







Languages
Pages
Legal


Caffeine modulates post-transcriptional regulation of SRSF2
1
Methylxanthines Increase Expression of the Splicing Factor SRSF2 by Regulating Multiple
Post-Transcriptional Mechanisms
Jia Shi, Kirk Pabon, Kathleen W. Scotto1
From the Rutgers Cancer Institute of New Jersey,
the Robert Wood Johnson Medical School and the Graduate School of Biomedical Sciences
Rutgers, The State University of New Jersey, New Brunswick, NJ 08903
*Running title: Caffeine modulates post-transcriptional regulation of SRSF2
To whom correspondence should be addressed: Kathleen W. Scotto. Rutgers Cancer Institute of New
Jersey, Rutgers, the State University of New Jersey, 195 Little Albany Street, New Brunswick, NJ 08903,
USA, Tel: (732) 235-4266; Fax: (732)-235-6596; Email: [email protected]
Keywords: SRSF2, alternative splicing, microRNA, caffeine, methylxanthines, translational efficiency,
nonsense-mediated decay, homeostasis
Background: Caffeine regulates alternative
splicing by increasing SRSF2, normally
constrained by a negative feedback loop.
Results: Caffeine blocks nonsense-mediated
decay, induces 3’UTR alternative splicing, and
down-regulates SRSF2-targeting microRNAs,
thereby breaking the negative feedback loop to
increase SRSF2.
Conclusion: Caffeine modulates multiple post-
transcriptional processes to increase SRSF2.
Significance: This study expands our
understanding of SRSF2 regulation, and may
provide insight into SRSF2 dysregulation in
disease.
ABSTRACT
We have previously reported that the
methylxanthine caffeine increases expression
of the splicing factor SRSF2, the levels of
which are normally controlled by a negative
auto-regulatory loop. In the present study we
have investigated the mechanism by which
methylxanthines induce this aberrant
overexpression. RT-PCR analyses suggested
little impact of caffeine on SRSF2 total
mRNA levels. Instead, caffeine induced
changes in the levels of SRSF2 3ʹUTR splice
variants. While some of these variant were
substrates for nonsense-medicated decay
(NMD), and could potentially have been
stabilized by caffeine-mediated inhibition of
NMD, downregulation of NMD by a genetic
approach was not sufficient to reproduce the
phenotype. Furthermore, cell-based assays
demonstrated that some of the caffeine-
induced variants were intrinsically more
efficiently translated than others; the
addition of caffeine increased the
translational efficiency of most SRSF2
transcripts. MicroRNA array analyses
revealed a significant caffeine-mediated
decrease in the expression of two SRSF2-
targeting miRs, both of which were shown to
repress translation of specific SRSF2 splice
variants. These data support a complex
model whereby caffeine downregulates
SRSF2-targeting microRNAs, leading to an
increase in SRSF2 translation, which in turn
induces SRSF2 splicing. SRSF2 splice
variants are then stabilized by caffeine-
mediated NMD inhibition, breaking the
normal negative feedback loop and allowing
the aberrant increase in SRSF2 protein levels.
These findings highlight the complexity of
SRSF2 gene regulation, and suggest ways in
which SRSF2 expression may be dysregulated
in disease.
Alternative splicing (AS) of pre-mRNA is a
fundamental cellular process that selectively
joins alternate exons together to produce
different mRNA variants from a single gene.
More than 90% of human genes undergo
alternative splicing, resulting in strikingly high
levels of mRNA complexity (1-3). In some
cases, AS leads to the selective inclusion or
http://www.jbc.org/cgi/doi/10.1074/jbc.M114.624254The latest version is at JBC Papers in Press. Published on March 28, 2015 as Manuscript M114.624254
Copyright 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Caffeine modulates post-transcriptional regulation of SRSF2
2
skipping of one or more protein-coding exons,
resulting in functionally different protein
isoforms with altered domains; this contributes
significantly to proteomic diversity. In other
cases, untranslated regions (UTRs) are altered or
premature termination codons (PTCs) are
introduced by AS, thereby generating complex
mRNA populations that are substrates for post-
transcriptional regulation at the level of
translational efficiency, mRNA localization
and/or mRNA stability (4-7).
The potential for AS to exert a complex and
profound effect on gene expression requires that
tightly controlled regulatory processes are in
place. This control is primarily executed by
positively- or negatively-acting splicing factors
that bind to cis-elements within pre-mRNA near
the regulated splice sites, modulating their
recognition efficiency by the multi-protein
spliceosome (8). Two major families of splicing
factors have been characterized: the
serine/arginine-rich (SR) protein family and the
HnRNP protein family, while additional RNA-
binding proteins have also been identified
(9,10). Splicing factors work antagonistically
such that their relative concentration in a given
cell is a critical determinant of the fate of
targeted exons (11,12). Accordingly, under
steady-state conditions the relative levels of
splicing factors are expected to remain constant,
while intrinsic (i.e. during differentiation and
development) or extrinsic (i.e. environmental
cues, stressor) signals can alter this ratio, thereby
altering AS choices and, potentially, cell fate.
An additional layer of control is exerted by
nonsense-mediated decay (NMD), an RNA
surveillance system that ensures the fidelity of
gene expression by degrading non-productive
mRNAs containing premature termination
codons (PTCs)(13), including those produced by
AS. Recent genome-scale studies revealed that
many splicing regulators employ a dual
mechanism, alternative splicing coupled with
nonsense-mediated decay (AS-NMD), to limit
their own expression and prevent excessive
accumulation that can be deleterious to the cell
(14,15).
SRSF2 (SC35) is a ubiquitous splicing factor
that plays a critical role in both constitutive and
alternative splicing (16,17), most often
functioning as an activator to enhance the
recognition of particular splice sites. Studies
using SRSF2 conditional knockout mice and
SRSF2-/-
MEFs have suggest that this protein is
essential for cell proliferation and the
maintenance of genomic stability, at least during
thymus and pituitary development (18). It was
also suggested that SRSF2 plays a role in the
regulation of transcription elongation (19,20).
Additionally, transcription of SRSF2 has been
shown to be regulated by E2F1, and is required
for E2F1-induced apoptosis (21). Alkylating
agent-induced apoptosis was shown to be
accompanied by a significant increase in both
E2F1 and SRSF2, suggesting a possible role for
SRSF2 in genotoxic stress response (21).
Interestingly, a recent study suggested that
SRSF2 is the most enriched splicing factor in
human pluripotent stem cells, where SRSF2 is
regulated by OCT4 and required for
pluripotency via its ability to regulate the AS of
MBD2, the methyl-CpG binding protein (22).
Given the key role of SRSF2 in multiple
regulatory pathways, its dysregulation is likely
to be pathological; indeed, several studies have
identified a strong correlation between SRSF2
mutations and subclassification and
prognostication of myelodysplastic syndrome
(MDS), a heterogeneous group of myeloid
neoplasms that predispose to acute myeloid
leukemia (23-26).
SRSF2 levels are carefully controlled. In
addition to transcriptional regulatory controls,
SRSF2 RNA is subjected to negative auto-
regulation via AS-NMD (27). In this scenario,
when levels of SRSF2 protein exceed the
tolerable cellular threshold, SRSF2 induces
alternative splicing of its own transcript,
resulting in the expression of two novel PTC-
containing splice variants at the expense of the
primary SRSF2 transcript. These variants are
destined for degradation by NMD; through this
feedback loop, overall SRSF2 mRNA levels are
reduced and SRSF2 protein expression returns to
steady state levels (27). Although a recent
study suggested that both HnRNP H and
TARDBP (TDP-43) act as antagonists to SRSF2
in regulating a retained intron in this
unproductive alternative splicing at the 3’UTR
(28), the complexity of SRSF2 mRNA variant
formation, its regulation and the functional
implications are poorly understood.
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Caffeine modulates post-transcriptional regulation of SRSF2
3
We have previously shown that caffeine can
alter the AS of a subset of genes associated with
the cancer phenotype. This is accomplished, at
least in part, by the induction of high levels of
SRSF2 (29). In the present report we extend our
analyses by addressing the mechanism(s) that
lead to increased levels of SRSF2 protein. We
now show that this caffeine-mediated SRSF2
increase involves a complex series of events that
includes down-regulation of SRSF2-targeting
microRNAs, inhibition of NMD, and induction
of novel SRSF2 splice variants with altered
translational efficiency. As a consequence, the
SRSF2 negative feedback loop is broken,
allowing for sustained increase of SRSF2
protein in the cell. This study advances our
understanding of SRSF2 gene regulation,
highlighting the multifaceted mechanisms at the
post-transcriptional level that regulate SRSF2-
mediated AS decisions, and provides a basis to
investigate dysfunction of AS during
pathogenesis.
EXPERIMENTAL PROCEDURES
Cell culture and chemical reagents -- The
human cervical carcinoma HeLa cells line
(ATCC®, CCL-2) was maintained in DMEM
supplemented with 10% (v/v) FBS and 2.0 mM
glutamine. Caffeine and cycloheximide were
purchased from Sigma and prepared
immediately prior to use.
Cytotoxicity Assay – HeLa cells were seeded
in 96-well plates at a density of 7,500 cells per
well. Various concentrations (1.75 ~21 mM) of
xanthine derivatives, including caffeine,
pentoxifylline, theophylline, and isocaffeine,
were added to wells with 8 repeats for each
experimental condition. MTT assays were
performed three days after treatment following
manufacturer’s instructions (Promega).
Semi-quantitative RT-PCR Assays -- Total
RNA was prepared with Trizol reagent
(Invitrogen) and analysed using SS One-step
RT-PCR reagents (Invitrogen) according to the
manufacturer’s recommendations. RNAse-free
DNAse I treatment was performed when needed.
The initial RNA input was 300 ng, and the
number of amplification cycles was pre-
determined to ensure a linear range
amplification of targeted transcripts (28 cycles
to assay total SRSF2 mRNA levels, 40 cycles to
assay individual transcript groups including GA,
GB, GC, and GD. Primers for SRSF2 total
mRNA: a: 5’-CTG AGG ACG CTA TGG ATG
CCA-3’ ; b: 5’-GAC TTG GAC TTG GAC CTT
CGT-3’; Primers for SRSF2 3’UTRs: c: 5’-CCA
AGT CTC CTG AAG AGG AAG G-3’; d: 5’-
CTG AGA AAA GCT AAC ACC AAG-3’; e:
5’-GAA AAT GGT AAT GTC TGG GAA TC-
3’; f: 5’-GTC AGG AGG CCA CAA ATT
AGG-3’.
Real-time quantitative RT-PCR -- Purified
RNA was first treated with RNAse-free DNase I
and then reverse-transcribed into cDNA using
the High-Capacity cDNA Reverse Transcription
Kit (AB Applied Biosystems/Life Technologies)
following manufacturer’s instructions. Real-
time PCR was performed using SYBR® Green
PCR Master Mix (AB Applied Biosystems/Life
Technologies) and individually designed primer
sets. For total SRSF2 mRNA, sense: 5’-CTG
AGG ACG CTA TGG ATG CCA-3’; antisense:
5’-GAC TTG GAC TTG GAC CTT CGT-3’; for
β2M, the same as previously reported (29); for
Flag-SRSF2, sense: 5’-CGA CTC ACT ATA
GGG AGA CC-3’, antisense: 5’-GAG GTG
CGG TAG GTC AGG TT-3’; for GFP, sense:
5’- GGG TGA AGG TGA TGC AAC ATA C-
3’, antisense: 5’- CTC GCA AAG CAT TGA
ACA CCA-3’. The final Ct value was an
average of 3 repeats of each assay, repeated
three times. The relative amount of mRNA in
samples was determined using the 2-ΔΔCt
method.
RNA Interference Assay -- 3 X 104 HeLa
cells in 0.5-mL DMEM without antibiotics were
seeded into each well of a 24-well plate and
incubated overnight. 100 nM siRNA
(SiGENOME SMARTpool reagent,
DHARMACON) was mixed with
Oligofectamine reagent (Invitrogen), incubated
at room temperature for 20 min and added drop-
wise to cells. 72 hours after siRNA transfection,
cells were harvested for either RNA isolation or
whole cell lysates, and assayed by either RT-
PCR or Western blot analysis.
