







Languages
Pages
Legal


Elaborado por Miguel Ángel Gamboa-Gaitán (MAGG)
Capítulo 8: Hoja
Introducción
La hoja es un órgano principalmente encargado de la fotosíntesis y del intercambio gaseoso con el ambiente, ya que por los
estomas se da la entrada de dióxido de carbono y la inevitable salida de vapor de agua. Su forma es altamente variable entre
especies e incluso entre individuos conespecíficos, por lo que presenta una elevada plasticidad fenotípica, de acuerdo con
las condiciones ambientales. En este capítulo se estudia la estructura básica de la hoja, así como las variaciones y las
adaptaciones morfoanatómicas y funcionales más notables presentes en las hojas de las plantas modernas.
Contenido
8.1. Las verdaderas hojas poseen vascularización interna.
8.2. Los primordios foliares originan las hojas en las yemas apicales de una manera ordenada.
8.3. La vascularización de la hoja se deriva del sistema vascular del tallo.
8.4. Hay una abundante variedad de formas y tamaños foliares.
8.5. La anatomía foliar es muy variada.
8.6. La vida de la hoja es de duración limitada.

MAGG – Botánica General: Introducción al estudio de las plantas
158
8.1. Las verdaderas hojas poseen vascularización
interna.
La hoja es un órgano vital para las plantas porque su
función primordial es hacer fotosíntesis. Sin embargo,
aunque parezca extraño, no todas las plantas vivientes
poseen verdaderas hojas, bien sea porque nunca las
tuvieron, o porque las perdieron secundariamente en el
transcurso de su historia natural. Un buen ejemplo de esta
situación se da en los cactus y otras plantas de ambientes
áridos, las cuales redujeron sus hojas a espinas y pasaron
la función fotosintética al tallo (Figura 8.1).
Por definición, los órganos vegetales poseen un
sistema vascular interno, lo que implica que sólo aquellas
plantas que presentan un sistema de conducción pueden
poseer verdaderos órganos como hojas, raíces y tallo. A
las plantas que poseen tejidos vasculares se les denomina
traqueófitas, mientras que los vegetales que carecen de
ellos son llamados en conjunto briófitas o plantas
avasculares. En ellas no hay hojas verdaderas, así que las
estructuras aplanadas de apariencia foliar son en realidad
filidios (Figura 8.2), que son láminas de células
encargadas de la fotosíntesis pero con una anatomía
completamente diferente a las hojas verdaderas.
Cuando el cuerpo vegetal posee órganos bien
diferenciados (raíz, tallo, hoja), se le denomina cormo, y
las plantas así constituidas son llamadas cormófitas. En
ellas, las hojas pueden ser clasificadas en dos tipos:
micrófilos y megáfilos, clasificación que depende,
precisamente, de su anatomía vascular. Los micrófilos se
presentan en las plantas más primitivas y aunque se creía
que eran de pequeño tamaño (de ahí su nombre), en
realidad pueden ser de más de un metro de longitud. Se
caracterizan porque presentan un solo haz vascular no
ramificado, que se deriva de la estela del tallo y penetra
la hoja sin dejar brecha foliar. Entre las plantas vivientes
los micrófilos sólo están presentes en algunas plantas
vasculares sin semillas (pteridófitas). Los megáfilos, de
otra parte, presentan brecha foliar y pueden tener varios
haces vasculares que se ramifican en la lámina foliar,
estando presentes en la gran mayoría de las plantas
actuales.
8.2. Los primordios foliares originan las hojas en las
yemas apicales de una manera ordenada.
En la base de los meristemos apicales, células situadas
bajo la protodermis empiezan a proliferar produciendo un
abultamiento cónico denominado primordio foliar
(Figura 8.3). Dicho primordio pronto alcanza una mayor
longitud que el meristemo apical y se encuentra
constituido de células meristemáticas diferenciadas en
protodermis y meristemo basal. En el centro del
primordio un grupo de células se diferencia en
procambio, tejido que originará xilema y floema
primarios. En las monocotiledóneas es particularmente
notable que el primordio foliar envuelve completamente
al meristemo, por lo tanto la futura hoja estará
envolviendo al tallo usando su base foliar (Figura 8.4). El
lugar exacto donde se forma el primordio foliar está
genéticamente determinado, es decir, no es al azar. Las
bases fisiológicas de este fenómeno no están bien
entendidas, pero lo cierto es que cada planta tiene una
manera constante de distribuir las hojas a lo largo de su
tallo, fenómeno que se conoce con el nombre de filotaxis.
La mejor explicación existente acerca de cómo se forman
los primordios foliares es la hipótesis de los campos
inhibitorios, que básicamente establece que un primordio
se desarrolla en el lugar donde no llegan los efectos
inhibitorios de los primordios ya formados.
Un par de conceptos importantes, relacionados
con el origen ordenado de los primordios foliares y con el
subsecuente desarrollo de la hoja, deben ser mencionados
aquí. El primero es el plastocrono, que se refiere al
tiempo transcurrido entre la formación de un primordio y

