







Languages
Pages
Legal


Association analyses based on false discovery rate implicate many new loci 1
for coronary artery disease 2
3
Christopher P Nelson1,2,60, Anuj Goel3,4,60 , Adam S Butterworth5,6,60, Stavroula Kanoni7,8,60, 4 Tom R Webb1,2, Eirini Marouli7,8, Lingyao Zeng9, Ioanna Ntalla7,8, Florence Y Lai1,2, Jemma C 5 Hopewell10, Olga Giannakopoulou7,8, Tao Jiang5, Stephen E Hamby1,2, Emanuele Di 6 Angelantonio5,6, Themistocles L Assimes11, Erwin P Bottinger12, John C Chambers13–15, Robert 7 Clarke10, Colin N A Palmer16,17 , Richard M Cubbon18, Patrick Ellinor19 , Raili Ermel20 , 8 Evangelos Evangelou13,21 , Paul W Franks22–24, Christopher Grace3,4, Dongfeng Gu25, 9 Aroon D Hingorani26, Joanna M M Howson5, Erik Ingelsson27, Adnan Kastrati9, Thorsten 10 Kessler9, Theodosios Kyriakou3,4, Terho Lehtimäki28, Xiangfeng Lu27, Yingchang Lu12,29, 11 Winfried März30–32, Ruth McPherson33, Andres Metspalu34, Mar Pujades-Rodriguez35, Arno 12 Ruusalepp20,36, Eric E Schadt37, Amand F Schmidt38, Michael J Sweeting5, Pierre A Zalloua39,40 13 , Kamal Al Ghalayini41, Bernard D Keavney42,43 , Jaspal S Kooner14,15,44, Ruth J F Loos12,45, Riyaz 14 S Patel46,47, Martin K Rutter48,49, Maciej Tomaszewski43,50, Ioanna Tzoulaki13,21 , Eleftheria 15 Zeggini51 , Jeanette Erdmann52–54, George Dedoussis55, Johan L M Björkegren36,37,56, EPIC-16 CVD Consortium57, CARDIoGRAMplusC4D57, The UK Biobank CardioMetabolic Consortium 17 CHD working group57, Heribert Schunkert9,60, Martin Farrall3,4,60, John Danesh5,6,58,60, Nilesh J 18 Samani1,2,60, Hugh Watkins3,4,60 & Panos Deloukas7,8,59,60 19 20 21 1Department of Cardiovascular Sciences, University of Leicester, Leicester, UK. 2National 22 Institute for Health Research Leicester Biomedical Research Centre, Leicester, UK. 3Division 23 of Cardiovascular Medicine, Radcliffe Department of Medicine, University of Oxford, Oxford, 24 UK. 4Wellcome Trust Centre for Human Genetics, University of Oxford, Oxford, UK. 25 5MRC/BHF Cardiovascular Epidemiology Unit, Department of Public Health and Primary 26 Care, University of Cambridge, Cambridge, UK. 6NIHR Blood and Transplant Research Unit in 27 Donor Health and Genomics, Department of Public Health and Primary Care, University of 28 Cambridge, Cambridge, UK. 7William Harvey Research Institute, Barts & the London Medical 29 School, Queen Mary University of London, London, UK. 8Centre for Genomic Health, Queen 30 Mary University of London, London, UK. 9German Heart Center Munich, Clinic at Technische 31 Universität München and Deutsches Zentrum für Herz- und Kreislauferkrankungen (DZHK), 32 partner site Munich Heart Alliance, Munich, Germany. 10CTSU, Nuffield Department of 33 Population Health, University of Oxford, Oxford, UK. 11Department of Medicine, Stanford 34 University School of Medicine, Stanford, California, USA. 12Charles Bronfman Institute for 35 Personalized Medicine, Icahn School of Medicine at Mount Sinai, New York, New York, USA. 36 13Department of Epidemiology and Biostatistics, Imperial College London, London, UK. 37 14Department of Cardiology, Ealing Hospital, London North West Healthcare NHS Trust, 38 Southall, UK. 15Imperial College Healthcare NHS Trust, London, UK. 16Molecular and Clinical 39 Medicine, Biomedical Research Institute, University of Dundee, Ninewells Hospital, Dundee, 40 UK. 17Pharmacogenomics Centre, Biomedical Research Institute, University of Dundee, 41 Ninewells Hospital, Dundee, UK. 18Leeds Institute of Cardiovascular and Metabolic Medicine, 42 University of Leeds, Leeds, UK. 19Cardiac Arrhythmia Service and Cardiovascular Research 43 Center, Broad Institute of Harvard and Massachusetts Institute of Technology, Boston, 44 Massachusetts, USA. 20Department of Cardiac Surgery, Tartu University Hospital, Tartu, 45
Nature Genetics: doi:10.1038/ng.3913

Estonia. 21Department of Hygiene and Epidemiology, University of Ioannina Medical School, 46 Ioannina, Greece. 22Department of Clinical Sciences, Genetic & Molecular Epidemiology 47 Unit, Lund University Diabetes Center, Skåne University Hospital, Lund University, Malmö, 48 Sweden. 23Department of Nutrition, Harvard T.H. Chan School of Public Health, Harvard 49 University, Boston, Massachusetts, USA. 24Department of Public Health and Clinical 50 Medicine, Unit of Medicine, Umeå University, Umeå, Sweden. 25State Key Laboratory of 51 Cardiovascular Disease, Fuwai Hospital, National Center of Cardiovascular Diseases, Chinese 52 Academy of Medical Sciences and Peking Union Medical College, Beijing, China. 26University 53 College London, London, UK. 27Department of Medicine, Division of Cardiovascular 54 Medicine, Stanford University School of Medicine, Stanford, California, USA. 28Department 55 of Clinical Chemistry, Fimlab Laboratories and Faculty of Medicine and Life Sciences, 56 University of Tampere, Tampere, Finland. 29Division of Epidemiology, Department of 57 Medicine, Vanderbilt-Ingram Cancer Center, Vanderbilt Epidemiology Center, Vanderbilt 58 University School of Medicine, Nashville, Tennessee, USA. 30Clinical Institute of Medical and 59 Chemical Laboratory Diagnostics, Medical University of Graz, Graz, Austria. 31Medical Clinic 60 V (Nephrology, Rheumatology, Hypertensiology, Endocrinology, Diabetology), Medical 61 Faculty Mannheim, University of Heidelberg, Mannheim, Germany. 32Academy, Synlab 62 Holding Deutschland GmbH, Mannheim, Germany. 33Ruddy Canadian Cardiovascular 63 Genetics Centre, University of Ottawa Heart Institute, Ottawa, Ontario, Canada. 34Estonian 64 Genome Center, University of Tartu, Tartu, Estonia. 35Leeds Institute of Biomedical and 65 Clinical Sciences, University of Leeds, Leeds, UK. 36Clinical Gene Networks AB, Stockholm, 66 Sweden. 37Department of Genetics and Genomic Sciences, Icahn Institute for Genomics and 67 Multiscale Biology, Icahn School of Medicine at Mount Sinai, New York, New York, USA. 68 38Institute of Cardiovascular Science, UCL, London, UK. 39Lebanese American University, 69 School of Medicine, Beirut, Lebanon. 40Harvard T.H. Chan School of Public Health, Boston, 70 Massachusetts, USA. 41Department of Medicine, King Abdulaziz University, Jeddah, Saudi 71 Arabia. 42Division of Cardiovascular Sciences, Faculty of Biology, Medicine and Health, 72 University of Manchester, Manchester, UK. 43Central Manchester University Hospitals NHS 73 Foundation Trust, Manchester Academic Health Science Centre, Manchester, UK. 74 44Cardiovascular Science, National Heart and Lung Institute, Imperial College London, 75 London, UK. 45Mindich Child Health Development Institute, Icahn School of Medicine at 76 Mount Sinai, New York, New York, USA. 46Farr Institute of Health Informatics, UCL, London, 77 UK. 47Bart’s Heart Centre, St Bartholomew’s Hospital, London, UK. 48Division of Diabetes, 78 Endocrinology and Gastroenterology, Faculty of Biology, Medicine and Health, University of 79 Manchester, Manchester, UK. 49Manchester Diabetes Centre, Central Manchester University 80 Hospitals NHS Foundation Trust, Manchester Academic Health Science Centre, Manchester, 81 UK. 50Division of Medicine, Central Manchester NHS Foundation Trust, Manchester 82 Academic Health Science Centre, Manchester, UK. 51Wellcome Trust Sanger Institute, 83 Hinxton, UK. 52Institute for Cardiogenetics, University of Lübeck, Lübeck, Germany. 53DZHK 84 (German Research Centre for Cardiovascular Research), partner site Hamburg/Lübeck/Kiel, 85 Lübeck, Germany. 54University Heart Center Lübeck, Lübeck, Germany. 55Department of 86 Nutrition-Dietetics, Harokopio University, Athens, Greece. 56Integrated Cardio Metabolic 87 Centre, Department of Medicine, Karolinska Institutet, Karolinska Universitetssjukhuset, 88 Huddinge, Sweden. 57A list of members and affiliations appears in the Supplementary Note. 89 58Wellcome Trust Sanger Institute, Wellcome Trust Genome Campus, Hinxton, UK. 59Princess 90 Al-Jawhara Al-Brahim Centre of Excellence in Research of Hereditary Disorders (PACER-HD), 91
Nature Genetics: doi:10.1038/ng.3913

King Abdulaziz University, Jeddah, Saudi Arabia. 60These authors contributed equally to this 92 work. 93
94
95
96
Supplementary Note 97
98
99
1. Definitions of cases and controls for studies in the CARDIoGRAMplusC4D 1000 Genomes 100
imputed GWAS and Exome chip meta-analyses. 101
102
Detailed information on the studies which participated in the 1000 Genomes GWAS and 103
Exome chip meta-analyses are provided in references 2 and 3 respectively. A brief account of 104
the definitions used to define CAD cases and controls in these studies is given below. 105
106
ADVANCE: The Atherosclerotic Disease VAscular functioN and genetiC Epidemiology study 107
enrolled patients with MI, angina with at least one angiographic stenosis of >50%, or 108
revascularization procedure in men 18 to 45 or women 18 to 55 years of age at the time of 109
the event, incident stable exertional angina at an older age, and incident non-fatal MI at an 110
older age. All controls were free of clinical CAD, cerebrovascular disease and peripheral 111
arterial disease at the time of recruitment; AGES: in the Reykjavik Study MI was defined 112
according to the MONICA criteria in the Icelandic Myocardial infarction registry maintained 113
by the Icelandic Heart Association for the Medical Directorate of Iceland. All MIs were 114
validated by examining all records from the participants‟ death certificates, Hospital records 115
and autopsy records; ARIC The Atherosclerosis Risk in Communities study is a prospective, 116
community-based sample of 15,792 white and African American adults aged 45 to 64 years 117
recruited from four communities in the United States between 1987 and 1989; ATVB: MI case-118
control study in men or women ≤ 45 years of age. Controls were free of thromboembolic 119
disease; BHF-FHS: CAD cases were recruited from the British Heart Foundation Family Heart 120
Study and supplemented by additional cases from WTCCC-CAD2. Controls were selected from 121
the UK 1958 Birth Cohort; BAS: The Beijing Atherosclerosis Study recruited cases with 122
Nature Genetics: doi:10.1038/ng.3913

validated history of MI. Controls were randomly selected from subjects participating in a 123
community-based survey of cardiovascular risk factors in Beijing and were judged to be free 124
of CAD by history, clinical examination, electrocardiography, and Rose questionnaire; BioMe 125
Biobank: CAD cases were ascertained from the BioMe Biobank using the electronic health 126
record with ICD9 codes 410.xx to 414.xx and abnormal stress test or abnormal coronary 127
angiography. Controls were individuals from the BioMe Biobank who did not meet the criteria 128
for cases; BioVU: cases with MI or CAD were ascertained from the Vanderbilt University 129
Medical Center Biorepository by searching the electronic medical record for ≥ 2 instances of 130
ICD-9 codes 410.x – 414. Controls were individuals from the Vanderbilt University 131
Biorepository who did not have any record of ICD-9 codes 410.x – 414.x; CARDIOGENICS: 132
cases from Germany and England were under the age of 65 with a confirmed primary MI 133
within the preceding 3-36 months. In France cases had acute coronary syndrome with one 134
stenosis >50% diagnosed in at least one major coronary artery. Controls were healthy blood 135
donors (aged 32 to 65 years) recruited in UK; CAS: The China Atherosclerosis Study (CAS) 136
recruited cases have a family history of CAD (83.8%). Diagnoses of MI cases was based on 137
electrocardiograms and cardiac enzymes whereas CAD subjects without known history of MI 138
had > 70% stenosis in at least one of a major epicardial vessel. Controls were individuals who 139
did not develop incident CAD and had no family history of CAD during the 8 year follow-up 140
period of the study from four northern field centers of InterASIA; CCGB: Cases had at least 141
one of MI, CABG, PTCA or a stenosis of at least 50% in at least one epicardial vessel. Diabetic 142
cases and cases aged greater than 55 for men or 65 for women were excluded. Controls had 143
a CTA or angiogram demonstrating no stenosis of greater than 50% and were >65 years old 144
for men and 70 years old for women at the time of recruitment; COROGENE: FINRISK is a 145
population cohort collected every 5 years (10 000 individuals recruited each time) form 5 146
different regions in Finland. Data is collected with an extensive questionnaire and baseline 147
measurements (physical examination, laboratory tests). The cohort is followed from national 148
registers (causes of death register, Hospital discharge register), which cover 100% of the 149
population. ICD codes from these registers are used to define cases and controls; Duke: cases 150
had an MI or coronary stenosis ≥ 50%. Controls were > 50 years old without coronary stenosis 151
> 30% and without history of MI, coronary artery bypass grafting, percutaneous coronary 152
intervention, or heart transplant. EGCUT: CAD or MI cases were ascertained from the Estonian 153
Biobank (Estonian Genome Center at the University of Tartu) using the medical history and 154
Nature Genetics: doi:10.1038/ng.3913

