







Languages
Pages
Legal


1. Introduction
The ecology of larval fish has been studied for many years because of their importance inunderstanding the population dynamics of commercially important fish species around the world.Their interesting morphology and ecology also has been studied to learn about the wide varietyof behavioral and morphological adaptations used by fish larvae, which are often quite differentin appearance than their juveniles and adults (Moser 1981; Leis and McCormick 2002; Leis2006; Masuda 2009). However, one major group of fish larvae that has received less attentionfor various reasons are leptocephali, which are the larvae of marine and freshwater eels and theirclose relatives that live in the ocean (Castle 1984; Smith 1989a; Mochioka 2003; Lecomte-Finiger 2004). There has been increasing concern recently though, that changes in the ocean due
Aqua-BioSci. Monogr. (ABSM), Vol. 2, No. 4, pp. 1–94 (2009)© 2009 TERRAPUB, Tokyo. All rights reserved. www.terrapub.co.jp/onlinemonographs/absm/
*Corresponding author at:Ocean Research Institute, The University of Tokyo1-15-1, Minamidai, Nakano-ku, Tokyo 164-8639, JapanPhone: +81-3-5351-6512Fax: +81-3-5351-6514e-mail: [email protected]
Received on May 12, 2009Accepted on August 3, 2009Published online on
October 31, 2009
Keywords• leptocephali• Anguilliformes• eels• fish larvae• early life history• larval ecology• larval growth rates• larval distribution• metamorphosis• recruitment
Ecology of Anguilliform Leptocephali:Remarkable Transparent Fish Larvaeof the Ocean Surface Layer
Michael J. Miller*
Ocean Research Institute, The University of Tokyo1-15-1, Minamidai, Nakano-ku, Tokyo 164-8639, Japan
AbstractThis review examines the present state of knowledge about the ecology of
anguilliform leptocephali, which are the unique but poorly understood larvae ofeels. All eels spawn in the ocean and their leptocephali live in the ocean surfacelayer. Their presence worldwide and basic biology have not been extensively studieddue to their strong ability to avoid standard plankton nets and their fragile trans-parent bodies. Leptocephali have laterally compressed bodies and contain a highproportion of transparent energy storage compounds. They have diverse morpho-logical features, but appear to feed only on particulate material, such as marinesnow or discarded larvacean houses. Some information on their chemical compo-sition, respiration, growth rates, depth distributions, swimming ability, metamor-phosis, and recruitment patterns has been reported, which highlights the interest-ing and unique aspects of leptocephalus larvae. Regional zoogeography and re-productive ecology of adults and ocean currents affect the spatial and temporaldistribution patterns of leptocephali, which have long larval durations, but mostlife histories and larval recruitment behaviors remain undocumented. Their trans-parency, feeding strategy, and large size seem to be a unique and successful larvalstrategy, but the abundance and ecological significance of leptocephali in the oceanappear to have been underestimated.

2 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
to alterations in the ocean–atmosphere system may be affecting the survival of anguillid lepto-cephali (see Miller et al. 2009a), so there is a need to gain a greater understanding of the ecologyof leptocephali.
Leptocephali are poorly known largely because they grow much larger than typical fishlarvae, and they are rarely collected by the standard-sized plankton nets used by fisheries scien-tists and biological oceanographers. As will be reviewed below, they have large eyes,mechanoreceptors, and can actively swim both forwards and backwards, so this in combinationwith their large size appears to make leptocephali well adapted to avoid small plankton nets(≤1 m diameter) or any sized trawl during the day (Castonguay and McCleave 1987a; Miller andMcCleave 1994; Miller and Tsukamoto 2004; Miller et al. 2006a). Another problem that hasslowed the progress in research on leptocephali is that these larvae typically show no resem-blance to the juvenile or adult forms of each species, so it is extraordinarily difficult to matchlarval forms to adult species using morphological characteristics.
Leptocephali differ so much from their adult forms that for about a century they werethought to be a unique type of marine fish (Smith 1989a). Eventually it was realized that lepto-cephali are actually the larval forms of the fishes of the superorder Elopomorpha, which in-cludes species with both eel-like and typical fish-like bodies. The true eels of the Anguilliformesall have elongate body forms and swim using typical anguilliform locomotion (Gray 1933) thatenables them to swim in both directions (D’Août and Aerts 1999). This order includes about 15families, with all but one family being almost entirely marine species throughout their life histo-ries (Böhlke 1989a). The eels of the Anguillidae are the catadromous eels that live in freshwaterand estuarine habitats as juveniles and adults, but spawn in the ocean and have leptocephaluslarvae (Tesch 2003; Aoyama 2009). The gulper and swallower eels of the historical orderSaccopharyngiformes are also eel-like in body form, and genetically appear to be containedwithin the Anguilliformes (Inoue et al. 2004). The bonefishes and spiny eels of the Albuliformes(including the historical Notacanthiformes), and the tarpons and ladyfishes of the Elopiformeshave more typical fish-like bodies and do not resemble eels even though they all share thecommon larval form of leptocephalus larvae. All of these elopomorph orders are distributedworldwide from tropical to temperate waters and in the deep sea for some species (Nelson 2006),although anguillids are absent in the South Atlantic and eastern Pacific oceans (Aoyama 2009).
Despite their global distribution and the existence of more than 800 species of eels (Nel-son 2006), little is known about the life histories of most species or the ecology of their lepto-cephali (Böhlke 1989a,b; Smith 1989a; Miller and Tsukamoto 2004). Adults are difficult tostudy due to the nocturnal and often fossorial behavior of most eels, or the deep depths at whichmany species live in the ocean. Leptocephali are difficult to collect unless large trawls are fishedat night (Miller and Tsukamoto 2004, 2006), but even if they are collected, leptocephali rarelysurvive being captured due to their fragile body form.
Leptocephali are unusual due to their highly laterally compressed bodies, which are al-most totally transparent (Fig. 1). They are transparent as a result of their bodies mostly contain-ing transparent energy storage material consisting primarily of glycosaminoglycan (GAG) com-pounds (Pfeiler 1999; Pfeiler et al. 2002), which also provide structural support for the bodyuntil they are converted into new body tissues when the leptocephali metamorphose into juve-nile eels at the end of their larval phase. These larvae are very fragile though, because their bodyis covered with a thin layer of tissue that is only a few cell layers thick (Hulet 1978; Suzuki andOtake 2000; Nakamura et al. 2002) and is easily damaged.
Leptocephali exhibit a wide variety of body shapes that range from very long and thin todeep, with rounded or pointed tails (Fig. 2). Head shapes also vary greatly (Fig. 3; but see be-low). Maximum sizes of leptocephali can range from about 50 mm to greater than 300 mm (totallength) (Castle 1984; Smith 1989a; Böhlke 1989b), but they remain transparent and very fragileregardless of their size or body shape until they metamorphose into juvenile eels.
Research on leptocephali primarily began after expeditions started searching for the spawn-ing place of the Atlantic eels by collecting their leptocephali in the early part of the last century(Schmidt 1922; Boëtius and Harding 1985; McCleave 2003). During these early surveys foranguillid leptocephali, which expanded into the Indo–Pacific (Jespersen 1942), various othertaxa were also collected, and some of these marine eel leptocephali were later studied (e.g.

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 3
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Bertin 1938; Bauchot 1959; Castle 1970, 1979, 1997; Castle and Raju 1975; Smith and Castle1982; Karmovskaya 1990; Castle and Smith 1999). Research on leptocephalus morphology andtheir species identifications achieved greater advances as a result of collections made in morerecent years in the western South Pacific (Castle 1963, 1964, 1965a,b,c), the Gulf of Guinea ofwestern Africa (Blache 1977), and to an even greater extent in the western North Atlantic (Smith1969, 1974), where most leptocephali were eventually identified to the species level (Smith1979; Böhlke 1989b). Other more recent studies compared regional catches of leptocephali tothe known distributions of adults (Richardson and Cowen 2004a,b; Ross et al. 2007). In mostparts of the world however, such as in the Indo–Pacific, the species identifications are still notknown for the majority of leptocephali (Mochioka et al. 1982, 1991; Tabeta and Mochioka1988a; Miller et al. 2002a, 2006a; Miller and Tsukamoto 2004, 2006).
Fig. 1. Photographs of leptocephali of the Japanese eel, Anguilla japonica, that were artificially spawned and reared in thelaboratory at the IRAGO Institute in Japan. The larvae are approximately 30–50 mm long, and about 200 days old. Photo-graphs are courtesy of Yoshiaki Yamada.

4 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
There has been research on the distributions, life history characteristics, and assemblagestructure of leptocephali in a variety of regions of the world using various levels of identifica-tion. In the western North Atlantic, the distributions of anguillid leptocephali were studied inmore detail to help define the spawning areas of the two species, the European eel, Anguillaanguilla, and the American eel, Anguilla rostrata, which have overlapping spawning areas inthe southern Sargasso Sea (Schoth and Tesch 1982; Kleckner et al. 1983; McCleave et al. 1987;Kleckner and McCleave 1988; Tesch and Wegner 1990). These surveys for anguillid lepto-cephali also collected many marine eel species whose life histories were examined (McCleaveand Miller 1994; Miller and McCleave 1994; Miller 1995, 2002a; Wippelhauser et al. 1996).Studies on the assemblages of leptocephali in these areas were conducted as well (Miller andMcCleave 1994, 2007; Miller 1995). Leptocephali also have been collected in the Gulf of Mexico(Smith 1989a; Crabtree et al. 1992), over or along the edge of the continental shelf of the south-eastern US east coast (Fahay and Obenchain 1978; Ross et al. 2007), and around Barbados inthe eastern Caribbean Sea region (Richardson and Cowen 2004a). In the northeastern NorthAtlantic, research has mostly focused on leptocephali of anguillids (Tesch 1980; Bast and Strehlow1990; McCleave et al. 1998) and a few other taxa of leptocephali (e.g. Strehlow et al. 1998;Correia et al. 2002a,b, 2003). A few taxa have been reported from the east coast of South Americain recent years (De Castro and Bonecker 2005; Figueroa and Ehrlich 2006).
Fig. 2. Photographs of freshly caught leptocephali of 13 families of the Anguilliformes, which show the wide variety of bodyshapes of leptocephali. Members of the three subfamilies of the Congridae are also shown. Sizes of leptocephali are notproportional.

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 5
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Various surveys for leptocephali in the Indo–Pacific region have been conducted recentlyto learn about the spawning areas of tropical and temperate anguillid species in the westernPacific and Indonesian Seas, which have resulted in many areas being sampled (e.g. Fig. 4).These studies have centered on the anguillid eels that spawn offshore (Tsukamoto 1992, 2006;Aoyama et al. 1999, 2007; Ishikawa et al. 2001; Miller et al. 2002b; Tsukamoto et al. 2003;Kuroki et al. 2009a) or those that spawn in the Indonesian Seas (Aoyama et al. 2003; Wouthuyzen
Fig. 3. Photographs showing typical head and jaw shapes of many types of anguilliform leptocephali. These taxa are Anguillamarmorata (A), Ariosoma major (B), Heteroconger hassi (C), Conger sp. (D), Bathycongrus sp. (E), Gnathophis sp. (F),Muraenesox cinereus (G), Serrivomeridae sp. (H), Muraenidae sp. (I), Chlopsidae sp. (J), Nettenchelys sp. (K), andSynaphobranchinae sp. (L), Ilyophinae, Synaphobranchidae (M), Eurypharynx pelecanoides (N), Cyema atrum (O). Photosare reprinted from Miller and Tsukamoto (2004).

6 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
et al. 2009). In 1991, the spawning area of the Japanese eel, Anguilla japonica, was discovered(Tsukamoto 1992), and subsequent surveys collected recently hatched preleptocephali near theWest Mariana Ridge, which includes several shallow seamounts (Tsukamoto 2006, 2009). Itwas also found that a population of the giant mottled eel, Anguilla marmorata, spawns in thesame general area as A. japonica (Miller et al. 2002b; Kuroki et al. 2009a), and spawning areasof other anguillids were detected in several other regions (Aoyama et al. 1999, 2003; Kuroki etal. 2008a). As a result of the surveys for anguillid leptocephali in the western Pacific region, thespecies composition, assemblage structure, and life history characteristics have been examinedfor marine eel leptocephali (Minagawa et al. 2004; Wouthuyzen et al. 2005; Miller et al. 2006a,b),and other research resulted from fisheries related collections (Takahashi et al. 2008).
In conjunction with some of these surveys for leptocephali in both the Pacific and Atlanticoceans, data on the depth distributions of leptocephali were gathered (Schoth and Tesch 1984;Castonguay and McCleave 1987a; Kajihara et al. 1988; Otake et al. 1998). These studies indi-cated that leptocephali live in the surface layer of the ocean at depths mostly in the upper 100 mat night, and likely deeper during the day to depths typically less than 250–300 m for somespecies that make diurnal vertical migrations.
Some leptocephali from recent surveys also have been used to study the age and growthand species identifications with otolith and gene-sequencing techniques. Gene-sequence analy-sis has become the standard technique for identifying anguillid leptocephali in the Indo–Pacificwhere counts of their numbers of myomeres (muscle segments that correspond to the number ofvertebrae in adults) often overlap among the different species (Aoyama et al. 1999, 2003, 2007;Kuroki et al. 2006a, 2008a). Genetic sequences have also been used recently to match lepto-cephali to their adult species (Ma et al. 2007, 2008a), which is a technique that will facilitatematching of adult and larval forms in the Indo–Pacific where little progress has been made inidentifying leptocephali. New data on the likely ages of Indo–Pacific leptocephali have beengained from studies on their otolith microstructure, with anguillid (Ishikawa et al. 2001;Tsukamoto et al. 2003; Kuroki et al. 2005, 2006a,b, 2007, 2008a), congrid (Lee and Byun 1996;Kimura et al. 2004) and other families of marine eel leptocephali having been studied (Ma et al.2005; Lee et al. 2008). These studies have found that the growth rates of different species of
Fig. 4. Species diversity of leptocephali at various stations for leptocephali from 1995–2003 (A). Diversity is plotted as Hill’sN1 index. Symbols correspond to N1 values that range between about 2 and 8. Reprinted with modifications from Minagawaand Miller. Distribution and diversity patterns of leptocephali. In: Watanabe Y (ed). Living Marine Resources: How Do TheyFluctuate in the Ocean? Tokai University Press, Tokyo. 2005; pp. 168–189. © 2005, Minagawa and Miller. Map of the Indo–Pacific region (B) showing the approximate distribution of coral reef areas in red, and bathymetry of the ocean in blue (lightblue is shallow, and dark blue is deep), which was modified from imagery obtained from the ReefBase project (http://www.reefbase.org; NASA and NOAA; see Stumpf et al. 1999).

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 7
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
leptocephali may vary based on differing life history characteristics (Ma et al. 2005; Kuroki etal. 2006a). Otolith increments also have been examined in a few species of leptocephali in theNorth Atlantic such as in anguillids (Castonguay 1987), conger eels (Correia et al. 2002a,b,2003), and a few other species (Bishop et al. 2000). Other studies have examined otolith depo-sition rates of leptocephali held in captivity as they metamorphose into juveniles (Chen andTzeng 1996; Powles et al. 2006). Variations in otolith increment widths and Sr:Ca ratio dataobtained from glass eel otoliths has provided information on the timing of metamorphosis invarious species (Otake 2003; Aoyama 2009).
Some studies have also examined the biology and physiology of leptocephali collected inthe wild to learn about their ecology. These studies have included examinations of gut contents,which were found to contain particulate matter such as marine snow, fecal pellets, or discardedlarvacean (appendicularian) houses (Otake et al. 1990, 1993; Mochioka and Iwamizu 1996).The types of material observed in the guts indicate that leptocephali feed on marine snow-likematerial, but not zooplankton or phytoplankton. Other types of studies on leptocephali haveincluded enzyme assays to examine levels of respiration (Pfeiler and Govoni 1993), oxygenconsumption experiments (Bishop and Torres 1999), or analyses of the chemical composition oftheir bodies (Donnelly et al. 1995; Pfeiler et al. 1998; Bishop et al. 2000). Physiological meas-urements and age estimates made on four species of leptocephali from the eastern Gulf of Mexicowere used to model the energetics of these species (Bishop and Torres 2001). However, some ofthe most extensive research on the physiology and chemical composition of leptocephali hasbeen done on bonefish larvae (Albuliformes), since they can often be easily collected in coastalwaters and quickly moved to the laboratory (e.g. Pfeiler 1986, 1996, 1997, 2001; Padrón et al.1996). Few studies on most taxa of leptocephali have been done though, due to the difficulty incollecting healthy specimens in nets at sea and keeping them alive on board research vessels.
The development of artificial culture techniques for anguillid species such as Anguillajaponica (Tanaka et al. 2001, 2003; Tanaka 2003), provides new potential for conducting basicresearch on the biology and behavior of leptocephali in the laboratory though. The ultimate goalis to provide a consistent supply of artificially spawned and reared glass eels for aquaculture, butthe recent success in rearing leptocephali through metamorphosis into the yellow eel stage (Tanakaet al. 2003) has enabled all stages of eels to be available for laboratory research. This has re-sulted in comparative studies on phototaxis (Yamada et al. 2009) and buoyancy (Tsukamoto etal. 2009) of different stages leptocephali and glass eels reared in the laboratory. Such studiesrepresent a new research frontier that can help improve our understanding of leptocephali untiltechniques can be developed to study wild-caught leptocephali.
Another recent development that is assisting research on leptocephali was the rapid devel-opment of digital imaging technology. The availability of digital cameras and digital imagingsystems for microscopes have enabled freshly caught leptocephali to be photographed duringsampling surveys (Miller and Tsukamoto 2004). Few photographs of leptocephali were pub-lished during the era of film cameras, so these new photographs have helped to document themorphology of leptocephali in several recent studies (Kuroki et al. 2005, 2008a; Kimura et al.2006; Tsukamoto 2006) or books and reviews (Miller and Tsukamoto 2004, 2006). This reviewof the present state of knowledge about the ecology of anguilliform leptocephali will also usemany digital photographs of freshly caught leptocephali to illustrate various aspects of how themorphology of leptocephali may be related to their ecology.
The objective of this review is to provide a broad overview of the ecology of leptocephalithat ranges from where leptocephali are found in the ocean, to what can be deduced about theirbiology and ecology from their morphology and from studies conducted on them using variousanalytical techniques. This begins with an overview of what is known about the basic morphol-ogy, biology, and behavior of leptocephali, which is followed by a section on the zoogeographyof leptocephali that describes where the various types of eels live, and where their larvae arelikely to be distributed. Then, aspects of the ecology of leptocephali are evaluated to provide anoverall picture of the present state of knowledge about leptocephali and on the possible ecologi-cal significance of these cosmopolitan, surface dwelling larvae.

8 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
2. Biology of leptocephali
2-1. Developmental stages
Leptocephali have several different morphologically distinguishable stages that seem tobe consistent among all anguilliform taxa. Eels have large eggs compared to many other fishes,which can range from about 1–4 mm in diameter, and have a large perivitelline space (Castle1984; Smith 1989a; Umezawa et al. 1991; Horie et al. 2002). The Anguilliformes is one of thefew orders of fishes that have the characteristic of yolk extension (Virta and Cooper 2009),which can be seen in the posterior extension of the yolk in the newly hatched larva in Fig. 5.Some eel larvae appear to be born in a morphologically very undeveloped state with almost nodevelopment of the features of the head, as has been observed in species hatched in the labora-tory, such as A. japonica (Fig. 5; Okamura et al. 2002), Muraenesox cinereus (Umezawa et al.1991), and Conger myriaster (Horie et al. 2002). Other species though, such as ophichthids mayhatch at a more developed state with the eyes and some early teeth already formed (Leiby 1979a,1989). Evidence of more advanced development before hatching was seen in what appears to bea muraenid larva collected while still within the egg case, which had fully developed eyes andearly jaws (Fig. 6). Larvae of A. japonica (Fig. 7A) and Serrivomeridae (Fig. 7C) collected in theocean, suggest these taxa, at least, hatch in a very undeveloped state and then develop the mor-phological features of the head during the preleptocephalus stage (Fig. 8).
Fig. 5. Drawings of recently hatched Anguilla japonica larvae hatched in the laboratory from 1–7 days after hatching,showing the location of various types of mechanosensory cells. Scale bars are 1 mm. Reprinted with permission of JohnWiley & Sons, Inc. from Okamura A, Oka HP, Yamada Y, Utoh T, Mikawa N, Horie N, Tanaka S. Development of lateral lineorgans in leptocephali of the freshwater eel Anguilla japonica (Teleostei, Anguilliformes). Journal of Morphology, 2002; 254:81–91. © 2002, Wiley-Liss, Inc., A Wiley Company.

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 9
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Fig. 6. Photograph of what appears to be a larva of the Muraenidae that was collected in the western North Pacific near theWest Mariana Ridge while it was still inside of an egg case before hatching.
Fig. 7. Photographs of preleptocephali of Anguilla japonica (A), Ophichthidae (B), and Serrivomeridae (C), showing theundeveloped morphology of recently hatched wild-caught anguilliform larvae. Scale bars are 1 mm.

10 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
The preleptocephalus stage is defined as the period just after hatching when the larva isstill absorbing its oil globule and not actively feeding on external food. These larvae have poorlydeveloped eyes, few or no teeth, and an oil globule. The size of preleptocephali of various taxamay range from about 3–7 mm, but no comparative analysis has been published. An oil globuleis clearly present in anguillid preleptocephali (Fig. 8; Tsukamoto 2006; Kuroki et al. 2009a) andin ophichthid and serrivomerid preleptocephali (Fig. 7). The oil globule of the ophichthid spe-cies shown in Fig. 7B and of another species collected along coastal Japan (Kimura et al. 2006)shows a golden color. At least some species are born with no teeth, or just rudimentary teeth thatmay be easily damaged in the net (Fig. 8). Teeth appear and grow during early posthatchingdevelopment. The eggs and preleptocephali of most species of marine eels have not been de-scribed, so much research remains to be done on the early development of each family of eels.
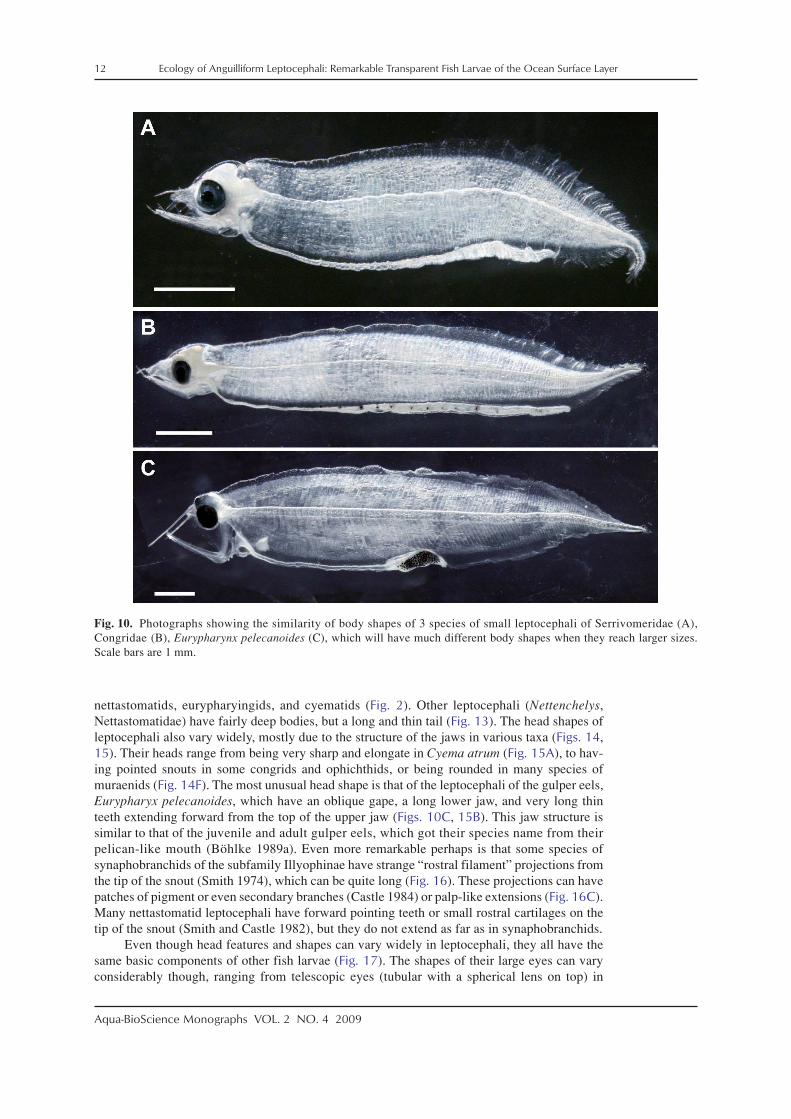
The leptocephalus stage begins after the oil globule is absorbed and the eyes and teeth areformed. Larvae at this early stage have relatively few, but very long, forward pointing teeth(Fig. 9), and this stage has been referred to by Leiby (1979b, 1989) as the engyodontic stage.Body shapes of these small leptocephali are generally similar (Fig. 10), even though they mayultimately develop more long and thin or deeper bodies. As leptocephali grow larger, the longteeth are replaced by shorter ones, more teeth are formed, and the teeth point less forward(Figs. 3, 11). These larvae have been referred to as the euryodontic stage (Leiby 1979b, 1989).This transition appears to occur at around 20 mm in ophichthids (Leiby 1989) and other lepto-cephali (Castle 1984), and can also be seen in the development series of Derichthys serpentinusof Castle (1970) (Fig. 11).
Fig. 8. Photographs of the head regions of preleptocephali of the Japanese eel, Anguilla japonica, at various stages ofdevelopment ranging from having unpigmented eyes and no teeth (A), to fully formed eyes and jaws with teeth (D), but alllarvae still having an oil globule. Scale bar in (C) is 1 mm. Reprinted by permission of Macmillan Publishers Ltd: Nature.2006; 439: 929. Tsukamoto K. Spawning of eels near a seamount.

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 11
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Leptocephali keep growing until they reach a species specific maximum size. This maxi-mum size varies from about 50–100 mm in many species and up to 300 mm or more in sometaxa such as Ariosoma and Nemichthys. The full size leptocephali then metamorphose into theglass eel or elver stage (Fig. 12). During the process of metamorphosis the laterally compressedbody of the leptocephalus changes to be more rounded. Towards the end of the transformationinto the glass eel and elver stages their bodies become pigmented. See Section 2-7 for a moredetailed discussion of metamorphosis in eels.
2-2. Morphological features
Although body shapes (Figs. 2, 13) and head shapes (Figs. 3, 14–17) can vary widelyamong taxa, leptocephali all have similar basic morphological features. Drawings and photo-graphs of the diverse body shapes of leptocephali and guides to identify them have been pub-lished (e.g. Blache 1977; Smith 1979; Fahay 1983; Tabeta and Mochioka 1988a; Böhlke 1989b;Miller and Tsukamoto 2004). All leptocephali are strongly laterally compressed and transparent,with a small tubular gut along the ventral margin. Dorsal, anal, and pectoral fins are presentstarting in the euryodontic leptocephalus stage, but the pectoral fin does not have rays untilmetamorphosis (Smith 1989a). Body shapes can vary from very long and thin as in some congrids,nemichthyids, and some nettastomatids, to deep bodied such as in chlopsids, some congrids,
Fig. 9. Photographs of the head and jaws of 4 species of engyodontic stage leptocephali showing the variation in thepatterns of length and angle of leptocephalus teeth. Anguilla marmorata (A), a likely serrivomerid (B), Eurypharynxpelecanoides (C), Muraenidae (D).
12 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
nettastomatids, eurypharyingids, and cyematids (Fig. 2). Other leptocephali (Nettenchelys,Nettastomatidae) have fairly deep bodies, but a long and thin tail (Fig. 13). The head shapes ofleptocephali also vary widely, mostly due to the structure of the jaws in various taxa (Figs. 14,15). Their heads range from being very sharp and elongate in Cyema atrum (Fig. 15A), to hav-ing pointed snouts in some congrids and ophichthids, or being rounded in many species ofmuraenids (Fig. 14F). The most unusual head shape is that of the leptocephali of the gulper eels,Eurypharyx pelecanoides, which have an oblique gape, a long lower jaw, and very long thinteeth extending forward from the top of the upper jaw (Figs. 10C, 15B). This jaw structure issimilar to that of the juvenile and adult gulper eels, which got their species name from theirpelican-like mouth (Böhlke 1989a). Even more remarkable perhaps is that some species ofsynaphobranchids of the subfamily Illyophinae have strange “rostral filament” projections fromthe tip of the snout (Smith 1974), which can be quite long (Fig. 16). These projections can havepatches of pigment or even secondary branches (Castle 1984) or palp-like extensions (Fig. 16C).Many nettastomatid leptocephali have forward pointing teeth or small rostral cartilages on thetip of the snout (Smith and Castle 1982), but they do not extend as far as in synaphobranchids.
Even though head features and shapes can vary widely in leptocephali, they all have thesame basic components of other fish larvae (Fig. 17). The shapes of their large eyes can varyconsiderably though, ranging from telescopic eyes (tubular with a spherical lens on top) in
Fig. 10. Photographs showing the similarity of body shapes of 3 species of small leptocephali of Serrivomeridae (A),Congridae (B), Eurypharynx pelecanoides (C), which will have much different body shapes when they reach larger sizes.Scale bars are 1 mm.

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 13
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Synaphobranchidae (Figs. 14C, 15E, 16, 17B), to perfectly round in others such as Anguilla andThalassenchelys (Figs. 15C, D). Other species have more oval eyes (Fig. 15). Leptocephali havelarge brains and a heart that is visible through the body wall (Fig. 17), but they lack well devel-oped gills until they approach metamorphosis. The operculum is open so water can be ejectedfrom the buccal cavity during feeding. There is very little ossification of the head or body otherthan the teeth until metamorphosis. Leiby (1979a, 1989) described the osteology of ophichthidleptocephali.
The gut and internal organs of leptocephali are generally quite small and inconspicuous.Their tubular gut can be completely straight with no visible swellings or there can be distinctswellings and curvatures along the gut (Figs. 2, 13, 18). The swellings appear to be liver tissue,but other organs also contribute in some cases. Anteriorly, the esophagus extends from the oralcavity, over the heart (Figs. 17A, B), and along the anteroventral margin of the body until itreaches the rudimentary stomach, which is located at the beginning of the intestine (Fig. 18).The lack of a distinguishable stomach is typical of most fish larvae (Govoni et al. 1986). Avisible swelling at this location often includes the liver and gallbladder (Fig. 18). The liver lobes
Fig. 11. Drawings on the body shapes and head morphology of different developmental stages of the leptocephali of Derichthysserpentinus, ranging from an early leptocephalus (7 mm) to a late metamorphosing larva (71 mm) (other stages: 18, 37, 54mm). Modified from with permission of the American Society of Ichthyologists and Herpetologists from Castle. Distribu-tion, larval growth, and metamorphosis of the eel Derichthys serpentinus Gill, 1884 (Pisces, Derichthyidae). Copeia 1970;1970: 444–452.

14 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Fig. 12. A developmental growth series of Anguilla japonica larvae reared in the laboratory from a newly hatched larva(preleptocephalus, 3.6 mm in total length, 0 days old) to the maximum size at the end of the leptocephalus stage (59.0 mm,260 days old), and the glass eel stage (58.5 mm, 268 days old). The body size of leptocephali diminished slightly during theprocess of metamorphosis. Specimens older than 260 days are the same individual. Reprinted with kind permission fromSpringer Science + Business Media: Marine Biology, Positive buoyancy in eel leptocephali: an adaptation for life in the oceansurface layer. 156, 2009, 835–846. Tsukamoto K, Yamada Y, Okamura A, Tanaka H, Miller MJ, Kaneko T, Horie N, Utoh T,Mikawa N, Tanaka S. Figure 2. © 2009, Springer-Verlag. (Original photographs provided by Hideki Tanaka of the NationalResearch Institute of Aquaculture).

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 15
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
can be split among several different gut swellings, and the kidney is a linear structure located ontop of the gut at various locations depending on the species (Fig. 17C).
All congrid leptocephali lack any major gut swellings, but in one group the straighttubular gut extends outside of the body and trails freely behind the body. This is referred toas an exterillium gut (see Ariosoma type leptocephalus in Fig. 2, with most of the exterilliumgut broken off), and they can be longer than 150 mm in some species of large bathymyridleptocephali (Ariosoma, Parabathymyrus) that have this type of gut (Mochioka et al. 1982;Tabeta and Mochioka 1988a; Smith 1989a). These extensions appear to simply lengthen the gutto increase absorptive surface area as they do in other fish larvae (Moser 1981), but they havenot been studied in leptocephali.
Fig. 13. Photographs of 5 spe-cies of leptocephali that havedifferent body shapes and gutmorphology.

16 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
The body of leptocephali is transparent mainly because it is comprised mostly of transpar-ent material contained in a mucinous pouch (Pfeiler 1999) and their circulatory system containscolorless blood. Extending through the center of the body are the spinal chord (Yoshida et al.1995), notochord, and dorsal aorta (Fig. 17). Other components of the circulatory system extendalong the gut, and there are several vertical blood vessels including the renal arteries that extendvertically up to the dorsal aorta (Hulet 1978). The blood is transparent because leptocephali lackred blood cells (Hulet and Robins 1989).
The integument of leptocephali is thin and fragile. The mucinous pouch is overlain by athin layer of muscle tissue that is organized into individual muscle segments, called myomeres.The myomeres are overlain by a thin layer of epidermal cells, which are only a few cells thick
Fig. 14. Photographs of 6 species of leptocephali that have different head shapes that range from very pointed snouts torounded and blunt snouts. Muraenidae (A), Congridae (B), Synaphobranchidae (C), Neenchelys (Ophichthidae) (D),Serrivomeridae (E), Muraenidae (F). Scale bars are 1 mm.

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 17
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
(Leonard and Summers 1976; Hulet 1978; Suzuki and Otake 2000; Nakamura et al. 2002). Theskin of A. japonica leptocephali generally consists of only one layer of club cells that includesecretory vacuoles or a few epidermal cells, which overlay the dermis (Suzuki and Otake 2000).The same was observed in late-stage Conger myriaster leptocephali, which during metamor-phosis also developed more layers of club cells, a layer of epidermal cells underneath, and somemucus cells (Fig. 19; Nakamura et al. 2002). Because the skin of leptocephali is so thin and theydo not develop substantial gills until after metamorphosis begins, it has generally been assumedthat they mostly respire through gas exchange across their integument (Pfeiler 1999).
Fig. 15. Photographs of 6 species of leptocephali that have different eye shapes that range from round to oval, or are tel-escopic with a spherical lens as in panel (E). Cyema atrum (Cyematidae) (A), Eurypharynx pelecanoides (B), Anguillamarmorata (C), Thalassenchelys (D), Synaphobranchinae (E), Congridae (F). Scale bars are 1 mm.

18 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
See Blache (1977), Smith (1979, 1989a), Böhlke (1989b), and Leiby (1989) for descrip-tions of leptocephalus morphology, Hulet (1978) for a detailed morphological analysis of theleptocephalus of Ariosoma balearicum, and Castle (1984) for an overview of the morphology ofeach family of anguilliform leptocephali.
2-3. Sensory organs
Leptocephali have several different sensory systems that they likely use for feeding orpredator avoidance. Visual and olfactory systems are obvious from their external morphology orthe size of different parts of the brain (Pfeiler 1989), and these two senses are thought to be thedominant senses of leptocephali (Smith 1989a). All leptocephali have large eyes even thoughtheir position can vary from being in about the middle of the head, to being raised on the top ofthe head such as in Cyema atrum (Fig. 15A). The consistently large eyes emphasize their likely
Fig. 16. Photographs of 3 species ofsynaphobranchid leptocephali of the subfamilyIlyophinae that have rostral filaments that ex-tend from the tip of their snouts, which canhave pigmentation or even other types of fila-ments extending from them as seen in the bot-tom panel. The top and bottom photographs arefrom Miller and Tsukamoto (2004).

