
Zootaxa,Review of the Neotropical blackfly subgenus...
96
ZOOTAXA Review of the Neotropical blackfly subgenus Inaequalium Coscarón & Wygodzinsky (Diptera: Simuliidae) based on adults and pupal morphology LUIS MIGUEL HERNÁNDEZ, ANTHONY JOHN SHELLEY, ANTONIO PAULINO ANDRADE DE LUNA DIAS & MARILZA MAIA-HERZOG Magnolia Press Auckland, New Zealand 1649 TERM OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.
Transcript of Zootaxa,Review of the Neotropical blackfly subgenus...

ZOOTAXA
Review of the Neotropical blackfly subgenus Inaequalium Coscarón & Wygodzinsky (Diptera: Simuliidae) based on adults and pupal morphology
LUIS MIGUEL HERNÁNDEZ, ANTHONY JOHN SHELLEY, ANTONIO PAULINO ANDRADE DE LUNA DIAS & MARILZA MAIA-HERZOG
Magnolia PressAuckland, New Zealand
1649
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.2 · Zootaxa 1649 © 2007 Magnolia Press
LUIS MIGUEL HERNÁNDEZ, ANTHONY JOHN SHELLEY, ANTONIO PAULINO ANDRADE DE
LUNA DIAS & MARILZA MAIA-HERZOGReview of the Neotropical blackfly subgenus Inaequalium Coscarón & Wygodzinsky (Diptera: Simuli-idae) based on adults and pupal morphology(Zootaxa 1649)
96 pp.; 30 cm.
28 November 2007
ISBN 978-1-86977-167-6 (paperback)
ISBN 978-1-86977-168-3 (Online edition)
FIRST PUBLISHED IN 2007 BY
Magnolia Press
P.O. Box 41-383
Auckland 1346
New Zealand
e-mail: [email protected]
http://www.mapress.com/zootaxa/
© 2007 Magnolia Press
All rights reserved.
No part of this publication may be reproduced, stored, transmitted or disseminated, in any form, or by any
means, without prior written permission from the publisher, to whom all requests to reproduce copyright
material should be directed in writing.
This authorization does not extend to any other kind of copying, by any means, in any form, and for any purpose
other than private research use.
ISSN 1175-5326 (Print edition)
ISSN 1175-5334 (Online edition)
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Accepted by J. Moulton: 19 Sept. 2007; published: 28 Nov. 2007 3
ZOOTAXAISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2007 · Magnolia Press
Zootaxa 1649: 1–96 (2007) www.mapress.com/zootaxa/
Review of the Neotropical blackfly subgenus Inaequalium Coscarón & Wygodzinsky (Diptera: Simuliidae) based on adults and pupal morphology
LUIS MIGUEL HERNÁNDEZ1, ANTHONY JOHN SHELLEY1, ANTONIO PAULINO ANDRADE DE
LUNA DIAS 2 & MARILZA MAIA-HERZOG 2
1 Diptera Division, Simuliidae and Onchocerciasis Research Programme, Department of Entomology, The Natural History Museum, Cromwell Road, SW7 5BD, London, UK; [email protected] Laboratório de Simulídeos e Oncocercose (LSO), Instituto Oswaldo Cruz, Rio de Janeiro, Brasil
Table of contents
Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3Materials & Methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4The subgenus Inaequalium . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
Diagnosis of the subgenus Inaequalium . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12Key to species of Inaequalium . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13Species descriptions, distribution and biology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15Species inquirendae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 51Unavailable names . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 54
Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 55References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 55Material Examined . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 80Appendix 1. Diagnosis of the subgenus Inaequalium by various authors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 95
Abstract
The species of the subgenus Inaequalium are reviewed based on adult and pupal morphology. All main taxonomic char-acters are fully illustrated together with a key to pupae for species identification. Discussions on the species’ taxonomyand brief summaries of their distribution and biology are also provided. As a result of this study, 14 species are nowincluded in the subgenus Inaequalium. They are placed into two species groups, the botulibranchium species group withthree species and the inaequale species group with 11 species, including two that are treated as species inquirendae: S.lurybayae Smart and S. parimaense Ramírez-Pérez, Yarzábal, Takaoka, Tada & Ramírez. Simulium lurybayae Smart, S.margaritatum Pepinelli, Hamada & Luz and S. maranguapense (Pessoa, Ríos-Velásquez & Py-Daniel) are now placed inthe inaequale species group. Two lectotypes are designated for the following species: S. clavibranchium Lutz and S.diversibranchium Lutz. Simulium hauseri (Coscarón & Coscarón-Arias) is regarded as an unavailable name.
Key words: Simuliidae, black flies, Neotropical region, taxonomy, genus Simulium, subgenus Inaequalium
Introduction
Species of the subgenus Inaequalium Coscarón & Wygodzinsky are widely distributed in the NeotropicalRegion, extending from Central America (Panama) to northeast Argentina (Coscarón 1987, 1991; Juñent &Coscarón, 2001). At present this subgenus includes 14 valid species (present paper), with the vast majority of
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.4 · Zootaxa 1649 © 2007 Magnolia Press
taxa commonly found in the coastal, mountainous areas of southern Brazil (Crosskey & Howard 2004;Coscarón 1991; Coscarón & Coscarón-Arias 1997, 2007; Pepinelli et al. 2006; Pessoa et al. 2005; Strieder,2004). Some of the species in this subgenus are potentially medically important because of their highanthropophily in northern Brazil and Guyana, and their presence in the endemic focus of pemphigus foliaceus(fogo selvagem) in the state of Mato Grosso, Brazil, e.g. S. inaequale (Eaton et al., 1998; Shelley et al., 2004).
