







Languages
Pages
Legal


Amino Acid Catabolism: Carbon Skeletons
Copyright © 1999-2007 by Joyce J. Diwan. All rights reserved.
Molecular Biochemistry II

Amino Acid Carbon Skeletons
Amino acids, when deaminated, yield -keto acids that, directly or via additional reactions, feed into major metabolic pathways (e.g., Krebs Cycle).
Amino acids are grouped into 2 classes, based on whether or not their carbon skeletons can be converted to glucose: glucogenic ketogenic.

Carbon skeletons of glucogenic amino acids are degraded to:
pyruvate, or
a 4-C or 5-C intermediate of Krebs Cycle. These are precursors for gluconeogenesis.
Glucogenic amino acids are the major carbon source for gluconeogenesis when glucose levels are low.
They can also be catabolized for energy, or converted to glycogen or fatty acids for energy storage.

Carbon skeletons of ketogenic amino acids are degraded to: acetyl-CoA, or acetoacetate.
Acetyl CoA, & its precursor acetoacetate, cannot yield net production of oxaloacetate, the gluconeogenesis precursor.
For every 2-C acetyl residue entering Krebs Cycle, 2 C leave as CO2.
Carbon skeletons of ketogenic amino acids can be catabolized for energy in Krebs Cycle, or converted to ketone bodies or fatty acids.
They cannot be converted to glucose.
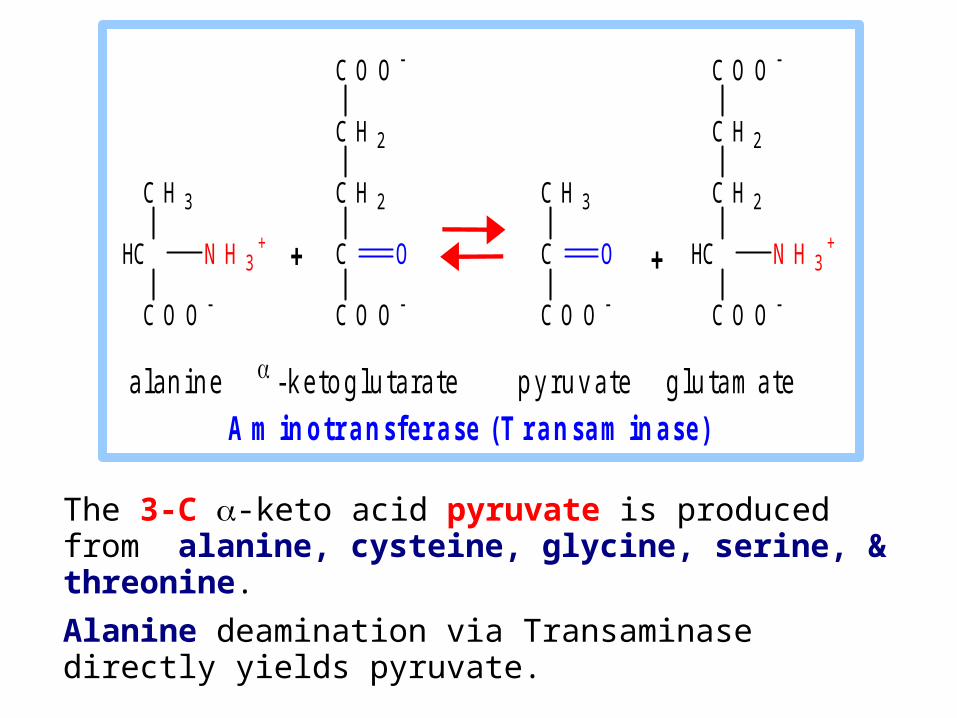
The 3-C -keto acid pyruvate is produced from alanine, cysteine, glycine, serine, & threonine.
Alanine deamination via Transaminase directly yields pyruvate.
a l a n i n e - k e t o g l u t a r a t e p y r u v a t e g l u t a m a t e
A m i n o t r a n s f e r a s e ( T r a n s a m i n a s e )
C O O
C H 2
C H 2
C
C O O
O
C H 3
HC
C O O
N H 3+
C O O
C H 2
C H 2
HC
C O O
N H 3+
C H 3
C
C O O
O + +
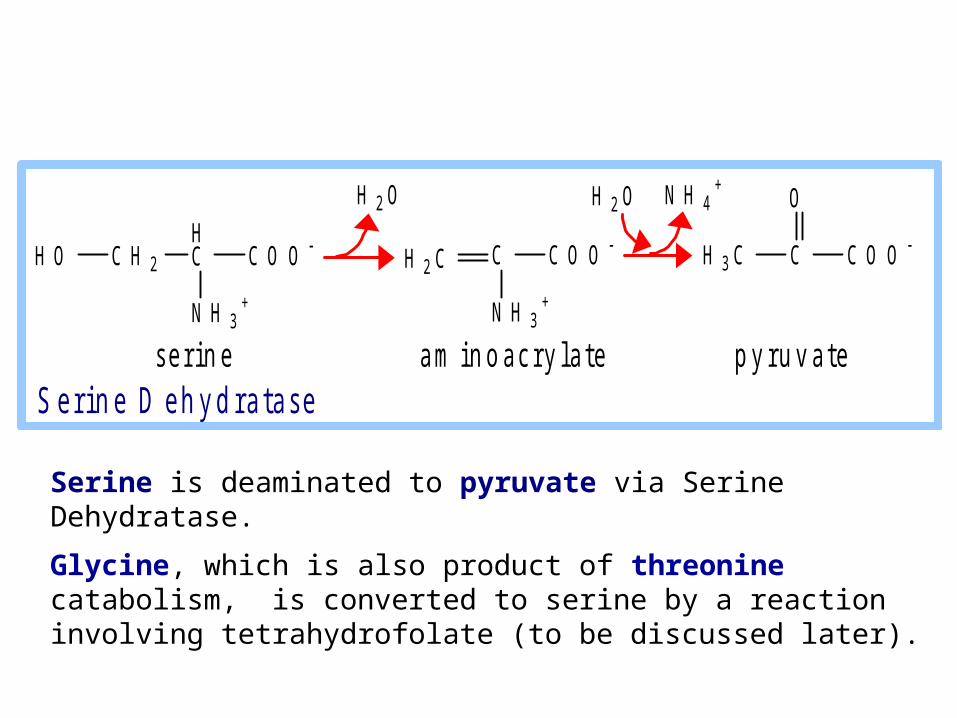
Serine is deaminated to pyruvate via Serine Dehydratase.
Glycine, which is also product of threonine catabolism, is converted to serine by a reaction involving tetrahydrofolate (to be discussed later).
H O C H 2
HC C O O
N H 3+
C C O O
OH 2 O N H 4+
C C O O
N H 3+
H 2 C H 3 C
H 2 O
s e r i n e a m i n o a c r y l a t e p y r u v a t e S e r i n e D e h y d r a t a s e

