






Languages
Pages
Legal


SKELETAL SYSTEM: APPENDICULAR
SKELETON
Mrs. Ofelia Solano Saludar
Department of Natural Sciences University of St. La Salle
Bacolod City

Appendages and supporting girdles: pectoral and pelvic

The appendicular skeleton has a different embryonic origin than the axial skeleton.
Somatic mesoderm of the lateral plate contributes to the limb bud mesenchyme.
Initially, mesenchyme of the limb bud consists solely of this lateral plate origin.
As the basic skeletal plan is laid out, mesenchymal cells from the somites migrate in to form muscle cells, and neural crest cells migrate in to form both nerves and pigment cells.

1. Gill arch hypothesis - fins derived from the last 2 gill arches
2. Fin spine hypothesis - fins derived from tissue attached to spines that may have evolved to provide protection from predators (Acanthodians)
3. Fin-fold hypothesis- continuous finfolds became discontinuous from loss
ORIGIN OF PAIRED APPENDAGES

EVIDENCES FOR THE FIN-FOLD HYPOTHESIS:
Development and early structure of paired & unpaired fins are identical
In Acanthodians, the fin fold became discontinuous with each segment supported by a rigid spine; placoderms had a pair in the front of the body, and a pair at the rear.
Ammocoetes larva had continuous dorsocaudal fin folds
Ventrolateral fin folds found in jawless ostracoderms helped stabilize body position during swimming.

*The number of muscle buds & nerve branches to the
embryonic paired fins involve more segments than in the
adult*Shape of the fins &
parallelism of fin rays
suggest origin from a
continuous fin fold.

Steering, braking, controlling inclination while horizontal swimming, & as stabilizers that prevent rolling; forward thrust by lateral undulation of body
Pterygiophores (bone or cartilage) provide base support for fin rays: Basalia- enlarged, proximal;
3 types: pro, meso, metapterygia
Radialia- slender, distal Found in all fishes except
Agnathans where there are no paired fins
Slight development of pectoral fins, small musculature & absence of jaws are features of bottom-dwelling forms.
FINS

Fins are stiffened by dermal rays:
1. Lepidotrichia- jointed bony dermal scales; teleosts
2. Ceratotrichia- cartilaginous unsegmented rays; chondrichthyes
3. Actinotrichia- delicate distal rays; found in both types

1. Archipterygial fins are symmetrical along the central axis (modern fishes); supporting skeleton and musculature are inside the body wall of the fish.
2. Metapterygial fins are assym- metrical along the central axis; the skeletal and muscular support are outside of the body wall; were preadapted to bear weight and used for locomotion by fishes wallowing in shallow mud flats or temporary water bodies.

1. Pectoral & Pelvic fins: Lobed fins- fleshy proximal lobe consisting
of fin skeleton, muscles, fin rays; sarcopterygians
Fin folds- broad base; basalia of pelvic fins are modified as claspers to transfer sperm to the female in male chondrichthyans
Ray fins- lost basals, flexible; teleosts
PAIRED FINS

Heterocercal
placoderms, sharks
HomocercalteleostsDiphycercal
Dipnoans & Latimeria
3. Caudal fins:
2. Median fins: Dorsal fins used as keels, rarely used for
locomotionAnal fins- modified as gonopodium;
analogous to claspers

OSTEICHTHYES
Pectoral fin
Pelvic fin
Anal fin
Caudal finDorsal fins


Fins on opposite
sides of the body attach to GIRDLES that in turn attach the fins to the
axial skeleton and to each
other.

PECTORAL GIRDLEOther than locomotion, it also shields the heart, forms the back of the oral cavity and acts as an attachment site for jaw musculature. Membrane bones: cleithrum, supracleithrum, postcleithrum, posttemporal (anchors to skull), clavicle, interclavicleReplacement bones: Coracoid, Scapula

PECTORAL GIRDLES: FROM FISH TO TETRAPOD PLACODERMS- first to have girdles. Pectoral
girdle was the scapulocoracoid with an articular fossa that received basal pterygiophores of fins. Pelvic girdle was a single endoskeletal element.
SARCOPTERYGIANS- scapulocoracoid, cleithrum, post- temporal, clavicle & interclavicle bones
CROSSOPTERYGIANS: clavicle, cleithrum, scapulocoracoid

FISHES: scapula & coracoid receive
the forces transmitted to the
trunk; posttemporal
braces the girdles against the caudal angles of the skull;
clavicle braces against its
opposite in a midventral symphysis

CHONDRICHTHYES Primitive sharks had pectoral & pelvic fins
consisting of basals & tightly packed radials
Girdles consisted of a single enlarged basal element similar to placoderms.

