







Languages
Pages
Legal


Evolutionary physiologytopics
1. Patterns
2. Processes

1. Patterns
• How and why of particular transitions
How and why did endothermic vertebrates evolve from ectothermic ancestors?
Evolutionary physiologytopics

Endothermy versus ectothermy
scala naturae

Endothermy versus ectothermy

Endothermy versus ectothermy
Advantages of endothermy:
• Stenothermy
• Aerobic metabolism
• Independent of environment

Endothermy versus ectothermy
Advantages of ectothermy:
• Low energetic requirements

0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
mammals
Passerine birds
reptiles
metabolism (Wg-1day-1)
0.1g 10g 1kg 100kg 1000kg

Endothermy versus ectothermy
Advantages of ectothermy:
• Low energy requirements
Low food habitats




Endothermy versus ectothermy
Advantages of ectothermy:
• Low energy requirements
Low food habitats Fluctuating food habitats

Mt Chappell Island
Flinders Island
Cape Barren Island

Chappell Island tiger snake(Notechis ater serventyi)
Short-tailed shearwater(Puffinus tenuirostris)

Gila monster (Heloderma suspectum)

Western banded gecko (Coleonyx variegatus)

Endothermy versus ectothermy
Advantages of ectothermy:
• Low energy requirements
Low food habitats Fluctuating food habitats Small body dimensions
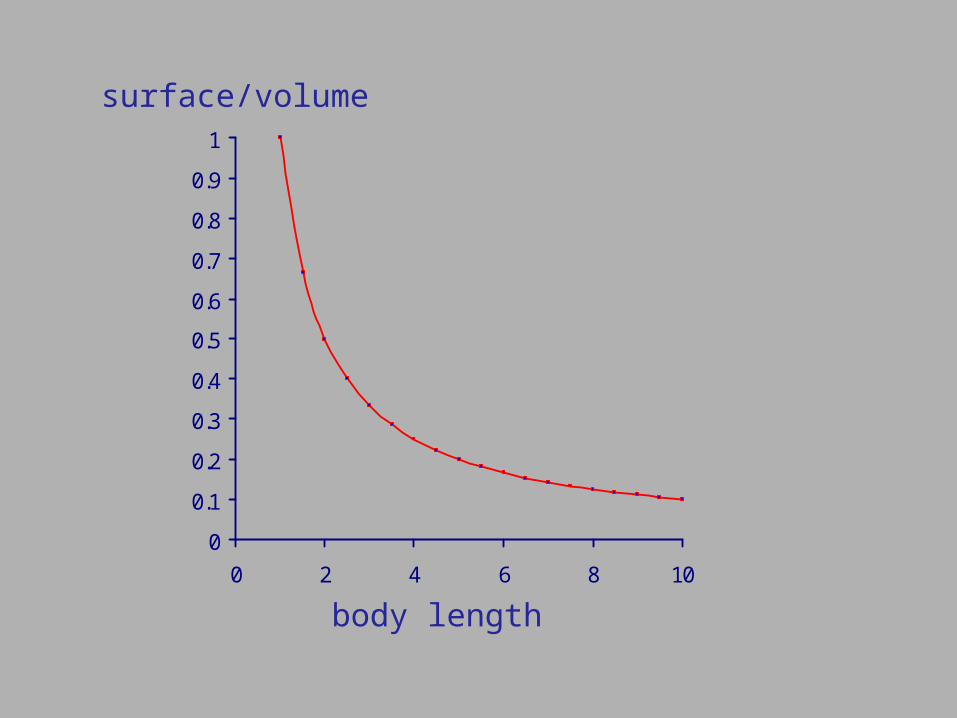
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
0 2 4 6 8 10
body length
surface/volume

mammals: >20 g
lizards: 8% spp. < 1 g 80% spp. < 20 g
salamanders: 20% spp. < 1 g90% spp. < 20 g

Dwarf chameleon
Monte Iberia EleuthDwarf gecko

Kitti’s hog-nosed bat
Etruscan shrewW: 1.5-2.5 gFR: 4xW/dayHR: 835 b/minRR: 661 p/min
L: 29-33 mm