MAGG – Botánica General: Introducción al estudio de las plantas
159
el siguiente. Como puede intuirse, se trata de una
característica propia de cada especie. En las
angiospermas, debe formarse una yema axilar de la hoja,
lo cual no sucede antes de que el primordio foliar esté en
su segundo o tercer plastocrono. El otro tema relevante es
el de los meristemos foliares que permiten el crecimiento
marginal y, a veces, el crecimiento adaxial. En el margen
la hoja debe crecer expandiéndose lateralmente hasta
obtener la típica forma aplanada, lo cual logra debido a
meristemos marginales que se forman tras cierto tiempo
en los lados del primordio foliar. De otra parte, el grosor
de la vena media y del pecíolo pueden aumentar
considerablemente en algunas dicotiledóneas debido a un
meristemo adaxial adicional, el cual desarrolla una
estructura laminar aplanada lateralmente que es la
responsable del aumento en grosor de la hoja. Tanto en
Acacia como en Acorus se han realizado notables
estudios en este tema.
Se supone que la filotaxis ha sido optimizada
durante el proceso evolutivo, de tal forma que el
solapamiento foliar sea mínimo y por lo tanto las
posibilidades fotosintéticas de cada hoja sean máximas.
Dicha filotaxis sirve para clasificar a las hojas en tres
grupos básicos: 1) opuestas, cuando dos hojas se originan
en un mismo nudo pero en los lados contrarios del tallo,
2) alternas, cuando sólo hay una hoja por nudo y 3)
verticiladas, cuando hay más de dos hojas por nudo
(Figura 8.5).
Al observar un tallo desde arriba, se puede ver
que las hojas alternas forman un espiral si se sigue la
secuencia de hojas desde la más cercana al ápice superior
hasta la hoja más basal. Se apreciará que hay grupos de
hojas que están solapadas, es decir, que forman una línea
vertical en la cual las hojas superiores están ubicadas
exactamente sobre algunas de las hojas situadas más
abajo. Todas las hojas que forman esta línea vertical de
solapamiento constituyen un ortostiquio u ortóstico,
mientras que la secuencia en espiral se denomina
parastiquio o parástico. Mediante la serie matemática de
Fibonacci es posible descifrar la filotaxis, ya que de dos
números continuos de la serie, el primero representa el
número de hojas que hay entre una hoja cualquiera y su
siguiente en el ortostiquio, mientras que el segundo
número es la cantidad de vueltas que hay que darle al
tallo siguiendo la espiral de hojas (el parastiquio), para
encontrar dicha hoja solapada.
8.3. La vascularización de la hoja se deriva del
sistema vascular del tallo.
Las hebras de procambio derivadas del meristemo apical
crecen y se diferencian dentro del primordio foliar a
medida que éste madura, razón por la cual el sistema
vascular de la hoja es continuo con el del tallo. Lo que se
observa en ramas maduras, es que en cada nodo uno o
más haces vasculares divergen del cilindro vascular del
tallo y penetran a las hojas originadas de ese nudo. Este
haz que diverge se conoce como trazo foliar y la región
por encima de él, que se rellena de tejido basal
parenquimático e interrumpe el haz vascular del tallo, se
denomina brecha foliar. Esta característica está presente
sólo en los megáfilos (Figura 8.6B), que es el tipo de hoja
predominante en la actualidad, ya que en los micrófilos
sólo hay un haz vascular que penetra a la hoja desde el
tallo y no deja brecha foliar (Figura 8.6A). Es notable que
en un nudo individual de un megáfilo, varias hebras
vasculares pueden penetrar en la lámina foliar,
constituyendo así una hoja multilacunar pues cada trazo
posee su propia laguna o brecha foliar.
8.4. Hay una abundante variedad de formas y
tamaños foliares.
La hoja presenta una gran variedad de estilos, formas y
tamaños, que ha generado abundantes términos y formas
de clasificación. Aquí sólo se revisará la terminología