Western Blot Analysis -- Cells were washed
twice with cold PBS and lysed in RIPA buffer
(50 mM Tris- HCl, pH 7.4, 150 mM NaCl, 1.0%
Triton X-100, 0.1% SDS, 1% deoxycholate plus
protease inhibitor cocktail (Roche Diagnostics),
1 mM sodium fluoride, 1 mM sodium
orthovanadate, and 100 mg/ml
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Caffeine modulates post-transcriptional regulation of SRSF2
4
phenylmethylsulfonyl fluoride. The protein
concentration of cell lysates was determined
using the Pierce BCA protein assay. Equal
amounts of total protein (15-25 ug) were
analyzed by 10% SDS-PAGE followed by
immunoblotting using the following antibodies:
rabbit anti-TARBP (GeneTex, 1:1000), mouse
anti-HnRNP F/H (Abcam, 1: 1000), goat anti-
hUpf1 (Abcam, 1:1000), mouse anti- SRSF2
(provided by Drs Cyril Bourgois and James
Stevenin, 1:20), mouse anti-Flag (Agilent,
1:2000) or mouse anti-tubulin (Santa Cruz
Biotechnology, 1:1000). The secondary
antibody was either HRP-conjugated donkey
anti-goat IgG (Santa Cruz Biotechnology,
1:2500), HRP-conjugated goat-anti-rabbit IgG
(Santa Cruz Biotechnology, 1:2500), or HRP-
conjugated goat-anti-mouse IgG (Upstate
Technology, 1:2500). Immunoreactive bands
were visualized using the Visualizer™ Western
Blot Detection Kit (Upstate Technology)
following manufacturer’s directions.
SRSF2 transcript expression vectors --
SRSF2 cDNAs with variations in their 3’UTR
were amplified by RT-PCR as described in
Results and Figures using primers with
restriction enzyme linker EcoR I (attached to
primers c and e) and Xba I (attached to primers
d and f). Each of the RT-PCR products were
gel-purified and digested with EcoR I and Xba I,
then individually cloned into pcDNA3.1-Flag-
SRSF2 downstream of the SRSF2 coding
region. Resulting constructs (pcDNA3.1-Flag-
SRSF2-3’UTRs: A1, A2, A3, D1, D2, D3, etc.)
were verified by sequencing.
In vitro translation assay -- Equal amounts
(125 ng) of each SRSF2 splice variant construct
(A1,A2, A3, D1, D2, or D3) was assayed in the
presence of EasyTag™ L-[35
S]-Methionine
(PerkinElmer) using the TNT® Quick Coupled
Transcription/translation System (Promega).
Translated protein was analyzed by SDS-PAGE
followed by radiography using the Bio-Rad
phosphor Imager. Signals were quantified using
Bio-Rad Quantity One software.
Cell-based translational assa -- HeLa cells
were seeded at 1.25 X 105 cells per well in 12-
well plates. The next day 200 ng plasmid DNA
of each SRSF2 variant construct was
individually co-transfected with 37.5 ng plasmid
DNA of GFP construct (pN3-GFP) in duplicate
using Lipofectamine 2000 (Invitrogen). Seven
hours post- transfection, cells were untreated or
treated with caffeine; 3 hrs later, cells were
collected from each well and divided in half.
One half was dissolved in RIPA buffer for
western blot analysis using anti-Flag (Agilent,
1:2000) and anti-GFP antibodies (Santa Cruz
Technologies, 1:500) in a single procedure. The
other half was subjected to RNA purification
using trizol (Invitrogen) for real-time RT-PCR
analyses of Flag-SRSF2 and GFP mRNAs.
Translational efficiency was determined by
normalizing Flag-SRSF2 expression levels to
corresponding GFP levels.
MicroRNA (miR) array analysis -- HeLa
cells were treated with caffeine (14 mM) for
either 3 hrs or 24 hrs. Treated samples and
untreated controls were collected for total RNA
using miRCURYTM
RNA Isolation Kits
(Exiqon), and labeled with appropriate dyes
using miRCURY LNATM
microRNA Array Hi-
power Labeling Kit. The miRCURY LNATM
microRNA Array, v11.0-human, was the
platform utilized. Hybridization and processing
of arrays were performed by the CINJ DNA
Core Facility. Each experimental condition was
repeated 6 times including three biological
replicates and two technical replicates for each
biological replicate. The average signal density
was normalized by standard protocols (CINJ
Bioinformatics Core Facility) as well as internal
controls provided on the miRCURY LNATM
microRNA Array platform using miRCURY
LNATM
microRNA Array Analysis software.
miR qRT-PCR -- Total RNA was isolated
using miRCURYTM
RNA Isolation Kits
(Exiqon), and cDNA was prepared using the
Universal cDNA Synthesis Kit (Exiqon)
following manufacturers’ instructions.
Individually pre-designed LNA™-enhanced
microRNA qPCR primer sets were purchased
from Exiqon. qPCR was performed using
SYBR® Green Master Mix Kit (Exiqon) and a
Stratagene MX3000p qPCR system. RNA input
was normalized by U1 snRNA and SNORD49a.
RT-PCR efficiency was normalized using the
RNA Spike-in kit (Exiqon). The final Ct value
was an average of 5 repeats of each assay,
repeated three times. The relative amount of
mRNA in samples was determined using the 2-
ΔΔCt method.
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Caffeine modulates post-transcriptional regulation of SRSF2
5
Anti-miR and miR-mimic co-transfections--
HeLa cells were seeded at ~80,000 cells/well in
12-well plates and incubated overnight. Either
mirVana™ miRNA mimics or inhibitors
(Ambion, Invitrogen), at 30~50 nM (determined
empirically), were co-transfected with 150 ng of
indicated SRSF2 3’UTR plasmid DNA as well
as 37.5 ng of a GFP expression construct (pN3-
GFP). Lipofectamine 2000 reagent (Invitrogen)
was used following manufacturer’s instructions.
18~22 hrs after transfection, whole cell lysates
were collected for western blot analyses of Flag-
SRSF2 expression. GFP was used to control for
transfection efficiency.
To assay the effect of anti-miR and miR
mimics on SRSF2 expression, HeLa cells were
seeded at ~80,000 cells/well in 12-well plates
and incubated overnight. For miR mimics
experiment, 45nM of mirVana™ miRNA mimic
(Ambion, Invitrogen) was co-transfected with
the KLF6 minigene (29) as described. Caffeine
was added 18 hrs after transfection, and 10 hrs
later, cells were harvested for either western blot
analysis of SRSF2 expression or RT-PCR of
KLF6. For miR inhibitor experiments, 45nM of
mirVana™ miRNA inhibitor (Ambion,
Invitrogen) was co-transfected with hSMG-1
siRNA (100 nM, Dharmacon, GE Healthcare)
and the KLF6 minigene as described. Following
a 46-hr incubation, cells were harvested for
western blot analysis and RT-PCR performed as
above.
Statistical analysis -- All statistical analyses
were performed using GraphPad PRISM version
6 software, nonparametric t tests. A difference
with a p value < 0.05 was considered significant.
Error bars represent the mean ± SD.
RESULTS
Methylxanthines differentially induce KLF6
splice variants and increase levels of SRSF2
protein. We have previously reported that
caffeine, a tri-methylxanthine derivative, can
induce alternative splicing of a subset of cancer-
associated genes, including the tumor suppressor
gene KLF6. This induction is due, at least in
part, to a caffeine-mediated increase (up to 6-
fold) of the splicing factor SRSF2. In HeLa
cells this increase can be observed as early as 1
hour post caffeine treatment and is sustained for
at least 24 hours (29). To evaluate the impact of
other members of this class on splicing, we
examined the ability of several xanthine
derivatives, including pentoxifylline, caffeine,
theophylline, and isocaffeine, on induction of
the KLF6 splice variant, SpKLF6 (Figure 1A).
Within 18hrs of treatment, pentoxifylline
exhibited the strongest induction among the
group, followed by caffeine and theophylline.
Isocaffeine had minimal effect on SpKLF6
expression. This corresponded to the degree of
induction of SRSF2 by these compounds (Figure
1B), further supporting our observation that
caffeine-induced SpKLF6 expression is
mediated by SRSF2. Although pentoxifylline
induced the highest level of SRSF2, it also
caused severe cell death at the concentrations
used (14 mM) (Figure 1C). Therefore, caffeine
was used as the model drug to investigate the
mechanisms by which methylxanthines increase
SRSF2.
Caffeine does not impact total SRSF2 mRNA
levels. To investigate the molecular basis for the
increase in SRSF2 protein, we first considered
the possibility that caffeine was working at the
level of SRSF2 transcription. To evaluate this,
we employed both semi-quantitative and
quantitative RT-PCR to examine the total
SRSF2 mRNA following caffeine treatment.
Previous studies have described three SRSF2
transcripts that differ in the splicing of exon 3
(E3) and/or intron 4/5 (I4/5) at the 3’UTR
(Figure 2A) (27). Public databases (NCBI-
Aceview and Ensembl) suggested the existence
of additional splice variants of SRSF2 mRNA
that differ in their 3’UTR splicing choices and
polyadenylation sites (Figure 2A).
Interestingly, there are no alternative splice sites
documented within the SRSF2 coding region
that spans Exon 1 (E1) and 2 (E2), indicating
that both the known and predicted SRSF2
transcript variants would encode the same
protein. To account for all known and predicted
transcripts, primers located within the two
protein coding exons, E1 and E2 (Figure 2A,
primers ―a‖ and ―b‖), were chosen for analysis
of RNAs isolated from HeLa cells either
untreated or treated with caffeine for times
indicated. After careful linear range
optimization and quantitative analyses, both
semi-quantitative RT-PCR (Figure 2B) and
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Caffeine modulates post-transcriptional regulation of SRSF2
6
quantitative RT-PCR (Figure 2C) identified no
significant change in total SRSF2 mRNA
following caffeine treatment, suggesting that
caffeine regulates SRSF2 protein levels post-
transcriptionally.
Caffeine influences splicing decisions at the
SRSF2 3’UTR. Since there was no apparent
increase in total SRSF2 mRNA levels
concomitant with the caffeine-induced increase
in SRSF2 protein, and cycloheximide
experiments did not indicate a caffeine-mediated
change in SRSF2 protein stability (data not
shown), we next considered the possibility that
caffeine may impact SRSF2 splicing choices at
the 3’UTR, yielding transcripts with different
stability and/or translatability.
Four possible groups of SRSF2 transcripts
have been either described before or predicted
by the aforementioned databases. The major
SRSF2 transcript (hereafter referred to as A1)
includes the canonical E1, E2 and
polyadenylation site, but skips E3 and retains
intron 4/5 (Figure 3A, diagram). Two additional
SRSF2 transcripts, A2 and A3, can be induced
by high levels of SRSF2 protein via AS at the
3’UTR (Figure 3A, diagrams). Since A2 and A3
each contain a PTC, they are potential substrates
for NMD (27,30) and therefore can be quickly
degraded to restore normal levels of SRSF2
protein. Together, we refer to these mRNAs as
Group A (GA) transcripts. The predicted Group
B (GB) transcripts are similar to GA transcripts
in that they skip E3, but they utilize an alternate
polyadenylation site (Figure 3B, diagram).
Group C (GC) transcripts are predicted to
resemble GA transcripts in that they use the
canonical polyadenylation site, but contain an
elongated E2 due to differential utilization of the
E2 5’ splice site (Figure 3C, diagram). Finally,
predicted Group D (GD) transcripts resemble
GC transcripts in their E2 5’splicing but use the
alternative polyadenylation site (Figure 3D,
diagram).
To systematically examine the impact of
caffeine on all the identified and predicted
variants of SRSF2 RNA, multiplex RT-PCR
assays were developed to detect transcripts in
the four distinct groups in untreated and
caffeine-treated HeLa cells. Multiplex RT-PCR
using primer ―c‖ and ―d‖ was used to detect the
GA transcripts. Following a one hour exposure
to caffeine, a ―switch‖ in the expression of
splice variants was observed, with a significant
increase in expression of A2 and, to a lesser
extent A3, at the expense of A1 (Figure 3A).