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 19
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
importance. Thalassenchelys leptocephali can have a remarkably large eye that fills the spacebetween the upper jaw and the top of the head (Fig. 15C). Relative eye size is not so largethough in Thallasenchelys leptocephali that reach very large body sizes of about 300 mm(Castle and Raju 1975). The apparently large size of the optic lobe of the brain of leptocephali,also supports the inference that vision is an important sense for eel larvae, because theleptocephali of Anguilla japonica (Tomoda and Uematsu 1996) and Ariosoma balearicum(Hulet 1978) have been reported to have large optic lobes. For A. japonica, the relative size
Fig. 17. Various morphological features labeled on two species of leptocephali with very different head shapes (A, B). Thetop larva is a garden eel leptocephalus (Gorgasia, Congridae), and the bottom is a synaphobranchid of the subfamily Ilyophinae.NO = Nasal organ. The lower side of the body of an anguillid leptocephalus showing various morphological features (C).

20 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
of the optic lobe decreases in the juvenile stage, when eels are no longer visual feeders(Tomoda and Uematsu 1996). A 50-mm long Anguilla leptocephalus was found to have onlyone type of rod-like retinal photoreceptor (Pankhurst 1984), which is similar to the observationsof only rods in the retina of A. balearicum leptocephali (Hulet 1978). These observations sug-gest that leptocephalus eyes are adapted to low light conditions, as is true of some adult eels(Hess et al. 1998). This is in contrast with many other types of fish larvae, which have beenfound to have pure cone retinas (Evans and Fernald 1990).
Two categories of eyes are present in leptocephali: normal and telescopic eyes. The latterare found only in the Synaphobranchidae (Figs. 14C, 15E, 16, 17B) and some notacanthids(Smith 1974, 1979). In the two subfamilies of synaphobranchids, a large spherical lens is posi-tioned on top of a tube containing the retina (Fig. 15E). This eye structure is thought to be relatedto the slightly deeper depths and lower light where these larvae live (Smith 1989a). Other deep-sea fishes have similar eyes (Warrant and Locket 2004). Perhaps the eyes look upward so theycan see the silhouettes of food particles against the downwelling light. The eyes of these eellarvae change into more normal shaped eyes during metamorphosis (Smith 1974), which is theopposite sequence to many deep-sea fishes that develop telescopic eyes as juveniles, after alarval stage in the surface layer with normal eyes (Evans and Fernald 1990).
Fig. 18. Photographs of the anteriorgut regions of Gorgasia (A) andOphichthidae (B) showing their livertissue (LV), gall bladder (GB), thebeginning of the main part of the gut(intestine: INT), and the general areawhere there is likely a small diverticu-lum that is the equivalent of the stom-ach (ST). The two main lobes of thelivers in the ophichthid are connectedwith liver tissue.

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 21
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Olfaction is also probably available to leptocephali. All leptocephali have a pair of olfac-tory rosettes just in front of the eyes (Figs. 14, 15, 17, 20), but they do not appear to be devel-oped until the late engyodontic stage (Figs. 9, 11). These structures vary in size among speciesand sizes of leptocephali, with Thalassenchelys being an example of having a small nasal organ(Fig. 15D) and Gorgasia having a large one (Fig. 17A). It is usually positioned just in front ofthe eye, but in synaphobranchids with a very long snout, it is positioned near the end of the snout(Fig. 17B). The nasal organs of most species appear to get larger with age and can be greatlyenlarged in metamorphosing leptocephali (see Section 2-7). Some species develop nostrils thatextend outside of the body during metamorphosis, which likely remain in the juveniles andadults. It appears that the nasal organs of many species may have two openings, one anterior and
Fig. 19. Sections showing the changes in thethickness of the skin of Conger myriaster larvaeat the stages of premetamorphic leptocephalus (A),metamorphosing leptocephalus (B), andpostmetamorphosis (C) using immunohistochemicalstaining. Arrows show club cells (mostly red color);mucous cells (mc); epidermal cells (ec); dermis (dm);muscle (m). Scale bars are 10 µm. Reprinted withpermission of Wiley-Blackwell from Nakamura etal. Development of epidermal and mucosal galectincontaining cells in metamorphosing leptocephali ofJapanese conger. Journal of Fish Biology, 2002;61: 822–833. © 2002, The Fisheries Society of theBritish Isles.

22 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
one posterior, based on scanning electron microscope (SEM) observations (Appelbaum and Riehl1993). This would allow water to flow over the olfactory rosette tissue as the leptocephalusswims through the water. However, Nemichthys scolopaceus leptocephali have only one open-ing to the nasal organ (Appelbaum and Riehl 1993).
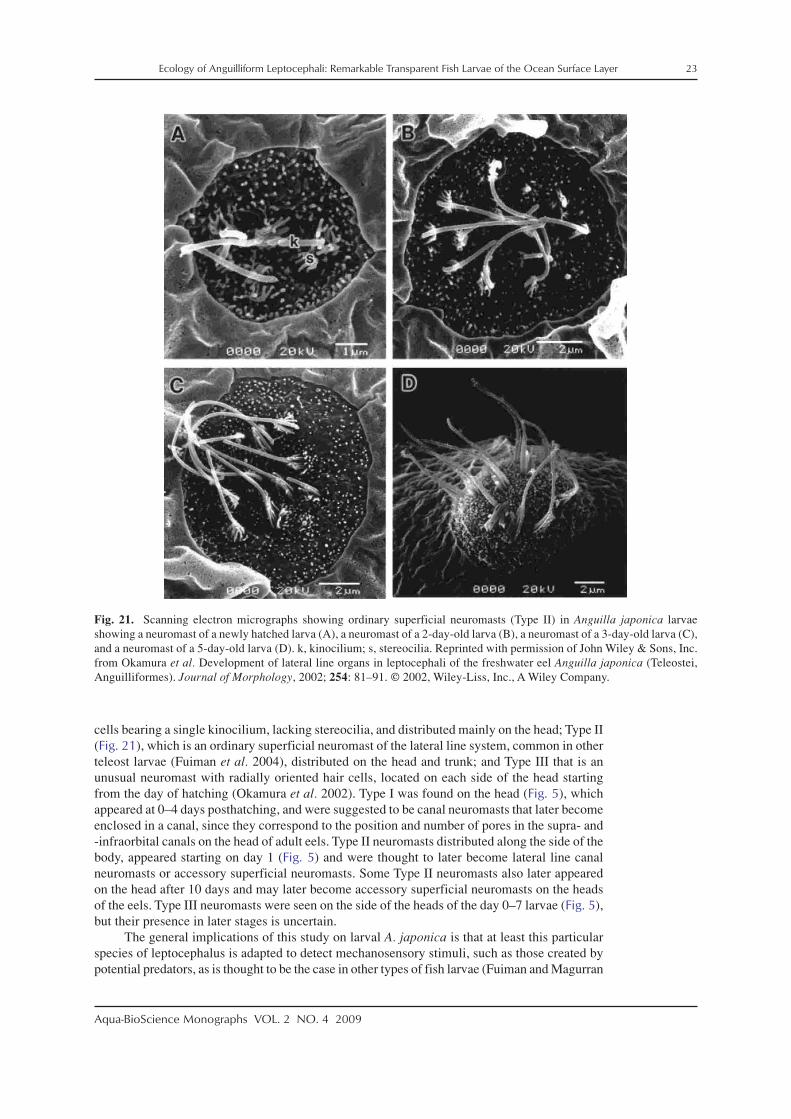
Leptocephali also appear to have a rudimentary lateral line system to provide mechanore-ception. Okamura et al. (2002) used SEM to examine the development of mechanoreceptor cellsin newly hatched larvae of A. japonica that were obtained from artificially matured and spawnedeels. Those larvae had three types of these sensory cells distributed on various parts of theirbody at different developmental stages (Fig. 5). These types were classified as Type I with hair
Fig. 20. Photographs of the head regionsshowing the nasal organ in both panelsand sensory pores in the lower jaw of ametamorphosing muraenid leptocephalusin (B). The three largest visible sensorypores are shown with black arrows, andthe velum (see Section 2-4) is shown witha red arrow.
Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 23
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
cells bearing a single kinocilium, lacking stereocilia, and distributed mainly on the head; Type II(Fig. 21), which is an ordinary superficial neuromast of the lateral line system, common in otherteleost larvae (Fuiman et al. 2004), distributed on the head and trunk; and Type III that is anunusual neuromast with radially oriented hair cells, located on each side of the head startingfrom the day of hatching (Okamura et al. 2002). Type I was found on the head (Fig. 5), whichappeared at 0–4 days posthatching, and were suggested to be canal neuromasts that later becomeenclosed in a canal, since they correspond to the position and number of pores in the supra- and-infraorbital canals on the head of adult eels. Type II neuromasts distributed along the side of thebody, appeared starting on day 1 (Fig. 5) and were thought to later become lateral line canalneuromasts or accessory superficial neuromasts. Some Type II neuromasts also later appearedon the head after 10 days and may later become accessory superficial neuromasts on the headsof the eels. Type III neuromasts were seen on the side of the heads of the day 0–7 larvae (Fig. 5),but their presence in later stages is uncertain.
The general implications of this study on larval A. japonica is that at least this particularspecies of leptocephalus is adapted to detect mechanosensory stimuli, such as those created bypotential predators, as is thought to be the case in other types of fish larvae (Fuiman and Magurran
Fig. 21. Scanning electron micrographs showing ordinary superficial neuromasts (Type II) in Anguilla japonica larvaeshowing a neuromast of a newly hatched larva (A), a neuromast of a 2-day-old larva (B), a neuromast of a 3-day-old larva (C),and a neuromast of a 5-day-old larva (D). k, kinocilium; s, stereocilia. Reprinted with permission of John Wiley & Sons, Inc.from Okamura et al. Development of lateral line organs in leptocephali of the freshwater eel Anguilla japonica (Teleostei,Anguilliformes). Journal of Morphology, 2002; 254: 81–91. © 2002, Wiley-Liss, Inc., A Wiley Company.

24 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
1994). The potential importance of this mechanosensory system for leptocephali in general issupported by observations of neuromasts located on the head of several other species ofanguilliform leptocephali that also have been observed using SEM (Appelbaum and Riehl 1993).Although these mechanoreceptors observed in leptocephali of A. japonica and other speciesusing SEM are not apparent while examining freshly caught leptocephali, large sensory poresthat presumably contain mechanoreceptors appear along the lower jaw of some species duringmetamorphosis (Figs. 11, 20B).
Future studies are needed on the sensory biology of leptocephali, but the basic observa-tions that have been made so far indicate that leptocephali have mostly similar sensory systemsof other marine fishes that enable them to detect mechanical, olfactory, and visual cues. How-ever, since anguillid glass eels, and yellow eels have been found to have a geomagnetic sense(Nishi et al. 2004; Nishi and Kawamura 2005), the function of which is most likely to guidemigrating adult eels back to their spawning area (see Tsukamoto 2009) using a geomagneticmap sense as hypothesized for other marine animals (Lohmann et al. 2008). If true, this wouldrequire that leptocephali also have the ability to imprint on the geomagnetic cues that character-ize the area where they were born. Therefore it is possible that at least some leptocephali, suchas anguillids, also have a geomagnetic sense.
2-4. Feeding ecology
What leptocephali use as a food source was a mystery for many years, and even now theirfeeding ecology is poorly known. It has been difficult to understand what leptocephali feed onbecause no identifiable objects such as small zooplankton have been seen in their guts afterbeing captured in nets and examined onboard research vessels. There is often an unidentifiablematerial inside the intestine, which can flow out the end of the gut while a leptocephalus is beingobserved after capture (Fig. 22C). However, the lack of visually identifiable organisms in theirguts and the thin layer of epidermis covering their body led to the hypothesis that leptocephaliabsorbed dissolved organic carbon (DOC) across the surface of their bodies (Pfeiler 1986;Hulet and Robins 1989). Hulet (1978) incorrectly thought that the gut was occluded and non-functional based on sections made of Ariosoma balearicum leptocephali, and the observation oftiny microvilli-like extensions of the epidermis of some leptocephali was seen as support for thedirect absorption of DOC hypothesis (Hulet 1978; Hulet and Robbins 1989). There were nosuccessful attempts to test this hypothesis, likely due to the difficulty of obtaining healthy lepto-cephali and the constraints of carrying out labeled isotope experiments that can exclude theuptake of the tracers by bacteria attached to the leptocephali.
Eventually the gut contents of leptocephali of various families that were collected in coastalregions of Japan were carefully examined, or they were directly observed to feed in the labora-tory. Leptocephali of Muraenesox cinereus were observed to eat squid paste (Mochioka et al.1993), and the red paste could clearly be seen in their guts after it was ingested (Fig. 23). Theyignored zooplankton in the tank and were not attracted to fish paste. Their feeding on squid pastestopped at metamorphosis when the gut started to move forward (Mochioka et al. 1993). Otakeet al. (1993) examined the gut contents of Conger myriaster, C. japonicus, and M. cinereus andfound zooplankton fecal pellets or smaller particles in the guts of most of the more than 600leptocephali examined. They also found that the nitrogen isotope ratio of the bodies of lepto-cephali (gut excluded) was very low, supporting the evidence found in their gut contents thatthey were feeding on particulate material and gaining their nutrition at the bacterial level of thefood chain (Otake et al. 1993). Otake et al. (1990) also suggested ciliates may have been presentin the guts of Anguilla japonica leptocephali, but this was not confirmed, and they may simplyhave been attached to ingested particulate matter. Govoni (in press) also reported ciliates in theguts of ophichthid leptocephali, so these organisms may also contribute to the diet of lepto-cephali in some areas.
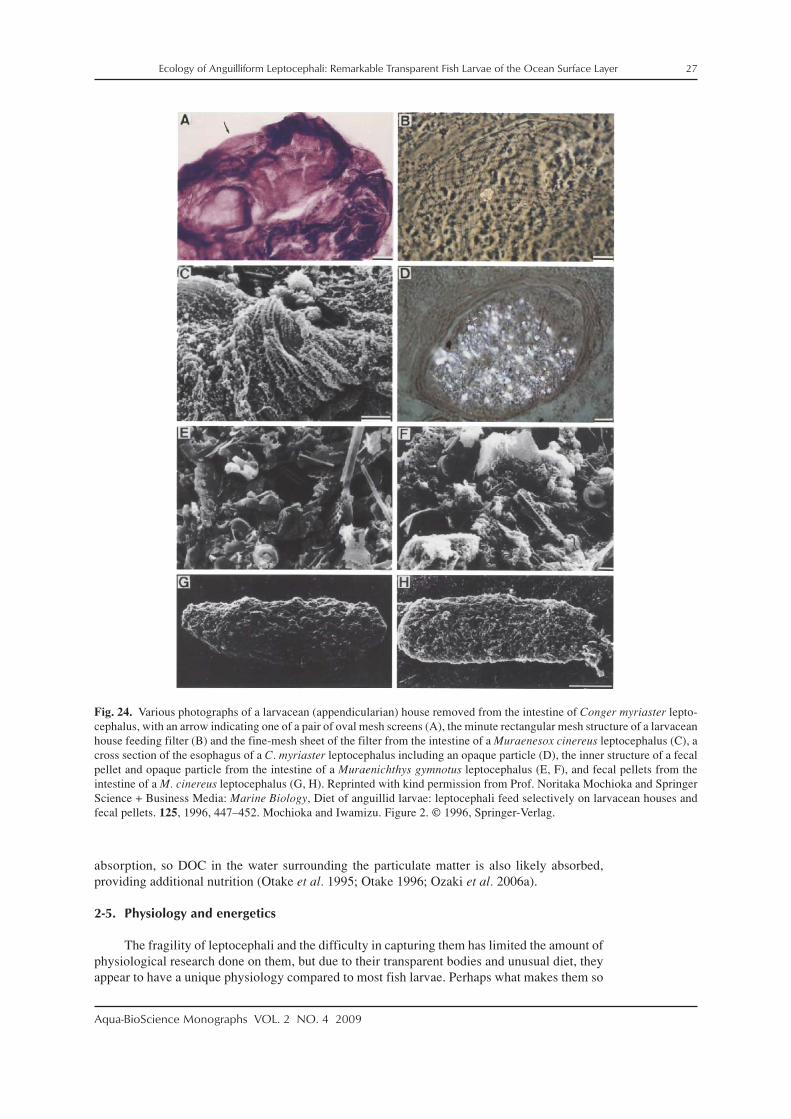
Mochioka and Iwamizu (1996) examined the guts of eight species of leptocephali(Congridae, Muraenesocidae, Muraenidae, Nettastomatidae, Ophichthidae) that were also caughtalong coastal Japan. The particles in their guts were identified as either larvacean houses, larvacean

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 25
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
fecal pellets, or zooplankton fecal pellets. More detailed SEM observations confirmed that ob-jects such as zooplankton fecal pellets and parts of discarded larvacean houses were present inthe guts of some leptocephali (Fig. 24). Mochioka and Iwamizu (1996) detected structures thatappeared to be the mesh screens (Figs. 24B, C) used as filters within the larvacean houses(Flood and Deibel 1998). Oval objects that are likely zooplankton fecal pellets are also occa-sionally seen inside the guts of leptocephali collected in offshore areas far from the higher pro-ductivity coastal waters where previous studies were conducted (Fig. 22). These types of obser-vations disproved the hypothesis that leptocephali do not feed and only get their nutrition byabsorbing DOC.
The eventual success in getting artificially spawned A. japonica leptocephali to grow un-der laboratory conditions (Tanaka et al. 2001) has provided further evidence that leptocephaliactively ingest food. Initially, some early stage leptocephali were observed to eat rotifers, butthis food source would not keep them alive very long (Tanaka et al. 1995). It was later found thatseemingly the only material that would stimulate the larvae to eat and grow was shark egg yolkpaste (Tanaka et al. 2001, 2003). Differentiation of the alimentary canal was complete by 7 daysafter hatching in A. japonica preleptocephali, indicating that they had developed the ability toabsorb food by this young age (Ozaki et al. 2006a). Other studies examined the digestive en-zymes of early stage A. japonica larvae and established the presence of trypsin in first feedinglarvae (Kurokawa et al. 1995; Kurokawa and Pedersen 2003; Pedersen et al. 2003) and in otherspecies (Kruse et al. 1996), but these studies did not reveal much about the likely natural food ofleptocephali.
Fig. 22. Photographs of the intestines of leptocephali that contain particles that appear to be fecal pellets (A, B, D), and aphotograph of the material contained in the gut of a leptocephalus that has flowed out of the gut (C).

26 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Although leptocephali will eat a prepared paste if it contains shark egg yolk, this materialis not a natural food. The yolk likely contains some chemical compound that stimulates a feed-ing response, which is positively reinforced by the nutrition provided by the food, resulting insuccessful feeding and growth. Other than the obvious possibility that the compound is relatedto what they naturally eat as larvae, another possibility is that it is related to an important fooditem they eat later in life, and they have an innate and irresistible attraction to that chemicalregardless of their life history stage, due to its significance in feeding during the juvenile stage.Fish or other types of eggs are one possibility for this, because it is well known among fisher-man that horseshoe crabs laden with eggs are the best bait for the American eel. Despite mucheffort though, it is presently unclear what special ingredient in shark egg yolk triggers a feedingresponse in larvae that seem to feed on particulate material such as marine snow or discardedlarvacean houses in the ocean.
It has been suggested that leptocephali may target larvacean houses as a food source in theocean (Westerberg 1990; Mochioka and Iwamizu 1996), but it is presently unclear whetherleptocephali are feeding specialists or if any type of available particulate matter, such as marinesnow, is sufficient as a food source. Larvacean houses were found in 80% of the guts of lepto-cephali that contained food in the study of Mochioka and Iwamizu (1996), suggesting that theywere an important part of the diet of leptocephali in that region of coastal Japan. The forwardpointing teeth appear to be well-designed to grasp larvacean houses (Westerberg 1990; Mochiokaand Iwamizu 1996) and to help squeeze the house material and any associated bacterial or otherparticles, such as fecal pellets, into the oral cavity. These forward pointing teeth also wouldenable excess material to be ejected away from the mouth to avoid fouling the teeth.
There are other morphological features that may be important for feeding by leptocephali.Many leptocephali have a velum that hangs down from the edges of the roof of the mouth (Hulet1978; Hulet and Robins 1989), which would provide a lining or a seal for the oral chamber whenthe mouth is closed (Figs. 20B, 25). This thin tissue may enable water containing a wide sizerange of particles to be forced into the esophagus. Another factor is that observations of the finestructure of leptocephalus guts have suggested that they are adapted for intestinal water
Fig. 23. A photograph of a Muraenesoxcinereus leptocephalus (familyMuraenesocidae) that just ingestedsome red colored squid paste in anaquarium. White arrow shows the endof the gut. Adapted with kind permis-sion from Prof. Noritaka Mochioka andSpringer Science + Business Media:Environmental Biology of Fishes, Lep-tocephalus eel larvae will feed inaquaria. 36, 1993, 381–384. Mochiokaet al., Fig. 1. © 1993, Kluwer Aca-demic Publishers.
Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 27
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
absorption, so DOC in the water surrounding the particulate matter is also likely absorbed,providing additional nutrition (Otake et al. 1995; Otake 1996; Ozaki et al. 2006a).
2-5. Physiology and energetics
The fragility of leptocephali and the difficulty in capturing them has limited the amount ofphysiological research done on them, but due to their transparent bodies and unusual diet, theyappear to have a unique physiology compared to most fish larvae. Perhaps what makes them so
Fig. 24. Various photographs of a larvacean (appendicularian) house removed from the intestine of Conger myriaster lepto-cephalus, with an arrow indicating one of a pair of oval mesh screens (A), the minute rectangular mesh structure of a larvaceanhouse feeding filter (B) and the fine-mesh sheet of the filter from the intestine of a Muraenesox cinereus leptocephalus (C), across section of the esophagus of a C. myriaster leptocephalus including an opaque particle (D), the inner structure of a fecalpellet and opaque particle from the intestine of a Muraenichthys gymnotus leptocephalus (E, F), and fecal pellets from theintestine of a M. cinereus leptocephalus (G, H). Reprinted with kind permission from Prof. Noritaka Mochioka and SpringerScience + Business Media: Marine Biology, Diet of anguillid larvae: leptocephali feed selectively on larvacean houses andfecal pellets. 125, 1996, 447–452. Mochioka and Iwamizu. Figure 2. © 1996, Springer-Verlag.

28 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
unique is the transparent GAG material filling their bodies, which unlike the muscle tissue thatcomprises a high proportion of the body of typical fish larvae, is not metabolically active mate-rial. As a result of this, their respiration rates are low compared to their body mass (Pfeiler andGovoni 1993; Bishop and Torres 1999), and become proportionally even lower as they increasein size by building a larger relative content of GAG material and water (Bishop et al. 2000).
In one study, the respiration rates of several species of leptocephali were determined usingelectron transport system (ETS) assays on fresh and frozen leptocephali, which included twocongrids, two ophichthids and a species of bonefish (Pfeiler and Govoni 1993). ETS activity isassumed to represent the maximum potential oxygen consumption rate (see Pfeiler and Govoni1993). This study clearly showed a decrease in wet-weight-specific ETS activity with increasein body length in premetamorphic leptocephali (Pfeiler and Govoni 1993), which was consistentwith a previous ETS activity study that included leptocephali (Schalk 1988). Bonefish lepto-cephali were also tested in respirometers, which showed that respiration rates increased duringmetamorphosis as GAG and other materials were converted into respiring tissues (Pfeiler andGovoni 1993).
Other estimates of leptocephalus respiration were obtained by testing anguilliform lepto-cephali in respirometry chambers or using enzyme assays (Bishop and Torres 1999). This studyshowed the same pattern of a decrease in wet-weight-specific respiration with increasing bodymass and water content using the respirometry chambers (Fig. 26) and in enzyme assays ofcitrate synthase and lactate dehydrogenase activities (Bishop and Torres 1999). These measure-ments in combination with estimates of excretion during the respirometry, and growth estimatesfrom otolith analyses (Bishop et al. 2000) were then used to model the energetics of these spe-cies of leptocephali (Bishop and Torres 2001). Those analyses indicated that leptocephali devotemost of their energy not to growth as is the case with most fish larvae, but to metabolism. It wasestimated that 60–92% of their total ingested energy was used for metabolism, which combinedwith their very low mass-specific respiration rates, made them physiologically very differentfrom other fish larvae.
In addition to respiration data for leptocephali, there are also basic data on their chemicalcomposition and RNA/DNA content. Pfeiler (1999) reviewed the literature on the chemicalcomposition, biology, and physiology of leptocephali, and some more recent studies have beenpublished. The gelatinous extracellular matrix of GAG comprises most of the bodies of lepto-cephali, but this matrix contains a high percentage of water that ranges from about 90–95% ofthe total body content (Pfeiler and Govoni 1993; Donnelly et al. 1995; Pfeiler 1999; Bishop etal. 2000; Wuenschel and Able 2008). The general chemical composition of leptocephali has
Fig. 25. Photographs of the heads of Avocettina (Nemichthyidae) (A) and Ariosoma (Congridae) (B) that show a velum(indicated by yellow arrows) extending downward from the roof of the mouth.

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 29
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
been examined in a few species such as the congrids, Ariosoma balearicum (Donnelly et al.1995) and Paraconger caudilimbatus, a muraenid (Gymnothorax saxicola), an ophichthid(Ophichthus gomesii) (Bishop et al. 2000), as well as in non-anguilliform bonefish leptocephali(Pfeiler 1988, 1991, 1993; Pfeiler et al. 1998, 2002). These studies have found that differentspecies of leptocephali are comprised of roughly similar proportions of protein, lipids, carbohy-drates, and nucleic acids. The GAG carbohydrate compounds are long-chain mucopolysaccharidesthat may comprise 10% or more of the dry mass. They provide both structural support for themuscle tissue needed for swimming and act as an energy storage material in preparation formetamorphosis (Pfeiler 1999; Pfeiler et al. 2002).
Pfeiler et al. (2002) recently reported that hyaluronan is the principal GAG in the bodymatrix of seven species of anguilliform and in ladyfish Elops saurus (Elopiformes) leptocephali,but it was a minor GAG component in the bonefish Albula sp. (Albuliformes). Previous reportsthat chondroitin and chondroitin sulfate were the primary eel GAG compounds (Pfeiler 1993)may have been incorrect due to technical difficulties in accurately identifying them (Pfeiler etal. 2002). Trace amounts of unsaturated disaccharides of chondroitin sulfate were also presentin the more recent study of eel leptocephali (Pfeiler et al. 2002). In addition to its presumedrole in maintaining the structural integrity and hydration of the gelatinous body of the lepto-cephalus, hyaluronan was postulated to function as a storage polysaccharide in those species inwhich it is the predominant GAG. In bonefish leptocephali, keratan sulfate appears to be thedominant form of GAG, with hyaluronan and chondroitin sulfate being minor components(Pfeiler et al. 1988, 1993).
Most biochemical and physiological research on leptocephali has been done on bonefishes(genus Albula) as they approach recruitment age and begin to metamorphose (Pfeiler 1986,1997, 2001; Padrón et al. 1996; Pfeiler et al. 1998). How precisely these findings apply toanguilliform leptocephali is unclear. Studies on some species of anguilliform leptocephali in thewestern North Atlantic have found that they have high blood osmolalities compared to most
Fig. 26. Plots of the respiration rates offour species of leptocephali collected inthe Gulf of Mexico by nets and quicklytested in respirometers on board the re-search vessel. Reprinted with permissionfrom Bishop and Torres. The Journal ofExperimental Biology, 1999. 202: 2485–2493. © 1999, Company of Biologists.

30 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
marine fishes, but have values below that of seawater (Hulet et al. 1972; Hulet and Robins1989). These values ranging from roughly 500–1000 mOsm kg–1 H2O suggest that somepremetamorphic leptocephali are closer to being isotonic with seawater than any other fishes(Hulet et al. 1972; Hulet and Robbins 1989). Mean blood osmolalities of premetamorphicA. balearicum samples, for example, were 792–910 mOsm kg–1 H2O, compared to a value ofabout 1058 kg–1 H2O for seawater (Hulet and Robbins 1989). Blood osmolalities of Rhynchocongerflavus may tend to decrease from about 804 to 611 mOsm kg–1 H2O as leptocephali increase insize from 50 to 100 mm, but the relationship between size and osmolality was not clear, and wasobtained from specimens collected during different cruises (Hulet and Robbins 1989). Otheranguilliform leptocephali showed osmolalities in a similar range, with the lowest value recordedfrom a nettastomatid (483), and the highest from a congrid (1057) (Hulet and Robbins 1989).Tsukamoto et al. (2009) reported a mean value of 450 mOsm kg–1 H2O for the extracellularmatrix of A. japonica leptocephali that were reared in the laboratory, and suggested that this lowosmolality is likely an important factor in their buoyancy.
Leptocephali have no swim bladder, but they appear to have high buoyancy compared tomany other marine organisms that live in the surface layer of the open ocean (Tsukamoto et al.2009). Eels do not develop a swim bladder until after metamorphosis near the end of the glassstage, based on observations of an anguillid (Zwerger et al. 2002), so their lipid content, highwater content, and almost complete lack of ossified structures may contribute to their buoyancy(Pfeiler 1999). A comparative study of the specific gravity of 25 taxa of 7 phyla of zooplanktonand 6 taxa of leptocephali found that leptocephali were among the species with the lowest spe-cific gravities and highest buoyancies (Tsukamoto et al. 2009). Most ocean-collected lepto-cephali had lower specific gravities (1.028–1.032) than a variety of crustaceans and pelagicmollusks (~1.040–1.450 for 13 taxa), but had values similar to the gelatinous zooplankton, in-cluding jellyfish, salps, ctenophores, and chaetognaths (~1.020–1.034 for 8 taxa).
Buoyancy of leptocephali during their ontogeny was also examined by Tsukamoto et al.(2009) using artificially spawned and reared eggs and larvae of A. japonica. Preleptocephali hadvery low specific gravities, probably due to their oil globule, but once the globule was absorbedand the larvae were ready to initiate feeding, their specific gravities increased (Fig. 27). As theleptocephali grew and accumulated higher proportions of extracellular matrix material, theirspecific gravities decreased steadily. During metamorphosis however, their specific gravitiesrapidly increased again, and the glass eel stage showed the highest values and were negativelybuoyant (Fig. 27). This apparent ontogenetic change in the buoyancy of A. japonica was hypoth-esized to have some adaptive value for this species (Tsukamoto et al. 2009), because the eggsand preleptocephali would tend to float towards shallow depths where food is more abundant,but as they grow they need to be free to migrate vertically on a diel cycle. As they grow neartheir maximum size, however, high buoyancy would tend to assist their migration back to theirrecruitment areas.
Chloride cells scattered over the surface of the body (Fig. 28A; Sasai et al. 1998; Kanekoet al. 2003, 2008) probably help maintain buoyancy by sustaining osmolalities lower than seawater(Tsukamoto et al. 2009). Apparent chloride cells have also been observed in Ariosoma balearicumand Anguilla rostrata leptocephali (Leonard and Summers 1976; Hulet 1978) and in bonefishlarvae (Pfeiler and Lindley 1989). In freshly caught undamaged leptocephali, small objects canbe seen scattered over the integument that may be chloride cells (Fig. 28B), but this needs to beconfirmed. Other fish larvae have osmoregulation abilities (Varsamos et al. 2005) and typicallyuse a swim bladder to obtain buoyancy, or sometimes have other structures such as expandedfins or fin folds to help reduce sinking (Moser 1981).
2-6. Growth of leptocephali
Growth rates of leptocephali have been estimated primarily from change in body sizeamong samples collected at different times and from analysis of otolith microstructure.Smith (1989a) was able to observe changes in size of the leptocephali of some species indifferent seasons in the Gulf of Mexico, and apparent growth of cohorts of some species can beseen in the catch data of Blache (1977). But examples of this type of data being available are

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 31
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
rare. The other method that has been used recently is the examination of increments in thesagittal otoliths of leptocephali, which are likely to be deposited daily as they are in other fishlarvae under normal circumstances (Campana and Neilson 1985).
Several studies have validated that daily rings are deposited in the otoliths of anguillidglass eels that have recruited to coastal areas in both temperate (Tsukamoto 1989; Martin 1995;Cieri and McCleave 2001) and tropical species (Arai et al. 2000; Sugeha et al. 2001a). Dailydeposition was indicated in a study of artificially spawned and reared A. japonica early stageleptocephali (Umezawa et al. 1989; Shinoda et al. 2004). Although there have been no valida-tion studies on larger premetamorphic stage leptocephali, all species of anguillid leptocephalithat have been examined from tropical or subtropical latitudes have otolith increments that canbe clearly seen in SEM imagery of otolith cross sections (Fig. 29). Clear increments have alsobeen observed in other taxa of anguilliform leptocephali such as Dysomma and Saurenchelys(Ma et al. 2005), and in Conger leptocephali (Lee and Byun 1996; Correia et al. 2002a, 2003,2004) at least during the time periods that the larvae were living in warm-water regions. Thesestudies suggest that leptocephali that are feeding and growing normally deposit daily otolithincrements.
Fig. 27. Ontogenetic changes in specific gravity of Anguilla japonica during its early life history. The specific gravity valuesof 6 batches of eggs (about 1 mm in diameter) that hatched 46 hours after fertilization at 22°C (green circles). The specificgravity values of preleptocephali are means of 10 specimens of 6 batches (light blue triangles). Symbols of the other stagesare leptocephalus (blue diamonds), metamorphosing leptocephalus (purple triangles) and glass eels (red circles), that repre-sent individuals from at least 3 larval batches. The two horizontal broken lines indicate the specific gravity of seawater at26°C (upper: 1.023) and 22°C (lower: 1.024), which roughly correspond to the water temperatures at depths of 100 and 200m in the North Equatorial Current (15°N, 140°E), respectively. Reprinted with kind permission from Springer Science +Business Media: Marine Biology, Positive buoyancy in eel leptocephali: an adaptation for life in the ocean surface layer. 156,2009, 835–846. Tsukamoto K, Yamada Y, Okamura A, Tanaka H, Miller MJ, Kaneko T, Horie N, Utoh T, Mikawa N, TanakaS. Figure 2. © 2009, Springer-Verlag.

32 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Additional evidence of daily increment deposition throughout the leptocephalus stage hasbeen obtained from back-calculated hatching date analyses in some species that indicate lunarspawning patterns. Studies on A. japonica leptocephali used the back-calculated hatching datesestimated from their otolith increments to formulate the hypothesis that the silver eels spawnonly during new moon periods within their spawning season (Ishikawa et al. 2001; Tsukamotoet al. 2003). This hypothesis has been subsequently validated by collections of recently hatchedpreleptocephali just after new moon periods in the spawning area during several different years(Tsukamoto 2006, 2009). A similar pattern was observed in the back-calculated hatching datesof a wide size range of Kaupichthys leptocephali (Chlopsidae) collected around Sulawesi Islandin the central Indonesian Seas (Fig. 30; Lee et al. 2008). However, estimated hatching dates of
Fig. 28. Extrabranchial chloride cells in an Anguilla japonica larva (10 days after hatching), detected by whole-mountimmunocytochemistry with anti-Na+/K+-ATPase fluorescently labeled and observed with a confocal laser scanning micro-scope (A). Chloride cells are distributed over the body surface of the larva, which lacks functional gills and gill chloride cells.Reprinted with kind permission from Springer Science + Business Media: Marine Biology, Positive buoyancy in eel lepto-cephali: an adaptation for life in the ocean surface layer. 156, 2009, 835–846. Tsukamoto K, Yamada Y, Okamura A, TanakaH, Miller MJ, Kaneko T, Horie N, Utoh T, Mikawa N, Tanaka S. Figure 4. © 2009, Springer-Verlag. (Original photographsprovided by Toyoji Kaneko of The University of Tokyo); A photograph of the anterior body region of a serrivomeridpreleptocephalus (see Fig. 7C for whole body photograph) showing small whitish spots on the skin of the dorsal surface of thebody, which could be chloride cells (B).

