





![Bacillus Cereus Seminario1[1]](https://static.fdocuments.net/doc/165x107/55cf92fd550346f57b9afb62/bacillus-cereus-seminario11.jpg)

Languages
Pages
Legal


République Algérienne Démocratique et Populaire
Ministère de l’Enseignement Supérieur et de la Recherche Scientifique
UNIVERSITE de TLEMCEN
Faculté des Sciences de la Nature et de la Vie et Sciences de la Terre et de l’Univers
Département de Biologie
LAMABE
MEMOIRE
En vue de l’obtention du
Diplôme de MASTER : En biologie
Option : Microbiologie et contrôle de qualité
Thème
Caractérisation du potentiel PGP de la flore
Bacillus cereus isolée du sol
Présenté par
GHITRI Imane
Soutenu le 04/07/2018, devant le jury composé de :
Présidente Mme BENSALEHF. M.C.B Université de Tlemcen
Encadreur Mme MALEK F. M.C.A Université de Tlemcen
Examinatrice Mme KHOLKHAL W. M.C.B Université de Tlemcen
Année universitaire 2017-2018

Dédicaces
C’est avec une profonde émotion que je dédie ce
mémoire :
A mes chers parents, pour tous leurs sacrifices, leur
amour, leur tendresse, leur soutien et leurs prières tout au
long de mes études,
A mes chères sœurs zineb et wassila pour leurs
encouragements permanents, et leur soutien moral,
A mon chers frères, karim pour leur appui et leur
encouragement,
A toute ma famille pour leur soutien tout au long de
mon parcours universitaire,
Que ce travail soit l’accomplissement de vos vœux tant
allégués, et le fuit de votre soutien infaillible,
Merci d’être toujours là pour moi.
Imane.

Remerciements
En préambule à ce mémoire nous remerciant ALLAH qui nous
aide et nous donne la patience et le courage durant ces langues années
d’étude.
La première personne que nous tenons à remercier est notre
encadrant Mme. MALEK Fadila maitre de conférences A à la Faculté
de SNV / STU (Université de Tlemcen) pour l’orientation, la confiance,
la patience qui ont constitué un apport considérable sans lequel ce
travail n’aurait pas pu être mené au bon port. Qu’elle trouve dans ce
travail un hommage vivant à sa haute personnalité.
On remercie chaleureusement Mme. BENSALAH Fatima,
Maître de conférences classe B, Département de Biologie, de nous
avoir fait l’honneur de présider le jury de ce mémoire. On la remercie
pour sa compréhension. Trouvez ici l’expression de notre profonde
gratitude.
Mon sincères remerciements vont également à Mme. Kholkhal
Wahiba Maitre de conférences classe B au département de Biologie à
l’Université de Tlemcen, d’avoir accepté d’examiner ce travail.
Nous souhaitant adresser nos remerciements les plus sincères
aux personnes qui nous ont apporté leur aide et qui ont contribué à
l’élaboration de ce mémoire ainsi qu’à la réussite de cette formidable
année universitaire, en particulier l’ingénieur du laboratoire
pédagogique N ° 02 Mr YAZID Amine.
Merci pour une autre fois.

Table des matières
1. Introduction générale 01
Synthèse bibliographique
Partie 01 : microflore rhizosphériques 03
1. Caractéristiques du sol 03
2. Présence microbienne dans le sol 03
3. La rhizosphère 04
3.1. Les interactions rhizosphérique 04
Partie 02 : L’effet bénéfique des bactéries du sol et de la rhizosphère 05
1. Introduction 05
2. l’effet phytostimulateur : mécanisme direct 05
2.1. La fixation de l’azote 05
2.2. La solubilisation du phosphate 06
2.3. La solubilisation du potassium 07
2.4. La solubilisation du Zinc 07
2.5. La production de phytohormone 08
2.5.1. L’acide indole acétique 08
2.5.2. Gibbérellines 08
2.5.3. Les cytokinines 09
2.5.4. L’éthylène 09
2.5.4.1. Les interactions de l’éthylène avec autre phytohormones 09
2.5.4.2. L’enzyme 1-aminocycloprone-1-carboxylate (ACC) désaminase 10
2.6. La production de sidérophores 10
2.7. Protection contre le stress abiotique et stress salin 11
3. L’effet phytoprotecteur : mécanismes indirects 11
3.1. Production d’enzymes lytique par les PGPR : chitinase et glucanase 12
3.2. L’antibiose 12
3.3. La compétition 13
3.4. Prédation et parasitisme 13
3.5. Production de cyanure d’hydrogène 13
3.6. L’induction de la résistance de la plante 13
4. La formation de biofilm 14

5. Les PGPR une approche biotechnologique durable 15
Partie 03 : Etude d’une bactérie PGPR Bacillus cereus 16
1. Les principaux caractères phénotypiques du groupe B. cereus 16
2.Taxonomie B. cereus 17
3. Les sept groupes phénotypiques du groupe B. cereus 17
4. cycle de vie B. cereus 19
Matériel et méthodes
1. l’échantillonnage 20
2. Détermination du pH du sol 21
3.1. L’isolement des bactéries du sol
3.2. L’isolement à partir des racines végétales
21
21
4. Les tests d’identification au genre Bacillus 21
4.1. La coloration de Gram 21
4.2. Le teste de la catalase 22
4.3. La mobilité 22
5. L’identification au groupe B. cereus 22
5.1. Croissance sur Chapman 22
5.2. Vérification de l’activité hémolytique 22
5.3. L’identification par galerie Api 20 E 23
6. Etude du pouvoir PGP in vitro 23
6.1. La production d’enzymes 23
6.1.1. Détermination de l’activité protéolytique 23
6.1.2. Détermination de l’activité amylolytique 23
6.2. La production de l’acide indole acétique 23
6.3. La solubilisation du phosphate 24
6.4. L’activité antagoniste 24
6.4.1. L’activité antifongique 24
6.4.2. L’activité antibactérienne 25
6.5. La formation de biofilm 25
6.5.1. La formation de biofilm sur microplaque de titration 25
6.5.1.1. Lecture des microplaques sur lecteur ELISA 25
6.5.2. La formation de biofilm sur des feuilles végétal autoclavé 26
6.5.3. La formation de biofilm sur des feuilles traitées par un désinfectant 26

6.6. Mesure de l’hydrophobicité des spores de B. cereus 26
6.6.1. Préparation des suspension sporales 26
6.6.2. La technique MATH 27
7. Détermination du pouvoir PGP in planta 28
7.1. La culture des graines de persille 28
7.1.1. Matériel végétal 28
7.1.2. L’inoculation du sol des pots 28
7.2. Test du pouvoir antagoniste 28
7.2.1 Préparation de l’inoculum de la souche B. cereus antagoniste 29
7.2.2. Préparation de la suspension de spores de Rhisopus 29
7.2.3. Mode de l’inoculation 29
Résultats et Discussions
1. Caractéristique du sol 30
2. Isolement et identification de Bacillus 30
3. Identification au genre Bacillus 32
4. Identification au groupe B. cereus 32
4.1. Détermination du biotype 34
5.1. La production d’enzymes : L’activité amylolytique et protéolytique 34
5.2. La production de l’acide indole acétique et solubilisation du phosphate 35
5.3. L’activité antagoniste 37
5.4. La formation de biofilm 38
5.5. L’hydrophobicité de surface des spores B. cereus 40
6. Résultats des tests in planta 41
Conclusion 44
Référence bibliographiques 46
Annexe

Liste des abréviations
ACC : 1-aminocyclopropane-1-carboxylate désaminase.
°C : Degré Celsius
EPS : exopolysacharides.
Fe3+ : Fer ferrique
GA : gibbérellines.
g : Gramme.
HCN : cyanure d’hydrogène
IRS : résistance systémique induit.
K : potassium
KSB : bactérie solubilisatrice de potassium
MATH : L’adhésion microbienne au solvant
N2 : Nitrogène
PGPR : plant growthpromotingrhizobactéria
pH : Potentiel d’Hydrogène
TIAC : toxi-infections alimentaire collectives
QS : qurom sensing

Listes des figures
Figure 1 : une représentation de la rhizosphère et leurs interactions. 04
Figure 2 : représentation du cycle d’azote selon Johann Dréo,2006. 06
Figure 3 : Rôle des rhizobactéries favorisant la croissance des plantes. 07
Figure 4 : interactions d’exsudats racinaire des plantes, PGPR contre le
pathogène.
14
Figure 5 : observation de B. cereus au microscope optique, et au
microscope environnemental.
16
Figure 6 : les stades de germination de B. cereus. 19
Figure 7 : les sites d’échantillonnages au niveau du jardin cultivés. 20
Figure 8 : les sites d’échantillonnages au niveau de la montagne. 20
Figure 09 : les microplaques colorées au cristal violet et remplit de
solvants d’extraction.
25
Figure 10 : Aspect des colonies de Bacillus sur gélose au sang (A) et sur
milieu TSA (B et C).
31
Figure 11 : observation macroscopique de la souche racinaire du romarin
(Sr2) (swarming mobilité).
31
Figure 12 : observation microscopique des souches après une coloration
de Gram (G=10x 100).
32
Figure 13 : Mise en évidence de la production de lécithinases sur milieu
Chapman à l’émulsion de jaune d’œuf.
33
Figure 14 : Mise en évidence de l’activité hémolytique des souches testé
(A : gélose au sang neuf).
34
Figure 15 : Résultats d’identification par galerie Api 20E, (1 : S13E3, 2 :
SrE2).
34
Figure 16 : l’activité amynolytique (A et B) et protéolytique (C) des
souches.
35
Figure 17 : la production de AIA et la solubilisation du phosphate par B.
cereus.
35
Figure 18 : Antagonisme de B. cereus à l’égard de souches de
Pseudomonas (A : control positif ; B : une colonisation ; C : inhibition).
37
Figure 19 : Antagonisme antifongique de la souche B. cereus SR1, contre
Rhizopus (1témoin positif et 2 témoin négative).
37

Figure 20 : la formation de biofilm sur microplaque de titration. 38
Figure 21 : Formation de biofilm in-vivo sur des feuilles de plante. 40

Liste des tableaux
Tableau 1 : échelle d’interprétation de l’acidité du sol 33
Tableau 2 : résultats de la mesure du pH du sol 31
Tableau 3 : Résultats d’identification au genre Bacillus isoler à partir
du sol cultivé et non cultivé.
33
Tableau 4 : les résultats d’identification par galerie Api 20E 34
Tableau 5 : Propriétés stimulatrice de la croissance des plantes des
souches isolées.
36
Tableau 6 : Résultats de l’antagonisme antibactérien 37
Tableau 7 : lecture qualitative de la formation de biofilm 39
Tableau 8 : lecture des microplaques de titration 40
Tableau 9 : résultat de l’hydrophobicité des souches 41

Introduction

Introduction
1
Introduction
Le sol est un système vivant, complexe et un réservoir de microorganismes. Il contient
tous les types microbiens, eucaryotes (champignons, algues et protozoaires), et procaryotes
(bactéries et cyanobactéries). En termes de diversité et de densité, sol est un milieu riche et il
n’est pas simplement le support dans lequel les plantes s’enracinent et puisent les éléments
nutritifs indispensables à leur développement.
La rhizosphère est la région du sol située sous les racines des plantes. De nombreuses
interactions sont observées entre la plante, le sol et les microorganismes au niveau des
racines. Parmi les micro-organismes bénéfiques aux plantes, la flore PGPR (plant growth
promoting rhizobacteria) exerce de nombreuses fonctions, améliorant la croissance des
plantes que ce soit directement, par l’augmentation de la fertilité du sol, ou indirectement on
lui conférant une meilleure résistance aux agents pathogènes ainsi qu’aux stress
environnementales (Souza et al., 2015a). Parmi les groupes microbiens du sol qui influant sur
la croissance des plantes, on peut citer : Rhizobium, Pseudomonas, Alcaligenes, Azospirillum,
Klebsiella, Burkholderia, Enterobacter et Azotobacter sp, Serratia aussi que la flore
bactérienne aérobie sporulée qui est largement répandue dans la nature telle que le genre
Bacillus(Kumar et al., 2012).
Le groupe Bacillus cereus, est parmi les bactéries cultivables les plus abondantes du
sol. Ce sont des bactéries à Gram positif (+), sous forme des bâtonnets, et qui forment des
endospores(Dromigny, 2008). Elle présente des capacités remarquables, peuvent synthétiser
une vaste gamme de substances bénéfiques (Stein, 2005), est sont caractérisées par une
grande capacité de formation de biofilm. B. cereus et les autre espèce du genre Bacillus
stimulant la croissance végétale par différents processus tels que la production d'IAA, la
solubilisation du phosphate, fixation de l'azote, et les attributs de la lutte biologique comme la
production de HCN, sidérophore, enzymes hydrolytiques et les antibiotiques (Kumar, et
al.,2009). Ces bactéries présentant de ce fait, un potentiel stimulateur de la croissance des
plantes ou potentiel PGP intéressant pouvant être exploité pour l’amélioration des culture
végétales.
L’objectif de ce travail est d’isoler des souches sporulante à partir de la rhizosphère
d’un sol cultivé, et la rhizosphère de sol a couvert végétal naturel et de les caractérisé par
rapport à leurs propriétés de stimulation de la croissance des plantes en vue de les utiliser in
vivo ; puis l’identifié au groupe B. cereus ; et enfin le conservé.

Introduction
2
La démarche expérimentale suivi comprend :
❖ L’échantillonnage à partir des parcelles des sols étudiées.
❖ Isolement de la flore bactérienne sporulée sur des milieux de culture appropriée.
❖ Identification phénotypique des isolas au genre Bacillus et au groupe B. cereus.
❖ Evaluation du potentiel PGP des souches isolées par l’étude de :
❖ La production de l’acide indole acétique(AIA),
❖ La solubilisation du phosphate,
❖ La phytoprotection par la production des enzymes (amylase et protéase) ; l’activité
antibactérienne et antifongique,
❖ La formation de biofilm, et caractérisation des propriétés de surface des spores.
Pour valoriser l’efficacité de ces biofertilisants nous avons réalisé des tests in planta,
via des cultures en pot de deux légumes : le persille et la tomate.

Synthèse
bibliographique

Synthèse bibliographique
3
Partie 01 : microflores rhizosphériques
1. Caractéristiques du sol
Un sol est un ensemble organisé en différents horizons, évolutif, ou la vie est présente
et dont le matériau est la terre. Il est le lieu de transferts de flux : eau, air, énergie et vie. Le
sol est le milieu d’interactions entre les différentes sphère conceptuelles constituant le globe
terrestre (Girard et al., 2011). La texture du sol se définit par ses proportions relatives en
argile, limon, sable fin, sable grossier. La texture du sol se définit par ses proportions
relatives en argile, limon, sable fin, sable grossier. Le pH, notion importante, mesure l’acidité
ou l’alcalinité du sol. Dans une échelle de 1 à 14, un milieu est neutre quand son pH est de 7.
En dessous, il est acide, au-dessus, il est basique ou alcalin. Les sols calcaires sont en général
basiques, alors que les sols sableux ou très riches en matière organique que s’assoit végétal ou
animale ou microorganismes(Calvet, 2013).
2. Présence microbienne dans le sol
Les microorganismes représentent une biomasse de 1%à 4% de la masse du carbone
organique du sol et de 75% à 90% de la biomasse vivante, cette proportion étant souvent plus
grande pour les sols prairie que pour les sols cultivés. La biodiversité microbienne des sols est
décrite à l’aide de plusieurs méthodes, que ce soient par les méthodes classiques (l’isolement
et l’identification), ou par les méthodes récemment développées (l’extraction quantitative de
l’ADN) (Girard et al., 2011).
L’abondance des microorganismes diminue avec la profondeur, surtout pour les
bactéries, et se diffèrent selon leur localisation. Dans le sol, une grande partie des
microorganismes se trouve au voisinage des racines, dans la rhizosphère, ou les substances
nutritives sont abondantes (Girardet al., 2011). Les bactéries sont les plus abondante d’où un
gramme de sol forestier contient en moyenne environ 4 x 10⁷ bactéries, tandis qu’un gramme
de prairie ou le sol cultivé en contient selon les estimations 2 x 10⁹ bactéries (Prescott et
al.,2013).
Les bactéries représentant plus de 1000 espèces, sont responsables de nombreux
processus : libération des éléments nutritifs à partir de la matière organique et des minéraux
du sol. Oxydation de l’ammonium en nitrates (nitrification - bactéries nitrosomonas et
nitrobacters). Production d’hormones de croissance qui favorisent le développement des

Synthèse bibliographique
4
racines. Compétition avec les micro-organismes pathogènes limitant ainsi les risques de
maladie (symbiotech.over-blog.com, 2017).
3. La rhizosphère
La rhizosphère est le volume de terre directement soumis à l’action des racines. Elle
correspond aux surfaces d’échanges entre les racines et les minéraux de la terre ainsi qu’entre
les racines et les microorganismes, c’est une zone où les relations entre plantes et micro-
organismes sont particulièrement actives (figure 1). Elle constitue une continuité entre le
milieu biologique et le milieu physique (Vacheron et al., 2013a).
3.1. Les interactions rhizosphériques
Dans la rhizosphère on distingue trois zones : l'endorhizosphère, le rhizoplan et le
ectorhizosphère. Dans cet environnement, il y a des interactions entre les racines des plantes,
les microbes, et le sol. Les propriétés physiques et chimiques du sol, peuvent modifient la
population microbienne dans la rhizosphère (Huang et al., 2014).
Figure 1 : une représentation de la rhizosphère et leurs interactions (Badri et al., 2009).
Les interactions de la rhizosphère sont souvent affectées par de nombreux signaux
régulateurs différents de nature chimiques, Communication moléculaire entre l'hôte et
pathogènes, ou la rhizoremediation (la dégradation des polluants par l'augmentation de
nombreux activité métaboliques des microorganismes rhizosphèriques) (Kuiper et al., 2004).
Ces signaux sont très dynamiques et peuvent être modifié. Les interactions entre les racines et
les microorganismes du sol sont souvent spécialisés, et jouent un rôle important dans un
certain nombre de processus écosystémiques vitaux, tels que la séquestration du carbone et les
cycles des éléments nutritifs. Les microorganismes en particulier les bactéries ont un effet
bénéfique sur la croissance des plantes (Badri et al., 2009).