Examination of putative GB transcripts using
primers ―c‖ and ―f‖ detected B2 but not B1 in
untreated samples, suggesting that SRSF2
transcripts that utilize the canonical E2 and the
alternative polyadenylation site selectively skip
E3; caffeine had a negligible effect on
expression of this group (Figure 3B). Multiplex
RT-PCR using primers ―e‖ and ―d‖ detected a
novel group of SRSF2 transcripts that contain an
elongated E2 with alternative 5’ splice site
choices at 1101 nt (C1), 389 nt (C2 and C4), 543
nt (C3), or 60 nt (C5) downstream of the stop
codon (Figure 3C). The impact of caffeine on
GC transcripts was similar to what was observed
for GA transcripts, in that alternative splicing of
intron 4/5 was induced (C1) at the expense of C2
(Figure 3C). In addition, utilization of novel 5’
splice sites 543 nt (C3) and 60 nt (C5) was
promoted (Figure 2C, note increased expression
of C3 and C5). Notably, GC transcripts were
not well represented in the population
(visualization required 5 additional PCR
amplification cycles). In contrast, GD
transcripts D1, D2 and D3, also novel splice
variants with similar alternative 5’ splice sites as
GC transcripts which utilize the alternative
polyadenylation site, are well expressed.
Caffeine increased the GD transcripts that utilize
5’ splice sites at 543 nt (D1) and 60 nt (D3), at
the expense of D2.
To summarize, caffeine significantly changed
the AS pattern of SRSF2 transcripts within the
3’UTR. This effect was fairly rapid; as shown
in Figure 3E, upper panel, the effect of caffeine
on A1 (decrease) and A2/A3 (increase) could be
observed as early as one hour, with maximum
levels achieved by 8 hrs. Similar kinetics was
obtained with GD transcripts (lower panel). As
basal GC transcript expression was negligible,
and caffeine had a minimal effect on the splicing
of GB transcripts, we focused on GA and GD
transcripts for the remainder of the study.
Caffeine does not change the levels of known
repressors of SRSF2 3’UTR alternative splicing.
It has been previously reported that two negative
splicing regulators, TARBP and HnRNP H, can
antagonize SRSF2-mediated splicing of intron
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Caffeine modulates post-transcriptional regulation of SRSF2
7
4/5 (28). We therefore considered the
possibility that the expression of one or both of
these negative regulators could be altered by
caffeine, leading to the skipping of intron 4/5.
To examine this possibility, TARBP and
HnRNP H levels were analysed following
treatment with or without caffeine; no
significant change in levels of either factor was
observed over a 24 hr period (Figure 4),
suggesting that they are not responsible for
caffeine-induced SRSF2 3’UTR alternative
splicing.
Inhibition of NMD is not sufficient to increase
SRSF2. One of the well-documented effects of
caffeine is inhibition of NMD, due to the
negative impact of caffeine on phosphorylation
of the essential factor hUpf1 (31-34). Since
several of the caffeine-induced SRSF2
transcripts (A2, A3, D1, D2, and D3) are
potential substrates for NMD, we next
considered the possibility that inhibition of
NMD by caffeine was sufficient to allow NMD-
sensitive isoforms to be stabilized and
accumulated. First, cycloheximide (CHX) was
used to inhibit the pioneer round of protein
translation that is required for NMD (33,35). As
shown in Figure 5A, CHX treatment alone
partially altered the GA and GD expression
profiles (compare lane 3 to lane 1); however,
treatment with CHX and caffeine resulted in a
distinctive splicing pattern within the 3’UTR
(compare lane 3 to lane 2), indicating that CHX-
mediated NMD inhibition could not reproduce
the effect of caffeine on SRSF2 variants
expression. To directly determine whether
NMD inhibition alone was sufficient to generate
alternate SRSF2 transcripts as well as increased
SRSF2 protein levels, an RNAi approach was
used to knockdown hUpf1 (36-38). As shown in
Figure 5B, hUpf1 levels were decreased by at
least ~ 85% using a targeted SiRNA smart pool
(Dharmacon), resulting in inhibition of NMD as
evidenced by the accumulation of a known
NMD substrate, the alternatively spliced, exon
11(34 nt)-skipped PTB1(39). Further RT-PCR
analyses on GA and GD transcripts revealed
that, although hUpf1 knockdown induced some
changes in SRSF2 splice variant expression
(Figure 5C, compare lane 3 to lane 1), it was not
sufficient to induce the full spectrum of changes
that is observed in the presence of caffeine.
Importantly, NMD inhibition was not sufficient
to induce an increase in SRSF2 protein (Figure
5D, lane 3 vs. lane 2). We have previously
shown that caffeine had no effect on other SR
proteins that employ AS-NMD as their auto-
regulatory mechanism (15,29,33), consistent
with our hypothesis that NMD inhibition itself is
not sufficient to induce SRSF2 protein and an
additional, gene-specific mechanism must be in
place.
SRSF2 transcripts with varied 3’UTRs exhibit
different translational efficiencies. Given that
caffeine altered the SRSF2 3’UTR alternative
splicing choices without changing the total
mRNA levels and that the 3’UTR is known to
play a critical role in the regulation of
translational efficiency (40-42), we considered
the possibility that the caffeine-regulated AS
transcripts had different translational
efficiencies. Both in vitro translation assays and
cell-based translational assays were employed to
evaluate this possibility. Initially, the individual
3’UTR of SRSF2 transcripts were inserted into a
renilla luciferase reporter vector to test for
relative translational efficiency, while firefly
luciferase vector served as a transfection control,
as is standard in the field. Unfortunately,
caffeine interfered with detection of luciferase
activity in this assay. Therefore, Flag-SRSF2
cDNA (created by fusing a Flag tag to the N-
terminal of each construct) was used instead of
renilla luciferase cDNA as a reporter and GFP
replaced firefly luciferase as a control for
transfection efficiency. Under these conditions,
transcription of each variant (A1, A2, A3, and
D1, D2, D3, complete mRNA) is controlled by
the same promoter, and products of each can be
distinguished from endogenous SRSF2 using the
anti-Flag antibody.
Plasmid DNA was prepared and carefully
quantitated for each construct. The same
amount of input DNA was assayed the TNT®
Quick Coupled Transcription/Translation
System (Promega). As shown in Figure 6A,
transcripts A2 and A3 exhibited significantly
higher intrinsic translational efficiency than A1
in this assay, while translation of GD group
members were similar. Next, cell-based
translation was assayed by co-transfecting each
of the SRSF2 transcript constructs with a GFP
expression construct to serve as a control for
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Caffeine modulates post-transcriptional regulation of SRSF2
8
transfection efficiency (43). Approximately 7
hours post transfection, cells were either
untreated or treated with caffeine. After 3 hours,
cells were harvested for western blot analysis of
both Flag-SRSF2 and GFP. The translational
efficiency of each SRSF2 transcript was
determined by normalizing Flag-SRSF2
expression to the expression of GFP. To assure
that the expression/stability of mRNA did not
differ among the transcripts, mRNA levels of
each Flag-SRSF2 and GFP were examined using
real-time quantitative RT-PCR assay. When
normalized to GFP, the levels of Flag-SRSF2
mRNA remained constant for all transcripts
tested (A1-A3, D1-D3) (Figure 6C).
As shown in Figure 6B, GA transcripts with
shorter 3’UTRs (A2, A3) exhibited higher
translation efficiency than A1 (left panel), while
the GD transcript D2 was translated with the
highest efficiency among the GD members
(right panel). Importantly, caffeine significantly
increased translational efficiency of every
variant tested by approximately 2-3 fold. This
moderate regulation of translation was
reminiscent of the impact of microRNAs (miRs)
on translation efficiency (44).
Caffeine-mediated downregulation of miR-183-
5p and miR-33a-5p alters protein translation
efficiency of specific SRSF2 transcripts.
MicroRNAs are small non-coding RNAs that
regulate gene expression by base-pairing with
specific mRNA molecules, usually resulting in
altered protein translation or mRNA stability
(45,46). MicroRNA-mediated translation
repression has recently been suggested as a
mechanism for SRSF2 regulation in amygdala
during acute stress (miR-183-5p) (47) and in
hepatocellular carcinoma during the acquisition
of drug resistance (miR-193a-5p) (48).
To determine whether caffeine regulates the
expression of miRs that could in turn impact
SRSF2 expression, microarrays were carried out
using the miRCURY LNATM
microRNA Array
platform and total RNA isolated from either
untreated HeLa cells or cells treated for 3 hrs or
24 hrs with caffeine. Each condition was
repeated three times and data was normalized to
internal controls present on the array. After
careful data mining using common criteria, ~
8% of the MiRs examined (99 out of ~1200)
were found to be downregulated by caffeine
(Supplementary Data Table 1 and 2). This
group of miRs was aligned with computationally
predicted SRSF2-targeting miRs (185 miRs,
based on the miRscan database,
http://genes.mit.edu/mirscan/), then further
limited to miRs that were abundantly expressed
in untreated control cells. Two microRNAs,
miR-183-5p and miR-33a-5p, were found to be
significantly down-regulated by caffeine; miR-
183-5p has previously been suggested to repress
SRSF2 translation (47) by interacting with the
SRSF2 transcript at position 1159 nt within the
3’UTR. MiR-33a-5p was suggested to target the
SRSF2 3'UTR at position 1746nt
(http://www.microrna.org/) (Figure 7A).
Therefore, miR-183-5p would likely target A1,
D1, D2, and D3 and miR-33a-5p could impact
translation of A1, A2 and A3 (Figure 7A). qRT-
PCR analysis of RNAs isolated from untreated
and treated HeLa cells was performed to validate
the downregulation of these miRs by caffeine.
As shown in Figure 7B, following 3-hr of
caffeine treatment, miR-183-5p was decreased
dramatically (5~6-fold) while miR-33q-5p levels
remain largely unchanged. However, following
24-hr of caffeine treatment, expression of miR-
183-5p was still decreased and miR-33a-5p was
decreased by almost half, similar to what was
observed in the microarray analysis.
To test the hypothesis that these miRs
impacted translation of specific SRSF2
transcripts, the levels of miR-183-5p and miR-
33a-5p were manipulated using either specific
miR mimics or specific miR inhibitors, which
were co-transfected into HeLa cells along with
one of the putative target SRSF2 transcripts. We
reasoned that if the miR indeed regulated the
translation of the co-transfected SRSF2
transcripts, differences should be observed in
their translational efficiency when compared to
controls. Similar to what was employed for the
cell-based transfection-translation assays in
Figure 6, a GFP expression plasmid was used to
control for transfection efficiency. 18-20 hours
post transfection, both flag-SRSF2 and GFP
levels were assessed by western blotting, and
translation efficiency was determined by
normalizing Flag-SRSF2 levels to GFP levels.
As showed in Figure 7C, the translation
efficiency of the wild-type A1 transcript (A1-
WT), predicted to be repressed by miR-183-5p,
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Caffeine modulates post-transcriptional regulation of SRSF2
9
was further decreased by miR-183-5p mimics
and increased by miR-183-5p inhibitors.
Importantly, when the putative miR-183-5p
binding site within A1 was mutated (A1-
183Mut), neither the miR-183-5p mimic nor the
inhibitor had a significant impact on A1-183
mutant translation. Moreover, neither the miR-
183-5p mimic nor inhibitor had an effect on
translation of A2, which lacks the miR-183-5p
binding site. Taken together, these results
identify miR-183-5p as an SRSF2 (A1)-
targeting miR that binds to the 3’UTR at 1159-nt
downstream of the stop codon to repress A1
translation. Similar assays using A2 WT and
A2-33aMut also identified miR-33a-5p as an
SRSF2-targeting miR imposing translational
regulation (Figure 7D), with D1 serving as a
negative control.
To further confirm the effect of miR-183-5p
and miR-33a-5p on caffeine-mediated SRSF2
translation, miR-183-5p and miR-33a-5p mimics
were co-transfected with the KLF6 minigene,
followed by caffeine treatment. As shown in
Figure 7E (left panel, top), while caffeine
increased endogenous SRSF2 by almost 4-fold
in the presence of non-specific miR mimics, this
effect was largely reduced in the presence of the
specific miR mimics with a concomitant
decrease in KLF6 alternative splicing (~ 3-fold,
from 89% to 32%). We next investigated the
effect that downregulation of miR-183-5p and
miR-33a-5p had on SRSF2 and KLF6, reasoning
that this should mimic at least some of the
effects of caffeine. Since our data suggested that
caffeine has two activities that regulate
SRSF2/KLF6 – downregulation of the SRSF2-
targeting miRs and inhibition of NMD, we
inhibited NMD by siRNA-mediated
downregulation of the key NMD protein,
hSMG1(37), which is also the direct target of
caffeine (31,32), in this experiment. As shown
in Figure 7E, right panel, when NMD is
inhibited, downregulation of the miRs increased
SRSF2 levels (~3-fold) and KLF6 alternative
splicing (~ 3-fold, from 23% to 68%). Taken
together, our results reveal a novel effect of
caffeine on miR regulation that in turn impacts
SRSF2 translation and KLF6 alternative
splicing, supporting the hypothesis that caffeine-
induced SRSF2 translational regulation
contributes significantly to caffeine-induced
increases in SRSF2 protein levels and alternative
splicing decisions.