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 33
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
this genus corresponded to full moon periods. This difference is likely related to the contrastingenvironments (subtropical open ocean versus coastal tropical seas) in which these two types ofeels spawn (Lee et al. 2008). The common lunar spawning patterns supports the hypothesis thatotolith rings are deposited daily in leptocephali, because if rings were not deposited daily, thepatterns of lunar spawning could not be detected in otoliths.
The clear increments in otoliths of several species of eel leptocephali (Fig. 29) have beenused to estimate their larval growth rates. These studies have found that anguillid leptocephaliappear to grow at mean rates from about 0.23 mm/d for Atlantic eels that experience lower watertemperatures during their early growth, to much faster growth rates of up to 0.61 mm/d in the
Fig. 29. Scanning electron microscope micrographs of cross sections of the otoliths of leptocephali of Anguilla celebesensis(37.5 mm) (A), Anguilla borneensis (42.0 mm) (B), and Saurenchelys stylura (42.0 mm), which show the likely daily otolithgrowth increments. HC: hatching check, FFC: first feeding check (C). (A, B) adapted with permission from Marine EcologyProgress Series, 309, Kuroki M, Aoyama J, Miller MJ, Wouthuyzen S, Arai T, Tsukamoto K. Contrasting patterns of growthand migration of tropical anguillid leptocephali in the western Pacific and Indonesian Seas. 233–246, 2006, Figures 5c, d.© 2006, Inter-Research; (C) reprinted with permission of Wiley-Blackwell from Ma T, Miller MJ, Shinoda A, Minagawa G,Aoyama J, Tsukamoto K. Age and growth of Saurenchelys (Nettastomatidae) and Dysomma (Synaphobranchidae) lepto-cephali in the East China Sea. Journal of Fish Biology, 2005; 67: 1619–1630. © 2005, The Fisheries Society of the BritishIsles.

34 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
tropical species A. celebesensis (Table 1). Castonguay (1987) examined the age and growth ofthe leptocephali of two Atlantic anguillids in the Sargasso Sea, and there have been studies onthe leptocephali of the other species of northern temperate anguillid species, A. japonica(Tsukamoto et al. 1992, 2003; Ishikawa et al. 2001) and A. australis (Kuroki et al. 2008a).There have also been studies on the age and growth of tropical species of anguillid lepto-cephali in the central Indo–Pacific (Kuroki et al. 2005, 2006a,b, 2007), and off west Sumatra(Kuroki et al. 2007). Some species for which only a few specimens have been examined forgrowth rates, such as A. interioris (0.46–0.54 mm/day, Kuroki et al. 2006b), and A. obscura andA. megastoma (0.34–0.49 mm/day, Kuroki et al. 2008a) are not shown in Table 1.
All mean growth rate values obtained from Indo–Pacific anguillid leptocephali have beenbetween 0.35 and 0.61 mm/day (Table 1), but it appears that individual growth rates slowdown as the larvae reach full size and begin to metamorphose. For example, A. marmorata andA. bicolor pacifica leptocephali reached a maximum size of about 50 mm and then appeared tostop increasing in total length even though their ages continued to increase (Fig. 31). A similarreduction in growth was seen in Kaupichthys leptocephali in the Indonesian Seas as they reachedearly metamorphosis (Fig. 30; Lee et al. 2008). It is possible that they may stop increasing intotal length if they reach their maximum size while still offshore away from the appropriate cuesto trigger metamorphosis.
Fig. 30. The relationship between total length and ageof the Kaupichthys leptocephali collected aroundSulawesi Island in September 2002 that was estimatedfrom their otolith increments. The leptocephali that mayhave begun the process of metamorphosis based on theirotolith increment widths are shown with black circles.The Von Bertalanffy’s growth curve that was fitted tothe data is shown. Reprinted with kind permission fromSpringer Science + Business Media: Marine Biology,Distribution and early life history of Kaupichthys lepto-cephali (family Chlopsidae) in the central IndonesianSeas, 153, 2008, 285–295. Lee TW, Miller MJ, HwangHB, Wouthuyzen S, Tsukamoto K. Figure 8. © 2008,Springer-Verlag.
Table 1. Mean growth rates (GR), sizes (total length: TL), and ages of leptocephali from the Indo–Pacific that have beenstudied using SEM observations of their sagittal otoliths. Mean growth rates are a mixture of values estimated from regres-sions of age and TL, or means of individual growth rates. Some large leptocephali that had reached their maximum sizes withslower growth rates were excluded. Maximum growth rates of individual leptocephali in each study (Max. Ind-GR) are alsoshown. WNP = western North Pacific. N = number of leptocephali examined for otolith microstructure.
Species by Region Area collected N TL range Age GR Max. References(mm) (d) (mm/d) Ind-GR
Anguilla japonica WNP 17 10.0–37.0 12–73 0.46 0.68 Ishikawa et al. (2001)Anguilla bicolor pacifica WNP/Indonesian Seas 43 24.0–54.1 40–128 0.42 0.62 Kuroki et al. (2006a)Anguilla marmorata WNP/Indonesian Seas 42 8.0–50.7 22–137 0.48 0.65 Kuroki et al. (2006a)Anguilla marmorata WSP 12 19.0–50.9 25–155 0.47 0.64 Kuroki et al. (2008a)Anguilla borneensis Indonesian Seas 5 8.5–49.0 20–106 0.55 0.57 Kuroki et al. (2006a)Anguilla celebesensis Indonesian Seas 40 12.3–47.8 17–110 0.61 0.74 Kuroki et al. (2006a)Anguilla bicolor bicolor E. Indian Ocean 34 44.1–55.5 114–158 0.35 0.39 Kuroki et al. (2007)Anguilla australis WSP 15 20.5–48.3 32–150 0.45 0.62 Kuroki et al. (2008a)Anguilla reinhardtii WSP 10 20.0–34.0 28–62 0.60 0.68 Kuroki et al. (2008a)Conger myriaster Eastern Japan 68 78.9–107.7 111–239 0.54–0.68 0.82 Kimura et al. (2004)Dysomma sp. E. China Sea 22 8.4–33.5 17–66 0.44 0.53 Ma et al. (2005)Saurenchelys stylura E. China Sea 21 10.0–48.6 16–75 0.68 0.83 Ma et al. (2005)Kaupichthys hyoproroides Indonesian Seas 24 24.7–60.8 39–161 0.46–0.65 0.67 Lee et al. (2008)

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 35
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
These studies also indicate that there are some differences in growth patterns betweentemperate and tropical anguillid leptocephali, which may be related to their different scales ofmigration. One obvious difference is that the maximum size of tropical leptocephali (50–55mm; Jespersen 1942; Kuroki et al. 2009b) is less than that of temperate species (typically about70 mm) that can reach about 85 mm in A. anguilla, which has the longest larval migration(Tesch 1980; Boëtius and Harding 1985; Kleckner and McCleave 1985; Bast and Strehlow 1990).Kuroki et al. (2006a) compared the growth rates of four different tropical anguillid leptocephali(Fig. 31) that recruit to habitats in the central Indonesian Seas. The leptocephali of the twospecies with short migrations from local spawning areas, A. borneensis and A. celebesensis(Aoyama et al. 2003), were found to have slightly higher growth rates than A. marmoratathat migrates from an offshore spawning area in an overlapping region with A. japonica(Kuroki et al. 2009a), and A. bicolor pacifica which is thought to spawn outside of the Indone-sian Seas (Kuroki et al. 2006a). These different growth rates were hypothesized to be re-lated to the distance of the larval migration of each species, with temperate species having theslowest growth rates. The influence of temperature is difficult to exclude in most comparisons,but in one case where similar sized leptocephali of a tropical (A. reinhardtii) and a temperate(A. australis) species were collected at the same stations to the south of the Solomon Islands inthe western South Pacific, the tropical species had a higher growth rate even though both specieslikely had experienced the same water temperatures (Kuroki et al. 2008a). Shiao et al. (2002)also suggested that A. reinhardtii had faster larval growth rates than A. australis based on exami-nations of the otolith microstructure of glass eels of the two species.
Marine eel leptocephalus otolith studies have found even greater variability in growthrates among different species or regions (Table 1). A study of the age and growth of two speciescollected during the KT-00-16 survey in the East China Sea (Fig. 31; Miller et al. 2002a)found different growth rates between species (Fig. 32). The otolith increments in these twospecies indicated that Saurenchely stylura had a mean growth rate of 0.68 mm/d compared to0.44 mm/d in Dysomma sp. (Ma et al. 2005). The maximum size of S. stylura (~150 mm) islikely considerably larger than that of Dysomma sp. (~80 mm), which is one possible explana-tion for its apparent faster growth rate even though they were collected at mostly the samestations (Fig. 32). Kimura et al. (2004) found that Conger myriaster leptocephali before meta-morphosis had mean growth that ranged from 0.54–0.68 at three different sites of eastern Japan.
Analyses of the otolith microstructure of the four species of leptocephali studied in theeastern Gulf of Mexico (muraenid, Gymnothorax saxicola, and ophichthid Ophichthus gomesii,
Fig. 31. Plots of the ages and total lengths of tropical species of anguillid leptocephali from the western Pacific and centralIndonesian Seas based on their otolith increments, showing Anguilla marmorata (triangles), Anguilla bicolor pacifica (cir-cles), Anguilla celebesensis (crosses) and Anguilla borneensis (diamonds), with metamorphosing leptocephali (M) and glasseels (G) of some species indicated by letters within the symbols. Reprinted with permission from Marine Ecology ProgressSeries, 309, Kuroki M, Aoyama J, Miller MJ, Wouthuyzen S, Arai T, Tsukamoto K. Contrasting patterns of growth andmigration of tropical anguillid leptocephali in the western Pacific and Indonesian Seas. 233–246, 2006, Figure 6. © 2006,Inter-Research.

36 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
two congrids Ariosoma balearicum, Paraconger caudilimbatus) found highly variable apparentgrowth rates among individuals within each species, with very fast mean growth rates ofgreater than 1 mm/d in all of the species (Bishop et al. 2000). The estimated growth rates ofP. caudilimbatus leptocephali were lower and less variable than the other species though, andtheir growth rates slowed down with increased size as has been observed in Indo–Pacific spe-cies. Growth rates of the other species in that study were much more variable, and if the ageswere accurate, would indicate very rapid growth rates (>2–3 mm/d). These leptocephali werenot collected during the same sampling survey as in the study by Ma et al. (2005), but werecollected during several surveys (Bishop et al. 2000) with potentially differing water tempera-ture conditions.
There are no published studies on the otolith microstructure of muraenids and ophichthidsfrom the Indo–Pacific that can help evaluate the aging results of Bishop et al. (2000), but ob-servations of large Ariosoma major leptocephali from the western North Pacific indicatethat at least some Ariosoma otolith increments may be difficult to accurately discern and count(A. Shinoda pers. comm.). Basic examinations of the age data for A. balearicum and O. gomesii(Bishop et al. 2000) for example, indicate that in the otoliths of some individuals of these twospecies (particularly in the 24–44 and 20–25 increment number ranges, respectively), they hadthe same number of otolith increments, but their body lengths varied by as much as about75–80 mm or more. It is unclear if such wide variation in size of individuals of apparently thesame age could be due to differences in temperatures they experienced, feeding success, orsome other factors. Therefore, it is difficult to determine if such apparently high growth rates insome of the specimens of each these species of leptocephali from the Gulf of Mexico are
Fig. 32. Map showing the sampling stations of the KT-00-16 survey for leptocephali in the East China Sea region from27 November to 7 December 2000 (left panel). Locations where leptocephali of both Saurenchelys and Dysomma werecollected are shown by solid circles, and circles filled in the left half indicate that only the former species was collected, andcircles filled in the right half indicate only the latter species was collected (open circles show stations with neither species).Right panels show the relationship between age and total length based on the otolith increments of the leptocephali ofSaurenchelys stylura and Dysomma sp. (probably D. anguillare). Adapted with permission of Wiley-Blackwell from Ma T,Miller MJ, Shinoda A, Minagawa G, Aoyama J, Tsukamoto K. Age and growth of Saurenchelys (Nettastomatidae) and Dysomma(Synaphobranchidae) leptocephali in the East China Sea. Journal of Fish Biology, 2005; 67: 1619–1630. © 2005, The Fisher-ies Society of the British Isles.

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 37
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
accurate or not, since they are significantly higher than those estimated by the otolith incre-ments of leptocephali of other species.
2-7. Metamorphosis
One of the remarkable features of anguilliform leptocephali is their drastic morphologicaltransformation during metamorphosis into the glass eel or elver stage. Such a major change inbody shape and structure is required because of the very different habitats and food materialsused by eels compared to their larvae. The laterally compressed and transparent bodies of lepto-cephali are adapted for a pelagic life in the surface layer of the ocean where they feed on particulatematter and must avoid predators. Once they transform into young eels they enter a wide range ofbenthic marine habitats, move into estuaries or freshwater in the case of Anguilla, or live in themesopelagic zone, where the fragile leptocephalus body form and feeding strategy would not beappropriate.
Several external morphological changes occur in leptocephali at the beginning of meta-morphosis such as a thickening of the head, enlargement of the olfactory organ, and a loss of theteeth (Figs. 20B, 33, 34). In many nettastomatid leptocephali the olfactory rosette becomes veryenlarged and elongate during metamorphosis (Smith and Castle 1982). In some metamorphos-ing muraenid leptocephali, the gills become larger and are visible (Fig. 34), whereas no gills aretypically seen in premetamorphic leptocephali (Figs. 14, 15). The other major changes thatoccur during metamorphosis are that the gut and the origins of the dorsal and anal fins moveforward, as can be seen in Ariosoma gilberti from the eastern Pacific (Fig. 35) and in Congeroceanicus from the Atlantic coast of New Jersey (Fig. 36). Because the gut is moving forwardwith tissue being reabsorbed, the larvae do not feed during metamorphosis (Otake 2003). Therelative position of the gut as it moves forward typically has been used as an indicator of themetamorphic stage of leptocephali (e.g. Lee and Byun 1996; Otake et al. 1997; Bell et al. 2003;Kimura et al. 2004). The sensory pores on the head and the nostrils of some species begin toform during the metamorphosing leptocephalus stage (Figs. 11, 20B, 33). In species withmarkedly different head shapes such as the mesopelagic Nemichthys that have long curvedjaws, the jaws appear to rapidly extend forward even when the larvae are still in the leptocepha-lus stage (Fig. 33B). This also occurs in serrivomerid leptocephali (Beebe and Crane 1936;Miller and Tsukamoto 2004). A major enlargement of the olfactory organ is apparent in themetamorphosing synaphobranchid in Fig. 33A. Red blood cells begin to form in at least somelate stage metamorphosing leptocephali such as Derichthys serpentinus (Fig. 37B), or during theglass eel stage in Conger myriaster collected in coastal waters of southern Japan that metamor-phosed while held in an aquarium (Fig. 37A). Toward the end of metamorphosis when the youngeels are about to start feeding again, there is increased ossification of the skull and vertebralcolumn, and this increased development of the skeleton generally coincides with depletion ofthe gelatinous support matrix (Pfeiler 1999).
What exactly causes metamorphosis to begin in leptocephali is not known (Otake 2003),but it is generally thought to occur after a certain size required for metamorphosis is reached andenvironmental cues trigger the process. A basic age or size may be important for mesopelagiceel larvae, since they may be able to metamorphose anywhere in deep water and successfullybegin their juvenile phase. But in the case of eel larvae recruiting to shallow marine habitats orcoastal areas, metamorphosis before nearing a suitable juvenile habitat could result in starvationand death, or being eaten by predators, since young eels are likely poorly adapted to live in theopen ocean. This suggests that there are environmental cues such as chemical compounds, bot-tom depth, or salinity, which may act as triggers to initiate metamorphosis when leptocephaliapproach continental shelves or islands. In the absence of such cues, most leptocephali maydelay metamorphosis, as has been indicated in other types of fish larvae such as Acanthuridaeand Labridae (Victor 1986; McCormick 1999).
A careful examination of the geographic distribution of most species of metamorphosingleptocephali has not been done. European eel leptocephali have been found to metamorphoseinto glass eels in waters deeper than 1000 m as they approach Europe (Antunes and Tesch 1997),

38 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
so the trigger for their metamorphosis is unclear. Collections of metamorphosing American eelleptocephali are limited, with a few being collected far offshore and some over or near the edgeof the continental shelf (Kleckner and McCleave 1985). Similarly, some metamorphosing lepto-cephali of A. japonica have been collected offshore along the western margin of the PhilippineSea (Arai et al. 1997; Tsukamoto 2006). By the time they enter the Kuroshio Current south of
Fig. 33. Photographs of the head regions of metamorphosing leptocephali of an 88.0 mm Dysomma (Ilyophinae,Synaphobranchidae) (A), a 243 mm Nemichthys (Nemichthyidae) (B), and a 74.3 mm Chlopsidae (C). Nostrils can be seen in(A) and (C), and the jaws have begun to extend in the nemichthyid leptocephalus before completing metamorphosis (B).

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 39
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Japan, some have almost completed their metamorphosis into glass eels (Otake et al. 2006), sometamorphosis in anguillids may be triggered well before they arrive at the continental shelves.Miller et al. (2006b) pointed out that the biggest catches of metamorphosing Ariosoma scheeleileptocephali in a survey around Sulawesi Island were made at stations that had a shallow layerof lower salinity water near the surface, and questioned if this could be a possible cue for trig-gering metamorphosis. Chen et al. (2008) suggested that the lower salinity of coastal waterscould be important to trigger metamorphosis in tarpon larvae.
Regardless of the trigger, thyroid hormones appear to be important in the process of meta-morphosis (Otake 2003). An abrupt increase in thyroid hormones was observed in C. myriasterleptocephali during the onset of metamorphosis (Yamano et al. 1991). Similarly, thyroid hor-mones were not detected in artificially reared A. japonica leptocephali until metamorphosis(Yamano et al. 2007). Experiments using a thyroid hormone inhibitor with tarpon leptocephalishowed that metamorphosis would not proceed when thyroid hormones were blocked (Shiaoand Hwang 2006), indicating they are a key component of metamorphosis in leptocephali. Con-versely, cortisol levels were high in premetamorphic C. myriaster leptocephali, but then de-creased to low levels throughout metamorphosis, suggesting that cortisol may be involved inmaintaining the premetamorphic state (Yamano et al. 1991). This suggests that the initiation ofmetamorphosis requires a decrease in cortisol production as thyroid hormones increase. Lowtemperature may also be able to delay metamorphosis based on findings with tarpon larvae(Chen et al. 2008).
Fig. 34. Photograph of the head of a 59.3 mm metamorphosing Uropterygiinae leptocephalus of the family Muraenidae,showing a large nasal organ, developing gills, and a large sagittal otolith.

40 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Although the factors that initiate metamorphosis are difficult to determine, studies onotolith microstructure of some species of leptocephali and glass eels have revealed informationabout the possible age at metamorphosis and duration of metamorphosis. Otolith microstructureof metamorphosing Anguilla japonica (Arai et al. 1997), A. marmorata (Kuroki et al. 2005) andC. myriaster leptocephali (Otake et al. 1997), and glass eels of A. japonica (Otake et al. 1994;Chen and Tzeng 1996; Arai et al. 1997) and the Atlantic eels (e.g. Wang and Tzeng 2000) havebeen examined using SEM observations of increments and Sr:Ca ratio measurements. Thesestudies suggested that there was a rapid increase in otolith increment widths during metamor-phosis that was accompanied by a decrease in Sr:Ca ratios (Fig. 38). Using these otolith charac-teristics, the timing of metamorphosis of the leptocephalus stage has been estimated in glass eelscollected in coastal waters for both temperate and tropical anguillid glass eels (Marui et al.2001; Otake 2003). Mean ages at metamorphosis for various anguillid species have been re-ported to range from less than a 100 days in A. celebesensis (Arai et al. 2001) to almost 250 daysin A. dieffenbachii (Marui et al. 2001). The other tropical and temperate species of glass eels thathave been examined generally have larval durations within this range (see Aoyama 2009 for arecent review), but the European eel may have a longer larval duration than any other species(see Kuroki et al. 2008b). The same pattern of increment widths during metamorphosis has beenobserved in the Atlantic species of Conger, whose ages at metamorphosis have been estimatedusing this method (Correia et al. 2003, 2004) and in a species of muraenid (Ling et al. 2005).
The number of increments in the zone of increased increment widths has also been used toestimate the duration of metamorphosis using glass eel otoliths. This duration is likely depend-ent on temperature based on metamorphosing tarpon larvae (Chen et al. 2008) and comparisonsof tropical and temperate anguillid larvae (Otake 2003). The apparent durations of metamorpho-sis in anguillid larvae have ranged from 9 days in a tropical species to more than 60–70 days in
Fig. 35. Drawings of various developmental stages of Ariosoma gilberti from the eastern Pacific ranging from a leptocepha-lus before metamorphosis to an elver after metamorphosis. Triangles show the anterior positions of the dorsal and anal fins,which move much farther forward along with the gut (just in front of anal fin) during metamorphosis. The scale bars show5 mm, but not all drawings are proportional in size. Modified from Raju SN. Congrid Eels of the Eastern Pacific and Key toTheir Leptocephali. NOAA Technical Report NMFS 22, U.S. Department of Commerce. 1985; 1–19.

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 41
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
temperate species (see Marui et al. 2001; Otake 2003; Aoyama 2009).The hypothesis that changes in the otolith can be used to determine the timing of metamor-
phosis has been supported by several studies that examined different stages of larvae before andafter metamorphosis. Kuroki et al. (2005) showed that the increase in increment width startedduring metamorphosis in the leptocephalus stage of A. marmorata, along with a decrease inSr:Ca ratios. By comparing premetamorphic leptocephali to a metamorphosing specimen andoceanic glass eels (see Fig. 39), it was found that there was an increase in increment width in themetamorphosing leptocephalus, and an increase followed by a decrease in the oceanic glasseels. Lee and Byun (1996) found the same pattern of increment width increase in metamorphos-ing C. myriaster leptocephali, but not in premetamorphic individuals. In glass eels of A. marmorataand other anguillid species, the increment widths decrease even further by the time they recruitto coastal areas and are collected (Marui et al. 2001; Kuroki et al. 2005). An increase in incre-ment width during metamorphosis was also found in tarpon leptocephali held in the laboratory(Shiao and Hwang 2006; Chen et al. 2008). Powles et al. (2006) showed that otoliths of ophichthidleptocephali, Myrophis punctatus, continued to deposit increments during metamorphosis, butthey did not analyze increment widths. No increase in increment widths were observed in anguillidglass eels that recruited to Iceland however, which may have been due to the cold temperaturesand slow metamorphosis experienced by those individuals during their leptocephalus stage
Fig. 36. Drawings of three stages of Conger oceanicus larvae that recruited to estuarine habitats in New Jersey of the US eastcoast. Modified with permission of the American Society of Ichthyologists and Hepetologists from Bell GW, Witting DA,Able KW. Aspects of metamorphosis and habitat use in the conger eel, Conger oceanicus. Copeia 2003; 2003: 544–552.

42 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
(Kuroki et al. 2008b). In general though, it appears that the increase in increment widths inlarval eel otoliths is a useful marker for the timing of onset of metamorphosis that can be used tocompare the larval durations among different eel species, as illustrated by Marui et al. (2001).
2-8. Swimming behavior
The swimming ability of leptocephali has been poorly documented, but they appear to bevery capable swimmers. The ability to swim is important for migrations to recruitment areas,speed of vertical migrations, and for being abile to avoid predators. General observations ofhealthy leptocephali held in aquaria suggest they are good swimmers (Miller and Tsukamoto2004) and are able to burst swim for at least short periods. A recent nighttime video observationof an ophichthid leptocephalus swimming over deep-water off the island of Hawaii (video avail-able at: http://www.australianmuseum.net.au/movie/Ophichthid-leptocephalus-Kona-Hawaii/)also showed a very active swimming style that included rapid changes of swimming direction(MJ Miller and MJ D’Avella unpublished manuscript). Because of their use of anguilliformswimming movements (Gray 1933; D’Août and Aerts 1999), they also are able to swim bothforwards and backwards. Artificially spawned and reared Anguilla japonica leptocephali of 30–60 mm lengths were negatively phototaxic, swimming at speeds of 3.6 ± 2.7 cm s–1 horizon-tally and 2.8 ± 1.1 cm s–1 upwards away from a sudden light stimulus (Yamada et al. 2009).Their ability for long term sustained swimming, such as during the long migration of A. anguillaleptocephali recruiting to Europe, is unknown though, and has been a matter of considerablespeculation (McCleave 1993; McCleave et al. 1998).
Wuenschel and Able (2008) measured critical swimming speed (Ucrit) of recently recruitedConger leptocephali and glass eels and anguillid glass eels in a laboratory swim channel.
Fig. 37. Photographs of the metamorphosing leptocephalus, late glass eel, and elver stages of Conger myriaster (A) fromMiller and Tsukamoto (2004), and a metamorphosing 96.3 mm leptocephalus of Derichthys serpentinus (B) showing thepresence of red blood cells in the glass eel and elver stage of C. myriaster and the D. serpentinus larva.

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 43
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Fig. 38. A diagrammatical represen-tation of the change in otolith incre-ment width and Sr:Ca ratio in theotoliths of anguillid larvae fromhatching to metamorphosis, and tothe transition to the glass eel stage.Reprinted with permission fromMarine Ecology Progress Series,213, Marui M, Arai T, Miller MJ,Jellyman DJ, Tsukamoto K. Com-parison of early life history betweenNew Zealand temperate eels and Pa-cific tropical eels revealed by otolithmicrostructure and microchemistry.273–284, 2001, Figure 7. © 2001,Inter-Research.
Fig. 39. Scanning electron microscope images of the otoliths of Anguilla marmorata leptocephali at the premetamorphicstage (top), metamorphosing leptocephalus stage (middle), and the oceanic glass eel stage (bottom). White circle shows themetamorphosis check. Adapted from Kuroki et al. (2005).

44 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Critical swimming speed is a useful index of swimming ability obtained by swimming fish atincreasing speed intervals until they reach exhaustion (Fisher 2005; Fisher et al. 2005). Congeroceanicus larvae were collected during April–June at water temperatures of 14.0–24.5°C, andwere separated into premetamorphic and metamorphosing leptocephali (ER–M1) and glass eels(M2–M3) as illustrated in Fig. 36 and described by Bell et al. (2003). Ucrit of C. oceanicusranged from 12.0–26.8 cm s–1 for ER–M1 stage metamorphosing leptocephali and 4.1–25.0cm s–1 for M2–M3 stage glass eels (Fig. 40C). Anguilla rostrata glass eels that recruited to thesame area from January to June at temperatures of 4.0–21.0°C were considerably smaller (48.7–68.1 mm) than the Conger leptocephali (80.2–117.8 mm) and glass eels (68.3–90.0 mm). Anguillarostrata glass eels were separated into stage 1 and stage 2 glass eels based on their degree ofpigmentation, and they had Ucrit values that were 6.5–21.1 cm s–1 and 8.2–14.1 cm s–1, respec-tively (Fig. 40B). Later stages (M2–M3) of C. oceanicus showed greater variability in Ucrit thanearlier stages (ER–M1) or A. rostrata glass eels (Fig. 40). The strongest swimmers of eitherspecies appeared to be the late stage metamorphosing C. oceanicus leptocephali and a few oftheir glass eels. Although these were shorter than the earlier stage leptocephali, their greaterpercent dry weights suggested they had greater muscle mass (Fig. 40A). These individuals withUcrit values greater than about 20 cm s–1 all had longer bodies than the A. rostrata glass eels,which may account for part of their stronger swimming ability.
Fig. 40. Plots of the percent dry weights ofAnguilla rostrata glass eels (light blue circles,early-stage; blue circles, late-stage) and Con-ger oceanicus late-stage or metamorphosingleptocephali (yellow squares, ER1–M1 stagesas shown in Fig. 36) or glass eels (black squares,M2–M3 stages) (A), and their critical swimmingspeeds (Ucrit) (B and C) that were measured inlarvae that had recruited to coastal New Jerseyof the US east coast. Modified with kind per-mission from Prof. Kenneth W. Able andDr. Mark Wuenschel, and from Springer Sci-ence + Business Media: Marine Biology, Swim-ming ability of eels (Anguilla rostrata, Congeroceanicus) at estuarine ingress: contrasting pat-terns of cross-shelf transport? 154, 2008, 775–786, Wuenschel MJ, Able KW, Figures 2, 4b,5b. © 2008, Springer-Verlag.

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 45
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Temperature was significantly related to Ucrit values for both the stage 1 A. rostrata glasseels and the C. oceanicus ER–M1 stage leptocephali. Most of the Anguilla glass eels were testedat temperatures of 4–16°C and most of the Conger at 16–24°C, so the faster critical speeds of themetamorphosing Conger leptocephali and early glass eels is difficult to interpret. Temperatureaffects both the physiology of fish larvae and the characteristics of the water they must swimthrough (Fuiman and Batty 1997; von Herbing 2002). The Ucrit values of various families ofprimarily tropical coral reef area fishes range higher, from about 20–60 cm s–1 at 20–30°C(Fisher 2005; Fisher et al. 2005).
Observations of live leptocephali and the findings of Wuenschel and Able (2008) suggestleptocephali are capable swimmers, but further swimming stamina experiments are needed toevaluate their long term swimming ability. The anguilliform swimming mode in adult silver eelshas been found to be extremely efficient (van den Thillart et al. 2009). This efficiency has beendemonstrated by silver eels that can swim for 6 months or more without feeding, and seeminglystill have enough fat reserves for reproduction (van den Thillart et al. 2009). This suggests thatleptocephali with a similar anguilliform swimming mode may be well-adapted for long-termslow swimming.
3. Zoogeography of leptocephali
3-1. Taxonomic groups of eels
Leptocephali are distributed worldwide from tropical to lower-temperate latitudes becausea variety of taxa of eels live at these latitudes. The best known eels are of the family Anguillidae,which live in freshwater and estuarine habitats in most regions of the world, but their lepto-cephali are only found in areas of the ocean close to their specific spawning areas (Miller 2003;Aoyama 2009). Three anguilliform families (Congridae, Muraenidae, Ophichthidae) are par-ticularly rich in species and are distributed worldwide, except at high latitudes. These familiesinclude species whose juveniles and adults range from shallow, nearshore marine habitats, outover the continental shelf and slope, and down to depths of 1000 m or more in a few species(Böhlke 1989a). Other less diverse families that also live in shallow marine habitats worldwideinclude the Chlopsidae, Moringuidae, and Muraenesocidae. Although many marine eels likelylive in shallow tropical areas, typically only a few species of muraenid eels (Gilbert et al. 2005),the colonial garden eels (Thresher 1984; Smith 1989b), or ophichthids with color patterns mim-icking sea snakes (Randall 2005) are commonly observed by divers during the day. Other eels ofthe Nettastomatidae and some Synaphobranchidae may live primarily over the outer continentalshelf or on the upper slope. Synaphobranchids are among the most common fish on the abyssalplain at depths of 1000–3000 m (Haedrich and Merrett 1988; King et al. 2006). Eels of theDerichthyidae, Eurypharyngidae, Nemichthyidae, and Serrivomeridae live entirely mesopelagiclives in the upper few thousand meters of the world’s oceans (Böhlke 1989a). Some species suchas the cyematids may live even deeper. Therefore, eels of at least a few taxa are found in almostall marine habitats worldwide, except perhaps pelagically over the continental shelf and in verycold waters in the polar regions.
3-2. Spawning areas of eels
Where eels spawn in the ocean in relation to currents is a critical determinant of how theirlarvae become distributed. But due to the difficulty in observing spawning eels, their spawningareas are mostly known only from catches of small leptocephali. Their spawning locations rangewidely from shallow water to offshore regions of the ocean due to the various life histories ofeach family or species of eels.
All anguillid species spawn over deep water (Tsukamoto et al. 2002; Miller 2003; Tesch2003; Aoyama 2009), but the distance offshore that they migrate can vary from less than about100 km in the case of the tropical species A. celebesensis (Aoyama et al. 2003), to many thou-sands of kilometers for temperate anguillid species in the northern and southern hemisphere

46 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
subtropical gyres (Fig. 41). Although all temperate anguillids have long migrations, tropical eelshave a wider range of migration distances and types of spawning areas. Their known spawningareas range from local spawning (A. celebesensis and A. borneensis in the Indonesian Seas;Fig. 42), to offshore spawning by the northern population of A. marmorata in the NorthEquatorial Current region in an area that overlaps with the temperate species A. japonica(Kuroki et al. 2009a) (Fig. 42). This population of A. marmorata is both genetically(Minegishi et al. 2008) and morphologically (Watanabe et al. 2008) distinct from otherpopulations of that species that spawn in different areas. A similar case of both tropical andtemperate anguillids spawning offshore in about the same area appears to exist for the tropicalA. reinhartii and the temperate A. australis. Both spawn somewhere in the South EquatorialCurrent of the western South Pacific (Kuroki et al. 2008a).
Fig. 41. The spawning areas of the northern temperate species of anguillid eels in the subtropical gyres of the western NorthAtlantic showing the Florida Current (FC), Gulf Stream, and North Atlantic Drift (A), and in the western North Pacificshowing the North Equatorial Current (NEC), Mindanao Current (MC) and Kuroshio Current (B). The region where spawn-ing likely occurs by the southern temperate anguillid species in the western South Pacific is also shown along with the SouthEquatorial Current and the East Australian Current (C). Adapted with permission of the American Fisheries Society fromChallenges for Diadromous Fishes in a Dynamic Global Environment. Haro A, Avery T, Beal K, Cooper J, Cunjak R, DadswellM, Klauda R, Moffitt C, Rulifson R, Smith K (eds). Review of ocean-atmospheric factors in the Atlantic and Pacific oceansinfluencing spawning and recruitment of anguillid eels. Miller MJ, Kimura S, Friedland KD, Knights B, Kim H, Jellyman JD,Tsukamoto K. Am. Fish. Soc. Symp. 69. Bethesda, Maryland. 2009a; pp. 231–249, Figure 2.
Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 47
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
In contrast to anguillids, most species of marine eels may typically spawn within or neartheir adult habitats (Miller 2002b). Mesopelagic eel species may just spawn where they live inoffshore areas of the open ocean, because they are often widely distributed (Castonguay andMcCleave 1987b; Karmovskaya 1990; Miller and McCleave 1994; Smith and Miller 1996;Wippelhauser et al. 1996; Miller et al. 2006a). How they choose where to spawn is not knownthough, and how their spawning locations affect the distribution of leptocephali has not beenstudied. Similarly, some muraenids likely spawn wherever they live in shallow water habitats(Moyer and Zaiser 1982; Ferraris 1985), as do garden eels (subfamily Heterocongrinae), whichlive colonially in burrows in shallow areas (Thresher 1984). Therefore, the many islands oratolls in the Indo–Pacific, which have coral reef and other types of habitats (Fig. 4B) used bycongrid, chlopsid, muraenid, and ophichthid eels, are source areas for their leptocephali to spreadthroughout the western Pacific and Indonesian Seas region. Some species may also spawnover the continental shelf or migrate towards the edge of the shelf (Cohen and Dean 1970;Ross and Rohde 2003). There is evidence of many marine eel species living and spawning inwarm tropical areas, such as the Indonesian Seas (Wouthuyzen et al. 2005), where the biodiversity
Fig. 42. Geographic distribution of tropical anguillid leptocephalicollected in the western Pacific and central Indonesian Seas from1995 to 2004, showing the leptocephali of Anguilla marmorata (redtriangles), which spawns offshore and recruits to Indonesia and otherareas in the region and Anguilla bicolor pacifica (yellow circles)whose spawning area is not known (A), and leptocephali of Anguillacelebesensis (blue circles) and Anguilla borneensis (orange trian-gles), whose adults spawn in the Indonesian Seas region (B). M =metamorphosing leptocephalus; G = oceanic glass eel. Adapted fromwith permission of the American Fisheries Society from Challengesfor Diadromous Fishes in a Dynamic Global Environment. HaroA, Smith K, Rulifson R, Moffitt C, Klauda R, Dadswell M, CunjakR, Cooper J, Beal K, Avery T (eds). Migration scales of catadromouseels: diversity and evolution of larval migration based on their dis-tribution, morphology and early life history. Kuroki M, Aoyama J,Kim H, Miller MJ, Kimura S, Tsukamoto K. Am. Fish. Soc. Symp.69. Bethesda, Maryland. 2009; pp. 887–889, Figure 1.

