







Languages
Pages
Legal


Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras
receptoras de embriones bovinos
Néstor Isaías Tovío Luna
Universidad Nacional de Colombia Facultad de Medicina Veterinaria y de Zootecnia
Maestria en Salud y Producción Animal Bogotá., D.C. Colombia
2011


Efectos de la aplicación de eCG (Día 5 u 8)
sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras
receptoras de embriones bovinos
Néstor Isaías Tovío Luna
Trabajo de investigación presentado como requisito parcial para optar al título de Magister en Ciencias – Salud y Producción Animal
Director: PhD., MSc. Henry Alberto Grajales Lombana
Línea de Investigación: Fisiología de la Reproducción
Grupo de investigación: Biología de la Adaptación de los Animales al Trópico
Universidad Nacional de Colombia Facultad de Medicina Veterinaria y de Zootecnia
Maestria en Salud y Producción Animal Bogotá., D.C. Colombia
2011


Especialmente a DIOS, quien por intermedio
de la VIRGEN MARÍA consagra toda su
atención en mi existencia.
A mis padres, EMMA CECILIA LUNA e ISAÍAS
RAFAEL TOVÍO ESPÍTIA, quienes con mucho
esfuerzo me han formado.
A mi esposa SANDRA REYES y mi hijo
SERGIO IVÁN TOVÍO REYES, los cuales son
el sosten de mi vida.
A mis hermanos ORLANDO, RAFAEL,
HECTOR, quienes siempre están pendientes
de mi.
A mis sobrinos CAMILO ANDRES, CARLOS
EDURDO, EMMANUEL y SARITA, quienes
son la alegría de la familia


Agradecimientos
Al Doctor HENRY ALBERTO GRAJALES LOMBANA, profesor asociado de la
Facultad de Medicina Veterinaria y de Zootecnia, de la Universidad Nacional de
Colombia, quien como tutor fue guía darante la formación en mi posgrado.
Al Doctor IVÁN MAURICIO GONZALEZ, director de CGR Biotecnología
Reproductiva, quien me apoyó en la parte práctica del proyecto.
Al Doctor JUAN JOSÉ MOLINA, director técnico de Schering- Plough Colombia,
quien gracias a sus sugerencias se logró concretar el proyecto de investigación


Resumen y Abstract IX
Resumen Se evalúa el efecto de la aplicación al día 5 u 8, de 400UI de Gonadotropina Coriónica
Equina (eCG), en un protocolo de sincronización para transferencia de embriones, sobre
desarrollo folicular (día 9), desarrollo luteal (día 17), concentración de progesterona “P4”
(días 9 y 17) y porcentaje de preñez (día 52), en 70 novillas raza Holsteín. Se analizan
independientemente a los tratamientos de eCG, las relaciones entre diámetro del folículo
dominante (DFD) “día 9” y el volumen luteal (VL) “día 17”, el VL (día 17) y concentración
de P4 (día 17). Se relacionan los tratamientos de eCG, el DFD (día 9), el VL (día 17) y
concentración de P4 (día 17) frente al procentaje de preñez. No hubo efecto (P>0.05) del
día de aplicación de eCG sobre número de FD, pero si (P<0,05) en cuanto a DFD. No
hubo efecto (P>0.05) del día de aplicación de eCG para número de cuerpos lúteos, VL,
concentración de P4, (días 9 y 17) y preñez. No hubo relación (P>0.05) entre el DFD y
VL; pero si existió (P<0,05) entre el VL y niveles de P4. Al analizar la relación entre los
tratamientos, el DFD, el VL y la concentración de P4 (día 17) frente a preñez, se observó
que solamente existió relación positiva (P<0,1) entre esta última (preñez) y la
concentración de P4. El día de aplicación de eCG (día 5 u 8) afecta solamente el DFD. La
preñez es afectada por la concentración de P4 la cual tiene relación con el VL.
Palabras clave: eCG, Cuerpo lúteo, Foliculo Dominante, Progesterona, Transferencia
de embriones.
Abstract The application effect of the 400 IU Equine Chorionic Gonadotrophin (eCG) was
evaluated through a synchronization protocol for embryo transfer on days 5 and 8 within a
population of 70 Holstein heifers. Measurements of follicle and corpus luteum
development were taken on days 9 and 17 respectively. Similarly progesterone P4
concentration was measured on days 9 and 17, as well as, the pregnancy rate on day 52.
Were analyzed independently of eCG treatments, the relationships between dominant
follicle diameter (DFD) "Day 9" and luteal volume (VL) "Day 17"; the VL (day 17) and
concentration of P4 (day 17). Relate eCG treatments, the DFD (day 9), VL (day 17) and

X Título de la tesis o trabajo de investigación
concentration of P4 (day 17) with the percentage of pregnancy. There was no effect (P>
0.05) the day of eCG on the number of FD, there were differences (P <0.05) in terms of
DFD. There was no effect (P> 0.05) the day of eCG for number of corpus luteum, VL, P4
concentration (9 and 17) and pregnancy. There was no relationship (P> 0.05) between
the DFD and VL, but there was (P <0.05) between the VL and P4 levels. Analyzing the
relationship between treatments, DFD, VL and the concentration of P4 against pregnancy,
it was noted that only existed positive correlation (P<0.1) between the pregnancy and
concentration of P4. The day of eCG administration (Day 5 or 8) affects only the DFD. The
pregnancy is affected by the concentration of P4 which is related to the VL.
Key words: eCG, Corpus luteum, Dominant Follicles, Progesterone, Embryo Transfer

Contenido XI
Contenido
Pág.
Resumen ...............................................................................................................................IX
Abstract .................................................................................................................................IX
Lista de figuras ................................................................................................................... XIV
Lista de tablas ..................................................................................................................... XV
Lista de graficas ................................................................................................................. XVI
Lista de esquemas ............................................................................................................ XVII
Introducción............................................................................................................................ 1
Bibliografía ............................................................................................................................. 6
1. Revisión de literatura ....................................................................................................... 11
1.1. Algunos aspectos relacionados al transporte y regulación hormonal ......................... 11
1.1.1. Transporte hormonal ................................................................................................. 11
1.1.2. Regulación hormonal ................................................................................................. 12
1.2. Mecanismo de acción hormonal ................................................................................... 12
1.2.1. Receptores de membrana ........................................................................................ 14
1.2.1.1. Receptores que provocan cambios de permeabilidad de la membrana .............. 14
1.2.1.2. Receptores acoplados a una proteína G .............................................................. 15
1.2.1.3. Receptores catalíticos ............................................................................................ 15
1.2.1.4. Mecanismos del segundo mensajero para comunicar funciones hormonales
intracelulares ........................................................................................................................ 15
1.2.1.4.1. Mecanismo del sistema segundo mensajero cAMP ........................................... 16
1.2.1.4.2. Lípidos de membrana como segundos mensajeros ........................................... 17
1.2.1.4.3. Iones de calcio y la calmodulina comosistema de segundo mesajero............... 17
1.2.2. Receptores intracelulares .......................................................................................... 18
1.3. Aspectos sobre dinamica folicular y hormonal ............................................................. 19
1.3.1. Factores que controlan el desarrollo folicular ........................................................... 25
1.3.1.1. Factores endocrinos ............................................................................................... 25
1.3.1.2. Factores exógenos ................................................................................................. 29
1.4. Aspectos relacionados al ciclo estral de la vaca.......................................................... 31

XII Título de la tesis o trabajo de investigación
1.4.1. Implicacion de las hormonas esteroides durante el ciclo estral ............................... 34
1.4.1.1. Vías implicadas en la síntesis de estrógenos ........................................................ 36
1.4.1.2. Vías implicadas en la síntesis de progesterona .................................................... 37
1.4.2. Fases del ciclo estral ................................................................................................. 40
1.5. Factores relacionados con el desarrollo embrionario .................................................. 50
1.6. Aspectos relacionados con el reconocimiento materno de la preñez ......................... 54
1.6.1. Señal luteotrópica ...................................................................................................... 56
1.6.2. Señal luteolítica ......................................................................................................... 57
1.7. Control del ciclo estral en programas de transplante embrionario .............................. 60
1.7.1 Estrategias antiluteolíticas en programas de transplante embrionario en bovinos ... 64
Bibliografía ........................................................................................................................... 68
2. Efectos de la aplicación de ecg (día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel
de progesterona y tasa de preñez en hembras receptoras de embriones bovinos .......... 89
Resumen .............................................................................................................................. 89
Introducción.......................................................................................................................... 90
2.1. Hipotesis ....................................................................................................................... 94
2.2. Objetivo ......................................................................................................................... 95
2.3. Objetivos específicos .................................................................................................... 95
2.4. Materiales y métodos.................................................................................................... 95
2.4.1. Localización ............................................................................................................... 96
2.4.2. Grupo experimental ................................................................................................... 96
2.4.2.1. Descripción del grupo de novillas evaluado ........................................................... 96
2.4.3. Condiciones de manejo ............................................................................................. 97
2.4.4. Tratamientos aplicados para control de ciclo estral .................................................. 97
2.4.4.1. Tratamiento 1 (Grupo eCG día 5) .......................................................................... 97
2.4.4.2. Tratamiento 2 (Grupo eCG día 8) .......................................................................... 99
2.4.5. Toma de muestras y ecografía ................................................................................ 102
2.4.6. Medición de variables .............................................................................................. 103
2.4.7. Análisis estadístico .................................................................................................. 105
2.5. Resultados y discusión ............................................................................................... 110
2.5.1. Efecto del día de la aplicación (día 5 u 8) de la eCG sobre el desarrollo del folículo
dominante (número y tamaño) .......................................................................................... 110
2.5.1.1. Número de folículos dominantes (día 9) .............................................................. 111
2.5.1.2. Diámetro folículo dominante (día 9) ..................................................................... 114

Contenido XIII
2.5.2. Efecto del día de la aplicación de la eCG sobre el desarrollo del cuerpo lúteo
(número y volumen), porcentaje de doble ovulación, porcentaje de utilización,
concentración plasmática de P4 y porcentaje de preñez .................................................. 117
2.5.2.1. Cuerpo lúteo (número) día 17 .............................................................................. 118
2.5.2.1.1. Porcentaje de doble ovulación (día 17) ............................................................ 121
2.5.2.2. Volumen luteal (mm3) día 17 ................................................................................ 121
2.5.2.2.1. Efecto del diámetro del folículo dominante (mm) día 9 sobre el volumen luteal
(mm3) día 17 ...................................................................................................................... 121
2.5.2.2.2. Porcentaje de utilización día 17 ........................................................................ 122
2.5.2.3. Concentración de progesterona ng/ml ................................................................. 124
2.5.2.3.1. Concentración de progesterona día 9 ............................................................... 124
2.5.2.3.2. Concentración de progesterona día 17 ............................................................. 126
2.5.2.3.2.1.. Efecto del volumen luteal (mm3) día 17 sobre la concentración plasmática
de progesterona (ng/ml) día 17 ......................................................................................... 126
2.5.2.3.2.2. Concentraciones de progesterona durante el ciclo estral sincronizado ........ 129
2.5.2.4. Porcentaje de preñez ........................................................................................... 130
2.5.2.4.1. Concentraciones plasmáticas de progesterona durante los días 5, 9 y 17
según estado reproductivo (Preñadas y no preñadas) ..................................................... 137
2.5.2.4.2. Concentración de progesterona durante el muestreo 2 (Día 9) en las novillas
no preñadas ....................................................................................................................... 140
2.5.2.4.3. Efecto de los tratamientos (eCG día 5 u 8), diámetro folicular (día 9),
volumenluteal (día 17) y concentración plasmática de progesterona (día 17) sobre la
presentación de preñez (día 52)........................................................................................ 141
3. Conclusiones y recomendaciones ................................................................................. 145
3.1 Conclusiones……………………………………………………………………………… 145
3.2 Recomendaciones ....................................................................................................... 146
Bibliografía ......................................................................................................................... 147

Contenido XIV
Lista de figuras
Pág. Figura 1. Algunos genes implicados en la síntesis de progesterona y estradiol ............... 35
Figura 2. Biosíntesis de la progesterona ............................................................................ 39 Figura 3. Cascada luteolítica en la vaca............................................................................. 42

Contenido XV
Lista de tablas Pág.
Tabla 1. Edad, peso y condición corporal de la muestra total y por tratamientos ............. 97
Tabla 2. Folículos dominantes presentes muestreo día 9 ................................................ 111
Tabla 3. . Comportamiento de estructuras luteales y su volumen, concentración de
progesterona y porcentaje de preñez ................................................................................ 118
Tabla 4. Contrastes entre perfiles de progesterona - Razón de probabilidad de preñez 143

Contenido XVI
Lista de gráficas
Grafica 1. Relación estimada de la concentración de progesterona (ng/ml) en función del
volumen luteal (día 17 - muestreo 3) ................................................................................. 127 Grafica 2. Comportamiento general de la progesterona durante los tres (3) muestreos
realizados en la totalidad de las novillas ........................................................................... 129
Grafica 3. Comportamiento general de la progesterona según estado reproductivo ..... 137
Grafica 4. Concentración de progesterona novillas no preñadas (muestreo 2 - día 9) 140
Grafica 5. Función logística para el rango de valores de concentración de progesterona ...... 142

Introducción Innumerables factores intervienen en los resultados de los programas de transplante
embrionario (TE) en bovinos, siendo relevantes los que tienen relación con las hembras
receptoras (Revisado por Tovío et al., 2008); esto se evidencia en las bajos porcentajes
de eficiencia, ya que normalmente en los núcleos de receptoras tratadas con protocolos
tradicionales de sincronización hormonal de ciclo estral, solamente del 40 al 50 % de las
hembras clasifican para él TE, el otro 50 o 60% son descartadas por su baja o inexistente
respuesta a los tratamientos de sincronización (Nasser et al., 2004). Considerando un
porcentaje de concepción del 40% del total de animales aprovechados, se obtendrá
apenas 20 % de gestaciones al final de todo el proceso (Thibier, 2002; Bó, 2003).
Indudablemente existen numerosos factores endocrinológicos, celulares y moleculares
entre otros, que determinan el mantenimiento o la pérdida de la preñez en etapas
iniciales, pero aún no han sido completamente aclarados (Spencer et al., 2004).
Teniendo en cuenta lo anterior, es fundamental mejorar los elementos de
fundamentación científica que puedan aclarar los diferentes procesos que encaminan el
desarrollo, reconocimiento y mantenimiento de la preñez en los programas de TE (Bó,
2003). De acuerdo a esto último, se evidencia que las secreciones hormonales
específicas interactúan con determinados factores para desencadenar el desarrollo del
embrión; especialmente se destaca la progesterona (P4), hormona secretada
principalmente por el cuerpo lúteo (CL), el cual tiene una duración activa y prolongada,
rasgo característico de la preñez en los mamíferos (Revisado por Tovío et al., 2008).
La P4 actúa en el útero estimulando y manteniendo funciones necesarias para el
desarrollo embrionario temprano; esto con la finalidad de llevar a cabo la implantación,
placentación y el exitoso desarrollo fetal (Spencer et al., 2004). De acuerdo a esto, se
afirma que el control endocrino para la síntesis de proteínas uterinas durante el desarrollo
embrionario temprano, lo tiene principalmente la P4, la cual es la responsable de los
cambios cualitativos y cuantitativos en el medio ambiente uterino, controlando la síntesis
y secreción de un sin número de sustancias (entre 1000 a 5000) como son factores de
crecimiento, citoquinas, proteínas etc. (Hafez, 2000); según esto, se asume que

2
Introducción
deficiencias de P4, podrían causar que el endometrio llegue a ser poco eficiente en
cuanto a la nutrición histotrópica, la cual es la primer fuente disponible para el
crecimiento, mantenimiento y supervivencia del conceptus (Revisado por Tovío et al.,
2008).
Las deficiencias de P4 causan alteraciones de origen endocrino, que contribuyen a
aumentar los niveles basales de mortalidad embrionaria (ME), probablemente asociados
a una selección natural ineludible, la cual busca la eliminación de genotipos poco
competentes (Rodríguez, 2001; Gouveia et al., 2004).
Alrededor de 25 al 40% de pérdidas embrionarias se han podido detectar durante los
primeros días de gestación en hembras receptoras de embriones bovinos (Santos et al.,
2004); se observa que la mayoría de estas hembras retornan a celo en la fecha prevista,
a los 20 – 22 días, manifestando un ciclo sexual normal y completo (vacas repetidoras)
(Palma, 2008), por lo que se sugiere que la ME pudo originarse entre los días 7 y 17, es
decir, el periodo comprendido entre el TE y el reconocimiento materno de la preñez
(RMP) (Binelli et al., 2001).
En cuanto a las pérdidas de preñez que suceden entre el día 28 y 98 (cuando ya ha
ocurrido el RMP), se han calculado porcentajes que oscilan entre el 7 y 33 % (Silke et
al., 2002).
De acuerdo a lo anterior, se sugiere que durante el establecimiento de la preñez se
presenta un "periodo crítico" muy definido entre los días 15 a 17 (Binelli et al., 2001); se
podría pensar que la biología durante este periodo es multifactorial y compleja, en donde
el endometrio puede estar recibiendo una señal antiluteolítica poco adecuada, que no
causa el bloqueo de la producción de Prostaglandina F2α (PGF2α) endometrial, lo que
desencadena la lisis del CL (el mantenimiento de la preñez es dependiente de la
funcionalidad del CL) (Santos et al., 2004). La señal antiluteolítica es generada por el
embrión en las células mononucleadas del trofoblasto, las cuales secretan interferón tau
(IFN-τ), bloqueando de esta manera la síntesis de PGF2α producida a nivel endometrial
(Binelli et al., 2001; Revisado por Grajales y Tovío, 2009). Esto último sugiere que las

Introducción
3
pérdidas del embrión podrían estar ocurriendo por una señal débil o inadecuada por parte
de éste para la inhibición de la síntesis de prostaglandina uterina (Palma, 2008).
La naturaleza critica alrededor del periodo de reconocimiento, aposición y adhesión del
embrión al endometrio uterino durante la implantación, determina la necesidad de un
estricto sincronismo entre el embrión transferido y su receptora, enfatizando la
importancia tanto del medio uterino como de las señales del conceptus que dan lugar al
RMP (Binelli et al., 2001), señales que deben ser emitidas en el momento y
concentración precisa, de tal manera que se garantice el mantenimiento de la estructura
y funcionalidad del CL, que genere una continua producción de P4 para el mantenimiento
del ambiente embriotrófico que apoye el normal desarrollo del conceptus (Revisado por
Grajales et al., 2011).
La comunicación entre el conceptus y el útero, no siempre tiene éxito, lo que
desencadena grandes pérdidas embrionarias (Binelli et al., 2001). Por eso mismo se
plantean estrategias que buscan minimizar el efecto luteolítico el cual desencadena la
pérdida temprana del embrión, acontecimiento más fácilmente evidenciado en los
programas de transplante embrionario, ya que con éstos se asegura la presencia del
embrión vivo desde el día 6 a 8 momento en el cual éste es trasplantado (Baruselli et al.,
2005).
Conocida la influencia de la P4 en determinados eventos relacionados con el
mantenimiento de la preñez desde estadios tempranos y la influencia de la PGF2α para
causar luteólisis, se ha propuesto y desarrollado una serie de estrategias hormonales
con la finalidad de mantener la preñez. Estas se basan en hacer más eficiente la
capacidad secretora de P4 por parte del CL, y que esta secreción ocurra en el momento
oportuno, garantizando de esta manera un ambiente uterino adecuado para el desarrollo
del embrión transplantado a hembras receptoras bovinas; todo esto encaminado a
incrementar la tasa de preñez en los programas de TE (Thatcher et al., 2001).
La relación entre el porcentaje de preñez y la concentración plasmática de P4 de acuerdo
al tamaño del CL en receptoras de embriones bovinos Bos Indicus y Bos Taurus, ha sido
objeto de controversia en varios estudios realizados. Algunos investigadores han

4
Introducción
verificado la correlación positiva entre estas variables, encontrando que a mayor área del
CL mayor es la concentración de P4 plasmática y consecuentemente mayor es la tasa de
preñez en diferentes razas (Thatcher et al., 2001; Vasconcelos et al., 2001; Bó et al.,
Mantovani et al., 2002; Baruselli et al., 2005; Lopes et al., 2007). Con estos resultados se
podría evidenciar que los incrementos en la concentración de P4 durante el "periodo
crítico" (Binelli et al., 2001), estimulan la secreción de los agentes antiluteolíticos con lo
cual se hace eficiente el RMP (Vasconcelos et al., 2001). Igualmente en diferentes
reportes se observa que el aumento en las concentraciones plasmáticas de P4 durante el
diestro se correlaciona positivamente con la capacidad del embrión en secretar IFN-τ,
provocando así un aumento en las tasas de preñez (Thatcher et al., 2001; Mann et al.,
2009).
Según los estudios se podría sugerir que a mayor concentración de P4 plasmática mejor
será el ambiente uterino para el conceptus en vías de desarrollo (Figueira et al., 2011).
Esto se entiende si se tiene en cuenta que cualquier variación en la concentración de P4
es determinante en la modulación de la expresión y secreción de factores de
crecimiento, citoquinas y proteínas, que condicionan el medio uterino para los procesos
de receptividad endometrial y de viabilidad embrionaria (Revisado por Peña et al., 2007).
De acuerdo a lo anterior, se podría sugerir que al brindar fuentes directas o indirectas de
P4 a hembras durante los primeros días de preñez, el porcentaje de perdidas
embrionarias disminuye, ya que se mejora el ambiente uterino en donde el conceptus
tendrá un desarrollo adecuado evidenciando mejor la síntesis y secreción de IFN-τ, pues
esta secreción está influenciada por el estado de desarrollo embrionario (Kebler et al.,
1997; Siqueira et al., 2009; Revisado por Tovío et al., 2010).
Con la finalidad de aumentar las tasas de preñez se han utilizado tratamientos
hormonales en hembras receptoras de embriones bovinos, utilizado Gonadotropina
Corionica Equina (eCG) en protocolos de transplante de embriones a tiempo fijo (TETF)
(Baruselli et al., 2005).
La eCG (antes conocida como Gonadotropina Sérica de Yegua Preñada PMSG) es una
hormona glicoproteica con peso molecular de 45 kDa. y promedio de vida de 3 días,

Introducción
5
producida por los cálices endometriales en la yegua preñada entre los días 40 a 130
(Bousfield y Butnev, 2001). Se compone de dos subunidades α-y β (la subunidad α está
codificada por un gen común en todas las glicoproteínas, mientras que estas hormonas
difieren en el gen que codifica para subunidad β-, la cual confiere la especificidad
hormonal) (Palma, 2008; Forcada et al., 2011).
Esta hormona se vincula a los receptores foliculares de FSH y de LH, y a los receptores
de LH del CL, creando de esta forma condiciones de crecimiento folicular, ovulación y
luteinización (Murphy and Martinuk, 1991 Baruselli et al., 2003; Filho et al., 2004; Filho et
al., 2010); pero su acción predominante es de FSH, lo que daría a lugar a la formación de
cuerpos lúteos accesorios característicos de la yegua gestante (Fuentes y de la Fuente,
2007; Forcada et al., 2011).
La aplicación de eCG en el momento esperado de una nueva onda de crecimiento
folicular, ha demostrado eficiencia en cuanto a superovulación (de acuerdo a la
dosificación) y/o desarrollo de un folículo dominante de mayor diámetro, determinando de
esta forma un mayor número de cuerpos lúteos o un CL grande (Baruselli et al., 2005).
Esto va acompañado de mayores concentraciones plasmáticas de P4 y mejor porcentaje
de utilización (hembras transferidas/hembras sincronizadas*100), concepción y de
preñez frente a tratamientos sin aplicación de esta hormona tanto en ganado Bos Indicus
como Bos Taurus y sus cruces (Santos et al., 2001; Marques et al., 2003; Everton and
Baruselli, 2003; Madureira et al., 2004; Rodrigues et al., 2004; Baruselli et al., 2005;
Mamani et al., 2007; Pita et al., 2009; Sousa et al., 2009; Figueira et al., 2011).
Se ha verificado que con la aplicación de eCG el día 8 dentro de un protocolo de
sincronización en hembras receptoras (Bos Indicus/Bos Taurus) de embriones bovinos se
consiguen cuerpos lúteos únicos de mayor tamaño frente a la aplicación de la misma
hormona el día 5, pero esto no ha determinado diferencias entre la concentración
plasmática de progesterona producida, ni en los porcentajes de preñez obtenidos entre
tratamientos (Nasser et al., 2004). Igualmente en otros reportes al respecto se ha
verificado que el porcentaje de utilización (hembras transferidas/hembras
sincronizadas*100), ha mejorado cuando la aplicación de eCG se suministra hacia el día

6
Introducción
8 dentro de un protocolo para transplante de embrión a tiempo fijo a receptoras Bos
Indicus (Quezada y Ortiz 2007).
Bibliografia 1. Baruselli P, Bó G, Reis E, Marques M, Sá M. Introducción de la IATF en el manejo
reproductivo de rebaños de ganado de engorde en Brasil. Congreso Internacional de
Reproducción Bovina. Bogotá 2005.
2. Baruselli P, Marques O, Nasser LF, Reis EL and Bó G. Effect of eCG on pregnancy
rates of lactating zebu beef cows treated with CIDR–B devices for timed artificial
insemination. Theriogenology. 2003. 59: 214.
3. Binelli M, Thatcher W, Mattos R, Baruselli P. Antiluteolytic Strategies to Improve
Fertility in Cattle. Theriogenology 2001. 56: 1451–1463.
4. Bó GA. Sincronización de celos para programas de Inseminación Artificial y
Transferencia de embriones Bovinos. Instituto de Reproducción Animal de Córdoba
Argentina (IRAC) 2003.
5. Bó GA. y Caccia, M. Ultrasonografía reproductiva en el ganado bovino. Facultad de
Ciencias Agropecuarias. Universidad Católica de Córdoba Argentina. Instituto de
Reproducción Animal de Córdoba (IRAC). 2003.
6. Bó GA, Baruselli PS, Moreno D, Cutaia L, Caccia M, Tribulo H. The control of
follicular wave development for self – pointed embryo transfer programs in cattle.
Theriogenology. 2002. 57: 53–72.
7. Bousfield GR; Butnev VY.Identificación of twelve O-glycosylation sites in equine
chorionic gonadtropin beta and equine luteinizing hormone ss by solid-phase Edman
degradation. Biol Reprod 2001, 64: 136-147.
8. Everton L, Baruselli P. Sincronización de receptoras cruce por cebú en condiciones
tropicales. Cuarto seminario de reproducción de grandes animales (memorias)
Bogotá, 2003.
9. Figueira M, Pegorer RL. Ereno, R.A. Satrapa, V.G. Pinheiro, L.A. Trinca, C.M. Barros
Neither plasma progesterone concentrations nor exogenous eCG affects rates of

Introducción
7
ovulation or pregnancy in fixed-time artificial insemination (FTAI) protocols for puberal
Nellore heifers. Theriogenology, Volume 75, Issue 1, 1 January 2011, Pages 17-23.
10. Filho MF, MO, Reis EL. Viel JO, Nichi M, Baruselli P. Dinámica follicular de vacas
Nelore lactantes em anestro tratadas com progestágeno, eCG e GnRH. Acta
Scientiae Veterinariae 32 (suplemento). 235, 2004
11. Filho MF, Ayres H, Ferreira RM, Marques MO, Reis EL. Silva RCP, Rodrigues CA,
Madureira EH, Bo´ GA, Baruselli P. Equine chorionic gonadotropin and gonadotropin-
releasing hormone enhance fertility in a norgestomet-based, timed artificial
insemination protocol in suckled Nelore (Bos indicus) cows. Theriogenology 2010.
73: 651–658
12. Forcada FM. Ait Amer-Meziane,J.A. Abecia M.C. Maurel J.A. Cebrián-Pérez T.
Muiño-Blanco B. Asenjo MI. Vázquez A. Casao. Repeated superovulation using a
simplified FSH/eCG treatment for in vivo embryo production in sheep Original
Research Article Theriogenology, Volume 75, Issue 4, 1 March 2011, Pages 769-
776.
13. Fuentes S, De la Fuente J. Tasas de gestación de novillas receptoras sincronizadas
con Gonadotropina Coriónica Equina u Hormona Folículo Estimulante. Acta Scientiae
Veterinariae 2007. 35 (Supl. 3): s 767 – s 772.
14. Gouveia MF, Nogueiraa, Melo DS, Carvalho LM, Fuck EJ, Trinca LA, Moraes Barros
C. Do high progesterone concentrations decreasepregnancy rates in embryo
recipients synchronized with PGF2a and eCG?. Theriogenology. 2004. 61, 1283–
1290.
15. Grajales H, Tovío N, Duica A. Fundamentos de fisiología reproductiva en la hembra
bovina. Primera edición. 2011.
16. Grajales, H.; Tovío N. Aspectos Básicos del ciclo estral de la vaca. Revista Genética
Bovina. 2009. 32-36. Volumen 15.
17. Hafez E. Reproducción e inseminación de los animales domésticos, 7ª ed.
Reproductive Health Center IVF/Andrology International. Kiawah Island, South
Carolina, USA. McGraw – Hill Interamericana. 2000. 33–144.
18. Kerbler TL, Buhr MM, Jordan LT, Leslie KE, Walton JS. Relationship between
maternal plasma progesterone concentration and interferon-tau synthesis by the
conceptus in cattle. Theriogenology 1997. 47: 703-714.

8
Introducción
19. Lopes AS, Butler ST, Gilbert RO, Butler WR. Relationship of pre-ovulatory follicle
size, estradiol concentrations and season to pregnancy outcome in dairy cows. Anim
Reprod Sci 2007. 99: 34-43.
20. Nasser L, Reis E, Oliveira M, Bó GA, Baruselli P. The use of hormonal treatments to
improve reproductive performance of anestrous beef cattle in tropical climates. Anim
Reprod Sci 2004. 82, 83: 479–486.
21. Madureira EH, Marques MO, Baruselli P, Nasser LF, Rodrigues CA. Efeito do
tratamiento com eCG na taxa de concepção de vacas nelore com diferentes escores
de condição corporal inseminadas em tempo fixo (analise Retrospectiva). Acta
Scientiae veterinarae .2004. 32 (suplemento): p 228.
22. Mann GE, Corpus luteum size and plasma progesterone concentration in cows. Anim
Rep Sci. 2009. 115: 296–299.
23. Mantovani A, Baruselli P, Bó GA, Cavalcante A, Gacek F. Aumento de las
dimensiones del folículo dominante y del cuerpo lúteo, concentración plasmática de
progesterona y de la tasa de aprovechamiento de receptoras de embriones bovinos
sincronizadas con CIDR-B® por tiempo prolongado. Revista Brasilera de
Reproducción Animal 2002.
24. Mamani E, Ortiz J, Quezada M. Evaluación de diferentes dosis de eCG en
tratamientos de sincronización de celos en receptoras de embriones. VII simposio de
reproducción animal – IRAC. 2007. p, 275.
25. Marques MO, Reis EL, Campos, Filho EP, Baruselli PS. Efeitos da adminiatracao de
eCG e de benzoato de estradiol para sincronizacao da ovulacao em vacas Bos
taurus taurus X Bos taurus indicus no periodo pós parto. In: Simposio Internacional
de Reproducción Animal, Huerta Grande. V. 392, 2003.
26. Murphy B, Martinuk S. Equine chorionic gonadotropin. Endocrine Rewiews 1991. 12:
27– 44.
27. Palma G. Biotecnología de la reproducción. Biotecnología de la reproducción, tercera
edición. Instituto Nacional de Tecnología agropecuaria–INTA. Argentina. 2008. p
693.
28. Peña M, Góngora A, Estrada J. Factores de crecimiento en el desarrollo folicular,
embrionario temprano e implantación. Implicaciones en la producción de embriones
bovinos. Journal MVZ Córdoba 2007. 12 (1): 942–954.

Introducción
9
29. Pita F, Matute R, Tribulo R, Tribulo P, Tribulo H, Bó G. Efecto de la aplicación de la
Gonadotropina Coriónica Equina (eCG) y PGF2 alfa sobre la tasa de
aprovechamiento y tasa de preñez en un protocolo de transferencia de embriones a
tiempo fijo. VIII Simposio internacional de reproducción animal -IRAC 2009.
30. Quesada JM y Ortiz JJ. Sincronización de receptoras de embriones tratadas con
CIDR, benzoato de estradio, D Cloprostenol asociado o no a eCG. VII simposio de
reproducción animal – IRAC. 2007. p, 277.
31. Rodrigues CA, Ayres H, Reis EL, Madureira EH, Baruselli PS. Aumento de taxa de
prenhez em vacas Nelore inseminadas em tempo fixo com o uso de eCG em
diferentes periodos pós-parto.; Acta Scientae veterinariae. V32 (Suplemento), 220.
2004.
32. Rodríguez J. Mecanismos para el reconocimiento materno de la preñez. En:
Reproducción Bovina. Editor González C. Maracaibo – Venezuela. Ed. Fundación
GIRARZ 2001. 31–39.
33. Santos J, Thatcher W, Chebel R, Cerri R, Galvão K. The effect of embryonic death
rates in cattle on the efficacy of estrus synchronization programs. Anim Reprod Sci
2004. 83: 513–535.
34. Santos JE, Thatcher WW, Pool L, Overton MW. Effect of human chorionic
gonadotropin on luteal function and reproductive performance of high-producing
lactating Holstein dairy cows. J Anim Sci 2001;79:2881–94.
35. Silke V, Diskin MG, Kenny DA, Boland MP, Mee JF, and Sreenan JM. Extent, pattern
and factors associated with late embryonic loss in dairy cows. Anim. Reprod. Sci.
2002. 71: 1–12.
36. Siqueira LGB, Torres CAA, Souza ED, Monteiro PLJ, Arashiro EKN, Camargo LSA,
Fernandes CAC, Viana JHM. Pregnancy rates and corpus luteum–related factors
affecting pregnancy establishment in bovine recipients synchronized for fixed-time
embryo transfer. Theriogenology 2009. 72: 949–958.
37. Souza AH, Viechnieski S, Lima FA, Silva FF, Araujo R, Bo´ G, Wiltbank C, Baruselli
P. Effects of equine chorionic gonadotropin and type of ovulatory stimulus in a timed-
AI protocol on reproductive responses in dairy cows. Theriogenology 2009. 72:10–21.
38. Spencer T, Burghardt R, Johnson G, Bazer FW. Conceptus signals for establishment
and maintenance of pregnancy. Anim Reprod Sci 2004. 83: 537– 550.

10
Introducción
39. Spencer T, Johnson GA, Bazer FW and Burghardt RC. Implantation mechanisms:
insights from the sheep Center for Animal Biotechnology and Genomics, Public
Health, Texas A&M University. 2004.128: 657–668.
40. Thatcher W, Guzeloglu A, Binelli M, Hansed TR, Pru JK. Uterine conceptus
interactions and reproductive failure in cattle. Theriogenology 2001. 56: 1435–1450.
41. Thibier M. More than half a millon bovine embryos transfered in 2002: a report from
the IETS data retrieval committee. IETS Newsletter, 2003.
42. Tovío N, Duica A, Grajales H. Mortalidad embrionaria en bovinos. Rev.Genética
bovina, vol.19, p. 34-38. 2010.
43. Tovío N, Duica A, Grajales H. Desarrollo embrionario y estrategias antiluteoliticas
hormonales en programas de transplante de embriones bovinos. Rev.MVZ Cordoba,
vol.13, no.1, p.1240-1251. 2008.
44. Vasconcelos J, Sartori R, Oliveira H, Guenther J, Wilbank M. Reduction is size of the
ovulatory follicle reduces subsequent luteal size and pregnancy rate. Theriogenology
2001. 56: 307–314.

1. Revisión de literatura
1.1. Algunos aspectos relacionados al transporte y regulación hormonal
1.1.1. Transporte hormonal Después de la secreción por parte de células especializadas, las hormonas pueden
encontrarse en el torrente circulatorio libres o unidas a transportadores proteicos. En
general, las hormonas proteicas y peptidicas, así como las catecolaminas, por su carácter
hidrosluble circulan en forma libre. Por el contrario las hormonas tiroideas y los esteroides
se unen a proteínas transportadoras. Esta forma de transporte tiene también un gran
significado fisiológico, ya que por ejemplo, evita las pérdidas de esteroides por filtración
glomerular. La cantidad de hormona libre y la unida a proteínas se encuentran en equilibrio,
pero solamente la hormona libre es la que produce las acciones fisiológicas (Driancourt et
al., 2000).
Existen varios transportadores de esteroides, como la transcortina o globulina ligante de
cortisol (CBG), la cual une con alta afinidad los corticosteroides y la P4. El estradiol y la
testosterona se unen a la globulina ligante de hormonas sexuales (SHBG). Es conveniente
saber que además existen sustancias que pueden influir en los niveles sanguíneos de
estas proteínas transportadoras, así como en su afinidad por los ligadores; por ejemplo, los
estrógenos aumentan los niveles de SHBG; en cambio, los andrógenos los disminuyen.
Esto constituye otro mecanismo de regulación de la acción hormonal (Toosi et al., 2010).
1.1.2. Regulación hormonal La secreción hormonal se relaciona con el papel que desempeñan en el mantenimiento de
la homeostasis. El mecanismo de regulación dominante es el llamado de retroalimentación
negativa; este es el caso típico observado por la acción de la hormona gonadotrófica

12 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
folículo estimulante (FSH), la cual actúa en asocio con la hormona luteinizante (LH) sobre
las células del folículo ovárico estimulando su crecimiento y la síntesis de estradiol (Sartori
et al. 2002); a su vez las células de la granulosa del folículo en desarrollo producirán
principalmente otras dos hormonas, estradiol e inhibina; un aumento de estas dos
hormonas en sangre actuarán sobre el hipotálamo y la hipófisis para suprimir la secreción
de FSH (Rabiee et al., 2001).
Ocasionalmente se observa retroalimentación positiva cuando un producto de la acción de
las hormonas estimula, en principio, secreción hormonal adicional. Este mecanismo de
regulación se observa principalmente en la fase preovulatoria del ciclo estral bovino. En
este momento del ciclo y cuando no hay niveles circulantes de progesterona se va a
producir un aumento de los niveles circulantes de estrógenos (producido por el folículo
preovulatorio). Este pico preovulatorio de estrógenos actuará a nivel del hipotálamo para
producir una descarga preovulatoria de hormona liberadora de gonadotropinas (GnRH) y
consecuente con esto el pico preovulatorio de LH (Starbuck et al. 2004).
La regulación por retroalimentación, ya sea positiva o negativa, se puede ejercer en todos
los niveles de la función celular endocrina, es decir, en la síntesis de las hormonas y en la
liberación de la hormona almacenada. Existen pautas de liberación hormonal integradas
que son dirigidas por ritmos circadianos, fases de sueño, variaciones estacionales y fases
del desarrollo (fetal, neonatal, puberal o de la vejez). Además, el dolor, las emociones, el
terror, las lesiones y el estrés pueden desencadenar la liberación de hormonas a través de
complejas vías neurales (Rabiee et al., 2001).
1.2. Mecanismo de acción hormonal Las células del organismo necesitan comunicarse entre sí con el fin de regular y coordinar
su desarrollo, diferenciación, división y acción. Las hormonas casi nunca actúan
directamente en la maquinaria celular sino que primeramente se deben unir a receptores
específicos, los cuales casi siempre son proteínas muy grandes, encontrándose alrededor
de 2.000 a 100.000 receptores aproximadamente en cada célula, los cuales son altamente
específicos para una determinada hormona. (Lüttgenau et al., 2011).
La ubicación de los receptores en la célula blanco van a estar directamente relacionados
con la naturaleza química de las hormonas y su capacidad para atravesar la membrana

Capítulo 1 13
celular. Cuando las moléculas son de naturaleza hidrosoluble (proteínas, péptidos y
catecolaminas), los receptores se van a encontrar en la membrana celular; en cambio,
cuando las moléculas son de naturaleza liposoluble los receptores se encontrarán dentro
de la célula. Dentro de estos últimos, hay receptores que se encuentran en el citoplasma
como lo son los receptores de las hormonas esteroideas; otros receptores están en el
núcleo, como lo son los receptores de las hormonas tiroideas (David and Michael, 2000).
La reacción entre hormona y receptor proporciona la especificidad y determina la magnitud
inicial de la acción hormonal, la cual está regida por la cantidad de receptores ocupados
por la hormona; esta magnitud va a aumentar cuando el receptor tiene una alta afinidad por
la hormona, cuando la célula se expone a concentraciones mayores de hormona y cuando
el número de receptores es muy alto (McDonald´s, 2003).
Es notable en algunas ocasiones en que la cantidad de hormona unida al receptor no
suele limitar la velocidad, pudiendo observarse respuestas biológicas máximas con sólo
una pequeña parte de los receptores disponibles ocupados por la hormona; las hormonas
peptídicas podrían presentar este sistema de ahorro de receptores (Ritindra, et al., 2001).
Las moléculas de los receptores se sintetizan, se trasladan a las zonas de asociación con
las moléculas hormonales y se degradan continuamente a tal punto de que el número de
receptores en una célula blanco no es constante de un momento a otro (Ritindra, et al.,
2001). En estos procesos pueden influir las respectivas hormonas específicas, las cuales
disminuyen la cantidad de sus propios receptores o ejercen sobre ellos una
desensibilización limitando así su acción sobre la célula. Este proceso se denomina
regulación hacia abajo “Down regulation”; durante este fenómeno disminuye la respuesta
del tejido blanco a la hormona en forma proporcional al grado de disminución del número
de receptores. Este proceso es especialmente observado con las hormonas peptídicas
(David and Michael, 2000).
Si la concentración en sangre de la hormona permanece elevada durante periodos
prolongados (días, semanas), los complejos hormona – receptor de la membrana
celular reaccionan con un movimiento lateral (migración lateral) que los ubica en
una región especial de la membrana denominada “coated pit” (área cubierta). Así
mismo, en caso de exceso crónico de hormonas se produce una concentración de
complejo hormona receptor (formación de racimos) en determinadas regiones de la

14 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
membrana celular, que reaccionan formando una invaginación que les permite
internalizar los complejos hormona - receptor en forma de endocitosis para seguir
metabolizándolos en el interior de la célula (David and Michael, 2000).
Tras el efecto prolongado por el exceso de hormona peptídica, las células diana
pueden hacerse totalmente insensibles a ella porque han internalizado todos los
receptores. La disminución por la densidad de receptores en las células diana
también podría ser un motivo por el cual las hormonas peptídicas se segregan con
un ritmo pulsátil (Intervalos). El incremento y la disminución del número de
moléculas peptídicas segregadas impide que la hormona provoque el “Down-
regulation” de sus propios receptores en la célula diana (Palma, 2008).
Otras hormonas reclutan sus propios receptores, amplificando así la acción de la hormona
sobre la célula. Este proceso se le conoce como regulación hacia arriba “Up – regulation”,
en donde la hormona estimulante induce a la síntesis de más moléculas receptoras en la
célula blanco. En este caso, el tejido blanco se vuelve gradualmente más sensible a los
efectos estimulantes de la hormona (Rabiee et al., 2001).
1.2.1. Receptores de membrana Especialmente las hormonas polipeptídicas, poseen receptores ubicados en la
superficie de la membrana celular, reconociendo las señales reguladoras del
exterior para después transmitirlas y amplificarlas. Los receptores de membrana se
subdividen a su vez en tres clases dependiendo principalmente de mecanismos de
transducción (una hormona puede activar más de un mecanismo de transducción).
La especificidad de las hormonas peptídicas se debe a la estructura de receptores
especiales en la membrana celular de las células diana, donde ejerce un efecto que
induce una señal que será transmitida hacia el interior de la célula (Palma, 2008).
1.2.1.1. Receptores que provocan cambios de permeabilidad de la membrana Prácticamente todas las sustancias neurotransmisoras, que son hormonas locales, se
combinan con receptores en la membrana post sináptica; esto provoca un cambio de la
estructura proteica del receptor y causa la apertura o cierre los canales para uno o más

Capítulo 1 15
iones de Na+, otros para iones de K+, otros para iones de Ca++, etc.; posteriormente el
movimiento de estos iones a través de los canales causa los efectos subsiguientes en las
células post sinápticas (David and Michael, 2000).
1.2.1.2. Receptores acoplados a una proteína G En este caso la interacción entre el receptor y el efector está mediada por una proteína
con capacidad para unir e hidrolizar el nucleótido guanosina trifosfato (GTP). Estos
receptores activan una cadena compleja de acontecimientos que alteran la concentración
de uno o más mensajeros intracelulares (Ritindra, et al., 2001). El más descrito de estos
mensajeros (llamado segundo mensajero, ya que no es la hormona quien produce
directamente los cambios intracelulares, sino el cAMP quien sirve como mensajero para
causar estos efectos) es el adenosin monofosfato cíclico (cAMP), el cual causa varios
efectos al interior de la célula para controlar su actividad (King et al., 2003).
1.2.1.3. Receptores catalíticos Estos receptores al ser activados directamente por las hormonas operan como enzimas.
La mayoría de ellos son proteínas de transmembrana con un dominio citoplasmático que
funciona como una Tirosín Cinasa, como el factor de crecimiento epidérmico (REGF). Un
ejemplo es la insulina y los factores cuya estructura es similar a la insulina (IGF) (Gerardd
et al., 2006). En este caso cuando la hormona se une con la porción externa del receptor
de membrana produce un cambio en la estructura de la molécula receptora, causando a
su vez que la porción interna de la molécula se convierta en una cinasa activada. Luego
esta cinasa provoca la fosforilación de varios sustratos proteicos dentro de la célula que
resulta en la respuesta celular (Roberts, 2007).
1.2.1.4 Mecanismos del segundo mensajero para comunicar funciones hormonales intracelulares El principal medio por el cual las hormonas que no pueden atravesar la membrana ejercen
acciones intracelulares es a través de la formación de una sustancia llamada segundo
mensajero. Este mensajero inducirá casi todos los efectos intracelulares de la hormona. De
esta manera, el único efecto directo que tiene la hormona sobre la célula es activar su propio
receptor, ya que el segundo mensajero hace el resto (Ritindra et al., 2001).