Endothermy versus ectothermy
Advantages of ectothermy:
• Low energy requirements
Low food habitats Fluctuating food habitats Small body dimensions Elongate body forms
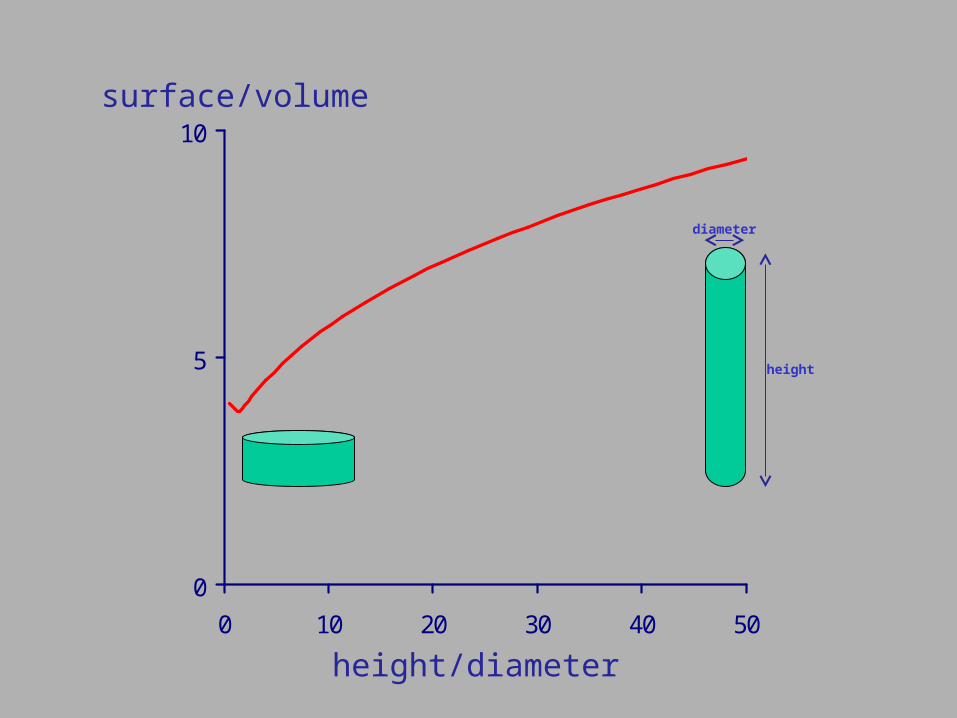
0
5
10
0 10 20 30 40 50
height/diameter
surface/volume
diameter
height

weasel (Mustela nivalis) wood rat (Neotoma sp.)
energy loss: x2

Afrocaecilia taitana Desmognathus ochrophaeus
Bipes bipes Anguis fragilis

Opheodrys aestivus

Endothermy versus ectothermy
Advantages of ectothermy:
• Low energy requirements
Low food habitats Fluctuating food habitats Small body dimensions Elongate body forms Low water habitats

Sauromalus obesus
Scaphiopus couchii

Endothermy versus ectothermy
Advantages of ectothermy:
• Low energy requirements
Low food habitats Fluctuating food habitats Small body dimensions Elongate body forms Low water habitats Low oxygen habitats

Amblyrhynchus cristatus
Iguana iguana

Neoseps reynoldsi

Scincus mitranus

Dilong paradoxus Xu et al. 2004. Nature 431: 680-684.

Dimetrodon (Pelycosauria)
Moschops (Therapsida)
Synapsida (mammal-like reptiles)

Endothermy in Mammalia:
1.RM x5
2.Tb > Ta, 28°C < Tb < 40°C
3. Tcore < 1-2°C
4.Maero x5

• Thermoregulation first physiological version
Synapsida evolve from small ectotherms
increase in size(30-100 kg)
become inertial homeotherms
evolve insulation
Tb constant,physiological benefits
decrease in size
increased metabolismimproved insulation
McNab 1978. Am. Nat. 112: 1-21.

• Thermoregulation first brain version
Synapsida evolvefrom small ectotherms
increase in sizeincrease in size(30-100 kg)(30-100 kg)
become inertial become inertial homeotherms homeotherms
evolve evolve insulationinsulation
Tb constant,physiological benefits
evolve larger, morecomplex brains
Hulbert 1980.

• Thermoregulation first ecological version
Synapsida evolvefrom small ectotherms
increase in sizeincrease in size(30-100 kg)(30-100 kg)
become inertial become inertial homeothermshomeotherms
evolve evolve insulationinsulation
Tb constant,physiological advantages
evolve nocturnalhabits
Crompton et al. 1978. Nature 272: 333-336.

• Aerobic capacity first sustained ativity version
Ruben 1995 Ann. Rev. Physiol. 57: 69-95.
small change inbasal metabolic rate
minimal effect on thermoregulatory capacity
large effect onmaximal aerobic metabolic rate

• Aerobic capacity first parental care version
Koteja 2000 Proc. R. Soc. Lond. 267: 479-484
small change in basal metabolic rate
minieme verandering inthermoregulatie-capaciteit
large effect onmaximal aerobic metabolic rate
necessary for locomotor costs related to parental care

1. Patterns
• How and why of particular transitions• Testing a-priori-hypotheses
plastic responses are adaptive
Evolutionary physiologytopics

Dicerandra linearifolia
Winn A.A. 1999. J. Evol. Biol. 12: 306-313.
• leaf length• leaf thickness• density of stomata

winter summer
Leaf
leng
th (
mm
)
5
10
15
20
25
30
35
winter summer
Lea
f thi
ckne
ss (
mm
)
0.140
0.145
0.150
0.155
0.160
0.165
winter summer
Den
sity
of s
tom
ata
(m
m-2
)
76
78
80
82
84
86
88
90
92
94
96
Winn A.A. 1999. J. Evol. Biol. 12: 306-313.