current health status that is recorded according to ICD-10 codes (CAD defined with ICD-10 155
I20-I25). Controls were selected from the Estonian Biobank (Estonian Genome Center at the 156
University of Tartu) who did not have any record of cardiovascular diseases (ICD-10 I10-I79). 157
EPIC-CAD: The EPIC (European Prospective Study into Cancer and Nutrition) study sub-cohorts 158
from the UK were used. Subjects were collected in collaboration with general practitioners, 159
mainly in Cambridgeshire and Norfolk. Cases were individuals who developed fatal or non-160
fatal CAD during an average follow-up of 11 years ending June 2006. Participants were 161
identified if they had a hospital admission and/or died with CAD as the underlying cause. CAD 162
was defined as cause of death codes ICD-9 410-414 or ICD-10 I20-I25, and hospital discharge 163
codes ICD-10 I20.0, I21, I22, or I23 according to the International Classification of Diseases, 164
9th and 10th revisions, respectively. Controls were study participants who remained free of 165
any cardiovascular disease during follow-up (defined as ICD-9 401-448 and ICD-10 I10-I79); 166
FamHS: The Family Heart Study includes prevalent CHD cases based on self report of coronary 167
bypass, MI, and/or any of the following: coronary angioplasty, balloon angioplasty, 168
atherectomy, stent, percutaneous transluminal coronary angioplasty, or percutaneous 169
coronary intervention; FGENTCARD: study subjects were selected from the coronary artery 170
disease database to have undergone cardiac catheterization following a single consistent and 171
stringent recruitment protocol. Cardiac catheterization was performed by Judkins technique, 172
based on clinical need such as MI as diagnosed by electrocardiogram and high troponin levels, 173
unstable or stable angina, heart failure or reversible ischemia. The four main coronary 174
arteries: the left main artery (LMCA), the left anterior descending artery (LAD), the left 175
circumflex artery (LCx), and the right coronary artery (RCA) were visualized from different 176
angles by angiography. The extent of stenosis in these vessels was assessed and recorded by 177
percentage; FHS: The Framingham Heart Study included prevalent MI cases at the time of 178
giving blood. MI in this project was defined as a recognized non-fatal MI diagnosed with 179
documented evidence from EKG or elevated enzymes; CHD was defined as recognized fatal 180
or non-fatal MI, coronary insufficiency or the first fatal CHD event; FIA3: MI cases of MI were 181
recruited in the Vasterbotten Intervention Program (VIP), WHO’s Multinational Monitoring of 182
Trends and Determinants in Cardiovascular Disease (MONICA) study in northern Sweden and 183
the Mammography Screening Project (MSP) in Vasterbotten. Controls were individuals free 184
of MI from VIP and MSP; GenRIC: Genomics Research in Cardiovascular Disease study 185
recruited Korean CAD cases confirmed by standard coronary angiography - MI (31%), stable 186
Nature Genetics: doi:10.1038/ng.3913

angina (41%) and unstable angina (28%). MI diagnosis was based on electrocardiographic 187
patterns and on elevated creatine kinase-MB and troponin I levels. Controls were randomly 188
selected as a shared control group for Korean cancer and CAD from 1,200,000 subjects aged 189
40-69; GoDARTS: Cases in the GoDARTS (Genetics of Diabetes Audit and Research in Tayside 190
Scotland) study were first-ever CAD event, defined as fatal and non-fatal myocardial 191
infarction, unstable angina, or coronary revascularization. Controls were free of CAD, stroke, 192
and peripheral vascular disease; German CAD North and South cohorts: cases were 193
individuals with MI or CAD. Controls were derived from population-based studies in Germany; 194
GerMIFS I-IV: Cases from the GerMIFS I-II study had a strong positive family history for CAD 195
and an early onset of disease, i.e. were enriched for a strong genetic component. GerMIFS III 196
cases were non-fatal MI identified in the KORA registry as hospitalized survivors of MI 26–74 197
years of age. MI diagnosis was done with the algorithm of WHO Multinational Monitoring of 198
Trends and Determinants in Cardiovascular Disease (MONICA) project. The GerMIFS IV cases 199
consist of consecutive patients referred for coronary angiography, classified as CAD or MI 200
cases based on the coronary angiogram (at least a 50% stenosis in one major coronary vessel) 201
and age of onset (<65 years in males and <70 years in females). Controls were subjects from 202
the MONICA/KORA Augsburg survey F4, the PopGen blood donor sample 2 (PopGen-BSP), the 203
Berlin Aging Study II (BASE-II); HSDS: The Helsinki Sudden Death Study had CAD cases defined 204
as coronary artery stenosis 50 % or greater, old or acute MI, or sudden cardiac death caused 205
by plaque rupture and thrombosis; HPS: The MRC/BHF Heart Protection Study was a large 206
UK-based cholesterol-lowering trial involving participants with a history of MI, unstable or 207
stable angina, CABG, or angioplasty. Cases had a history of MI, unstable or stable angina, 208
CABG, or angioplasty. Population controls were used from the UK Twins Study and National 209
Blood Service collections; HUNT: MI Cases were retrospectively identified as HUNT 2 and 210
HUNT 3 participants diagnosed with acute MI (ICD-10 I21 or ICD-9 410) in the medical 211
departments at the two local hospitals in Nord-Trøndelag County from December 1987 to 212
June 2011. Controls were selected among HUNT 2 and HUNT 3 participants with available 213
DNA (N = 70,300) after excluding individuals with the following hospital diagnosed or self-214
reported conditions in themselves or known 1st and/or 2nd degree family members: MI, 215
angina, heart failure, stroke, aortic aneurysm, atherosclerosis, intermittent claudication, and 216
registered percutaneous coronary angioplasty procedures or bypass surgery; ITH: The 217
INTERHEART study used worldwide cases and controls of European ethnicity. Incident acute 218
Nature Genetics: doi:10.1038/ng.3913

MI, presenting to a hospital within 24 hours of symptom onset. Age and sex matched hospital 219
and community based, with no previous diagnosis of heart disease or history of exertional 220
chest pain; LIFE-Heart: is an observational study that recruits patients undergoing first-time 221
diagnostic coronary angiography due to suspected stable CAD with previously untreated 222
coronary arteries, patients with stable left main coronary artery disease and patients with 223
acute MI. CAD was defined as luminal reduction of >50% in any vessel. Normal angiograms 224
were considered as controls. MI cases were defined as the subset of CAD cases with either 225
anamnestic MI, acute MI (requiring revascularization via percutaneous coronary intervention) 226
or elevated troponine levels. MI controls were defined as normal angiograms with neither of 227
the above mentioned criteria of MI; LOLIPOP: The London Life Sciences Prospective 228
Population (LOLIPOP) study is composed of men and women aged 35-75 years, and of North 229
European or Indian Asian origin, identified from the lists of 58 GPs in West London. Cases had 230
a history of MI, CABG or PTCA or angiographically confirmed coronary artery stenosis greater 231
than 50%. Controls were free from CAD; LURIC: The Ludwigshafen Risk and Cardiovascular 232
Health study consists of patients hospitalized for coronary angiography. CAD was defined as 233
the presence of a visible luminal narrowing (>50% stenosis) in at least one of 15 coronary 234
segments according to a classification of the American Heart Association; MAYOVDB: in the 235
he Mayo Clinic Vascular Disease Biorepository CHD was defined as history of MI, coronary 236
revascularization, angina with positive stress test, or presence of >50% stenosis of an 237
epicardial coronary artery on coronary angiography. Controls were defined based on absence 238
of case status for CHD as well as absence of other forms of atherosclerotic vascular disease 239
(peripheral arterial disease, AAA, cerebrovascular disease); MedStar: The MedStar study is a 240
Washington Hospital Center based study that recruited a consecutive cohort of patients 241
undergoing cardiac catheterization at Washington Hospital. Enrolment criteria include any 242
clinical indication for cardiac catheterization and ability to give informed consent. Coronary 243
angiograms were scored on the day by the interventional cardiologist who performed the 244
procedure and reviewed by a second cardiologist at a later date. CAD cases identified with 245
one or more coronary vessels with ≥50% stenosis were divided into stable CAD cases without 246
history of MI and CAD cases with a history of MI; controls showed no evidence of CAD on 247
coronary angiography; MDC: cases were prevalent and incident nonfatal or fatal MI. Controls 248
were participants free of CHD at baseline and during follow-up; MIGen: Myocardial Infarction 249
Genetics includes cases of early-onset MI (in men less than or equal to 50 y old or women less 250
Nature Genetics: doi:10.1038/ng.3913

than or equal to 60 y old) and age- and sex-matched controls free of MI. MI was diagnosed 251
on the basis of autopsy evidence of fatal MI or a combination of chest pain, 252
electrocardiographic evidence of MI, or elevation of one or more cardiac biomarkers (creatine 253
kinase or cardiac troponin); MHI: cases were ascertained from the Montreal Heart Institute 254
Biobank. CAD was defined as the presence of MI, percutaneous coronary intervention, or 255
coronary artery bypass grafting. Controls were individuals from the Montreal Heart Institute 256
Biobank who were free of history of MI, percutaneous coronary intervention, or coronary 257
artery bypass grafting; OHS: cases had angiographically confirmed coronary artery disease (>1 258
coronary artery with >50% stenosis) and did not have type 2 diabetes; ≤ 50 years old for males 259
and ≤ 50 years old for females. Controls were asymptomatic males > 65 and females > 70; 260
PAS-AMC: symptomatic CAD before 51 years of age, defined as MI, coronary 261
revascularization, or evidence of at least 70% stenosis in a major epicardial coronary artery. 262
Controls were asymptomatic individuals with over 95% of them from the same region as 263
cases; PennCath: cases had angiographically confirmed coronary artery disease (>1 coronary 264
artery with 50% stenosis); ≤ 55 years old for males and ≤ 60 years old for females. Controls 265
were individuals with normal coronary angiography, men > 40 years old and women > 45 266
years old; PIVUS: Prospective Investigation of the Vasculature in Uppsala Seniors is a 267
community-based study where all men and women at age 70 living in Uppsala, Sweden were 268
invited to participate in 2001. Information about coronary heart disease was collected 269
through the Swedish Patient Register and Cause of Death Register; PREDICTCVD: a case-270
cohort sample that comprises of incident cardiovascular disease cases and matched controls 271
selected from the 1992, 1997, 2002 and 2007 FINRISK surveys. CHD was defined as MI, 272
unstable angina pectoris, CABG, PTCA, or death due to CHD; PROCARDIS: symptomatic CAD 273
before age 66. CAD was defined as clinically documented evidence of myocardial infarction, 274
coronary artery bypass grafting, acute coronary syndrome, coronary angioplasty, or stable 275
angina. Controls were individuals with no personal or sibling history of CAD before age 66; 276
PROMIS: the Pakistan Risk of Myocardial Infarction Study is an incident case-control study of 277
acute MI. MI cases had symptoms within 24 hours of hospital presentation, typical ECG 278
changes; and positive troponin-I test. Controls were identified contemporaneously in the 279
same hospitals as the index cases and selected from among people who had no history of CVD 280
and were frequency-matched to MI cases by sex and age (5-year bands); PROSPER: 281
PROspective Study of Pravastatin in the Elderly at Risk was a multicenter randomized placebo-282
Nature Genetics: doi:10.1038/ng.3913

controlled trial to assess whether treatment with pravastatin diminishes the risk of major 283
vascular events in elderly; RS: The Rotterdam Study (RS) is a prospective, population-based 284
cohort study on determinants of several chronic diseases in older adults. Information on CAD 285
and MI, medical history, and use of medication were obtained during the interview; 286
SDS/AIDHS: in the Sikh Diabetes Study/Asian Indian Diabetic Heart Study, CAD was 287
considered if there was use of nitrate medication (nitroglycerine), electrocardiographic 288
evidence of angina pain, coronary angiographic evidence of severe (greater than 50%) 289
stenosis, or echocardiographic evidence of MI. Diagnosis was based on date of CABG or 290
angioplasty, and medication usage obtained from patient records; THISEAS: The Hellenic 291
Study of Interactions between SNPs and Eating in Atherosclerosis Susceptibility recruited 292
subjects with a first-ever MI before age of 70 yrs presenting with either ACS or stable CAD 293
defined as >50% stenosis in at least one of the three main coronary vessels assessed by 294
coronary angiography. Controls were free of any cardiovascular disease, cancer, or 295
inflammatory diseases; TWINGENE: is a longitudinal study nested within the Swedish Twin 296
Register initiated to examine associations between genetic factors and cardiovascular disease 297
in Swedish twins. Information about coronary heart disease was collected through the 298
Swedish Patient Register and Cause of Death Register; ULSAM: Men born between 1920 and 299
1924 in Uppsala, Sweden were invited to participate at age 50 (N=2,841) in this longitudinal 300
cohort study. Information about CAD was collected through the Swedish Patient Register and 301
Cause of Death Register; VHS: documented MI, coronary artery bypass grafting, CAD (by 302
angiography) in males ≤ 45 years old and females ≤ 50 years old. Controls were individuals 303
with normal coronary angiography in males > 60 years old or females > 65 years old; WGHS: 304
in the Women's Genome Health Study incident CVD was assessed by physician review of 305
medical records following self-report of a CVD event; WHI: cases were individuals from the 306
Women’s Health Initiative who had incident MI, coronary revascularization, hospitalized 307
angina or death due to coronary disease. Controls were participants free of CHD on follow-308
up. 309
310
2. Functional annotation of candidate CAD risk genes in the 13 new genome-wide significant 311
loci 312
313
TDRKH 314
Nature Genetics: doi:10.1038/ng.3913