The 4-C Krebs Cycle intermediate oxaloacetate is produced from aspartate & asparagine.
Aspartate transamination yields oxaloacetate.
Aspartate is also converted to fumarate in Urea Cycle. Fumarate is converted to oxaloacetate in Krebs cycle.
a s p a r t a t e - k e t o g l u t a r a t e o x a l o a c e t a t e g l u t a m a t e
A m i n o t r a n s f e r a s e ( T r a n s a m i n a s e )
C O O
C H 2
C H 2
C
C O O
O
C O O
C H 2
HC
C O O
N H 3+
C O O
C H 2
C H 2
HC
C O O
N H 3+
C O O
C H 2
C
C O O
O + +

Asparagine loses the amino group from its R-group by hydrolysis catalyzed by Asparaginase.
This yields aspartate, which can be converted to oxaloacetate, e.g., by transamination.
C
C H 2
H C
C O O
N H 3+
OH 2 N
C O O
C H 2
H C
C O O
N H 3+
H 2 O N H 4+
a s p a r a g in e a s p a r t a t e A s p a r a g in a s e

The 4-C Krebs Cycle intermediate succinyl-CoA is produced from isoleucine, valine, & methionine.
Propionyl-CoA, an intermediate on these pathways, is also a product of -oxidation of fatty acids with an odd number of C atoms.

The branched chain amino acids initially share in part a common pathway.
Branched Chain -Keto Acid Dehydrogenase (BCKDH) is a multi-subunit complex homologous to Pyruvate Dehydrogenase complex.
Genetic deficiency of BCKDH is called Maple Syrup Urine Disease (MSUD).
High concentrations of branched chain keto acids in urine give it a characteristic odor.

Propionyl-CoA is carboxylated to methylmalonyl-CoA.
A racemase yields the L-isomer essential to the subsequent reaction.
Methylmalonyl-CoA Mutase catalyzes a molecular rearrangement: the branched C chain of methylmalonyl-CoA is converted to the linear C chain of succinyl-CoA.
The carboxyl that is in ester linkage to the thiol of coenzyme A is shifted to an adjacent carbon atom, with opposite shift of a hydrogen atom.
C C H 3
C S-C o A
O
C C H 3
C S-C oA
O
C O O
C
C S-C oA
O
C O O
C C
C O O
C
C
O
H
H
CoA-S H
HH HH
H
H
H
H
H CO 3
A T P A D P
+ P i
p r o p i o n y l - C o A D - m e t h y l m a l o n y l - C o A L - m e t h y l m a l o n y l - C o A s u c c i n y l - C o A
P r o p i o n y l - C o A M e t h y l m a l o n y l - C o A M e t h y l m a l o n y l - C o A C a r b o x y l a s e ( B i o t i n ) R a c e m a s e M u t a s e ( B 1 2 )
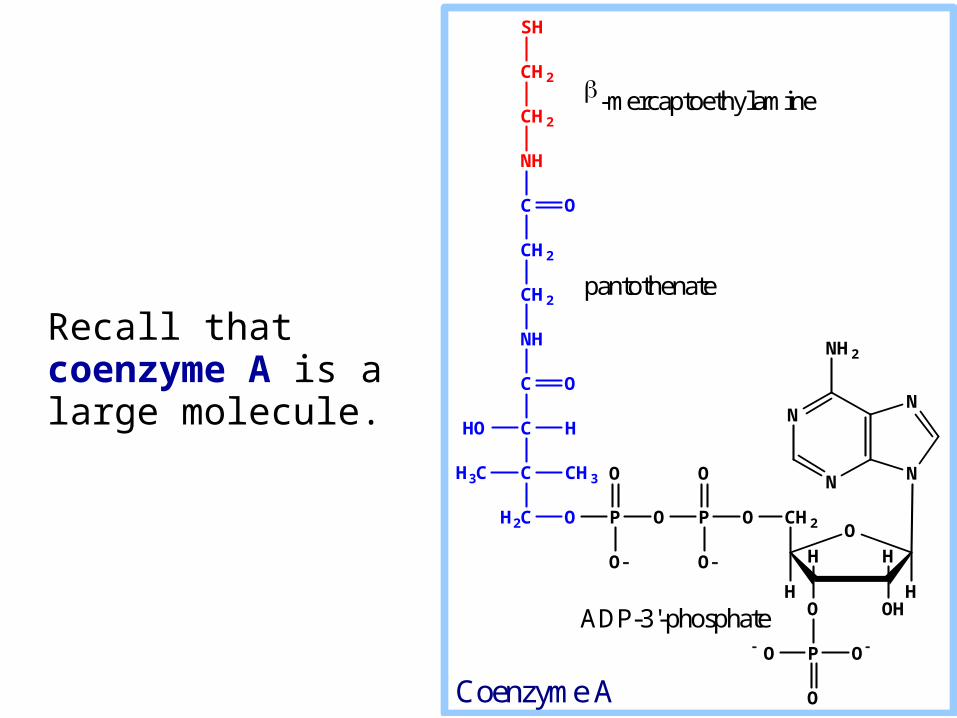
Recall that coenzyme A is a large molecule.
N
N N
N
NH2
O
OHO
HH
H
CH2
H
OPOPOH2C
O
O O
O
P
O
O O
C
C
C
NH
CH2
CH2
C
NH
CH3H3C
HHO
O
CH2
CH2
SH
O
-mercaptoethylamine
pantothenate
ADP-3'-phosphate
Coenzyme A

Coenzyme B12, a derivative of vitamin B12 (cobalamin), is the prosthetic group of Methylmalonyl-CoA Mutase.
C
C S-CoA
O
COO
C C
COO
C
C
O
H
H
CoA-S H
HHH
H
H
L-methylmalonyl-CoA succinyl-CoA
Methylmalonyl-CoA Mutase (B12)