DISCUSSION
We have previously reported that caffeine
can impact alternative splicing (AS) of a subset
of cancer-related genes. Using KLF6 as a
model, we demonstrated that the change in
KLF6 splicing pattern could not be mimicked by
inhibition of nonsense-mediated decay (NMD),
even though the caffeine-induced KLF6 splice
variant was a potential target for this RNA
surveillance mechanism. Instead, the altered
splicing of KLF6 was a result, at least in part, of
a caffeine-induced increase (~6-fold) in levels of
the splicing factor SRSF2 (29). In this report,
we show that additional methylxanthines can
also induce AS of KLF6, with a concomitant
increase in SRSF2 levels. SRSF2 gene
expression is known to be guarded by a negative
feedback loop, and a low steady level of SRSF2
is important for homeostasis of isoform
expression as well as cell proliferation and
genome stability (18). Thus, it was important to
understand how methylxanthines, using caffeine
as a prototype, had affected this normally
unsustainable increase of SRSF2. We
demonstrate that, in addition to its ability to
inhibit NMD, caffeine can also regulate the
expression of specific microRNAs, resulting in
changes in translation efficiency of SRSF2
transcripts. Together, these modifications result
in increased translation of certain SRSF2
variants and stabilization of NMD-sensitive
SRSF2 transcripts with higher intrinsic
translational efficiency; together these caffeine-
induced changes lead to a surge of SRSF2
protein levels and an increase in SRSF2 splice
variants designated for degradation by NMD,
and the cycle continued. Thus, caffeine breaks
the negative feedback loop controlling SRSF2
gene expression and further enforces a positive
feed-forward loop that fuels the increased
production of SRSF2 (See Model, Figure 8).
While we have not yet determined whether
caffeine plays a similar role in vivo, it is likely
that it is mimicking an endogenous process or
processes that are responsible for fine-tuning
SRSF2 gene expression in response to
developmental or environmental signals, or
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Caffeine modulates post-transcriptional regulation of SRSF2
10
during pathogenesis. Thus, caffeine represents a
valuable tool to dissect this mechanism.
Previous studies have identified two SRSF2
3’UTR variants induced by increased SRSF2
levels or depletion of the splicing repressor
HnRNP H (27,28,30). These SRSF2 variants of
the major transcript, A1, correspond to A2 and
A3 in this study. In this study, we identified
additional groups of transcripts, GB, GC, and
GD. Most of these novel variants are potential
targets of NMD, indicating that they are
probably part of the AS-NMD circuit regulating
SRSF2 protein levels (3,49). For the major GA
transcripts, translation of the variants in cultured
cells fit the general dogma – variants with
shorter 3’UTRs were translated with the highest
efficiency (50). However, translation of GD
variants was less conventional, in that the
variant with the intermediate size 3’UTR (D2)
was translated with the highest efficiency
(Figure 6). Interestingly, this difference among
GD transcripts was not obvious in cell-free
transcription/translation assays (Figure 6A),
suggesting that other cellular processes/signals,
most likely interacting with the 3’UTR
sequence between 60 to 543nt downstream of
the stop codon, are involved. The GD
transcripts account for a relatively small portion
of the total SRSF2 mRNAs in HeLa cells, but
appear to make a marked contribution to SRSF2
gene regulation, which may be more critical in
tissue types or cell lines that preferentially
utilize the alternative polyadenylation site (51).
Our study is the most thorough examination of
SRSF2 transcripts and their translational
efficiency to date. While the expression and role
of these variants in different cells/tissues, and
under different physiological and pathological
conditions, has yet to be determined, this study,
together with the comprehensive analysis of
SRSF1 by the Krainer group (52), contributes to
our understanding of the complex regulation of
SR proteins gene expression.
One unexpected result of these studies was
that depletion of hUpf1, best known as an
essential factor in the NMD pathway, had little
effect on SRSF2 levels alone, but prevented
induction of SRSF2 by caffeine (Figure 5C).
Interestingly, recent studies suggest additional
functions of hUpf1 other than in NMD,
including a role in the regulation of protein
translation. Yoneda and colleagues recently
reported that hUpf1 was required for Stau2
overexpression-mediated induction of translation
(53), while Moore and colleagues suggested that
hUpf1, when associated with Exon junction
complex (EJC), may contribute to the translation
of spliced mRNAs (54). Indeed, NMD factors
may be involved in both RNA degradation and
enhanced translation when splicing efficiency is
improved (55). Given that our model indicates a
substantial contribution from caffeine-
stabilized/NMD-targeted SRSF2 transcripts to
caffeine-mediated SRSF2 increase, we speculate
that hUpf1 may be required in translation of
these transcripts. Further studies will be needed
to validate this hypothesis.
While searching for a second mechanism for
the caffeine induction of SRSF2, we identified a
novel action of this methylxanthine, the
regulation of miR expression. MiR-mediated
gene expression regulation is a well-recognized
post-transcriptional mechanism that either
represses translation and/or destabilizes mRNA
transcripts to control protein production (44,56-
58). Nearly 60% of protein-coding genes are
predicted to be targeted by microRNAs and most
microRNAs act by base-pairing with the 3’UTR
of the targeted mRNA (59). One gene could
potentially be targeted by multiple microRNAs;
however, it is unlikely that all of them would be
functional at the same time because miRs are
expressed in a tissue- and development-specific
manner (44,56-58,60). For SRSF2, algorithms
including microRNA (www.microrna.org) and
miRScan (http://genes.mit.edu/mirscan) predict
~185 putative miR binding sites within the
3’UTR. Only two of these have been previously
validated: miR-183-5p in stress response (47)
and miR-193a-3p in cancer drug resistance (48).
We now show that caffeine can regulate a subset
of miRs, including miR-183-5p, which targets
A1 and possibly all GD members (Figure 7). The down-regulation of miR-183-5p by caffeine
was rapid, dramatic, and persistent (Figure 7B
and Supplement Table 1 and 2), supporting a
significant role of this downregulation in
caffeine-mediated SRSF2 protein induction. In
addition, we have for the first time validated
miR-33a-5p as an SRSF2-targeting miR, and
shown that this miR is also downregulated by
caffeine. Because the putative binding site for
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Caffeine modulates post-transcriptional regulation of SRSF2
11
miR-33a-5p is located 1746 nt downstream of
the stop codon, this miR primarily impacted
protein production of transcripts using the
canonical polyadenylation site such as the GA
group, the major group of SRSF2 transcripts.
It should be noted that the miR-183-5p
binding site, shared by A1 and D1-D3, overlaps
a putative ARE (A-U rich element)(56). AREs
are found in the 3’UTR of many mRNAs and
can interact with a number of proteins to either
stabilize (the Hu family of proteins) or
destabilize (AUF1, TTP, BRF1, TIA-1, TIAR,
and KSRP) the mRNA (57-60), a common
determinant of RNA stability. To date, the
putative AREs within SRSF2 3’UTR have not
been functionally tested, nor have their cognate
binding proteins, either stabilizing or
destabilizing, been identified. Nevertheless, it is
feasible that their proximity to the miR binding
site may impact miR activity under certain
conditions. The function of these AREs and
their potential impact on the miR activity and
SFSF2 RNA stability (or vice versa) is currently
under investigation.
Our results suggest that aberrant expression
of either miR-183-5p or miR-33a-5p would have
a marked impact on SRSF2 protein production,
which in turn could influence alternative
splicing choices in SRSF2-targeted genes,
including many cancer-related genes. Indeed,
abnormal levels of miR-183-5p have been
observed in various tumors (61), where it
appears to play a role in tumor progression due
to its impact on proliferation, apoptosis, and
metastasis (62-65). Altered regulation of miR-
183-5p has also made it an attractive biomarker
candidate (66-72). miR-33a-5p has also been
suggested to play a role in tumorigenesis (73-
75), as well as in controlling cholesterol and
lipid metabolism (76,77). However, to date
there has been no evidence to link aberrant
expression of these miRs to AS in cancers.
Therefore, our observation that alteration of
these miRs can have a profound effect on the
expression of SRSF2, a critical splicing factor
regulating genes involved in cell proliferation,
cell death and metastasis, warrants further
studies.
It is not yet known how caffeine modulates
levels of miR-183-5p, miR-33a-5p, or other
miRs. Since transcriptional regulation can be
influenced by caffeine (78,79 ,80), we compared
the effects of caffeine on miR-183-5p levels to
that of two additional miRs within the miR-183-
96-182 cluster, miR-96 or miR-182, because
they share the same promoter (81,82). No
significant effect was observed (Supplement,
Table 1 and 2), suggesting that transcriptional
regulation is unlikely to be the underlying
mechanism. Given the pleiotropic effect of
caffeine on cells (83-85), it is possible that
multiple mechanisms may be involved in
caffeine-mediated miR regulation.
In summary, we have shown that caffeine
regulates expression of SRSF2 through a
complex set of post-transcriptional mechanisms
that work together to break the AS-NMD
feedback loop and foster a feed-forward loop
that increases SRSF2 protein levels. Our studies
to date underscore the complicated interplay of
multiple mechanisms needed to control SRSF2
expression that in turn controls the expression of
a large subset of target genes. This knowledge
may provide a foundation for exploring the
aberrant regulation of SRSF2 and target genes in
pathogenesis.
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Caffeine modulates post-transcriptional regulation of SRSF2
12
REFERENCES
1. Takeda, J., Suzuki, Y., Sakate, R., Sato, Y., Gojobori, T., Imanishi, T., and Sugano, S. (2010) H-
DBAS: human-transcriptome database for alternative splicing: update 2010. Nucleic Acids Res
38, D86-90
2. Bauman, J. A., and Kole, R. (2011) Modulation of RNA splicing as a potential treatment for
cancer. Bioengineered bugs 2, 125-128
3. McGlincy, N. J., and Smith, C. W. (2008) Alternative splicing resulting in nonsense-mediated
mRNA decay: what is the meaning of nonsense? Trends Biochem Sci 33, 385-393
4. Glisovic, T., Bachorik, J. L., Yong, J., and Dreyfuss, G. (2008) RNA-binding proteins and post-
transcriptional gene regulation. FEBS Lett 582, 1977-1986
5. Ajay, S. S., Athey, B. D., and Lee, I. (2010) Unified translation repression mechanism for
microRNAs and upstream AUGs. BMC Genomics 11, 155
6. Galban, S., Kuwano, Y., Pullmann, R., Jr., Martindale, J. L., Kim, H. H., Lal, A., Abdelmohsen,
K., Yang, X., Dang, Y., Liu, J. O., Lewis, S. M., Holcik, M., and Gorospe, M. (2008) RNA-
binding proteins HuR and PTB promote the translation of hypoxia-inducible factor 1alpha. Mol
Cell Biol 28, 93-107
7. Kato, Y., and Nakamura, A. (2009) [Mechanisms underlying maternal RNA translation and
localization during Drosophila oogenesis]. Tanpakushitsu kakusan koso. Protein, nucleic acid,
enzyme 54, 2159-2164
8. Kalsotra, A., and Cooper, T. A. (2011) Functional consequences of developmentally regulated
alternative splicing. Nat Rev Genet 12, 715-729
9. Braeutigam, C., Rago, L., Rolke, A., Waldmeier, L., Christofori, G., and Winter, J. (2014) The
RNA-binding protein Rbfox2: an essential regulator of EMT-driven alternative splicing and a
mediator of cellular invasion. Oncogene 33, 1082-1092
10. Coelho, M. B., and Smith, C. W. (2014) Regulation of alternative pre-mRNA splicing. Methods
Mol Biol 1126, 55-82
11. Smith, C. W., and Valcarcel, J. (2000) Alternative pre-mRNA splicing: the logic of combinatorial
control. Trends Biochem Sci 25, 381-388
12. Caceres, J. F., and Kornblihtt, A. R. (2002) Alternative splicing: multiple control mechanisms and
involvement in human disease. Trends Genet 18, 186-193
13. Mendell, J. T., Sharifi, N. A., Meyers, J. L., Martinez-Murillo, F., and Dietz, H. C. (2004)
Nonsense surveillance regulates expression of diverse classes of mammalian transcripts and
mutes genomic noise. Nat Genet 36, 1073-1078
14. McIlwain, D. R., Pan, Q., Reilly, P. T., Elia, A. J., McCracken, S., Wakeham, A. C., Itie-Youten,
A., Blencowe, B. J., and Mak, T. W. (2010) Smg1 is required for embryogenesis and regulates
diverse genes via alternative splicing coupled to nonsense-mediated mRNA decay. Proc Natl
Acad Sci U S A 107, 12186-12191
15. Saltzman, A. L., Kim, Y. K., Pan, Q., Fagnani, M. M., Maquat, L. E., and Blencowe, B. J. (2008)
Regulation of multiple core spliceosomal proteins by alternative splicing-coupled nonsense-
mediated mRNA decay. Mol Cell Biol 28, 4320-4330
16. Fu, X. D., Mayeda, A., Maniatis, T., and Krainer, A. R. (1992) General splicing factors SF2 and
SC35 have equivalent activities in vitro, and both affect alternative 5' and 3' splice site selection.
