







Languages
Pages
Legal


UNIVERSIDADE TECNOLÓGICA FEDERAL DO PARANÁ
PROGRAMA DE PÓS-GRADUAÇÃO EM TECNOLOGIA
DE PROCESSOS QUÍMICOS E BIOQUÍMICOS
GABRIELLE CRISTINA CALEGARI
(1→6)-β-D-GLUCANA (LASIODIPLODANA) SULFATADA: OBTENÇÃO, CARACTERIZAÇÃO E PROPRIEDADES BIOATIVAS
Dissertação de Mestrado
Pato Branco, 2019

2
GABRIELLE CRISTINA CALEGARI
(1→6)-β-D-GLUCANA (LASIODIPLODANA) SULFATADA:
OBTENÇÃO, CARACTERIZAÇÃO E PROPRIEDADES BIOATIVAS
Dissertação de Mestrado apresentada ao Programa de Pós-graduação em Tecnologia de Processos Químicos e Bioquímicos da Universidade Tecnológica Federal do Paraná como requisito parcial para obtenção do título de “Mestre em Tecnologia de Processos Químicos e Bioquímicos” Orientador: Dr. Mário Antônio Alves da Cunha. Coorientadora: Drª. Vidiany Aparecida Queiroz Santos.
Pato Branco, 2019

3

4
TERMO DE APROVAÇÃO Nº 82
TÍTULO DA DISSERTAÇÃO
“(1→6)-β-D-GLUCANA (LASIODIPLODANA) SULFATADA: OBTENÇÃO,
CARACTERIZAÇÃO E PROPRIEDADES BIOATIVAS”
Autora
Gabrielle Cristina Calegari
Esta dissertação foi apresentada às 13 horas e 30 minutos do dia 22 de fevereiro de 2019,
como requisito parcial para a obtenção do título de MESTRE EM TECNOLOGIA DE
PROCESSOS QUÍMICOS E BIOQUÍMICOS – Linha de pesquisa em biotecnologia – no
Programa de Pós-Graduação em Tecnologia de Processos Químicos e Bioquímicos. A autora
foi arguida pela Banca Examinadora abaixo assinada, a qual, após deliberação, considerou
o trabalho aprovado.
____________________________________ Prof. Dr. Mario Antonio Alves da Cunha
UTFPR/PB Orientador
______________________________________ Profa. Dra. Aneli De Melo Barbosa Dekker
UEL/Londrina Examinadora
__________________________________ Profa. Dra. Sirlei Dias Teixeira
UTFPR/PB Examinadora
Visto da Coordenação
Prof. Dr. Edimir Andrade Pereira
Coordenador do Programa de Pós-Graduação em Tecnologia de Processos Químicos e Bioquímicos - PPGTP
O Termo de Aprovação assinado encontra-se na Coordenação do PPGTP
MINISTÉRIO DA EDUCAÇÃO
Universidade Tecnológica Federal do Paraná Câmpus Pato Branco
Programa de Pós-Graduação em Tecnologia de Processos Químicos e Bioquímicos

5
“Os outros... Os que amaram sem pedir compensações, os que não lhe exigiram nada e lhe deram tudo.”
Olhai os lírios do campo Érico Veríssimo

6
RESUMO CALEGARI, Gabrielle Cristina. (1→6)-β-D-glucana (lasiodiplodana) sulfatada: obtenção, caracterização e propriedades bioativas. 2019. 111 f. Dissertação – Programa de Pós-Graduação em Tecnologia de Processos Químicos e Bioquímicos, Universidade Tecnológica Federal do Paraná. Pato Branco, 2019. A lasiodiplodana é uma (1→6)-β-D-glucana fúngica que apresenta diferentes
funcionalidades biológicas, incluindo atividades antioxidante, hipocolesterolêmica,
hipoglicemiante e anticarcinogênica. Suas propriedades biológicas podem ser
potencializadas por meio de modificações químicas na estrutura da macromolécula,
como por exemplo, a sulfatação. Neste contexto, no presente trabalho foram obtidos
derivados sulfatados com diferentes graus de substituição (DS), os quais foram
caracterizados por Espectroscopia de Infravermelho com Transformada de Fourier
(IV-TF), Microscopia Eletrônica de Varredura (MEV), Análise Térmica (DTG, TGA e
DTA), Difração de Raios-X (DRX) e solubilidade. Algumas propriedades biológicas
foram avaliadas, tais como atividade antimicrobiana (MIC), antioxidante (FRAP, OH,
ABTS e DPPH) e potencial citotóxico. Dentre os derivados sulfatados com diferentes
DS (1,61; 1,42; 1,02 e 0,15) observou-se que não houve uma correlação direta entre
DS e concentração do reagente sulfatante empregado no protocolo de derivatização.
A derivatização promoveu aumento na solubilidade da macromolécula em relação a
lasiodiplodana nativa. A sulfatação foi confirmada por IV-TF, através do surgimento
de bandas em 1250 cm-1 (S=O) e 810 cm-1 (C-O-S), oriundas da introdução de
grupamentos sulfônicos. As imagens de MEV demonstraram que a sulfatação
promoveu mudanças morfológicas na superfície do biopolímero, levando ao
surgimento de estruturas fibrilares e heterogêneas ao longo da área superficial. A
lasiodiplodana sulfatada apresentou maior estabilidade térmica, ocorrendo oxidação
do biopolímero em temperaturas próximas a 460 °C. A análise por DRX demonstrou
que as amostras de lasiodiplodana nativa e sulfatadas apresentaram perfil
difratográfico predominantemente amorfo, característico de polissacarídeos. A
sulfatação potencializou a atividade antimicrobiana da lasiodiplodana, especialmente
contra a Candida albicans (atividade fungicida) e Salmonella enterica Typhimurium
(atividade bacteriostática). A amostra nativa apresentou maior capacidade de
remoção do radical hidroxila, enquanto os derivados sulfatados apresentaram maior

7
potencial FRAP. As lasiodiplodanas nativa e sulfatadas não demonstraram
citotoxicidade letal contra cepas selvagem e mutantes de Saccharomyces cerevisiae,
nas concentrações estudadas. As amostras com maiores DS (1,42 e 1,61)
apresentaram menor potencial de indução ao estresse oxidativo. Os resultados do
presente estudo indicam que o processo de sulfatação pode ser uma ferramenta
atrativa para a potencialização de atividades biológicas da lasiodiplodana, podendo
contribuir para a diversificação de suas aplicações biotecnológicas.
Palavras-chave: bioatividade, polissacarídeos fúngicos, sulfatação.

8
ABTRACT CALEGARI, Gabrielle Cristina. (1→6)-β-D-glucan (lasiodiplodan) sulfated: obtaining, characterization and bioactive properties. 2019. 111 f. Dissertation - Post-Graduation Program in Chemical and Biochemical Processes Technology, Universidade Tecnológica Federal do Paraná. Pato Branco, 2019. Lasiodiplodan is a fungal (1→6)-β-D-glucan which exhibits different biological
functionalities, including antioxidant, hypocholesterolemic, hypoglycemic and
anticarcinogenic activities. Their biological properties may be potentiated by chemical
modifications in the macromolecule structure, such as sulfation. In this context, sulfate
derivatives with different degrees of substitution (DS) were obtained and characterized
by Fourier Transform Infrared Spectroscopy (FT-IR), Scanning Electron Microscopy
(SEM), Thermal Analysis (DTG, TGA and DTA), X-ray Diffraction (XRD) and solubility.
Biological properties were evaluated, including antimicrobial (MIC) and antioxidante
(FRAP, OH, ABTS and DPPH) activities and cytotoxic potential. Sulfated derivatives
with different DS (1.61, 1.42, 1.02 and 0.15) were obtained, and there was no direct
correlation between DS and the concentration of the sulfating reagent used in the
derivatization protocol. Derivatization promoted an increase in the solubility of the
macromolecule regarding to native lasiodiplodan. Sulfation was confirmed by FT-IR
through the emergence of bands at 1250 cm-1 (S=O) and 810 cm-1 (C-O-S), derived
from the introduction of sulfonic groups. SEM showed that sulfation promoted
morphological changes on the surface of the biopolymer, leading to the appearance of
heterogeneous fibrillar structures along the surface area. The sulfated lasiodiplodan
showed higher thermal stability (oxidation temperatures close to 460 °C). XRD analysis
demonstrated that the native and sulfated lasiodiplodan samples present a
predominantly amorphous diffractographic profile, characteristic of polysaccharides.
Sulphation enhanced the antimicrobial activity of lasiodiplodan, especially against
Candida albicans (fungicidal activity) and Salmonella enterica Typhimurium
(bacteriostatic activity). The native sample presented higher hydroxyl radical removal
capacity, while the sulfated derivatives showed higher FRAP potential. Native and
sulfated lasiodiplodan did not demonstrate lethal cytotoxicity against wild and mutants
Saccharomyces cerevisiae strains at the concentrations studied. Samples with higher
DS (1.42 and 1.61) had lower potential for induction to oxidative stress. The results of
the present study indicate that the sulphation process can be an attractive tool to

9
improve the lasiodiplodan biological activities and could contribute to the diversification
of its biotechnological applications.
Key-words: bioactivity, fungal polysaccharides, sulfation.

10
AGRADECIMENTOS
Agradeço primeiramente a Deus pela vida, pela força que me foi concedida
nesta caminhada, principalmente nos momentos mais difíceis.
A minha família, meu pai Celso, minha mãe Izabel, e aos meus irmãos Matheus
e Marcelle, por sempre estarem ao meu lado e me apoiarem. Obrigada pela força,
carinho, paciência (e muita) e amor, ao lado de vocês esta caminhada tornou-se mais
leve. Amo vocês.
Agradeço as minhas amigas Carlise, Anna Paulla e Renata pela amizade e
apoio durante esses dois anos de mestrado. As manhãs de aula, listas, provas, dias
no laboratório, jantas mensais e conversas fizeram diferença nos meus dias. As
minhas amigas Marcieli, Jéssica, Larissa e Giulia que mesmo longe estiveram
presente em todos os momentos. Também as minhas amigas de vida, Raquel e
Àckilis. Obrigada por sempre me ouvirem, mesmo não convivendo mais dia-a-dia,
sempre me deram força. A Débora, que nesses últimos anos tornou-se uma grande
amiga e companheira de açaí, obrigada.
Aos meus colegas e amigos de laboratório de biotecnologia e bioprocessos,
que sempre me deram apoio, incentivo e que tornaram a rotina do laboratório, seja
em dias da semana, noites, fins de semana e feriados mais agradáveis e felizes.
A nossa assistente administrativa do PPGTP, Neide, que sempre foi atenciosa
em me ajudar. Meu sincero agradecimento e reconhecimento.
A minha co-orientadora, amiga, e segunda mãe Dra. Vidiany, por tudo.
Obrigada pelos ensinamentos, por sempre (sempre mesmo) estar ao meu lado, por
toda ajuda e orientação nesta caminhada. Obrigada pela palavra amiga, pelos
momentos pertinentes e intransferíveis de amizade.
Ao meu orientador Dr. Mário por confiar a mim este trabalho. Obrigada pelos
ensinamentos, atenção e orientações que foram, com certeza, fundamentais em
minha formação acadêmica e profissional.
Este estudo foi financiado em partes pela Coordenação de Aperfoiçoamento de
Pessoal de Nível Superior – Brasil (CAPES) – Código de Financiamento 001.

11
LISTA DE FIGURAS
Figura 1. Representação da formação do dissacarídeo lactose (β-D-galactopiranosil-
(1,4)-D-glicopiranose), por meio da ligação glicosídica entre os monossacarídeos β-
1,4-galactose e α-1,4-glicose. ................................................................................... 22
Figura 2. Estrutura química dos polissacarídeos vegetais amido (a) e celulose (b). . 23
Figura 3. Fungo Lasiodiplodia theobromae MMPI cultivado em ágar sabouraud com
cloranfenicol, evidenciando o abundante crescimento de micélio aéreo. .................. 25
Figura 4. Estrutura básica da parede celular fúngica. ............................................... 27
Figura 5. Estrutura química da (a) dextrana (1,6-α—D-glicosil), (b) levana (2,6-β-D-
frutosil) e (c) xantana. ................................................................................................ 28
Figura 6. Exemplos de tipos de conformações das β-glucanas, (a) β-1,3, (b) β-1,4 e
(c) β-1,6. .................................................................................................................... 30
Figura 7. Representação estrutural de β-glucanas modificadas por: (a) sulfatação, (b)
carboximetilação, (c) fosforilação, (d) acetilação, (e) selenização e (f) alquilação .... 33
Figura 8. Representação estrutural de uma glucana sulfatada no carbono 3. .......... 34
Figura 9. Representação esquemática dos diferentes alvos das substâncias
antimicrobianas na estrutura celular e mecanismos de resistência. .......................... 36
Figura 10. Estrutura química da heparina. ................................................................ 37
Figura 11. Representação esquemática do mecanismo de ação anticoagulante de
polissacarídeos sulfatados. ....................................................................................... 38
Figura 12. Reação de oxirredução do radical livre TPTZ durante avalição
antioxidante (FRAP). ................................................................................................. 40
Figura 13. Reação de hidroxilação do salicilato de sódio. ......................................... 41
Figura 14. Reação de oxirredução do radical livre DPPH durante avalição
antioxidante de DPPH. .............................................................................................. 41
Figura 15. Reação de oxirredução do radical livre ABTS durante avalição
antioxidante de ABTS. ............................................................................................... 42
Figura 16. Fluxograma da produção biotecnológica da lasiodiplodana nativa e do
processo de derivatização química por sulfatação .................................................... 46
Figura 17. Fluxograma das caracterizações estruturais e biológicas realizadas nas
amostras.................................................................................................................... 47

12
Figura 18. Sistema de microcultivo fúngico em lâmina. ............................................ 48
Figura 19. Representação esquemática do processo de produção e recuperação da
lasiodiplodana nativa (LAS-N) ................................................................................... 49
Figura 20. Processo de preparo do reagente sulfatante, utilizando ácido
clorossulfônico e piridina. .......................................................................................... 50
Figura 21. Processo de sulfatação da lasiodiplodana. .............................................. 51
Figura 22. Cultivo do fungo L. theobromae MMPI em ágar PDA: 7 dias de incubação
(A) e 14 dias de incubação (B). ................................................................................. 58
Figura 23. Microscopia ótica do fungo L. theobromae MMPI: (A) micélio aéreo na
superfície do meio PDA; (B) microcultivo em lâmina; (C) visualização das hifas com
amplitudes de 40 x e (D) 1000 x. .............................................................................. 59
Figura 24. Mecanismo reacional da sulfatação da lasiodiplodana. ........................... 61
Figura 25. Espectros de infravermelho da LAS-N, LAS-S1:10, LAS-S1:6, LAS-S1:4 e
LAS-S1:5. .................................................................................................................. 66
Figura 26. Sobreposição dos espectros de infravermelho da LAS-N e LAS-S1:5
(DS=1,61). ................................................................................................................. 67
Figura 27. Ampliações nas regiões entre (A) 4000 e 2700 cm-1, (B) 1600 e 1100 cm-
1 e (C) 1100 e 400 cm-1, comparando a LAS-N e a lasiodiplodana sulfatada (LAS-
S1:5). ......................................................................................................................... 67
Figura 28. Micrografias das amostras LAS-N, LAS-S1:10, LAS-S1:6, LAS-S1:4 e
LAS-S1:5, nas amplitudes de 300x, 800x e 1200x. ................................................... 69
Figura 29. Perfil térmico da amostra de lasiodiplodana nativa (LAS-N), pelas curvas
de DTA, DTG e TGA. ................................................................................................ 71
Figura 30. Curvas de DTA, DTG e TGA das amostras sulfatadas: (a) LAS-S1:10, DS
de 0,15, (b) LAS-S1:6, DS de 1,02, (c) LAS-S1:4, DS de 1,42 e (d) LAS-S1:5 com
DS de 1,61. ............................................................................................................... 72
Figura 31. Difratogramas das amostras de lasiodiplodana nativa (LAS-N), e das
sulfatadas (LAS-S1:5, LAS-S1:4, LAS-S1:6 e LAS-S1:10. ........................................ 74
Figura 32. Potencial de redução do íon férrico (FRAP) das amostras de LAS-N e
LAS-S. LAS-S1:5, DS=1,61; LAS-S1:4, DS=1,42; LAS-S1:6, DS=1,02; LAS-S1:10,
DS=0,15. ................................................................................................................... 79
Figura 33. Potencial de capacidade de sequestro do radical OH• das amostras de
LAS-N e LAS-S. LAS-S1:5, DS=1,61; LAS-S1:4, DS=1,42; LAS-S1:6, DS=1,02; LAS-
S1:10, DS=0,15. ........................................................................................................ 81

13
Figura 34. Potencial de captura do radical DPPH das amostras de LAS-N e LAS-S.
LAS-S1:5, DS=1,61; LAS-S1:4, DS=1,42; LAS-S1:6, DS=1,02; LAS-S1:10, DS=0,15.
.................................................................................................................................. 83
Figura 35. Potencial de captura do radical ABTS das amostras de LAS-N e LAS-S.
LAS-S1:5, DS=1,61; LAS-S1:4, DS=1,42; LAS-S1:6, DS=1,02; LAS-S1:10, DS=0,15.
.................................................................................................................................. 84

14
LISTA DE TABELAS
Tabela 1. Proporções utilizadas de ácido clorossulfônico e piridina no preparo das
amostras.................................................................................................................... 50
Tabela 2. Composição elementar (C e S) das amostras sulfatadas e seus
respectivos graus de substituição. ............................................................................ 62
Tabela 3. Solubilidade das amostras de lasiodiplodana nativa e sulfatadas em água
e DMSO 20%. ........................................................................................................... 64
Tabela 4. Potencial antimicrobiano das amostras de lasiodiplodana nativa e
sulfatada contra diferentes cepas de bactérias e leveduras. ..................................... 76
Tabela 5. Estresse oxidativo frente as cepas de leveduras testadas das amostras de
lasiodiplodana nativa e sulfatadas. ............................................................................ 86

15
LISTA DE ABREVIATURAS
5-Fu 5-Fluorouracil
ABTS Ácido 2,2’-azino-bis(3-etilbenzotiazolina-6-sulfônico
ATCC American Type Culture Collection
BHI Brain Heart Infusion
CSA Ácido Clorossulfônico
DMSO Dimetilsulfóxido
DNA Ácido Desoxirribonucleico
DPPH 2,2-difenil-1-picrilhidrazila
DRX Difratometria de Raios-X
DS Grau de Substituição
DTA Análise Térmica Diferencial
DTG Análise de Termogravimetria Derivada
DV2 Dengue Sorotipo 2
EPS Exopolissacarídeo
FRAP Poder de Redução do Íon Férrico
FTIR Espectroscopia de Infravermelho com Transformada de Fourier
GSH1 Glutamato Citeína Ligase
GST Glutationas Tranferases
GTT1 Glutationa Transferase 1
GTT2 Glutationa Transferase 2
HIV-1 Vírus da Imunodeficiência Humana
IMC Índice de Massa Corporal
IPS Polissacarídeos Intracelulares
LAS-N Lasiodiplodana Nativa
LAS-S Lasiodiplodana Sulfatada
MEV Microscopia Eletrônica de Varredura
MIC Concentração Mínima Inibitória
MRB Modificadores de Respostas Biológicas
NK Natural Killer
OH• Radical Hidroxila
Py Piridina

16
PDA Ágar Batata Dextrose
RNA Ácido Ribonucleico
RNS Espécies Reativas de Nitrogênio
ROS Espécies Reativas de Oxigênio
TGA Análise Termogravimétrica
TPTZ 2,4,6-Tri(2-pyridyl)-s-triazine
UFC Unidade de Formação de Colônias
VMSM Meio de Sais Minerais de Vogel
YPD Yeast Extract Peptone Dextrose

17
SUMÁRIO
1 INTRODUÇÃO ....................................................................................................... 19
2 OBJETIVOS ........................................................................................................... 21
2.1 Objetivo geral .................................................................................................... 21
2.2 Objetivos específicos........................................................................................ 21
3 REFERENCIAL TEÓRICO ..................................................................................... 22
3.1 Polissacarídeos ................................................................................................. 22
3.1.2 Polissacarídeos microbianos ............................................................................ 24
3.2 Lasiodiplodia theobromae ................................................................................ 25
3.3 Exopolissacarídeos (EPS) ................................................................................ 26
3.4 Glucanas ............................................................................................................ 29
3.4.1 Lasiodiplodana ................................................................................................. 31
3.5 Modificações químicas de polissacarídeos .................................................... 32
3.5.1 Modificação química por sulfatação ................................................................. 34
3.5.2 Atividades biológicas relacionadas à sulfatação .............................................. 35
3.6 Potencial antioxidante ...................................................................................... 39
3.6.1 Poder de redução do íon férrico (FRAP) .......................................................... 39
3.6.2 Sequestro do radical hidroxila .......................................................................... 40
3.6.3 Captura do radical DPPH ................................................................................. 41
3.6.4 Captura do radical ABTS .................................................................................. 42
3.7 Estresse oxidativo ............................................................................................. 42
4 MATERIAIS E MÉTODOS ..................................................................................... 46
4.1 Fluxograma das atividades desenvolvidas ..................................................... 46
4.2 Produção do exopolissacarídeo lasiodiplodana e derivatização química ... 47
4.2.1 Caracterização macro e micro-morfológica da cepa Lasiodiplodia theobromae
MMPI ......................................................................................................................... 47
4.2.2 Preparo do inóculo para produção de lasiodiplodana ...................................... 48
4.2.3 Produção da lasiodiplodana em cultivo submerso ........................................... 48
4.2.4 Derivatização química da lasiodiplodana por sulfatação .................................. 49
4.3 Caracterização química da lasiodiplodana nativa e sulfatada ...................... 51
4.3.1 Determinação do grau de sulfatação (DS) ....................................................... 51
4.3.2 Avaliação da solubilidade ................................................................................. 52

18
4.3.3 Espectroscopia no infravermelho com transformada de Fourier (IV-TF) .......... 52
4.3.4 Microscopia eletrônica de varredura (MEV) ..................................................... 53
4.3.5 Análise Térmica ................................................................................................ 53
4.3.6 Difração de raios-X (DRX) ................................................................................ 53
4.4 Caracterização biológica da lasiodiplodana nativa e sulfatada .................... 53
4.4.1 Determinação do potencial antimicrobiano ....................................................... 53
4.4.2 Atividade antioxidante ...................................................................................... 54
4.4.2.1 Poder de redução do íon férrico (FRAP) ....................................................... 54
4.4.2.2 Sequestro do radical hidroxila ....................................................................... 55
4.4.2.3 Captura do radical DPPH .............................................................................. 55
4.4.2.4 Captura do radical cátion ABTS .................................................................... 56
4.4.3 Citotoxicidade sobre células de Saccharomyces cerevisiae ............................ 56
5 RESULTADOS E DISCUSSÃO ............................................................................. 58
5.1 Aspectos morfológicos da cepa Lasiodiplodia theobromae MMPI .............. 58
5.2 Grau de sulfatação ............................................................................................ 59
5.3 Solubilidade dos derivados obtidos ................................................................ 63
5.4 Análises dos espectros de Infravermelho (IV-TF) .......................................... 65
5.5 Análise das imagens de microscopia eletrônica de varredura (MEV) .......... 68
5.6 Perfil térmico das amostras de lasiodiplodana .............................................. 71
5.7 Análise por difração de raios-X (DRX) ............................................................. 73
5.8 Atividade antimicrobiana (MIC) ........................................................................ 75
5.9 Potencial Antioxidante ...................................................................................... 79
5.9.1 Avaliação do poder de redução do íon férrico (FRAP) ..................................... 79
5.9.2 Avaliação da capacidade de sequestro do radical hidroxila ............................. 80
5.9.3 Avaliação da capacidade de captura do radical DPPH .................................... 82
5.9.4 Avaliação da captura do radical cátion ABTS ................................................... 84
5.10 Avaliação da citotoxicidade sobre Saccharomyces cerevisiae .................. 85
6 CONCLUSÕES ...................................................................................................... 88
7 REFERÊNCIAS ...................................................................................................... 90
8 APÊNDICES ........................................................................................................ 103

19
1 INTRODUÇÃO
Polissacarídeos provenientes de bactérias, leveduras, fungos filamentosos e
algas têm sido descritos na literatura científica como materiais com diferentes
potencialidades biológicas. Muitas destas macromoléculas têm atraído atenção do
setor farmacêutico por apresentarem atividades imunomoduladora, antioxidante, anti-
inflamatória, antimicrobiana e antitumoral, além de algumas contribuírem para a
redução de riscos de desenvolvimento de doenças cardiovasculares e diabetes.
Dentre os polissacarídeos microbianos, destacam-se as β-glucanas que são
biopolímeros constituídos por unidades de glucose unidas por ligações glicosídicas e
diferenciam-se pelo tamanho das cadeias poliméricas, estrutura molecular, tipo de
conformação e ramificações. É importante mencionar que as diferenças
conformacionais e estruturais interferem diretamente nas funcionalidades biológicas
que estas biomacromoléculas podem apresentar. Neste contexto, é estratégica a
busca por novas glucanas, incluindo a obtenção de derivados quimicamente
modificados, os quais podem apresentar novas funcionalidades biotecnológicas ou
ainda terem suas propriedades potencializadas.
A lasiodiplodana, uma D-glucana extracelular com estrutura linear do tipo
β(1→6) produzida pelo fungo ascomiceto Lasiodiplodia theobromae MMPI, tem
demonstrado diferentes funções biológicas, incluindo atividade antioxidante,
hipocolesterolêmica, hipoglicemiante e anticarcinogênica.
Dentre as modificações químicas que podem potencializar as funcionalidades
biológicas dos polissacarídeos, é destacada a sulfatação. A sulfatação é uma reação
que envolve a substituição nucleofílica nos grupos hidroxilas presentes nas unidades
monoméricas, por grupamentos sulfatos (O-SO3H). Tal derivatização promove
principalmente a potencialização das atividades anticoagulante, antitrombótica,
antiviral e antimicrobiana.
Polissacarídeos modificados quimicamente têm atraído cada vez mais a
atenção de pesquisadores e do setor industrial, em função de suas interessantes
propriedades tecnológicas e funcionalidades biológicas. Estes biopolímeros
constituem hoje um nicho de mercado que movimenta milhões de dólares, com
perspectivas de expansão especialmente nos setores farmacêutico e alimentício. O
mercado global de biopolímeros foi avaliado em US$ 2.465,4 milhões no ano de 2017

20
e há perspectivas de um crescimento de aproximadamente 192%, com taxa de
crescimento anual composta (CAGR) de 15,2% atingindo então um valor de US$
7.196,8 milhões até 2024 (NAGI, 2018).
Com base nas informações apresentadas, o presente trabalho teve por objetivo
a bioprodução e a derivatização química por sulfatação da lasiodiplodana ((1→6)-β-
D-glucana). Macromoléculas com diferentes graus de sulfatação foram obtidas,
caracterizadas quimicamente e submetidas a avaliação de potencialidades biológicas,
como atividade antimicrobiana e antioxidante. Além disso, foi avaliado o estresse
oxidativo dos derivados sobre cepas recombinantes de Saccharomyces cerevisiae.

21
2 OBJETIVOS
2.1 Objetivo geral
Obter e caracterizar derivados sulfatados de (1→6)-β-D-glucana.
2.2 Objetivos específicos
Produzir a (1→6)-β-D-glucana (lasiodiplodana) pelo fungo L. theobromae MMPI
em cultivo submerso;
Modificar químicamente por sulfatação, utilizando diferentes concentrações do
reagente sulfatante;
Avaliar o grau de substituição e solubilidade dos derivados obtidos;
Caracterizar por espectroscopia na região do infravermelho com transformada
de Fourier (IV-TF), microscopia eletrônica de varredura (MEV), análise térmica
(DTG, TGA e DTA) e difratometria de raios-X (DRX);
Avaliar os potenciais antioxidante e antimicrobiano;
Avaliar o estresse oxidativo da lasiodiplodana nativa e dos derivados usando
Saccharomyces cerevisiae como modelo biológico.