48 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
of fishes is generally the highest in the world (Randall 1998; Allen and Adrim 2003; Carpenterand Springer 2005). Fewer species of eels appear to live and spawn in colder water areas, suchas the eastern North Pacific though, based on the limited number of leptocephali reported there(Raju 1985; Matarese et al. 1989).
Most surveys for leptocephali have been in offshore waters due to the oceanic spawn-ing of anguillid eels, and these collections have shown that very few marine eels migratevery far offshore to spawn. Congrids such as Conger oceanicus (McCleave and Miller 1994)and some Ariosoma balearicum (Miller 2002a) appear to migrate offshore to spawn, but mostother marine eels that live in continental shelf habitats do not appear to migrate much pastthe edge of the shelf. Collections of leptocephali over or near shelf areas or around islands alsohave shown that most shallow water marine eels do not migrate very far offshore to spawn. Theexact spawning areas of marine eels are generally not known, but the smallest leptocephali ofmost shallow water eels are typically collected over or near the edge of the continental shelf(Castle and Roberston 1974; Miller et al. 2002a; Kimura et al. 2006) or in close proximity toshallow water habitats (Castle 1968; Miller 1995, 2002b; Miller et al. 2002a, 2006a; Miller andMcCleave 2007), suggesting nearby spawning areas.
A survey in the late fall along the outer shelf of the East China Sea (Fig. 32) found evi-dence of spawning by many species of eels (Miller et al. 2002a). The two most abundant speciesof leptocephali were Gnathophis nystromi nystromi (Congridae) and Dysomma anguillare(Synaphobranchidae), which based on their distributions of small leptocephali were spawning atthe outer edge of the shelf (Fig. 43). The exact stations at which the smallest leptocephali of thetwo species were caught differed slightly though, suggesting that D. anguillare was spawning alittle farther out over the slope than G. nystromi nystromi. All sizes of leptocephali of theMuraenidae, Ophichthidae, and Saurenchelys (Nettastomatidae) were also abundant over theouter shelf (Fig. 44), indicating that spawning occurred near the edge of the shelf. Small lepto-cephali of G. nystromi nystromi and a few other species were also collected along the outeredge of the continental shelf of northeast Japan (Kimura et al. 2006), which is similar to obser-vations from other studies on various species of Gnathophis around the world (Castle 1968;Castle and Roberston 1974).
3-3. Distribution and abundance of leptocephali
The abundance of leptocephali in any particular area is likely directly related to theproximity of that area to the spawning locations of the species and the regional patterns ofocean currents that disperse the larvae. The leptocephali of eels that live over the continen-tal shelf have been found to be very abundant near their spawning areas such as along theedge of the shelf in the East China Sea due to large catches of small leptocephali (Figs. 43, 44;Miller et al. 2002a). Similarly, leptocephali of congrid, chlopsid, muraenid, and ophichthid eelswere very abundant in the Northwest Providence Channel of the Northern Bahamas at the south-west edge of the Sargasso Sea in the western North Atlantic (Fig. 45; Miller and McCleave2007). However, there are often strong currents adjacent to the these high density concentrationsof leptocephali, such as the Kuroshio Current in the East China Sea and the Florida Current(southern part of the Gulf Stream) adjacent to the northern Bahamas, which likely transportmany leptocephali further offshore.
Some species of leptocephali can be quite abundant offshore in the open ocean, as a resultof local spawning or offshore transport by frontal jets (Miller and McCleave 1994). Lepto-cephali of the mesopelagic eels that are abundant in some offshore areas during their spawningseasons include Derichthyidae (Castonguay and McCleave 1987b), Nemichthyidae(Karmovskaya 1990; Wippelhauser et al. 1996) and Serrivomeridae (Miller and McCleave 1994;Miller et al. 2006a). Anguillid leptocephali also are sometimes abundant in regions near theiroffshore spawning areas (Kleckner and McCleave 1988; Tsukamoto 1992) and then they be-come widely distributed in their larval development and migration areas (Kleckner and McCleave1985; McCleave and Kleckner 1987; Kettle and Haines 2006; Kuroki et al. 2009a,b). In moreenclosed areas such as the Indonesian Seas, leptocephali of all types of eels mix together with
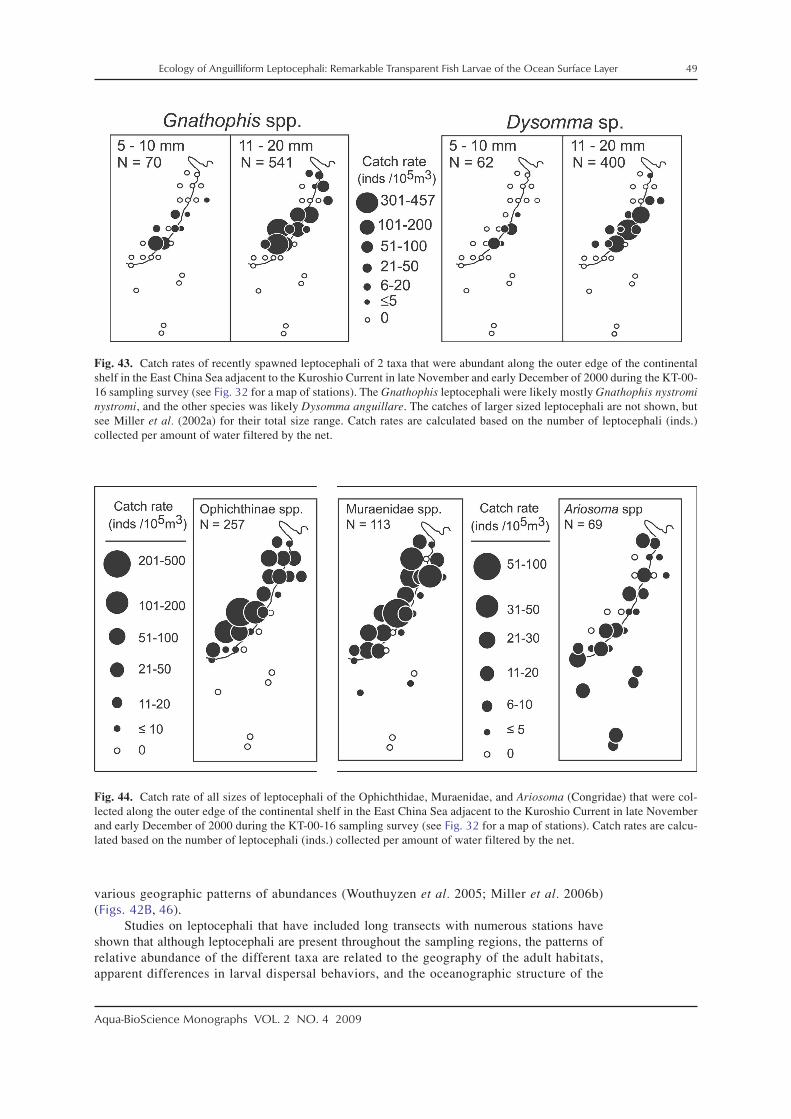
Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 49
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
various geographic patterns of abundances (Wouthuyzen et al. 2005; Miller et al. 2006b)(Figs. 42B, 46).
Studies on leptocephali that have included long transects with numerous stations haveshown that although leptocephali are present throughout the sampling regions, the patterns ofrelative abundance of the different taxa are related to the geography of the adult habitats,apparent differences in larval dispersal behaviors, and the oceanographic structure of the
Fig. 43. Catch rates of recently spawned leptocephali of 2 taxa that were abundant along the outer edge of the continentalshelf in the East China Sea adjacent to the Kuroshio Current in late November and early December of 2000 during the KT-00-16 sampling survey (see Fig. 32 for a map of stations). The Gnathophis leptocephali were likely mostly Gnathophis nystrominystromi, and the other species was likely Dysomma anguillare. The catches of larger sized leptocephali are not shown, butsee Miller et al. (2002a) for their total size range. Catch rates are calculated based on the number of leptocephali (inds.)collected per amount of water filtered by the net.
Fig. 44. Catch rate of all sizes of leptocephali of the Ophichthidae, Muraenidae, and Ariosoma (Congridae) that were col-lected along the outer edge of the continental shelf in the East China Sea adjacent to the Kuroshio Current in late Novemberand early December of 2000 during the KT-00-16 sampling survey (see Fig. 32 for a map of stations). Catch rates are calcu-lated based on the number of leptocephali (inds.) collected per amount of water filtered by the net.
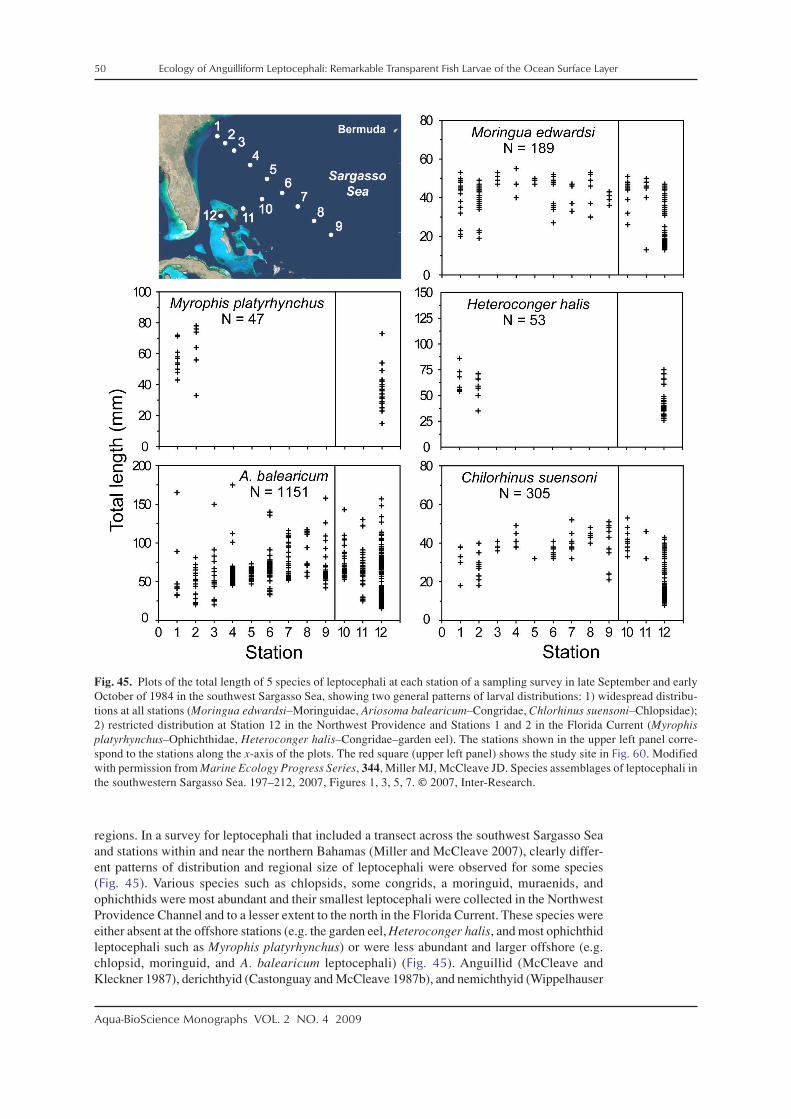
50 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
regions. In a survey for leptocephali that included a transect across the southwest Sargasso Seaand stations within and near the northern Bahamas (Miller and McCleave 2007), clearly differ-ent patterns of distribution and regional size of leptocephali were observed for some species(Fig. 45). Various species such as chlopsids, some congrids, a moringuid, muraenids, andophichthids were most abundant and their smallest leptocephali were collected in the NorthwestProvidence Channel and to a lesser extent to the north in the Florida Current. These species wereeither absent at the offshore stations (e.g. the garden eel, Heteroconger halis, and most ophichthidleptocephali such as Myrophis platyrhynchus) or were less abundant and larger offshore (e.g.chlopsid, moringuid, and A. balearicum leptocephali) (Fig. 45). Anguillid (McCleave andKleckner 1987), derichthyid (Castonguay and McCleave 1987b), and nemichthyid (Wippelhauser
Fig. 45. Plots of the total length of 5 species of leptocephali at each station of a sampling survey in late September and earlyOctober of 1984 in the southwest Sargasso Sea, showing two general patterns of larval distributions: 1) widespread distribu-tions at all stations (Moringua edwardsi–Moringuidae, Ariosoma balearicum–Congridae, Chlorhinus suensoni–Chlopsidae);2) restricted distribution at Station 12 in the Northwest Providence and Stations 1 and 2 in the Florida Current (Myrophisplatyrhynchus–Ophichthidae, Heteroconger halis–Congridae–garden eel). The stations shown in the upper left panel corre-spond to the stations along the x-axis of the plots. The red square (upper left panel) shows the study site in Fig. 60. Modifiedwith permission from Marine Ecology Progress Series, 344, Miller MJ, McCleave JD. Species assemblages of leptocephali inthe southwestern Sargasso Sea. 197–212, 2007, Figures 1, 3, 5, 7. © 2007, Inter-Research.
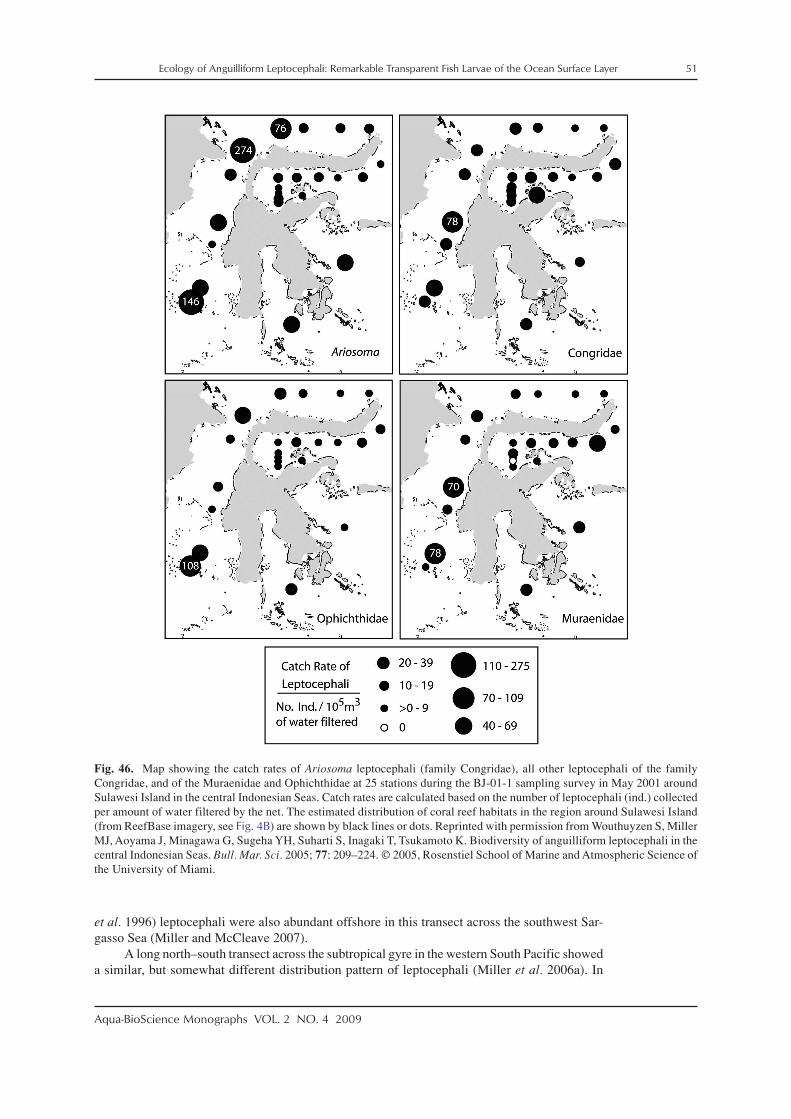
Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 51
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
et al. 1996) leptocephali were also abundant offshore in this transect across the southwest Sar-gasso Sea (Miller and McCleave 2007).
A long north–south transect across the subtropical gyre in the western South Pacific showeda similar, but somewhat different distribution pattern of leptocephali (Miller et al. 2006a). In
Fig. 46. Map showing the catch rates of Ariosoma leptocephali (family Congridae), all other leptocephali of the familyCongridae, and of the Muraenidae and Ophichthidae at 25 stations during the BJ-01-1 sampling survey in May 2001 aroundSulawesi Island in the central Indonesian Seas. Catch rates are calculated based on the number of leptocephali (ind.) collectedper amount of water filtered by the net. The estimated distribution of coral reef habitats in the region around Sulawesi Island(from ReefBase imagery, see Fig. 4B) are shown by black lines or dots. Reprinted with permission from Wouthuyzen S, MillerMJ, Aoyama J, Minagawa G, Sugeha YH, Suharti S, Inagaki T, Tsukamoto K. Biodiversity of anguilliform leptocephali in thecentral Indonesian Seas. Bull. Mar. Sci. 2005; 77: 209–224. © 2005, Rosenstiel School of Marine and Atmospheric Science ofthe University of Miami.

52 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
this transect, serrivomerid leptocephali were very abundant at a wide range of sizes in the warmerwater regions in the north from latitudes 5–17°S, but were less abundant and generally larger inthe colder water of the southern part of the gyre (Fig. 47). Leptocephali of anguillids and shal-low water eels were also rare in the colder water, but they were abundant in a more limitedregion in the central part of the transect at the latitude of the southern branch of the SouthEquatorial Current near the southern edge of the warm water (12–16°S, Fig. 47). The anguillidabundance there may have been due to spawning areas at that latitude, and the abundance ofshallow water marine eel families of leptocephali may have been due to the presence of adultson the larger island groups, such as Fiji, and Tonga, Samoa. The westward flow of the SouthEquatorial Current likely transported some of the leptocephali offshore where they were cap-
Fig. 47. Catch rates of 9 families of leptocephali at each station along 175˚E transects north and south of the Fiji Islands (A–C), and temperature and salinity sections down to 500 m made from the CTD profiles at each sampling station in the westernSouth Pacific (bottom panels), in August 1995 during the KH-95-2 sampling survey. The black lines below the station labelsof the catch rate plots indicate which stations were sampled at night. The Fiji islands are located from 16–18˚S, to the east ofStn. 73–76. These transects can be seen in the bottom right side of Fig. 4A. Catch rates are calculated based on the number ofleptocephali collected per amount of water filtered by the net. Reprinted from Deep-Sea Research I, 53, Miller MJ, AoyamaJ, Mochioka N, Otake T, Castle PHJ, Minagawa G, Inagaki T, Tsukamoto K. Geographic variation in the assemblages ofleptocephali in the western South Pacific, 776–794, 2006, with permission from Elsevier. © 2006, Elsevier Ltd.

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 53
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
tured at some stations of the transect, but they were rare to the north where there are few islands,and to the south in the colder water that was likely flowing to the east (Miller et al. 2006a).Serrivomerid, muraenid and other leptocephali were abundant and present at a wide range ofsizes at each station just to the north of Fiji (Fig. 48). Similar geographic differences inabundances of some species of leptocephali were observed in transects that crossed temperaturefronts in the Sargasso Sea (Miller and McCleave 1994).
The detailed distributions of leptocephali in the world’s oceans is beyond the scope of thisreview, since there are many less extensive reports on leptocephali. These reports indicate that atleast some leptocephali can be collected anywhere that appropriate sampling effort is applied attropical and subtropical latitudes. The taxonomic composition of leptocephali in each area mayvary widely however, depending on the geography of adult habitats or the time of year, as dis-cussed in the next section. The geographic variation of presence of species of leptocephali is
Fig. 48. Plots of the total length of the individual leptocephali of several taxa collected at each station along 175˚E transectsnorth and south of the Fiji Islands (same stations as Fig. 47). Reprinted from Deep-Sea Research I, 53, Miller MJ, Aoyama J,Mochioka N, Otake T, Castle PHJ, Minagawa G, Inagaki T, Tsukamoto K. Geographic variation in the assemblages of lepto-cephali in the western South Pacific, 776–794, 2006, with permission from Elsevier. © 2006, Elsevier Ltd.

54 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
illustrated by a plot of species diversity presented by Minagawa and Miller (2005) that shows ahigh species diversity in the Indonesian Seas, as was indicated by Wouthuyzen et al. (2005)and Minagawa et al. (2004), but also high diversity near the Solomon Islands in the westernSouth Pacific (Fig. 4A), as found by Miller et al. (2006a). The lowest diversities were observedin far offshore areas where the leptocephali of shallow water eels are only present in lowabundances. However, another factor is that the leptocephali of some species such as ophichthidsand garden eels seem to be rarely transported offshore into these areas compared to other spe-cies, except in areas with very strong currents (Miller 1995; Miller and McCleave 1994, 2007;Miller et al. 2006a).
3-4. Seasonal occurrence of leptocephali
Another factor that clearly affects the distribution, abundance and taxonomic compositionof leptocephali in some regions is the seasonality of spawning. Most of what is known aboutseasonality of spawning has been gathered about anguillid eels. Spawning seasons of temperateanguillids have been roughly estimated from the timing of the downstream migrations of silvereels into the ocean (Jellyman 2003; Tesch 2003) and the catches of their small leptocephali(McCleave 2008). Temperate anguillids typically migrate in the late summer or fall and theirleptocephali are abundant in their offshore spawning areas and larval migration routes in thenext spring and summer (e.g. McCleave and Kleckner 1987; Kleckner and McCleave 1988;Tsukamoto 1992; Tsukamoto et al. 2003). The timing of spawning of tropical anguillids is muchless understood, however, even though seasonal differences have been observed in the presenceor size of leptocephali in the central Indonesian Seas (Wouthuyzen et al. 2009) and off westSumatra in the eastern Indian Ocean (Jespersen 1942; Aoyama et al. 2007).
Even less is known about the seasonality of spawning of most marine eels though, sincereproductive behaviors of most species can not be monitored. Therefore, catches of leptocephaliare presently the best way to learn about the general seasonal patterns of spawning by marineeels. In tropical areas, such as the Indonesian Seas, published and unpublished data from varioussurveys have shown that a wide variety of species of leptocephali appear to be present therethroughout the year (Miller et al. 2002c, 2004; Wouthuyzen et al. 2005; Miller et al. unpubl.data). Although leptocephali are present all year, the seasonality of spawning of most species ofeels is not known, so it can not be assumed that most species of marine eels spawn all year.Chlopsid eels have been suggested to have spawned over five consecutive months in the centralIndonesian Seas based on analyses of otoliths from leptocephali collected in September(Fig. 30; Lee et al. 2008), but more research is needed to evaluate the spawning seasons of eelsin tropical regions.
At higher subtropical and in temperate areas, such as near Japan, there is some evidencethat various taxa of eels have seasonal patterns of spawning based on catches of leptocephali.Seasonal variation in taxonomic composition, relative abundance, size, and geographic distribu-tion of leptocephali collected in late spring (May–Jun) and late autumn (Oct–Dec) were com-pared in the East China Sea and in Suruga Bay of eastern Japan (Fig. 49; Minagawa et al. 2007).There were indications that some taxa of eels were spawning all year in the more southernregion of the East China Sea, although species such as Dysomma anguillare were absent there inthe late spring (Fig. 49B). Further north in Suruga Bay, it appeared that there was a much strongerpattern of seasonality of spawning, with far fewer leptocephali being present in the late spring(Figs. 49C, D). This suggested that at the temperate latitudes, such as at Suruga Bay (34°N),most marine eels may stop spawning during the winter months when water temperatures arecolder. Studies on a few species of marine eels in East Asia, such as Ariosoma meeki (Ishii et al.2003), Conger myriaster (Okamura et al. 2000; Utoh et al. 2005), and Muraenesox cinereus(Takai 1959, 1979), have found seasonal patterns of reproductive maturation, suggesting thatthese eels have spawning seasons in fall.
There is also some information available about the seasonality of spawning of marine eelsin other regions of the world based on catches of leptocephali. In the Gulf of Mexico, at a similarlatitude as the East China Sea, catches of leptocephali suggest seasonal spawning of the congrids

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 55
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Rhynchoconger flavus and Xenomystax congroides, but other congrids such as Urocongersyringinus, appeared to spawn all year (Smith 1989a). Occurrences of small Gnathophis lepto-cephali in coastal waters of New Zealand, eastern Australia, and South Africa suggested sea-sonal spawning, but spawning off western Australia may occur year round (Castle 1968; Castleand Roberson 1974). Castle (1969) showed that leptocephali of various families were collectedat all times of year along a north–south transect that was repeatedly sampled in the easternIndian Ocean off western Australia and south of Java Island of Indonesia. Catches were greatestfrom May to August with higher species richness occurring in the north closer to Java andgreater abundances occurring further south near Australia due to large catches of Ariosoma andGnathophis leptocephali.
In the tropical region of the Gulf of Guinea off western Africa, Blache (1977) reported thatin extensive sampling over a 10-year period, many species, including several congrids, showedevidence of seasonal spawning at latitudes of about 5°N to 18°S. Based on a lack of catches of
Fig. 49. Catch rates of 3 families of leptocephali at each station during 2 sampling surveys in the East China Sea during lateautumn (Oct–Dec) and late spring (May–June) (A, B), and during two surveys in Suruga Bay during late autumn (Oct–Nov)and late spring (May–June) (C, D), which show seasonal differences in catches between surveys. Catch rates are calculatedbased on the number of leptocephali (ind.) collected per amount of water filtered by the net. Modified with permission fromEstuarine, Coastal and Shelf Science, 71, Minagawa G, Miller MJ, Kimura Y, Watanabe S, Shinoda A, Aoyama J, TsukamotoK. Seasonal differences in catches of leptocephali in the East China Sea and Suruga Bay, Japan, 730–740, 2007, with permis-sion from Elsevier. © 2007, Elsevier Ltd.

56 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
leptocephali, there was an apparent absence of spawning activity from July to October (Blache1977), which corresponds directly to the period of wind-driven upwelling of cold water in theregion (Okumura and Xie 2004). This suggests that although this region is close to the equator,the colder water temperatures during the upwelling season may cause eels to stop their spawn-ing activity, as has been suggested at higher latitudes.
Seasonal patterns in the catches of leptocephali of mesopelagic eels in the Sargasso Seahave indicated that at least some of these species only spawn during part of the year. For exam-ple, although derichthyid leptocephali were abundant in the summer and fall across the south-west Sargasso Sea (Castonguay and McCleave 1987b), they were absent in the same generalregion in the winter and spring (Miller and McCleave 1994). The opposite pattern was observedfor Serrivomer leptocephali, which were abundant in the winter and spring (Miller and McCleave1994), but almost completely absent in the summer and fall (Miller and McCleave 2007). Otherspecies are present in both seasons, so more research is needed to gain a better understanding ofthe many factors that can affect the zoogeography and temporal occurrence of leptocephali.
4. Ecology of leptocephali
4-1. Depth distribution and vertical migration
There have only been a few published reports on the depth distribution of leptocephali,primarily due to the difficulty of using large enough nets that can sample discrete layers of theocean. All of the studies that have been done, however, have indicated that leptocephali are mostabundant in the upper 100 m at night, but their exact depth distributions during the day haveremained difficult to determine because of likely net avoidance. Studies on leptocephali in thewestern North Pacific (Kajihara et al. 1988) and on A. japonica (Otake et al. 1998) have indi-cated a shallow depth distribution of leptocephali in the upper 100 m at night. Small A. japonicaleptocephali were most abundant between 60 and 90 m at night during two surveys in the west-ern North Pacific (Otake et al. 1998). Schoth and Tesch (1984) examined the depth distributionof small anguillid leptocephali in the Sargasso Sea and found evidence of vertical migrationfrom about 30–70 m during night to 70–280 m during the day.
The most extensive study on the depth distribution of leptocephali was conducted byCastonguay and McCleave (1987a) during several surveys for leptocephali in the Sargasso Sea,in the late winter/spring spawning season and in the summer/fall when anguillid leptocephaliare larger in size. Data from this study indicated that the smaller recently spawned leptocephaliof A. rostrata and A. anguilla were present at a range of depths between 50 and 350 m duringboth day and night (Fig. 50). As the leptocephali grew they were present only in the upper 100 mat night and showed increasing evidence of vertical migration to greater depths during the day(Fig. 50; Castonguay and McCleave, 1987a). At one site, where >40 mm anguillid leptocephaliwere collected in the southwest Sargasso Sea in August (Fig. 50, bottom right panel), mostleptocephali during nighttime were collected within the thermocline at 50–70 m in tempera-tures of about 24–25°C (see McCleave and Kleckner 1987), but none were collected in themixed layer of 27°C water in the upper 30 m, or below the thermocline at 100–125 m depths(Castonguay and McCleave 1987a).
A similar pattern of depth distribution in the upper 100 m at night was reported byCastonguay and McCleave (1987a) for various species of marine eel leptocephali, but therewere indications of differences in distributions among taxa within each season. Leptocephali ofthe mesopelagic eel, Derichthys serpentinus, showed evidence of vertical migrations similar toAnguilla, with most being collected from 30–70 m at night in the summer and fall (Fig. 51). Avariety of other species of leptocephali were also collected including those of mesopelagic andshallow water eels (Fig. 52). Some species such as Avocettina infans and Serrivomer brevidentatus(mesopelagic) were most abundant in the upper 50 m depth at night, but various other specieswere more abundant at 50–100 m (Fig. 52). In contrast, Ariosoma balearicum was distributedevenly between those two depth ranges at night (Castonguay and McCleave, 1987a). The catchdata also showed that in contrast to other species, such as Anguilla and D. serpentinus, Nemichthys

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 57
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
scolopaceus leptocephali < 80 mm did not seem to migrate vertically and could be caught in theshallow layers during the day, but at least some of the larger sizes did migrate vertically. Little orno evidence was found that some species such as A. balearicum, A. infans, or C. oceanicus weremigrating vertically.
These observations that leptocephali are generally found in the upper 100 m at night andwithin the upper 300 m during day has been confirmed by catches of leptocephali in manydeployments of the much larger multiple-net MOCNESS-10 trawl system (10 m2 mouthopening, Wiebe et al. 1985) in studies on mesopelagic fishes (e.g. Backus and Craddock 1982)or salps (e.g. Madin et al. 2006) during a wide variety of cruises in the western North Atlantic.Most leptocephali were collected in nets that fished at least part of the time in the upper 100 mat night, and few were ever collected deeper than 300 m in the many nets that fished in deeplayers during both day and night (Miller unpubl. data).
Fig. 50. Vertical distribution of six size ranges of anguillid leptocephali (Anguilla rostrata and Anguilla anguilla combined)that were collected in the Sargasso Sea during spring, summer, and fall of 1983–1985 using a 2-m ring net during both day(white bars) and night (black bars) in the study of Castonguay and McCleave (1987a). Catches are plotted as a percentage oftotal density of leptocephali within each size class based on the amount of water filtered by the net during all the tows. Zerosshow the depth ranges where no leptocephali were collected, and based on clear patterns of catches in the earlier cruises, someshallower depths during the day and deeper depths at night where no leptocephali were collected, were not sampled during thelater cruises. Redrawn from Castonguay M, McCleave JD, Vertical distributions, diel and ontogenetic vertical migrations andnet avoidance of leptocephali of Anguilla and other common species in the Sargasso Sea, Journal of Plankton Research,1987, 9(1), 195–214, by permission of Oxford University Press.
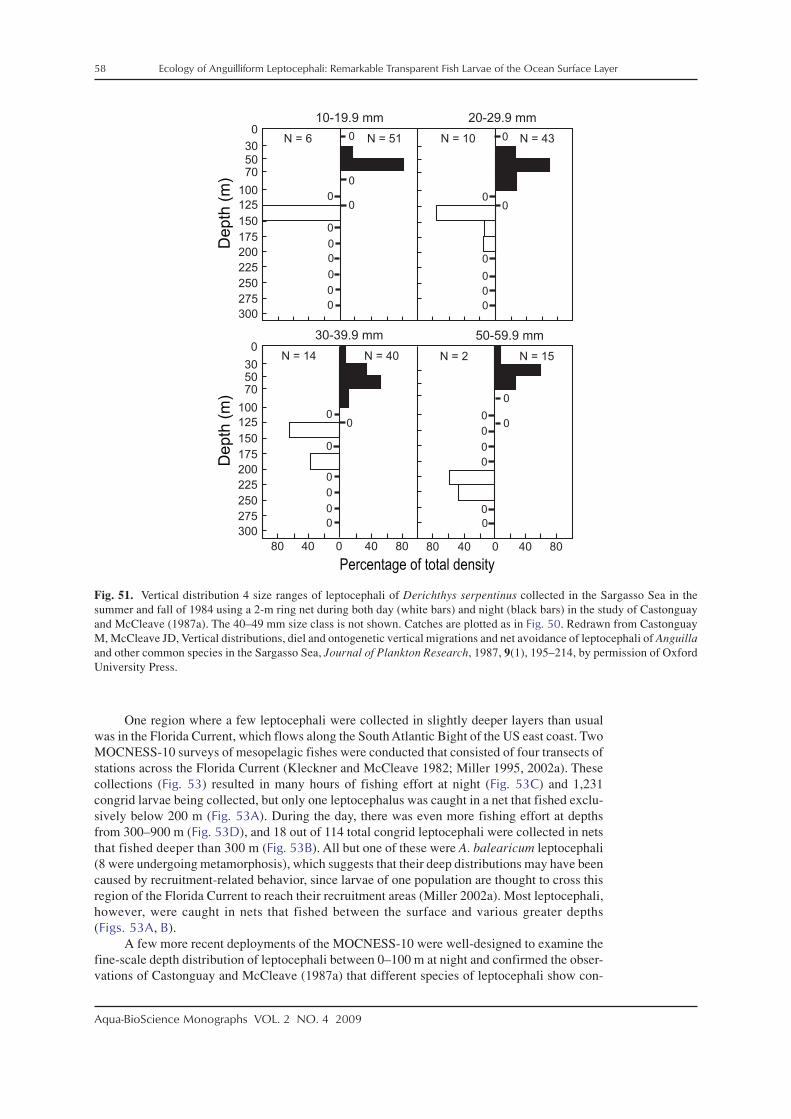
58 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
One region where a few leptocephali were collected in slightly deeper layers than usualwas in the Florida Current, which flows along the South Atlantic Bight of the US east coast. TwoMOCNESS-10 surveys of mesopelagic fishes were conducted that consisted of four transects ofstations across the Florida Current (Kleckner and McCleave 1982; Miller 1995, 2002a). Thesecollections (Fig. 53) resulted in many hours of fishing effort at night (Fig. 53C) and 1,231congrid larvae being collected, but only one leptocephalus was caught in a net that fished exclu-sively below 200 m (Fig. 53A). During the day, there was even more fishing effort at depthsfrom 300–900 m (Fig. 53D), and 18 out of 114 total congrid leptocephali were collected in netsthat fished deeper than 300 m (Fig. 53B). All but one of these were A. balearicum leptocephali(8 were undergoing metamorphosis), which suggests that their deep distributions may have beencaused by recruitment-related behavior, since larvae of one population are thought to cross thisregion of the Florida Current to reach their recruitment areas (Miller 2002a). Most leptocephali,however, were caught in nets that fished between the surface and various greater depths(Figs. 53A, B).
A few more recent deployments of the MOCNESS-10 were well-designed to examine thefine-scale depth distribution of leptocephali between 0–100 m at night and confirmed the obser-vations of Castonguay and McCleave (1987a) that different species of leptocephali show con-
Fig. 51. Vertical distribution 4 size ranges of leptocephali of Derichthys serpentinus collected in the Sargasso Sea in thesummer and fall of 1984 using a 2-m ring net during both day (white bars) and night (black bars) in the study of Castonguayand McCleave (1987a). The 40–49 mm size class is not shown. Catches are plotted as in Fig. 50. Redrawn from CastonguayM, McCleave JD, Vertical distributions, diel and ontogenetic vertical migrations and net avoidance of leptocephali of Anguillaand other common species in the Sargasso Sea, Journal of Plankton Research, 1987, 9(1), 195–214, by permission of OxfordUniversity Press.
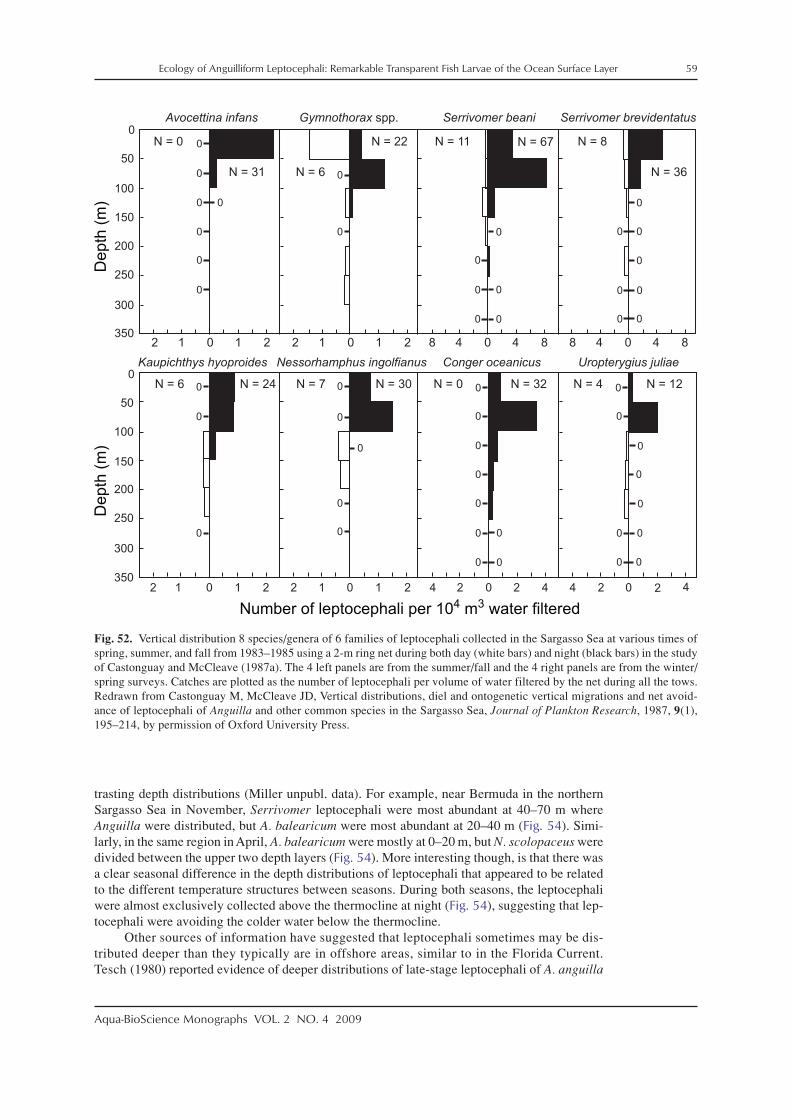
Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 59
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
trasting depth distributions (Miller unpubl. data). For example, near Bermuda in the northernSargasso Sea in November, Serrivomer leptocephali were most abundant at 40–70 m whereAnguilla were distributed, but A. balearicum were most abundant at 20–40 m (Fig. 54). Simi-larly, in the same region in April, A. balearicum were mostly at 0–20 m, but N. scolopaceus weredivided between the upper two depth layers (Fig. 54). More interesting though, is that there wasa clear seasonal difference in the depth distributions of leptocephali that appeared to be relatedto the different temperature structures between seasons. During both seasons, the leptocephaliwere almost exclusively collected above the thermocline at night (Fig. 54), suggesting that lep-tocephali were avoiding the colder water below the thermocline.
Other sources of information have suggested that leptocephali sometimes may be dis-tributed deeper than they typically are in offshore areas, similar to in the Florida Current.Tesch (1980) reported evidence of deeper distributions of late-stage leptocephali of A. anguilla
Fig. 52. Vertical distribution 8 species/genera of 6 families of leptocephali collected in the Sargasso Sea at various times ofspring, summer, and fall from 1983–1985 using a 2-m ring net during both day (white bars) and night (black bars) in the studyof Castonguay and McCleave (1987a). The 4 left panels are from the summer/fall and the 4 right panels are from the winter/spring surveys. Catches are plotted as the number of leptocephali per volume of water filtered by the net during all the tows.Redrawn from Castonguay M, McCleave JD, Vertical distributions, diel and ontogenetic vertical migrations and net avoid-ance of leptocephali of Anguilla and other common species in the Sargasso Sea, Journal of Plankton Research, 1987, 9(1),195–214, by permission of Oxford University Press.