MAGG – Botánica General: Introducción al estudio de las plantas
160
básica, necesaria para comprender la naturaleza foliar
elemental, la cual es el sustento de todas las demás
variantes. Se puede empezar por el término nomófilo, que
se emplea para referirse a la hoja típica o normal, la cual
es un megáfilo cuyo prototipo es la hoja de las
angiospermas dicotiledóneas (Figura 8.7). Dicho
nomófilo está constituido típicamente de una parte
aplanada, de amplia superficie, llamada la lámina y un
cilindro sólido que conecta la lámina con el tallo, al cual
se denomina pecíolo. El pecíolo se prolonga en la lámina
foliar formando la vena media, mientras que en su
extremo opuesto se une a la rama que sostiene la hoja,
presentando en dicha unión unos pequeños apéndices
foliosos denominados estípulas. La parte de la lámina que
se une al pecíolo se denomina base foliar y el extremo
opuesto se llama ápice foliar. Todas estas partes pueden
estar ausentes o muy modificadas según la especie. La
superficie superior de la lámina se denomina haz y recibe
directamente la luz solar, la inferior se denomina el
envés.
Las hojas pueden ser simples o compuestas
según la estructura de su lámina (Figuras 8.7 y 8.8).
Cuando la lámina no presenta invaginaciones que
alcancen la vena media, se dice que es una hoja simple,
pero cuando las invaginaciones alcanzan la vena media,
dando a la lámina la apariencia de estar partida en
pequeñas subunidades que parecen hojas pequeñas, se
dice que es una hoja compuesta. Dichas subunidades se
denominan foliolos y están adheridas al raquis, o eje
central, por pequeños tallos llamados peciólulos. Es
posible que haya divisiones de segundo y tercer orden,
constituyendo hojas muy complejas. En algunas especies
las hojas compuestas tienen sus foliolos en una
disposición palmada, por lo que carecen de raquis. Las
hojas compuestas son paripinnadas si poseen un par de
foliolos en el ápice del raquis, o imparipinnadas si poseen
un número impar.
El borde de la lámina foliar, así como su base y
su ápice, presentan numerosas variaciones morfológicas
que sirven para clasificar las hojas (Figuras 8.9 - 8.12).
Adicionalmente a las variaciones locales mencionadas,
las hojas también pueden modificar notablemente su
forma total, como por ejemplo cuando forman zarcillos.
Éstos son órganos delgados y alargados que se pueden
enrollar alrededor de objetos sólidos, con el fin de
permitir un mejor posicionamiento del vástago con
respecto a la luz solar (Figura 8.13). Otras variaciones
notables pueden estar presentes en los pecíolos, como las
pequeñas prolongaciones laterales de la lámina, razón por
la cual se les denomina pecíolos alados. El raquis de las
hojas compuestas también puede presentar estas
prolongaciones y se denomina raquis alado. Incluso
puede presentar glándulas de néctar en el haz (Figura
8.14), que aparentemente funcionan atrayendo hormigas
hacia las hojas para evitar la visita de potenciales
herbívoros. Estas modificaciones son muy comunes en
las leguminosas del género Inga.
Algunas variaciones morfofuncionales de las
hojas son dignas de ser mencionadas (Figura 8.15). Por
ejemplo, los catáfilos son hojas inferiores, ubicadas entre
los cotiledones y los nomófilos, de forma y estructura
más sencillas, generalmente escuamiformes, sin clorofila,
membranosas o coriáceas. El ejemplo típico de ellos se
encuentra en los bulbos de la cebolla cabezona (Allium,
figura 8.15B). Los hipsófilos son hojas superiores
ubicadas entre los nomófilos y los órganos florales
(Figura 8.15A), también con morfoanatomía más
sencilla. Algunos ejemplos son las espatas, brácteas,
bracteolas, estípulas, involucros y escamas, llamadas en
conjunto hojas espúreas por algunos autores. Los
antófilos, de otra parte, son las hojas florales, es decir, las
hojas modificadas que conforman los verticilos de la flor
completa y que sirven a la función reproductiva. Los más
externos son los sépalos y los pétalos, que conforman el
cáliz y la corola, respectivamente. Se trata de antófilos