The CAD associated region spans a region of around 40kb that contains 4 genes; OAZ3 315
(Ornithine decarboxylase antizyme 3), TDRKH (Tudor and KH domain containing), LINGO4 316
(Leucine rich repeat and Ig domain containing 4) and RORC (RAR related orphan receptor C). 317
Several variants overlap annotated regulatory elements, including 3 SNPs in the proximal 318
promoter region of TDRKH. Another variant (rs6673002) in high LD with the CAD lead variant 319
(rs11810571) is associated with increased expression of TDRKH in skeletal muscle. TDRKH 320
encodes a protein involved in the primary piRNA biogenesis pathway and is required during 321
spermatogenesis to repress transposable elements and prevent their mobilization, which is 322
essential for the germline integrity. The piRNA metabolic process mediates the repression of 323
transposable elements during meiosis by forming complexes composed of piRNAs and Piwi 324
proteins and govern the methylation and subsequent repression of transposons. 325
326
FN1 327
The lead variant at this locus, rs1250259, causes an asparagine to leucine change at amino 328
acid position 15 of FN1 (Fibronectin 1). FN1 is a glycoprotein that interacts with other 329
extracellular matrix proteins and cell-surface receptor proteins to regulate cell adhesion and 330
migration. FN1 expression is increased in plaques (PMID:2663879) and mouse models have 331
demonstrated a causal role for FN1 in the development and progression of atherosclerosis17-332
18. There are several variants in high LD with the coding variant at the 5’ end of FN1 which 333
overlap active promoter elements and show associations with increased expression of FN1 in 334
subcutaneous fat (Supplementary Figure 6, Supplementary Table 9). The CAD lead variant is 335
also associated with plasma LDL-cholesterol (PMID:24097068) (Supplementary Table 10). 336
Diseases associated with FN1 include glomerulopathy with fibronectin deposits 2 and plasma 337
fibronectin deficiency. 338
339
RHOA 340
The lead variant at this locus, rs7623687, falls in intron 1 of RHOA (Ras Homolog Family 341
Member A), which encodes a member of the Rho family of small GTPases. rs7623687 is 342
predicted to be the most likely functional variant by CADD and GWAVA and overlaps a 343
predicted promoter, which is active in the majority of cells and tissues (Supplementary Figure 344
6). When activated, RHOA acts through a number of target proteins to transduce signals to 345
Nature Genetics: doi:10.1038/ng.3913

the actin cytoskeleton and nucleus and regulate cell phenotype. In smooth muscle, RHOA is 346
important for inactivating myosin phosphatase and sustaining smooth muscle contraction 347
(PMID:9635436, PMID:19403695) and signalling through RHOA controls actin remodelling, 348
cellular proliferation and migration and the transition of vascular smooth muscle cells from a 349
contractile to a proliferative phenotype (PMID:22467316). Pathological activity of the RHOA 350
effector ROCK (Rho-associated protein kinase) has been reported in experimental 351
hypertension (PMID: 9353125), asthma, atherosclerosis (PMID:1975438), stroke, heart failure 352
and other cardiovascular diseases (PMID:2392130). There are 8 further variants in high LD 353
with the lead SNP. The CAD associated region covers approximately 120kb and, as well as 354
RHOA, overlaps; GPX1 (Glutathione peroxidase 1), which encodes an antioxidant enzyme, 355
TCTA (T-cell leukemia translocation altered), which encodes a protein of unknown function 356
implicated in leukaemia, AMT (Aminomethyltransferase) which encodes a mitochondrial 357
enzyme involved in glycine catabolism, NICN1 (Nicolin 1), which is of unknown function and 358
the region upstream of DAG1 (Dystrophin associated glycoprotein 1), mutations in which 359
cause several forms of muscular dystrophy. The absence of any association with gene 360
expression at this locus makes the causal gene difficult to predict. 361
362
UMPS/ITGB5 363
The lead variant and its high LD proxies span a 35kb region containing UMPS (Uridine 364
Monophosphate Synthetase), which encodes an enzyme that catalyses the final two steps of 365
the de novo pyrimidine biosynthetic pathway, and the 3’ end of ITGB5 (Integrin Subunit Beta 366
5). The risk allele of rs150512726, CADD score of 20.7 (a CADD score over 20 means the 367
variant is predicted to be within the 1% most deleterious variants), causes a 3 amino acid 368
deletion within the cytoplasmic tail of ITGB5 (Supplementary Figure 6). Integrins are cell 369
surface-proteins which regulate and are regulated by extracellular matrix components such 370
as fibronectin, laminins, collagens and vitronectin. ITGB5 forms a heterodimer with ITGA5 and 371
along with other αν dimers regulates the activation of latent TGFB1 from the extracellular 372
matrix (PMID: 16142753, PMID:18086923, PMID:24216753). Interestingly, a recent study 373
showed that cells expressing a ανβ5 containing the rs150512726 deletion allele were unable 374
to activate latent TGFB1 (PMID: 22025551). 375
Nature Genetics: doi:10.1038/ng.3913

376
ARHGEF26 377
At this locus, 140 variants in high LD with the lead variant span a region of approximately 378
170kb containing ARHGEF26 (Rho Guanine Nucleotide Exchange Factor 26) and its antisense 379
transcript ARGEF26-AS1. The lead variant rs12493885 causes a valine to leucine change at 380
amino acid position 29 in ARHGEF26, a member of the Rho-guanine nucleotide exchange 381
factor (Rho-GEF) family which regulate Rho GTPases by catalyzing the exchange of GDP for 382
GTP. However this amino acid change is not conserved and is not predicted to be damaging. 383
The high LD variants are non-coding with several of the variants showing evidence for 384
regulatory functionality (Supplemental Figure 6). Functionally, ARHGEF26 is a strong 385
candidate gene for CAD. It is expressed in endothelial cells where it acts downstream of 386
adhesion molecules to control the formation on membrane docking structures in leukocyte 387
trans-endothelial migration (PMID: 17875742). On an atherosclerosis prone hyperlipidemic 388
background, Arhgef26 knockout in mice resulted in reduced atherosclerosis with plaques also 389
showing reduced macrophage content (PMID: 23372835). There is also evidence implicating 390
another gene at this locus. The risk allele of rs62276842 (r2=0.95 with the CAD lead SNP) is 391
associated with reduced expression of DHX36 (DEAH-Box Helicase 36) in skeletal muscle. 392
DHX36, which is immediately 3’ of ARHGEF26, encodes an RNA-helicase involved in mRNA 393
stability and is implicated in the type I interferon response (PMID: 21703541). 394
395
PRDM8/ FGF5 396
The CAD associated lead variant and its high LD proxies fall in an intergenic region between 397
PRDM8 (PR/SET domain 8), which encodes a member of a conserved family of histone 398
methyltransferases that predominantly act as negative regulators of transcription and FGF5 399
(Fibroblast growth factor 5), which encodes a member of the fibroblast growth factor family. 400
Four variants overlap regulatory elements, with rs1458038, which overlaps elements active 401
in skin fibroblasts and heart, having the highest functional prediction score in both CADD and 402
GWAVA. PRDM8 contains an N-terminal Su(var)3-9, enhancer-of-zeste, a Trithorax (SET) 403
domain and a double zinc-finger domain. In mouse knockouts there is mis-targeting by 404
neurons of the dorsal telencephalon, abnormal itch-like behavior, and impaired 405
Nature Genetics: doi:10.1038/ng.3913

differentiation of rod bipolar cells (PMID: 26023183). In humans, the protein has been shown 406
to interact with the phosphatase laforin and the ubiquitin ligase malin, which regulate 407
glycogen construction in the cytoplasm (PMID: 22961547). PRDM8 may be involved in the 408
control of steroidogenesis through transcriptional repression of steroidogenesis marker 409
genes such as CYP17A1 and LHCGR. Diseases associated with PRDM8 include epilepsy, 410
progressive myoclonic 10 and early-Onset Lafora Body Disease. FGF family members possess 411
broad mitogenic and cell survival activities, and are involved in a variety of biological 412
processes, including embryonic development, cell growth, morphogenesis, tissue repair, 413
tumor growth and invasion. FGF5 is a secreted peptide that regulates cell proliferation, 414
particularly the growth of hair follicles. FGF5 has been associated with blood pressure traits 415
in Hispanics and African Americans (PMID: 27736895) as well as trichomegaly. 416
417
PDE5A/MAD2L1 418
The lead variant and its two high LD proxies fall in an intergenic region between a non-coding 419
RNA, LOC100996694, and MAD2L1 (MAD2 mitotic arrest deficient-like 1 (yeast)), which 420
encodes a component of the mitotic spindle assembly checkpoint that prevents the onset of 421
anaphase until all chromosomes are properly aligned at the metaphase plate. The CAD 422
associated variants are associated with increased expression of PDE5A (Phosphodiesterase 423
5A), which is located 350kb upstream, in the atherosclerotic wall suggesting that these 424
variants might reside in a long-range enhancer element. The two proxy variants do overlap 425
an enhancer active in colon smooth muscle and the eQTL suggests that regulatory activity in 426
the blood vessel might only be apparent in disease. PDE5A encodes a cGMP-binding, cGMP-427
specific phosphodiesterase, a member of the cyclic nucleotide phosphodiesterase family. 428
PDE5A is involved in the regulation of intracellular concentrations of cyclic nucleotides and is 429
important for smooth muscle relaxation in the cardiovascular system where it regulates nitric-430
oxide-generated cGMP (PMID: 15489334). Diseases associated with PDE5A 431
include Priapism and Nonarteritic Anterior Ischemic Optic Neuropathy. 432
433
HDGFL1 434
Nature Genetics: doi:10.1038/ng.3913

The lead variant, rs6909752, falls in an intergenic region approximately 20kb downstream of 435
HDGFL1 (Hepatoma derived growth factor-like 1), which encodes an uncharacterised protein 436
with homology to HDGF; a regulator of cell growth, proliferation and migration. The lead 437
variant and several of the high LD proxy SNPs overlap regulatory elements that are active in 438
a range of different cells and tissue types (Supplementary Figure 6). The variant with highest 439
CADD and GWAVA score, rs6909574, is in an enhancer active in stem cells, while the lead 440
variant is in an enhancer active in smooth muscle from different tissue types. 441
442
ARNTL 443
The lead variant, rs3993105, and its high LD proxies are clustered within a 20kb region 444
containing the proximal promoter and first exon of ARNTL (Aryl hydrocarbon receptor nuclear 445
translocator like). Several variants overlap regulatory elements active in most cell and tissue 446
types (Supplementary Figure 6). ARNTL encodes a transcriptional activator which forms a 447
core component of the circadian clock. The circadian clock is an important regulator of 448
physiological functions such as metabolism, sleep, body temperature, blood pressure, 449
endocrine, immune, cardiovascular, and renal function. Disruptions in the circadian rhythms 450
contribute to the pathology of cardiovascular diseases, cancer, metabolic syndromes and 451
aging. ARNTL positively regulates myogenesis and negatively regulates adipogenesis via the 452
transcriptional control of the canonical Wnt signaling pathway. ARNTL has been associated 453
with major depressive disorder and accelerated response to antidepressant drug 454
treatment as well as schizophrenia. 455
HNF1A 456
The lead variant, rs2244608, and its high LD proxies span a 30kb region around HNF1A (HNF1 457
Homeobox A). Several variants overlap regulatory elements, however, the most likely 458
functional variant is rs1169288, which causes an isoleucine to leucine amino acid change at 459
amino acid position 27 in the HNF1A protein. This variant has a CADD score of 25.6 and is 460
predicted to be among the top 1% most deleterious variants. HNF1A is a transcription factor 461
that regulates the expression of genes involved in lipid and glucose metabolism. The CAD 462
associated variants are also associated with plasma lipids, C-reactive protein and several 463
other metabolites (Supplementary Table 10). Mutations in HNF1A cause maturity onset 464
Nature Genetics: doi:10.1038/ng.3913