In modern sharks the paired basal components extend to
the body midline which fuse to form a U-shaped
scapulocoracoid bar for the pectoral girdle and a
puboischiadic bar for the pelvic girdle.

The pectoral girdle is cartilaginous, with no dermal elements: coracoid, scapula, suprascapula (scapulacoracoid)
It does not connect to axial skeleton, but fused at midline

TETRAPODS: Clavicle- missing in limbless forms; present in
most mammals; furculum (wishbone) of birds Coracoids- procoracoids & coracoids ossify in
coracoid plate; assist or replace the clavicles in bracing against the reptilian & avian sternum; vestigial coracoid process of the scapula in eutherians
Scapula- present in all tetrapods; suprascapula typically fuses with scapula
Dermal bones predominate in the pectoral girdle of bony fishes; replacement bones predominate in tetrapods.
Tetrapods never brace their pectoral girdles against the skull or vertebral column.

AMPHIBIANS: Acquired interclavicle (episternum in
tetrapods), lost posttemporal, supracleithrum & cleithrum
Replacement bones: coracoid, scapula & suprascapula
Urodeles have no membrane bones Anurans have clavicle, no interclavicle
REPTILES: Scapula, coracoid, sometimes clavicle, &
interclavicle Crocodiles have reduced clavicle Turtles have interclavicle fused with shell Snakes have no girdle Lizards have a significant clavicle,
interclavicle

BIRDS: 2 clavicles plus interclavicle form
furcula; bladelike scapula; procoracoid
articulates with sternum

MAMMALS A new coracoid is formed. The scapula enlarges & the coracoid is
reduced as the coracoid process. The interclavicle bone persists in therapsid
reptiles & monotremes, but is lost in marsupials and eutherians.

The clavicle is large in digging, climbing, or flying forelimbs
Reduced or absent in felines (leaping dissipates impact on muscles), cetaceans (fishlike), ungulates (facilitates grazing)

THERAPSIDS, MONOTREMES: clavicle, procoracoid, coracoid, scapula
EUTHERIAN MAMMALS: scapula is divided by scapular spine into supraspinous & infraspinous fossae to accommodate origins of strong muscles that insert on humerus; acromion process articulates with clavicle

PHYLOGENY
PECTORAL
GIRDLE

PECTORAL GIRDLES OF SELECTED VERTEBRATES

FISHES Consists of 2 cartilaginous or bony plates
(ischiopubic plates) that articulate with the pelvic fins )
Usually meet ventrally in a symphysis, or form a median bar in sharks & lungfishes

Arose from pterygiophores supporting fins
Brace posterior paired appendages & enclose pelvic cavity organs
No dermal components (unlike pectoral girdle)
Sarcopterygians: one bone embedded in the body wall. The left & right pelvic girdles do not meet at midline & there is no connection to the axial skeleton
Crossopterygians: single element with processes
PELVIC GIRDLE

Ichthyostega, an amphibian in the fossil record, had 3 fused bones in the pelvic girdle: pubis, ischium & the ilium, attached to vertebral column by sacral ribs.
In frogs & toads, the ilia are elongated & extend from sacral vertebra to urostyle; sacroiliac joint between ilium & sacral vertebra is freely moveable & moves when a frog or toad jumps

REPTILESStronger sacroiliac joint; ilium & ischium
are expanded to accommodate musculature
for more muscle attachment & stability needed for bipedal locomotion

BIRDS: Girdles braced
against lumbar & sacral vertebrae
Pubic bones are typically long & thin
Synsacrum – Ilium is braced against fused vertebrae
Limited pubic symphysis provides a larger outlet for eggs

MAMMALS Ilium, ischium,
and pubis unite to form the innominate bone (coxa)
Encompass a pelvic cavity
Epipubic bone in marsupials is unique for pouch support

PHYLOGENY
PELVIC
GIRDLE

Terrestial vertebrates need stronger girdles and limbs. Pectoral girdle
became detached from the skull providing a neck region for better head mobility
Limbs plus lateral undulations, should provide points of pivot for trunk to move
TRANSITION TO LAND:

The Labyrinthodont amphibians probably evolved from a Crossopterygian ancestor. When the fresh
water pools in which these fish lived became stagnant, they may have crawled up the bank to
breath air using primitive lungs. As the lobed fins of these fish evolved into stronger limbs, the first
tetrapods appeared.
ANCESTRAL TETRAPOD LIMBS

EARLY TETRAPOD LIMBS ARE
MODIFICATIONS OF RHIPIDISTIAN FINS:
Loss of fin rays and distal radials produced the first tetrapod limb.
Digits are a novel feature of tetrapods and are not modifications of radials.
Girdles remained fishlike.