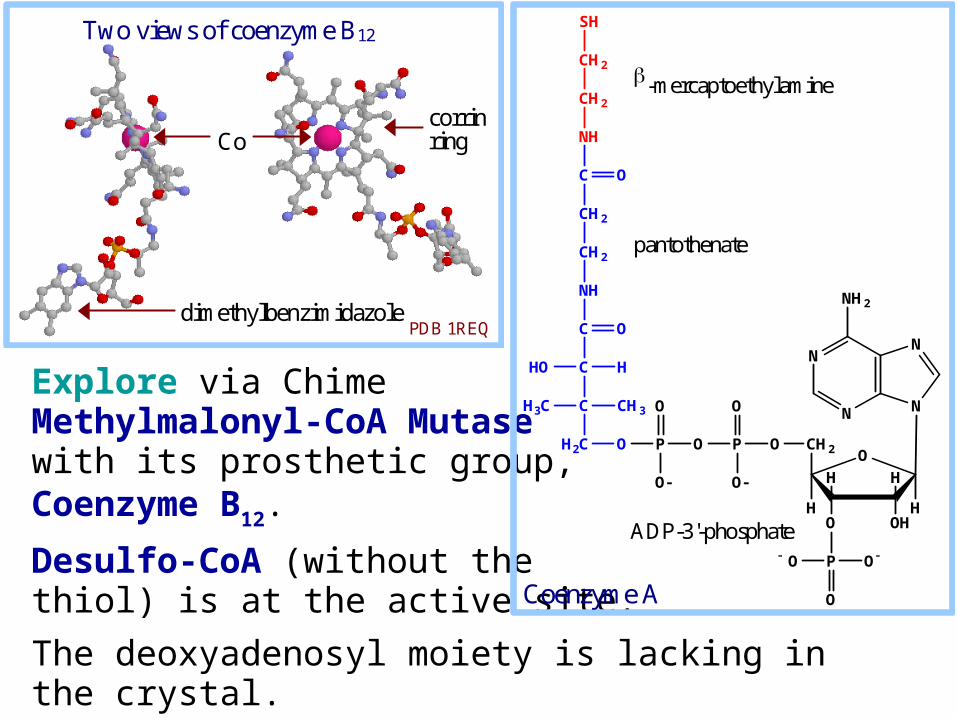
A crystal structure of the enzyme-bound coenzyme B12.
Coenzyme B12 contains a heme-like corrin ring with a cobalt ion coordinated to 4 ring N atoms.
Co
dimethylbenzimidazole
Two views of coenzyme B12
corrin ring
PDB 1REQ
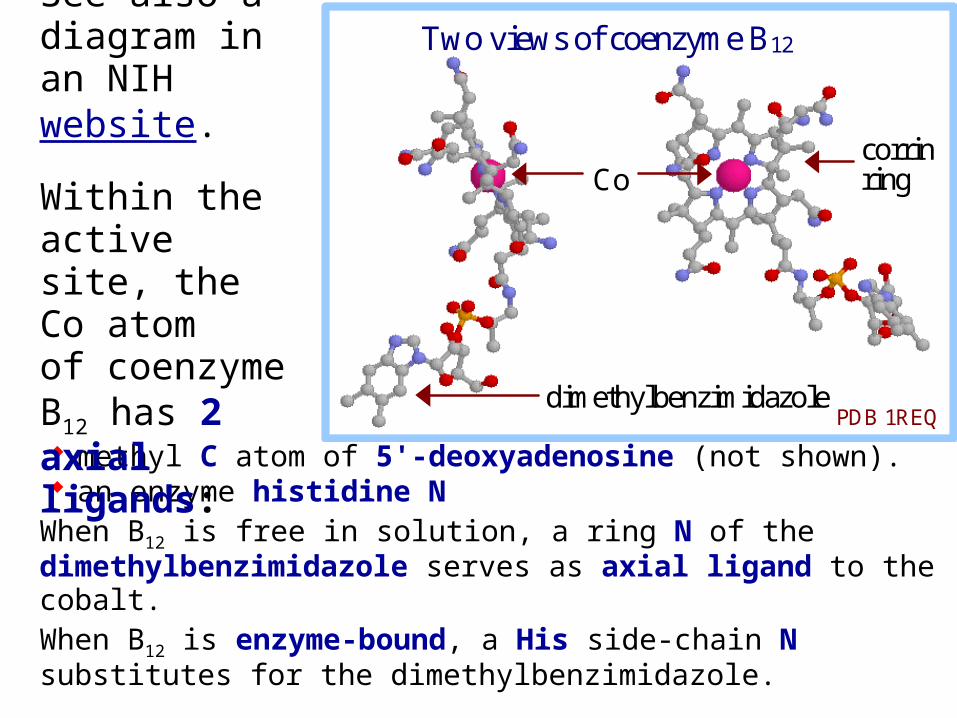
methyl C atom of 5'-deoxyadenosine (not shown). an enzyme histidine N
When B12 is free in solution, a ring N of the dimethylbenzimidazole serves as axial ligand to the cobalt.When B12 is enzyme-bound, a His side-chain N substitutes for the dimethylbenzimidazole.
Co
dimethylbenzimidazole
Two views of coenzyme B12
corrin ring
PDB 1REQ
See also a diagram in an NIH website.
Within the active site, the Co atom of coenzyme B12 has 2 axial ligands:

Homolytic cleavage of the deoxyadenosyl C-Co bond during catalysis yields a deoxyadenosyl carbon radical, as Co3+ becomes Co2+.
Reaction of this with methylmalonyl-CoA generates a radical substrate intermediate and 5'-deoxyadenosine.
Following rearrangement of the substrate, the product radical abstracts a H atom from the methyl group of 5'-deoxyadenosine.
This yields succinyl-CoA and the 5'-deoxyadenosyl radical, which reacts with coenzyme B12 to reestablish the deoxyadenosyl C-Co bond.

Methyl group transfers are also carried out by B12 (cobalamin).
Methyl-B12 (methylcobalamin), with a methyl axial ligand substituting for the deoxyadenosyl moiety of coenzyme B12, is an intermediate of such transfers.
E.g., B12 is a prosthetic group of the mammalian enzyme that catalyzes methylation of homocysteine to form methionine (to be discussed later).

Vitamin B12 is synthesized only by bacteria.
Ruminants get B12 from bacteria in their digestive system.
Humans obtain B12 from meat or dairy products.
Vitamin B12 bound to the protein gastric intrinsic factor is absorbed by cells in the upper part of the human small intestine via receptor-mediated endocytosis.
B12 synthesized by bacteria in the large intestine is unavailable.
Strict vegetarians eventually become deficient in B12 unless they consume it in pill form.
Vitamin B12 is transported in the blood bound to the protein transcobalamin, which is recognized by a receptor that mediates uptake into body cells.
Explore via ChimeMethylmalonyl-CoA Mutase with its prosthetic group, Coenzyme B12.
Desulfo-CoA (without the thiol) is at the active site.
The deoxyadenosyl moiety is lacking in the crystal.
N
N N
N
NH2
O
OHO
HH
H
CH2
H
OPOPOH2C
O
O O
O
P
O
O O
C
C
C
NH
CH2
CH2
C
NH
CH3H3C
HHO
O
CH2
CH2
SH
O
-mercaptoethylamine
pantothenate
ADP-3'-phosphate
Coenzyme A
Co
dimethylbenzimidazole
Two views of coenzyme B12
corrin ring
PDB 1REQ
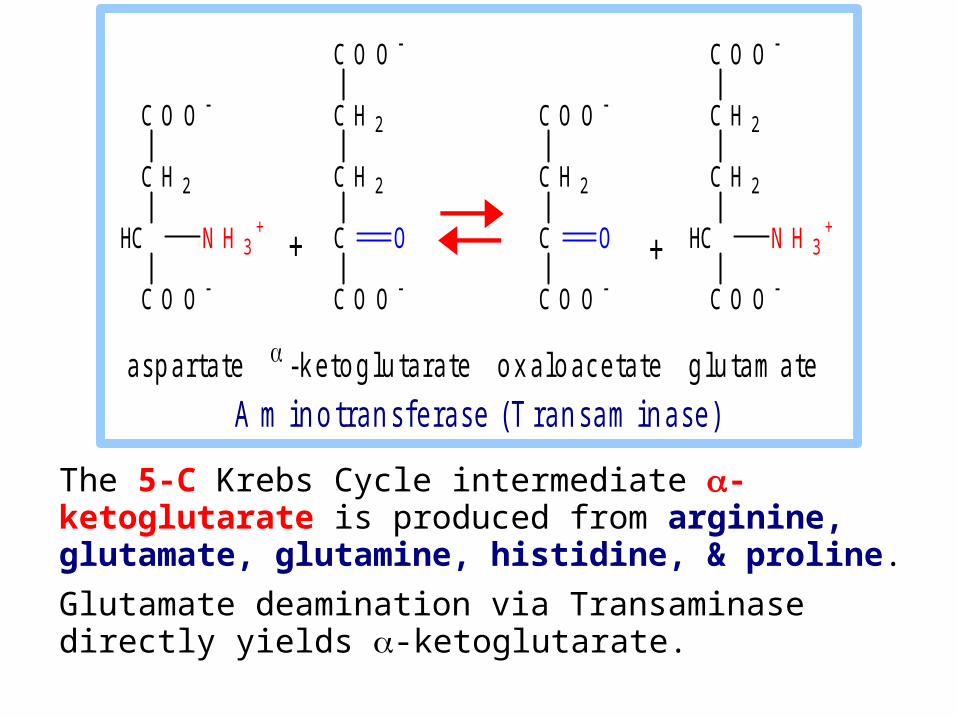
The 5-C Krebs Cycle intermediate -ketoglutarate is produced from arginine, glutamate, glutamine, histidine, & proline.
Glutamate deamination via Transaminase directly yields -ketoglutarate.
a s p a r t a t e - k e t o g l u t a r a t e o x a l o a c e t a t e g l u t a m a t e
A m i n o t r a n s f e r a s e ( T r a n s a m i n a s e )
C O O
C H 2
C H 2
C
C O O
O
C O O
C H 2
HC
C O O
N H 3+
C O O
C H 2
C H 2
HC
C O O
N H 3+
C O O
C H 2
C
C O O
O + +