Winn A.A. 1999. J. Evol. Biol. 12: 306-313.
winter summer
Se
lect
ion
gra
dië
nt f
or
lea
ve le
ng
th
0.30
0.35
0.40
0.45
0.50
0.55
0.60
0.65
winter summer
Se
lect
ion
gra
dië
nt f
or
leav
e th
ickn
ess
-0.04
-0.02
0.00
0.02
0.04
0.06
0.08
winter summer
Se
lect
ion
gra
dië
nt f
or
stom
ata
de
nsity
-0.06
-0.05
-0.04
-0.03
-0.02
-0.01

Beneficial acclimation hypothesis

Beneficial acclimation hypothesis
Colder is better Hotter is better

Beneficial acclimation hypothesis
Deleterious acclimation hypothesis
Beneficial acclimation hypothesis
Escherichia coli
Leroi et al. 1994.Proc. Natl. Acad. Sci. USA 91: 1917-1921.
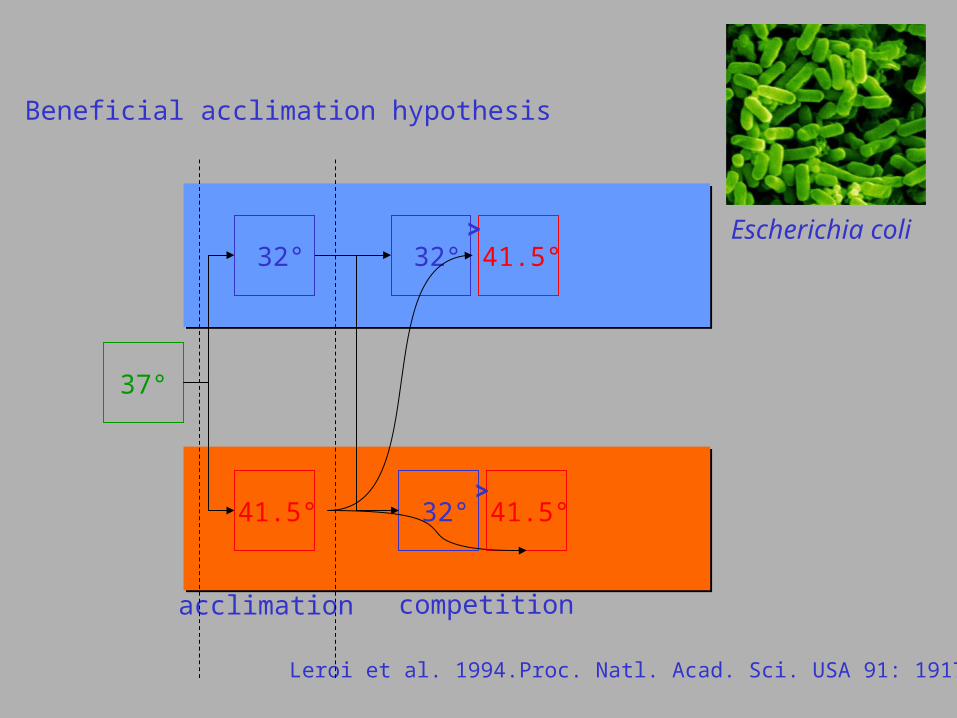
Beneficial acclimation hypothesis
Escherichia coli
37°
32°
32°
competition
41.5°
41.5°
>
>
Leroi et al. 1994.Proc. Natl. Acad. Sci. USA 91: 1917-1921.
32°
41.5°
acclimation