Synthèse bibliographique
5
Partie 02 : l’effet bénéfique des bactéries du sol et de la rhizosphère
1. Introduction
Les rhizobactéries stimulatrices de la croissance des plantes (PGPR : Plant growth-
promoting rhizobacteria) : sont les bactéries du sol qui colonisent les racines des plantes et
améliorent leur croissance directement ou indirectement ; et à leur tour les racines des plantes
sécrètent des métabolites qui peuvent être utilisés comme éléments nutritifs. La stimulation
directe peut inclure la fixation de l'azote atmosphérique, synthèse de diverses phytohormones
et des enzymes, ainsi que la solubilisation des minéraux du sol. Tandis que la stimulation
indirecte qui comprend l'inhibition des phytopathogènes via trois types d'interactions, la
compétition, l’antagonisme et l’induction de la défense de la plante (la résistance systémique
induit : IRS)(Gopalakrishnanet al., 2012; Huang et al., 2014; Osman et al., 2017).
Parmi les groupes microbiens du sol qui s’influx sur la croissance des plantes, on peut
citer : Rhizobium, Pseudomonas, Alcaligenes, Azospirillum, Klebsiella, Burkholderia,
Enterobacter et Azotobacter sp, Serratia, et en particulier le genre Bacillus qui forme des
spores qui lui donne une certaine résistance à la température, et à la concentrations élevées
des produits chimiques, impliquant un potentiel PGPR particulier (Kumar et al., 2012;
Osman et al., 2017).
Le genre Bacillus présente des capacités remarquables, peut synthétiser une vaste
gamme de substances bénéfiques (Stein, 2005). Le genre Bacillus stimule la croissance
végétale par différant processus tels que la production d'IAA, la solubilisation du phosphate,
fixation de l'azote, et les attributs de la lutte biologique comme la production de HCN,
sidérophore, enzymes hydrolytiques et les antibiotiques (Kumar et al., 2009).
2. L’effet phytostimulateur : mécanismes directs
2.1. La fixation de l’azote
L'azote est un élément chimique très disponible dans la nature et indispensable pour le
fonctionnement des organismes. L'atmosphère est constituée d'environ 80% de gaz N2. Mais
la plupart des végétaux ne peuvent l'utiliser sous cette forme inerte. Les plantes absorbent
seulement les formes ioniques solubles dans l'eau qui se trouvent sous les formes nitriques ou
nitrates (NO3-) et ammoniacales ou ammonium (NH4+), au niveau des racines (bio en
ligne.com). Il existe plusieurs types de bactéries fixatrice de l’azote (les bactéries
associatives, symbiotes et libres, vivant dans la rhizosphère) ; mais toutes les bactéries

Synthèse bibliographique
6
fixatrices d’azote comme Azospirillum, Burkholderia, Gluconacetobacter et
Pseudomonasutilisent forme un complexe enzymatique appelé la nitrogénase. Et donc la
fixation associative de l’azote et la promotion de la croissance végétale par les rhizobactéries
se fait sur des organes végétaux spéciaux appelés nodules de racine ou de tige (figure 2). Les
rhizobactéries fixatrice de l’azote sont importantes pour une bonne fertilisation du sol et un
système agricole durable (Pereg et McMillan, 2015; Wdowiak-Wróbel et al., 2017).
Figure 2 : représentation du cycle d’azote selon Johann Dréo,2006.
2.2. La solubilisation du phosphate
Le phosphore est le deuxième élément nutritif le plus important pour les plantes, après
l’azote, et il représente environ 0,2% du poids sec d'une plante. Le phosphate existe dans le
sol sous forme de sels minéraux ou incorporé dans des composés organiques. Bien que ces
composés du phosphore soient abondants dans les sols agricoles, la majorité d'entre eux se
présente sous une forme insoluble ( Oteino et al., 2015;Alori et al., 2017).
Les bactéries solubilisatrices du phosphate comme Rhizobium, Enterbacter, Serratia,
Citrobacter, Proteus, Klebsiella, Pseudomonas, ou encore Bacillus jouent un rôle pour la
nutrition des plantes puisqu’elles ont la capacité à convertir P insoluble au P soluble en
produisant des acides organiques, la chélation et l'échange d'ions ce qui augmente la fertilité
du sol. De plus, les sols contiennent des acides organiques de bas poids moléculaire avec un
ou plusieurs groupes carboxyliques et certains acides. Le rôle des acides organiques dans la
solubilisation du P dépend fortement du sol (Gupta et al., 2012; Panhwar et al., 2013).

Synthèse bibliographique
7
2.3. La solubilisation du potassium
Le potassium (K) fait partie des trois éléments majeurs nécessaires à la croissance des
plantes avec l’azote et le phosphore. Il joue un rôle important dans le métabolisme de l’azote
et la fabrication des protéines et contribue à la résistance aux maladies et à la sécheresse
(Faucher, 2017). Le potassium est disponible sous quatre formes dans le sol, et certains
microorganismes du sol (bactéries, champignons), comme les Enterobacter et Bacillus sont
capables de dissoudre le potassium insoluble (KSB) car elles produisent de l'acide oxalique et
des acides citriques et des enzymes ; parce que la plus grande partie du potassium n'est pas
disponible pour l'absorption par les plantes. L'efficacité de la solubilisation du potassium par
différentes bactéries varie selon la nature des minéraux porteurs de potassium et les conditions
aérobies (Parmar etSindhu, 2013; Setiawati et Mutmainnah, 2016).
2.4. La solubilisation du Zinc
Le zinc est un élément métallique ou métalloïdes dits « traces » car il est présent en
faible concentration dans la croûte terrestre ou les organismes vivants. Il désigne un oligo-
élément indispensable aux processus biologiques, mais toxiques à des teneurs élevées même
dans des concentrations relativement faibles. Le zinc est requis par de nombreuses enzymes
en quantités infime(Habibi et Karimi, 2014). Le zinc est tout jour associé au phosphate pour
être utile par la plante, et les PGPR ont la capacité de le solubiliser (Kumar et al., 2012). La
figure 3 représentent les différents mécanismes de stimulation de la croissance des plantes.
Figure 3 : Rôle des rhizobactéries favorisant la croissance des plantes (Gouda et al.,
2018a).

Synthèse bibliographique
8
2.5. La production de phytohormones
Les plantes sont soumises à divers stress abiotiques, tels que la sécheresse, la
température extrême, la salinité et les métaux lourds. Les phytohormones sont parmi les
régulateurs de croissance les plus importants ; ils sont connus pour avoir un impact important
sur le métabolisme et la croissance des plantes, et en outre, ils jouent un rôle essentiel dans la
stimulation des mécanismes de réponse de la défense des plantes contre les
stress(Egamberdieva et al., 2017).
Les microorganismes associés aux racines, y compris les microorganismes libres,
symbiotiques ou endophytes, peuvent produire divers types de phytohormones ; par exemple
des isolats bactériens provenant de la rhizosphère d'un légume appartenant aux genres
Bacillus, Klebsiella, Leifsoniaet Enterobacter montre une capacité à produire de l'IAA et
d'améliorer la croissance du maïs dans le sol contaminé (Egamberdieva et al., 2017).
2.5.1. L’acide indole acétique
La biosynthèse de l’acide indole acétique « IAA : Indole-3-acetic acid » a été corrélée
avec la stimulation et la prolifération racinaire par les bactéries de la rhizosphère, ce qui a
amélioré l'assimilation des nutriments par les plantes associées (Malik et Sindhu, 2011). Le
groupe B. cereus est également capable de produire des quantités physiologiquement actives
d'auxines (IAA)(M. Usha Rani, 2011). Les bactériens producteurs d'IAA comme les
Enterobacter taylorae, Klebsiella planticola, Alcaligenes faecalis, Xanthomonas maltophila,
Pseudomonas sp. et Flavobacteriumsp ont le potentiel d'interférer avec n'importe lequel de
ces processus en introduisant l'IAA dans le pool d'auxine de la plante (Leveau ,
Lindow,2005).On a constaté que l'effet de l'AIA dépend de la concentration, c'est-à-dire que
de faibles concentrations d'AIA exogènes peuvent favoriser la croissance des racines, tandis
que des concentrations élevées peuvent l’inhiber (Malik et Sindhu, 2011).
2.5.2. Gibbérellines
Gibbérellines sont diterpénoïdes, régulant la croissance des plantes. Elles sont
couramment utilisées dans l'agriculture moderne. Les hormones végétales gibbérellines sont
nécessaires à la germination des graines. Les voies de signalisation de l'hormone peuvent
stimuler la germination de la graine par la libération de la dormance de l’endosperme et
l’expansion de la cellule embryonnaire. En outre, les gibbérellines peuvent influencer la
production de protéines du stress oxydatif, et des métaux lourds(Miransari et Smith,

Synthèse bibliographique
9
2014).Mais très peu de rhizobactéries favorisant la croissance des plantes (PGPR) sont
connues pour produire des gibbérellines (GA)(Kang et al., 2014).
2.5.3. Les cytokinines
Les cytokinines ce sont des hormones végétales dérivées de molécule d'adénine qu’ont
la capacité d’améliorer la division cellulaire des plantes et la résistance des plantes aux
facteurs nuisibles. Ils peuvent également affecter l’activités des cellules du méristèmes dans
les racines, ainsi que la sénescence des feuilles(Chanclud, 2015). De plus, ils sont efficaces
dans la formation de nodule au cours de la mise en place de la symbiose N2-fixing et d'autres
interactions entre la plante et les microbes. Parmi les souches bactériennes productrices de
cytokinines associées aux racines Arthrobacter, Bacillus, Azospirillum et Pseudomonas, qui
peut augmenter la biomasse des racines, ainsi que la teneur en proline dans les tissus végétaux
(Miransari et Smith, 2014 ; Egamberdievaet al., 2017). Les cytokinines sont également
largement utilisées dans la bio-ingénierie et la production agricole pour des cultures
cellulaires dans des bioréacteurs, pour la micropropagation de plantes cultivées, l’obtention
des plantes transgéniques, et pour contrôler le sexe des plantes (Andrabi et al., 2018).
2.5.4. L'éthylène
L'éthylène est une phytohormone gazeuse simple avec de multiples rôles dans la
régulation du métabolisme au niveau moléculaire, cellulaire, et à l'échelle de la plante entière
(Khan et al., 2017). L'éthylène régule les réponses des plantes aux conditions de stress
abiotique, et biotique ; un faible dégagement d'éthylène déclenche des réponses adaptatives et
influence d'autres voies de signalisation hormonale qui induit la résistance des plantes
(Ravanbakhsh et al., 2018).
2.5.4.1 Les interactions de l’éthylène avec autre phytohormones
-Éthylène et Auxine : l'action de l'éthylène sur la croissance des feuilles peut être dépendante
ou indépendante de l'auxine. La coordination hormonale est un aspect important qui régule les
processus de croissance des feuilles. L'auxine induit la production d'éthylène, et de nombreux
effets des auxines exogènes sont, en fait, des réponses à l'éthylène (Iqbal et al., 2017).
-Éthylène et cytokinine : les rapports sur l'interaction entre l'éthylène et la cytokinine sont
rares. Cependant, la cytokinine peut jouer un rôle important dans l'initiation des feuilles grâce
au maintien de la S-adénosyl-méthionine (SAM) ; un métabolite immédiat pour la biosynthèse
de l'éthylène (Iqbal et al., 2017).

Synthèse bibliographique
10
-Ethylène et gibbérellines : Gibbérellines (GA) jouent un rôle dans l'expansion des feuilles, et
l'augmentation des taux d'éthylène induira une gamme d'effets pléiotropes, tels que l'inhibition
de la croissance et la floraison tardive, en plus de la réponse cible ; et donc l’éthylène et
gibbérellines en un effet contradictoire (Iqbal et al., 2017 ; Ravanbakhsh et al., 2018).
-Ethylène et acide abscissique : ont une relation étroite, et les deux hormones ont un effet
inhibiteur de la croissance des plantes (Benschop et al., 2005).
2.5.4.2. L'enzyme 1-aminocyclopropane-1-carboxylate (ACC) désaminase
Un certain nombre de PGPR contient l'enzyme 1-aminocyclopropane-1-carboxylate
(ACC) désaminase, le précurseur de cette enzyme ACC peut cliver l'usine d'éthylène, et ainsi
abaisser le niveau d'éthylène pour qui ne devient pas élevé au point où la croissance initiale
des racines est altérée, dans une plante en développement ou stressée (Penrose et Glick,
2003).
La bactérie Pseudomonas sp, et la levure Cyberlindnerasaturnus étaient les deux premiers
microorganismes signalés pour synthétiser l'ACC désaminase ; par la suite, l'activité de la
désaminase de l'ACC a été observée chez de nombreuses bactéries, à Gram positives et à
Gram négatives, avec différents types de métabolisme. Des souches appartenant au genre
Mesorhizobium, ne sont capables de produire l'enzyme que pendant la phase symbiotique,
lorsqu'elles sont localisées à l'intérieur d'un nodule racinaire (Gamalero and Glick, 2015).
2.6. La production de sidérophores
Le fer est l'un des micronutriments les plus importants pour la croissance microbienne
dans divers environnements. Bien que le fer soit présent sur terre en grande quantité, il est
insoluble. Donc les microorganismes ont développé un système efficace d'acquisition de Fe
des « sidérophores » (liaison Fe (III) de bas poids moléculaire composés (200e2000 Da), pour
acquérir des quantités suffisantes d’élément(Złoch et al., 2016). Le mécanisme du
sidérophore consiste d'abord à se lier à une forme ferrique de fer et à former un complexe de
sidérophore-fer qui pénètre dans les cellules par l'intermédiaire de récepteurs sidérophores
spécifiques présents dans la membrane cellulaire. Pour les bactéries gram-positives, le
transport du complexe sidérophore-fer est réalisé par l'implication de protéines sidérophores
irrévocables, de perméases et d'ATPases. Alors que dans les bactéries gram-négatives, le
mécanisme de transport est très différent en raison de leur structure membranaire complexe.
Ici, ils transfèrent le complexe sidérophore-fer à travers une protéine de liaison périplasmique

Synthèse bibliographique
11
et une protéine membranaire cytoplasmique correspondant à un transporteur de cassette se
liant à l'ATP (S. kumaret al., 2017).
De nombreuses bactéries associées à des plantes et qui peuvent synthétiser des
sidérophores tel que Bradyrhizobium, Pseudomonas, Rhizobium, Streptomyces, Serratia,
Azospirillum, et Bacillus, à donner aux plantes un avantage concurrentiel dans la colonisation
des tissus végétaux en les aidant à exclure d'autres micro-organismes de la même niche
écologique, et peuvent également contribuer à la suppression de pathogènes dans la
rhizosphère. De plus, il a été montré que les bactéries productrices de sidérophores résistant
aux métaux, et jouent un rôle important dans la croissance et la survie de la plante, ainsi que
l’induction de la résistance des plantes(Pereg et McMillan, 2015; Złoch et al., 2016).
2.7. Protection contre le stress abiotique et stress salin
Le stress d'aridité causé « par la sécheresse, la température élevée, la salinité qui est
l'un des graves problèmes environnementaux qui causent le stress osmotique », sont les stress
abiotiques les plus dominant qui limitent la croissance et la productivité des plantes(Kumar
et Verma, 2018). La tolérance à ce stress est multigénique et quantifiable et comprend
l'accumulation de certains métabolites de stress, tels que les poly-sucres, la proline, la glycine-
bétaïne, l'acide abscissique et la régulation positive de la synthèse d'antioxydants
enzymatiques et non enzymatiques, comme la superoxyde dismutase (SOD), la catalase
(CAT), l'ascorbate peroxydase (APX), la glutathion réductase, l'acide ascorbique, l'α-
tocophérol et le glutathion. L'application des microorganismes producteurs de ces métabolites
tel que certains rhizobactéries stimulatrices de la croissance des plantes (PGP) et certains sont
des champignons mycorhiziens favorisent la croissance des plantes, dans de telles conditions.
Il offre un moyen économiquement fascinant et écologique de protéger les plantes contre les
conditions de stress ( Tank et Saraf, 2009 ; Gouda et al., 2018).
3. L’effet phytoprotecteur : mécanismes indirects
Contrairement aux mécanismes directs des PGPR, les mécanismes indirects se
produisant à l'extérieur du corps de la plante permettant de réduire ou arrêter les effets
nuisibles des phytopathogènes en produisant différents signaux. Les plantes répondent aux
signaux envoyés par la bactérie soit en produisant des substances antagonistes ou en induisant
une résistance aux agents pathogènes. Pour le stress biotique, les rhizobactéries jouent un rôle
dans l’induction de la résistance(Tabassum et al., 2017).