22
3 REFERENCIAL TEÓRICO
3.1 Polissacarídeos
Polissacarídeos são estruturas de condensação em que unidades
monossacarídicas como D-ribose, D-glicose e frutose, estão unidas entre si por
ligações glicosídicas. Eles podem diferir entre si em relação à unidade monomérica
constituinte, o grau de ramificação, massa molecular, tipo de ligações e no
comprimento de suas cadeias (XIAO; GRINSTAFF, 2017; KAGIMURA et al., 2015a).
As ligações glicosídicas ocorrem pela condensação ou desidratação do grupo
hidroxila hemiacetal de um açúcar e um grupo hidroxila de outra unidade de açúcar,
como representado na figura 1.
Figura 1. Representação da formação do dissacarídeo lactose (β-D-galactopiranosil-(1,4)-D-glicopiranose), por meio da ligação glicosídica entre os monossacarídeos β-1,4-galactose e α-1,4-glicose. Fonte: Autoria própria.
Tais macromoléculas desempenham importantes funções em sistemas
biológicos, atuando no armazenamento de energia, lubrificação e transdução de sinal
celular, bem como função estrutural (XIAO; GRINSTAFF, 2017). Polissacarídeos
podem ser encontrados em plantas e vegetais (amido e celulose), células animais
(glicogênio), algas (ágar e alginato), e ainda em sementes e exsudados de árvores
(gomas guar e arábica).
Polissacarídeos de origem vegetal têm se destacado como materiais que
possuem propriedades biotecnológicas. KUMA e KUMAR (2017), avaliaram a

23
utilização de um polissacarídeo oriundo das sementes de Albizia lebbeck L. (faveiro)
como excipiente de fármacos. Tal estudo mostrou que o polissacarídeo de faveiro
possui propriedades adequadas para uso como estabilizante de espumas
alimentícias, além de ser um material termicamente estável e sensível ao pH, podendo
ser utilizado como veículo de liberação de drogas. Similarmente, TANG et al. (2018),
extraíram três tipos de polissacarídeos de batata doce roxa e avaliaram seus efeitos
imunológicos. Neste estudo os autores verificaram que estes polissacarídeos são
capazes de estimular a resposta imune através de macrófagos e regular a imunidade
adaptativa, aumentando a produção de imunoglobulina, como revelou os testes
realizados em camundongos.
Entre os polissacarídeos vegetais tradicionalmente utilizados pelas indústrias
alimentícia e farmacêutica, destaca-se o amido, um homopolissacarídeo constituído
por unidades de D-glicopiranose (figura 2a), o qual é empregado na elaboração de
filmes biodegradáveis, utilizados em embalagens alternativas de alimentos
LUCHESE, SPADA e TESSARO (2017) e no revestimento de papéis a fim de
aumentar a resistência à umidade (LIN et al., 2017).
A celulose vegetal, um polissacarídeo constituído por subunidades de glicose
unidas por ligações do tipo (14), como demonstrado na figura 2b, também tem sido
empregada como agente transportador de medicamentos (WIJAYA et al., 2017),
filmes biodegradáveis para embalagens, como material absorvente e fibra
(PHINICHKA; KAENTHONG, 2016), além de serem utilizadas em sistemas óleo-gel
com grande campo de aplicação nas indústrias de alimentos, farmacêutica e de
cosméticos (JIANG et al., 2017).
Figura 2. Estrutura química dos polissacarídeos vegetais amido (a) e celulose (b). Fonte: Autoria própria.

24
3.1.2 Polissacarídeos microbianos
Além dos polissacarídeos de origem vegetal, os polissacarídeos de origem
microbiana obtidos de leveduras, bactérias, algas e fungos filamentosos (SOUZA;
GARCIA-CRUZ, 2004) tem ganhado espaço no mercado mundial. O interesse em
polissacarídeos microbianos está associado ao fato de tais moléculas além de
apresentarem diferentes propriedades tecnológicas (espessante, umectante,
texturizante, etc) mostram-se promissoras no tratamento de algumas doenças,
apresentando também atividades antimicrobianas, antioxidantes e antitumorais,
dentre outras bioatividades (ALZORQI et al., 2016; XU et al., 2017).
A localização celular é um fator de classificação destes biopolímeros. Há
polissacarídeos que permanecem ligados à parede celular (polissacarídeos
estruturais); outros são encontrados intracelularmente (grânulos ou inclusões
celulares), e os exopolissacarídeos, os quais são liberados para o meio extracelular
(MAHAPATRA; BANERJEE, 2013). Outro aspecto a ser considerado, é que os
polissacarídeos microbianos podem ser sintetizados em diferentes fases do
crescimento microbiano, sendo o mecanismo biossintético, particularmente específico
para cada tipo de microrganismo (SOUZA; GARCIA-CRUZ, 2004).
DI et al. (2017) caracterizaram quimicamente e avaliaram algumas atividades
biológicas de exopolissacarídeos produzidos por espécies de Lactobacillus. Os
autores verificaram que, além da possibilidade de utilização em produtos alimentícios
funcionais, tais polissacarídeos desempenharam efeito inibitório significativo contra a
proliferação de células tumorais colorretais HT-29.
Estudos desenvolvidos por HAN et al. (2016), mostraram a produção de
polissacarídeo exocelular (exopolissacarídeo) pela levedura Sporidiobolus pararoseus
JD-2. Esta molécula demonstrou caráter aniônico capaz de potencializar funções
imunológicas, além de poder ser empregado como espessante em indústrias de
alimentos.
Os fungos filamentosos são os principais produtores de polissacarídeos, os
quais apresentam 75% de sua parede celular composta por tais macromoléculas
(FUKUDA et al., 2009). Diferentes fungos têm sido mencionados na literatura científica
como produtores de polissacarídeos, dentre os quais destacam-se o Cordyceps
kyushuensis (ZHANG et al., 2015), Botryosphaeria rhodina (GIESE et al., 2011),

25
Lasiodiplodia theobromae MMPI (KAGIMURA et al., 2015b), Aureobasidium pullulans
(AOKI et al., 2015), Botrytis cinerea (JADHAV; GUPTA, 2016) e Epicoccum nigrum
(SCHMID et al., 2006).
3.2 Lasiodiplodia theobromae
O Lasiodiplodia theobromae é um fungo filamentoso pertencente à família
Botryosphaeriaceae, gênero Lasiodiplodia, anteriormente designado como
Botryosphaeria theobromae (MUNIZ et al., 2011). Este fungo é a representação do
estado assexuado do Botryosphaeria rhodina porém, há isolados diferentes que
podem apresentar características variáveis quanto a coloração e velocidade de
crescimento (SOMENSI, 2014). O L. theobromae apresenta abundante micélio aéreo
e a coloração das colônias, como demonstrado na figura 3, pode variar de branco à
cinza e até negro, dependendo do substrato e tempo de incubação (RODRIGUES,
2003).
Figura 3. Fungo Lasiodiplodia theobromae MMPI cultivado em ágar sabouraud com cloranfenicol, evidenciando o abundante crescimento de micélio aéreo. Fonte: Autoria própria.
L. theobromae é um fitopatógeno típico das regiões tropicais e subtropicais, que
causa sérios prejuízos a numerosas espécies vegetais cultivadas. Este fungo infecta
os caules levando à redução no transporte de nutrientes e água, com consequente
redução da fotossíntese e morte do cultivar (CIPRIANO et al., 2015). O L. theobromae

26
tem sido associado a manchas foliares, necrose e gomose de plantas, incluindo
herbáceas e plantas lenhosas. Estudos revelam que o L. theobromae pode causar
doenças em flores como a Rosa rugosa (CHEN et al., 2016), cajueiros (CIPRIANO et
al., 2015), Falcataria moluccana (JI et al., 2017) e abacateiros (MAFTOONAZAD et
al., 2007). Menos frequentemente, também tem sido associado a doenças
subcutâneas em humanos (PAPACOSTAS et al., 2015).
Em contrapartida, este fungo tem se mostrado um eficiente produtor de
exopolissacarídeo (EPS), como a lasiodiplodana ((1→6)-β-glucana) quando cultivado
sob fermentação (KAGIMURA et al., 2015a).
Similarmente, (OLIVEIRA et al., 2015) reportaram a produção de dois tipos de
exopolissacarídeos, uma (1→3)(1→6)-β-glucana e uma (1→6)-β-glucana pela cepa
L. theobromae MMBJ em cultivo submerso. Estes autores avaliaram a atividade
imunomoduladora e verificaram a presença de citocinas como TFN-α (pró-
inflamatória) e IL-10 (anti-inflamatória) envolvidas na atividade de modulação, sendo
verificada resposta dose-dependente em relação a citocina pró-inflamatória.
3.3 Exopolissacarídeos (EPS)
Os polissacarídeos microbianos comumente são produzidos como constituintes
da parede celular, onde desempenham funções estruturais e de proteção contra
estresses exteriores. A composição polissacarídica da parede celular fúngica típica
está apresentada na figura 4. Entretanto, alguns polissacarídeos podem ser
excretados para o meio extracelular, sendo estes denominados exopolissacarídeos
(EPS). Os EPS apresentam vantagens em relação aos polissacarídeos intracelulares
e de parede celular, quanto a sua obtenção ou disponibilidade, pois apresentam
elevada produtividade, maior facilidade de isolamento e purificação, além da
possibilidade de serem produzidos a partir de resíduos agroindustriais como matéria
prima (MAHAPATRA; BANERJEE, 2013; DONOT; FONTANA; SCHORR-GALINDO,
2017).

27
Figura 4. Estrutura básica da parede celular fúngica. Fonte: Adaptado de GEOGHEGAN, STEINBERG e GURR, 2017).
A biossíntese de exopolissacarídeos por microrganismos pode estar
relacionada tanto a capacidade de sobrevivência quando em condições adversas,
quanto em condições ótimas de incubação, onde produzem estas biomoléculas como
reserva energética. Tais biomoléculas podem apresentar uma grande variedade de
combinações estruturais, e são classificadas em relação a sua composição
monomérica, em homopolissacarídeos e heteropolissacarídeos.
Homopolissacarídeos são constituídos por um único tipo de monossacarídeo na forma
de unidade repetida, como as dextranas e levanas (Figuras 5 a e b). Já os
heteropolissacarídeos, são constituídos por vários tipos de unidades
monossacarídicas, como a goma xantana (Figura 5 c) (DONOT; FONTANA;
SCHORR-GALINDO, 2012).
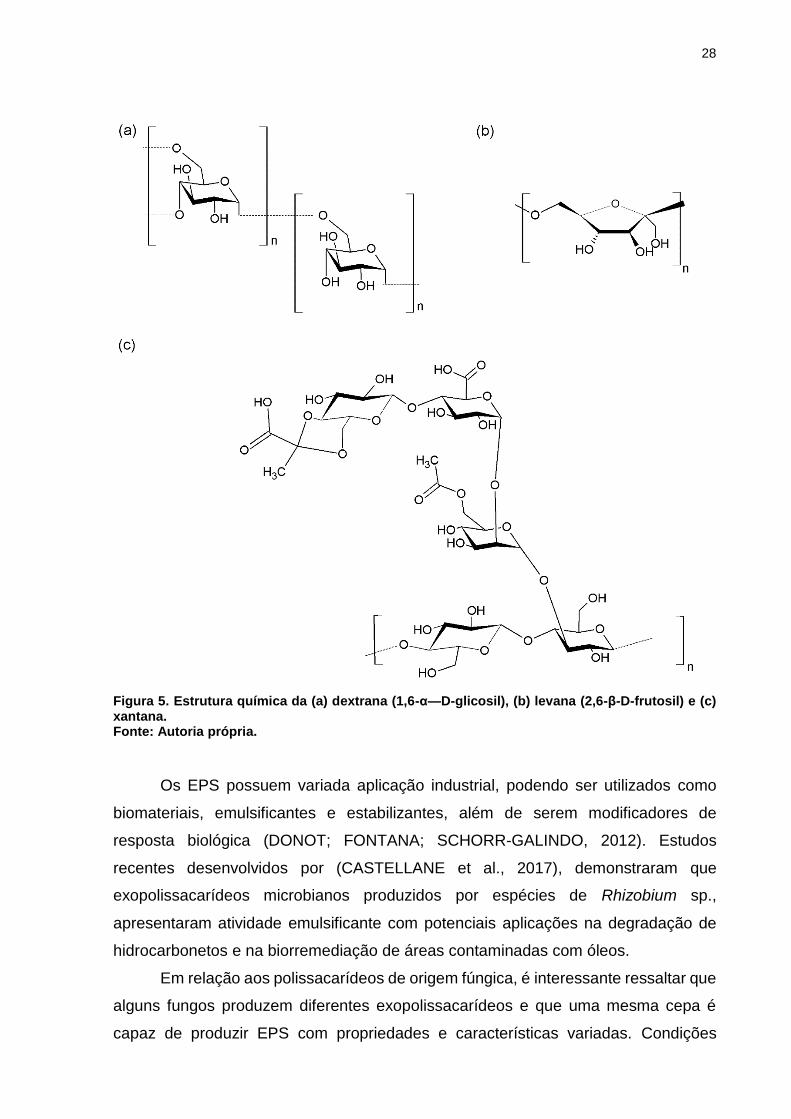
28
Figura 5. Estrutura química da (a) dextrana (1,6-α—D-glicosil), (b) levana (2,6-β-D-frutosil) e (c) xantana. Fonte: Autoria própria.
Os EPS possuem variada aplicação industrial, podendo ser utilizados como
biomateriais, emulsificantes e estabilizantes, além de serem modificadores de
resposta biológica (DONOT; FONTANA; SCHORR-GALINDO, 2012). Estudos
recentes desenvolvidos por (CASTELLANE et al., 2017), demonstraram que
exopolissacarídeos microbianos produzidos por espécies de Rhizobium sp.,
apresentaram atividade emulsificante com potenciais aplicações na degradação de
hidrocarbonetos e na biorremediação de áreas contaminadas com óleos.
Em relação aos polissacarídeos de origem fúngica, é interessante ressaltar que
alguns fungos produzem diferentes exopolissacarídeos e que uma mesma cepa é
capaz de produzir EPS com propriedades e características variadas. Condições

29
fermentativas específicas interferem diretamente na produção dos exopolissacarídeos
fúngicos, dentre as quais destacam-se o pH, temperatura (entre 22 e 30 °C), aeração,
velocidade de agitação, tempo de incubação, composição do meio de cultura, fonte
de carbono, concentração da fonte de carbono (glicose, maltose, entre outros), fonte
de nitrogênio (orgânica como peptona e inorgânica como sais de amônio) e ainda a
adição de sais contendo fosfato como fosfato monopotássico (KH2PO4), fosfato
dipotássico (K2HPO4) e sulfato de magnésio heptahidratado (MgSO4.7H2O) (SOUZA,
GARCIA-CRUZ, 2004; MAHAPATRA, BANERJEE, 2013).
Neste sentido, CUNHA et al. (2012), avaliaram a influência de parâmetros
fermentativos como fonte de carbono e nitrogênio e forma de condução da
fermentação para a produção do EPS lasidiplodana ((1→6)-β-D-glucana). Maiores
rendimentos foram obtidos utilizandondo-se glicose e extrato de levedura como fontes
de carbono e nitrogênio, respectivamente, e em fermentações conduzidas em frascos
agitados. Da mesma forma, ZHU, DU, XU, (2016), avaliaram os efeitos da composição
do meio de cultura em relação a fonte de carbono (sacarose, glicose e amido solúvel)
e nitrogênio (peptona, extrato de carne e extrato de levedura) sobre o crescimento
micelial, massa molecular e potencial antitumoral de polissacarídeos intracelulares
(IPS) oriundos de Cordyceps gunnii. Estes autores verificaram que as fontes de
carbono e nitrogênio mais eficientes foram sacarose e peptona, respectivamente.
Comumente, as fontes de carbono e nitrogênio empregadas nos meios de produção
de EPS tem grande influência tanto no crescimento celular, como na biossíntese dos
EPS.
3.4 Glucanas
As glucanas são polímeros de glucose encontradas nas paredes celulares de
microrganismos (leveduras e fungos filamentosos), algas e cereais, como aveia e
cevada, ou ainda podem ser produzidas e excretadas (EPS) para o meio de cultivo
por algumas bactérias e fungos filamentosos (KAGIMURA et al., 2015b). Podem
diferenciar entre si em relação ao tamanho da cadeia, massa molecular, solubilidade
em água e tipo de ligação glicosídica, podendo apresentar-se na configuração α ou β
e estrutura linear ou ramificada (MORENO-MENDIETA et al., 2017).

30
-Glucanas de origem vegetal encontram-se geralmente sob a forma de
cadeias lineares unidas por ligações glicosídicas (1→4) com algumas ligações (1→3),
já as obtidas de algas e bactérias comumente apresentam cadeias lineares do tipo
(1→3), enquanto que as de origem fúngica e produzidas por líquens apresentam
ligações do tipo (1→3) e (1→6) (MORENO-MENDIETA et al., 2017), como
representado na figura 6.
Figura 6. Exemplos de tipos de conformações das β-glucanas, (a) β-1,3, (b) β-1,4 e (c) β-1,6. Fonte: Autoria própria.
As β-glucanas são carboidratos que predominam em fungos, as quais atuam
como agentes estruturais de parede celular e destacam-se na área biotecnológica por
atuarem como modificadores de respostas biológicas (MRB) (MORENO-MENDIETA
et al., 2017 ;KAGIMURA et al., 2015b). Tais macromoléculas têm apresentado
também propriedades antioxidantes (JIANG et al., 2016) e antimicrobianas (KHAN et
al., 2016), dentre várias outras.
Além disso, estas biomoléculas têm grande aplicabilidade em diferentes
setores industriais, por possuírem propriedades físicas como capacidade de
espessamento, estabilização, emulsificação e gelificação (KAGIMURA et al., 2015a).
SARTESHNIZI et al. (2014) estudaram a formulação de salsicha prebiótica
contendo amido, β-glucana de aveia e amido resistente, avaliando as interações entre
tais macromoléculas na salsicha. No embutido, a β-glucana exerceu efeito tecnológico
positivo em relação a dureza, cor e suculência.
No setor de cosméticos, as β-glucanas destacam-se por apresentarem
propriedades antioxidantes e anti-idade (KAGIMURA et al., 2015a) e têm ganhado
importância no mercado mundial por apresentarem importantes propriedades
biológicas. No estudo desenvolvido por LIU et al. (2016), foi avaliado o mecanismo de

31
ação de uma β-D-glucana extraída de aveia, sobre o controle dos níveis de glicose
sanguínea e efeito hepatogênico em camundongos. Estes autores, verificaram que tal
-glucana, em doses de médias a altas, demonstrou grande efeito hipoglicemiante em
ratos induzidos a hiperglicemia com estreptozotocina e nicotinamida.
HUSSAIN, RATHER, SURADKAR (2017) avaliaram uma β-D-glucana extraída
de aveia e irradiada com raios gama contra diferentes tipos de células cancerígenas
(células cancerígenas de cólon humano (Colo-205), células de câncer de mama ductal
humana, células tumorais epiteliais (T47D), células de adenocarcinoma de mama
humana (MCF7) e queratinócitos humanos). As β-D-glucanas irradiadas
apresentaram efeito antiproliferativo frente a linhagens celulares de câncer de Colo-
205 e MCF-7. Além disso, as β-D-glucanas irradiadas tiveram sua solubilidade e
capacidade de absorção de água aumentadas.
AOE et al. (2017), investigaram a adoção de uma dieta baseada na utilização
de cevada com elevado teor de β-glucana, a fim de verificar se esta reduziria a
obesidade e a gordura visceral em indivíduos japoneses. A ingestão de cevada com
alto teor de β-glucana levou a reduções significativas da gordura visceral, além da
redução do IMC (índice de massa corporal), peso corporal e circunferência da cintura;
indicando que a implementação de polissacarídeos como as β-glucanas na dieta,
podem contribuir para a prevenção da obesidade.
3.4.1 Lasiodiplodana
A lasiodiplodana é uma (1→6)-β-D-glucana extracelular produzida pelo
ascomiceto Lasiodiplodia theobromae MMPI, quando cultivado em meio submerso
rico em açúcares, especialmente glicose e sacarose (CUNHA et al., 2012).
Glucanas com configuração β(1→6) são as mais incomuns entre as β-
glucanas, sendo encontradas como constituintes de paredes celulares de fungos
filamentosos (CUNHA et al., 2012). Entretanto, a lasiodiplodana ((1→6)-β-D-glucana)
produzida pelo L. theobromae MMPI é uma glucana extracelular que pode ser
precipitada com etanol 95% (v/v) e facilmente recuperada do caldo de cultivo. O
processo de produção da lasiodiplodana é mais simples, mais econômico, e

32
ambientalmente menos agressivo quando comparado aos processos tradicionais de
extração de glucana de parede celular microbiana e vegetal.
Esta macromolécula tem demonstrado diferentes funções biológicas, dentre as
quais destacam-se atividades antioxidante, hipoglicemiante, hipocolesterolêmica e
anticarcinogênica (CUNHA et al., 2012; TÚRMINA et al., 2012; KAGIMURA et al.,
2015b).
Outra propriedade biológica da lasiodiplodana é seu efeito protetor contra
danos ao DNA induzido pela droga anticarcinogênica doxorrubicina, descrito
recentemente por MELLO et al. (2017). Estes autores investigaram os efeitos
genotóxicos da lasiodiplodana, sua potencial capacidade protetora contra danos ao
DNA e seu impacto na expressão de genes associados aos danos e as vias de
resposta inflamatória. Foi verificado que a lasiodiplodana não induziu distúrbios na
estabilidade do DNA e foi capaz de reduzir significativamente os danos ao DNA e o
desenvolvimento de processos inflamatórios. Além disso, TÚRMINA et al. (2012),
avaliaram os efeitos toxicológicos do tratamento sub-crônico de camundongos de
ambos os gêneros, com lasiodiplodana (1→6)-β-D-glucana, por meio da avaliação dos
efeitos bioquímicos, hematológicos e histopatológicos. Tais autores descreveram que
houve redução significativa da glicemia (35%) no grupo dos camundongos machos; e
que o tratamento com a lasiodiplodana não promoveu alterações hematológicas e
histológicas que levassem a sinais de toxicidade nos animais, independente do
gênero.
3.5 Modificações químicas de polissacarídeos
Nem todos os polissacarídeos naturais demonstram propriedades biológicas.
Desta forma, derivatizações químicas na estrutura primária da macromolécula têm
sido uma importante ferramenta para potencializar e/ou criar funcionalidades
biológicas (LI, DAI, SHAH, 2017; LI et al., 2016). Estas atividades estão diretamente
relacionadas à estrutura molecular do polissacarídeo, incluindo a composição
monomérica, ligação glicosídica da cadeia principal, grau de ramificação, grau de
substituição, conformação espacial das cadeias e solubilidade da biomolécula (MA et
al., 2012).
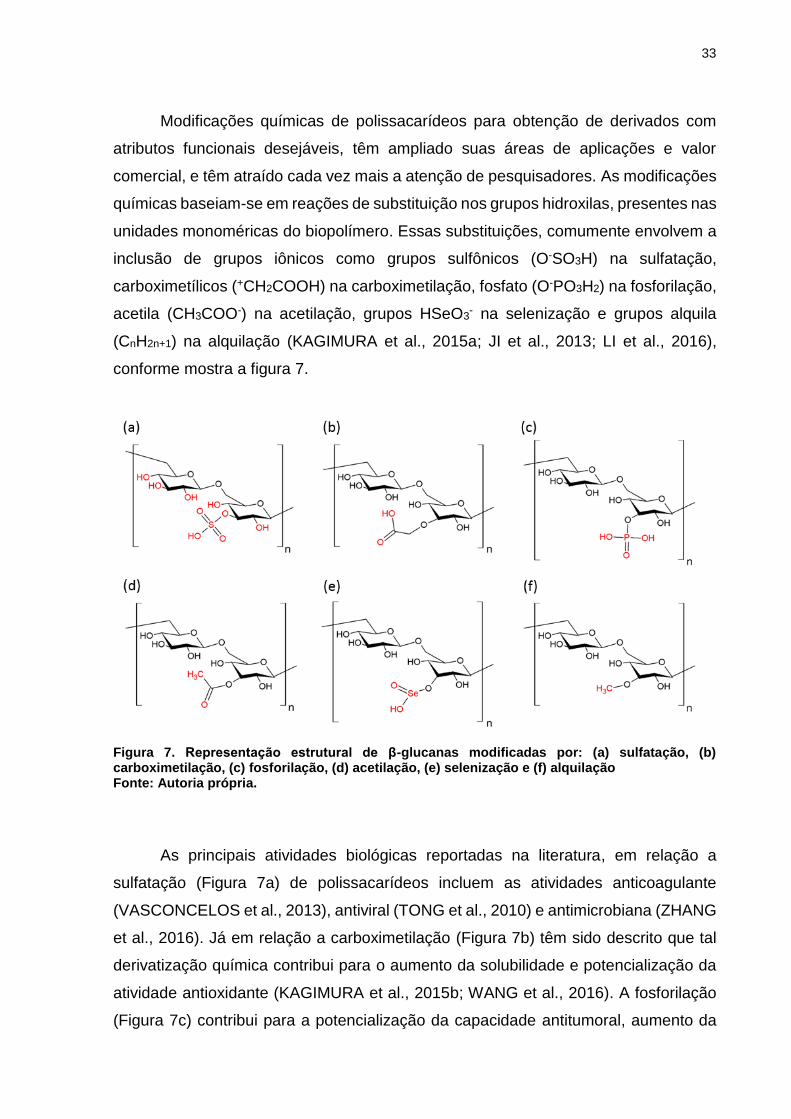
33
Modificações químicas de polissacarídeos para obtenção de derivados com
atributos funcionais desejáveis, têm ampliado suas áreas de aplicações e valor
comercial, e têm atraído cada vez mais a atenção de pesquisadores. As modificações
químicas baseiam-se em reações de substituição nos grupos hidroxilas, presentes nas
unidades monoméricas do biopolímero. Essas substituições, comumente envolvem a
inclusão de grupos iônicos como grupos sulfônicos (O-SO3H) na sulfatação,
carboximetílicos (+CH2COOH) na carboximetilação, fosfato (O-PO3H2) na fosforilação,
acetila (CH3COO-) na acetilação, grupos HSeO3- na selenização e grupos alquila
(CnH2n+1) na alquilação (KAGIMURA et al., 2015a; JI et al., 2013; LI et al., 2016),
conforme mostra a figura 7.
Figura 7. Representação estrutural de β-glucanas modificadas por: (a) sulfatação, (b) carboximetilação, (c) fosforilação, (d) acetilação, (e) selenização e (f) alquilação Fonte: Autoria própria.
As principais atividades biológicas reportadas na literatura, em relação a
sulfatação (Figura 7a) de polissacarídeos incluem as atividades anticoagulante
(VASCONCELOS et al., 2013), antiviral (TONG et al., 2010) e antimicrobiana (ZHANG
et al., 2016). Já em relação a carboximetilação (Figura 7b) têm sido descrito que tal
derivatização química contribui para o aumento da solubilidade e potencialização da
atividade antioxidante (KAGIMURA et al., 2015b; WANG et al., 2016). A fosforilação
(Figura 7c) contribui para a potencialização da capacidade antitumoral, aumento da

34
solubilidade e atividade antioxidante (YE et al., 2013; DENG et al., 2015). A
modificação por acetilação (Figura 7d) tem a capacidade de melhorar a capacidade
antioxidante de alguns polissacarídeos (SONG et al., 2013). A selenização (Figura 7e)
auxilia na melhora da atividade imunológica (QIN et al., 2013) e a alquilação (Figura
7f) demostra potencializar atividades antimicrobianas (XIE et al., 2002).
3.5.1 Modificação química por sulfatação
Polissacarídeos sulfatados podem ser naturalmente encontrados como
componente de parede celular de algas marinhas, ou obtidos por reações de
derivatização química (KRICHEN et al., 2015; PEASURA et al., 2015; SUN et al.,
2016). A sulfatação (Figura 8) consiste em uma substituição nucleofílica nos grupos
hidroxilas por grupamentos sulfatos (O-SO3H) e tem sido realizada em polissacarídeos
como glucanas, pectinas e quitosanas (WANG et al., 2015; LI, DAI, SHAH, 2017).
Figura 8. Representação estrutural de uma glucana sulfatada no carbono 3. Fonte: Autoria própria.
Na sulfatação utiliza-se um solvente orgânico e um agente sulfatante, sendo
empregados como agentes sulfatantes o ácido sulfúrico, trióxido de enxofre-piridina,
ácido clorossulfônico-piridina ou trióxido de enxofre-dimetilacetamida e
dimetilsulfóxido (DMSO), a dimetilformamida e formamida tem sido utilizadas como
solventes (LU et al., 2008). A eficiência da reação, ou seja, o número de hidroxilas do
polissacarídeo substituídas por grupos sulfonatos é avaliada pelo grau de substituição
(DS), o qual é influenciado principalmente por parâmetros como temperatura, tempo

35
de reação, concentração do agente derivatizante e proporção entre agente sulfatante
e solvente orgânico (LU et al., 2008; LI, DAI, SHAH, 2017).
A sulfatação comumente é feita pela solubilização ou suspensão do
polissacarídeo em dimetilsulfóxido (DMSO), formamida ou dimetilformamida, sendo
adicionando na sequência o catalisador, geralmente piridina, e então, o agente
sulfatante. O catalisador e o agente sulfatante podem ser adicionados separadamente
ou em conjunto, formando um complexo catalisador - agente sulfatante (ZHANG et al.,
2011; VASCONCELOS et al., 2013). A reação é então neutralizada, com bicarbonato
de sódio ou hidróxido de sódio e a mistura reacional é submetida a intensa diálise para
pré-purificação e seleção de massa molecular desejada (LI; DAI; SHAH, 2017).
O grau de substituição (DS) que representa o número de sítios (hidroxilas)
substituídos por unidade de monômero do polímero sulfatado é avaliado pela
determinação do conteúdo de enxofre inserido na macromolécula, sendo comumente
empregado o método cloreto de bário-gelatina ou avaliação instrumental por análise
elementar (VASCONCELOS et al., 2013; WAN-MOHTAR et al., 2016).
3.5.2 Atividades biológicas relacionadas à sulfatação
A sulfatação tem sido amplamente utilizada para potencializar propriedades
biológicas, principalmente, as propriedades anticoagulante, antiviral, antitrombótica e
antimicrobiana (VASCONCELOS et al., 2013; ZHANG et al., 2016).
BUDDANA, VARANASI, SHETTY (2015) analisaram as atividades biológicas
de uma α-(1→3)-glucana extraída de Streptococcus mutans e derivatizada por
sulfatação. Os autores verificaram que a amostra derivatizada com grau de
substituição (DS) de 1,19, apresentou aumento nas atividades biológicas, incluindo
maior efeito antimicrobiano frente a bactérias Gram negativas como Pseudomonas
aeruginosa MTCC 6458, Stenotrophomonas mutans MTCC 497 e Escherichia coli
MTCC 1680. Além disso, também verificaram atividade anti-inflamatória e fibrinolítica
na glucana sulfatada.
LI e SHAH (2014) avaliaram os efeitos antimicrobianos e antioxidantes de
polissacarídeos obtidos de Streptococcus thermophilus e Pleurotus eryngii após
sulfatação com DS de 0,31 e 0,69, respectivamente. Tais autores reportaram que a
sulfatação potencializou a atividade antioxidante, a qual foi avaliada por diferentes

36
protocolos (DPPH, superóxido, radical hidroxila e FRAP). Similarmente após a
sulfatação os polissacarídeos demonstraram atividade inibitória contra Escherichia
coli, Staphylococcus aureus e Listeria monocytogenes.
Em relação à atividade antimicrobiana, o mecanismo de ação de
polissacarídeos ainda não está claro, entretanto HE et al. (2010) propôs que estas
biomoléculas promovem o rompimento da parede celular e da membrana
citoplasmática, levando ao extravasamento de moléculas essenciais, além de causar
decomposição do DNA do microrganismo (Figura 9). É importante destacar que
diferentes substâncias antimicrobianas podem atuar em diferentes alvos da estrutura
celular, como na parede celular, síntese de DNA e RNA, síntese de folato, membrana
celular e síntese de proteínas. Já o mecanismo de resistência das células a agentes
antimicrobianos pode ocorrer, por exemplo, por efluxo e inativação de enzimas,
conforme demostrado no esquema criado por LEWIS (2013) e representado na figura
9.
Figura 9. Representação esquemática dos diferentes alvos das substâncias antimicrobianas na estrutura celular e mecanismos de resistência. Fonte: Adaptado de LEWIS, 2013.
VASCONCELOS et al. (2013), modificaram quimicamente por sulfonação, uma
(1→6)-β-D-glucana, produzida pelo fungo Lasiodiplodia theobromae MMRL. Os

37
autores obtiveram derivado (glucana sulfonada) com DS de 0,95 e verificaram que o
polissacarídeo exibiu atividade anticoagulante após a sulfonação.
LIANG et al. (2018) descreveram um polissacarídeo sulfatado com
características anticoagulantes similares as das heparinas (Figura 10), atuando
através de cargas negativas de grupos sulfatos, que neutralizam os resíduos de
aminoácidos carregados positivamente por ligações eletrostáticas à antitrombina,
melhorando assim a atividade anticoagulante, como ilustrado por POMIN (2012) no
esquema apresentado na figura 11.
Figura 10. Estrutura química da heparina. Fonte: Autoria própria.