diabetes of the young type 3 (MODY3) and can also result in the appearance of hepatic 465
adenomas. HNF1A has been associated with insulin-dependent diabetes mellitus. 466
467
CDH13 468
The lead variant, rs750048, falls in intron 2 of CDH13 (Cadherin 13), has no high LD proxies 469
and is associated with increased expression of CDH13 and a number of nearby non-coding 470
transcripts in internal mammary artery and atherosclerotic aortic root. rs750048 overlaps an 471
enhancer active in mesenchymal stem cells, muscle satellite cells and skeletal muscle. CDH13 472
is an atypical member of the cadherin superfamily of calcium-dependent cell adhesion 473
proteins. CDH13 lacks the transmembrane domain and cytoplasmic domain of the other 474
cadherins and mediates cell signalling rather than adhesion (PMID: 9650591). CDH13 is highly 475
expressed in the vasculature and shows increased expression in atherosclerosis (PMID: 476
11326751). CDH13 can mediate the cellular response to lipoproteins including LDL (PMID: 477
9468307) and adiponectin (PMID: 15210937) and the CAD lead variant is associated with 478
circulating adiponectin levels (PMID:22479202). Other variants in CDH13 have also been 479
linked to blood pressure, arterial stiffness (PMID: 17903302) and cardiometabolic traits 480
(PMID: 24142632). 481
482
TGFB1 483
The lead variant at this locus, rs8108632, is in intron 1 of TGFB1 (Transforming growth factor 484
beta 1) and is not in high LD with any other variants (Supplementary Figure 6). The SNP is in 485
a regulatory element, which is active in various blood cells. rs4803455 (r2=0.68 with the CAD 486
lead SNP) is associated with migraine (Supplementary Table 10). TGFB1 is a cytokine 487
expressed by various cell types in atherosclerotic plaques. It induces expression of 488
extracellular matrix proteins and inhibits proliferation of smooth muscle cells (PMID: 489
3471760, PMID: 2065041) and reduces adhesion molecule expression and inhibits cell 490
proliferation and migration of endothelial cells (PMID: 7683321). TGFB1 also reduces 491
macrophage foam cell formation (PMID: 11742878). In addition, several studies have 492
Nature Genetics: doi:10.1038/ng.3913

demonstrated that TGFB1 inhibits atherogenesis in vivo (PMID: 7489365, PMID: 12067907, 493
PMID: 11701621, PMID: 14568988). 494
495
SNRPD2 496
The lead variant at this locus, rs1964272, is in an intergenic region immediately 3’ of SNRPD2 497
(Small Nuclear Ribonucleoprotein D2 Polypeptide), which encodes a member of the small 498
nuclear ribonucleoprotein core protein family required for pre-mRNA splicing and small 499
nuclear ribonucleoprotein biogenesis. There are just two high LD proxies, both of which are 500
in introns of QPCTL (Glutaminyl-peptide cyclotransferase like), which encodes an enzyme 501
responsible for the biosynthesis of pyroglutamyl peptides, a posttranslational modification 502
present at the N-terminus of some peptide hormones and secretory proteins. The most likely 503
functional variant, rs2302593, is in the bi-directional promoter, active in most cells and 504
tissues, of SNPRD2 and QPCTL in a region where several transcription factors have been 505
shown to bind. The absence of an association with gene expression means that it is not 506
possible to predict which of the genes might be involved in disease. The CAD associated 507
variants also associate with BMI (Supplementary Table 10). 508
509
2. Consortia 510
511
EPIC-CVD 512
Kim Overvad1,2, Anne Tjønneland3, Francoise Clavel-Chapelon4, Rudolf Kaaks5, Heiner 513 Boeing6, Antonia Trichopoulou7,8, Pietro Ferrari9, Domenico Palli10, Vittorio Krogh11, 514 Salvatore Panico12, Rosario Tumino13, Giuseppe Matullo14,15, Jolanda Boer16, Yvonne van. 515 der. Schouw17, Elisabete Weiderpass18,19,20,21, J. Ramon Quiros22, María-José Sánchez23,24, 516 Carmen Navarro25,26, Conchi Moreno-Iribas2,3, Larraitz Arriola27, Olle Melander28, Patrik 517 Wennberg29, Nicholas J. Wareham30, Timothy J. Key31, Elio Riboli32, Adam S. Butterworth33,34, 518 John Danesh33,34,35 519
520 1. Department of Public Health, Section for Epidemiology, Aarhus University, Aarhus, 521 Denmark 522 2. Department of Cardiology, Aalborg University Hospital, Aalborg, Denmark 523 3. Diet, Genes and Environment, Danish Cancer Society Research Center, Copenhagen, 524 Denmark 525 4. INSERM, Centre for Research in Epidemiology and Population Health (CESP), U1018, 526 Nutrition, Hormones, and Women's Health Team, Institut Gustave Roussy, Villejuif, France 527
Nature Genetics: doi:10.1038/ng.3913

5. Division of Cancer Genetic Epidemiology, German Cancer Research Centre (DKFZ), im 528 Neuenheimer Feld 581, 69121 Heidelberg, Germany 529 6. Department of Epidemiology, German Institute of Human Nutrition (DIfE), Potsdam-530 Rehbrücke, Germany 531 7. WHO Collaborating Center for Nutrition and Health, Unit of Nutritional Epidemiology and 532 Nutrition in Public Health, Department of Hygiene, Epidemiology and Medical Statistics, 533 University of Athens Medical School, Athens, Greece 534 8. Hellenic Health Foundation, Athens, Greece 535 9. IARC, Lyon, France 536 10. Molecular and Nutritional Epidemiology Unit, Centro per lo Studio e la Prevenzione 537 Oncologica-Scientific Institute of Tuscany, Florence, Italy 538 11. Epidemiology and Prevention Unit, Fondazione IRCCS Istituto Nazionale dei Tumori, 539 Milan, Italy 540 12. Dipartimento di Medicina Clinica e Chirurgia, Federico II University, Naples, Italy 541 13. Cancer Registry and Histopathology Unit, Civic- M.P.Arezzo Hospital, ASP Ragusa, Italy 542 14. Human Genetics Foundation, Turin, Italy 543 15. Department of Medical Sciences, University of Turin, Italy 544 16. Centre for Nutrition, Prevention and Health Services, National Institute for Public Health 545 and the Environment (RIVM), Bilthoven, the Netherlands 546 17. Julius Center for Health Sciences and Primary Care, University Medical Center Utrecht, 547 Utrecht, the Netherlands 548 18. Department of Community Medicine, Faculty of Health Sciences, University of Tromsø, 549 The Arctic University of Norway, Tromsø, Norway 550 19. Department of Research, Cancer Registry of Norway, Institute of Population-Based 551 Cancer Research, Oslo, Norway 552 20. Department of Medical Epidemiology and Biostatistics, Karolinska Institutet, Stockholm, 553 Sweden 554 21. Genetic Epidemiology Group, Folkhälsan Research Center, Helsinki, Finland 555 22. Public Health Directorate, Asturias, Spain 556 23. Public Health Institute of Navarra, Pamplona, Spain 557 24. Red de Investigación en Servicios de Salud en Enfermedades Crónicas, Madrid, Spain 558 25. Epidemiology Department, Murcia Health Authority, Murcia, Spain 559 26. Centro de Investigación Biomédica en red en Epidemiología y Salud Pública, Barcelona, 560 Spain 561 27. Public Health Division of Gipuzkoa, Instituto Bio-Donostia, Basque Government, 562 CIBERESP, Spain 563 28. CRR, Lund University, 20502 Malmo ̈564 29. Department of Public Health and Clinical Medicine, Family Medicine, Umeå University, 565 Umeå, Sweden 566 30. Medical Research Council Epidemiology Unit, University of Cambridge, Cambridge, UK 567 31. Cancer Epidemiology Unit, Nuffield Department of Population Health, University of 568 Oxford 569 32. School of Public Health, Imperial College London, UK 570 33. Cardiovascular Epidemiology Unit, Department of Public Health & Primary Care, 571 University of Cambridge, UK 572 34. The National Institute for Health Research Blood and Transplant Unit (NIHR BTRU) in 573
Nature Genetics: doi:10.1038/ng.3913

Donor Health and Genomics at the University of Cambridge, UK 574 35. Wellcome Trust Sanger Institute, Genome Campus, Hinxton, UK 575
576
CARDIoGRAMplusC4D 577 578 Themistocles L. Assimes1, Adam S. Butterworth2,3, John C. Chambers4,5,6, Robert Clarke7, 579 Rory Collins7, John Danesh2,3,8, George Dedoussis9, Panos Deloukas10,11,12, Jeanette 580 Erdmann13,14,15, Martin Farrall16,17, Paul W. Franks18,19,20, Anuj Goel16,17, Dongfeng Gu21, 581 Jemma C. Hopewell22, G Kees Hovingh23, Erik Ingelsson24, Stavroula Kanoni10,11, Sekar 582 Kathiresan25,26,27, Bong-Jo Kim28, Jaspal S. Kooner5,6,29, Terho Lehtimäki30, Lars Lind31, 583 Xiangfeng Lu21, Eirini Marouli10,11, Winfried März32,33,34, Ruth McPherson35, Andres 584 Metspalu36, Christopher P. Nelson37,38, Markku S. Nieminen39, Majid Nikpay40, Christopher 585 O'Donnell41, Colin Palmer42, Markus Perola43,44,45, Muredach P. Reilly46, Samuli Ripatti47, 586 Robert Roberts48, Danish Saleheen49,50, Nilesh Samani37,38, Manjinder S. Sandhu3,8,51, 587 Heribert Schunkert52, Agneta Siegbahn53, Unnur Thorsteinsdottir54,55, Hugh Watkins16,17, 588 Tom R. Webb37,38, Christina Willenborg56, Cristen J. Willer57,58,59, Hong H. Won60, Pierre A. 589 Zalloua61,62, Natalie R. van Zuydam42 590
1. Department of Medicine, Stanford University School of Medicine, Stanford, CA 591 94305, USA 592
2. MRC/BHF Cardiovascular Epidemiology Unit, Department of Public Health and 593 Primary Care, University of Cambridge,, Cambridge CB1 8RN, UK 594
3. NIHR Blood and Transplant Research Unit in Donor Health and Genomics, 595 Department of Public Health and Primary Care, University of Cambridge, Cambridge 596 CB1 8RN, UK 597
4. Department of Epidemiology and Biostatistics, Imperial College London, London W2 598 1PG, UK 599
5. Department of Cardiology, Ealing Hospital, London North West Healthcare NHS 600 Trust, Middlesex UB1 3HW, UK 601
6. Imperial College Healthcare NHS Trust, London W12 0HS, UK 602 7. Clinical Trial Service Unit and Epidemiological Studies Unit, Nuffield Department of 603
Population Health, University of Oxford, Oxford, UK 604 8. Wellcome Trust Sanger Institute, Wellcome Trust Genome Campus, Hinxton, 605
Cambridge CB10 1RQ, UK 606 9. Department of Nutrition-Dietetics,Harokopio University, Athens, Greece 607 10. William Harvey Research Institute, Barts & the London Medical School, Queen Mary 608
University of London, London EC1M 6BQ, UK 609 11. Centre for Genomic Health, Queen Mary University of London, London EC1M 6BQ, 610
UK 611 12. Princess Al-Jawhara Al-Brahim Centre of Excellence in Research of Hereditary 612
Disorders (PACER-HD), King Abdulaziz University, Jeddah, Saudi Arabia 613 13. Institute for Cardiogenetics, University of Lübeck, Lübeck, 23562, Germany 614 14. DZHK (German Research Centre for Cardiovascular Research), partner site 615
Hamburg/Lübeck/Kiel, Lübeck 23562, Germany 616 15. University Heart Center Lübeck, Lübeck, 23562, Germany 617
Nature Genetics: doi:10.1038/ng.3913

16. Division of Cardiovascular Medicine, Radcliffe Department of Medicine, University of 618 Oxford, Oxford OX3 9DU, UK 619
17. Wellcome Trust Centre for Human Genetics, University of Oxford, Oxford OX3 7BN, 620 UK 621
18. Department of Clinical Sciences, Genetic & Molecular Epidemiology Unit, Lund 622 University Diabetes Center, Skåne University Hospital, Lund University, 623 Malmö/Skåne SE-205 02, Sweden 624
19. Department of Nutrition, Harvard T. H. Chan School of Public Health, Harvard 625 University, Boston/Massachusetts MA 02115, USA 626
20. Department of Public Health and Clinical Medicine, Unit of Medicine, Umeå 627 University, Umeå/Västerbotten SE-901 85, Sweden 628
21. State Key Laboratory of Cardiovascular Disease, Fuwai Hospital, National Center of 629 Cardiovascular Diseases, Chinese Academy of Medical Sciences and Peking Union 630 Medical College, Beijing 100037, China 631
22. CTSU, Nuffield Department of Population Health, University of Oxford, Oxford OX3 632 7LF, UK 633
23. Department of Vascular Medicine, Academic Medical Center, the Netherlands 634 24. Department of Medicine, Division of Cardiovascular Medicine, Stanford University 635
School of Medicine, Stanford, CA 94305, Stanford, CA 94305, US 636 25. Massachusetts General Hospital, Boston, MA 02114, USA 637 26. Broad Institute, Cambridge, MA 2142, USA 638 27. Harvard Medical School, Boston, MA 2115, USA 639 28. Center for Genome Science, Korea National Institute of Health, Chungcheongbuk-do 640
28159, Korea 641 29. Cardiovascular Science, National Heart and Lung Institute, Imperial College London, 642
London W12 0NN, UK 643 30. Department of Clinical Chemistry, Fimlab Laboratories and Faculty of Medicine and 644
Life Sciences, University of Tampere, Tampere 33520, Finland 645 31. Department of Medical Sciences, Cardiovascular Epidemiology, Uppsala University, 646
Uppsala 751 85, Sweden 647 32. Clinical Institute of Medical and Chemical Laboratory Diagnostics, Medical University 648
of Graz, Graz 8036, Austria 649 33. Medical Clinic V (Nephrology, Rheumatology, Hypertensiology, Endocrinology, 650
Diabetology), Medical Faculty Mannheim, University of Heidelberg, Mannheim 651 68167, Germany 652
34. Academy, Synlab Holding Deutschland GmbH, Mannheim 68161, Germany 653 35. Ruddy Canadian Cardiovascular Genetics Centre University of Ottawa Heart Institute, 654
Ottawa K1Y 4W7, Canada 655 36. Estonian Genome Center, University of Tartu, Tartu 51010, Estonia 656 37. Department of Cardiovascular Sciences, University of Leicester, Leicester LE3 9QP, 657
UK 658 38. National Institute for Health Research Leicester Cardiovascular Biomedical Research 659
Unit, Leicester LE3 9QP, UK 660 39. Division of Cardiology, Department of Medicine, Helsinki University Central Hospital 661
(HUCH), Helsinki, Finland 662 40. Ruddy Canadian Cardiovascular Genetics Centre, Ottawa K1Y 4W7, Canada 663
Nature Genetics: doi:10.1038/ng.3913