Beneficial acclimation hypothesis
Bicyclus anynana Geister T.L. & Fischer 2007. Behav. Ecol. 18: 658-664.
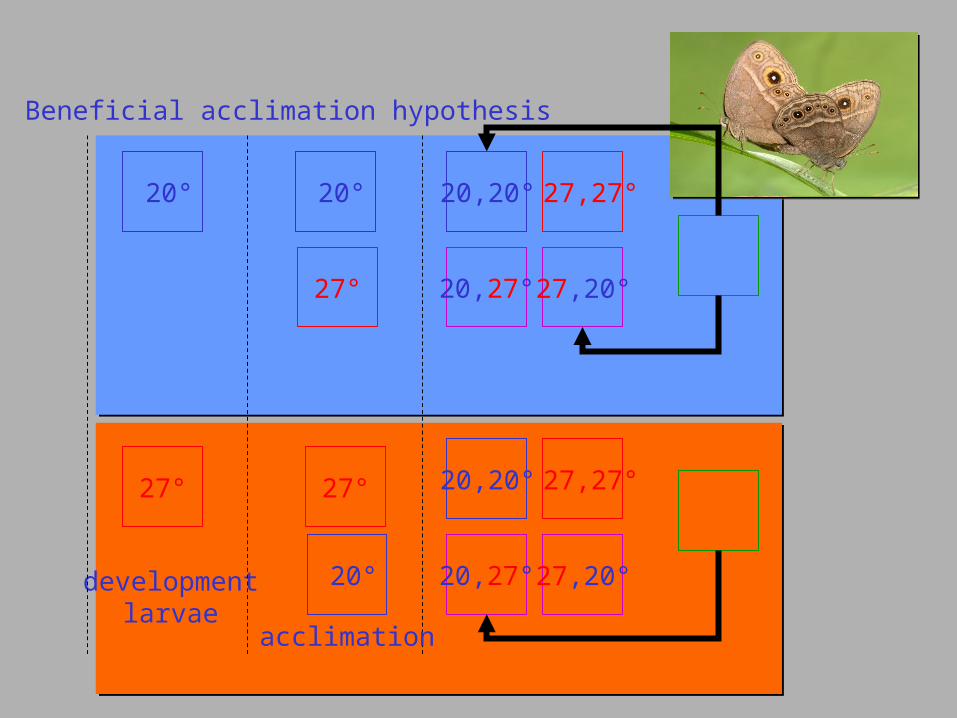
Beneficial acclimation hypothesis
20°
27°
developmentlarvae
20,20°
20,27°
27,27°
27,20°
20,20°
20,27°
27,27°
27,20°
20°
27°
27°
20°
acclimation

Beneficial acclimation hypothesis
Oribatida Deere J.A. & Chown S.L. 2006. Am. Nat. 168: 630-644.

Marion Island,Prince Edward Islands


Beneficial acclimation hypothesis
Deere J.A. & Chown S.L. 2006. Am. Nat. 168: 630-644.
10°
acclimation7 days
5°
0°
15°
Halozetes marinus
Halozetes marionensis
Halozetes belgicae
Halozetes fulvus
Podacarus auberti
Locomotor tests -5° up to 35°

Beneficial acclimation hypothesis
Deere J.A. & Chown S.L. 2006. Am. Nat. 168: 630-644.
Halozetes marinus
Halozetes marionensis
Halozetes belgicae
Halozetes fulvus
Podacarus auberti
deleterious acclimation

Beneficial acclimation hypothesis
Deere J.A. & Chown S.L. 2006. Am. Nat. 168: 630-644.
15°C10°C5°C0°C

Beneficial acclimation hypothesis
Deere J.A. & Chown S.L. 2006. Am. Nat. 168: 630-644.
Halozetes marinus
Halozetes marionensis
Halozetes belgicae
Halozetes fulvus
Podacarus auberti
deleterious acclimation
beneficial acclimation
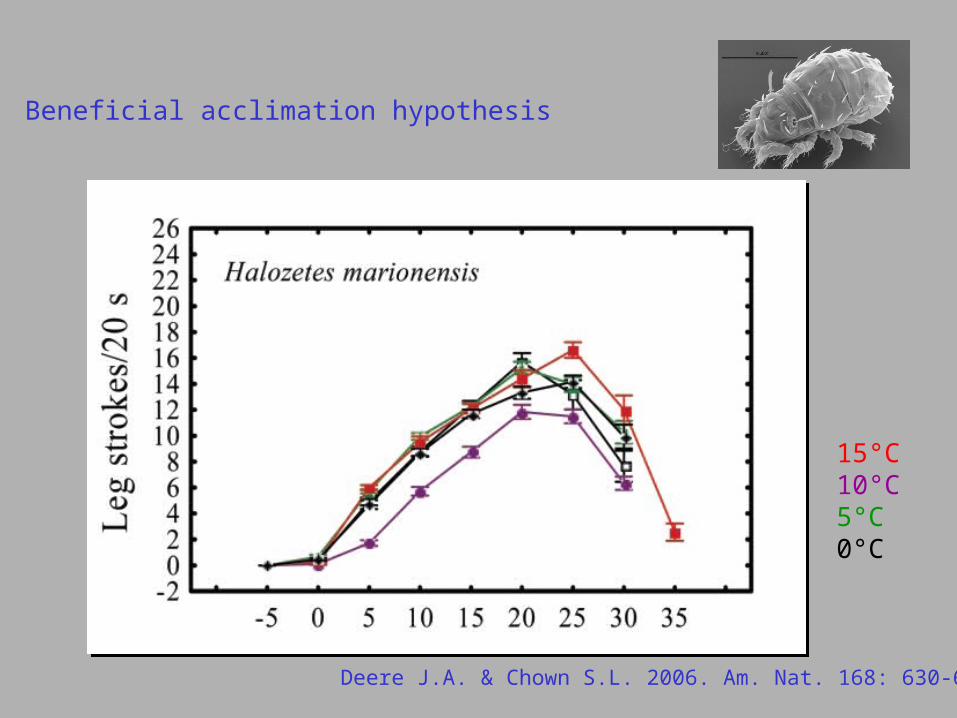
Beneficial acclimation hypothesis
Deere J.A. & Chown S.L. 2006. Am. Nat. 168: 630-644.
15°C10°C5°C0°C