38
Figura 11. Representação esquemática do mecanismo de ação anticoagulante de polissacarídeos sulfatados. Fonte: Adaptado de POMIN (2012).
Em relação a capacidade antiviral, estudos desenvolvidos por TONG et al.
(2010) avaliaram a atividade antiviral de uma α-D-glucana sulfonada frente a infecção
do vírus da dengue sorotipo 2 (DV2), incluindo a infecção inicial e a replicação
intracelular. Os resultados revelaram que a glucana sulfatada exerceu efeito inibitório
potente sobre o DV2, através da interferência entre a interação do vírus com as células
alvo. Da mesma forma, MURRY et al. (2014) avaliaram os inibidores químicos via
sulfatação contra o vírus HIV-1. Neste estudo, os autores verificaram que esses tipos
de inibidores bloqueiam eficientemente a iniciação da transcrição do vírus, durante a
reativação da latência.
SUN et al. (2009), estudaram a atividade antitumoral após sulfatar a glucana
((1→3)-β-D-glucana) extraída de corpos de frutificação do basidiomiceto Russula
virescens. Tais autores descreveram que todos os derivados sulfatados (um total de
cinco derivados) exibiram atividade antitumoral aprimorada contra Sarcoma 180. Além
disso, este estudo revelou que os derivados apresentaram toxicidade menor que o 5-
Fu (agente anticancerígeno conhecido), atuando apenas nas células doentes e não
nas células saudáveis, estimulando assim o mecanismo de resposta imune do
organismo.

39
Como descrito por SHI (2016) os polissacarídeos atuam ativando o sistema
imunológico de um organismo, ou seja, promovendo a maturação, diferenciação e
reprodução de três tipos de células: os linfócitos, macrófagos e as células NK (Natural
Killer), porém, o mecanismo de ação da atividade antitumoral por estas células ainda
não está completamente esclarecido.
3.6 Potencial antioxidante
Os antioxidantes são moléculas capazes de diminuir ou prevenir a oxidação de
outras moléculas e quando atuam em sistemas biológicos, evitam o estresse oxidativo
que causam danos celulares ao organismo e podem levar a doenças cardiovasculares
e inflamatórias (BOROSKI et al., 2015).
O aumento na busca de antioxidantes naturais, tem crescido em todo mundo
em função da toxicidade dos antioxidantes sintéticos, especialmente quando utilizados
em grandes quantidades. Além disso, antioxidantes naturais são capazes de fornecer
diversos outros efeitos benéficos à saúde devido a outras propriedades biológicas
presentes (BOROSKI et al., 2015).
Em estudos científicos é importante conduzir diferentes ensaios para a
avaliação da atividade antioxidante, a fim de avaliar adequadamente o potencial de
remoção de substâncias radicalares e potencial redutor. Cada protocolo de avaliação
de atividade antioxidante tem suas especificidades e a determinação da capacidade
antioxidante de uma molécula, pode ser feita por métodos in vivo e in vitro.
Neste estudo, foram abordadas cinco metodologias in vitro para determinação
do potencial antioxidante da lasiodiplodana nativa (LAS-N) e dos derivados sulfatados
(LAS-S). As metodologias empregadas foram: determinação do poder de redução do
íon férrico (FRAP), sequestro do radical hidroxila, captura do radical DPPH, eliminação
de peróxido de hidrogênio (H2O2) e captura do radical ABTS, cujos fundamentos
metodológicos são descritos a seguir.
3.6.1 Poder de redução do íon férrico (FRAP)
Este método de verificação do potencial antioxidante utiliza o complexo férrico
de 2,4,6-tripiridil-1,3,5-triazina (Fe3+ - TPTZ), na presença de uma substância

40
antioxidante redutora e em meio ácido. O complexo recebe um elétron e é reduzido a
(Fe2+ - TPTZ), como ilustrado na figura 12. O complexo (Fe2+ - TPTZ) apresenta
intensa coloração azul a qual é lida em 593 nm (BOROSKI et al., 2015; LI, DAI, SHAH,
2017).
Figura 12. Reação de oxirredução do radical livre TPTZ durante avalição antioxidante (FRAP). Fonte: Adaptado de LI, DAI, SHAH, 2017.
3.6.2 Sequestro do radical hidroxila
Nesta metodologia empregada, é determinada a capacidade que uma amostra
possui de sequestrar os radicais hidroxilas que são gerados na Reação Fenton, entre
sulfato ferroso (FeSO4) e peróxido de hidrogênio (H2O2), como representado na
equação (1).
𝑭𝒆𝟐+ + 𝑯𝟐𝑶𝟐 → 𝑭𝒆𝟑+ + °𝑶𝑯 + 𝑶𝑯− (Equação 1)
Os antioxidantes podem ser capazes de prevenir a formação de radicais
hidroxila por meio da inativação de íons metálicos livres por quelação ou pela
conversão de H2O2 em compostos inofensivos, como água e oxigênio molecular (LI,
DAI, SHAH, 2017). Os radicais hidroxilas que não são sequestrados participam da
reação de hidroxilação do salicilato de sódio, também presente neste procedimento,
desta forma, quanto maior a capacidade antioxidante da amostra, menos salicilato
hidroxilado estará presente no meio (Figura 13).
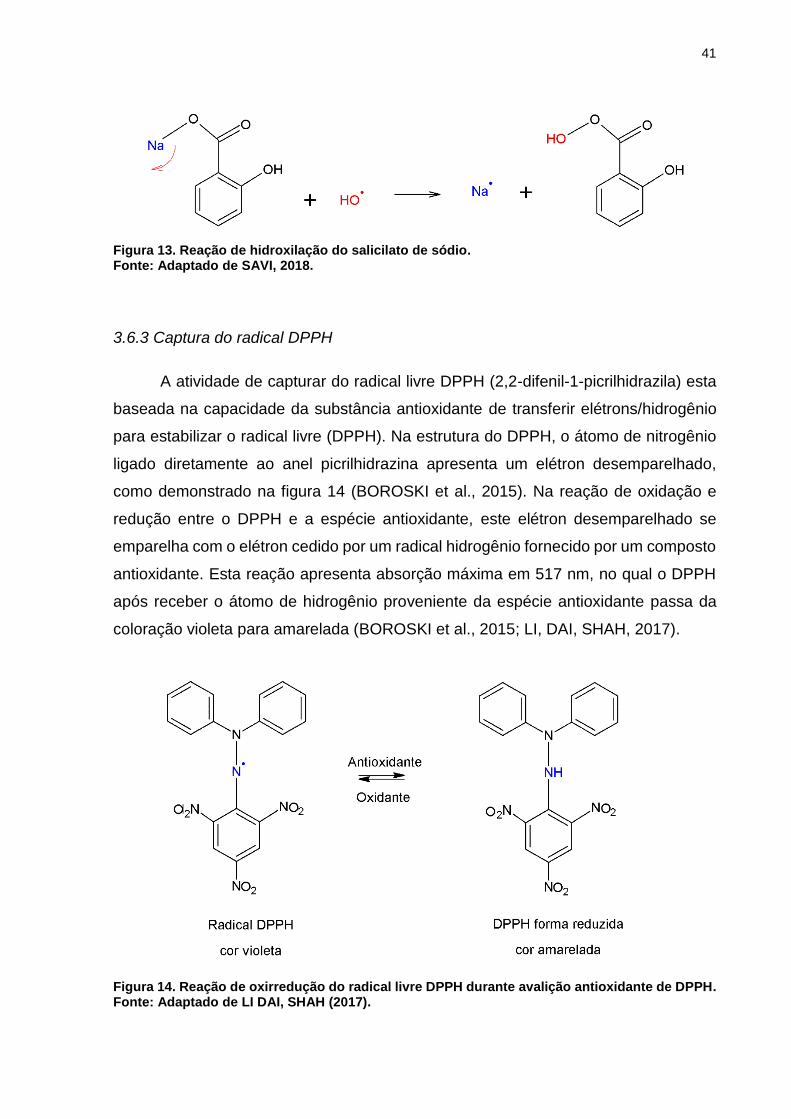
41
Figura 13. Reação de hidroxilação do salicilato de sódio. Fonte: Adaptado de SAVI, 2018.
3.6.3 Captura do radical DPPH
A atividade de capturar do radical livre DPPH (2,2-difenil-1-picrilhidrazila) esta
baseada na capacidade da substância antioxidante de transferir elétrons/hidrogênio
para estabilizar o radical livre (DPPH). Na estrutura do DPPH, o átomo de nitrogênio
ligado diretamente ao anel picrilhidrazina apresenta um elétron desemparelhado,
como demonstrado na figura 14 (BOROSKI et al., 2015). Na reação de oxidação e
redução entre o DPPH e a espécie antioxidante, este elétron desemparelhado se
emparelha com o elétron cedido por um radical hidrogênio fornecido por um composto
antioxidante. Esta reação apresenta absorção máxima em 517 nm, no qual o DPPH
após receber o átomo de hidrogênio proveniente da espécie antioxidante passa da
coloração violeta para amarelada (BOROSKI et al., 2015; LI, DAI, SHAH, 2017).
Figura 14. Reação de oxirredução do radical livre DPPH durante avalição antioxidante de DPPH. Fonte: Adaptado de LI DAI, SHAH (2017).

42
É interessante ressaltar que a acessibilidade estérica do DPPH é um fator de
limitação da reação, de forma que moléculas maiores apresentam menor acesso ao
sítio ativo do radical, do que aquelas menores, apresentando dessa forma diferença
na capacidade antioxidante nestas moléculas.
3.6.4 Captura do radical ABTS
A metodologia de captura do ABTS (ácido 2,2’-azino-bis(3-etilbenzotiazolina-6-
sulfônico) (Figura 15), é baseada na capacidade da substância antioxidante de
estabilizar o radical cátion ABTS presente na solução, que volta à forma de composto
neutro ABTS, através de uma reação de oxidação. Este deslocamento de equilíbrio
entre essas duas formas do ABTS, ocasiona a descoloração da solução verde escura,
que vai ficando mais clara à medida que o ABTS vai sendo convertido em sua forma
neutra, ocasionando dessa forma a diminuição da absorbância em 734 nm (BOROSKI
et al., 2015).
Figura 15. Reação de oxirredução do radical livre ABTS durante avalição antioxidante de ABTS. Fonte: Adaptado de BOROSKI et al. (2015).
3.7 Estresse oxidativo
O estresse oxidativo é definido como um desequilíbrio entre a geração de ROS
(espécies reativas de oxigênio), RNS (espécies reativas de nitrogênio) ou
antioxidantes celulares, que podem causar danos à membranas celulares, proteínas,
lipídios, carboidratos e à ácidos nucleicos (CASTRO et al., 2008; CHIARELLO et al.,
2018).

43
Tais estruturas oxidantes as quais constituem os denominados ROS e RNS
(superóxido, radicais hidroxila, peróxido de hidrogênio e óxido cítrico) fazem parte do
metabolismo celular, ocorrendo principalmente nas mitocôndrias como produto de
reações enzimáticas da cadeia respiratória e da fagocitose, e também em reações
não enzimáticas como aquelas que envolvem oxigênio e compostos orgânicos,
(CHIARELLO et al., 2018). Dessa forma o processo de estresse oxidativo tem sido
relacionado ao processo natural de envelhecimento celular e com doenças
neurodegenerativas como o Alzheimer, diabetes, aterosclerose, doenças
inflamatórias e câncer (CHEIGNON et al., 2018).
Recentemente células de leveduras vêm sendo utilizadas como modelo
biológico para estudos de mecanismos moleculares de resistência ao estresse
oxidativo, por apresentarem algumas semelhanças com células de mamíferos, como
a síntese de algumas proteínas (COSTA, MORADAS-FERREIRA, 2001;
SUBHASWARAJ et al., 2017). Muitos genes que codificam proteínas mitocondriais
são altamente conservados entre eucariotos, o que possibilita o uso deste
microrganismo como um modelo para estudos fisiológicos de proteína a nível
mitocondrial (KRUFT et al., 2001).
Leveduras podem crescer tanto em condições anaeróbicas como aeróbicas
sendo, portanto, expostas a espécies reativas de oxigênio geradas como subprodutos
do metabolismo celular. A principal fonte de espécies reativas de oxigênio é a cadeia
respiratória mitocondrial, que é responsável por 85 a 90% do oxigênio consumido nas
células. Em condições fisiológicas normais, os danos celulares são evitados por
mecanismos de defesa que neutralizam as espécies reativas de oxigênio. Entretanto,
em condições específicas de estresse, os níveis de ROS e a capacidade antioxidante
das células são desequilibrados, podendo resultar na diminuição dos antioxidantes
e/ou aumento na produção de ROS. Neste contexto, o acúmulo de moléculas
oxidadas são associadas à morte celular (COSTA, MORADAS-FERREIRA, 2001).
Células de levedura podem detectar o estresse oxidativo, sendo capazes de
construir uma resposta a nível molecular envolvendo a indução de antioxidantes
primários e secundários. A levedura Saccharomyces cerevisiae é um sistema
biológico ideal para ser estudado como modelo na avaliação de vias de desintoxicação
celular, devido à facilidade de manipulação bioquímica e genética. De fato, grande
número de genes que medeiam a resistência a xenobióticos, bem como vias

44
metabólicas, foram identificados em células de leveduras (COSTA, MORADAS-
FERREIRA, 2001; CASTRO et al., 2007; SUBHASWARAJ et al., 2017).
Glutationas tranferases (GST) compreende uma família de isoenzimas
metabólicas de fase II que desempenham um papel crucial na proteção de células
contra xenobióticos, catalisando a conjugação da forma reduzida de glutationa à
substratos xenobióticos. Nos mamíferos as GST estão amplamente distribuídas
dentro das células e classificadas como citosólica, mitocondriais e associada à
membrana proteínas.
Similarmente como nos mamíferos, nas células de S. cerevisiae duas
glutationas transferases (Gtt1 e Gtt2) já foram identificadas (CASTRO et al., 2008).
Mais recentemente microrganismos como leveduras têm sido empregadas como
modelos biológicos em ensaios para a avalição dos efeitos tóxicos de diferentes
substâncias com potencial de promoção de estresse oxidativo. Em especial leveduras
mutantes do gênero S. cerevisiae, deficientes em enzimas envolvidas em mecanismos
antioxidantes endógenos são empregadas em tais estudos (SUBHASWARAJ et al.,
2017; CHEIGNON et al., 2018).
As células de levedura podem crescer na presença e ausência de oxigênio e
esse recurso pode ser utilizado para avaliar o papel do ciclo redox na toxicidade de
determinada substância (RODRIGUEZ et al., 2004). KERDSOMBOON et al. (2016)
avaliaram a ação protetora de um extrato solúvel de folha de Moringa oleifera contra
a toxicidade do cádmio, utilizando como modelo biológico cepas de S. cerevisiae. Os
autores verificaram que este extrato exibiu um efeito protetor contra o estresse
oxidativo induzido por cádmio e H2O2 através da redução de espécies reativas
intracelulares de oxigênio.
BAYLIAK e LUSHCHAK (2011), estudaram o efeito do extrato aquoso da raiz
de Rhodiola rosea sobre o tempo de vida e atividade de enzimas antioxidantes na
levedura S. cerevisiae. Tais autores verificaram que a suplementação do meio de
crescimento com extrato de R. rosea diminuiu a sobrevivência de células na fase
exponencial sob estresse oxidativo induzido por H2O2, mas aumentou a viabilidade e
o sucesso reprodutivo das células na fase estacionária de crescimento. Dessa forma,
os resultados encontrados sugeriram que R. rosea atua como um estressor para as
células de S. cerevisiae, o que sensibiliza as células de levedura ao estresse oxidativo
na fase exponencial, mas induz a adaptação em células em fase estacionária,

45
demonstrando o efeito positivo na sobrevivência das leveduras sem ativação das
principais enzimas antioxidantes.
46
4 MATERIAIS E MÉTODOS
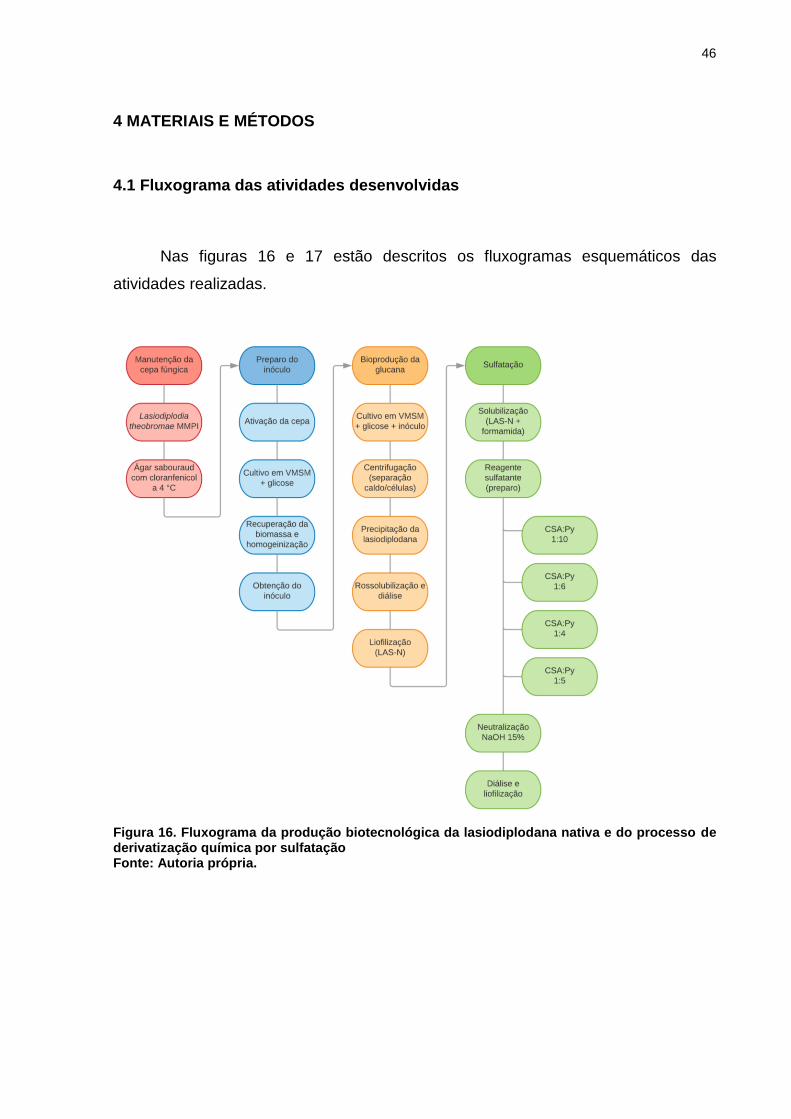
4.1 Fluxograma das atividades desenvolvidas
Nas figuras 16 e 17 estão descritos os fluxogramas esquemáticos das
atividades realizadas.
Figura 16. Fluxograma da produção biotecnológica da lasiodiplodana nativa e do processo de derivatização química por sulfatação Fonte: Autoria própria.

47
Figura 17. Fluxograma das caracterizações estruturais e biológicas realizadas nas amostras. Fonte: Autoria própria.
4.2 Produção do exopolissacarídeo lasiodiplodana e derivatização química
4.2.1 Caracterização macro e micro-morfológica da cepa Lasiodiplodia theobromae
MMPI
Neste trabalho foi utilizado o fungo ascomiceto Lasiodiplodia theobromae MMPI
isolado de fruta do conde (Annona squamosa). O fungo foi conservado em meio ágar
sabouraud com cloranfenicol a 4 °C por repiques periódicos.
Para avaliação morfológica da cultura fúngica, foram conduzidos cultivos em
placas de Petri em meio PDA com cloranfenicol (cultivo superficial) e também
microcultivo em lâmina usando meio PDA com cloranfenicol.
No cultivo superficial, uma porção de micélio (cultura estoque) do fungo foi
repicada para placas de Petri contendo meio PDA cloranfenicol, as quais foram
incubadas em estufa bacteriológica à 28 °C por 4 dias para ativação da cultura. Na
sequência, novo repique foi feito para meio PDA cloranfenicol e então as placas foram
incubadas por 7 dias à 28 °C.
No microcultivo, duas lâminas esterilizadas foram sobrepostas em forma de
cruz e dispostas em placa de Petri contendo um chumaço de algodão umidificado para
manutenção da umidade (Figura 18). O fungo foi inoculado em uma gota de ágar PDA
cloranfenicol sobre a lâmina, a qual foi coberta com uma lamínula, e então a placa foi

48
fechada assepticamente e incubada à 28 °C por 7 dias. Após este período, a lamínula
foi cuidadosamente retirada com auxílio de uma pinça, sendo então gotejado corante
azul de metileno 1% e examinada em microscópio ótico modelo (Bioval, Brasil).
Figura 18. Sistema de microcultivo fúngico em lâmina. Fonte: Autoria própria.
4.2.2 Preparo do inóculo para produção de lasiodiplodana
Uma porção de micélio fúngico foi transferido assepticamente para placas de
Petri contendo ágar sabouraud com cloranfenicol. As placas foram incubadas a 28 °C
por 72 horas em estufa bacteriológica. Em seguida, o fungo foi repicado para frascos
Erlenmeyer contendo 100 mL de Meio de Sais Minerais de Vogel – VMSM (VOGUEL,
1956) e 5 g L-1 de glicose e incubados a 28 °C sob agitação de 150 rpm por 48 horas.
Esta pré-cultura foi centrifugada para recuperação do micélio fúngico e então, os
pellets de biomassa foram homogeneizados e ressuspendidos em solução salina
0,9% para obtenção de um inóculo padronizado com leitura de absorção entre 0,4 e
0,5 a 400 nm (CUNHA et al., 2012).
4.2.3 Produção da lasiodiplodana em cultivo submerso
Lasiodiplodana foi produzida em frascos Erlenmeyer de 250 mL por cultivo
submerso conduzido em agitador orbital de bancada (shaker). O volume de trabalho
utilizado foi de 150 mL, sendo constituído por meio de fermentação (135 mL) à base
de sais minerais de Vogel (VOGUEL, 1956), 20 g L-1 de glicose e 15 mL de inóculo

49
padronizado. Os frascos foram incubados sob agitação de 150 rpm, a 28 °C por 72
horas (CUNHA et al., 2012).
Após 72 horas o meio de cultivo foi separado da biomassa celular por
centrifugação (1500 x g, 15 min.), e então a lasiodiplodana foi precipitada do caldo
com 3 volumes de etanol resfriado a 4 °C (overnight). A lasiodiplodana precipitada foi
ressolubilizada em água destilada (60 °C), e então submetida a intensa diálise contra
água destilada por aproximadamente cinco dias (com três trocas diárias de água),
como demonstrado na figura 19. Por fim, o material dialisado foi desidratado por
liofilização, nomeado como LAS-N e armazenado adequadamente para posteriores
análises de caracterização e sulfatação.
Figura 19. Representação esquemática do processo de produção e recuperação da lasiodiplodana nativa (LAS-N) Fonte: Autoria própria.
4.2.4 Derivatização química da lasiodiplodana por sulfatação
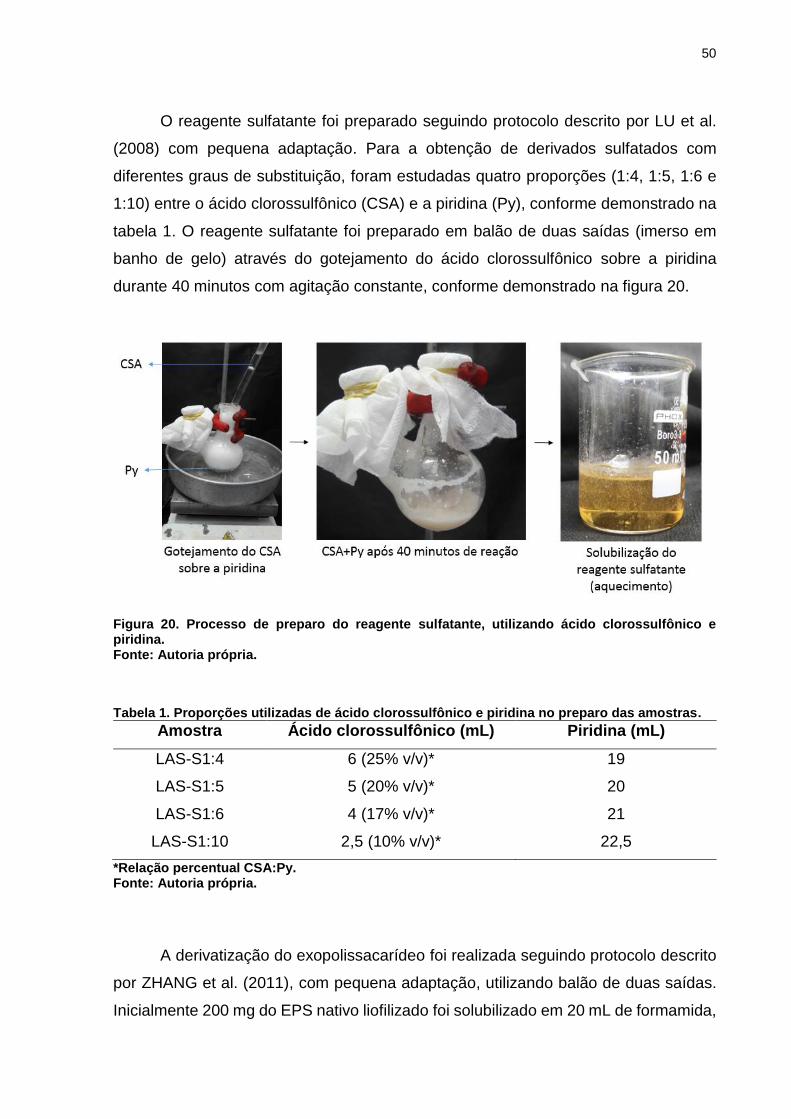
50
O reagente sulfatante foi preparado seguindo protocolo descrito por LU et al.
(2008) com pequena adaptação. Para a obtenção de derivados sulfatados com
diferentes graus de substituição, foram estudadas quatro proporções (1:4, 1:5, 1:6 e
1:10) entre o ácido clorossulfônico (CSA) e a piridina (Py), conforme demonstrado na
tabela 1. O reagente sulfatante foi preparado em balão de duas saídas (imerso em
banho de gelo) através do gotejamento do ácido clorossulfônico sobre a piridina
durante 40 minutos com agitação constante, conforme demonstrado na figura 20.
Figura 20. Processo de preparo do reagente sulfatante, utilizando ácido clorossulfônico e piridina. Fonte: Autoria própria.
Tabela 1. Proporções utilizadas de ácido clorossulfônico e piridina no preparo das amostras.
Amostra Ácido clorossulfônico (mL) Piridina (mL)
LAS-S1:4 6 (25% v/v)* 19
LAS-S1:5 5 (20% v/v)* 20
LAS-S1:6 4 (17% v/v)* 21
LAS-S1:10 2,5 (10% v/v)* 22,5
*Relação percentual CSA:Py. Fonte: Autoria própria.
A derivatização do exopolissacarídeo foi realizada seguindo protocolo descrito
por ZHANG et al. (2011), com pequena adaptação, utilizando balão de duas saídas.
Inicialmente 200 mg do EPS nativo liofilizado foi solubilizado em 20 mL de formamida,

51
sob intensa agitação (agitador magnético) por 72 horas à temperatura ambiente. Em
seguida, foram adicionados, gota-a-gota, 20 mL do reagente sulfatante previamente
preparado, e então, a solução permaneceu sob agitação por 3 horas à temperatura
de 60 °C. A sulfatação foi interrompida pela neutralização do meio (pH entre 7 e 8)
com adição de solução NaOH (15% m v-1). A solução resultante foi extensivamente
dialisada contra água destilada, por seis dias, com várias trocas diárias de água
(Figura 21). Por fim, o produto dialisado foi submetido a liofilização e nomeado como
LAS-S.
Figura 21. Processo de sulfatação da lasiodiplodana. Fonte: Autoria própria.
4.3 Caracterização química da lasiodiplodana nativa e sulfatada
4.3.1 Determinação do grau de sulfatação (DS)
O grau de substituição por sulfatação da macromolécula foi determinado com
base na correlação entre os conteúdos de enxofre e carbono da amostra seguindo
protocolo descrito por DE MOURA NETO et al. (2011). O grau de sulfatação é definido

52
como o número médio de grupos sulfatos (O-SO3H) inseridos em cada unidade
monossacarídica da macromolécula lasiodiplodana. Os conteúdos de enxofre e
carbono presentes nas amostras de lasiodiplodana foram determinados em analisador
elementar CHNS/O 2400 série II (Perkin Elmer, USA), e o grau de substituição foi
estimado de acordo com equação 2.
𝑫𝑺 = (𝑺%
𝒎𝒂𝒔𝒔𝒂 𝒂𝒕ô𝒎𝒊𝒄𝒂 𝒅𝒐 𝑺⁄ )
(𝑪%𝒎𝒂𝒔𝒔𝒂 𝒂𝒕ô𝒎𝒊𝒄𝒂 𝒅𝒐 𝑪 𝒙 𝟔⁄ )
∴ 𝑫𝑺 = 𝟐, 𝟐𝟓 (𝑺%
𝑪%), (Equação 2)
Onde, S% e C% corresponde aos conteúdos percentuais de enxofre e de carbono
presentes na amostra, respectivamente. O número 6 corresponde ao número de
átomos de carbono por unidade monomérica.
4.3.2 Avaliação da solubilidade
Para avaliação da solubilidade em água, foi empregado o protocolo descrito por
WANG et al. (2012), com adaptações. As amostras de lasiodiplodana (10 mg) foram
suspensas em água destilada (10 mL) e agitadas por 24 horas à 25 °C. Em seguida,
foram centrifugadas a 1500 x g por 10 minutos. O sobrenadante então, foi coletado e
utilizado para quantificação de açúcares totais, sendo estes, diretamente relacionados
com a quantidade de amostra solúvel. A solubilidade em água foi expressa em
porcentagem de massa solúvel de EPS. A solubilidade em solução de DMSO 20%,
também foi avaliada seguindo protocolo similar ao empregado na avaliação da
hidrossolubilidade, apenas substituindo a água por solução de DMSO 20%
(GALEANE et al., 2017). A solubilidade em DMSO 20% foi realizado visando posterior
utilização nos ensaios de atividade biológica.
4.3.3 Espectroscopia no infravermelho com transformada de Fourier (IV-TF)
Os espectros de infravermelho das amostras de LAS-N e LAS-S foram obtidos
em espectrômetro Frontier (Perkin Elmer®, USA) na faixa espectral compreendida
entre 400 e 4000 cm-1, com resolução de 2 cm-1, número de acumulações de 16
varreduras para cada espectro, utilizando a técnica de disco de KBr.