In the pectoral fins of rhipidistian crossopterygians, a single basal bone
(humerus), articulates with the scapula and distally with a pair of radials (radius & ulna).

Trot- diagonally opposite feet meet the ground, the center of mass lies on the line connecting the 2 pts. of support; a 3rd point of support on a long tail further stabilizes gait.
Lateral-sequence gait- center of mass inside triangle of support
Limb rotation- muscles rotate long bones to retract feet & propel body forward.
In early tetrapods, limbs were short & first
segment extended
straight out from the body.
As such, their primitive gaits include:

From a sprawled position, tetrapods can change their position by drawing limbs under the body. The sprawled posture brings a medially directed force towards the pectoral girdle. As limbs are brought under the body, forces shift vertically, accounting for phylogenetic loss of some pectoral elements.

This change in limb posture results from torsion of the distal ends of long bones so that they are nearly parallel the vertebral column. This brings the digits
forward and in line with direction of travel.

TETRAPOD LIMBS
Some have lost one or both pairs; in others, one pair is modified as arms, wings, or paddles; typically have 5
segments: Stylopodium: proximal Zeugopodium: middle Mesopodium: carpals & tarsals Metapodium: metacarpals &
metatarsals Phalanges: bones of the digits
(fingers, toes, claws)

PROPODIUM or Anterior Limb:
1. Brachium (upper arm) – humerus2. Antebrachium (forearm) - radius &
ulna 3. Carpus (wrist) – proximal row:
radiale, ulnare, intermedium, & pisiform ; middle row: 3 centrals; distal row: starting on radial side: 1-5 distals
4. Metacarpus (palm) – metacarpals 5. Digits or phalanges - general
formula starting at thumb: 2,3,4,5,3
Manus, autopodium, forefoot or hand: includes mesopodium, metapodium, phalanges

FORELIMBS
OF
SELECTED VERTEBRATES

EPIPODIUM or Posterior
limb1. femur (thigh) - femur 2. crus (shank) - tibia &
fibula 3. tarsus (ankle) - tarsals 4. metatarsus (instep) -
metatarsals 5. digits - phalanges
Pes, autopodium, hindfoot or foot: includes
mesopodium, metapodium, phalanges

Some lack both pairs of limbs: caecilians (apodans), most snakes; snake-like lizards
Some have forelimbs only: manatees & dugongs, dolphins; cetaceans, sirenians have vestigial elements embedded in body wall

A reduced number of ankle bones in birds have fused with the tibia & metatarsals to add an elongated segment to the hindlimbs.
An intratarsal joint adds to the flexibility of the pes.
Digit 2 is long digit of wing
Phalangeal formula for foot is 2,3,4,5,0

PES OF SELECTED VERTEBRATES

• Walking, running, cursorial (terrestrial, usually quadrupedal)
• Many specialized for moving quickly in a terrestrial habitat
• Pattern of footfalls depends on species and speed of travel
• Humans are the only truly bipedal species
TYPES OF TERRESTIAL LOCOMOTION

1. Plantigrade: flat-footed, all bones of manus and/or pes on the ground; amphibians, most reptiles & some mammals (insectivores, monkeys, apes, humans, & bears)
2. Digitigrade: 1st digit is reduced or lost, manus & pes are elevated (rabbits, rodents, & many carnivores)

3. Unguligrade: reduced number of digits, walk on tips of remaining digits, claws become hooves

As the fastest North American mammal,
the pronghorn antelope
(unguligrades) can reach speeds of 60
miles per hour. At high speed they cover the ground in strides of 14 to 24
feet, and are known to run for long distances at speeds of 30 to 40
miles per hour.