Proc Natl Acad Sci U S A 89, 11224-11228
17. Tacke, R., and Manley, J. L. (1995) The human splicing factors ASF/SF2 and SC35 possess
distinct, functionally significant RNA binding specificities. Embo J 14, 3540-3551
18. Xiao, R., Sun, Y., Ding, J. H., Lin, S., Rose, D. W., Rosenfeld, M. G., Fu, X. D., and Li, X.
(2007) Splicing regulator SC35 is essential for genomic stability and cell proliferation during
mammalian organogenesis. Mol Cell Biol 27, 5393-5402
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Caffeine modulates post-transcriptional regulation of SRSF2
13
19. Ji, X., Zhou, Y., Pandit, S., Huang, J., Li, H., Lin, C. Y., Xiao, R., Burge, C. B., and Fu, X. D.
(2013) SR proteins collaborate with 7SK and promoter-associated nascent RNA to release paused
polymerase. Cell 153, 855-868
20. Lin, S., Coutinho-Mansfield, G., Wang, D., Pandit, S., and Fu, X. D. (2008) The splicing factor
SC35 has an active role in transcriptional elongation. Nat Struct Mol Biol 15, 819-826
21. Merdzhanova, G., Edmond, V., De Seranno, S., Van den Broeck, A., Corcos, L., Brambilla, C.,
Brambilla, E., Gazzeri, S., and Eymin, B. (2008) E2F1 controls alternative splicing pattern of
genes involved in apoptosis through upregulation of the splicing factor SC35. Cell death and
differentiation 15, 1815-1823
22. Lu, Y., Loh, Y. H., Li, H., Cesana, M., Ficarro, S. B., Parikh, J. R., Salomonis, N., Toh, C. X.,
Andreadis, S. T., Luckey, C. J., Collins, J. J., Daley, G. Q., and Marto, J. A. (2014) Alternative
splicing of MBD2 supports self-renewal in human pluripotent stem cells. Cell stem cell 15, 92-
101
23. Muller-Thomas, C., Rudelius, M., Rondak, I. C., Haferlach, T., Schanz, J., Huberle, C., Schmidt,
B., Blaser, R., Kremer, M., Peschel, C., Germing, U., Platzbecker, U., and Gotze, K. (2014)
Response to azacitidine is independent of p53 expression in higher-risk myelodysplastic
syndromes and secondary acute myeloid leukemia. Haematologica 99, e179-181
24. Larsson, C. A., Cote, G., and Quintas-Cardama, A. (2013) The changing mutational landscape of
acute myeloid leukemia and myelodysplastic syndrome. Molecular cancer research : MCR 11,
815-827
25. Lasho, T. L., Jimma, T., Finke, C. M., Patnaik, M., Hanson, C. A., Ketterling, R. P., Pardanani,
A., and Tefferi, A. (2012) SRSF2 mutations in primary myelofibrosis: significant clustering with
IDH mutations and independent association with inferior overall and leukemia-free survival.
Blood 120, 4168-4171
26. Yoshida, K., Sanada, M., Shiraishi, Y., Nowak, D., Nagata, Y., Yamamoto, R., Sato, Y., Sato-
Otsubo, A., Kon, A., Nagasaki, M., Chalkidis, G., Suzuki, Y., Shiosaka, M., Kawahata, R.,
Yamaguchi, T., Otsu, M., Obara, N., Sakata-Yanagimoto, M., Ishiyama, K., Mori, H., Nolte, F.,
Hofmann, W. K., Miyawaki, S., Sugano, S., Haferlach, C., Koeffler, H. P., Shih, L. Y., Haferlach,
T., Chiba, S., Nakauchi, H., Miyano, S., and Ogawa, S. (2011) Frequent pathway mutations of
splicing machinery in myelodysplasia. Nature 478, 64-69
27. Sureau, A., Gattoni, R., Dooghe, Y., Stevenin, J., and Soret, J. (2001) SC35 autoregulates its
expression by promoting splicing events that destabilize its mRNAs. Embo J 20, 1785-1796
28. Dreumont, N., Hardy, S., Behm-Ansmant, I., Kister, L., Branlant, C., Stevenin, J., and Bourgeois,
C. F. (2010) Antagonistic factors control the unproductive splicing of SC35 terminal intron.
Nucleic Acids Res 38, 1353-1366
29. Shi, J., Hu, Z., Pabon, K., and Scotto, K. W. (2008) Caffeine regulates alternative splicing in a
subset of cancer-associated genes: a role for SC35. Mol Cell Biol 28, 883-895
30. Lareau, L. F., Brooks, A. N., Soergel, D. A., Meng, Q., and Brenner, S. E. (2007) The coupling of
alternative splicing and nonsense-mediated mRNA decay. Adv Exp Med Biol 623, 190-211
31. Usuki, F., Yamashita, A., Higuchi, I., Ohnishi, T., Shiraishi, T., Osame, M., and Ohno, S. (2004)
Inhibition of nonsense-mediated mRNA decay rescues the phenotype in Ullrich's disease. Ann
Neurol 55, 740-744
32. Ivanov, I., Lo, K. C., Hawthorn, L., Cowell, J. K., and Ionov, Y. (2007) Identifying candidate
colon cancer tumor suppressor genes using inhibition of nonsense-mediated mRNA decay in
colon cancer cells. Oncogene 26, 2873-2884
33. Moriniere, M., Delhommeau, F., Calender, A., Ribeiro, L., Delaunay, J., and Baklouti, F. (2010)
Nonsense-mediated mRNA decay (NMD) blockage promotes nonsense mRNA stabilization in
protein 4.1R deficient cells carrying the 4.1R Coimbra variant of hereditary elliptocytosis. Blood
cells, molecules & diseases 45, 284-288
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Caffeine modulates post-transcriptional regulation of SRSF2
14
34. Johnson, J. K., Waddell, N., and Chenevix-Trench, G. (2012) The application of nonsense-
mediated mRNA decay inhibition to the identification of breast cancer susceptibility genes. BMC
cancer 12, 246
35. Dang, Y., Low, W. K., Xu, J., Gehring, N. H., Dietz, H. C., Romo, D., and Liu, J. O. (2009)
Inhibition of nonsense-mediated mRNA decay by the natural product pateamine A through
eukaryotic initiation factor 4AIII. J Biol Chem 284, 23613-23621
36. Mendell, J. T., ap Rhys, C. M., and Dietz, H. C. (2002) Separable roles for rent1/hUpf1 in altered
splicing and decay of nonsense transcripts. Science 298, 419-422
37. Usuki, F., Yamashita, A., Kashima, I., Higuchi, I., Osame, M., and Ohno, S. (2006) Specific
inhibition of nonsense-mediated mRNA decay components, SMG-1 or Upf1, rescues the
phenotype of Ullrich disease fibroblasts. Molecular therapy : the journal of the American Society
of Gene Therapy 14, 351-360
38. Akaike, Y., Kurokawa, K., Kajita, K., Kuwano, Y., Masuda, K., Nishida, K., Kang, S. W.,
Tanahashi, T., and Rokutan, K. (2011) Skipping of an alternative intron in the srsf1 3'
untranslated region increases transcript stability. The journal of medical investigation : JMI 58,
180-187
39. Wollerton, M. C., Gooding, C., Wagner, E. J., Garcia-Blanco, M. A., and Smith, C. W. (2004)
Autoregulation of polypyrimidine tract binding protein by alternative splicing leading to
nonsense-mediated decay. Mol Cell 13, 91-100
40. Jia, J., Yao, P., Arif, A., and Fox, P. L. (2013) Regulation and dysregulation of 3'UTR-mediated
translational control. Current opinion in genetics & development 23, 29-34
41. Ratti, A., Fallini, C., Colombrita, C., Pascale, A., Laforenza, U., Quattrone, A., and Silani, V.
(2008) Post-transcriptional regulation of neuro-oncological ventral antigen 1 by the neuronal
RNA-binding proteins ELAV. J Biol Chem 283, 7531-7541
42. Khabar, K. S. (2010) Post-transcriptional control during chronic inflammation and cancer: a focus
on AU-rich elements. Cellular and molecular life sciences : CMLS 67, 2937-2955
43. Muroski, M. E., Kogot, J. M., and Strouse, G. F. (2012) Bimodal gold nanoparticle therapeutics
for manipulating exogenous and endogenous protein levels in mammalian cells. J Am Chem Soc
134, 19722-19730
44. Baek, D., Villen, J., Shin, C., Camargo, F. D., Gygi, S. P., and Bartel, D. P. (2008) The impact of
microRNAs on protein output. Nature 455, 64-71
45. Farazi, T. A., Hoell, J. I., Morozov, P., and Tuschl, T. (2013) MicroRNAs in human cancer. Adv
Exp Med Biol 774, 1-20
46. Risso, G., Pelisch, F., Quaglino, A., Pozzi, B., and Srebrow, A. (2012) Regulating the regulators:
serine/arginine-rich proteins under scrutiny. IUBMB life 64, 809-816
47. Meerson, A., Cacheaux, L., Goosens, K. A., Sapolsky, R. M., Soreq, H., and Kaufer, D. (2010)
Changes in brain MicroRNAs contribute to cholinergic stress reactions. J Mol Neurosci 40, 47-55
48. Ma, K., He, Y., Zhang, H., Fei, Q., Niu, D., Wang, D., Ding, X., Xu, H., Chen, X., and Zhu, J.
(2012) DNA methylation-regulated miR-193a-3p dictates resistance of hepatocellular carcinoma
to 5-fluorouracil via repression of SRSF2 expression. J Biol Chem 287, 5639-5649
49. Ni, J. Z., Grate, L., Donohue, J. P., Preston, C., Nobida, N., O'Brien, G., Shiue, L., Clark, T. A.,
Blume, J. E., and Ares, M., Jr. (2007) Ultraconserved elements are associated with homeostatic
control of splicing regulators by alternative splicing and nonsense-mediated decay. Genes Dev 21,
708-718
50. Sandberg, R., Neilson, J. R., Sarma, A., Sharp, P. A., and Burge, C. B. (2008) Proliferating cells
express mRNAs with shortened 3' untranslated regions and fewer microRNA target sites. Science