Glutamate deamination by Glutamate Dehydrogenase also directly yields -ketoglutarate.
O O CH 2C
H 2C C C O O
O
+ N H 4+
N A D (P )+
N AD(P)H
O O CH 2C
H 2C C C O O
N H 3+
Hglu tam ate
-ke toglu tara te
G lu tam ate D ehydrogenase
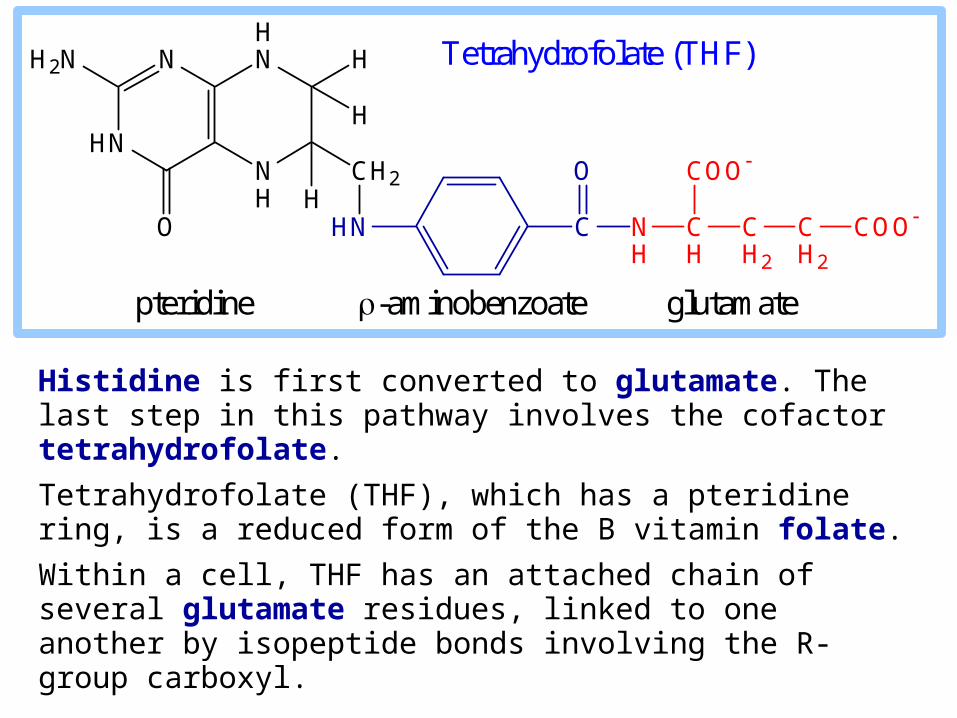
Histidine is first converted to glutamate. The last step in this pathway involves the cofactor tetrahydrofolate.
Tetrahydrofolate (THF), which has a pteridine ring, is a reduced form of the B vitamin folate.
Within a cell, THF has an attached chain of several glutamate residues, linked to one another by isopeptide bonds involving the R-group carboxyl.
NH
HNN
HN
H2N H
H
H
CH2
HNO C
O
NH
CH
COO
CH2
CH2
COO
Tetrahydrofolate (THF)
pteridine -aminobenzoate glutamate
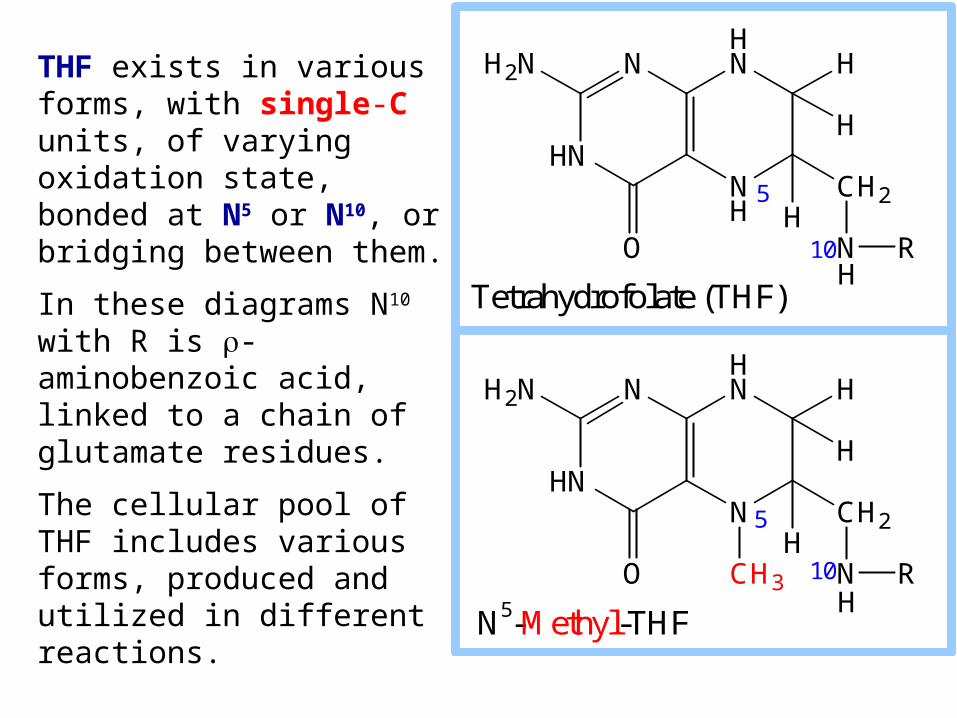
THF exists in various forms, with single-C units, of varying oxidation state, bonded at N5 or N10, or bridging between them.
In these diagrams N10 with R is -aminobenzoic acid, linked to a chain of glutamate residues.
The cellular pool of THF includes various forms, produced and utilized in different reactions.
NH
HNN
HN
H2N H
H
H
CH2
N RO
N
HNN
HN
H2N H
H
H
CH2
N RO CH3
H
H
Tetrahydrofolate (THF)
N5-Methyl-THF
5
10
5
10
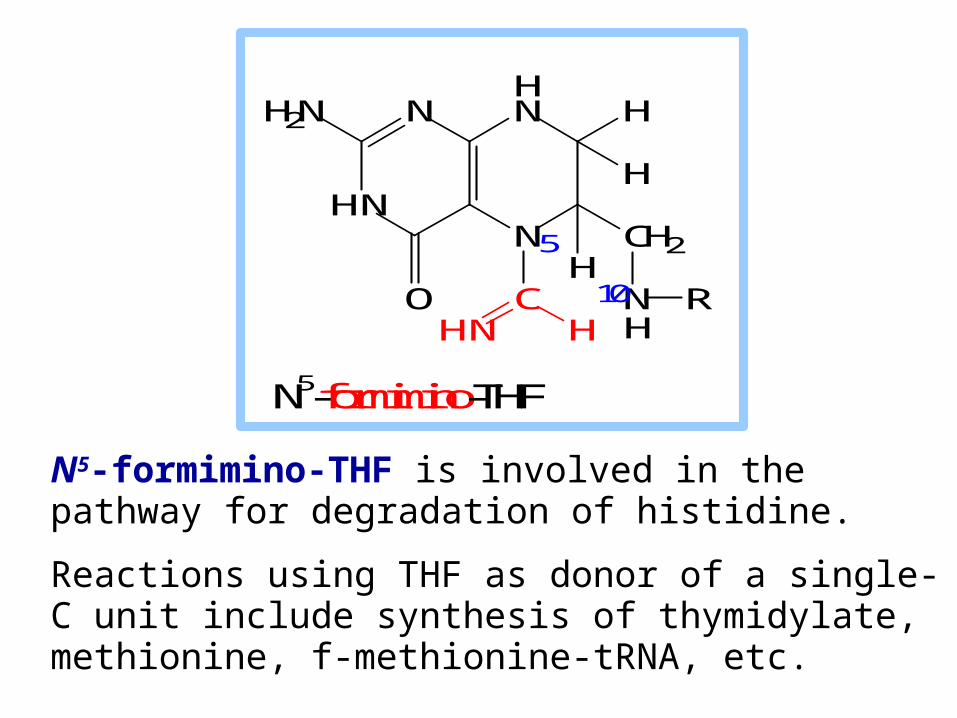
N5-formimino-THF is involved in the pathway for degradation of histidine.
Reactions using THF as donor of a single-C unit include synthesis of thymidylate, methionine, f-methionine-tRNA, etc.
NH
HNN
HN
H2N H
H
H
CH2
N RO
N
HNN
HN
H2N H
H
H
CH2
N RO C
H
HHHN
N5-formimino-THF
5
10
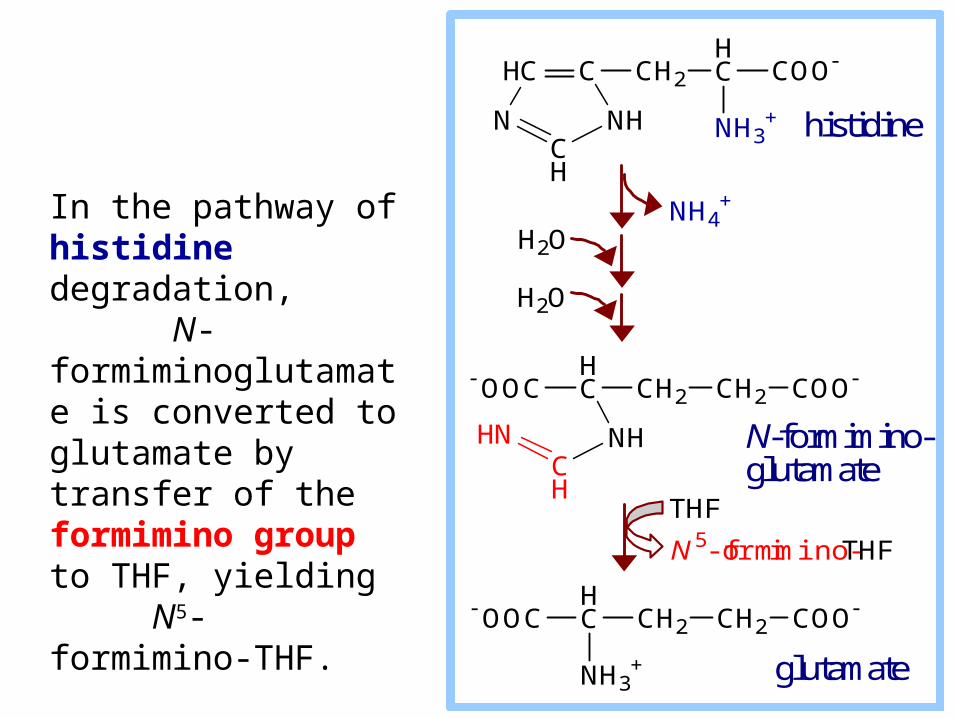
In the pathway of histidine degradation, N-formiminoglutamate is converted to glutamate by transfer of the formimino group to THF, yielding N5-formimino-THF.
HC C CH2
HC COO
NH3+N NH
CH
OOCHC CH2 CH2 COO
HN NHCH
OOCHC CH2 CH2 COO
NH3+
THF
N 5-formimino-THF
NH4+
H2O
H2O
histidine
N-formimino-glutamate
glutamate