Beneficial acclimation hypothesis
Deere J.A. & Chown S.L. 2006. Am. Nat. 168: 630-644.
Halozetes marinus
Halozetes marionensis
Halozetes belgicae
Halozetes fulvus
Podacarus auberti
colder is better
deleterious acclimation
beneficial acclimation

Beneficial acclimation hypothesis
Deere J.A. & Chown S.L. 2006. Am. Nat. 168: 630-644.
15°C10°C5°C0°C

Beneficial acclimation hypothesis
Deere J.A. & Chown S.L. 2006. Am. Nat. 168: 630-644.
Halozetes marinus
Halozetes marionensis
Halozetes belgicae
Halozetes fulvus
Podacarus auberti geen plasticiteit
geen plasticiteit
colder is better
deleterious acclimation
beneficial acclimation

Beneficial acclimation hypothesis
Deere J.A. & Chown S.L. 2006. Am. Nat. 168: 630-644.
15°C10°C5°C0°C

1. Patterns
• How and why of particular transitions• Testing a-priori-hypotheses
plastic responses are adaptive phenotypic plasticity ~ environmental variability
Evolutionary physiologytopics

Rana temporaria Lind & Johansson 2006. J. Evol. Biol. 20: 1288-1297

• 14 small islands• 10 clutches < 20-50 eggs• depth pools• variability drying / island• lab: 4 tadpoles / container• 2 regimes: Constant & Drying
• developmental time ~ regime (D<C)• developmental time ~ island• phenotypic plasticity ~ variability island
Lind & Johansson 2006. J. Evol. Biol. 20: 1288-1297

constant
drying
developmental time
28
17
island 1(homo)
plasticity=11
28
10
island 2(hetero)
plasticity=18
• devolopmental time ~ regime (D<C)• developmental time ~ island• phenotypic plasticity ~ variability island

Lind & Johansson 2006. J. Evol. Biol. 20: 1288-1297

1. Patterns
• How and why of particular transitions• Testing a-priori-hypotheses
plastic responses are adaptive phenotypic plasticity ~ environmental variability a jack-of-all-trades is a master of none
Evolutionary physiologytopics

0
5
10
15
20
25
30
35
40
6 8 10 14 18 22 26 30 34 38
sprint speed‘specialist’
‘generalist’
lichaamstemperatuur
sprint speed
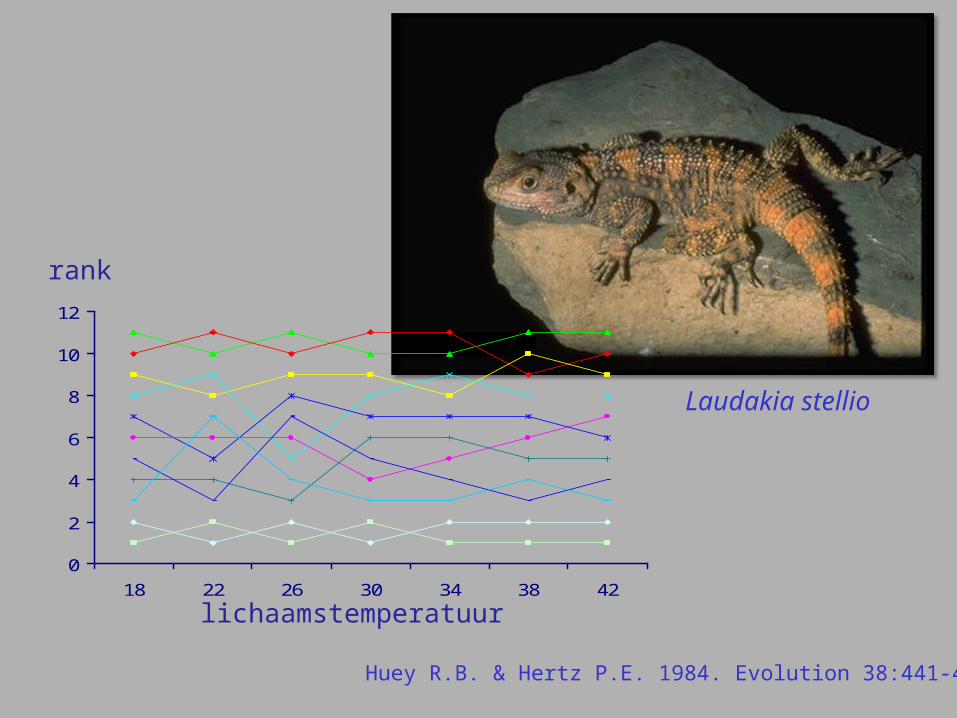
0
2
4
6
8
10
12
18 22 26 30 34 38 42
Laudakia stellio
lichaamstemperatuur
rank
Huey R.B. & Hertz P.E. 1984. Evolution 38:441-444.