320, 1643-1647
51. Zhang, H., Lee, J. Y., and Tian, B. (2005) Biased alternative polyadenylation in human tissues.
Genome Biol 6, R100
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Caffeine modulates post-transcriptional regulation of SRSF2
15
52. Sun, S., Zhang, Z., Sinha, R., Karni, R., and Krainer, A. R. (2010) SF2/ASF autoregulation
involves multiple layers of post-transcriptional and translational control. Nat Struct Mol Biol 17,
306-312
53. Miki, T., Kamikawa, Y., Kurono, S., Kaneko, Y., Katahira, J., and Yoneda, Y. (2011) Cell type-
dependent gene regulation by Staufen2 in conjunction with Upf1. BMC molecular biology 12, 48
54. Nott, A., Le Hir, H., and Moore, M. J. (2004) Splicing enhances translation in mammalian cells:
an additional function of the exon junction complex. Genes Dev 18, 210-222
55. Gudikote, J. P., Imam, J. S., Garcia, R. F., and Wilkinson, M. F. (2005) RNA splicing promotes
translation and RNA surveillance. Nat Struct Mol Biol 12, 801-809
56. Bhattacharya, S., Giordano, T., Brewer, G., and Malter, J. S. (1999) Identification of AUF-1
ligands reveals vast diversity of early response gene mRNAs. Nucleic Acids Res 27, 1464-1472
57. von Roretz, C., Di Marco, S., Mazroui, R., and Gallouzi, I. E. (2011) Turnover of AU-rich-
containing mRNAs during stress: a matter of survival. Wiley interdisciplinary reviews. RNA 2,
336-347
58. Zhuang, R., Rao, J. N., Zou, T., Liu, L., Xiao, L., Cao, S., Hansraj, N. Z., Gorospe, M., and
Wang, J. Y. (2013) miR-195 competes with HuR to modulate stim1 mRNA stability and regulate
cell migration. Nucleic Acids Res 41, 7905-7919
59. Lee, H. H., Kim, W. T., Kim, D. H., Park, J. W., Kang, T. H., Chung, J. W., and Leem, S. H.
(2013) Tristetraprolin suppresses AHRR expression through mRNA destabilization. FEBS Lett
587, 1518-1523
60. Maitra, S., Chou, C. F., Luber, C. A., Lee, K. Y., Mann, M., and Chen, C. Y. (2008) The AU-rich
element mRNA decay-promoting activity of BRF1 is regulated by mitogen-activated protein
kinase-activated protein kinase 2. RNA 14, 950-959
61. Zhang, Q. H., Sun, H. M., Zheng, R. Z., Li, Y. C., Zhang, Q., Cheng, P., Tang, Z. H., and Huang,
F. (2013) Meta-analysis of microRNA-183 family expression in human cancer studies comparing
cancer tissues with noncancerous tissues. Gene 527, 26-32
62. Zhu, J., Feng, Y., Ke, Z., Yang, Z., Zhou, J., Huang, X., and Wang, L. (2012) Down-regulation of
miR-183 promotes migration and invasion of osteosarcoma by targeting Ezrin. Am J Pathol 180,
2440-2451
63. Zhang, Z., Li, S., and Cheng, S. Y. (2013) The miR-183 approximately 96 approximately 182
cluster promotes tumorigenesis in a mouse model of medulloblastoma. Journal of biomedical
research 27, 486-494
64. Shi, X. Y., Gu, L., Chen, J., Guo, X. R., and Shi, Y. L. (2014) Downregulation of miR-183
inhibits apoptosis and enhances the invasive potential of endometrial stromal cells in
endometriosis. International journal of molecular medicine 33, 59-67
65. Yoshino, H., Seki, N., Itesako, T., Chiyomaru, T., Nakagawa, M., and Enokida, H. (2013)
Aberrant expression of microRNAs in bladder cancer. Nature reviews. Urology 10, 396-404
66. Silva-Santos, R. M., Costa-Pinheiro, P., Luis, A., Antunes, L., Lobo, F., Oliveira, J., Henrique,
R., and Jeronimo, C. (2013) MicroRNA profile: a promising ancillary tool for accurate renal cell
tumour diagnosis. British journal of cancer 109, 2646-2653
67. Vosa, U., Vooder, T., Kolde, R., Vilo, J., Metspalu, A., and Annilo, T. (2013) Meta-analysis of
microRNA expression in lung cancer. Int J Cancer 132, 2884-2893
68. Rizos, E., Siafakas, N., Koumarianou, A., Katsantoni, E., Filippopoulou, A., Ntounas, P.,
Touloumis, C., Kastania, A., and Zoumpourlis, V. (2012) miR-183 as a molecular and protective
biomarker for cancer in schizophrenic subjects. Oncol Rep 28, 2200-2204
69. Lin, Q., Mao, W., Shu, Y., Lin, F., Liu, S., Shen, H., Gao, W., Li, S., and Shen, D. (2012) A
cluster of specified microRNAs in peripheral blood as biomarkers for metastatic non-small-cell
lung cancer by stem-loop RT-PCR. Journal of cancer research and clinical oncology 138, 85-93
70. Zhu, W., Liu, X., He, J., Chen, D., Hunag, Y., and Zhang, Y. K. (2011) Overexpression of
members of the microRNA-183 family is a risk factor for lung cancer: a case control study. BMC
cancer 11, 393
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Caffeine modulates post-transcriptional regulation of SRSF2
16
71. Yamada, Y., Enokida, H., Kojima, S., Kawakami, K., Chiyomaru, T., Tatarano, S., Yoshino, H.,
Kawahara, K., Nishiyama, K., Seki, N., and Nakagawa, M. (2011) MiR-96 and miR-183
detection in urine serve as potential tumor markers of urothelial carcinoma: correlation with stage
and grade, and comparison with urinary cytology. Cancer science 102, 522-529
72. Schaefer, A., Jung, M., Mollenkopf, H. J., Wagner, I., Stephan, C., Jentzmik, F., Miller, K., Lein,
M., Kristiansen, G., and Jung, K. (2010) Diagnostic and prognostic implications of microRNA
profiling in prostate carcinoma. Int J Cancer 126, 1166-1176
73. Kuo, P. L., Liao, S. H., Hung, J. Y., Huang, M. S., and Hsu, Y. L. (2013) MicroRNA-33a
functions as a bone metastasis suppressor in lung cancer by targeting parathyroid hormone related
protein. Biochimica et biophysica acta 1830, 3756-3766
74. Cirera-Salinas, D., Pauta, M., Allen, R. M., Salerno, A. G., Ramirez, C. M., Chamorro-Jorganes,
A., Wanschel, A. C., Lasuncion, M. A., Morales-Ruiz, M., Suarez, Y., Baldan, A., Esplugues, E.,
and Fernandez-Hernando, C. (2012) Mir-33 regulates cell proliferation and cell cycle progression.
Cell Cycle 11, 922-933
75. Thomas, M., Lange-Grunweller, K., Weirauch, U., Gutsch, D., Aigner, A., Grunweller, A., and
Hartmann, R. K. (2012) The proto-oncogene Pim-1 is a target of miR-33a. Oncogene 31, 918-928
76. Wijesekara, N., Zhang, L. H., Kang, M. H., Abraham, T., Bhattacharjee, A., Warnock, G. L.,
Verchere, C. B., and Hayden, M. R. (2012) miR-33a modulates ABCA1 expression, cholesterol
accumulation, and insulin secretion in pancreatic islets. Diabetes 61, 653-658
77. Najafi-Shoushtari, S. H., Kristo, F., Li, Y., Shioda, T., Cohen, D. E., Gerszten, R. E., and Naar,
A. M. (2010) MicroRNA-33 and the SREBP host genes cooperate to control cholesterol
homeostasis. Science 328, 1566-1569
78. Hardingham, G. E., Cruzalegui, F. H., Chawla, S., and Bading, H. (1998) Mechanisms controlling
gene expression by nuclear calcium signals. Cell Calcium 23, 131-134
79. Svenningsson, P., Nomikos, G. G., and Fredholm, B. B. (1999) The stimulatory action and the
development of tolerance to caffeine is associated with alterations in gene expression in specific
brain regions. The Journal of neuroscience : the official journal of the Society for Neuroscience
19, 4011-4022
80. Dassesse, D., Vanderwinden, J. M., Goldberg, I., Vanderhaeghen, J. J., and Schiffmann, S. N.
(1999) Caffeine-mediated induction of c-fos, zif-268 and arc expression through A1 receptors in
the striatum: different interactions with the dopaminergic system. The European journal of
neuroscience 11, 3101-3114
81. Tang, X., Zheng, D., Hu, P., Zeng, Z., Li, M., Tucker, L., Monahan, R., Resnick, M. B., Liu, M.,
and Ramratnam, B. (2014) Glycogen synthase kinase 3 beta inhibits microRNA-183-96-182
cluster via the beta-Catenin/TCF/LEF-1 pathway in gastric cancer cells. Nucleic Acids Res 42,
2988-98
82. Mihelich, B. L., Khramtsova, E. A., Arva, N., Vaishnav, A., Johnson, D. N., Giangreco, A. A.,
Martens-Uzunova, E., Bagasra, O., Kajdacsy-Balla, A., and Nonn, L. (2011) miR-183-96-182
cluster is overexpressed in prostate tissue and regulates zinc homeostasis in prostate cells. J Biol
Chem 286, 44503-44511
83. Kuranda, K., Leberre, V., Sokol, S., Palamarczyk, G., and Francois, J. (2006) Investigating the
caffeine effects in the yeast Saccharomyces cerevisiae brings new insights into the connection
between TOR, PKC and Ras/cAMP signalling pathways. Molecular microbiology 61, 1147-1166
84. Rao, F. V., Andersen, O. A., Vora, K. A., Demartino, J. A., and van Aalten, D. M. (2005)
Methylxanthine drugs are chitinase inhibitors: investigation of inhibition and binding modes.
Chemistry & biology 12, 973-980
85. Tornaletti, S., Russo, P., Parodi, S., and Pedrini, A. M. (1989) Studies on DNA binding of
caffeine and derivatives: evidence of intercalation by DNA-unwinding experiments. Biochimica
et biophysica acta 1007, 112-115
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Caffeine modulates post-transcriptional regulation of SRSF2
17
Acknowledgments—The authors thank Drs Cyril F. Bourgeois and James Stevenin at Institut de
Je´ne´tique et de Biologie Mole´culaire et Cellulaire, France, for generously providing SRSF2(SC35)
monoclonal antibody. This research was supported by the Microarray Core facility and Bioinformatics
Core facility of the Rutgers Cancer Institute of New Jersey (P30CA072720).
FOOTNOTES
*This work was supported by NCI-R01 CA 122573 (K.W. S.) and NCI-P30CA072720 to the Rutgers
Cancer Institute of New Jersey.
1 To whom correspondence should be addressed: Kathleen W. Scotto. Rutgers Cancer Institute of New
Jersey, Rutgers, the State University of New Jersey, 195 Little Albany Street, New Brunswick, NJ 08903,
USA, Tel: (732) 235-4266; Fax: (732)-235-6596; Email: [email protected]
FIGURE LEGENDS
Figure 1. Methylxanthines induce alternative splicing of KLF6 and increase levels of SRSF2. (A)
RT-PCR analyses on samples treated with methylxanthines (14 mM), including pentoxifylline, caffeine,
theophylline, and isocaffeine, revealed various degrees of SpKLF6 induction following 18hrs of treatment.
(B) Western blotting analyses detected increased levels of SRSF2 in samples treated with pentoxifylline,
caffeine, and theophylline; isocaffeine had a minimal effect. The order of magnitude of SRSF2 increase is
pentoxifylline>caffeine>theophylline>isocaffeine, which coincided with the degree of SpKLF6 AS
induction. (C) Cytotoxicity assays revealed considerable cell death following pentoxifylline treatment
when compared to caffeine, at the effective concentration (14 mM) used to induce maximal alternative
splicing of KLF6.
Figure 2. Caffeine does not change total SRSF2 mRNA levels. (A) Schematic illustration of SRSF2
gene structure. ―E1‖: exon 1; ―E2‖: exon 2; The canonical polyadenylation site is found 2046 nt
downstream of the stop codon, while an alternative polyadenylation site is located at 1341 nt downstream
of the stop codon. (B) Semi-quantitative RT-PCR analysis using primers ―a‖ and ―b‖ on RNA samples
collected from Hela cells with or without caffeine treatment at different time points. No change in SRSF2
total mRNA was observed following caffeine (14 mM) treatment. β2M RNA was used as a loading
control. This assay was repeated at least three times with consistent results. Statistical analyses were
performed using GraphPad Prism 6 software, and significance was determined by the non-parametric t
tests. A P < 0.05 was considered significant. (C) SYBR® real-time RT-PCR analyses confirmed the
results from semi-quantitative RT-PCR, i.e. that no significant change in total SRSF2 mRNA was induced
by caffeine. Error bars represent Mean ± SD.