Because of the essential roles of THF as acceptor and donor of single carbon units, dietary deficiency of folate, genetic deficiencies in folate metabolism or transport, and the increased catabolism of folate seen in some disease states, result in various metabolic effects leading to increased risk of developmental defects, cardiovascular disease, and cancer.
NH
HNN
HN
H2N H
H
H
CH2
N ROHTetrahydrofolate (THF)
5
10

Aromatic Amino Acids
Aromatic amino acids phenylalanine & tyrosine are catabolized to fumarate and acetoacetate.
Hydroxylation of phenylalanine to form tyrosine involves the reductant tetrahydrobiopterin. Biopterin, like folate, has a pteridine ring.
Dihydrobiopterin is reduced to tetrahydrobiopterin by electron transfer from NADH.
Thus NADH is secondarily the e donor for conversion of phenylalanine to tyrosine.
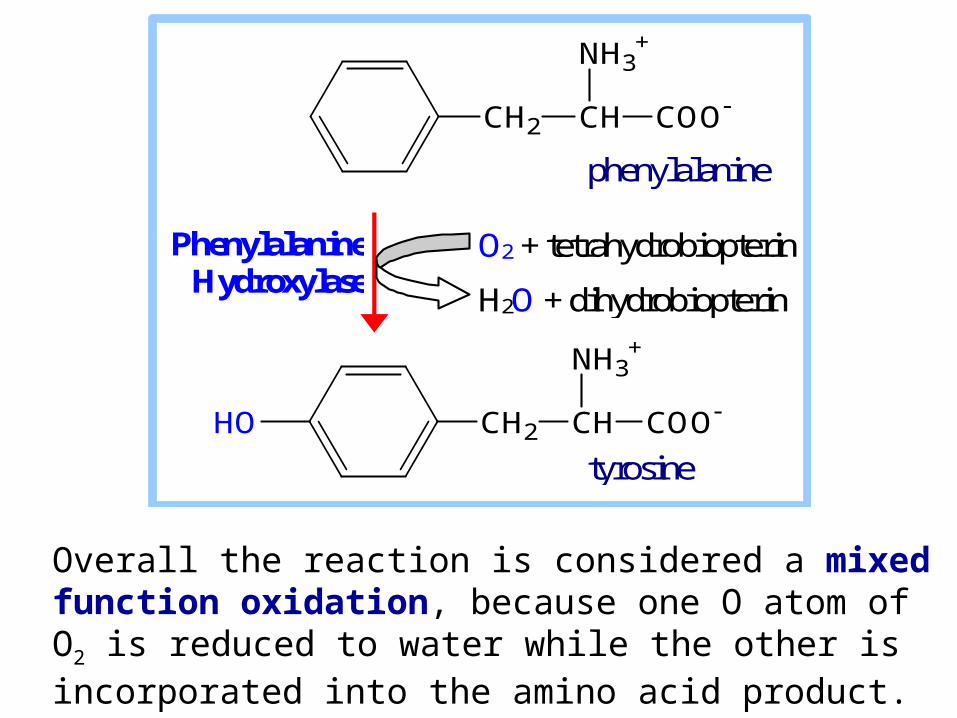
Overall the reaction is considered a mixed function oxidation, because one O atom of O2 is reduced to water while the other is incorporated into the amino acid product.
CH2 CH COO
NH3+
CH2 CH COO
NH3+
HO
phenylalanine
tyrosine
O2 + tetrahydrobiopterin
H2O + dihydrobiopterin
Phenylalanine Hydroxylase
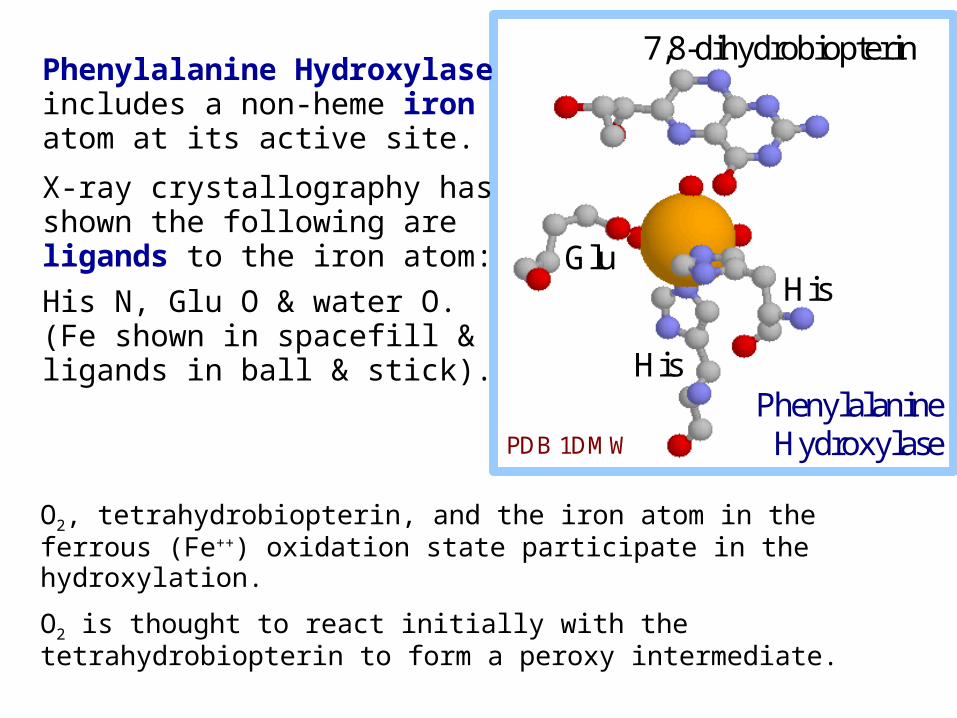
O2, tetrahydrobiopterin, and the iron atom in the ferrous (Fe++) oxidation state participate in the hydroxylation.
O2 is thought to react initially with the tetrahydrobiopterin to form a peroxy intermediate.
His
His Glu
7,8-dihydrobiopterin
Phenylalanine Hydroxylase PDB 1DMW
Phenylalanine Hydroxylase includes a non-heme iron atom at its active site.
X-ray crystallography has shown the following are ligands to the iron atom:
His N, Glu O & water O. (Fe shown in spacefill & ligands in ball & stick).
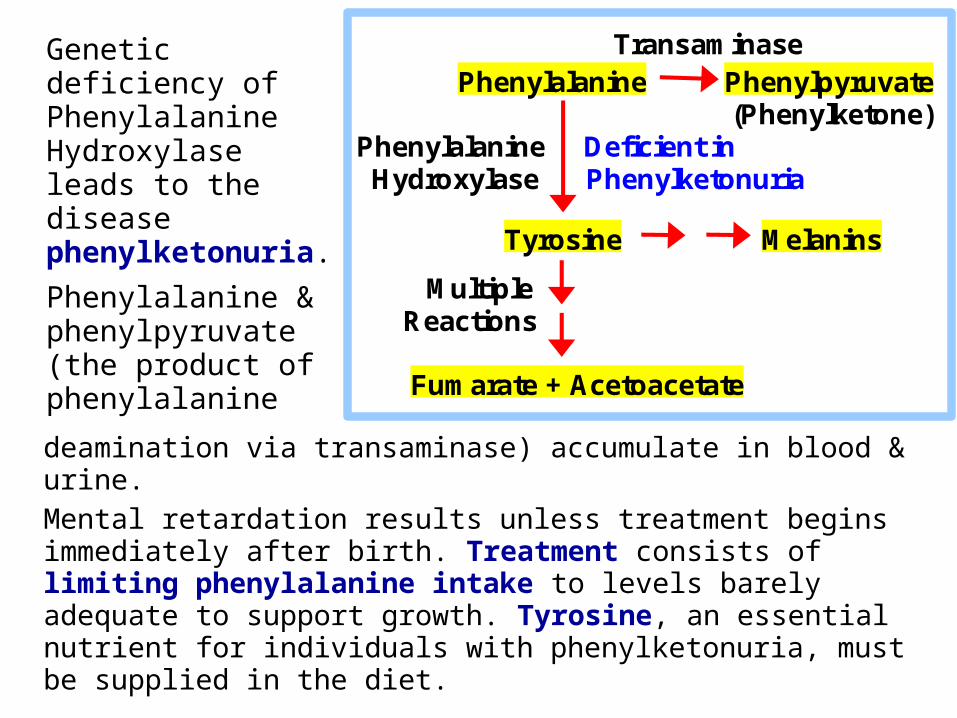
deamination via transaminase) accumulate in blood & urine.
Mental retardation results unless treatment begins immediately after birth. Treatment consists of limiting phenylalanine intake to levels barely adequate to support growth. Tyrosine, an essential nutrient for individuals with phenylketonuria, must be supplied in the diet.
Transaminase Phenylalanine Phenylpyruvate (Phenylketone) Phenylalanine Deficient in Hydroxylase Phenylketonuria
Tyrosine Melanins
Multiple Reactions
Fumarate + Acetoacetate
Genetic deficiency of Phenylalanine Hydroxylase leads to the disease phenylketonuria.
Phenylalanine & phenylpyruvate (the product of phenylalanine
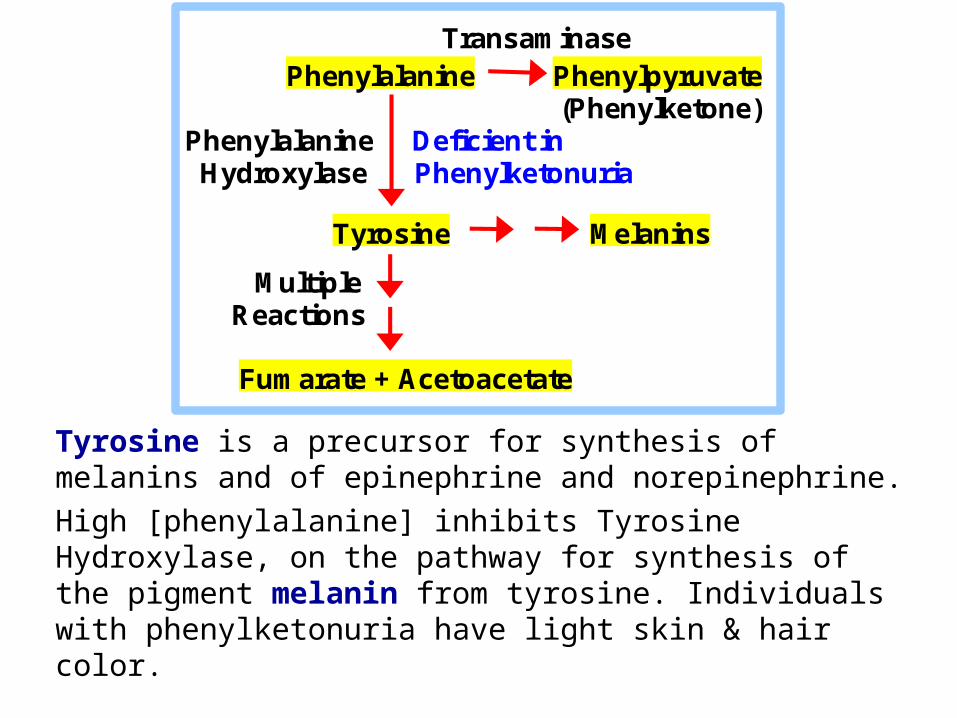
Tyrosine is a precursor for synthesis of melanins and of epinephrine and norepinephrine.
High [phenylalanine] inhibits Tyrosine Hydroxylase, on the pathway for synthesis of the pigment melanin from tyrosine. Individuals with phenylketonuria have light skin & hair color.
Transaminase Phenylalanine Phenylpyruvate (Phenylketone) Phenylalanine Deficient in Hydroxylase Phenylketonuria
Tyrosine Melanins
Multiple Reactions
Fumarate + Acetoacetate
H3C SH2C
H2C
HC COO
NH3+CH2
+
O
OHOH
HH
HH
AdenineH3C S
H2C
H2C
HC COO
NH3+
HSH2C
H2C
HC COO
NH3+
SH2C
H2C
HC COO
NH3+CH2
O
OHOH
HH
HH
Adenine
methionine
homocysteine
S-adenosyl-methionine(SAM)
S-adenosyl-homocysteine
ATP PPi + Pi
adenosine H2O
acceptor
methylated acceptorTHF
N5-methyl-THF
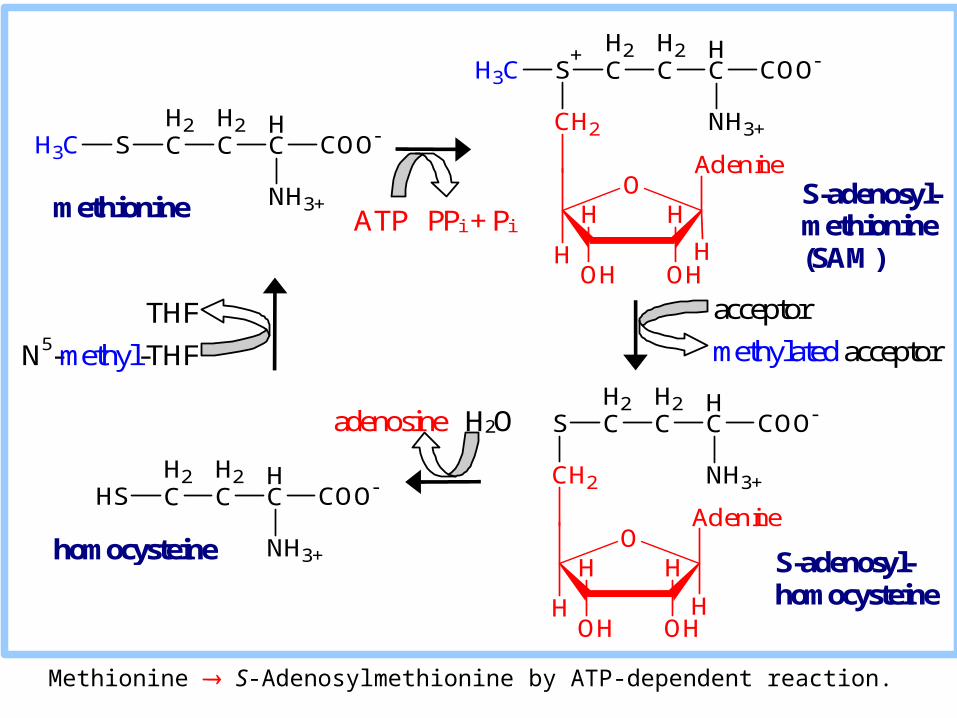
Methionine S-Adenosylmethionine by ATP-dependent reaction.