16 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
El cAMP no es el único sistema utilizado como segundo mensajero, otros de especial
importancia son los iones de calcio (Ca++), la calmodulina y productos de fosfolípidos de
la membrana (David et al., 2000).
1.2.1.4.1. Mecanismo del sistema segundo mensajero cAMP La hormona estimulatoria o inhibitoria se une a su receptor específico en la superficie de
membrana de la célula blanco. La especialización del receptor determina qué hormona
afectará a la célula. La unión de la hormona receptor activará la enzima adenil ciclasa a
través de un transductor denominado proteína G (estimulatoria o inhibitoria) (Scott And
Pawson, 2000).
La adenil ciclasa causa conversión inmediata de gran parte del ATP citoplasmático en
cAMP (mensajero intracelular – transmisor más habitual utilizado), que es el compuesto
3´, 5´ - adenosina monofosfato cíclico. Una vez formado el cAMP dentro de la célula,
activa una enzima forforilante llamada proteína cinasa cAMP dependiente o simplemente
cinasa A. La activación de la cinasa A ocurre cuando el cAMP se une a la subunidad
reguladora de la cinasa A y queda libre la subunidad catalítica; entonces la cinasa A
activa puede usar el ATP como fuente de grupos fosfatos de alta energía para fosforilar
otras proteínas celulares y de esta manera se constituye la respuesta celular
(McDonald´s, 2003).
Este mecanismo tiene la particularidad de que cada reacción generalmente activa una
cascada de enzimas. Es decir, una primera enzima es activada y luego esta activa otra
enzima, que a su vez activa una tercera enzima y así sucesivamente. La importancia de
este mecanismo es que solo unas pocas moléculas de adenil ciclasa activada pueden
causar que más moléculas de cAMP sean activadas y esto a su vez generar la activación
de muchas más moléculas de la proteína cinasa A y así sucesivamente. De esta manera,
una mínima cantidad de hormonas actuando en la superficie de la célula puede iniciar una
cascada muy poderosa con la fuerza para activar toda la célula (Campbell et al., 2006).
La acción específica que se produce en respuesta al cAMP en cada tipo de célula blanco
depende de la naturaleza de la maquinaria intracelular ya que las células varían en
cuanto a las enzimas que poseen; por lo tanto, se activan distintas funciones en las

Capítulo 1 17
células blanco. Estas funciones pueden ser: iniciar la síntesis de químicos intracelulares
específicos, causar la contracción o relajación muscular, alterar la permeabilidad de la
célula, y otros efectos. Las hormonas de la reproducción que actúan utilizando
principalmente el cAMP como segundo mensajero son la FSH y la LH (Cunningham and
Klein, 2009).
1.2.1.4.2. Lípidos de membrana como segundos mensajeros Algunas hormonas se unen a sus receptores y el complejo hormona receptor a su vez
activa (vía una proteína G estimulatoria) una enzima fosfodiesterasa (PDE) llamada
Fosfolipasa C o fosfodiesterasa. Esta enzima causa que algunos de los fosfolípidos en la
membrana celular se dividan en sustancias más pequeñas que tienen efectos
intracelulares de segundo mensajero muy difundidos. Una de las hormonas que actúa a
través de este mecanismo es la GnRH (Ritindra, et al., 2001).
Uno de los fosfolípidos de membrana que se rompe de esta manera es el Fosfatidil
Inositol Bifosfato (PIP2). Los productos más importantes que sirven como segundos
mensajeros son Inositol Trifosfato (IP3) y Diacilglicerol (DAG). El IP3 moviliza los iones
de Ca++ desde la mitocondria y el retículo endoplásmico, luego los iones de Ca++
producen sus propios efectos de segundo mensajero como la contracción de músculos
lisos, cambios en la secreción de las células secretoras, cambios en la acción ciliar, etc.
Por su parte el DAG, el otro lípido segundo mensajero, activa la enzima proteína Cinasa
C (Campbell et al., 2006). Esta activación se intensifica con el aumento de iones de Ca++
que se han liberado en respuesta al IP3. Llegado el momento, la proteína Cinasa C
activada tiene un papel especialmente importante en la división y proliferación celular. Es
importante hacer notar que la porción lipídica del DAG es el ácido araquidónico el cual es
precursor de las prostaglandinas (Filmore, 2004).
1.2.1.4.3. Iones de calcio y la Calmodulina como sistema de segundo mensajero Otro sistema de segundo mensajero opera en respuesta al ingreso de iones de Ca++ en
las células. El ingreso de Ca++ puede iniciarse por cambios en el potencial eléctrico de la
membrana que abren los canales de Ca++ en las membranas, o por hormonas que

18 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
interactúan con los receptores de la membrana que también abren canales de ca++
(Ritindra, et al., 2001).
Al entrar en la célula los iones de Ca++ se unen a una proteína llamada calmodulina.
Esta proteína tiene cuatro sitios de de unión al Ca++, separados; cuando tres o cuatro de
estos sitios se han unido con Ca++, ocurre un cambio conformacional que activa la
calmodulina produciendo efectos múltiples dentro de la célula de la misma manera que
funciona el cAMP. Por ejemplo, activa un conjunto de enzimas (diferentes de aquellas
activadas por cAMP), causando a su vez un conjunto adicional de reacciones
metabólicas intracelulares. Una de las funciones específicas de la calmodulina es la de
activar la miosina cinasa que luego actúa directamente sobre la miosina del músculo liso
para causar la contracción de este (Chin and Means 2000).
La concentración normal de iones de Ca++ en la mayoría de células del cuerpo es de 10-
7 a 10-8 mol/litro, lo cual no es suficiente para activar el sistema de calmodulina, pero
cuando la concentración de iones Ca++ se eleva hasta 10–6 a 10–5 mol/litro, hay
suficientes uniones para que ocurran todas las acciones intracelulares de la calmodulina.
Esta es casi la misma cantidad de iones Ca++ que se requiere para activar la troponina
C, la cual causa la contracción de los músculos del esqueleto. Esta troponina C es muy
similar a la calmodulina tanto en su función como en su estructura proteica (Campbell et
al., 2006).
1.2.2. Receptores intracelulares Varios tipos de hormonas, especialmente las esteroideas y las tiroideas, se unen con
receptores proteicos no en la membrana celular sino dentro de la misma célula. Las
hormonas esteroideas son solubles en el medio lipídico de la membrana celular
(liposolubles) y por lo tanto difunden a través de la membrana celular por difusión pasiva
(Cunningham and Klein, 2009).
Las proteínas receptoras de esteroides son asimétricas presentes en un número de
3.000 a 100.000 copias por célula efectora. Las respuestas son proporcionales al número
de receptores con hormona esteroidea unida, sugiriendo que no hay segundos
mensajeros intermediarios como el caso de las hormonas polipeptídicas. El complejo

Capítulo 1 19
hormona receptor activado luego va a dirigirse al núcleo celular donde se une y activa
puntos específicos para formar el ARNm; por lo tanto, en estos casos pasa un tiempo
desde que la hormona ha ingresado en la célula hasta que las proteínas recién formadas
aparecen en la célula y pasan a controlar las funciones celulares nuevas o aumentadas
(Senger, 2005).
La secuencia y mecanismos de acción de las hormonas esteroideas podría resumirse de
la siguiente manera: La hormona entra en el citoplasma en donde se une a la proteína
receptora específica, por lo cual se ocasiona una alteración física en el receptor de modo
que se va a unir fuertemente a la cromatina en el núcleo; mientras el receptor no esté
unido a nada, estos puntos de unión de la cromatina se inactivan mediante proteínas de
choque térmico que van asociadas con la molécula receptora; estas proteínas de choque
térmico se sintetizan en mayor cantidad en situaciones de estrés como por ejemplo calor
y ayudan en determinadas funciones fisiológicas como en la regulación de la actividad de
los factores de unión al ADN (Peres, 2005). Luego se presenta una combinación activa
en el proceso de transcripción de genes específicos para formar el ARN mensajero
(ARNm), el cual se difunde en el citoplasma, donde conduce el proceso de traducción en
los ribosomas para formar nuevas proteínas (McDonald´s, 2003).
La acción de una hormona esteroidea solo dura mientras el complejo hormona receptor
está unido a la cromatina. La actividad biológica dependerá de la tasa de disociación de
la hormona esteroidea de su receptor y por otro lado, la actividad biológica dependerá de
la semivida del complejo hormona receptor con su dominio de la unión a la cromatina
(Ritindra, et al., 2001).
1.3. Aspectos sobre dinámica folicular y hormonal Rajakoski en 1960, propuso la existencia de dos ondas de crecimiento folicular en las
vacas, las cuales comenzaban a evidenciarse entre los días 3 a 12 y 13 a 0 del ciclo
estral bovino. Posteriormente con el uso de la ultrasonografía como método efectivo para
realizar el seguimiento y evaluación de la actividad folicular ovárica diversos
investigadores confirmaron la presencia de estas ondas de crecimiento y patrones de
desarrollo de folículos antrales (Buratini et al., 2000; Gintherg et al., 2001).

20 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
El número de ondas de crecimiento folicular presentes en cada ciclo varía de 1 a 6;
presentándose en la mayor parte de las hembras 2 a 3 ondas. En un ciclo con 2 ondas,
la maduración del segundo folículo dominante coincide con la regresión del cuerpo lúteo
y culmina con la ovulación del mismo. En otros casos se presentan 3 ondas de
crecimiento folicular y coincide con un ciclo estral de mayor duración, debido a que se
posterga el celo al no ovular el segundo folículo dominante y en consecuencia, el tercer
folículo requiere de un tiempo para madurar y poder ovular (Martinez et al., 2000; Peres,
2005).
En un estudio realizado por Cardozo et al., (1994), con novillas Holsteín en la Sabana de
Bogotá, se halló que el 20% de ellas presentaban constantemente ciclos de 25 días de
duración, presumiblemente con tres ondas de crecimiento folicular.
Durante el crecimiento de una onda folicular se podrían describir claramente cuatro
fases:
- Reclutamiento
Durante el CE un grupo de 3 a 6 folículos (de 2 a 5 mm.) comienzan a desarrollarse a
partir de una cohorte de folículos antrales pequeños que comienzan a madurar bajo un
aporte adecuado de Gonadotropinas, especialmente por un pico transitorio de FSH, que
le permiten avanzar en su desarrollo (Palma, 2008). Los niveles circulantes de FSH antes
del reclutamiento de un grupo de folículos aumentan transitoriamente y esto se
caracteriza por la expresión de mRNA que codifica para la elaboración de las aromatasas
P450arom y P450scc en las células foliculares (Revisado por Grajales, et al, 2011).
El desarrollo folicular desencadena el inicio de la primera onda de crecimiento folicular,
estimulada por el segundo pico de FSH el cual ocurre posterior a la ovulación (Liu et al.,
2007).
Existe un claro patrón durante el crecimiento de la primera onda folicular; los folículos
exitosos responden al incremento transitorio de FSH aumentando su crecimiento y la
síntesis de estradiol, así como una elevación en la producción de inhibinas de alto peso
molecular y activina, mientras que la folistatina y el dimero de inhibina de bajo peso
molecular se mantienen en bajas concentraciones. A pesar de esto los dos mayores

Capítulo 1 21
competidores por la dominancia no se pueden distinguir hasta después de ocurrida la
selección (Mihm et al., 2000; Austin et al., 2001)
Frecuentemente al momento del reclutamiento los diferentes folículos de la cohorte son
desiguales; los más pequeños son ricos en EGF y más sensibles al efecto inhibitorio de
la EGF en la diferenciación celular. Por tanto, el efecto estimulatorio de la FSH sobre la
actividad aromatasa es más marcado en los folículos más grandes de la cohorte
(Safdarian et al., 2006).
Se ha reportado que se establece una competencia por la dominancia entre los folículos
reclutados en cada onda folicular, por lo cual solamente un folículo de la cohorte adquiere
un desarrollo funcional y estructural, lo que le permite seguir su crecimiento en un
ambiente bajo en concentraciones de gonadotropinas, al tiempo en que sus compañeros
de cohorte sufren atresia (Revisado por Montaño y Ruiz, 2005).
- Selección
Durante los días 2, 3 y 4 del ciclo estral, se detectan por medio de ultrasonografía uno o
varios folículos (provenientes de la etapa de reclutamiento) de un tamaño promedio de 6
a 9 mm., con lo cual la fase selección comienza a ejercerse (Avila et al., 2005). A medida
que los folículos maduran, comienzan a depender de la LH, lo cual puede ser parte del
mecanismo de selección del folículo dominante (Mihm et al., 2000; Fortune et al., 2001).
Los cambios en el patrón de expresión del mRNA para los receptores de la
gonadotropinas y enzimas esteroidales dentro de las células foliculares, parecen estar
muy ligados a las modificaciones en las concentraciones sanguíneas de gonadotropinas
(Rubianes, 2000).
La selección se relaciona con la interferencia del folículo más grande sobre la capacidad
de los folículos más pequeños de recibir un adecuado soporte gonadotrópico. Esto podría
ser llevado a cabo mediante dos vías. La vía pasiva por la cual el folículo mayor inhibe
indirectamente el crecimiento de los folículos menos maduros reduciendo las
concentraciones de FSH por debajo del umbral necesario para mantener a los otros
folículos. En la vía activa, el folículo mayor inhibe directamente el crecimiento de los
demás folículos secretando en la sangre sustancias que reducen su sensibilidad a la FSH
(Recabarren et al., 2003).

22 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
- Dominancia
La fase de dominancia, es el proceso por el cual el folículo seleccionado ejerce un efecto
inhibitorio sobre el reclutamiento de una nueva cohorte de folículos. Para el
establecimiento de esta dominancia se requiere que se presente divergencia o
desviación, que corresponde al tiempo en el cual el folículo dominante y el (los)
subordinado (s) más desarrollado (s) crecen a una tasa diferente, antes de que el
subordinado manifieste atresia (Revisado por Montaño y Ruiz, 2005).
Los estímulos hormonales durante esta etapa se dirigen hacia el folículo dominante con
lo cual aumenta su irrigación sanguínea. La maduración de este folículo se relaciona con
los niveles altos de proteína reguladora esteroidogenica aguda (StAR), síntesis de
receptores para FSH y LH (principalmente de receptores de LH – LHR), así como la
elaboración de proteínas y enzimas aromatasas necesarias para la elaboración de
andrógenos y progestágenos (Axel et al., 2002).
El folículo dominate alcanza un tamaño marcadamente superior a los demás (diámetro
mayor a 10 mm) y es responsable de la secreción de estradiol (Belkys et al., 2005). Esta
actividad estrogenica está relacionada con el incremento en la expresión de genes para
aromatasas (P450 arom y P450 17 – α hidroxilasa), 3β-hidroxi-esteroide deshidrogenasa
(3βHSD) y receptor de FSH, así como la adquisición de receptores para LH en las
células de la granulosa (Meduri et al., 2008). Igualmente esta actividad estrogénica se
correlaciona negativamente con las cantidades intrafoliculares de IGFBPs (IGFBP2,
IGFBP4 y IGFBP5) < 40 kDa (Mazerbourg et al., 2001).
Una vez el folículo dominante ha sido seleccionado la IGFBP 4 y IGFBP 5, se mantienen
en niveles bajos, y el folículo dominante incrementa la biodisponibilidad de IGF (Austin et
al., 2001).
Se ha encontrado que el IGF-I está presente en altas concentraciones en los folículos de
mayor tamaño y su concentración se va incrementando paralelamente con el incremento
en la receptividad de las células de la granulosa a IGF-I, lo cual estimula el proceso de
aromatización así como el incremento de los receptores de LH en las células de la
granulosa (Meduri et al., 2008). De acuerdo a esto, se ha encontrado que la expresión de
genes que codifican para PAPA-A está relacionada con la expresión de aromatasas y

Capítulo 1 23
receptores para LH en las células de la granulosa; esto podría ser importante para las
etapas finales de selección del folículo dominante (Mazerbourg et al., 2001).
Bodensteiner et al. (1996), realizaron un estudio en donde marcaron radiactivamente y
contaron los receptores para FSH (FSH-R) y LH (LH-R) en células de la granulosa de
folículos dominantes y subordinados los días dos y cuatro de la onda, encontrando que el
día dos no se presentaron diferencias en el número de FSH-R y LH-R por célula y por
folículo, pero el día cuatro las diferencias entre ambos tipos de folículos eran altamente
significativas para FSH-R y significativas para LH-R en favor de los folículos dominantes.
Sin embargo, Xu et al. (1997) no encontraron diferencias en los niveles de expresión
génica para FSH-R en células de la granulosa de folículos bovinos que medían entre 0.5
y 14 mm de diámetro y solo hallaron expresión de genes para LH-R en folículos mayores
de 9 mm, lo cual sugiere que los FSH-R no limitan el establecimiento de la dominancia
(Revisado por Henao y Trujillo, 2000).
Se sugiere igualmente que el mismo aumento de los receptores (LHR), posiblemente
incrementan la actividad aromatasa en respuesta a las gonadotropinas, o posiblemente
provoca un cambio de dependencia de FSH a LH; esto aumentaría o mantendría la
capacidad del folículo dominante para producir más estradiol que los folículos
subordinados (Revisado por Montaño y Ruiz, 2005).
La respuesta diferencial a la FSH es fundamental en la selección del folículo dominante y
por tanto las acciones intrafoliculares de factores de crecimiento ováricos inducidos por la
FSH juegan un papel importante en la modulación de la respuesta de la cohorte folicular
a las gonadotropinas. Cambios en las cantidades intrafoliculares de proteínas
pertenecientes a la familia inhibina e IGF podrían ser marcadores predictivos de la
sobrevivencia folicular durante la caída de la FSH (Mihm et al., 2000), en donde las
inhibinas que abundan en el fluido folicular las cuales tienen diferentes formas diméricas
(subunidades α y β) con distintos pesos moleculares ejercen su acción supresora de esta
gonadotropina (Knight y Glister, 2001).
Para explicar la causa por la cual el folículo dominante de las primeras ondas (1 de 2
ondas, 2 de 3 ondas y 3 de 4 ondas) sufre regresión, se ha propuesto que se debería a
una disminución en la frecuencia de los pulsos de LH debido a los altos niveles de P4

24 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
circulante (Revisado por Montaño y Ruiz, 2005). Ello provocaría una menor síntesis de
andrógenos y en consecuencia de estradiol. Manikkam y Rajamahendran (1997)
sugierieron que la atresia del folículo dominante estaría relacionada con una menor
actividad de la enzima aromatasa en las células de la granulosa y no por falta de sustrato
(andrógenos). El autor concluye que puede existir un aumento de IGFBPs (Proteína
fijadora de Factor de Crecimiento Similar a la Insulina) con una consecuente disminución
en la disponibilidad de IGF – I y II, lo cual provocaría una disminución de la actividad de
la enzima aromatasa y en consecuencia una menor síntesis de estrógenos.
Durante la fase de crecimiento el folículo dominante mantiene bajos niveles de IGFBPs
de bajo peso molecular, mientras que la pérdida de dominancia está acompañada por el
incremento de estas, dando lugar a apoptosis de las células de la granulosa (Revisado
por Olivera et al., 2007).
En folículos preovulatorios se incrementan las concentraciones de IGF-I, así como la
actividad estrogénica, mientras que las cantidades intrafoliculares de IGFBPs 2, 4 y 5 se
encuentran reducidas o indetectables. Incrementos de IGFBPs 2 y 5 se han observado
en folículos preovulatorios después del pico de gonadotropinas y justo antes de la
ovulación, cuando la P4 intrafolicular se eleva y la producción de estradiol y andrógenos
disminuye (Knight y Glister, 2001).
- Atresia
La gran mayoría de los folículos presentes al nacimiento degeneran a través del proceso
conocido como atresia (clausura u obliteración de un orificio o conducto del cuerpo), la
cual ha sido descrita para diversas especies mamíferas (Huanca, 2001).
La fase de atresia, consiste en la desaparición de los folículos que no son seleccionados
como dominantes, o del folículo dominante el cual no llega a ser ovulatorio (cuando la
lisis del cuerpo lúteo no coincide con la dominancia folicular) (Braw-Tal y Roth, 2005).
Se debe aclarar que la atresia se presenta en cualquier estadio del desarrollo folicular,
aunque es más frecuente en folículos antrales y su incidencia está directamente
relacionada con el tamaño de los folículos; los más grandes presentan un índice
proliferativo mayor, que hace a sus células más susceptibles a la muerte por apoptosis y
por lo tanto a la atresia (Ranferi et al., 2010).

Capítulo 1 25
Cuando los folículos sufren atresia cesa la síntesis de estradiol y las concentraciones de
P4 intrafolicular aumenta. Igualmente durante este proceso, se destacan algunos cambios
morfológicos e histológicos, como son: núcleos picnóticos y fragmentación nuclear en las
células de la granulosa, desprendimiento de las células de la granulosa por la pérdida de
la matriz intercelular, desprendimiento del complejo cumulus-ovocito y en algunos casos
hipertrofia de las células de la teca (Sharma, 2000). Además, ocurren procesos
bioquímicos como la reducción en la síntesis y por ende en la cantidad de ADN en
células de la granulosa (Rosales et al., 2000; Olhagaray 2004), pérdida de uniones
comunicantes, pérdida de receptores a gonadotropinas (Ranferi et al., 2010), así como la
disminución en la síntesis y expresión de mRNA para aromatasas y receptores a
gonadotropinas (Sharma, 2000).
Es importante resaltar el aumento en la expresión de varios genes, entre ellos, el de las
proteínas de unión del Factor de Crecimiento Isulínico (IGBPs) (Pascual-Leone 2001), el
de la Glicoproteína -2-Sulfatada (TRPM-2) (Igwebuike, 2010), el de una Aspartil
Endopeptidasa, la Catepsina-D y el del receptor para la Angiotensina II (Hastie and
Haresign, 2006). Además, en la atresia participan enzimas lisosomales (Fosfatasa Ácida
y Glucosaminidasa), con actividad Anti-Tripsina (Rosales et al., 2000), las involucradas
en la remodelación tisular como Colagenasas, Gelatinasas, Pz-Peptidasa (Shimizu et al.,
2008) y el activador del Plasminógeno entre otras (Hastie and Haresign, 2010).
La presencia de núcleos picnóticos y cariorrexis en las células de la granulosa y teca de
folículos atrésicos (Hastie and Haresign, 2008), así como estudios bioquímicos
posteriores que demostraron la fragmentación internucleosomal del ADN en estas
células, han permitido conocer que la apoptosis es el tipo de muerte preponderante en la
atresia (Igwebuike, 2010), aunque no el único (Rosales-Torres et al., 2010).
1.3.1. Factores que controlan el desarrollo folicular 1.3.1.1. Factores endocrinos Los mecanismos de selección folicular no son aún completamente claros, pero se conoce
que involucran complejas relaciones entre esteroides intrafoliculares, péptidos ováricos,
factores de crecimiento y gonadotropinas hipofisarias (Katagiri and Takahashi, 2006).

26 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
El contenido del fluido folicular tiene una serie de factores secretados por las células de
la granulosa y de la teca. Estos factores probablemente interactúan constantemente, así
el balance entre la acción estimuladora e inhibidora dentro del folículo en sus diferentes
estadios determinará el desarrollo o la atrésia folicular (Rosales et al., 2010).
Estudios reportados por Ireland et al., (2000), sugieren la existencia de factores que se
encuentran en el fluido folicular, especialmente factores de crecimiento, tales como
inhibina, activina, factor insulínico de crecimiento (IGF) y sus proteínas transportadoras,
que juegan un papel importante en la regulación del crecimiento, diferenciación y función
folicular. La variación entre ellos determinaría cual es el folículo dominante en cada onda
folicular.
Mihm et al., (2000), hallaron que los futuros folículos dominantes producen más cantidad
de E2 y P4, mientras las concentraciones de proteínas transportadoras de IGF fueron
bajas, siendo estos marcadores bioquímicos para determinar que folículo de una cohorte
de similar tamaño se volverá dominante. Los niveles intrafoliculares de P4, inhibina,
activina, folistatina y proteínas transportadoras para IGF fueron similares en todos los
folículos de la cohorte.
Campbell (1999), estudió los factores que pueden incrementar o atenuar la acción de las
gonadotropinas para estimular el desarrollo folicular. Usando modelos In vitro, observó
como el factor insulínico de crecimiento (IGF) estimula la proliferación y la producción de
hormonas por parte de las células de la teca y de la granulosa. Igualmente observó “in
vivo” el estímulo para desarrollo folicular en el ovario y la secreción de andrógenos y
estradiol; mientras que el Factor Transformador de Crecimiento α (TGF α) y el Factor
Epidérmico de Crecimiento (EGF) inhibió la diferenciación de las células de la teca y de
la granulosa. Estos reportes coinciden con los de Lucy (2000), al respecto de que el IGF
actúa en la diferenciación, crecimiento y proliferación de las células foliculares, siendo la
más importante función el sinergismo con las gonadotropinas para cumplir variadas
funciones, entre ellas la mitogénesis y la esteroidogénesis. Este sinergismo es logrado
gracias a la habilidad del IGF para incrementar el número de receptores para las
gonadotropínas y el incremento de receptores de los segundos mensajeros. Al mismo
tiempo las gonadotropinas incrementan la expresión de receptores para IGF y pueden

Capítulo 1 27
incrementar la síntesis de IGF en las células de la granulosa. (Padmanabhan et al.,
2002).
La emergencia de una onda folicular precedida por un pico de FSH coincide con el cese
de crecimiento del folículo dominante, el cual posee concentraciones más altas de
estradiol que los folículos subordinados; por lo tanto, un incremento rápido en los niveles
de estradiol es una característica crucial del folículo dominante (Fortune et al., 2004).
El incremento en las concentraciones de FSH que preceden a la emergencia de la onda
ovulatoria se atribuyen a la declinación de las concentraciones de sustancias inhibitorias
(estradiol, inhibina, folistatina y otras sustancias) (Fortune, et al., 2004; Donadeu y
Ginther, 2002). Este pico de FSH culmina cuando el folículo dominante o más grande
está alrededor de 5 mm en hembras bovinas y 13 mm en yeguas (Ginther et al., 2003).
El Control endocrino de la maduración folicular se basa en las células de la granulosa y la
teca. Si los folículos alcanzan su fase de desarrollo reactivo a las gonadotropinas, las
células de la granulosa presentan receptores para FSH y las células de la teca presentan
receptores para LH. La LH en su segregación en cantidades basales en la adenihipofisis,
al llegar y unirse a los receptores de las células de la teca estimula la síntesis de
andrógenos por mediación del cAMP. Los andrógenos sintetizados en las células de la
teca a partir del colesterol en donde intervienen las enzimas P450scc (enzima de clivaje
de la cadena lateral de colesterol) y 3β – hidroxiesteroide deshidrogenasa (3β-HSD),
pasan a través de la membrana basal hacia las células de la granulosa en donde sucede
una aromatización causada por el citocromo P450 arom (aromatasa) y P450 17 – α
hidroxilasa, de esta manera se convierten los andrógenos en estrógenos, proceso el cual
se presenta por la estimulación hecha por la FSH la cual cuenta con sus receptores en
dichas células. (Niswender et al., 2000). La pérdida del proceso de aromatización
descrito, el cual es causado por enzimas específicas marca el inicio de la luteinizaciòn
(Murphy, 2000).
El número cada vez mayor de células de la granulosa provoca la síntesis continuamente
creciente de estrógenos y la secreción del folículo en fase de maduración (Rosales et al.,
2010). Estos estrógenos segregados tienen dos efectos muy importantes a nivel folicular
e hipofisiario. A nivel folicular inducen la invasión de vasos sanguíneos (angiogénesis)
por la teca interna; así este folículo recibe más cantidad de sangre que el resto de
folículos de su oleada de desarrollo (Beg et al., 2002). A nivel de hipófisis, la

28 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
concentración creciente de estrógenos junto con el aumento simultáneo de inhibina
(segregada por las células de la granulosa) impide que siga incrementándose la
secreción de FSH (feed back negativo) (Guinther et al., 2003). Esto mejora las
probabilidades de desarrollo del folículo que llegará a ser dominante frente a los otros
folículos competidores de la cohorte, y ejerce una dominancia pasiva sobre ellos,
impidiendo que a sus receptores se una la FSH cada vez más escasa, gracias a su mejor
riego sanguíneo y a su número mucho mayor de receptores (Revisado por Jiménez y
Hernández, 2000).
El incremento del estradiol está asociado con un incremento en la expresión de genes
en células de la granulosa para receptores de aromatasa y receptores para FSH y LH
(Garverick et al., 2010).
En la fase final de la maduración (desarrollo preovulatorio), los estrógenos ejercen un
tercer efecto significativo sobre el folículo, puesto que se induce la formación de
receptores de LH en las células de granulosa (Crowe et al., 2001). De esta forma el
folículo preovulatorio dispondría de los puntos de unión hormonal para reaccionar a la
secreción de LH que provocará la ovulación (Murphy, 2000).
El mRNA para receptores de LH en las células de la granulosa ha sido detectado en
folículos > 8mm, mas no en los folículos subordinados. Se ha reportado recientemente
que el mRNA para proteína receptora de LH es cerca de 8 veces mayor en el folículo
más grande que en el segundo durante el día del inicio de la desviación. Por tanto, la
inducción de receptores de LH en las células de la granulosa es uno de los eventos que
determinan la selección de un folículo dominante (Beg y Ginther, 2006).
La frecuencia alta de los pulsos de LH es necesaria para mantener la dominancia durante
la onda folicular, mientras que una frecuencia reducida pero de mayor amplitud marca la
pérdida de la dominancia (Perera et al., 2005).
La expresión de enzimas esteroidogénicas y de los genes receptores de gonadotropinas
desciende en los folículos preovulatorios después del pico de LH, esto acompañado con
descenso en la concentración de androstenediona y de 17 β estradiol y con el incremento
en la concentración de P4 en el fluido folicular. Este aumento después del pico

Capítulo 1 29
preovulatorio de LH estaría relacionado con el inicio de la luteinización de las células
tecales y de la granulosa o con la reducción en la conversión de progestinas a
andrógenos (Fike et al., 2004).
Al comienzo el número de receptores de FSH de una célula de la granulosa son
limitados, de manera que un folículo en crecimiento solamente puede multiplicar hasta
cierto punto los receptores de FSH de una célula de la granulosa, ya que su
retroalimentación sería limitada. A medida del crecimiento del folículo se va aumentando
el número de células de la granulosa, lo que le permite aumentar rápidamente el número
de receptores de FSH. Puesto que el estrógeno sintetizado tiene un efecto mitógeno
(estimulante de la mitosis), el destino final del folículo dependerá de si es capaz de
sintetizar estrógenos en cantidad suficiente para aumentar el número de sus células,
aumentando así indirectamente el número de receptores para FSH. Solamente cuando
es capaz de convertir los andrógenos procedentes de la teca en estrógenos, consigue
mantener un microambiente impregnado de estrógeno. Si no lo consigue sufre una
transformación andrógena y la consecuente atresia (Murphy, 2000; Hafez, 2000).
1.3.1.2. Factores exógenos Existen factores que aunque son independientes de la función ovárica pueden llegar a
alterar de manera directa su normal funcionamiento. La nutrición juega un papel
importante en la reproducción. Se establece que el 80% de las vacas en lactancias
tempranas tienen un balance de energía negativo; pierden peso, disminuyen la
producción láctea y deprimen la función reproductiva (Gwazdauskas et al., 2000).
Las vacas con severo estrés nutricional fallan en la ovulación del primer folículo
dominante, posiblemente debido a que la frecuencia en el pulso de LH es reducida para
estimular la suficiente secreción de estrógenos por parte del folículo dominante e inducir
el pico preovulatorio de LH. En novillas de carne se evidencia que la restricción
nutricional aguda actúa tanto a nivel ovárico como del eje hipotalámico – hipofisiario,
disminuyendo la tasa de crecimiento del folículo dominante que parece estar suprimida
por la declinación en la concentración de IGF (Mackey et al., 2000).

30 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
La condición corporal (CC) es un factor el cual se puede ver afectado ya sea por
condiciones en el periodo posparto, lactancia, estrés calórico (descenso en ingesta de
alimento), nutricionales, patológicas etc. Las vacas que sufren pérdida de CC desde el
parto hasta el servicio tienen los índices de concepción más bajos (Wiltbank, 2001). En
relación con lo anterior, se puede decir que el balance de energía está directamente
relacionado con la CC y esta a su vez con el número de folículos pequeños presentes en el
ovario, siendo mayor en número en vacas con CC entre 3 y 5 (Gwazdauskas et al., 2000).
Las vacas que mantienen el peso y la condición corporal presentan mayor concentración
de LH basal y gran liberación de LH inducida por la GnRH comparadas con aquellas con
menor condición corporal (Mackey et al., 2000).
La leptina una citoquina que recientemente se ha propuesto como modulador del eje
hipotalámico – hipofisiario, es secretada por los adipositos y su concentración plamática
se relaciona con pérdida de condición corporal, evidenciándose que en restricción
nutricional incrementa la expresión de receptores de leptina en el núcleo hipotalámico
(Amstalden et al., 2000).
Las concentración de leptina en plasma disminuye durante el balance energético
negativo, puesto que el consumo de nutrientes influye sobre las cantidades de mRNA
para leptina en grasa (Amstalden et al., 2000).
El hipotálamo convierte las señales de la leptina en respuestas neuronales sobre el
consumo de alimento. El Neuropéptido Y (NPY) el cual es un neurotransmisor que se
encuentra en el núcleo Arquato y que afecta la secreción de las hormonas hipofisiarias
especialmente la LH, parece ser importante en la regulación del consumo. La leptina
inhibe las señales del NPY afectando en baja el consumo de alimento (Jang et al, 2000).
La administración de leptina también estimula la producción de gonadotropinas en la
hipófisis por medio de estimulación de GnRH en el hipotálamo (Zieba et al, 2003). En
rumiantes, la administración de leptina recombinante ovina en vacas de carne adultas
sometidas a ayuno estimuló la secreción de LH (Amstalden et al, 2000). En vacas de
leche ovariectomizadas afectó la secreción de LH en relación directa con la dosis
aplicada (Zieba et al, 2003).

Capítulo 1 31
Durante la preñez los niveles de leptina son altos y disminuyen rápidamente luego del
parto. Esta caída es debida a los costos energéticos de la producción de leche (Block et
al, 2001). El estimulo de la lactancia por sí mismo no parece influenciar la disminución de
las concentraciones de leptina (Zieba et al, 2003).
1.4. Aspectos relacionados al ciclo estral de la vaca El ciclo estral (CE) se ha definido como el intervalo entre la presentación de un celo,
estro o calor (receptividad sexual) y la aparición del siguiente, cuyo rango de duración
está entre 17 y 21 días, dependiendo de factores como la raza entre otros (Revisado por
Grajales y Tovío 2009).
El ciclo estral está controlado por la interacción del eje hipotálamo – hipófisis – ovario –
útero, lo cual se enmarca un sistema jerárquico, en cuya cima estaría el hipotálamo, en el
centro la hipófisis y en la base los ovarios y útero (Beltmana et al., 2011).
El hipotálamo es una estructura que se encuentra estrechamente unida a la parte inferior
del cerebro. Entre sus neurosecreciones se encuentra la hormona liberadora de las
gonadotropinas (GnRH), la cual se sintetiza básicamente en las células que se
encuentran en el núcleo arqueado, y de allí se secreta al sistema portahiposifisiario. Esta
hormona es un pequeño péptido de 10 aminoácidos (decapéptido), el cual tiene
funciones autócrinas y parácrinas, posee una semivida de apenas 2 a 4 minutos, por lo
que se entiende que el mantenimiento del ciclo estral depende de la secreción
permanente de esta hormona. La importancia de la GnRH radica esencialmente en la
estimulación para la secreción simultánea de otras hormonas, la luteinizante (LH) y
foliculoestimulante (FSH) (Cattle et al., 2011).
La secreción de las moléculas de GnRH de las neuronas del núcleo arqueado está
sometida a una regulación muy fina, influida por la propia GnRH, por las gonadotropinas
segregadas y por las hormonas esteroides del ovario, las cuales siguen un bucle largo de
retroalimentación (long feed - back loop) (Webb et al., 2004). Puesto que las neuronas
productoras de GnRH no tienen receptores para estrógenos, esta señal hormonal
esteroidea se transformará en un mediador de dopamina, adrenalina y endorfina antes de
ser transmitida a una neurona productora de GnRH (Findlay et al., 2004).

32 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
La GnRH se libera de una manera pulsátil y continua debido a múltiples comunicaciones
de retroalimentación. Esta forma de secreción impide entre otras que las células diana
del lóbulo anterior de la hipófisis internalicen sus receptores de la membrana celular lo
que provocaría que la molécula de GnRH perdiese su actividad. Seguido a su liberación,
la GnRH pasa hacia los capilares del sistema porta hipofisiario y de allí a las células de la
hipofisis en la porción anterior (adenohipofisis), en donde se une al receptor de la célula
gonadotrofa ubicado en la membrana celular para producir cambios moleculares (Crowe
et al., 2001).
En la adenohipoisis se producen varios tipos de hormonas, entre la cuales la LH y la FSH
cumplen papel importante en la regulación neuroendocrina del ciclo estral. Su
mecanismo de acción se ejerce a través de la unión de la hormona a receptores ubicados
en la membrana celular de las células diana y estimulación del adenilato ciclasa como
segundo mensajero (Canty et al., 2006).
Las gonadotropinas hipofisiarias FSH y LH, reducen su propia síntesis y secreción
mediante un bucle corto de retroalimentación (Short feedback loop) que inhibe la
secreción de la GnRH; y finalmente la GnRH segregada reduce su propia síntesis al
retroalimentar a sus propias neuronas (ultra – short feedback loop) (Webb et al., 2004).
Cuando las moléculas de GnRH alcanzan las células gonadotrofas de la adenohipófisis,
estas segregan FSH y LH, las cuales se almacenan en forma de gránulos en el
citoplasma de la célula gonadotrofa (la síntesis y almacenamiento van variando durante
el ciclo).Tras recibir la señal de la GnRH a través de los receptores, se transportan hasta
la membrana plasmática a través de la cual se segregan los gránulos, es decir, además
de la síntesis también la secreción de gonadotrofinas está regulado por el control de la
GnRH; si el suministro de GnRH a la adenohipófisis es insuficiente, no solamente se
reducirá la secreción, sino también la síntesis de gonadotropinas (Palma, 2008).
La GnRH es capaz de sensibilizar su propia célula diana, y solamente se consigue esta
sensibilización de las células de la adenohipófisis cuando aumenta la concentración de
estrógenos en plasma sanguíneo; con lo cual se consigue secreción de grandes
cantidades de LH lo que provoca la ovulación (Austin et al., 2001).

Capítulo 1 33
En las primeras fases foliculares cuando la concentración de estrógenos aún es baja, se
almacena y segrega poca cantidad de FSH y LH (retroalimentación negativa). A medida
que aumenta la concentración de estrógenos, la secreción de FSH y LH aumenta
lentamente así como su almacenamiento. En esta fase del ciclo, el estrógeno, incluso a
bajas concentraciones, tiene un efecto positivo sobre la síntesis y el almacenamiento de
gonadotropinas. Simultáneamente el efecto negativo de los estrógenos sobre la
secreción de la FSH y LH impide que las gonadotropinas almacenadas se segreguen
antes de hora (Stocco et al., 2007).
A medida que va avanzando la fase folicular, la mayor concentración de estrógenos
sensibiliza las células gonadotropas a los pulsos de GnRH que se producen con mayor
intensidad. Cada impulso de GnRH no solamente provoca la secreción de los gránulos
que en ese momento se encuentran junto a la membrana, también se activa la reserva
intracelular de gránulos de gonadotropina para el siguiente pulso. Este proceso también
se apoya con la multiplicación de los receptores de GnRH, lo cual también es
dependiente de los estrógenos (Webb et al., 2004).
A medida que avanza la maduración folicular, la concentración de estrógenos en sangre
aumenta hasta alcanzar un umbral crítico que se mantiene durante un periodo también
crítico. En este periodo aumenta también la frecuencia y amplitud de GnRH. A cada pulso
que va llegando se responde con mayor intensidad que el anterior, por que se dispone
de más cantidad de FSH y LH almacenada. Todo el sistema se apoya en el efecto
positivo de los estrógenos sobre la movilización de los depósitos de gonadotropinas y la
síntesis adicional de receptores de GnRH. Por tal motivo se observa la secreción de
grandes cantidades de LH (Revisado por Grajales y Tovío 2009).
El folículo en fase de maduración sigue produciendo estrógenos y adicionalmente
inhibina; estas dos, inhiben la secreción de FSH (retroalimentación negativa), por lo que
se observa que durante la secreción preovulatoria de gonadotropinas se segrega
esencialmente LH (retroalimentación positiva). De esta forma se puede pensar que el
propio folículo determina mediante su concentración de estrógenos si ovula o no (Palma,
2008).

34 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
1.4.1 Implicación de las hormonas esteroides durante el ciclo estral No solamente son producidas por las gónadas, las glándulas suprarrenales y la placenta
secretan igualmente esteroides. Estas hormonas se dividen en tres grupos de acuerdo al
número de átomos de carbono que presentan: andrógenos, estrógenos y progestágenos;
el ovario puede sintetizar estos tres tipos de hormonas, las cuales poseen un núcleo
básico conocido como anillo ciclopentanoperhidrofenantreno (Stevenson, 2001). Un
esteroide de 18 carbonos tiene actividad de estrógeno, uno de 19 carbonos tiene
actividad de andrógeno (esteroide de 19 carbonos con un hidroxilo o un oxígeno en las
posiciones 3 y 17 y un enlace doble en la posición 4) y uno de 21 tiene actividad de
progestágeno (Knight and Glister, 2003).
En el transcurso de la hormogénesis esteroidea del ovario, el número de átomos de
carbono de la estructura del colesterol o de cualquier otra hormona esteroidea puede
reducirse, pero nunca aumentarse. De allí se entiende que todos los estrógenos ováricos
se producen a partir de precursores androgénicos (Stocco et al., 2007).
La síntesis de todos los esteroides depende del transporte del colesterol (esteroide de 27
carbonos) a la mitocondria y de la membrana mitocondrial externa a la interna, en donde
el complejo enzimático rompe la cadena del colesterol para separar la cadena lateral
convertida en pregnenolona la cual tiene 20 carbonos, que a su vez se convierte en
andrógenos y en estrógenos (Niswender, 2002).
Bajo condiciones normales la mayoría del colesterol es sintetizado en el hígado y
transportado a los tejidos esteroidogénicos (corteza adrenal, folículo, cuerpo lúteo) en
forma de lipoproteínas de alta y baja densidad (HDL y LDL), las cuales son la fuente más
común del colesterol utilizado por el CL para la producción de hormonas esteroides
(Knight and Glister, 2003).
El consumo de lipoproteínas de baja densidad por parte de las células luteales se
presenta bajo el esquema de endocitosis mediada por receptores; el mecanismo por el
cual las células luteales captan lipoproteínas de alta densidad aún no es conocido
claramente (Stocoo et al., 2007).

Capítulo 1 35
El paso limitante en la vía esteroidogénica parece ser el transporte de colesterol de la
membrana mitocondrial externa a la interna, este paso también parece ser el principal
sitio de regulación positivo y negativo de la esteroidogénesis por el sistema de segundos
mensajeros (Stocco et al., 2007).
Se ha propuesto que la proteína reguladora de la esteroidogénesis aguda (StAR),
receptores mitocondriales de benzodiacepina y sus ligandos endógenos, al parecer son
requeridos para el transporte normal del colesterol desde la membrana mitocondrial
externa a la interna en donde sucede la ruptura de la cadena del colesterol (Niswender et
al., 2000).
Figura 1. Algunos genes implicados en la síntesis de progesterona, y estradiol (Tomado de Stocco et al., 2007).
Las vías biosintéticas en todos los órganos endocrinos que producen hormonas
esteroides son similares, solamente se diferencian por los sistemas enzimáticos que
contienen. El ovario se distingue de las glándulas adrenales en que carece de 21 –
hidroxilasa y 11 – β hidroxilasa; por tal motivo el tejido ovárico normal no puede sintetizar
glucocorticoides ni mineralocorticoides (Niswender, 2002).

36 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
1.4.1.1. Vías implicadas en la síntesis de estrógenos De todos los esteroides, los estrógenos producidos por los folículos ováricos tienen el
rango más amplio de funciones fisiológicas. Entre estas se destaca la acción sobre el
sistema nervioso central para inducir signos de celo, la acción en el útero para aumentar
la amplitud y frecuencia de las contracciones potencializando los efectos de la oxitocina y
la PGF2α y la participación en el desarrollo caracteres secundarios femeninos y control
de retroalimentación negativa y positiva en la liberación de LH y FSH a través del
hipotálamo (Sartori and Barros 2011).
En el modelo que explica su síntesis se observa que la LH interacciona con su receptor
ubicado en las células de la teca interna productoras de androsteniediona a partir de la
progesterona y ésta a partir del colesterol, fenómeno en el cual intervienen las enzimas
P450 scc (Enzima de clivaje de la cadena lateral de colesterol – la cual tiene acción
20,22 desmolasa: por lo cual rompe los dobles enlaces y reduce la molécula del
colesterol) y 3β – hidroxiesteroide deshidrogenasa (3β-HSD). La androsteniediona pasa a
través de la membrana basal a las células de la granulosa en donde es aromatizada a
partir de las enzimas P450 arom (Tiene acción aromatasa - interviene en el paso de
andrógenos a estrógenos) y P450 17 – α (Tiene acción 17 -hidroxilasa y 17,20 liasa),
convirtiéndose de esta forma en estradiol. Este estrógeno pasa al líquido folicular y de allí
a la circulación general para posteriormente llegar a su órgano blanco y ejercer múltiples
efectos (Mollo et al., 2007).
A medida que el folículo dominante sigue creciendo la FSH además de muchos efectos
inducirá la síntesis de receptores para la LH en la célula de la granulosa, aumentando
aún más la producción de andrógenos que luego son transformados en estrógenos por
parte del folículo dominante (Rosales y Guzmán, 2008; Revisado por Olivera, et al.,
2007).
Los estrógenos son transportados por proteínas de enlace en la circulación, como la
globulina fijadora de las hormonas sexuales (SHBG), (específicamente la β2 -globulina)
para poder llegar a los órganos blanco, entre los cuales se encuentra, el sistema nervioso
central (SNC), la vulva, la vagina, el útero y el oviducto. En el SNC se estimula la
conducta de celo y en el hipotálamo ejercen un ´´feed back´´ negativo sobre el control de

Capítulo 1 37
la actividad tónica y positivo sobre la actividad cíclica. En la vulva y vagina se produce un
aumento de flujo sanguíneo (hiperemia), congestión y aumento de color, como también
extravasación de líquido (agua y sales) al espacio extracelular con la consecuente
aparición de edema. En el útero los estrógenos actúan como hormona del crecimiento
produciendo proliferación de las células y glándulas endometriales las cuales aumentan
su secreción. En el miométrio se observa hipertrofia de la capa muscular circular y
longitudinal y se sensibilizan sus células a la acción de la oxitocina, por lo que se
favorece la contractilidad y conductibilidad de las mismas. En el cérvix se produce
relajación, aumenta su diámetro y aparece abundante secreción mucosa, filante y
transparente. En el oviducto se incrementa la actividad del músculo liso y de la secreción
de un líquido seroso junto con estimulación de la división celular (Fernandes et al., 2001).
Durante la fase folicular, los estrógenos ejercen efectos de retroalimentación positiva en
el control de la liberación de LH y FSH en la adenohipófisis, esto ocurre por la
estimulación ejercida por esta hormona esreroidea en el hipotálamo para la liberación de
la GnRH. Durante la fase luteal, es decir cuando tenemos un CL funcional y por lo tanto
altos niveles de P4 circulante, se causa inhibición en la secreción de GnRH (Stocco, et al.,
2007).
Existen efectos no reproductivos causados por los estrógenos en los cuales se incluye la
estimulación de la asimilación del calcio y la osificación de los huesos, causan
maduración del cartílago intersticial de los huesos largos e inhiben su crecimiento.
También originan un efecto anabólico de ganancia de peso y de crecimiento; el posible
mecanismo de acción de este fenómeno parece estar basado en la estimulación por los
estrógenos de la liberación adicional de hormona de crecimiento a partir de la hipófisis
(Palma, 2008).
1.4.1.2. Vías implicadas en la síntesis de progesterona La P4 es el progestágeno natural más importante, el cual es secretado por la placenta,
las glándulas suprarrenales y por las células luteales; estas últimas por estimulo de la
actividad luteotrópica de la LH y en menor proporción por otras hormonas como la GnRH,
la prolactina, GH y la catecolamina (Revisado por Olivera et al. 2007).