Huey R.B. & Hertz P.E. 1984. Evolution 38:441-444.
Amoeba
0
1
2
3
4
5
10 15 20 25 30 38lichaamstemperatuur
rank

Escherichia coli
Hughes et al. 2007. Physiol. Biochem. Zool. 80: 406-421.

Escherichia coli
Hughes et al. 2007. Physiol. Biochem. Zool. 80: 406-421.
5.3
6.3
7.0
7.8
2000 generations
non-active
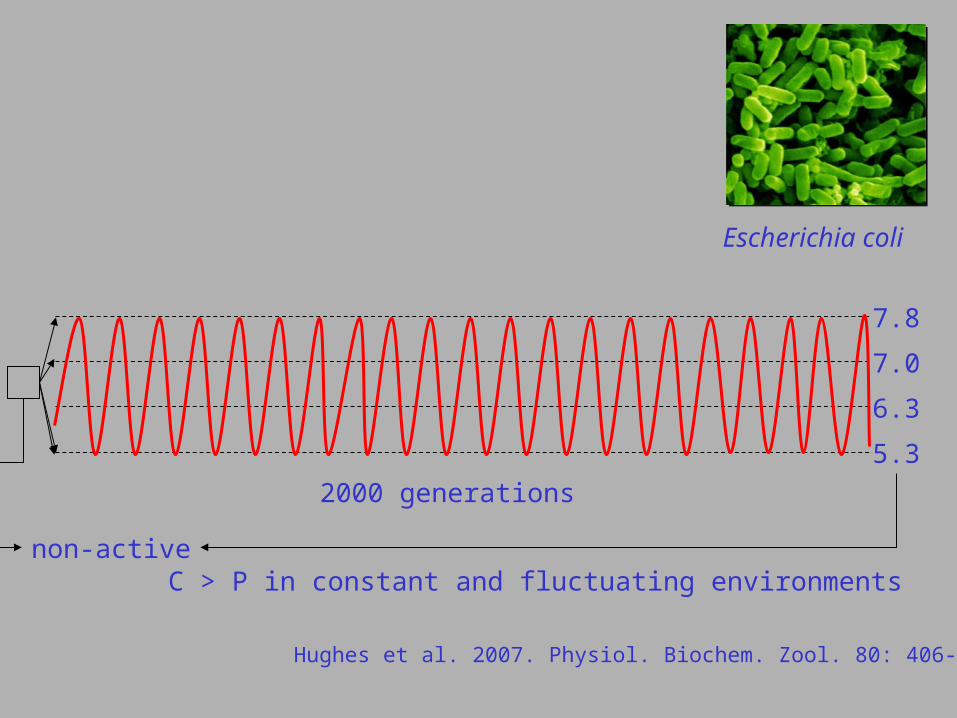
Escherichia coli
Hughes et al. 2007. Physiol. Biochem. Zool. 80: 406-421.
5.3
6.3
7.0
7.8
2000 generations
non-activeC > P in constant and fluctuating environments

Escherichia coli
Hughes et al. 2007. Physiol. Biochem. Zool. 80: 406-421.
5.3
6.3
7.0
7.8
2000 generations
non-activeR > P in some fluctuating and constant environments