The subgenus Inaequalium was erected by Coscarón & Wygodzinsky (1984) to accommodate 10 speciesmorphologically similar to ones in the subgenus Psaroniocompsa Enderlein. The adults of most of the speciesin Inaequalium are morphologically similar, and species recognition relies mainly on the configuration of thepupal gill filaments. Coscarón (1987) divided the 15 species (excluding three species inquirendae) that herecognized in this subgenus into two groups: the inaequale group with 12 species and the botulibranchiumgroup with three species. Crosskey & Howard’s list of Simuliidae (2004) included 17 species in the subgenusInaequalium, which was divided into the botulibranchium and inaequale species groups, with four and 13species, respectively. These authors did not assign S. pseudoexiguum Nunes de Mello & Barbosa de Almeidato any species group. More recently, two species of Inaequalium described from Brazil were not included inany species group in the original descriptions because they shared characters of species in both the inaequaleand botulibranchium species groups. These species are S. maranguapense (Pessoa, Ríos-Velásquez & Py-Daniel) [described as Inaequalium maranguapense by Pessoa et al. (2005) from Ceará State], and S.margaritatum Pepinelli, Hamada & Luz collected in the state of Bahía (Pepinelli et al. 2006). The taxonomicposition of the subgenus Inaequalium is controversial because several authors, for example Py-Daniel &Moreira Sampaio (1994a, b; 1995) and Strieder & Py-Daniel (2000, 2002), considered it a genus, whileCoscarón (1987, 1991) and Crosskey & Howard (2004) regarded it as a subgenus within Simulium Latreille.Coscarón & Coscarón-Arias (2007) listed 14 species in Inaequalium, neglecting to include the three species ofthe botulibranchium group. They also omitted in this total the two recently published species cited in theiraddendum and three species inquirendae.
This paper provides a biosystematic study of the species of the subgenus Inaequalium in the NeotropicalRegion that includes species descriptions for females, males and pupae, and a key for identification of thepupal stage. All main taxonomic characters are fully illustrated and notes on the distribution and basic biologyare given where details are available.
Materials & Methods
The techniques for collection, rearing, measurement of specimens and terminology used are those detailed byShelley et al. (1997). Dissections of specimens were carried out using a modified technique detailed inHernández & Shelley (2005) and Hernández et al. (2005). Only adults obtained from pupae and, in somecases, man-biting specimens have been used in this paper and they are listed in Material Examined. Timeconstraints and unavailability of larval material precluded the description of this stage. Therefore, the readersshould consult the review papers of Coscarón & Coscarón-Arias (2007), Coscarón & Wygodzinsky (1984),Hernández et al. (2006), Pepinelli et al. (2006), Pessoa et al. (2005) and Strieder & Py-Daniel (1999, 2000)for descriptions of the larval stage, when available. The larva is unknown for two species considered here asspecies inquirendae: S. lurybayae Smart and S. parimaense Ramírez-Pérez, Yarzábal, Takaoka, Tada &Ramírez. Some digital images here presented (see Figs. 49, 50, 53, 54) are of inferior quality. This is becausethe first author of this paper only had access to a low-resolution digital camera during a visit to INPA toexamine type material of Simuliidae under the care of Victor Py-Daniel. However, we have decided to includethese photographs in the paper because they illustrate the general morphology of type material for some of thespecies described in this paper, which are not easily accessible for study.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 5NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
Our analysis of the subgenus Inaequalium has been based on type material wherever possible and for manyspecies from Brazil we have examined long series of specimens collected by us in order to assess intraspecificvariation. In all other cases when no specimens were available for examination we have relied on descriptionsand figures produced by other authors. Many of these were made when the taxonomy of the NeotropicalSimuliidae was less advanced and hence some of our key characters were not described and a certain amountof calculated guesswork has been used. We have placed less emphasis on larval morphology because of thehomogeneity of Inaequalium species in this life stage (see also Strieder & Py-Daniel, 2000), the relativedearth of modern descriptions, and the lack of material in our collections.
All images illustrating the morphology were obtained directly from specimens using a Synoptics digitalimaging system at The Natural History Museum, London (BMNH) as detailed in Hernández & Shelley (2005)and Hernández et al. (2005). We follow the classification of Crosskey & Howard (2004) for the placement ofInaequalium as a subgenus of Simulium Latreille. The following acronyms are used for depositories of speci-mens referred to in this paper in the text and under Material Examined.
