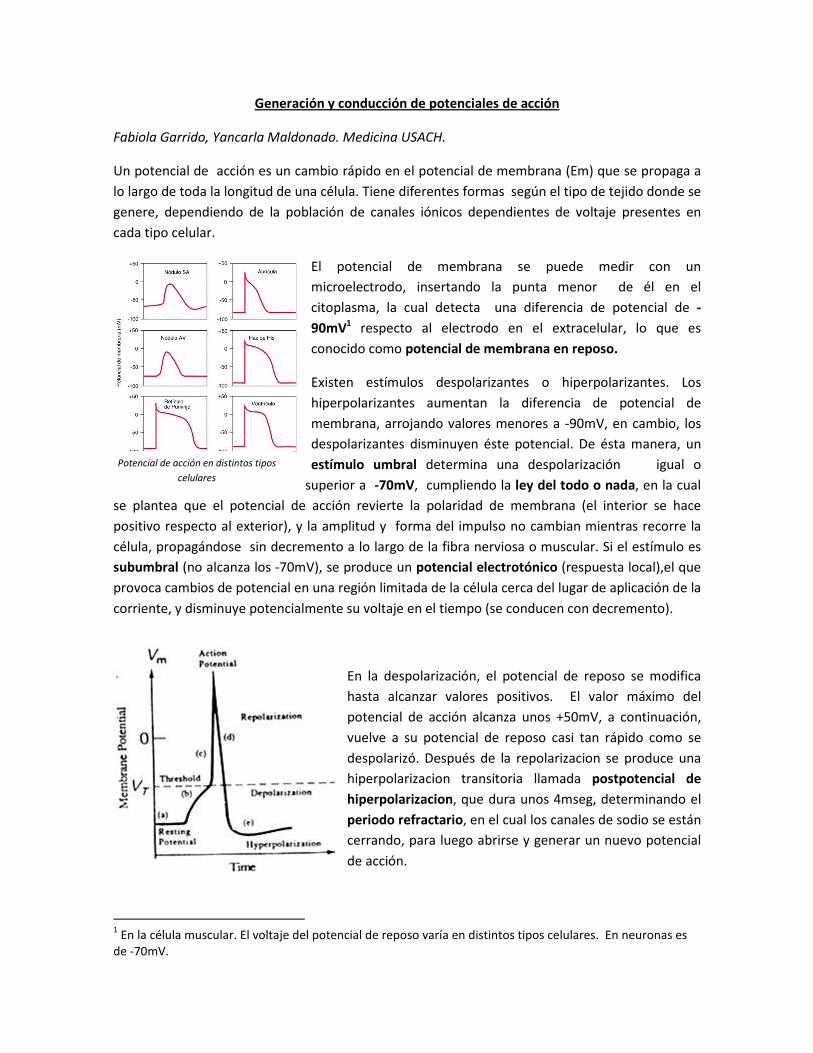
Un potencial de acción es un cambio rápido en el potencial de membrana (Em) que se propaga a
lo largo de toda la longitud de una célula. Tiene diferentes formas según el tipo de tejido donde se
genere, dependiendo de la población de canales iónicos dependientes de voltaje presentes en
cada tipo celular.
El potencial de membrana se puede medir con un
microelectrodo, insertando la punta menor de él en el
citoplasma, la cual detecta una diferencia de potencial de -
90mV1 respecto al electrodo en el extracelular, lo que es
conocido como potencial de membrana en reposo.
Existen estímulos despolarizantes o hiperpolarizantes. Los
hiperpolarizantes aumentan la diferencia de potencial de
membrana, arrojando valores menores a -90mV, en cambio, los
despolarizantes disminuyen éste potencial. De ésta manera, un
estímulo umbral determina una despolarización igual o
superior a -70mV, cumpliendo la ley del todo o nada, en la cual
se plantea que el potencial de acción revierte la polaridad de membrana (el interior se hace
positivo respecto al exterior), y la amplitud y forma del impulso no cambian mientras recorre la
célula, propagándose sin decremento a lo largo de la fibra nerviosa o muscular. Si el estímulo es
subumbral (no alcanza los -70mV), se produce un potencial electrotónico (respuesta local),el que
provoca cambios de potencial en una región limitada de la célula cerca del lugar de aplicación de la
corriente, y disminuye potencialmente su voltaje en el tiempo (se conducen con decremento).