53
4.3.4 Microscopia eletrônica de varredura (MEV)
As micrografias foram obtidas em microscópio eletrônico de varredura de
bancada TM3000 (Hitachi, USA) a partir das amostras liofilizadas de LAS-N e LAS-S.
Imagens com amplitudes de 300, 800 e 1200 x, foram adquiridas das amostras fixadas
em fita de carbono.
4.3.5 Análise Térmica
Amostras liofilizadas de LAS-N e LAS-S foram submetidas à Análise de
Termogravimetria Derivada (DTG), Análise Térmica Diferencial (DTA) e Análise
Termogravimétrica (TGA), realizadas em equipamento SDT Q600 TA (TA Instruments,
USA). A perda de massa foi acompanhada entre 25 °C e 800 °C, com taxa de
aquecimento de 10 °C min-1, em atmosfera de ar sintético com fluxo de 50 mL min-1.
4.3.6 Difração de raios-X (DRX)
Para obtenção dos difratogramas das amostras foi utilizado difratômetro
MiniFlex600 (Rigaku, USA), usando fonte de radiação de lâmpada de cobre
(CuKα=1,5418 Å), corrente de 15 mA, 40 kV de voltagem, faixa de varredura de 10° a
60° (2θ) em modo step scan ao passo de 0,05°/2 segundos.
4.4 Caracterização biológica da lasiodiplodana nativa e sulfatada
4.4.1 Determinação do potencial antimicrobiano
O potencial antimicrobiano foi determinado conforme procedimento descrito por
KRICHEN et al., (2015). Os ensaios de atividade antimicrobiana foram conduzidos em
placas de Elisa com 96 poços. Em cada cavidade foram adicionados 100 µL da
amostra (LAS-N e LAS-S) nas concentrações de 0,26; 0,20; 0,15; 0,10 e 0,05 mg mL-
1 juntamente com 100 µL de caldo Mueller Hinton (bactérias) ou caldo Sabouraud com
cloranfenicol (leveduras) e 5 µL da suspensão microbiana previamente padronizada

54
em escala de McFarland 0,5 (1,5x108 UFC mL-1). Nestes ensaios, foram testadas
quatro cepas bacterianas (Escherichia coli ATCC 25922, Listeria monocytogenes
ATCC 19111, Staphylococcus aureus ATCC 25923 e Salmonella enterica
Typhimurium ATCC 0028) e duas cepas de leveduras (Candida albicans ATCC 10231
e Candida tropicalis 13803). As placas foram incubadas por 24 horas à 28 °C e 37 °C,
para leveduras e bactérias, respectivamente. Após incubação, foram adicionados em
cada cavidade, 20 µL do corante-reagente resazurina (0,01%) para verificação da
presença de células viáveis. Células viáveis são indicadas pelo aparecimento de
coloração rósea pela formação do composto intermediário rizorufina, oriundo da
reação de oxi-redução do corante resazurina (azul). Ou seja, o desenvolvimento de
coloração rósea no meio indica presença de células viáveis e a ausência indica
inibição celular. Após duas horas em repouso, a primeira leitura foi realizada, na qual
foi verificada a inibição dos microrganismos testados. O material dos poços que
apresentaram resultados positivos, ou seja, inibição dos microrganismos
(permanência da coloração azul do meio), foram repicados para placas de Petri
contendo ágar BHI (bactérias) ou ágar sabouraud com cloranfenicol (leveduras), as
quais foram incubadas em estufa bacteriológica à 28 °C (leveduras) ou 37 °C
(bactérias) por 24 horas, para verificação da atividade bactericida ou fungicida da
amostra. Como controles positivos, foram utilizados os antimicrobianos tetraciclina e
fluconazol para as bactérias e leveduras, respectivamente. Água peptonada 0,1%
esterilizada foi utilizada como controle negativo.
4.4.2 Atividade antioxidante
O potencial antioxidante das amostras foi avaliado por diferentes protocolos in
vitro, sendo analisado o poder de redução do íon férrico pelo método FRAP, a
capacidade de sequestro do radical OH e a capacidade de eliminação dos radicais
DPPH e ABTS.
4.4.2.1 Poder de redução do íon férrico (FRAP)
Esta análise foi baseada nos protocolos descritos por RUFINO et al., (2007) e
WOOTTON-BEARD, MORAN e RYAN (2011). No preparo do reagente FRAP foram
misturados 25 mL de solução tampão acetato de sódio (300 mmol L-1) pH 3,6, 2,5 mL
de solução TPTZ (10 mmol L-1) dissolvido em HCl 40 mmol L-1 e 2,5 mL de solução

55
de cloreto férrico (20 mmol L-1). O reagente foi armazenado por 15 minutos a 37 °C
antes do uso. Para determinação do potencial antioxidante FRAP, 90 µL da amostra
nas concentrações de 0,10, 0,18 e 0,26 mg mL-1 foram misturados com 270 µL de
água destilada e 2,7 mL de solução FRAP e esta solução foi armazenada na ausência
de luz por 30 minutos a 37 °C e então procedida a leitura a 595 nm. Solução FRAP foi
utilizada como branco. As análises foram realizadas em triplicata e os resultados
expressos em µmol FeSO4.7H2O através de correlação com curva de calibração de
sulfato ferroso (0,2 - 2,0 mmol L-1).
4.4.2.2 Sequestro do radical hidroxila
O potencial de sequestro de OH• foi avaliado com base no protocolo descrito
por LIU et al., (2010). Mistura reacional (2 mL) contendo 0,5 mL de FeSO4 (1,5 mmol
L-1), 0,35 mL de H2O2 (6 mmol L-1), 0,15 mL de salicilato de sódio (20 mmol L-1) e 1
mL da amostra em diferentes concentrações (0,10, 0,18 e 0,26 mg mL-1) foi obtida.
Ácido ascórbico foi utilizado como controle positivo. Após incubação por 1 hora a 37
°C as leituras de absorbância foram realizadas em espectrofotômetro a 562 nm. As
análises foram realizadas em triplicata e o percentual de sequestro do radical hidroxila
foi determinado, segundo equação 3.
% 𝒅𝒆 𝑹𝒆𝒎𝒐çã𝒐 𝒅𝒆 𝑶𝑯 = [𝟏 −(𝑨𝟏−𝑨𝟐)
𝑨𝟎] 𝒙𝟏𝟎𝟎% (Equação 3)
Onde, A0 corresponde a absorbância do controle, A1 a absorbância da amostra ou
ácido ascórbico e A2 a absorbância do branco com salicilato de sódio.
4.4.2.3 Captura do radical DPPH
O potencial de captura do radical DPPH foi analisado seguindo metodologia
descrita por LOCATELLI et al., (2009), com adaptações. Em tubo de ensaio foram
misturados 500 µL da amostra (Aamostra) nas concentrações de 0,10, 0,18 ou 0,26 mg
mL-1, 3 mL de etanol P.A e 300 µL de solução etanólica de DPPH (0,5 mmol L-1). As
leituras espectrofotométricas foram obtidas em 517 nm após 80 minutos de reação. O
aparelho foi zerado com solução etanólica 80% e como controle (Acontrole) foi utilizado
a mesma mistura reacional apenas sendo substituído o volume da amostra por igual

56
volume de solução etanólica 80%. A porcentagem de atividade de captura do radical
hidroxila foi estimada segundo equação 4.
% 𝑨𝑨 = 𝟏𝟎𝟎 −(𝑨𝒂𝒎𝒐𝒔𝒕𝒓𝒂𝒙𝟏𝟎𝟎)
𝑨𝒄𝒐𝒏𝒕𝒓𝒐𝒍𝒆 (Equação 4)
4.4.2.4 Captura do radical cátion ABTS
O potencial de captura do radical cátion ABTS foi determinado seguindo
metodologia descrita por RUFINO et al., (2007) e WOOTON-BEARD, MORAN e
RYAN (2007) com adaptações. Radical ABTS foi obtido pela mistura de 5 mL de
solução de ABTS (7 mmol L-1) e 88 µL da solução de persulfato de potássio (140 mmol
L-1) em tubo de ensaio, o qual foi mantido ao abrigo de luz em temperatura ambiente
durante 16 horas. Em seguida, foram diluídos 1 mL da solução ABTS em álcool etílico
até obtenção de absorbância de 0,7 em 734 nm. Em tubos de ensaio foram misturados
30 µL de amostra nas concentrações de 0,10, 0,18 e 0,26 mg mL-1 e 3 mL da solução
de ABTS diluída em etanol. Os tubos foram agitados e deixados ao abrigo luz por 6
minutos até leitura de absorbância em 734 nm. Como branco foi utilizado solução de
DMSO 20%. As análises foram realizadas em triplicata e os resultados expressos em
mmol L-1 de Trolox equivalente mL-1 pela correlação com curva padrão de Trolox (0,1,
0,25, 0,5, 1,0, 1,5 e 2,0 mmol L-1).
4.4.3 Citotoxicidade sobre células de Saccharomyces cerevisiae
O potencial de citotoxicidade da lasiodiplodana nativa e sulfatada foi avaliada
em relação a capacidade de indução ao estresse oxidativo, em cepas de
Saccharomyces cerevisiae. Foram utilizadas as linhagens Saccharomyces cerevisiae
f.r bayanus (Fermol Perlage, AEB Biochemistry Latin American AS, Brazil) e leveduras
mutantes Saccharomyces cerevisiae (YIR038C∆GTT1, YLL060C∆GTT2 e
YSL101C∆GSH1).
As linhagens de S. cerevisiae YIR038C, YLL060C e YSL101C são mutantes
cujos genes GTT1 (Glutationa Transferase 1), GTT2 (Glutationa Transferase 2) e
GSH1 (Glutamato Cisteína Ligase) foram respectivamente interrompidos pelo gene
KanMX4 (Euroscarf, Frankfurt, Alemanha) e apresentam deficiência na enzima

57
Glutationa (GSH/GSSG). As cepas de levedura foram mantidas em meio YPD à 4 °C.
As cepas foram reativadas em caldo nutriente e repicadas para ágar YPD antes dos
ensaios.
O potencial de estresse oxidativo pelas amostras de lasiodiplodana nativa e
sulfatada foi avaliado com base nos protocolos descritos por CASTRO et al., (2008),
e SUBHASWARAJ et al., (2017) com adaptações. Os ensaios foram conduzidos em
placas de Elisa de 96 poços. Em cada cavidade foram adicionados 100 µL da amostra
de lasiodiplodana nativa ou sulfatada nas concentrações de 0,10; 0,20 ou 0,26 mg mL-
1, juntamente com 100 µL de caldo YPD e 20 µL da suspensão microbiana com
concentração padronizada em escala de McFarland 0,5 (1,5x108 UFC mL-1). As placas
foram incubadas por 24 horas a 28 °C; após incubação foram adicionados em cada
cavidade 20 µL do corante-reagente resazurina (0,01%) e deixadas em repouso por 2
horas para verificação da presença de células viáveis (coloração rósea).
O aparecimento de coloração azul nos poços das placas de Elisa indicavam
inatividade celular. Os meios de tais poços foram repicados para placas de Petri
contendo ágar YPD, as quais foram incubadas em estufa bacteriológica à 28 °C, para
comprovação da ausência de viabilidade celular (morte celular) ou apenas efeito
microbiostático das amostras sobre as leveduras avaliadas. Como controle positivo foi
utilizado menadiona, e água peptonada 0,1% esterilizada foi utilizada como controle
negativo.

58
5 RESULTADOS E DISCUSSÃO
5.1 Aspectos morfológicos da cepa Lasiodiplodia theobromae MMPI
Conforme pode ser verificado nas figuras 22 A e 23A, após 7 dias de incubação
em meio PDA foi observado intenso crescimento micelial, tanto crescimento radial
quanto prolongamento aéreo das hifas, sendo toda placa recoberta pelo micélio
fúngico. Após 14 dias de incubação algumas mudanças macroscópicas podem ser
observadas (Figura 22 B) na cultura, incluindo redução do comprimento do micélio
aéreo e escurecimento do micélio vegetativo, destacando a coloração branca
acinzentada do micélio aéreo.
Figura 22. Cultivo do fungo L. theobromae MMPI em ágar PDA: 7 dias de incubação (A) e 14 dias de incubação (B). Fonte: Autoria própria.
O microcultivo do L. theobramae permitiu a visualização morfológica de suas
hifas (Figuras 23 C e D), sendo observados hifas hialinas septadas com poucas
ramificações. Não foram observados esporos fúngicos em tais condições de cultivo,
possivelmente em função de condições adequadas e não estressoras de cultivo.
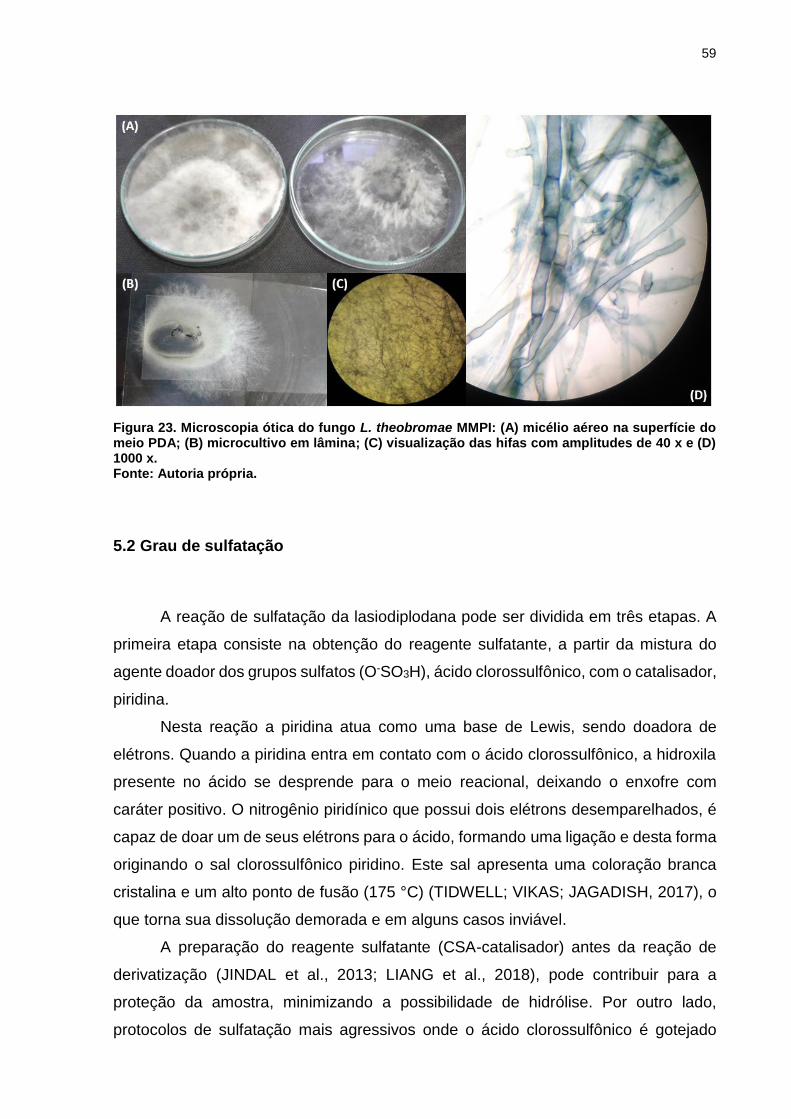
59
Figura 23. Microscopia ótica do fungo L. theobromae MMPI: (A) micélio aéreo na superfície do meio PDA; (B) microcultivo em lâmina; (C) visualização das hifas com amplitudes de 40 x e (D) 1000 x. Fonte: Autoria própria.
5.2 Grau de sulfatação
A reação de sulfatação da lasiodiplodana pode ser dividida em três etapas. A
primeira etapa consiste na obtenção do reagente sulfatante, a partir da mistura do
agente doador dos grupos sulfatos (O-SO3H), ácido clorossulfônico, com o catalisador,
piridina.
Nesta reação a piridina atua como uma base de Lewis, sendo doadora de
elétrons. Quando a piridina entra em contato com o ácido clorossulfônico, a hidroxila
presente no ácido se desprende para o meio reacional, deixando o enxofre com
caráter positivo. O nitrogênio piridínico que possui dois elétrons desemparelhados, é
capaz de doar um de seus elétrons para o ácido, formando uma ligação e desta forma
originando o sal clorossulfônico piridino. Este sal apresenta uma coloração branca
cristalina e um alto ponto de fusão (175 °C) (TIDWELL; VIKAS; JAGADISH, 2017), o
que torna sua dissolução demorada e em alguns casos inviável.
A preparação do reagente sulfatante (CSA-catalisador) antes da reação de
derivatização (JINDAL et al., 2013; LIANG et al., 2018), pode contribuir para a
proteção da amostra, minimizando a possibilidade de hidrólise. Por outro lado,
protocolos de sulfatação mais agressivos onde o ácido clorossulfônico é gotejado

60
diretamente sobre amostra solubilizada e contendo o catalisador (MENDES et al.,
2009), pode levar a hidrólise parcial da amostra.
O segundo passo da sulfatação corresponde a reação entre o sal
clorossulfônico piridino e a amostra (lasiodiplodana nativa). Nesta etapa reacional
ocorre a retirada do hidrogênio de uma das hidroxilas da lasiodiplodana nativa e a
inserção do grupo sulfônico na macromolécula, o qual é oriundo do sal clorossulfônico
piridino, previamente formado. Paralelamente o catalisador piridina é liberado para o
meio reacional.
Na terceira e última etapa a mistura reacional é neutralizada com NaOH,
ocorrendo a formação do cloreto de sódio, ligação da hidroxila do álcali à
macromolécula e geração do produto sulfatado estável, como descrito na figura 26.
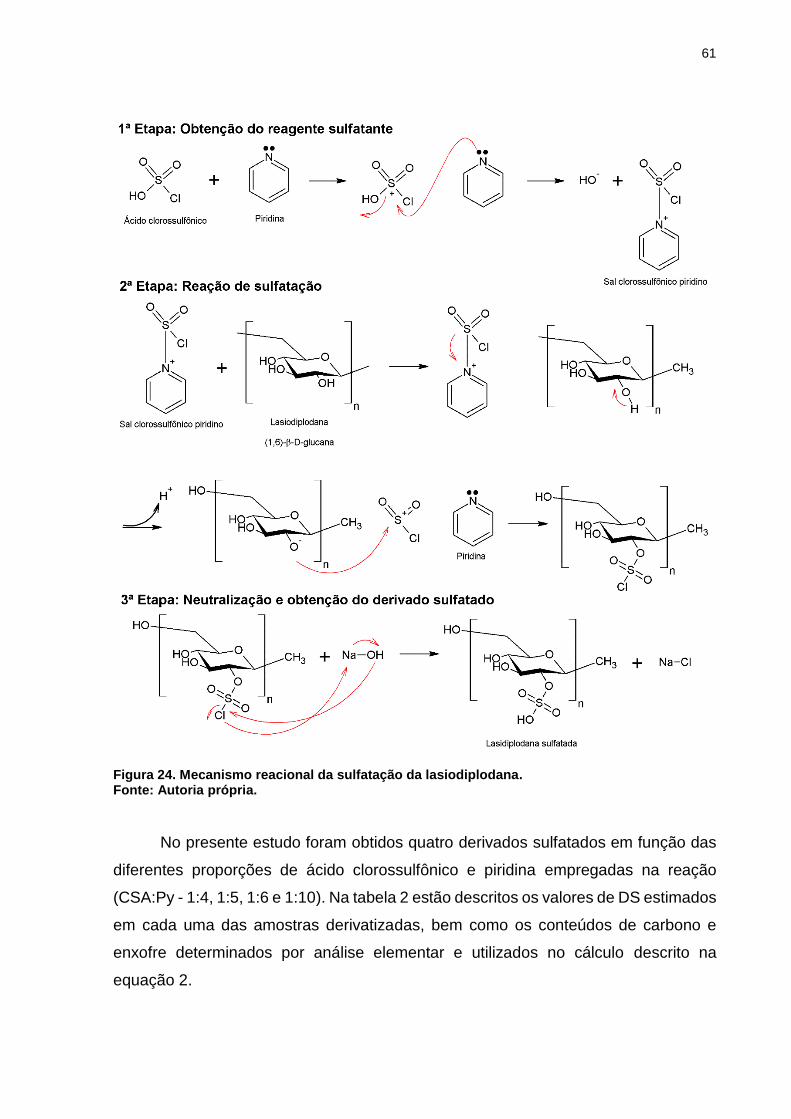
61
Figura 24. Mecanismo reacional da sulfatação da lasiodiplodana. Fonte: Autoria própria.
No presente estudo foram obtidos quatro derivados sulfatados em função das
diferentes proporções de ácido clorossulfônico e piridina empregadas na reação
(CSA:Py - 1:4, 1:5, 1:6 e 1:10). Na tabela 2 estão descritos os valores de DS estimados
em cada uma das amostras derivatizadas, bem como os conteúdos de carbono e
enxofre determinados por análise elementar e utilizados no cálculo descrito na
equação 2.
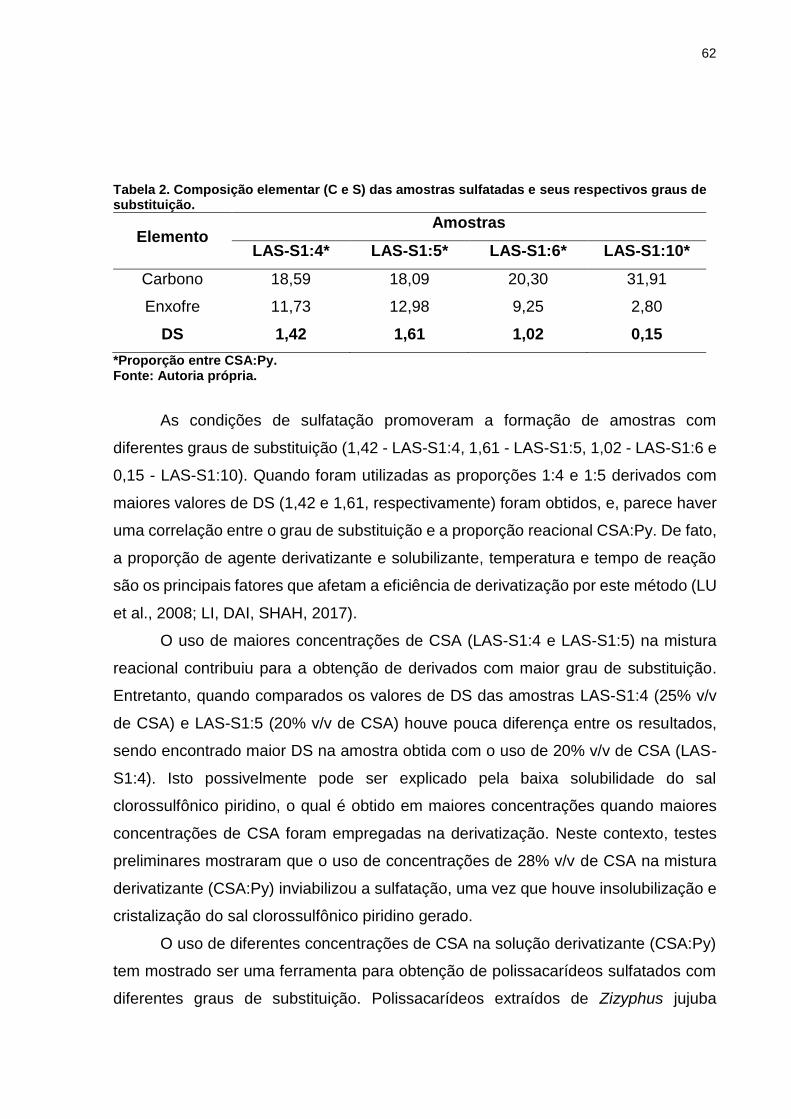
62
Tabela 2. Composição elementar (C e S) das amostras sulfatadas e seus respectivos graus de substituição.
Elemento Amostras
LAS-S1:4* LAS-S1:5* LAS-S1:6* LAS-S1:10*
Carbono 18,59 18,09 20,30 31,91
Enxofre 11,73 12,98 9,25 2,80
DS 1,42 1,61 1,02 0,15
*Proporção entre CSA:Py. Fonte: Autoria própria.
As condições de sulfatação promoveram a formação de amostras com
diferentes graus de substituição (1,42 - LAS-S1:4, 1,61 - LAS-S1:5, 1,02 - LAS-S1:6 e
0,15 - LAS-S1:10). Quando foram utilizadas as proporções 1:4 e 1:5 derivados com
maiores valores de DS (1,42 e 1,61, respectivamente) foram obtidos, e, parece haver
uma correlação entre o grau de substituição e a proporção reacional CSA:Py. De fato,
a proporção de agente derivatizante e solubilizante, temperatura e tempo de reação
são os principais fatores que afetam a eficiência de derivatização por este método (LU
et al., 2008; LI, DAI, SHAH, 2017).
O uso de maiores concentrações de CSA (LAS-S1:4 e LAS-S1:5) na mistura
reacional contribuiu para a obtenção de derivados com maior grau de substituição.
Entretanto, quando comparados os valores de DS das amostras LAS-S1:4 (25% v/v
de CSA) e LAS-S1:5 (20% v/v de CSA) houve pouca diferença entre os resultados,
sendo encontrado maior DS na amostra obtida com o uso de 20% v/v de CSA (LAS-
S1:4). Isto possivelmente pode ser explicado pela baixa solubilidade do sal
clorossulfônico piridino, o qual é obtido em maiores concentrações quando maiores
concentrações de CSA foram empregadas na derivatização. Neste contexto, testes
preliminares mostraram que o uso de concentrações de 28% v/v de CSA na mistura
derivatizante (CSA:Py) inviabilizou a sulfatação, uma vez que houve insolubilização e
cristalização do sal clorossulfônico piridino gerado.
O uso de diferentes concentrações de CSA na solução derivatizante (CSA:Py)
tem mostrado ser uma ferramenta para obtenção de polissacarídeos sulfatados com
diferentes graus de substituição. Polissacarídeos extraídos de Zizyphus jujuba