Escherichia coli
Hughes et al. 2007. Physiol. Biochem. Zool. 80: 406-421.
5.3
6.3
7.0
7.8
2000 generations
non-activeB > P in fluctuating environments, but not in 7.8
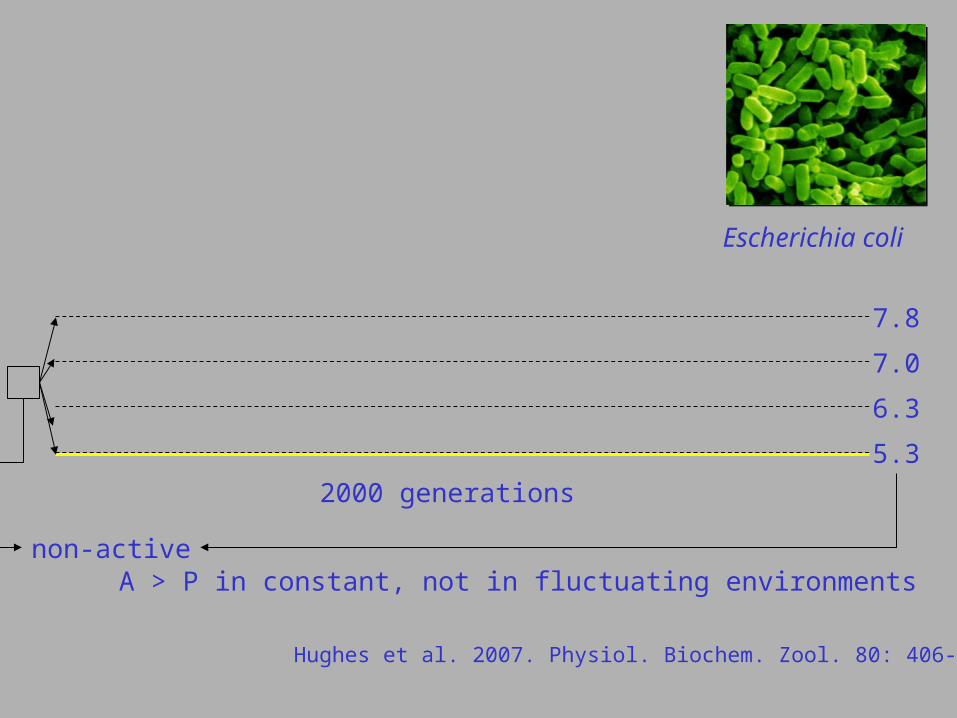
Escherichia coli
Hughes et al. 2007. Physiol. Biochem. Zool. 80: 406-421.
5.3
6.3
7.0
7.8
2000 generations
non-activeA > P in constant, not in fluctuating environments

Escherichia coli
Hughes et al. 2007. Physiol. Biochem. Zool. 80: 406-421.
(1) adaptation to cycling pH, randomly changing pH and constante pH follows different patterns
(2) in variable environments generalists evolve, in constant environments specialists evolve;
(3) in variable environments the ‘cycling’ lines have a higher fitness than the ‘random changes’ lines;
(4) an acclimation benefit (BAH) was not always detected.

Goodman et al. 2007. Evol. Ecol. Res. 9: 527-546.
• 18 Lygosominae• sprinting, jumping, clinging, climbing

1. Patterns
• How and why of particular transitions• Testing a-priori-hypotheses
plastic responses are adaptive phenotypic plasticity ~ environmental variability a jack-of-all-traits is a master of none symmorphosis: design satisfies need
Evolutionary physiologytopics

Evolutionary physiologytopics

Evolutionary physiologytopics
king pin

one half rule

V02max
mitochondriain muscle cells
capillary design(volume, surface)
hematocrite
heartstroke volume
surface pulmonary vesiclesdiffusion capacity membrane
Weibel et al. 1991. Proc.Natl. Acad. Sci. USA 88: 10357-10361

1. Patterns
2. Processes• natural selection
Evolutionary physiologytopics
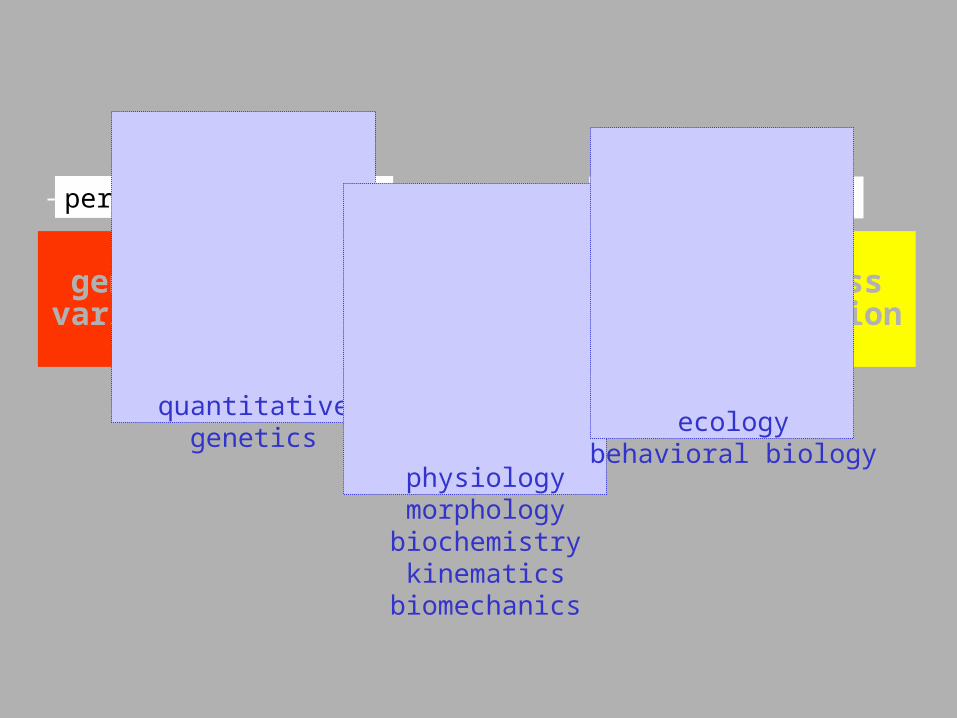
performancevariation
fitnessvariation
design variation
geneticvariation ???? ??
performance gradient fitness gradient
quantitativegenetics
physiologymorphologybiochemistrykinematics
biomechanics
ecologybehavioral biology