Synthèse bibliographique
12
L'environnement rhizosphérique et antagoniste bactérien peuvent être mises en évidence
selon(Beneduzi et al., 2012), via les mécanismes suivants :
-La synthèse des enzymes hydrolytiques, telles que les chitinases, les glucanases, les
protéases, et les lipases, qui peuvent lyser les cellules fongiques pathogènes.
-la concurrence pour les nutriments et la colonisation appropriée des niches à la surface
radiculaire.
-la production de sidérophores et des antibiotiques.
3.1. Production d’enzymes lytique par PGPR : chitinase et de glucanase
L'un des principaux mécanismes utilisés par les agents de lutte biologique contre les
pathogènes du sol consiste la production d'enzymes dégradant les parois cellulaires, telles que
la β-1,3-glucanase, la chitinase, la cellulase et la protéase sécrétées par les souches de
biocontrôle de PGPR, exersant un effet inhibiteur direct sur la croissance des hyphes des
pathogènes fongiques en dégradant leur paroi cellulaire. Les agents de lutte biologique ayant
les potentiels des activités chitinolytiques, la chitine est convertie principalement en chitosan,
(RaafatandSahl, 2009), cette activité comprennent B. licheniformis, B. cereus, B. circulans,
B. subtilis et B. thuringiensis. Parmi les bactéries à Gram négatif possèdent des activités
chitinolytiques en distingue, Serratia marcescens, Enterobacter agglomerans, Pseudomonas
aeruginosaet P. fluorescens(Goswamiet al., 2016a).
3.2. L’antibiose
Les bactéries de la rhizosphères sont sous-utilisées comme des agents de lutte
biologique, sur tous les bactéries qui tuent les pathogènes par l'antagonisme, et les bactéries
qui produisent des antibiotiques, et des biosurfactants. La synthèse des antibiotiques par les
bactéries et un biocontrôle négatif par rapport à la plante (Lugtenberget Kamilova, 2009;
Mota et al., 2017). Les antibiotiques sont des toxines de bas poids moléculaire produites par
les communautés bactériennes qui peuvent tuer et / ou empoisonner d'autres microbes.
Cependant, les biosurfactants sont des composés amphiphiles qui peuvent causer une fuite des
composant cellulaire, en interagissant avec les membranes de micro-organismes nuisibles.
Plusieurs antibiotiques et des toxines ont été identifiées à partir de la population bactérienne
vivant dans la rhizosphère(Tabassum et al., 2017). Le groupe B. cereus produit les
antibiotiques zwittermicine A (aminopolyol) et kanosamine (aminoglycoside), contribuent à
la lutte biologique(Beneduzi et al., 2012).

Synthèse bibliographique
13
3.3. La compétition
La compétition est l'interaction indirecte dans laquelle les pathogènes présents dans la
rhizosphère sont en concurrence les uns avec les autres pour la nourriture et pour l’occupation
physique du site. L’environnement de la rhizosphère est souvent limité en nutriments, et pour
que les microbes réussissent à coloniser et à survivre, ils occupent les sites où l'eau et les
nutriments sont assimilables, et contenant du carbone facilement disponible. Ainsi, pour que
les microbes puissent survivre dans un tel environnement, ils sécrètent des sidérophores qui
ont une forte affinité avec le fer et rendent moins disponible aux pathogènes et inhibent leur
croissance dans la rhizosphère (Tabassum et al., 2017).
3.4. Prédation et parasitisme
La prédation et le parasitisme, le mécanisme majeur de biocontrôle utilisé par certaines
espèces (fongiques) de Trichoderma, mais ce mécanisme existe actuellement chez les
bactéries (Lugtenberg et Kamilova, 2009).
3.5. Production de cyanure d'hydrogène
Les micro-organismes phytopathogènes constituent une menace majeure et chronique
pour l'agriculture durable et la stabilité des écosystèmes dans le monde entier. Le cyanure
d'hydrogène (HCN) est le composé volatil le plus puissant produit par de nombreuses
rhizobactéries favorisant la croissance des plantes, et possédant une activité contre les
phytopathogènes. Le cyanure d'hydrogène est généralement produit par le genre Bacillus, y
compris B. amyloliquefaciens, B. cereus, B. mycoides, B. pumilus, B. sphaericus, et B.
subtilis(Ahemad et Kibret, 2014a).
3.6. L’induction de la résistance de la plante
Les PGPR fournissent une stratégie alternative pour protéger la plante contre les
maladies via la résistance systématique induite (ISR). Le processus par lequel le traitement
des plantes par PGPR déclenche la défense de l'hôte se traduisent par la réduction de la
sévérité ou l'incidence de la maladie causée par des pathogènes qui sont séparés dans l'espace
est appelé ISR (Goswami et al., 2016b). Cette « immunisation » de la plante, ne provoquant
pas de symptômes visibles de maladie. L’ISR s'est avérée être induite par des
microorganismes variés et plus particulièrement par des rhizobactéries. Celles-ci incluent des
bactéries Gram+ comme Bacillus pumilus, Bacillus subtilis, Bacillus amyloquefaciens et
Bacillus thuringiensis ou des bactéries Gram- qui sont les plus étudiées dans le contexte de
Synthèse bibliographique
14
l'ISR, bien que des entérobactéries. Ainsi, l'induction des rhizobactéries produisent un signal
qui se propage de façon systémique à l'intérieur de la plante et augmente la défense, et la
capacité des tissus d’éloigner l'infection causer par les agents pathogènes (figure 04),
(Jourdan et al, 2008 ; Benhamou etRey, 2012).
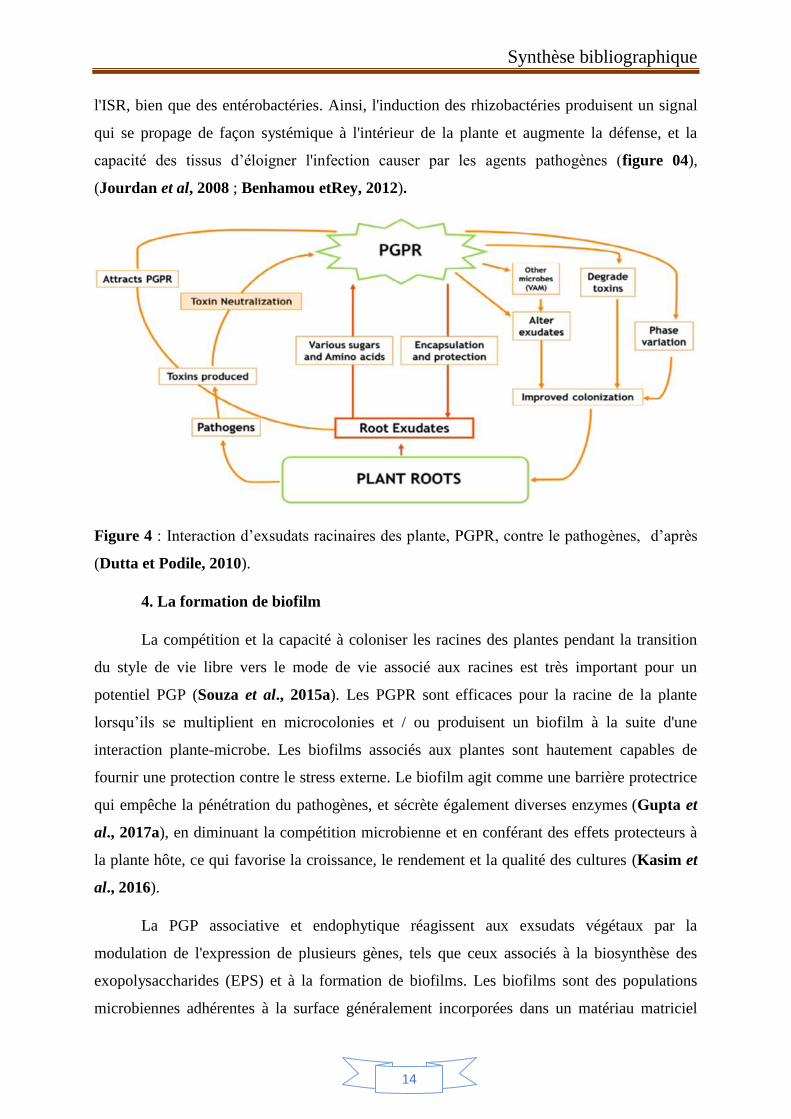
Figure 4 : Interaction d’exsudats racinaires des plante, PGPR, contre le pathogènes, d’après
(Dutta et Podile, 2010).
4. La formation de biofilm
La compétition et la capacité à coloniser les racines des plantes pendant la transition
du style de vie libre vers le mode de vie associé aux racines est très important pour un
potentiel PGP (Souza et al., 2015a). Les PGPR sont efficaces pour la racine de la plante
lorsqu’ils se multiplient en microcolonies et / ou produisent un biofilm à la suite d'une
interaction plante-microbe. Les biofilms associés aux plantes sont hautement capables de
fournir une protection contre le stress externe. Le biofilm agit comme une barrière protectrice
qui empêche la pénétration du pathogènes, et sécrète également diverses enzymes (Gupta et
al., 2017a), en diminuant la compétition microbienne et en conférant des effets protecteurs à
la plante hôte, ce qui favorise la croissance, le rendement et la qualité des cultures (Kasim et
al., 2016).
La PGP associative et endophytique réagissent aux exsudats végétaux par la
modulation de l'expression de plusieurs gènes, tels que ceux associés à la biosynthèse des
exopolysaccharides (EPS) et à la formation de biofilms. Les biofilms sont des populations
microbiennes adhérentes à la surface généralement incorporées dans un matériau matriciel

Synthèse bibliographique
15
autoproduit(Souza et al., 2015a).La plupart des espèces bactériennes utilisent la détection du
quorum pour coordonner leur expression du gène selon la densité locale de leur population
(QS), (Seneviratne et al., 2010). Cette communication moléculaire intense entourant les
racines fournit une large gamme d'interactions microbe-microbe, rendant cet environnement
hautement compétitif parmi les bactéries du sol. La compétition microbienne et les activités
comprennent, par exemple, la motilité, l'attachement, la croissance, la résistance au stress, la
production de métabolites secondaires, et la détection du quorum (Souza et al., 2015b).
5. Les PGPR : une approche biotechnologique durable
Les PGPR en été classer selon leurs activités fonctionnelles comme des biofertilisants
(augmentation de la disponibilité des nutriments pour la plante), des phytostimulateurs
(promotion de la croissance des plantes), des rhizorémédiateurs, et les biopesticides (contrôle
des maladies, principalement par la production d'antibiotiques et de métabolites
antifongiques) (Ahemad et Kibret, 2014).
Les Biostimulants végétaux, parfois appelés biostimulants agricoles, sont une
classification diverse des substances qui peut être ajouté à l'environnement autour d’une
plante « la rhizosphère » et ont des effets positifs sur la croissance des plantes, et aussi sur la
tolérance au stress abiotique et biotique. Les Biostimulants ne sont pas des nutriments, mais
ils facilitent l'adoption des nutriments ou contribuer de manière bénéfique à la croissance ou
la résistance au stress, c’est pour cette raison les PGPR sont utilisé dans les nouvelles
approches biotechnologiques(Van Oosten et al., 2017).
Les PGPR producteurs de biofilm sont utilisées comme des biofertilisants du sol afin
de réduire les effets néfastes du stress sur les plantes(Gupta et al., 2017b)Les produits
biologiques à base des PGPR qui forme des spores bactériennes en une durée de conservation
qui varie de 1 à 3 ans, et un inconvénient de temps de revenir au stade métabolique actif d'une
cellule végétative après leur application. Cependant, il est probable que les souches de
contrôle biologique les plus efficaces agissent via plusieurs mécanismes (Kumar et al.,
2012).

Synthèse bibliographique
16
Partie 03 : Etude d’une bactérie PGPR : Bacillus cereus
1. Les Principaux caractères phénotypiques du groupe B. cereus
B. cereus est considérées comme une bactérie du sol, et comprend des espèces
étroitement apparentées, qui ont une très grande importance pour l’homme. B. cereus est un
groupe de bactéries à Gram positif (+), sous forme des bâtonnets assez volumineux, mesurant
de 1à1,8 µm de diamètre par 4à8 µm de long, mobile avec une ciliature péritriche, aérobies ou
aroanaérobies facultatifs, et qui forme des endospores(Dromigny, 2008). Les spores de B.
cereus ont une forme non déformante, en règle générale ovales, et en position subterminale,
ou parfois en position paracentrale à raison d'une spore par cellule. En effet les endospores et
la formation de biofilm en considéré comme des propriétés de survie et de résistance envers
les conditions défavorables. B. cereus est capable de croitre à une large gamme de
température allant de 4°C à 55°C avec une température optimale de croissance comprise entre
30°C et 40°C (Bottone, 2010; Lin et al., 2017).
Ce groupe de bactéries appartenant au genre Bacillus ayant une grande importance
économique et clinique ; c’est un contaminons fréquent des produits alimentaires, mais il peut
vivre en saprophyte dans le sol(Jensen et al., 2003). Le groupe B. cereus est un groupe de
microorganisme très homogène, réuni sous le nom de « B. cereus sensu lato » (Malek et al.,
2013)qui se diffère des autres Bacille de morphologie similaire, par l’activité hémolytique, et
la production de lécithinase sur milieu Mossel, ainsi l'incapacité à fermenter le mannitol,
selon la norme AFNOR (2005).
Figure 5 : observation de B. cereus au microscope optique, et au microscope électronique
environnemental.

Synthèse bibliographique
17
2. Taxonomie du groupe B. cereus selon (Dromigny, 2008).
Domène des eubactéries
Phylum des firmicutes
La classe des Bacilli
L’ordre des bacillales
La famille des Bacillaceae
Le genre Bacillus
Le groupe B. cereus
3. Les sept groupes phylogénétiques du groupe B. cereus
Le groupe B. cereus comprend six espèces qui ne sont pas considérées comme des
espèces génomiques distinctes: B. cereus, B. thuringiensis, B. anthracis, B. mycoides, B.
pseudomycoides et B. weihenstephanensi, et une nouvel espèce Bacillus cytotoxicus(Jensen et
al., 2003),et répartis entre sept groupes phylogénétiques selon la classification de
(Guinebertière et al.,2008).
Le groupe I regroupe les souches de l'espèce pseudomycoides, dont la température de
croissance est comprise entre 10°C et 43°C. Les souches psychrotrophes de B. cereus et B.
thuringiensis du groupe II ne présentent pas de croissance en dessous de 7°C et peuvent
croître jusqu'à 40°C. Les groupes III, IV et V regroupent les souches de B. cereus sensu
stricto et B. thuringiensis, les plus mésophiles du groupe B. cereus. Le caractère mésophile est
de plus en plus marqué du groupe V vers le groupe III qui regroupe les souches de B.
anthracis. Le groupe VI contient les souches psychrotrophes de B. weihenstephanensis et B.
mycoides qui sont capables de croître dès 5°C mais sont incapables de croître au-delà de
37°C. Le groupe VII contient les souches les plus thermophiles, dont la température de
croissance varie de 20°C à 50°C ( Malek et al., 2013; Brillard et al., 2015).
B. cereus sensu stricto : est fréquemment isolée à partir du sol, de plantes et de
l'intestin de différents animaux, et elle est capable de coloniser différents habitats. Elle est
capable de synthétiser des enzymes extracellulaires d'intérêt industriel, des toxines et des
antibiotiques. B. cereus est un agent de TIAC (toxi-infections alimentaires collectives), sa
pathogénicité provient de sa capacité à produire plusieurs facteurs de virulence tels que des
phospholipases, des entérotoxines et des toxines émétiques (céreulides) entrainant des toxi-
infections alimentaires (Granum etLund, 1997; Kotiranta et al., 2000).

Synthèse bibliographique
18
Bacillus thuringiensis : est utilisées dans la lutte biologique au sa qualité de
biopesticides. Les bactéries synthétisent des toxines cristallines actives contre diverses
espèces d'insectes nuisibles des ordres Lepidoptera, Coleoptera et Diptera. De plus, les
insecticides contenant des spores et des cristaux de B. thuringiensis révèlent une activité pour
les hémocytes des insectes (Cinar et al., 2007).
Bacillus anthtacis: est constituée de grands bacilles à germes Gram positif aérobie, et
le seul pathogène obligatoire du genre Bacillus, le responsable de l’anthrax, et qui tire son
nom du mot grec charbon. B anthracis, en raison de sa capacité à provoquer des escarres
cutanées noires semblables au charbon. Bien que B.anthracis a été signalé comme sensibles à
un nombre limité d'agents antimicrobiens, elle est caractérisée par la résistance à la
pénicilline, l'érythromycine et les quinolones a été noté(Koehler, 2009; Lin et al., 2017).
Bacillus mycoides et pseudomycoides: est un nombre de B.cereus le moins étudié car il
est le moins nocif pour homme, par rapport à B. anthracis et B. cereus ou l’agent insecticide
B. thuringiensis. B. mycoides a été étudié pour la première fois par le naturaliste russe Gause
(1939), qui étudie la genèse des colonies typiques de ces bacilles, dont la croissance sur
milieu gélosé forme une forme rhizoïdale ressemblant aux hyphes fongiques, groupés en
faisceaux courbes à droite ou à gauche(Turchi et al., 2012). B. mycoides a montré une
capacité à produire des biosurfactants (Composé actif biologique des détergents qui met en
solution ou en suspension les souillures adhérentes à des surfaces. Il favorise l'humidification,
la solubilisation et l'émulsion de composés organiques), dans une gamme de pH de 5 à 9 et à
des températures variables de 20°C à 50 ° C, qui lui donne un grand intérêt biotechnologique
(Romanowska et al., 2015).
Bacillus weihenstephanensis : se distingue par sa capacité à croître à 7 ° C mais pas à
43 ° C, et à différant pH, et aussi des séquences de signatures spécifiques de Bacillus
weihenstephanensis ont été trouvées dans certaines souches de B. cereus et de B. mycoides
suggérant une psychotolérance. Elle peut produire une toxine émétique (cereulide). Cette
toxine est stable à la chaleur et aux acides qui provoque une intoxication alimentaire
accompagnée de vomissements ( Soufiane et Côté, 2013; Guérin et al., 2017).
Bacillus cytotoxicus : une nouvelle espèce qu’a été placé dans le groupe phylogénique
VII, et caractérisé par sa toxicité élevée, sa thermotolérance particulier (croissance à 50 ° C),
et son apparition relativement rare in (Malek et al., 2013).