AMNH American Museum of Natural History, New York, USABMNH Department of Entomology, Natural History Museum, London, United
KingdomDERM* Laboratório de Entomología de la División de Endemias Rurales, Mara-
cay, Aragua State, VenezuelaICBUSP Coleção Entomológica do Departamento de Parasitologia do Instituto de
Ciências Biomédicas da Universidade de São Paulo, BrazilIDVC* Instituto de Dermatología, Villa de Cura, Aragua, Venezuela [Previously
recorded as IND, Instituto Nacional de Dermatología, Villa de Cura, Ara-gua in Shelley et al., 1997]
INPA Instituto Nacional de Pesquisas da Amazônia, Manaus, BrazilIOC Laboratório de Simuliidae e Oncocercose (LSO), Instituto Oswaldo Cruz,
Rio de Janeiro, BrazilLEU Laboratório de Entomologia, Centro de Ciências da Saúde, Universidade
do Vale do Rio dos Sinos, São Leopoldo, Rio Grande do Sul, BrazilMCNFZ Museu de Ciências Naturais da Fundação Zoobotânica do Rio Grande do
Sul, Porto Alegre, BrazilMEPG* Museo Entomológico “Dr Pablo Garcia”, Instituto de Altos Estudios en
Salúd Pública “Dr Arnoldo Gabaldón”, Maracay, Estado Aragua, Vene-zuela
MCZ Museum of Comparative Zoology, Harvard, USAMLP Museo de Ciencias Naturales, La Plata, ArgentinaMZUSP Museu de Zoologia, Universidade de São Paulo, BrazilNM Naturhistoriches Museum, Vienna (Wien), AustriaNMHU Museum für Naturkunde der Humboldt-Universität, Berlin, Germany
[Previously recorded as Zoologisches Museum der Humboldt-Universität(ZM) by Shelley et al. (1989) and ZMHU in Shelley et al. (1997)]
SMT Staatliche Museum für Tierkunde, Dresden, GermanyUSNM [also NMNH] National Museum of Natural History, Washington, USA [Also previously
recorded as United States National Museum of Natural History (USNM)in Shelley et al. (1997)]
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.6 · Zootaxa 1649 © 2007 Magnolia Press
*Note on Jaime Ramírez Pérez Simuliidae collection. Jaime Ramírez Pérez comprehensively studied theSimuliidae of Venezuela during the 1970s and 1980s, and compiled all the species taxonomic and biologicalinformation in his latest monograph in 1983 “Los Jejénes de Venezuela”. The types of all species that hedescribed as well as non-type material are recorded in the literature as being deposited either in his privatecollection or in the collections of DERM and IDVC. These two institutions were within the same organiza-tion, which ceased to exist four years ago. The Simuliidae collection of Ramírez-Pérez was kept in his housefor many years, but has now been donated to the MEPG. The collection is being reviewed by Hortencia Fron-tado under the supervision of Maria Eugenia Grillet and the situation is as follows: the collection holds somespecimens in alcohol, but mainly consists of pinned and slide material. Unfortunately, it was not previouslykept in good condition and all the alcohol vials have now dried. However, the proportion of specimens kept inalcohol was minimal. The pinned material is also in a very poor condition. Currently, the slide collection isbeing examined and some of the slides are in good condition, particularly the preparations containing imma-ture stages. It is not clear at this juncture how many type specimens are in the collection because specimensgenerally do not have type labels, although they are often from the type locality (Maria Eugenia Grillet, pers.comm. to L. M. Hernández). Recent information provided by Hortencia Frontado to Luis Hernández has con-firmed that the type of S. parimaense is not present in the Ramírez-Pérez Simuliidae collection.
The subgenus Inaequalium
Literature reviewIn this section we summarize the previous work on the subgenus Inaequalium followed by our own
conclusions and our proposed grouping of species within this subgenus. We have largely drawn uponCoscarón’s treatment of the Neotropical fauna (1987) and based our arrangement on Crosskey’s classificationin his annotated world list (Crosskey & Howard, 2004).
Previous work Coscarón (1980) during a revision of some Simuliidae species inhabiting the coastal region of Brazil
redescribed S. botulibranchium Lutz and described two new species, S. souzalopesi and S. petropoliense. Inthis paper, he mentioned that these species “constituted a small group with an affinity to the subgenusInaequalium”. The three species described in the paper were similar to species of Inaequalium in adultcoloration, setation of the thorax and abdominal chaetotaxy of the pupa as well as the relative proportions ofthe antennal segments, and the form of the hypostomium in the larva. Differences in the form of the femaleparaproct, gonocoxite (as basímero), gonostyle (as distímero) and ventral plate of the male and also the pupalgill form precluded their formal inclusion in this subgenus. However, it was suggested that these speciesmight possibly be included as a separate group within Inaequalium. Unfortunately, he failed to record thespecies present in the subgenus Inaequalium or give a diagnosis for this subgenus. As the name Inaequaliumwas published with no named species, designation of type species or subgeneric diagnosis Crosskey (1988)considered Inaequalium as an unavailable name [see also International Code of Zoological nomenclature(1999), articles 11 and 13].
The subgenus Inaequalium was formally described by Coscarón & Wygodzinsky in 1984 in their revisionof the species of the subgenera Psaroniocompsa and Inaequalium. In this paper, the authors described allmorphological characters necessary to recognize the species in this subgenus, selected Eusimulium inaequale(described by Paterson & Shannon in 1927) as the type species of Inaequalium, and presented a cladrogramoutlining the phylogenetic relationships between the species that they assigned to Inaequalium. Othertaxonomic features in this paper are as follows: S. mariavulcanoae was described as a new species; twospecies, S. baiense Pinto and S. pseudoexiguum Nunes de Mello & Barbosa de Almeida (see also Table 1)
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 7NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
were included as species inquirendae; and five new synonyms were proposed for S. inaequale (=Trichodagmia manicata Enderlein, S. jundiaiensis D’Andretta & González B), S. subnigrum Lutz (= S.diversifurcatum Lutz, S. mbarigui Coscarón) and S. diversibranchium Lutz (= S. missionum Coscarón).