LeGalliard et al. 2004. Nature 432: 502-505.
Zootoca vivipara
juvenile survival
initial endurance
limited food supply
abundant food supply

1. Patterns
2. Processes• natural selection• sexual selection
• intrasexual selection (male-male combat)• intersexual selection (female choice)
Evolutionary physiologytopics

deCarvalho et al. 2004. Anim. Behav. 68: 473-482.
Neriene litigiosa
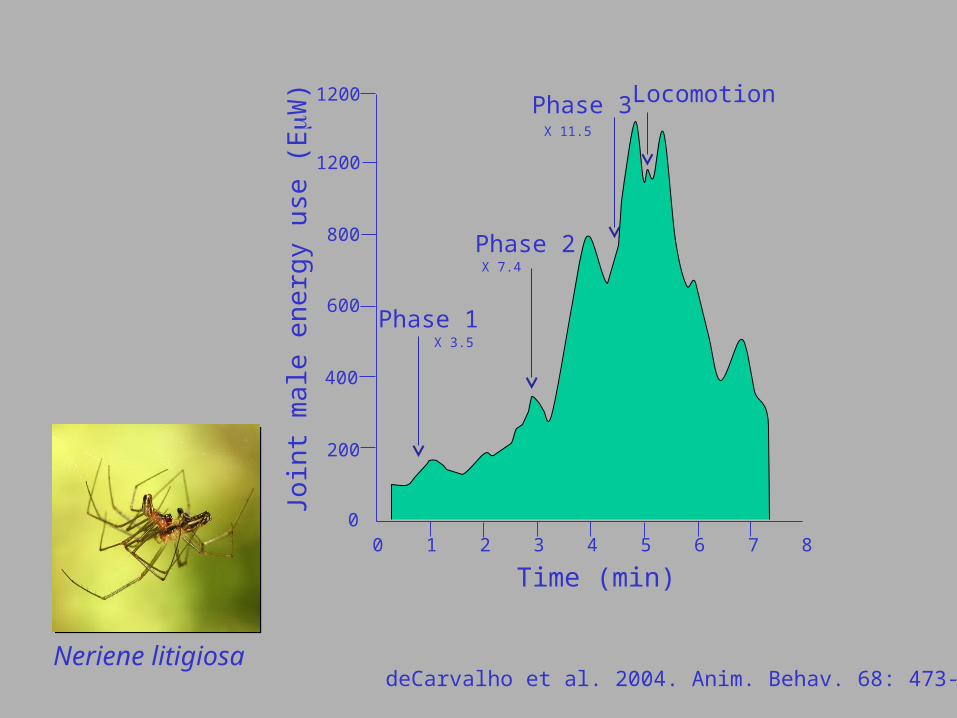
deCarvalho et al. 2004. Anim. Behav. 68: 473-482.Neriene litigiosa
Time (min)
Join
t m
ale
en
erg
y u
se (
EW
)
1200
1200
800
600
400
200
00 1 2 3 4 5 6 7 8
Phase 1
Phase 2
Phase 3 Locomotion
X 3.5
X 7.4
X 11.5

Necora puber Uca lactea
Thorpe et al. 1995. Anim. Behav. 50: 1657-1666
Matsuma & Murai 1995. Anim. Behav. 69: 569-577
anaerobic respiration

Agkistrodon contortix
Schuett & Grober 2000 Physiol & Behav 71: 335-341.

Agkistrodon contortix
Schuett & Grober 2000 Physiol & Behav 71: 335-341.

Anolis sagrei


Evolutionary physiologyimplications

Evolutionary physiologyimplications

Evolutionary physiologyimplications

Evolutionary physiologyimplications
http://www.sfecologie.org/blog/2011/09/30/evolrescueonline-topic-1/
Top Related