60 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
(mostly 60–85 mm) as they were approaching Europe for recruitment, which were possibly asdeep as about 300–650 m during the day. They were most abundant at 50–100 m at night. Thesecollections were made in horizontal tows of an IKMT though, so some contamination fromshallower depths could not be ruled out. Ross et al. (2007) also indicated that some marine eelleptocephali collected along the southeast US coast just inshore of the Florida Current were atgreater depths (about 300–800 m) than typically observed, but at least part of their data werecollected with open nets that may have unavoidably collected some leptocephali in shallowerlayers than the targeted fishing depth. However, since a few leptocephali were collected at greaterdepths by several nets of the MOCNESS-10 in the Florida Current (Fig. 53), which should nothave any contamination from other depths, it is possible that when leptocephali are approachingthe continental shelves where they have the potential to recruit, they migrate vertically to greaterdepths, perhaps searching for the bottom.
At the other extreme, it is clear that leptocephali in the open ocean at least, can also some-times be found in the neuston layer at or near the surface at night, within about the upper 1 m ofthe ocean. Ariosoma scheelei leptocephali were collected at the surface at 11 of the 25 stationsaround Sulawesi Island in May (1.5 m ring net towed next to the ship), but no metamorphosingindividuals were caught at the surface (Miller et al. 2006b). Leptocephali were also frequentlycollected in the same way at the surface to the northwest in the Sulu Sea in both Februaryand November (Miller et al. 2004; Miller and Tsukamoto unpubl. data). These taxa included
Fig. 53. Plots of catch rates of congrid leptocephali collected by the MOCNESS-10 opening and closing trawl in 4 transectsof stations across the Florida Current that are plotted as the number of leptocephali (ind.) caught per minute each net fished,at night (A) and during daytime (B), during August 1978 and February 1979 surveys for mesopelagic fishes. Data are plottedas catch rates and the minimum depth fished by each net (nets fished at various depth ranges deeper than each minimumdepth). Note the very different y-axis scales for catch rates during night and day. See Kleckner and McCleave (1982), andMiller (1995) for maps of the stations sampled during these surveys.
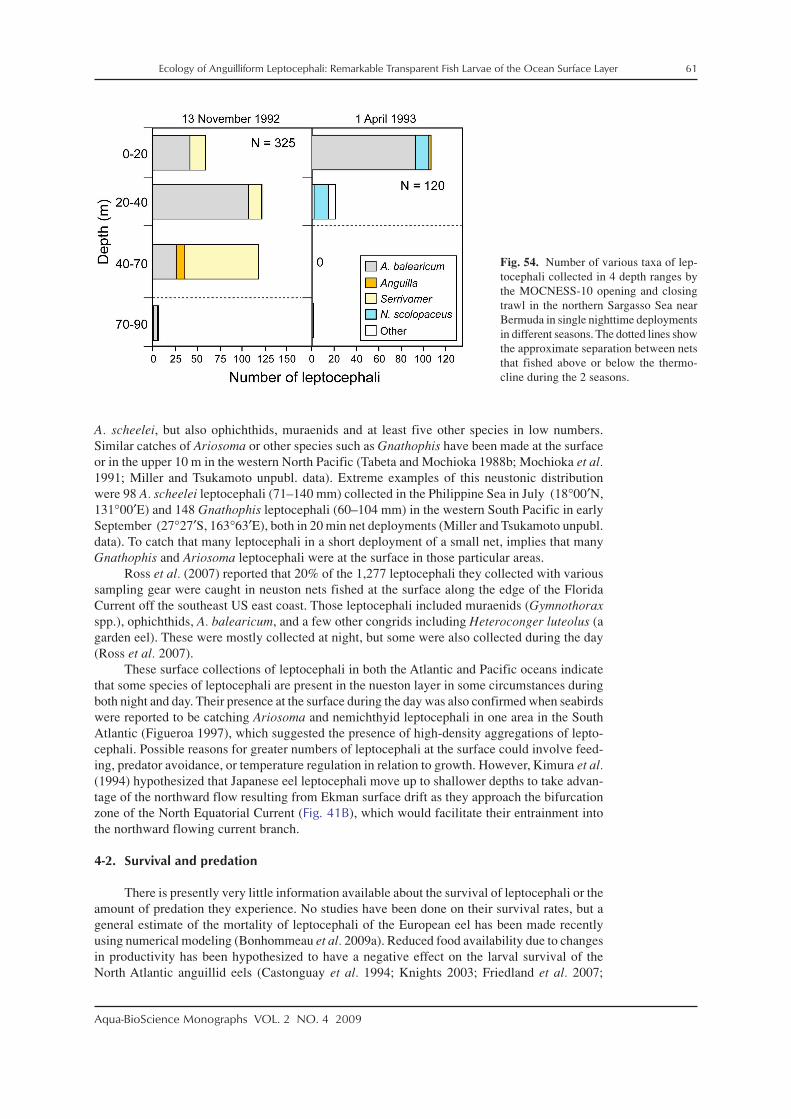
Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 61
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
A. scheelei, but also ophichthids, muraenids and at least five other species in low numbers.Similar catches of Ariosoma or other species such as Gnathophis have been made at the surfaceor in the upper 10 m in the western North Pacific (Tabeta and Mochioka 1988b; Mochioka et al.1991; Miller and Tsukamoto unpubl. data). Extreme examples of this neustonic distributionwere 98 A. scheelei leptocephali (71–140 mm) collected in the Philippine Sea in July (18°00′N,131°00′E) and 148 Gnathophis leptocephali (60–104 mm) in the western South Pacific in earlySeptember (27°27′S, 163°63′E), both in 20 min net deployments (Miller and Tsukamoto unpubl.data). To catch that many leptocephali in a short deployment of a small net, implies that manyGnathophis and Ariosoma leptocephali were at the surface in those particular areas.
Ross et al. (2007) reported that 20% of the 1,277 leptocephali they collected with varioussampling gear were caught in neuston nets fished at the surface along the edge of the FloridaCurrent off the southeast US east coast. Those leptocephali included muraenids (Gymnothoraxspp.), ophichthids, A. balearicum, and a few other congrids including Heteroconger luteolus (agarden eel). These were mostly collected at night, but some were also collected during the day(Ross et al. 2007).
These surface collections of leptocephali in both the Atlantic and Pacific oceans indicatethat some species of leptocephali are present in the nueston layer in some circumstances duringboth night and day. Their presence at the surface during the day was also confirmed when seabirdswere reported to be catching Ariosoma and nemichthyid leptocephali in one area in the SouthAtlantic (Figueroa 1997), which suggested the presence of high-density aggregations of lepto-cephali. Possible reasons for greater numbers of leptocephali at the surface could involve feed-ing, predator avoidance, or temperature regulation in relation to growth. However, Kimura et al.(1994) hypothesized that Japanese eel leptocephali move up to shallower depths to take advan-tage of the northward flow resulting from Ekman surface drift as they approach the bifurcationzone of the North Equatorial Current (Fig. 41B), which would facilitate their entrainment intothe northward flowing current branch.
4-2. Survival and predation
There is presently very little information available about the survival of leptocephali or theamount of predation they experience. No studies have been done on their survival rates, but ageneral estimate of the mortality of leptocephali of the European eel has been made recentlyusing numerical modeling (Bonhommeau et al. 2009a). Reduced food availability due to changesin productivity has been hypothesized to have a negative effect on the larval survival of theNorth Atlantic anguillid eels (Castonguay et al. 1994; Knights 2003; Friedland et al. 2007;
Fig. 54. Number of various taxa of lep-tocephali collected in 4 depth ranges bythe MOCNESS-10 opening and closingtrawl in the northern Sargasso Sea nearBermuda in single nighttime deploymentsin different seasons. The dotted lines showthe approximate separation between netsthat fished above or below the thermo-cline during the 2 seasons.

62 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Bonhommeau et al. 2008a,b; Miller et al. 2009a). One possible factor affecting survival may bethe availability of appropriately sized particles for feeding by the early stage larvae, since theymay be especially vulnerable to mortality after they deplete their yolk reserves. These factorshave been suggested to be potentially responsible for the declining populations of northern tem-perate anguillid eels, because correlations have been found between recruitment and variousfactors such a the North Atlantic Oscillation index (Knights 2003; Friedland et al. 2007; Kettleet al. 2008), temperature as a proxy for productivity (Bonhommeau et al. 2008a,b), or the likelylatitude of frontal features (Kimura et al. 2001; Kimura and Tsukamoto 2006; Friedland et al.2007) as is discussed in Section 5-5.
Another factor that would be important in determining the survival of leptocephali in theocean is their immune system. They appear to have components of an immune system based onresearch on A. japonica leptocephali, which detected the presence of lectin in the club cells thatline their bodies (Suzuki and Otake 2000). This may provide some defense against pathogensthat come in contact with their skin, but some specimens also showed free leukocyte-like cells inthe connective tissue of their guts. Other cells potentially related to the immune system wereobserved, but since most organs including those related to immune cell production are poorlydeveloped in leptocephali, the extent of their defenses against pathogens is unclear (Suzuki andOtake 2000).
It is also unclear how much predation occurs on leptocephali by other marine organisms.Their high degree of transparency likely reduces the ability of predators to see them. But thefragile and gelatinous nature of leptocephali, with few or no ossified hard parts, probably makesthem difficult to detect or identify in stomach content studies of pelagic fishes. There is someevidence though that predators can sometimes see them and catch them. Seabirds were observedfeeding their young large leptocephali off the coast of Brazil (Figueroa 1997), and “eels and eellarvae” were reported as being among the gut contents of dolphin fish (Coryphaena hippurus)collected east of Taiwan during winter (Wu et al. 2006); but the latter case appears to be unusualcompared to other studies of the food habits of dolphin fish. Some types of sea snakes are knownto target small moray eels as prey (Reed et al. 2002; Ineich et al. 2007; Brischoux et al. 2009),and marine mammals such as dolphins sometimes feed on eels when they are abundant (Amir etal. 2005), but there is apparently no predator that is known to feed regularly on leptocephali.
Transparency is an important strategy of a wide variety of organisms in the open ocean toreduce predation, by making it difficult for predators to distinguish the outline of the organism(Johnsen 2001). Leptocephali appear to have evolved a strategy of being almost totally transpar-ent. Although it is not evident in persevered or even recently deceased leptocephali in which thethin layer of muscle tissue on each side of the body is no longer transparent, live leptocephaliappear almost completely transparent (Figs. 1, 55). Selection pressure for transparency is onepossible reason that leptocephali do not form red blood cells until late in the metamorphic proc-ess, since red colored blood might be very visible during the daytime. The body surface ofleptocephali also appears to be very reflective (Fig. 55), which could provide crypsis in somelight conditions as reflective coloration does for many pelagic fishes (Johnsen 2003).
Another feature that may have evolved to reduce the ability of predators to see and captureleptocephali is their wide variety of pigmentation patterns. Some leptocephali have melanophoreson their heads, guts, side of their bodies (Figs. 13, 14, 56), internally, or even on the iris(Fig. 57E). The size and arrangement of these spots can range from large dendritic melanophoresor large patches of small melanophores, to tiny melanophores scattered all over the body (seeBlache 1977; Smith 1979; Böhlke 1989b; Miller and Tsukamoto 2004). One common pattern isa single row of melanophores or small rows of pigment on the myosepta between the myomeresthat extend along the midline of the body. Others patterns are melanophores spaced out alongthe entire length of the gut (e.g. congrids, Fig. 56), or pigment on top of the liver swellings(e.g. ophichthids, Fig. 18B). A remarkable new video observation of a swimming ophichthidleptocephalus at night off Hawaii (MJ Miller and MJ D’Avella unpublished manuscript) indi-cated that the pigment spots along its gut were enlarged dorsally and were paired structures(http://www.australianmuseum.net.au/movie/Ophichthid-leptocephalus-Kona-Hawaii/). Thismarked difference in shape and size compared to those of other ophichthid leptocephali after

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 63
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
being collected and preserved (Leiby 1989), suggests that the size and shape of some of theseinternal pigment patches containing melanophores and possibly other types of chromatophores,may be changeable and controlled by the nervous/and or endocrine systems in some ophichthidsas they frequently are in the skin of other fishes (Fujii 2000; Grether et al. 2004).
The various spots or pigment patches in leptocephali are likely designed to break up thevisual outline of the body, or the less transparent features such as the head, gut, and liverswellings while the leptocephali are moving, by a process referred to as flicker fusion (Moser1981). Many dark spots moving quickly within the body outline of a leptocephalus may disruptthe ability of the predator’s visual system to distinguish the outline of the leptocephalus. Simi-larly, black spots on various regions of the gut would likely break up the linear image of thetubular gut containing food, thus making it less visible within the rapid body undulations duringtheir anguilliform swimming motions.
A few taxa of leptocephali also have various kinds of yellow (xanthic) pigmentation, butthe function of this type of pigment is unclear. Yellow pigmentation is found on the anterior partof the esophagus and liver swellings of Neenchelys (Ophichthidae) and Saurenchelys(Nettastomatidae) (Figs. 57A, C), and internally around the eyes of some muraenids (Figs. 57B,F). At least one species of muraenid also has small yellow xanthophores scattered all over theside and top of its head, mixed with melanophores (Fig. 57E). Yellow pigment also commonly
Fig. 55. Frame-capture photographs of a chlopsid leptocephalus filmed after being captured and placed in an aquarium onboard a research vessel, showing it curling up in what is hypothesized to be behavioral mimicry of gelatinous zooplankton,which are typically avoided by predators.

64 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
occurs on the dorsal surface of the eye in some congrids and ophichthids (Fig. 57D). This yellowpigment is mixed with black spots and may be more common than can be judged from photo-graphs.
The function of this yellow pigment in leptocephali may be related to the wavelengths oflight that it absorbs. For example Warrant and Locket (2004) pointed out that the yellow lenses
Fig. 56. Photographs of various types of pigment spots (melanophores) on the guts or side of the body of leptocephali ofcongrids (A, F), chlopsids (B, C), a synaphobranchid (Synaphobranchinae) (D), and Leptocephalus holti (Cyematidae) (E).

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 65
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
of the eyes of some deep-sea fishes selectively absorb light in the blue part of the spectrum, thuscutting out most of the light that could have reached the retina. Removing most of the bluebackground light enhances the slightly greener counter-illumination, which could highlight theoutline of prey organisms. The yellow pigmentation in leptocephali may also selectively absorbblue light, which could serve to contribute towards camouflaging the larvae by making somebody features less visible. Yellow pigment has also been suggested to provide some protection
Fig. 57. Photographs of Neenchelys (Ophichthidae) (A), Muraenidae (B, E, F), and Saurenchelys (Nettastomatidae) (C),Ophichthidae (D), that show various types of yellow pigmented tissue on the tip of the snout, esophagus, liver swellings,around the eye, above the eye, or apparent chromatophores scattered all over the head.

66 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
against damage to fish larvae by ultraviolet (UV) and visible light (Moser 1981). It may functionthis way around their eyes, or on their liver tissue. In the case of the species with yellow pigmentpatches above their eye (Fig. 57D), it may absorb light shining down from above.
These types of questions and the issue of the survival rates of leptocephali are presentlyjust a matter of speculation due to a lack of information. Observations of live leptocephali makeit clear that their ability to swim both forwards and backwards is likely an important factor inavoiding predators. Having the choice to immediately evade an approaching predator in twopossible directions makes it difficult for the predator to know which direction the leptocephaluswill go when attacked. In addition, it is possible that leptocephali can use a form of behavioralmimicry to confuse some predators as they rapidly approach them. A leptocephalus of Chilorhinussuensoni (Chlopsidae) observed in an aquarium on board a research vessel in the Sargasso Seaexhibited a curling up behavior to form a tight ball (Fig. 55). Many fish species are known tomimic other organisms that most fishes avoid eating (Randall 2005), so it is possible that lepto-cephali curl up so that they will appear to be a jellyfish, salp, siphonopore, or ctenophore(Fig. 55). These transparent gelatinous invertebrates are typically found at the same depths asleptocephali but are unpalatable. A species of congrid leptocephalus has also been observedperforming this same behavior (pers. observ.), so more information is needed to determine ifthis curling behavior may be a common predator avoidance strategy of leptocephali.
Leptocephali are also occasionally vulnerable to ectoparasites, including those that appearto be some type of parasitic hydroid (Fig. 58). This type of parasite is rare on leptocephali in theIndo–Pacific, but it also has been observed on a few leptocephali collected in the western NorthAtlantic (pers. observ.). How many leptocephali may die from this parasitism and thus not becollected is unknown. It is also possible that some larger invertebrates have the ability to prey onsmall leptocephali considering that a small nemichthyid leptocephalus was recently documentedto have been eaten by a chaetognath (Fig. 59; Johnson et al. 2006). Whether or not the lepto-cephalus was captured and ingested while within the net is not known, but it shows thatchaetognaths may be predators of small leptocephali. Future research is needed to evaluate pre-dation pressure on leptocephali and to determine what other factors, such as poor feeding condi-tions, can affect their survival.
4-3. Recruitment behavior
All different types of leptocephali live in the surface layer of the ocean ranging from theouter edge of the continental shelf to far out in the open ocean, but each species must return to itsown particular juvenile growth habitat. For anguillid eels, recruitment (defined here as enteringtheir juvenile habitats) occurs after metamorphosis into glass eels, which then enter freshwaterand estuarine habitats. Various studies have examined the recruitment patterns of anguillid glasseels in both temperate and tropical regions (e.g. Sugeha et al. 2001b; Sullivan et al. 2006, 2009).Less studied are the larvae of mesopelagic eels, which presumably metamorphose at a certainsize and move into deeper water to start their juvenile stage, based on studies of the life historiesof these eels (e.g. Beebe and Crane 1936; Böhlke 1989a). In contrast, leptocephali of families ofshallow water and continental slope eels must move away from the deep ocean and recruit to theappropriate depths for each species. Species that live as juveniles and adults in very shallowwater coastal areas over the shelf, such as around tropical coral reefs, in sandy areas, or onbanks, must enter these habitats from deeper water.
Although the leptocephali that recruit to those habitats have never been studied in particu-lar to determine the factors affecting their recruitment patterns, some useful data have beenobtained in general studies on fish larvae that recruit to coastal areas in tropical habitats. Theselarval recruitment studies were conducted using either channel nets anchored to the bottom, butfishing just below the surface, or crest nets attached to the bottom in small channels in the outerreef crests. Both types of nets have collected substantial numbers of recruiting leptocephali insome areas.
One of these areas that was sampled using channel nets was part of research designed tostudy the recruitment patterns of grouper larvae moving onto the large shallow area referred to

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 67
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
as the Great Bahama Bank (GBB) of the northern Bahamas (Shenker et al. 1993). The GBBis a distinctively shaped bank located at the southwest corner of the Sargasso Sea, which dropsoff very rapidly to depths >1000 m on all sides (Figs. 45, 60B, C). Shenker et al. (1993) usedchannel nets primarily fished in the upper 1 m that were deployed at 4 locations near Lee Stock-ing Island on the west side of Exuma Sound of the GBB (Fig. 60A) to sample fish larvae flowingonto the banks during flood tides. During two years of sampling from December to February,8,067 leptocephali were collected, which was far more that any other family of fish larvae(Thorrold et al. 1994a). Almost all (95.7%) of those leptocephali were collected at night, and thecatches showed clear periodicity (Fig. 60D) correlated to the number of hours of no moonlightwith strong flood tide and wind (Thorrold et al. 1994b). These catches had a periodicity of28 days (Thorrold et al. 1994b) and the catches at all 4 locations showed the same temporalpatterns (Thorrold et al. 1994a), with location 2 providing the most leptocephali and location 4providing the fewest (Shenker et al. 1993).
In the same kind of sampling in summer, leptocephali of the Ophichthidae (N = 2605)were most abundant, but moringuids (N = 807), congrids (N = 400), chlopsids (N = 343) andmuraenids (N = 157) were also collected (Thorrold et al. 1994c). During the summer though,labrid and clupeid larvae were more abundant than all leptocephali combined. The catches in thesummer also showed a clear periodicity with a 14-day semilunar cycle that was clearly seen inthe catches of ophichthids (Fig. 60E; Thorrold et al. 1994c).
Other studies on the recruitment of fish larvae have used crest nets, which sampled tidalflows over the outer edge of barrier reefs in tropical coral reef areas in western Australia (McIlwain2003), French Polynesia (Dufour et al. 1996), and the Caribbean Sea (Nolan and Danilowicz2008). In western Australia, 1,430 leptocephali were collected in two Oct/Nov to March sam-pling seasons, with four families of other larval fishes having higher abundances. The objectiveswere to examine the magnitude of larval supply between years and its periodicity, and to com-pare the synchrony in larval supply at scales of 500 m and 5 km. In the area sampled in bothyears, 1.7 times as many leptocephali were collected in the first year. Leptocephali showed asignificant correlation in arrival data over 5 km, but at a 1-day lag. Interestingly, of all the taxaof fish larvae collected, leptocephali showed the strongest lunar cycle of catches, with a perio-dicity of 27 days (McIlwain 2003). In French Polynesia, leptocephali were the fifth mostabundant taxon of large fish larvae collected in crest nets at 5 sites on different sides of Moorea
Fig. 58. Photographs of an unidentified parasite attached to a congrid leptocephalus.

68 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Fig
. 59.
Pho
togr
aphs
of a
25
mm
cha
etog
nath
(Sag
itta
hex
apte
ra) t
hat h
ad in
gest
ed a
n ap
prox
imat
ely
17 m
m N
emic
hthy
s sc
olop
aceu
s le
ptoc
epha
lus.
The
lept
ocep
halu
s is
fol
ded
and
both
its
head
and
tail
poi
nt to
war
ds th
e an
teri
or e
nd o
f th
e ch
aeto
gnat
h. R
epri
nted
fro
m J
ohns
on e
t al.
(200
6).

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 69
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Island, with 2,399 being collected (Dufour et al. 1996). Various patterns of lunar cycles of catcheswere observed at these sites also. A more recent study compared the catches of both channel netsand crest nets at Turneffe Atoll off Belize to evaluate which type of net is better for collectingrecruiting fish larvae (Nolan and Danilowicz 2008). Crest nets were found to consistentlycollect more larvae than the channel nets fished at the surface in mangrove channels, and thiswas clearly true for leptocephali, with 81% being caught in the crest nets. This study also seemedto be generally consistent with previous studies, with higher catches being related to the lunarcycle and the strength and direction of winds, but leptocephali were not analyzed separately ordistinguished into different taxa.
Fig. 60. Maps of the locations of 4 channel nets (red squares) that were used to collect ingressing leptocephali and otherlarval fishes around Lee Stocking Island on the Great Bahama Bank (GBB) (A), the location of the study area in relation to thelarger region of the GBB (B) and in a satellite image of the GBB that shows the deep water areas of Exuma Sound and theTongue of the Ocean (TO) on either side of the study area (C); and plots of the catches of all species of leptocephali in thewinter season (D) and of the family Ophichthidae in the summer season (E). (A) Adapted with permission from MarineEcology Progress Series, Onshore transport of settlement-stage Nassau grouper Epinephelus striatus and other fishes inExuma Sound, Bahamas. 98, 1993, 31–43. Shenker JM, Maddox ED, Wishinski E, Pearl A, Thorrold SR, Smith N, Figure 1.© 1993, Inter-Research.; (D) adapted with kind permission from Dr. Shenker and Springer Science + Business Media: MarineBiology, Larval supply of shorefishes to nursery habitats around Lee Stocking Island, Bahamas. II. Lunar and ocanographicinfluences. 118, 1994, 567–578. Thorrold SR, Shenker JM, Maddox ED, Mojica R, Wishinski E, Figure 2. © Springer-Verlag;(E) adapted with permission from Marine Ecology Progress Series, Temporal patterns in the larval supply of summer-recruit-ing reef fishes to Lee Stocking Island, Bahamas. 112, 1994, 75–86. Thorrold SR, Shenker JM, Mojica R, Maddox ED,Wishinski E, Figure 4. © 1994, Inter-Research.

70 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
These various studies that have documented leptocephali recruiting into tropical habitats,raise some interesting questions. The cyclical patterns of recruitment suggest the possibility thatleptocephali are abundant in the waters just offshore of the banks where they have been col-lected, and that they may wait for the appropriate environmental conditions to move onshore. Itis also possible that passive oceanographic processes combined with net avoidance during moon-light could explain these recruitment processes if stronger tides simply transport more lepto-cephali onshore. However, it is possible that leptocephali are adapted to time their inshore move-ments during periods with the strongest tidal transport and the lowest risk of predation. If this isthe case, then it is also possible that leptocephali from offshore areas may accumulate just off-shore near the shelf break and then remain nearby using active swimming behavior until it is theappropriate time to move inshore.
The observations of leptocephali recruiting into shallow water indicate that many speciesenter these habitats as leptocephali, and not as glass eels as in anguillids. A similar patternoccurs with Conger oceanicus, which is spawned offshore in the Sargasso Sea, but recruits backto temperate latitudes as leptocephali (Able and Fahay 1998; Bell et al. 2003). These Congerleptocephali have been captured while entering estuarine habitats in New Jersey (Fig. 36; stageER–M1) from April–June (Wuenschel and Able 2008). Leptocephali of the common ophichthidMyrophis punctatus also enter this estuarine system (Able and Fahay 1998), and probably alongthe entire coastline to the south and into the Gulf of Mexico, since for example, they are alsocaptured in tidal currents flowing into estuarine habitats in North Carolina (Pfeiler and Govoni1993). In East Asia, Conger myriaster also are commonly collected in shallow coastal areas asthey begin to metamorphose (e.g. Otake et al. 1994; Lee and Byun 1996). Future research usingappropriate sampling gear in carefully selected locations will likely find that leptocephali ofmany species of common coastal marine eels can be collected as they enter their growth habitatsin areas ranging from tropical to southern temperate latitudes worldwide.
5. General discussion and future perspectives
5-1. Biology of leptocephali
Much remains to be learned about the biology of leptocephali, because in some respects,what has been learned about their morphology and behavior creates more questions than itanswers. Their highly compressed transparent bodies seem to act as energy storage reservoirsthat also provide structural support (Pfeiler 1999; Pfeiler et al. 2002), buoyancy (Tsukamoto etal. 2009) and a strong swimming ability (Wuenschel and Able 2008) until metamorphosis.They live in the surface layer (Figs. 50–54) where there is the highest degree of biologicalactivity and food availability and at least some of them show vertical migrations like many othermarine organisms (Lampert 1989). Due to a minimum amount of respiring tissue, their meta-bolic demands are low (Pfeiler and Govoni 1993; Bishop and Torres 1999), thus enabling themto feed on readily available bacteria or protozoa attached to particulate matter that contain littleenergy (Otake et al. 1993; Mochioka and Iwamizu 1996); yet they apparently grow quite fast(Table 1) and reach large sizes.
The hypothesis that leptocephali feed exclusively on particulate matter or its associatedDOC, is supported by two general factors. The first is that no zooplankton have ever been seenin their guts, but their guts typically contain a detritus-like material. Second, stable isotopestudies indicated the trophic level of leptocephali is at the lowest levels of the food chain (Otakeet al. 1993; Kimura and Tsukamoto 2006). This type of feeding strategy is logical, however,because marine snow and discarded larvacean houses are typically very abundant in the surfacelayer of the ocean, and by feeding on this material, they will not directly compete for food withmost other fish larvae. Zooplankton fecal pellets (Turner 2002), discarded larvacean houses(Alldredge 1976), and marine snow (Alldredge and Silver 1988) are common components of thewater column in the upper few hundred meters of the ocean. These particulate materials formaggregates containing bacteria, phytoplankton, and protozoa, as well as the mucus-like larvaceanhouses that collectively form marine snow (Alldredge and Silver 1988), which is a food source

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 71
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
of various types of zooplankton (Alldredge 1976). Some fish larvae also feed directly on dis-carded larvacean houses, because they are often abundant and they are immediately replaced bya new house when their filters clog (Alldredge 1972, 1976; Sato et al. 2003). Other fish feeddirectly on the larvaceans themselves (Capitanio et al. 2005; Llopiz and Cowen 2009), but byfeeding only on discarded larvacean houses or marine snow, leptocephali do not directly impactthe organisms producing their food source.
Particulate organic material that includes marine snow and discarded larvacean houses istypically most abundant in the upper 100 m of the water column in regions such as the NorthPacific subtropical gyre and along its western margins (Ichikawa and Nishizawa 1975;Ichikawa 1982; Hebel and Karl 2001; Pilskaln et al. 2005). Because marine snow is most abun-dant in the surface layer where leptocephali live, it is a logical food source for them. Somelarvacean houses (Galt et al. 1985) and some types of marine snow (Herren et al. 2004) havebioluminescent properties that could make it possible to be seen and eaten at night by lepto-cephali which appear to be adapted for night vision. It remains to be determined, however, underwhat types of light conditions and depths leptocephali feed on particulate material.
Another major question is whether there are any differences in feeding strategies or thematerials that different species eat. The greatly different lengths and orientations of theteeth of small leptocephali compared to the larger ones suggest that there may be a specificrange of size of particles that they primarily target. Based on the relative teeth size, these parti-cles may be large compared to the mouth of the small engyodontic stage leptocephali with largeteeth (Figs. 9, 11), and smaller relative to the head of larger euryodontic stage leptocephali withsmall teeth (Figs. 11, 14, 15). The morphological diversity of head and jaw structures suggestthe possibility that various species may use different tactics to locate food or may target differ-ent food sources. For example, are the wide range of jaw shapes among taxa of leptocephalirelated to feeding on different types of particulate matter? Or are they simply related to thedevelopmental constraints during metamorphosis with the larvae needing to have similar jawstructures to the adults, as may be the case in the gulper eels (Eurypharynx pelecanoides, Figs. 9,10, 15). The leptocephali of Thalassenchelys that reach very large sizes, but whose adults areunknown (apparently related to Colocongridae, Lopez et al. 2007), have two larger front teeththat differ from other species (Fig. 15C), so it is possible that these teeth are related to feeding ona special type of food source consumed by the larger leptocephali that can reach >300 mm.
The most extreme examples of morphological specializations likely related to feedingare the upward pointing telescopic eyes of the Synaphobranchidae (Fig. 15E), and the rostralfilaments possessed by some species of the synaphobranchid subfamily Ilyophinae (Fig. 16).Telescopic eyes are commonly seen in deep-sea fishes and are thought to be used in feeding forlooking upward to see the silhouettes of prey above in very low light conditions (Warrant andLocket 2004). More difficult to understand, though, are the apparently cartilaginous extensionsfrom the tip of the rostrum of some species. They can be quite long or even have branchingstructures or palp-like extensions. They have never been studied in particular, so their structureand function is unknown. One idea is that the long filament could be used to locate food in verylow light conditions by waving it back and forth to feel for larvacean houses or other largepieces of particulate matter. Using them to stimulate bioluminescent organisms to produce lightthat they could use for visualizing food items is another speculative idea. The palp-like struc-tures seen in Fig. 16C look very similar to the bioluminescent barbels of many types of deep-seafishes, but presently the function of these interesting structures cannot be determined withoutdetailed morphological analyses, or chance observations of their behavior in the ocean.
Other more achievable research problems still remain regarding the biology of leptocephali,however, such as their comparative anatomy, physiology, depth distribution and feeding behavior.For example why do leptocephali almost exclusively reside in the upper 100 m at night and whydo some species vertically migrate during the day and others apparently do not? If leptocephaliare visual feeders as appears to be likely, feeding in daytime is perhaps most likely, unlessthey can target food particles with bioluminescent properties, as mentioned previously. Analternative hypothesis is that they reside in the upper layers at night to be in the warmest waterto facilitate faster growth, but some vertically migrate to deeper, lower light regions presumably