Synthèse bibliographique
19
4. Cycle de vie B. cereus
B. cereus l’espèce saprophyte du sol est capable de produire des spores qui peuvent
survivre pendant des années dans leur état de dormance dans le sol. Les spores dormantes
retour à leur cycle de croissance végétative dans un procédé comprenant la germination et
l'excroissance qui convertit la spore germée dans une cellule en croissance (figure 6). Il existe
un certain nombre de types différents d'agents qui déclenchent la germination des spores. Les
spores germées ont une résistance à la chaleur inférieure au spores dormantes et peuvent être
inactivées à des températures plus basses (LøVdal et al., 2011; Setlow, 2014).
Figure 6 : les stades de germination de B. cereus.

Matériel et
méthodes

Matériel et méthodes
20
Cette étude expérimentale a été réalisé au niveau du laboratoire de pédagogie N°2 du
département de biologie, faculté SNV-STU université de Tlemcen.
1. L’échantillonnage
Les échantillons de sol sont prélevés dans la région de Ain el Houtz, les sites de
prélèvements sont divisés en deux grands types de sol : le sol d’un jardin cultivé, et le sol
d’une montagne recouvert d’une végétation naturelle.
Deux échantillons sont prélevés à partir du jardin au niveau de la rhizosphère d’un
citronnier quatre saison arrosé par l’eau de rinçage des fruits et des légumes domestiques, et à
partir de la rhizosphère d’un romarin traité par des cendres d’origine végétale) (figure7).
Trois autres échantillons ont été prélevés à partir du sol de montagne au niveau de la
rhizosphère d’un sapin associé à une plante aromatique du genre Thymus, et au niveau de la
rhizosphère des mauvaises herbes au-dessous d’un arbre ainsi que de celle d’un amandier)
(figure8). Les prélèvements ont été réalisés à l’aide d’une cuillère préalablement stérilisée, la
technique consiste à enfoncer verticalement la cuillère dans le sol au niveau de la rhizosphère
des végétaux, à une profondeur de 10 à 20 cm. Les échantillons sont placés dans des boites de
pétri recouvertes avec du papier aluminium et conservées à 4°C.
Figure 07 : Les sites d’échantillonnage au niveau du jardin cultivés (Ain El Houtz 2018).
Figure 08 : Les sites d’échantillonnage au niveau de la montagne (Ain El Houtz 2018).

Matériel et méthodes
21
2. Détermination du pH du sol
Le pH du sol a été déterminé à l’aide d’un pH-mètre. On fait dissoudre 10g de sol dans
25ml de l’eau distillée, ce mélange doit être homogénéisé par un agitateur magnétique
pendant 3-5 min, laissé décanter puis le pH du surnageant est mesuré. La lecture des résultats
se fait par comparaison aux valeurs de référence (Baize, 2000).
3.1. L’isolement des bactéries du sol
L’isolement des bactéries du sol a été réalisé sur gélose au sang et sur milieu TSA
(Tryptone-soja-agar), après avoir fait un traitement thermique (80°C pendant 10min) pour
détruire les formes végétatives et sélectionner les endospores. Une série de dilution (de la
solution mère 1g dans 9 ml de l’eau physiologique, jusqu’ à la dilution 10-³) a été réalisée,
puis 100µl de l’inoculum sont ensemencés sur gélose au sang, ou bien TSA par étalement à
l’aide d’un râteau.
3.2. L’isolement à partir des racines végétales
-Les racines sont lavées trois fois avec de l’eau distillée stérile, séchées devant un bec
bunsen puis prélevées à l’aide d’une pince stérile et déposée sur milieu TSA.
4. les tests d’identifications au genre Bacillus
Les quatre critères recherchés, et jugés suffisants selon Gordon et al., (1973) et Claus
et Berkely, (1986) in (Bouali et al., 2016) sont vérifiées pour affirmer l’appartenance au
genre Bacillus :
- Réaction positive à la coloration de Gram
- formation de l’endospore par observation microscopique, grâce à sa réfringence à la lumière,
- la mobilité à l’état vital
- La catalase positive
4.1. La coloration de Gram
Des frottis de culture jeune sont réalisés et soumis à la coloration de Gram classique.
Les frottis colorés sont observés au microscope optique au grossissement x1000.

Matériel et méthodes
22
4.2. Le test de catalase
La catalase est une enzyme antioxydante qui convertit le peroxyde d’hydrogène en eau
et molécule d’oxygène.
H2O2 H2O + ½ O2
La technique de mise en évidence l’enzyme « catalase », qui caractérise les aérobies et les
aéro-anaérobies facultatives consiste à déposer une goutte de l’eau oxygénée a 10 volume sur
une petite quantité d’une culture bactérienne jeune. Le résultat positif se traduit par un
dégagement de gaz.
4.3. La mobilité
Le milieu mannitol mobilité, est utilisé pour tester la mobilité des souches. La mobilité
est un caractère morphologique du genre Bacillus. L’ensemencement du milieu a été effectué
par une piqûre centrale à l’aide d’une pipette pasteur, l’incubation se fait à 30°C pendant 24h.
La mobilité se traduit par une culture envahissante du milieu, alors que les souches immobiles
ne se développent qu’au niveau de la piqure.
5. Identification au groupe B. cereus
5.1. Croissance sur Chapman
La mise en évidence de l’absence de fermentation du mannitol et la présence de
l’enzyme lécithinase ont été réalisées sur milieux Chapman additionné d’émulsion de jaune
d’œuf, en remplacement du milieu Mossel. Le milieu Chapman permet en outre la mise en
évidence de l’halotolérance.
Le groupe B. cereus est caractérisé par l’absence de fermentation du mannitol, et la
production de lécithinase, pour tester les souches isolées, on réalise un ensemencement par
strie sur milieu Chapman supplémenté d’une émulation de jaune d’œuf. L’incubation à 30°C
pondant 24h et 72h. La croissance sur Chapman se traduit par la présence des colonies, et la
production de lécithinase se traduit par la formation d’un halo clair tous autour de la strie.
5.2. Vérification de l’activité hémolytique
La vérification de l’activité hémolytique a été réalisée sur gélose au sang par
ensemencement d’une strie, pour tester l’activité hémolytique des souches racinaires, et les
souches isolées sur TSA.

Matériel et méthodes
23
5.3. L’identification par galerie Api 20E
L’identification du groupe B. cereus par galerie Api 20E concerne que les douze
premiers caractères. Le mode d’inoculation de la galerie Api est comme suit :
-Préparation de la galerie
-préparation de l’inoculum : à l’aide d’une pipette pasteur, on prend une ou deux colonies à
partir d’une boite gélosée de 24h, et on prépare des suspensions dans 5 ml de l’eau
physiologique stérile, dont la DO est comprise entre 0.6 et 0.8.
-l’inoculation de la galerie selon le protocole galerie Api 20E.
6. Etude du pouvoir PGP in vitro
Les propriétés stimulatrices de la croissance des plantes étudiées comprennent la
production d’enzymes extracellulaires (proétases, et amylases), la prodcution d’hormone de
croissance (acide indole acétique), la solubilisation du phosphate, l’activité antagoniste et le
pouvoir de colonisation du système racinaire par l’évaluation de la formation de biofilm.
6.1. La production d’enzymes
6.1.1. Détermination de l’activité protéolytique
L’activité protéolytique a été a été étudiée sur gélose nutritive contenant 5% de lait
écrémé. Après 48 heures d’incubation à 30°C, la présence de cette activité est détectée par un
halo clair autour de la strie indiquant l’hydrolyse de la caséine, par contre un résultat négatif
ne montre aucune zone d’hydrolyse autour de la culture (Marchand et al., 2009).
6.1.2. Détermination de l’activité amylolytique
La mise en évidence l’activité amylolytique est réalisée sur gélose nutritif contenant
1% de l’amidon soluble. L’ensemencement par strie a été réalisé, puis l’incubation à 30°C.
Après l’obtention d’une bonne culture la gélose a été recouverte d’une solution de Lugol.
L’hydrolyse de l’amidon se traduit par l’apparition d’une zone claire autour de la colonie, par
contre un résultat négatif se traduit par une couleur brune autour de la culture (Gupta et al.,
2004).
6.2. La production de l’acide indole acétique
La production de l’acide indole acétique est mise en évidence sur milieu TSB
additionné de 1g/l du tryptophane comme seule source de carbone, selon la technique de

Matériel et méthodes
24
(Etesami et al., 2015). Le milieu est inoculé par 100µl d’une culture bactérienne de 24h et
incubé pendant 72h à 30°C. Après incubation, les cultures sont centrifugées à 5000 rpm/
20min. Un volume de 1ml de surnageant est mélangé à 1ml du réactif de Salkowski (50 ml,
35% de l’acide sulfurique H₂SO₄, 1 ml de la solution du chlorure férique 0.5 M FeCl₃). La
présence de l’acide indole acétique est indiquée par l’apparition de la couleur rose (Goswami
et al., 2014).
6.3. La solubilisation du phosphate
La capacité de solubilisation des phosphates est réalisée selon la technique décrite
par(Beneduzi et al., 2008) sur milieu glucose TGEA (tryptone glucose extractagare). Les
deux autres solutions ont été préparées séparément ; On dissout d'abord 5 g de K2HP04 dans
50 ml d'eau distillée et ensuite 10 g de CaCl2 dans 100 ml d'eau distillée. Ces deux solutions
ont été ajoutées à 1 L de TGEA. Un volume de 10μl de chaque culture bactérienne de 24h est
déposé à la surface du milieu puis incubé à 30°C pendant 5 à 7 jours, la solubilisation des
phosphates se traduit par un halo clair autour de la colonie(Bouali et al., 2016).
6.4. L’activité antagoniste
6.4.1. L’activité antifongique
L’activité antifongique est testée sur milieu PDA (Potato-Dextrose- Agar), et sur
milieu TSA. Les agents pathogènes (champignon), sont isolées : à partir d’une feuille d’un
citronnier ayant une apparence de brûlée. Le champignon responsable de cette altération, est
identifié par un examen macroscopique(Souza et al., 2017).
Le test de l’antagonisme est réalisé comme suit :
-La souche de B. cereus est ensemencée par une strie centrale sur milieu PDA ou TSA, et
incubée à 30°C pendant 48h.
-A l’issue de ce délai, on dépose à une distance de 2,5 cm, un disque de champignon mature
de 7 jours ; puis on ré-incube à 30°C pendant 7à 10 jours.
Une boite témoin est ensemencée par le disque de champignon seul sans la strie de l’agent
antagoniste.

Matériel et méthodes
25
6.4.2. L’activité antibactérienne
L’activité antibactérienne est également un des caractères PGP phyto-protecteur, le
test est réalisé sur milieu TSA, et vérifié sur milieu Muller Hinton, par ensemencement d’une
strie centrale de la souche antagoniste et, incubation à 30°C pondant 24h. Après incubation on
ensemence par strie les 3 souches (Staphylococcus, Klebsiella, Pseudomonas aeruginosa),
obtenus par Mr. Belyagoubi, et on réincube à 30°C pondant 24h. Le résultat positif se traduit
par l’apparition d’une zone d’inhibition.
6.5. La formation de biofilm
6.5.1. La formation de biofilm sur microplaque de titration : 96 puits
La formation de biofilm a été réalisée selon la technique d’O’toole (O’Toole, 2011)
-sur milieu TSB on prépare une culture bactérienne à partir d’un milieu gélosé, incubation à
30°Cpendant 24h. Après incubation on mesure la DO qui doit être entre 0,6-0,8.
-les puits sont remplit avec 150µl de l’inoculum, puis incuber pendant 24h, à quatre
température différente 4°C, 30°C, 37°C et 42°C.
-Après incubation, les puits sont vidés, et rincés 3 fois à l’eau pour éliminer les cellules libres
ou faiblement attachées. Les microplaques sont ensuite, séchées en position renversée puis
colorées avec une solution de cristal violet à 0,1% pendant 30 mn, puis lavées avec de l’eau
pour éliminer les résidus du colorant.
6.5.1.1. Lecture des microplaques sur lecteur ELISA
La lecture des microplaques a pour but de quantifier la formation de biofilm ; pour
réaliser cette lecture, on remplit les puits avec de l’acide acétique à 33%.
Figure 9 : les microplaques colorées au cristal violet et rempli du solvant d’extraction.
30°C 37 °C 45 ° C

Matériel et méthodes
26
6.5.2. La formation de biofilm sur des feuilles végétales autoclavées
- Des feuilles d’un oranger sain sont envelopper dans du papier d’aluminium, et autoclavées
pour les stériliser.
-Les feuilles ainsi stérilisées sont placées dans des boites de pétri ; puis inoculées par 150µl
d’une pré-culture de 18h dans le milieu TSB, après avoir délimité la zone de l’inoculum.
-Les feuilles bactérisées sont incubées à 30°C pendant 24h. Après incubation, les feuilles sont
lavées laver les feuilles avec de l’EDS, puis écouvillonnées. L’écouvillon contaminé est
plongé dans un bouillon nutritif, bien vortexé pour détacher les bactéries de l’écouvillon ; puis
prendre 150µl du BN et ensemencer une gélose nutritive par étalement à l’aide d’un râteau ; et
incuber à 30°C pendant 24h.
6.5.3. La formation de biofilm sur feuilles traitées par un désinfectant
Pour former le biofilm sur un tissu végétal le protocole suivant est adopté :
-couper les feuilles en carré de 2cm², et plonger les dans un premier temps dans un bécher qui
contient de l’alcool, pour les désinfecter, puis dans l’eau distillé stérile, pour les rincer.
-récupérer les feuilles à l’aide d’une pince stérile, les mettre dans une boite de pétri, et les
laisser séchés devant un bec bunsen.
-les feuilles sont ensemencées avec 100µl de l’inoculum de 24h, ajusté à une DO comprise
entre 06-08, puis elles sont incubées à 30°C pondent 20h.
-écouvillonner les feuilles, puis plonger l’écouvillon dans un bouillon nutritif, bien agiter au
vortex pour détacher les bactéries de l’écouvillon ; prendre 100µl du bouillon nutritif et
ensemencer une gélose nutritive par étalement à l’aide d’un râteau, et enfin incuber à 30°C
pendant 24h.
6.6. Mesure de l’hydrophobicité des spores de B. cereus
6.6.1. Préparation des suspensions sporales
L’hydrophobicité est une propriété importante de la surface des spores de B. cereus,
qui a une relation étroite avec leur pouvoir d’adhésion aux surfaces inertes (Malek et al.,
2013). Pour cela des suspensions sporales sont préparées dans l’eau distillée stérile. Les
souches testées sont d’abord ensemencées sur gélose nutritive par des stries serrées, pour
avoir un tapis microbien dense et homogène. L’incubation se fait à 30°C jusqu’à ce que le

Matériel et méthodes
27
taux de sporulation atteigne les 90% (le temps d’incubation se varie de 5 à 7
jours)(Simmonds et al., 2003). Le taux de sporulation est vérifié par un examen
microscopique de frottis colorés à la fushine. La récupération des spores se fait comme suit :
devant un bec bunsen, inonder la boite de pétri avec de l’eau distiller stérile, racler la culture
bactérienne à l’aide d’un râteau et récupérer la suspension dans un bécher stérile, puis verser
le contenu dans un tube stérile.
Les suspensions sporales ainsi obtenues sont centrifugées trois fois à 2000 g pendant
20 min, le surnageant est remplacé à chaque fois avec de l’eau distillé stérile. Ces opérations
sont terminées par un traitement thermique à 80°C pendant 10min, qui permet d’éliminer les
formes végétatives. Les suspensions sporales sont conservées à 4 °C jusqu’à leur utilisation.
6.6.2. La technique MATH
L’adhésion microbienne au solvant ou technique MATH : microbial adhesion to
solvent(Faille et al., 2002), a pour but de déterminer l’hydrophobicité de la surface des
cellules bactériennes, par mesure de leur affinité à l’hexadécane. Le protocole utilisé pour
mesure cette propriété de surface chez les spores des souches de B. cereus, est décrit dans
Malek (2013).
-dans un tube à hémolyse : prendre 2ml de la suspension sporale dont la DO à 590 nm est
comprise entre 0.6-0.8 et lui rajouter 400µl du solvant apolaire l’hexadécane.
-le mélange est vortexé, pendant de 30 secondes, pour obtenir une émulsion.
-Le mélange subit une décantation pendant 15 min pour l'obtention de deux phases séparées,
et une mesure de la densité optique de la phase aqueuse est réalisée dans un colorimètre à
590nm.
-Le pourcentage d'adhésion au solvant est alors donné par la relation suivante :
A (%) = (Di-Df) / fDi x 100
Di: représente la densité optique initiale,
Df: représente la densité optique de la phase aqueuse après décantation finale,
A(%) : 20˂hydrophyle˂40-60 hydrophobe˃60˂ hautement hydrophobe.