41. Framingham Heart Study, National Heart, Lung and Blood Institute, Framingham MA 664 01702, USA 665
42. Pat McPherson Centre for Pharmacogenetics and Pharmacogenomics, The School of 666 Medicine, University of Dundee, Ninewells Hospital and Medical School, Dundee DD1 667 9SY 668
43. National Institute for Health and Welfare, Helsinki FI00290, Finland 669 44. University of Helsinki, Institute for Molecular Medicine (FIMM) and Diabetes and 670
Obesity Research Program, Helsinki FI00014, Finland 671 45. University of Tartu, Estonian Genome Center, Tartu, Estonia, Tartu 51010, Estonia 672 46. Cardiovascular Institute, Perelman School of Medicine at the University of 673
Pennsylvania, Philadelphia, USA 674 47. Department of Chronic Disease Prevention, National Institute for Health and 675
Welfare, Helsinki FIN-00271, Finland 676 48. University of Ottawa Heart Institute, Cardiovascular Research Methods Centre and 677
Ruddy Canadian Cardiovascular Genetics Centre, Ottawa, Canada 678 49. Department of Biostatistics and Epidemiology, University of Pennsylvania, USA 679 50. Center for Non-Communicable Diseases, Karachi, Pakistan 680 51. MRC Epidemiology Unit, Institute of Metabolic Science, Addenbrooke's Hospital, 681
Cambridge, UK 682 52. German Heart Center Munich, Clinic at Technische Universität München, Munich 683
80636, Germany 684 53. Uppsala Clinical Research Center, Uppsala University, Uppsala, Sweden 685 54. deCODE Genetics/Amgen inc., Reykjavik 101, Iceland 686 55. Faculty of Medicine, University of Iceland, Reykjavik 101, Iceland 687 56. Institut für Integrative und Experimentelle Genomik, Universität zu Lübeck, Lübeck, 688
Germany 689 57. Department of Internal Medicine, University of Michigan, Ann Arbor, MI 48109, USA 690 58. Department of Computational Medicine and Bioinformatics, University of Michigan, 691
Ann Arbor, MI 48109, USA 692 59. Department of Human Genetics, University of Michigan, Ann Arbor, MI 48109, USA 693 60. Samsung Advanced Institute for Health Sciences and Technology (SAIHST), 694
Sungkyunkwan University, Samsung Medical Center, Seoul, Korea 695 61. Lebanese American University, School of Medicine, Beirut 13-5053, Lebanon 696 62. Harvard T.H. Chan School of Public Health, Boston 02115, USA 697
698
699
The UK-Biobank CardioMetailic Consortium - CHD working group 700
Adam S. Butterworth1,2, John Danesh1,2,3, Panos Deloukas4,5,6, Emanuele Di Angelantonio1,2, 701 Evangelos Evangelou7,8, Martin Farrall9,10, Olga Giannakopoulou4,5, Anuj Goel9,10, Christopher 702 Grace9,10, Harry Hemingway11, Aroon Hingorani12, Jemma C. Hopewell13, Stavroula Kanoni4,5, 703 Bernard Keavney14,15, Jaspal S. Kooner16,17,18, Theodosios Kyriakou9,10, Florence Lai19,20, Ruth 704 J. F. Loos21,22, Eirini Marouli4,5, Christopher P. Nelson19,20, Ioanna Ntalla4,5, Riyaz S. Patel23,24, 705 Mar Pujades-Rodriguez25, Martin Rutter26,27, Nilesh Samani19,20, Amand F. Schmidt28, John R. 706
Nature Genetics: doi:10.1038/ng.3913

Thompson29, Maciej Tomaszewski30,31, Ioanna Tzoulaki32,33, Hugh Watkins9,10, Tom R. 707 Webb19,20, 708
1. MRC/BHF Cardiovascular Epidemiology Unit, Department of Public Health and 709 Primary Care, University of Cambridge,, Cambridge CB1 8RN, UK 710
2. NIHR Blood and Transplant Research Unit in Donor Health and Genomics, 711 Department of Public Health and Primary Care, University of Cambridge, 712 Cambridge CB1 8RN, UK 713
3. Wellcome Trust Sanger Institute, Wellcome Trust Genome Campus, Hinxton, 714 Cambridge CB10 1RQ, UK 715
4. William Harvey Research Institute, Barts & the London Medical School, Queen 716 Mary University of London, London EC1M 6BQ, UK 717
5. Centre for Genomic Health, Queen Mary University of London, London EC1M 718 6BQ, UK 719
6. Princess Al-Jawhara Al-Brahim Centre of Excellence in Research of Hereditary 720 Disorders (PACER-HD), King Abdulaziz University, Jeddah, Saudi Arabia 721
7. Epidemiology and Biostatistics, London W2 1PG, UK 722 8. Hygiene and Epidemiology, Ioannina 45110, Greece 723 9. Division of Cardiovascular Medicine, Radcliffe Department of Medicine, University 724
of Oxford, Oxford OX3 9DU, UK 725 10. Wellcome Trust Centre for Human Genetics, University of Oxford, Oxford OX3 726
7BN, UK 727 11. The Farr Institute of Health Informatics Research and the National Institute for 728
Health Research, Biomedical Research Centre, University College London, London, 729 UK 730
12. University College London, London, UK 731 13. CTSU, Nuffield Department of Population Health, University of Oxford, Oxford 732
OX3 7LF, UK 733 14. Division of Cardiovascular Sciences, Faculty of Biology, Medicine and Health, The 734
University of Manchester, Manchester M13 9PT, UK 735 15. Central Manchester University Hospitals NHS Foundation Trust, Manchester 736
Academic Health Science Centre, Manchester M13 9WL, UK 737 16. Cardiovascular Science, National Heart and Lung Institute, Imperial College 738
London, London W12 0NN, UK 739 17. Department of Cardiology, Ealing Hospital, London North West Healthcare NHS 740
Trust, Middlesex UB1 3HW, UK 741 18. Imperial College Healthcare NHS Trust, London W12 0HS, UK 742 19. Department of Cardiovascular Sciences, University of Leicester, Leicester LE3 9QP, 743
UK 744 20. National Institute for Health Research Leicester Cardiovascular Biomedical 745
Research Unit, Leicester LE3 9QP, UK 746 21. Charles Bronfman Institute for Personalized Medicine, Icahn Shool of Medicine at 747
Mount Sinai, NewYork City,NY 10029, USA 748 22. Mindich Child Health Development Institute, Icahn Shool of Medicine at Mount 749
Sinai, New York, NY 10069, USA, NewYork City,NY 10029, USA 750 23. Farr Institute of Health Informatics, UCL, London NW1 2DA, UK 751 24. Bart's Heart Centre, St Bartholomew's Hospital, London Ec1A 7BA, UK 752
Nature Genetics: doi:10.1038/ng.3913

25. Leeds Institute of Biomedical and Clinical Sciences, The University of Leeds, Leeds 753 LS2 9JT, UK 754
26. Cardiovascular, Metabolic and Nutritional Sciences Research Domain/University 755 of Manchester, Manchester M13 9PL, UK 756
27. Manchester Diabetes Centre, Central Manchester University Hospitals NHS 757 Foundation Trust, Mancehster Academic Health Science Centre, Manchester M13 758 0JE, England 759
28. Institute of cardiovascular science, UCL, London, UK 760 29. Department of Health Sciences, University of Leicester, Leicester, UK 761 30. Division of Cardiovascular Sciences Faculty of Biology, Medicine and Health The 762
University of Manchester, Manchester M13 9PT, UK 763 31. Division of Medicine, Central Manchester NHS Foundation Trust, Manchester 764
Academic Health Science Centre, Manchester M13 9PT, UK 765 32. Department of Epidemiology and Biostatistics, Imperial College London, London 766
W2 1PG, UK 767 33. Department of Hygiene and Epidemiology, University of Ioannina Medical School, 768
Ioannina 45110, Greece 769
Nature Genetics: doi:10.1038/ng.3913

Supplementary Table 1 – Comparing the statistical power between the two CAD phenotype 770 definitions in the UK Biobank 771
Supplementary Table 2 –Known CAD loci. Estimates from the SOFT meta-analysis, listing the 772 published and the top variant for the region in the meta-analysis (separate Excel file) 773
Supplementary Table 3 – Comparison of the lambda statistic across meta-analyses 774
Supplementary Table 4 – Variants with an FDR Q-value <5% from the meta-analysis with the SOFT 775 and HARD phenotypes (separate Excel file) 776
Supplementary Table 5 –UK Biobank and replication sample demographics 777
Supplementary Table 6 – Association data per analytical stage for the 13 new lead variants reaching 778 genome-wide significance (separate Excel file) 779
Supplementary Table 7 – GCTA joint model results for independent variants with a 5% FDR 780 (separate Excel file) 781
Supplementary Table 8 – Comparison of genetic risk scores for prediction of CAD in an independent 782 sample 783
Supplementary Table 9 – eQTL data in STARNET and GTEx (separate Excel file) 784
Supplementary Table 10 – Phenome scan for the 13 novel genome-wide significant loci (separate 785 Excel file) 786
Supplementary Table 11 – DEPICT analysis – gene enrichment sets (1% FDR) (separate Excel file) 787
Supplementary Table 12 – Gene networks derived from 5% FDR set 788
Supplementary Table 13 – 61 independent variants from known loci estimated using GCTA 789
790
Supplementary Figure 1 - Manhattan plots of the UK Biobank GWAS using the SOFT (left) and HARD 791 (right) CAD definition 792
Supplementary Figure 2 - Meta-analyses flow chart 793
Supplementary Figure 3 – Replication of overlapping 5% FDR variants from 1000G GWAS in UK 794 Biobank SOFT analysis 795
Supplementary Figure 4 – Cox regression analysis 796
Supplementary Figure 5 – LocusZoom plots of the novel loci 797
Supplementary Figure 6 – Functional annotation of the novel loci 798
Supplementary Figure 7 – Tissue distribution of eQTLs linked to 5% FDR variants in STARNET and 799 GTEx 800
Supplementary Figure 8 – Canonical pathways showing enrichment 801
Supplementary Figure 9 – Depict 802
Supplementary Figure 10 – Odds ratio (OR) vs risk allele frequency plot for independent variants 803 reaching genome-wide significance (p < 5 x10-8) in the joint analysis with GCTA 804
Nature Genetics: doi:10.1038/ng.3913