63
sulfatados com o uso de solução CSA:Py nas proporções de 1:2, 1:4, 1:6 e 1:8
originaram derivados sulfatados com DS variando de 0,36 a 2,30 (CAI et al., 2018).
Similarmente, XIE et al. (2016) derivatizaram por sulfatação polissacarídeo
extraído de Cyclocarya paliurus e verificaram que o aumento da proporção utilizada
de ácido clorossulfônico-piridina (1:1, 1:4, 1:6 e 1:8) possibilitou a obtenção de
derivados com maior DS (0,52, 0,33, 0,20 e 0,06).
O grau de substituição das amostras de lasiodiplodana foi estimado,
considerando que cada unidade monossacarídica da macromolécula possui três
hidroxilas livres, ou seja, é possível a obtenção de um DS máximo de três (DS= 3).
Foram obtidas eficiências de 47,33% (LAS-S1:4, DS: 1,42), 53,67% (LAS-S1:5,
DS:1,61), 34% (LAS-S1:6, DS: 1,02) e 5% (LAS-S1:10, DS:0,15).
Polissacarídeos sulfatados com DS superior a 0,8 têm sido destacados na
literatura científica como macromoléculas bioativas (VASCONCELOS; et al., 2013;
(ZHANG et al., 2010; LIANG et al., 2018). Neste sentido, é importante o
desenvolvimento de protocolos de sulfatação eficientes que promovam a obtenção de
derivados com elevados valores de DS.
5.3 Solubilidade dos derivados obtidos
Modificações estruturais em polímeros podem promover diversas alterações
em suas propriedades químicas, físico-químicas e biológicas. A inserção de
heteroátomos ou grupamentos químicos na estrutura dos polissacarídeos pode levar
a mudanças nas interações com a água ou outros solventes, podendo aumentar ou
diminuir sua solubilidade.
Neste trabalho, foram avaliadas a solubilidade da lasiodiplodana nativa e
sulfatada em água e DMSO 20% em temperatura ambiente e os resultados estão
descritos na tabela 3.
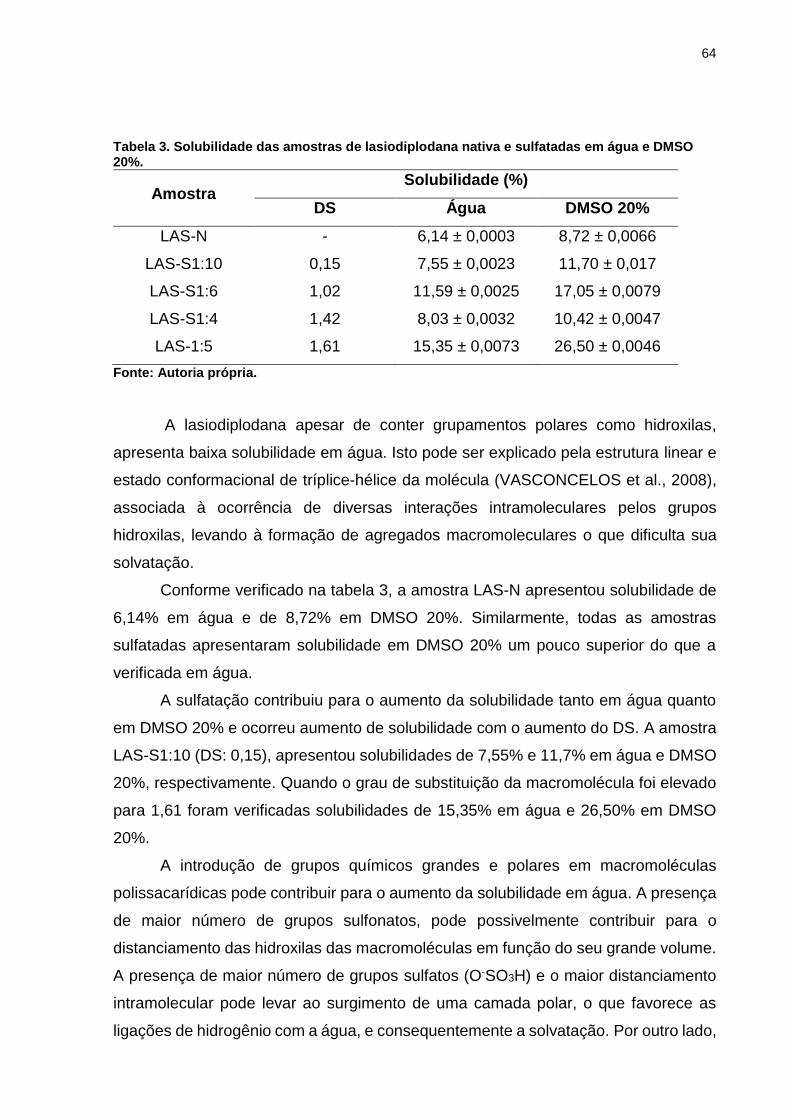
64
Tabela 3. Solubilidade das amostras de lasiodiplodana nativa e sulfatadas em água e DMSO 20%.
Amostra Solubilidade (%)
DS Água DMSO 20%
LAS-N - 6,14 ± 0,0003 8,72 ± 0,0066
LAS-S1:10 0,15 7,55 ± 0,0023 11,70 ± 0,017
LAS-S1:6 1,02 11,59 ± 0,0025 17,05 ± 0,0079
LAS-S1:4 1,42 8,03 ± 0,0032 10,42 ± 0,0047
LAS-1:5 1,61 15,35 ± 0,0073 26,50 ± 0,0046
Fonte: Autoria própria.
A lasiodiplodana apesar de conter grupamentos polares como hidroxilas,
apresenta baixa solubilidade em água. Isto pode ser explicado pela estrutura linear e
estado conformacional de tríplice-hélice da molécula (VASCONCELOS et al., 2008),
associada à ocorrência de diversas interações intramoleculares pelos grupos
hidroxilas, levando à formação de agregados macromoleculares o que dificulta sua
solvatação.
Conforme verificado na tabela 3, a amostra LAS-N apresentou solubilidade de
6,14% em água e de 8,72% em DMSO 20%. Similarmente, todas as amostras
sulfatadas apresentaram solubilidade em DMSO 20% um pouco superior do que a
verificada em água.
A sulfatação contribuiu para o aumento da solubilidade tanto em água quanto
em DMSO 20% e ocorreu aumento de solubilidade com o aumento do DS. A amostra
LAS-S1:10 (DS: 0,15), apresentou solubilidades de 7,55% e 11,7% em água e DMSO
20%, respectivamente. Quando o grau de substituição da macromolécula foi elevado
para 1,61 foram verificadas solubilidades de 15,35% em água e 26,50% em DMSO
20%.
A introdução de grupos químicos grandes e polares em macromoléculas
polissacarídicas pode contribuir para o aumento da solubilidade em água. A presença
de maior número de grupos sulfonatos, pode possivelmente contribuir para o
distanciamento das hidroxilas das macromoléculas em função do seu grande volume.
A presença de maior número de grupos sulfatos (O-SO3H) e o maior distanciamento
intramolecular pode levar ao surgimento de uma camada polar, o que favorece as
ligações de hidrogênio com a água, e consequentemente a solvatação. Por outro lado,

65
a menor solubilidade de derivados com baixo grau de sulfatação comparado com
derivados de maior DS, pode ser explicado pela possibilidade de ligações
intramoleculares pela ocorrência de interações entre o oxigênio do grupo sulfatos e
átomos de hidrogênio das hidroxilas não substituídas do anel de glicose.
5.4 Análises dos espectros de Infravermelho (IV-TF)
A análise de infravermelho permite obter informações fundamentais sobre as
estruturas de moléculas químicas e é uma técnica que baseia-se na absorção da
radiação no espectro do infravermelho pelas moléculas, as quais são excitadas para
atingir um estado de maior energia. No processo de absorção, as frequências de
radiação absorvidas são aquelas que equivalem as frequências vibracionais naturais
da molécula em questão. Esta energia absorvida serve para aumentar a amplitude
dos movimentos vibracionais das ligações na molécula, movimentos estes, que
podem ser de estiramento e dobramento das ligações na maioria das moléculas mais
covalentes (PAVIA et al., 2010).
A figura 25 mostra os espectros de infravermelho (normalizados) das amostras
de lasiodiplodana nativa (LAS-N) e sulfatadas (LAS-S).
Os espectros gerados entre 4000 e 400 cm-1, apresentaram sinais típicos de
polissacarídeos com bandas características de glucanas, como mostra a figura 25. A
banda larga de intensidade forte na região de 3400 cm-1 é atribuída a vibração de
estiramento O-H (PAVIA et al., 2010; BUDDANA, VARANASI, SHETTY, 2015;
KRICHEN et al., 2015), presente nas glucanas nativas e modificadas como hidroxilas
livres.
A banda em aproximadamente 2900 cm-1 é característica de estiramento C-H
sp3 (PAVIA et al., 2010; BUDDANA, VARANASI, SHETTY, 2015), presente em
polissacarídeos como as glucanas, que são constituídas principalmente por ligações
entre carbono e hidrogênio. A banda de média intensidade em 1600 cm-1 é atribuída
à vibração de estiramento do anel de glicose (XU et al., 2009).
Bandas de baixa intensidade entre 1400 e 1200 cm-1 indicam vibrações de
deformação de C-H e O-H, típicas de carboidratos (CALEGARI et al., 2017), as quais
podem apresentar pequenas variações de intensidade e número de onda em função
da natureza do polissacarídeo (CALEGARI et al., 2017). Além disso, há um sinal na
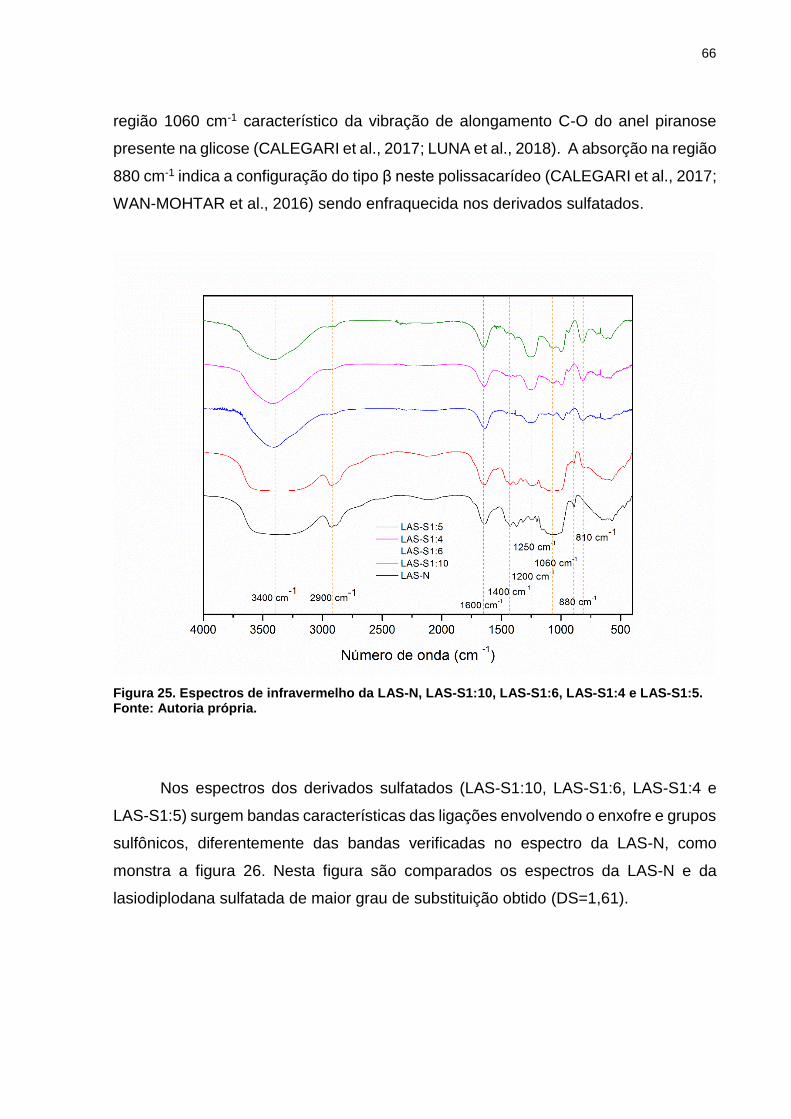
66
região 1060 cm-1 característico da vibração de alongamento C-O do anel piranose
presente na glicose (CALEGARI et al., 2017; LUNA et al., 2018). A absorção na região
880 cm-1 indica a configuração do tipo β neste polissacarídeo (CALEGARI et al., 2017;
WAN-MOHTAR et al., 2016) sendo enfraquecida nos derivados sulfatados.
Figura 25. Espectros de infravermelho da LAS-N, LAS-S1:10, LAS-S1:6, LAS-S1:4 e LAS-S1:5. Fonte: Autoria própria.
Nos espectros dos derivados sulfatados (LAS-S1:10, LAS-S1:6, LAS-S1:4 e
LAS-S1:5) surgem bandas características das ligações envolvendo o enxofre e grupos
sulfônicos, diferentemente das bandas verificadas no espectro da LAS-N, como
monstra a figura 26. Nesta figura são comparados os espectros da LAS-N e da
lasiodiplodana sulfatada de maior grau de substituição obtido (DS=1,61).
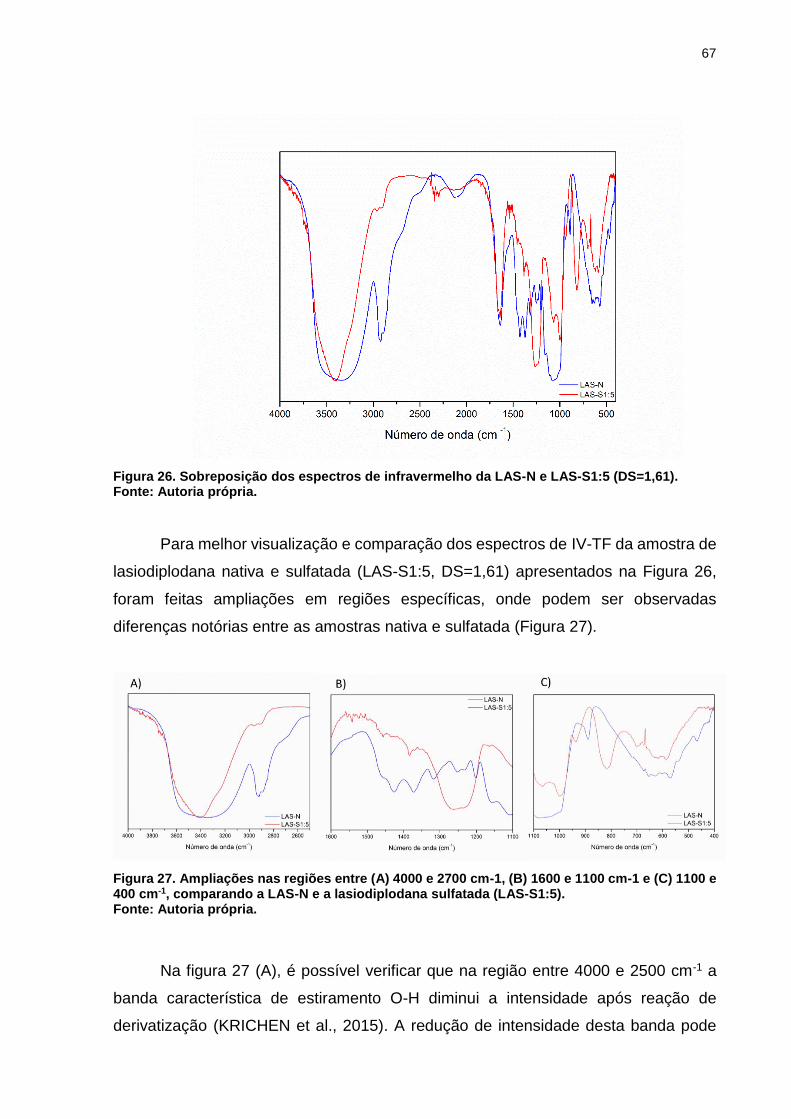
67
Figura 26. Sobreposição dos espectros de infravermelho da LAS-N e LAS-S1:5 (DS=1,61). Fonte: Autoria própria.
Para melhor visualização e comparação dos espectros de IV-TF da amostra de
lasiodiplodana nativa e sulfatada (LAS-S1:5, DS=1,61) apresentados na Figura 26,
foram feitas ampliações em regiões específicas, onde podem ser observadas
diferenças notórias entre as amostras nativa e sulfatada (Figura 27).
Figura 27. Ampliações nas regiões entre (A) 4000 e 2700 cm-1, (B) 1600 e 1100 cm-1 e (C) 1100 e 400 cm-1, comparando a LAS-N e a lasiodiplodana sulfatada (LAS-S1:5). Fonte: Autoria própria.
Na figura 27 (A), é possível verificar que na região entre 4000 e 2500 cm-1 a
banda característica de estiramento O-H diminui a intensidade após reação de
derivatização (KRICHEN et al., 2015). A redução de intensidade desta banda pode

68
ser atribuída a diferenças de absorção na região do infravermelho entre as hidroxilas
constituintes da lasiodiplodana e as hidroxilas presentes no grupamento sulfatos (O-
SO3H).
Após sulfatação há desaparecimento das bandas características de vibrações
de deformação de C-H e O-H (WAN-MOHTAR et al., 2016) verificadas em 1400 cm-1
na amostra LAS-N (Figura 27 B), indicando a ocorrência de uma mudança estrutural
na molécula, causada pela derivatização realizada.
Outro aspecto observado é o surgimento de uma banda na região de 1250 cm- 1
nas amostras sulfatadas (Figuras 25 e 27B), a qual é atribuída a vibração de
alongamento assimétrico (forte) S=O, associado à distribuição de grupos sulfonato na
molécula (ANAND et al., 2018). Na figura 27 também pode ser observado aumento na
intensidade da banda em 1250 cm-1 concomitante ao aumento do grau de substituição
da amostra.
Na região de 810 cm-1 é verificada banda correspondente à vibração simétrica
C-O-S, associado ao grupo C-O-SO3, o que comprova a sulfatação das amostras
(CALEGARI et al., 2017; VASCONCELOS et al., 2013). Maior intensidade desta
banda é verificada nas amostras com maiores graus de substituição, conforme pode
ser visto nas figuras 25 e 27 C.
Similarmente, CALEGARI et al., (2017), verificaram em amostras de
lasiodiplodana sulfatada com baixo DS (0,24) o surgimento de sinais característicos
de grupos sulfônicos, como as bandas na região de 1240 e 810 cm-1 correspondentes
a vibrações assimétricas (forte) S=O e vibração simétrica C-O-S, respectivamente.
LIU et al., (2017) produziram um derivado sulfatado com alto DS (1,6) a partir
da glucana extraída do basidiomiceto Phellinus ribis, utilizando formamida e ácido
clorosulfônico como agente sulfatante. Tais autores também confirmaram a sulfatação
da amostra pelas bandas características nas regiões em 1251 cm-1 (vibração
assimétrica S=O) e 808 cm-1 (simétrica das ligações C-O-S associada ao grupo C-O-
SO3).
5.5 Análise das imagens de microscopia eletrônica de varredura (MEV)

69
As micrografias das amostras de lasiodiplodana nativa e dos derivados
sulfatados estão apresentadas na figura 28, nas amplitudes de 300x, 800x e 1200x.
Figura 28. Micrografias das amostras LAS-N, LAS-S1:10, LAS-S1:6, LAS-S1:4 e LAS-S1:5, nas amplitudes de 300x, 800x e 1200x. Fonte: Autoria própria.

70
A microscopia eletrônica de varredura mostra que a lasiodiplodana nativa (LAS-
N) apresenta uma superfície desuniforme contendo dobras e bordas irregulares
(Figura 28, LAS-N A e B) e apresentando porções semelhantes a filmes finos
translúcidos com torções ao longo da extensão (Figura 28 LAS-N A e C).
Similarmente, THEIS et al., (2017) verificaram que amostras de lasiodiplodana nativa
apresentaram macroestrutura irregular lembrando placas com dobras e torções em
sua extensão.
Por outro lado, em estudo desenvolvido por KAGIMURA et al., (2015b), as
micrografias de amostras de lasiodiplodana nativa, evidenciaram a presença de
grânulos esféricos em toda a superfície das placas que compunham a macroestrutura
do polímero. Tal diferença pode ser justificada pela forma em que se deu o cultivo
submerso. KAGIMURA et al., (2015b) conduziu a fermentação em biorreator de
bancada com condições fermentativas diferentes daquelas utilizadas no presente
estudo e também por THEIS et al., (2017).
A sulfatação promoveu mudanças morfológicas na superfície do biopolímero
levando ao surgimento de estruturas fibrilares e heterogêneas ao longo da área
superficial, conforme monstra a figura 28. O grau de substituição (DS) parece
influenciar na morfologia superficial da molécula, tendo sido observado nas amostras
com os maiores DS (LAS-S1:4 e LAS-S1:5) maior quantidade de estruturas fibrilares
associada a porções similares a filmes finos e translúcidos. Por outro lado, nas
amostras com menores graus de substituição (LAS-S1:10 e LAS-S1:6) há um menor
número de estruturas fibrilares ao longo da área superficial. As diferenças
morfológicas entre as amostras com maiores DS comparadas com as de menores DS,
possivelmente estão associadas com a intensidade da reação de sulfatação, visto que
para maior sulfatação, maiores quantidades de ácido clorosulfônico no reagente
sulfatante são utilizadas.
CALEGARI et al., (2017), reportaram intensas mudanças na área superficial de
amostras de lasiodiplodana sulfonada com DS de 0,24, comparadas à amostra não
derivatizada. Similarmente, WANG et al., (2005), também relataram mudanças
morfológicas na superfície de uma (1→3)-β-D-glucana sulfatada com DS de 0,36,
observando rupturas na área superficial e o surgimento de fibras ao longo da sua
extensão.
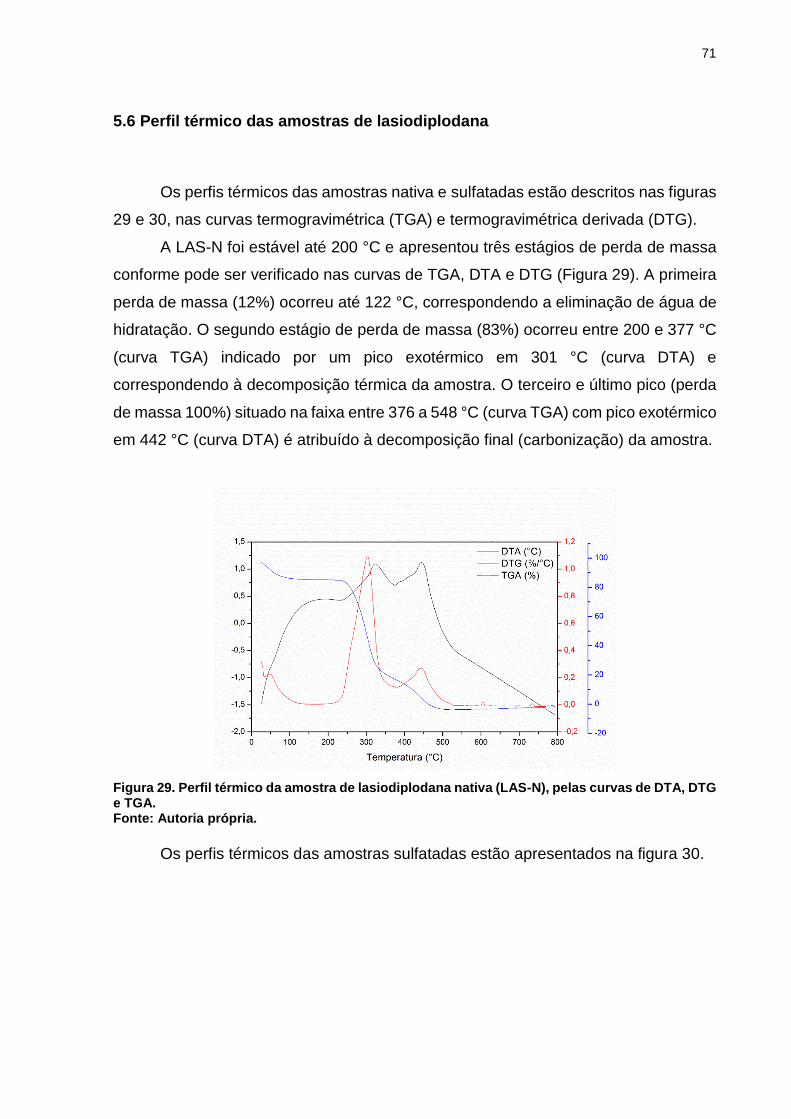
71
5.6 Perfil térmico das amostras de lasiodiplodana
Os perfis térmicos das amostras nativa e sulfatadas estão descritos nas figuras
29 e 30, nas curvas termogravimétrica (TGA) e termogravimétrica derivada (DTG).
A LAS-N foi estável até 200 °C e apresentou três estágios de perda de massa
conforme pode ser verificado nas curvas de TGA, DTA e DTG (Figura 29). A primeira
perda de massa (12%) ocorreu até 122 °C, correspondendo a eliminação de água de
hidratação. O segundo estágio de perda de massa (83%) ocorreu entre 200 e 377 °C
(curva TGA) indicado por um pico exotérmico em 301 °C (curva DTA) e
correspondendo à decomposição térmica da amostra. O terceiro e último pico (perda
de massa 100%) situado na faixa entre 376 a 548 °C (curva TGA) com pico exotérmico
em 442 °C (curva DTA) é atribuído à decomposição final (carbonização) da amostra.
Figura 29. Perfil térmico da amostra de lasiodiplodana nativa (LAS-N), pelas curvas de DTA, DTG e TGA. Fonte: Autoria própria.
Os perfis térmicos das amostras sulfatadas estão apresentados na figura 30.
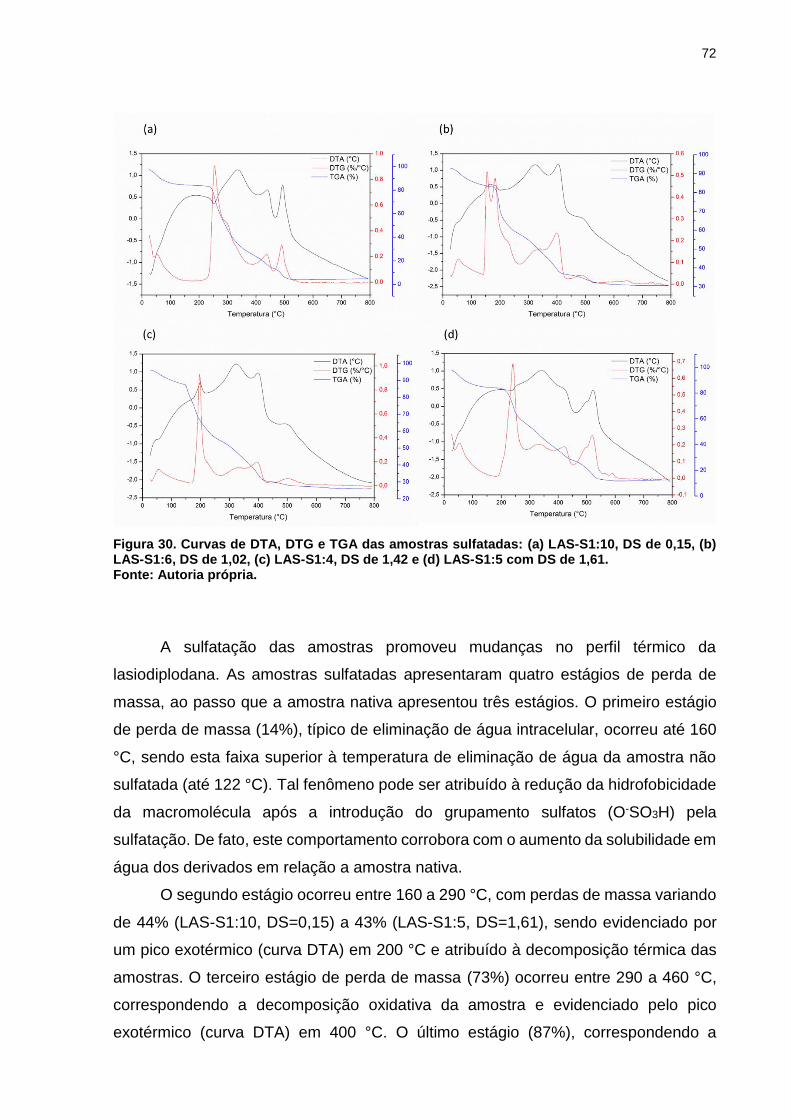
72
Figura 30. Curvas de DTA, DTG e TGA das amostras sulfatadas: (a) LAS-S1:10, DS de 0,15, (b) LAS-S1:6, DS de 1,02, (c) LAS-S1:4, DS de 1,42 e (d) LAS-S1:5 com DS de 1,61. Fonte: Autoria própria.
A sulfatação das amostras promoveu mudanças no perfil térmico da
lasiodiplodana. As amostras sulfatadas apresentaram quatro estágios de perda de
massa, ao passo que a amostra nativa apresentou três estágios. O primeiro estágio
de perda de massa (14%), típico de eliminação de água intracelular, ocorreu até 160
°C, sendo esta faixa superior à temperatura de eliminação de água da amostra não
sulfatada (até 122 °C). Tal fenômeno pode ser atribuído à redução da hidrofobicidade
da macromolécula após a introdução do grupamento sulfatos (O-SO3H) pela
sulfatação. De fato, este comportamento corrobora com o aumento da solubilidade em
água dos derivados em relação a amostra nativa.
O segundo estágio ocorreu entre 160 a 290 °C, com perdas de massa variando
de 44% (LAS-S1:10, DS=0,15) a 43% (LAS-S1:5, DS=1,61), sendo evidenciado por
um pico exotérmico (curva DTA) em 200 °C e atribuído à decomposição térmica das
amostras. O terceiro estágio de perda de massa (73%) ocorreu entre 290 a 460 °C,
correspondendo a decomposição oxidativa da amostra e evidenciado pelo pico
exotérmico (curva DTA) em 400 °C. O último estágio (87%), correspondendo a