Matériel et méthodes
28
7. Détermination du pouvoir PGP in planta
La détermination du potentiel PGP chez les souches de B. cereus isolés, a permis de
choisir les souches les plus actives pour les essais in-vivo, sur des cultures en pot (la souche
isolée à partir de l’échantillon 3 « S11E3 », et la souche isolée à partir de l’échantillon 5 «
S23E5 »). Le persil a été choisi comme matériel végétal à tester.
7.1. La culture des graines de persil
7.1.1. Matériel végétal
Les graines de persil sont désinfectées et semées selon la technique décrite par
Daayf,1993, avec quelques modifications. Les graines sont lavées à l’hydrochlorite de sodium
à 15% pendant 30 secondes, puis elles sont placées dans les suspensions sporales des souches
à tester (une souche productrice de l’acide indole acétique, et une souche qui solubilise le
phosphate). Les graines ainsi contaminées sont placées dans des pots contenant de la terre
tamisée et stérilisée à 120°C pendant 1 heure, deux fois à 24 heures d’intervalle. La
croissance se fait à la température ambiante. Un témoin négatif consiste en l’utilisation de la
même culture dans les conditions de croissance normale.
7.1.2. L’inoculation du sol des pots
Les pots semés ont été arrosés par une culture de 3 jours de B. cereus dans le milieu
TSB contenant de tryptophane à 1g/L.
7.2. Test du pouvoir antagoniste
L’antagonisme a été étudié chez une souche de B. cereus isolés des racines de la
citronnier (Sr01), contre une souche fongique appartenant au genre Rhizopus isolé au niveau
des feuilles de la même arbre.
7.2.1. Matériel végétal
Les graines de tomate sont désinfectées pendant 30 secondes avec de l’hydrochlorite
de sodium à 15%, semées dans le sol stérile, et la croissance se fait à la température ambiante,
(Daayf, 1993) in Bensaleh.

Matériel et méthodes
29
7.2.2. Préparation de l’inoculum de la souche B. cereus antagoniste
La souche de B. cereus est ensemencée dans le milieu TSB pour avoir une culture jeune de 24
h. la charge microbienne est déterminée par un colorimètre et la densité optique à 590 nm est
ajustée à 0.8 (Aleysinghe,2007).
7.2.3. Préparation de la suspension de spores de Rhisopus
La suspension est récupérée selon la technique décrite par Essalmani et Lahloo, 2003,
avec des modifications adaptées. La suspension des spores est obtenue après grattage de la
surface de la culture du champignon de 15 jours sur milieu PDA, et mélangé à l’eau distillé.
Après agitation, le mélange est filtré sur papier filtre.
7.2.4. Mode de l’inoculation
Après apparition des quatre premières feuilles vraies, la jeune plantule est
délicatement déterrée puis trempées pendant 30 minutes dans une suspension de Bacillus
selon Stall et walter (1966) ; elle est ensuite repiquée dans son pot. Après 48h, la plantule
traitées et trempée dans une suspension de Rhisopus pendant 30minutes. Une plantule témoin
positif trempée dans l’inoculum pathogène sans la bactérisassions bénéfique de la souche de
B. cereus. Une plantule témoin négatif trempée dans de l’eau distillé

Résultats et
discussions

Résultats et discussion
30
1. Caractéristiques du sol
Le pH du sol est une propriété chimique qui influe sur les microorganismes. Les
résultats de la mesure du pH des sols étudiés sont donnés dans le tableau 2. Les valeurs de pH
des sol cultivés et non cultivés varient entre 8 et 8,33 à l’exception du sol de l’échantillon de
la rhizosphère du romarin dont le pH est relativement acide (6,20) et qui a une apparence
jaunâtre et contient des cendres d’origine végétale. Les résultats obtenus ont été comparés à
un tableau de référence (tableau 1).
Tableau 1 : Echelle d’interprétation de l’acidité du sol (Baize, 2000).
PH Sol
˂3,5 Hyper acide
3,5-5 Très acide
5-6,5 Acide
6,5-7,5 Neutre
7,5-8,5 Basique
˃8,5 Très basique
Tableau 2 : Résultats de la mesure du pH du sol
L’échantillon PH Apparence Interprétation
ECJ1 8,20 Maron Basique
E R J2 6,20 Jaunâtre Relativement acide
ESTM3 8,00 Maron pale Basique
EHM4 8,30 Maron foncer Basique
EAM5 8,33 Maron foncer Basique
E : échantillon S : sapin H : herbe
C : citronnier T : Thymus A : amende
J : jardin M : montagne
2. Isolement et identification de souches Bacillus
Les techniques et les milieux de culture utilisés, le milieu TSA et la gélose au sang ont
permis l’isolement de 28 souches bactériennes à partir des sols rhizosphériques et des racines
végétales (tableau 3). Sept souches ont été isolées à partir de la rhizosphère du citronnier,

Résultats et discussion
31
trois souches à partir de la rhizosphère du romarin, neuf souches à partir de la rhizosphère de
sapin, deux souches à partir de la rhizosphère des mauvaises herbes, et deux à partir de celle
de l’amandier.
Les souches se développant en biofilm sur le système racinaire, sont isolées par simple
diffusion à partir des racines végétales préalablement lavées et déposées à la surface de la
gélose TSA (figure 11 c). De chaque échantillon on a pu isoler une souche formant des
colonies dont l’aspect est homogène, miel, et envahissant (Figure 11). L’isolement des
souches sur gélose au sang permet de mettre en évidence le pouvoir hémolytique des souches
appartenant au genre Bacillus notamment le groupe B. cereus.
Figure 10 : Aspect des colonies de Bacillus sur gélose au sang (A) et sur milieu TSA (B et
C).
La souche racinaire 2 isolé du biofilm de la racine de romarin a montré, sur gélose au
sang un aspect cultural particulier, caractérisé par l’envahissement de la gélose, suggérant un
mode de mobilité de type swarmingou déplacement en groupe.
SrE02.
Figure 11 : observation macroscopique de la souche racinaire du romarin (Sr2) (swarming
mobilité).
Les bactéries sporuleés appartenant à Bacillus et au genres proches sont des
représentants importants du sol, ils font de la flore PGP (Ding et al., 2016)en particulier le
groupe B. cereus.
Résultats et discussion
32
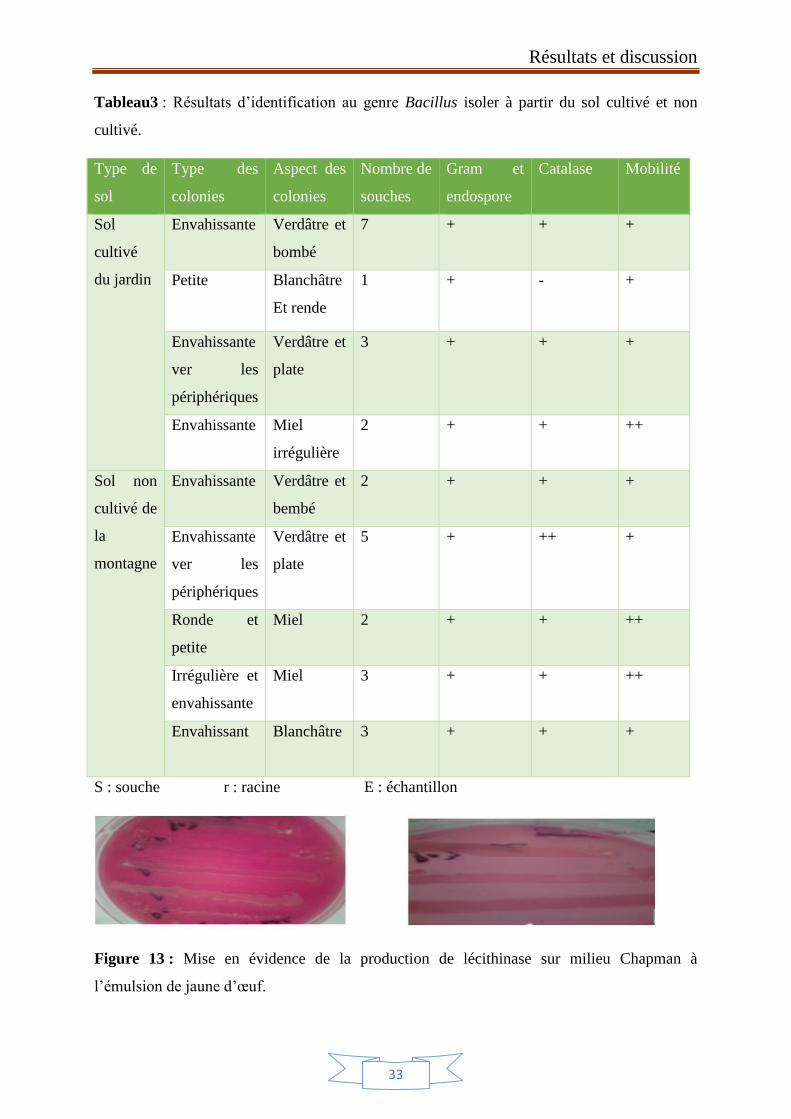
3. Identification au genre Bacillus
Les résultats des tests d’identification sont résumés dans les tableau 3, qui contient les
critères d’orientation vers le genre Bacillus à savoir une réaction positive à la coloration de
Gram, la présence des endospores, la présence de la catalase, et la mobilité.
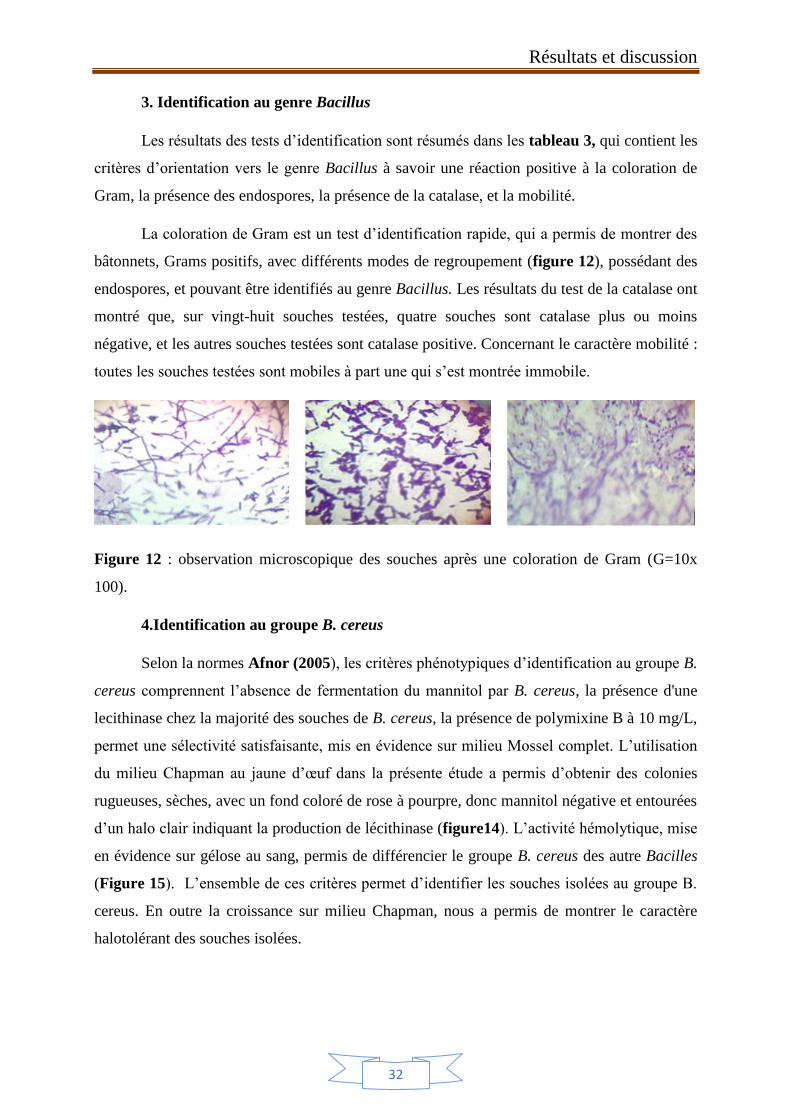
La coloration de Gram est un test d’identification rapide, qui a permis de montrer des
bâtonnets, Grams positifs, avec différents modes de regroupement (figure 12), possédant des
endospores, et pouvant être identifiés au genre Bacillus. Les résultats du test de la catalase ont
montré que, sur vingt-huit souches testées, quatre souches sont catalase plus ou moins
négative, et les autres souches testées sont catalase positive. Concernant le caractère mobilité :
toutes les souches testées sont mobiles à part une qui s’est montrée immobile.
Figure 12 : observation microscopique des souches après une coloration de Gram (G=10x
100).
4.Identification au groupe B. cereus
Selon la normes Afnor (2005), les critères phénotypiques d’identification au groupe B.
cereus comprennent l’absence de fermentation du mannitol par B. cereus, la présence d'une
lecithinase chez la majorité des souches de B. cereus, la présence de polymixine B à 10 mg/L,
permet une sélectivité satisfaisante, mis en évidence sur milieu Mossel complet. L’utilisation
du milieu Chapman au jaune d’œuf dans la présente étude a permis d’obtenir des colonies
rugueuses, sèches, avec un fond coloré de rose à pourpre, donc mannitol négative et entourées
d’un halo clair indiquant la production de lécithinase (figure14). L’activité hémolytique, mise
en évidence sur gélose au sang, permis de différencier le groupe B. cereus des autre Bacilles
(Figure 15). L’ensemble de ces critères permet d’identifier les souches isolées au groupe B.
cereus. En outre la croissance sur milieu Chapman, nous a permis de montrer le caractère
halotolérant des souches isolées.
Résultats et discussion
33
Tableau3 : Résultats d’identification au genre Bacillus isoler à partir du sol cultivé et non
cultivé.
Type de
sol
Type des
colonies
Aspect des
colonies
Nombre de
souches
Gram et
endospore
Catalase Mobilité
Sol
cultivé
du jardin
Envahissante Verdâtre et
bombé
7 + + +
Petite Blanchâtre
Et rende
1 + - +
Envahissante
ver les
périphériques
Verdâtre et
plate
3 + + +
Envahissante Miel
irrégulière
2 + + ++
Sol non
cultivé de
la
montagne
Envahissante Verdâtre et
bembé
2 + + +
Envahissante
ver les
périphériques
Verdâtre et
plate
5 + ++ +
Ronde et
petite
Miel 2 + + ++
Irrégulière et
envahissante
Miel 3 + + ++
Envahissant Blanchâtre
3 + + +
S : souche r : racine E : échantillon
Figure 13 : Mise en évidence de la production de lécithinase sur milieu Chapman à
l’émulsion de jaune d’œuf.

Résultats et discussion
34
Figure 14 : Mise en évidence de l’activité hémolytique des souches testé (A : gélose au sang
neuf).
4.1. Détermination du biotype
L’identification au sein du groupe B. cereus par la galerie Api 20 E, ne prend en
considération que les douze premiers caractères. Les résultats obtenus ont été résumé dans le
tableau 4 ; et figure15. D’après ces résultats, la souche isolée à partir du sol, (S11E3) est
identifiée comme B. thuringensis, et la souche isolée à partir de la racine du romarin (Sr2) a
été identifiée à l’espèce Enterobacter aerogènes.
Tableau 4 : les résultats d’identification par galerie Api 20E
Figure 15 : Résultats d’identification par galerie Api 20E, (1 : S13E3, 2 : SrE2).
5.1. La production d’enzyme : l’activité amylolytique et protéolytique
Les souches isolées ont montré une bonne capacité protéolytique, et amylolytique,
(figure 16), les caractéristiques des sites de prélèvement (sol 1 du jardin, arrosé par l’eau de
rinçage des fruit et légumes, le sol 2 qui contient de la matière organique, et les sols 3,4 et 5
qui contiennent les feuilles des arbres morte en décomposition, ont influencé les caractères
métaboliques des souches. Les résultats sont en conformité avec S.Umayaparvathi et al.,
(2013), qui ont trouvé que les souches B. cereus sont caractérisées par une grande activité
protéolytique.