72 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
to avoid predation during the day. Many marine fishes have UV vision (Johnson and Widder2001) as do some larval fishes (Britt et al. 2001; Browman et al. 2006), but it is unclear if thishas any relationship to the ecology of leptocephali. There appears to be enough light for UVvision down to 200 m (Losey et al. 1999), which could facilitate feeding at deeper depths duringthe day, or alternatively the UV vision of predators could somehow make leptocephali morevisible at shallow depths during the day. More data are needed on the daytime depth distribu-tions of leptocephali, because existing data are clearly biased by net avoidance.
Other basic issues that need to be examined are the ecology of metamorphosis and recruit-ment. The triggers of metamorphosis are presently unknown for leptocephali and the recruit-ment dynamics of marine eels have never been examined. Can all leptocephali delay metamor-phosis if they reach their maximum size but are drifting far offshore and how many actually waitnearby their recruitment areas before swimming onshore when they reach a certain size? Byapplying otolith aging techniques to a wider range of species and recruiting leptocephali inaddition to those collected offshore, the mysteries of what happens when these fragile, transpar-ent larvae change into threatening, voracious predators may be revealed.
5-2. Leptocephalus growth
Progress has been made towards understanding the growth rates of leptocephali, whichgrow up to much larger sizes than most fish larvae, but relatively few anguilliform species havebeen studied. Analyses of otolith microstructure of leptocephali and glass eels have typicallyobserved well-defined increments, with the exception of the outer opaque zone that apparentlyoccurs when leptocephali move through cold waters and experience slow growth (Lee and Byun1996; Antunes and Tesch 1997). Species collected in the mostly warm Indo–Pacific region havegrowth rates <1 mm/d and linear relationships between age and body length, with growth slow-ing down as maximum size is reached (Table 1; Figs. 30–32). These relationships and the find-ing of apparent lunar cycles of spawning based on backcalculated hatching dates (Ishikawa etal. 2001; Tsukamoto et al. 2003; Lee et al. 2008), suggest that increments are deposited daily inthese species (Powles et al. 2006).
There are differences in growth rates among species, however, which may be related totheir maximum size and their distances of migration, as proposed by Kuroki et al. (2006a) foranguillid leptocephali. Larval growth rates and timing of metamorphosis may be key factors inthe early life histories of anguillid eels for defining their recruitment areas (Chen and Tzeng1996; Wang and Tzeng 2000; Shiao et al. 2002; Kuroki et al. 2006a, 2008a). There also may bedifferent growth strategies of marine eel leptocephali, related to their maximum body sizes ordispersal strategies as discussed below. This hypothesis is supported by the observation that thegrowth rate of the leptocephali of a synaphobranchid species with a smaller maximum size waslower than the growth rate of a nettastomatid species with a larger maximum size, which werecollected during the same survey in the East China Sea (Fig. 32).
These types of inter-taxon differences could be a contributing factor to the findings of theage and growth study on leptocephali from the Gulf of Mexico, which found similar results tothose of Indo–Pacific species for one species, but found much higher variability and some veryhigh apparent growth rates for the three other species that were studied (Bishop et al. 2000). Thecause of this variability is unknown because otolith microstructure of these taxa has not beenexamined in the Indo–Pacific. It is possible that the conditions for leptocephalus growth arebetter (or sometimes worse) in the eastern Gulf of Mexico, or that the species studied frequentlyhave particularly high growth rates compared to other species that have been studied in theIndo–Pacific. However, it can not be assumed that visible otolith increments are deposited dailyin all leptocephali or in all larval stages of eels, since so little is known about the biology andbehavior of most species.
Deposition of daily increments is thought to be related to physiological cycles that causeincrement formation, which in the case of fish larvae, could be linked to a cessation of feeding atnight or vertical migration into lower temperatures in deeper water. However, there is evidencethat otolith increments might stop being deposited in some anguillid glass eels, based on the

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 73
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
mismatch of many of their back-calculated hatching dates and the known spawning season oftemperate species such as Anguilla rostrata, as indicated by collections of small leptocephali intheir spawning area (McCleave 2008). This period when increments are not deposited likelyoccurs during the glass eel stage because studies on metamorphosing anguilliform and elopiformleptocephali have found that otolith deposition continues during metamorphosis (Chen and Tzeng1996; Powles et al. 2006) and wider increments are formed (Figs. 38, 39). In addition, experi-ments have shown that glass eels held at temperatures of 12°C or lower stop depositing otolithincrements (Umezawa and Tsukamoto 1991; Fukuda et al. 2009).
Since glass eels typically do not feed until they begin their upstream migration, if theyhide in the substrate and wait for the appropriate water temperature or other cues to start theirupstream migration (see Linton et al. 2007), a hypothesis can be proposed that there may be areduction or cessation of otolith deposition as a result of their lack of feeding or activity. Thiscould occur during such a period of inactivity, because otolith deposition requires the secretionof otolith matrix proteins, and the entry of calcium into the endolymph that is then crystallizedonto the otolith surface, with increments being formed by alternation between mineral-rich andorganic-matrix-rich layers (Campana 1999; Morales-Nin 2000; Takagi et al. 2005). Theseprocesses may be partially under hormonal control (Morales-Nin 2000), and a lack of feeding,or stress to the fish, has been shown to disrupt normal increment formation in some fishes (seereferences in Fukuda et al. 2009). So the lack of feeding or activity in glass eels that may hide inthe substrate to conserve energy until they begin their upstream migration, or hormonal changesdue to inactivity, could result in minimal otolith deposition in the case of glass eels, which maybe particularly adapted to conserve energy until they migrate upstream. This period of lack ofdeposition may correspond to what has been referred to as the “freshwater check” (Fig. 38) asmentioned previously (Kuroki et al. 2008b; Fukuda et al. 2009). If this check is actually formedwhen glass eels stop migrating and wait in the substrate for a variable period of time, the fresh-water check would represent an accurate age at first recruitment if daily increments were formedprior to its formation. But due to the potential missing time not recorded by visible incrementswithin the zone of the check, the backcalculated hatching dates of glass eels estimated from theotolith rings would not be accurate, as was indicated by McCleave (2008).
In contrast to inactive anguillid glass eels, species of leptocephali that have a dailycycle of feeding and growth or vertical migration between different temperatures (Figs. 50, 51;Castonguay and McCleave 1987a) will likely deposit daily increments in their otoliths. Thismay occur in many species of leptocephali, but in some metamorphosing Conger leptocephali(Lee and Byun 1996; Correia et al. 2002a, 2003) or in the otoliths of some late stage A. anguillaleptocephali or glass eels (Antunes and Tesch 1997), there is a diffuse zone in the outer regionsof their otoliths where increments are irregular or are not clearly visible using standard etchingtechniques. This zone may correspond to a region of slow but continuous otolith growth beforeand after metamorphosis, possibly due to the fish experiencing low water temperatures (Antunesand Tesch 1997). Both A. anguilla and the temperate species of Conger would likely experiencea period of cold water temperature during the last stages of their migration to coastal areas thatmay cause a much slower growth rate in the leptocephali or reduced metabolism in the glass eelsthat disrupts the formation of clearly visible daily increments. With a technique of otolith etch-ing at a different angle though, Lee and Byun (1996) were able to discern faint increments, eventhough efforts using various other etching techniques could not (Correia et al. 2006).
A better understanding of otolith daily increments will help to resolve the question ofwhether or not A. anguilla leptocephali can actually cross the North Atlantic and recruit at agesof one year or less as suggested by increment counts (Lecomte-Finiger 1992, 1994), or if it takesconsiderably longer as discussed by McCleave et al. (1998). Modeling studies of larval trans-port of A. anguilla leptocephali suggest it would take about 2 years or less to cross the Atlanticbasin using currents (Kettle and Haines 2006; Bonhommeau et al. 2009a,b). However, the pos-sibility of these leptocephali using directional swimming behavior to speed up the migration cannot be excluded, since leptocephali are capable swimmers (Wuenschel and Able 2008). Thesetypes of questions highlight the need for more knowledge about the factors regulating growthand otolith deposition in leptocephali and glass eels. Further otolith studies are needed on a widerange of anguillid and marine eel leptocephali to document their growth rates and larval durations.

74 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
5-3. Zoogeography and diversity of leptocephali
Because leptocephali have a long larval duration, their patterns of distribution and abun-dance can be complex and determined by a variety of factors. Three key factors are likely todetermine the spatial and temporal abundance of anguilliform leptocephali in each particulararea of the world’s tropical and subtropical ocean regions. The first is the biodiversity of eels inthe region and the distribution of available habitat for anguillid eels and for shallow water orcontinental slope marine eel species. For example, the biodiversity of leptocephali is very highin the central Indonesian Seas (Wouthuyzen et al. 2005), and to a lesser degree near the GBB ofthe northern Bahamas (Fig. 45; Miller and McCleave 2007), but is much lower in offshore areasof subtropical gyres (Fig. 4; Miller and McCleave 1994; Miller et al. 2006a).
A second factor that determines abundance is where the various species spawn in relationto ocean surface currents. Many species appear to spawn along outer continental shelves, suchas in the East China Sea (Miller et al. 2002a; Minagawa et al. 2005; Minagawa and Miller 2005)and the southwest Sargasso Sea (Miller 1995; Miller and McCleave 2007), where densities andspecies richness of leptocephali are likely much higher than offshore where fewer species spawn.In some areas, strong currents pass by the edge of continental shelves where eels spawn, such asthe western boundary currents of subtropical gyres (the Florida Current/Gulf Stream, the KuroshioCurrent, and the East Australian Current). These currents and their recirculations likely have amajor effect on the distributions of leptocephali in the western regions of these gyres. Moreprecise information on spawning areas for each type of marine eel is needed, since there appearsto be a range of distances of spawning migrations, as is discussed below.
The third factor in leptocephalus abundance is the seasonality of spawning of differentspecies of eels. Temperate anguillid eels clearly have seasonal cycles of migration and offshorespawning that result in their leptocephali being abundant in the spawning areas and larval mi-gration routes at certain times of year. Less is known about the spawning seasons of tropicalanguillids that appear to spawn at various times of the year (Arai et al. 2001; Kuroki et al.2006a), but may have seasonal cycles of downstream migration and spawning linked to tropicalmonsoon cycles (Wouthuyzen et al. 2009). Similarly little is known about seasonal patterns ofspawning of most marine eels, but some information is available in a few areas of the worldwhere collections have been made at multiple times of year, such as off South Africa (Castle1968), the Sargasso Sea (Miller and McCleave 1994, 2007), Gulf of Mexico (Smith 1989a),Gulf of Guinea (Blache 1977), or at least during two times of the year near Japan (Miller et al.2002a; Kimura et al. 2006; Minagawa et al. 2007) and in the Indonesian Seas (Miller et al.2002b,c, 2004; Wouthuyzen et al. 2005). These studies have found that some taxa of marine eelsshow clear seasonal patterns of spawning, but others do not. In offshore areas the presence ofleptocephali of at least some mesopelagic eels such as derichthyids and serrivomerids is sea-sonal (Castonguay and McCleave 1987b; Miller and McCleave 1994, 2007; Miller et al. 2006a;Miller and Tsukamoto unpubl. data). Much more research is needed to determine the spawningseasons of each type of marine eel and for tropical anguillids in order to gain a better under-standing of the seasonal cycles of distribution and abundance of leptocephali in each region ofthe world.
In the Indo–Pacific and in other regions outside of the western North Atlantic, a majorobstacle to progress in understanding the seasonal presence and geographic distribution patternsof leptocephali is the inability to identify most leptocephali to the species level. This is due to analmost complete absence of morphological features or non-overlapping meristic characters pos-sessed by both the larval and adult forms of each species. This is especially true for highlydiverse families, such as Muraenidae, which appear to have many leptocephali within the genusGymnothorax that are very similar morphologically (Castle 1965a; Smith 1989b). In addition,the larvae of some muraenid eels have never been distinguished, possibly a result of differentpatterns of habitat use or larval behavior, as suggested by an extraordinarily large greenish meta-morphosing leptocephalus, which was photographed recently as it was apparently recruiting toa shallow water coral reef area in southern Indonesia (Fig. 61; Miller et al. 2009b).
Due to the greater number of species of marine eels in the Indo–Pacific, the problem ofleptocephalus identification represents a major challenge that will be greatly facilitated using

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 75
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
genetic techniques for matching larvae with adults of the same species (Miller and Tsukamoto2006). DNA sequences have already been used to identify some congrid leptocephali (Ma et al.2007, 2008a) or to report cryptic genetic lineages within widely distributed species, such asUroconger lepturus (Ma et al. 2008b) or bonefishes (Colborn et al. 2001). As adult geneticsequences become available, greater use of genetic identification techniques can be made, likethose being used to identify anguillid leptocephali (e.g. Aoyama et al. 1999, 2003, 2007) andproposed for fish larvae in general (Richardson et al. 2007; Teletchea 2009). For example, DNAbarcoding techniques have recently been applied to many fish species in Australia (Ward et al.2005). Real-time polymerase chain reaction (PCR) techniques can also be used to identify eggsand small leptocephali onboard research vessels to enable targeted sampling strategies duringcruises (Watanabe et al. 2004; Minegishi et al. 2009). Genetic sequence analyses also have beenused to suggest the possible phylogenetic relationship of the large leptocephali of Thalassenchelys(Fig. 62A) for which there are no known adults (Lopez et al. 2007); and similar studies canreveal new understanding about other unknown leptocephali such as the Type I of Smith (1979)(Fig. 62B). Using genetic sequencing techniques in conjunction with morphological descrip-tions of leptocephali, a better understanding of the biodiversity, zoogeography, and life historiesof anguilliform eels will be possible in the Indo–Pacific and other regions of the world.
5-4. The leptocephalus larval strategy
A leptocephalus larva is the seemingly the only shared character of the ancient and diversefishes of the Elopomorpha (Inoue et al. 2004), which may have existed for about 220–260 mil-lion years (Inoue et al. 2005), and the leptocephalus larval strategy may be a key component of
Fig. 61. Photographs of a large metamorphosing leptocephalus with an apparent greenish coloration that was observedmoving slowly over the bottom in a coarse sand and coral clump area near Sangeang Island in the Flores Sea of southernIndonesia. This leptocephalus appeared to be larger than 400 mm, and was likely a never-before collected species of theMuraenidae. Reprinted with permission from Miller MJ, Powell J, Tsukamoto K. Observation of a large metamorphosingleptocephalus in a coral reef habitat at Sangeang Island, Indonesia. Zool. Stud. 2009b; 48: 107.
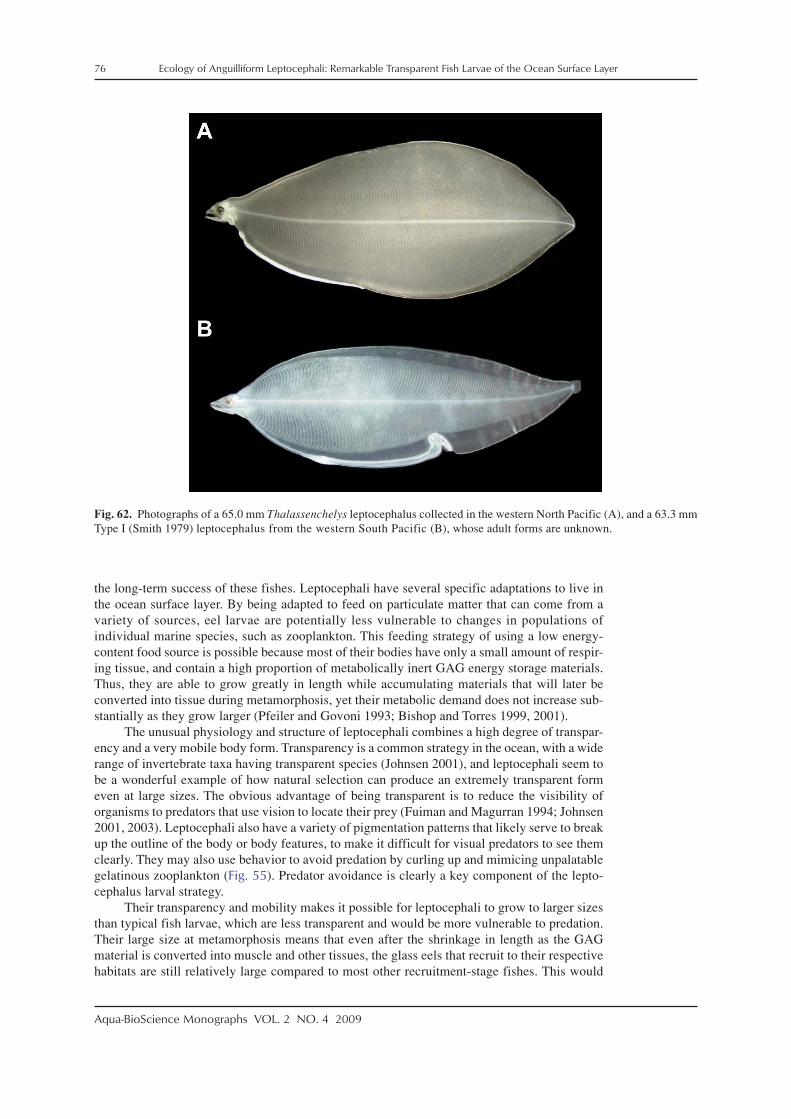
76 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Fig. 62. Photographs of a 65.0 mm Thalassenchelys leptocephalus collected in the western North Pacific (A), and a 63.3 mmType I (Smith 1979) leptocephalus from the western South Pacific (B), whose adult forms are unknown.
the long-term success of these fishes. Leptocephali have several specific adaptations to live inthe ocean surface layer. By being adapted to feed on particulate matter that can come from avariety of sources, eel larvae are potentially less vulnerable to changes in populations ofindividual marine species, such as zooplankton. This feeding strategy of using a low energy-content food source is possible because most of their bodies have only a small amount of respir-ing tissue, and contain a high proportion of metabolically inert GAG energy storage materials.Thus, they are able to grow greatly in length while accumulating materials that will later beconverted into tissue during metamorphosis, yet their metabolic demand does not increase sub-stantially as they grow larger (Pfeiler and Govoni 1993; Bishop and Torres 1999, 2001).
The unusual physiology and structure of leptocephali combines a high degree of transpar-ency and a very mobile body form. Transparency is a common strategy in the ocean, with a widerange of invertebrate taxa having transparent species (Johnsen 2001), and leptocephali seem tobe a wonderful example of how natural selection can produce an extremely transparent formeven at large sizes. The obvious advantage of being transparent is to reduce the visibility oforganisms to predators that use vision to locate their prey (Fuiman and Magurran 1994; Johnsen2001, 2003). Leptocephali also have a variety of pigmentation patterns that likely serve to breakup the outline of the body or body features, to make it difficult for visual predators to see themclearly. They may also use behavior to avoid predation by curling up and mimicing unpalatablegelatinous zooplankton (Fig. 55). Predator avoidance is clearly a key component of the lepto-cephalus larval strategy.
Their transparency and mobility makes it possible for leptocephali to grow to larger sizesthan typical fish larvae, which are less transparent and would be more vulnerable to predation.Their large size at metamorphosis means that even after the shrinkage in length as the GAGmaterial is converted into muscle and other tissues, the glass eels that recruit to their respectivehabitats are still relatively large compared to most other recruitment-stage fishes. This would

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 77
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
likely reduce the range of predators as the young eels enter their juvenile growth habitats, as hasbeen indicated for other types of fish larvae (Miller et al. 1988).
Another aspect of the leptocephalus larval strategy is that it appears to be adapted for usingthe relatively predator-free open ocean for feeding and growth. The threat of predation is likelymuch lower offshore because productivity is lower and there is typically less food to supportlarge predator populations. So by feeding on a ubiquitous food source such as particulate mate-rial, eels are able to use the open ocean for an extended period of larval development to reachlarge sizes before recruitment.
There are variations in the geographic patterns of how leptocephali use the open ocean,though, because the distributions of various taxa are quite different. Temperate and some tropi-cal anguillid larvae have a strategy of using westward flowing currents to transport them back totheir recruitment areas while they feed and grow to their maximum larval size (Tsukamoto et al.2002; Kuroki et al. 2008a, 2009a,b). The alternative strategy of local spawning and larval reten-tion near their adult growth habitats would probably not be successful in temperate regions, dueto the accumulations of large numbers of leptocephali along the continental margins where thereare many predators and colder temperatures. A different strategy used by some marine eelsappears to be based on using the recirculation patterns of the western regions of subtropicalgyres, as in a population of Ariosoma balearicum that lives adjacent to the Sargasso Sea(Miller 2002a) and Ariosoma major that lives along the margin of the western North Pacific(Miller et al. unpl. manuscript). A short migration offshore to spawn on the eastern side of thewestern boundary current of each subtropical gyre, appears to result in the larvae of these Ariosomaspecies using the entire western regions of each gyre as larval development areas (Miller 2002a),and not just the low latitude parts of the gyres like in anguillids. A similar strategy, but at asmaller scale, is used by tropical anguillids that migrate short distances offshore to spawn overdeep water in regions where their leptocephali will remain near the species range and can recruitback to that same general area. Species such as A. borneensis and A. celebesensis in the centralIndonesian Seas are examples of this (Aoyama et al. 2003; Kuroki et al. 2006a). The otherapparent strategy is to spawn over or near the edge of the shelf so that at least a portion of thelarvae will remain near their adult habitats. Most muraenids, ophichthids, and garden eels ap-pear to use this spawning and larval distribution strategy.
There may be variations in the larval dispersal patterns resulting from this local spawningpattern, however, that could be due to different behavioral tactics used by some leptocephali.The distinctly lower offshore abundances of garden eel leptocephali and of most ophichthidspecies suggest these taxa of leptocephali may use swimming behavior or some other mecha-nism to stay near the continental shelf or around islands where they were born. There is increas-ing evidence that larval swimming behavior and self-recruitment are important for many coralreef fishes (see Leis 2002, 2006; Leis and McCormick 2002; Leis and Carson-Ewart 2003), so itis likely important for some marine eels as well. Olfactory cues or the sound of coral reefs havebeen investigated as possible cues for fish larvae to find recruitment habitats when they reachsettlement stage (Atema et al. 2002; Leis and Lockett 2005). The strong swimming ability ofleptocephali in conjunction with their large size could enable them to swim back to the shelfeven if they were transported offshore during their early larval stage. Future studies on the fine-scale distribution of leptocephali in relation to the shelf break and ocean currents and on theswimming ability of different species will help to reveal which species appear to use larvalretention strategies and which are more adapted for dispersal.
5-5. Oceanic changes and leptocephalus recruitment
The leptocephalus strategy appears to be well adapted to life in the ocean surface layer, butthere are increasing indications that the population dynamics of some eel species may be linkedto changes in the ocean that affect their larvae. Declines of northern temperate anguillids havebeen hypothesized to be related at least in part to changes in the ocean (Castonguay et al. 1994;Knights 2003). Studies testing for correlations between recruitment and various physical andbiological parameters in the ocean suggest that the leptocephalus stage may be vulnerable to

78 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
changes in the ocean resulting from fluctuations of the ocean–atmosphere system (see Miller etal. 2009a for a recent review).
Changes in spawning location or alteration of currents could affect recruitment by pre-venting successful larval migration back to their recruitment areas (Kimura et al. 2001; Kimuraand Tsukamoto, 2006; Friedland et al. 2007; Kim et al. 2007; Tsukamoto 2009; Zenimoto et al.2009), but these larval transport factors are not directly related to the biology of leptocephali.Changes in productivity in the surface layer, however, could affect larval survival by reducingfeeding success of leptocephali (Knights 2003; Friedland et al., 2007; Bonhommeau et al.2008a,b). In particular, several recent studies have found correlations between recruitment andproductivity-related factors. Correlations have been found between the North Atlantic Oscilla-tion index and recruitment of A. anguilla to the Netherlands (Knights 2003; Friedland et al.2007) or other parts of Europe (Kettle et al. 2008). The North Atlantic Oscillation may be re-lated to a variety of changes in the North Atlantic, so an effect on productivity or communitystructure that could impact the feeding success of leptocephali may be the mechanism behindthese correlations. Other studies have found direct correlations between sea surface temperatureor productivity fluctuations and recruitment in both of the northern hemisphere subtropical gyres(Bonhommeau et al. 2008a,b). Increasing ocean temperatures tend to reduce productivity, so seasurface temperature increases in the global ocean (e.g. Levitus et al. 2000) have the potential toreduce productivity, which could affect the feeding success of leptocephali. Not enough is knownabout the feeding ecology of leptocephali to know at which stage these effects could occur. Itmay be during the critical first feeding period after yolk absorption when the young larvae maybe most vulnerable to starvation. These factors highlight the need for a better understanding ofleptocephalus ecology in the ocean, not only to help conserve the economically importantanguillids and conger eels, but also marine eels whose role in marine ecosystems is still un-known.
5-6. Ecological significance of leptocephali in the surface layer
Knowledge about leptocephali is perhaps a unique story in the history of marine biology,since there have been extensive surveys for fishes, fish larvae and zooplankton in most regionsof the world’s oceans, but the sampling gear in the vast majority of cases has been unable tocollect many leptocephali. The primary problem is that leptocephali are able to avoid the stand-ard plankton nets that are adequate to collect representative samples of most fish larvae andzooplankton, such as the 1-m plankton net, twin bongo nets, or the 1-m MOCNESS (seeWiebe and Benefield 2003 for a review of plankton net technology). The visual system of lepto-cephali during the day or their mechanosensory system at night in combination with the poten-tial for bioluminescent organisms to make nets visible at night (Ross et al. 2007), probably giveleptocephali the chance to avoid plankton nets in most circumstances, in the same ways that fishlarvae in general are adapted to avoid predators (Fuiman and Magurran 1994). Leptocephali,however, have the added advantage of being able to evade the net by either swimming forwardsor backwards, or being able to quickly change swimming directions due to their flexible body.Their larger size is also likely a major factor in their net avoidance abilities. In addition, many ofthe larger nets deployed to catch pelagic marine fishes had mesh sizes that were too large, or thefishing effort was too deep. These factors seemingly have resulted in underestimates of theabundance of leptocephali in tropical and subtropical regions.
This underestimation would not be particularly important if leptocephali werezooplanktivores, like most other fish larvae. However, the existence of large numbers of fishlarvae that feed on particulate matter and reach large sizes, may have greater implications forunderstanding the carbon cycle of the ocean than simply an underestimation of grazing onzooplankton by fish larvae. There have been extensive efforts to understand the dynamic fluxesof particulate material (technically referred to as particulate organic carbon: POC) in the surfacelayer of the ocean, since this is a critical factor in understanding the carbon cycle in the ocean(e.g. Hebel and Karl 2001; Pilskaln et al. 2005; Honjo et al. 2008; Lee et al. 2009). Many studieshave used sediment traps to determine how much POC sinks out of the surface layer where

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 79
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
primary production occurs and POC is produced by various biological processes (Buesseler etal. 2007; Honjo et al. 2008). One recurring theme in studies trying to model the dynamics ofPOC is that the estimated amount of POC produced but not consumed by animals in the surfacelayer typically is much greater than what is actually collected in sediment traps. Siegel et al.(2008) summarized the general results of experiments to assess the production of sinking parti-cles and their vertical flux during major biogeochemical research programs by stating: “Theseexperiments nearly always resulted in a poor correspondence between determinations of pri-mary production rates measured in the euphotic zone and the sinking flux at depth. This raisesthe question why are the observations between primary production and export flux often poorlyrelated?”
Various hypotheses have been proposed to account for this mismatch, such as materialescaping from the traps due to hydrodynamic factors and POC being broken up by the swim-ming activities of zooplankton before reaching the traps or even inside the traps (Buesseler et al.2007; Honjo et al. 2008). However, none of these studies considered the fact that there arehundreds of species leptocephali found everywhere in the tropical and subtropical oceans, thatreach large sizes and appear to be to feeding on POC year-round. A recent modeling studynarrowed down the likely requirement to make models of particle flux match the observed data.Stemmann et al. (2004) stated that to stop the high flux of large particles to deeper layers asappears to be happening, “… requires the mesozooplankton to feed preferentially on large set-tling particles using remote detection, totally ingesting the particles.” Although they pointed outthat this does not match the capabilities of the zooplankton that were considered to be the graz-ers of particulate matter, this description corresponds well with the likely feeding behavior ofleptocephali.
It seems possible that due to the indications that leptocephali are widely distributed andoften quite abundant in many regions, that they may have at least some level of impact on thecycling of POC in the ocean surface layer that needs to be considered in models of the oceancarbon cycle. The high abundances of leptocephali along continental shelves (Miller et al. 2002a;Miller and McCleave 2007), in anguillid or mesopelagic eel spawning areas (Castonguay andMcCleave 1987b; Kleckner and McCleave 1988; Tsukamoto 1992; Miller and McCleave 1994;Wippelhauser et al. 1996; Miller et al. 2006a), in areas such as convergence zones associatedwith fronts (Miller and McCleave 1994), of species such as Ariosoma in some regions (Miller2002b; Miller et al. 2006b; Miller and McCleave 2007), and the relative abundance of lepto-cephali among the fish larvae recruiting to shallow tropical habitats (e.g. Shenker et al. 1993;McIlwain 2003), argue for the likelihood that leptocephali may be important compo-nents of many marine ecosystems worldwide. This is especially true if their abundances areunderestimated by the large trawls presently used to collect them due to some degree of netavoidance. Further data on the abundance of anguilliform leptocephali will be obtained in ongo-ing research on samples from the northwest Coral Sea made with a 70 m2 mouth opening midwatertrawl (see Dennis et al. 2001), which indicate that large numbers of leptocephali were present inthat region within the upper 200 m during both day and night (MJ Miller and JM Leis unpubl.data).
The various types of information that are presently available about leptocephali, suggestthat more knowledge about all aspects of these transparent larvae is needed to obtain a betterunderstanding of their role in the dynamics of the ocean. Leptocephali clearly transport carbonand other elements from the ocean surface layer to many different ecosystems, which seeminglyhas not been discussed previously in the literature, except perhaps for anguillids, which enterfreshwater. For example, the larvae of the Cyematidae, Eurypharyngidae, Derichthyidae,Nemichthyidae, and Serrivomeridae recruit to the mesopelagic layer and deeper waters. Larvaeof the Synaphobranchidae recruit to deep benthic habitats where their juveniles and adults areimportant components of the deep-sea fauna. Nettastomatid and a variety of species of larvae ofother families recruit to continental slope habitats. Carbon is also transported in the form ofmetamorphosing leptocephali or glass eels to the shallow water regions of continental shelves oraround islands worldwide. Presently, these components of the global ocean carbon cycle and therole of leptocephali and their recruiting larvae in marine communities in general are completely

80 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
unquantified, and seemingly unrecognized for the most part. However, Honjo et al. (2008) pointedout the need to gain a better understanding of the biological influences on carbon transport fromorganisms, such as vertically migrating zooplankton, so research efforts are also needed to evaluatethe ecological importance of eels and leptocephali in the many marine ecosystems in which theylive worldwide.
5-7. Future research perspectives
This review points out some of the many questions that remain to be answered about theseremarkable fish larvae. Leptocephali are present in the surface layer of the ocean all over theworld, except at very high latitudes, but they have been rarely seen or caught compared to otherfish larvae or zooplankton, so they represent a new research frontier in helping to increase un-derstanding of marine ecosystems and regional biodiversity (Miller and Tsukamoto 2006). Adulteels whose larvae can be collected using large trawls fished at night are themselves even harderto study due to their cryptic behaviors, so studies on leptocephali can provide useful informationabout the biodiversity of eels and the potential variations in their life histories.
Studies on the biology and behavior of leptocephali are quite limited, but may be able toincrease using recently developed techniques to spawn and rear leptocephali in the laboratory(Tanaka et al. 2001, 2003). This could lead to more experimental studies related to their ecology(Tsukamoto et al. 2009; Yamada et al. 2009) and physiology (Ozaki et al. 2000, 2006b). Lepto-cephali can be collected and held in aquaria as they recruit to coastal areas, but they typicallymetamorphose rapidly, thus providing a limited period of time for research compared to artifi-cially reared leptocephali.
Much of the biochemical and physiological research on leptocephali has been done onbonefishes (Pfeiler 1999), so more studies on a wider range of anguilliform species are needed.These will build on previous research (Hulet and Robins 1989; Donnelly et al. 1995; Pfeiler andGovoni 1993; Bishop and Torres 1999, 2001; Bishop et al. 2000), which has found that lepto-cephali are remarkably different physiologically than other fish larvae. These types of studiesalong with a greater understanding of the distributional ecology of leptocephali in relation tohydrographic features and ocean currents will be essential for knowing how eels and their larvaefit into the ecology of the oceans. With changing ocean-atmosphere conditions causing a warm-ing of the worlds oceans (Levitus et al. 2000), there is a need to understand all components ofmarine ecosystems, especially those, such as leptocephali, that appear to be abundant, but whoseecological role in the ocean is poorly understood. Future research will likely reveal many detailsabout the mysterious biology of leptocephali that have remained hidden in the ocean, wherethese remarkable glass-like larvae have always been swimming just outside of our view.
AcknowledgementsThe efforts of many people made this review possible, and I especially thank Prof. Katsumi Tsukamoto
for providing the chance to examine and photograph freshly caught leptocephali in the western Pacific,Indian Ocean, and Indonesian Seas, and for many years of productive research collaboration on lepto-cephali. I also thank Prof. James D. McCleave for starting my interest in leptocephali and for collabora-tion on leptocephali from his Sargasso Sea collections, Jim Craddock and Karsten Hartel for providingaccess to the WHOI leptocephali collections, and Larry Madin for providing the chance to collect lepto-cephali with the MOCNESS-10 and to film the live chlopsid leptocephalus. Tae Won Lee, Mari Kuroki,and Tao Ma conducted much of the otolith research overviewed here, and the pioneering leptocepha-lus research of Tsuguo Otake and Noritaka Mochioka are also acknowledged. Jun Aoyama, SatoshiIshikawa, Shun Watanabe, Takaomi Arai, Akira Shinoda, Yobuo Kimura, Gen Minagawa, Yuki Minegishi,Tatsuki Yoshinaga, Yuzuru Suzuki, Atsushi Fukui, Machiko Oya and many others also have assistedwith research efforts on leptocephali over the years and have sorted through many plankton samples toprovide the chance to study these remarkable transparent larvae. Sam Wouthuyzen, Hagi Yulia Sugeha,and Ono Kurnaen Sumadhiharga of LIPI, Indonesia helped make the collaborative sampling surveys inthe Indonesian Seas a success. The captains and crew of the research vessels Hakuho Maru, Tansei Maru,and Baruna Jaya VII also helped our sampling efforts during many cruises. The scientists at the IRAGOInstitute have conducted a new generation of ecologically related research on artificially cultured lepto-cephali, and I especially thank Yoshiaki Yamada for providing photographs showing the transparency of

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 81
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
leptocephali. Jerry Powel kindly provided the photographs of the large greenish leptocephalus, and Mat-thew D’Avella provided the interesting video of the ophichthid leptocephalus. I also thank David G. Smithand the late Peter Castle who were some of the pioneers of leptocephalus identification, and their lastingcontributions to this field will continue to be useful as research on leptocephali progresses around theworld in the future. David G. Smith and an anonymous reviewer provided suggestions that significantlyimproved the paper.
ReferencesAble KW, Fahay MP. The first year in the life of estuarine fishes in the Middle Atlantic Bight. Rutgers
University Press, New Brunswick. 1998.Alldredge AL. Abandoned larvacean houses, a unique food source in the pelagic environment. Science
1972; 177: 885–887.Alldredge AL. Discarded appendicularian houses as sources of food, surface habitats, and particulate
organic matter in planktonic environments. Limnol. Oceanogr. 1976; 21: 14–23.Alldredge AL, Silver MW. Characteristics, dynamics and significance of marine snow. Progr. Oceanogr.
1988; 20: 41–82.Allen GR, Adrim M. Coral reef fishes of Indonesia. Zool. Stud. 2003; 42: 1–72.Amir OA, Berggren P, Ndaro SGM, Jiddawi NS. Feeding ecology of the Indo–Pacific bottlenose dolphin
(Tursiops aduncus) incidentally caught in the gillnet fisheries off Zanzibar, Tanzania. Est. Coast.Shelf Sci. 2005; 63: 429–437.
Antunes C, Tesch F-W. A critical consideration of the metamorphosis zone when identifying daily rings inotoliths of European eel, Anguilla anguilla (L.). Ecol. Freshw. Fish 1997; 6: 102–107.
Aoyama J. Life history and evolution of migration in catadromous eels (genus Anguilla). Aqua-BioSci.Monogr. (ABSM) 2009; 2: 1–42.
Aoyama J, Mochioka N, Otake T, Ishikawa S, Kawakami Y, Castle PHJ, Nishida M, Tsukamoto K. Distri-bution and dispersal of anguillid leptocephali in the western Pacific Ocean revealed by molecularanalysis. Mar. Ecol. Progr. Ser. 1999; 188: 193–200.
Aoyama J, Wouthuyzen S, Miller MJ, Inagaki T, Tsukamoto K. Short-distance spawning migration oftropical freshwater eels. Biol. Bull. 2003; 204: 104–108.
Aoyama J, Wouthuyzen S, Miller MJ, Minegishi Y, Minagawa G, Kuroki M, Suharti SR, Kawakami T,Sumardiharga KO, Tsukamoto K. Distribution of leptocephali of the freshwater eels, genus Anguilla,in the waters off west Sumatra in the Indian Ocean. Environ. Biol. Fish. 2007; 80: 445–452.
Appelbaum S, Riehl R. Scanning electron microscopic observations on the head morphology of sevendifferent leptocephali belonging to six eel families (Anguilliformes). Helgoländer Meeresunters1993; 47: 113–124.
Arai T, Otake T, Tsukamoto K. Drastic changes in otolith microstructure and microchemistry accompany-ing the onset of metamorphosis in the Japanese eel Anguilla japonica. Mar. Ecol. Progr. Ser. 1997;161: 17–22.
Arai T, Limbong D, Tsukamoto K. Validation of otolith daily increments in the tropical eel Anguillacelebesensis. Can. J. Zool. 2000; 78: 1078–1084.
Arai T, Limbong D, Otake T, Tsukamoto K. Recruitment mechanisms of tropical eels Anguilla spp. andimplications for the evolution of oceanic migration in the genus Anguilla. Mar. Ecol. Progr. Ser.2001; 216: 253–264.
Atema J, Kingsford MJ, Gerlach G. Larval reef fish could use odour for detection, retention and orienta-tion to reefs. Mar. Ecol. Progr. Ser. 2002; 241: 151–160.
Backus RH, Craddock JE. Mesopelagic fishes in Gulf Stream cold-core rings. J. Mar. Res. 1982; 40: 1–19.Bast HD, Strehlow B. Length composition and abundance of eel larvae, Anguilla anguilla (Anguilliformes:
Anguillidae), in the Iberian Basin (northeastern Atlantic) during July–September 1984. HelgoländerMeeresunters 1990; 44: 353–361.
Bauchot ML. Étude des larves leptocéphales du groupe Leptocephalus lanceolatus Strömman et identifi-cation à la famille des Serrivomeridae. Dana Rep. 1959; 48: 1–148 (in French).
Beebe W, Crane J. Deep-sea fishes of the Bermuda Oceanographic Expeditions. Family Serrivomeridae,Part I: Genus Serrivomer. Zoologica 1936; 20: 53–102.
Bell GW, Witting DA, Able KW. Aspects of metamorphosis and habitat use in the conger eel, Congeroceanicus. Copeia 2003; 2003: 544–552.
Bertin L. Formes nouvelles et formes larvaires de poisons apodes appartenant au sous- ordere des Lyomeres.Dana Rep. 1938; No. 15: 26 pp. (in French).
Bishop RE, Torres JJ. Leptocephalus energetics: metabolism and excretion. J. Exper. Biol. 1999; 202:2485–2493.