Coscarón & Wygodzinsky (1984) recognized the difficulty in identifying the adults of Inaequalium andPsaroniocompsa species and stated that “…both subgenera are composed of morphologically similarsympatric species…” (see also Appendix 1 for more details). They had reviewed both subgenera togetherbecause of the similarity in coloration of the thorax (black laterally and grey posteriorly) and abdomen ofadults, female paraprocts short and wider than long, pupal gill with six long, fine filaments (exceptionally fouror 2–3 thick filaments), and similar coloration and morphology of the hypostomium, mandibular teeth, andanal gills in the larva. They concluded that both subgenera must have shared a common lineage, but stillregarded Psaroniocompsa and Inaequalium as separate taxa. The authors also provided a table to distinguishInaequalium from Psaroniocompsa. Their main characters to distinguish Inaequalium were as follows: adultswith hairs on basal section of R; female with gonapophyses distally narrow and with or without scatteredmicrotrichiae and internal margins divergent and weakly sclerotized; male with subtriangular tosubtrapezoidal gonostyle with terminal spine; pupa with trifid to quadrifid trichomes; larva with cephalicapotome “not always positive”, “not very distinct”, sometimes delimiting a subbasal, median clearer window;larval third antennal segment much longer than first and second segments. In the same paper the authors(Coscarón & Wygodzinsky, 1984) also emphasized the variation in size, adult thoracic pubescence and thepupal gill configuration in these two subgenera. Their species group analysis of Inaequalium separated“…with certain effort…” S. travassosi from the other species because of its pupa with three filaments anddifferent proportions of the genital fork.
Several species now considered to belong to Inaequalium had been previously placed in differentsubgenera within Simulium. The major changes are listed in Table 1, with a detailed diagnosis for thesubgenus by different authors given in Appendix 1. The first comprehensive paper on Inaequalium species isthat of Coscarón & Wygodzinsky (1984) who provided species descriptions, but did not allocate them tospecies groups. Simulium nahimi, described by Py-Daniel (1984) from Mato Grosso State in Brazil, wasassigned by the author to Inaequalium. Coscarón in 1987 produced the most comprehensive treatment of thesubgenera and species of Simuliidae in the Neotropical Region. He divided species of the subgenusInaequalium into two species groups, the inaequale group (11 species) and botulibranchium group (threespecies) based on adult genitalia, pupal gill configuration and girth, and morphology of the larvalhypostomium. He also placed S. baiense, S. lurybayae and S. pseudoexiguum as species inquirendae.However, Crosskey (1988) in his world list of Simulium species apparently followed Coscarón &Wygodzinsky (1984) in their concept of Inaequalium, but considered S. nahimi as a member of the subgenusCerqueirellum Py-Daniel [now a synonym of Psaroniocompsa], and left the following species “unassigned tosubgenus”: S. botulibranchium, S. clarki, S. lurybayae, S. nogueirai D’Andretta & González, S. petropoliense,S. rappae Py-Daniel & Coscarón, and S. souzalopesi. In both publications, the name of S. parimaense[described by Ramírez-Pérez et al. (1986) from Venezuela as S. parimaensis] is missing from the species listin Simulium. A further species, S. nogueirai, described by D’Andretta & González B. (1964), was presumedmisplaced by Py-Daniel & Moreira (1989), who redescribed it and assigned it to the subgenus Inaequalium.These authors also added some morphological characters to recognize the members of this subgenus (seeAppendix 1).
Coscarón (1991) reviewed all subgenera and species occurring in the “Cono Sur” region of South America[Argentina, Chile, southern Bolivia, Paraguay, Uruguay and S.E. Brazil] and provided a diagnosis ofInaequalium (see Table 1, Appendix 1). He presented a key to separate the adults and pupal stages for 15species of Inaequalium [not 14 as cited in the paper] in the Neotropical Region, but only fully treated thosespecies (11 in total) that occur in the Cono Sur. However, he did not include in the key S. pseudoexiguum andS. nogueirai, despite stating that it included all species for the Neotropical region. The major changes in this
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.8 · Zootaxa 1649 © 2007 Magnolia Press
paper are as follows: he only recorded Inaequalium extending from Venezuela to Argentina [S. clarkisynonymized with S. inaequale by Hernández et al. (2007) occurs in Panama]; S. petropoliense and S.parimaense were not included in the key to females; he assigned S. lurybayae to the botulibranchium speciesgroup; and regarded S. baiense as a junior synonym of S. inaequale.
TABLE 1. Previous assignment of Simulium species now considered to belong to the subgenus Inaequalium.