Résultats et discussion
35
Figure 16 : l’activité amynolytique (A et B) et protéolytique (C) des souches.
L’ensemble des résultats portant sur les caractères stimulateurs de la croissance des
plantes sont résumés dans le tableau 5.
5.2. La production de l’acide indole acétique et la solubilisation du phosphate
Les résultats obtenus montrent que le groupe B. cereus a la capacité de produire de
l’AIA (une phytohormone qui accélère la croissance des racines). En effet neuf souches sur
vingt-huit produisent cette phytohormone, dans des proportions différentes (figure 17).
Juniore et al., (2015) ; ont montré que le genre Bacillus a la capacité de produire l’AIA
synthétisé à partir du tryptophane. L’effet de l'IAA dépend de la concentration. A faibles
concentrations, l’IAA exogène peut favoriser la croissance des racines, tandis que de fortes
concentrations peuvent l’inhiber (Deepak,2011). La solubilisation du phosphate par les
souches B. cereus isolées se traduit par la formation d’un halo claire au tour de la culture
bactérienne, comme montré par(Maheswar et Sathiyavani, 2012). Tandis que, comme le
montre la figure 18, la production de l’AIA, se traduit par l’apparition d’une coloration rose
due à la réaction de l’acide indole acétique synthétisé par la souche avec le réactif de
Salkowski(Goswamiet al., 2014).
Figure 17 : la production de AIA et la solubilisation du phosphate par B. cereus.
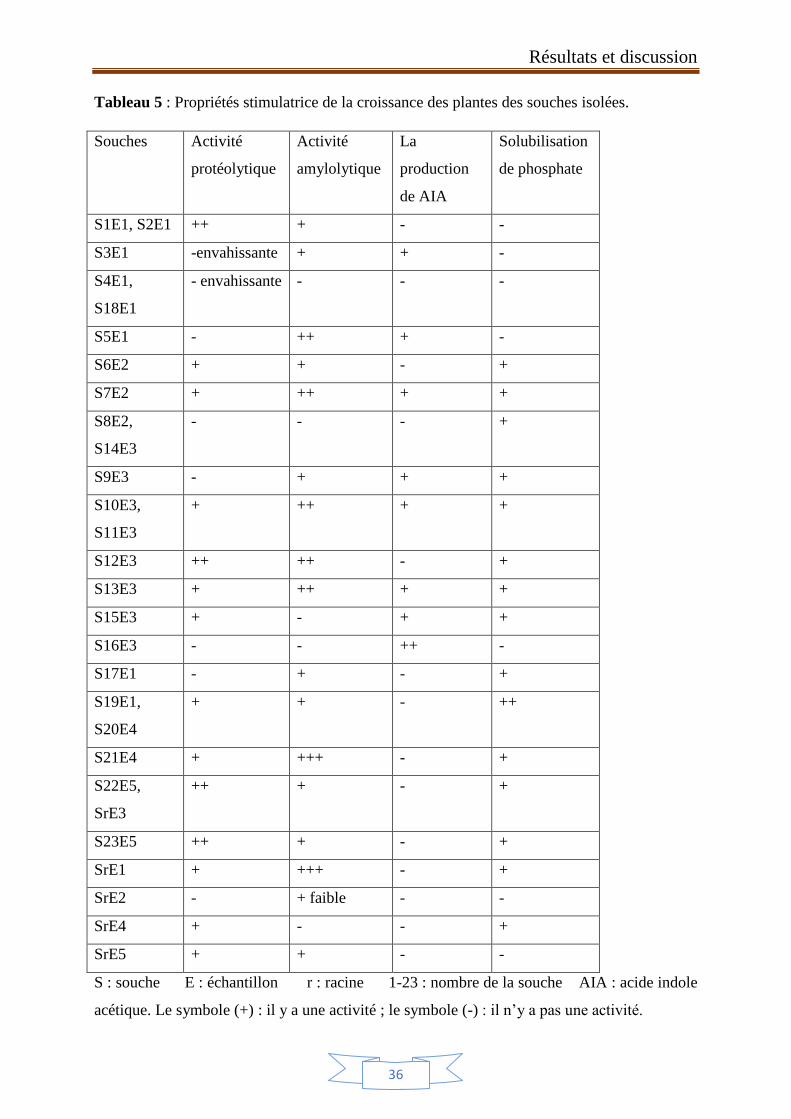
Résultats et discussion
36
Tableau 5 : Propriétés stimulatrice de la croissance des plantes des souches isolées.
Souches Activité
protéolytique
Activité
amylolytique
La
production
de AIA
Solubilisation
de phosphate
S1E1, S2E1 ++ + - -
S3E1 -envahissante + + -
S4E1,
S18E1
- envahissante - - -
S5E1 - ++ + -
S6E2 + + - +
S7E2 + ++ + +
S8E2,
S14E3
- - - +
S9E3 - + + +
S10E3,
S11E3
+ ++ + +
S12E3 ++ ++ - +
S13E3 + ++ + +
S15E3 + - + +
S16E3 - - ++ -
S17E1 - + - +
S19E1,
S20E4
+ + - ++
S21E4 + +++ - +
S22E5,
SrE3
++ + - +
S23E5 ++ + - +
SrE1 + +++ - +
SrE2 - + faible - -
SrE4 + - - +
SrE5 + + - -
S : souche E : échantillon r : racine 1-23 : nombre de la souche AIA : acide indole
acétique. Le symbole (+) : il y a une activité ; le symbole (-) : il n’y a pas une activité.
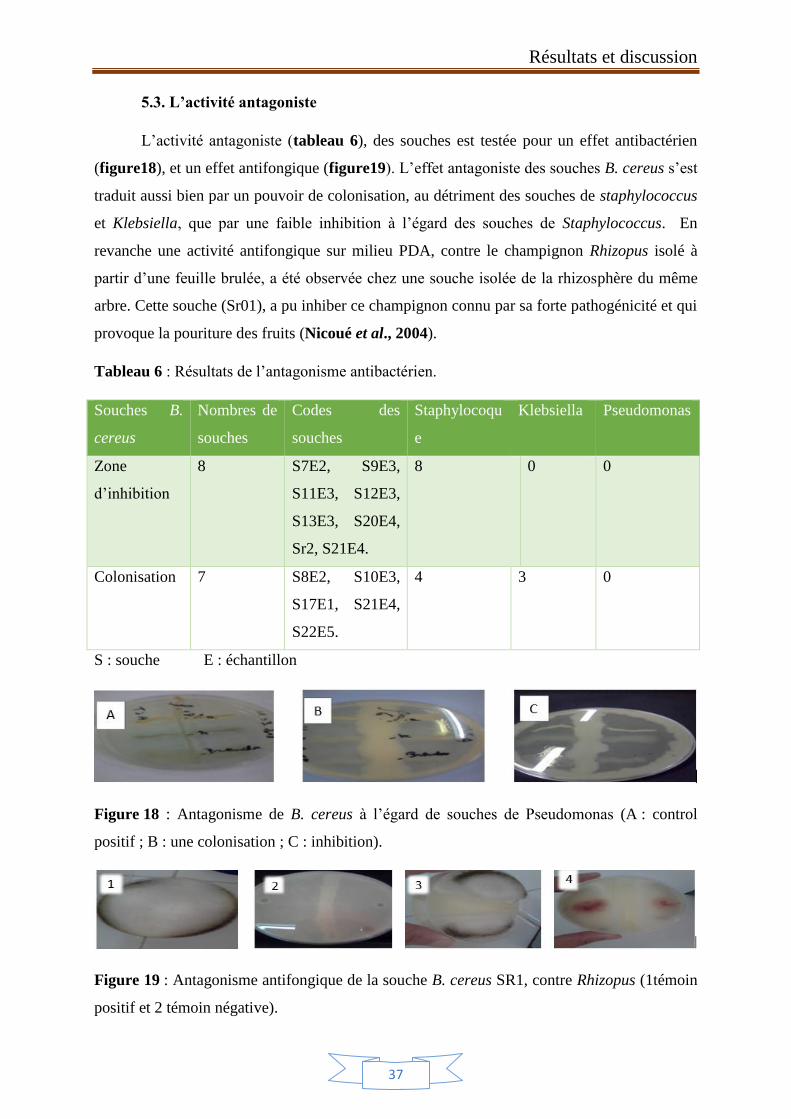
Résultats et discussion
37
5.3. L’activité antagoniste
L’activité antagoniste (tableau 6), des souches est testée pour un effet antibactérien
(figure18), et un effet antifongique (figure19). L’effet antagoniste des souches B. cereus s’est
traduit aussi bien par un pouvoir de colonisation, au détriment des souches de staphylococcus
et Klebsiella, que par une faible inhibition à l’égard des souches de Staphylococcus. En
revanche une activité antifongique sur milieu PDA, contre le champignon Rhizopus isolé à
partir d’une feuille brulée, a été observée chez une souche isolée de la rhizosphère du même
arbre. Cette souche (Sr01), a pu inhiber ce champignon connu par sa forte pathogénicité et qui
provoque la pouriture des fruits (Nicoué et al., 2004).
Tableau 6 : Résultats de l’antagonisme antibactérien.
Souches B.
cereus
Nombres de
souches
Codes des
souches
Staphylocoqu
e
Klebsiella Pseudomonas
Zone
d’inhibition
8 S7E2, S9E3,
S11E3, S12E3,
S13E3, S20E4,
Sr2, S21E4.
8 0 0
Colonisation 7 S8E2, S10E3,
S17E1, S21E4,
S22E5.
4 3 0
S : souche E : échantillon
Figure 18 : Antagonisme de B. cereus à l’égard de souches de Pseudomonas (A : control
positif ; B : une colonisation ; C : inhibition).
Figure 19 : Antagonisme antifongique de la souche B. cereus SR1, contre Rhizopus (1témoin
positif et 2 témoin négative).

Résultats et discussion
38
5.4. La formation de biofilm
La plupart des souches de B. cereus isolé du sol n’ont pas pu former un bon biofilm
dans les microplaques de titration. Ceci peut être dû à une incapacité à adhérer au polystyrène,
comme démonté par (Karunakaran and Biggs, 2011), chez des souches de B. cereus. Par
contre toutes les souches isolées du système racinaire sont caractérisées par un bon potentiel
de formation du biofilm sur les parois des puits des microplaques (figure 21). La formation de
biofilm se traduit par la présence d’un anneau à la surface du puits, après coloration au cristal
violet à 1%.
Figure 20 : la formation de biofilm sur microplaque de titration selon la technique décrit par
O’toole 2011.
La formation de biofilm est influencée par la température d’incubation, tel que montré
par la figure 21 et le tableau 7. La température s’influencer sur le rapport d’attraction des
cellules végétales et des spores (Ryu et Beuchat, 2005).
La formation de biofilm sur un tissus végétal a pour but de tester le potentiel de
colonisation du système racinaire des plantes par les PGPR en particulier B. cereus. Les
résultats ont montré que toutes les souches testées ont pu former un biofilm, que ce soit sur les
feuille désinfectées à l’alcool, ou sur les feuilles autoclavées (figure 21). La particularité de la
formation de biofilm sur les feuilles autoclavées, c’est que le milieu de culture n’a pas été
renouvelé, les souches se sont développées directement sur les feuilles, qui ont servi de source
de nutriments pour les bactéries.
4° C 30° C 37° C 45° C
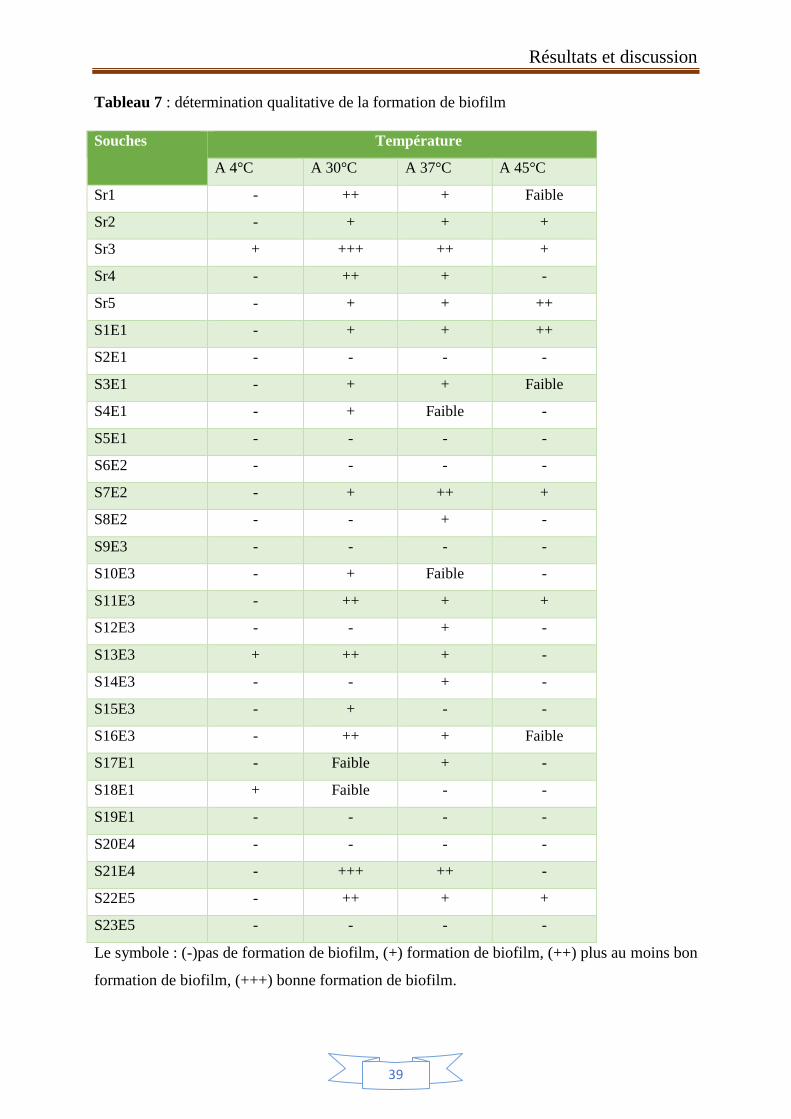
Résultats et discussion
39
Tableau 7 : détermination qualitative de la formation de biofilm
Souches Température
A 4°C A 30°C A 37°C A 45°C
Sr1 - ++ + Faible
Sr2 - + + +
Sr3 + +++ ++ +
Sr4 - ++ + -
Sr5 - + + ++
S1E1 - + + ++
S2E1 - - - -
S3E1 - + + Faible
S4E1 - + Faible -
S5E1 - - - -
S6E2 - - - -
S7E2 - + ++ +
S8E2 - - + -
S9E3 - - - -
S10E3 - + Faible -
S11E3 - ++ + +
S12E3 - - + -
S13E3 + ++ + -
S14E3 - - + -
S15E3 - + - -
S16E3 - ++ + Faible
S17E1 - Faible + -
S18E1 + Faible - -
S19E1 - - - -
S20E4 - - - -
S21E4 - +++ ++ -
S22E5 - ++ + +
S23E5 - - - -
Le symbole : (-)pas de formation de biofilm, (+) formation de biofilm, (++) plus au moins bon
formation de biofilm, (+++) bonne formation de biofilm.
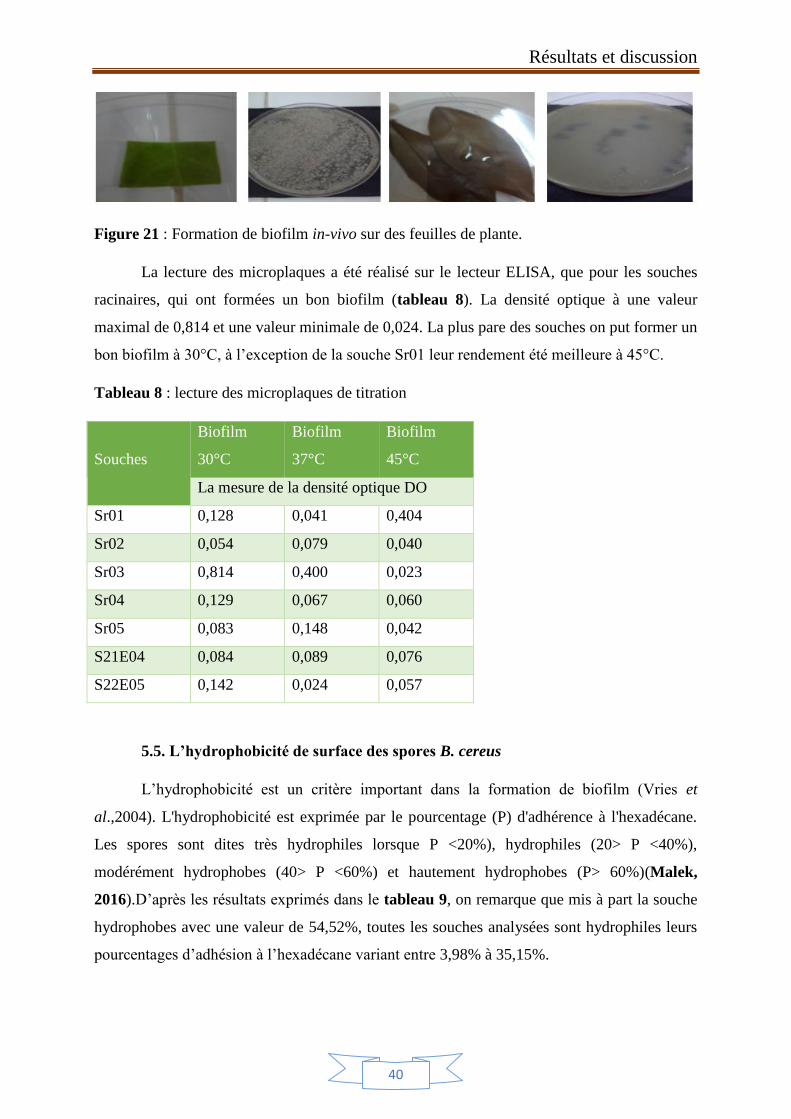
Résultats et discussion
40
Figure 21 : Formation de biofilm in-vivo sur des feuilles de plante.
La lecture des microplaques a été réalisé sur le lecteur ELISA, que pour les souches
racinaires, qui ont formées un bon biofilm (tableau 8). La densité optique à une valeur
maximal de 0,814 et une valeur minimale de 0,024. La plus pare des souches on put former un
bon biofilm à 30°C, à l’exception de la souche Sr01 leur rendement été meilleure à 45°C.
Tableau 8 : lecture des microplaques de titration
Souches
Biofilm
30°C
Biofilm
37°C
Biofilm
45°C
La mesure de la densité optique DO
Sr01 0,128 0,041 0,404
Sr02 0,054 0,079 0,040
Sr03 0,814 0,400 0,023
Sr04 0,129 0,067 0,060
Sr05 0,083 0,148 0,042
S21E04 0,084 0,089 0,076
S22E05 0,142 0,024 0,057
5.5. L’hydrophobicité de surface des spores B. cereus
L’hydrophobicité est un critère important dans la formation de biofilm (Vries et
al.,2004). L'hydrophobicité est exprimée par le pourcentage (P) d'adhérence à l'hexadécane.
Les spores sont dites très hydrophiles lorsque P <20%), hydrophiles (20> P <40%),
modérément hydrophobes (40> P <60%) et hautement hydrophobes (P> 60%)(Malek,
2016).D’après les résultats exprimés dans le tableau 9, on remarque que mis à part la souche
hydrophobes avec une valeur de 54,52%, toutes les souches analysées sont hydrophiles leurs
pourcentages d’adhésion à l’hexadécane variant entre 3,98% à 35,15%.