38 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Durante el pico preovulatorio de la LH, las células de la granulosa adquieren la capacidad
de producir progesterona y pierden la capacidad de producir estrógenos debido a la
inhibición de la enzima aromatasa P450 arom y P450 17 - α, este fenómeno de
diferenciación celular es conocido como luteinización (Rosales y Guzmán 2008).
El mecanismo por el cual se estimula la secreción de P4 involucra la unión de la LH a su
receptor (receptor con siete segmentos transmembranales – receptores acoplados a
proteína G), con lo cual se activa el sistema adenilato ciclasa, lo que convierte el ATP en
AMP cíclico (AMPc), por lo cual se activa el sistema proteína kinasa A (PKA), con lo cual
se desarrolla la fosforilación de las enzimas involucradas en la esteroidogénesis como la
colesterol-esterasa, lo que mejora el transporte de colesterol desde el citoplasma a la
membrana mitocondrial externa (el colesterol no pueden difundir libremente en el citosol
y llegar a la mitocondria sin antes unirse a proteínas transportadoras como la proteína
transportadora de esteroles - SCP-2), y de allí a la membrana mitocondrial interna de las
células luteales pequeñas (Niswender et al., 2000). En este sitio por acción de la enzima
P450 scc, la adrenodoxina y la adrenodoxina reductasa, ocurre la ruptura de la cadena
lateral del colesterol para producir pregnenolona, la cual es transportada al retículo
endoplasmático liso que está asociado a la mitocondria, en donde la 3β –
hidroxiesteroide deshidrogenasa (3β-HSD) participa en la conversión de pregnenolona a
P4 (Barros et al., 2009); esta última es liberada de la célula mediante un proceso de
difusión (Rosales y Guzmán 2008).
Estudios en bovinos han evidenciado que el IGF-I y la GH incrementan la secreción de P4
por parte de los tejidos luteales. Referente a ésto se ha encontrado mRNA que codifica
para receptores de GH en el tejido luteal de bovinos. Esta hormona puede tener un
efecto directo en la función luteal a través de la unión a su receptor y la activación de la
enzima tirosina kinasa asociada a membrana (JAK2). Igualmente la GH podría
influenciarla función lútea indirectamente por el incremento en la expresión del IGF-I, el
cual estimula la secreción de P4 a través de la modificación del citoesqueleto y puede
estar involucrado en la prevención de la muerte celular ayudando a mantener el peso
luteal (Niswender et al., 2000; Schams and Berisha, 2004).
Se ha sugerido que las prostaglandinas I y E (PGI, PGE) se producen en mayores
cantidades durante la fase luteal temprana y podrían jugar un papel importante en el

Capítulo 1 39
desarrollo luteal. Estas prostaglandinas aumentan los niveles de AMPc, el cual activa la
PKA, por lo cual se aumenta la síntesis de P4 (Niswender et al., 2000).
Figura 2. Biosíntesis de la Progesterona. (Tomado de Niswender, 2002)
Posterior a su síntesis la P4 es transportada en la sangre por una globulina de enlace de
hormonas sexuales (SHBG), específicamente la globulina fijadora del cortisol (CGB). Su
acción se observa después de que el tejido blanco ha estado expuesto durante cierto
tiempo a la estimulación de los estrógenos. A nivel de hipotálamo ejerce un efecto de
´´feed back´´ negativo sobre el control de la actividad tónica de la secreción de GnRH
(Hafez, 2000). En el endometrio causa aumento de espesor del epitelio (Columnar y
alto) y las glándulas uterinas alcanzan su máximo desarrollo (largas, enrolladas y
ramificadas). Las glándulas son activamente secretoras; producen un líquido formado por
proteínas séricas y específicas del útero, glucoproteínas y minerales, los cuales son
esenciales para la nutrición del embrión durante la fase de preadhesión. En el miometrio
inhibe las contracciones permitiendo que se lleve a cabo la gestación y en el cérvix se
produce la formación de un tapón mucoso formado por una sustancia densa, opaca y de
poca cantidad, esto prácticamente transforma y convierte al útero en una cámara de
incubación (Palma, 2008).

40 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Resumiendo la acción principal de la P4 se sabe que prepara el endometrio para la
implantación embrionaria y mantenimiento de la preñez, actúa en relación con los
estrógenos para inducir el comportamiento estral, desarrolla tejido secretor (alveolos) en
la glándula mamaria y en concentraciones mayores a 1ng/ml inhibe el estro y la oleada
ovulatoria de LH (Kayser et al., 2006).
1.4.2. Fases del ciclo estral - Proestro
Se caracteriza por comenzar con la regresión del cuerpo lúteo (CL) del ciclo anterior y
finaliza con los primeros signos que se manifiestan durante el celo. Su rango de duración
es aproximadamente de 3 días (Knobil, and Neill, 2006)
Si la vaca no llega a quedar preñada, hacia los días 10 y 15 del ciclo estral comienza a
disminuir la producción de progesterona (P4) y aumenta la producción de estrógenos (E2)
(producidos por los folículos en crecimiento), lo que induce a su vez la producción de
oxitocina (OX) (producida en las células del hipotálamo y del cuerpo lúteo (Wiltbank,
2001); tal situación estimula el clivaje de ácido araquidónico, el cual se transforma bajo
complejidad de mecanismos en PGF2α, la cual es producida en el epitelio endometrial y
en menor proporción por el cuerpo lúteo (Niswender et al. 1994). Esta hormona es vertida
a la vena uterina, de donde por difusión pasa a la arteria ovárica y por un mecanismo de
contracorriente alcanza al CL, en donde se une a receptores ubicados en las células
luteales grandes y causa aumento del mRNA que codifica para la prostaglandina
sintetasa – 2 (PGHS – 2), necesaria para la producción de PGF2α luteal, posiblemente
necesaria para completar el proceso de la luteolisis (La oxitocina luteal y Prostaglandina
uterina están interrelacionadas mediante un doble “Feed back” positivo el cual permite
desencadenar la luteolisis) (Carvalho etal., 2008). Allí en las células luteales, gracias a
eventos intracelulares (Disminución en la fluidez de las membranas, disminución de
antioxidantes, aumento en la formación de radicales superoxidos y aumento en de la
actividad de la fosfolipasa y enzimas proteolíticas), se presenta la muerte por apoptosis
de las células mediante el desencadenamiento de cascadas de señalización que
involucran hormonas como la prolactina; citoquinas como el Factor de Necrosis Tumoral
alpha (TNFα), el Interferón gamma (IFNγ) y el Fas ligando (FasL); especies reactivas de
oxígeno (ROS), endotelina-1(ET-1) (Péptido vasoconstrictor de 21 aminoácidos

Capítulo 1 41
producido por las células endoteliales el cual aumenta luego de inyectar PGF2α en el CL
en regresión y en las venas ováricas, inhibiendo la secreción de P4 de las células luteales
cultivadas in vitro), la proteína HSP70 de choque térmico y macrófagos, los cuales
podrían estar involucrados en un proceso de fagocitosis (Olexikova et al. 2010;
Rosenkrans et al., 2010).
Hehnke – Vagnoni etal., (1995) propusieron que el factor de necrosis tumoral (TNFα)
interviene en la remoción de desechos y células degeneradas, participando de esta forma
en la última fase de la disminución de la P4. Al ingresar células productoras del TNFα
(Leucocitos y macrófagos) en el CL podría explicarse el aumento de TNFα que ocurre
durante la regresión luteal (Sartorelli et al., 2005).
Girsh et al., (1996), encontraron que la PGF2α estimula la expresión del mRNA para ET–
1 en cultivo de células endoteliales derivadas de la microvasculatura del CL bovino,
dando base a la hipótesis que la ET – 1 luteal es un mediador promotor luteolítico local
en el proceso de regresión lútea.
Al proceso de luteolisis se ha asociado un aumento en la expresión de la Proteína 1 –
Quimiotáctica para Monocitos (MCP – 1) y la infiltración de monocitos/macrófagos
(Niemann et al., 2003). Benyo y Pate, (1992) observaron una rápida inducción de mRNA
para el MCP – 1 después de un tratamiento con PGF2α, lo que podría inducir un
reclutamiento de las células inmunes involucradas en la remoción de células no
funcionales durante la regresión del CL. Estas células inmunes secretan citoqinas, las
cuales disminuyen la esteroidogenesis de las células luteales y su viabilidad, en
consecuencia se produce una regresión funcional caracterizada por una disminución de
P4, y una regresión estructural determinada por la degradación de tejido (Sangsritavong
et al., 2002). En el primer caso, el efecto antiesteroidogenico estaría mediado por el
sistema del segundo mensajero de la proteína quinasa C (PQC); en el segundo, el
efecto luteolítico (Perdida de tejido luteal) estaría mediado por el aumento de la
concentración del Ca ++ libre intracelular (En el momento de la regresión luteal se
presenta una rápida disminución en el peso del cuerpo lúteo, en el contenido de ADN y
en el tamaño de las células). (Wiltbank, 2001). Este calcio estimularía endonucleasas que
fragmentan el ADN, mecanismo común en los procesos de apoptosis e importante en la
regresión del CL en la vaca y en la oveja (Hussein, 2005).

42 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Se ha sugerido que la LH podría estar participando también en el proceso de luteolisis,
ya que se han detectado sitios de fijación para esta hormona en el endometrio bovino y
un aumento en la secreción endometrial y luteal de PGF2α en respuesta a esta hormona
durante el periodo luteolítico (Juengel et al., 1993). Igualmente se han detectado
receptores para la LH en la vena uterina los cuales se evidencian por un aumento en la
expresión de la ciclooxigenasa y la producción de PGF2α y E2 por la vena en respuesta a
la LH (Shams y Berisha, 2004).
Figura 3. Cascada luteolítica en la vaca (Tomado de Shams and Berisha 2004)
La disminución de la P4 a niveles inferiores a 1 ng./ml., suprimen el “feed back” negativo
sobre la secreción de gonadotropinas. Consecuentemente se produce un aumento en la
frecuencia pulsátil en los niveles de LH y en menor magnitud de FSH. Por otro lado la
hipófisis aumenta la respuesta a GnRH. El aumento en la frecuencia pulsátil de las
gonadotropinas durante la fase folicular del ciclo estral estimula el desarrollo de un
folículo ovárico; a medida que este se desarrolla, se produce un aumento de los niveles
de estradiol que podrían provocar una disminución de los niveles basales de FSH y un
estimulo en la secreción de LH. Los niveles de estradiol continúan aumentando y cuando
se alcanza cierto nivel se estimula la receptividad al macho y comienza el periodo de
estro (Thatcher et al., 2002).

Capítulo 1 43
- Estro
Esta etapa involucra todos los cambios que permiten la ovulación y el comienzo de la
formación del CL.
Cuando la GnRH es liberada, pasa hacia los capilares del sistema porta hipofisiario y de
allí a las células de la hipófisis en la porción anterior (adenohipofisis), en donde se une a
receptores ubicados en la membrana celular de las células gonadotrofas para producir
cambios moleculares que contribuyen a la producción de las hormonas LH y FSH, las
cuales cumplen un papel importante en la regulación neuroendocrina del ciclo estral.
Estas dos hormonas se almacenan en forma de gránulos en el citoplasma de la célula
gonadotrofa y de allí posteriormente se segregan tras seguir recibiendo la señal de la
GnRH; es decir, además de la síntesis también la secreción de gonadotropinas está
regulada por el control de la GnRH. Si el suministro de GnRH a la adenohipófisis es
insuficiente, no solamente se reducirá la secreción, sino también la síntesis de
gonadotropinas, las cuales regulan el desarrollo folicular, con lo cual comienza a
aumentar la concentración de estrógenos (Shams y Berisha, 2004).
En las primeras fases foliculares cuando la concentración de estrógenos aún es baja, se
almacena y segrega poca cantidad de FSH y LH (retroalimentación negativa). A medida
que aumenta la concentración de estrógenos, la secreción de FSH y LH aumenta
lentamente así como su almacenamiento. En esta fase del ciclo, el estrógeno incluso a
bajas concentraciones, tiene un efecto positivo sobre la síntesis y el almacenamiento de
gonadotropinas (Revisado por Grajales, 2011).
Con el aumento gradual de concentración estrogénica comienzan a aparecer los
primeros signos de celo, estro o calor. Durante esta etapa, los estrógenos en altas
concentraciones provocan un pico de la GnRH y en consecuencia un pico preovulatorio
de LH, este es el resultado del aumento en la frecuencia y amplitud de los pulsos de
dicha hormona (Revisado por Grajales y Tovío, 2009).
La FSH disminuye su secreción consecuencia del “feed back” negativo estrogénico y de
la inhibina. Posteriormente del pico de LH, 4 a 12 horas se produce incremento en la
concentración basal y amplitud de los pulsos de FSH, relacionándose esto con la primera
onda de crecimiento folicular. Luego de 12 a 24 horas de comenzado el celo, el sistema

44 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
nervioso se torna refractario al estradiol y cesan todas las manifestaciones psíquicas del
mismo (Shams y Berisha, 2004).
El estro se caracteriza por la receptividad de la hembra hacia el macho, dejándose
montar por éste y por otras hembras (conducta homosexual). Esta etapa tiene un rango
de duración de 2 a 24 horas, en donde actúa la hormona estradiol, la cual conlleva a que
la vaca presente cambios en el tracto reproductivo y alteraciones de conducta,
destacándose entre estos la inquietud, ansiedad, pérdida de apetito, descarga vaginal de
moco con mínima viscosidad formado por filamentos de diferente grosor que forman una
estructura tridimensional en forma de red cuyo olor atrae al macho, la vulva se
edematiza, se produce aumento de tono miometrial en el útero y el canal cervical sufre
ablandamiento y apertura, lo cual es regulado por la oxitocina y la P4. En referencia a
esto último, Fuchs et al., (1996) comenta que las prostaglandinas E2 y E1 podrían estar
involucradas también en el mecanismo de reblandecimiento cervical.
- Metaestro
El metaestro corresponde al periodo inmediatamente posterior al estro, cuyo promedio de
duración es de 6 días. Durante esta etapa ocurre la ovulación (ruptura de la pared
folicular y salida del ovocito con la finalidad de ser fecundado por el espermatozoide), lo
que desencadena en la formación del llamado cuerpo hemorrágico, que posteriormente
se convierte en cuerpo lúteo y cuerpo lúteo funcional, estructura fundamental en muchos
de los procesos reproductivos y en cada etapa fisiológica, incluyendo la ovulación, la
duración del ciclo estral, el reconocimiento de la preñez y la sobrevivencia embrionaria en
la mayoría de las especies mamíferas (Revisado por Grajales y Tovío, 2009).
El folículo en fase de maduración sigue produciendo estrógenos y adicionalmente otra
hormona llamada inhibina, las cuales, suprimen la secreción de FSH (retroalimentación
negativa), por lo que se observa que durante la secreción preovulatoria de
gonadotropinas se segrega esencialmente LH (retroalimentación positiva). De esta forma
se podría pensar que el propio folículo determina mediante su concentración de
estrógenos si ovula o no (Revisado por Olivera et al., 2007).
El pico preovulatorio de LH es una señal que causa despolarización de la membrana
plasmática de uno o más de los tipos celulares presentes en el folículo ovárico maduro. A
medida que el folículo avanza hacia su ovulación, se produce una inflamación más

Capítulo 1 45
aguda, aumenta la despolarización de las células foliculares y se generan potenciales de
acción y cambios en la concentración de Ca++ citosólico que contribuirían a la actividad
degradativa que disminuyen el espesor de la pared folicular (Carvalho et al., 2008).
Poco antes de la ovulación, durante el pico preovulatorio de la LH, las células de la
granulosa que recubren el folículo preovulatorio adquieren la capacidad de producir P4 a
partir de colesterol y pierden la capacidad de producir estrógenos debido a la inhibición
de los factores necesarios para tal fin como lo son las enzimas aromatasas (Revisado por
Olivera et al., 2007).
Con la luteinización se producen cambios endocrinológicos y bioquímicos que permiten
que las células foliculares se transformen en luteales. La membrana basal, ubicada entre
las células de la granulosa y de la teca interna, pierde su funcionalidad y los capilares
invaden y forman una red en el antro folicular. Las células de la teca se hipertrofian,
hiperplasian y migran hacia el interior de la cavidad folicular, dispersándose entre las
células de la granulosa luteinizadas. (Hafez, 2000).
Las células foliculares sufren una reprogramación irreversible que las van sometiendo al
proceso de luteinización, en el cual dejan de dividirse y comienzan a expresar un nuevo
conjunto de moléculas que le permiten a las células sobrevivir en un ambiente hormonal
nuevo. Por lo tanto, la función de estas células depende de una determinada
combinación de genes que codifican para proteínas reguladoras y de señalización,
receptores y factores de transcripción (Stocco et al., 2007).
La ovulación es un proceso que ocurre de 10 a 20 horas despoés de la terminación del
celo, y es desencadenado por el pico preovulatorio de la LH, lo que provoca como primer
evento un aumento en el flujo sanguíneo en el folículo ovulatorio, mediado posiblemente
por agentes vasoactivos tales como histamina, quininas, prostaglandinas y
lipooxigenasas. Posterior a la ovulación se produce edema en las células de la teca
interna y externa, se disocia el cúmulos oophorus, se interrumpe la inhibición de la
meiosis y se elimina el primer corpúsculo polar (Hafez, 2000).
Posterior a la ovulación, la cavidad antral del folículo se llena de sangre, por tal razón a la
estructura se le denomina cuerpo hemorrágico, de allí se forma el CL a partir de la

46 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
hipertrofia de las células de la granulosa y de la teca que por su cambio morfológico
comienzan a denominarse células luteales grandes y células luteales pequeñas,
respectivamente. El órgano formado tiene una estructura maciza y puede o no presentar
una cavidad central no luteinizada (Niswender et al, 2000; Rathbone et al., 2001).
Entre dos a tres días post-ovulación, el CL desarrolla una intensa angiogénesis
(formación de vasos sanguíneos), lo que lo convierte, proporcionalmente, en uno de los
órganos más vascularizados del organismo (Revisado por Olivera, et al., 2007). La
intensidad de los procesos angiogénicos dentro del CL alcanzan su pico 2 a 3 días
después de la ovulación, así la mayoría de las células esteroidogénicas del CL maduro
están en contacto con uno o más capilares sanguíneos los cuales se multiplican para dar
soporte al desarrollo de nuevas células luteales, lo cual parece ser potenciado localmente
por la angiotensina II y por factores de crecimiento que inducen angiogénesis como el
VEGF; FGF y IGF, con lo cual se da soporte a la síntesis de P4 en las células luteales
(Acosta and Miyamoto, 2004; Berisha and Schams, 2005).
Este CL es considerado como una glándula endocrina transitoria, cuyo principal producto
de secreción es la P4, la cual participa en múltiples procesos como el reconocimiento, la
adhesión e implantación del conceptus, el mantenimiento de la gestación en sus estadios
tempranos y la regulación de la dinámica folicular (Revisado por Olivera et al., 2007).
A medida que aumenta el proceso de ovulación, aumenta la actividad de las enzimas,
activador del plasminógeno, kalicreinas y colagenasas. Por último se produce la ruptura
del folículo, lo cual sería dependiente de la activación de los fibroblastos en el tejido
conectivo de la teca del folículo maduro. La PGF2α podría estar produciendo
contracciones ováricas que provocaría la ruptura, el folículo se contraería bajo el mismo
efecto de la prostaglandina y así se liberaría el ovocito (Revisado por Grajales et al,
2011).
La actividad degradativa durante este proceso se caracteriza por un aumento de la
actividad de la colagenasa la cual provendría de los fibroblastos de la teca externa. Es
probable también que eosinófilos y macrófagos migren por medio de un efecto
quimiotáctico hacia el tejido ovárico inflamado y contribuyan con su actividad hidrolítica.
Además el aumento en la actividad de sustancias similares a las kalicreinas que ocurre

Capítulo 1 47
en el folículo ovulatorio contribuiría a la degradación del tejido conectivo folicular. Las
Kalicreínas generan kalidina y kininas las cuales como se mencionó provocan la
respuesta vascular característica de los tejidos inflamados y estimulan a las colagenasas
latentes. A medida que avanza el proceso de degradación del tejido conectivo que
separa el folículo de la superficie ovárica se inicia la formación del estigma (Área
extremadamente delgada del ápice folicular (Hafez, 2000).
Otras sustancias involucradas en el proceso ovulatorio son los ácidos grasos derivados
del ácido araquidónico por la vía enzimática de la lipoxigenasa (leucotrienos) o de la
ciclooxigenasa (prostaglandinas). La prostaglandina E2 estimula el activador del
plasminógeno que permite el paso del plasminógeno, ubicado en el fluido folicular y en el
edema extracelular a plasmina. Esta ejerce su efecto sobre la colagenasa latente
transformándola en colagenasa. La PGF2α provoca ruptura de los lisosomas con la
liberación de las enzimas contenidas en su interior, las que sumadas a la actividad de las
plasmina, culminan con la formación del estigma (Acosta and Miyamoto, 2004).
En la regulación de la síntesis de las prostaglandinas participan la testosterona y la P4,
sintetizadas en las células luteales. Esta última se ha determinado que solo es necesaria
en la primer mitad del proceso ovulatorio (Acosta and Miyamoto, 2004).
Por último se produce la ruptura del folículo, lo cual sería dependiente de la activación de
los fibroblastos en el tejido conectivo de la teca del folículo maduro. La PGF2α podría
estar produciendo contracciones ováricas que provocarían la ruptura, el folículo se
contraería bajo el mismo efecto de la prostaglandina y así se liberaría el ovocito
(Revisado por Tovío y Grajales, 2009).
- Diestro
Este periodo tiene un rango de duración de 10 a 12 días. Se caracteriza por la
consolidación del cuerpo lúteo funcional, el cual comienza a producir concentraciones
altas de P4, con la finalidad de provocar cambios cualitativos y cuantitativos en el medio
ambiente uterino, controlando la síntesis y secreción de proteínas, por lo cual se
mantienen las funciones necesarias para el desarrollo embrionario temprano; esto con la
finalidad de llevar a cabo la implantación, placentación, y el exitoso desarrollo fetal
(Rathbone et al., 2001).

48 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
De acuerdo a lo anterior se asume que deficiencias de P4 podrían causar que el
endometrio llegue a ser deficiente en cuanto a la nutrición histotrópica la cual es la
primera fuente disponible para el crecimiento, mantenimiento y supervivencia del
conceptus (embrión con todas sus membranas asociadas) (Revisado por Gonella, 2010).
Durante el diestro temprano la P4 proveniente del CL recientemente formado estimula la
acumulación de fosfolípidos en las células del epitelio del lumen endometrial y el epitelio
glandular superficial que pueden liberar ácido araquidónico para la posterior síntesis y
secreción de PGF2α (Spencer et al., 2004a).
Durante la fase luteal media la LH se secreta en menor frecuencia pulsátil y mayor
amplitud comparada con la fase luteal temprana (Castilho et al., 2007). En la regulación
de su secreción interviene la P4, estradiol y los neuropéptidos opioides, los cuales
combinan sus efectos para suprimir la frecuencia pulsátil de dicha gonadotropina. Esta
hormona interviene en el desarrollo y mantenimiento del CL, y su secreción pulsátil es
necesaria para mantener los niveles de P4 durante los primeros 12 días del ciclo estral,
(Revisado por Grajales y Tovío, 2009). Posteriormente esa secreción pulsátil no sería
necesaria y sería regulada por bajas concentraciones de LH o por mecanismos
independientes de dicha hormona (Wiltbank, 2001).
El pico preovulatorio de LH provoca la luteinización de las células de la teca y granulosa,
y altera la vía esteroidogénica para que la P4 sea la principal hormona esteroide
producida por este tipo de células (Niswender et al., 2000).
El mecanismo por el cual la LH estimula la secreción de P4 por parte de las células
luteales pequeñas involucra la formación de AMPc y la activación de la proteína quinasa
A. En el caso de las células luteales grandes éstas incrementan su tamaño, pero se
mantienen constantes en cantidad; aún no es claro el mecanismo de acción utilizado por
la LH en estas células. Se sabe que estas células expresan mRNA para IGF – I y tienen
receptores para IGF – I. También se conoce que responden al estímulo extremo de IGF
– I con un aumento en la secreción de P4 (Wiltbank, 2001).

Capítulo 1 49
Se plantea que varios tipos de genes son inducidos debido al pico de LH, los cuales
intervienen en la ovulación y la inducción hacia la luteinización. Entre estos genes se
encuentran los que codifican para receptores de P4 (Spencer et al., 2004b), potenciador
vinculante de proteína β (C/EBP β), factor de crecimiento de respuesta temprana (Egr-1)
(también conocido como NGFI-A) (Revisado por peña et al., 2007; Espey et al., 2000),
receptor nuclear 77 (Nur77) (también conocido como NGFI-B) (Park et. al., 2001),
ciclooxigenasa 2 (COX-2) (Lim et al., 1997), P450arom y P450scc. Esta última se
mantiene expresada mucho tiempo después del comienzo de la luteinización, mientras
que la COX-2 y P450 arom declinan varias horas después del pico de LH (Burkhart et al.,
2010).
De acuerdo a lo anterior se ha sugerido que C/EBP β, da lugar a cuatro isoformas de
proteínas; proteína enriquecida activadora del hígado – LAP (de 38 y 34 kDa
respectivamente) y proteína enriquecida inhibidora del hígado LIP (21 y 14 kDa
respectivamente), esta última actúa como inhibidor de la acción de la LAP, ejerciendo
función supresora transcripcional en las células para la expresión de COX-2 y P450
arom; esto sucede solamente durante el proceso de luteinización, lo cual sugiere que
este proceso no está involucrado en el mantenimiento de la función lútea (Wiltbank,
2001).
La FSH y la Prostaglandina I2 (PGI2) también podrían estar interviniendo de cierta forma
en la síntesis de la P4. La FSH se uniría a receptores ubicados en el CL y provocaría un
aumento en la secreción de P4. La PGI2, por su parte, estimularía a las células luteales
para la síntesis de P4, y aumentaría el flujo sanguíneo en el ovario con un estímulo
positivo sobre la síntesis y secreción de dicha hormona (Burkhart et al., 2010).
El cuerpo lúteo no solamente sintetiza progesterona, otras hormonas y factores son
producidos por esta estructura ovárica, incluyendo la oxitocina, relaxina, factor de
crecimiento insulínico (IGF) - 1, y prostaglandinas entre otros. (Revisado por Tovío y
Grajales, 2009).
Se ha encontrado que la noradrenalina estimula la secreción de oxitocina y P4 en la mitad
de la fase luteal mediante beta adrenoreceptores ubicados en el CL del bovino,

50 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
específicamente en las células luteales pequeñas. No obstante la secreción de oxitocina
ocurriría por parte de las células luteales grandes (Dos Santos et al., 2009).
Si la vaca llega a quedar preñada, se presenta hacia el final de la mitad de la fase luteal
del ciclo estral disminución de los receptores de progesterona (P4) en el útero, por lo cual
comienzan a disminuir la concentración de esta hormona (regulación en baja),
desencadenando los factores necesarios para causar luteolisis (lisis del cuerpo lúteo)
(Gabriel et al., 2011). En ese momento el interferón tau (IFN-τ) producido por las células
mononucleares del trofoblasto embrionario, desencadena el bloqueo de la expresión de
los factores necesarios para evitar la producción de estrógenos y oxitocina, impidiendo
de esta forma el desencadenamiento del mecanismo endometrial luteolítico (PGF2α).
Sucedido esto, la vaca entra en un estado de anestro por preñez, el cual cesará posterior
al parto, con la finalidad de continuar con el desarrollo del ciclo estral con sus etapas
completas (Bilby et al., 2004).
1.5. Factores relacionados con el desarrollo embrionario
Los factores que comprometen el desarrollo embrionario y el subsiguiente crecimiento
fetal son complejos ya que involucra procesos como la proliferación celular, el desarrollo
y mantenimiento del CL, las funciones del oviducto y del útero, la implantación y
vascularización del embrión entre otros (Thatcher et al., 2001).
Durante los primeros estadios de división celular desde una célula hasta el blastocisto
temprano alrededor del día 7 - 8, el embrión se encuentra encerrado en la llamada zona
pelúcida, donde sus requerimiento para mantenimiento se basan en el piruvato y
oxalacetato (Araújo et al., 2005). En estas primeras etapas antes de la fertilización, se ha
evidenciado la presencia de factores de crecimiento (FC) como el transformador alfa
(TGF-α), el transformador beta (TGF-β) y el derivado de plaquetas (PDGF), lo cual
sugiere que están involucrados en el proceso de desarrollo temprano del ovocito
(Thatcher et al., 1997).
Entre el tercer y cuarto día posterior a la fertilización (fase de 8 a 16 células), el embrión
migra del oviducto hacia el cuerno uterino en donde la glucosa es utilizada como sustrato
energético (Gomez y Diez 2000). El IGF-I podría estar involucrado en este proceso de

Capítulo 1 51
descenso embrionario, ya que el ARNm que codifica para IGF-I ha sido encontrado en el
oviducto bovino durante esta fase de migración (Kimura et al., 2008).
A los 5 o 6 días de vida embrionaria (fase de 16 a 32 células), se lleva a cabo el proceso
de compactación celular, formándose contactos que desarrollan uniones firmes entre las
células (Thatcher et al., 2004). En esta etapa el embrión comienza a funcionar como un
organismo llamado mórula, el cual es relativamente independiente del ambiente uterino y
su supervivencia parece depender de su programación genética. Durante este estadio
se expresan las anormalidades cromosómicas procedentes del padre, lo que podría
desencadenar en mortalidad embrionaria (ME) (Araújo et al., 2005).
El desarrollo de uniones intracelulares estrechas en el estadio de mórula durante la
compactación, es seguido de la acumulación de líquido formando una cavidad central
que recibe el nombre de blastocele, la cual también acumula líquido proveniente del
metabolismo mitocondrial (Araújo et al., 2005).
La localización de la bomba sodio–potasio (Na+/K+) activa en la membrana basal del
trofoblactodermo para transporte iónico activo, esto establece un gradiente que induce
movimiento de líquido hacia adentro del blastocele, provocando expansión que hace que
las células de la mórula compactada se ubiquen hacia la zona externa o interna de las
vesículas llenas de líquido, con lo cual se forma una capa celular externa que recibe el
nombre de trofoblasto y una masa celular interna llamada embrioblasto, originándose de
esta manera el estadio de blastocisto (Pereira et al., 2008); el cual alrededor de los 8 días
de vida posee aproximadamente 120 células asociadas con la masa celular interna
(embrioblasto) (25%) y el trofoblasto (75%) (Thatcher et al., 2002).
Aproximadamente al 9º o 10º día de vida (fase de 160 células) el blastocisto eclosiona de
la zona pelúcida por combinación de acciones físicas y enzimáticas de la vesícula en
expansión (Johnson, 1979). Esta liberación podría involucrar síntesis de prostaglandinas
por parte del embrión ya que la P4 estaría evitando la expansión y liberación del
blastocisto (Thatcher et al., 2001). Las enzimas plasmina y tripsina activan al blastocisto
lo cual causa reblandecimiento de su zona matriz permitiendo la expansión y ruptura a lo
largo del plano ecuatorial para la salida de la masa celular (Brayman et al. 2004).
Inmediatamente se establece el primer contacto del embrión y el epitelio uterino materno,

52 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
lo cual desencadena un intercambio de nutrientes y prepara el ambiente para un estado
de preimplantación embrionaria (Pereira et al., 2008). Tras la liberación el embrión
comienza a cubrir físicamente parte del endometrio para poder comenzar a regular la
producción de PGF2α uterina la cual cumple funciones de luteólisis (Kaplan, 2002).
El estado de preimplantación embrionaria “in vivo”, estaría regulado por factores de
crecimiento de origen materno y embrionario, como el TGF-α, TGF-β1, PDGF-α, IGF-I,
IGF-II, TGF-β2, TGF-β3, CSF-1, Interleucina 3 (IL-3) e IL-6; además citoquinas como el
factor inhibidor de la leucemia (LIF) (Revisado por Peña et al., 2007).
La regulación de la receptividad del útero a la implantación del blastocisto, podría estar
ejercida por una glucoproteína transmembranosa denominada Musina 1 (Muc–1), la
cuales expresada en la superficie de gran parte de epitelio del tracto reproductivo
(Brayman et al. 2004). Esta glucoproteína es abundante en la fase no receptiva y se
reduce notablemente o puede estar ausente para el momento de la implantación, con lo
cual se abre paso a la cascada de adhesión del trofoblasto embrionario, acción regulada
por las integrinas en el lumen del epitelio endometrial, donde se involucran proteínas
como la LGALS15, SPP1 y la fibronectina oncofetal en el caso de ovinos (Johnson et al.,
2003; Brayman et al., 2004).
Como la síntesis epiteliar de Muc–1 está regulada por la P4, la perdida de receptores
nucleares para esta hormona en el epitelio uterino (día 8 – 10) reduciría la producción de
Muc–1 y se abriría un estado receptivo para la adhesión del embrión. Para el día 10 se
reduce la producción de la P4, lo que conlleva igualmente a la reducción de la
concentración de Muc-1 (Spencer et al., 2007). Esta reducción permite la interacción y
contacto entre factores adhesivos como las integrinas y sus receptores, acercando el
embrión a la superficie uterina y formando un tipo de placentación epiteliocorial (Spencer
et al., 2004a).
Johnson et al., (1999) reportan la expresión de integrinas en rumiantes, las cuales son
secretadas por las glándulas endometriales y detectadas a lo largo de la superficie del
lumen uterino hacia los días 19 a 21 de preñez (periodo temprano de adhesión con
puntos visibles entre carúnculas y cotiledones).

Capítulo 1 53
Se sugiere que la αvβ3 es una de las más versátiles integrinas capaz de vincular
proteínas como la fibronectina, fibronectina oncofetal, vitronectina, tenascina, galactina -
15 y osteopontin de la cual se reporta que el epitelio glandular de ovejas preñadas
expresa su mRNA (Yang y Liu, 2003; Gray et al., 2004; Spencer et al., 2004a).
También se ha observado que la expresión endometrial de esta proteína es regulada por
la P4 y no por el IFN-τ (Johnson et al., 2000). En vacas se reporta específicamente la
expresión de subunidades de integrinas β1, α3, α6 y αvβ3 (Spencer et al., 2007).
Otra molécula vinculada a la adhesión embrionaria, es la molécula 1 de adhesión celular
1 (GLYCAM -1), la cual es secretada por el endometrio y ha sido detectada en vacas
preñadas, encontrado sus máximas concentraciones hacia el día 15 y 17 de gestación
(Klisch et al., 2009).
Se ha sugerido que el trofoblasto comienza a adherirse firmemente al lumen del epitelio
endometrial hacia el día 16, observándose durante esta adhesión del embrión bovino y
ovino el desarrollo de microvellosidades, las cuales son proyectadas hacia el interior de
la luz de las glándulas uterinas (Spencer et al., 2004b). Estas vellosidades proporcionan
un anclaje transitorio y una estructura absorbente para el embrión mientras sigue
completándose la implantación. Posteriormente estas microvellosidades se reducen en
longitud lo que conlleva a un contacto más cercano entre estas y el epitelio uterino, el
cual termina comprimiéndose hacia la superficie trofoblástica interbloqueándose con las
proyecciones citoplásmicas en la superficie del trofoblasto hasta que sus
microvellosidades vuelven a desarrollarse, de esta forma generando una adhesión
compleja (Spencer et al., 2004a).
Hoffman y Wooding (1993) sugirieron que solamente las células mononucleadas del
trofoblasto embrionario son las que se adhieren al lumen del epitelio endometrial. En
cuanto que las células binucleadas del trofoblasto podrían estar participando de alguna
forma en la formación del sincitio maternofetal, escencial para la implantación exitosa y
posterior crecimiento placentomal; en donde se forman paquetes de células trinucleadas.
Igualmente las células binucleadas podrían estar involucradas de alguna manera en la

54 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
síntesis y secreción de proteínas (lactógeno placentario) y hormonas esteroideas (P4)
entre otras (Spencer et al. 2004b).
Se ha planteado por algunos autores que los mecanismos que intervienen en el
desarrollo embrionario, implantación y desarrollo placentario son aún complejos, lo que
hace que se desconozca el papel de muchas de las moléculas presentes en el proceso
de la gestación. Las proteínas asociadas a la gestación (PAGs) son, junto con moléculas
conocidas como el lactógeno placentario y el IFN-τ, producto de las células trofoblásticas
(mononucleadas y binucleadas) del embrión y tejido placentario de mamíferos ungulados
(Klisch et al., 2009)
Varios estudios han revelado la expresión de ARNm en la placenta de rumiantes la
existencia de multitud de genes de PAG. La elevada homología que caracteriza estas
moléculas dificulta la purificación y aislamiento de formas simples, por lo que sólo se ha
conseguido analizar la secuencia de un número muy reducido de PAGs (Wooding, 2005).
Hacia el día 42 después de la fertilización termina el período embrionario al completarse
el periodo de diferenciación, evidenciándose el feto en el cual la mayoría de los tejidos,
sistemas y órganos se encuentran ya formados (Hafez, 2000).
1.6. Aspectos relacionados con el reconocimiento materno de la preñez El RMP se define como el periodo crítico en el cual el conceptus da señales de su
presencia a la madre (Spencer et al., 2004a). Este reconocimiento requiere que el
conceptus se transforme de esférico hacia una forma elongada para generar una mayor
superficie de contacto con el epitelio uterino, con la finalidad de desencadenar la
producción del factor antiluteolítico IFN-τ; producción regida entre otros por el estado de
desarrollo embrionario (Binelli et al., 2001). Este interferón es producido en las células
mononucleares del trofoblasto embrionario entre los días 10 y 25, con producción
máxima alrededor de los días 14 y 19 de gestación, tiempo en el cual el embrión se
encuentra en estado de precontacto (Fleming et al., 2001; Dailey et al., 2002).
La síntesis y secreción de IFN-τ se relaciona con la presencia en el fluido luminal uterino
del FC IGF- I y IGF-II. La presencia de un solo FC no afectaría la presencia de interferón,

Capítulo 1 55
sin embargo ambos factores podrían aumentar significativamente su secreción, lo que
podría indicar un papel importante para los FC similares a la insulina en el RMP. El IFN-τ
comienza a establecer efecto antiluteolítico a nivel de la región intercaruncular, en donde
las células responden más a concentraciones de Oxitocina (OT) (Roberts, 1999). La
expresión génica de IFN-τ se ha detectado en la región de contacto de la implantación
celular, entre las células mononucleadas del trofoblasto embrionario y las células epitelio
del lumen endometrial (Spencer et al., 2007).
De acuerdo a la clasificación de la Sociedad Internacional de Interferones, éstos son
glucoproteínas sintetizadas durante la respuesta inmunitaria así como bajo influencia de
múltiples estímulos antigénicos o mitógenos; igualmente poseen propiedades antivíricas
y antiproliferativas (Rodríguez, 2001).
El IFN-τ se clasifica como una proteína ácida con peso molecular de 19 kDa y cuenta con
172 aminoácidos. Es el primer interferón clasificado en la familia tipo I cuyas propiedades
no solamente son antivíricas y es expresado por una familia de genes que está
restringida a especies de rumiantes como vacas, ovejas y cabras (Bilby et al., 2004;
Roberts, 2007).
Durante la fase lútea la P4 tiene una doble función en la regulación de la expresión de
receptores para la OT en el útero, induciendo la presencia de altos niveles de receptores
para P4 en el endometrio (modulación en alta), con lo cual se inhibe la expresión de los
receptores para estrógenos (RE) y para oxitocina (ROT), fenómeno que se conoce como
el "bloqueo de la P4" (Roberts, 1999). No obstante, la misma P4 comienza a ejercer
gradualmente una regulación para la disminución de la expresión de sus propios
receptores (modulación baja) en el epitelio luminal, perdiendo así su habilidad para
suprimir la expresión endometrial de los RE y ROT, por lo cual en el caso de existir
preñez el embrión debe extender el "bloqueo de la P4", previniendo así la producción
pulsátil de PGF2α mediante la secreción de IFN-τ (factor luteotrópico) (Klisch et al., 2009).
Este efecto antiluteolítico del IFN-τ, resulta en el mantenimiento estructural y funcional
del CL, por lo tanto se asegura la secreción de P4 que es esencial para mantener un
ambiente uterino que soporte los acontecimientos críticos para el desarrollo exitoso del
conceptus (Bilby et al., 2004).

56 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
1.6.1. Señal luteotrópica La formación del CL activa la síntesis y secreción de P4 por parte de esta glándula
endocrina transitoria, con ello se incrementan sus receptores en el útero hacia los
primeros días del diestro (regulación en alta), lo que desencadena el bloqueo de la
expresión del gen para RE, así mismo se previene que los estrógenos incrementen la
expresión de genes para ROT (Rodríguez, 2001; Olivera, 2006).
Hacia el final de la mitad de la fase luteal del CE los receptores de P4 en el útero
comienzan a disminuir (regulación en baja), desencadenando un incremento en la
expresión de RE y ROT, con lo cual se desencadenan los factores necesarios para
causar luteolisis, evidenciándose un nuevo ciclo estral en el caso de no existir preñez
(Olivera, 2006).
El IFN-τ producido por las células mononucleares del trofoblasto embrionario, actúa de
forma parácrina ligándose a sus receptores en las células endometriales, lo que
desencadena el bloqueo de la expresión de ARNm que codifica para la expresión del gen
de receptores alfa de E2 (RE) y la expresión del gen para ROT, impidiendo de esta forma
el desencadenamiento del mecanismo endometrial luteolítico. Sin embargo el interferón
no inhibe la producción basal de la PGF2α (Igwebuike, 2006; Green et al., 2010).
Algunos autores sugieren que al ligarse el IFN-τ a su receptor de membrana, induce la
vía de transcripción citoplasmática (StAR), proteína que fosforilada forma un complejo
que migra al núcleo, allí se une a la región reguladora de los genes inductores del
interferón que podrían estar bloqueando uno o más pasos de la vía de síntesis de la
prostaglandina (Spicer et al., 2011). Igualmente se ha propuesto que el IFN-τ también
ejerce un efecto antiluteolítico incrementando los niveles de ácido linoléico, el cual inhibe
por competencia de cox 2 al ácido araquidónico, con lo cual la síntesis de PGF2α se ve
afectada (Rodríguez, 2001).
En el mecanismo antiluteolítico se sugiere que participan además del IFN-τ la
prostaglandina E2 la cual ejercería protección del CL, el factor activador de las plaquetas
(FAP), este último posiblemente en sinergismo con el interferón y quizás otras proteínas
de origen trofoblástico entre otros (Campanile et al., 2007).

Capítulo 1 57
Básicamente el RMP está regulado por la capacidad del endometrio en responder a las
señales generadas por el embrión, lo que desencadena el bloqueo en la producción de
PGF2α. Este bloqueo depende básicamente de la elongación del conceptus el cual debe
cubrir una adecuada cantidad de puntos de cuerno uterino para comenzar a generar la
síntesis de IFN-τ (Campanile et al., 2007). Teniendo en cuenta lo anterior se ha
evidenciado que el conceptus bovino varía de tamaño que va de 15 a 250 mm. para el
día 17, con lo cual se resalta la importancia del desarrollo embrionario que es regulado
en gran parte por interacciones guiadas por la P4 (Binelli et al., 2001).
1.6.2. Señal luteolítica La vaca ovula espontáneamente, lo cual es dependiente del ciclo estral que es repetitivo
de acuerdo a su duración hasta el establecimiento de la preñez (Hafez, 2000). Por tal
razón se podría decir que el ciclo estral es dependiente de la acción del endometrio
(epitelio luminal y glandular superficial), si se tiene en cuenta que este es la fuente del
factor antiluteal (PGF2α) cuando la vaca no se encuentra preñada o cuando no se
reconoce su preñez (Barnea et al., 2000).
Durante el diestro temprano, la síntesis de P4 producida por el CL recién formado
comienza a disponer el útero para el establecimiento de la preñez, pero igualmente activa
mecanismos determinantes para la producción endometrial del factor luteolítico
(Rodríguez, 2001).
Hacia los días 10 y 15 del ciclo estral, cuando comienza a disminuir el número de
receptores de P4 en el útero (modulación baja), comienzan a aumentar los RE
endometriales los cuales inducen la expresión de ROT (Johnson et al., 2000). Los E2
provienen de los folículos en crecimiento y estimulan el aumento de sus receptores alfa.
La OT proviene de las células luteales grandes; igualmente es producida en el
hipotálamo y almacenada en la neurohipófisis (Spencer et al., 2004a).
La alta concentración de OX igualmente comienza a provocar la pérdida de sensibilidad
del endometrio, por lo cual luego de un lapso de tiempo se deja de secretar PGF2α. El
intervalo entre pulsos de PGF2α está determinado por la pérdida de la sensibilidad del
endometrio a la OX, y por el tiempo en que tarda en recuperarla. Así, el mecanismo de

58 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
retroalimentación positiva se interrumpe y vuelve a establecerse hasta que pasan 6
horas, tiempo suficiente para que el endometrio recupere la sensibilidad a la OX; por tal
motivo los pulsos de PGF2α se presentan con un intervalo de 6 a 8 horas (Thatcher et
al., 2002).
En la célula endometrial la unión de la OT con su receptor estimula el clivaje de ácido
araquidónico a partir de la fosfolipasa C (PLC), la cual activa la fosfolipasa A2 (PLA2)
(localizadas en la membrana plasmática) (Roberts et al., 1999). Intervienen para este fin
mensajeros como el inositol trifosfato (IP3) y el diacylglicerol (DAG) (sistema de segundo
mensajero fosfatidil – inositol – diacilglicerol – proteína kinasa C) (Rodríguez, 2001). El
IP3 estimula la liberación de Ca ++ de las reservas intracelulares (Thatcher et al., 2002). El
DAG activa la proteína Kinasa C (PKC) la cual estimula a la PLA2 en presencia de Ca ++
para que posteriormente se induzca la liberación de ácido araquidónico de los depósitos
fosfolipidicos en el epitelio endometrial (la P4 en etapas tempranas de CE induce
incremento en las reservas de fosfolípidos (Thatcher et al., 2002).
La transformación del ácido araquidónico en PGF2α, es regulada por la isoforma
ciclooxigenaxa 2 (COX 2) de la prostaglandina sintetasa, enzima identificada en el
epitelio luminal de vacas preñadas, la cual limita la síntesis de las prostaglandinas
(Hassan et al., 2007). La actividad de esta enzima se incrementa durante el diestro
temprano por acción de la P4 la cual es inducida por el estradiol, factores de crecimiento
e interleuquinas (Rodríguez, 2001; Peña et al., 2007). La concentración de COX 2 se
reduce cuando el IFN-τ comienza a ser secretado por parte de las células del trofoblasto
embrionario (Thatcher et al., 2002).
La ciclooxigenasa (Prostaglandina G/H sintetasa) cataliza el paso limitante en la
biosíntesis de prostaglandinas, que es la conversión del ácido araquidónico a
prostaglandina H2 (PGH2), la cual es convertida rápidamente a PGF2α gracias a la
prostaglandina F sintetasa entre otras (Niswender et al., 2000).
Al final de cada ciclo estral hacia el día 16 - 17 el CL sufre luteólisis (inhibición de la
esteroidogénesis - inducción de la apoptosis) por la acción de la PGF2α, la cual es
producida en el epitelio endometrial y en menor proporción por el CL, esta es vertida a la

Capítulo 1 59
vena uterina, de donde por difusión pasa a la arteria ovárica y por contracorriente alcanza
al CL (Esto permite a la PGF2α viajar a la arteria ovárica sin entrar a la circulación
pulmonar, en donde podría ser enzimáticamente inactivada en los pulmones), en donde
se une a receptores ubicados en las células luteales (Niswender et al., 2000). Allí por
eventos intracelulares se presenta la muerte de las células por apoptosis mediante el
desencadenamiento de cascadas de señalización que involucran hormonas como la
prolactina; citoquinas como el Factor de Necrosis Tumoral alpha (TNFα), el Interferón
gamma (IFNγ) y el Fas ligando (FasL); especies reactivas de oxígeno (ROS), endotelina -
1(E1) y la proteína HSP70 de choque térmico, entre otros (Nakamura y Sakamoto, 2001;
Roberts, 2007).
La endotelina -1 se ha identificado como posible mediadora de los efectos de la PGF2α
en el flujo sanguíneo luteal. La PGF2α estimula a las células endoteliales del CL a
producir endotelina – 1 in vitro e in vivo. Esta endotelina posee actividad vasoconstrictora
e inhibe la actividad esteroidogénica de las células luteales (Niswender et al., 2000).
El proceso apoptotico parece estar mediado por el ARNm para FAS y FalsL, a través de
la activación de la caspasa-3 (Pru et al., 2002). El receptor de FasL es un dimero proteico
con un solo segmento transmembranal que en su porción intracitoplasmática tiene un
dominio de unión de muerte celular, al cual se une la proteína FADD, que a la vez tiene
un dominio efector de muerte, que es el sitio de unión de la procaspasa – 8 para formar
el complejo señalizador para la inducción de muerte. Las procaspasas – 8 reclutadas, se
clivan para activarse en caspasa – 8 (Rueda et al., 2000)
La luteolisis ha sido planteada en dos pasos: Luteolisis funcional, que consiste en la
pérdida de la capacidad de sintetizar P4, y luteolisis estructural, que es la involución del
CL acompañada de la pérdida en la integridad de sus células (Pretheeban et al., 2010).
Se sugiere que para que se produzca luteólisis, el CL debe ser expuesto
aproximadamente de 5 a 8 pulsos de PGF2α con intervalos de 6 a 8 horas (Mann and
Lamming 2006a). Esta pulsatilidad ocurre en respuesta a la unión de la OT con su
receptor en las células del endometrio uterino (Rodríguez, 2001). Igualmente se plantea
que el CL debe alcanzar cierto grado de madurez caracterizado por una amplia
vascularización y por la producción de P4 que lleva a niveles por encima de 1 ng/ml., esto
ocurre aproximadamente hacia el día 5 del ciclo estral (Revisado por Olivera et al., 2007).