Authors Subgenus Species group Species
Coscarón & Wygodzin-sky (1984)
Inaequalium–
Species inquirendae
beaupertuyi, clavibranchium, diversibranchium, inae-quale, mariavulcanoae, subclavibranchium, subnigrum, travassosi
baiense, pseudoexiguum
Coscarón (1987) Inaequalium botulibranchium group
inaequale group
Species inquirendae
botulibranchium, petropoliense, souzalopesi
beaupertuyi, clavibranchium, clarki, diversibranchium, inaequale, mariavulcanoae, nahimi , nogueirai, rappae, subclavibranchium, subnigrum, travassosi
baiense, lurybayae, pseudoexiguum
Crosskey (1988) Inaequalium
Cerqueirellum
Species unassigned to genus
–
–
–
baiense, beaupertuyi, clavibranchium, diversibranchium, inaequale, mariavulcanoae, pseudoexiguum, subclavi-branchium, subnigrum, travassosi
nahimi
botulibranchium, clarki, lurybayae, nogueirai, petropo-liense, rappae, souzalopesi
Coscarón (1991) [The author only fully treated 11 species for the Cono Sur]
Inaequalium botulibranchium group
inaequale group
botulibranchium, lurybayae , petropoliense, souzalopesi
beaupertuyi, clavibranchium, clarki, diversibranchium, inaequale, mariavulcanoae, nahimi, rappae, subclavi-branchium, subnigrum, travassosi
Py-Daniel & Moreira (1995) [Only for Brazilian fauna]
Inaequalium [As Genus]
-botulibranchium, clavibranchium, diversibranchium, inaequale, mariavulcanoae, nahimi, nogueirai, petropo-liense, pseudoexiguum, rappae, souzalopesi, subclavi-branchium, subnigrum, travassosi
Crosskey & Howard (1997) [This world inventory developed from the checklist of Crosskey (1988)]
Inaequalium
Psaroniocompsa
botulibranchium group
inaequale group
Inaequalium species unplaced to group
Psaroniocompsa species unplaced to group
botulibranchium, lurybayae, petropoliense, souzalopesi
beaupertuyi, clarki, clavibranchium, diversibranchium, inaequale, mariavulcanoae, nahimi, nogueirai, rappae, subclavibranchium, subnigrum, travassosi
pseudoexiguum
parimaense
..... continued on the next page
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 9NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
Py-Daniel & Moreira Sampaio (1994a) proposed recognition of only two higher categories in Simuliidae,and they elevated all subgenera to genera. This action was then applied by Py-Daniel & Moreira Sampaio
TABLE 1 (continued)
Authors Subgenus Species group Species
Coscarón & Coscarón Arias (1997)
Inaequalium botulibranchium group
inaequale group
Species names men-tioned but not analyzed Because of lack of mate-rial
botulibranchium, petropoliense, souzalopesi
clarki, clavibranchium, diversibranchium, inaequale, mariavulcanoae, nahimi, parimaensis, rappae, subclavi-branchium, subnigrum, travassosi
nogueirai, beaupertuyi [related to S. subnigrum]clarki, lurybayae [related to S. inaequale]
Strieder & Py-Daniel (1999, 2000)
Inaequalium [As Genus]
-
Species inquirendae
beaupertuyi, botulibranchium, clavibranchium, diversi-branchium, inaequale, mariavulcanoae, nahimi, noguei-rai, rappae, subclavibranchium, subnigrum, travassosi
clarki, lurybayae, parimaense, pseudoexiguum
Crosskey & Howard (2004) [First published by Crosskey & Howard (1997) followed by two updates Crosskey (1999, 2002).]
Inaequalium
Psaroniocompsa
botulibranchium group
inaequale group
Species unplaced to group
Psaroniocompsa species unplaced to group
botulibranchium, lurybayae, petropoliense, souzalopesi
beaupertuyi, clavibranchium, clarki, diversibranchium, inaequale, leopoldense, mariavulcanoae, nahimi, noguei-rai, rappae, subclavibranchium, subnigrum, travassosi
pseudoexiguum
parimaense
Strieder & Py-Daniel (2002)
Inaequalium [As Genus]
botulibranchium group
inaequale group
Species not analyzed as uncertain taxonomic situ-ation or insufficient data
botulibranchium, petropoliense, souzalopesi, travassosi
beaupertuyi, clavibranchium, diversibranchium, inae-quale, leopoldense, mariavulcanoae, nahimi, nogueirai, rappae, subclavibranchium, subnigrum
clarki, lurybayae, parimaense, pseudoexiguum
Coscarón & Coscarón Arias (2007)[Species list as in Coscarn (1991, 1997) except for two addi-tional species]
Inaequalium Inaequale group hauseri, leopoldense
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.10 · Zootaxa 1649 © 2007 Magnolia Press
(1995) to the Brazilian fauna [see also Py-Daniel & Moreira Sampaio (1994b) for new specific name changesin Simuliidae] and included only 14 species for this country. However, this system has not been followed byCrosskey & Howard (1997) in their “World Inventory of Simuliidae” and three subsequent updates thatfollowed (Crosskey, 1999, 2002; Crosskey & Howard, 2004). It is evident that the latter authors followed theprevious papers of Coscarón (1987, 1991) for the classification of Inaequalium and treated S. pseudoexiguumas “species unplaced to group”. However, they regarded S. parimaense as a “species unplaced to group” in thesubgenus Psaroniocompsa. In a subsequent paper Coscarón & Miranda-Esquivel (1998) published a scientificnote dealing with synonyms of S. inaequale. They maintained those proposed in Coscarón & Wygodzinsky(1984) and Coscarón (1991) for S. inaequale, and also synonymized Thyrsopelma argentata Enderlein withthis species.