En la despolarización, el potencial de reposo se modifica
hasta alcanzar valores positivos. El valor máximo del
potencial de acción alcanza unos +50mV, a continuación,
vuelve a su potencial de reposo casi tan rápido como se
despolarizó. Después de la repolarizacion se produce una
hiperpolarizacion transitoria llamada postpotencial de
hiperpolarizacion, que dura unos 4mseg, determinando el
periodo refractario, en el cual los canales de sodio se están
cerrando, para luego abrirse y generar un nuevo potencial
de acción.
1 En la célula muscular. El voltaje del potencial de reposo varía en distintos tipos celulares. En neuronas es
de -70mV.
Potencial de acción en distintos tipos
celulares
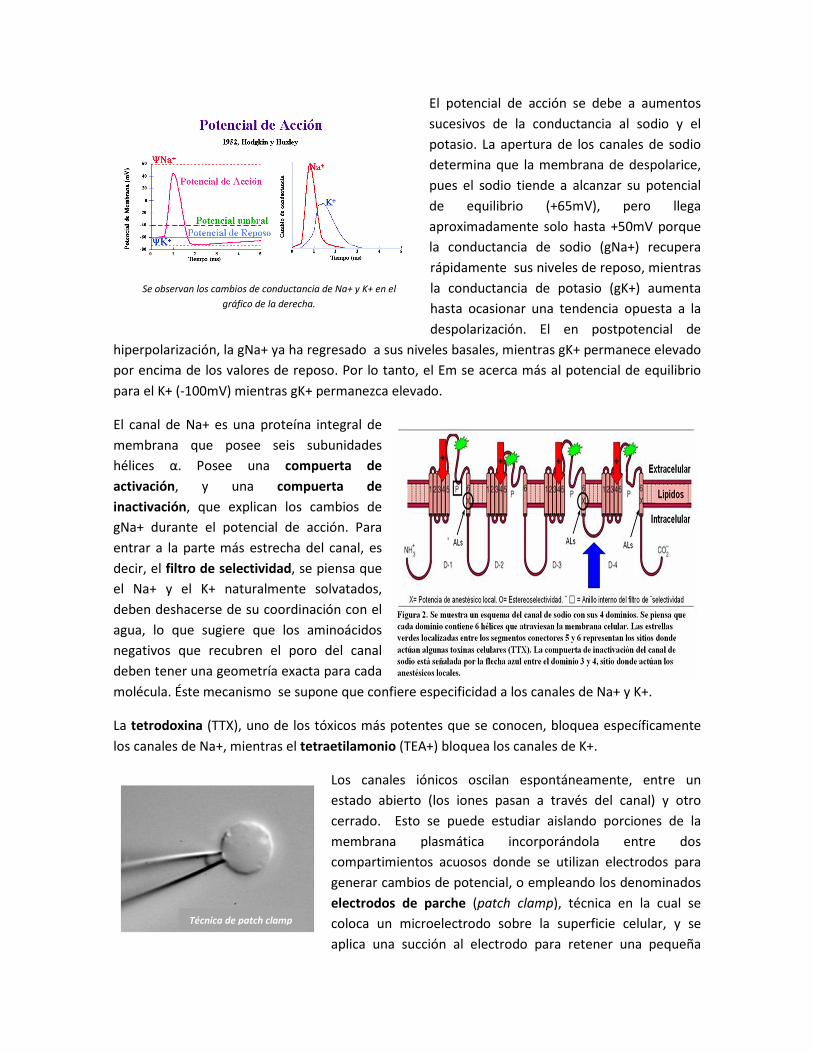
El potencial de acción se debe a aumentos
sucesivos de la conductancia al sodio y el
potasio. La apertura de los canales de sodio
determina que la membrana de despolarice,
pues el sodio tiende a alcanzar su potencial
de equilibrio (+65mV), pero llega
aproximadamente solo hasta +50mV porque
la conductancia de sodio (gNa+) recupera
rápidamente sus niveles de reposo, mientras
la conductancia de potasio (gK+) aumenta
hasta ocasionar una tendencia opuesta a la
despolarización. El en postpotencial de
hiperpolarización, la gNa+ ya ha regresado a sus niveles basales, mientras gK+ permanece elevado
por encima de los valores de reposo. Por lo tanto, el Em se acerca más al potencial de equilibrio
para el K+ (-100mV) mientras gK+ permanezca elevado.