MAGG – Botánica General: Introducción al estudio de las plantas
161
estériles, que atraen polinizadores o protegen las
estructuras reproductivas más internas, por lo que su
contribución a la reproducción es indirecta. Por su parte,
los antófilos más internos están muy modificados y
conforman los estambres y el pistilo, que en conjunto son
denominados a veces androceo y gineceo, y son antófilos
fértiles.
Otra sorprendente adaptación foliar se da en las
plantas carnívoras (algunos autores prefieren llamarlas
insectívoras), las cuales poseen hojas muy modificadas
(Figura 8.13B), para atrapar principalmente insectos, si
bien es posible que pequeños vertebrados mueran en su
interior (de ahí lo de carnívoras). La descomposición de
los animales allí atrapados provee sustancias nutritivas
para la planta, ya que generalmente están presentes en
especies vegetales que viven en suelos con deficiencias
en nutrientes.
8.5. La anatomía foliar es muy variada.
Los tres tipos de sistemas de tejidos, basal, vascular y
dérmico, están presentes en prácticamente todas las
hojas, pero su disposición anatómica puede variar entre
grupos de plantas. En las angiospermas, por ejemplo,
sigue básicamente dos patrones: el de las plantas
dicotiledóneas y el de las monocotiledóneas. En las hojas
de las dicotiledóneas las venas adoptan un patrón en red
en la hoja, con venas mayores y un sistema de
interconexión constituido por venas menores (Figura
8.7), al cual se denomina venación reticulada. Por su
parte, las monocotiledóneas poseen un patrón de
venación en paralelo, donde las venas se extienden
longitudinalmente desde la base hacia el ápice de la hoja
(Figura 8.16B). La única excepción entre las
monocotiledóneas la constituye el grupo de las aráceas
(Figura 8.16C), familia que posee el mismo patrón
reticulado de las dicotiledóneas. Estas diferencias
anatómicas de las hojas, junto a otros caracteres, eran
usados para sustentar la clasificación de las angiospermas
en dos grandes subgrupos, magnoliópsidas y liliópsidas.
Actualmente, sin embargo, no es posible sostener dicha
clasificación ya que los descubrimientos sistemáticos
recientes no la respaldan. A pesar de ello, los términos
monocotiledóneas y dicotiledóneas siguen siendo usados
de manera informal con fines didácticos.
Es pertinente mencionar aquí que la anatomía no
es el único criterio usado para clasificar las hojas, ya que
también se les clasifica con base en aspectos
ecofisiológicos, según los cuales puede haber hojas
hidrófilas, mesófilas y xerófilas, según las adaptaciones
que presenten a ambientes ricos en agua, mésicos o
áridos, respectivamente. En este tipo de clasificación
también se puede detectar el conjunto de características
anatómicas particulares de cada tipo foliar. A
continuación se revisará cada uno de los tres elementos
básicos que constituyen las hojas de las plantas
modernas: la epidermis, el mesófilo y el sistema vascular
(Figuras 8.17 - 8.19).
La epidermis de la hoja está compuesta por un
grupo de células compactamente agrupadas, que carecen
de cloroplastos y poseen una capa gruesa de compuestos
lipídicos denominada cutícula, la cual se deposita en la
pared celular que está en contacto directo con el ambiente
externo y ayuda a prevenir la deshidratación. Tanto la
superficie superior (haz) como la inferior (envés), poseen
una epidermis, en la cual hay varios tipos celulares
especializados. En la epidermis de las hojas de los pastos,
por ejemplo, están las denominadas células buliformes
(Figura 5.17B), que son mucho más grandes que las
células epidérmicas comunes y se arreglan en filas
continuas longitudinales. Ellas están involucradas en el
plegamiento de las hojas, generalmente en respuesta a
cambios en la de disponibilidad de agua.