Since then species in Inaequalium have received a lot of attention and several authors have publishedreviews and phylogenetic analyses independently. Coscarón & Coscarón-Arias (1997) produced aphylogenetic analysis of 13 species of the subgenus Inaequalium together with the subgenera Chirostilbia andPsaroniocompsa. For Inaequalium they used the subgenus Notolepria as the outgroup. They did not includein their analysis S. beaupertuyi Ramírez-Pérez, Rassi & Ramírez, S. nogueirai [related to S. subnigrum], andS. clarki, S. lurybayae [related to S. inaequale] because of lack of material. However, in a distribution map forall species in Inaequalium they figured the distribution of S. clarki and S. beaupertuyi. They recognized twospecies groups: the botulibranchium and inaequale species groups. Within the inaequale group, S.parimaense formed an independent clade, and they concluded that S. travassosi and S. rappae formed asubgroup within the inaequale group. Strieder et al. (1992) provided a key to identify those species found inthe state of Rio Grande Sul and treated Inaequalium as a genus. Strieder and Py-Daniel (1999, 2000) alsoregarded Inaequalium as a genus and studied those characters that define this taxon based on immature stages.In their latest paper Strieder & Py-Daniel (2000) described S. leopoldense [as I. leopoldense] based only onlarvae and pupae from São Leopoldo, Rio Grande do Sul. Another phylogenetic tree was constructed by thesame authors, Strieder & Py-Daniel (2002), in which they only recognized two species groups as before, andtwo subgroups within the inaequale group. In this paper S. travassosi was transferred from the inaequale tothe botulibranchium species group. More recently, in a paper on the Simuliidae of Belize Shelley et al. (2002)discussed the taxonomic position of S. quadrivittatum Loew, and mistakenly attributed its placement in thesubgenus Inaequalium to Coscarón (1987). In the latter paper the quadrivittatum group followed on from thesubgenus Inaequalium, but was considered by the author to have subgeneric rank and the term speciessubgroup was used in place of species group. In subsequent publications the author used the universallyaccepted definition for species group.
Two new species have been recently described that pertain to Inaequalium: S. maranguapense and S.margaritatum by Pessoa et al. (2005) and Pepinelli et al. (2006), respectively. Neither species was allotted toa species group. More recently, Hernández et al. (2006) reviewed the taxonomy of S. petropoliense in whichthe larva and the male were described for the first time, the female and pupa were redescribed, andillustrations of the pupal gill variation and comparisons with other species in the botulibranchium wereprovided.
The most recent classification of Inaequalium was provided in the overview of the Neotropical Simuliidaeby Coscarón & Coscarón-Arias (2007). These authors included 14 species in the subgenus [actually 17 andthree as species inquirendae not here included in the total] of which 14 were allocated to the inaequale andthree to the botulibranchium species group (Table 1). In the same publication the authors added the name, S.hauseri [as I. hauseri] described by Strieder (1998) from Brazil. Their new diagnosis for this subgenus isessentially the same as those provided previously (e.g., Coscarón, 1987, 1991; Table 1) with the omission ofinformation on coloration of antennae, pleura, legs, description of Lutz’s organ, and additional information onthe adult genitalia, pupal cocoon and larva. They separated the two species groups as in previous publicationswith additional information on features of the cibarium and gill (see Table 1).
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 11NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
More recently, Hernández et al. (2007) dealt with new synonymies of Simuliidae in the Neotropical regionand proposed the following synonyms in the subgenus Inaequalium: for S. inaequale (T. manicata, S. clarki,S. pseudoexiguum) and for S. subnigrum (S. subclavibranchium Lutz, S. nahimi, S. nogueirai, S. beaupertuyi,and I. leopoldense). In the same paper, these authors designated a neotype for S. inaequale, a lectotype for S.subclavibranchium, clarified which life stage was designated as the holotype of S. [as Inaequalium]leopoldense, and provided additional information on the preservation and condition of all type species thatthey had examined.
Our conclusionsWe prefer the broader approach of Coscarón (1987), Coscarón & Coscarón-Arias (2007) and Crosskey &
Howard (1997, 2004) in the treatment of Inaequalium as a subgenus, instead of that of Py-Daniel & MoreiraSampaio (1994a, b), Py-Daniel (1995), and Strieder & Py-Daniel (2000, 2002) who consider Inaequalium as agenus. We have maintained species group names of Coscarón (Coscarón, 1987, 1991; Coscarón & Coscarón-Arias, 2007). In our classification of the subgenus Inaequalium not all species clearly fit into groups becauseof the overlap in characters.
We have based our species groups only on characters that we consider to generally show relativelyminimal intraspecific variation in the Neotropical fauna. These are: scutal pattern, cibarium, adult genitalia,and cocoon and gill morphology of pupae. We have used a suite of these characters and species have beenplaced in a species group when most of the characters in that species coincide with those of the species group.We maintain the division of the subgenus Inaequalium into the botulibranchium and inaequale speciesgroups. The most obvious characters separating the two groups are the form of the female cercus andparaproct, the male gonostyle and ventral plate and the pupal gill. The use of these characters in determiningphylogenetic relationships may only be adequately resolved when considered in combination with cytologicaland molecular characters. It has been suggested by several authorities, e.g. Crosskey (1988), Shelley (1988),Crosskey & Howard (1997), that Inaequalium might be better subsumed within Psaroniocompsa due to thetheir morphological similarity. We support this suggestion here. However, this still needs further investigationin relation to species of other subgenera in the Neotropical Region and a final decision will be made followingthis analysis in the forthcoming revision of “The Simuliidae of Brazil by A.J. Shelley, L.M. Hernández, M.Maia-Herzog, A.P.A. Luna Dias & P.R. Garitano’.