60 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
1.7. Control del ciclo estral en programas de transplante embrionario
Para poder obtener mejores resultados en cualquier técnica de reproducción asistida
debe realizarse un control efectivo del ciclo estral de la hembra bovina. Actualmente, se
vienen desarrollando una serie de protocolos de sincronización del estro, proceso por el
cual se controla el ciclo estral de la hembra por medio de la utilización de hormonas
exógenas que ejercen un directo control sobre el ciclo. Estos procedimientos están
influenciados principalmente por la acción de la prostaglandina F2α que produce la
regresión de una manera precoz del cuerpo lúteo y por la acción de la progestágenos los
que actúan en la hembra simulando un cuerpo lúteo (Palma, 2008).
La prostaglandina F2α es uno de los métodos utilizados tradicionalmente en protocolos
de control del ciclo estral. El inconveniente de ésta, es que es efectiva únicamente
durante la fase luteal del ciclo estral; una aplicación de prostaglandina F2α entre los días
6 y 16 inducirá la regresión de cuerpo lúteo lo que permitirá la finalización de la fase
luteal, dando paso al inicio otro ciclo. Al efectuar una aplicación de prostaglandina F2α,
se encuentra que solo un 50 y 70% de animales responderán al tratamiento entrando en
calor. Si se administran dos aplicaciones de prostaglandina F2α con un intervalo de 11 a
14 días (Gómez, 2005), se permitirá que todas las vacas tengan una fase lútea presente
en el momento de la segunda aplicación por lo que se logra la sincronización completa
del ciclo estral, realizando la detección del celo 5 días después de la segunda aplicación.
Sin embargo la acción de prostaglandina F2α se ve limitada por que solo actúa en
animales que se encuentren ciclando y que presenten un cuerpo lúteo normal (Hafez,
2000; Bó, 2002; Gouveira, 2004).
Otra manera para Sincronizar el ciclo estral de la hembra bovina es con la ayuda de la
progestágenos, por medio de la cual se trata de simular la fase luteínica del ciclo estral,
prolongándola (Gordon, 1999). El beneficio de este sistema de control del ciclo estral, es
que no se necesita que el animal presente un cuerpo lúteo funcional. Al encontrarse
niveles de P4 exógena en el sistema del animal va a generarse un bloqueo hormonal,
debido a que éste simula la presencia de un cuerpo lúteo; pero es importante contar con
un sistema en el que los niveles de P4 permanezcan por un espacio de tiempo

Capítulo 1 61
determinado en el cuerpo del animal, como pueden ser implantes subcutáneos,
dispositivos intravaginales, esponjas, entre otros (Barreto and MacManus, 2002; Gómez,
2005).
Las nuevas aplicaciones de tratamientos hormonales muestran que la interacción de
hormonas generan mejores resultados en el control del ciclo estral (sincronización de
celos). Así se encuentra que existen varios protocolos como el Ovsynch en el que se
aplica GnRH + prostaglandina F2α + GnRH para después de 16 a 24 horas post -
aplicación de la segunda dosis de GnRH proceder a Inseminar a tiempo fijo (Gouveira,
2004; Bó y Cutaia, 2005). Al hacerse comparaciones entre el método Ovsynch y una
inseminación artificial a celo detectado, Stevenson (1999), encontró diferencias
significativas (P<0.05) ya que el porcentaje de preñez con el método Ovsynch fue del
35% mientras que al inseminar a celo detectado presentó un 26% de efectividad. En
1997, Pursley encontró diferencias significativas (P<0.05), ya que el porcentaje de preñez
con el método Ovsynch fue del 39% mientras que al inseminar a celo detectado presentó
un porcentaje de preñez del 36%. En 2000, De la Sota encontró diferencias significativas
(P<0.05) ya que el porcentaje de preñez con el método Ovsynch fue del 14% mientras
que al inseminar a celo detectado presentó un porcentaje de preñez del 5%. En otro
estudio realizado, se encontraron diferencias significativas (P<0.05) ya que el porcentaje
de preñez con el método Ovsynch fue del 50% mientras que al inseminar a celo
detectado presentó un porcentaje de preñez del 40% (Colazo et al, 1996). De esta
manera se evidencia un aumento en el porcentaje de preñez después de la IA, mediante
el uso del protocolo en el que se utilizan dos dosis de GnRH y una dosis de
prostaglandina F2α contra la IA a celo detectado.
Para la aplicación de las técnicas de sincronización del estro también se encuentran
protocolos en los que se emplean dispositivos de liberación lenta de P4. Esta hormona ha
sido empleada hace aproximadamente cuarenta años para el control del ciclo estral
(Wiltbank, 1965). Actualmente se está trabajando con estos dispositivos que al utilizarlos
en el animal por vía intravaginal, al día cero del protocolo sumado con la aplicación de
Benzoato de Estradiol, va a producir un control del desarrollo folicular, ya que al
aumentarse los niveles de estradiol se estimula la secreción de inhibina, pero genera un
déficit de los niveles de folistatina, lo cual genera una disminución de la acción de FSH
(Bó et al., 2002).

62 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
El dispositivo de liberación lenta de P4 al ser utilizado solo, genera unos niveles muy
bajos de ésta y al estar presente estos niveles en el cuerpo del animal se va a producir
una sobremaduración por la persistencia del folículo lo cual hace que cuando ovule este
folículo se obtenga un oocito alterado o envejecido (Sartori, 2002; Mufeed et al., 2008). Al
aplicar el dispositivo de liberación de P4 junto con una dosis de estradiol se genera una
retroalimentación negativa para la liberación de GnRH actuando a nivel de hipotálamo y
suprimiendo el desarrollo de los folículos presentes en el ovario, como la vida media del
estradiol exógeno es de aproximadamente 4 días, el día 4.3 caen los niveles de éste
haciendo que se produzca una liberación de GnRH en el hipotálamo, la cual va a la
hipófisis liberando FSH y LH, permitiendo que se inicie una nueva onda de crecimiento
folicular (Bó et al., 1995; Bó et al., 2002; Tribulo, 2002). De esta manera combinando la
regulación de la fase folicular con la de la fase luteal, es posible predecir el surgimiento
de una nueva onda de desarrollo folicular (Twagiramungu et al., 1995; Bó et al., 1996). El
día 7 contado desde el inicio del protocolo de sincronización se retira el dispositivo y se
realiza una aplicación de prostaglandina F2α (para inducir luteolisis); el día 8 se aplica
Benzoato de Estradiol (para sincronizar el momento de la ovulación) y treinta horas
después proceder a Inseminar a tiempo fijo (Barreto and MacManus, 2002; Barret et al.,
2006), ya que la mayoría de los animales ovulan 66 horas después de retirado el
implante de liberación lenta de P4 (Acosta, 2004). La variabilidad de resultados obtenidos
mediante las diferentes técnicas de sincronización hace que se permita avanzar en los
protocolos que controlan el desarrollo folicular; de manera que los animales presenten un
folículo en crecimiento, con capacidad de ovular en el momento del retiro del dispositivo
de liberación lenta de P4 y de la aplicación de la prostaglandina (Bó et al., 1991; Kastelic
et al., 1996; Colazo et al., 1996).
Las ventajas del tratamiento para la sincronización del celo por medio de la asociación
del dispositivo de liberación lenta de P4, estradiol, prostaglandina y GnRH, se evidencia
por medio de trabajos realizados por Bó et al., (1995), en el que a un grupo de hembras
bovinas, n=34, se les sincronizó el estro con el tratamiento descrito anteriormente, el cual
fue comparado contra una sincronización efectuada con dos inyecciones de PGF2α. Los
resultados tras la aplicación de los tratamientos se reportaron así, el 75% de las hembras
sincronizadas con el tratamiento P4, estradiol, PGF2α, GnRH presentaron una ovulación
efectiva 72 a 84 horas después del tratamiento y un 40% de las hembras sincronizadas

Capítulo 1 63
con dos inyecciones de PGF2α ovularon en el mismo tiempo, encontrando diferencias
significativas entre estos tratamientos (P<0.05). De la misma forma se evidencia la
efectividad de este tratamiento cuando Anderson (1999) lo comparó contra una
sincronización que consistía en una inyección aplicada el día cero de GnRH, una
inyección de PGF2α el día seis y se insemina el animal 12 horas después de la
observación del celo. Los resultados reportados al realizar esta comparación,
demuestran que el tratamiento en el que se usa el dispositivo de liberación lenta de P4
generó un 64% de preñez mientras que el otro tratamiento resultó en un 44% de preñez;
otra ventaja de la utilización de este tratamiento es la posibilidad de efectuar una
inseminación artificial a tiempo fijo (IATF). En otra fase del mismo estudio se
sincronizaron 3145 vacas para comparar el uso del tratamiento del dispositivo de
liberación lenta de P4 contra un control no tratado. Se reportó que los animales tratados
con el dispositivo de progesterona presentaron un porcentaje de preñez del 60%
enfrentado con el grupo control que fue del 53%.
Así mismo, se evaluaron las variaciones del protocolo en el que se emplea el dispositivo
progestágeno CIDR-B; se aplicó por 7 u 8 días obteniéndose tasas de preñez del 60%
(en el tratamiento CIDR-B) por 7 días y de 50% (en el tratamiento CIDR-B) por 8 días
(P<0.05), encontrándose una mayor eficiencia en el tratamiento de 7 días (Bó et al.,
1996).
La alternativa más efectiva para realizar un efectivo control del ciclo estral y de la
ovulación resultó ser el tratamiento con el CIDR-B más 17-Beta estradiol, prostaglandina
F2α y una aplicación de GnRH. Se presentó un porcentaje de preñez del 77%. Este fue
enfrentado contra la aplicación del CIDR-B más una aplicación de GnRH, el cual
presentó un porcentaje de preñez del 42% (P<0.05%). Por este motivo actualmente se
presentan óptimos resultados en el control del ciclo estral mediante aplicación de estas
terapias hormonales en las hembras bovinas tratadas con estos dispositivos de liberación
de P4 (Baruselli et al., 2001a, b; Sakase et al., 2006;), inclusive en el trópico alto
colombiano, se confirman estos resultados haciendo las comparaciones entre estos
métodos de control del ciclo estral (Celemin y Cifuentes, 2003).

64 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
1.7.1 Estrategias antiluteolíticas en programas de transplante embrionario en bovinos La naturaleza crítica alrededor del periodo de reconocimiento, aposición y adhesión del
embrión al endometrio uterino durante la implantación, determina la necesidad de un
estricto sincronismo entre el embrión trasplantado y su receptora, enfatizando la
importancia tanto del medio uterino como de las señales del conceptus que dan lugar al
RMP (Revisado por Tovío et al., 2008), señales que deben ser emitidas en el momento y
concentración precisa, de tal manera que se garantice el mantenimiento de la estructura
y funcionalidad del CL, que genere una continua producción de P4 para el mantenimiento
del ambiente embriotrófico que apoye el normal desarrollo del conceptus (Binelli et al.,
2001; Mann and Lamming, 2001).
La hembra debe ajustar sus propiedades fisiológicas dependiendo de si está o no
preñada, si lo está, debe generarse un bloqueo efectivo antiluteolítico el cual depende de
la habilidad del conceptus en enviar señales para que el endometrio uterino responda a
ese bloqueo en la producción de PGF2α (Thatcher et al., 2002).
La comunicación entre el conceptus y el útero, no siempre tiene éxito, lo que
desencadena grandes pérdidas embrionarias (Binelli et al., 2001). Por eso mismo se
plantean estrategias que buscan minimizar el efecto luteolítico el cual desencadena la
pérdida temprana del embrión, acontecimiento más fácilmente evidenciado en los
programas de transplante embrionario, ya que con éstos se asegura la presencia del
embrión vivo desde el día 6 a 8 momento en el cual este es trasplantado (Baruselli et al.,
2005).
La relación entre el porcentaje de preñez y la concentración plasmática de P4 de acuerdo
al tamaño del CL en receptoras de embriones bovinos, ha sido objeto de controversia en
varios estudios realizados. Algunos investigadores han verificado la correlación positiva
entre estas variables, encontrando que a mayor área del CL mayor es la concentración
de P4 plasmática, y consecuentemente mayor es la tasa de preñez (Kastelic et al., 1990;
Vasconcelos et al., 2001; Bó et al., 2002; Thatcher et al., 2002; Mantovani et al., 2002;
Baruselli et al., 2005; Lopes et al., 2007). Con estos resultados se podría evidenciar que
los incrementos en la concentración de P4 durante el "periodo crítico" estimula la

Capítulo 1 65
secreción de los agentes antiluteolíticos con lo cual se hace eficiente el RMP
(Vasconcelos et al., 2001).
Contrariamente autores como Spell et al. (2001), no han encontrado correlación entre las
variables tamaño del CL, concentración de P4 plasmática y porcentaje de preñez en
hembras receptoras con embrión transplantado.
En varios estudios realizados el aumento en concentraciones plasmáticas de P4 durante
el diestro ha sido correlacionado positivamente con la capacidad del embrión en secretar
IFN-τ, provocando así aumento en las tasas de preñez (Thatcher et al., 2002; Mann et
al., 2006b), pero en otros reportes no se ha observado esta relación y efecto (Moreira et
al., 2004).
Kerbler et al. (1997), encontraron una correlación positiva entre la concentración de P4
plasmática y la síntesis de IFN-τ producido por embriones de 18 días de edad, en donde
se halló la concentración de interferón utilizando una prueba antiviral con virus de
estomatítis vesicular. Según estos resultados se podría sugerir que a mayor
concentración de P4 plasmática en hembras preñadas mejor será el ambiente uterino
para el conceptus en vías de desarrollo (Revisado por Gonella, 2010). Esto se entiende si
se tiene en cuenta que la P4 es la precursora de los diferentes componentes que
conforman este ambiente. Cualquier variación en la concentración de P4 es determinante
en la modulación de la expresión y secreción de factores de crecimiento, citoquinas y
proteínas, que condicionan el medio uterino para los procesos de receptividad
endometrial y de viabilidad embrionaria (Mann et al., 2006; Pinheiro et al. 2009; Beltman
et al., 2009).
De acuerdo a lo anterior se podría sugerir que al brindar fuentes directas o indirectas de
P4 a hembras durante los primeros días de preñez el porcentaje de pérdidas
embrionarias disminuye, ya que se mejora el ambiente uterino en donde el conceptus
tendrá un desarrollo adecuado evidenciando mejor síntesis y secreción de IFN-τ, pues
esta secreción está influenciada por el estado de desarrollo embrionario (Walker et al.,
2009).

66 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Con la finalidad aumentar la tasa de preñez se han utilizado tratamientos hormonales en
hembras receptoras de embriones bovinos, utilizado Gonadotropina Corionica Equina
(eCG) en protocolos de transplante de embriones a tiempo fijo (TETF) (Santos et al.,
2001; Rodrigues et al., 2004; Baruselli et al., 2005; Fuentes y De La fuente 2007).
La eCG (antes conocida como Gonadotropina Sérica de Yegua Preñada PMSG) es una
hormona glicoproteica con peso molecular de 45 kDa. y promedio de vida de 3 días,
producida por los cálices endometriales en la yegua preñada entre los días 40 a 130
(Bousfield y Butnev, 2001). Se compone de dos subunidades α-y β (la subunidad α está
codificada por un gen común en todas las glicoproteínas, mientras que estas hormonas
difieren en el gen que codifica para subunidad β-, la cual confiere la especificidad
hormonal) (Pinheiro et al. 2009; Small et al., 2009).
Esta hormona se vincula a los receptores foliculares de FSH y de LH, y a los receptores
de LH del CL, creando de esta forma condiciones de crecimiento folicular, ovulación y
luteinización (Baruselli et al., 2003; Rezende et al., 2006); Pero su acción predominante
es de FSH, lo que daría a lugar a la formación de cuerpos lúteos accesorios
característicos de la yegua gestante (Bousfield y Butnev, 2001).
La aplicación de eCG en el momento esperado de una nueva onda de crecimiento
folicular, ha demostrado eficiencia en cuanto a superovulación y/o desarrollo de un
folículo dominante de mayor diámetro, determinando de esta forma un mayor número de
cuerpos lúteos o un CL grande (Baruselli et al., 2005). Esto va acompañado de mayores
concentraciones plasmáticas de P4 y mejores tasas de aprovechamiento, concepción y
de preñez frente a tratamientos sin aplicación de esta hormona (Tribulo et el., 2002;
Everton and Baruselli, 2003; Madureira et al., 2004; Baruselli et al., 2005).
Investigaciones realizadas por Melanie et al. (2004), no reportaron diferencia entre la
concentración plasmática de P4 y la cantidad de cuerpos lúteos presentes en hembras
preñadas, por el contrario encontraron mayor pérdida de preñeces en las hembras con
doble ovulación, lo que sugiere que demasiada P4 plasmática podría estar alterando el
balance hormonal uterino perjudicando el ambiente embriotrófico para el embrión en
desarrollo.

Capítulo 1 67
Nasser et al., (2004), han verificado que con la aplicación de eCG el día 8 de
sincronización, determina apenas un 2% de doble ovulación en receptoras de embriones,
pero evidenciaron que con la aplicación de esta hormona se consiguen cuerpos lúteos
únicos de mayor tamaño, incrementando así la tasa de preñez. Igualmente Quezada y
Ortiz (2007), encontraron que con la aplicación de la eCG el día 8 se mejora la tasa de
aprovechamiento, sin embargo no encontraron que esta hormona mejore el área del CL.
En un estudio realizado por Mamani et al. (2007) de acuerdo a la dosificación de la eCG,
no se reporta diferencia de la tasa de aprovechamiento y tasa de preñez con la utilización
de diferentes dosis de eCG (200, 300 y 400 UI) en hembras receptoras cruzadas Bos
Indicus X Bos Taurus.
Se han realizado estudios utilizando eCG en donde se ha observado el incremento en las
tasas de preñez en programas de inseminación artificial a tiempo fijo (IATF) (Baruselli et
al., 2003; Bó, 2003; Sousa et al., 2009; Filho et al., 2010).
Con la finalidad de incrementar la P4 plasmática en búfalas y vacas, se han utilizado
tratamientos con GnRH, LH, hCG (esta última producida en el sincitio trofoblasto de las
mujeres embarazadas) y dispositivos de liberación lenta de P4 aplicados el día 7 del ciclo
estral (Campanile et al., 2007).
La hCG se ha aplicado en tratamientos de sincronización el día 6, obteniéndose tasas de
preñez más altas frente a grupos sin la aplicación de esta hormona. Estos resultados
igualmente sugieren que la hCG, la cual tiene acción LH, dependiendo del día de su
aplicación induce la ovulación y la formación de cuerpos lúteos accesorios, los cuales
incrementan la concentración de P4 plasmática y la tasa de preñez en hembras
receptoras de embriones bovinos (Baruselli et al., 2003).

68 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Bibliografía
1. Acosta TJ and Miyamoto A. Vascular control of ovarian function: Ovulation, corpus
luteum formation and regression. Animal Reproduction Science 2004. 82-83: 127-
140.
2. Anderson, J. Resultados de trabajos utilizando CIDR-B. Segundo Workshop de
Reproducción Bovina Boehringer Ingelheim S.A, Tandil, Argentina. 1999. 1-17.
3. Amstalden M, García, S, Williams R, Stanko S, Nizielski C, Morrison D, Keisler And
Williams GL. Leptin gene expression, circulating leptin, and luteinizing hormone
pulsatility are acutely responsive to short – term fasting in prepuberal heifers:
Relationsships to circulating insuling and insuling – like growth factor I. Biol Reprod
2000. 63: 127–133.
4. Araújo M, Vale V, Ferreira A, Sá W, Barreto J, Filho L. Secreção de interferon-tau em
embriões bovinos produzidos in vitro frescos e congelados. Interferon tau secretion in
cattle embryos in vitro fertilized before and after cryopreservation. Arq Bras Med Vet
Zootec 2005. 57: 751-756.
5. Austin EJ, Mihm M, Evans ACO, Knight PG, Ireland JLH, Ireland JJ, Roche JF.
Alterations in intrafollicular regulatory factors and apoptosis during selection of
follicles in the first follicular wave of the bovine estrous cycle. Biol Reprod 2001. 64:
839-848.
6. Avila M, Madeira, Lucci MC, Aquino S, Bao N, Morphometric and ultrastructural
characterization of Bos indicus preantral follicles. Animal Reproduction Science 2005.
87: 45–57.
7. Axel p. Mathieu, Lavigne P and Lehoux J: Molecular modeling and structure-based
thermodynamic analysis of the star protein. Endocrine research, 2002, vol. 28, no. 4
pages 419-423
8. Baruselli P, Bó G, Reis E, Marques M, Sá M. Introducción de la IATF en el manejo
reproductivo de rebaños de ganado de engorde en Brasil. Congreso Internacional de
Reproducción Bovina. Bogotá 2005.
9. Baruselli P, Marques O, Nasser LF, Reis EL and Bó G. Effect of eCG on pregnancy
rates of lactating zebu beef cows treated with CIDR–B devices for timed artificial
insemination. Theriogenology. 2003. 59: 214.

Capítulo 1 69
10. Baruselli P, Marques MO, Hothnann EM, Costa Neto WP, Grandinetti RR, B6 G.
Increased pregnancy rates in embryo recipients treated with CIDR-B devices.
Theriogenology. 2001. 55:157 (abstr).
11. Baruselli, P., Marques, M., Madureira, E., Costa Neto, W., Gradinettie, R., Bo, G.
Increased pregnancy rates in embryo recipients treated whith CIDR-B devices.
Theriogenology. 2001. 55: Abstract 55 – 355.
12. Barrett D.W, Bartlewski PM, Duggaathi R, Davies K.L, Rawlings NC. Suppression of
follicle wave emergence in cycle ewes by Supraphysiologic concentrations of
estradiol 17 beta and induction with a physiologic dose of exogenous ovine follicle
stimulating hormone. Biol Reprod 2006. 75: 633-641.
13. Barros CM, Ereno RL, Machado MF, Buratini Jr, Pegorer MF, Simões, RAL, Satrapa
RA. Gene expression of luteinizing hormone receptor (LHr) isoforms in granulosa
cells of follicles from Nelore heifers before, during and after follicular deviation.
Reprod. Fertil. 2009.Dev. 21, 187 (abstract).
14. Barreto, A. and MacManus, C. The effect of progestagen CIDR device on pregnancy
rates in bovine embryo transfer recipients. Theriogenology. 2002. 57: 1974.
15. Barnea E, Choi Y, Leavis P. Embryo maternal signaling prior to implantation. En:
textbook of obstetrics ginecology I. Munteanu. Ed. SIEP. Boston 2000.
16. Belkys J. Vásquez M, Bastidas P. Comportamiento reproductivo de vacas Brahman
de primera lactancia suplementadas con proteína no degradable Zootecnia Trop.
2005. v.23 n.4.
17. Beg MA. and Guither OJ. Follicle selection in cattle and horses: Role of intrafollicular
factors Reproduction. 2006. 132: 365-377.
18. Beg MA, Bergfelt DR, Kot K. and Guither OJ. Follicle selection in cattle: Dinamics of
follicular fluid factors during development of follicle dominance. Biol. Reprod. 2002.
66: 120–126.
19. Beltman ME, Lonergan P, Diskin MG, Roche JF, Crowe MA. Effect of progesterone
supplementation in the first week post conception on embryo survival in beef
heifers Original Research Article Theriogenology, Volume 71, Issue 7, 15 April 2009,
Pages 1173-1179.
29. Beltmana P, Lonergana BM, Diskinc M, Rochea JF, Crowea MA. Oestrous cycles in
Bos taurus cattle. Contents lists available at. Animal Reproduction Science 124
(2011) 163–169.

70 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
20. Benyo DF and Pate JL. Tumor necrosis facto alfa alter bovine luteal cell synthetic
capacity and viability. Endocrinology. 1992. 43: 130. 854–860.
21. Bergfelt D.R. and Ginther O.J. Relationships between FSH surges and follicular
waves during the estrous cycle in mares. Theriogenology 1993. 39: 781–796.
22. Berisha B and Schams D. Ovarian function in ruminants. Domestic Animal
Endocrinology 2005. 29: 305-317.
23. Bilby TR, Guzelogtu A, kamimura S, Pancarci SM, Michel F, And Head HH.
Pregnancy and bovine somatotropin in nonlactaing dairy cows. I. Ovarian, conceptus,
and insulin – like growth factor system responses. J. Dairy Sci. 2004. 87, 32: 56–67.
24. Binelli M, Thatcher W, Mattos R, Baruselli P. Antiluteolytic Strategies to Improve
Fertility in Cattle. Theriogenology 2001. 56: 1451–1463.
25. Block SS, Butler WR, Ehrhardt RA, Bell AW, Van Amburgh ME, Boisclair YR.
Decreased concentration of plasma leptin in periparturient dairy cows is caused by
negative energy balance. J Endocrinol 2001. 171(2):339–48.
26. Bó GA. Sincronización de celos para programas de Inseminación Artificial y
Transferencia de embriones Bovinos. Instituto de Reproducción Animal de Córdoba
Argentina (IRAC) 2003.
27. Bó GA, Baruselli PS, Moreno D, Cutaia L, Caccia M, Tribulo H. The control of
follicular wave development for self – pointed embryo transfer programs in cattle.
Theriogenology. 2002. 57: 53–72.
28. Bó, G., Berfegfet, D., Brogliatti, G., Pierson, R. and Mapletoft, R. Sistemic Vs. local
effect of exogenous estradiol on follicular development in heifers. Theriogenology.
1996. 45, 1: 333.
29. Bó, G., Berfegfet, D. y Mapletoft, R. 1996. Manipulación de la dinámica folicular en
ganado bovino: Su aplicación en programas de Transferencia de embriones.
Memorias, II Simposio internacional de reproducción animal. Cordoba, Argentina.
1996. 53-68.
30. Bó, G., Caccia, M., Tribulo, H., Adams, G., Pierson, R. and Mapletoft, R. 1995. Estrus
synchronization in cattle with estradiol-17B and CIDR-B vaginal devices.
Theriogenology. 1995. 43, 1: 340.
31. Bó, G., Cutaia, L. Implementación de programas de inseminación artificial en rodeos
de cría. Argentina. Memorias del VI Simposio Internacional de Reproducción Animal.
Instituto de Reproducción Animal de Córdoba Argentina. 2005. 326-332.

Capítulo 1 71
32. Bó, G., Pierson, R. and Mapletoft, R. 1991. The effect of estradiol valerate on
follicular dynamics and superovulatory response in cows with Synchro-Mate-B
implants. Theriogenology. 1991. 36, 1: 169-183.
33. Bodensteiner KJ. Wiltbank MC, Bergfelt DR. and Ginther OJ. Alterations in follicular
estradiol and gonadotropin receptors during development of bovine antral follicle. En:
Theriogenology. Vol. 45.1996. p. 499–512
34. Bousfield GR; Butnev VY.Identificación of twelve O-glycosylation sites in equine
chorionic gonadtropin beta and equine luteinizing hormone ss by solid-phase Edman
degradation. Biol Reprod 2001, 64: 136-147.
35. Braw-Tal R, Roth Z. Gene expression for LH receptor, 17 alpha-hydroxylase and
StAR in the theca interna of preantral and early antral follicles in the bovine ovary.
Reproduction 2005. 129: 453-61.
36. Brayman M, Thathiah A and Carson DD. MUC1: A multifunctional cell surface
component of reproductive tissue epithelia.Reproductive Biology and Endocrinology 2
4.Cooper DN 2002 Galectinomics: finding themes in complexity.Biochimica et
Biophysica Acta 2004. 1572: 209–231
37. Buratini Jr, Price J, Visintin CA, Bó JA. Effects of dominant follicle aspiration and
treatment with recombinant bovine somatotropin (BST) on ovarian follicular
development in nelore (Bos indicus) heifers. Theriogenology. 2000. 54, 421–432.
38. Burkhart MN, Juengel JL, SmithPR, Heath DA, PerryGA, Smith MF, Garverick HA.
Morphological development and characterization of aromatase and estrogen
receptors alpha and beta in fetal ovaries of cattle from days 110 to 250. Animal
Reproduction Science, Volume 117, Issues 1-2, January 2010, Pages 43-54.
39. Campbell P, Anthony A, Peters T. Bioquímica y biología molecular en la era
posgenómica. Elsevier. 2006.
40. Campbell BK. The modulation of gonadotrophic hormone action on the ovary by
paracrine and autocrine factors. Reprod. Dom. Anim. 1999. 34: 147-152.
41. Campanile G, Di Palo R, Neglia G, Vecchio D, Gasparrini B. Corpus luteum and
embryonic mortality in buffaloes treated with a GnRH agonist, hCG and
progesterone. Theriogenology 2007. 67: 1393–1398.
42. Canty MJ, Boland MP, Evans AC, Crowe MA. Alterations in follicular IGFBP mRNA
expression and follicular fluid IGFBP concentrations during the first follicle wave in
beef heifers. Anim. Reprod. 2006.Sci. 93, 199–217.

72 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
43. Cardoso J, Diaz F, Hernandez A. Estroes cycle characteristics and blood
progesterone levels in Holstein heifers under altitude and tropical conditions in
Colombia. Tropicultura 1994. 12: 148–151.
44. Carvalho JB, Carvalho NA, Reis EL, Nichi M, Souza AH, Baruselli PS. Effect of early
luteolysis in progesterone-based timed AI rotocols in Bos indicus, Bos indicus x Bos
taurus, and Bos taurus heifers. Theriogenology 2008. 69, 167–175.
45. Castilho C, Garcia JM, Renesto A, Nogueira GP, Brito LF. Follicular dynamics and
plasma FSH and progesterone concentrations during follicular deviation in the first
post-ovulatory wave in Nelore (Bos indicus) heifers. Anim. Reprod. Sci. 2007. 98,
189–196.
46. Cattle R. Sartori A, Barrosb CM. Reproductive cycles in Bos indicus Contents lists
available at Animal Reproduction Science 124 (2011). 244–250.
47. Celemin, A., Cifuentes, E. 2003. Sincronización de celos y la ovulación para IATF en
vacas Holstein en anestro posparto en la sabana de Bogotá. Memorias IV Seminario
Internacional de reproducción en grandes animales, CGR Biotecnología
Reproductiva. Bogotá, Colombia. 2003.
48. Chin D, Means AR. "Calmodulin: a prototypical calcium sensor". Trends Cell Biol.
2000. 10 (8): 322–8.
49. Colazo, M., Bartolomé, J., Schmitdt, E., illuminati, H., Serrano, B., Moyano, B.,
Moralejo, R. 1996. Sincronización de los celos combinando progestagenos, GnRH,
PGF2α, e inseminacion a tiempo fijo en vacas para carne. Memorias, II Simposio
internacional de reproducción animal. Cordoba, Argentina. 1996. 254
50. Crowe MA, Enright WJ, Boland MP, Roche JF, Follicular growth and serum follicle-
stimulating hormone (FSH) responses to recombinant bovine FSH in GnRH-
immunized anoestrous heifers. Anim. Sci. 2001. 73, 115–122.
51. Crowe MA, Kelly P, Driancourt MA, Boland MP and Roche JF. Effects of follicle-
stimulating hormone with and without luteinizing hormone on serum hormone
concentrations, follicle growth, and intrafollicular. 2001.
52. Cunningham J,Klein Bradley. Fisiología veterinaria. cuarta edición. Elsevier
Saunders. 2009.
53. Dailey RA, Inskeep EK, Lewis PL. Pregnancy failures in cattle: a perspective on
embryo loss. In: Proceedings of the XVIIIth International Conference on Reproduction
of Farm Animals, Slovakia, 2002. p, 1–8.

Capítulo 1 73
54. David L and Michael M. Cox. Lehninger Principles of Biochemistry. 2000, third
edition. Ed. Wort. Cap. 5.
55. De la Sota RL, Crudeli GA, Torres Jimenez G. Repeated synchronization of ovulation
and timed insemination in commercial brahman and bradford cows. 14th ICAR.
Stockholm, Sweden. 2000. 2-6 July. Vol 2:97.
56. Donadeu FX and Ginther OJ. Follicular waves and circulating concentrations of
gonadotropins, inhibin, and estradiol during the anovulatory season in mares.
Reproduction 2002. 124: 875–885.
57. Dos Santos RM, Goissis MD, Fantini DA, Bertan CM, Moraes JL, Vasconcelos,
Binelli Elevated progesterone concentrations enhance prostaglandin F2α synthesis in
dairy cows. Animal Reproduction Science, Volume 114, Issues 1-3, August 2009,
Pages 62-71.
58. Driancourt MA, Reynaud K, Cortvrindt R, Smitz J. Roles of KIT and KIT LIGAND in
ovarian function. Rev. Reprod. 2000. 5: 143–152.
59. Driancourt MA. Follicular dynamics in sheep and cattle. Theriogenology 1991. 35, 55-
79.
60. Espey LL, Ujioka T, Russell DL, Skelsey M, Vladu B, Robker RL, Okamura H,
Richards JS. Induction of early growth response protein-1 gene expression in the rat
ovary in response to an ovulatory dose of human chorionic gonadotropin.
Endocrinology 2000. 141: 2385–2391
61. Everton L, Baruselli P. Sincronización de receptoras cruce por cebú en condiciones
tropicales. Cuarto seminario de reproducción de grandes animales (memorias)
Bogotá, 2003.
62. Fernandes P, Teixeira AB, Crocci AJ, Barros CM. Timed artificial insemination in beef
cattle using GnRH agonist, PGF2alpha and estradiol benzoate (EB). Theriogenology
2001. 55, 1521–1532.
63. Fike KE, Kojima FN, Lindsey BR, Bergfeld EG, Quintal-Franco JA, Melvin EJ, Zanella
EL, Wehrman ME, Kinder JE. Regulation of frequency of luteinizing hormone pulses
by magnitude of acute change in circulating concentration of progesterone of female
cattle. Animal Reproduction Science, Volume 84, Issues 3-4, September 2004,
Pages 279-291.
64. Filmore D. Modern Drug Discovery (American Chemical Society). 2004 (November):
pp. 24-28.

74 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
65. Filho MF, Ayres H, Ferreira RM, Marques MO, Reis EL. Silva RCP, Rodrigues CA,
Madureira EH, Bo´ GA, Baruselli P. Equine chorionic gonadotropin and gonadotropin-
releasing hormone enhance fertility in a norgestomet-based, timed artificial
insemination protocol in suckled Nelore (Bos indicus) cows. Theriogenology 2010.
73: 651–658.
66. Findlay JK, Drummond AE, Dyson ML, Baillie AJ, Robertson, DM, Ethier JF,
Recruitment and development of the follicle; the roles of the transforming growth
factor-beta superfamily. Mol. Cell.Endocrinol. 2004.191, 35–43.
67. Fleming JA, Choi Y, Johnson GA, Spencer TE, and Bazer FW. Cloning of the ovine
estrogen receptor-alpha promoter and functional regulation by ovine interferon-tau.
Endocrinology. 2001. 142: 2879–2887.
68. Fortune JE, Rivera GM and Yang MY. Follicular development: the role of the follicular
microenvironment in selection of the dominant follicle. Animal Reproduction Science
2004. 82, 83: 109–126.
69. Fortune EJ, Rivera MG, Evans ACO, Turzillo AM. Differentiation of dominant versus
subordinate follicles in cattle.Biol Reprod 2001. 65: 648-654.
70. Biol. Reprod. 1995. 52: 1020–1026.
71. Fuchs AR, Ivell R, Fielt PA, Chang SM And Fields MJ. Oxitocin receptors in the
bovine cervix: Distribution and gene expression during the estrous cycle. Biol.
Reprod. 1996. 54: 700–708.
72. Fuentes S, De la Fuente J. Tasas de gestación de novillas receptoras sincronizadas
con Gonadotropina Coriónica Equina u Hormona Folículo Estimulante. Acta Scientiae
Veterinariae 2007. 35 (Supl. 3): s 767 – s 772.
73. Gabriel HG, Wallenhorst S, Dietrich E, Holtz W. The effect of prostaglandin F2α
administration at the time of insemination on the pregnancy rate of dairy
cows. Animal Reproduction Science, Volume 123, Issues 1-2, January 2011, Pages
1-4.
74. Garverick HA, Juengel JL, Smith P, Heath DA, Burkhart MN, Perry GA, Smith MF,
McNatty KP. Development of the ovary and ontongeny of mRNA and protein for P40
aromatase (arom) and estrogen receptors (ER) α and β during early fetal life in cattle.
Animal Reproduction Science, Volume 117, Issues 1-2, January 2010, Pages 24-33.
75. Gerardd J. Tortora Bryan Derrickson. Principios de anatomía y fisiología. 11edicion
2006 editorial medica panamericana, S.A. de C.V.

Capítulo 1 75
76. Ginther OJ, Beg MA, Donadeu FX and Bergfelt DR. Mechanism of follicle deviation in
monovular farm species. Animal Reproduction Science 2003. 78: 239 –257.
77. Ginther OJ, Beg MA, Bergfelt DR, Donadeu FX, Kot K. Follicle selection in monovular
species. Biol. Reprod. 2001. 65: 638–647.
78. Girsh E, Wang W, Mamluk R, Arditi F, Friedman A, Milvae RA And Median R.
Regulation of endothelin – 1 expression in the bovine corpus luteum: elevation by
prostaglandin F2 alfa. Endocrinology 1996. 137: 5191–5196.
79. Gómez, C. 2005. Transferencia de embriones experiencias en Colombia. Memorias,
congreso internacional de reproducción bovina (Intervet), Bogotá Colombia. 2005.
155-158.
80. Gomez E, Diez C. Effects of glucose and protein sources on bovine embryo
development in vitro. Animal Reproduction Science, Volume 58, Issues 1-2, 28
February 2000, Pages 23-37.
81. Gonella, A., Posible relación de los niveles séricos ováricos (17 Beta estradiol y
Progesterona), sus respectivos receptores endometriales (ER-1 alfa y PGR) y
loscambios histológicos del endometrio durante el ciclo estral en hembras cíclicas de
tres razas bovinas. Tesis de Maestría. Facultad de Medicina Veterinaria y de
Zootecnia. Universidad Nacional de Colombia – Sede Bogotá D.C. 2010.
82. Gordon, I. Reproducción controlada del ganado vacuno y búfalos. Zaragoza España.
Ed. Acribia. 1999.
83. Gouveira, M., Melo, D., Carvalho, L., Fuck, E., Trinca, L., Moraes, C. 2004. Do high
progesterone concentrations decrease pregnancy rates in embryo recipients
synchronized with PGF2α and eCG. Theriogenology. 61, 7-8: 1283-1290.
84. Grajales H, Tovío N, Duica A. Fundamentos de fisiología reproductiva en la hembra
bovina. Primera edición. 2011.
85. Grajales, H.; Tovío N. Aspectos Básicos del ciclo estral de la vaca. Revista Genética
Bovina. 2009. 32-36. Volumen 15.
86. Gray CA, Adelson DL, Bazer FW, Burghardt RC, Meeusen EN and Spencer TE.
Discovery and characterization of an epithelialspecificgalectin in the endometrium
that forms crystals in thetrophectoderm. PNAS 2004. 101: 7982–7987
87. Green JC, Okamura CS, Poock SE, Lucy MC. Measurement of interferon-tau (IFN-τ)
stimulated gene expression in blood leukocytes for pregnancy diagnosis within 18–
20 d after insemination in dairy cattle. Animal Reproduction Science, Volume 121,
Issues 1-2, August 2010, Pages 24-33.

76 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
88. Gwazdauskas, F.V.; Kendrick, K.W.; Pryor, A.W. and Balley, T.I. Symposium:
Folliculogenesis in the bovine ovary. J. Dairy Sci. 2000; 83:1625 – 1634.
89. Hafez E. Reproducción e inseminación de los animales domésticos, 7ª ed.
Reproductive Health Center IVF/Andrology International. Kiawah Island, South
Carolina, USA. McGraw – Hill Interamericana. 2000. 33–144.
90. Hanrahan, JP, Gregan SM, Mulsant P, Mullen M, Davis GH, Powell R and Galloway
SM. Mutations in the genes for oocyte-derived growth factors GDF9 and BMP15 are
associated with both increased ovulation rate and sterility in Cambridge and Belclare
sheep (Ovis Aries). Biol. Reprod. 2004. 70: 900–909.
91. Hastie PM, Haresign W. Modulating peripheral gonadotrophin levels affects follicular
expression of mRNAs encoding insulin-like growth factor binding proteins in sheep.
Animal Reproduction Science, Volume 119, Issues 3-4, June 2010, Pages 198-204.
92. Hastie PM, Haresign W. Modulating peripheral gonadotrophin levels affects follicular
expression of mRNAs encoding insulink-like growth factors and receptors in sheep.
Animal Reproduction Science, Volume 109, Issues 1-4, December 2008, Pages 110-
123.
93. Hastie PM, Haresign W. Expression of mRNAs encoding insulin-like growth factor
(IGF) ligands, IGF receptors and IGF binding proteins during follicular growth and
atresia in the ovine ovary throughout the oestrous cycle. Animal Reproduction
Science, Volume 92, Issues 3- May 2006, Pages 284-299.
94. Hehnke–Vagnoni, K, Clark C, Taylor M and Ford S. Presence and localization of
tumor necrosis factor – Alfa in the corpus luteum of monpregnant pig. Biol. Reprod.
1995. 53: 1339–1344.
95. Henao G, Trujillo L. Establecimiento y desarrollo de la dominancia folicular bovina.
Revista Colombiana de Ciencias Pecuarias. 2000 Vol. 13 No.2: 108-120.
96. Hoffman LH and Wooding FB. Giant and binucleate trophoblast cells of mammals.
Journal of Experimental Zoology. 1993. 266: 559–577.
97. Huanca W. Inseminación artificial a tiempo fijo en vacas lecheras. Rev. investig. vet.
Perú, 2001. v.12 n.2 Lima jul./dic.
98. Hussein MR. Apoptosis in the ovary: molecular mechanisms. Hum Reprod Update
2005. 11:162-178.
99. Igwebuike UM. Impact of maternal nutrition on ovine foetoplacental development: A
review of the role of of insulin-like growth factors. Animal Reproduction Science,
Volume 121, Issues 3-4, September 2010, Pages 189-196.

Capítulo 1 77
100. Igwebuike UM. Trophoblast cells of ruminant placentas—A minireview. Animal
Reproduction Science, Volume 93, Issues 3-4, July 2006, Pages 185-198.
101. Ireland JJ, Mihn M, Austin E, Diskin MG and Roche JF. Historical perspective of
turnover of dominant follicles during the bovine estrous cycle: Key concepts, studies,
advancements and terms. J. Dairy Sci. 2000. 83: 1648 -1658.
102. Jang M, Mistry A, Swick AG, Romsos DR. Leptin rapidly inhibits hypothalamic
neuropeptide Y secretion and stimulates corticotropin-releasing hormone secretion in
adrenalectomized mice. J Nutr 2000. 130 (11): 2813–20.
103. Jiménez C. y Hernández A. Lecturas sobre reproducción bovina. II. El ciclo estral
de la vaca. Facultad de Medicina Veterinaria y de Zootecnia. Universidad Nacional
de Colombia. 2000.
104. Johnson M. Intrinsec and extrinsec factors in preimplantation development Reprod
Fert 1979. 55: 255.
105. Johnson GA, Burghardt RC, Spencer TE, Newton GR, Ott TL, Bazer FW. Ovine
osteopontin II. Osteopontin and αvβ3 integrin expression in the ovine uterus and
conceptus during the peri-implantation period. Biol Reprod 1999. 61: 892–899.
106. Johnson GA, Spencer TE, Burghardt RL, Taylor KM, Gray CM, BazerFW.
Proyesterone modulation of osteopontin gives expression in ovine uterus. Biol
Reprod 2000. 42: in press. 104.
107. Johnson GA, Burghardt RL, Bazer FW, and Spencer TE. Osteopontin: roles in
implantation and placentation. Biol Reprod 2003. 69: 1458–1471.
108. Joyce IM, Pendola FL, Wigglesworth K, Eppig JJ. Oocyte regulation of kit ligand
expression in mouse ovarian follicles. Dev. Biol. 1999. 214: 342–353.
109. Juengel JL, Garverick HA, Johnson AL, Youngquist RS. And Smith MF. Apoptosis
during luteal regressión in cattle. Endocrinology. 1993; 132. 249–254.
110. Kastelic P, Bergfelt DR, Ginther OJ. Relationship between ultrasonic assessment
of the corpus luteum and plasma progesterone concentration in heifers.
Theriogenology. 1990. 33, 6: 1269 – 1278.
111. Kastelic J, Pierson R and Ginther O. Ultrasonic Morphology of a corpora lutea and
central luteal cavities during the estrous cycle and early pregnancy in heifers.
Theriogenology. 1990. 34, 3: 487-498.
112. Katagiri S, Takahashi Y. Potential relationship between normalization of
endometrial epidermal growth factor profile and restoration of fertility in repeat

78 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
breeder cows. Animal Reproduction Science, Volume 95, Issues 1-2, September
2006, Pages 54-66.
113. Kayser JPR, Kim JG, Cerny RL, Vallet J. Global characterization of porcine
intrauterine proteins during early pregnancy. Reproduction. 2006. 132: 379-388.
114. Kaplan JH. Biochemistry of K.ATPase. Annu Rev Biochem 2002. 71: 511–535.
115. Kerbler TL, Buhr MM, Jordan LT, Leslie KE, Walton JS. Relationship between
maternal plasma progesterone concentration and interferon-tau synthesis by the
conceptus in cattle. Theriogenology 1997. 47: 703-714.
116. Kimura K, Iwata H, Thompson JG. The effect of glucosamine concentration on the
development and sex ratio of bovine embryos. Animal Reproduction Science, Volume
103, Issues 3-4, 30 January 2008, Pages 228-238.
117. Klisch, K, Wooding, FBP and Jones, CJP. The Glycosylation Pattern of Secretory
Granules in Binucleate Trophoblast Cells is Highly Conserved in Ruminants.
Placenta. 2009..
118. King N, Hittinger CT, Carroll SB. Evolution of key cell signaling and adhesion
protein families predates animal origins». Science 2003. 301 (5631): pp. 361-3.
119. Knight PG, Glister C, Local roles of TGF-beta superfamily members in the control
of ovarian follicle development. Anim. Reprod. Sci. 2003. 78, 165–183.
120. Knight PG, Glister C. Potential local regulatory functions of inhibins, activins and
follistatin in the ovary. Reproduction. 2001. 121: 503-512.
121. Knobil E. and Neill J. Physiology of reproduction. Third edition. Academic Press is
an imprint of El sevier. 2006.
122. Lim H, Paria BC, Das SK, Dinchuk JE, Langenbach R, Trzaskos JM, Dey SK.
Multiple female reproductive failures in cyclooxygenase2-deficient mice. Cell. 1997.
91:197–208.
123. Liu X, Q Dai, Rawlings NC. Ultrasonographic image attributes of non-ovulatory
follicles and follicles with different luteal outcomes in gonadotropin-releasing hormone
(GnRH)-treated anestrous ewes. Theriogenology. 2007. 67, 957-969.
124. Lopes AS, Butler ST, Gilbert RO, Butler WR. Relationship of pre-ovulatory follicle
size, estradiol concentrations and season to pregnancy outcome in dairy cows. Anim
Reprod Sci 2007. 99: 34-43.
125. Lucy MC. Regulation of ovarian follicular growth by somatotropin and insuline like
growth factors in cattle. J. Dairy Sci. 2000. 83: 1635–1647.