82 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Bishop RE, Torres JJ. Leptocephalus energetics: assembly of the energetics equation. Mar. Biol. 2001;138: 1093–1098.
Bishop RE, Torres JJ, Crabtree RE. Chemical composition and growth indices in leptocephalus larvae.Mar. Biol. 2000; 137: 205–214.
Blache J. Leptocephales des poissons Anguilliformes dans la zone sud du Golfe de Guinée. Faune Tropicale1977; 10: 1–381 (in French).
Boëtius J, Harding EF. A re-examination of Johannes Schmidt’s Atlantic eel investigations. Dana 1985; 4:129–162.
Böhlke EB (ed). Fishes of the Western North Atlantic. Orders Anguilliformes and Saccopharyngiformes.Part 9, Vol. 1, pp. 1–655. Sears Foundation for Marine Research, New Haven. 1989a.
Böhlke EB (ed). Fishes of the Western North Atlantic. Leptocephali. Part 9, Vol. 2, pp. 657–1055. SearsFoundation for Marine Research, New Haven. 1989b.
Bonhommeau S, Chassot E, Rivot E. Fluctuations in European eel (Anguilla anguilla) recruitment result-ing from environmental changes in the Sargasso Sea. Fish. Oceanogr. 2008a; 17: 32–44.
Bonhommeau S, Chassot E, Planque B, Rivot E, Knap AH, Le Pape O. Impact of climate on eel populationsof the Northern Hemisphere. Mar. Ecol. Progr. Ser. 2008b; 373: 71–80.
Bonhommeau S, Le Pape O, Gascuel D, Blanke B, Tréguier A-M, Grima N, Vermard Y, Castonguay M,Rivot E. Estimates of the mortality and the duration of the trans-Atlantic migration of European eelAnguilla anguilla leptocephali using a particle tracking model. J. Fish Biol. 2009a; 74: 1891–1914.
Bonhommeau S, Blanke B, Tréguier A-M, Grima N, Rivot E, Vermard Y, Greiner E, Le Pape O. How fastcan the European eel (Anguilla anguilla) larvae cross the Atlantic Ocean? Fish. Oceanogr. 2009b;18: 371–385.
Brischoux F, Bonnet X, Shine R. Determinants of dietary specialization: a comparison of two sympatricspecies of sea snakes. Oikos 2009; 118: 145–151.
Britt LL, Loew ER, McFarland WN. Visual pigments in the early life stages of Pacific northwest marinefishes. J. Exp. Biol. 2001; 204: 2581–2587.
Browman HI, Skiftersvik AB, Kuhn P. The relationship between ultraviolet and polarized light and growthrate in the early larval stages of turbot (Scophtalmus maximus), Atlantic cod (Gadus morhua) andAtlantic herring (Clupea harengus) reared in intensive culture conditions. Aquaculture 2006; 256:296–301.
Buesseler KO, Antia AN, Chen M, Fowler SW, GardnerWD, Gustaffson Ö, Harada K, Michaels AF,Rutgers van der Loeff M, Sarin M, Steinberg DK, Trull T. An assessment of the use of sedimenttraps for estimating upper ocean particle fluxes. J. Mar. Res. 2007; 65: 345–416.
Campana SE. Chemistry and composition of fish otoliths: pathways, mechanisms and applications. Mar.Ecol. Progr. Ser. 1999; 188: 263–297.
Campana SE, Neilson JD. Microstructure of fish otoliths. Can. J. Fish. Aquat. Sci. 1985; 42: 1014–1032.Capitanio FL, Pájaro M, Esnal GB. Appendicularians: an important food supply for the Argentine an-
chovy Engraulis anchoita in coastal waters. J. Appl. Ichthyol. 2005; 21: 414–419.Carpenter KE, Springer VG. The center of the center of shore fish biodiversity: the Philippine Islands.
Environ. Biol. Fish 2005; 72: 467–480.Castle PHJ. Anguillid leptocephali in the southwest Pacific. Zool. Publ. Victoria Univ. Wellington 1963;
33: 1–14.Castle PHJ. Congrid leptocephali in Australasian waters with descriptions of Conger wilsoni (Bl. and
Schn.) and C. verreauxi Kaup. Zool. Publ. Victoria Univ. Wellington 1964; 37: 1–45.Castle PHJ. Muraenid leptocephali in Australasian waters. Trans. R. Soc. N.Z. Zool. 1965a; 7: 57–84.Castle PHJ. Ophichthid leptocephali in Australasian waters. Trans. R. Soc. N.Z. Zool. 1965b; 7: 97–123.Castle PHJ. Leptocephali of the Nemichthyidae, Serrivomeridae, Synaphobranchidae and Nettastomatidae
in Australasian waters. Trans. R. Soc. N.Z. Zool. 1965c; 5: 131–146.Castle PHJ. Larval development of the congrid eel Gnathophis capensis (Kaup), off southern Africa, with
notes on the identity of Congermuraena australis Barnard. Zoologica Africana 1968; 3: 139–154.Castle PHJ. Species structure and seasonal distribution of leptocephali in the eastern Indian Ocean (110°E).
Cah. ORSTOM sér. Oceanogr. 1969; 7: 53–88.Castle PHJ. Distribution, larval growth, and metamorphosis of the eel Derichthys serpentinus Gill, 1884
(Pisces, Derichthyidae). Copeia 1970; 1970: 444–452.Castle PHJ. Early life-history of the eel Moringua edwardsi (Pisces, Moringuidae) in the western North
Atlantic. Bull. Mar. Sci. 1979; 29: 1–18.Castle PHJ. Notacanthiformes and Anguilliformes: Development, In: Moser HG, Richards WJ (eds). On-
togeny and Systematics of Fishes. Amer. Soc. Ichthyol. Herpetol. Spec. Publ. 1. pp. 62–93. Ameri-can Society of Ichthyologists and Hepetologists, New York. 1984.

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 83
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Castle PHJ. Garden eel leptocephali: characters, generic identification, distribution, and relationships.Bull. Mar. Sci. 1997; 60: 6–22.
Castle PHJ, Robertson DA. Early life history of the congrid eels Gnathophis habenatus and G. incognitusin New Zealand waters. N.Z. J. Mar. Freshw. Res. 1974; 8: 95–110.
Castle PHJ, Raju NS. Some rare leptocephali from the Atlantic and Indo–Pacific Oceans. Dana Rep.1975; 85: 1–25.
Castle PHJ, Smith DG. A reassessment of the eels of the genus Bathycongrus in the Indo–West Pacific.J. Fish Biol. 1999; 54: 973–995.
Castonguay M. Growth of American and European eel leptocephali as revealed by otolith microstructure.Can. J. Zool. 1987; 65: 875–878.
Castonguay M, McCleave JD. Vertical distributions, diel and ontogenetic vertical migrations and net avoid-ance of leptocephali of Anguilla and other common species in the Sargasso Sea. J. Plankt. Res.1987a; 9: 195–214.
Castonguay L, McCleave JD. Distribution of leptocephali of the oceanic species Derichthys serpentinusand Nessorhamphus ingolfianus (Family Derichthyidae) in the western Sargasso Sea in relation tophysical oceanography. Bull. Mar. Sci. 1987b; 41: 807–821.
Castonguay M, Hodson PV, Moriarty C, Drinkwater KF, Jessop BM. Is there a role of ocean environmentin American and European eel decline? Fish. Oceanogr. 1994; 3: 197–203.
Chen HL, Shen KN, Chang CW, Iizuka Y, Tzeng WN. Effects of water temperature, salinity and feedingregimes on metamorphosis, growth and otolith Sr:Ca ratios of Megalops cyprinoides leptocephali.Aquat. Biol. 2008; 3: 41–50.
Chen PW, Tzeng WN. Timing of metamorphosis and estuarine arrival across the dispersal range of theJapanese eel Anguilla japonica. Mar. Ecol. Progr. Ser. 1996; 131: 87–96.
Cieri MD, McCleave JD. Validation of daily otolith increments in glass-phase American eels Anguillarostrata (Lesueur) during estuarine residency. J. Exp. Mar. Biol. Ecol. 2001; 257: 219–227.
Cohen DM, Dean D. Sexual maturity and migratory behaviour of the tropical eel, Ahlia egmontis. Nature1970; 227: 189–190.
Colborn J, Crabtree RE, Shaklee JB, Pfeiler E, Bowen BW. The evolutionary enigma of bonefishes (Albulaspp.): cryptic species and ancient separations in a globally distributed shorefish. Evolution 2001;55: 807-820.
Correia AT, Antunes C, Coimbra J. Aspects of the early life history of the European conger eel (Congerconger) inferred from the otolith microstructure of metamorphic larvae. Mar. Biol. 2002a; 140:165–173.
Correia, AT, Isidro EJ, Antunes C, Coimbra J. Age, growth, distribution and ecological aspects of Congerconger leptocephali collected in the Azores, based on otolith analysis of premetamorphic speci-mens. Mar. Biol. 2002b; 141: 1,141–1,151.
Correia AT, Antunes C, Isidro EJ, Coimbra J. Changes in otolith microstructure and microchemistry dur-ing larval development of the European conger eel (Conger conger). Mar. Biol. 2003; 142: 777–789.
Correia AT, Correia AT, Able KW, Antunes C, Coimbra J. Early life history of the American conger eel(Conger oceanicus) as revealed by otolith microstructure and microchemistry of metamorphosingleptocephali. Mar. Biol. 2004; 145: 477–488.
Correia AT, Antunes C, Wilson JM, Coimbra J. An evaluation of the otolith characteristics of Congerconger during metamorphosis. J. Fish Biol. 2006; 68: 99–119.
Crabtree RE, Cyr EC, Bishop RE, Falkenstein LM, Dean JM. Age and growth of tarpon, Megalopsatlanticus, larvae in the eastern Gulf of Mexico, with notes on relative abundance and probablespawning areas. Environ. Biol. Fish. 1992; 35: 361–370.
D’Août P, Aerts K. A kinematic comparison of forward and backward swimming in the eel Anguillaanguilla. J. Exper. Biol. 1999; 202: 1511–1521.
De Castro MC, Bonecker ACT. Leptocephali collected off the eastern coast of Brazil (12°–23°S). Zootaxa2005; 935: 1–28.
Dennis DM, Pitcher CR, Skewes TD. Distribution and transport pathways of Panulirus ornatus (Fabricius,1776) and Panulirus spp. larvae in the Coral Sea, Australia. Mar. Freshw. Res. 2001; 52: 1175–85.
Donnelly J, Torres JJ, Crabtree RE. Proximate composition and nucleic acid content of premetamorphicleptocephalus larvae of the congrid eel Ariosoma balearicum. Mar. Biol. 1995; 123: 851–858.
Dufour V, Riclet E, Lo-Yat A. Colonization of reef fishes at Moorea Island, French Polynesia: temporaland spatial variation of the larval flux. Mar. Freshw. Res. 1996; 47: 413–422.
Evans BI, Fernald RD. Metamorphosis and fish vision. J. Neurobiol. 1990; 21: 1037–1052.Fahay MP. Anguilliformes: leptocephalus larvae. Pages 46–61 in Guide to the early stages of marine

84 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
fishes occurring in the western North Atlantic Ocean, Cape Hatteras to the Southern Scotian Shelf.J. Northwest Atl. Fishery Sci. 1983; 4: 3–423.
Fahay MP, Obenchain CL Leptocephali of the ophichthid genera Ahlia, Myrophis, Ophichthus,Pisodonophis, Callechelys, Letharchus, and Apterichtus on the Atlantic continental shelf of theUnited States. Bull. Mar. Sci. 1978; 28: 442–486.
Ferraris CJ. Redescription and spawning behavior of the muraenid eel Gymnothorax herrei. Copeia 1985;1985: 518–520.
Figueroa DE. A curious record of leptocephali from Ilha Do Farol, Atol das Rocas, Brazil. Bull. Mar. Sci.1997; 61: 935–936.
Figueroa DE, Ehrlich M. Systematics and distribution of leptocephali in the western South Atlantic. Bull.Mar. Sci. 2006; 78: 227–242.
Fisher R. Swimming speeds of larval coral reef fishes: impacts on self-recruitment and dispersal. Mar.Ecol. Progr. Ser. 2005; 285: 223–232.
Fisher R, Leis JM, Clark DL, Wilson SK. Critical swimming speeds of late-stage coral reef fish larvae:variation within species, among species and between locations. Mar. Biol. 2005; 147: 1201–1212.
Flood PR, Deibel D. The appendicularian house. In: Bone Q (ed). The Biology of Pelagic Tunicates.Oxford University Press, Oxford. 1998; pp. 105–124.
Friedland KD, Miller MJ, Knights B. Oceanic changes in the Sargasso Sea and declines in recruitment ofthe European eel. ICES J. Mar. Sci. 2007; 64: 519–530.
Fuiman LA, Batty RS. What a drag it is getting cold: Partitioning the physical and physiological effects oftemperature on fish swimming. J. Exp. Biol. 1997; 200: 1745–1755.
Fuiman LA, Magurran AE. Development of predator defences in fishes. Rev. Fish Biol. Fisheries 1994; 4:145–183.
Fuiman LA, Higgs DM, Poling KR. Changing structure and function of the ear and lateral line system offishes during development. In: Govoni JJ (ed). The Development of Form and Function in Fishesand the Question of Larval Adaptation. Am. Fish. Soc. Symp. 40. Bethesda, Maryland. 2004; 117–144.
Fujii R. The regulation of motile activity in fish chromatophores. Pigm. Cell. Res. 2000; 13: 300–319.Fukuda N, Kuroki M, Shinoda A, Yamada Y, Okamura A, Aoyama J, Tsukamoto K. Influence of water
temperature and feeding regime on otolith growth in Anguilla japonica glass eels and elvers: Doesotolith growth cease at low temperatures? J. Fish Biol. 2009; 74: 1915–1933.
Galt CP, Grober MS, Sykes PF. Taxonomic correlates of bioluminescence among appendicularians(Urochordata: Larvacea). Biol. Bull. 1985; 168: 125–134.
Gilbert M, Rasmusssen JB, Kramer DL. Estimating the density and biomass of moray eels (Muraenidae)using a modified visual census method for hole-dwelling reef fauna. Environ. Biol. Fishes 2005; 73:415–426.
Govoni J. Feeding on protists and particulates by the leptocephali of the worm eels Myrophis spp. (Teleostei,Anguilliformes, Ophichthidae), and the potential energy contribution of large aloricate protozoa.Scientia Marina (in press).
Govoni J, Boehlert GW, Watanabe Y. The physiology of digestion in fish larvae. Environ. Biol. Fishes1986; 16: 59–77.
Gray J. Studies in animal locomotion. I. The movement of fish with special reference to the eel. J. Exp.Biol. 1933; 10: 88–103.
Grether GF, Kolluru GR, Nersissian K. Individual colour patches as multicomponent signals. Biol. Rev.2004; 79: 583–610.
Haedrich RL, Merrett NR. Summary atlas of deep-living demersal fishes in the North Atlantic Basin. J.Natural Hist. 1988; 22: 1325–1362.
Hebel DV, Karl DM. Seasonal, interannual and decadal variations in particulate matter concentrations andcomposition in the subtropical North Pacific Ocean. Deep-Sea Res. II 2001; 48: 1669–1695.
Herren CM, Alldredge AL, Case JF. Coastal bioluminescent marine snow: fine structure of biolumines-cence distribution. Cont. Shelf. Res. 2004; 24: 413–429.
Hess M, Melzer RR, Smola U. The photoreceptors of Muraena helena and Ariosoma balearicum—acomparison of multiple bank retinae in anguilliform eels (Teleostei). Zool. Anz. 1998; 237: 127–137.
Honjo S, Manganini SJ, Krishfield RA, Francois R. Particulate organic carbon fluxes to the ocean interiorand factors controlling the biological pump: A synthesis of global sediment trap programs since1983. Progr. Oceanogr. 2008; 76: 217–285.
Horie N, Utoh T, Yamada Y, Okamura A, Zhang H, Mikawa N, Akazawa A, Tanaka S, Oka H. Develop-ment of embryos and larvae in the common Japanese conger Conger myriaster. Fish. Sci. 2002; 68:972–983.

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 85
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Hulet WC. Structure and functional development of the eel leptocephalus Ariosoma balearicum (De LaRoche, 1809). Phil. Trans. Roy. Soc. London, Biol. Sci. 1978; 82: 107–138.
Hulet WH, Robins CR. The evolutionary significance of the leptocephalus larva. In: Böhlke EB (ed).Fishes of the Western North Atlantic. Leptocephali. Part 9, Vol. 2. Sears Foundation for MarineResearch, New Haven. 1989; pp. 669–677.
Hulet WH, Fischer J, Rietberg BJ. Electrolyte composition of anguilliform leptocephali from the Straits ofFlorida. Bull. Mar. Sci. 1972; 22: 432–448.
Ichikawa T. Particulate organic carbon and nitrogen in the adjacent seas of the Pacific Ocean. Mar. Biol.1982; 68: 49–60.
Ichikawa T, Nishizawa S. Particulate organic carbon and nitrogen in the eastern Pacific Ocean. Mar. Biol.1975; 29: 129–138.
Ineich I, Bonnet X, Brischoux F, Kulbicki M, Séret B, Shine R. Anguilliform fishes and sea kraits: ne-glected predators in coral-reef ecosystems. Mar. Biol. 2007; 151: 793–802.
Inoue JG, Miya M, Tsukamoto K, Nishida M. Mitogenomic evidence for the monophyly of elopomorphfishes (Teleostei) and the evolutionary origin of the leptocephalus larva. Mol. Phyl. Evol. 2004; 32:274–286.
Inoue JG, Miya M, Venkatesh B, Nishida M. The mitochondrial genome of Indonesian coelacanth Latimeriamenadoensis (Sarcopterygii: Coelacanthiformes) and divergence time estimation between the twocoelacanths. Gene 2005; 349: 227–235.
Ishii S, Yoshiura Y, Kajimura S, Mochioka N, Aida K. Gonadal development and expression profiles ofgonadotropin genes in wild sea conger, Ariosoma meeki. Fish Physiol. Biochem. 2003; 28: 95–96.
Ishikawa S, Suzuki K, Inagaki T, Watanabe S, Kimura Y, Okamura A, Otake T, Mochioka N, Suzuki Y,Hasumoto H, Oya M, Miller MJ, Lee TW, Fricke H, Tsukamoto K. Spawning time and place of theJapanese eel, Anguilla japonica, in the North Equatorial Current of the western North Pacific Ocean.Fish. Sci. 2001; 67: 1097–1103.
Jellyman DJ. The distribution and biology of the South Pacific species of Anguilla. In: Aida K, TsukamotoK, Yamauchi K (eds). Eel Biology. Springer, Tokyo. 2003; pp. 275–292.
Jespersen P. Indopacific leptocephalids of the genus Anguilla: Systematic and biological studies. DanaRep. 1942; 22: 1–128.
Johnsen S. Hidden in plain sight: the ecology and physiology of organismal transparency. Biol. Bull.2001; 201: 301–318.
Johnsen S. Lifting the cloak of invisibility: the effects of changing optical conditions on pelagic crypsis.Integr. Comp. Biol. 2003; 43: 580–590.
Johnsen S, Widder EA. Ultraviolet absorption in transparent zooplankton and its implications for depthdistribution and visual predation. Mar. Biol. 2001; 138: 717–730.
Johnson TB, Miller MJ, Tsukamoto K, Terazaki M. First record of a leptocephalus larva ingested by achaetognath. Coastal Mar. Sci. 2006; 30: 469–472.
Kajihara T, Tsukamoto K, Otake T, Mochioka N, Hasumoto H, Oya M, Tabeta O. Sampling leptocephaliwith reference to the diel vertical migration and the gears. Nippon Suisan Gakkaishi 1988; 54: 941–946.
Kaneko T, Hasegawa S, Sasai S. Chloride cells in the Japanese eel during their early life stages anddownstream migration. In: Aida K, Tsukamoto K, Yamauchi K (eds). Eel Biology. Springer, Tokyo.2003; pp. 457–468.
Kaneko T, Watanabe S, Lee KM. Functional morphology of mitochondrion-rich cells in euryhaline andstenohaline teleosts. Aqua-BioSci. Monogr. (ABSM) 2008; 1(1): 1–62.
Karmovskaya ES. Leptocephali of eels of the genus Nemichthys (Nemichthyidae, Osteichthyes). Voprovosyikhtiolgii 1990; 30: 551–563 (Translated from Russian by Scripta Technica, Inc.).
Kettle AJ, Haines K. How does the European eel (Anguilla anguilla) retain its population structure duringits larval migration across the North Atlantic Ocean? Can. J. Fish. Aquat. Sci. 2006; 63: 90–106.
Kettle AJ, Bakker DCE, Haines K. Impact of the North Atlantic Oscillation on the trans-Atlantic migra-tions of the European eel (Anguilla anguilla). J. Geophys. Res. 2008; 113: G03004, doi:10.1029/2007JG000589.
Kim H, Kimura S, Shinoda A, Kitagawa T, Sasai Y, Sasaki H. Effect of El Niño on migration and larvaltransport of the Japanese eel, Anguilla japonica. ICES J. Mar. Sci. 2007; 64: 1387–1395.
Kimura S, Tsukamoto K. The salinity front in the North Equatorial Current: A landmark for the spawningmigration of the Japanese eel (Anguilla japonica) related to the stock recruitment. Deep-Sea Res. II2006; 53: 315–325.
Kimura S, Tsukamoto K, Sugimoto T. A model for the larval migration of the Japanese eel: roles of thetrade winds and salinity front. Mar. Biol. 1994; 119: 185–190.

86 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Kimura S, Inoue T, Sugimoto T. Fluctuation in the distribution of low-salinity water in the North Equato-rial Current and its effect on the larval transport of the Japanese eel. Fish. Oceanogr. 2001; 10: 51–60.
Kimura Y, Ishikawa S, Tokai T, Nishida M, Tsukamoto K. Early life history characteristics and genetichomogeneity of Conger myriaster leptocephali along the east coast of central Japan. Fisheries Res.2004; 70: 61–69.
Kimura Y, Miller MJ, Minagawa G, Watanabe S, Shinoda A, Aoyama J, Tsukamoto K. Evidence of a localspawning site of marine eels along northeastern Japan, based on the distribution of small lepto-cephali. Fish. Oceanogr. 2006; 15: 183–190.
King NJ, Bagley PM, Priede IG. Depth zonation and latitudinal distribution of deep-sea scavenging demersalfishes of the Mid-Atlantic Ridge, 42 to 53°N. Mar. Ecol. Progr. Ser. 2006; 319: 263–274.
Kleckner RC, McCleave JD. Entry of migrating American eel leptocephali into the Gulf Stream system.Helgoländer Meeresunters 1982; 35: 329–339.
Kleckner RC, McCleave JD. Spatial and temporal distribution of American eel larvae in relation to NorthAtlantic Ocean current systems. Dana 1985; 4: 67–92.
Kleckner RC, McCleave JD. The northern limit of spawning by Atlantic eels (Anguilla spp.) in the Sar-gasso Sea in relation to thermal fronts and surface water masses. J. Mar. Res. 1988; 46: 647–667.
Kleckner RC, McCleave JD, Wippelhauser GS. Spawning of American eel, Anguilla rostrata, relative tothermal fronts in the Sargasso Sea. Env. Biol. Fishes 1983; 9: 289–293.
Knights B. A review of the possible impacts of long-term oceanic and climate changes and fishing mortal-ity on recruitment of anguillid eels of the Northern Hemisphere. Science Total Environ. 2003; 310:237–244.
Kruse C, Strehlow B, Schmidt H, Müller PK. Presence of trypsin in distinctive body segments of lepto-cephalus larvae of Anguilliformes. Aquaculture 1996; 142: 237–244.
Kurokawa T, Pedersen BH. The digestive system of eel larvae. In: Aida K, Tsukamoto K, Yamauchi K(eds). Eel Biology. Springer, Tokyo. 2003; pp. 435–444.
Kurokawa T, Kagawa H, Ohta H, Tanaka H, Okuzawa K, Hirose K, Development of digestive organs andfeeding ability in larvae of Japanese eel (Anguilla japonica). Can. J. Fish. Aquat. Sci. 1995; 52:1030–1036.
Kuroki M, Aoyama J, Miller MJ, Arai T, Sugeha HY, Minagawa G, Wouthuyzen S, Tsukamoto K. Corre-spondence between otolith microstructural changes and early life history events in Anguillamarmorata leptocephali and glass eels. Coast. Mar. Sci. 2005; 29: 154–161.
Kuroki M, Aoyama J, Miller MJ, Wouthuyzen S, Arai T, Tsukamoto K. Contrasting patterns of growth andmigration of tropical anguillid leptocephali in the western Pacific and Indonesian Seas. Mar. Ecol.Progr. Ser. 2006a; 309: 233–246.
Kuroki M, Aoyama J, Wouthuyzen S, Sumardhiharga K, Miller MJ, Minagawa G, Tsukamoto K. Age andgrowth of Anguilla interioris leptocephali collected in Indonesian waters. Coastal Mar. Sci. 2006b;30: 264–268.
Kuroki M, Aoyama J, Wouthuyzen S, Sumardiharga K, Miller MJ, Tsukamoto K. Age and growth ofAnguilla bicolor bicolor leptocephali in the eastern Indian Ocean. J. Fish Biol. 2007; 70: 538–550.
Kuroki M, Aoyama J, Miller MJ, Watanabe S, Shinoda A, Jellyman DJ, Tsukamoto K. Distribution andearly life history characteristics of anguillid leptocephali in the western South Pacific Ocean. Mar.Freshw. Res. 2008a; 59: 1035–1047.
Kuroki M, Kawai M, Jónsson B, Aoyama J, Miller MJ, Noakes DLG, Tsukamoto K. Inshore migrationand otolith microstructure/microchemistry of anguillid glass eels recruited to Iceland. Environ. Biol.Fish. 2008b; 83: 309–325.
Kuroki M, Aoyama J, Miller MJ, Yoshinaga T, Shinoda S, Hagihara S, Tsukamoto K. Sympatric spawningof Anguilla marmorata and Anguilla japonica in the western North Pacific Ocean. J. Fish Biol.2009a; 74: 1853–1865.
Kuroki M, Aoyama J, Kim H, Miller MJ, Kimura S, Tsukamoto K. Migration scales of catadromous eels:diversity and evolution of larval migration based on their distribution, morphology and early lifehistory. In: Haro A, Smith K, Rulifson R, Moffitt C, Klauda R, Dadswell M, Cunjak R, Cooper J,Beal K, Avery T (eds). Challenges for Diadromous Fishes in a Dynamic Global Environment. Am.Fish. Soc. Symp. 69. Bethesda, Maryland. 2009b; pp. 887–889.
Lampert W. The adaptive significance of diel vertical migration of zooplankton. Funct. Ecol. 1989; 3: 21–27.
Lecomte-Finiger R. Growth history and age at recruitment of European glass eels (Anguilla anguilla) asrevealed by otolith microstructure. Mar. Biol. 1992; 114: 205–210.
Lecomte-Finiger R. The early life of the European eel. Nature 1994; 370: 424.Lecomte-Finiger R. Leptocephali, these unappreciated larvae. Cybium 2004; 28: 83–95 (in French).

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 87
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Lee C, Armstrong RA, Cochran JK, Engel A, Fowler SW, Goutx M, Masqué P, Miquel JC, Peterson M,Tamburini C, Wakeham S. MedFlux: Investigations of particle flux in the Twilight Zone. Deep-SeaRes. II 2009; 56: 1363–1368.
Lee TW, Byun JS. Microstructural growth in otoliths of conger eel (Conger myriaster) leptocephali dur-ing the metamorphic stage. Mar. Biol. 1996; 125: 259–268.
Lee TW, Miller MJ, Hwang HB, Wouthuyzen S, Tsukamoto K. Distribution and early life history ofKaupichthys leptocephali (family Chlopsidae) in the central Indonesian Seas. Mar. Biol. 2008; 153:285–295.
Leiby MM. Morphological development of the eel Myrophis punctatus (Ophichthidae) from hatching tometamorphosis, with emphasis on the developing head skeleton. Bull. Mar. Sci. 1979a; 29: 509–521.
Leiby MM. Leptocephalus larvae of the eel family Ophichthidae. I. Ophichthus gomesi Castelnau. Bull.Mar. Sci. 1979b; 29: 329–343.
Leiby MM. Family Ophichthidae: Leptocephali. In: Böhlke EB (ed). Fishes of the Western North Atlantic.Leptocephali. Part 9, Vol. 2. Sears Foundation for Marine Research, New Haven. 1989; pp. 764–897.
Leis JM. Pacific coral-reef fishes: the implications of behaviour and ecology of larvae for biodiversity andconservation, and a reassessment of the open population paradigm. Environ. Biol. Fish. 2002; 65:199–208.
Leis JM. Are larvae of demersal fishes plankton or nekton? Adv. Mar. Biol. 2006; 51: 57–141.Leis JM, Carson-Ewart MBM. Orientation of pelagic larvae of coral-reef fishes in the ocean. Mar. Ecol.
Progr. Ser. 2003; 252: 239–253.Leis JM, McCormick MI. The biology, behaviour, and ecology of the pelagic, larval stage of coral reef
fishes. In: Sale PF (ed). Coral Reef Fishes: Dynamics and Diversity in a Complex Ecosystem. Aca-demic, San Diego, 2002; pp 171–199.
Leis JM, Lockett MM. Localization of reef sounds by settlement-stage larvae of coral-reef fishes(Pomacentridae). Bull. Mar. Sci. 2005; 76: 715–724.
Leonard JB, Summers RG. The ultrastructure of the integument of the American eel, Anguilla rostrata.Cell Tissue Res. 1976; 171: 1–30.
Levitus S, Antonov JI, Boyer TP. Warming of the world ocean. Science 2000; 287: 2,225–2,229.Ling YJ, Iizuka Y, Tzeng WN. Decreased Sr/Ca ratios in the otoliths of two marine eels, Gymnothorax
reticularis and Muraenesox cinereus, during metamorphosis. Mar. Ecol. Progr. Ser. 2005; 304: 201–206.
Linton ED, Jónsson B, Noakes DLG. Effects of water temperature on the swimming and climbing behav-iour of glass eels, Anguilla spp. Environ. Biol. Fish 2007; 78: 189–192.
Llopiz JK, Cowen RK. Variability in the trophic role of coral reef fish larvae in the oceanic plankton. Mar.Ecol. Progr. Ser. 2009; 381: 259–272.
Lohmann KJ, Putman NF, Lohmann CMF. Geomagnetic imprinting: A unifying hypothesis of long-dis-tance natal homing in salmon and sea turtles. Proc. Nat. Acad. Sci. 2008; 105; 19,096–19,101.
Lopez JA, Westneat MW, Hanel R. The phylogenctic affinities of the mysterious anguilliform generaColoconger and Thalassenchelys as supported by mtDNA sequences. Copeia 2007; 2007: 959–966.
Losey GS, Cronin TW, Goldsmith TH, Hyde D, Marshall, NJ, McFarland WN. The UV visual world offishes: a review. J. Fish Biol. 1999; 54: 921–943.
Ma T, Miller MJ, Shinoda A, Minagawa G, Aoyama J, Tsukamoto K. Age and growth of Saurenchelys(Nettastomatidae) and Dysomma (Synaphobranchidae) leptocephali in the East China Sea. J. FishBiol. 2005; 67: 1619–1630.
Ma T, Miller MJ, J Aoyama, K Tsukamoto. Genetic identification of Conger myriaster leptocephali in theEast China Sea region. Fish. Sci. 2007; 73: 989–994.
Ma T, Miller MJ, Aoyama J, Minagawa G, Inoue JG, Watanabe S, Tsukamoto K. Genetic identification oftwo species of Ariosoma leptocephali. Coast. Mar. Sci. 2008a; 32: 48–53.
Ma T, Aoyama J, Miller MJ, Minegishi Y, Inoue JG, Tsukamoto K. Genetic differentiation in the genusUroconger in the Indo–Pacific region. Aquat. Biol. 2008b; 2: 29–35.
Madin LP, Kremer P, Wiebe PH, Purcell JE, Horgan EH, Nemazie DA. Periodic swarms of the salp Salpaaspera in the Slope Water off the NE United States: biovolume, vertical migration, grazing, andvertical flux. Deep-Sea Res. I 2006; 53: 804–819.
Martin MH. Validation of daily growth increments in otoliths of Anguilla rostrata (Lesueur) elvers. Can.J. Zool. 1995; 73; 208–211.
Marui M, Arai T, Miller MJ, Jellyman DJ, Tsukamoto K. Comparison of early life history between NewZealand temperate eels and Pacific tropical eels revealed by otolith microstructure and microchem-istry. Mar. Ecol. Progr. Ser. 2001; 213: 273–284.