H3C S CH2
CH2
HC COO
NH3+CH2
+
O
OHOH
HH
HH
Adenine
HS CH2
CH2
HC COO
NH3+
S CH2
CH2
HC COO
NH3+CH2
O
OHOH
HH
HH
Adenine
homocysteine
S-adenosyl-methionine (SAM)
S-adenosyl-homocysteine
adenosine H2O
acceptor
methylated acceptor
SAM is a methyl group donor in synthetic reactions.
The resulting S-adenosylhomocysteine is hydrolyzed to homocysteine.
Homocysteine may be catabolized via a complex pathway to cysteine & succinyl-CoA.
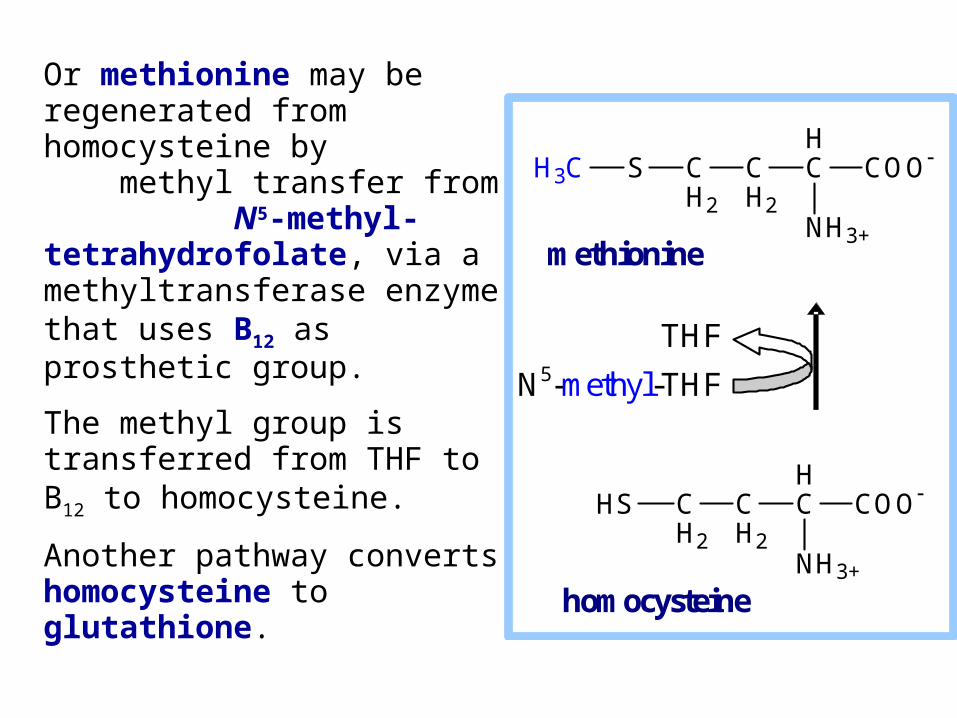
Or methionine may be regenerated from homocysteine by methyl transfer from N5-methyl-tetrahydrofolate, via a methyltransferase enzyme that uses B12 as prosthetic group.
The methyl group is transferred from THF to B12 to homocysteine.
Another pathway converts homocysteine to glutathione.
H3C S CH2
CH2
HC COO
NH3+
HS CH2
CH2
HC COO
NH3+
methionine
homocysteine
THF
N5-methyl-THF