El canal de Na+ es una proteína integral de
membrana que posee seis subunidades
hélices α. Posee una compuerta de
activación, y una compuerta de
inactivación, que explican los cambios de
gNa+ durante el potencial de acción. Para
entrar a la parte más estrecha del canal, es
decir, el filtro de selectividad, se piensa que
el Na+ y el K+ naturalmente solvatados,
deben deshacerse de su coordinación con el
agua, lo que sugiere que los aminoácidos
negativos que recubren el poro del canal
deben tener una geometría exacta para cada
molécula. Éste mecanismo se supone que confiere especificidad a los canales de Na+ y K+.
La tetrodoxina (TTX), uno de los tóxicos más potentes que se conocen, bloquea específicamente
los canales de Na+, mientras el tetraetilamonio (TEA+) bloquea los canales de K+.
Los canales iónicos oscilan espontáneamente, entre un
estado abierto (los iones pasan a través del canal) y otro
cerrado. Esto se puede estudiar aislando porciones de la
membrana plasmática incorporándola entre dos
compartimientos acuosos donde se utilizan electrodos para
generar cambios de potencial, o empleando los denominados
electrodos de parche (patch clamp), técnica en la cual se
coloca un microelectrodo sobre la superficie celular, y se
aplica una succión al electrodo para retener una pequeña
Se observan los cambios de conductancia de Na+ y K+ en el
gráfico de la derecha.
Técnica de patch clamp

porción de la membrana donde puede haber uno o más canales iónicos funcionales. El estudio con
ésta técnica ha permitido determinar que el comportamiento de los canales individuales de Na+ es
aleatorio. La corriente macroscópica de Na+ es la corriente promedio generada a partir de miles
de éstos canales, de ésta manera, el “canal promedio” se abre como respuesta a la
despolarización, y luego de un breve periodo se cierra, aun cuando se mantenga aplicada la
despolarización.
La conducción de un potencial de acción a lo largo de un axón se basa en el flujo de corriente local. Para que un impulso eléctrico pueda circular por toda la longitud de un axón sin sufrir una reducción de intensidad, el potencial de acción se debe regenerar a sí mismo durante su conducción por la fibra, por lo tanto el potencial no solo de conduce sino que también se propaga. La propagación implica la generación de potenciales de acción “nuevos” conforme se va transmitiendo por la longitud de la célula.
En resumen, la propagación se produce por ciclos repetidos de despolarización que generan un flujo de corriente local suficiente para que una región adyacente de la membrana celular cree un potencial de acción. Así, el potencial de acción se propaga a larga distancia conservando la misma forma y tamaño.
En el SN el potencial de acción se genera primero en el segmento inicial del axón y se conduce al extremo terminal porque éste está rodeado de una elevada densidad de canales de Na+ regulados por voltaje, lo que permite que sea el punto de menos umbral en la célula.
Cualquier potencial de acción que se propague por el tercio medio del axón podrá generar otro en la dirección hacia el soma porque las zonas precedentes son refractarias.
Un solo axón no puede transmitir la información codificada de forma adecuada sobre sucesos que acontecen con mayor frecuencia que su capacidad de conducir potenciales de acción. Para eso pueden ser necesarias varias neuronas combinadas.
Efecto del Diámetro de la fibra sobre la velocidad de conducción: En las fibras amielínicas, la velocidad de conducción es proporcional a la raíz cuadrada del diámetro. Este efecto se relaciona con la resistencia longitudinal. Por lo tanto, el potencial de acción se conduce con mayor rapidez por las fibras de mayor diámetro. Sin embargo, la necesidad de mantener una mayor capacitancia tiende a retrasar la velocidad de conducción y a mitigar el aumento de la velocidad de conducción conseguido mediante el incremento del diámetro.
Mielinización
La mielina corresponde a las membranas plasmáticas de las células de Schwann o de la oligodendroglia que rodean y aíslan la fibra nerviosa. En la vaina de mielina se producen hendiduras cada 1-2 mm (Nódulos de Ranvier) y miden una micra de anchura.

Un axón mielinizado conduce a una velocidad muy superior a uno amielinizado del mismo calibre, porque la vaina de mielina aumenta la constante de longitud, reduce la capacitancia y limita la generación de potenciales de acción a los nódulos de Ranvier. La menor capacitancia de los axones mielinizados implica una despolarización más rápida.