Capítulo 1 79
126. Lüttgenau J, Beindorff N, Ulbrich SE, Kastelic JP, Bollwein H. Low plasma
progesterone concentrations are accompanied by reduced luteal blood flow and
increased size of the dominant follicle in dairy cows.Theriogenology, In Press,
Corrected Proof, Available online 6 May 2011.
127. Nakamura T, Sakamoto K. Reactive oxygen species up-regulates
cyclooxygenase-2, p53, and Bax mRNA expression in bovine luteal cells. Biochem
Biophys Res Commun 2001. 284: 203-210.
128. Nasser L, Reis E, Oliveira M, Bó GA, Baruselli P. The use of hormonal treatments
to improve reproductive performance of anestrous beef cattle in tropical climates.
Anim Reprod Sci 2004. 82, 83: 479–486.
129. Niemann H, Wilfried A. Kues Application of transgenesis in livestock for agriculture
and biomedicine. Animal Reproduction Science, Volume 79, Issues 3-4, 15
December 2003, Pages 291-317.
130. Niswender GD. Molecular control of luteal secretion of progesterone.
Reproduction. 2002. 123: 333–339. Fort Collins, CO 80523-1683, USA
131. Niswender GD, Juengel JL, Silva PJ, Rollyson MK and McIntush EW. Mechanisms
controlling the function and life span of the corpus luteum. Physiological. Reviews.
2000. 222. 80: 1-29.
132. Niswender GD, Juengel JL, McGuire WJ, Belifore CJ. And Witbank MC. Luteal
function: The estrous cycle andearly pregnancy. Biol. Reprod. 1994. 50: 239-247.
133. McDonald´s. Edi. Pineda MH. Veterinary endocrinology and reproduction, Fifth
edition. 2003.
134. Mackey, DR, Wylie ARG, Sreenan JM, Roche JF and Diskin MG. The effect of
acute nutritional change on follicle wave turnover, gonadotropin and steroid
concentration in beef heifers. J. Ani. Sci. 2000. 78: 429–442.
135. Madureira EH, Marques MO, Baruselli P, Nasser LF, Rodrigues CA. Efeito do
tratamiento com eCG na taxa de concepção de vacas nelore com diferentes escores
de condição corporal inseminadas em tempo fixo (analise Retrospectiva). Acta
Scientiae veterinarae .2004. 32 (suplemento): p 228.
136. Mann GE, Lamming GE. Relationship between maternal endocrine environment,
early embryo development and inhibition of the luteolytic mechanism in cows. J.
Reprod. Fertil. 2001. 121, 175–180.

80 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
137. Mann GE. Timing of prostaglandin F2α release episodes and oxytocin receptor
development during luteolysis in the cow.Animal Reproduction Science, 2006.
Volume 93, Issues 3-4, Pages 328-336.
138. Mann G, Fray M. Lamming, G. Effects of time of progesterone supplementation on
embryo development and interferon-tau production in the cow. The Veterinary
Journal 2006. 171: 500–503.
139. Manikkam M, Rajamahendran R. Progesterone induced atresia of the proestrus
dominantfollicle in the bovine ovary: changesin diameter, insulin-like growth factor
ystem, aromatase activity, steroid hormones and apoptotic index. Biol Repod 1997.
57: 580-587.
140. Mantovani A, Baruselli P, Bó GA, Cavalcante A, Gacek F. Aumento de las
dimensiones del folículo dominante y del cuerpo lúteo, concentración plasmática de
progesterona y de la tasa de aprovechamiento de receptoras de embriones bovinos
sincronizadas con CIDR-B® por tiempo prolongado. Revista Brasilera de
Reproducción Animal 2002.
141. Mamani E, Ortiz J, Quezada M. Evaluación de diferentes dosis de eCG en
tratamientos de sincronización de celos en receptoras de embriones. VII simposio de
reproducción animal – IRAC. 2007. p, 275.
142. Martinez, M., Adams, G., Kastelic, J., Bergfelt, D., Mapletoft, R. Induction of
follicular wave emergence for estrus synchronization and artificial insemination in
heifers. Theriogenology. 2000. 54: 757-769.
143. Mazerbourg S, Overgaard MT, Oxvig C, Christiansen M, Conover CA, Laurendeau
I, Vidaud M, Tosser-Klopp G, Zapf J, Monget P, Pregnancy-associated plasma rotein
– A (PAPP-A) in ovine, bovine, porcine and equine ovarian follicles: involvement in
IGF binding protein – 4 proteolytic degradation and mRNA expression during follicular
development. Endocrinology 2001.142: 5243-5253.
144. Mc Donald LE. and Capen CC. Endocrinología Veterinaria y Reproducción.
Editado por Mc Donald LE y Pineda MH. Editorial Interamericana-McGraw-Hill.
Mexico. 1991.
145. Meduri G, Bachelot A, Cocca MP, Vasseur C, Rodien P, Kuttenn F,Touraine P and
Misrahi M. Molecular pathology of the FSH receptor: New insights into FSH
physiology Molecular and Cellular Endocrinology Volume 282, Issues 1-2, 30 January
2008, Pages 130-142.

Capítulo 1 81
146. Melanie J, Starbuck A, Dailey E, Keith I. Factors affecting retention of early
pregnancy in dairy cattle Anim Reprod Sci 2004. 84: 27–39.
147. Mihm M, Austin EJ, Good TEM, Ireland JLH, Knight PG, Roche JF and Iteland JJ.
Identification of potential intrafollicular factors involved in selection of dominant
follicles in heifers. Biol. Reprod. 2000. 63: 811-819.
148. 146. Milvae RA, Hinckley ST, Carlson JC. Luteotropic and luteolytic mechanisms
in the bovine corpus luteum. Theriogenology 1996. 45:1327–49.
149. Mollo MR, Rumpf R, Martins AC, Mattos MCC, Lopes Jr, Carrijo LHD. Sartori R.
Ovarian function in Nelore heifers under low or high feed intake. Acta Sci. Vet. 2007.
35 (Suppl. 3), 958 (abstract).
150. Montaño E, Ruiz Z. ¿Por qué no ovulan los primeros folículos dominantes de las
vacas cebú posparto en el trópico colombiano? Rev. Col. Cienc. Pec. 2005 Vol. 18:2.
151. Moreira F, Badinga L, Burnley C, Thatcher WW. Bovine somatotropin increases
embryonic development in superovulated cows and improves post-transfer
pregnancy rates when given to lactating recipient cows. Theriogenology 2004. 57:
1371–87.
152. Mufeed A. Alnimer, Wadie F. Lubbadeh. Effect of progesterone (P4) intravaginal
device CIDR to reduce embryonic loss and to synchronize return to oestrus of
previously timed inseminated lactating dairy cows. Animal Reproduction Science,
Volume 107, Issues 1-2, August 2008, Pages 36-47.
153. Murphy BD. Models of luteinization. Biol. Reprod. 2000. 63: 2-11.
154. Murray R. Harper Bioquimica ilustrada. 17 edición. Manual Moderno 2007.
155. Olexikova L, Makarevich AV, Pivko J, Chrenek P. Antibody to Hsp70 alters
response of rabbit preimplantation embryos to hyperthermia in vitro. Animal
Reproduction Science, Volume 119, Issues 1-2, May 2010, Pages 130-136.
156. Olhagaray R. Comparacion de cuatro protocolos de sincronizacion de celo en
vacas holstein ovsynch, cidr-b, pgf2α dosis reducida, natural (prov. warnes, dpto.
santa cruz). Tesis de grado Universidad Autonoma Gabriel Rene Moreno. Facultad
de Medicina Veterinaria y Zootecnia Santa Cruz de la Sierra – Bolivia 2004.
157. Olivera AM, Tarazona MA, Ruíz CT, Giraldo EC. Modelo de luteólisis bovina. Rev
Col Cienc Pec 2007. 20: 387-393.
158. Olivera M. Gestación. En Galina C. (Ed), Reproducción de animales domésticos.
Edi. Limusa. México. 2006. p, 165–187.

82 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
159. Padmanabhan V, Battaglia D, Brown B, Karsch FJ, Lee JS, Phillips DJ and Cleeff
V. Neuroendocrine control of follicle stimulating hormone FSH secretion mediated via
changes in activin availability and does in involve changes in gonadotropin –
releasing hormone secretion. Biol. Repro. 2002. 66: 1395–1402.
160. Palma, G. Biotecnología de la reproducción, tercera edición. Buenos Aires,
Argentina. Ed. INTA. 2008.
161. Pru JK, Hendry IR, Davis JS, Rueda BR. Soluble Fas ligand activates the
sphingomyelin pathway and induces apoptosis in luteal steroidogenic cells
independently of stress-activated p38 MAPK). Endocrinology 2002. 143: 4350-4357.
162. Park JI, Park HJ, Choi HS, Lee K, Lee WK, Chun SY. Gonadotropin regulation of
NGFI-B messenger ribonucleic acid expressionduring ovarian follicle development in
the rat. Endocrinology 2001. 142:3051–3059.
163. Pate JL. Cellular components involved in luteolysis. J. Anim. Sci. 1994. 72. 1884–
1890.
164. Pascual-Leone AM. Regulación del crecimiento: axis GH/IGFs e hipótesis
neuroendocrina1. Anal. Real Acad. Farm.: 2001, 67.
165. Peixoto MGCD, Bergmann JAG, Suyama E, Carvalho MRS, Penna VM. Logistic
regression analysis of pregnancy rate following transfer of Bos indicus embryos into
Bos indicus _ Bos taurus heifers. Theriogenology 2007. 67: 287–292
166. Peña M, Góngora A, Estrada J. Factores de crecimiento en el desarrollo folicular,
embrionario temprano e implantación. Implicaciones en la producción de embriones
bovinos. Journal MVZ Córdoba 2007. 12 (1): 942–954.
167. Perera-Marín G, Murcia C, Rojas S, Hernández-Cerón J, González-Padilla E.
Pattern of circulating luteinizing hormone isoforms during the estrous and luteal
phases in Holstein heifers. Animal Reproduction Science, Volume 86, Issues 1-2,
March 2005, Pages 53-69.
168. Pereira RM, Carvalhais I, Pimenta J, Baptista MC, Vasques MI, Horta AEM,
Santos IC, Marques MR, Reis A, Silva M, Pereira C, Marques C. Biopsied and
vitrified bovine embryos viability is improved by trans10, cis12 conjugated linoleic
acid supplementation during in vitro embryos culture. Animal Reproduction Science,
Volume 106, Issues 3-4, July 2008, Pages 322-332.
169. Peres LC. Tratamientos de sincronización de la ovulación y factores que afectan
las tasas de preñez en receptoras de embriones bovinos congelados en etilenglicol y

Capítulo 1 83
transferidos a tiempo fijo. Tesis de Maestría, Universidad Nacional de Córdoba.
2005/06.
170. Pinheiro VG, Souza MF, Pegorer RA. Satrapa RL. Ereno LA, Trinca CM. Barros.
Effects of temporary calf removal and eCG on pregnancy rates to timed-insemination
in progesterone-treated postpartum Nellore cows. Theriogenology, Volume 71, Issue
3, February 2009, Pages 519-524.
171. Pretheeban T, Balendran A, Gordon MB, Rajamahendran R. mRNA of luteal
genes associated with progesterone synthesis, maintenance, and apoptosis in dairy
heifers and lactating dairy cows. Animal Reproduction Science, Volume 121, Issues
3-4, September 2010, Pages 218-224.
172. Pursley JR, Wiltbank MC, Stevenson JS, Ottobre JS, Garverick HA and Anderson
II. pregnancy rates per ai for cows and heifers inseminated at a synchronized
ovulation or synchronizes estrus. j. dairy sci. 1997.80:295.
173. Quesada JM y Ortiz JJ. Sincronización de receptoras de embriones tratadas con
CIDR, benzoato de estradio, D Cloprostenol asociado o no a eCG. VII simposio de
reproducción animal – IRAC. 2007. p, 277.
174. Rabiee ARK. Macmillan L and. Schwarzenberger F. Evaluation the effect of feed
intake on progesterone clearance rate by measuring faecal progesterone metabolites
in grazing dairy cows. Anim. Reprod. Asci. 2001. 67: 205–214.
175. Rajakoski E. The ovarian follicular system in sexually mature heifers with special
reference to seasonal, cyclical, and left–right variations. Acta Endocrinol 1960; 34:7–
68.
176. Ranferi G, Cejudo E, Carranza L, Hernández G. Síndrome de hiperestimulación
ovárica. Revista Mexicana de Medicina de la Reproducción 2010; 2(3):67-73
177. Rathbone MJ, Kinder JE, Fike K, Kojima F, Clopton D, Ogle CR, Bunt CR. Recent
advances in bovine reproductive endocrinology and physiology and their impact on
drug delivery system design for the control of the estrous cycle in cattle. Adv Drug
Deliv Rev 2001. 50: 277-320.
178. Recabarren SE, Lobos A, Poblete O, Muñoz P, Parilo J, Pulsatile follicle
stimulating hormone (FSH) secretion in prepubertal female sheep with and
without food restriction. Arch. med. vet. 2003. v.35 n.2.
179. Rezende M, Dalva M, Binelli M, Lopes M. Canine copus luteum regression:
Apoptosis and caspase-3 activity
Theriogenology, Volume 66, Issues 6-7, October 2006, Pages 1448-1453.

84 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
180. Ritindra N. Khan, Edward G. Tall, Rebecchi M. Effect of Maitotoxin on Guanine
Nucleotide Interaction with G-Protein subunits. International Journal of Toxicology
2001.20: 39-44.
181. Roberts M. Interferon-tau, a Type 1 interferon involved in maternal recognition of
pregnancy. Cytokine & Growth Factor Reviews 2007. 18: 403–408.
182. Roberts RM, Ealy AD, Alexenko AP, Han CS, Ezashi T. Trophoblast interferons.
Placenta 1999. 20: 259–264.
183. Rodrigues CA, Ayres H, Reis EL, Madureira EH, Baruselli PS. Aumento de taxa
de prenhez em vacas Nelore inseminadas em tempo fixo com o uso de eCG em
diferentes periodos pós-parto.; Acta Scientae veterinariae. V32 (Suplemento), 220.
2004.
184. Rodríguez J. Mecanismos para el reconocimiento materno de la preñez. En:
Reproducción Bovina. Editor González C. Maracaibo – Venezuela. Ed. Fundación
GIRARZ 2001. 31–39.
185. Rosales AM y Guzmán A. Apoptosis en la atresia folicular y regresión del CL.
Técnica Pecuaria Mex. 2008. 46 (2): 159–182.
186. Rosales-Torres AM, Alonso I, Vergara M, Romano MC, Castillo-Juárez H, Ávalos
A, Rosado A, Gutiérrez GC. Vascular endothelial growth factor isoforms 120, 164 and
205 are reduced with atresia in ovarian follicles of sheep. Animal Reproduction
Science, Volume 122, Issues 1-2, October 2010, Pages 111-117.
187. Rosales AM, Avalos A, Vergara M, Hernández O, Ballesteros LM, García R, et al.
Multiparametric study of atresia in ewe antral follicles: Histology, flow cytometry,
internucleosomal ADN fragmentation and lysosomal enzyme activities in granulosa
cells and follicular fluid. Molec Reprod Develop 2000. 55: 270-281.
188. Rosenkrans C Jr., Banks A, Reiter S, LooperM. Calving traits of crossbred
Brahman cows are associated with Heat Shock Protein 70 genetic polymorphisms.
Animal Reproduction Science, Volume 119, Issues 3-4, June 2010, Pages 178-182.
189. Rubianes E. Avances en el conocimiento de la fisiología ovárica de los pequeños
rumiantes y su aplicación para el manejo reproductivo Actas de Fisiología, 2000. 6:
93-103.
190. Rueda BR, Hendry IR, Hendry IW, Stormshak F, Slayden OD, et al. Decreased
progesterone levels and progesterone receptor antagonists promote apoptotic cell
death in bovine luteal cells. Biol Reprod 2000. 62: 269-276.

Capítulo 1 85
191. Safdarian M, M Kafi, M Hashemi. Reproductive performance of Karakul ewes
following different oestrous synchronization treatment outside the natural breeding
season. South Afric J Anim Sci. 2006. 36, 229-234.
192. Sangsritavong S, Combs DK, Sartori R, Armentano LE, Wiltbank MC. High feed
intake increases liver blood flow and metabolism of progesterone and estradiol 17
beta in dairy cattle. J. Dairy Sci. 2002. 85, 2831–2842.
193. Santos JE, Thatcher WW, Pool L, Overton MW. Effect of human chorionic
gonadotropin on luteal function and reproductive performance of high-producing
lactating Holstein dairy cows. J Anim Sci 2001;79:2881–94.
194. Sakase, M., Kawate, N., Nakagawa, C., Fukushima, M., Noda, M., Takeda, K.,
Ueno, S., Inaba, T., Kida, K., Tamada, H., Sawada. 2006. Preventive effects of CIDR-
based protocols on premature ovulation before timed-AI in Ovsynch in cycling beef
cows. The Veterinary Journal. In press.
195. Sartorelli ES, Carvalho LM, Bergfelt DR, Ginther OJ, Barros CM. Morphological
characterization of follicle deviation in Nelore (Bos indicus) heifers and cows.
Theriogenology. 2005. 63, 2382–2394.
196. Sartori R and Barros. Reproductive cycles in Bos indicus cattle. Animal
Reproduction Science. 2011.Volume 124, Issues 3-4, Pages 244-250
197. Sartori R, Haughian J, Shaver RD, Rosa GJ, Wiltbank MC. Comparison of Ovarian
Function and Circulating Steroids in Estrous Cycles of Holstein Heifers and Lactating
Cows. . Dairy Sci. 2004.87: 905–920.
198. Sartori GJ, Rosa M and Wiltbank MC. Ovarian Structures and Circulating Steroids
in Heifers and Lactating Cows in Summer and Lactating and Dry Cows in Winter J.
Dairy Sci. 2002. 85:2813–2822
199. Scott J. And Pawson T. Cell communication: The Inside Story. Sci. Am. 2000. 282
(6): 72-79.
200. Sharma RK. Follicular atresia in goat: A review. Indian J Anim Sci 2000. 70: 1035-
1046.
201. Shimizu T, Murayama C, Sudo N, Masa K, Akio T. Involvement of insulink and
growth hormone (GH) during follicular development in the bovine ovary. Animal
Reproduction Science, Volume 106, Issues 1-2, June 2008, Pages 143-152.
202. Senger PL. Pathways to pregnancy and parturition. Second edition revised. 2005.

86 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
203. Small JA, Colazo MG, Kastelic JP, Mapletoft RJ. Effects of progesterone
presynchronization and eCG on pregnancy rates to GnRH-based, timed-AI in beef
cattle. Theriogenology, Volume 71, Issue 4, 1 March 2009, Pages 698-706.
204. Shams D., Berisha B. Regulation of corpus luteum function in cattle – and
overview. Repro Dom Anim 2004. 39: 241-251.
205. Souza AH, Viechnieski S, Lima FA, Silva FF, Araujo R, Bo´ G, Wiltbank C,
Baruselli P. Effects of equine chorionic gonadotropin and type of ovulatory stimulus in
a timed-AI protocol on reproductive responses in dairy cows. Theriogenology 2009.
72:10–21.
206. Spell AR, Beal WE, Corah LR, Lamb GC. Evaluating recipients and embryo
factors that affeect pregnancy rates of embryo transfer in beef cattle. Theriogenology
2001. 56: 287–297.
207. Spencer T, Johnson G, Bazer F, Burghardt R. and Palmarini M. Pregnancy
recognition and conceptus implantation in domestic ruminants: roles of progesterone,
interferons and endogenous retroviruses. Reproduction Fertility and Development
2007.19: 65-78.
208. Spencer T, Burghardt R, Johnson G, Bazer FW. Conceptus signals for
establishment and maintenance of pregnancy. Anim Reprod Sci 2004. 83: 537– 550.
209. Spencer T, Johnson GA, Bazer FW and Burghardt RC. Implantation mechanisms:
insights from the sheep Center for Animal Biotechnology and Genomics, Public
Health, Texas A&M University. 2004.128: 657–668.
210. Spicer LJ, Schreiber NB, Lagaly DV, Aad PY, Douthit LB, Grado-Ahuir JA. Effect
of resistin on granulosa and theca cell function in cattle. Animal Reproduction
Science, Volume 124, Issues 1-2, March 2011, Pages 19-27.
211. Starbuck MJ, Dailey RA and Inskeep EK. Factors affecting retention of early
pregnancy in dairy cattle. Anim. Epro. Sci. 2004. (In press).
212. Stevenson JS. Areview of oestrous behaviour and detection in dairy cows. In:
Diskin, M.G. (Ed.), Proc. BSAS Occasional Publication No. 26, Fertility in the High-
Producing Dairy Cow. 2001. vol. 1, pp. 43–62.
213. Stevenson, J., Kobayashi, Y. and Thompson, K. Reproductive performance of
dairy cows in various programmed breeding systems including Ovsynch and
combinations of gonadotropinas releasing hormone and prostaglandin F2a. Journal
of Dairy Science.1999. 82: 506-515.

Capítulo 1 87
214. Stocco C.; Telleria C; Gibori G. The Molecular Control of Corpus Luteum
Formation, Function, and Regression. Endocrine Reviews 2007. 28(1): 117–149.
215. Thatcher W, Binelli M, Burke C, Staples JD and Coelho S. Antiluteolytic signals
between the conceptus and endometrium. Theriogenology. 1997. 47: 131-140.
216. Thatcher W, Guzeloglu A, Binelli M, Hansed TR, Pru JK. Uterine conceptus
interactions and reproductive failure in cattle. Theriogenology 2001. 56: 1435–1450.
217. Thatcher W, Moreira F, Santos J, Mattos R, lopez F, Pancarci S, Risco C. Effects
of hormonal treatments on reproductive performance and embryo production.
Theriogenology 2002. 55: 75– 90.
218. B.M. Toosi, S.V. Seekallu, D.M.W. Barrett, K.L. Davies, R. Duggavathi, E.T. Bagu,
N.C. Rawlings. Characteristics of peaks in serum concentrations of follicle-stimulating
hormone and estradiol, and follicular wave dynamics during the interovulatory interval
in cyclic ewes. Theriogenology, Volume 73, Issue 9, June 2010, Pages 1192-1201.
219. Tovío N, Duica A, Grajales H. Desarrollo embrionario y estrategias antiluteoliticas
hormonales en programas de transplante de embriones bovinos. Rev.MVZ Cordoba,
vol.13, no.1, p.1240-1251. 2008.
220. Tribulo, H. 2002. Curso de post-grado en reproducción bovina. Modulo IV.
Transferencia de embriones. CGR. Biotecnología reproductiva. Cap. 1 y 2.
221. Tribulo H, Moreno D, Cutaia L, Gatti G, Tribulo R, Caccia M, et al. Pregnancy
rates in embryo recipients treated with progesterone vaginal device and eCG and
transferred without estrus detection. Theriogenology 2002; 57: 563 [abstract].
222. Twagiramungu, H., Guilbault, L., Dufor, J. 1995. Synchronization of ovarian
follicular waves with a gonadotropin-releasing hormone agonist to increase the
precision of estrus in cattle. Journal Animal Science. 1995. 73: 3141-3151.
223. Vasconcelos J, Sartori R, Oliveira H, Guenther J, Wilbank M. Reduction is size of
the ovulatory follicle reduces subsequent luteal size and pregnancy rate.
Theriogenology 2001. 56: 307–314.
224. Walker A, Kimura K, Michael R. Expression of bovine interferon-tau variants
according to sex and age of conceptuses. Theriogenology, 2009. Volume 72, Issue
1,1, Pages 44-53.
225. Webb R, Garnsworthy PC, Gong J, Armstrong D. Control of follicular growth: local
interactions and nutritional influences. J. Anim. 2004. Sci. 82 (E-Suppl.), E63–E74.
Sci. 82 (E-Suppl.), E63–E74.

88 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
226. Wiltbank, J., Zimmerman, D., Ingalls, J., Rowden, W. Use of progestional
compounds alone or in combination with estrogen for synchronization of estrus.
Journal of Animal Science. 1965. 24: 990-994.
227. Wiltbank MC. Cell Types and hormonal mechanims associated wiht Mid – Cycle
corpus Luteum Function. J. Animal Sci. 2001. 72. 1873–1883.
228. Wooding FBP, Roberts RM, Green JA. Light and electron microscope
immunocytochemical studies of the distribution of Pregnancy-Associated
Glycoproteins (PAGs) throughout pregnancy in the cow: possible functional
implications. Placenta. 2005. 26: 807-827.
229. Xu YP, Chedrese PJ and Thacker PA. Growth hormone as an amplifier of insulin-
like growth factor-1 action: Potenciated estradiol accumulation. En: Journal of
Endocrinology. Vol.152 1997. p 201-209.
230. Yang RY and Liu FT. Galectins in cell growth and apoptosis. Cellular and
Molecular Life Sciences 2003. 60: 267–276.
231. Zieba DA, Amstalden M, Morton S, Gallino JL, Edwards JF, Harms PG, et al.
Effects of leptin on basal and GHRH-stimulated GH secretion from the bovine
adenohypophysis are dependent upon nutritional status. J Endocrinol 2003. 178(1):
83–9.

2. Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Resumen Se evalúa el efecto de la aplicación al día 5 u 8, de 400UI de Gonadotropina Coriónica
Equina (eCG), en un protocolo de sincronización para transferencia de embriones, sobre
desarrollo folicular (día 9), desarrollo luteal (día 17), concentración de progesterona “P4”
(días 9 y 17) y porcentaje de preñez (día 52), en 70 novillas raza Holsteín. Se analizan
independientemente a los tratamientos de eCG, las relaciones entre diámetro del folículo
dominante (DFD) “día 9” y el volumen luteal (VL) “día 17”, el VL (día 17) y concentración
de P4 (día 17). Se relacionan los tratamientos de eCG, el DFD (día 9), el VL (día 17) y
concentración de P4 (día 17) frente al procentaje de preñez. No hubo efecto (P>0.05) del
día de aplicación de eCG sobre número de FD, pero si (P<0,05) en cuanto a DFD. No
hubo efecto (P>0.05) del día de aplicación de eCG para número de cuerpos lúteos, VL,
concentración de P4, (días 9 y 17) y preñez. No hubo relación (P>0.05) entre el DFD y
VL; pero si existió (P<0,05) entre el VL y niveles de P4. Al analizar la relación entre los
tratamientos, el DFD, el VL y la concentración de P4 (día 17) frente a preñez, se observó
que solamente existió relación positiva (P<0,1) entre esta última (preñez) y la
concentración de P4. El día de aplicación de eCG (día 5 u 8) afecta solamente el DFD. La
preñez es afectada por la concentración de P4 la cual tiene relación con el VL.
Palabras clave: eCG, Cuerpo lúteo, Foliculo Dominante, Progesterona, Transferencia de
embriones.

90 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
Introducción Innumerables factores intervienen en los resultados de los programas de transplante
embrionario (TE) en bovinos, siendo relevantes los que tienen relación con las hembras
receptoras (Revisado por Tovío et al., 2008); esto se evidencia en las bajos porcentajes
de eficiencia, ya que normalmente en los núcleos de receptoras tratadas con protocolos
tradicionales de sincronización hormonal de ciclo estral, solamente del 40 al 50 % de las
hembras clasifican para él TE, el otro 50 o 60% son descartadas por su baja o inexistente
respuesta a los tratamientos de sincronización (Nasser et al., 2004). Considerando un
porcentaje de concepción del 40% del total de animales aprovechados, se obtendrá
apenas 20 % de gestaciones al final de todo el proceso (Thibier, 2002; Bó, 2003).
Indudablemente existen numerosos factores endocrinológicos, celulares y moleculares
entre otros, que determinan el mantenimiento o la pérdida de la preñez en etapas
iniciales, pero aún no han sido completamente aclarados (Spencer et al., 2004).
Teniendo en cuenta lo anterior, es fundamental mejorar los elementos de
fundamentación científica que puedan aclarar los diferentes procesos que encaminan el
desarrollo, reconocimiento y mantenimiento de la preñez en los programas de TE (Bó,
2003). De acuerdo a esto último, se evidencia que las secreciones hormonales
específicas interactúan con determinados factores para desencadenar el desarrollo del
embrión; especialmente se destaca la progesterona (P4), hormona secretada
principalmente por el cuerpo lúteo (CL), el cual tiene una duración activa y prolongada,
rasgo característico de la preñez en los mamíferos (Revisado por Tovío et al., 2008).
La P4 actúa en el útero estimulando y manteniendo funciones necesarias para el
desarrollo embrionario temprano; esto con la finalidad de llevar a cabo la implantación,
placentación y el exitoso desarrollo fetal (Spencer et al., 2004). De acuerdo a esto, se
afirma que el control endocrino para la síntesis de proteínas uterinas durante el desarrollo
embrionario temprano, lo tiene principalmente la P4, la cual es la responsable de los
cambios cualitativos y cuantitativos en el medio ambiente uterino, controlando la síntesis
y secreción de un sin número de sustancias (entre 1000 a 5000) como son factores de
crecimiento, citoquinas, proteínas etc. (Hafez, 2000); según esto, se asume que
deficiencias de P4, podrían causar que el endometrio llegue a ser poco eficiente en cuanto

Capítulo 2 91
a la nutrición histotrópica, la cual es la primer fuente disponible para el crecimiento,
mantenimiento y supervivencia del conceptus (Revisado por Tovío et al., 2008).
Las deficiencias de P4 causan alteraciones de origen endocrino, que contribuyen a
aumentar los niveles basales de mortalidad embrionaria (ME), probablemente asociados
a una selección natural ineludible, la cual busca la eliminación de genotipos poco
competentes (Rodríguez, 2001; Gouveia et al., 2004).
Alrededor de 25 al 40% de pérdidas embrionarias se han podido detectar durante los
primeros días de gestación en hembras receptoras de embriones bovinos (Santos et al.,
2004); se observa que la mayoría de estas hembras retornan a celo en la fecha prevista,
a los 20 – 22 días, manifestando un ciclo sexual normal y completo (vacas repetidoras)
(Palma, 2008), por lo que se sugiere que la ME pudo originarse entre los días 7 y 17, es
decir, el periodo comprendido entre el TE y el reconocimiento materno de la preñez
(RMP) (Binelli et al., 2001).
En cuanto a las pérdidas de preñez que suceden entre el día 28 y 98 (cuando ya ha
ocurrido el RMP), se han calculado porcentajes que oscilan entre el 7 y 33 % (Silke et
al., 2002).
De acuerdo a lo anterior, se sugiere que durante el establecimiento de la preñez se
presenta un "periodo crítico" muy definido entre los días 15 a 17 (Binelli et al., 2001); se
podría pensar que la biología durante este periodo es multifactorial y compleja, en donde
el endometrio puede estar recibiendo una señal antiluteolítica poco adecuada, que no
causa el bloqueo de la producción de Prostaglandina F2α (PGF2α) endometrial, lo que
desencadena la lisis del CL (el mantenimiento de la preñez es dependiente de la
funcionalidad del CL) (Santos et al., 2004). La señal antiluteolítica es generada por el
embrión en las células mononucleadas del trofoblasto, las cuales secretan interferón tau
(IFN-τ), bloqueando de esta manera la síntesis de PGF2α producida a nivel endometrial
(Binelli et al., 2001; Revisado por Grajales y Tovío, 2009). Esto último sugiere que las
pérdidas del embrión podrían estar ocurriendo por una señal débil o inadecuada por parte
de éste para la inhibición de la síntesis de prostaglandina uterina (Palma, 2008).

92 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
La naturaleza critica alrededor del periodo de reconocimiento, aposición y adhesión del
embrión al endometrio uterino durante la implantación, determina la necesidad de un
estricto sincronismo entre el embrión transferido y su receptora, enfatizando la
importancia tanto del medio uterino como de las señales del conceptus que dan lugar al
RMP (Binelli et al., 2001), señales que deben ser emitidas en el momento y
concentración precisa, de tal manera que se garantice el mantenimiento de la estructura
y funcionalidad del CL, que genere una continua producción de P4 para el mantenimiento
del ambiente embriotrófico que apoye el normal desarrollo del conceptus (Revisado por
Grajales et al., 2011).
La comunicación entre el conceptus y el útero, no siempre tiene éxito, lo que desencadena
grandes pérdidas embrionarias (Binelli et al., 2001). Por eso mismo se plantean estrategias
que buscan minimizar el efecto luteolítico el cual desencadena la pérdida temprana del
embrión, acontecimiento más fácilmente evidenciado en los programas de transplante
embrionario, ya que con éstos se asegura la presencia del embrión vivo desde el día 6 a 8
momento en el cual éste es trasplantado (Baruselli et al., 2005).
Conocida la influencia de la P4 en determinados eventos relacionados con el
mantenimiento de la preñez desde estadios tempranos y la influencia de la PGF2α para
causar luteólisis, se ha propuesto y desarrollado una serie de estrategias hormonales
con la finalidad de mantener la preñez. Estas se basan en hacer más eficiente la
capacidad secretora de P4 por parte del CL, y que esta secreción ocurra en el momento
oportuno, garantizando de esta manera un ambiente uterino adecuado para el desarrollo
del embrión transplantado a hembras receptoras bovinas; todo esto encaminado a
incrementar la tasa de preñez en los programas de TE (Thatcher et al., 2001).
La relación entre el porcentaje de preñez y la concentración plasmática de P4 de acuerdo
al tamaño del CL en receptoras de embriones bovinos Bos Indicus y Bos Taurus, ha sido
objeto de controversia en varios estudios realizados. Algunos investigadores han
verificado la correlación positiva entre estas variables, encontrando que a mayor área del
CL mayor es la concentración de P4 plasmática y consecuentemente mayor es la tasa de
preñez en diferentes razas (Thatcher et al., 2001; Vasconcelos et al., 2001; Bó et al.,
Mantovani et al., 2002; Baruselli et al., 2005; Lopes et al., 2007). Con estos resultados se
podría evidenciar que los incrementos en la concentración de P4 durante el "periodo

Capítulo 2 93
crítico" (Binelli et al., 2001), estimulan la secreción de los agentes antiluteolíticos con lo cual
se hace eficiente el RMP (Vasconcelos et al., 2001). Igualmente en diferentes reportes se
observa que el aumento en las concentraciones plasmáticas de P4 durante el diestro se
correlaciona positivamente con la capacidad del embrión en secretar IFN-τ, provocando así
un aumento en las tasas de preñez (Thatcher et al., 2001; Mann et al., 2009).
Según los estudios se podría sugerir que a mayor concentración de P4 plasmática mejor
será el ambiente uterino para el conceptus en vías de desarrollo (Figueira et al., 2011).
Esto se entiende si se tiene en cuenta que cualquier variación en la concentración de P4
es determinante en la modulación de la expresión y secreción de factores de
crecimiento, citoquinas y proteínas, que condicionan el medio uterino para los procesos
de receptividad endometrial y de viabilidad embrionaria (Revisado por Peña et al., 2007).
De acuerdo a lo anterior, se podría sugerir que al brindar fuentes directas o indirectas de
P4 a hembras durante los primeros días de preñez, el porcentaje de perdidas
embrionarias disminuye, ya que se mejora el ambiente uterino en donde el conceptus
tendrá un desarrollo adecuado evidenciando mejor la síntesis y secreción de IFN-τ, pues
esta secreción está influenciada por el estado de desarrollo embrionario (Kebler et al.,
1997; Siqueira et al., 2009; Revisado por Tovío et al., 2010).
Con la finalidad de aumentar las tasas de preñez se han utilizado tratamientos
hormonales en hembras receptoras de embriones bovinos, utilizado Gonadotropina
Corionica Equina (eCG) en protocolos de transplante de embriones a tiempo fijo (TETF)
(Baruselli et al., 2005).
La eCG (antes conocida como Gonadotropina Sérica de Yegua Preñada PMSG) es una
hormona glicoproteica con peso molecular de 45 kDa. y promedio de vida de 3 días,
producida por los cálices endometriales en la yegua preñada entre los días 40 a 130
(Bousfield y Butnev, 2001). Se compone de dos subunidades α-y β (la subunidad α está
codificada por un gen común en todas las glicoproteínas, mientras que estas hormonas
difieren en el gen que codifica para subunidad β-, la cual confiere la especificidad
hormonal) (Palma, 2008; Forcada et al., 2011).

94 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
Esta hormona se vincula a los receptores foliculares de FSH y de LH, y a los receptores
de LH del CL, creando de esta forma condiciones de crecimiento folicular, ovulación y
luteinización (Murphy and Martinuk, 1991 Baruselli et al., 2003; Filho et al., 2004; Filho et
al., 2010); pero su acción predominante es de FSH, lo que daría a lugar a la formación de
cuerpos lúteos accesorios característicos de la yegua gestante (Fuentes y de la Fuente,
2007; Forcada et al., 2011).
La aplicación de eCG en el momento esperado de una nueva onda de crecimiento
folicular, ha demostrado eficiencia en cuanto a superovulación (de acuerdo a la
dosificación) y/o desarrollo de un folículo dominante de mayor diámetro, determinando de
esta forma un mayor número de cuerpos lúteos o un CL grande (Baruselli et al., 2005).
Esto va acompañado de mayores concentraciones plasmáticas de P4 y mejor porcentaje
de utilización (hembras transferidas/hembras sincronizadas*100), concepción y de
preñez frente a tratamientos sin aplicación de esta hormona tanto en ganado Bos Indicus
como Bos Taurus y sus cruces (Santos et al., 2001; Marques et al., 2003; Everton and
Baruselli, 2003; Madureira et al., 2004; Rodrigues et al., 2004; Baruselli et al., 2005;
Mamani et al., 2007; Pita et al., 2009; Sousa et al., 2009; Figueira et al., 2011).
Se ha verificado que con la aplicación de eCG el día 8 dentro de un protocolo de
sincronización en hembras receptoras (Bos Indicus/Bos Taurus) de embriones bovinos se
consiguen cuerpos lúteos únicos de mayor tamaño frente a la aplicación de la misma
hormona el día 5, pero esto no ha determinado diferencias entre la concentración
plasmática de progesterona producida, ni en los porcentajes de preñez obtenidos entre
tratamientos (Nasser et al., 2004). Igualmente en otros reportes al respecto se ha
verificado que el porcentaje de utilización (hembras transferidas/hembras
sincronizadas*100), ha mejorado cuando la aplicación de eCG se suministra hacia el día
8 dentro de un protocolo para transplante de embrión a tiempo fijo a receptoras Bos
Indicus (Quezada y Ortiz 2007).
2.1. Hipótesis El día de aplicación de la hormona Gonadotropina Coriónica Equina (eCG) determina el
desarrollo folicular y luteal, su capacidad secretora de progesterona y la tasa de preñez
en novillas de la raza Holstein receptoras de embriones bovinos.

Capítulo 2 95
2.2. Objetivo Evaluar el efecto de la aplicación de un tratamiento con Gonadotropina Coriónica Equina
(eCG) el día 5 o el día 8, dentro de un protocolo de sincronización para transplante de
embriones a tiempo fijo (TETF), sobre el desarrollo folicular y luteal, la concentración de
progesterona plasmática y el porcentaje de preñez, en novillas de la raza Holstein
receptoras de embriones bovinos en el trópico alto colombiano.
2.3. Objetivos especificos a) Determinar si existe efecto del día de la aplicación (día 5 u 8) de Gonadotropina
Coriónica Equina (eCG) sobre el desarrollo del folículo dominante (número y tamaño)
el día nueve (9) del protocolo de sincronización.
b) Determinar si existe efecto del día de la aplicación (día 5 u 8) de la Gonadotropina
Coriónica Equina (eCG) en el desarrollo del Cuerpo Lúteo (número y tamaño) para el
día diecisiete (17) del protocolo de sincronización.
c) Determinar si existe efecto del día de la aplicación (día 5 u 8) de Gonadotropina
Coriónica Equina (eCG) sobre la concentración de progesterona (P4) plasmática para
los días nueve (9) y diecisiete (17) del protocolo de control de ciclo para trasplante
embrionario en hembras receptoras bovinas.
d) Determinar si existe efecto del día de la aplicación (día 5 u 8) de Gonadotropina
Coriónica Equina (eCG) sobre el porcentaje de preñez en hembras receptoras
bovinas al día 52.
e) Analizar independientemente a los tratamientos (eCG día 5 ó eCG día 8) las
relaciones entre:
- Diámetro (mm) del folículo dominante (día 9) y el volumen luteal “mm3” (día 17)
- Volumen luteal “mm3” (día 17) y la concentración plasmática de P4 “ng/ml” (día 17).
f) Analizar la relación de los tratamientos utilizados (eCG día 5 ó eCG día 8), el diámetro
del folículo dominante (día 9), el volumen luteal “mm3” (día 17) y la concentración
plasmática de P4 “ng/ml” (día 17) frente al porcentaje de preñez (día 52).
2.4. Materiales y métodos
El trabajo se realizó bajo el aval del Comité de Bioética de la Facultad de Medicina
Veterinaria y de Zootecnia – UNAL Bogotá; igualmente, bajo los parámetros de manejo

96 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
animal de la empresa ganadera particular, en la cual se desarrollo el experimento
(hacienda Gaviotas, adscrita a la Central de Reproducción Animal C.G.R –
Cundinamarca – Colombia).
2.4.1. Localización El trabajo de campo se desarrollo en una central de transferencia de embriones bovinos
ubicada en el municipio de Zipaquirá departamento Cundinamarca, localizado a los 4º 56’
50’’ latitud norte y 74º 2’ 30’’ de longitud oeste; altura sobre el nivel del mar 2652 metros;
temperatura media de 14 ºC; humedad relativa 60% y precipitación 824.2 mm/año.
El trabajo de laboratorio se realizó en el Laboratorio de Hormonas de la Facultad de
Medicina Veterinaria y de Zootecnia de la Universidad Nacional de Colombia – Sede
Bogotá D.C., en el cual se midieron y calcularon las concentraciones plasmáticas de
progesterona (P4.), mediante la técnica de Radioinmunoanálisis (RIA) utilizando un
contador de radiación gamma, marca Beckman 500®, freezer para bajas temperaturas,
centrífugas, micropipetas, baños serológicos, estufas y cabina de flujo laminar.
Igualmente se utilizó el material y equipos necesarios, reglamentarios para llevar a cabo
los procedimientos adecuadamente.
2.4.2. Grupo experimental Se utilizaron 70 novillas de la raza Holstein Friesian, con edades entre 15 a 26 meses,
pesos entre 300 y 420 kilogramos y condición corporal entre 3 y 4, en escala calificatoria
de 1 a 5 (Edmonson et al., 1989), las cuales fueron sometidas a examen por palpación
rectal del útero y estructuras ováricas (folículos y cuerpos lúteos), con la finalidad de
confirmar ciclicidad reproductiva y normalidad uterina. Igualmente se sometieron a
exámen clínico general para garantizar un buen estado nutricional y sanitario.
La metodología utilizada para la distribución de las novillas en los grupos de tratamientos
correspondió a la generación de números aleatorios, los cuales provienen de una
distribución uniforme continua (0,1).
2.4.2.1. Descripción del grupo de novillas evaluado En la tabla 1 se describen las variables edad, peso y condición corporal (promedios,
desviaciones y coeficientes de variación), en donde se muestra el estado para el total del
grupo de novillas seleccionadas, así mismo como para el grupo de hembras

Capítulo 2 97
seleccionadas para la aplicación del tratamiento 1 (eCG día 5) y tratamiento 2 (eCG día
8).
Tabla 1. Edad, peso y condición corporal de la muestra total y por tratamientos.
eCG día 5 (n=42) 17 26 12 19.7 ± 0.42 300 420 6 385 ± 3.63 3 4 9 3.5 ± 0.04
eCG día 8 (n=28) 15 23 14 18.5 ± 0.30 340 420 6 381 ± 4.03 3 4 8 3.7 ± 0.07
Total muestra (n=70) 15 26 9 19.2 ± 0.29 300 420 6 383 ± 2.69 3 4 9 3.6 ± 0.04
CV
Condición corporal (1-5)
Min. Max. Promedio¹
Edad (meses) Peso (kg.)
Promedio¹ Promedio¹ Min.Max. CVCV Min. Max.Tratamiento
¹Todos los resultados se expresan como promedio (media) ± DS.
De acuerdo a los datos descritos se denota la homogeneidad de la muestra utilizada en
cada uno de los dos grupos tratados (eCG día 5 y eCG día 8), en cuanto a las tres
variables descritas (edad, peso y condición corporal), variables de las cuales se observa
que sus valores son semejantes a los de animales utilizados en estudios similares
(Fuentes y De la Fuente, 2007; Sartori et al., 2004; Nasser et al., 2004; Sartori et al.,
2002; Vasconcelos et al., 2001). Igualmente, estos valores se consideran normales, de
acuerdo al nivel de desarrollo y características de la raza utilizada (López, 2006).
2.4.3. Condiciones de manejo Los animales fueron manejados bajo pastoreo en praderas sembradas con una mezcla
de Lolium Peremne (Rye Grass) y Pennisetum clandestinum (Kikuyo). Igualmente se
suministró sal mineralizada y agua a voluntad.
2.4.4. Tratamientos aplicados para control de ciclo estral El protocolo de sincronización tuvo una duración de diez (10) días, por lo tanto, se
expresa el día de inicio del tratamiento, como día “0” o como día “-10”. El día asumido de
la presentación de celo igualmente se expresa como día “10” o como día del celo “01” (Bó

98 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
et al., 2002), con la finalidad de una mejor comprensión, de acuerdo a varios estudios
discutidos.
Las hembras fueron repartidas en dos grupos a los cuales se aplicó un protocolo de
sincronización, que se diferenció entre grupos por el día de aplicación (día 5 o día 8) de
400UI de Gonadotropina Coriónica equina (eCG). El día 5 de iniciado el protocolo, al azar
42 hembras (n=42) recibieron la dosis correspondiente de eCG. Las otras 28 hembras
(n=28) recibieron la dosis el día 8 de iniciado el protocolo.
Los protocolos de sincronización utilizados fueron bajo el modelo de Transferencia de
Embrión a Tiempo Fijo (TETF), en donde se observaron los celos, con la finalidad de
evidenciar hasta cierto punto la eficiencia de la respuesta al protocolo de sincronización
empleado (Bó y Caccia, 2003). Los embriones fueron transferidos a todas las novillas en
una sola jornada en el mes de junio de 2009.
Los embriones transferidos correspondieron a la raza Holsteín, los cuales se colectaron,
manipularon, clasificaron, congelaron en etilenglicol 1.5 M (transferencia directa) y
posteriormente se transfirieron por el mismo técnico a todas las novillas en codígo de
estadío cuatro (morula) y código de calidad uno (excelente o bueno). Este procedimiento
se realizó de acuerdo a lo reglamentado por la Sociedad Internacional de Transferencia
de Embriones (IETS, 2008).
2.4.4.1. Tratamiento 1 (Grupo 1) Aplicación de la terapia de apoyo hormonal con eCG día 5 (Día 5 con respecto al inicio del protocolo. Día - 5 con respecto al celo) - Día 0 (-10): Aplicación de Dispositivo Intravaginal Bovino (DIB® - P4. 1 gr) + 2 mg de
benzoato de estradiol IM (Syntex 2 ® ml).
- Día 5 (-5): Aplicación de Gonadotropina Corionica Equina (eCG - 400 UI IM -
Novormon® 2 ml.) + Prostaglandina F2α (1.5 mg IM - Prolise ® 2 ml) +
Muestreo sanguíneo.
- Día 8 (-2): Retiro del Dispositivo intravaginal Bovino (DIB ®)
- Día 9 (-1): Aplicación de benzoato de estradiol (1 mg IM - Syntex 1 ml ®) + Muestreo
sanguíneo + Ecografía.