In various reactions, S-adenosylmethionine (SAM) is a donor of diverse chemical groups including methylene, amino, ribosyl and aminoalkyl groups, and a source of 5'-deoxyadenosyl radicals.
But SAM is best known as a methyl group donor.
O
OHOH
HH
HH
Adenine
H3C S CH2
CH2
HC COO
NH3+CH2
+
S-adenosylmethionine (SAM)

Examples:S-adenosylmethionine as methyl group donor methylation of bases in tRNA methylation of cytosine residues in DNA methylation of norepinephrine epinephrine
CH2 NH3+CHHO
HOOH
CH2
HNCHHO
HOOH
CH3
S-adenosylmethionine
S-adenosylhomocysteine
norepinephrine
epinephrine
O
OHOH
HH
HH
Adenine
H3C S CH2
CH2
HC COO
NH3+CH2
+
S-adenosylmethionine (SAM)
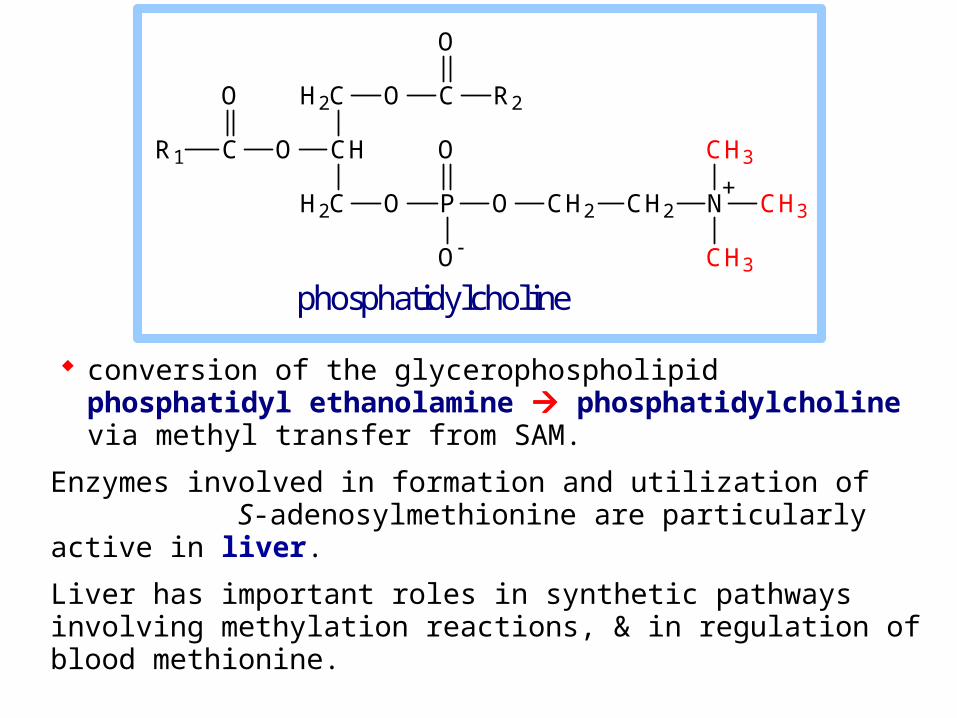
conversion of the glycerophospholipid phosphatidyl ethanolamine phosphatidylcholine via methyl transfer from SAM.
Enzymes involved in formation and utilization of S-adenosylmethionine are particularly active in liver.
Liver has important roles in synthetic pathways involving methylation reactions, & in regulation of blood methionine.
O P O
O
O
H2C
CH
H2C
OCR1
O O C
O
R2
CH2 CH2 N CH3
CH3
CH3
+
phosphatidylcholine

Methyl Group Donors
Methyl group donors in synthetic reactions include:
methyl-B12
S-adenosylmethionine (SAM)
N5-methyl-tetrahydrofolate (N5-methyl-THF)
Lysine & Tryptophan
The complex pathways for degradation of lysine and tryptophan will not be covered.

Check out the OMIM website for an example of an inborn error of metabolism, phenylketonuria, a disease resulting from deficiency of Phenylalanine Hydroxylase.