Résultats et discussion
41
Tableau 9 : Résultat de l’hydrophobicité des souches.
Souches Hydrophobicité Critères
Sr02 17,22% Très hydrophile
S23 3,98% Très hydrophile
S18 54,52% Modéré hydrophobe
Sr01 33,04% Hydrophile
S21 35,15% Hydrophile
S04 13,94% Très hydrophile
S11 26,05% Hydrophile
Sr03 14,25% Très hydrophile
Le potentiel d'adhésion élevé des spores de B. cereus est lié à l'hydrophobicité de la
surface des spores qui repose sur des structures morphologiques, notamment l'exosporium et
les appendices (Wijmanet al., 2007). Les résultats indiquent que nos souches sont de
hydrophiles à modéré hydrophobes ; et le pH du sol mesuré est alcalin. Les spores de B.
cereus sont de nature hydrophobe, mais le stress alcalin conduisait à une diminution de
l'hydrophobicité de la surface des spores, selon (Malek, 2016).
6. Résultats des tests in planta
L’étude du potentiel PGP in planta visait à évaluer le potentiel de biocontrôle des
bactéries isolées à partir de différentes espèces de plantes et de sols(Mota et al., 2017)La
lecture des résultats de la culture en pots du persils, a été réalisé en trois temps : après 10
jours, 21 jours, et 45 jours de l’inoculation des pots. La première lecture : le pot inoculé avec
la souche productrice de l’acide indole acétique été le plus haut, et dans le pot inoculé avec la
souche qui solubilise le phosphate, le maximum des graines ont germé par rapport au témoin.
L’IAA exogène contrôle le développement et la croissance des plantes : de faibles
concentrations d'IAA peuvent stimuler l'élongation primaire des racines, tandis que des
niveaux élevés d'IAA le diminuent (Vacheron et al., 2013) ; et on remarque qu’à un certain
stade les plantules inoculées avec la souche (S11E3), ont arrêté de croitre.
Les résultats de l’activité antagoniste :
La souche (Sr1) a montré une activité in vitro contre le champignon Rhizopus, elle a
pu l’inhibé dans la plante. La plantule de tomate témoin positif a été desséchée après 5 jour

Résultats et discussion
42
d’incubation, puis elle est morte. Par contre la plantule traitée, a résisté au champignon. Le
genre Bacillus a montré une forte activité phytoprotectrice contre les champignon, Kumar et
al., (2012).

Conclusion

Conclusion
44
Conclusion
Les micro-organismes en particulier les bactéries rhizosphérique connus sous le terme
de PGPR, ont un grand impact dans le domaine de l’agriculture. L’objectif globale de notre
travailles été de sélectionner à partir de la rhizosphère de différentes plantes et différents sols,
des bactérien sporulé « B. cereus », et de tester leur activité stimulatrice de la croissance des
végétaux.
Ce travail nous a permis d’identifier et de caractériser 28 souches appartenant au genre
Bacillus dont des souches été identifié au groupe B.cereus selon une approche phénotypique,
morphologique et biochimique. Les activités stimulatrices de la croissance des plantes ont été
étudié chez les souches isolé sont : la production de l’acide indole acétique (AIA), et la
solubilisation du phosphate, la production d’enzymes pour une activité amylolytique et
protéolytique, l’antagonisme bactérien et anti fongique, et enfin la formation de biofilm et
hydrophobicité des spores.
Les résultats in vitro montrent que neuf souches ont la capacité de produire la
phytohormone (AIA), dix-neuf souches ont pu solubiliser le phosphate, vingt souches ont une
activité amylolytique, seize souches ont une activité protéolytique, et dont l’intensité varie
d’une souche à l’autre. Un autre aspect important est le biocontrôle des agents
phytopathogènes : les souches isolées ont montré une capacité a colonisé l’agent pathogène
Staphylococus et Klebsiella. En revanche une soule souche appartient au groupe Bacillus
cereus a pu inhiber le champignon Rhizopus.
La formation de biofilms est influencée par le type de support inerte ou vivant
(polystyrène des microplaques de titration, ou tissu végétal vivant), et la température
d’incubation. L’étude de l’hydrophobicité des spores des 8 souches de B. cereus a révélé un
caractère majoritairement hydrophile.
La culture en pot permit de tester in vivo les souches isolées. Les résultats montrent
que le groupe B. cereus a un effet bénéfique en stimulant la croissance du persille et la tomate,
ou les protégant contre les agents pathogènes.
Au terme de ce travail qui ouvre plusieurs perspectives de recherche il serait
intéressant d’approfondir les investigations sur le groupe B. cereus de la région de Ain El
Houtz. Il serait intéressant de poursuivre la caractérisation de ces souches en réalisant d’autres
tests physiologiques et surtout moléculaires, ainsi que la caractérisation d’autres propriétés

Conclusion
45
stimulatrices de la croissance végétale telle que, la fixation d’azote, l’antagonisme
phytopathogène, et l’identificationdes métabolites actifs à effet inhibiteur, tels que la
production des sidérophores, la cyanogénèse (HCN), l’activité ACC désaminase, et le mieux
comprendre le pouvoir de colonisation qu’il a un effet direct et indirect sur la croissance des
plantes.

Références
bibliographiques

Références bibliographiques
46
Afnor. 2005. Norme NF/EN/ISO 7932 [Version 2004] : Microbiologie des aliments -
Méthode horizontale pour le dénombrement de Bacillus cereus présomptifs - Technique par
comptage des colonies à 30 °C.
Ahemad, M., and Kibret, M. (2014). Mechanisms and applications of plant growth
promoting rhizobacteria: Current perspective. J. King Saud Univ. - Sci. 1–20.
Alori, E.T., Glick, B.R., and Babalola, O.O. (2017). Microbial Phosphorus Solubilization
and Its Potential for Use in Sustainable Agriculture. Front. Microbiol. 8.
Andrabi, S.B.A., Tahara, M., Matsubara, R., Toyama, T., Aonuma, H., Sakakibara, H.,
Suematsu, M., Tanabe, K., Nozaki, T., and Nagamune, K. (2018). Plant hormone
cytokinins control cell cycle progression and plastid replication in apicomplexan parasites.
Parasitol. Int. 67, 47–58.
Badri, D.V., Weir, T.L., van der Lelie, D., and Vivanco, J.M. (2009). Rhizosphere
chemical dialogues: plant–microbe interactions. Curr. Opin. Biotechnol. 20, 642–650.
Baize, D. (2000). Guide des analyses en pédologie: choix, expression, présentation,
interprétation (Paris: Institut national de la recherche agronomique).
Beneduzi, A., Peres, D., Vargas, L.K., Bodanese-Zanettini, M.H., and Passaglia, L.M.P.
(2008). Evaluation of genetic diversity and plant growth promoting activities of nitrogen-
fixing bacilli isolated from rice fields in South Brazil. Appl. Soil Ecol. 39, 311–320.
Beneduzi, A., Ambrosini, A., and Passaglia, L.M.P. (2012). Plant growth-promoting
rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genet. Mol. Biol.
35, 1044–1051.
Benhamou, N., and Rey, P. (2012). Stimulateurs des défenses naturelles des plantes : une
nouvelle stratégie phytosanitaire dans un contexte d’écoproduction durable.: I. Principes de la
résistance induite. Phytoprotection 92, 1.
Bensaleh, F., Gaouar-Benyelles,N., Choukri Beghdad, M. (2014). High-performance liquid
chromatography (HPLC) Identification of five new phenolic compounds involved in the olive
tree (Olea europea var. Sigoise) resistance to Verticillium dahliae. African Journal of
Microbiology Research (8),2, 192-199.
Benschop, J.J., Jackson, M.B., Gühl, K., Vreeburg, R.A.M., Croker, S.J., Peeters,
A.J.M., and Voesenek, L.A.C.J. (2005). Contrasting interactions between ethylene and

Références bibliographiques
47
abscisic acid in Rumex species differing in submergence tolerance: ABA-regulated growth in
Rumex. Plant J. 44, 756–768.
Bottone, E.J. (2010). Bacillus cereus, a Volatile Human Pathogen. Clin. Microbiol. Rev. 23,
382–398.
Bouali, wafa, Malek, F., Sahin, F., and Abdelouahid, D.E. (2016). Morphological,
physiological and biochemical characterizations of some soil isolates of Bacillus cereus group
from Algeria. Afr. J. Microbiol. Res. 10, 1094–1103.
Brillard, J., Dupont, C.M.S., Berge, O., Dargaignaratz, C., Oriol-Gagnier, S., Doussan,
C., Broussolle, V., Gillon, M., Clavel, T., and Bérard, A. (2015). The Water Cycle, a
Potential Source of the Bacterial Pathogen Bacillus cereus. BioMed Res. Int. 2015, 1–15.
Calvet, R. (2013). Le Sol (France Agricole).
Chanclud, E. (2015). Etude du rôle des cytokinines végétales et fongiques dans l’interaction
riz-Magnaporthe oryzae. BGPI - Biol. Génétique Interact. Plantes-Parasites Pour Prot.
Intégrée 1.
Cinar, C., Apaydin, O., Yenidunya, A.F., Harsa, S., and Gunes, H. (2007). Isolation and
characterization of Bacillus thuringiensis strains from olive-related habitats in Turkey. J.
Appl. Microbiol. 0, 071010063119014-???
Ding, H., Niu, B., Fan, H., Li, Y., and Wang, Q. (2016). Draft Genome Sequence of
Bacillus cereus 905, a Plant Growth-Promoting Rhizobacterium of Wheat. Genome Announc.
4, e00489-16.
Dromigny, E. (2008). Bacillus cereus (LAVOISIER). page 1-7, 39.
Dutta, S., and Podile, A.R. (2010). Plant Growth Promoting Rhizobacteria (PGPR): the bugs
to debug the root zone. Crit. Rev. Microbiol. 36, 232–244.
Egamberdieva, D., Wirth, S.J., Alqarawi, A.A., Abd_Allah, E.F., and Hashem, A. (2017).
Phytohormones and Beneficial Microbes: Essential Components for Plants to Balance Stress
and Fitness. Front. Microbiol. 8.
Etesami, H., Alikhani, H.A., and Hosseini, H.M. (2015). Indole-3-acetic acid (IAA)
production trait, a useful screening to select endophytic and rhizosphere competent bacteria
for rice growth promoting agents. MethodsX 2, 72–78.

Références bibliographiques
48
Faille, C., Jullien, C., Fontaine, F., Bellon-Fontaine, M.-N., Slomianny, C., and
Benezech, T. (2002). Adhesion of Bacillus spores and Escherichia coli cells to inert surfaces:
role of surface hydrophobicity. Can. J. Microbiol. 48, 728–738.
Faucher, Y. (2017). Parlons potassium!
Gamalero, E., and Glick, B.R. (2015). Bacterial Modulation of Plant Ethylene Levels. Plant
Physiol. 169, 13–22.
Girard, M.-C. (2011). Etude des sol description, cartographie, utilisation (Dunod).
Gopalakrishnan, S., Upadhyaya, H., Vadlamudi, S., Humayun, P., Vidya, M.S., Alekhya,
G., Singh, A., Vijayabharathi, R., Bhimineni, R.K., Seema, M., et al. (2012). Plant
growth-promoting traits of biocontrol potential bacteria isolated from rice rhizosphere.
SpringerPlus 1, 71.
Goswami, D., Dhandhukia, P., Patel, P., and Thakker, J.N. (2014). Screening of PGPR
from saline desert of Kutch: Growth promotion in Arachis hypogea by Bacillus licheniformis
A2. Microbiol. Res. 169, 66–75.
Goswami, D., Thakker, J.N., and Dhandhukia, P.C. (2016). Portraying mechanics of plant
growth promoting rhizobacteria (PGPR): A review. Cogent Food Agric. 2.
Gouda, S., Kerry, R.G., Das, G., Paramithiotis, S., Shin, H.-S., and Patra, J.K. (2018).
Revitalization of plant growth promoting rhizobacteria for sustainable development in
agriculture. Microbiol. Res. 206, 131–140.
Granum, P.E., and Lund, T. (1997). Bacillus cereus and its food poisoning toxins. FEMS
Microbiol. Lett. 157, 223–228.
Guérin, A., Rønning, H.T., Dargaignaratz, C., Clavel, T., Broussolle, V., Mahillon, J.,
Granum, P.E., and Nguyen-The, C. (2017). Cereulide production by Bacillus
weihenstephanensis strains during growth at different pH values and temperatures. Food
Microbiol. 65, 130–135.
Gupta, G., Snehi, S.K., and Singh, V. (2017a). Role of PGPR in Biofilm Formations and Its
Importance in Plant Health. In Biofilms in Plant and Soil Health, I. Ahmad, and F.M. Husain,
eds. (Chichester, UK: John Wiley & Sons, Ltd), pp. 27–42.

Références bibliographiques
49
Gupta, M., Kiran, S., Gulati, A., Singh, B., and Tewari, R. (2012). Isolation and
identification of phosphate solubilizing bacteria able to enhance the growth and aloin-A
biosynthesis of Aloe barbadensis Miller. Microbiol. Res. 167, 358–363.
Gupta, S., Kaushal, R., Spehia, R.S., Pathania, S.S., and Sharma, V. (2017b).
Productivity of capsicum influenced by conjoint application of isolated indigenous PGPR and
chemical fertilizers. J. Plant Nutr. 40, 921–927.
Habibi, M.H., and Karimi, B. (2014). Preparation, Characterization, and Application of Zinc
Oxide Nanoparticles by Sol-Gel Pyrrolysis Method: Influence of Annealing Temperature on
Crystalline Phases. Synth. React. Inorg. Met.-Org. Nano-Met. Chem. 44, 1291–1298.
Huang, X.-F., Chaparro, J.M., Reardon, K.F., Zhang, R., Shen, Q., and Vivanco, J.M.
(2014). Rhizosphere interactions: root exudates, microbes, and microbial communities.
Botany 92, 267–275.
Iqbal, N., Khan, N.A., Ferrante, A., Trivellini, A., Francini, A., and Khan, M.I.R. (2017).
Ethylene Role in Plant Growth, Development and Senescence: Interaction with Other
Phytohormones. Front. Plant Sci. 08.
Jensen, G.B., Hansen, B.M., Eilenberg, J., and Mahillon, J. (2003). The hidden lifestyles
of Bacillus cereus and relatives. Environ. Microbiol. 5(8), 631–640.
Kang, S.-M., Khan, A.L., You, Y.-H., Kim, J.-G., Kamran, M., and Lee, I.-J. (2014).
Gibberellin production by newly isolated strain Leifsonia soli SE134 and its potential to
promote plant growth. J. Microbiol. Biotechnol. 24, 106–112.
Karunakaran, E., and Biggs, C.A. (2011). Mechanisms of Bacillus cereus biofilm
formation: an investigation of the physicochemical characteristics of cell surfaces and
extracellular proteins. Appl. Microbiol. Biotechnol. 89, 1161–1175.
Kasim, W.A., Gaafar, R.M., Abou-Ali, R.M., Omar, M.N., and Hewait, H.M. (2016).
Effect of biofilm forming plant growth promoting rhizobacteria on salinity tolerance in barley.
Ann. Agric. Sci. 61, 217–227.
Khan, N.A., Khan, M.I.R., Ferrante, A., and Poor, P. (2017). Editorial: Ethylene: A Key
Regulatory Molecule in Plants. Front. Plant Sci. 8.
Koehler, T.M. (2009). Bacillus anthracis physiology and genetics. Mol. Aspects Med. 30,
386–396.

Références bibliographiques
50
Kotiranta, A., Lounatmaa, K., and Haapasalo, M. (2000). Epidemiology and pathogenesis
of Bacillus cereus infections. Microbes Infect. 2, 189–198.
Kuiper, I., Lagendijk, E.L., Bloemberg, G.V., and Lugtenberg, B.J.J. (2004).
Rhizoremediation: A Beneficial Plant-Microbe Interaction. Mol. Plant. Microbe Interact. 17,
6–15.
Kumar, A. (2009). EFFECT OF ALKALINITY ON GROWTH PERFORMANCE OF
JATROPHA CURCAS INOCULATED WITH PGPR AND AM FUNGI. J. Phytol. 177–184.
Kumar, A., and Verma, J.P. (2018). Does plant—Microbe interaction confer stress
tolerance in plants: A review? Microbiol. Res. 207, 41–52.
Kumar, P., Dubey, R.C., and Maheshwari, D.K. (2012). Bacillus strains isolated from
rhizosphere showed plant growth promoting and antagonistic activity against phytopathogens.
Microbiol. Res. 167, 493–499.
Lin, Y., Ren, F., Zhao, L., and Guo, H. (2017). Genotypes and the persistence survival
phenotypes of Bacillus cereus isolated from UHT milk processing lines. Food Control 82, 48–
56.
LøVdal, I.S., Hovda, M.B., Granum, P.E., and Rosnes, J.T. (2011). Promoting Bacillus
cereus Spore Germination for Subsequent Inactivation by Mild Heat Treatment. J. Food Prot.
74, 2079–2089.
Lugtenberg, B., and Kamilova, F. (2009). Plant-Growth-Promoting Rhizobacteria. Annu.
Rev. Microbiol. 63, 541–556.
M. Usha Rani (2011). Bacillus cereus and Enterobacter cancerogenus screened for their
efficient plant growth promoting traits rhizobacteria (PGPR) and antagonistic traits among
sixteen bacterial isolates from rhizospheric soils of Pigeon Pea. Afr. J. Microbiol. Res. 5.
Maheswar, N.U., and Sathiyavani, G. (2012). Solubilization of phosphate by Bacillus Sps,
from groundnut rhizosphere (Arachishypogaea L). J. Chem. Pharm. Res. 4(8):4007-4011.
Malek, F. (2016). Hydrophobicity and specific biofilm features of Bacillus cereus spores
subjected to pH stresses. Afr. J. Microbiol. Res. 10, 744–751.