Supplementary Table 1 - Comparing the statistical power between the two CAD phenotype 808 definitions in the UK Biobank 809
CAD phenotype Odds Ratio No. cases No. controls Power (%) Neff
SOFT 1.059 10,801 137,371 93.3 40,055 HARD 1.072 6,482 137,371 91.4 24,760
810
Forty-six lead variants for GWAS-significant (p-value < 5 x 10-8) loci in the 1000 Genomes CAD 811 meta-analysis2 were selected as informative markers for an exploratory power analysis of two 812 CAD phenotype definitions in the UK Biobank. Median odds ratio across the 46 variants; 813 power for a susceptibility allele frequency = 0.2 and type 1 error (alpha) = 0.05; Neff = effective 814 number of cases and controls = (4 × no. cases × no. controls)/(no. cases + no. controls). The 815 median effect size for the 46 variants in the 48 participating studies in 1000-Genomes CAD 816 meta-analysis was 1.089; range 1.022 – 1.159.817
818
819
820
821
822
823
Supplementary Table 2 - Known CAD loci. Estimates from the SOFT meta-analysis, listing the 824 published and the top variant for the region in the meta-analysis (separate Excel file) 825
826
827
828
829
830
831
832
833
834
835
836
837
838
Nature Genetics: doi:10.1038/ng.3913
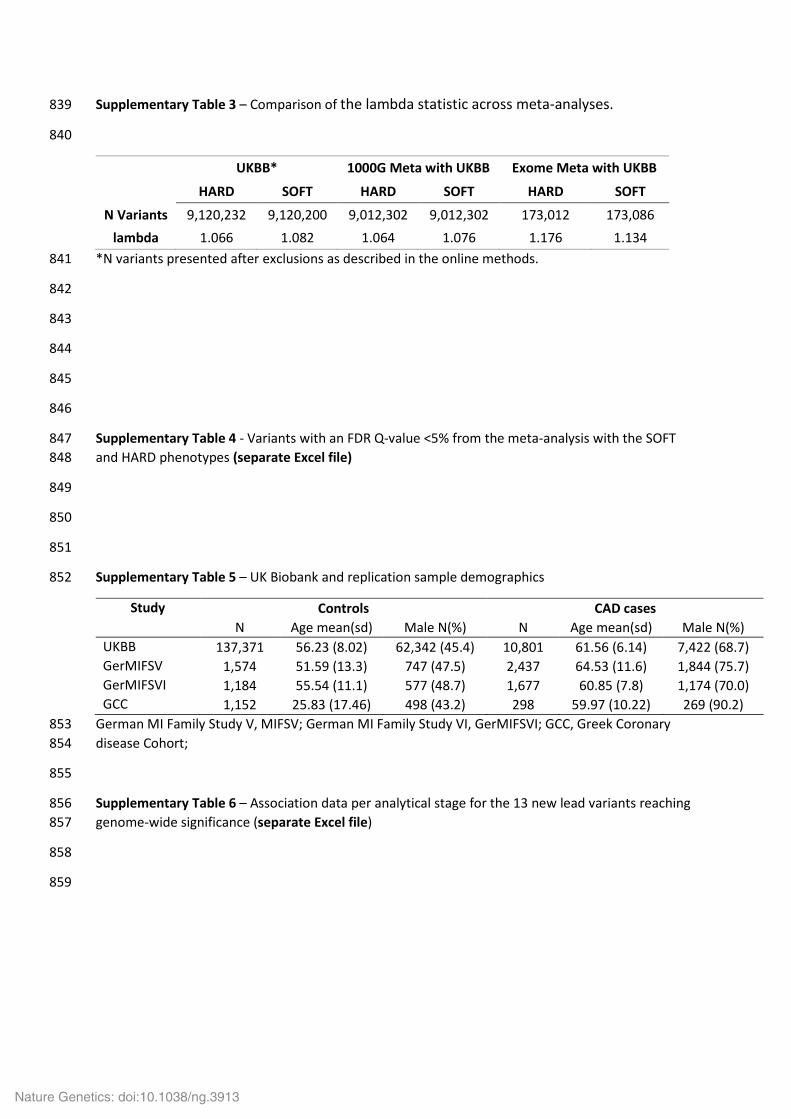
Supplementary Table 3 – Comparison of the lambda statistic across meta-analyses. 839
840
UKBB* 1000G Meta with UKBB Exome Meta with UKBB
HARD SOFT HARD SOFT HARD SOFT
N Variants 9,120,232 9,120,200 9,012,302 9,012,302 173,012 173,086 lambda 1.066 1.082 1.064 1.076 1.176 1.134
*N variants presented after exclusions as described in the online methods. 841
842
843
844
845
846
Supplementary Table 4 - Variants with an FDR Q-value <5% from the meta-analysis with the SOFT 847 and HARD phenotypes (separate Excel file) 848
849
850
851
Supplementary Table 5 – UK Biobank and replication sample demographics 852
Study Controls CAD cases N Age mean(sd) Male N(%) N Age mean(sd) Male N(%)
UKBB 137,371 56.23 (8.02) 62,342 (45.4) 10,801 61.56 (6.14) 7,422 (68.7) GerMIFSV 1,574 51.59 (13.3) 747 (47.5) 2,437 64.53 (11.6) 1,844 (75.7) GerMIFSVI 1,184 55.54 (11.1) 577 (48.7) 1,677 60.85 (7.8) 1,174 (70.0) GCC 1,152 25.83 (17.46) 498 (43.2) 298 59.97 (10.22) 269 (90.2)
German MI Family Study V, MIFSV; German MI Family Study VI, GerMIFSVI; GCC, Greek Coronary 853 disease Cohort; 854
855
Supplementary Table 6 – Association data per analytical stage for the 13 new lead variants reaching 856 genome-wide significance (separate Excel file) 857
858
859
Nature Genetics: doi:10.1038/ng.3913

27
Supplementary Table 7 – GCTA joint model results across 5% FDR SNPs (separate Excel file) 860
861
Nature Genetics: doi:10.1038/ng.3913
28
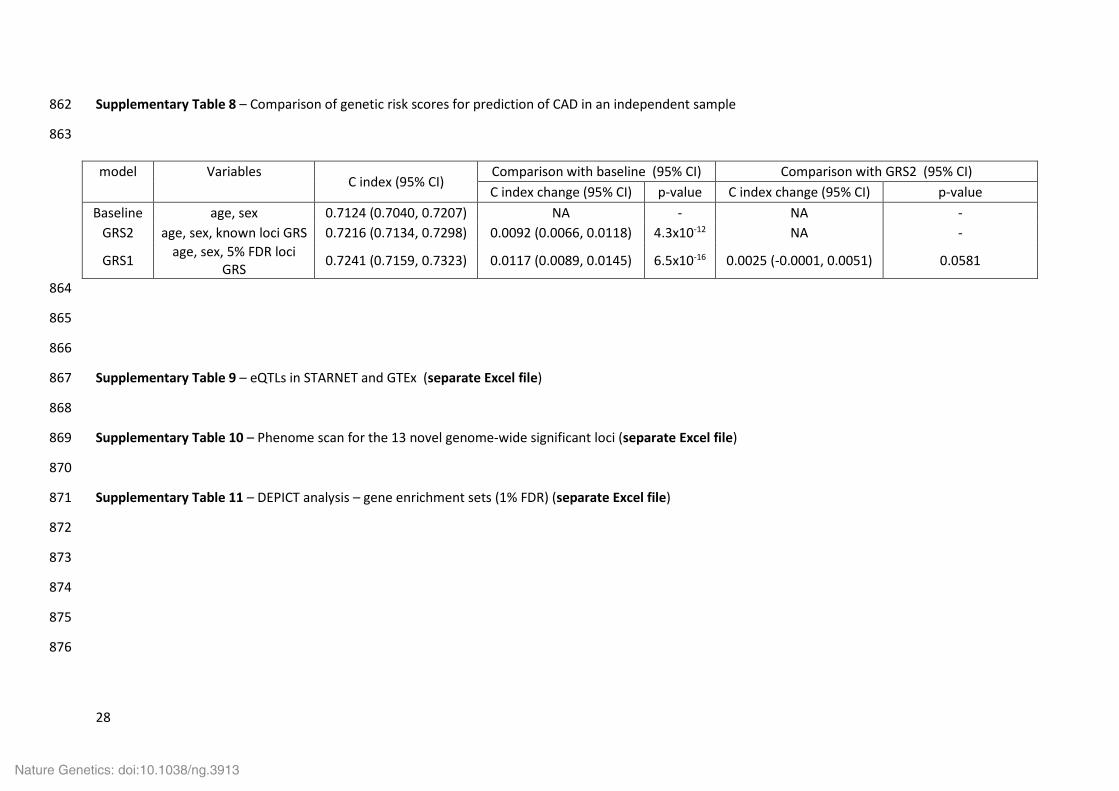
Supplementary Table 8 – Comparison of genetic risk scores for prediction of CAD in an independent sample 862
863
864
865
866
Supplementary Table 9 – eQTLs in STARNET and GTEx (separate Excel file) 867
868
Supplementary Table 10 – Phenome scan for the 13 novel genome-wide significant loci (separate Excel file) 869
870
Supplementary Table 11 – DEPICT analysis – gene enrichment sets (1% FDR) (separate Excel file) 871
872
873
874
875
876
model Variables C index (95% CI)
Comparison with baseline (95% CI) Comparison with GRS2 (95% CI) C index change (95% CI) p-value C index change (95% CI) p-value
Baseline age, sex 0.7124 (0.7040, 0.7207) NA - NA - GRS2 age, sex, known loci GRS 0.7216 (0.7134, 0.7298) 0.0092 (0.0066, 0.0118) 4.3x10-12 NA -
GRS1 age, sex, 5% FDR loci GRS 0.7241 (0.7159, 0.7323) 0.0117 (0.0089, 0.0145) 6.5x10-16 0.0025 (-0.0001, 0.0051) 0.0581
Nature Genetics: doi:10.1038/ng.3913

29
Supplementary Table 12 – Gene networks derived from FDR set. Candidate genes in known CAD risk loci (black) and the 13 new genomewide loci (red) are 877 indicated in bold. Network 1 is composed of 6 interlinked groups (A-F) initially defined by IPA (shaded green). 878
879
ID Molecules in Network Score Focus Molecules Top Diseases and Functions
1A ABCA1,ABCG8,Akt,ANGPTL4,APOA1,APOA5,APOB,APOC1,APOE,ARNTL,C1S, CALCRL,CETP,COL4A1,Collagen(s),CSF3R,ERK1/2,FGF1,FLT1,Focal adhesion kinase,GCK,Growth hormone,HDL,HDL-cholesterol,HMG CoA synthase,HNF1A,IGF2R, IL23,IL6R,IRS1,ITGB5,JINK1/2,KDR,LDL,LDL-cholesterol,LDLR,LIPA,LIPG, LPA,LPL,LRP,LRP1,MAP2K1/2,MFGE8,MMP13,MYH11,N-cor,NEDD9,NOS3, NPC1L1,Nr1h,P38 MAPK,PCSK9,PEMT,PLAUR,PLG,PRKAA,SAA,SCARB1,SERPINA1, SH2B3,Smad1/5/8,SORT1,SRC (family),SWAP70,SYK/ZAP,TPO,TRIB1, VLDL-cholesterol,ZEB2
66 47 Lipid Metabolism, Molecular Transport, Small Molecule Biochemistry
1B ADCY8,Alp,Alpha catenin,AMPK,Ap1,ARNT,CABIN1,Calcineurin protein(s),calpain, CD3,Cg,CLDN1,Collagen Alpha1,Creb,CXCL12,CXCR4,Cyclin A,CYP17A1,DPYSL2, EDN1,EDNRA,ERK,F Actin,Fcer1,FES,FN1,FURIN,Gsk3,Ige,IL12 (complex), Immunoglobulin,Jnk,KSR2,LOX,MAP3K1,Mapk,Mek,MTORC1,NDST2,NFkB (complex), p70 S6k,PAK1,PARP,PDGF BB,PDGFD,PI3K (complex),PI3K (family),Pka,Pkc(s), PLPP3,PRKD2,PTX3,pyruvate kinase,Rac,Ras,Ras homolog,RHOA,RNF216,Rock, RUNX1,SMAD3,TCR,Tgf beta,TGFB1,TWIST1,Ubiquitin,USP10,Vegf,VEGFA,ZFPM2
35 31
Cardiovascular System Development and Function, Organismal Development, Organismal Injury and Abnormalities
1C ABR,ADAMTS7,AGO2,AKT1,AR,ARHGAP26,ATP2B3,BOP1,CACNA1A,CACNA1B,CEP63, CNNM2,COL4A2,Collagen type I,CUL1,Cyfip2,DISC1,EIF3G,GRPEL1,HDAC2,IGF2BP1,IL4, ITGB1,JPH3,KCNJ2,LMOD1,MAP6,MIA3,mir-365,mir-383,mir-548,miR-365-3p (and other miRNAs w/seed AAUGCCC),MORF4L1,MRAS,MRFAP1,MYC,MYOG,NMRK2,p85 (pik3r), PAFAH1B1,PALLD,PHACTR1,PPP2R3A,PRIM2,PTHLH,PTPA,RAB21,RAB39A,RAB3B,RB1, RHOA,RNF14,SERPINH1,SLC22A3,SLC4A10,SLC8A2,SNRPD2,SPECC1L,SRM,SYNJ1,Syt7, THOP1,TMEM131,TRPM3,TSHZ3,USP5,WDR7,WDR12,WDR37,ZC3HC1
29 27 Cellular Function and Maintenance, Molecular Transport, Cellular Assembly and Organization
Nature Genetics: doi:10.1038/ng.3913