Figure 3. Caffeine induces rapid changes in the 3’UTR of SRSF2 RNA (A-D). The known Group A
(GA) transcripts as well as the putative Group B, C and D (GB, GC, GD) transcripts are schematically
depicted in the left panels. Semi-quantitative RT-PCR was performed with different primers to query the
existence of both known and putative SRSF2 transcripts in HeLa cells. (A) Analyses using primers ―c‖
and ―d‖ on samples with or without 1hr-caffeine treatment revealed rapid changes in levels of the GA
splice variants (right panel). (B) Analysis using primers ―c‖ and ―f‖ confirmed the existence of putative
GB SRSF2 transcripts that utilize the alternative polyadenylation site instead of the canonical
polyadenylation site. Caffeine treatment (14 mM) had minimal effect on the B1 transcript (inclusion of
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Caffeine modulates post-transcriptional regulation of SRSF2
18
E3) (right panel). Note that primers ―c’ and ―f‖ will also amplify A1. (C) Analysis using primers ―e‖ and
―d‖ confirmed the presence of GC transcripts that utilize the canonical polyadenylation site but select
alternative E2 5’splice sites at positions 1101 nt (C1), 389 nt (C2 and C4), 543 nt (C3), or 60 nt (C5)
downstream of the stop codon. Caffeine induced changes in GC transcripts. GC transcripts are poorly
expressed because they required 5 additional cycles of PCR to be comparably visualized. (D) Analysis
using primers ―e‖ and ―f‖ confirmed novel GD transcripts that utilize the alternative polyadenylation site
as well as alternative E2 5’splice sites. Caffeine (1 hr) induced D1 and D3 at the expense of D2. (E)
Time course analyses of caffeine-induced alternative splicing changes within the 3’UTR among GA and
GD transcripts.
Figure 4. Caffeine (14 mM) does not change the levels of TARDBP and HnRNP F/H to influence
SRSF2 AS at the 3’UTR. Western blotting analysis was performed on samples collected at indicated
time points. Tubulin was used as the internal control.
Figure 5. Inhibition of NMD is not sufficient to increase SRSF2 levels. (A) Cycloheximide-mediated
NMD inhibition only resulted in partial mimics of the effect of caffeine on alternative splicing pattern in
GA and GD transcripts. (B) RNAi reduced the hUpf1 level by ~85%, inhibiting NMD as evidenced by
the accumulation of a PTB1 splice variant (exon 11-skipped PTB1), known to be an NMD target. Note
that caffeine treatment alone (lane 2) also resulted in accumulation of exon 11-skipped PTB1, indicating
some degree of inhibition of NMD by caffeine. (C) The SRSF2 3’UTR splicing pattern induced by
caffeine was only partially reproduced by hUpf1 knockdown-mediated NMD inhibition. (D) Western
blot analysis revealed that hUpf1 RNAi-mediated NMD inhibition was not sufficient to increase levels of
SRSF2 protein. Quantitation was normalized to input control tubulin. Statistical analysis was performed
based on data from at least three experiments. Error bars represent Mean ± SD.
Figure 6. GA and GD SRSF2 variants exhibit different translational efficiencies and caffeine (14
mM) increases their translation. (A) In vitro translation assays detected a higher intrinsic translational
efficiency of A2 and A3 when compared to A1 transcripts. No significant difference was detected among
GD SRSF2 variants by this assay. (B) Cell-based translation assays also indicated that the caffeine-
induced transcripts A2 and A3 were translated at a higher rate as compared to A1. Caffeine increased
translational efficiency of all SRSF2 variants. SRSF2 cDNA in frame with an N-terminal FLAG tag was
tethered with individual SRSF2 3’UTRs and inserted into a mammalian expression plasmid. Co-
transfection assays introduced both SRSF2 and GFP constructs into HeLa cells. Western blot analysis
was utilized to quantitate relative translation efficiency. GFP was used as a control for transfection
efficiency. (C) Real-time RT-PCR analyses of Flag-SRSF2 and GFP mRNA in the cell-based
translational efficiency assay. The relative levels of Flag-SRSF2 mRNA of each SRSF2 variants were
normalized to levels of GFP mRNA from the same cell populations/experimental conditions. Statistical
analysis was performed based on data from at least three repeats using GraphPad PRISM 6 software, non-
parametric t tests. A difference with a P < 0.05 was considered significant. Error bars represent Mean ±
SD.
Figure 7. Caffeine decreases levels of SRSF2-targeting miRs to increase translational efficiency of
certain SRSF2 transcripts. (A) Strategy for identifying putative caffeine-decreased SRSF2-targeting
miRs in HeLa cells (lower, left) and schematic of miR binding sites within the SRSF2 3’UTR (top). Two
candidates were selected (miR-183-5p and miR-33a-5p) and mapped to GA and GD transcripts. (B)
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Caffeine modulates post-transcriptional regulation of SRSF2
19
MicroRNA array analyses revealed caffeine-mediated decreases in the levels of miR-183-5p and miR-
33a-5p (left). This decrease was validated by quantitative RT-PCR (right). The final Ct value was an
average of 5 repeats of each assay, and each assay was repeated three times. A difference with a P < 0.05
was considered significant. (C) The miR-183-5p binding site in the Flag-SRSF2 A1 transcript was
disabled by site-directed mutagenesis (top). Either A-1WT or A1-183Mut was co-transfected with either
miR-183-5p mimics or inhibitor. GFP was included in the transfection cocktail as transfection efficiency
control. Each assay was repeated three times. A difference at P < 0.05 was considered significant. (D)
The miR-33a-5p binding site in Flag-SRSF2 A2 transcript was disabled by site-directed mutagenesis (top).
Either A-2WT or A2-33aMut was co-transfected together with miR-33a-5p mimics or inhibitors. GFP
was included as the transfection efficiency control. (E) Levels of endogenous SRSF2 and alternative
splicing of KLF6 minigene were analyzed after cells were transfected with miR mimics in the presence
of caffeine (left panel) or miR inhibitors in the presence of hSMG1 siRNA to block NMD (right panel).
Non-specific miRs served as controls. Each assay was repeated three times. Statistical analyses were
performed using GraphPad Prism version 6 software, and significance was determined by the non-
parametric t test. A difference with a P < 0.05 was considered significant. Error bars represent Mean ±
SD.
Figure 8. Proposed model illustrating the mechanisms by which caffeine increases SRSF2 protein
expression. Under normal conditions, SRSF2 homeostasis is maintained by a complex interplay of post-
transcriptional mechanisms including microRNA-mediated translation repression (left panel) and an
alternative splicing associated NMD (AS-NMD)--the autoregulatory feedback loop (middle panel).
Translation of the major SRSF2 transcripts A1 and D2 are suppressed by specific miRs binding to the
3’UTR. When SRSF2 levels are increased due to intrinsic or environmental signals, the increased SRSF2
promotes alternative splicing at the 3’UTR, resulting in multiple splice variants such as A2, A3, D1, and
D3. These transcripts are destined for NMD, thereby decreasing the level of SRSF2 mRNA and the
production of SRSF2 protein. Caffeine has two key effects on SRSF2 homeostasis regulation. First,
caffeine decreases the levels of SRSF2-targeting miRs, releasing translational repression of the major
SRSF2 transcripts, allowing a surge of SRSF2 protein synthesis. This increased SRSF2 triggers the
synthesis of splice variants A2, A3, D1, and D3, normally substrates for NMD. However, caffeine also
inhibits NMD, blocking the degradation of SRSF2 transcript variants. Thus, caffeine affects multiple
regulatory mechanisms breaking the negative feedback loop resulting in a sustained increase of SRSF2
protein.
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from
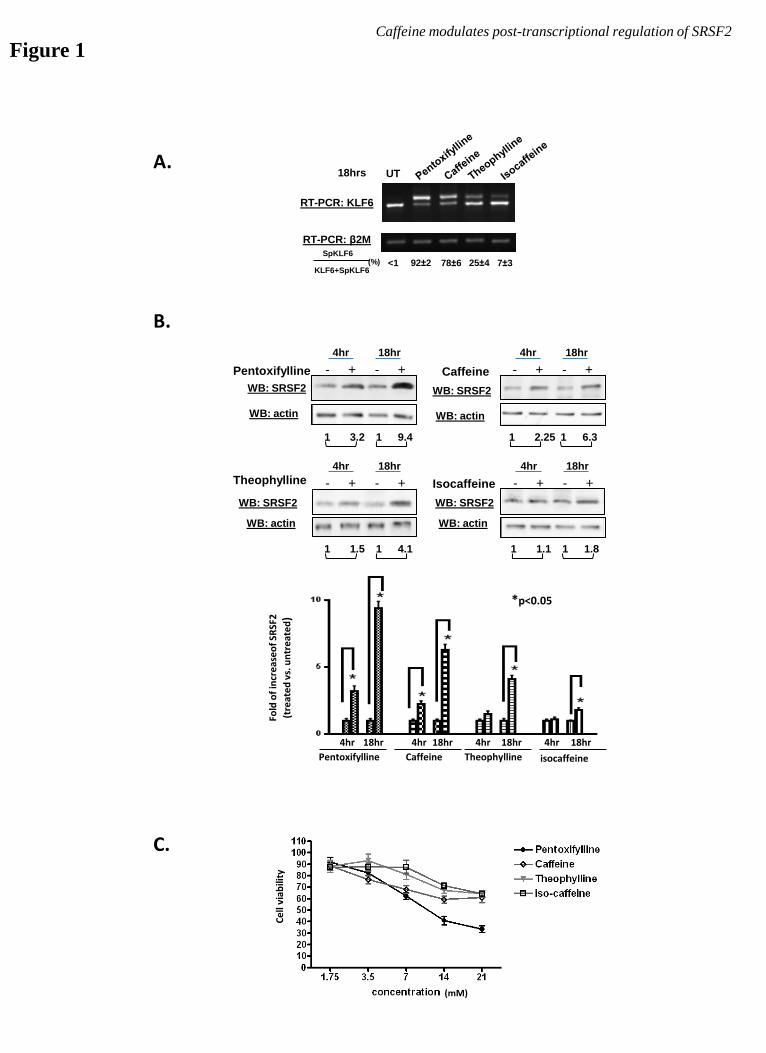
B.
A. UT
RT-PCR: KLF6
RT-PCR: β2M
SpKLF6
KLF6+SpKLF6 <1 (%) 92±2 78±6
18hrs
25±4 7±3
C.
Pentoxifylline Caffeine
Theophylline Isocaffeine
WB: SRSF2
WB: actin
- + - +
4hr 18hr
WB: SRSF2
WB: actin
- + - +
4hr 18hr
- + - +
4hr 18hr
WB: SRSF2
WB: actin
- + - +
4hr 18hr
WB: SRSF2
WB: actin
1 3.2 1 9.4 1 2.25 1 6.3
1 1.5 1 4.1 1 1.1 1 1.8
Figure 1 Caffeine modulates post-transcriptional regulation of SRSF2
Pentoxifylline Caffeine Theophylline isocaffeine
Fo
ld o
f in
cre
ase
of
SRSF
2
(tr
eat
ed
vs.
un
tre
ate
d)
4hr 18hr 4hr 18hr 4hr 18hr 4hr 18hr
*p<0.05
(mM)
Ce
ll vi
abili
ty
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Figure 2
B.
A.
+
8hr
RT-PCR with primers a and b:
- +
1hr
- +
2hr
- +
4hr
- +
6hr
- Caffeine
β2M
SRSF2
-
0hr
Caff +/-
Caffeine modulates post-transcriptional regulation of SRSF2
1 286 389 1003 1101 1600 ATG
STOP E1 E2 E3 E4 E5 I 4/5
Alt. PolyA
1341-nt
Canonical
PolyA
2046-nt a b
C. SYBR Real-time qPCR with primers a and b following universal RT reactions
Re
lati
ve le
vels
of
tota
l SR
SF2
mR
NA
N
orm
aliz
ed
by
leve
ls o
f β
2M
mR
NA
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Figure 3A/3B
A.
B.