88 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Masuda R. Behavioral ontogeny of marine pelagic fishes with the implications of the sustainable manage-ment of fisheries resources. Aqua-BioSci. Monogr. (ABSM) 2009; 2(2): 1–56.
Matarese AC, Kendall AW, Blood DM, Vinter BM. Laboratory guide to early life history stages of north-east Pacific fishes. 1989; NOAA Technical Report NMFS 80.
McCleave JD. Physical and behavioural controls on the oceanic distribution and migration of leptocephali.J. Fish Biol. 1993; 43(Suppl A): 243–273.
McCleave JD. Spawning areas of Atlantic eels. In: Aida K, Tsukamoto K, Yamauchi K (eds). Eel Biology.Springer, Tokyo. 2003; pp. 141–156.
McCleave JD. Contrasts between spawning times of Anguilla species estimated from larval sampling atsea and from otolith analysis of recruiting glass eels. Mar. Biol. 2008; 155: 249–262.
McCleave JD, Kleckner RC. Distribution of leptocephali of the catadromous Anguilla species in the west-ern Sargasso Sea in relation to water circulation and migration. Bull. Mar. Sci. 1987; 41: 789–806.
McCleave JD, Miller MJ. Spawning of Conger oceanicus and Conger triporiceps (Congridae) in theSargasso Sea and subsequent distribution of leptocephali. Environ. Biol. Fish. 1994; 39: 339–355.
McCleave JD, Kleckner RC, Castonguay M. Reproductive sympatry of American and European eels andimplications for migration and taxonomy. In: Dadswell MJ, Klauda RJ, Moffitt CM, Saunders RL,Rulifson RA, Cooper JE (eds). Common Strategies of Anadromous and Catadromous Fishes. Am.Fish. Soc. Symp. 1, Bethesda, Maryland. 1987; pp. 286–297.
McCleave JD, Brickley PJ, O’Brien KM, Kistner-Morris DA, Wong MW, Gallagher M, Watson SM. Doleptocephali of the European eel swim to reach continental waters? Status of the question. J. Mar.Biol. Assoc. UK 1998; 78: 285–306.
McCormick MI. Delayed metamorphosis of a tropical reef fish (Acanthurus triostegus): a field experi-ment. Mar. Ecol. Progr. Ser. 1999; 176: 25–38.
McIlwain JL. Fine-scale temporal and spatial patterns of larval supply to a fringing reef in Western Aus-tralia. Mar. Ecol. Progr. Ser. 2003; 252: 207–222.
Miller MJ. Species assemblages of leptocephali in the Sargasso Sea and Florida Current. Mar. Ecol. Progr.Ser. 1995; 121: 11–26.
Miller MJ. Distribution and ecology of Ariosoma balearicum (Congridae) leptocephali in the westernNorth Atlantic. Environ. Biol. Fish. 2002a; 63: 235–252.
Miller MJ. Contrasting migratory strategies of marine and freshwater eels. Fish. Sci. 2002b; 68(Suppl. I):37–40.
Miller MJ. The worldwide distribution of anguillid leptocephali. In: Aida K, Tsukamoto K, Yamauchi K(eds). Eel Biology. Springer, Tokyo. 2003; pp. 157–168.
Miller MJ, McCleave JD. Species assemblages of leptocephali in the subtropical convergence zone of theSargasso Sea. J. Mar. Res. 1994; 52: 743–772.
Miller MJ, McCleave JD. Species assemblages of leptocephali in the southwestern Sargasso Sea. Mar.Ecol. Progr. Ser. 2007; 344: 197–212.
Miller MJ, Tsukamoto K. An Introduction to Leptocephali: Biology and Identification. Ocean ResearchInstitute, University of Tokyo, Tokyo. 2004.
Miller MJ, Tsukamoto K. Studies on eels and leptocephali in Southeast Asia: A new research frontier.Coastal Mar. Sci. 2006; 30: 283–292.
Miller MJ, Otake T, Minagawa G, Inagaki T, Tsukamoto K. Distribution of leptocephali in the KuroshioCurrent and East China Sea. Mar. Ecol. Progr. Ser. 2002a; 235: 279–288.
Miller MJ, Mochioka N, Otake T, Tsukamoto K. Evidence of a spawning area of Anguilla marmorata inthe western North Pacific. Mar. Biol. 2002b; 140: 809–814.
Miller MJ, Wouthuyzen S, Minagawa G, Tsukamoto K. Distribution and species richness of leptocephaliin the Indonesian Seas. Oseanologi dan Limnol. di Indon. 2002c; 34: 55–69.
Miller MJ, Minegishi Y, Kuroki M, Inagaki T. Leptocephali collected in the Sulu Sea and surroundingareas. In: Nishida S, Gamo T (eds). Preliminary report of the Hakuho Maru Cruise KH-02-4 (TheSulu Sea and Western Pacific). Ocean Research Institute, University of Tokyo. 2004; pp 59–63.
Miller MJ, Aoyama J, Mochioka N, Otake T, Castle PHJ, Minagawa G, Inagaki T, Tsukamoto K. Geo-graphic variation in the assemblages of leptocephali in the western South Pacific. Deep-Sea Res. I2006a; 53: 776–794.
Miller MJ, Wouthuyzen S, Minagawa G, Aoyama J, Tsukamoto K. Distribution and ecology of lepto-cephali of the congrid eel, Ariosoma scheelei, around Sulawesi Island, Indonesia. Mar. Biol. 2006b;148: 1101–1111.
Miller MJ, Kimura S, Friedland KD, Knights B, Kim H, Jellyman JD, Tsukamoto K. Review of ocean-atmospheric factors in the Atlantic and Pacific oceans influencing spawning and recruitment ofanguillid eels. In: Haro A, Smith K, Rulifson R, Moffitt C, Klauda R, Dadswell M, Cunjak R,

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 89
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Cooper J, Beal K, Avery T (eds). Challenges for Diadromous Fishes in a Dynamic Global Environ-ment. Am. Fish. Soc. Symp. 69. Bethesda, Maryland. 2009a; pp. 231–249.
Miller MJ, Powell J, Tsukamoto K. Observation of a large metamorphosing leptocephalus in a coral reefhabitat at Sangeang Island, Indonesia. Zool. Stud. 2009b; 48: 107.
Miller T, Crowder LB, Rice JA, Marschall EA. Larval size and recruitment mechanisms in fishes: towarda conceptual framework. Can. J. Fish. Aquat. Sci. 1988; 45: 1657–1670.
Minagawa G, Miller MJ. Distribution and diversity patterns of leptocephali. In: Watanabe Y (ed). LivingMarine Resources: How Do They Fluctuate in the Ocean? Tokai University Press, Tokyo. 2005;pp. 168–189 (in Japanese).
Minagawa G, Miller MJ, Aoyama J, Wouthuyzen S, Tsukamoto K. Contrasting assemblages of lepto-cephali in the western Pacific. Mar. Ecol. Progr. Ser. 2004; 271: 245–259.
Minagawa G, Miller MJ, Kimura Y, Watanabe S, Shinoda A, Aoyama J, Tsukamoto K. Seasonal differ-ences in catches of leptocephali in the East China Sea and Suruga Bay, Japan. Est. Coast. Shelf Sci.2007; 71: 730–740.
Minegishi Y, Aoyama J, Tsukamoto K. Multiple population structure of the giant mottled eel, Anguillamarmorata. Molec. Ecol. 2008; 17: 3109–3122.
Minegishi Y, Yoshinaga T, Aoyama J, Tsukamoto K. Species identification of Anguilla japonica by real-time PCR based on a sequence detection system: a practical application to eggs and larvae. ICES J.Mar. Sci. 2009; 66: 1915–1918.
Mochioka N. Leptocephali. In: Aida K, Tsukamoto K, Yamauchi K (eds). Eel Biology. Springer, Tokyo.2003; pp. 51–60.
Mochioka N, Iwamizu M. Diet of anguillid larvae: leptocephali feed selectively on larvacean houses andfecal pellets. Mar. Biol. 1996; 125: 447–452.
Mochioka N, Kakuda S, Tabeta O. Congrid leptocephali in the western North and Middle Pacific—IExterilium Ariosoma-type larvae. J. Fac. Appl. Biol. Sci. Hiroshima Univ. 1982; 21: 35–66.
Mochioka N, Tabeta O, Kakuda S, Tsukahara H. Congrid leptocephali in the western north and middlePacific—II. Non-exterilium Ariosoma type larvae. Bull. Mar. Sci. 1991; 48: 606–622.
Mochioka N, Iwamizu M, Kanda T. Leptocephalus eel larvae will feed in aquaria. Environ. Biol. Fish.1993; 36: 381–384.
Morales-Nin B. Review of the growth regulation processes of otolith daily increment formation. FisheriesRes. 2000; 46: 53–67.
Moser HG. Morphological and functional aspects of marine fish larvae. In: Lasker R (ed). Marine FishLarvae: Morphology, Ecology, and Relation to Fisheries. Washington Sea Grant Program, Seattle,WA. 1981; pp. 89–131.
Moyer JT, Zaiser MJ. Reproductive behavior of moray eels at Miyade-jima, Japan. 1982; Jpn. J. Ichthyol.28: 466–468.
Nakamura O, Saeki M, Kamiya H, Muramoto K, Watanabe T. Development of epidermal and mucosalgalectin containing cells in metamorphosing leptocephali of Japanese conger. J. Fish Biol. 2002;61: 822–833.
Nelson JS. Fishes of the World. 4th edn. John Wiley and Sons, Hoboken, NJ. 2006.Nishi T, Kawamura G. Anguilla japonica is already magnetosensitive at the glass eel phase. J. Fish Biol.
2005; 67: 1213–1224.Nishi T, Kawamura G, Matsumoto K. Magnetic sense in the Japanese eel, Anguilla japonica as deter-
mined by conditioning and electrocardiography. J. Exper. Biol. 2004; 207: 2965–2970.Nolan CJ, Danilowicz BS. Advantages of using crest nets to sample presettlement larvae of reef fishes in
the Caribbean Sea. Fishery Bull. 2008; 106: 213–221.Okamura A, Utoh T, Zhang H, Yamada Y, Horie N, Mikawa N, Tanaka S, Motonobu T, Oka HP. Seasonal
changes in maturity in the conger eel Conger myriaster at the Pacific coast of Atsumi peninsula,central Japan. Nippon Suisan Gakkaishi 2000; 66: 412–416.
Okamura A, Oka HP, Yamada Y, Utoh T, Mikawa N, Horie N, Tanaka S. Development of lateral lineorgans in leptocephali of the freshwater eel Anguilla japonica (Teleostei, Anguilliformes). J. Morphol.2002; 254: 81–91.
Okumura Y, Xie SP, Interaction of the Atlantic equatorial cold tongue and the African monsoon. J. Cli-mate 2004; 17: 3589–3602.
Otake T. Fine structure and function of the alimentary canal in leptocephali of the Japanese eel Anguillajaponica. 1996; Fish. Sci. 62: 28–34.
Otake T. Metamorphosis. In: Aida K, Tsukamoto K, Yamauchi K (eds). Eel Biology. Springer, Tokyo.2003; pp. 61–74.
Otake T, Nogami K, Maruyama K. Possible food sources of leptocephali. La mer 1990; 28: 218–224.

90 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Otake T, Nogami K, Maruyama K. Dissolved and particulate organic matter as possible food sources foreel leptocephali. Mar. Ecol. Progr. Ser. 1993; 92: 27–34.
Otake T, Ishii T, Nakahara M, Nakamura R. Drastic changes in otolith strontium/calcium ratios in lepto-cephali and glass eels of Japanese eel Anguilla japonica. Mar. Ecol. Progr. Ser. 1994; 112: 189–193.
Otake T, Hirokawa J, Fujimoto H, Imaizumi K. Fine structure and function of the gut epithelium of pikeeel larvae. J. Fish Biol. 1995; 47: 126–142.
Otake T, Ishii T, Ishii T, Nakahara M, Nakamura R. Changes in otolith strontium:calcium ratios in meta-morphosing Conger myriaster leptocephali. Mar. Biol. 1997; 128: 565–572.
Otake T, Inagaki T, Hasumoto H, Mochioka N, Tsukamoto K. Diel vertical distribution of Anguilla japonicaleptocephali. Ichthyol. Res. 1998; 45: 208–211.
Otake T, Miller MJ, Inagaki T, Minagawa G, Shinoda A, Kimura Y, Sasai S, Oya M, Tasumi S, Suzuki Y,Uchida M, Tsukamoto K. Evidence for migration of metamorphosing larvae of Anguilla japonica inthe Kuroshio. Coast. Mar. Sci. 2006; 30: 453–458.
Ozaki Y, Okumura H, Kazeto Y, Ikeuchi T, Ijiri S, Nagae M, Adachi S, Yamauchi K. Developmentalchanges in pituitary-thyroid axis, and formation of gonads in leptocephali and glass eels of Anguillaspp. Fish. Sci. 2000; 66: 1115–1122.
Ozaki Y, Tanaka H, Kagawa H, Ohta H, Adachi S, Yamauchi K. Fine structure and differentiation of thealimentary canal in captive-bred Japanese eel Anguilla japonica preleptocephali. Fish. Sci. 2006a;72: 13–19.
Ozaki Y, Fukada H, Tanaka H, Kagawa H, Ohta H, Adachi S, Hara A, Yamauchi K. Expression of growthhormone family and growth hormone receptor during early development in the Japanese eel (Anguillajaponica). Comp. Biochem. Physiol. Part B: Biochem. Mol. Biol. 2006b; 145: 27–34.
Padrón D, Lindley VA, Pfeiler E. Changes in lipid composition during metamorphosis of bonefish (Albulasp.) leptocephali. Lipids 1996; 31: 513–519.
Pankhurst NW. Retinal development in larval and juvenile European eel, Anguilla anguilla (L.). Can. J.Zool. 1984; 62: 335–343.
Pedersen BH, Ueberscha B, Kurokawa T. Digestive response and rates of growth in pre-leptocephaluslarvae of the Japanese eel Anguilla japonica reared on artificial diets. Aquaculture 2003; 215: 321–338.
Pfeiler E. Towards an explanation of the developmental strategy in leptocephalus larvae of marine fishes.Environ. Biol. Fish. 1986; 15: 3–13.
Pfeiler E. Isolation and partial characterization of a novel keratan sulfate proteoglycan from metamor-phosing bonefish (Albula) larvae. Fish Physiol. Biochem. 1988; 4: 175–187.
Pfeiler E. Sensory systems and behavior of premetamorphic and metamorphic leptocephalus larvae. BrainBehav. Evol. 1989; 34: 25–34.
Pfeiler E. Glycosaminoglycan composition of anguilliform and elopiform leptocephali. J. Fish Biol. 1991;38: 533–540.
Pfeiler E. Characterization and distribution of undersulfated chondroitin sulfate and chondroitin in lepto-cephalus larvae of elopomorph fishes. Fish Physiol. Biochem. 1993; 12: 143–148.
Pfeiler E. Energetics of metamorphosis in bonefish (Albula sp.) leptocephali: Role of keratan sulfateglycosaminoglycan. Fish Physiol. Biochem. 1996; 15: 359–62.
Pfeiler E. Effect of Ca2+ on survival and development of metamorphosing bonefish (Albula sp.) lepto-cephali. Mar. Biol. 1997; 127: 571–578.
Pfeiler E. Developmental physiology of elopomorph leptocephali. Comp. Biochem. Physiol. 1999; A123:113–128.
Pfeiler E. Changes in hypoxia tolerance during metamorphosis of bonefish leptocephali. J. Fish Biol.2001; 59: 1677–1681.
Pfeiler E, Govoni JJ. Metabolic rates in early life history stages of elopomorph fishes. Biol. Bull. 1993;185: 277–283.
Pfeiler E, Lindley V. Chloride-type cells in the skin of the metamorphosing bonefish (Albula sp.) lepto-cephalus. J. Exp. Zool. 1989; 250: 11–16.
Pfeiler E, Lindley VA, Elser JJ. Elemental (C, N and P) analysis of metamorphosing bonefish (Albula sp.)leptocephali: relationship to catabolism of endogenous organic compounds, tissue remodeling, andfeeding ecology. Mar. Biol. 1998; 132: 21–28.
Pfeiler E, Toyoda H, Williams MD, Nieman RA. Identification, structural analysis and function ofhyaluronan in developing fish larvae (leptocephali). Comp. Biochem. Physiol. B, Biochem. Molec.Biol. 2002; 132: 443–451.
Pilskaln CH, Villareal TA, Dennett M, Darkangelo-Wood C, Meadows G High concentrations of marinesnow and diatom algal mats in the North Pacific Subtropical Gyre: Implications for carbon andnitrogen cycles in the oligotrophic ocean. Deep-Sea Res. I 2005; 52: 2315–2332.

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 91
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Powles PM, Hare JA, Laban EH, Warlen SM. Does eel metamorphosis cause a breakdown in the tenets ofotolith applications? A case study using the speckled worm eel (Myrophis punctatus, Ophichthidae).Can. J. Fish. Aquat. Sci. 2006; 63: 1460–1468.
Raju SN. Congrid eels of the eastern pacific and key to their leptocephali. NOAA Technical Report NMFS22, U.S. Department of Commerce. 1985; 1–19.
Randall JE. Zoogeography of shore fishes of the Indo–Pacific region. Zool. Stud. 1998; 37: 227–267.Randall JE. A review of mimicry in marine fishes. Zool. Stud. 2005; 44: 299–328.Reed RN, Shine R, Shetty S. Sea kraits (Squamata: Laticauda spp.) as a useful bioassay for assessing local
diversity of eels (Muraenidae, Congridae) in the western Pacific Ocean. Copeia 2002; 2002: 1098–1101.
Richardson DE, Cowen RK. Diversity of leptocephalus larvae around the island of Barbados (West Indies):relevance to regional distributions. Mar. Ecol. Progr. Ser. 2004a; 282: 271–284.
Richardson DE, Cowen RK. New leptocephalus types collected around the Island of Barbados (WestIndies). Copeia 2004b; 2004: 888–895.
Richardson DE, Vanwye JD, Exum AM, Cowen RK, Crawford DL. High-throughput species identifica-tion: from DNA isolation to bioinformatics. Molec. Ecol. Notes 2007; 7: 199–207.
Ross SW, Rohde FC. Collections of ophichthid eels on the surface at night off North Carolina. Bull. Mar.Sci. 2003; 72: 241–246.
Ross SW, Casazza TL, Quattrini AM, Sulak KJ. Anguilliform larvae collected off North Carolina. Mar.Biol. 2007; 150: 681–695.
Sasai S, Kaneko T, Tsukamoto K. Extrabranchial chloride cells in early life stages of the Japanese eel,Anguilla japonica. Ichthyol. Res. 1998; 45: 95–98.
Sato R, Tanaka Y, Ishimaru T. Species-specific house productivity of appendicularians. Mar. Ecol. Progr.Ser. 2003; 259: 163–172.
Schalk PH. Respiratory electron transport system (ETS) activities in zooplankton and micronekton of theIndo–Pacific region. Mar. Ecol. Progr. Ser. 1988; 44: 25–35.
Schmidt J. The breeding places of the eel. Phil. Trans. R. Soc. Lond. 1922; 211: 179–208.Schoth M, Tesch F-W. Spatial distribution of 0-group eel larvae (Anguilla sp.) in the Sargasso Sea.
Helgoländer Meeresunters 1982; 35: 309–320.Schoth M, Tesch F-W. Vertical distribution of 0-group Anguilla larvae in the Sargasso Sea with reference
to other anguilliform leptocephali. Meeresforsch 1984; 30: 188–195.Shenker JM, Maddox ED, Wishinski E, Pearl A, Thorrold SR, Smith N. Onshore transport of settlement-
stage Nassau grouper Epinephelus striatus and other fishes in Exuma Sound, Bahamas. Mar. Ecol.Progr. Ser. 1993; 98: 31–43.
Shiao JC, Hwang PP. Thyroid hormones are necessary for the metamorphosis of tarpon Megalopscyprinoides leptocephali. J. Exper. Mar. Biol. Ecol. 2006; 331: 121–132.
Shiao JC, Tzeng WN, Collins A, Iizuka Y. Role of marine larval duration and growth rate of glass eels indetermining the distribution of Anguilla reinhardtii and A. australis on Australian eastern coasts.Mar. Freshw. Res. 2002; 53: 687–695.
Shinoda A, Tanaka H, Kagawa H, Ohta H, Tsukamoto K. Otolith microstructural analysis of reared larvaeof the Japanese eel Anguilla japonica. Fish. Sci. 2004; 70: 340–342.
Siegel DA, Fields E, Buesseler KO. A bottom-up view of the biological pump: Modeling source funnelsabove ocean sediment traps. Deep-Sea Res. I 2008; 55: 108–127.
Smith DG. Xenocongrid eel larvae in the western North Atlantic. Bull. Mar. Sci. 1969; 19: 377–408.Smith DG. Dysommid eel larvae in the western North Atlantic. Copeia 1974; 1974: 671–680.Smith DG. Guide to the Leptocephali (Elopiformes, Anguilliformes, and Notacanthiformes). NOAA Tech-
nical Report NMFS Circular 424, U.S. Department of Commerce. 1979; 39 pp.Smith DG. Introduction to leptocephali. In: Böhlke EB (ed). Fishes of the Western North Atlantic. Lepto-
cephali. Part 9, Vol. 2. Sears Foundation for Marine Research, New Haven. 1989a; pp. 657–668.Smith DG. Family Muraenidae: Leptocephali. In: Böhlke EB (ed). Fishes of the Western North Atlantic.
Leptocephali. Part 9, Vol. 2. Sears Foundation for Marine Research, New Haven. 1989b; pp. 900–916.
Smith DG, Castle PHJ. Larvae of the nettastomatid eels: systematics and distribution. Dana Rep. 1982;90: 44 pp.
Smith DG, Miller MJ Cyematid larvae of the Leptocephalus holti group in the Atlantic and Pacific oceans(Pisces: Saccopharyngiformes). Breviora 1996; 503: 1–12.
Stemmann L, Jackson GA, Gorsky G. A vertical model of particle size distributions and fluxes in themidwater column that includes biological and physical processes—Part II: application to a threeyear survey in the NW Mediterranean Sea. Deep-Sea Res. I 2004; 51: 885–908.

92 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Strehlow B, Antunes C, Niermann U, Tesch F-W. Distribution and ecological aspects of leptocephalicollected 1979–-1994 in North- and Central Atlantic. I. Congridae. Helgoländer Meeresunters 1998;52: 85–102.
Stumpf RP, Feldman G, Kuring N, Franz B, Green E, Robinson J. SeaWiFS spies reefs. Reef Encounter1999; 26: 29–30.
Sugeha HY, Shinoda A, Marui M, Arai T, Tsukamoto K. Validation of otolith daily increments in thetropical eel Anguilla marmorata. Mar. Ecol. Progr. Ser. 2001a; 220: 291–294.
Sugeha, HY, Arai T, Miller MJ, Limbong D, Tsukamoto K. Inshore migration of the tropical eels, Anguillaspp., recruiting to the Poigar River estuary on Sulawesi Island. Mar. Ecol. Progr. Ser. 2001b; 221:233–243.
Sullivan MC, Able KW, Hare JA, Walsh HJ. Anguilla rostrata glass eel ingress into two, US east coastestuaries: patterns, processes and implications for adult abundance. J. Fish Biol. 2006; 69: 1081–1101.
Sullivan MC, Wuenschel MJ, Able K. Inter and intra-estuary variability in ingress, condition and settle-ment of the American eel Anguilla rostrata. J. Fish Biol. 2009; 74: 1949–1969.
Suzuki Y, Otake T. Skin lectin and the lymphoid tissues in the leptocephalus larvae of the Japanese eelAnguilla japonica. Fish. Sci. 2000; 66: 636–643.
Tabeta O, Mochioka N. Leptocephali. In: Okiyama M (ed). An Atlas of the Early Stage Fishes in Japan.Tokai University Press, Tokyo, 1988a; pp. 15–64 (in Japanese).
Tabeta O, Mochioka N. Anguilliform and elopiform leptocephali. In: Kajihara T (ed). Preliminary reportof the Hakuho Maru Cruise KH-86-4. Ocean Research Institute, University of Tokyo. 1988b;pp. 22–25.
Takagi Y, Tohse H, Murayama E, Ohira T, Nagasawa H. Diel changes in endolymph aragonite saturationrate and mRNA expression of otolith matrix proteins in the trout otolith organ. Mar. Ecol. Progr.Ser. 2005; 294: 249–256.
Takahashi M, Mochioka N, Shinagawa S, Yatsu A, Nakazono A. Distribution patterns of leptocephali inthe Kuroshio–Oyashio transitional region of the western North Pacific. Fish. Oceanogr. 2008; 17:165–177.
Takai T. Studies on the morphology, ecology and culture of the important apodal fishes, Muraenesoxcinereus (Folskal) and Conger myriaster (Brevoort). J. Shimonoseki Coll. Fisheries. 1959; 8: 209–555 (in Japanese).
Takai T. Review: Reproduction of the pike eel in the Seto Inland Water. Saibaigiken 1979; 8: 77–82 (inJapanese).
Tanaka H. Techniques for larval rearing. In: Aida K, Tsukamoto K, Yamauchi K (eds). Eel Biology. Springer,Tokyo. 2003; pp. 427–434.
Tanaka H, Kagawa H, Ohta H, Okuzawa K, Hirose K. The first report of eel larvae ingesting rotifers. Fish.Sci. 1995; 61: 171–172.
Tanaka H, Kagawa H, Ohta H. Production of leptocephali of Japanese eel (Anguilla japonica) in captivity.Aquaculture 2001; 201: 51–60.
Tanaka H, Kagawa H, Ohta H, Unuma T, Nomura K. The first production of glass eel in captivity: fishreproductive physiology facilitates great progress in aquaculture. Fish Physiol. Biochem. 2003; 28:493–497.
Teletchea F. Molecular identification methods of fish species: reassessment and possible applications.Rev. Fish Biol. Fisheries 2009; 19: 265–293.
Tesch F-W. Occurrence of eel Anguilla anguilla larvae west of the European continental shelf, 1971–1977. Env. Biol. Fish. 1980; 5: 185–190.
Tesch F-W. The Eel. Blackwell Science, Oxford. 2003; 408 pp.Tesch F-W, Wegner G. The distribution of small larvae of Anguilla sp. related hydrographic conditions
1981 between Bermuda and Puerto Rico. Int. Rev der Ges Hydrobiol. 1990; 75: 845–858.Thorrold SR, Shenker JM, Wishinski E, Mojica R, Maddox ED. Larval supply of shorefishes to nursery
habitats around Lee Stocking Island, Bahamas. I. Small-scale distribution patterns. Mar. Biol. 1994a;118: 555–566.
Thorrold SR, Shenker JM, Maddox ED, Mojica R, Wishinski E. Larval supply of shorefishes to nurseryhabitats around Lee Stocking Island, Bahamas. II. Lunar and oceanographic influences. Mar. Biol.1994b; 118: 567–578.
Thorrold SR, Shenker JM, Mojica R, Maddox ED, Wishinski E. Temporal patterns in the larval supply ofsummer-recruiting reef fishes to Lee Stocking Island, Bahamas Mar. Ecol. Progr. Ser. 1994c; 112:75–86.
Thresher RE. Reproduction in Reef Fishes. Tropical Fish Hobbyist Publications, Neptune City, New Jer-sey. 1984.

Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer 93
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Tomoda H, Uematsu K. Morphogenesis of the brain in larval and juvenile Japanese eels, Anguilla japonica.Brain Behav. Evol. 1996; 47: 33–41.
Tsukamoto K. Otolith daily increments in the Japanese eel. Nippon Suisan Gakkaishi 1989; 55: 1017–1021.
Tsukamoto K. Discovery of the spawning area for the Japanese eel. Nature. 1992; 356: 789–791.Tsukamoto K. Spawning of eels near a seamount. Nature. 2006; 439: 929.Tsukamoto K. Oceanic migration and spawning of anguillid eels. J. Fish Biol. 2009; 74: 1833–1852.Tsukamoto K, Umezawa A, Ozawa T. Age and growth of Anguilla japonica leptocephali collected in
western North Pacific in July 1990. Nippon Suisan Gakkaishi 1992; 58: 457–459.Tsukamoto K, Aoyama J, Miller MJ. Migration, speciation and the evolution of diadromy in anguillid
eels. Can. J. Fish. Aquat. Sci. 2002; 59: 1989–1998.Tsukamoto K, Otake T, Mochioka N, Lee TW, Fricke H, Inagaki T, Aoyama J, Ishikawa S, Kimura S,
Miller MJ, Hasumoto H, Oya M, Suzuki Y. Seamounts, new moon and eel spawning: the search forthe spawning site of the Japanese eel. Environ. Biol. Fish. 2003; 66: 221–229.
Tsukamoto K, Yamada Y, Okamura A, Tanaka H, Miller MJ, Kaneko T, Horie N, Utoh T, Mikawa N,Tanaka S. Positive buoyancy in eel leptocephali: an adaptation for life in the ocean surface layer.Mar. Biol. 2009; 156: 835–846.
Turner JT. Zooplankton fecal pellets, marine snow and sinking phytoplankton blooms. Aquat. MicrobialEcol. 2002; 27: 57–102.
Umezawa A, Tsukamoto K. Factors influencing otolith increment formation in Japanese eel, Anguillajaponica T. & S., elvers. J. Fish Biol. 1991; 39: 211–223.
Umezawa A, Tsukamoto K, Tabeta O, Yamakawa H. Daily growth increments in the larval otolith of theJapanese eel, Anguilla japonica. Jpn. J. Ichthyol. 1989; 35: 440–444.
Umezawa A, Otake T, Hirokawa J, Tsukamoto K, Okiyama M. Development of the eggs and larvae of thepike eel, Muraenesox cinereus. Jpn. J. Ichthyol. 1991; 38: 35–39.
Utoh T, Horie N, Mikawa N, Okamura A, Yamada Y, Akazawa A, Tanaka S, Oka HP. Annual changes inovarian development and plasma estradio-17β level in reared female common Japanese conger,Conger myriaster. Fish. Sci. 2005; 71: 38–47.
van den Thillart G, Dufour S, Rankin JC (eds). Spawning Migration of the European Eel. Fish and Fisher-ies Series, Vol. 30. Springer. 2009.
Varsamos S, Nebel C, Charmantier G. Ontogeny of osmoregulation in postembryonic fish: a review. Comp.Biochem. Physiol. A 2005; 141: 401–429.
Victor BC. Delayed metamorphosis with reduced larval growth in a coral reef fish (Thalassoma bifasciatum).Can. J. Fish. Aquat. Sci. 1986; 43: 1208–1213.
Virta VC, Cooper MS. Ontogeny and phylogeny of the yolk extension in embryonic cypriniform fishes.J. Exp. Zool. (Mol. Dev. Evol.) 2009; 312B: 196–223.
von Herbing H. Effects of temperature on larval fish swimming performance: the importance of physics tophysiology. J. Fish Biol. 2002; 61: 856–876.
Wang CH, Tzeng WN. The timing of metamorphosis and growth rates of American and European eelleptocephali: A mechanism of larval segregative migration. Fisheries Res. 2000; 46: 191–205.
Ward RD, Zemlak TS, Innes BH, Last PR, Herbert PDN. DNA barcoding Australia’s fish species. Phil.Trans. R. Soc. B 2005; 360: 1847-1857.
Warrant EJ, Locket NA. Vision in the deep sea. Biol. Rev. 2004; 79: 671–712.Watanabe S, Minegishi Y, Yoshinaga T, Aoyama J, Tsukamoto K. A quick method for species identifica-
tion of Japanese eel (Anguilla japonica) using real-time PCR: an onboard application for use duringsampling surveys. Mar. Biotech. 2004; 6: 566–574.
Watanabe S, Aoyama J, Miller MJ, Ishikawa S, Feunteun E, Tsukamoto K. Evidence of population struc-ture in the giant mottled eel, Anguilla marmorata, using total number of vertebrae. Copeia 2008;2008: 680–688.
Westerberg H. A proposal regarding the source of nutrition of leptocephalus larvae. Int. Revue ges Hydrobiol.1990; 75: 863–864.
Wiebe PH, Benfield MC. From the Hensen net toward four-dimensional biological oceanography. Progr.Oceanogr. 2003; 56: 7–136.
Wiebe PH, Morton AW, Bradley AM, Backus RH, Craddock JE, Barber V, Cowles TJ, Flierl GR. Newdevelopments in the MOCNESS, an apparatus for sampling zooplankton and micronekton. Mar.Biol. 1985; 87: 313–323.
Wippelhauser GS, Miller MJ, McCleave JD. Evidence of spawning and the larval distribution of nemichthyideels in the Sargasso Sea. Bull. Mar. Sci. 1996; 59: 298–309.

94 Ecology of Anguilliform Leptocephali: Remarkable Transparent Fish Larvae of the Ocean Surface Layer
Aqua-BioScience Monographs VOL. 2 NO. 4 2009
Wouthuyzen S, Miller MJ, Aoyama J, Minagawa G, Sugeha YH, Suharti S, Inagaki T, Tsukamoto K.Biodiversity of anguilliform leptocephali in the central Indonesian Seas. Bull. Mar. Sci. 2005; 77:209–224.
Wouthuyzen S, Aoyama J, Sugeha YH, Miller MJ, Kuroki M, Minegishi Y, Suharti S, Tsukamoto K.Seasonality of spawning by tropical anguillid eels around Sulawesi Island, Indonesia.Naturwissenshaften 2009; 96: 153–158.
Wu CC, Lin JC, Su WC. Diet and feeding habits of dolphin fish (Coryphaena hippurus) in the waters offeastern Taiwan. J. Taiwan Fisheries Res. 2006; 14: 13–27.
Wuenschel MJ, Able KW. Swimming ability of eels (Anguilla rostrata, Conger oceanicus) at estuarineingress: contrasting patterns of cross-shelf transport? Mar. Biol. 2008; 154: 775–786.
Yamada Y, Okamura A, Mikawa N, Utoh T, Horie N, Tanaka S, Miller MJ, Tsukamoto K. Ontogeneticchanges in phototaxis behavior during metamorphosis of artificially reared Anguilla japonica lar-vae. Mar. Ecol. Progr. Ser. 2009; 379: 241–251.
Yamano K, Tagawa M, De Jesus EG, Hirano T, Miwa S, Inui Y. Changes in whole body concentrations ofthyroid hormones and cortisol in metamorphosing conger eel. J. Comp. Physiol. B 1991; 161: 371–375.
Yamano K, Nomura K, Tanaka H. Development of thyroid gland and changes in thyroid hormone levels inleptocephali of Japanese eel (Anguilla japonica). Aquaculture 2007; 270: 499–504.
Yoshida M, Tomoda H, Uematsu K. Spinal and parachordal neurons in the eel leptocephali, Anguillajaponica revealed by labeling with a fluorescent dye. Neurosci. Lett. 1995; 195: 206–208.
Zenimoto K, Kitagawa T, Miyazaki S, Sasai T, Sasaki, H, Kimura S. The effects of seasonal and interannualvariability of oceanic structure in the western Pacific North Equatorial Current on larval transportof the Japanese eel (Anguilla japonica). J. Fish Biol. 2009; 74: 1878–1890.
Zwerger P, Nimeth K, Würtz J, Salvenmoser W, Pelster B. Development of the swimbladder in the Euro-pean eel (Anguilla anguilla). Cell Tissue Res. 2002; 307: 155–164.
Top Related