30
1D ABCG5,AFF3,ALDH3A2,ANKS1A,ANLN,ANO1,CDKN1A,CLIC5,COL27A1,CREB5,CREBL2, CTNNB1,CTNNBIP1,DCLRE1B,DNAJB4,E2F3,EDARADD,EP300,Eph Receptor,FAM46A, FNDC3B,FOXO1,FRMD4B,GPR137B,GRIK1,HAL,HIST3H2A,HNF1A,HSD17B2,INS, KIAA1462,LBR,LNX1,MAP3K8,MC4R,MSH5,MSI1,MT1X,N4BP2L2,NAB2,NFIB,NUMB, PARP12,PHIP,PIPOX,PLEKHA1,PLEKHF1,Proinsulin,Ptprv,RAB37,RAB3A,RCSD1,RDH10,REL, SEMA5A,SLC6A19,SMG6,SVEP1,TAT,TDRKH,TERC,TGFBR2,TGS1,TINF2,TMPRSS4,TNS1, UBE2Z,Ubiquitin,WNT16,ZHX3
26 25 Cellular Assembly and Organization, Endocrine System Disorders, Organ Morphology
1E ABLIM1,ADAM19,AGTR1,AHR,BTG3,CELSR2,CEP170,Cg,CKAP2,CLEC4G, Cma2/Mcpt9,CNPY4,CTPS1,CTRB2,DGKH,DMAP1,DNMT1,DYM,EPHA3,ERLIN2,FBN2, FCHO1,FIP1L1,GATA2,GCH1,HMGCR,HR,IFNG,Jnk,KCNA4,KCTD3,LBR,LCN2,LIMA1, LRP12,LRP1B,LYZL1,MAGI3,MAP3K2,MBNL1,MCTP1,MRPS6,NDE1,NDFIP2,Pak2, PAX7,PKIA,PMM1,PPARG,PRSS2,RAB11FIP5,RHBDD3,SDC3,SEMA3E,SLC44A1, SLC52A2,SRC,TAGLN2,TESPA1,THRB,TNFSF18,TUBA8,UHRF1BP1,USP24,VAMP5, VAMP8,VANGL2,WNT2,YWHAG,ZNF703
23 23 Neurological Disease, Psychological Disorders, Hereditary Disorder
1F A4GALT,ALDH,AMPK,ANXA13,BCL2,C1GALT1,CDC123,CDH13,CENPH,coagulation factor,CSF2,CTR9,DMD,EGFR,ERO1B,FBXL17,FMO1,GGCX,GIGYF2,GKAP1,HCAR3, HERC1,IL6,IL34,IL1RAPL1,INSR,IRS1,ITLN1,KCNK5,KIAA0430,KIAA1324,LGALS4,LILRA5, mir-127,mir-7578,miR-434-3p (miRNAs w/seed UUGAACC),MRVI1,NEU3,NIPSNAP1, NPY5R,NSG1,NT5C2,OGDH,PASK,PCDH7,PDX1,PLCB3,PTGDR,PTPRZ1,PXK,RASD1,RCN3, RECQL4,RHBDF2,RNF149,RSPO3,SIK2,Sos,SYT11,TCR,TNF,TNR,TPST1,WTAP,XBP1,YY1, ZC3H12D,ZFP64,ZHX2,ZPR1
23 23 Cell Cycle, Gastrointestinal Disease, Inflammatory Disease
2 ABLIM,Actin,ADCY9,AGAP1,ARVCF,ATP2B1,Calmodulin,CAMK1D,CORO6,creatine kinase,DAB2IP,DUSP3,ERG,FGF5,FHL3,FOXC1,FSH,GNLY,GOT,GUCY1A3,Hdac,HDAC9, Histone h3,Histone h4,HTRA1,Iga,IgG,IgG1,Igl,Igm,ING1,KCNE2,KLF4,Lh, LOC81691, MAD2L1,MAP9,MAPRE2,MT3,Notch,OSR1,PADI3,PAPPA,PECAM1,POLQ,POP5,PPFIA4, PREX1,PRKX,PSG5,PTPN21,RBM15,RCOR1,REST,RNA polymerase II,Secretase gamma,SH3BP4,SKI,SLC22A1,SLC22A5,SMARCA4,STK17A,TCF,TCF21,TIMELESS,TLK1, TMED10,UTP14C,WLS,ZFP36L2
42 35 Cardiac Hypertrophy, Cardiovascular Disease, Developmental Disorder
Nature Genetics: doi:10.1038/ng.3913

31
3 ABHD2,ANGPTL6,ANXA8/ANXA8L1,ARPP19,ASL,ATXN2,BBS10,BCAS3,BRS3,CAV1, CD300LF,CEBPB,CERS6,CLDN5,CRBN,CRTC3,ELL,FFAR3,FHL5,FKBP6,FSTL3,GSTP1, IL12 (family),IL1F10,INHBC,Ins1,IP6K2,KPNA4,KRAS,LEP,LGALS12,MAFB,MCHR1,mir-155, mir-182,miR-146a-5p (and other miRNAs w/seed GAGAACU),miR-292b-5p (and other miRNAs w/seed CUCAAAA),MTRR,NAB1,NMU,NPBWR1,Npy4r,PDE10A,PGLYRP2,PPFIBP1, PRDX4,PRKAB2,PRLH,PTRF,RAI1,RNF2,RPN2,SCMH1,SERP1,SETD4,SIM1,SLC19A2,SLC22A4,SLC2A12,SLC5A3,SLC7A11,SLURP1,SP1,SRP54,SSFA2,ST5,TIMP2,TINAGL1,TP53, ZNF652
24 24 Protein Synthesis, Endocrine System Development and Function, Molecular Transport
4 CEBPA,THOC5 1 1 Cellular Development, Hematological System Development and Function, Hematopoiesis
5 ARHGEF26,Dlg 1 1 Cardiovascular System Development and Function, Cellular Function and Maintenance, Tissue Development
880
Nature Genetics: doi:10.1038/ng.3913

32
Supplementary Table 13 – 61 Independent variants from known loci estimated using GCTA 881
Markername Chr:Pos EA/OA EAF Joint OR (95% CI)
Joint Pvalue RefSeq name
rs10840293 11:9751196 A/G 0.552 1.05 (1.03 , 1.07) 6.71E-09 SWAP70 rs6689306 1:154395946 A/G 0.435 1.05 (1.03 , 1.07) 1.63E-09 IL6R rs13200993 6:22612912 T/C 0.348 1.05 (1.03 , 1.07) 3.00E-09 HDGFL1 rs2244608 12:121416988 G/A 0.335 1.05 (1.03 , 1.07) 2.81E-09 HNF1A rs10139550 14:100145710 G/C 0.421 1.05 (1.04 , 1.07) 1.79E-09 HHIPL1 rs112470402 1:57016950 I/D 0.746 1.05 (1.03 , 1.07) 4.99E-08 PPAP2B rs1870634 10:44480811 G/T 0.648 1.05 (1.04 , 1.07) 7.03E-10 LOC283033 rs10857147 4:81181072 T/A 0.277 1.06 (1.04 , 1.08) 8.73E-09 FGF5 rs4299376* 2:44072576 G/T 0.319 1.06 (1.04 , 1.08) 5.50E-10 ABCG8 rs1887318 10:30321598 T/C 0.428 1.06 (1.04 , 1.08) 3.99E-12 KIAA1462 rs11810571 1:151762308 G/C 0.807 1.06 (1.04 , 1.08) 2.47E-08 TDRKH rs7177201 15:79065380 T/C 0.320 1.06 (1.04 , 1.08) 4.89E-08 ADAMTS7 rs4773141 13:110954353 G/C 0.360 1.06 (1.04 , 1.08) 1.13E-09 COL4A1 rs7500448 16:83045790 A/G 0.763 1.06 (1.04 , 1.08) 5.01E-09 CDH13 rs2839812 11:103673294 T/A 0.307 1.06 (1.04 , 1.08) 1.92E-11 MIR4693 rs7164479 15:79123054 T/C 0.578 1.06 (1.04 , 1.08) 4.20E-12 ADAMTS7 rs2954029* 8:126490972 A/T 0.541 1.06 (1.04 , 1.08) 5.06E-13 TRIB1 rs7568458 2:85788175 A/T 0.452 1.06 (1.05 , 1.08) 2.30E-13 GGCX rs9515203 13:111049623 T/C 0.752 1.06 (1.04 , 1.08) 7.32E-10 COL4A2 rs2071382 15:91428197 T/C 0.464 1.06 (1.05 , 1.08) 6.90E-13 FES rs7435973 4:156436507 G/A 0.139 1.07 (1.04 , 1.09) 4.85E-08 MAP9 rs4593108 4:148281001 C/G 0.805 1.07 (1.04 , 1.09) 5.79E-10 EDNRA rs10774625* 12:111910219 A/G 0.491 1.07 (1.05 , 1.08) 1.02E-13 ATXN2 rs585967 2:21270554 C/A 0.844 1.07 (1.04 , 1.09) 3.11E-08 APOB rs2681472 12:90008959 G/A 0.189 1.07 (1.05 , 1.09) 7.38E-11 ATP2B1 rs12202017 6:134173151 A/G 0.702 1.07 (1.05 , 1.09) 5.78E-14 MGC34034 rs11556924 7:129663496 C/T 0.657 1.07 (1.05 , 1.09) 6.04E-13 ZC3HC1 rs3796587 4:156638073 C/G 0.816 1.07 (1.05 , 1.09) 1.13E-10 GUCY1A3 rs62265630 3:124475201 G/T 0.141 1.07 (1.05 , 1.09) 9.82E-09 ITGB5 rs11057830* 12:125307053 A/G 0.147 1.07 (1.05 , 1.10) 4.45E-09 SCARB1 rs1657345 10:44779078 A/G 0.865 1.07 (1.05 , 1.10) 5.95E-09 CXCL12 rs1250229* 2:216304384 T/C 0.262 1.07 (1.05 , 1.09) 1.78E-13 FN1 rs8068952* 17:59286644 G/C 0.228 1.07 (1.05 , 1.10) 1.38E-09 BCAS3 rs56131196 19:45422846 A/G 0.174 1.07 (1.05 , 1.10) 2.72E-09 APOC1 rs12493885* 3:153839866 C/G 0.863 1.07 (1.05 , 1.10) 3.22E-08 ARHGEF26 rs67180937 1:222823743 G/T 0.684 1.07 (1.05 , 1.09) 8.13E-14 MIA3 rs72743461 15:67441750 C/A 0.783 1.07 (1.05 , 1.10) 4.64E-12 SMAD3 rs7623687 3:49448566 A/C 0.859 1.07 (1.05 , 1.10) 3.63E-09 RHOA rs11191416 10:104604916 T/G 0.890 1.08 (1.05 , 1.10) 5.45E-09 CYP17A1 rs6841581 4:148401190 A/G 0.153 1.08 (1.05 , 1.10) 2.95E-11 EDNRA rs507666* 9:136149399 A/G 0.192 1.08 (1.06 , 1.10) 1.29E-12 ABO rs2107595 7:19049388 A/G 0.182 1.08 (1.06 , 1.10) 3.29E-13 HDAC9 rs139016349 3:138099161 I/D 0.163 1.08 (1.05 , 1.10) 8.92E-12 MRAS rs2246942* 10:91004886 G/A 0.349 1.08 (1.06 , 1.10) 3.36E-16 LIPA rs3130683* 6:31888367 T/C 0.860 1.08 (1.05 , 1.11) 2.31E-08 C2 rs11617955 13:110818102 T/A 0.891 1.09 (1.06 , 1.12) 7.11E-10 COL4A1 rs16986953* 2:19942473 A/G 0.073 1.11 (1.07 , 1.15) 5.36E-10 FLJ12334 rs9349379* 6:12903957 G/A 0.407 1.11 (1.09 , 1.13) 6.11E-36 PHACTR1
Nature Genetics: doi:10.1038/ng.3913

33
rs56170783 1:57016131 A/C 0.915 1.11 (1.08 , 1.15) 5.81E-13 PPAP2B rs7528419 1:109817192 A/G 0.782 1.11 (1.09 , 1.14) 3.57E-27 CELSR2 rs114123510 2:203831212 A/T 0.116 1.13 (1.10 , 1.15) 2.74E-19 ALS2CR8 rs3918226 7:150690176 T/C 0.071 1.13 (1.09 , 1.17) 1.53E-12 NOS3 rs6511720* 19:11202306 G/T 0.884 1.14 (1.11 , 1.17) 7.51E-22 LDLR rs7412* 19:45412079 C/T 0.922 1.14 (1.11 , 1.18) 2.01E-16 APOE rs28451064 21:35593827 A/G 0.124 1.14 (1.11 , 1.17) 2.49E-23 NCRNA00310 rs180803 22:24658858 G/T 0.977 1.18 (1.12 , 1.24) 6.89E-10 POM121L9P rs2891168 9:22098619 G/A 0.487 1.19 (1.17 , 1.21) 1.51E-101 CDKN2B-AS1 rs11591147* 1:55505647 G/T 0.984 1.25 (1.17 , 1.34) 1.61E-10 PCSK9 rs10455872 6:161010118 G/A 0.065 1.32 (1.28 , 1.37) 8.13E-53 LPA rs147555597 6:160911596 A/G 0.009 1.37 (1.24 , 1.51) 2.09E-10 LPAL2 rs186696265 6:161111700 T/C 0.014 1.62 (1.51 , 1.75) 1.45E-38 PLG
*Exome marker 882
Nature Genetics: doi:10.1038/ng.3913

34
Supplementary Figure 1 - Manhattan plots of the UK Biobank GWAS using the SOFT (left) and HARD (right) CAD definition respectively 883
884
885
Nature Genetics: doi:10.1038/ng.3913