73
decomposição final da amostra (carbonização) ocorreu entre 440 a 570 °C e foi
evidenciado pelo pico exotérmico em 500 °C (curva DTA).
De maneira geral a sulfatação da lasiodiplodana contribuiu para um pequeno
aumento na estabilidade térmica. Este comportamento é evidenciado pelo aumento
na faixa de temperatura correspondente a oxidação do biopolímero (de 377 °C para
460 °C), bem como aumento na temperatura final de carbonização da amostra (548
°C a 570 °C).
Estudos desenvolvidos por DE MOURA NETO et al. (2011), avaliaram o perfil
térmico de amostras de polissacarídeos extraídos de cajueiros e derivatizados por
sulfatação. Neste trabalho, os autores também observaram que a amostra permanecia
estável termicamente até aproximadamente 200 °C, e verificaram três estágios de
perda de massa.
ZHANG et al. (2016) avaliou o perfil térmico de um exopolissacarídeo
microbiano, produzido por Streptococcus thermophilus GST-6 e seu derivado
sulfatado (DS 0,26). Estes autores reportaram a presença de apenas dois estágios de
perda de massa e destacam uma pequena diminuição da estabilidade térmica da
amostra.
5.7 Análise por difração de raios-X (DRX)
O perfil difratográfico das amostras de lasiodiplodana nativa e sulfatada são
apresentados na figura 31.
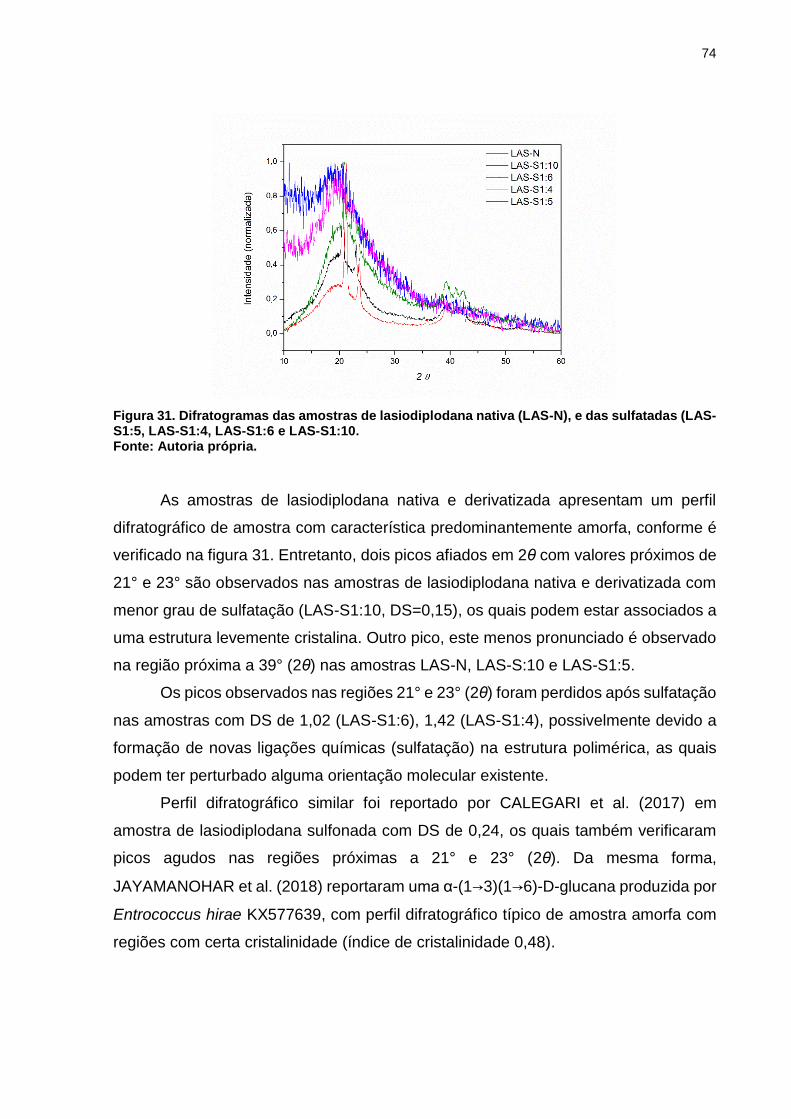
74
Figura 31. Difratogramas das amostras de lasiodiplodana nativa (LAS-N), e das sulfatadas (LAS-S1:5, LAS-S1:4, LAS-S1:6 e LAS-S1:10. Fonte: Autoria própria.
As amostras de lasiodiplodana nativa e derivatizada apresentam um perfil
difratográfico de amostra com característica predominantemente amorfa, conforme é
verificado na figura 31. Entretanto, dois picos afiados em 2θ com valores próximos de
21° e 23° são observados nas amostras de lasiodiplodana nativa e derivatizada com
menor grau de sulfatação (LAS-S1:10, DS=0,15), os quais podem estar associados a
uma estrutura levemente cristalina. Outro pico, este menos pronunciado é observado
na região próxima a 39° (2θ) nas amostras LAS-N, LAS-S:10 e LAS-S1:5.
Os picos observados nas regiões 21° e 23° (2θ) foram perdidos após sulfatação
nas amostras com DS de 1,02 (LAS-S1:6), 1,42 (LAS-S1:4), possivelmente devido a
formação de novas ligações químicas (sulfatação) na estrutura polimérica, as quais
podem ter perturbado alguma orientação molecular existente.
Perfil difratográfico similar foi reportado por CALEGARI et al. (2017) em
amostra de lasiodiplodana sulfonada com DS de 0,24, os quais também verificaram
picos agudos nas regiões próximas a 21° e 23° (2θ). Da mesma forma,
JAYAMANOHAR et al. (2018) reportaram uma α-(1→3)(1→6)-D-glucana produzida por
Entrococcus hirae KX577639, com perfil difratográfico típico de amostra amorfa com
regiões com certa cristalinidade (índice de cristalinidade 0,48).

75
5.8 Atividade antimicrobiana (MIC)
Na tabela 4 e apêndice 1 estão apresentados os resultados referentes ao
potencial antimicrobiano das amostras de lasiodiplodana nativa e sulfatadas.
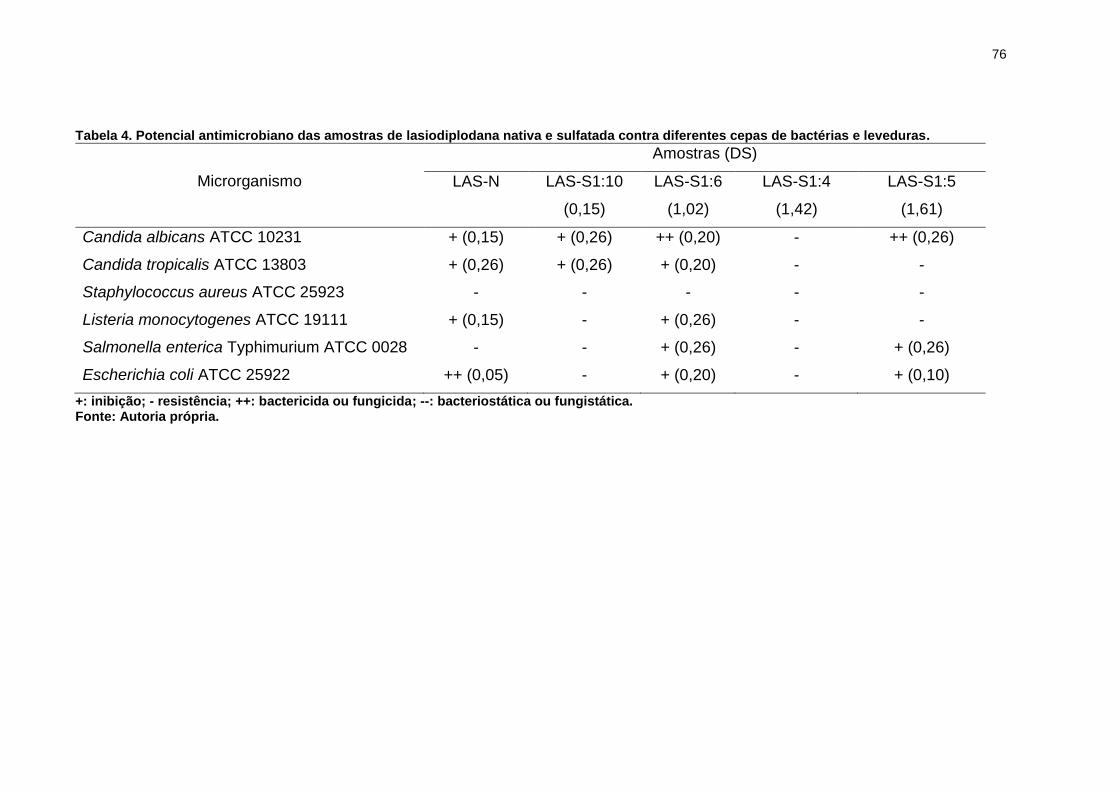
76
Tabela 4. Potencial antimicrobiano das amostras de lasiodiplodana nativa e sulfatada contra diferentes cepas de bactérias e leveduras.
Microrganismo
Amostras (DS)
LAS-N LAS-S1:10
(0,15)
LAS-S1:6
(1,02)
LAS-S1:4
(1,42)
LAS-S1:5
(1,61)
Candida albicans ATCC 10231 + (0,15) + (0,26) ++ (0,20) - ++ (0,26)
Candida tropicalis ATCC 13803 + (0,26) + (0,26) + (0,20) - -
Staphylococcus aureus ATCC 25923 - - - - -
Listeria monocytogenes ATCC 19111 + (0,15) - + (0,26) - -
Salmonella enterica Typhimurium ATCC 0028 - - + (0,26) - + (0,26)
Escherichia coli ATCC 25922 ++ (0,05) - + (0,20) - + (0,10)
+: inibição; - resistência; ++: bactericida ou fungicida; --: bacteriostática ou fungistática. Fonte: Autoria própria.

77
É possível verificar que as amostras de lasiodiplodanas nativa (LAS-N) e
sulfatadas apresentaram potencial de inibição microbiano tanto contra leveduras como
contra bactérias.
LAS-N apresentou efeito fungistático contra as leveduras Candida albicans e
Candida tropicalis nas concentrações de 0,15 mg mL-1 e 0,26 mg mL-1,
respectivamente. Similarmente, THEIS et al., (2017) encontraram efeito fungistático
da lasiodiplodana não modificada contra as mesmas leveduras em concentrações de
0,175 mg mL-1.
LAS-N também apresentou capacidade inibitória contra bactérias, sendo
verificado efeito bacteriostático contra L. monocytogenes (Gram positiva) na
concentração mínima de 0,15 mg mL-1, e efeito bactericida (MIC=0,05 mg mL-1) contra
Escherichia coli (Gram negativa).
Estudos anteriores desenvolvidos por THEIS et al., (2017), reportaram que
lasiodiplodana nativa demonstrou efeito bacteriostático (MIC = 17,5 mg mL-1), frente a
três cepas Gram negativas, Salmonella enterica Typhimurium ATCC 0028, E. coli
ATCC 25922 e Salmonella bongori ATCC 43975.
Da mesma forma, o polissacarídeo extracelular Lasiosana, produzido pelo
ascomiceto Lasiodiplodia sp. estirpe B2 demonstrou potencial antimicrobiano contra
diversas leveduras e bactérias (Staphylococcus aureus MTCC 96, Staphylococcus
aureus MLS16 MTCC 2940, Staphylococcus aureus ATCC 6538P, Micrococcus luteus
MTCC 2470, Escherichia coli MTCC 739, Klebsiella planticola MTCC 530,
Chromobacterium violaceum ATCC 12472, Enterococcus faecalis MTCC 2729,
Bacillus subtilis MTCC 121, e antifúngicas contra Candida albicans MTCC 183,
Candida albicans MTCC 227, Candida albicans MTCC 854, Candida albicans MTCC
1637, Candida albicans MTCC 3017, Candida albicans MTCC 3958, Candida albicans
MTCC 4748, Candida albicans MTCC 7315, Candida albicans MTCC 3018, Candida
parapsilosis MTCC 1744, Candida aaseri TCC 1962, Candida glabrata MTCC 3019,
Issatchenkia orientalis MTCC 3020 e Issatchenkia hanoiensis MTCC 4755) (KUMAR;
MONGOLLA; POMBALA, 2018).
As amostras sulfatadas também apresentaram atividade antimicrobiana contra
os microrganismos estudados, com exceção da cepa Staphylococcus aureus, a qual
demonstrou resistência frente a todas as amostras.

78
O grau de sulfatação da amostra parece influenciar seu potencial
antimicrobiano, entretanto, sem haver uma correlação linear entre o grau de
substituição e a capacidade inibitória. Neste contexto, pode-se observar que a
amostra com DS 1,42 não apresentou atividade inibitória contra os microrganismos
avaliados, ao passo que, a amostra com menor grau de substituição (DS = 0,15)
apresentou atividade fungistática (MIC = 0,26 mg mL-1) contra C. albicans e C.
tropicalis. Da mesma forma, as amostras com DS 1,02 (MIC = 0,20 mg mL-1) e 1,61
(MIC = 0,26 mg mL-1) foram capazes de exercer atividade fungicida contra C. albicans,
porém a espécie C. tropicalis não foi inibida pela amostra com maior DS (1,61).
A sulfatação parece não ter efeito muito efetivo contra bactérias Gram positivas
(S. aureus e L. monocytogenes), havendo inibição apenas da bactéria L.
monocytogenes pela amostra com DS de 1,02 (MIC = 0,26 mg mL-1). Dentre todas as
amostras sulfatadas estudadas, a amostra com DS 1,02 foi a que demonstrou maior
potencial de atividade antimicrobiana (tabela 4 e apêndice 1).
Importante destacar que o processo de sulfatação (DS 1,02 e 1,61) promoveu
o desenvolvimento de atividade antimicrobiana contra Salmonella enterica
Typhimurium (Gram negativa), efeito este não observado na molécula nativa (LAS-N).
Outro aspecto a ser considerado, é que comumente as bactérias Gram negativas são
mais resistentes do que as Gram positivas em função da sua parede celular
diferenciada (CALO et al., 2015).
CALEGARI et al., (2017), obteve resultados interessantes em relação à
atividade antimicrobiana contra diferentes bactérias. A lasiodiplodana modificada por
sulfatação com DS 0,24, demonstrou atividade bacteriostática contra S. enterica
Typhimurium ATCC 0028 e E. coli ATCC 25922. Já frente a bactérias Gram positivas
não foi verificado atividade inibitória. Entretanto, da mesma forma, não são elucidados
neste trabalho o mecanismo de ação destas moléculas na célula microbiana.
LI e SHAH (2014), analisaram o potencial antimicrobiano de dois
polissacarídeos oriundos de Pleurotus eryngii (DS de 0,69) e Streptococcus
thermophilus (DS de 0,31) após sulfatação e verificaram a potencialização da
atividade antimicrobiana pela derivatização por sulfatação.
Os resultados de atividade antimicrobiana obtidos após sulfatação da
lasiodiplodana no presente trabalho, revela que esta macromolécula pode ser uma
boa alternativa ao combate de infecções fúngicas, como aquelas relacionadas a
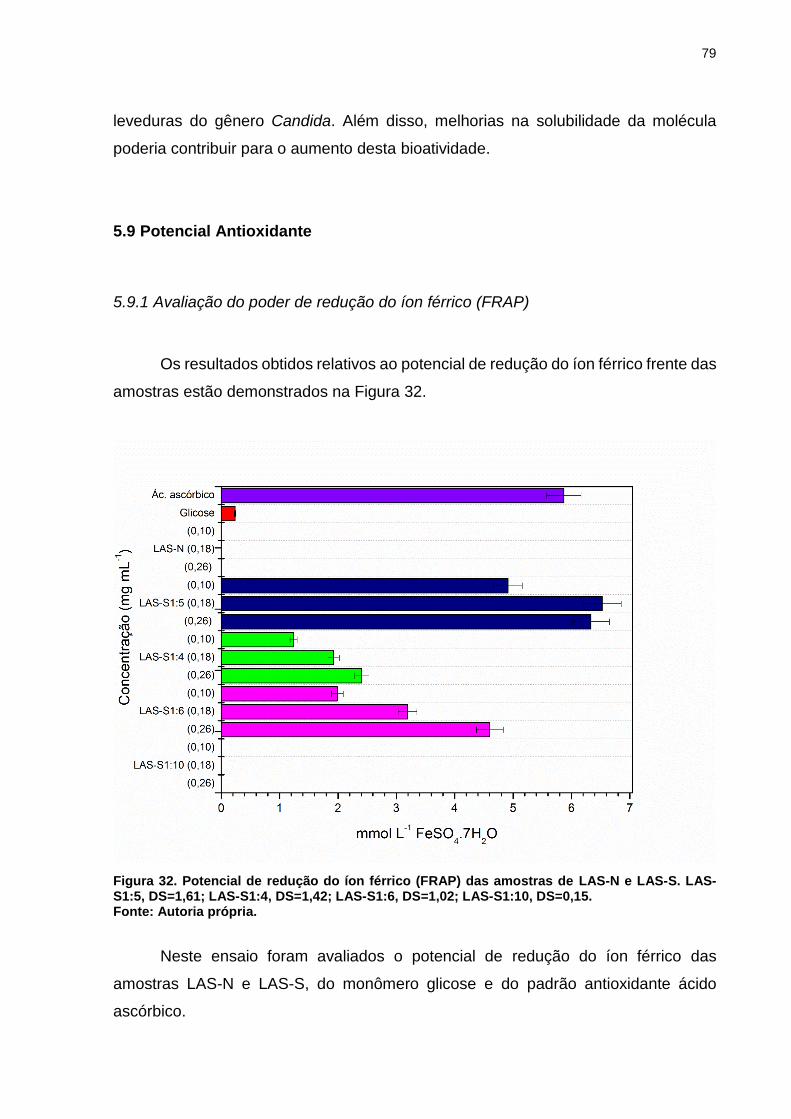
79
leveduras do gênero Candida. Além disso, melhorias na solubilidade da molécula
poderia contribuir para o aumento desta bioatividade.
5.9 Potencial Antioxidante
5.9.1 Avaliação do poder de redução do íon férrico (FRAP)
Os resultados obtidos relativos ao potencial de redução do íon férrico frente das
amostras estão demonstrados na Figura 32.
Figura 32. Potencial de redução do íon férrico (FRAP) das amostras de LAS-N e LAS-S. LAS-S1:5, DS=1,61; LAS-S1:4, DS=1,42; LAS-S1:6, DS=1,02; LAS-S1:10, DS=0,15. Fonte: Autoria própria.
Neste ensaio foram avaliados o potencial de redução do íon férrico das
amostras LAS-N e LAS-S, do monômero glicose e do padrão antioxidante ácido
ascórbico.

80
O monômero glicose apresentou baixo potencial de redução do íon férrico à
ferroso (0,24 mmol L-1 FeSO4.7H2O) e a lasiodiplodana nativa não demonstrou
capacidade de redução do íon férrico em todas as concentrações estudadas. Estes
resultados indicam que há um comportamento de atividade antioxidante diferente
entre a glicose na sua forma isolada e como constituinte da glucana lasiodiplodana. O
padrão ácido ascórbico apresentou elevado potencial FRAP (5,87 mmol L-1
FeSO4.7H2O), o que já era esperado considerando que ele é um padrão antioxidante
de referência.
A sulfatação da lasiodiplodana promoveu a potencialização da capacidade de
redução do íon férrico, com exceção da amostra com baixo grau de substituição (0,15),
que não demonstrou atividade redutora. A introdução de um maior número de
grupamentos sulfatos na macromolécula contribuiu para maior potencial antioxidante
redutor. As amostras com maiores DS (LAS-S1:5, DS=1,61 e LAS-S1:4, DS=1,42)
foram as que apresentaram os maiores potenciais antioxidante redutor, sendo
verificados valores de 6,53 mmol L-1 FeSO4.7H2O (0,18 mg mL-1) e 4,6 mmol L-1
FeSO4.7H2O (0,26 mg mL-1), respectivamente.
Outro aspecto a ser destacado, é uma correlação entre concentração do
polissacarídeo sulfatado e a atividade redutora de íon férrico nas amostras com
diferentes valores de DS. Relação dose dependente entre concentração do
polissacarídeo sulfatado e poder redutor também foi reportado por LI e SHAH (2014),
os quais avaliaram o potencial FRAP de derivados sulfatados de um polissacarídeo
extraído Pleurotus enryngii e de um exopolissacarídeo obtido de Streptococcus
thermophilus ASCC 1275.
5.9.2 Avaliação da capacidade de sequestro do radical hidroxila
A capacidade de sequestro do radical hidroxila testado frente as amostras de
lasiodiplodana nativa (LAS-N) e sulfatadas (LAS-S), bem como glicose e ácido
ascórbico, estão demonstradas na Figura 33.
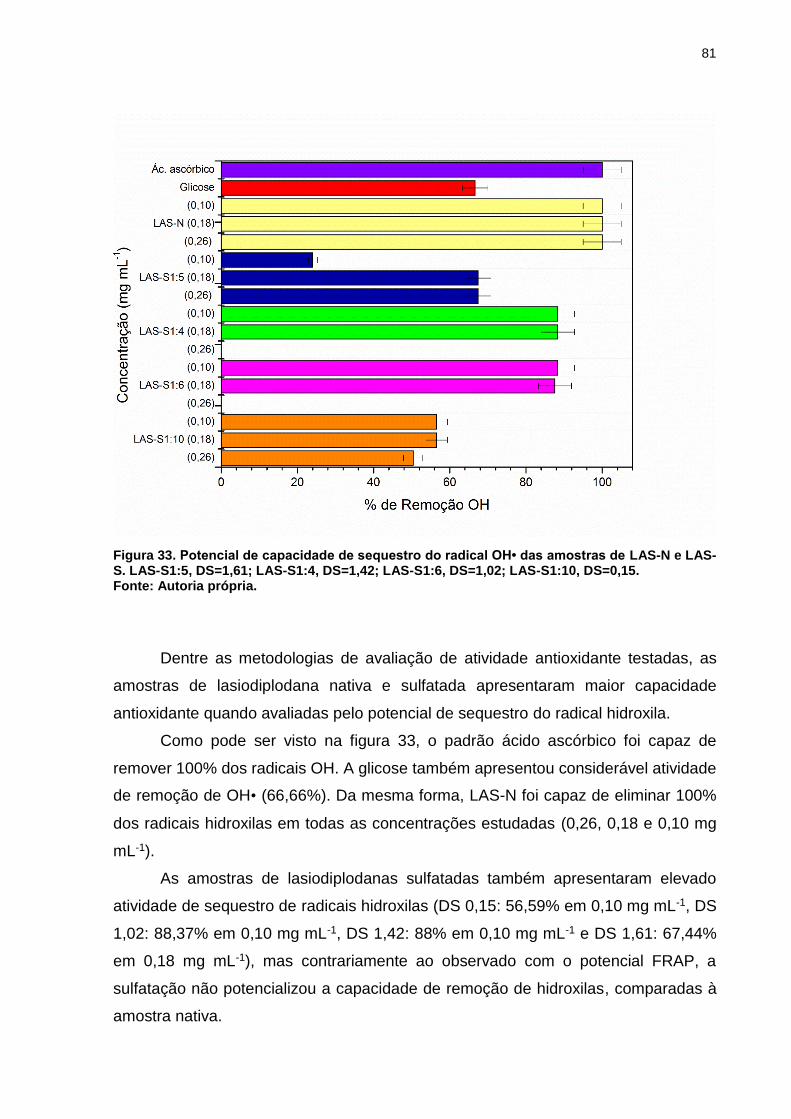
81
Figura 33. Potencial de capacidade de sequestro do radical OH• das amostras de LAS-N e LAS-S. LAS-S1:5, DS=1,61; LAS-S1:4, DS=1,42; LAS-S1:6, DS=1,02; LAS-S1:10, DS=0,15. Fonte: Autoria própria.
Dentre as metodologias de avaliação de atividade antioxidante testadas, as
amostras de lasiodiplodana nativa e sulfatada apresentaram maior capacidade
antioxidante quando avaliadas pelo potencial de sequestro do radical hidroxila.
Como pode ser visto na figura 33, o padrão ácido ascórbico foi capaz de
remover 100% dos radicais OH. A glicose também apresentou considerável atividade
de remoção de OH• (66,66%). Da mesma forma, LAS-N foi capaz de eliminar 100%
dos radicais hidroxilas em todas as concentrações estudadas (0,26, 0,18 e 0,10 mg
mL-1).
As amostras de lasiodiplodanas sulfatadas também apresentaram elevado
atividade de sequestro de radicais hidroxilas (DS 0,15: 56,59% em 0,10 mg mL-1, DS
1,02: 88,37% em 0,10 mg mL-1, DS 1,42: 88% em 0,10 mg mL-1 e DS 1,61: 67,44%
em 0,18 mg mL-1), mas contrariamente ao observado com o potencial FRAP, a
sulfatação não potencializou a capacidade de remoção de hidroxilas, comparadas à
amostra nativa.