Capítulo 2 99
- Día 10 (01): Observación de Celos
- Día 17 (7): Transferencia de embrión congelado + Muestreo sanguíneo + Ecografía.
- Día 52 (42): Ecografía para diagnostico de preñez.
Esquema 1. Tratamiento de sincronización con aplicación de eCG día 5 (-5).
Esquema 1. Tratamiento de sincronización con aplicación de eCG día 5 (-5).
2.4.4.2. Tratamiento 2 (grupo 2) Aplicación de la terapia de apoyo hormonal con eCG día 8 (Día 8 con respecto al inicio del protocolo. Día - 8 con respecto al celo) - Día 0 (-10): Aplicación de Dispositivo Intravaginal Bovino (DIB® - P4. 1 gr) + 2 mg de
benzoato de estradiol IM (Syntex 2 ® ml).
- Día 5 (-5): Aplicación de Prostaglandina F2α (1.5 mg IM - Prolise ® 2 ml) + Muestreo
sanguíneo.
- Día 8 (-2): Retiro del Dispositivo Intravaginal Bovino (DIB ®) + Aplicación de
Gonadotropina Corionica Equina (eCG - 400 UI IM - Novormon® 2
ml.).
- Día 9 (-1): Aplicación de benzoato de estradiol (1 mg IM - Syntex 1 ml ®) + Muestreo
sanguíneo + Ecografía.
- Día 10 (01): Observación de Celos
- Día 17 (7): Transferencia de embrión congelado + Muestreo sanguíneo + Ecografía.
- Día 52 (42): Ecografía para diagnostico de preñez.
BE
eCG + PF2α
BE Transplante embrionario
0 4 5 8 9 10 13 17 21 25 30 52 -10 -6 -5 -2 -1 01 3 7 11 15 20 42
DX. Preñez
Celos
Dispositivo Intravaginal DIB®
Muestreo sanguíneo
* **
* Día referencial inicio del protocolo como día cero (o)
** Día referencial inicio del protocolo como día menos 10 (-10)
Ecografía de estructuras ováricas

100 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
Esquema 2. Tratamiento de sincronización con aplicación de eCG día 8 (-2).
- Planteamiento del protocolo de sincronización utilizado
Se tomó como día 0 (cero) el día de inicio de los protocolos, en donde se buscó generar
un bloqueo de las gonadotropinas hipofisiarias. Este bloqueo se logró gracias a la
combinación de la P4. con el benzoato de estradiol (BE). El implante con P4. Inhibe la
liberación de LH (sin afectar niveles basales), con lo cual se genera la regresión de
cualquier folículo dominante. El BE inhibe la secreción de FSH, la cual no ha sido inhibida
completamente por la acción sola de la P4. (Barret et al., 2006). Como la concentración a
nivel sanguíneo del BE dura aproximadamente 4.3 días, una nueva onda de crecimiento
folicular se generará en este momento ya que los niveles plasmáticos de estradiol
descienden, con lo cual se sincronizará el crecimiento de la onda folicular nueva. (Bó y
Caccia, 2003)
Al colocar la prostaglandina el día cinco (5) (ciclo sincronizado) se causa luteolisis de
cualquier cuerpo lúteo de la onda anterior, con lo cual se elimina una posible fuente de
P4. que llegue a causar atresia folicular (Baruselli et al., 2005).
Con la aplicación de la eCG el día cinco (5) (Trtamiento 1, con referencia al día de inicio
de la sincronización), se busca generar reclutamiento folicular, gracias a la acción FSH
de la eCG, por lo cual se podría generar mas de un folículo dominante, generándose así
mas de un cuerpo luteo que generen concentraciones de P4 por encima a 1 ng/ml., que
preparen al útero para la adecuada implantación y desarrollo embrionario (Baruselli et al.,
2005); esto basados en que la P4 es estimulada principalmente por la actividad
luteotrópica de la LH (Olivera et al., 2007).
* Día referencial inicio del protocolo como día cero (o)
** Día referencial inicio del protocolo como día menos 10 (-10)
BE
eCG
BE Transplante embrionario
0 4 5 8 9 10 13 17 21 25 30 52 -10 -6 -5 -2 -1 01 3 7 11 15 20 42
DX. Preñez
Celos
Dispositivo Intravaginal DIB®
Muestreo sanguíneo
PF2α
* **
Ecografía de estructuras ováricas

Capítulo 2 101
Por otro lado con la aplicación de la eCG (la cual tiene acción FSH y LH) el día ocho (8)
(Tratamiento 2, con referencia al día de inicio de la sincronización), se busca generar que
el folículo (s) dominante (s) mantenga (n) su desarrollo y crecimiento en la fase final
(Evans et al., 1997), esto con la finalidad de que llegue a ser un folículo preovulatorio. El
desarrollo de este folículo se relaciona con la síntesis de receptores para FSH y LH en el
folículo, así como la elaboración de las aromatasas (Bao et al., 1997). La expresión de
un mayor número de receptores para la LH en las células de la granulosa se considera
como un mecanismo para la adquisición de dominancia (Sartori et al., 2001). Se ha
encontrado mRNA de receptores de LH en el folículo dominante, y su crecimiento está
asociado con mayor expresión de mRNA en las células de la granulosa (Mihm et al.,
2000); esto se relaciona con la preparación celular para formar el CL (Knobil and Neil,
2006), argumentan que la IGFBP participa en la luteinización del folículo dominante
inducida por la LH, por lo cual también se esperaría la síntesis de P4.
Al retirar el dispositivo intravaginal el día ocho (8) se desencadena una disminución en la
concentración de P4 plasmática, con lo cual se elimina el “Feed back” negativo para la
secreción de la LH, que es estimulada por acción de la concentración de estrógenos
generados por los folículos en crecimiento de la onda que se inició el día 4.3
aproximadamente (Bó et al., 2002).
Con la aplicación el día nueve (9) de benzoato de estradiol (BE) se espera generar el
desencadenamiento hipofisiario de LH (pico de LH), esto con la finalidad de generar la
sincronía de la ovulación (Bó y Caccia, 2003).
La observación de celos el día nueve (9) en la tarde y el día diez (10) en la mañana y en
la tarde se realiza con la finalidad de estimar un comportamiento referencial a la
respuesta de los animales ante el protocolo de sincronización (respuesta de eficiencia al
tratamiento de sincronización).
El día diecisiete (17) de la sincronización se transplantó el embrión lo cual coincide con la
edad y estadio de la estructura a transferir (mórula – Blastocisto).

102 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
2.4.5. Toma de muestras y ecografía A los dos (2) grupos de hembras sincronizadas (eCG día 5 y eCG día 8) se les tomó tres
(3) muestras de sangre los siguientes días con respecto al ciclo sincronizado:
- Día cinco “5” (inicio de la nueva onda folicular)
- Día nueve “9” (día de máximo desarrollo folicular)
- Día diecisiete “17” (día de la transferencia embrionaria)
La muestra de sangre se colectó directamente por medio de punción de los vasos
sanguíneos coccígeos hacia un tubo sin anticoagulante con capacidad para 6 ml.
(Vacuette®, Greiner Bio-one, Monroe, NC 28110, USA), el cual fue rotulado con los
datos de la hembra. Estas muestras se centrifugaron a 800 X g durante 15 minutos con la
finalidad de separar el suero, el cual se almacenó en viales, que se mantuvieron en
congelación a -20 °C, en el laboratorio de Hormonas de la Facultad de Medicina
Veterinaria y de Zootecnia de la Universidad Nacional de Colombia sede Bogotá, hasta el
momento de la cuantificación de los niveles de P4 por la técnica de radioinmunoanálisis
en fase sólida (RIA), utilizando un equipo Beckman 500® para lectura de rayos Gamma.
Para esta cuantificación se utilizaron kits Coat-A-Count Progesterone de Diagnostics
Products Corporation (DPC®), con una sensibilidad de 0.02 ng/ml. (Duica, 2011; Gonella,
2011; Báez, 2010; Grajales, 2000).
Los muestreos ecográficos se realizaron con la finalidad de observar y medir las
estructuras ováricas (folículos y cuerpos lúteos – número y tamaño). Estos se realizaron
los siguientes días con respecto al ciclo sincronizado:
- Día nueve “9” (día de máximo desarrollo folicular)
- Día diecisiete “17” (día de la transferencia embrionaria)
Para estas ecografías se utilizó un cecógrafo Aquila Pro con transductor lineal de 6.0 a
8.0 Mhz, Pie Medical ®, con el cual se trabajó graduándolo en 8.0 Mhz siguiendo las
recomendaciones de Duica, 2011. Las imágenes tanto de folículo (s) dominante (s) (día
9) como de cuerpo (s) luteo (s) (día 17), se congelaron inmediatamente de acuerdo a su
adecuada ubicación, esto con la finalidad de referenciar el dato de medida.

Capítulo 2 103
El día 52 (con respecto al día de inicio del protocolo) se procedió a realizar chequeo
ultrasonografico con la finalidad de determinar las hembras preñadas (día 42 de
gestación).
2.4.6. Medición de variables. Para la realización de este estudio se establecieron las siguientes variables respuesta:
a. Número de folículos dominantes presentes (Foliculos ≥ 9 mm - Onda folicular
sincronizada) (día 9 – muestreo 2).
Mediante ecografía se procedió a contar el número de folículo (s) dominante (s)
(Folículos ≥ 9 mm) (Sartori et al., 2004), presentes el día 9 en las horas de la mañana
(con respecto al día de inicio del tratamiento de sincronización – máximo desarrollo
folicular).
Para la determinación de la estructra folicular se tuvo en cuenta que los folículos
ováricos, como son estructuras que estan llenas de líquido, aparecen en la pantalla
del ecógrafo como áreas de color negro, también denominadas no ecogénicas.
Igualmente, antes de ecografiar se determinó la textura folicular a la palpación (Bó y
Caccia, 2003).
b. Diámetro (mm) de folículo dominante (Foliculos ≥ 9 mm - Onda folicular sincronizada)
presente (día 9 – muestreo 2).
La ecografía de (los) folículo (s) dominante (s) se realizó el día (nueve) 9 en horas de
la mañana, es decir, el día de máximo desarrollo folicular de acuerdo al tratamiento de
sincronización aplicado. Las imágenes del folículo (s) dominante (s) se congelaron en
momento más oportuno para realizar la medición inmediata del diámetro de la
estructura folicular siguiendo las recomendaciones de Bó, y Caccia (2000), en cuanto
a la no inclusión del espesor de la pared. Se midió el diámetro del (los) folículo (s) de
mayor tamaño (≥ 9 mm., Sartori et al., 2004), teniendo en cuenta que el día 9
(crecimiento de onda sincronizada) se encontró un folículo dominante (día de máximo
desarrollo folicular de acuerdo al tratamiento de sincronización aplicado).
c. Número de cuerpos lúteos presentes el día 17 (muestreo 3).
Mediante ecográfia se visualizaron y congelaron las imágenes para proceder a contar
el número de cuerpo (s) luteo (s), esto se realizó el día (diecisiete) 17 en las horas de

104 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
la mañana (de acuerdo al tratamiento de sincronización aplicado) con respecto al día
de inicio del tratamiento de sincronización (cuerpo luteo consolidado de 7 días) (Bó y
Caccia, 2003).
Antes de ecografiar se determinó la textura luteal a la palpación. Los cuerpos lúteos
poseen una ecotextura diferente a la del estroma ovárico e identificable por
ultrasonografía a partir del día de la ovulación (promedio: día 0,5) y a partir del día 3
con bordes bien definidos. Normalmente se distingue a lo largo de casi todo el ciclo
estral e inclusive hasta cerca de la siguiente ovulación (en promedio: 1,4 días antes de
que la misma) (Bó y Caccia, 2003).
*De acuerdo al número de estructuras luteales encontradas en día 17 y su relación
con el número de folículos dominantes presentes el día 9, también se determinó la
presencia de ovulaciones dobles (día 17).
d. Volumen (mm3) luteal presente el día 17 (muestreo 3)
Mediante visualización ecográfica se congelaron las imágenes y se procedió a medir
el largo por el ancho del (los) cuerpo (s) lúteo (s) presentes para el momento del
transplante embrionario el día 17 en las horas de la mañana (con respecto al día de
inicio del tratamiento de sincronización).
El volumen luteal se calculó con la fórmula planteada por Vasconcelos et al. (2001) V
= 4/3 x π x R3. En donde: R = (Largo del cuerpo lúteo / 2 + Ancho del cuerpo lúteo
/ 2) / 2.
*De acuerdo a las estructuras luteales (cantidad y volumen) evidenciadas el día 17
(transplante embrionario) se determinaron las hembras a las cuales se debía
trasplantar un embrión, de allí se calculó el porcentaje de utilización: (hembras
transferidas/hembras sincronizadas)*100.
e. Concentración plasmática de progesterona (ng/ml.) (día 5-muestreo 1, día 9-muestreo
2 y día 17-muestreo 3)
f.
g. Porcentaje de preñez (hembras preñadas/hembras transferidas*100. (día 52).
* Igualmente para la realización de los análisis se determinaron las siguientes relaciones:

Capítulo 2 105
- Relación entre el tamaño (diámetro mm) del folículo dominante (día 9) y el tamaño
(volumen mm3) del cuerpo lúteo (día 17) (independientemente a los tratamientos eCG
día 5 u 8).
- Relación entre el tamaño (volumen mm3) del cuerpo lúteo (día 17) y la concentración
(ng/ml) de P4 plasmática reportado para el mismo día (día 17 transplante embrionario).
- Relaciones de los tratamientos utilizados (eCG día 5 u 8), el diámetro (mm) del
folículo dominante (día 9), el volumen (mm3) luteal (día 17) y la concentración (ng/ml)
plasmática de P4 (día 17) sobre el porcentaje de preñez (día 52).
2.4.7. Análisis estadístico. - Distribución de los animales en los grupos de tratamientos. La metodología
utilizada para la distribución de las novillas en los grupos de tratamientos (eCG día 5 y
eCG día 8), correspondió a la generación de números aleatorios, los cuales provienen de
una distribución uniforme continua (0,1). (Resultado de una variable al azar especificada
por una distribución) (Rienzo et al., 2001).
- Homogeneidad de varianzas. Para todas las variables que se analizaron mediante el
modelo de covarianza, la homegeneidad de varianzas de los tratamientos se probó
mediante una prueba F (Steel and Torrie, 1980).
- Efecto de los tratamientos (eCG día 5 o eCG día 8) sobre el número de folículos dominantes (día 9). Se empleó un modelo completamente aleatorizado, con el peso,
condición corporal y edad inicial como covariables. Debido a que esta variable es un
conteo, se empleó la transformación logarítmica propuesta por Ali y Shook (1980):
Variable transformada = Ln (variable orgiginal +10)
El modelo utilizado fue el siguiente:
En donde:
: Es el número de folículos dominantes de la iésima vaca tratada con el j-ésimo
tratamiento.
: Es la media general.

106 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
: Con i=1, 2, 3 son los estimadores de mínimos cuadrados ordinarios de los efectos de
las covariables, que son respectivamente: condición corporal inicial (CC), peso inicial (P),
y edad inicial (E).
: Es el efecto del j-ésimo tratamiento, j=1,2.
: Es el efecto residual aleatório.
- Efecto de los tratamientos (eCG día 5 y eCG día 8) sobre el díametro del folículo
dominante (día 9). Se empleó un modelo completamente aleatorizado, con el peso,
condición corporal y edad inicial como covariables. El modelo utilizado fue el siguiente:
En donde:
: Es el diámetro del folículo dominante de la iésima vaca tratada con el j-ésimo
tratamiento.
: Es la media general.
: Con i=1, 2, 3 son los estimadores de mínimos cuadrados ordinarios de los efectos de
las covariables, que son respectivamente: condición corporal inicial (CC), peso inicial (P),
y edad inicial (E).
: Es el efecto del j-ésimo tratamiento, j=1,2.
: Es el efecto residual aleatório.
- Efecto de los tratamientos (eCG día 5 o eCG día 8) sobre el número de cuerpos lúteos (día 17). Se empleó un modelo completamente aleatorizado, con el peso,
condición corporal y edad inicial como covariables. Debido a que esta variable es un
conteo, se empleó la transformación logarítmica propuesta por Ali y Shook (1985):
.
El modelo utilizado fue el siguiente:
En donde:
: Es el número de cuerpos lúteos de la iésima vaca tratada con el j-ésimo
tratamiento.
: Es la media general.

Capítulo 2 107
: Con i=1, 2, 3 son los estimadores de mínimos cuadrados ordinarios de los efectos de
las covariables, que son respectivamente: condición corporal inicial (CC), peso inicial (P),
y edad inicial (E).
: Es el efecto del j-ésimo tratamiento, j=1,2.
: Es el efecto residual aleatório.
* De acuerdo al número de estructuras luteales encontradas en día 17 y su relación con
el número de folículos dominantes presentes el día 9, se determinó el número de ovulaciones dobles, las cuales también fueron conparadas entre tratamientos. Para
esto se realizó la prueba chi cuadrado para detectar diferencias entre los tratamientos,
con un nivel de significancia de 0.05.
- Efecto de los tratamientos (eCG día 5 y eCG día 8) sobre el volumen del cuerpo lúteo (día 17). Se empleó un modelo completamente aleatorizado, con el peso, condición
corporal y edad inicial como covariables. El modelo utilizado fue el siguiente:
En donde:
: Es el volumen luteal de la iésima vaca tratada con el j-ésimo tratamiento.
: Es la media general.
: Con i=1, 2, 3 son los estimadores de mínimos cuadrados ordinarios de los efectos de
las covariables, que son respectivamente: condición corporal inicial (CC), peso inicial (P),
y edad inicial (E).
: Es el efecto del j-ésimo tratamiento, j=1,2.
: Es el efecto residual aleatório.
* Igualmente el porcentaje de utilización: (hembras transferidas/hembras
sincronizadas)*100, fue analizado y comparado entre tratamientos mediante la prueba chi

108 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
cuadrado mediante el procedimiento FREQ de SAS, no se encontraron diferencias
significativas a un nivel de significncia de 0.05.
- Efecto de los tratamientos (eCG día 5 y eCG día 8) sobre la concentración de progesterona (días 9 y 17). Se utilizó un modelo mixto a una vía de clasificación con
arreglo de medidas repetidas en el tiempo con el volumen del cuerpo lúteo, el diámetro
folicular medio, diámetro del folículo dominante, la edad, peso inicial y condición corporal
inicial como covariables. Además de estas, como efectos fijos se tuvieron el tratamiento
(dos niveles, tratamiento 1 - eCG día 5 y Tratamiento 2 - eCG día 8), el tiempo (2 niveles,
Muestreo 2 - día 9 y Muestreo 3 – día 17) y su interacción; y como efecto aleatorio se
tuvieron en cuenta el animal y el residual.
El siguiente fue el modelo empleado:
En donde:
: Es la respuesta tomada el i-ésimo día a la k-ésima vaca, que fue tratada con el j-
ésimo tratamiento.
: Es el efecto del j-ésimo tratamiento, k=1,2.
: Es el efecto de la k-ésima vaca anidada en el j-ésimo tratamiento (error
experimental).
: Es el efecto del i-ésimo tiempo de medida.
: Es el efecto de la interacción del j-ésimo nivel del tratamiento con el i-ésimo
nivel del tiempo.
: Con i=1,2,…,6 son las estimaciones de mínimos cuadrados generalizados de los
efectos de las covariables, en su orden: Volumen del cuerpo lúteo (VCL), diámetro
folicular medio (TPFT), diámetro del folículo dominante (DPFD), edad inicial (E), peso
inicial (P) y condición corporal inicial (CC).
: Es el efecto residual.

Capítulo 2 109
Se compararon tres estructuras de la matriz de varianza-covarianza del residual (R);
estas fueron: simetría compuesta, no estructurada y primer orden autoregresivo. Por lo
tanto, se compararon tres modelos, que difirieron solamente en la estructura de R. Estas
comparaciones se realizaron mediante el estadístico BIC (Bayesian Index Criterion). El
modelo con la estructura de simetría compuesta mostró un mejor ajuste (menor BIC).
- Efecto del tratamiento (eCG día 5 y eCG día 8) sobre el porcentaje de preñez (hembras preñadas/hembras transferidas*100. Para esto se realizó la prueba chi
cuadrado mediante el procedimiento FREQ de SAS, esto con la finalidad de detectar
diferencias entre los tratamientos, con un nivel de significancia de 0.05
* Con la finalidad de observar la posible relación de diámetro (mm) del folículo dominante (día 9) sobre el volumen (mm3) del cuerpo luteal (día 17), se realizó un
modelo de regresión. Este análisis se realizó mediante el procedimiento REG de SAS, en
el cual se modelaron efectos lineales y cuadráticos del DF sobre VCL. De acuerdo a los
resultados del análisis preliminar de residuales fue necesario transformar la variable
dependiente; se empleó la transformación raíz cuadrada. De acuerdo a lo anterior el
modelo que se empleó fue el siguiente:
En donde es la raíz cuadrada del VCL, , y son los estimadores de
mínimos cuadrados del intercepto, efecto lineal y cuadrático de DF respectivamente y
es el error aleatorio. , ,
corresponde a la varianza del error.
* Con la finalidad de observar la posible relación de volumen (mm3) luteal (día 17) sobre la concentración de P4 (día 17), se realizó un modelo de regresión. Este análisis
se realizó mediante el procedimiento REG de SAS, en el cual se modelaron efectos
lineales y cuadráticos del VCL sobre P4. El siguiente fue el modelo empleado:
En donde , y , , y , son los estimadores de los mínimos cuadrados ordinarios
del intercepto y de los efectos lineales y cuadráticos de P4 y VCL respectivamente.

110 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
, , es la varianza
del error.
** Con el objeto de relacionar en conjunto, el efecto del tratamiento (eCG día 5 y eCG día 8), diámetro folicular (folículo dominante - día 9), volumen total luteal (día 17) y concentración de progesterona (día 17) sobre el diagnóstico de preñez, se planteó y
realizó una regresión logística por ser la preñez una variable de naturaleza dicotómica
(1= positivo, 0=negativo). Este modelo de regresión no lineal estima la probabilidad de
ocurrencia de un evento; en este caso se modeló la probabilidad de que el diagnóstico de
preñez fuera positivo: ; por lo tanto el modelo empleado fue el siguiente:
El análisis se efectuó mediante el procedimiento LOGISTIC de SAS; para la selección de
las variables que probablemente afectan el diagnostico de preñez y que por lo tanto se
incluyeron en el modelo bajo la metodología FORWARD. El nivel de significancia para
que cada una de estas variables fuera incluida en el modelo correspondió a 0.05.
Utilizando esta metodología se muestra la estimación del Odds y su respectivo intervalo
de confianza; se dice que si el intervalo contiene al uno (1), no existe mayor probabilidad
de que se presente el evento 1 (preñez positiva), si este no contiene al uno y su límite
inferior es mayor que uno, se concluye que es más probable que se dé el evento
favorable (es decir que la variable respuesta valga 1), si el intervalo de confianza no
contiene al uno y su límite superior es menor a 1, entonces es más probable que se
presente el evento 0 (no preñez en este caso). El intervalo de confianza que se obtuvo
fue del 95%.
2.5. Resultados y discusión 2.5.1. Efecto del día de la aplicación (día 5 u 8) de la eCG sobre el desarrollo del folículo dominante (número y tamaño) En la tabla 2, se muestra la comparación entre los tratamientos utilizados (eCG día 5 y
eCG día 8), en cuanto a las variables de número de folículos dominantes (≥ 9 mm)
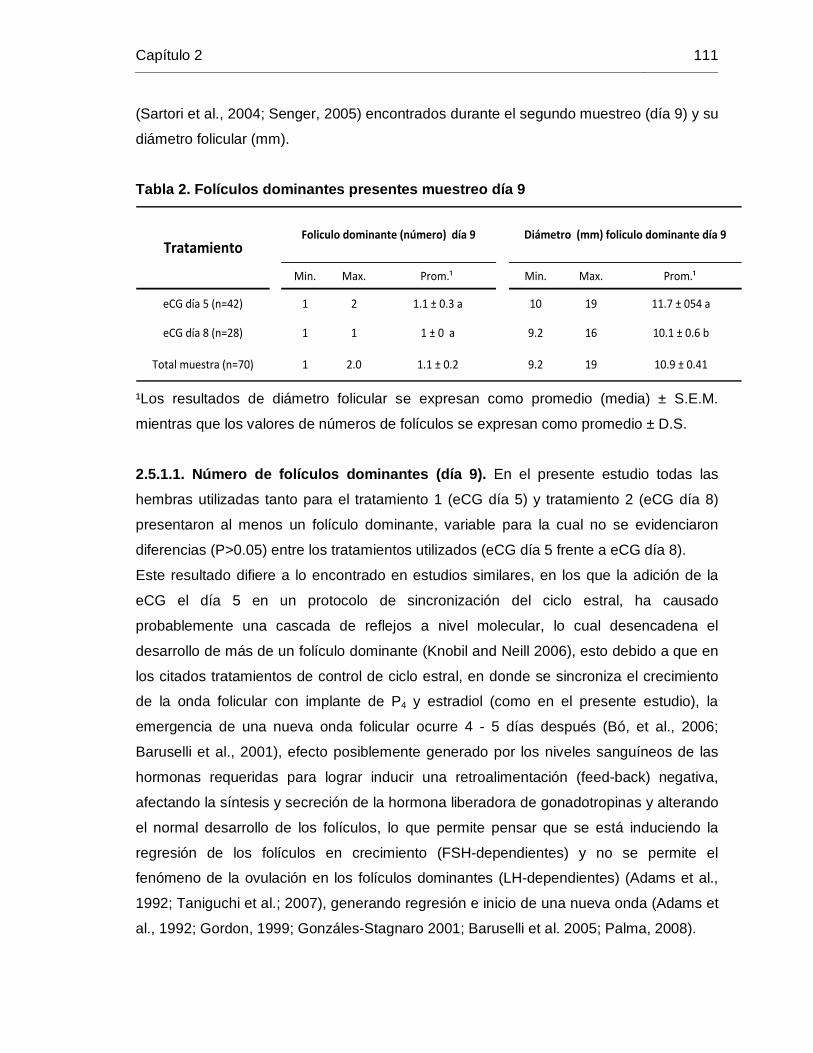
Capítulo 2 111
(Sartori et al., 2004; Senger, 2005) encontrados durante el segundo muestreo (día 9) y su
diámetro folicular (mm).
Tabla 2. Folículos dominantes presentes muestreo día 9
Min. Max. Prom.¹ Min. Max. Prom.¹
eCG día 5 (n=42) 1 2 1.1 ± 0.3 a 10 19 11.7 ± 054 a
TratamientoFoliculo dominante (número) día 9 Diámetro (mm) foliculo dominante día 9
Total muestra (n=70) 1
1 ± 0 a 9.2 16 10.1 ± 0.6 beCG día 8 (n=28) 1 1
2.0 1.1 ± 0.2 9.2 19 10.9 ± 0.41
¹Los resultados de diámetro folicular se expresan como promedio (media) ± S.E.M.
mientras que los valores de números de folículos se expresan como promedio ± D.S.
2.5.1.1. Número de folículos dominantes (día 9). En el presente estudio todas las
hembras utilizadas tanto para el tratamiento 1 (eCG día 5) y tratamiento 2 (eCG día 8)
presentaron al menos un folículo dominante, variable para la cual no se evidenciaron
diferencias (P>0.05) entre los tratamientos utilizados (eCG día 5 frente a eCG día 8).
Este resultado difiere a lo encontrado en estudios similares, en los que la adición de la
eCG el día 5 en un protocolo de sincronización del ciclo estral, ha causado
probablemente una cascada de reflejos a nivel molecular, lo cual desencadena el
desarrollo de más de un folículo dominante (Knobil and Neill 2006), esto debido a que en
los citados tratamientos de control de ciclo estral, en donde se sincroniza el crecimiento
de la onda folicular con implante de P4 y estradiol (como en el presente estudio), la
emergencia de una nueva onda folicular ocurre 4 - 5 días después (Bó, et al., 2006;
Baruselli et al., 2001), efecto posiblemente generado por los niveles sanguíneos de las
hormonas requeridas para lograr inducir una retroalimentación (feed-back) negativa,
afectando la síntesis y secreción de la hormona liberadora de gonadotropinas y alterando
el normal desarrollo de los folículos, lo que permite pensar que se está induciendo la
regresión de los folículos en crecimiento (FSH-dependientes) y no se permite el
fenómeno de la ovulación en los folículos dominantes (LH-dependientes) (Adams et al.,
1992; Taniguchi et al.; 2007), generando regresión e inicio de una nueva onda (Adams et
al., 1992; Gordon, 1999; Gonzáles-Stagnaro 2001; Baruselli et al. 2005; Palma, 2008).

112 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
Cuando la eCG es aplicada el día 5, la onda folicular está en una etapa muy temprana de
desarrollo, en consecuencia, la eCG podría dar lugar a la estimulación en el
reclutamiento y selección de más de un folículo dominante, ya que favorece la síntesis de
mRNA que codifica para los receptores de hormonas gonadotropicas FSH y LH a nivel de
las células de la teca y de la granulosa (Este efecto en el presente estudio solamente fue
observado en 4 de las 42 novillas, por lo cual no se comprobaría la hipótesis planteada al
respecto, ya que no se evidenciaron diferencias estadísticas entre los tratamientos
utilizados).
Con referencia a esto último, Nasser et al., (2004), quienes al aplicar 400 UI de eCG a
dos grupos de novillas receptoras cruzadas Nelore/Angus (eCG día 5 frente a eCG día 8)
con edades entre 22 y 30 meses, las cuales fueron suplementadas con 2 kilogramos/día
de concentrado, encontraron diferencias (P<0.05) de acuerdo al número de folículos
dominantes presentes hacia el día 8, siendo mayor el número de folículos cuando la eCG
se aplicó el día 5 (inicio de una nueva onda de crecimiento folicular), demostrando de
esta forma la acción folículo estimulante de la eCG, la cual estimula la síntesis del mRNA
que codifica para los receptores foliculares de FSH en los folículos en desarrollo (Kuran
et al., 1996; Baruselli et al., 2005), por lo cual se selecciona más de un folículo debido a
que no hay folículos dominantes en ese momento (día 5 de aplicación con referencia al
inicio de protocolo de control de ciclo estral) que inhiban a los demás folículos en
desarrollo; cuando la eCG se aplicó el día 8 (cuando ya la dominancia folicular está
establecida) solamente se presentó una estimulación del folículo dominante (vinculación
de la eCG a los receptores foliculares de FSH y LH) (Cameron and Batt, 1991), por tal
motivo se observaron diferencias estadísticas en el citado artículo.
Baruselli et al (2001), quienes sincronizaron receptoras cruzadas tipo Cebú, a las cuales
aplicaron 800 UI de eCG el día 5 con referencia al ciclo sincronizado (día de inicio del
crecimiento de nueva onda folicular); encontraron diferencias (P<0.05) para el número
promedio de folículos dominantes el día 8 de iniciada la sincronización, 2.04 ± 1.4 (mayor
a lo reportado en el presente estudio) esto frente a un tratamiento control (sin eCG) 0.6 ±
0.5. Este resultado podría indicar que la eCG participa de manera activa favoreciendo la
síntesis de mRNA que codifica para receptores de hormonas gonadotrópicas a nivel
folicular; permitiendo el aumento de la densidad de este tipo de receptores a nivel de
membrana en las células de la teca y de la granulosa de varios folículos (Sartori et al.,

Capítulo 2 113
2001), lo cual podría estar asociado con el aumento de estructuras foliculares
encontradas (Murphy and Martinuk 1991; Sartori et al., 2001). Con este resultado del
citado artículo se podría deducir la posible ventaja al aplicar la eCG frente a un
tratamiento control.
Al no encontrar diferencias estadísticas en esta variable en el presente estudio,
posiblemente podría deberse a el factor raza, en donde existen diferencias en cuanto a
factores como la sensibilidad hormonal (Knobil and Neill 2006), con lo cual se diferencian
las razas de las especies Bos Indicus y sus cruces, que son más sensibles que las razas
de las especies Bos Taurus (Sartori et al., 2004). Si se tiene en cuenta esto último, se
podría deducir que concentraciones de 400 UI de eCG van a desencadenar efecto
diferente en las razas de las dos especies, por lo cual se observa que dicha sensibilidad
no fue tan marcada en el presente estudio en donde se trabajó con novillas de la especie
Bos Taurus. De esto, último se deduce que se debería trabajar con diferentes
dosificaciones de acuerdo a los factores expuestos, ya que dosis de 400 UI de eCG,
como la utilizada en el presente estudio tendría menos probabilidad de estimular el
crecimiento de una gran número de folículos como si lo fue en otros estudios al respecto,
en donde han utilizado 1000 UI (Fuentes y De la Fuente, 2007) y 800 UI (Buruselli et al.,
2001) de eCG, utilizando novillas y vacas lo cual posiblemente también influye
drásticamente en los resultados (Dieleman et al., 1993; Vos et al.; 1994; Nogueira et al.,
2003).
Por otra parte, cuando el tratamiento de eCG se suministra el día 8, posiblemente el
dominio folicular ya está establecido (Buaruselli et al., 2001) y sólo puede tener como
resultado la estimulación de sólo el folículo dominante (en algunos casos el mayor
folículo subordinado) (Knobil and Neill 2006). En el presente estudio esto fue observado,
ya que solamente se evidencia un folículo dominante como promedio para el grupo de
novillas pertenecientes al tratamiento 2 (eCG día 8), máximo uno (1) y mínimo uno (1),
por lo cual se podría asumir el estímulo realizado a este único folículo dominante.
Concordante con lo anterior, la misma variable ha sido medida en otros estudios como en
el de Sartori et al., (2002), en el cual utilizando vacas Holstein a las cuales aplicaron
PGF2α el día 7 (en presencia de Cuerpo lúteo), encontraron en promedio 1.0 ± 0.0
folículos ovulatorios en vacas con ovulación sencilla, y 2.5 ± 0.1 folículos ovulatorios en
vacas con ovulación múltiple (no aplicó eCG, en ninguno de los casos), se observa en el

114 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
primer caso del estudio en referencia la similitud a lo reportado en el presente estudio en
cuanto a los dos tratamientos y el total. Con otros estudios no se ha evidenciado esta
tendencia (Nasser et al., 2004; Machado et al., 2006; Siqueira et al, 2009), debido
posiblemente a la variedad en cuanto a la tipología racial utilizada (Nogueira et al., 2003).
Los resultados encontrados de acuerdo a la variable en mensión el día 9 (nueve), es
similar a los encontrado en los citados estudios, en donde se nota que mínimo un folículo
dominante está presente hacia el día 8 o 9, es decir cuando el folículo ha alcanzado su
máximo desarrollo y se encuentra próximo a ovular.
2.5.1.2. Diámetro folículo dominante (día 9). Todas las novillas utilizadas
(tratamiento 1 – eCG día 5 y tratamiento 2 – eCG día 8) en el presente estudio
presentaron al menos un folículo dominante (folículo ≥ 9 mm) (Sartori et al., 2004;
Senger, 2005) con la aplicación de los protocolos para control de ciclo estral (Sartori et
al., 2002; Perry et al., 2005; Lynch et al., 2010). De acuerdo a esto, se evidenciaron
diferencias (P<0.05) entre los dos tratamientos, siendo mayor el diámetro folicular del
grupo eCG día 5 (tratamiento 1) frente al grupo eCG día 8 (tratamiento 2) (Véase tabla
2).
De acuerdo a los resultados obtenidos se podría decir que son similares a los reportados
por Machado et al., (2006), quienes al comparar tres protocolos para el control del ciclo
estral reportaron diámetros foliculares similares para el día 9 de 9.6±1.7 mm. (Protocolo
1), 9.5±0.6 mm. (Protocolo 2) y 9.8±0.6 mm. (Protocolo 3) en hembras cruzadas (Bos
taurus x Bos indicus). En otros estudios al respecto en donde han utilizado eCG durante
los protocolos de control de ciclo estral, se ha reportado valores diferentes de diámetro
folicular. En tal sentido Filho et al., (2010) reportan para vacas de la raza Nelore,
utilizando dos tratamientos de sincronización para IATF, en donde incluyeron eCG en un
grupo y eCG más GnRH en otro (aplicadas el día 9 con respecto al día de inicio del
protocolo de sincronización), un promedio de diámetro para el folículo dominante de 8.2 ±
0.9 mm, (grupo eCG), y 8.5 ± 0.8 mm (grupo eCG + GnRH), valores algo menores a los
reportados en el presente estudio; los cuales en otras investigaciones han sido similares
sobre todo cuando comparamos a los resultados obtenidos con el tratamiento 1 (eCG día
5). Así lo reportan Machado et al., (2006), los cuales trabajaron con la raza Nelore, en
donde utilizaron un protocolo para control de ciclo y aplicaron 400 UI de eCG el día 9, en

Capítulo 2 115
donde obtuvieron 12.72 ± 0.39 mm de diámetro folicular. Estos resultados son similares a
los reportados por Filho et al., (2004) quienes no encontraron diferencias significativas en
el diámetro folicular en un grupo de vacas Nelore sincronizadas para Inseminación
Artificial a Tiempo Fijo (IATF), en donde suministraron eCG el día 9 (12.4 ± 0.05 eCG vs,
12.1 ± 0.04 mm sin eCG).
En otros estudios en donde se ha trabajado con el mismo tipo racial utilizado en el
presente estudio, se han reportado valores superiores a los encontrados en la presente
investigación., es el caso del reporte de Sousa et al., (2009), en donde aplicaron a un
grupo de vacas de la raza Holstein 400 UI de eCG el día 8 (con referencia al día de inicio
de la sincronización), en donde encontraron un tamaño folículo dominante de 14.7 ± 0.6
mm y de 13.1 ± 0.6 mm en las vacas sin eCG. Igualmente Sartori et al., (2004), utilizando
específicamente novillas y vacas lactantes Holstein encontraron tamaños de folículos
ovulatorios de 14.9 ± 0.2 mm en las novillas y 16.8 ± 0.5 mm para las vacas. (Los
animales con CL presente no fueron tratados con ningún tipo de hormona, los otros con
GnRH). El mismo autor, Sartori et al., (2002) comparando vacas con ovulación sencilla y
vacas con ovulación múltiple de la misma raza, en las cuales aplicaron PGF2α el día 7
(en presencia Cl), encontraron diámetros foliculares de 15.8 ± 0.4 mm (en ovulación
sencilla), y 13.7 ± 0.3 mm (ovulación doble).
Con la aplicación de PGF2α con intervalo de 11 días, se han conocido reportes, como es
el caso de Lynch et al., (2010), los cuales utilizando vacas cruzadas tipo carne Bos
Taurus, sincronizadas con la aplicación de dos dosis de PF2α (11 días de intervalo),
encontraron folículos preovulatorios de 14.1 ± 1.9 mm de diámetro.
También se han realizado estudios en donde se ha tratado de sincronizar el crecimiento
de la onda folicular sin la utilización de hormonas, en donde se han encontrado
diferencias estadísticas. Este es el caso del reporte de Vasconcelos et al., (2001) en
donde se observan diámetros foliculares dominantes de 12.53 ± 0.37 mm en vacas
lecheras sin tratamiento alguno, frente a vacas tratadas con aspiración folicular 8.69 mm
± 0.35.
Se observa la gran variedad de resultados reportados en los diferentes estudios
referenciados, en tal sentido, se plantea que la tipología racial, Bos Indicus y sus cruces,

116 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
poseen menor tamaño ovárico, lo cual se reflejaría de igual forma en sus estructuras
producidas (folículos y cuerpos lúteos), esto frente a las encontradas en razas Bos
Taurus como la utilizada en el presente estudio (Gordon, 1999 Gonzáles – Stagnaro,
2001; Palma; 2008), pero esto no sería definitivo, ya que se evidencian otros elementos
como los tratamientos o dosificaciones utilizadas, estados de desarrollo (novillas, vacas),
número de partos, estado nutricional etc., lo cual podría explicar de alguna manera los
diferentes efectos encontrados al respecto en los reportes referenciados. (Dieleman et
al., 1993; Vos et al.; 1994; Nogueira et al., 2003; Rekwolt, 2004; Sartori et al., 2004;
Roche, 2006; Knobil and Neill 2006). De acuerdo a los diversos factores nutricionales que
indudablemente podrían incidir sobre la dinámica folicular de las novillas (Roche, 2006),
se podría de alguna manera pensar que las novillas utilizadas en el presente estudio se
encontraban en pastoreo, sin suplemento adicional que garantizara el cubrimiento total
de sus necesidades fisiológicas, por lo cual sería evidente un desbalance nutricional, lo
cual generaría efecto inicial a nivel del hipotálamo y consecuentemente de la hipófisis
anterior (adhenohipofisis), que afectaría la síntesis y secreción de las hormonas
gonadotrópicas, por lo cual se podría ver afectado el desarrollo y crecimiento folicular
(Rekwolt, 2004; Roche, 2006).
Debe tenerse en cuenta que en el presente estudio las mediciones de los folículos se
realizaron en la mañana del día 9 (antes de la aplicación de BE); posterior a esto a cada
novilla se le aplicó 1 ml (5 mg.) de benzoato de estradiol, con la finalidad de estimular la
síntesis y secreción de FSH y LH, para favorecer el desarrollo final del folículo, así como
su ovulación (Baruselli et al., 2005; Spey et al., 2006), por lo cual podría pensarse que
aún no se ha completado la etapa de desarrollo final del folículo preovulatorio antes de su
ovulación, en la cual por la acción de las hormonas gonadotrópicas, este incrementa su
diámetro final, motivo por el cual los promedios de diámetro folicular podrían ser
inferiores comparados con los resultados de la mayoría de estudios referenciados, en
donde las mediciones se realizaron en el momento de máximo desarrollo folicular (día 9)
(Vasconcelos, 2001; Sartori et al., 2002; Sartori et al., 2004; Perry et al., 2005; Perry et
al., 2007).
En cuanto al efecto encontrado (P<0.05) con el tratamiento 1 (eCG día 5 – Día de inicio
de la onda folicular del ciclo sincronizado) en el presente estudio, posiblemente se deba a
que la eCG ha causado el desencadenamiento de la cascada de eventos a nivel

Capítulo 2 117
molecular que provoca un mayor desarrollo del folículo dominante (Knobil and Neill
2006).
La eCG estimula la formación de mRNA que codifica para receptores de FSH y LH,
estímulo que sigue incrementándose debido a que el folículo se encuentra en etapa muy
temprana de dominancia y durante este periodo sigue actuando la eCG la cual tiene una
vida media de 3 días (Bó et al, 2004), por lo cual prácticamente se garantiza su acción
durante el tiempo que el folículo se va haciendo dominante, permitiendo el aumento de la
densidad de los receptores de FSH y LH, lo cual se asocia a crecimiento folicular. Por
otro lado, comparado con las novillas del grupo del tratamiento 2, (eCG día 8), cuya
aplicación se realizó cuando la dominancia folicular prácticamente ya estaba establecida,
el efecto estimulador por parte de la eCG sería muy corto.
2.5.2. Efecto del día de la aplicación de la eCG sobre el desarrollo del cuerpo lúteo (número y volumen), porcentaje de doble ovulación, porcentaje de utilización, concentración plasmática de P4 y porcentaje de preñez En la tabla 3, se reportan los resultados de las variables cuerpo lúteo (número) día 17,
porcentaje de doble ovulación día 17, volumen luteal (mm3) día 17, porcentaje de
utilización día 17, concentración plasmática de P4 (ng/ml) días nueve (9) y diecisiete (17)
y porcentaje de preñez (%) día cincuenta y dos (52), comparados entre tratamientos
(eCG día 5 frente a eCG día 8).
La discusión correspondiente a las variables mostradas en la tabla 3, volumen luteal día
17, concentración de P4, día 17 y porcentaje de preñez día 52, se realizará en conjunto al analizar el porcentaje de preñez (numeral 2.5.2.4), esto con el fin de comprender un
poco más los reportes de estudios al respecto en donde la mayoría tratan estas tres (3)
variables en conjunto, lo cual podría facilitar la comprensión de los resultados.