MAGG – Botánica General: Introducción al estudio de las plantas
162
Otro grupo de células epidermales muy
especializadas son las células guardas, que se disponen
en pares permitiendo la aparición de un espacio entre
ellas denominado poro estomático (Figuras 8.20 y 8.21).
Estas células y el espacio entre ellas son denominados en
conjunto estoma o aparato estomático, el cual funciona
regulando el aumento o la disminución del poro,
mediante los cambios de turgencia de las células guardas,
en un proceso complejo en el que intervienen los iones
K+ y Cl- (Sección 12.7). Justo por debajo del estoma
existe una cavidad formada por células del mesófilo
denominada cámara subestomática, en la cual hay una
elevada concentración de vapor de agua y a donde entra
el CO2 cuando se abre el poro.
El control estomático es de vital importancia
para la planta, ya que a través de esta estructura es que se
realiza la incorporación del CO2 atmosférico que se usará
en la fotosíntesis. Los estomas generalmente se ubican en
el envés, donde la temperatura es menor debido a que no
hay radiación solar directa, por lo cual se disminuye la
pérdida de agua cuando se abren para captar CO2. En las
plantas acuáticas, sin embargo, donde las hojas flotan en
la superficie del agua, los estomas se hallan en el haz
para estar en contacto directo con el aire. Una
modificación notable de la superficie foliar de algunas
plantas que viven en zonas áridas, son los estomas
hundidos, adaptación que consiste en la ubicación de
éstos en cavidades de la superficie de la hoja para evitar
que el vapor de agua se pierda rápidamente luego de la
transpiración. Asociadas a las células guardas del estoma
están las células subsidiarias, las cuales se usan para
clasificar los diferentes aparatos estomáticos.
Otra característica notable de la epidermis es la
presencia de sobrecrecimientos que le confieren una
textura y apariencia particulares a cada especie. El
término indumento hace referencia justo a dicha
apariencia general de la hoja (aunque también se aplica a
las ramas y al tallo), habiéndose desarrollado una
terminología extensa, no exenta de subjetividad, para
describir la forma en que luce externamente. Papilas,
escamas y tricomas son las estructuras más importantes
para el indumento superficial, las cuales ayudan a la
defensa foliar, la disipación de la radiación solar
excesiva, la secreción de sustancias y el ahorro de agua.
Todos ellos pueden ser tanto unicelulares como
pluricelulares, con variadas formas y tamaños,
particularmente los tricomas (Figura 8.22), lo que los
hace útiles en estudios taxonómicos. Las formas más
simples de sobrecrecimientos son las papilas, seguidas de
los tricomas simples, de los cuales son a veces
difícilmente distinguibles. Los tricomas ramificados son
un poco más complejos (poseen 2 a 5 brazos), y son
seguidos por los tricomas estelados, las escamas, los
tricomas dendríticos (muchos brazos) y las estructuras
especiales, tales como las glandulares. Los elementos
más complejos del indumento pueden ser sésiles o tener
un tallo que las eleva sobre la superficie epidermal.
El mesófilo corresponde al cuerpo propiamente
dicho de la hoja, es decir, todo lo encontrado entre las
dos epidermis. Allí se encuentran el parénquima
asimilador (clorénquima) y el sistema conductor (venas)
(Figuras 8.17 y 8.18). En las plantas dicotiledóneas la
función fotosintética está distribuida entre dos tipos de
clorénquima, el parénquima en empalizada y el
esponjoso. El parénquima en empalizada posee células de
forma columnar, abundantes cloroplastos y puede
presentar granos que almacenan almidón debido al
exceso de productos de la fotosíntesis (fotosintatos),
mientras que el parénquima esponjoso tiene abundantes
espacios intercelulares que se conectan con la cámara
subestomática, a la cual llega el CO2 necesario para la
fotosíntesis.
Inmersos en el mesófilo están los haces
vasculares, generalmente compuestos por xilema hacia

MAGG – Botánica General: Introducción al estudio de las plantas
163
arriba y floema hacia abajo. Los haces vasculares están
cubiertos por una capa especial de células de mesófilo
denominadas en conjunto la envoltura (o vaina) del haz,
que impide el contacto directo de los tejidos vasculares
con el aire de los espacios intercelulares del parénquima
esponjoso. Funciona de manera similar a la endodermis
de la raíz, ya que todo lo que entre o salga del haz
vascular debe atravesar esta capa de células.
En plantas monocotiledóneas, particularmente
en especies con fotosíntesis C4 (ej: caña de azúcar, maíz,
sorgo), la vaina del haz es mucho más conspicua que en
las C3, pues se encuentra rodeada por células del
mesófilo que ayudan a darle una apariencia redondeada,
notable en el corte transversal, a la cual se denomina
anatomía Kranz (Figura 8.18). A lo largo de los haces
vasculares, o aun en los bordes de las hojas, hay además
otra característica importante, como es la presencia de
tejidos de sostén (fibras de esclerénquima), que ayudan a
dar compactación y resistencia a la hoja. Las fibras
asociadas a las venas pueden formar un casquete al lado
del floema o también pueden formar un anillo que rodea
completamente al haz vascular.
8.6. La vida de la hoja es de duración limitada.
El término abscisión se usa para describir la separación
normal de la hoja (o de cualquier otro órgano) del cuerpo
vegetal, como consecuencia de cambios estructurales y
bioquímicos inducidos por la edad. En las hojas estos
cambios están bajo la influencia directa de la actividad
fitohormonal y crean, en la base del pecíolo, la
denominada capa de abscisión (Figura 8.23). En ella
ocurre un debilitamiento de las lamelas medias de las
paredes celulares, lo que conlleva a una pérdida de
cohesión intercelular que produce la caída de la hoja por
su propio peso. Subsecuentemente se creará una cicatriz
foliar en el lugar donde estaba adherida la hoja a la rama,
en la que se desarrolla una capa protectora con depósitos
de suberina.
El momento de la abscisión depende de la
longevidad de la hoja (Figura 8.24), la cual varía entre
especies y puede ser corta en especies que viven en zonas
con cambios climáticos bruscos (regiones templadas con
estaciones), o aun en plantas tropicales que tienen hábitos
caducifolios (como algunas especies de la familia
Bombacaceae). En algunas especies, sin embargo, la
longevidad foliar es larga, incluso de varios años, como
en algunas cícadas tropicales. Si bien la base molecular
de la caída foliar no es bien entendida, se sabe que hay
factores ambientales y fisiológicos que pueden afectar el
momento de inicio de la abscisión, lo cual debe estar muy
bien regulado ya que se requiere desencadenar eventos de
muerte celular programada y de reabsorción de nutrientes
de la hoja antes que se desprenda de la rama.
BIBLIOGRAFÍA
Clark DB, Clark DA & Grayum MH. 1992. Leaf
demography of a neotropical rain forest cycad, Zamia
skinneri (Zamiaceae). American Journal of Botany 79
(1): 28-33.
Cutter EG. 1971. Plant anatomy. Experiment and
interpretation. Part 2 Organs. Edward Arnold Ltd.,
Londres, Inglaterra.
Esau K. 1976. Anatomía vegetal. Ediciones Omega,
S.A., Barcelona, España.
González F. 1999. Monocotiledóneas y dicotiledóneas:
un sistema de clasificación que acaba con el siglo.
Revista de la Academia Colombiana de Ciencias 23: 195-
204.