35
Supplementary Figure 2 – Meta-analyses & Replication flow chart 886
887
Nature Genetics: doi:10.1038/ng.3913
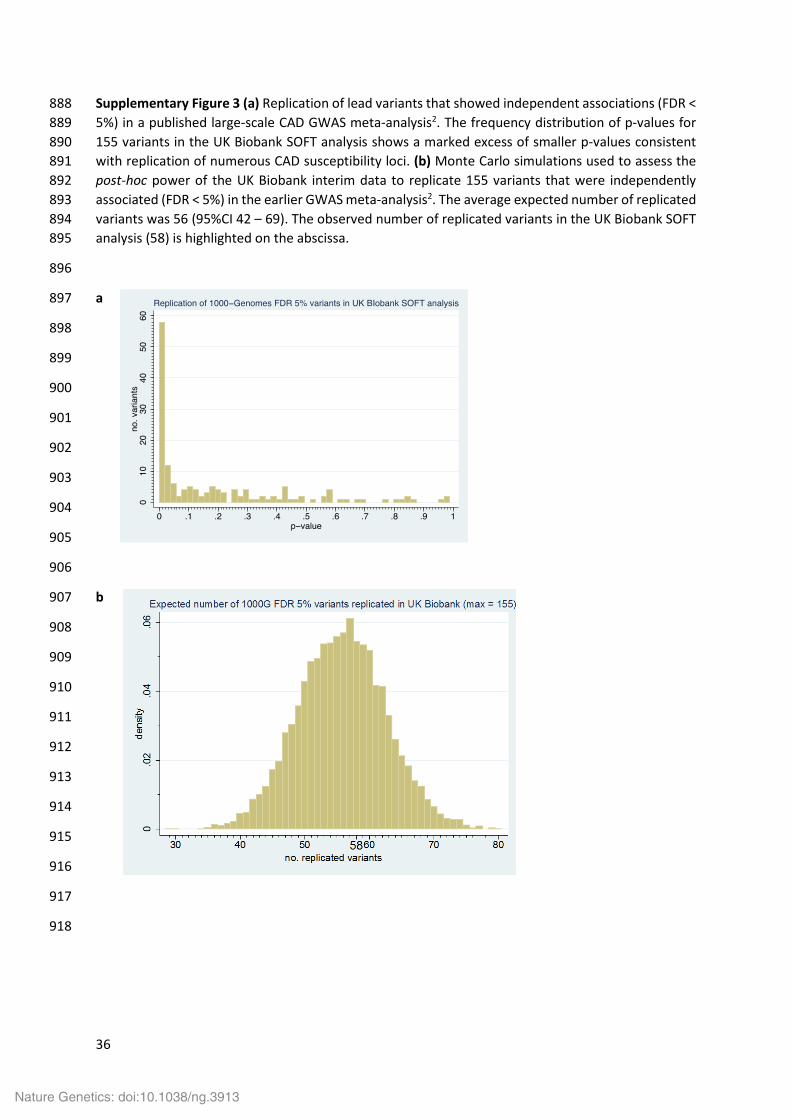
36
Supplementary Figure 3 (a) Replication of lead variants that showed independent associations (FDR < 888 5%) in a published large-scale CAD GWAS meta-analysis2. The frequency distribution of p-values for 889 155 variants in the UK Biobank SOFT analysis shows a marked excess of smaller p-values consistent 890 with replication of numerous CAD susceptibility loci. (b) Monte Carlo simulations used to assess the 891 post-hoc power of the UK Biobank interim data to replicate 155 variants that were independently 892 associated (FDR < 5%) in the earlier GWAS meta-analysis2. The average expected number of replicated 893 variants was 56 (95%CI 42 – 69). The observed number of replicated variants in the UK Biobank SOFT 894 analysis (58) is highlighted on the abscissa. 895
896
a 897
898
899
900
901
902
903
904
905
906
b 907
908
909
910
911
912
913
914
915
916
917
918
Nature Genetics: doi:10.1038/ng.3913

37
Supplementary Figure 4 – Cox regression analysis using a Genetic Risk Score (GRS) based on either all genome-wide significant SNPs (known and 13 new 919 CAD risk loci) or the 5% FDR SNPs (Supplementary Table 7) in the EPIC-CVD study. Both GRS were adjusted for age and sex. HR, Hazard ratio; CI, Confidence 920 Interval 921
922
923
924
925
HR (95% CI) p-value HR (95% CI) p-valueQ1 vs ref 0.690 (0.619, 0.770) 3.00E-11 0.729 (0.653, 0.813) 1.50E-08Q2 vs ref 0.912 (0.818, 1.016) 9.50E-02 0.831 (0.746, 0.926) 7.80E-04Q4 vs ref 1.044 (0.935, 1.165) 4.50E-01 1.176 (1.055, 1.311) 3.40E-03Q5 vs ref 1.316 (1.181, 1.467) 7.20E-07 1.315 (1.179, 1.468) 9.90E-07
Genome-wide SNPs (new & known)
5% FDR SNPs
926
927
Nature Genetics: doi:10.1038/ng.3913

38
Supplementary Figure 5 – Regional association plots of the novel loci drawn with LocusZoom 928
929
930
Nature Genetics: doi:10.1038/ng.3913

39
931
932
933
Nature Genetics: doi:10.1038/ng.3913
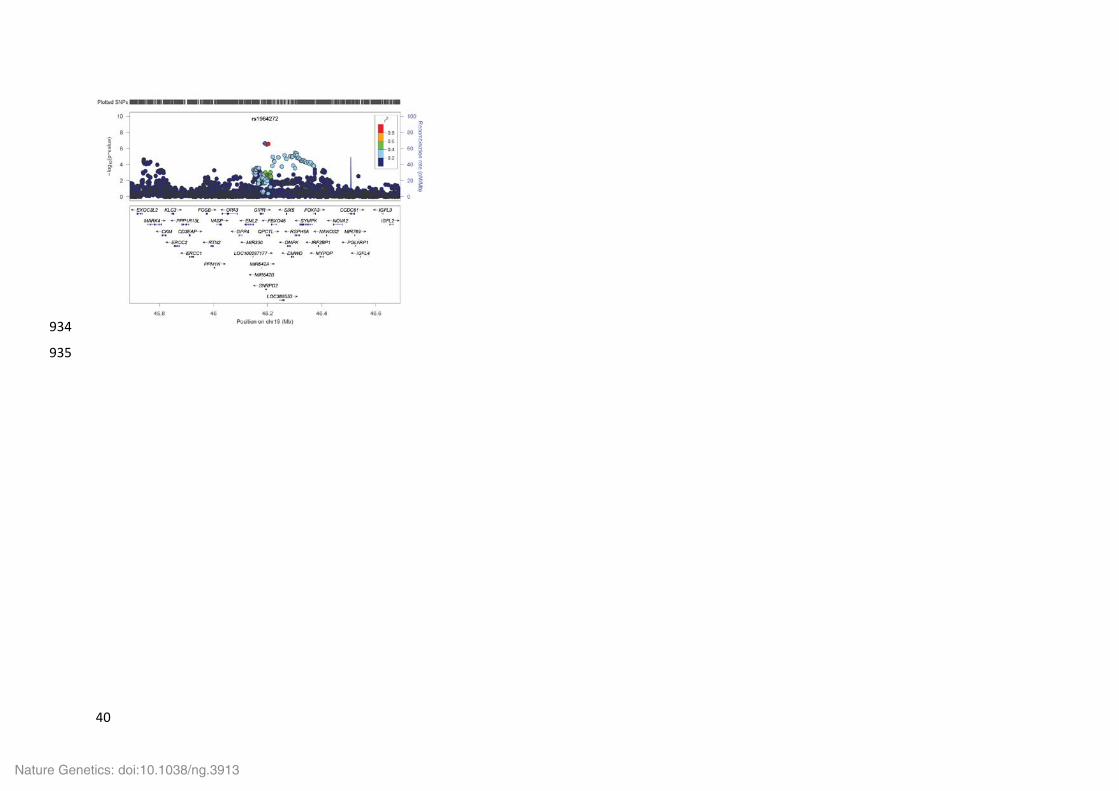
40
934
935
Nature Genetics: doi:10.1038/ng.3913

41
Supplementary Figure 6 – Functional annotation of the novel loci. Each panel shows a UCSC genome browser (hg19) screengrab of the CAD 936 associated variants and nearby genes. Regulatory annotation generated using Haploreg V4 is shown in tabular form and summarises the 937 regulatory features overlapping each CAD associated variant including promoter and enhancers based on chromatin state segmentation from 938 ENCODE and Roadmap epigenome, DNase hypersensitivity, bound proteins and predicted disruption of transcription factor binding motifs. 939 Functional prediction scores for each variant from CADD, GWAVA and RegulomeDB are also shown. Due to the high number of variants at 940 some loci only selected SNPs are shown. 941
Nature Genetics: doi:10.1038/ng.3913

42
942
Nature Genetics: doi:10.1038/ng.3913

43
943
Nature Genetics: doi:10.1038/ng.3913

44
944
Nature Genetics: doi:10.1038/ng.3913
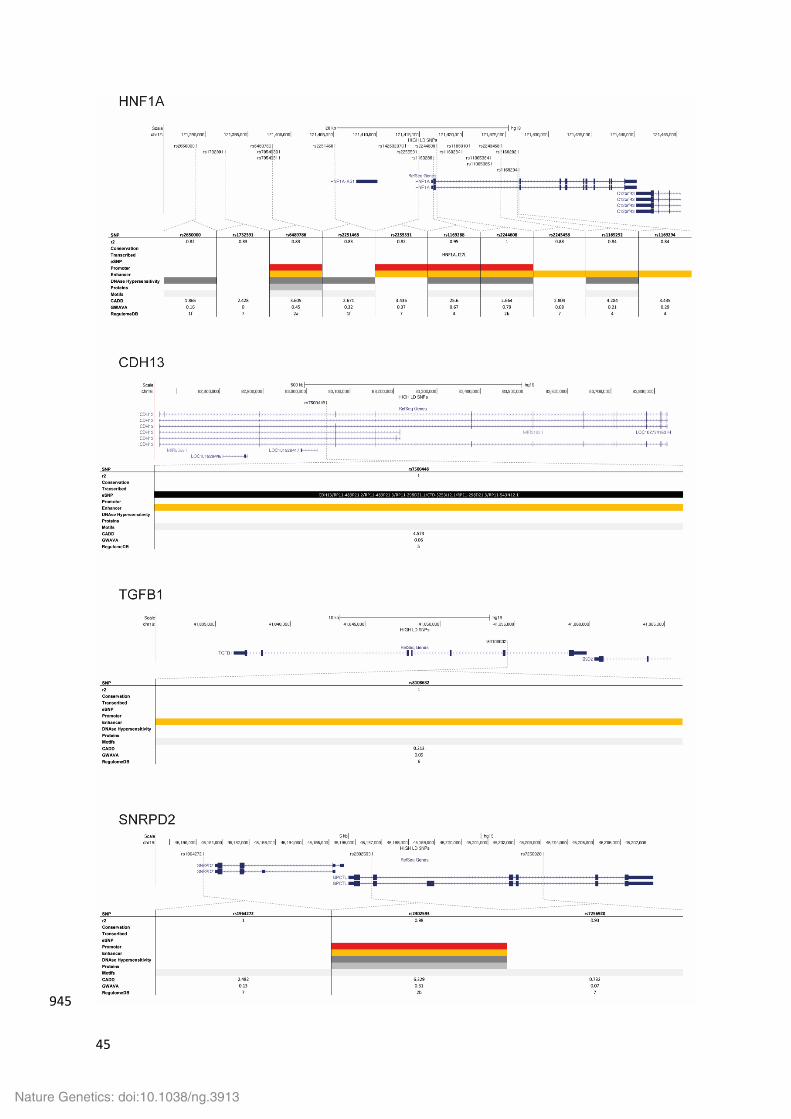
45
945
Nature Genetics: doi:10.1038/ng.3913

46
Supplementary Figure 7 – Tissue distribution of eQTLs linked to 5% FDR variants in STARNET and 946 GTEx. eQTLs were from the Stockholm-Tartu Atherosclerosis Reverse Network Engineering 947 Task (STARNET) RNA-seq dataset13 and the Genotype-Tissue Expression15 (GTEx) portal. 948 AOR, aortic arterial wall; SF, subcutaneous fat; MAM, mammary artery; SKLM, skeletal muscle; 949 VAF, visceral abdominal fat; LIV, liver. eQTLs were included if the best eSNP (i.e. the variant 950 with the most significant association with gene expression in cis) was in high LD (r2>0.8) with 951 the CAD lead SNP. 952
953
Nature Genetics: doi:10.1038/ng.3913

47
Supplementary Figure 8 – Canonical Pathways showing enrichment among the 357 genes derived from the 5% FDR SNP list. Pathways involved in lipid 954 metabolism and angiogenesis are indicated by red and green arrows respectively. 955
956
957
958
959
960
961
962
Nature Genetics: doi:10.1038/ng.3913
48
Supplementary Figure 9 – Heat map showing DEPICT gene set enrichment analysis results. (a) Full set 963 of 556 gene sets with evidence of enrichment at 1% FDR as listed in Supplementary Table 11. The x – 964 axis shows the gene name, which is predicted to be included to the reconstituted gene set indicated 965 in the y – axis. The color red indicates higher Z-score, where Z-score is a value representing each gene’s 966 inclusion in DEPICT’s reconstituted gene sets. Clustering was made based on complete linkage 967 method. (b-c) Zoom-ins to two additional heat map clusters to the one shown in Figure 4. 968
a 969
970
971
Nature Genetics: doi:10.1038/ng.3913

49
b 972
973
974
975
976
977
978
979
980
981
982
983
984
985
986
987
988
989
990
991
992
Nature Genetics: doi:10.1038/ng.3913

50
c 993
994
995
996
997
998
Nature Genetics: doi:10.1038/ng.3913

51
Supplementary Figure 10 – Odds ratio (OR) vs risk allele frequency plot for independent variants 999 reaching genome-wide significance (p < 5 x10-8) in the joint analysis with GCTA (Supplementary 1000 Table 13). For intergenic SNPs flanking genes are shown. 1001
1002
Nature Genetics: doi:10.1038/ng.3913
Top Related