82
Diferentemente do comportamento observado no potencial FRAP, uma
correlação direta entre a capacidade de remoção do radical hidroxila e o grau de
substituição não foi observado. Da mesma forma, a concentração do polissacarídeo
sulfatado não demonstrou influenciar a capacidade de remoção do radical hidroxila,
visto que as maiores concentrações testadas não promoveram aumento na atividade.
XIE et al., (2016) derivatizaram um polissacarídeo extraído de Cyclocarya
paliurus por sulfatação pelo método CSA:Py, no qual obtiveram quatro derivados com
graus de substituição de 0,55 (1:1), 0,42 (1:4), 0,25 (1:6) e 0,12 (1:8). Nesta pesquisa
os autores avaliaram as amostras frente a algumas análises biológicas, como a
capacidade de sequestro do radical hidroxila. Como resultados, verificaram que as
amostras com menores DS promoveram maior remoção de OH•, 44,44% (DS=0,55)
e 90,16% (DS=0,25), o que sugere que não houve uma correlação entre DS e
capacidade de remoção do radical hidroxila.
5.9.3 Avaliação da capacidade de captura do radical DPPH
A capacidade de captura do radical DPPH das amostras LAS-N e LAS-S,
glicose e Trolox, estão monstrados na Figura 34.
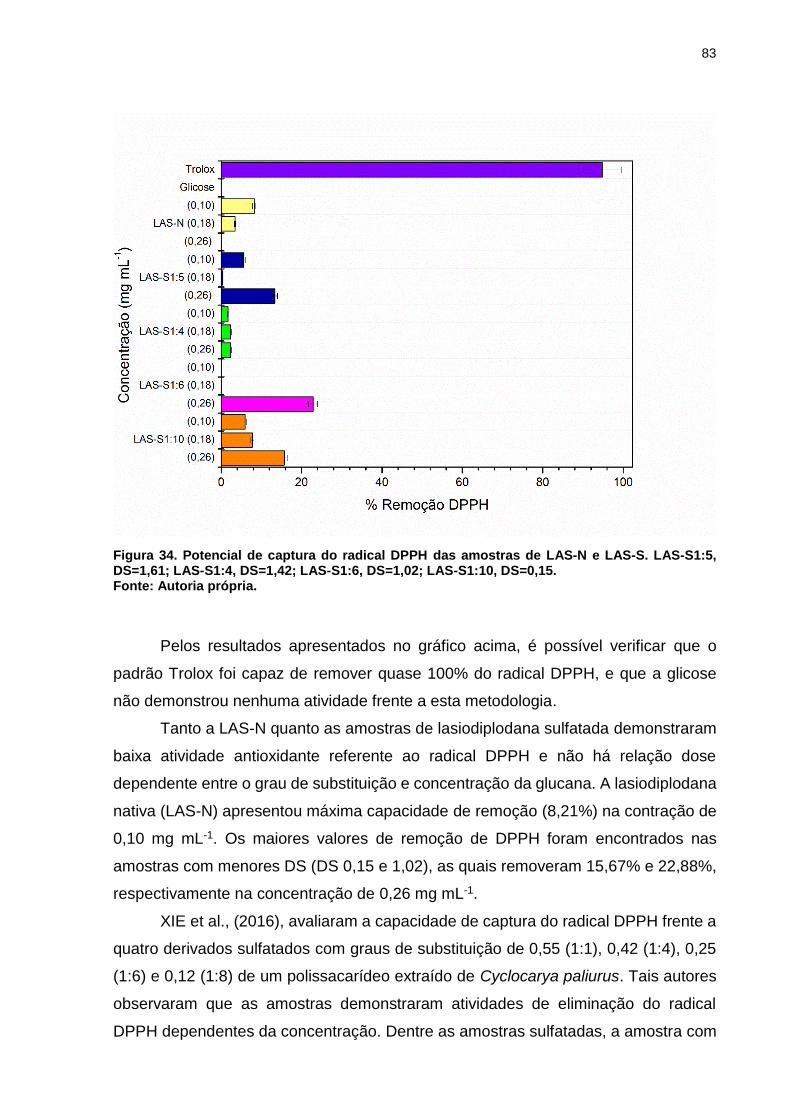
83
Figura 34. Potencial de captura do radical DPPH das amostras de LAS-N e LAS-S. LAS-S1:5, DS=1,61; LAS-S1:4, DS=1,42; LAS-S1:6, DS=1,02; LAS-S1:10, DS=0,15. Fonte: Autoria própria.
Pelos resultados apresentados no gráfico acima, é possível verificar que o
padrão Trolox foi capaz de remover quase 100% do radical DPPH, e que a glicose
não demonstrou nenhuma atividade frente a esta metodologia.
Tanto a LAS-N quanto as amostras de lasiodiplodana sulfatada demonstraram
baixa atividade antioxidante referente ao radical DPPH e não há relação dose
dependente entre o grau de substituição e concentração da glucana. A lasiodiplodana
nativa (LAS-N) apresentou máxima capacidade de remoção (8,21%) na contração de
0,10 mg mL-1. Os maiores valores de remoção de DPPH foram encontrados nas
amostras com menores DS (DS 0,15 e 1,02), as quais removeram 15,67% e 22,88%,
respectivamente na concentração de 0,26 mg mL-1.
XIE et al., (2016), avaliaram a capacidade de captura do radical DPPH frente a
quatro derivados sulfatados com graus de substituição de 0,55 (1:1), 0,42 (1:4), 0,25
(1:6) e 0,12 (1:8) de um polissacarídeo extraído de Cyclocarya paliurus. Tais autores
observaram que as amostras demonstraram atividades de eliminação do radical
DPPH dependentes da concentração. Dentre as amostras sulfatadas, a amostra com
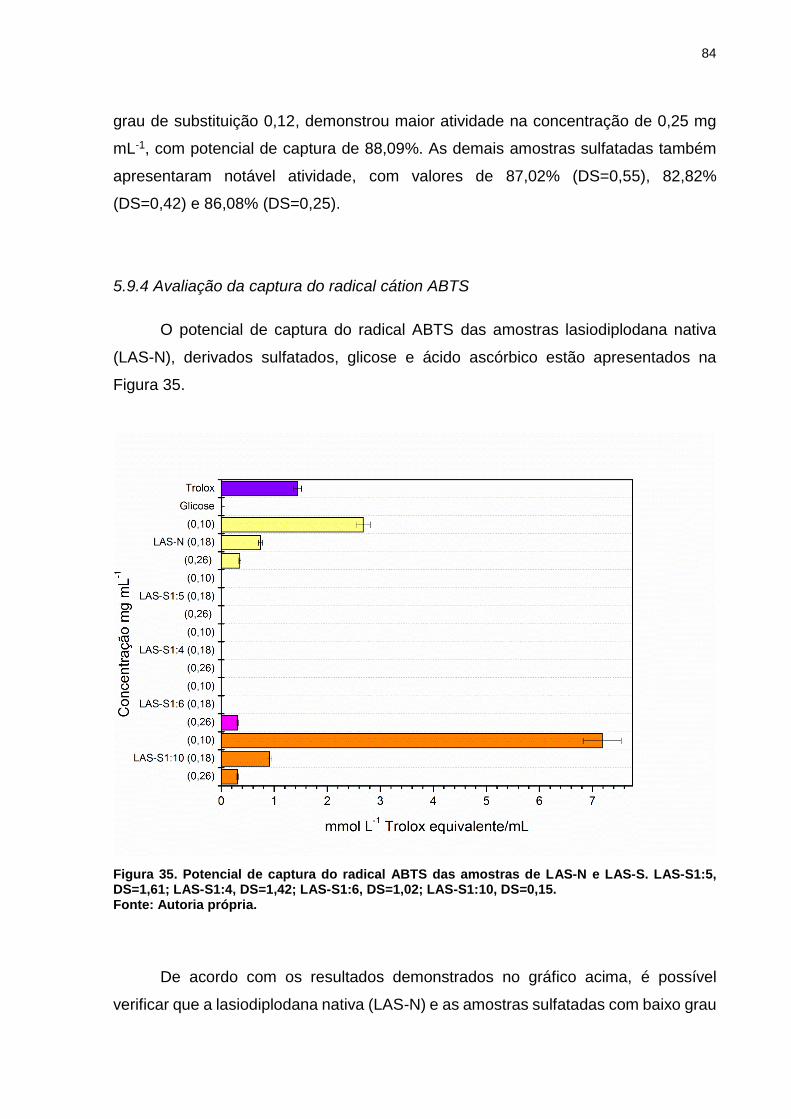
84
grau de substituição 0,12, demonstrou maior atividade na concentração de 0,25 mg
mL-1, com potencial de captura de 88,09%. As demais amostras sulfatadas também
apresentaram notável atividade, com valores de 87,02% (DS=0,55), 82,82%
(DS=0,42) e 86,08% (DS=0,25).
5.9.4 Avaliação da captura do radical cátion ABTS
O potencial de captura do radical ABTS das amostras lasiodiplodana nativa
(LAS-N), derivados sulfatados, glicose e ácido ascórbico estão apresentados na
Figura 35.
Figura 35. Potencial de captura do radical ABTS das amostras de LAS-N e LAS-S. LAS-S1:5, DS=1,61; LAS-S1:4, DS=1,42; LAS-S1:6, DS=1,02; LAS-S1:10, DS=0,15. Fonte: Autoria própria.
De acordo com os resultados demonstrados no gráfico acima, é possível
verificar que a lasiodiplodana nativa (LAS-N) e as amostras sulfatadas com baixo grau

85
de substituição (LAS-S1:10, DS 0,15) demonstraram potencial antioxidante contra o
radical ABTS.
As maiores atividades de captura do radical ABTS foram verificadas nos
ensaios com menores concentrações (0,10 mg mL-1) da glucana nativa (2,68 mmol L-
1 trolox equivalente/mL) ou sulfatada (7,18 mmol L-1 trolox equivalente/mL), ou seja, o
aumento da concentração do polissacarídeo não promoveu maior capacidade
antioxidante. A lasiodiplodana sulfatada com menor DS (0,15) apresentou potencial
de captura do ABTS 168% superior comparada a amostra LAS-N.
LI e SHAH (2017), reportaram que a sulfatação dos polissacarídeos obtidos de
Pleurotus eryngii (DS=0,73) e Streptococcus thermopilus ASCC 1725 (DS=0,37),
potencializou a capacidade de captura do radical ABTS, sendo verificadas remoções
de 70,8% e 61,6%, respectivamente. Tal aumento da atividade de captura do radical
ABTS foi justificado pelos autores em função das amostras sulfatadas terem maior
capacidade de doação de prótons para radicais ABTS.
As atividades antioxidantes da lasiodiplodana nativa e sulfatadas avaliadas pelo
método de remoção dos radicais DPPH e ABTS, foram inferiores as atividades
determinadas pelos métodos FRAP e remoção de OH•. Tal comportamento
possivelmente pode ser justificado em função do uso do etanol como solvente nos
protocolos de remoção dos radicais DPPH e ABTS, uma vez que polissacarídeos
podem ser insolubilizados frente a álcoois.
5.10 Avaliação da citotoxicidade sobre Saccharomyces cerevisiae
Os resultados do estresse oxidativo das amostras testadas (LAS-N e LAS-S)
estão apresentados na tabela 6.

86
Tabela 5. Estresse oxidativo frente as cepas de leveduras testadas das amostras de lasiodiplodana nativa e sulfatadas.
S. cerevisiae
Amostras (DS)
LAS-N 0,15 1,02 1,42 1,61 Menadiona
(mg mL-1) (m mol L-1)
Padrão* -- (0,20) -- (0,26) -- (0,20) - - ++ (18,5)
YLL060C∆GTT2 -- (0,20) -- (0,26) -- (0,20) - -- (0,26) ++ (18,5)
YIR038C∆GTT1 -- (0,20) -- (0,20) -- (0,20) - - ++ (18,5)
YSL101C∆GSH1 -- (0,20) -- (0,20) -- (0,20) - -- (0,26) ++ (18,5)
+: inibição; - resistência; ++: atividade biocida; --: atividade biostática.
*Padrão= Saccharomyces cerevisiae f.r bayanus; LAS-S. LAS-S1:5, DS=1,61; LAS-S1:4, DS=1,42;
LAS-S1:6, DS=1,02; LAS-S1:10, DS=0,15.
Fonte: Autoria própria.
Conforme verificado na tabela 6 e apêndice 2, a naftoquinona menadiona
(vitamina K3) demonstrou citotoxicidade letal contra as cepas de S. cerevisiae, mesmo
em concentrações bastante baixas. De fato, a menadiona é um importante agente
indutor do estresse oxidativo, o qual contribui para o aumento da produção de
espécies químicas reativas (ROS), e leva os níveis de ROS a excederem a capacidade
antioxidante endógena.
Por outro lado, a lasiodiplodana nativa (LAS-N) e as amostras sulfatadas não
demonstraram citotoxicidade letal contra as cepas de S. cerevisiae, nas
concentrações estudadas. Apenas a LAS-N demonstrou efeito inibitório (biostático)
durante 24 horas de incubação na concentração de 0,20 mg mL-1, tanto sobre a cepa
selvagem como as mutantes.
Similarmente, as amostras sulfatadas com os menores DS (0,15 e 1,02) não
demonstraram atividade biocida, apenas inibindo a atividade celular durante 24 horas
nas concentrações de 0,20 e 0,26 mg mL-1. Os derivados com maior grau de

87
substituição (1,42 e 1,61) apresentaram menor capacidade de indução ao estresse
oxidativo. Neste sentido, a amostra sulfatada com DS 1,42 não promoveu inibição à
viabilidade celular de nenhuma das cepas avaliadas. A amostra com maior grau de
sulfatação (1,61) promoveu atividade biostática em concentrações de 0,26 mg mL-1
apenas sobre os mutantes YLL060C e YSL101C.
Embora LAS-N e LAS-S demonstram atividade antioxidante, especialmente em
relação a capacidade de eliminação dos radicais hidroxila e atividade redutora do íon
férrico, possivelmente não há inibição da produção e acúmulo de determinadas
espécies reativas de oxigênio (ROS) pela lasiodiplodana. De fato, lasiodiplodana
possui atividade antimicrobiana contra determinadas espécies microbianas, como E.
coli e C. tropicalis estudadas neste trabalho. Entretanto, é importante notar que
mesmo não havendo eliminação de um desbalanço entre acúmulo de ROS e atividade
antioxidante, não houve letalidade das células das estirpes mutantes, as quais são
incapazes de produzir glutationa, sugerindo uma baixa toxicidade da macromolécula
lasiodiplodana, em relação a indução ao estresse oxidativo.
WAN-MOHTAR et al. (2016) avaliram a citotoxicidade de uma β-glucana nativa
(G) extraída do cogumelo Ganoderma lucidum BCCM 31549 e seu derivado sulfatado
(GS) frente a células de próstata humana (PN2TA) e linfócitos caucasianos de
linfomas histiocítico humano (U937). Tais autores verificaram que tanto os polímeros
G e GS não apresentaram efeito citotóxico sobre as células de próstata humana
(PN2TA). Em relação as células U937, as amostras demonstraram efeito
antiproliferativo dose dependente. A glucana sulfatada (GS) demonstrou ser mais
potente quanto a atividade antiproliferativa na concentração de 60 µg mL-1,
promovendo uma inibição de crescimento celular de 40%, quando comparada com a
glucana nativa (10%).

88
6 CONCLUSÕES
O protocolo analítico empregado no processo de sulfatação da lasiodiplodana
foi efetivo e promoveu a obtenção de derivados com diferentes graus de substituição
(1,61, 1,42, 1,02 e 0,15). A proporção ácido clorossulfônico-piridina empregadas no
protocolo de sulfatação demonstrou ter influência direta sobre o grau de substituição
da macromolécula.
A derivatização por sulfatação contribuiu para pequeno aumento na
solubilidade da lasiodiplodana, ocorrendo aumento de solubilidade com o aumento do
DS.
Os espectros de IV-TF confirmaram a sulfatação da lasiodiplodana, sendo
verificada bandas em 1250 cm-1 atribuída a vibração de alongamento assimétrico
(forte) S=O e 810 cm-1 correspondente a vibração simétrica C-O-S, associado ao
grupo C-O-SO3.
A sulfatação promoveu mudanças morfológicas na superfície do biopolímero,
levando ao surgimento de estruturas fibrilares e heterogêneas ao longo da área
superficial. O grau de substituição demonstrou ter influência nas mudanças
morfologicas da macromolécula.
Lasiodiplodana nativa apresentou elevada estabilidade térmica, ocorrendo
oxidação do biopolímero em temperaturas próximas a 377 °C. A sulfatação promoveu
aumento da estabilidade térmica, ocorrendo oxidação do biopolímero em
temperaturas próximas a 460 °C.
Análise por DRX demonstrou que as amostras de lasiodiplodana nativa e
sulfatadas apresentam perfil cristalográfico predominantemente amorfo, característico
de polissacarídeos.
Lasiodiplodana nativa (LAS-N) e derivados sulfatados demonstraram potencial
de inibição microbiana. LAS-N apresentou efeito fungistático contra C. albicans e C.
tropicalis, efeito bacteriostático contra L. monocytogenes e bactericida contra E. coli.
A sulfatação da macromolécula ampliou sua atividade antimicrobiana, especialmente
contra a C. albicans e S. enterica Typhimurium, exercendo efeito fungicida e
bacteriostático, respectivamente.

89
Lasiodiplodana nativa e sulfatada demonstraram potencial antioxidante. LAS-N
apresentou maior capacidade de remoção do radical hidroxila, enquanto os derivados
sulfatados apresentaram maior potencial FRAP.
Lasiodiplodana nativa e sulfatadas não demonstraram citotoxicidade letal
contra as cepas selvagem e mutantes de S. cerevisiae, nas concentrações estudadas.
Amostras com maiores DS (1,42 e 1,61) apresentaram menor potencial de indução ao
estresse oxidativo promotor de letalidade celular.

90
7 REFERÊNCIAS
ALZORQI, I. et al. Ultrasonically extracted β D glucan from artificially cultivated
mushroom , characteristic properties and antioxidant activity. ULTRASONICS
SONOCHEMISTRY, p. 3–5, 2016.
ANAND, J. et al. Bioactive potential and composition analysis of sulfated
polysaccharide from Acanthophora spicifera (Vahl) Borgeson. International Journal
of Biological Macromolecules, v. 111, p. 1238–1244, 2018.
AOE, S. et al. Effects of high β-glucan barley on visceral fat obesity in Japanese
individuals: A randomized, double-blind study. Nutrition, v. 42, p. 1–6, 2017.
AOKI, S. et al. Oral administration of the B-glucan produced by Aureobasidium
pullulans ameliorates development of atherosclerosis in apolipoprotein E deficient
mice. Journal of Functional Foods, v. 18, p. 22–27, 2015.
BAYLIAK, M. M.; LUSHCHAK, V. I. The golden root, Rhodiola rosea, prolongs lifespan
but decreases oxidative stress resistance in yeast Saccharomyces cerevisiae.
Manufacturing Technology, v. 18, p. 1262–1268, 2011.
BOROSKI, M.; VISENTAINER, J. V.; COTTICA, S. M.; MORAIS, D. R. Antioxidantes:
princípios e métodos analíticos. 1 ed. Curitiba: Appris, 2015.
BUDDANA, S. K.; VARANASI, Y. V. N.; SHETTY, P. R. Fibrinolytic, anti-inflammatory
and anti-microbial properties of alpha-(1-3)-glucans produced from Streptococcus
mutans (MTCC 497). Carbohydrate Polymers, v. 115, p. 152–159, 2015.
CAI, Y. et al. Sulfated modification and biological activities of polysaccharides derived
from Zizyphus jujuba cv. Jinchangzao. International Journal of Biological
Macromolecules, v. 120, p. 1149–1155, 2018.
CALEGARI, G. C. et al. Sulfonation of (1→6)-β-D-glucan (lasiodiplodan) and its

91
antioxidant and antimicrobial potential. Journal of Pharmacy and Pharmacology, v.
5, n. 1, p. 1–12, 2017.
CALO, J. R. et al. Essential oils in food systems: A systemic review. International
Journal of PharmTech Research, v. 9, n. 6, p. 409–416, 2015.
CASTELLANE, T. C. L. et al. Characterization of new exopolysaccharide production
by Rhizobium tropici during growth on hydrocarbon substrate. International Journal
of Biological Macromolecules, v. 96, p. 361–369, 2017.
CASTRO, F. A. V. et al. Menadione stress in Saccharomyces cerevisiae strains
deficient in the glutathione transferases. Biochimica et Biophysica Acta - General
Subjects, v. 1770, n. 2, p. 213–220, 2007.
CASTRO, F. A. V. et al. Cytotoxicity mechanism of two naphthoquinones (menadione
and plumbagin) in Saccharomyces cerevisiae. PLoS ONE, v. 3, n. 12, 2008.
CHEIGNON, C. et al. Oxidative stress and the amyloid beta peptide in Alzheimer’s
disease. Redox Biology, v. 14, n. October 2017, p. 450–464, 2018.
CHEN, S. et al. Characteristics of Lasiodiplodia theobromae from Rosa rugosa in
South China. Crop Protection, v. 79, n. 2016, p. 51–55, 2016.
CHIARELLO, D. I. et al. Oxidative stress: Normal pregnancy versus preeclampsia.
Molecular Basis of Disease.p. 1-33 , 2018.
CIPRIANO, A. K. A. L. et al. Proteomic analysis of responsive stem proteins of resistant
and susceptible cashew plants after Lasiodiplodia theobromae infection. Journal of
Proteomics, v. 113, p. 90–109, 2015.
COSTA, V.; MORADAS-FERREIRA, P. Oxidative stress and signal transduction in
Saccharomyces cerevisiae: Insights into ageing, apoptosis and diseases. Molecular
Aspects of Medicine, v. 22, n. 4–5, p. 217–246, 2001.

92
CUNHA, M. A. A. DA et al. Lasiodiplodan, an exocellular (1→6)-β-D-glucan from
Lasiodiplodia theobromae MMPI: Production on glucose, fermentation kinetics,
rheology and anti-proliferative activity. Journal of Industrial Microbiology and
Biotechnology, v. 39, n. 8, p. 1179–1188, 2012.
DE MOURA NETO, É. et al. Preparation and characterization of a chemically sulfated
cashew gum polysaccharide. Journal of the Brazilian Chemical Society, v. 22, n.
10, p. 1953–1960, 2011.
DENG, C. et al. Physiochemical and biological properties of phosphorylated
polysaccharides from Dictyophora indusiata. International Journal of Biological
Macromolecules, v. 72, p. 894–899, 2015.
DI, W. et al. Physicochemical characterization and antitumour activity of
exopolysaccharides produced by Lactobacillus casei SB27 from yak milk.
Carbohydrate Polymers, 2017.
DONOT, F.; FONTANA, A.; SCHORR-GALINDO, S. Microbial exopolysaccharides :
Main examples of synthesis , excretion , genetics and extraction. Carbohydrate
Polymers, v. 87, n. 2, p. 951–962, 2012.
DONOT, F.; FONTANA, A.; SCHORR-GALINDO, S. The Role of the Fungal Cell Wall
in the Infection of Plants. Bioresource Technology, v. 87, p. 951–962, 2017.
FUKUDA, E. K. et al. Polissacarídeos de parede celular fúngica : purificação e
caracterização. Semina:Ciências Agrárias, v. 30, n. July, p. 117–134, 2009.
GALEANE, M. C. et al. Phytochemical screening of Azadirachta indica A. Juss for
antimicrobial activity. African Journal of Microbiology Research, v. 11, n. 4, p. 117–
122, 2017.
GEOGHEGAN, I.; STEINBERG, G.; GURR, S. The Role of the Fungal Cell Wall in the
Infection of Plants. Trends in Microbiology, p. 1–11, 2017.

93
GIESE, E. C. et al. Comparison of β-1,3-glucanase production by Botryosphaeria
rhodina MAMB-05 and Trichoderma harzianum Rifai and its optimization using a
statistical mixture-design. Biochemical Engineering Journal, v. 53, n. 2, p. 239–243,
2011.
HAN, M. et al. Rheological properties of phosphorylated exopolysaccharide produced.
International Journal of Biological Macromolecules, v. 88, p. 603–613, 2016.
HE, F. et al. Studies on antibacterial activity and antibacterial mechanism of a novel
polysaccharide from Streptomyces virginia H03. Food Control, v. 21, n. 9, p. 1257–
1262, 2010.
HUSSAIN, P. R.; RATHER, S. A.; SURADKAR, P. P. Structural characterization and
evaluation of antioxidant, anticancer and hypoglycemic activity of radiation degraded
oat (Avena sativa) β- glucan. Radiation Physics and Chemistry, p. 0–1, 2017.
JADHAV, S. B.; GUPTA, A. Studies on application of β-1,3 glucanase in the
degradation of glucans produced by Botrytis cinerea and inhibition of fungal growth.
Biocatalysis and Agricultural Biotechnology, v. 7, p. 45–47, 2016.
JAYAMANOHAR, J. et al. Characterization of α-D-glucan produced by a probiont
Enterococcus hirae KX577639 from feces of south Indian Irula tribals. International
Journal of Biological Macromolecules, v. 118, p. 1667–1675, 2018.
JI, C. et al. A report on canker disease of Falcataria moluccana caused by
Lasiodiplodia theobromae in China. Crop Protection, v. 91, p. 89–92, 2017.
JI, Y.B. et al. Optimization of Synthesis of Seleno- Sargassum fusiforme (Harv.) Setch.
Polysaccharide by Response Surface Methodology, Its Characterization, and
Antioxidant Activity. Journal of Chemistry, v. 2013, p. 1–9, 2013.
JIANG, J. et al. Purification , structural characterization and in vitro antioxidant activity
of a novel polysaccharide from Boshuzhi. Carbohydrate Polymers, v. 147, p. 365–
371, 2016.

94
JIANG, Y. et al. Cellulose-rich oleogels prepared with an emulsion-templated
approach. Food Hydrocolloids, p. 1–5, 2017.
JINDAL, M. et al. Sulfation of Aegle marmelos gum: Synthesis, physico-chemical and
functional characterization. Carbohydrate Polymers, v. 92, n. 2, p. 1660–1668, 2013.
KAGIMURA, F. Y. et al. Biological activities of derivatized D-glucans: A review.
International Journal of Biological Macromolecules, v. 72, n. 2015, p. 588–598,
2015a.
KAGIMURA, F. Y. et al. Carboxymethylation of (1,6)-B-glucan (lasiodiplodan):
Preparation, characterization and antioxidant evaluation. Carbohydrate Polymers, v.
127, p. 390–399, 2015b.
KERDSOMBOON, K. et al. Soluble Moringa oleifera leaf extract reduces intracellular
cadmium accumulation and oxidative stress in Saccharomyces cerevisiae. Journal of
Bioscience and Bioengineering, v. 121, n. 5, p. 543–549, 2016.
KHAN, A. A. et al. Structural, thermal, functional, antioxidant & antimicrobial properties
of B-D-glucan extracted from baker’s yeast (Saccharomyces cereviseae)— Effect of
gamma-irradiation. Carbohydrate Polymers, v. 140, p. 442–450, 2016.
KRICHEN, F. et al. Extraction, characterization and antimicrobial activity of sulfated
polysaccharides from fish skins. International Journal of Biological
Macromolecules, v. 75, p. 283–289, 2015.
KUMA, C. A. K.; KUMAR, K. J. Structural, functional and pH sensitive release
characteristics of water-soluble polysaccharide from the seeds of Albizia lebbeck L.
Carbohydrate Polymers, 2017.
KUMAR, C. G.; MONGOLLA, P.; POMBALA, S. Lasiosan, a new exopolysaccharide
from Lasiodiplodia sp. strain B2 (MTCC 6000): Structural characterization and
biological evaluation. Process Biochemistry, v. 72, n. June, p. 162–169, 2018.

95
KRUFT, V.; EUBEL, H.; JÄNSCH, L.; WERHAHN, W.; BRAUN, H. P. Proteomic
Approach to Identify Novel Mitochondrial Proteins in Arabidospsis. Plant Physiol., v.
127, p. 1694-1710, 2001.
LEWIS, K. Platforms for antibiotic discovery. Nature Reviews Drug Discovery, v. 12,
n. 5, p. 371–387, 2013.
LI, S. et al. Molecular Modification of Polysaccharides and Resulting Bioactivities.
Comprehensive Reviews in Food Science and Food Safety, v. 15, n. 2, p. 237–
250, 2016.
LI, S.; DAI, S.; SHAH, N. P. Sulfonation and Antioxidative Evaluation of
Polysaccharides from Pleurotus Mushroom and Streptococcus thermophilus Bacteria:
A Review. Comprehensive Reviews in Food Science and Food Safety, v. 16, n. 2,
p. 282–294, 2017.
LI, S.; SHAH, N. P. Antioxidant and antibacterial activities of sulphated
polysaccharides from Pleurotus eryngii and Streptococcus thermophilus ASCC 1275.
Food Chemistry, v. 165, p. 262–270, 2014.
LI, S.; SHAH, N. P. Sulphonated modification of polysaccharides from Pleurotus eryngii
and Streptococcus thermophilus ASCC 1275 and antioxidant activities investigation
using CCD and Caco-2 cell line models. Food Chemistry, v. 225, p. 246–257, 2017.
LIANG, L. et al. Sulfated modification and anticoagulant activity of pumpkin (Cucurbita
pepo, Lady Godiva) polysaccharide. International Journal of Biological
Macromolecules, v. 106, p. 447–455, 2018.
LIN, D. et al. Enhancing moisture resistance of starch-coated paper by improving the
film forming capability of starch film. Industrial Crops and Products, v. 100, p. 12–
18, 2017.
LIU, M. et al. The anti-diabetic activity of oat β-D-glucan in streptozotocin–nicotinamide

96
induced diabetic mice. Biological Macromolecules, p. 1–25, 2016.
LIU, W. et al. Characterization and antioxidant activity of two low-molecular-weight
polysaccharides purified from the fruiting bodies of Ganoderma lucidum. International
Journal of Biological Macromolecules, v. 46, n. 4, p. 451–457, 2010.
LIU, Y. et al. Anti-angiogenic activity and mechanism of a chemically sulfated natural
glucan from Phellinus ribis. International Journal of Biological Macromolecules, v.
107, p. 2475–2483, 2017.
LOCATELLI, M. et al. Study of the DPPH-scavenging activity: Development of a free
software for the correct interpretation of data. Food Chemistry, v. 114, n. 3, p. 889–
897, 2009.
LU, Y. et al. Sulfated modification of epimedium polysaccharide and effects of the
modifiers on cellular infectivity of IBDV. Carbohydrate Polymers, v. 71, n. 2, p. 180–
186, 2008.
LUCHESE, C. L.; SPADA, J. C.; TESSARO, I. C. Starch content affects
physicochemical properties of corn and cassava starch-based films. Industrial Crops
and Products, v. 109, p. 619–626, 2017.
LUNA, W. S. et al. O-Acetylated (1→6)-β-D-Glucan (Lasiodiplodan): Chemical
Derivatization, Characterization and Antioxidant Activity. Journal of Pharmacy and
Pharmacology 6, v. 6, p. 320–332, 2018.
MA, L. et al. Chemical modification and antioxidant activities of polysaccharide from
mushroom Inonotus obliquus. Carbohydrate Polymers, v. 89, n. 2, p. 371–378, 2012.
MAFTOONAZAD, N. et al. Effect of pectin-based edible emulsion coating on changes
in quality of avocado exposed to Lasiodiplodia theobromae infection. Carbohydrate
Polymers, v. 68, n. 2, p. 341–349, 2007.
MAHAPATRA, S.; BANERJEE, D. Fungal Exopolysaccharide : Production ,

97
Composition and Applications. Microbiology Insights, v. 6, p. 1–16, 2013.
MELLO, M. B. et al. Protective effects of the exopolysaccharide Lasiodiplodan against
DNA damage and inflammation induced by doxorubicin in rats: Cytogenetic and gene
expression assays. Toxicology, v. 376, p. 66–74, 2017.
MENDES, S. F. et al. Sulfonation and anticoagulant activity of botryosphaeran from
Botryosphaeria rhodina MAMB-05 grown on fructose. International Journal of
Biological Macromolecules, v. 45, n. 3, p. 305–309, 2009.
MORENO-MENDIETA, S. et al. Potential of glucans as vaccine adjuvants: A review of
the alpha-glucans case. Carbohydrate Polymers, v. 165, p. 103–114, 2017.
MUNIZ, C. R. et al. Colonization of cashew plants by Lasiodiplodia theobromae:
Microscopical features. Micron, v. 42, n. 5, p. 419–428, 2011.
MURRY, J. P. et al. Sulfonation pathway inhibitors block reactivation of latent HIV-1.
Virology, v. 471, p. 1–12, 2014.
NAGI, M. Global Biopolymers Market to witness a CAGR of 15.2% during 2018-2024:
Energias Market Research Pvt. Ltd. 2018. Disponível em:
<https://globenewswire.com/news-release/2018/07/11/1535796/0/en/Global-
Biopolymers-Market-to-witness-a-CAGR-of-15-2-during-2018-2024-Energias-Market-
Research-Pvt-Ltd.html>. Acesso em: 07 fev. 2018.
OLIVEIRA, K. S. M. et al. (1→6)-and (1→3)(1→6)-β-glucans from Lasiodiplodia
theobromae MMBJ: Structural characterization and pro-inflammatory activity.
Carbohydrate Polymers, v. 133, n. 2015, p. 539–546, 2015.
PAPACOSTAS, L. J. et al. An unusual skin lesion caused by Lasiodiplodia
theobromae. Medical Mycology Case Reports, v. 8, p. 44–46, 2015.