4. Saltatorial (jumping, springing, bounding)
a. quadrupedal b. bipedal (ricochetal)
enlarged feet on propulsive limbs
center of gravity shifted toward rear of body
big hindquarters long tail (acts as
counterbalance)

5. Swimming Amphibious (semiaquatic):
webbed feet, flattened tail (used for propulsion); increase in number, & size of phalanges
Aquatic: limbs modified into flippers for propulsion; basically no externally visible tail
Marine (fully aquatic): fusion of cervical vertebrae; forelimbs modified into flippers (provides stability); elimination of hind limbs and most of pelvic girdle (not visible externally, but remnant bones may remain); tail flattened dorso-ventrally into a fluke (provides propulsion)

6. Gliding & Flying (volant)
Gliding has evolved independently at least 5 times
in marsupials, rodents and dermopterans
Birds - loss of digits & bones plus fusion of some bones
Bats - 5 digits; elongated metacarpals (II through V) & phalanges support the patagium
Pterosaurs - 4th digit is elongated to support patagium


7. Climbing (arboreal, scansorial)
Prehensility: larger body size relative to substrate • Occurs in hands,
feet, tail with friction pads
• Loss of claws (nails instead)
Suspensory (hanging) • Long, curved
digits • Fusion of cervical
vertebrae

• Usually dig with forelimbs, but sometimes with incisor teeth (e.g., mole rats), large claws
• Short, broad forelimbs and splayed to the sides (if forelimbs are used for digging)
• More vibrissae than terrestrial mammals, in more locations on the body
• Small eyes; often lack pinnae
6. Digging and burrowing (fossorial)

Brachiation- arm swings using gripping hands
Grasping- opposable thumb; thumb at wider angle from index finger; strong thumb muscles Saddle joint- at base of thumb where it meets palm

ADJUSTMENTS FOR
BIPEDALISM
The primate foot is strongly arched to absorb forces.
The large toe is no longer opposable, but flexes downward and is a major contributor to the propulsive force of locomotion.

The angle of articulation at the hip & knee joint is shifted medially in humans so that the feet are located directly below the body's center of mass.
Without this shift, humans would fall medially at each step, causing the side-to-side wobble motion observed when non-human primates walk bipedally.
The plane of the human pelvis is perpendicular to the vertebral column & forms a broad bowl with wide sides and a relatively narrow opening at its base.
A narrow opening of the pelvis & a large brain size results in difficult birthing. Babies are born with an incompletely developed brain because further brain development in utero would make passage through the birth canal impossible.

Posterior expansion of the ilium portion of the os coxae allows the gluteus maximus muscle to draw the femur posteriorly in the final part of the stride. This contributes to a smooth, efficient gait.
In non-human primates, the spine articulates with the posterior region of the skull. The human skull articulates with the spine on the ventral side, allowing the head to perch effortlessly atop the vertebral column.
The vertebral column in mammals, including non-human primates, is arched broadly to provide a beam to support the viscera against gravity.
When upright, the arched spine places the center of mass anterior to the pelvis causing non-human primates to fall forward onto their forelimbs.

The human spine has a convex curvature in the lumbar region that shifts the center of mass
posteriorly and places it directly over the pelvic girdle.

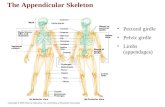
Human Skeletal System

HUMAN APPENDICULAR SKELETON

PECTORAL
GIRDLE

HUMAN PELVIC GIRDLE

FORELIMB

HINDLIMB
A sesamoid bone, the patella or kneecap
develops in birds &
mammals which protects the knee joint
from the abrasive
action of the tendon.

• Phalangeal formula of 2,3,3,3,3 on human hands and feet
• Very prominent olecranon process

QUESTIONS?

1. Provide evidences for the various hypotheses that attempt to explain the origin of paired fins.
2. Give the functions of vertebrate fins, and their supporting elements in various fish groups.
3. Describe the structural patterns that distinguish among shark fins, ray fins, and lobe fins.
4. Tabulate: dermal bones and replacement bones in the pectoral girdle starting from a basal vertebrate, fish, amphibian, reptile, mammal.
5. Compare and contrast: bones in the pelvic girdle of a basal vertebrate, fish, amphibian, reptile, mammal.
6. Give the functional significance of the following: different shapes of caudal fins; detachment of tetrapod pectoral girdle from skull; reduced clavicle in mammals; presence of scapular fossae; opposable thumb.
7. Describe how a tetrapod limb evolved from fins, and the modifications required for locomotion in a terrestrial habitat.
8. Give the different types of gaits, and limb modifications associated with each type.
9. Describe the mechanistic adjustments required for the transition of a tetrapod limb into bipedalism.
10. Describe the structural pattern of a vertebrate limb, and account for whatever modifications became evident in phylogenetic history.

QUIZ NEXT MEETING
Top Related