Références bibliographiques
51
Malek, F., Boudjemaa, B.M., Aouar-Métri, A., and Kihal, M. (2013). Identification and
genetic diversity of Bacillus cereus strains isolated from a pasteurized milk processing line in
Algeria. Dairy Sci. Technol. 93, 73–82.
Malik, D.K., and Sindhu, S.S. (2011). Production of indole acetic acid by Pseudomonas sp.:
effect of coinoculation with Mesorhizobium sp. Cicer on nodulation and plant growth of
chickpea (Cicer arietinum). Physiol. Mol. Biol. Plants 17, 25–32.
Miransari, M., and Smith, D.L. (2014). Plant hormones and seed germination. Environ.
Exp. Bot. 99, 110–121.
Mota, M.S., Gomes, C.B., Souza Júnior, I.T., and Moura, A.B. (2017). Bacterial selection
for biological control of plant disease: criterion determination and validation. Braz. J.
Microbiol. 48, 62–70.
Nicoué, E.É., Emond, J.-P., Vuillemard, J.-C., and Nunes, M.C. do N. (2004). Destruction
de Rhizopus stolonifer et Botrytis cinerea par des traitements ozone/ions. Phytoprotection 85,
81.
Osman, J.R., Fernandes, G., and DuBow, M.S. (2017). Bacterial diversity of the
rhizosphere and nearby surface soil of rice ( Oryza sativa ) growing in the Camargue (France).
Rhizosphere 3, 112–122.
Oteino, N., Lally, R.D., Kiwanuka, S., Lloyd, A., Ryan, D., Germaine, K.J., and Dowling,
D.N. (2015). Plant growth promotion induced by phosphate solubilizing endophytic
Pseudomonas isolates. Front. Microbiol. 6.
O’Toole, G.A. (2011). Microtiter Dish Biofilm Formation Assay. J. Vis. Exp.
Panhwar, Q.A., Jusop, S., Naher, U.A., Othman, R., and Razi, M.I. (2013). Application of
Potential Phosphate-Solubilizing Bacteria and Organic Acids on Phosphate Solubilization
from Phosphate Rock in Aerobic Rice. Sci. World J. 2013, 1–10.
Parmar, P., and Sindhu, S.S. (2013). Potassium Solubilization by Rhizosphere Bacteria:
Influence of Nutritional and Environmental Conditions. J. Microbiol. Res. 25–31.
Penrose, D.M., and Glick, B.R. (2003). Methods for isolating and characterizing ACC
deaminase-containing plant growth-promoting rhizobacteria. Physiol. Plant. 118, 10–15.

Références bibliographiques
52
Pereg, L., and McMillan, M. (2015). Scoping the potential uses of beneficial
microorganisms for increasing productivity in cotton cropping systems. Soil Biol. Biochem.
80, 349–358.
Prescott., Willey., Sherwood.,Woolvertton (2013). Microbiologie.page:693-697.
Raafat, D., and Sahl, H.-G. (2009). Chitosan and its antimicrobial potential - a critical
literature survey. Microb. Biotechnol. 2, 186–201.
Ravanbakhsh, M., Sasidharan, R., Voesenek, L.A.C.J., Kowalchuk, G.A., and Jousset,
A. (2018). Microbial modulation of plant ethylene signaling: ecological and evolutionary
consequences. Microbiome 6.
Romanowska, I., Strzelecki, B., and Bielecki, S. (2015). Biosolubilization of Polish brown
coal by Gordonia alkanivorans S7 and Bacillus mycoides NS1020. Fuel Process. Technol.
131, 430–436.
Ryu, J.-H., and Beuchat, L.R. (2005). Biofilm Formation and Sporulation by Bacillus cereus
on a Stainless Steel Surface and Subsequent Resistance of Vegetative Cells and Spores to
Chlorine, Chlorine Dioxide, and a Peroxyacetic Acid–Based Sanitizer. J. Food Prot. 68,
2614–2622.
S, U., S, M., M, A., and T, B. (2013). Purification and characterization of protease from
Bacillus cereus SU12 isolated from oyster Saccostrea cucullata. Afr. J. Biotechnol. 12, 5897–
5908.
S., V.K., Menon, S., Agarwal, H., and Gopalakrishnan, D. (2017). Characterization and
optimization of bacterium isolated from soil samples for the production of siderophores.
Resour.-Effic. Technol. 3, 434–439.
Seneviratne, G., Weerasekara, M.L.M.A.W., Seneviratne, K.A.C.N., Zavahir, J.S.,
Kecskés, M.L., and Kennedy, I.R. (2010). Importance of Biofilm Formation in Plant
Growth Promoting Rhizobacterial Action. In Plant Growth and Health Promoting Bacteria,
D.K. Maheshwari, ed. (Berlin, Heidelberg: Springer Berlin Heidelberg), pp. 81–95.
Setiawati, T.C., and Mutmainnah, L. (2016). Solubilization of Potassium Containing
Mineral by Microorganisms From Sugarcane Rhizosphere. Agric. Agric. Sci. Procedia 9,
108–117.

Références bibliographiques
53
Setlow, P. (2014). Germination of Spores of Bacillus Species: What We Know and Do Not
Know. J. Bacteriol. 196, 1297–1305.
Simmonds, P., Mossel, B.L., Intaraphan, T., and Deeth, H.C. (2003). Heat Resistance of
Bacillus Spores When Adhered to Stainless Steel and Its Relationship to Spore
Hydrophobicity. J. Food Prot. 66, 2070–2075.
Soufiane, B., and Côté, J.-C. (2013). Bacillus weihenstephanensis characteristics are present
in Bacillus cereus and Bacillus mycoides strains. FEMS Microbiol. Lett. 341, 127–137.
Souza, A.R.C. de, Baldoni, D.B., Lima, J., Porto, V., Marcuz, C., Machado, C., Ferraz,
R.C., Kuhn, R.C., Jacques, R.J.S., Guedes, J.V.C., et al. (2017). Selection, isolation, and
identification of fungi for bioherbicide production. Braz. J. Microbiol. 48, 101–108.
Souza, R. de, Ambrosini, A., and Passaglia, L.M.P. (2015). Plant growth-promoting
bacteria as inoculants in agricultural soils. Genet. Mol. Biol. 38, 401–419.
Stein, T. (2005). Bacillus subtilis antibiotics: structures, syntheses and specific functions:
Bacillus subtilis antibiotics. Mol. Microbiol. 56, 845–857.
Tabassum, B., Khan, A., Tariq, M., Ramzan, M., Iqbal Khan, M.S., Shahid, N., and
Aaliya, K. (2017). Bottlenecks in commercialisation and future prospects of PGPR. Appl.
Soil Ecol. 121, 102–117.
Tank, N., and Saraf, M. (2009). Enhancement of plant growth and decontamination of
nickel-spiked soil using PGPR. J. Basic Microbiol. 49, 195–204.
Turchi, L., Santini, T., Beccari, E., and Di Franco, C. (2012). Localization of new
peptidoglycan at poles in Bacillus mycoides, a member of the Bacillus cereus group. Arch.
Microbiol. 194, 887–892.
Vacheron, J., Desbrosses, G., Bouffaud, M.-L., Touraine, B., Moënne-Loccoz, Y.,
Muller, D., Legendre, L., Wisniewski-Dyé, F., and Prigent-Combaret, C. (2013). Plant
growth-promoting rhizobacteria and root system functioning. Front. Plant Sci. 4.
Van Oosten, M.J., Pepe, O., De Pascale, S., Silletti, S., and Maggio, A. (2017). The role of
biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol.
Technol. Agric. 4.

Références bibliographiques
54
Wdowiak-Wróbel, S., Marek-Kozaczuk, M., Kalita, M., Karaś, M., Wójcik, M., and
Małek, W. (2017). Diversity and plant growth promoting properties of rhizobia isolated from
root nodules of Ononis arvensis. Antonie Van Leeuwenhoek 110, 1087–1103.
Złoch, M., Thiem, D., Gadzała-Kopciuch, R., and Hrynkiewicz, K. (2016). Synthesis of
siderophores by plant-associated metallotolerant bacteria under exposure to Cd 2+.
Chemosphere 156, 312–325.
https://www.google.dz/search?q=la+germination+du+spore+bacillus+cereus&source=lnms&t
bm=isch&sa=X&ved=0ahUKEwiRtuW0rrfbAhVShaYKHUxnAHUQ_AUICigB&biw.
https://www.google.dz/search?q=germination+bacillus+cereus&source=lnms&tbm=isch&sa=
X&ved=0ahUKEwje2mU2ODbAhVHfywKHRjbAW4Q_AUICigB&biw=1242&bih=557#i
mgrc=d1WFhq2JykKUGM
Annexes

Annexes
Annexe 01 : les milieux de cultures
L’eau physiologique :
Na cl 9g
Eau distillé 1000 ml
1. gélose au sang (base)
Mélange de peptone 16g
Extrait de levure 2g
D-glucose 0,5g
Chlorure de sodium 7g
Agar 12g
L'eau distilée 1000Ml
pH 7,3±0,2
2. TSA (Gélose Trypticase de soja)
Trypticase 17g
Soytone 3g
Chlorure de sodium 5g
Agar 15g
Eau distillée 1000Ml
pH 7,2
3. Gélose nutritif
Beef extrait 1g/l
Least extrait 2g/l
Peptone 5g/l
Sodium chloride 5g/l
Agar 15g/l
pH 6,8±0,2
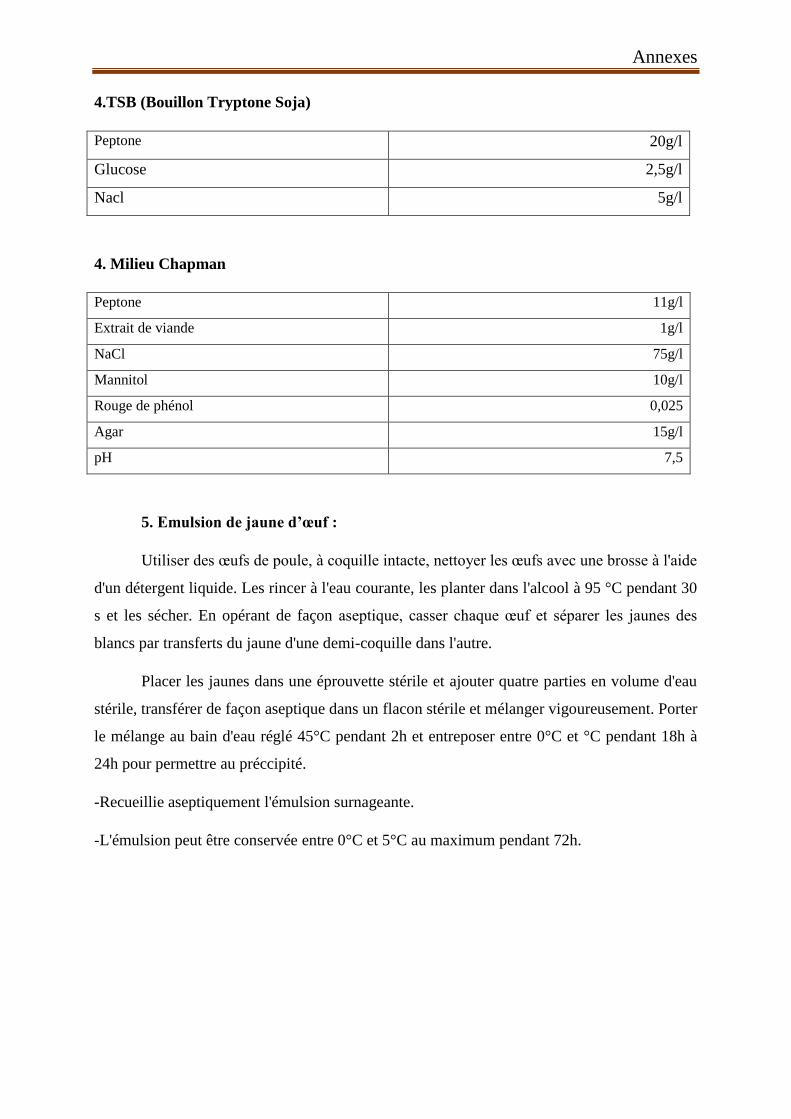
Annexes
4.TSB (Bouillon Tryptone Soja)
Peptone 20g/l
Glucose 2,5g/l
Nacl 5g/l
4. Milieu Chapman
Peptone 11g/l
Extrait de viande 1g/l
NaCl 75g/l
Mannitol 10g/l
Rouge de phénol 0,025
Agar 15g/l
pH 7,5
5. Emulsion de jaune d’œuf :
Utiliser des œufs de poule, à coquille intacte, nettoyer les œufs avec une brosse à l'aide
d'un détergent liquide. Les rincer à l'eau courante, les planter dans l'alcool à 95 °C pendant 30
s et les sécher. En opérant de façon aseptique, casser chaque œuf et séparer les jaunes des
blancs par transferts du jaune d'une demi-coquille dans l'autre.
Placer les jaunes dans une éprouvette stérile et ajouter quatre parties en volume d'eau
stérile, transférer de façon aseptique dans un flacon stérile et mélanger vigoureusement. Porter
le mélange au bain d'eau réglé 45°C pendant 2h et entreposer entre 0°C et °C pendant 18h à
24h pour permettre au préccipité.
-Recueillie aseptiquement l'émulsion surnageante.
-L'émulsion peut être conservée entre 0°C et 5°C au maximum pendant 72h.
Annexes
Annexe 02: Code des souches
Code de souches Signification
S1E1, S2E1, S3E1, S4E1, S5E1, S17E1,
S18E, S19E1, Sr1
Les souches isolées à partir de l'échantillon
1 : la rhizosphère du citronnier quatre saison
S6E2, S7E2, S8E2 Les souches isolées à partir de l'échantillon
2 : la rhizosphère du romarin
S9E3, S10E3, S11E3, S12E3, S14E3,
S15E3, S16E3
Les souches isolées à partir de l'échantillon
3 : la rhizosphère d’un sapin associé à une
plante aromatique du genre Thymus
S20E4, S21E4 Les souches isolées à partir de l'échantillon
4 : la rhizosphère des mauvaises herbes au-
dessous d’un arbre
S22E5, S23E5 Les souches isolées à partir de l'échantillon
5 : la rhizosphère d’un amandier
S : souche E : échantillon.

Résumé
La flore bactérienne de la rhizosphère ou la flore PGPR est caractérisées par ses effets bénéfiques
sur la croissance des plantes et peut être utilisé en tant que biofertilisant et biopesticide. Dans le
cadre de ce mémoire, 28 souches été isolées puis, identifier au genre Bacillus dent au groupe B.
cereus, via une approche phénotypique (morphologique et biochimique). Les propriétés PGPR
analysés dans cette étude sont : la production de la phytohormone l’acide indole acétique (AIA),
la solubilisation du phosphate, la production d’enzymes amylolytique et protéolytique,
l’antagonisme bactérien et antifongique, la formation de biofilm et l’hydrophobicité des spores.
Les résultats montrent que neuf souches sont productrices de l’AIA, dix-neuf souches solubilisent
le phosphate, la plupart ont une activité enzymatique, deuze souches ont une activité
antibactérienne, une seule souche qu’inhibe le champignon Rhizopus. L’étude de la formation de
biofilm sur les feuilles d’oranger, et sur les microplaques de titration, a montré une colonisation
importante du tissu vivant comparé au polystyrène Les résultats révélait dans PGPR intéressantes
chez les souches caractérisées, elles sont confirmées par les essais in vivo. La bactérisassions des
gaines de persilles et des plantules de tomate ont montré des effets bénéfiques sur la croissance
des plantules cultivé en pot.
Mot clé : Bacillus cereus, sol, identification, PGP, biofilm, antagonisme.
Abstract
The bacterial flora of the rhizosphereor the PGPR flora ischaracterized by itsbeneficialeffects on
plant growth and can beused as a biofertilizer and good biopesticide. In the context of thispaper,
28 strainswereisolated and thenidentifiedwith the genus Bacillus tooth B. cereus group, via a
phenotypicapproach (morphological and biochemical). The PGPR propertiesanalyzed in
thisstudyare: phytohormone production indole aceticacid (AIA), phosphate solubilization,
amylolytic and proteolytic enzyme production, bacterial and antifungalantagonism, biofilm
formation, and hydrophobicity of the spores. The results show thatninestrainsproduce AIA,
nineteenstrainssolubilize phosphate, most have enzymaticactivity, twostrains have an
antibacterialactivity, a single strainthatinhibits the fungus Rhizopus. The study of biofilm
formation on orange leaves, and on titration microplates, showed a significantcolonization of
living tissue compared to polystyrene. The resultsrevealed in PGPR of interest in the
characterizedstrains, they are confirmed by the tests in vivo. The bacterization of parsleysheaths
and tomatoseedlingsshowedbeneficialeffects on the growth of seedlingsgrown in pots. Key word :
Bacillus cereus, soil, identification, PGP, biofilm, antagonism.

ملخص
. ومبيدات حيويةالموجودة في طبقة الريزوسفير للنبات تتميز بأثارها المفيدة على نمو النباتات. يمكن استخدامها كمخصبات البكتيريا
سلالة من نوع بسيلوس من بينها نوع بسيلوس سيريوس. خصائص هذه البكتيريا تم تحليلها في هذه 28تم عزل ا العملفي سياق هذ
اذابة الفوسفات, انتاج انزيمات محللة البروتين و النشاء, العداء ضد اسيد اندول اسيتيك ,الدراسة هي قدرتها على انتاج هرمون نباتي
على انتاج سلالات اهم القدرة 9وتشكل البيوفيلم, وأخيرا كره الماء من طرف الجراثيم. اظهرت النتائج ان ’ البكتيريا و الفطريات
اهم نشاط مضاد للجراثيم الا سلالة واحدة 12انزيمي, سلالة لها القدرة على اذابة الفوسفور , الأغلبية لديهم نشاط 19الهرمون,
مضادة للفطر. دراسة تشكل البيوفيلم على أوراق البرتقال و البوليستيران أظهرت ان السلالة لها القدرة على احتلال الانسجة الحية.
الطماطم أظهرت اثار وشتلات لبقدونس عليها في الجسم الحي. البكتيريا الموضوعة في بذور ا ةالنتائج المتحصل عليها تم المصادق
مفيدة على نمو الشتلات المزروعة في الاصيص.
بسيلوس سيريوس, بيوفيلم, العداء, نمو النبات. الكلمات المفتاحية تحديد السلالات,
Top Related