Los canales de Na+ que generan el potencial de acción están densamente concentrados en los nódulos por eso el potencial solo se regenera en los propios nódulos. La resistencia al flujo de iones a través de la vaina es tan elevada que las corrientes transmembrana quedan limitadas a cortos trayectos de membrana plasmática desnuda (en los nódulos); por eso el potencial de acción se regenera en cada nódulo sucesivo. Pareciera que el potencial saltara de un nódulo a otro, lo que se conoce como conducción saltatoria.
Consecuencias funcionales de la mielinización
La elevada velocidad de conducción permite reflejos rápidos y también es responsable de que el procesamiento mental sea complejo y eficiente.
También tienen una mayor eficacia metabólica que los amielínicos porque las corrientes iónicas se limitan a los nódulos, por lo tanto muchos menos iones atraviesan una unidad de longitud de membrana de fibra y se necesita mucha menos acción de bombeo y por lo tanto menos gasto energético para mantener la gradiente.
Transducción y Codificación Sensitiva
Para que el SN reciba información, debe estimularse mediante la aplicación de energía que debe transducirse en un acontecimiento neural (potencial de acción). La estimulación es la acción de una energía ambiental mediante la activación de uno o más receptores sensitivos. Un estímulo es el acontecimiento ambiental (mecánico, térmico, químico, etc.) que excita los receptores sensitivos, aportando la información posteriormente acerca del estímulo al SNC.

La respuesta frente al estímulo es el efecto que éste ejerce sobre el organismo y puede producirse a nivel de:
• Potenciales receptores en los receptores sensitivos
• Transmisión de potenciales de acción a lo largo de los axones en las vía sensitivas.
• Acontecimientos sinápticos en las redes neuronales centrales.
• Actividad motora desencadenada por la estimulación sensitiva y que determina el comportamiento.
El proceso que permite al receptor sensitivo responder de forma útil ante un estímulo es la
Transducción Sensitiva.
En los quimiorreceptores, la unión de una sustancia química estimuladora a la molécula receptora abre un canal iónico que permite la entrada de una corriente iónica que despolariza la célula receptora sensitiva. En el mecanorreceptor es una fuerza mecánica la que abre el canal.
La Transducción Sensitiva suele producir un Potencial Receptor en la neurona aferente primaria, que suele ser un acontecimiento despolarizante, que se debe a una corriente de entrada que consigue que el potencial de membrana del receptor sensitivo se aproxime al umbral para generar un potencial de acción.
Un estímulo umbral es el estímulo más débil que se puede detectar de un modo fiable. Los estímulos de intensidad más débil puede generar potenciales de receptor subumbral, que no excita las células sensitivas centrales (no son detectados).
La Adaptación, un cambio en la forma de respuesta del receptor ante una estimulación prolongada o secuencial, es una propiedad característica de los receptores sensitivos, que condiciona que se adapten mejor para la transmisión de unos tipos particulares de información sensitiva. Es preciso mencionar que los receptores de dolor no son adaptativos.
Las distintas características temporales de un estímulo se pueden transmitir por los receptores mediante diversas velocidades de adaptación.
El campo receptor de una neurona sensitiva (campo receptor excitador) es la región que condiciona el comportamiento de la misma cuando se estimula. El campo receptor de una neurona del SNC es la suma de los campos de los receptores sensitivos que influyen sobre ella.
La localización del campo receptor está determinada por la localización del aparato de transducción sensitiva responsable de transmitir la información acerca del estímulo a la neurona sensitiva.
En general los campos son excitadores pero una neurona sensitiva central puede tener un campo excitador o inhibidor o uno complejo (incluye ambas áreas).

Codificación Sensitiva
Las neuronas sensitivas codifican estímulos. Durante el proceso de transducción sensitiva se debe producir la codificación de uno o más aspectos (modalidad de la sensación, localización espacial, umbral, intensidad, frecuencia y duración) del estímulo para que el SNC lo pueda interpretar. La información codificada es una abstracción basada en:
• Qué receptores sensitivos se activan.
• Las respuestas de estos receptores sensitivos frente al estímulo.
• El procesamiento de la información en la vía sensitiva.
La modalidad sensitiva es fácil de distinguir y su codificación se realiza mediante canales marcados en la mayor parte de los sistemas sensitivos y se origina en los receptores específicos donde comienza.
La localización espacial de un estímulo se indica mediante la activación de una población determinada de neuronas sensitivas, cuyos campos receptores se afectan por el estímulo.