Classification Our classification of Inaequalium has been derived from that proposed by Crosskey & Howard (2004)with two subsequent species descriptions by Pessoa et al. (2005) and Pepinelli et al. (2006). The 20 speciesunder Inaequalium have now been reduced to 14 valid species (including two species inquirendae) throughthe recent synonyms proposed by Hernández et al. (2007) and in this paper. All species are listed in alphabet-ical order within each species group. Valid species are given in bold and italics, with synonyms in italics.
BOTULIBRANCHIUM species group
botulibranchium Lutz, 1910petropoliense Coscarón, 1980 souzalopesi Coscarón, 1980
INAEQUALE species group
clavibranchium Lutz, 1910 diversibranchium Lutz, 1910 missionum Coscarón, 1976
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.12 · Zootaxa 1649 © 2007 Magnolia Press
inaequale Paterson & Shannon, 1927 baiense Pinto, 1932
manicatum (Enderlein, 1934)clarki Fairchild, 1940 argentatum (Enderlein, 1936)
jundiaiense D’Andretta & González B., 1964 pseudoexiguum Nunes de Mello & Barbosa de Almeida, 1974
maranguapense (Pessoa, Ríos-Velásquez & Py-Daniel, 2005)margaritatum Pepinelli , Hamada & Luz, 2006mariavulcanoae Coscarón & Wygodzinsky, 1984 rappae Py-Daniel& Coscarón, 1982 subnigrum Lutz, 1910
subclavibranchium Lutz, 1910 diversifurcatum Lutz, 1910 nogueirai D’Andretta & González B., 1964 beaupertuyi Ramírez-Pérez, Rassi & Ramírez, 1977 mbarigui Coscarón & Wygodzinsky, 1973 nahimi Py-Daniel, 1984 leopoldense (Strieder & Py-Daniel, 2000)
travassosi D’Andretta & D’Andretta, 1947
Species inquirendae within the inaequale species group
lurybayae Smart, 1944 parimaense Ramírez-Pérez, Yarzábal, Takaoka, Tada & Ramírez , 1986
Unavailable name within the inaequale species group
hauseri Coscarón & Coscarón-Arias, 2007
Diagnosis of the subgenus Inaequalium
This subgenus now includes 14 valid species of which 12 occur in Brazil. Female. Nudiocular area developedto varying degrees (Figs. 1–14). Cibarium with developed, sclerotized cornuae and well-sclerotized marginarmed with several rows of sharp teeth; median part protruded, except in S. souzalopesi [a similar conditionwas found in one specimen of S. margaritatum by Pepinelli et al. (2006)] (Figs. 15–30). Scutum black or greypruinose, although sometimes brown, with numerous, recumbent, golden or golden greenish hairs interspersedwith semi-erect black hairs; pattern weak when present. Thoracic pattern varies slightly with light direction.With light source anterior to specimen, 1+1 median and 1+1 submedian, grey, pruinose vittae divergingposteriorly and extending from anterior third of thorax to near posterior margin. With light posterior, tospecimen, pattern reverses with silver pruinose areas becoming black in species with a pattern; in species withno pattern only indistinct, silver pruinose areas are seen on anterior margin of thorax (Figs. 31–66). Eighthsternite with sclerotized posterior margins with groups of setae; gonapophyses subtriangular and protrudingposteriorly at inner margins, membranous except internal margins sclerotized (Figs. 107–119). Cercisubhemispherical, paraprocts poorly developed and almost same length as cercus (Figs. 123–133) ordeveloped and extending well beyond junction with cercus (Figs. 120–122). Genital fork (Figs. 134–147)
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 13NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
sclerotized, with developed lateral arms and anterior processes. Male. Scutum velvet black. With lightanterior to specimen, scutum either without pattern or with 1+1 submedian, silver, pruinose areas; thoraxcovered with golden, recumbent hairs irregularly distributed. With light posterior, to specimen thorax black(Figs. 148–167). Gonocoxite subrectangular; gonostyle subcylindrical, subtriangular or subtrapezoidal,usually with one apical spine, but 2–10 subapical spines present in some species (Figs. 168–182). Ventralplate lightly sclerotized, crescent-shaped, with many fine setae and median part of anterior margin bluntlypointed, rounded or developed anteriorly, and basal arms variously developed (Figs. 183–193). Parameresclerotized, usually with well-developed spines (Figs. 194–204). Median sclerite pear-shaped, with apicalincision. Pupa. Cocoon generally slipper shaped. Gill white to light brown consisting of 4–6 forwardlydirected filaments branching at various heights in basal half or bulbous, irregular primary filaments usuallycovered by hairs or spinules (Figs. 205–239). Larva. We have not been able to examine sufficiently largenumbers of specimens of different species to be able to provide a comprehensive diagnosis at the subgenericor species group level. The reader is referred to Table 1 for diagnoses of previous authors. Our diagnosticcharacters of the larva stem from that of Coscarón & Coscarón-Arias (2007).
The subgenus Inaequalium is divided into two species groups: the botulibranchium species group withthree species, and the inaequale species group with 11 species.
BOTULIBRANCHIUM species group
Diagnostic features of this species group are as follows. Female. Paraprocts well developed, subtriangular,extending well beyond junction with cercus by up to twice length of cercus (Figs. 120–121). Male.Gonocoxite subrectangular; gonostyle subcylindrical narrowing apically, same length as gonocoxite, andterminating in one apical spine (Figs. 168–171). Ventral plate with distinct shoulders; main body of ventralplate with median region distinctly developed and rounded apically. Pupa. Gill filaments distinctly swollencovered by spicules or hairs, or bare and arranged irregularly (Figs. 205–211). Larva. Postgenal cleft small,nearly extending to one third of length of head capsule; hypostomium nearly same length as postgenal bridge.