98
PAVIA, Donald L.; LAMPMAN, Gary M.; KRIZ, George S.; VYVYAN, James R.
Introdução à Espectroscopia. Revisão técnica Paulo Sergi Santos; [tradução Pedro
Barros]. – São Paulo: Cengage Learning, 2010.
PEASURA, N. et al. Characteristics and antioxidant of Ulva intestinalis sulphated
polysaccharides extracted with different solvents. International Journal of Biological
Macromolecules, v. 81, p. 912–919, 2015.
PHINICHKA, N.; KAENTHONG, S. Regenerated cellulose from high alpha cellulose
pulp of steam-exploded sugarcane bagasse. Journal of Materials Research and
Technology, p. 1–11, 2016.
POMIN, V. H. Structure-Function Relationship of Anticoagulant and
Antithrombotic Well-Defined Sulfated Polysaccharides from Marine
Invertebrates.1 ed. Elsevier Inc., 2012.
QIN, T. et al. Selenylation modification can enhance immune-enhancing activity of
Chuanminshen violaceum polysaccharide. Carbohydrate Polymers, v. 153, n. 1, p.
302–311, 2013.
RODRIGUES, R. Caracterização morfológica e patológica de lasiodiplodia
theobromae (pat.) griffon & maubl., agente causal das podridões de tronco e raízes da
videira. 2003. 53 f. Dissertação (Mestrado em Agricultura Tropical e Subtropical) -
Instituto Agronômico de Campinas, Campinas, 2003.
RODRIGUEZ, C. E. et al. An examination of quinone toxicity using the yeast
Saccharomyces cerevisiae model system. Toxicology, v. 201, n. 1–3, p. 185–196,
2004.
RUFINO, M. S. M. et al. Metodologia Científica: Determinação da Atividade
Antioxidante Total em Frutas pela Captura do Radical Livre ABTS+, EMBRAPA,
Comunicado Técnico, on line Fortaleza, 2007.
SARTESHNIZI, R. A. et al. LWT - Food Science and Technology Optimization of

99
prebiotic sausage formulation : Effect of using b -glucan and resistant starch by D-
optimal mixture design approach. LWT - Food Science and Technology, p. 1–7,
2014.
SAVI, A. Caracterização química, potencial antimicrobiano e antioxidante de
polissacarídeo extraído de cará-moela (Discorea bulbifera). 2018. 80 f. Dissertação
(Mestrado em Tecnologia de Processos Químicos e Bioquímicos) - Universidade
Tecnológica Federal do Paraná, Pato Branco, 2018.
SCHMID, F. et al. Structure and assembly of epiglucan, the extracellular (1→3;1→6)-
β-glucan produced by the fungus Epicoccum nigrum strain F19. Carbohydrate
Research, v. 341, n. 3, p. 365–373, 2006.
SHI, L. Bioactivities, isolation and purification methods of polysaccharides from natural
products: A review. International Journal of Biological Macromolecules, v. 92, p.
37–48, 2016.
SOMENSI, F.Y.K. Bioprodução de β-(1→6)-D-glucana e obtenção de derivado por
carboximetilação visando atividade biológica. 2014. 97 f. Dissertação (Mestrado em
Tecnologia de Processos Químicos e Bioquímicos) - Universidade Tecnológica
Federal do Paraná, Pato Branco, 2014.
SONG, Y. et al. Effect of acetylation on antioxidant and cytoprotective activity of
polysaccharides isolated from pumpkin (Cucurbita pepo, lady godiva). Carbohydrate
Polymers, v. 98, n. 1, p. 686–691, 2013.
SOUZA, D. M. DE; GARCIA-CRUZ, C. H. Fermentative production of exocellular
polysaccharides by bacteria. v. 25, n. January 2009, p. 331–340, 2004.
SUBHASWARAJ, P. et al. Determination of antioxidant activity of Hibiscus sabdariffa
and Croton caudatus in Saccharomyces cerevisiae model system. Journal of Food
Science and Technology, v. 54, n. 9, p. 2728–2736, 2017.
SUN, K. et al. An exopolysaccharide isolated from a coral-associated fungus and its

100
sulfated derivative activates macrophages. International Journal of Biological
Macromolecules, v. 82, p. 387–394, 2016.
SUN, Z. et al. Sulfation of (1,3)-B-d-glucan from the fruiting bodies of Russula
virescens and antitumor activities of the modifiers. Carbohydrate Polymers, v. 77, n.
3, p. 628–633, 2009.
TANG, C. et al. Immune-enhancing effects of polysaccharides from purple sweet
potato Chao. Journal of Fish Biology, p. 1–34, 2018.
THEIS, T. V. et al. Exocellular (1→6)-β-D-glucan (lasiodiplodan): Carboxymethylation,
thermal behavior, antioxidant and antimicrobial activity. American Journal of
Immunology, v. 13, n. 1, p. 19–33, 2017.
TIDWELL, T. T.; VIKAS, S.; JAGADISH, P. Sulfur Trioxide–Pyridine. 2017.
TONG, X. K. et al. WSS45, a sulfated α-D-glucan, strongly interferes with Dengue 2
virus infection in vitro. Acta Pharmacol Sin, v. 31, n. 5, p. 585–592, 2010.
TÚRMINA, J. A. et al. Toxicological assessment of β-(1→6)-glucan (lasiodiplodan) in
mice during a 28-day feeding study by gavage. Molecules, v. 17, n. 12, p. 14298–
14309, 2012.
VASCONCELOS, A. F. D. et al. Sulfonation and anticoagulant activity of fungal
exocellular β-(1→6)-D-glucan (lasiodiplodan). Carbohydrate Polymers, v. 92, n. 2, p.
1908–1914, 2013.
VASCONCELOS, A. F. D. et al. Three exopolysaccharides of the B-(1,6)-d-glucan type
and a B-(1,3;1,6)-d-glucan produced by strains of Botryosphaeria rhodina isolated from
rotting tropical fruit. Carbohydrate Research, v. 343, n. 14, p. 2481–2485, 2008.
VOGEL, H. J. A convenient growth médium for Neurospora crassa. Genetic Bulletin.,
v. 13, p. 42-47, 1956.

101
WAN-MOHTAR, W. A. A. Q. I. et al. Antimicrobial Properties and Cytotoxicity of
Sulfated (1,3)-β-D-Glucan from the Mycelium of the Mushroom Ganoderma lucidum.
Journal of Microbiology and Biotechnology, v. 26, n. 6, p. 999–1010, 2016.
WANG, Y. et al. Effects of degree of carboxymethylation on physicochemical and
biological properties of pachyman. International Journal of Biological
Macromolecules, v. 51, n. 5, p. 1052–1056, 2012.
WANG, Y. J. et al. A novel process for preparation of (1,3)-B-D-glucan sulphate by a
heterogeneous reaction and its structural elucidation. Carbohydrate Polymers, v. 59,
n. 1, p. 93–99, 2005.
WANG, Z. et al. Sulfated polysaccharides from Cyclocarya paliurus reduce H2O2-
induced oxidative stress in RAW264.7 cells. International Journal of Biological
Macromolecules, v. 80, n. 2015, p. 410–417, 2015.
WANG, Z. J. et al. Carboxymethylation of polysaccharide from Cyclocarya paliurus and
their characterization and antioxidant properties evaluation. Carbohydrate Polymers,
v. 136, n. 2016, p. 988–994, 2016.
WIJAYA, C. J. et al. Cellulose nanocrystals from passion fruit peels waste as antibiotic
drug carrier. Carbohydrate Polymers, v. 175, p. 370–376, 2017.
WOOTTON-BEARD, P. C.; MORAN, A.; RYAN, L. Stability of the total antioxidant
capacity and total polyphenol content of 23 commercially available vegetable juices
before and after in vitro digestion measured by FRAP, DPPH, ABTS and Folin–
Ciocalteu methods. Food Research International, v. 44, n. 1, 217–224, 2011.
XIAO, R.; GRINSTAFF, M. W. Chemical Synthesis of Polysaccharides and
Polysaccharide Mimetics. Progress in Polymer Science, p. 1–132, 2017.
XIE, J. H. et al. Sulfated modification, characterization and antioxidant activities of
polysaccharide from Cyclocarya paliurus. Food Hydrocolloids, v. 53, p. 7–15, 2016.

102
XIE, W. et al. Preparation and antibacterial activity of a water-soluble chitosan
derivative. Carbohydrate Polymers, v. 50, n. 1, p. 35–40, 2002.
XU, J. et al. Carboxymethylation of a polysaccharide extracted from Ganoderma
lucidum enhances its antioxidant activities in vitro. Carbohydrate Polymers, v. 78, n.
2, p. 227–234, 2009.
XU, Y. et al. International Journal of Biological Macromolecules Isolation ,
characterization and bioactivities of the polysaccharides from Dicliptera chinensis ( L
.) Juss . International Journal of Biological Macromolecules, v. 101, p. 603–611,
2017.
YE, M. et al. Phosphorylation and anti-tumor activity of exopolysaccharide from
Lachnum YM120. Carbohydrate Polymers, v. 97, n. 2, p. 690–694, 2013.
ZHANG, G. et al. Purification and antioxidant effect of novel fungal polysaccharides
from the stroma of Cordyceps kyushuensis. Industrial Crops & Products, v. 69, p.
485–491, 2015.
ZHANG, J. et al. Physicochemical characteristics and bioactivities of the
exopolysaccharide and its sulphated polymer from Streptococcus thermophilus GST-
6. Carbohydrate Polymers, v. 146, p. 368–375, 2016.
ZHANG, Y. et al. Modification, characterization and structure-anticoagulant activity
relationships of persimmon polysaccharides. Carbohydrate Polymers, v. 82, n. 2, p.
515–520, 2010.
ZHANG, Y. et al. Sulphated modification of a polysaccharide obtained from fresh
persimmon (Diospyros kaki L.) fruit and antioxidant activities of the sulphated
derivatives. Food Chemistry, v. 127, n. 3, p. 1084–1090, 2011.
ZHU, F.; DU, B.; XU, B. A critical review on production and industrial applications of
beta- glucans. Food hydrocolloids, v. 52, p. 275–288, 2016.

103
8 APÊNDICES
Apêndice 1.
Resultados referentes à análise antimicrobiana (MIC) da amostra de lasiodiplodana
nativa (LAS-N) frente às cepas de leveduras, à esquerda C. albicans e à direita C.
tropicalis. A: 0,26 mg mL-1; B: 0,20 mg mL-1; C: 0,15 mg mL-1; D: 0,10 mg mL-1; E: 0,05
mg mL-1; F: 5 mg mL-1 de fluconazol (controle positivo); G: salina 0,9% (controle
negativo).
Fonte: Autoria própria.
Resultados referentes à análise antimicrobiana (MIC) da amostra de lasiodiplodana
nativa (LAS-N) frente às cepas bactérias Gram positivas, à esquerda S. aureus e à

104
direita L. monocytogenes. A: 0,26 mg mL-1; B: 0,20 mg mL-1; C: 0,15 mg mL-1; D: 0,10
mg mL-1; E: 0,05 mg mL-1; F: 5 mg mL-1 de tetraciclina (controle positivo); G: salina
0,9% (controle negativo).
Fonte: Autoria própria.
Resultados referentes à análise antimicrobiana (MIC) da amostra de lasiodiplodana
nativa (LAS-N) frente às cepas bactérias Gram negativas (S. enterica Typhimurium à
esquerda e E. coli à direita). A: 0,26 mg mL-1; B: 0,20 mg mL-1; C: 0,15 mg mL-1; D:
0,10 mg mL-1; E: 0,05 mg mL-1; F: 5 mg mL-1 de tetraciclina (controle positivo); G:
salina 0,9% (controle negativo).
Fonte: Autoria própria.
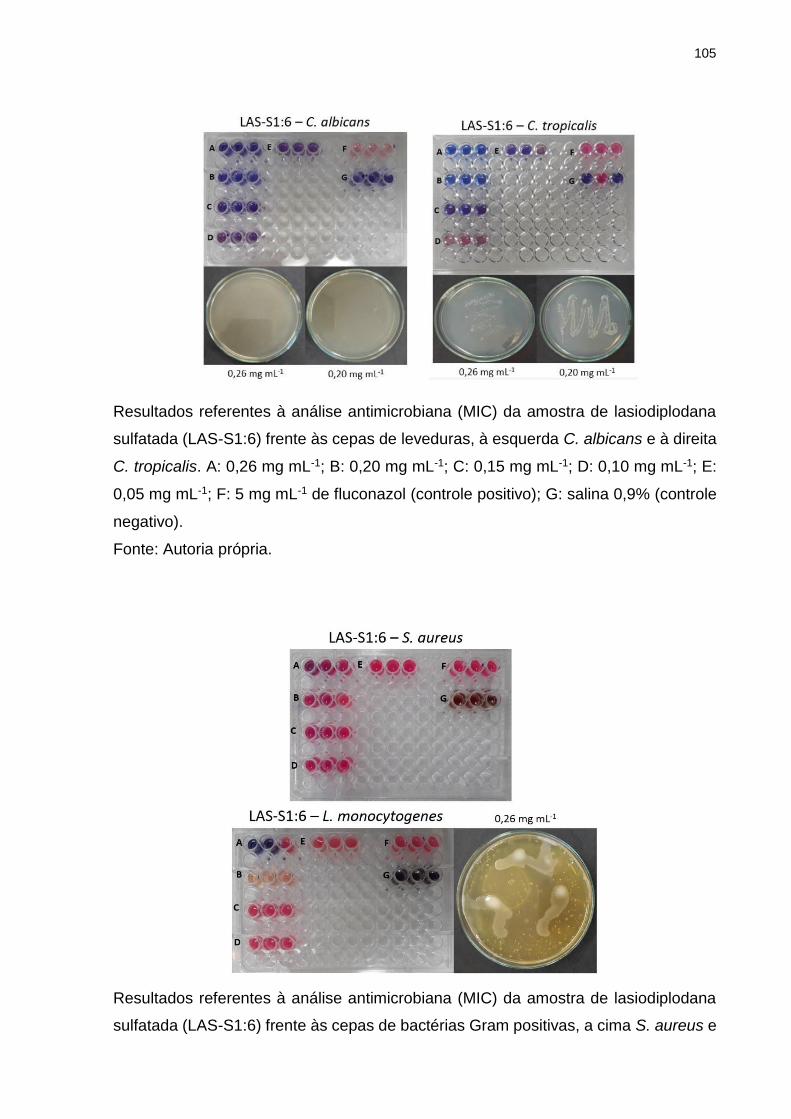
105
Resultados referentes à análise antimicrobiana (MIC) da amostra de lasiodiplodana
sulfatada (LAS-S1:6) frente às cepas de leveduras, à esquerda C. albicans e à direita
C. tropicalis. A: 0,26 mg mL-1; B: 0,20 mg mL-1; C: 0,15 mg mL-1; D: 0,10 mg mL-1; E:
0,05 mg mL-1; F: 5 mg mL-1 de fluconazol (controle positivo); G: salina 0,9% (controle
negativo).
Fonte: Autoria própria.
Resultados referentes à análise antimicrobiana (MIC) da amostra de lasiodiplodana
sulfatada (LAS-S1:6) frente às cepas de bactérias Gram positivas, a cima S. aureus e

106
abaixo L. monocytogenes. A: 0,26 mg mL-1; B: 0,20 mg mL-1; C: 0,15 mg mL-1; D: 0,10
mg mL-1; E: 0,05 mg mL-1; F: 5 mg mL-1 de tetraciclina (controle positivo); G: salina
0,9% (controle negativo).
Fonte: Autoria própria.
Resultados referentes à análise antimicrobiana (MIC) da amostra de lasiodiplodana
sulfatada (LAS-S1:6) frente às cepas de bactérias Gram negativas, à esquerda S.
enterica Typhimurium e à direita E. coli. A: 0,26 mg mL-1; B: 0,20 mg mL-1; C: 0,15 mg
mL-1; D: 0,10 mg mL-1; E: 0,05 mg mL-1; F: 5 mg mL-1 de tetraciclina (controle positivo);
G: salina 0,9% (controle negativo).
Fonte: Autoria própria.
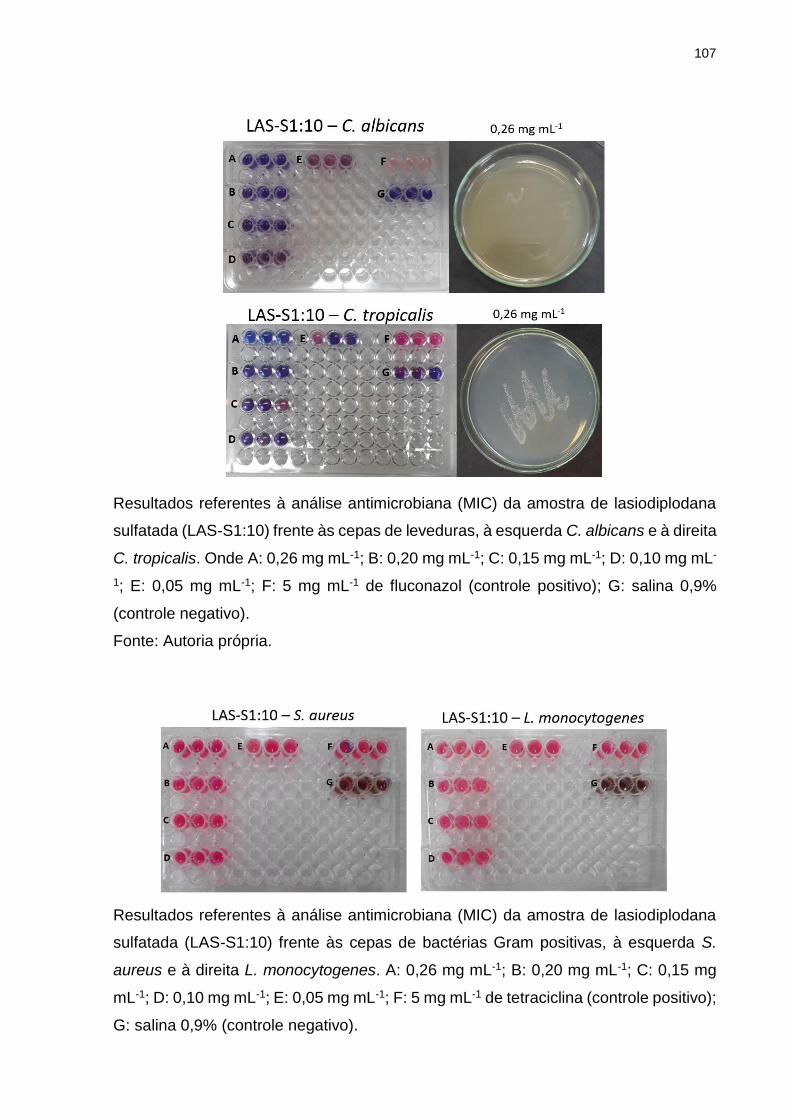
107
Resultados referentes à análise antimicrobiana (MIC) da amostra de lasiodiplodana
sulfatada (LAS-S1:10) frente às cepas de leveduras, à esquerda C. albicans e à direita
C. tropicalis. Onde A: 0,26 mg mL-1; B: 0,20 mg mL-1; C: 0,15 mg mL-1; D: 0,10 mg mL-
1; E: 0,05 mg mL-1; F: 5 mg mL-1 de fluconazol (controle positivo); G: salina 0,9%
(controle negativo).
Fonte: Autoria própria.
Resultados referentes à análise antimicrobiana (MIC) da amostra de lasiodiplodana
sulfatada (LAS-S1:10) frente às cepas de bactérias Gram positivas, à esquerda S.
aureus e à direita L. monocytogenes. A: 0,26 mg mL-1; B: 0,20 mg mL-1; C: 0,15 mg
mL-1; D: 0,10 mg mL-1; E: 0,05 mg mL-1; F: 5 mg mL-1 de tetraciclina (controle positivo);
G: salina 0,9% (controle negativo).

108
Fonte: Autoria própria.
Resultados referentes à análise antimicrobiana (MIC) da amostra de lasiodiplodana
sulfatada (LAS-S1:10) frente às cepas de bactérias Gram negativas, à esquerda S.
enterica Typhimurium e à direita E. coli. A: 0,26 mg mL-1; B: 0,20 mg mL-1; C: 0,15 mg
mL-1; D: 0,10 mg mL-1; E: 0,05 mg mL-1; F: 5 mg mL-1 de tetraciclina (controle positivo);
G: salina 0,9% (controle negativo).
Fonte: Autoria própria.
Resultados referentes à análise antimicrobiana (MIC) da amostra de lasiodiplodana
sulfatada (LAS-S1:4) frente às cepas de leveduras, à esquerda C. albicans e à direita
C. tropicalis. A: 0,26 mg mL-1; B: 0,20 mg mL-1; C: 0,15 mg mL-1; D: 0,10 mg mL-1; E:
0,05 mg mL-1; F: 5 mg mL-1 de fluconazol (controle positivo); G: salina 0,9% (controle
negativo).
Fonte: Autoria própria.
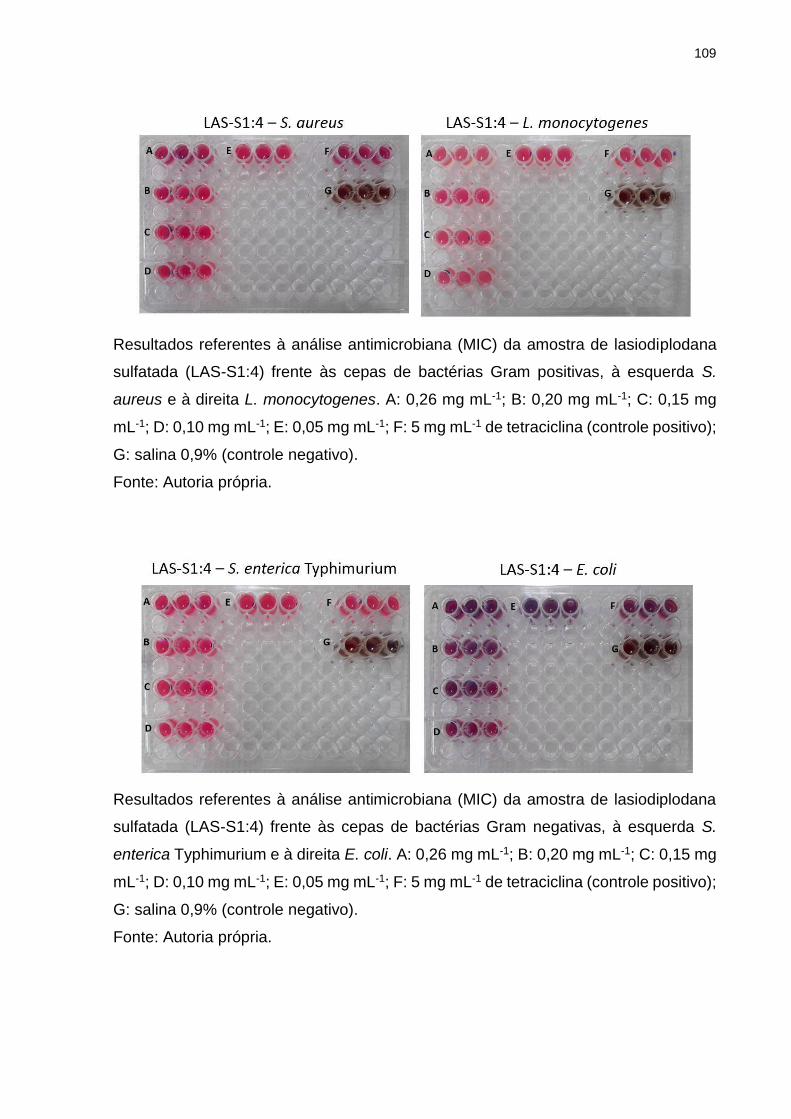
109
Resultados referentes à análise antimicrobiana (MIC) da amostra de lasiodiplodana
sulfatada (LAS-S1:4) frente às cepas de bactérias Gram positivas, à esquerda S.
aureus e à direita L. monocytogenes. A: 0,26 mg mL-1; B: 0,20 mg mL-1; C: 0,15 mg
mL-1; D: 0,10 mg mL-1; E: 0,05 mg mL-1; F: 5 mg mL-1 de tetraciclina (controle positivo);
G: salina 0,9% (controle negativo).
Fonte: Autoria própria.
Resultados referentes à análise antimicrobiana (MIC) da amostra de lasiodiplodana
sulfatada (LAS-S1:4) frente às cepas de bactérias Gram negativas, à esquerda S.
enterica Typhimurium e à direita E. coli. A: 0,26 mg mL-1; B: 0,20 mg mL-1; C: 0,15 mg
mL-1; D: 0,10 mg mL-1; E: 0,05 mg mL-1; F: 5 mg mL-1 de tetraciclina (controle positivo);
G: salina 0,9% (controle negativo).
Fonte: Autoria própria.
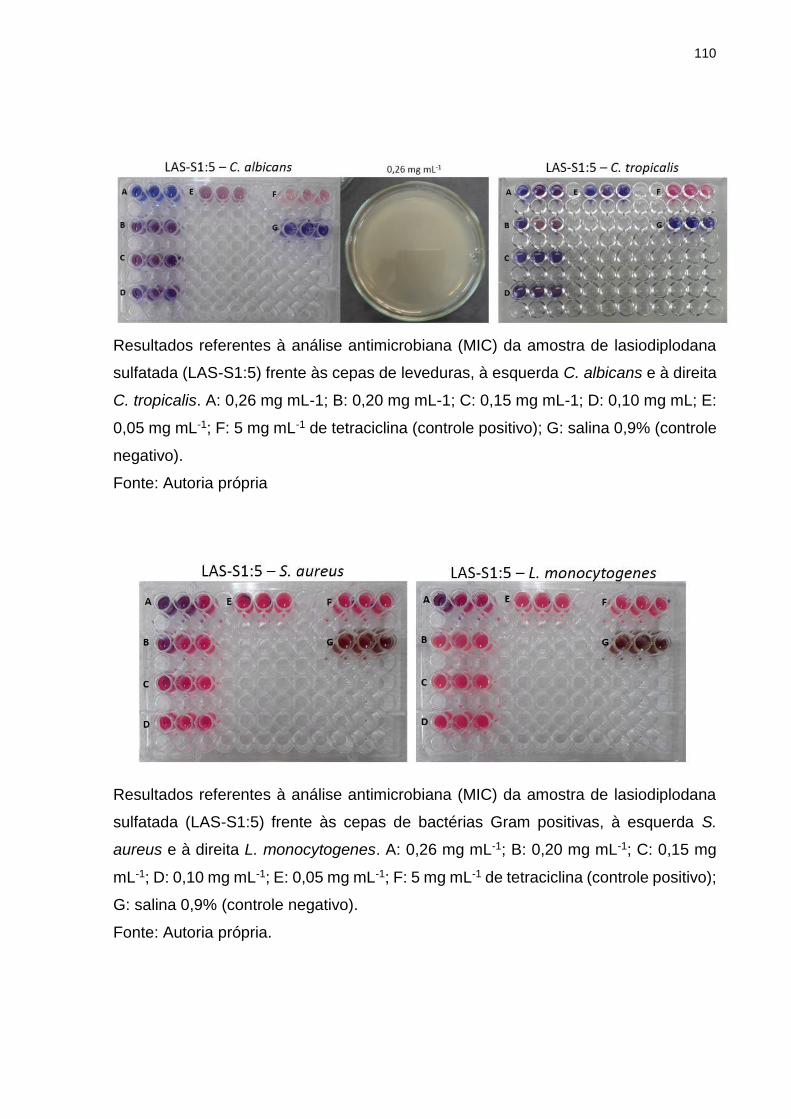
110
Resultados referentes à análise antimicrobiana (MIC) da amostra de lasiodiplodana
sulfatada (LAS-S1:5) frente às cepas de leveduras, à esquerda C. albicans e à direita
C. tropicalis. A: 0,26 mg mL-1; B: 0,20 mg mL-1; C: 0,15 mg mL-1; D: 0,10 mg mL; E:
0,05 mg mL-1; F: 5 mg mL-1 de tetraciclina (controle positivo); G: salina 0,9% (controle
negativo).
Fonte: Autoria própria
Resultados referentes à análise antimicrobiana (MIC) da amostra de lasiodiplodana
sulfatada (LAS-S1:5) frente às cepas de bactérias Gram positivas, à esquerda S.
aureus e à direita L. monocytogenes. A: 0,26 mg mL-1; B: 0,20 mg mL-1; C: 0,15 mg
mL-1; D: 0,10 mg mL-1; E: 0,05 mg mL-1; F: 5 mg mL-1 de tetraciclina (controle positivo);
G: salina 0,9% (controle negativo).
Fonte: Autoria própria.

111
Resultados referentes à análise antimicrobiana (MIC) da amostra de lasiodiplodana
sulfatada (LAS-S1:5) frente às cepas de bactérias Gram negativas, à cima S. enterica
Typhimurium, e a baixo E. coli. A: 0,26 mg mL-1; B: 0,20 mg mL-1; C: 0,15 mg mL-1; D:
0,10 mg mL-1; E: 0,05 mg mL-1; F: 5 mg mL-1 de tetraciclina (controle positivo); G:
salina 0,9% (controle negativo).
Fonte: Autoria própria.

112
Apêndice 2
Análise de citotoxicidade da lasiodiplodana nativa e derivados sulfatados sobre a S.
cerevisiae.
Fonte: Autoria própria.
Top Related