MAGG – Botánica General: Introducción al estudio de las plantas
164
Metcalfe CR & Chalk L. 1979. Anatomy of the
dicotyledons. Volume I: Systematic anatomy of leaf and
stem, with a brief history of the subject. 2 ed. Oxford
University Press, Oxford, Inglaterra.
Mora-Osejo LE. 2004. Morfología, sistemática y
evolución de las Angiospermae. Universidad Nacional de
Colombia, Facultad de Ciencias, Bogotá, DC, Colombia.
Raven PH, Evert RF & Curtis H. 1981. Biology of
plants, 3 ed. Worth Publishers, Inc. Nueva York, E.U.A.
Yoshida S. 2003. Molecular regulation of leaf
senescence. Current Opinion in Plant Biology 6: 79-84.
Figura de la portada del capítulo

MAGG – Botánica General: Introducción al estudio de las plantas
165
Figura 8.1. Tallo de un cactus. Algunas plantas modernas perdieron las hojas y pasaron la función fotosintética al tallo, lo
cual se ha interpretado como una respuesta adaptativa a ambientes secos, donde una amplia superficie foliar favorece la
pérdida de agua, lo cual es desventajoso.
Figura 8.2. Estructuras foliosas de una planta briófita (Pleurozium schereberi). A) Las estructuras aplanadas, conformadas
a veces por una sola capa de células, no son hojas verdaderas aunque poseen la función fotosintética, son filidios, B) en el
recuadro se muestra un corte transversal de un filidio, mostrando su parte media (a) y su borde (b). (Fotos cortesía de A.
Barón).
MAGG – Botánica General: Introducción al estudio de las plantas
166
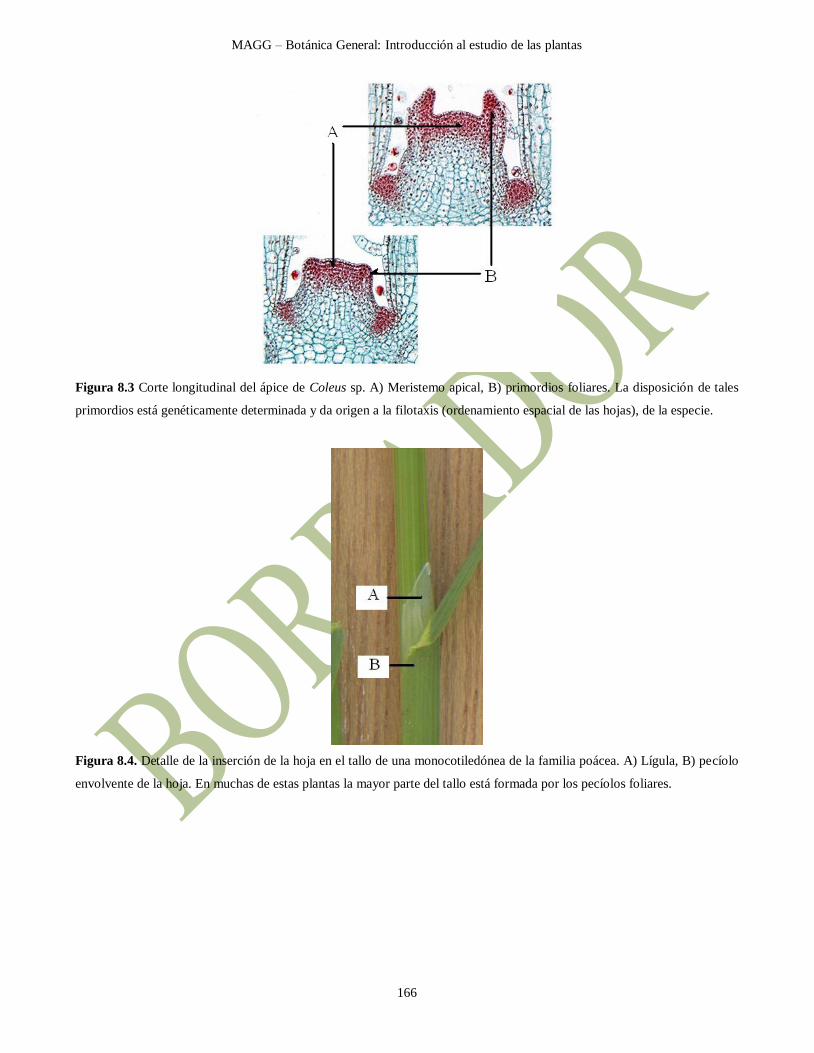
Figura 8.3 Corte longitudinal del ápice de Coleus sp. A) Meristemo apical, B) primordios foliares. La disposición de tales
primordios está genéticamente determinada y da origen a la filotaxis (ordenamiento espacial de las hojas), de la especie.
Figura 8.4. Detalle de la inserción de la hoja en el tallo de una monocotiledónea de la familia poácea. A) Lígula, B) pecíolo
envolvente de la hoja. En muchas de estas plantas la mayor parte del tallo está formada por los pecíolos foliares.