La intensidad del estímulo puede codificarse de varias formas. Dados que los potenciales de acción tienen una magnitud uniforme, algunas neuronas sensitivas codifican la intensidad a través de la frecuencia de descarga, o con el número de receptores sensitivos que se activan. Las neuronas sensitivas centrales que reciben los estímulos de una clase concreta de receptor sensitivo se activan de forma más potente cuantas más neuronas aferentes primarias descarguen. Los estímulos de distinta intensidad pueden activar distintos conjuntos de receptores sensitivos.
La frecuencia del estímulo se codifica en ocasiones por potenciales de acción. Una frecuencia de descarga no permite indicar de forma inequívoca la frecuencia y la intensidad en el mismo sistema.
La duración de estímulo puede codificarse en las neuronas sensitivas de adaptación lenta mediante la duración de disparos potenciados. El principio y el final de un estímulo se pueden codificar por descargas transitorias de los receptores sensitivos de adaptación rápida.

ANEXO
Estímulo Umbral: Estímulo que por su intensidad, alcanza el voltaje umbral para que se produzca
un potencial de acción.
Estímulo Subumbral: Estímulo que no alcanza el voltaje umbral de excitación. Produce un potencial
electrotónico.
Estímulo Supraumbral: Estímulo que supera el voltaje umbral de una célula excitable. A medida
que aumenta su valor, aumenta la intensidad del potencial de acción, hasta llegar a un límite, dado
por el estímulo supramáximo.
Estímulo Supramáximo: Estímulo muy superior al estímulo umbral, en el cual la intensidad de la
respuesta no varía ya que se alcanzó la saturación de los canales iónicos.
Potencial electrotónico: Variación de diferencias de potencial que se producen entre el interior y el
exterior de la membrana plasmática, determinadas por un estímulo subumbral. Algunas de sus
características son:
• Son locales y no se propagan.
• Son pasivos: Esto quiere decir que no cambia la polaridad de la membrana.
• Son sumables en el tiempo y en el espacio.
• Su amplitud varía, decrece en el tiempo y espacio, a diferencia de un potencial de acción en
una celula nerviosa en el cual la amplitud es constante.
• Puede ser hiperpolarizante o despolarizante. En caso de ser hiperpolarizante nunca se
llegará a dar un potencial de acción; en caso de ser despolarizante, si la sumatoria espacial
y temporal supera el potencial umbral se dará el potencial de acción.
•
Sumación Espacial: Se produce cuando dos señales aferentes llegan casi simultáneamente, por lo
que los dos potenciales post-sinápticos se suman, de forma que las dos señales aferentes
excitadoras simultáneas despolarizarán la célula post-sináptica aproximadamente al doble de lo
que sucedería con cada una de ellas por separado
Sumación Temporal: Tiene lugar cuando se producen dos o más potenciales de acción en una
única neurona pre-sináptica y en una sucesión rápida, de forma que los potenciales post-sinápticos
resultantes se superponen en el tiempo.
Reclutamiento de unidades motoras: El Sistema Nervioso para controlar el grado de contracción
muscular no puede modificar el potencia de acción, puesto que éste obedece a la ley del “todo o
nada”, por lo que presenta la misma amplitud y forma. Para ello regula la cantidad de unidades
motoras reclutadas según el movimiento que se desea ejecutar. Por ejemplo, para movimientos
que requieren de mucho esfuerzo, se recluta una mayor cantidad de unidades motoras,
aumentando, por ende, el número de fibras musculares que se contrae.
Tetania: Hiperexcitabilidad del sistema nervioso central o periférico. Se manifiesta por
contracciones musculares involuntarias a distintos niveles (laringoespasmo, espasmo carpopedeal,
convulsiones). La causa fundamental de esta patología radica en desequilibrio de iones calcio
(hipocalcemia), hidrógeno o magnesio, para surtir sus efectos a nivel de la placa motora
fundamentalmente.

Sacudidas: Son provocadas por las fasciculaciones o sacudidas musculares producto de irritación
de los cuerpos neuronales o de sus prolongaciones, detectadas en EM. Pueden aparecer
aisladamente sin trascendencia o bien por alteraciones metabólicas, por patologías radiculares,
nerviosas y enfermedades neuromusculares.
Constante de longitud: Se define como [ rm/rint ]1/2
donde rm es la resistencia de la membrana
plasmática y rint,, es la del citoplasma. Si la relación rm/rint es alta, menor es la corriente que se
pierde a lo largo de la membrana, funcionando así la célula “como un cable”, y recorriendo así, una
mayor distancia, la señal electrotónica sin una reducción apreciable de conducción y velocidad.