118 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
Tabla 3. Comportamiento de estructuras luteales y su volumen, concentración de progesterona y porcentaje de preñez
Min. Max. Prom.¹ Min. Max. Prom.¹ Min. Max. Prom.¹ Min. Max. Prom.¹
eCG día 5 (n=42) 1 2 1.1 ± 0.3 a 375 16443.6 5023.6 ± 512.4 a 0 0.84 0.28 ± 0.03 b 1.98 10.1 5.12 ± 0.31 a
375.4
5554.9 ± 758.6 a
2 10 5.2 ± 0.22 67 (47/70)5236.1 ± 429.7 100 (70/70) 0
Día 9
0.9 0.3 ± 0.03
0.33 ± 0.05 b 2 9.6 5.52 ± 0.30 a 64 (18/28) a
1 2 1.1 ± 0.2 5.7 18057.3
100 (28/28) a
Preñez (%) día 52
9,5 a 100 (42/42) a 69 (29/42) a
1 1 1 ± 0 a 0 a 1150.35 18057.3
Día 17% de
utilización día 17
Concentración de Progesterona (ng/ml)
0 0.86
Cuerpo lúteo (número) día 17 % Doble
ovulación (día 17)
Volumen luteal (mm3) día 17Tratamiento
eCG día 8 (n=28)
Total muestra (n=70)
¹Los resultados volumen luteal y concentración de progesterona se expresan como
promedio (media) ± S.E.M. mientras que los otros valores se expresan como promedio ±
D.S.
2.5.2.1. Cuerpo lúteo (número) día 17. Todas las novillas (70/70) de los dos
grupos experimentales del presente estudio (42 novillas pertenecientes al tratamiento 1 –
eCG día 5 y 28 novillas pertenecientes al tratamiento 2 – eCG día 8) presentaron al
menos un cuerpo lúteo consolidado y funcional para el día 17 (etapa de diestro) con la
aplicación de los protocolos de control de ciclo estral planteados, por lo cual se podría
pensar que igualmente todas las novillas ovularon, esto si tenemos en cuenta que todas
presentaron al menos un folículo dominante hacia el día 9 (tabla 2).
De acuerdo al promedio de esta variable, no se evidenciaron diferencias (P>0.05) entre
los tratamientos utilizados (eCG día 5 frente a eCG día 8), esto fue paralelo en cuanto al
promedio de folículos dominantes evidenciados (tabla 2) durante el experimento.
Contrariamente, en un estudio similar al presente, realizado por Nasser et al., (2004), se
reportan diferencias significativas entre los dos tratamientos utilizados (eCG el día 5 u 8),
en los cuales suministraron 400 UI de dicha hormona a receptoras cruzadas
Nelore/Angus, sincronizadas para la transferencia de embriones a tiempo fijo, en donde
se obtuvieron 1.44 ± 0.18 cuerpos lúteos (eCG día 5) frente a 1.03 ± 0.03 cuerpos lúteos
(eCG día 8), utilizando animales con tipología racial más sensible en cuanto a las
dosificaciones hormonales utilizadas (Spey et al., 2006; Baruselli et al., 2005). Sin
embargo los valores encontrados en el citado estudio cuando la eCG se aplicó el día 8,

Capítulo 2 119
son similares a los del presente reporte, lo cual posiblemente se deba a la similitud de
tratamientos de sincronización utilizados entre los dos estudios.
En otro estudio realizado por Baruselli et al. (2001), en el cual utilizaron tratamiento
control (sin eCG), encontraron diferencias estadísticas, utilizando receptoras para
embriones cruzadas tipo cebú, a las cuales aplicaron 800 UI de eCG (día 5 con
referencia al ciclo sincronizado – crecimiento de nueva onda folicular). En este estudio se
reporta 2.58 ± 2.93 cuerpos lúteos en animales con eCG y 0.5 ± 0.5 cuerpos lúteos en
animales sin eCG. Estos hallazgos podrían evidenciar la acción superovulatoria de la
eCG causada por la dosis de 800 UI suministrada el día de inicio de la onda folicular (Bó
et al, 2004). Este efecto superovulatorio también se ha visto en otros estudios como el
realizado por Fuentes y De la Fuente (2007), los cuales trabajaron con novillas Holsteín a
las que suministraron 1000 UI de eCG el día 5 (con referencia al inicio del tratamiento).
En este estudio observaron que dicha hormona aumentaba significativamente el número
de cuerpos lúteos (entre 2 y 5 estructuras en cada ovario en el momento de la
transferencia).
Estos resultados de acuerdo a la variable en cuestión, también podrían estar indicando la
diferente acción de la eCG de acuerdo a la dosificación utilizada, tipo racial y el estado
productivo del animal (vaca o novilla), lo cual podría influenciar como se explicó
anteriormente los resultados obtenidos en cuanto al número de folículos (Dieleman et al.,
1993; Vos et al.; 1994; Nogueira et al., 2003; Rekwolt, 2004; Sartori et al., 2004; Roche,
2006; Knobil and Neill 2006).
2.5.2.1.1. Porcentaje de doble ovulación (día 17). En cuanto a esta variable la cual se
deriva de los folículos dominantes presentes (día 9) y de los cuerpos lúteos evidenciados
(día 17), no se evidenciaron diferencias (P>0.05) entre los dos tratamientos utilizados
(eCG día 5 frente a eCG día 8).
La doble ovulación solamente fue observada en cuatro (4) de las cuarenta y dos (42)
novillas (9.5%) pertenecientes al tratamiento 1 (eCG día 5). Este porcentaje es menor al
encontrado por Sousa et al., (2009), quienes encontraron encontraron 12.5% del total de
las vacas Holstein con doble ovulación aplicando 400 UI de eCG (día 8 con referencia al
día de inicio del protocolo).

120 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
Igualmente Vasconcelos et al., (2001), reportan un 19.1% de dobles ovulaciones en
novillas de la raza Holstein bajo un protocolo Ovsynch y sincronizando el crecimiento de
la onda folicular utilizando aspiración folicular (folículos > 4 mm), esto frente a un grupo
control (sin aspiración) 9.5% (2/21), siendo este último porcentaje similar al porcentaje
reportado en el presente estudio.
De otro lado, los resultados de doble ovulación del presente estudio son mayores al 1.3%
(1/75) reportado por Sartori et al., (2002), utilizando novillas Bos Taurus sincronizadas
con PGF2α el día 7 (únicamente se aplicó en presencia de cuerpo lúteo), pero no ocurrió
lo mismo con vacas para lo cual se reporta 63.5% (33/52) de dobles ovulaciones. Esto
contrasta a otros reportes encontrados igualmente por Sartori et al., (2004) en vacas
Holstein 1.9 % (1/54) y en novillas el 17.9 % (5/28) utilizando solamente tratamientos de
GnRH (sin eCG) en hembras con cuerpo lúteo presente.
Como se puede observar, los resultados de esta variable (porcentaje de doble ovulación)
son contradictorios entre los citados reportes y el presente estudio. En tal sentido se
observa que la adición de la eCG el día 5 en un protocolo de sincronización del ciclo
estral, podría estar causando el desarrollo de más de un folículo dominante. Esto se
fundamenta ya que el tratamiento de sincronización con P4 y BE sincroniza el crecimiento
de una nueva onda folicular aproximadamente 4 a 5 días. Por tal motivo, al suministrar la
eCG (día 5), la onda folicular está en una etapa temprana de crecimiento, en
consecuencia permite la estimulación de más de un folículo dominante, lo cual en el
presente estudio solamente fue observado en 4 de las 42 novillas, lo cual no comprueba
estadísticamente la posible acción de la eCG para poder dar una conclusión valida al
respecto.
Se plantea que la dosificación de 400 UI de eCG suministrada a los dos grupos de
novillas del presente estudio, podría tener una probabilidad menor de estimular el
crecimiento de un número amplio de folículos; esto frente a lo observado en otros
estudios, en donde las dosificaciones han sido mayores (Buaruselli et al., 2001; Fuentes
y De la Fuente, 2007), estudios en donde también podría estar influyendo la edad, el
estado de desarrollo (novillas y vacas) y la raza (Bos Indicus y Bos Taurus), siendo más
sensibles a los tratamiento hormonales las razas Indicas, frente a las Taurinas (Randel
and Moseley, 1997).

Capítulo 2 121
Por otra parte cuando el tratamiento de eCG se suministra el día 8, el dominio folicular ya
está establecido y sólo podría tener como resultado la estimulación del folículo dominante
(Lynch et al., 2010; Perry et al., 2005; Sartori et al., 2002), lo cual posiblemente explica el
resultado obtenido al respecto en el presente trabajo con el grupo de novillas
pertenecientes al tratamiento 2 (eCG día 8), las cuales reflejan lo anterior de acuerdo a
los folículos ovulados.
2.5.2.2. Volumen luteal (mm3) día 17. Para esta variable no se evidenciaron
diferencias (P>0.05) entre los grupos tratados (eCG día 5 frente eCG día 8).
Se destaca la sincronía causada en cuanto alaevidencia de estructuras luteales
encontradas el día 17, por lo que se evidencia que el 100% de las novillas en los dos
tratamientos (tratamiento 1 – eCG día 5, 42/42 y eCG día 8, 28/28) presentaron por lo
menos una estructura luteal consolidada.
2.5.2.2.1. Efecto del diámetro del folículo dominante (mm) día 9 sobre el volumen luteal (mm3) día 17. Independientemente a los tratamientos de eCG utilizados (eCG día 5 y eCG día 8), el
análisis de regresión del diámetro (mm) del folículo dominante (día 9 del ciclo
sincronizado) sobre el volumen (mm3) luteal (día 17 del ciclo sincronizado), en la totalidad
de las novillas del estudio (n=70) no encontró significancia (P>0,05).
Estos resultados coinciden con lo reportado por Lynch et al., (2010), quienes hacen
referencia al tamaño del folículo preovulatorio frente al volumen luteal. En dicho estudio
no se encontró relación entre el diámetro folicular y el diámetro luteal en un grupo de
novillas (Bos Taurus), las cuales recibieron un tratamiento con prostaglandina para el
control del ciclo estral. En la misma investigación, Lynch et al., (2010), refuerzan lo
encontrado al analizar los niveles de hormonas esteroideas circulantes y sugieren que no
hay relación entre la concentración de estrógenos producidos durante el periodo
periovulatorio y los niveles de progesterona presentes durante el día 7 posterior a la
ovulación. Esto permitiría pensar que la capacidad esteroideogénica del cuerpo lúteo
consolidado no está asociada con la capacidad esteroideogénica de folículo dominante
del cual se formó.

122 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
Igualmente Vasconcelos et al., (2006), muestran bajos porcentajes de sobrevivencia
embrionaria en hembras que presentan folículos preovulatorios por encima de la media
del grupo. Esto permitiría pensar que al presentarse folículos grandes durante el
momento de la ovulación podría generar una cavidad ovulatoria (fosa de ovulación),
haciendo que se presenten cuerpos lúteos cavitarios; los cuales van a presentar una
menor cantidad de estroma luteal (volumen luteal) y menor cantidad de células luteales
productoras de progesterona (Kastelic et al., 1990a), siendo así, menor su síntesis de
progesterona frente a cuerpos lúteos no cavitarios, esto dependiendo de la cantidad de
volumen luteal presente (Duica 2011), por lo cual el ambiente embriotrofico será
diferente entre las dos situaciones, con lo cual se podría afectar la sobrevivencia
embrionaria y la tasa de concepción (1997; Lynch, 2010; Grygar).
2.5.2.2.2. Porcentaje de utilización día 17. En cuanto a esta variable
(hembras transferidas/hembras sincronizadas*100), no se presentaron diferencias
(P>0.05) entre los dos grupos tratados (eCG día 5 frente a eCG día 8).
Los porcentajes reportados en cuanto a esta variable permiten evidenciar que la totalidad
de las novillas tratadas (70/70 - 100%) con los protocolos de sincronización de ciclo estral
tanto para el tratamiento 1 (eCG día 5 – 42/42) como el tratamiento 2 (eCG día 8 –
28/28), presentaron adecuado desarrollo de estructura luteal (cuerpo luteo consolidado)
(Nasser et al., 2004) para que fuera transferido el embrión el día 17 (cuerpo lúteo
consolidado).
Menores porcentajes de utilización han sido reportados en varios estudios, es así que
investigadores como Pita et al., (2009) utilizando novillas cruce por Cebú, encontraron 84
%, de utilización con la aplicación de la eCG el día 5 del inicio del protocolo y 65 %
aplicando la eCG el día 8; en este estudio se evidenciaron diferencias estadísticas.
Igualmente se reportan menores porcentajes de utilización en un estudio realizado por
Nasser et al., (2004), en el cual utilizaron novillas receptoras Nelore/Angus sincronizadas
con 400 UI de eCG, reportan 90.7% (eCG día 5) y 82.7% (eCG día 8), estudio en el cual
tampoco se reportaron diferencias estadísticas. Igualmente Fuentes y De la Fuente
(2007), reportan menores porcentajes de utilización, 83%, utilizando novillas Holstein a

Capítulo 2 123
las cuales aplicaron 1000 UI de eCG el día 5 con referencia al inicio del tratamiento de
control de ciclo estral.
En general se observa que los porcentajes reportados en las investigaciones citadas
tienen en común que los menores porcentajes de utilización se han obtenido en los
grupos de hembras tratadas con eCG el día 8 con respecto a los grupos tratados el día 5,
lo cual no fue evidenciado en el presente estudio en el cual el porcentaje de utilización
fue 100% en cada uno de los tratamientos (eCG día 5 y eCG día 8).
Los resultados de la presente variable confirman los estudios anteriores (Fuentes y De la
Fuente, 1997; Tribulo et, al 2002; Bó et al, 2002) en los que la adición de la eCG en el
periodo de dominancia folicular (día 8) o de inicio de onda folicular (día 5) en un protocolo
de sincronización del estro, aumenta el porcentaje de hembras transferidas de acuerdo a
las sincronizadas (Bó y Caccia, 2003; Baruselli et al., 2005; Barret et al., 2006), en donde
la aplicación de la eCG (la cual tiene acción FSH y LH) durante el periodo de dominancia
(día 8) podría generar que el folículo dominante mantenga su desarrollo y crecimiento en
la fase final con la finalidad de que llegue a ser un folículo preovulatorio (Evans et al.,
1997). Este desarrollo de este folículo se relaciona con la síntesis de receptores para
FSH y LH, así como la elaboración de las aromatasas (Bao et al., 1997).
La expresión de un mayor número de receptores para la LH en las células de la
granulosa se considera como un mecanismo para la adquisición de dominancia (Sartori
et al., 2001), en referencia a esto, se ha encontrado mRNA que codifica para receptores
de LH en el folículo dominante; el crecimiento de este folículo está asociado con la mayor
expresión de este mRNA en las células de la granulosa (Mihm et al., 2000); esto
igualmente se relaciona con la preparación celular para la formación del cuerpo lúteo
(Revisado por Jiménez en: Hernández 2008). Por otro lado, con la aplicación de la eCG
hacia los días de inicio de la onda folicular (día 5), podría generar selección de varios
folículos, gracias a la acción FSH de la eCG, con lo cual podría generarse más de un
folículo dominante, lo que podría conllevar a la formación de varios cuerpos lúteos
(Revisado por Olivera et al., 2007). Esto no ocurrió en el presente estudio, pero
posiblemente la eCG favoreció la formación y consolidación del folículo en sus etapas de
selección y dominancia de las novilas pertenecientes al tratamiento 1 (eCG día 5).

124 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
2.5.2.3. Concentración de progesterona ng/ml Se debe aclarar que el día del muestreo 1 (día 5) definitivamente no podría haber efecto
del tratamiento con eCG para esta variable, ya que en el momento del muestreo se
colocó dicha hormona en el grupo del tratamiento 1; y en el grupo del tratamiento 2 se
colocó el día 8. Sin embargo se reporta el comportamiento de la concentración de P4 día
5 en la gráfica 2 y 3 con la finalidad de analizar la tendencia entre los grupos escogidos.
En estas gráficas se observan rangos de valores entre mínimos y máximos que pueden
ser asociados a dispersión en los datos causada por la irregularidad en las fases del ciclo
estral en la que se encontraban las novillas de los grupos experimentales utilizados al
momento de iniciar los tratamientos de sincronización, lo que indicaría que dentro del
grupo de novillas se encontraba un porcentaje importante de estas con cuerpos lúteos
presentes al inicio del tratamiento de sincronización, lo cual respaldaría lo reportado por
la literatura, en donde se habla que aproximadamente entre el 40 y 50% de un grupo de
hembras aptas reproductivamente podrían presentar estructuras luteales al realizar una
exploración reproductiva (Palma, 2008; Gordon et al., 1999). Por lo cual era de
esperarse en el presente estudio que un porcentaje importante de las novillas
presentaran niveles basales de P4 superiores a 1ng/ml (Grafica 2). Igualmente debe
tenerse en cuenta que el dispositivo de liberación lenta de P4 aplicado el día cero “0” del
protocolo para el control del ciclo estral, favoreció de alguna manera el incremento de
esta hormona evidenciándose durante el primer muestreo (día 5) (Baruselli, 2003; Bó,
2003).
2.5.2.3.1. Concentración de progesterona día 9. Para este día de
muestreo tampoco se evidenciaron diferencias (P>0.05) en cuanto a la concentración de
P4 entre tratamientos (eCG día 5 frente a eCG día 8).
Se debe recordar que el día el cinco (5) del inicio del protocolo de sincronización se
aplicó prostaglandina F2α la cual desencadenó la regresión funcional y estructural de
cualquier cuerpo lúteo existente (Barnea et al., 2000). Por tal motivo, al realizar la
exploración ultrasonográfica ovárica durante este día 9 (muestreo 2), no se encontraron
estructuras compatibles con cuerpos lúteos, lo cual se corrobora al observar las
concentraciones de P4 plasmática de los grupos de novillas en seguimiento, ya que al no
encontrarse estructuras luteales es de esperarse que los niveles sanguíneos de P4

Capítulo 2 125
durante los días 9 y 10 (ciclo sincronizado) se encuentren muy cercanos a niveles
basales: ≤ 0.3ng/ml (Ohtani, 1998).
Las concentraciones basales de P4 del presente estudio (véase gráfica 3), han coincidido
con las reportadas por algunos autores, como Kastelic et al., (1990), quienes en un grupo
de novillas de la raza Holstein relacionaron las medidas del cuerpo lúteo, obtenidas por
medio de ultrasonografia, con las concentraciones plasmáticas de P4 obteniendo como
resultado para el día 9, una máxima caída en los niveles de P4: ≤ 0.3ng/ml. Igualmente
Perry et al., (2005), en un grupo de vacas cruzadas con características cárnicas, a las
que aplicaron terapia hormonal para sincronizar el ciclo estral (GnRH, PGF2α, GnRH),
reportaron valores de concentración de P4 de 0.2 ng/ml, tanto para el grupo de hembras
gestante como para el grupo de no gestantes. Valores similares han sido reportados por
Chagas et al., (2002), trabajando con novillas de la raza Holstein receptoras de
embriones las cuales no fueron sometidas a tratamiento de control de ciclo estral
(embrión transferido con celo referencial), 0.21 ± 0.01 ng/ml. Igualmente Aguirre et al.,
(2006), quienes trabajaron con vacas de la raza Brahman, 0.224 ng/ml.
Otros autores han reportado niveles de P4 inferiores a los del presente estudio. Es así
como Sakase et al., (2006), en un trabajo en el que compararon 3 protocolos para el
control del ciclo estral, reportaron como valores mínimos de progesterona plasmática, en
el primer y tercer tratamiento, de 0.09 ng/ml y en el segundo de 0.08 ng/ml, durante el
noveno día del ciclo estral sincronizado.
Similares concentraciones de P4 en la fase folicular han sido referenciadas a nivel de
trópico bajo colombiano, es así que Grajales (2000), al momento del celo de acuerdo a
diferentes grupos raciales encontró para vacas tipo Cebú 0.165 ± 0.11 ng/ml.,
Romosinuano 0.221 ± 0.20 ng/ml., Holstein por Cebú 0.331 ± 0.36 ng/ml. y Simmental
por Cebú 0.347 ± 0.31 ng/ml.).
Los anteriores hallazgos, obtenidos por diferentes autores podrían considerarse acordes
con los datos arrojados por el presente estudio, ya que se presentaron tendencias
numéricas que permiten observar que el día en que se presentó máximo desarrollo
folicular, los niveles de P4 plasmática cayeron a niveles basales (Kastelic, 1990; Ohtani,
1998; Chagas, 2002; Perry, 2005; Sakase, 2006). Esto está asociado a la aplicación de

126 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
prostaglandina F2α (el día 5 del ciclo sincronizado), la cual favorece la dramática
disminución de la irrigación sanguínea que va al ovario, desencadenando la cascada de
mecanismos luteolíticos. De esta manera, para el día 9 se espera que a nivel ovárico se
encuentre un cuerpo albicans en el lugar en que se desarrollo el cuerpo lúteo y por lo
tanto niveles basales de P4 (Niswender, 2000; Schams and Berisha, 2004). En el
presente estudio la prostaglandina F2α tuvo efecto luteolítico, motivo por el cual
posiblemente no se encontraron diferencias estadísticas la concentración de P4 (día 9).
2.5.2.3.2. Concentración de progesterona día 17. No se presentaron
diferencias (P>0.05) en cuanto a la concentración de P4 entre tratamientos (eCG día 5
frente a eCG día 8).
Durante este muestreo, como era de esperarse a nivel ovárico se encuentra por lo menos
un cuerpo lúteo desarrollado y funcional, cuyos niveles de progesterona plasmática
tuvieron tendencia a la dispersión (Grafica 2), lo cual es consecuente a la diferencia entre
los tamaños de estructuras luteales evidenciadas.
2.5.2.3.2.1. Efecto del volumen luteal (mm3) día 17 sobre la concentración plasmática de progesterona (ng/ml) día 17.
La gráfica 1, muestra la función estimada mediante el análisis de regresión, la cual
resultó significativa (P<0,05), para la relación entre el volumen del cuerpo lúteo y la
concentración plasmática de progesterona para el día 17 (Muestreo 3)
independientemente a los tratamientos de eCG utilizados (eCG día 5 y eCG día 8), en la
totalidad de las novillas utilizadas durante el estudio (n=70). El análisis de regresión
mostró correlación positiva entre estas dos variables, lo cual indica que si aumenta el
volumen del cuerpo lúteo, aumenta la concentración de P4, y a mayor P4 implica un
mayor volumen luteal el cual la produjo, lo cual probablemente amentará la posibilidad de
adecuar un mejor ambiente embriotrofico necesario para que el conceptus se desarrolle,
con lo cual enviará señales más apropiadas para ser reconocido maternamente, lo que
podría desencadenar posiblemente en mayores porcentajes de preñez (Revisado por
Gonella, 2011).

Capítulo 2 127
Grafica 1. Relación estimada de la concentración de progesterona (ng/ml) en función del
volumen luteal (día 17).
y = 364.1x + 2970.R² = 0.053
0.0
1000.0
2000.0
3000.0
4000.0
5000.0
6000.0
7000.0
8000.0
9000.0
10000.0
11000.0
12000.0
13000.0
14000.0
1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5 5.0 5.5 6.0 6.5 7.0 7.5 8.0 8.5 9.0 9.5 10.0 10.5 11.0
Vo
lum
en
de
cu
erp
o l
ute
o (
mm
3)
Concentración de progesterona (ng/ml)
Concordante con lo anterior, España et al., (2004), encontraron correlación (P<0.001)
entre la concentración plasmática de progesterona y el área luteal, en vacas Holstein.
Los autores proponen que el área luteal podría asociarse con los niveles de progesterona
plasmática y esta a su vez, con el diagnóstico de gestación. Igualmente Spell et al.,
(2001) sincronizando receptoras Angus con tratamiento tradicional de dos
prostaglandinas separadas por 11 días y con implante de P4 por siete días (sin eCG)
encontraron una correlación significativa y positiva entre el volumen de tejido luteal y la
concentración de P4.
Sartori et al., (2002), utilizando vacas lactando, vacas secas y novillas Holstein a las
cuales sumunistraron PGF2α el día 7 del ciclo estral, igualmente encontraron correlación
entre el volumen luteal y la concentración plasmática de P4.
De la misma forma Rodríguez et al. (2007), en un estudio realizado en el trópico medio y
bajo colombiano, utilizando varios tipos raciales y cruces, reportaron correlación positiva
entre estas dos variables, lo cual coincide igualmente con el reporte de Duica (2011),
quien trabajó con novillas Holstein receptoras de embriones a las cuales se les aplicó
eCG hacia el día 5 (con referencia al ciclo sincronizado).

128 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
En otros estudios se ha relacionado el peso del cuerpo lúteo y la concentración
plasmática de P4. En tal sentido autores como Mann (2009) utilizando vacas multíparas
Holstein encontró para el día 5 (con referencia al celo) una fuerte relación entre el peso
del cuerpo lúteo y la concentración plasmática de progesterona. Sin embargo, esta
relación no estaba presente en el día 8 (con relación al celo), lo cual es contrario a lo
encontrado en el presente estudio.
En otros estudios se ha encontrado variación de esta correlación, es así que autores
como Howell et al., (1994), encontraron variación según la estación del año en que se
realizó el experimento. Otros autores también han demostrado correlación positiva entre
el área total del CL y la P4 en leche (Sprecher et al., 1989). Otros, sin embargo, han
observado índices de correlación insuficientes entre estas dos variables (Badinga et al.,
1994; Viana et al., 1998).
De acuerdo a los resultados de los diferentes estudios autores como Kastelic et al.,
(1990b), recomiendan que la valoración por medio de ultrasonografia de los cuerpos
lúteos se convierte en una alternativa viable para la determinación de las
concentraciones de progesterona por la valoración de la función luteal en novillas de la
raza Holstein.
Con respecto a las variaciones encontradas en algunos estudios podría decirse que la
cantidad de tejido luteal, formado por las células luteales grandes y pequeñas
productoras de P4, está relacionada con la concentración de dicha hormona en plasma,
pero no siempre los CL son funcionales, ya que posibles alteraciones de las células
mencionadas o modificaciones en algunos de sus componentes podrían alterar su
secreción (Revisado por Duica, 2011). Igualmente debe tenerse en cuenta que la
correlación entre área luteal y progesterona plasmática no es constante a lo largo de todo
el ciclo estral (Tom et al., 1998; Niswender, 2000), de manera que durante la fase de
regresión este índice perderá significancia, lo cual no fue el caso del presente estudio, en
el cual si se encontró correlación, posiblemente debido a que el día del muestreo 3 (día
17 con respecto al inicio del tratamiento de sincronización – transferencia embrionaria) se
detectó volumen luteal perteneciente a uno o varios CL (s) funcional (es) pos ovulación
en todas las novillas, así como igualmente lo han descrito en varios estudios al respecto

Capítulo 2 129
(Gonzales – Stagnaro et al., 1993; Howell et al., 1994; Ribadu et al., 1994; Battocchio et
al., 1999).
2.5.2.3.2.2. Concentraciones de progesterona durante el ciclo estral sincronizado En la grafica 2 se observa la curva de comportamiento en cuanto a la concentración de
P4 encontrada durante los tres muestreos realizados en la totalidad de las novillas (n=70)
(Muestreo 1 – día 5, Muestreo 2 – día 9, Muestreo 3 – día 17).
Grafica 2. Comportamiento general de la progesterona durante los tres (3) muestreos
realizados en la totalidad de las novillas
-0.50.31.11.92.73.54.35.15.96.77.58.39.19.9
10.711.512.313.113.9
4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
PR
OG
ESTE
RO
NA
(ng
/ml)
DÍAS DE MUESTREO
De acuerdo a la grafica podría deducirse, como se explicó anteriormente de forma
detallada, que el nivel de P4 para el día 5, es debido posiblemente a cuerpos lúteos
funcionales presentes antes de la aplicación de la prostaglandina F2α, ya que el 98.6%
(69/70) de la totalidad de las novillas del estudio presentaron concentraciones de P4 ≥ 1
ng/ml durante el primer muestreo (día 5). Igualmente se podría plantear que es causado
por el efecto residual causado por los implantes de liberación lenta de P4, los cuales
estaban presentes para el día del primer muestreo (día 5).

130 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
Teniendo en cuenta que el día 5 inmediatamente posterior al muestreo 1 se aplicó
prostaglandina F2α, se observa en la curva (muestreo 2 – día 9) la caída en los niveles
de P4 debido a la luteolisis funcional y estructural causada por el factor luteolítico de la
prostaglandina (Baruselli et al., 2005; Barnea et al, 2000), lo que posiblemente permitió el
desarrollo y dominancia folicular (Baruselli et al., 2003; Vasconcelos, 2001).
Igualmente se observa en la gráfica 2 un patrón de comportamiento, en el que al inicio y
al final (días 5 y 17) existe dispersión en el rango de datos; esto, si se tiene en cuenta la
explicación anterior, en cuanto a los posibles cuerpos lúteos encontrados al inicio del
protocolo de control de ciclo estral (día cero) y a la sincronización de crecimiento de la
onda lograda “día 5” (Bó et al., 2002; Bó, 2003). Adicionalmente la ovulación controlada
permitió que se generaran los cuerpos lúteos referentes al día 17 (muestreo 3), los
cuales difieren en su volumen (5236.1 ± 429.7) y que se refleja al final de la curva de
acuerdo a las diferentes concentraciones de P4 encontrada (> 1 ng/ml).
2.5.2.4. Porcentaje de preñez. En cuanto al porcentaje de preñez no se
encontraron diferencias (P>0.05) entre los dos tratamientos (eCG día 5 frente a eCG día
8), lo cual concuerda con el comportamiento encontrado entre las dos anteriores
variables analizadas (volumen luteal y concentración plasmática de P4). Por lo tanto la
mayor parte de la hipótesis planteada “El día de aplicación de la hormona Gonadotropina
Coriónica Equina (eCG) determina el desarrollo luteal, la concentración de progesterona
y la tasa de preñez en hembras receptoras de embriones bovinos” no recibió sustento
estadístico. Se podría resaltar que numéricamente el grupo de novillas pertenecientes al
tratamiento 1 (eCG día 5) obtuvieron un 5 %, más de preñez, esto frente a las novillas
correspondientes al tratamiento 2 (eCG día 8), pero válidamente no se podría concluir al
respecto.
Resultados contrarios en cuanto a las diferencias estadísticas han sido referenciados en
varios estudios. Es así que autores como Nasser et al., (2004), quienes utilizaron
receptoras cruzadas para embriones (Nelore/Angus - Bos Indicus/Bos Taurus),
sincronizadas con protocolo para Transferencia de Embrión a Tiempo Fijo (TETF), en el
cual varió el día de aplicación de la eCG para cada uno de los dos grupos de animales
utilizados (400 UI día 5 u 8), encontraron diferencias estadísticas en referencia a la

Capítulo 2 131
concentración de P4 plasmática, 2.69 ± 0.38 ng/ml (grupo eCG día 5) vs 1.63 ± 0.21
ng/ml (grupo eCG día 8); igualmente encontraron diferencias estadísticas en cuanto al
porcentaje de preñez (preñadas/transferidas), siendo más alto en el grupo eCG día 5
(63.4%) frente a eCG día 8 (36.1%) (Utilizando embriones in vitro, transferidos en fresco).
Se evidencia las menores concentraciones plasmáticas de P4 encontradas en el estudio
referenciado, en el cual posiblemente son producidas por cuerpos lúteos pequeños de
ovarios de menor tamaño característico a la tipología de las hembras utilizadas cruzadas
con Nelore (característica de los Bos indicus) frente a los de Bos Taurus utilizados en el
presente estudio (novillas Holstein).
Lo anterior no sería definitivo si se observa los resultados obtenidos por Machado et al.,
(2006) quienes utilizando vacas de la raza Nelore (Bos Indicus) en las cuales utilizaron
un protocolo para sincronización de la ovulación con eCG (400 UI) aplicada el día 9 (ciclo
sincronizado), obtuvieron 6927,49 ± 05,86 mm3 de volumen luteal y una concentración
plasmática de P4 de 8,15 ± 0,64 ng/ml., mayores que las encontradas en el presente
estudio, aclarando que la condición de las hembras de el citado artículo eran vacas y no
novillas, lo cual podría estar igualmente influenciando los resultados (Nogueira et al.,
2003; Vos et al.; 1994; Dieleman et al., 1993).
Mayores concentraciones plasmáticas de P4 que las encontradas en el presente reporte
se han observado en otros estudios como el realizado por Marques et al., (2003) quienes
buscando verificar los efectos de la eCG administrada en el momento de retiro del
implante con P4 en vacas Brangus, observaron diferencias estadísticas 8.6 ± 0.4 ng/ml en
las vacas con eCG, frente a las vacas sin eCG 6.4 ng/ml, confirmando prácticamente lo
evidenciado anteriormente por Baruselli et al., (2001), los cuales sincronizaron receptoras
cruzadas para embriones, a las cuales aplicaron 800 UI de eCG el día 5 con referencia al
ciclo sincronizado (crecimiento de nueva onda folicular), encontrando que con la
aplicación de la eCG se presentaban cuerpos lúteos de mayor tamaño (> 13 mm) para el
día del trasplante embrionario (84% de los animales) y con un porcentaje de preñez de
42% (con eCG) frente a 34% (sin eCG). Igualmente Sousa et al., (2009), quienes como
en el presente estudio utilizaron hembras de la raza Holstein, pero en condición de
vacas, sincronizadas para inseminación artificial, enciontraron porcentajes de preñez de
33.8% (eCG día 8 - 400 UI eCG) frente a 30.9%. (Sin eCG). (Menores a los obtenidos en
el presente estudio).

132 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
Estos resultados de alguna manera evidencian que al aplicar la eCG los efectos
encaminados hacia el desarrollo luteal podrían ser adecuados, consecuentemente la
síntesis de P4 se podría incrementar, colaborando de esta forma conl desarrollo
embrionario. De esta manera podría beneficiar el periodo de reconocimiento materno de
la preñez. Esto podría evidenciarse de alguna manera en el presente estudio si se
observan los porcentajes de preñez obtenidos en citados artículos (Dieleman et al., 1993;
Vos et al.; 1994; Baruselli et al., 2003; Nogueira et al., 2003; Naser et al., 2004; Bó y
Cutaia, 2005).
Podrían existir discrepancias a lo anteriormente expuesto de acuerdo a otros
investigadores: Es así que estudios como el realizado por Siqueira et al., (2009), no
encontraron diferencias significativas con vacas cruzadas (Bos Taurus x Bos Indicus), a
las cuales se les aplicó eCG el día 5 (con referencia al inicio del protocolo). En el citado
estudio la concentraciones de P4 encontradas para el día 17 (con respecto al ciclo
sincronizado) fueron 5.2 ± 5.0 ng/ml. y un área luteal de 72.4 ± 12.0 cm2 en vacas
preñadas, mientras que en las vacas vacias el área luteal fué 71.4 ± 11.3 cm2. Se nota en
este estudio referencial, que los valores de la concentración de P4 son muy similares al
presente trabajo, contrariamente a lo que posiblemente se esperaría, ya que en el
estudio referencial se usaron cruces (Bos Indicus x Bos Taurus), de lo cual se asumiría
que sus ovarios, estructuras y concentración de P4 fueran menores a las encontradas en
el presente trabajo “5.12 ± 0.31 ng/ml” ( (Naser et al., 2004; Bó y Cutaia, 2005), en donde
como se ha expuesto, se utilizaron hembras puras Bos Taurus (novillas Holstein),
entendiendo de igual forma que las estructuras en novillas podrían ser menores a las
encontradas en hembras en condición de vaca (Machado et al., 2006).
En vacas Bos Indicus, sin programas de transferencia embrionaria ni tratamientos
hormonales Aguirre et al., (2006), reportaron 45 % de preñez, porcentaje similar al
encontrado en otros diversos estudios como en el de Fuentes y De la Fuente (2007), en
el que utilizaron novillas Holsteín, las cuales si fueron tratadas hormonalmente (1000 UI
eCG el día 5 con referencia al inicio del tratamiento), los cuales reportan tasas de preñez
con embriones congelados en glicerol de 54%, en etilenglicol del 48% y frescos 52% (sin
diferencias estadísticas).

Capítulo 2 133
Igualmente Rodrigues et al., (2004) en vacas receptoras de embriones de la raza Nelore,
reporta porcentajes de preñez de 56% (con eCG) frente a 37.8% (sin eCG), los cuales
son mayores a lo reportado por Pita et al., (2009) con novillas cruce por Cebú, en donde
obtuvieron tasas de preñez de 44% con embriones congelados utilizando eCG día 5 del
protocolo y 30 % utilizando eCG el día 8 del protocolo. Estodatos son menores a lo
reportado por Peixoto et al., (2007), utilizando novillas receptoras Holstein y Cebú
cruzadas (Bos Indicus x Bos Taurus) en donde no utilizó eCG en el tratamiento de
sincronización, 63.7%, porcentaje similar al obtenido en el presente estudio como total.
Otros estudios han verificado el número de cuerpos lúteos con respecto a la tasa de
preñez, es así que autores como Siqueira et al., (2009), utilizando en receptoras
cruzadas (Bos Taurus Taurus x Bos Taurus Indicus) un tratamiento de sincronización
para transferencia de embrión a tiempo fijo (TETF) con embriones en fresco (eCG
aplicada el día 5 con referencia al inicio del protocolo), encontraron una tasa de preñez
de de 42.9 % (en receptoras que tuvieron cuerpo lúteo único) frente a 61.9 % (en
receptoras con múltiples cuerpos lúteos). El estudio referenciado podría concluir que a
mayor tejido luteal, mayor síntesis de P4 y consecuentemente mayor porcentaje de
preñez, sin embargo, este efecto no fue evidenciado en el presente estudio.
En cuanto al efecto de la P4 sobre la tasa de preñez se evidencia en otros estudios como
el de Filho et al., (2004), en el cual utilizó vacas Nelore para IATF, encontrando
diferencias estadísticas para la tasa de preñez, 63.2% (con eCG día 9) frente a 41.7 (sin
eCG). Estas diferencias significativas tambienfueron observadas por Silva et al., (2004),
utilizando vacas Nelore, 51.7% (con eCG) frente a 33.8% (sin eCG) y Baruselli et al.,
(2003) con vacas de la misma raza, 55.1% (con eCG día 8) frente a 38% (sin eCG).
Estos resultados, teniendo en cuenta la aplicación de la eCG el día 9, son superiores al
obtenido por Filho et al., (2010), 46.2% con tratamiento de eCG aplicada este mismo día
9.
De acuerdo a estos resultados referenciados de porcentajes de preñez, se puede
observar que son menores a los encontrados en el presente estudio (Vésase tabla 3). Ya
que los estudios referenciados son en programas de IA, en donde los porcentajes de
preñez usualmente son más altos que los porcentajes obtenidos en programas de
transferencia de embriones (Barret et al., 2006; Bó y Caccia, 2003).

134 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
También resultados controversiales se han evidenciado en estudios en donde se ha
tratado de sincronizar el inicio de crecimiento de las ondas foliculares con la aspiración
de los folículos ováricos. Vasconcelos et al., (2001) encontraron diferencias estadísticas
aspirando folículos reclutados > 4 mm en vacas lecheras, sincronizadas con Ovsynch en
la concentración de P4 para el día 7 (17 del presente estudio) 1.16 ± 0.12 ng/ml. y 2862 ±
228 mm3 de volumen luteal (sin aspirar), mientras que para el grupo aspirado fue de 1.42
± 0.11 ng/ml con 5363 ± 342 mm3 de volumen luteal. En dicho estudio solamente se
reportó diferencia significativas en cuanto al volumen luteal en el grupo aspirado son
similares a los resultados encontrados en la presente investigación.
También se han realizado trabajos con la finalidad de sincronizar el ciclo estral utilizando
PGF2α, esto con la finalidad de ejercer acción sobre el cuerpo lúteo. Es así que autores
como Lynch et al., (2010), utilizando hembras tipo carne (Bos Taurus) sincronizadas con
dos dosis de PF2α (11 días de intervalo), encontraron cuerpos lúteos con diámetros de
21.61 ± 4.51 mm. Así mismo Sartori et al., (2002), utilizando dos grupos de vacas
Holstein en las cuales aplicaron PGF2α el día 7 (en presencia de cuerpo lúteo) y en el
otro grupo no aplicó (no presencia de cuerpo lúteo), reportaron volúmenes luteales
máximos de 6178.7 ± 516.4 mm3 y 8542.3 ± 453.8 mm3 respectivamente. Igualmente la
concentración de P4 encontrada fue de 1.8 ± 0.2 ng/ml y 2.2 ± 0.2 ng/ml. Los volúmenes
encontrados en el estudio referenciado fueron mayores a los encontrados en el presente
trabajo, tanto en el tratamiento 1 (eCG día 5) como en el tratamiento 2 (eCG día 8), en
donde se utilizaron novillas, y no vacas como en el estudio referenciado. En tanto que la
concentración plasmática de P4 encontrada en el estudio en referencia fue mucho menor
a la encontrada en el presente estudio, en donde posiblemente se evidencia la acción de
la eCG causada al respecto para los dos tratamientos, logrando así mayor síntesis de P4
luteal; en otros estudios, en donde también han utilizado PGF2α, como el realizado por
Mann (2009), utilizando vacas multíparas Holstein han referenciado concentraciones de
P4 encontradas hacia el día 8 del ciclo estral (día 18 de nuestro estudio) de 5.82 ± 0.39
ng/ml, las cuales son muy similares a las referenciadas en el presente estudio en los dos
tratamientos utilizados.
En cuanto a estudios que reporten porcentaje de preñez en receptoras de embriones las
cuales han sido sincronizadas con tratamiento tradicional de dos prostaglandinas
separadas por 11 días y con implante de P4 por siete días (sin eCG), se ha evidenciado

Capítulo 2 135
en el caso de Spell et al., (2001), trabajando con hembras de la raza Angus porcentajes
de preñez de 83% con embriones frescos y 69% con embriones congelados, porcentajes
mayores a lo reportado en el presente estudio.
En cuanto a la condición de desarrollo de la hembra (novilla o vaca) se ha verificado por
varios autores que las novillas podrían estar produciendo más P4 que las vacas,
independientemente del tamaño luteal. Es así que Sartori et al., (2004), utilizando novillas
y vacas lactantes Holstein, encontraron volúmenes luteales de 7303 ± 308 mm3 en
novillas, y 11248 ± 776 en vacas con una concentración de P4 de 7.3 ± 0.4 ng/ml en
novillas y 5.8 ± 0.6 ng/ml en vacas, evidenciando diferencias significativas. Estos valores
son contradictorios, ya que se esperaría que a mayor volumen luteal más P4 sintetizada
(Baruselli et al., 2005; Madureira et al., 2004; Everton and Baruselli, 2003; Niswender et
al., 2000), lo cual no se evidencia si se analizan estos datos. Pero más contradictorio
resulta si se analizan los datos reportados por Chagas et al., (2002), quienes trabajaron
con receptoras novillas y vacas Holstein (transfiriendo con celo referencial - no
sincronizadas), encontraron concentraciones promedio de P4 de 3.0 ± 0.1 ng/ml en
novillas y 2.0 ± 0.1 ng/ml en vacas, con un volumen luteal de 5515.2 ± 273.4 mm3 y
7293.6 ± 304.8 mm3 respectivamente.en cuanto al porcentaje de preñez con embriones
congelados para las novillas fue de 52.3 %. En este caso los porcentajes de preñez
reprotados fueron inferiores al reportado en el presente estudio.
En Colombia, se han reportado resultados utilizando hembras receptoras de embriones
de diferente tipología racial y cruces. Rodríguez et al., (2007) en un estudio realizado en
el trópico medio y bajo colombiano reportan promedios generales de P4 de 3.6 ± 2.6
ng/ml., con diámetros luteales de 19.6 ± 1.8 mm. y tasas de preñez con embriones en
fresco de 31%, y con congelados de 46%. Los datos de estas tres varibles son menores
a lo encontrado en el presente estudio, lo cual podría deberse a las condiciones de los
embriones in vitro transferidos en alguna proporción en el estudio referencial entre otros,
y en menor grado a las tipologías raciales (Bos Indicus) trabajadas, ya que según lo
referenciado anteriormente, diferentes porcentajes de preñez se han obtenido utilizando
tipologías raciales Bos Indicus y Bos Taurus (Nogueira et al., 2003; Vos et al.; 1994;
Dieleman et al., 1993).