MAGG – Botánica General: Introducción al estudio de las plantas
167
Figura 8.5. Filotaxis básica en las angiospermas. A) Hojas simples alternas en una hierba trepadora (Bomarea sp.), a1 y a2
son hojas maduras que se insertan a diferente altura en el mismo tallo, B) hojas simples opuestas en el cafeto, b1 es opuesta
a b2, C) hojas simples opuestas decusadas, lo cual es consecuencia que el plano en que c1 y c2 son opuestas, es
perpendicular al plano de oposición de c3 y c4, D) hojas verticiladas, d1, d2, d3 y d4 son hojas maduras que nacen a la
misma altura en el mismo tallo.
Figura 8.6. Representación esquemática de la venación en tallo y hoja. A) Micrófilo con un solo haz vascular que no deja
brecha foliar y entra a la hoja, B) megáfilo con brecha foliar y haces ramificados en la lámina.

MAGG – Botánica General: Introducción al estudio de las plantas
168
Figura 8.7. Detalle de la parte terminal de la rama de una morácea silvestre. A) Lámina o limbo de la hoja, se notan las
venas secundarias y terciarias interconectadas formando un patrón reticulado, B) pecíolos foliares, C) yema apical, en la
cual se desarrollan las nuevas hojas.
Figura 8.8. Varios tipos de hojas de angiospermas. A) Hojas simples alternas, nótese que la inserción de los pecíolos se da
en lugares diferentes alrededor de la rama, B) hoja compuesta imparipinnada, C) hoja palmadamente compuesta, en la cual
los peciólulos de los foliolos se insertan en un mismo lugar del pecíolo.

169
Figura 8.9. Esquema de los principales tipos de hojas simples (A-L) y compuestas (M-R). A) Cordada, B) cuneada, C)
deltoide, D) elíptica, E) perfoliada, F) lanceolada, G) subulada, H) peltada, I) oblanceolada, J) elíptica, K) obovada, L)
ovada, M) bifoliada, N) trifoliada, O) imparipinnada, P) paripinnada, Q) bipinnada, R) palmaticompuesta.

170
Figura 8.10. Bordes foliares en las angiospermas. A) Borde entero, B) borde dentado, C) borde lobulado, D) margen con
profundas entradas a manera de lóbulos. Hay una gran diversidad de variaciones en el borde foliar, las cuales son útiles en
estudios taxonómicos y morfológicos.
Figura 8.11. Esquema de los principales tipos de borde y venación foliares. A) Borde entero, B) borde dentado, C) borde
serrado, D) borde crenado, E) borde ciliado, F) borde sinuado, G) borde hendido, H) borde partido, I) venación dicótoma, J)
venación palmadamente reticulada, K) venación pinnadamente reticulada, L) venación paralela.