RT-PCR with primers c and d :
650bp
1 kb A1
A2
A3
850bp
1-hr Caff - +
Group A transcripts:
A2
A3
A1
2046 1 286 389 1003 1101 1600
c d ATG
STOP E1 E2 E3 E4 E5 I 4/5
RT-PCR with primers c and f:
- + 1-hr Caff
B1
B2+A1
650bp
500bp
400bp
Group B transcripts:
B1
B2
2046 1 389 1003 1101 1600
c
ATG
STOP
Alt. PolyA
1341
f
286
E1 E2 E3 E4 E5 I 4/5
Caffeine modulates post-transcriptional regulation of SRSF2
0%
20%
40%
60%
80%
100%
UT Caff
A1 80% 61%
A2 3% 14%
A3 17% 25%
A1
A2
A3
1-hr
0%
20%
40%
60%
80%
100%
UT Caff
B1 0% 0%
B2+A1 100% 100%
B1
B2+A1
1-hr
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from
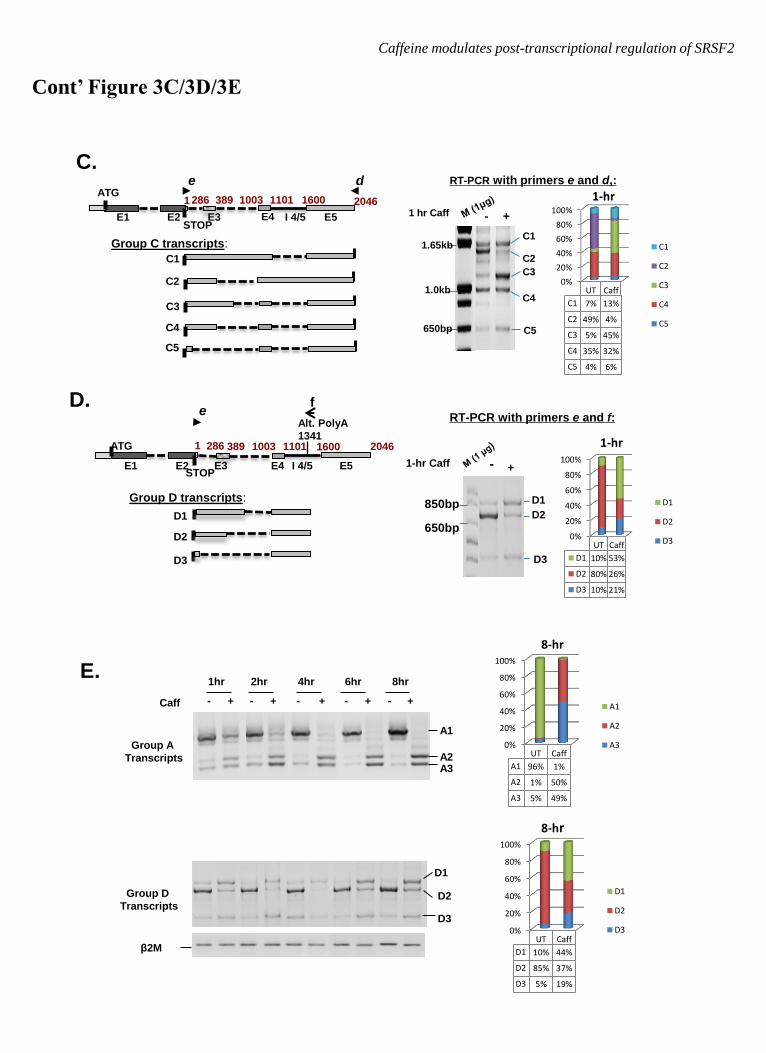
Cont’ Figure 3C/3D/3E
C.
D.
Group D transcripts:
D1
D2
D3
2046 1 389 1003 1101 1600 ATG
STOP
Alt. PolyA
1341 286
e
E1 E2 E3 E4 E5 I 4/5
f
RT-PCR with primers e and d,:
- + 1 hr Caff
C1
C2
C3
C4
C5 650bp
1.0kb
1.65kb Group C transcripts:
C5
C4
C3
C2
C1
2046 1 286 389 1003 1101 1600
d ATG
STOP E1 E2 E3 E4 E5 I 4/5
e
RT-PCR with primers e and f:
E.
Group A
Transcripts
Caff - + - + - + - +
2hr 4hr 6hr 8hr
- +
1hr
A3 A2
A1
β2M
Group D
Transcripts
D1
D2
D3
1-hr Caff -
D1
D2
D3
650bp
850bp
+
Caffeine modulates post-transcriptional regulation of SRSF2
0%
20%
40%
60%
80%
100%
UT Caff
A1 96% 1%
A2 1% 50%
A3 5% 49%
A1
A2
A3
8-hr
0%
20%
40%
60%
80%
100%
UT Caff
D1 10% 44%
D2 85% 37%
D3 5% 19%
D1
D2
D3
8-hr
0%
20%
40%
60%
80%
100%
UT Caff
D1 10% 53%
D2 80% 26%
D3 10% 21%
D1
D2
D3
1-hr
0%
20%
40%
60%
80%
100%
UT Caff
C1 7% 13%
C2 49% 4%
C3 5% 45%
C4 35% 32%
C5 4% 6%
C1
C2
C3
C4
C5
1-hr
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Figure 4
Caffeine modulates post-transcriptional regulation of SRSF2
WB:TARDBP
WB: HnRNP H/F
WB: Tubulin
2-hr 6-hr 24-hr Caff
- + - + - +
1.8 2.0 1.9 1.8 1.7 1.8
1.1 1.3 1.2 1.3 1.4 1.3
6.5 7.1 6.3 6.1 6.6 6.7
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Figure 5
A.
Caffeine modulates post-transcriptional regulation of SRSF2
B.
RT-PCR: Exon 11
skipped PTB1
WB: Upf1
WB: tubulin
SiRNA
Caff
Control
- + -
hUpf1
+
1 1 0.13 0.15
1 2 3 4
24-hr
D.
1
WB: SRSF2
WB: tubulin
1 2 3 4
siRNA
Caff
Control
- + -
hUpf1
+
24-hr
C.
A1
A2
A3
- + Caff
10-hr
+ + CHX
-
-
D1
D2
D3
+
-
1 2 3 4
RT-PCR: GA
A1
A2
A3
RT-PCR: GD
D1
D2
D3
1 2 3 4
SiRNA
Caff
Control
- + -
hUpf1
+
24-hr
Rel
ativ
e le
vels
of
SR
SF2
pro
tein
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Figure 6 Caffeine modulates post-transcriptional regulation of SRSF2
A. In vitro translation assay:
Flag-SRSF2
1 1.8 1.9 1 0.88 0.73
A1 A2 A3 D1 D2 D3
Re
lati
ve q
uan
tita
tio
n
of
Flag
-SR
SF2
Re
lati
ve q
uan
tita
tio
n
of
Flag
-SR
SF2
B. Cell-based translation assay:
A2 A1 A3
- + - + - + D2 D1 D3
- + - + - +
WB: Flag-SRSF2
WB: GFP
Caffeine
Re
lati
ve q
uan
tita
tio
n o
f
Flag
-SR
SF2
No
rmal
ize
d b
y G
FP
Re
lati
ve q
uan
tita
tio
n
of
Flag
-SR
SF2
no
rmal
ize
d b
y G
FP
2.8 4.9 3.6 10.8 4.6 9.8 2.9 7.8 6.4 12.3 3.5 6.5
Rela
tive l
evels
of
Fla
g-S
C35 m
RN
A
no
rmalized
by l
evels
of
GF
P m
RN
A
C.
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Figure 7A/7B
Caffeine modulates post-transcriptional regulation of SRSF2
A.
99 MicroRNAs
down-
regulated
by caffeine
154 MicroRNAs
abundantly
expressed in HeLa
185 MicroRNAs
target for SRSF2
as predicted
miR-183
miR-33a 3' acgUUACGUUGA--UGUU-ACGUg 5' hsa-miR-33a
| | | | | | | | | | | | | | | 1746:5' uaaAAUGC----CUGACAACUGCAc 3' SRSF2
3' ucacuUAAGAUGGUCACGGUAu 5' hsa-miR-183
| | | : | | : | | | | | | | 1159:5’ acuucAUUUUAU---GUGCCAUu 3' SRSF2
2046 1 389 1003 1101 1600 ATG
STOP
Alt. PolyA
1341 286
miR-183
(1159 nt-)
miR-33a
(1746nt-)
Canonical
PolyA
D1
D2
D3
A2
A3
A1
B. ARRAY ANALYSIS
3hr 24hr 3hr 24hr
qRT-PCR
Rela
tiv
e lev
els
of
miR
s
3hr 24hr 3hr 24hr
Rela
tiv
e lev
els
of
miR
s
miR-183-5p
miR-33a-5p miR-33a-5p
miR-183-5p
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

3’ ucacuUAAGAUGGUCACGGUAu 5' hsa-miR-183
| | | : | | : | | | | | | | 1159: 5’ acuucAUUUUAU----GUGCCAUu 3' A1- WT
A1-183Mut
hsa-miR-183 3’ ucacuUAAGAUGGUCACGGUAu 5’
c a gg tUu t U--- acuucAaaU-a c
: :
2046 1 389 1003 1101 1600 ATG
STOP
Alt. PolyA
1341 286
miR-183
(1159 nt-)
Canonical
PolyA
A2
A1
C.
Mimic Inhibitor Mimic Inhibitor NS NS 183 183
A1-WT A1-183Mut
Caffeine modulates post-transcriptional regulation of SRSF2
Figure 7C
A2
NS 183 NS 183
WB: Flag-SRSF2
WB: GFP
Rela
tive
tran
sla
tio
na
l eff
icie
nc
y
NS NS 183 183
A1-WT A1-183Mut
1.0 0.3 1.1 1.1 1.0 3.6 0.9 1.1 1.0 1.0 1.1 1.0
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

3' acgUUACGUUGA-UGUU--ACGUg 5' hsa-miR-33a
| | | | | | | | | | | | | | | 1746: 5' uaaAAUGC----CUGACAACUGCAc 3' A2-WT
A2-33aMut uaaAta c-g----gUGtg t tC ac-gAc 3' acgUUACGUUGA-UGUU-ACGUg 5' hsa-miR-33a
2046 1 389 1003 1101 1600 ATG
STOP
Alt. PolyA
1341 286
miR-33a
(1746nt-)
Canonical
PolyA
D1
A2
Figure 7D/E
Caffeine modulates post-transcriptional regulation of SRSF2
Mimic Inhibitor
NS NS 33a 33a
A2-WT A2-33aMut
NS NS 33a 33a
A2-WT A2-33aMut D1
33a 33a NS NS Mimic Inhibitor
Rela
tive
tran
sla
tio
na
l eff
icie
nc
y
1.0 0.4 1.2 1.2 1.0 2.1 1.2 1.2 1.0 0.9 1.1 1.1
D.
WB: Flag-SRSF2
WB: GFP
E.
*p=0.04
*
Rel
ativ
e q
uan
tity
of
SRSF
2 *p=0.04
*
Rel
ativ
e q
uan
tity
of
SRSF
2
SpKLF6
KLF6
NS NS 183+33a
Caff (14 mM)
miR Mimic
- + +
WB: SRSF2
RT-PCR: KLF6minigene
WB: tubulin
None NS 183+33a
miR Inhibitor
hSMG1 siRNA (blocking NMD)
1.0 3.5 1.5 1.0 0.9 2.8
SpKLF6
KLF6+SpKLF6 (%)
15 89 32 28 23 64
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

5’ 3’
SRSF2 pre-mRNA miR
Normal SRSF2 Homeostasis
5’ 3’
High Levels of SRSF2 Protein
5’ 3’
SRSF2 AS mRNA (A2, A3, D1,D3)
5’ 3’
PTC
5’
3’
NMD
PTC
PTC
PTC
5’ 3’
5’ 3’
SRSF2 major mRNA (A1, D2)
Low Levels of SRSF2 Protein
SRSF2 protein
Caffeine
5’ 3’
5’ 3’
5’ 3’
5’ 3’
SRSF2 AS mRNA (A2, A3, D1,D3)
5’ 3’
5’ 3’
NMD SRSF2 protein
SRSF2 protein
miR
Figure 8
Caffeine modulates post-transcriptional regulation of SRSF2
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Jia Shi, Kirk Pabon and Kathleen W. ScottoMultiple Post-Transcriptional Mechanisms
Methylxanthines Increase Expression of the Splicing Factor SRSF2 by Regulating
published online March 28, 2015J. Biol. Chem.
10.1074/jbc.M114.624254Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2015/04/09/M114.624254.DC1
by guest on August 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Top Related