136 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
Con relación a lo anterior (variables planteadas en la tabla 3 (Volumen luteal “día 17”,
concentración de P4 “día 17” y tasa de preñez “día 52), se hace evidente que existe
variedad en cuanto a estudios al respecto. Los resultados en su gran mayoría son
contradictorios en lo referente a que a mayor volumen o área luteal mayor la
concentración de progesterona plasmática sintetizada, lo que a su vez podría conllevar a
mayores porcentajes de preñez (Moura et al., 2001; Marques et al., 2002). En los citados
estudios se han utilizado diversos protocolos, en algunos de los cuales se usó como
apoyo hormonal Gonadotropina Corionica Equina (eCG). En el presente estudio, cuando
la eCG se suministró el día cinco (5) se buscaba reclutamiento y selección folicular
(Barret et al., 2006), que permitiera la estimulación de más de un folículo, lo que
culminaría con ovulaciones múltiples y esto a su vez con mayor tejido luteal (Bó y Caccia,
2003), el cual sintetizaría mayor concentración de P4 (Baruselli et al., 2005), con lo que se
generaría un mejor ambiente embriotrófico (Revisado por Gonella et al., 2010) en el cual
el desarrollo embrionario hubiera sido adecuado, lo que favorecería el reconocimiento
materno de la preñez. Por otra parte, el tratamiento con eCG aplicada el día 8, cuando ya
el dominio folicular está establecido (Evans et al., 1997), se pretendía una mayor
estimulación del folículo dominante (en algunos casos el mayor folículo subordinado)
(Revisado por Olivera et al., 2006), generar mayor tejido luteal, mayor concentración de
P4 plasmática y mayor porcentaje de preñez (Bó y Caccia, 2003). Según esto, de
acuerdo a los porcentajes de preñeces conseguidos tanto con el tratamiento 1 (eCG día
5) y tratamiento 2 (eCG día 8), se podría decir que fueron beneficiosos, si tenemos en
cuenta los porcentajes de preñez reportados en los múltiples estudios citados. Este
resultado pudo deberse al aumento de las concentraciones plasmáticas de progesterona
causada por la acción eCG (Cameron and Batt, 1991; Berisha and Schams, 2005), lo
cual pudo haber estimulado el crecimiento embrionario y optimizar la secreción de
interferón tau y por tanto, el reconocimiento materno de la gestación se dio en los
momentos oportunos (Mann et al., 1999). Este resultado pudo deberse a la acción
luteotrópica causada al aplicar las 400 UI de eCG (Baruselli et al., 2005; Kuran et al.,
1996) a los dos grupos de novillas de los grupos experimentales.
Al no encontrar diferencias (P>0.05) entre tratamientos (eCG día 5 Vs eCG día 8) en la
presente investigación, en cuanto a las variables analizadas en la tabla 3 (Volumen luteal
“día 17”, concentración de P4 “día 17” y tasa de preñez “día 52), se podría especular
sobre los posibles factores determinates. Dichos factores podrían jugar un importante

Capítulo 2 137
papel en el desempeño de los taratamientos aplicados en los diferentes estudios citados.
Estos podrían ser factores dietarios entre otros, los cuales indudablemente afectan el
desempeño fisiológico reproductivo (Rekwolt, 2004; Roche, 2006). Se debe tener en
cuenta el factor raza, en donde existen diferencias en cuanto a sensibilidad hormonal
(Sartori et al., 2004); Se podría deducir que concentraciones de 400 UI de eCG van a
desencadenar efecto diferente en las razas de las dos especies (Indicas, Taurinas y sus
cruces) (Nogueira et al., 2003; Vos et al.; 1994; Dieleman et al., 1993).
2.5.2.4.1. Concentraciones plasmáticas de progesterona durante los días 5, 9 y 17 según estado reproductivo (Preñadas y no preñadas) En la gráfica 3 se evidencia el comportamiento general promedio de la P4 en la totalidad
de las novillas utilizadas en el estudio durante los tres muestreos realizados (día 5; día 9
y día 17). Se evidencia este comportamiento según el estado reproductivo de las novillas
(preñada o no preñada), en donde notablemente se observa que las mayores
concentraciones durante el tercer muestreo (día 17) están en las novillas que se lograron
preñar, las cuales presentaron en promedio 40.6% (1.585 ng/ml) mas de P4 plasmática.
Grafica 3. Comportamiento general de la progesterona según estado reproductivo
5.025
0.278
5.117
5.591
0.312
5.490
4.076
0.275
3.905
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0
5.5
6.0
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
PRO
GES
TERO
NA
(ng/
ml)
DÍAS DE MUESTREO
Total novillas
Novillas preñadas
Novillas no preñadas

138 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
Con respecto a estas concentraciones, Siqueira et al., (2009), utilizando un tratamiento
de sincronización para Transferencia de Embrión a Tiempo Fijo (TETF), en donde
utilizaron eCG aplicada el día 5 (con referencia al inicio del protocolo), en receptoras
cruzadas (Bos taurus taurus x Bos taurus indicus), encontraron para el día de la
transferencia (día 17 con respecto al ciclo sincronizado) una concentración de P4 de 5.2 ±
5.0 ng/ml. en las novillas preñadas frente a 3.8 ± 2.4 ng/ml en las novillas no preñadas,
(con diferencias estadísticas), concentraciones plasmáticas similares a las encontradas
en el presente estudio.
En otros estudios en donde no se ha utilizado eCG, como en el realizado por Chagas et
al., (2002), quienes trabajaron con receptoras vacas y novillas Holstein, a las cuales
trasfirieron embriones utilizando celo referencial, encontraron concentraciones de P4 el
día cero (día de máximo desarrollo folicular) para las preñadas de 0.22 ± 0.01 ng/ml y
para las no preñadas 0.21 ± 0.02 ng/ml, valores similares a los reportados en el presente
estudio; estos autores encontraron para el día de la transferencia (día 17 del presente
estudio) 2.92 ± 0.08 ng/ml para las receptoras preñadas frente 2.88 ± 0.08 ng/ml para las
no preñadas. Estos valores numéricamente son menores a los reportados en el presente
estudio, en donde probablemente la eCG podría estar favoreciendo el proceso de
luteinización e interviniendo en la síntesis y liberación de P4 (Murphy, 2000; Niswender,
2000).
Spell et al., (2001), sincronizando receptoras Angus con tratamiento tradicional de dos
prostaglandinas separadas por 11 días y con implante de P4 por siete días (sin eCG)
encontraron una concentración de P4 de 4.1 ng/ml para las preñadas y 3.9 ng/ml para las
no preñadas, estudio en donde no se reportaron diferencias estadísticas. Las
concentraciones de P4 para las no preñadas fueron similares a las encontradas en el
presente estudio, pero superiores para las novillas preñadas.
En otro estudio realizado por Lopes et al., 2007, en donde utilizaron vacas Holstein
sincronizadas con Ovsynch encontraron concentraciones de P4 el día de la inseminación
artificial en las vacas preñadas de 0.19 ± 0.01 ng/ml. y para las no preñadas 0.24 ± 0.02
ng/ml, sugiriendo que esta hormona inclusive a estas bajas concentraciones podría estar
afectando la fertilidad. Con respecto a esto, Moreira et al., (2000), sugiere que se
presentan bajas tasas de preñez para vacas con insuficienciente regresión del cuerpo

Capítulo 2 139
lúteo después de una inyección de prostaglandina F2α, ya que las concentraciones de P4
podrían causar maduración incompleta del folículo preovulatorio, debido a los bajos
niveles de LH circulantes (alteración en el modelo de liberación), comprometiendo de
esta forma la ovulación folicular, alteración en la sensibilización de las células foliculares
que cambian a luteales, alterando de esta manera la liberación del ovocito. Igualmente se
podría presentar alteración en la secreción de P4, modificando el medio ambiente uterino
en donde el desarrollo embrionario podría no ser el más adecuado y por lo cual la
concentración de interferon Tau podría no ser la apropiada para que cause la señal
eficiente de reconocimiento materno de la preñez.
En el presente estudio, posiblemente la prostaglandina aplicada el día 5 (con respecto al
ciclo sincronizado) tuvo efecto en la totalidad de las novillas (tratamiento 1 eCG día 5 y
tratamiento 2 eCG día 8), ya que no hubo evidencia de estructuras lúteas el día que se
realizó el muestreo 2 (día 9 – máximo desarrollo folicular), por lo tanto se podría asumir
que el pico preovulatorio de LH se presentó, claro está, que si tenemos en cuenta que se
evidenció concentración medible de P4 para ese día (día 9), podría asumirse que esta
provino del proceso de la luteinización durante la etapa de dominancia del folículo
ovulatorio (Revisado por Olivera et al., 2006), o por residulaidad del implante de P4
utilizado (Bó et al., 2006), entre otras. Por lo tanto se pensaría que la P4 pudo haber
afectado directamente la síntesis y liberación de LH (Hafez, 2000), efecto que
posiblemente se evidencia al analizar la grafica 4.
Se observa que las concentraciones plasmáticas de P4 el día de la trnasferencia
embrionaria (día 17) son en algunos casos superiores (numéricamente) en las hembras
que se lograron preñar frente a las que no lo fueron. Referente a esto, Duica (2011) ha
evidenciado este posible efecto en novillas Holstein receptoras, a las cuales aplicó eCG
el día 5 (referenciado de acuerdo al inicio del tratamiento de sincronización), encontrando
diferencia (P<0,05) en hembras gestantes y no gestantes, lo cual podría dar a entender
de alguna forma en el aporte adecuado en el medio ambiente uterino brindado por la P4 a
los embriones transferidos, los cuales gracias a su desarrollo son eficientes en cuanto a
la síntesis y secreción de Interferon Tau (Revisado por Gonella, 2010), con lo cual el
reconocimiento materno de la preñez se estaría dando de una manera adecuada
(Niswender et al., 2000; Vasconcelos et al, 2001).
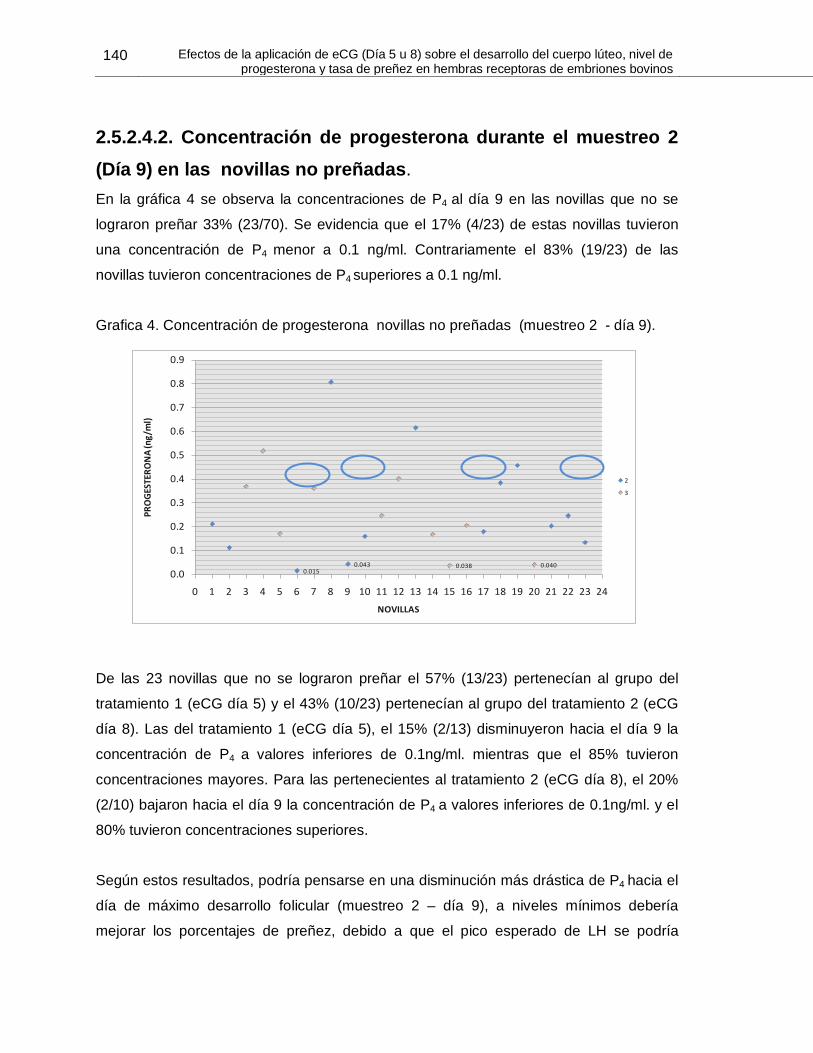
140 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
2.5.2.4.2. Concentración de progesterona durante el muestreo 2 (Día 9) en las novillas no preñadas. En la gráfica 4 se observa la concentraciones de P4 al día 9 en las novillas que no se
lograron preñar 33% (23/70). Se evidencia que el 17% (4/23) de estas novillas tuvieron
una concentración de P4 menor a 0.1 ng/ml. Contrariamente el 83% (19/23) de las
novillas tuvieron concentraciones de P4 superiores a 0.1 ng/ml.
Grafica 4. Concentración de progesterona novillas no preñadas (muestreo 2 - día 9).
De las 23 novillas que no se lograron preñar el 57% (13/23) pertenecían al grupo del
tratamiento 1 (eCG día 5) y el 43% (10/23) pertenecían al grupo del tratamiento 2 (eCG
día 8). Las del tratamiento 1 (eCG día 5), el 15% (2/13) disminuyeron hacia el día 9 la
concentración de P4 a valores inferiores de 0.1ng/ml. mientras que el 85% tuvieron
concentraciones mayores. Para las pertenecientes al tratamiento 2 (eCG día 8), el 20%
(2/10) bajaron hacia el día 9 la concentración de P4 a valores inferiores de 0.1ng/ml. y el
80% tuvieron concentraciones superiores.
Según estos resultados, podría pensarse en una disminución más drástica de P4 hacia el
día de máximo desarrollo folicular (muestreo 2 – día 9), a niveles mínimos debería
mejorar los porcentajes de preñez, debido a que el pico esperado de LH se podría
0.0150.043 0.038 0.040
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
PRO
GES
TERO
NA
(ng/
ml)
NOVILLAS
2
3

Capítulo 2 141
presentar sin dificultad con la consecuente luteinización y ovulación adecuada (Lucy et
al., 1992). De allí se plantea la importancia en cuanto a los niveles suprabasales de P4 en
las vacas, ya que al no presentarse la ovulación a tiempo del folículo dominante se
presenta envejecimiento del ovocito, el cual posiblemente no llegue a ser fertilizado.
adicionalmente se podría afectar la luteinización del folículo ovulatorio y con ello la
estructuración o programación de la vida del cuerpo lúteo y la síntesis de P4. Finalmente a
nivel uterino, este podría también perjudicarse debido a la evidencia de desbalance entre
las hormonas esteroides (P4 y Estradiol), entre otros (Darwash and Lamming 1998).
2.5.2.4.3. Efecto de los tratamientos (eCG día 5 u 8), diámetro folicular (día 9), volumen luteal (día 17) y concentración plasmática de progesterona (día 17) sobre la presentación de preñez (día 52). De acuerdo al análisis del efecto del tratamiento (eCG día 5 y eCG día 8), diámetro
folicular (folículo dominante - día 9), volumen total luteal (día 17) y concentración de
progesterona (día 17) sobre el diagnóstico de preñez, en la función logística utilizada,
solamente la variable concentración de progesterona plasmática del día 17 (tercer
muestreo) tuvo efecto significativo (P<0.1) sobre la probabilidad de que la preñez se
presentara.
En la gráfica 5 se muestra la función logística para el rango de valores de concentración
de la progesterona plasmática en cuanto a la presentación de la preñez, variable para la
cual se trabajó con un nivel de significancia de 0.1, debido a que la preñez es una
característica que podría estar influenciada por innumerables factores, los cuales sería
casi imposible de controlar.
La función logística estimada que describe la probabilidad de que el diagnostico de
preñez sea positivo en función de la concentración de progesterona para el día 17 es la
siguiente:

142 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
Grafica 5. Función logística para el rango de valores de concentración de progesterona.
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
1 2 3 4 5 6 7 8 9 10
P (D
P =
1)
Concentración de Progesterona (ng/ml.)
De acuerdo a la función logistica se podría deducir que el incremento en la probabilidad
de preñez se presenta hasta el máximo valor de concentración de P4 observado en el
estudio (10,1 ng/ml). Se debe aclarar que este incremento en la probabilidad de preñez
podría aumentar hasta el nivel máximo encontrado de P4 (10,1 ng/ml), para la totalidad
de animales utilizados en el estudio (Tratamiento 1 – eCG día 5 y Tratamiento 2 eCG día
8), en el rango de valores reportados; sin embrago, no siempre se podrá esperar que un
incremento en el valor de P4 incremente la probabilidad de preñez (Melanie et al., 2004).
esto podría llegar a ser independiente de la afirmación que indica que los niveles de P4
en sangre y el medio ambiente uterino durante la fase luteal favorecen el establecimiento
y mantenimiento de la gestación (Schams, 2004).
Contrariamente a lo encontrado en el presente estudio, en otros no se ha demostrado el
efecto significativo de la variable P4 sobre la preñez. Siqueira et al., (2009), utilizaron un
tratamiento de sincronización para transplante de embrión a tiempo fijo (embriones In
vivo e In vitro), utilizando eCG aplicada el día 5 (con referencia al inicio del protocolo), en
receptoras cruzadas (Bos taurus taurus x Bos taurus indicus), en donde aplicaron un
modelo de regresión logística, se evidenció que la única variable que significativamente
(P = 0,0002) afectaba la preñez era el tipo de embrión producido (In vivo e In vitro), pero
no la concentración de P4, ecotextura y área del cuerpo lúteo, categoría de los animales,
calidad del embrión y etapa de desarrollo embrionario.

Capítulo 2 143
Rodríguez et al., (2007), en un estudio en donde utilizaron varios tipos raciales y cruces
de receptoras, utilizando embriones transferidos en fresco y congelados, evaluaron las
variables independientes: diámetro del cuerpo lúteo, concentración de P4, estadio
embrionario, calidad embrionaria y la raza sobre el diagnostico de preñez, y no
encontraron dependencia de alguna de estas variables sobre el diagnostico de preñez.
Otros estudios han tratado de relacionar otras variables con la preñez. Peixoto et al.,
(2007), en el cual utilizaron receptoras de embriones novillas Holstein y Cebú cruzadas
(Bos Indicus x Bos Taurus) para determinar que variables explicativas (años de la
transferencia 1992-1999, temporada - otoño, invierno y primavera, raza del embrión, fase
del embrión, calidad del embrión y sincronía entre donantes y receptores) influían
directamente sobre la preñez, encontrando que el mejor modelo logístico que explicó la
preñez incluyó los efectos del año, época de la transferencia, etapa embrionaria, calidad
y sincronía del estro entre donante y receptora.
En tal sentido y analizando otras variables explicativas, Perry et al., (2007), quienes
trabajaron con novillas mestizas sincronizadas bajo protocolo CO-Synch para
inseminación artificial, encontraron que el tamaño folicular preovulatorio (≥12,8 mm)
predecía la preñez en 68,0 ± 4,9%, preñez que disminuía a medida que el folículo
disminuye de tamaño. Dicha disminución no se observó en el presente estudio.
La tabla 4 muestra algunas comparaciones de perfiles de P4 (ng/ml) del tercer muestreo
(día 17) mediante la razón de probabilidad de preñez (razón de los Odds).
Tabla 4. Contrastes entre perfiles de progesterona - Razón de probabilidad de preñez
Perfil 1 Perfil 26 4 2.72
8 4 7.4
Probabilidad de preñezPerfiles de concentración de
Progesterona plasmática (ng/ml)
4 2 2.72
2 4.485
De acuerdo a los resultados mostrados en la tabla, se analiza que al comparar una
novilla con una concentración de P4 de 6 ng/ml (perfil 1), con una novilla con una

144 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación
concentración de P4 de 4 ng/ml (perfil 2), se encuentra un valor de la razón de
probabilidad (Odds) de 2.72, lo cual indica que es 2.72 veces más probable que la vaca
con el perfil 1 quede preñada, comparada con la vaca del perfil 2, mostrando así un
aumento en la probabilidad de preñez con el aumento en la concentración de P4.

3. Conclusiones y recomendaciones 3.1 Conclusiones - No Existió efecto en el día de aplicación (día 5 o día 8) de la Gonadotropina Corionica
Equina (eCG) sobre el número de folículos dominantes presentes. Sin embargo, la eCG
si influye en el diámetro folicular (mm3) cuando es aplicada el día 5 con referencia al
inicio de un protocolo para control de ciclo estral en novillas de la raza Holstein.
- De acuerdo al día de aplicación de la Gonadotropina Corionica Equina (día 5 o día 8),
no existió efecto sobre el desarrollo luteal (volumen mm3 y número de cuerpos luteos),
medido para el día del día 17 dentro de un protocolo para control de ciclo estral en
novillas de la raza Holstein.
- No se evidencio efecto de acuerdo al día de aplicación (5 u 8) de la Gonadotropina
Coriónica Equina (eCG) sobre la concentración de progesterona (P4) plasmática ng/ml
medida para los días, nueve (9) y diecisiete (17) dentro de un protocolo para control de
ciclo estral en novillas de la raza Holstein.
- El día de la aplicación de la Gonadotropina Corionica Equina (eCG día 5 o eCG día 8)
dentro de un protocolo para control de ciclo estral no influye sobre el porcentaje de
preñez evidenciado para el día 52 en novillas de la raza Holstein receptoras para
embriones.
- Independientemente de la aplicación de la Gonadotropina Corionica Equina (eCG)
dentro del protocolo para control de ciclo estral en novillas de la raza Holstein, no se
evidenció relación entre el diámetro (mm) del folículo dominante (día 9) y el volumen
(mm3) luteal (día 17). Contrariamente este volumen luteal (mm3), determinado para el día
17, sí tiene relación sobre los niveles plasmáticos de progesterona encontrados para ese
mismo día (17), es decir, que por cada mm3 cúbico que aumenta el volumen luteal, se
aumenta la secreción (ng/ml.) de progesterona plasmática.

146 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
Título de la tesis o trabajo de investigación Título de la tesis o trabajo de investigación
- Los tratamientos con eCG (día 5 o día 8), el diámetro del folículo dominante (día 9) y el
volumen luteal (día 17) no afecta la preñez; solamente existe relación en cuanto a la
concentración plasmática de progesterona (ng/ml) evaluada el día 17 sobre la
determinación de la preñez, de tal manera, que al aumentar los niveles de progesterona
plasmática aumenta la probabilidad de preñez, dentro del rango de valores encontrados en
el estudio (1,98 a 10,1 ng/ml.).
3.2 Recomendaciones Es necesario diseñar estudios que permitan comparar las diferentes hormonas utilizadas
en los programas para el control del ciclo estral en la hembra receptora de embriones
bovinos, especialmente en lo referente a la Gonadotropina Corionica Equina (eCG), en
cuanto a su dosificación en los diferentes genotipos raciales y cruces, estados fisiológicos
(novilla, vaca de primer o varios partos), en diferentes latitudes, sistemas de manejo
(pastoreo, suplementación etc). Es importante comparar esta hormona con grupos
controles, lo cual permitiría realizar ajustes con la finalidad de incrementar los porcentajes
de preñez y a su vez la eficiencia reproductiva en los programas de transferencia
embrionaria en bovinos.
Sería pertinente realizar investigaciones a nivel molecular, en las que se profundice acerca
de los factores determinantes celulares que involucran el desarrollo de las estructuras
luteales a nivel ovárico, esto cuando se aplican terapias hormonales con la finalidad de
favorecer el proceso luteotrópico, que generen niveles suficientes de progesterona, para el
establecimiento de un medio ambiente uterino que favorezca el desarrollo embrionario así
como el mantenimiento de la preñez.
Es importante realizar investigaciones que permitieran conocer más acerca de los efectos
de los niveles suprabasales y la residualidad de la progesterona durante la fase folicular en
las hembras bovinas a las cuales se les va a trasplantar un embrión, más aún a las que se
controla el ciclo estral con diferentes tratamientos de sincronización, y aún más pertinente
a las cuales se les suministra apoyo hormonal como la Gonadotropina Coriónica Equina
(eCG), ya que este fenómeno podría ser un punto crítico que afecta la eficiencia
reproductiva en la hembra receptora de embriones.

Bibliografía 1. Adams GP, Matteri RL, Kastelic JP, Ko JCH And Ginther. Association between surges
of follicle – simulating hormone and the emergence of follicular waves in J. Reprod.
Fert. 1992. 94: 177-188.
2. Aguirre G, Pardo C, Góngora A. Inicio del celo, tasa de gestación y relación del tiempo
de inseminación con los niveles de progesterona en vacas brahmán. Rev. MVZ
Córdoba 2006. 11 (1): 766 – 772.
3. Ali, A. K. A., Shook, G. E..An optimun transformation of somatic cell concentration in
milk production. J. Dairy Sci. 1980. 63:487
4. Badinga L, Thatcher WW, Wilcox CJ, Morris G, Entwistle K, Wolfenson D. Effect of
season on follicular dynamics and plasma concentrations of estradiol-17beta,
progesterone and luteinizing hormone in lactating holstein cows. Theriogenology 1994.
42:1263- 1274.
5. Baez G. Relaciones hormonales y dinámica folicular durante el periodo posparto en
vacas sanmartinero. Tesis de Maestría. Facultad de Medicina Veterinaria y de
Zootecnia. Universidad Nacional de Colombia – Sede Bogotá D.C. 2010.
6. Bao B, Garverick AH, Smit GW, Smith MF, Salfen BE, Youngquist RS. Changes in
messenger ribonucleic and acid encoding luteinizing hormone receptor, citochrome
P50 side chain cleavage, and aromatase are associated with recruitment and selection
of bovine ovaian follicles. Biol Reprod 1997. 56 (5): 1158–1168.
7. Baruselli P, Bó G, Reis E, Marques M, Sá M. Introducción de la IATF en el manejo
reproductivo de rebaños de ganado de engorde en Brasil. Congreso Internacional de
Reproducción Bovina. Bogotá 2005.
8. Baruselli P, Marques O, Nasser LF, Reis EL and Bó G. Effect of eCG on pregnancy
rates of lactating zebu beef cows treated with CIDR–B devices for timed artificial
insemination. Theriogenology. 2003. 59: 214.
9. Baruselli P, Marques MO, Hothnann EM, Costa Neto WP, Grandinetti RR, B6 G.
Increased pregnancy rates in embryo recipients treated with CIDR-B devices.
Theriogenology. 2001. 55:157 (abstr).
10. Barrett D.W, Bartlewski PM, Duggaathi R, Davies K.L, Rawlings NC. Suppression
of follicle wave emergence in cycle ewes by Supraphysiologic concentrations of

148 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
estradiol 17 beta and induction with a physiologic dose of exogenous ovine follicle
stimulating hormone. Biol Reprod. 2006. 75: 633-641.
11. Barnea E, Choi Y, Leavis P. Embryo maternal signaling prior to implantation. En:
textbook of obstetrics ginecology I. Munteanu. Ed. SIEP. Boston 2000.
12. Battocchio M, Gabai G, Mollo A, Veronesi MC, Soldano F, Bono G, Cairoli F.
Agreement between ultrasonographic classification of the CL and the plasma
progesterone concentration in dairy cows. Theriogenology 1999. 51:1059-1069.
13. Battye K, Parkinson T, Lenner L, Evans G, O´Neill C, lamming G.
Potentialsynergism between platelet-activating factor and 1 – recombinant interferon in
promoting luteal maintenance in cyclic ewes. Reprod Fer 1993. 97: 21.
14. Berisha B and Schams D. Ovarian function in ruminants. Domestic Animal
Endocrinology 2005. 29: 305-317.
15. Binelli M, Thatcher W, Mattos R, Baruselli P. Antiluteolytic Strategies to Improve
Fertility in Cattle. Theriogenology 2001. 56: 1451–1463.
16. Bó G, Colazo M, Martínez MF, Kastelic JP. Mapletoft. Sincronizacion de la
emergencia de la onda folicular y la ovulación en animales tratados con
progestagenos y diferentes esteres de estradiol.2o simpósio internacional de
reprodução animal aplicada. 2006.
17. Bó GA, Cutaia L. Implementación de programas de inseminación artificial en
rodeos de cría. Argentina. Memorias del VI Simposio Internacional de Reproducción
Animal. Instituto de Reproducción Animal de Córdoba Argentina. 2005. 326-332.
18. Bó GA, Alonso A, Carcedo J, Caccia M. Modulo I, Actualización sobre fisiología de
la reproducción de la vaca. Especialización en reproducción bovina. IRAC, CGR,
Universidad de Cordoba Argentina. 2004.
19. Bó GA. Sincronización de celos para programas de Inseminación Artificial y
Transferencia de embriones Bovinos. Instituto de Reproducción Animal de Córdoba
Argentina (IRAC) 2003.
20. Bó GA. y Caccia, M. Ultrasonografía reproductiva en el ganado bovino. Facultad
de Ciencias Agropecuarias. Universidad Católica de Córdoba Argentina. Instituto de
Reproducción Animal de Córdoba (IRAC). 2003.
21. Bó GA, Baruselli PS, Moreno D, Cutaia L, Caccia M, Tribulo H. The control of
follicular wave development for self – pointed embryo transfer programs in cattle.
Theriogenology. 2002. 57: 53–72.

Bibliografía 149
22. Bousfield GR; Butnev VY.Identificación of twelve O-glycosylation sites in equine
chorionic gonadtropin beta and equine luteinizing hormone ss by solid-phase Edman
degradation. Biol Reprod 2001, 64: 136-147.
23. Cameron AWN, Batt PA. PMSG may directly stimulate ovulation in female goats.
Anim Reprod Sci 1991; 25:233–9.
24. Chagas J, Lopes L, Robalo J. Plasma progesterone profiles and factors affecting
embryo-fetal mortality following embryo transfer in dairy cattle. Theriogenology. 2002.
58: 51- 59.
25. Combarnous Y. Molecular basis of the specificity of binding of glycoprotein
hormones to their receptors. Endocr Rev 1992. 13: 670–691.
26. Drawash, A., Lamming.The importance of milk progesterone concentrations during
early pregnancy in the cow. Journal of Animal Breding. 1998, 2: 41-43.
27. Dieleman SJ, Bevers MM, Vos PLAM, de Loos FAM. PMSG/anti-PMSG in cattle: a
simple and efficientsuperovulatory treatment. Theriogenology 1993;39:25–41.
28. Duica A. 2010. Efecto del diámetro del folículo ovulatorio, tamaño del cuerpo
lúteo y perfiles de progesterona sobre la tasa de preñez en la hembra receptora de
embriones bovinos. Tesis Maestría en Salud Animal. Facultad de Medicina Veterinaria
y de Zootecnia. Universidad Nacional de Colombia, Sede Bogotá D.C.
29. Edmonson AJ, Lean IJ, Weaver LD, Farver T, Webster GA. Body condition chart
for Holstein dairy cows. J Dairy Sci 1989. 72: 68–78.
30. España F, Pérez C, Rodríguez I, Dorado J, Hidalgo M. estudio comparativo de la
eficiencia del diagnostico precoz de gestación en vacuno mediante ecografía luteal y
progesterona plasmática. Revista Científica 2004, 14: 1-12.
31. Evans ACO and Fortune JE. Selection of the dominant follicle in cattle occurs in
the absence of differences in the expression of messenger ribonucleic acid for
gonadotropin receptors. Endocrinology 1997. 138: 2963-2971.
32. Evans AC, Komar CM, Wandji SA. Fortune JE. Changes in androgen secretion
and luteinizing hormone pulse amplitude are associated with the recruitment and
growth of ovarian follicles during the luteal phase of the bovine estrous cycle. Biol
reprod 1997. 57 (2): 394–401.
33. Everton L, Baruselli P. Sincronización de receptoras cruce por cebú en
condiciones tropicales. Cuarto seminario de reproducción de grandes animales
(memorias) Bogotá, 2003.

150 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
34. Filho MF, MO, Reis EL. Viel JO, Nichi M, Baruselli P. Dinámica follicular de vacas
Nelore lactantes em anestro tratadas com progestágeno, eCG e GnRH. Acta Scientiae
Veterinariae 32 (suplemento). 235, 2004
35. Filho MF, Ayres H, Ferreira RM, Marques MO, Reis EL. Silva RCP, Rodrigues CA,
Madureira EH, Bo´ GA, Baruselli P. Equine chorionic gonadotropin and gonadotropin-
releasing hormone enhance fertility in a norgestomet-based, timed artificial
insemination protocol in suckled Nelore (Bos indicus) cows. Theriogenology 2010. 73:
651–658
36. Fuentes S, De la Fuente J. Tasas de gestación de novillas receptoras
sincronizadas con Gonadotropina Coriónica Equina u Hormona Folículo Estimulante.
Acta Scientiae Veterinariae 2007. 35 (Supl. 3): s 767 – s 772.
37. Fuentes S, De la Fuente J. Different synchronization treatments for direct embryo
transfer to recipients heifers, In: Proceedings of the XIII Annual Meeting AETE, Lyon,
France; 1997. p. 148.
38. Gonzales-Stagnaro, C. Reproducción bovina. Venezuela. Ed. Fundación Girarz.
2001.
39. Gonzales -Stagnaro C, Madrid-Bury N, Morales J, Marín D. Efecto luteoprotector
del tratamiento GnRH en vacas mestizas repetidoras con cuerpo lúteo sub-funcional.
Rev. Científ. FCV-LUZ. 1993 1:14-20.
40. Gordon, I. Reproducción controlada del ganado vacuno y búfalos. Zaragoza
España. Ed. Acribia. 1999.
41. Grajales H. Comportamiento reproductiva del Ganado bovino en el trópico cálido
húmedo colombiano; pubertad, ciclo estral, niveles de hormonas esteroides y su
relación con la tasa de concepción y fertilidad. Tesis de Ph.D. Facultad de Medicina
veterinaria y de Zootecnia. Universidad nacional de Colombia 2000.
42. Grygar I, Kudlac E, Dolezel R, Nedbalkova J. Volume of luteal tissue and
concentration of serum progesterone in cows bearing homogeneous corpus luteum or
corpus luteum with cavity. Animal Reproduction Science. 1997. 49: 77-82.
43. Gonella, A., Posible relación de los niveles séricos ováricos (17 Beta estradiol y
Progesterona), sus respectivos receptores endometriales (ER-1 alfa y PGR) y
loscambios histológicos del endometrio durante el ciclo estral en hembras cíclicas de
tres razas bovinas. Tesis de Maestría. Facultad de Medicina Veterinaria y de
Zootecnia. Universidad Nacional de Colombia – Sede Bogotá D.C. 2010.

Bibliografía 151
44. Gonella, A., Grajales, H., Hernández, A. 2010. Ambiente receptivo uterino: control
materno, control embrionario, muerte embrionaria. Rev. MVZ. Córdoba. 15, 1: 1976-
1984.
45. Hafez E. Reproducción e inseminación de los animales domésticos, 7ª ed.
Reproductive Health Center IVF/Andrology International. Kiawah Island, South
Carolina, USA. McGraw – Hill Interamericana. 2000. 33–144.
46. Hernández L y Zarco A. Función del cuerpo lúteo y muerte embrionaria en
rumiantes. Ciencia veterinaria. 1998. 8: 1-28.
47. Howell JL, Fuquay JW, Smith AE. Corpus luteum growth and function in lactating
Holstein cows during spring and summer. J. Dai. Sci. 1994. 77:735-739.
48. IETS (Manual de la Sociedad Internacional de Transferencia de Embriones):
Tercera edición. Editores Stringfellow D, Scidel, S. Junio de 2000.
49. Jiménez C. y Hernández A. Lecturas sobre reproducción bovina. II. El ciclo estral
de la vaca. Facultad de Medicina Veterinaria y de Zootecnia. Universidad Nacional de
Colombia. 2000.
50. Kastelic P, Bergfelt DR, Ginther OJ. Relationship between ultrasonic assessment
of the corpus luteum and plasma progesterone concentration in heifers.
Theriogenology. 1990. 33, 6: 1269 – 1278.
51. Kastelic J, Pierson R and Ginther O. Ultrasonic Morphology of a corpora lutea and
central luteal cavities during the estrous cycle and early pregnancy in heifers.
Theriogenology. 1990. 34, 3: 487-498.
52. Kerbler TL, Buhr MM, Jordan LT, Leslie KE, Walton JS. Relationship between
maternal plasma progesterone concentration and interferon-tau synthesis by the
conceptus in cattle. Theriogenology 1997. 47: 703-714.
53. Knobil E and Neill J. Physiology of reproduction. Third edition. Academic Press is
an imprint of El sevier. 2006.
54. Krisans SK. Cell compartmentalization of cholesterol byosynthesis. Annals of the New
York academy of Science 1996. 804: 142–164.
54. Kuran M, Hutchinson JSM, Broadbent PJ. The response of bovine granulosa cells
to different gonadotrophins in culture. Anim Reprod Sci 1996; 45:1–12.
55. Lopes AS, Butler ST, Gilbert RO, Butler WR. Relationship of pre-ovulatory follicle
size, estradiol concentrations and season to pregnancy outcome in dairy cows. Anim
Reprod Sci 2007. 99: 34-43.

152 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
56. Lopez, F.Relación entre condición corporal y eficiencia reproductiva en vacas
Holstein. Revista Facultad de Ciencias Agrarias 2006. V4, No 1, 77 - 86.
57. Lucy MC, Savio JD, Badinga L., De La Sota RL. And Thatcher WW. Factors that
affect ovarian follicular dynamics in cattle. J. Anim. Sci. 1992. 70: 3615–3626.
58. Lynch CO, Kenny DA, Childs S and Diskin MG. The relationship between
periovulatory endocrine and follicular activity on corpus luteum size, function, and
subsequent embryo survival. Theriogenology. 2010. 73: 190 – 198.
59. Nasser L, Reis E, Oliveira M, Bó GA, Baruselli P. The use of hormonal treatments
to improve reproductive performance of anestrous beef cattle in tropical climates. Anim
Reprod Sci 2004. 82, 83: 479–486.
60. Nieder GL and Corder CN. Effects of opiate antagonists on early pregnancy and
pseudopregnancy in mice. Journal of Reproduction and Fertility. 1982, 65 341-346.
61. Niswender GD, Juengel JL, Silva PJ, Rollyson MK and McIntush EW. Mechanisms
controlling the function and life span of the corpus luteum. Physiological.
Reviews.2000. 222. 80: 1-29.
62. Nogueira, M., Melo, D., Carvalho, L., Fuck, E., Trinca, L. and Moraes, C. Do
progesterone concentrations decrease pregnancy rates in embryo recipients
synchronized with PGF2 and eCG? Theriogenology. 2003. 61. 7-8: 1283-1290.
63. Machado R, Taveira R, Aurelio M, Bergamaschi CM, Alvarenga C and Binelli M.
Sincronização da ovulação em vacas da raça Nelore e seus efeitos na função
ovariana Boletim de pesquisa e desenvolvimento 7. Diciembre de 2006.
64. Madureira EH, Marques MO, Baruselli P, Nasser LF, Rodrigues CA. Efeito do
tratamiento com eCG na taxa de concepção de vacas nelore com diferentes escores
de condição corporal inseminadas em tempo fixo (analise Retrospectiva). Acta
Scientiae veterinarae .2004. 32 (suplemento): p 228.
65. Mann GE, Lamming GE. The influence of progesterone during early pregnancy in
cattle. Reprod Dom Anim 1999. 34: 269–274.
66. Mann GE, Lamming GE, Robinson RS, Wathes DC. The regulation of interferon-t
production and uterine hormone receptors during pregnancy. J Reprod Fertil 1999.
54(Suppl):317–28.
67. Mann GE, Corpus luteum size and plasma progesterone concentration in cows.
Anim Rep Sci. 2009. 115: 296–299.

Bibliografía 153
68. Mann G, Fray M. Lamming, G. Effects of time of progesterone supplementation on
embryo development and interferon-tau production in the cow. The Veterinary Journal
2006. 171: 500–503.
69. Mantovani A, Baruselli P, Bó GA, Cavalcante A, Gacek F. Aumento de las
dimensiones del folículo dominante y del cuerpo lúteo, concentración plasmática de
progesterona y de la tasa de aprovechamiento de receptoras de embriones bovinos
sincronizadas con CIDR-B® por tiempo prolongado. Revista Brasilera de
Reproducción Animal 2002.
70. Marques MO, Madureira EH; Oliveira CA, Bó GA, Baruselli PS. Ovarian
ultrasonography and plasma progesterone concentration in Bos Taurus x Bos Indicus
heifer administered different treatments on Day 7 of the estrous cycle. Theriogenology
2002; submitted
71. Marques MO, Reis EL, Campos, Filho EP, Baruselli PS. Efeitos da adminiatracao
de eCG e de benzoato de estradiol para sincronizacao da ovulacao em vacas Bos
taurus taurus X Bos taurus indicus no periodo pós parto. In: Simposio Internacional de
Reproducción Animal, Huerta Grande. V. 392, 2003.
72. Melanie J, Starbuck A, Dailey E, Keith I. Factors affecting retention of early
pregnancy in dairy cattle Anim Reprod Sci 2004. 84: 27–39.
73. Mihm M, Austin EJ, Good TEM, Ireland JLH, Knight PG, Roche JF and Iteland JJ.
Identification of potential intrafollicular factors involved in selection of dominant follicles
in heifers. Biol. Reprod. 2000. 63: 811-819.
74. Moreira F, Risco CA, Pires MFA, Ambrose JD, Drost M, Thatcher WW. Use of
bovine somatotropin in lactating dairy cows receiving timed artificial insemination. J.
Dairy Sci. 2000. 83: 1237–1247.
75. Moura MT. Marques MO, Frare J, Madureira EH, Bó GA, Baruselli PS.
Sincronizacao da ovulacao com Crestar ® e CIDR ® para inovulacao de embrioes
bovinos em tempo fixo. 4° Simpósio Internacional de Reproducción Animal 2001; 269
(abstr).
76. Murphy B, Martinuk S. Equine chorionic gonadotropin. Endocrine Rewiews 1991.
12: 27– 44.
77. Murphy BD. Models of luteinization. Biol. Reprod. 2000. 63: 2-11.
78. Olivera AM, Tarazona MA, Ruíz CT, Giraldo EC. Modelo de luteólisis bovina. Rev
Col Cienc Pec 2007. 20: 387-393.

154 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
79. Olivera M. Gestación. En Galina C. (Ed), Reproducción de animales domésticos.
Edi. Limusa. México. 2006. p, 165–187.
80. Ohtani M, Kobayashi S, Miyamoto A, Hayashi K, Fukui Y. Real-time relatioships
between intraluteal and plasma concentrations of endotelin, oxytocin, and
progesterone during prostaglandin F2α-induced luteolysis in the cow. Biology of
Reproduction. 1998. 58: 103-108.
81. Palma G. Biotecnología de la reproducción. Biotecnología de la reproducción,
tercera edición. Instituto Nacional de Tecnología agropecuaria–INTA. Argentina. 2008.
p 693.
82. Peixoto MGCD, Bergmann JAG, Suyama E, Carvalho MRS, Penna VM. Logistic
regression analysis of pregnancy rate following transfer of Bos indicus embryos into
Bos indicus _ Bos taurus heifers. Theriogenology 2007. 67: 287–292
83. Perry GA, Smith MF, Roberts AJ, Mac Neil MD and Geary TW. Relationship
between size of the ovulatory follicle and pregnancy success in beef. J Anim Sci
2007.85: 684-689.
84. Perry, G., Smith, M., Lucy, M., Green, J., Parks, T., MacNeil, M., Roberts, A.
Geary, T. Proceedings of the national academy of sciences of the United States of
Amarica Current (PNAS). 2005. 102, 14: 5268-5273.
85. Pierce JG, Parsons TF. Glycoprotein hormones: structure and function. Annu Rev
Biochem 1982. 50: 465–495.
86. Pita F, Matute R, Tribulo R, Tribulo P, Tribulo H, Bó G. Efecto de la aplicación de
la Gonadotropina Coriónica Equina (eCG) y PGF2 alfa sobre la tasa de
aprovechamiento y tasa de preñez en un protocolo de transferencia de embriones a
tiempo fijo. VIII Simposio internacional de reproducción animal -IRAC 2009.
87. Quesada JM y Ortiz JJ. Sincronización de receptoras de embriones tratadas con
CIDR, benzoato de estradio, D Cloprostenol asociado o no a eCG. VII simposio de
reproducción animal – IRAC. 2007. p, 277.
88. Randel RD, Moseley WM. Serum luteinizing hormone surge and progesterone
near estrus in Brahman, Brahman x Hereford and Hereford heifers. J. anim. Sci. 1997;
(Supl. 1), 45, 199.
89. Rekwot P. Effects of fedding maize stover and cottonseed cake on onset of
puberty in Bunaji (Bos indicus) heifers. Tropical Animal Health and Production. 2004.
36: 637-644.

Bibliografía 155
90. Ribadu AY, Ward WR, Dobson H. Comparative evaluation of ovarian structures in
cattle by palpation per rectum, ultrasonography and plasma progesterone
concentration. Vet. Rec.1994. 5:452-457.
91. Rodríguez J. Mecanismos para el reconocimiento materno de la preñez. En:
Reproducción Bovina. Editor González C. Maracaibo – Venezuela. Ed. Fundación
GIRARZ 2001. 31–39.
92. Rekwot P. Effects of fedding maize stover and cottonseed cake on onset of
puberty in Bunaji (Bos indicus) heifers. Tropical Animal Health and Production. 2004.
36: 637-644.
93. Rienzo J, Casanoves F, Gonzalez L. Tablada, E. Margot; Díaz, María del P.;
Robledo, C. Walter; Balzarini, Mónica G. Estadística para las ciencias agropecuarias.
Triunfar. Córdoba, Córdoba. AR. 2001.
94. Roche J.The effect of nutritional management of the dairy cow on reproductive
efficiency. Animal Reproduction Science. 2006. 96: 282-296.
95. Rodríguez, Juan, Giraldo, Carlos, Castañeda, Susana et al. Análisis multifactorial
de las tasas de preã‘ez en programas de transferencia de embriones en colombia.
Rev.MVZ Cordoba, Jul. 2007, vol.12, no.2, p.978.
96. Rodrigues CA, Ayres H, Reis EL, Madureira EH, Baruselli PS. Aumento de taxa
de prenhez em vacas Nelore inseminadas em tempo fixo com o uso de eCG em
diferentes periodos pós-parto.; Acta Scientae veterinariae. V32 (Suplemento), 220.
2004.
97. Rodríguez J. Mecanismos para el reconocimiento materno de la preñez. En:
Reproducción Bovina. Editor González C. Maracaibo – Venezuela. Ed. Fundación
GIRARZ 2001. 31–39.
98. Santos J, Thatcher W, Chebel R, Cerri R, Galvão K. The effect of embryonic death
rates in cattle on the efficacy of estrus synchronization programs. Anim Reprod Sci
2004. 83: 513–535.
99. Sakase, M., Kawate, N., Nakagawa, C., Fukushima, M., Noda, M., Takeda, K.,
Ueno, S., Inaba, T., Kida, K., Tamada, H., Sawada. 2006. Preventive effects of CIDR-
based protocols on premature ovulation before timed-AI in Ovsynch in cycling beef
cows. The Veterinary Journal. In press.

156 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
100. Sartori R, Haughian J, Shaver RD, Rosa GJ, Wiltbank MC. Comparison of Ovarian
Function and Circulating Steroids in Estrous Cycles of Holstein Heifers and Lactating
Cows. . Dairy Sci. 2004.87: 905–920.
101. Sartori GJ, Rosa M and Wiltbank MC. Ovarian Structures and Circulating Steroids
in Heifers and Lactating Cows in Summer and Lactating and Dry Cows in Winter J.
Dairy Sci. 2002. 85:2813–2822
102. Sartori R., Fricke P.M. Ferreira J.C., Ginther O.J. Wilbank M.C. Follicular deviation
and acquisition of ovulatory capacity in bovine follicles. Biol Reprod 2001; 65: 1403 –
1409.
103. Senger PL. Pathways to pregnancy and parturition. Second edition revised. 2005.
104. Silke V, Diskin MG, Kenny DA, Boland MP, Mee JF, and Sreenan JM. Extent,
pattern and factors associated with late embryonic loss in dairy cows. Anim. Reprod.
Sci. 2002. 71: 1–12.
105. Shams D., Berisha B. Regulation of corpus luteum function in cattle – and
overview. Repro Dom Anim 2004. 39: 241-251.
106. Siqueira LGB, Torres CAA, Souza ED, Monteiro PLJ, Arashiro EKN, Camargo
LSA, Fernandes CAC, Viana JHM. Pregnancy rates and corpus luteum–related factors
affecting pregnancy establishment in bovine recipients synchronized for fixed-time
embryo transfer. Theriogenology 2009. 72: 949–958.
107. Silva R C P, Rodriguez C A, Marques M O, Ayres H, Reis E L, Nichi M, Madureira
E H y Baruselli P S. Efeito do eCG e do GnRH na taxa de prenhez de vacas Nelore
lactantes inseminadas em tempo fixo. Acta Scientiae Veterinariae 2004; 32
(Suplemento): 221.
108. Spey LL. and Lipner H. Ovulation In: The Physiology of Reproduction Nobil, .and J
Neill (Eds). 2006. 13: 725–780.
109. Souza AH, Viechnieski S, Lima FA, Silva FF, Araujo R, Bo´ G, Wiltbank C,
Baruselli P. Effects of equine chorionic gonadotropin and type of ovulatory stimulus in
a timed-AI protocol on reproductive responses in dairy cows. Theriogenology 2009.
72:10–21.
110. Spell AR, Beal WE, Corah LR, Lamb GC. Evaluating recipients and embryo
factors that affeect pregnancy rates of embryo transfer in beef cattle. Theriogenology
2001. 56: 287–297.
111. Spencer T, Burghardt R, Johnson G, Bazer FW. Conceptus signals for
establishment and maintenance of pregnancy. Anim Reprod Sci 2004. 83: 537– 550.

Bibliografía 157
112. Spencer T, Johnson GA, Bazer FW and Burghardt RC. Implantation mechanisms:
insights from the sheep Center for Animal Biotechnology and Genomics, Public Health,
Texas A&M University. 2004.128: 657–668.
113. Sprecher DJ, Nebel RL, Whitmn SS. The predictive value, sensitivity and
specificity of palpation per rectum and transrectal ultrasonography for the
determination of bovine luteal status. Theriogenology 1989. 31:1165-1172.
114. Sreenan J, Diskin M. Early embryonic mortality in the cow: its relationship with
progesterone concentration. Vet Rec 1982. 112: 517-521.
115. Stringfellow D, Seidel S, Manual of the International embryo transfer society. Third
edition.April 1998.
116. Steel RGD, Torrie JH. 1980. Analysis of covariance, In: Principles and Procedures
of Statistics: a Biometrical Approach, pp. 401-437. McGraw-Hill, New York.
117. Taniguchi, M., Ikeda, A., Arikawa, E., Shimizu, R., Seki, M., Karaki, M.,
Rajamahendran, R. Otoi, T. Ovarian follicular and corpus luteum changes,
progesterone concentrations, estrus and ovulation following estradiol
benzoate/progesterone based treatment protocol in cross-bred cows. Animal
Reproduction Science. 2007. 99: 389-394.
118. Thatcher W, Guzeloglu A, Binelli M, Hansed TR, Pru JK. Uterine conceptus
interactions and reproductive failure in cattle. Theriogenology 2001. 56: 1435–1450.
119. Thatcher W, Moreira F, Santos J, Mattos R, lopez F, Pancarci S, Risco C. Effects
of hormonal treatments on reproductive performance and embryo production.
Theriogenology 2002. 55: 75– 90.
120. Thibier M. More than half a millon bovine embryos transfered in 2002: a report
from the IETS data retrieval committee. IETS Newsletter, 2003.
121. Tom JW, Pierson RA, Adams GP. Quantitative echotexture analysis of bovine
ovarian follicles. Theriogenology 1998 50:339-346.
122. Tribulo H, Moreno D, Cutaia L, Gatti G, Tribulo R, Caccia M, et al. Pregnancy
rates in embryo recipients treated with progesterone vaginal device and eCG and
transferred without estrus detection. Theriogenology 2002; 57: 563 [abstract].
123. Vasconcelos J, Demétrio D, Santos R, Chiari J, Rodrigues C, Sá Filho O. Factors
potentially affecting fertility of lactating dairy cow recipients. Theriogenology 2006.65:
192–200.

158 Efectos de la aplicación de eCG (Día 5 u 8) sobre el desarrollo del cuerpo lúteo, nivel de progesterona y tasa de preñez en hembras receptoras de embriones bovinos
124. Vasconcelos J, Sartori R, Oliveira H, Guenther J, Wilbank M. Reduction is size of
the ovulatory follicle reduces subsequent luteal size and pregnancy rate.
Theriogenology 2001. 56: 307–314.
125. Viana JHM, Torres CAA, Fernandes CA, Ferreira AM. Avaliaçao ultra- sonográfica
do corpo lúteo em novilhas mestiças utilizadas como receptoras de embriao. Archiv.
De Reprod. Anim. 1998. 5:42-47.
126. Vos PLAM, van der Schans A, de Wit AAC, Willemse AH, Bevers MM, Dieleman
SJ. Effect of neutralization of pregnant mare’s serum gonadotrophin (PMSG) shortly
before or at the preovulatory LH surge in PMSG-superovulated heifers on follicular
function and development. J Reprod Fertil 1994;100: 387–93.
Top Related