171
Figura 8.12. Representación esquemática de los principales tipos de ápices (A-I) y bases (J-Q) foliares. A) Ápice
acuminado, B) agudo, C) aristado, D) cuspidado, E) emarginado, F) mucronado, G) obtuso, H), redondeado, I) truncado. J)
Base auriculada, K) cordada, L) cuneada, M) hastada, N) oblicua, O) redondeada, P) sagitada, Q) truncada.
Figura 8.13. Hoja modificada de Nepenthes sp. A) Zarcillos foliares con los cuales la planta se enrolla alrededor de objetos
sólidos, B) ápice de la hoja modificado para crear una estructura hueca en la que se deposita agua y se descomponen
pequeños animales.

172
Figura 8.14. Fragmento de una hoja compuesta de Inga sp. A) Glándulas sobre el raquis de la hoja, los cuales atraen
hormigas que protegen la planta, B) raquis alado entre los foliolos, compuesto de una lámina que posee tejido fotosintético y
vascular.
Figura 8.15. Hojas modificadas en angiospermas. A) Parte superior de una acantácea del género Justicia, en la que se
muestran las hojas normales abajo (nomófilos), además de las brácteas en la base de las flores que corresponden a hipsófilos
(a), así como las flores cuyos pétalos son antófilos (b). B) Corte transversal de una cebolla común (Allium sp.), en la que se
muestra el conjunto de hojas modificadas para el almacenamiento (catáfilos).

173
Figura 8.16. Hojas con diferente patrón de venación. A) Típica hoja de planta dicotiledónea, en las cuales existe un patrón
reticulado de venas, B) fragmento de una hoja de monocotiledónea (orquídea), en la que se aprecia la dirección longitudinal
de las venas. Este tipo de venación está presente en casi todas las monocotiledóneas, con una sola excepción notable, la
familia Araceae (C), en la cual las venas están dispuestas reticuladamente.
Figura 8.17. Corte transversal de la hoja de una dicotiledónea de la familia melastomatácea. A) Epidermis foliar, B)
mesófilo, C) haz vascular de la vena media, D) parénquima en empalizada (células columnares), E) parénquima esponjoso.

174
Figura 8.18. Corte transversal de la hoja de una monocotiledónea (maíz), mostrando anatomía Kranz. A) Cutícula, B)
células de la epidermis, C) célula gigante de la vaina del haz con cloroplastos muy grandes, D) xilema, E) floema F)
parénquima del mesófilo. Foto cortesía de X. Marquínez.
Figura 8.19. Corte transversal de la hoja acicular de un pino. A) Epidermis monostratificada, B) hipodermis, C) estoma
hundido, D) endodermis, E) parénquima de transferencia, F) ducto de resina. Un haz vascular se encuentra en un círculo.

175
Figura 8.20. Estomas en la hoja de Arabidopsis, mostrados mediante una fotografía electrónica de barrido. A) Células
guardas del estoma, B) poro estomático. Foto cortesía del Dartmouth Electron Microscope Facility, Dartmouth College.
Figura 8.21. Representación esquemática del estoma. A) Estoma anomocítico, en el que las células guardas se diferencian
claramente de las otras células de la epidermis; no hay células subsidiarias, B) estoma tetracítico con cuatro células
subsidiarias, dos de ellas paralelas al estoma, C) estoma anisocítico, rodeado por tres subsidiarias desiguales, D) estoma
ciclocítico, en el que las subsidiarias forman anillos alrededor de las células guardas, E) estoma diacítico, con dos
subsidiarias cuyas paredes limitantes son perpendiculares al estoma, F) estoma paracítico, en el que las subsidiarias son
paralelas al eje mayor del estoma. G) Estoma visto desde arriba, las células guardas tapan parcialmente a las subsidiarias y
la cutícula forma un reborde externo, mejor visto en el corte longitudinal (H): h1) cámara subestomática o cavidad del
estoma, h2) poro, h3) reborde de la cutícula que puede formar una cavidad externa.

176
Figura 8.22. Representación esquemática de los principales tipos de tricomas. A) Tricoma estelado con tallo pluricelular,
B) tricoma ramificado de dos brazos, C) escama en vista lateral, D) escama en vista superior, E) tricoma simple con tallo, F)
tricoma simple unicelular, G) tricoma ramificado de tres brazos, H e I) tricomas simples, J) tricoma estelado, K) tricoma
dendrítico.

177
Figura 8.23. Vista al microscopio de un corte longitudinal por la capa de abscisión foliar. A) Tejido de la rama a la que está
adherida la hoja, B) capa de abscisión, donde las células experimentan el debilitamiento de su lamela media, C) pecíolo de
la hoja.
Figura 8.24. Plantas con hojas de variada duración. A) En algunas plantas tropicales como la ceiba (Bombacaceae), las
hojas son cambiadas periódicamente, lo cual sucede antes de la floración, B) en otras especies, como las cícadas, las hojas
son mucho más longevas, pudiendo durar incluso varios años.
Top Related