INAEQUALE species group
Diagnostic features of this species group are as follows. Female. Paraprocts slightly developed,subrectangular, only extending beyond the junction with cercus by one-half length of cercus (Figs. 122–132).Male. Gonocoxite subrectangular; gonostyle subtriangular or subtrapezoidal, nearly same length asgonocoxite, and terminating in one apical spine [some species with 2–10 subapical spines] (Figs. 171–180).Ventral plate without distinct shoulders; main body of ventral plate crescent-shaped, often pointed centrally.Pupa. Gill filaments fine and rarely swollen, either bare or covered by small spinules and arranged regularly(Figs. 212–239). Larva. Postgenal cleft long, nearly reaching middle of head capsule; hypostomium 1.5 timeslonger than postgenal bridge.
Key to species of Inaequalium
Several keys exist that purport to separate the females and males of all species of the Inaequalium speciesgroup. However, we were unable to reliably separate the adult life stage of this homogenous group, except tospecies group using the structure of the paraproct (Figs. 120–133) and the gonostyle (Figs. 168–182). Fromthere on species separation is not possible because of their morphological homogeneity and so recourse has tobe made to reared specimens with attention given to pupal morphology. The reader should consult themorphological descriptions and digital images of adults as an aid to species identification. The number and
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.14 · Zootaxa 1649 © 2007 Magnolia Press
configuration of the pupal gill filaments is usually the most reliable taxonomic character to identify species inthis subgenus. However, variation in this character has been found in several taxa making their identificationdifficult for non-taxonomists. Simulium lurybayae and S. parimaense are not included in the key to speciesbecause the pupa is unknown in the former species, and because of the confusion on the status of the femaleand pupa of the latter (see also Taxonomic discussion under each taxon).
PUPAE
1. Gill bifid or trifid, swollen, with finger-like or fine secondary filaments and usually with bulbous protru-sions .............................................................................................................................................................2
– Gills with 3–6, fine or swollen filaments, without finger-like secondary filaments and no bulbous protru-sions ............................................................................................................................................................5
2. Gill trifid, forwardly directed from base, regular in shape with dorsal primary branch narrowing apicallyand sometimes with two finger-like projections. Gill covered with fine spicules (Figs. 236–239) ................................................................................................................................................................. S. travassosi
– Gill bifid, primary branches initially orientated in dorsal and ventral planes and then forwardly directed,irregular in shape with dorsal primary branch narrowing apically with or without finger-like projections.Gill glabrous or covered with spicules and/or hairs (Figs. 205–211) ...........................................................3
3. Gill surface glabrous. Dorsal primary branch bifurcating basally forming tapering secondary branches;upper branch bifurcating distally and lower branch bifurcating basally. Ventral primary branch short andbulbous with two short, pointed filaments apically (Fig. 211) ................................................S. souzalopesi
– Gill surface with spicules and/or hair. Dorsal and ventral primary branches short and swollen, either taper-ing apically or not, with pointed ends (Figs. 205–210) ................................................................................4
4. Dorsal primary branch with small bulbous protrusion basally, one or two ventrally directed short, stout fil-aments beyond mid point on ventral surface and 1–4 short, stout terminal filaments. Ventral primarybranch shorter with bulbous protrusion near base. Filaments covered in spicules. (Figs. 205, 206) ....................................................................................................................................................S. botulibranchium
– Dorsal primary branch with small bulbous protrusion at right angle from where it points forward from itsbasal vertical orientation; apex of primary branch with 3–4 finger-like projections. Ventral primary branchof similar form except tapering apically with no division. Filaments covered in hairs and spicules. (Figs.207–210) ............................................................................................................................... S. petropoliense
5. Gill with four filaments ................................................................................................................................6– Gill with six filaments .................................................................................................................................86. Gill filaments fine (Fig. 217) ............................................................................................... S. margaritatum– Gill filaments stout (Figs. 216, 229–234) .....................................................................................................77. Gill filaments all arising from base and tapering distally (Fig. 216) ............................... S. maranguapense– Gill bifurcating basally with a further bifurcation in each of the dorsal and ventral primary branches.
Bifurcation of dorsal primary branch varying in position from basal to apical; bifurcation of ventral pri-mary branch near mid-point of gill (Figs. 229–234) ..................................................................... S. rappae
8. Gill filaments lightly to strongly swollen with basal division into three primary branches, each of whichbifurcates at varying distances from base (Figs. 219–228) ............................................................ S. rappae
– Gill filaments narrow and dividing into primary branches basally (Figs. 212–215, 218, 235)................... 99. Apices of filaments club-shaped and pointed (Figs. 212, 213) ....................................... S. clavibranchium – Apices of filaments narrow and pointed (Figs. 214, 215, 218, 235) ......................................................... 1010. Gill divided basally into three primary branches (Fig. 215) .....................................................S. inaequale – Gill divided basally into dorsal and ventral primary branches (Figs. 214, 218, 235)................................ 1111. Dorsal primary branch thicker than ventral primary branch. Dorsal primary branch bifurcates basally into
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 15NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
two thick secondary branches, each of which bifurcates into finer filaments at about same level in basalhalf of gill (Figs. 214, 218) ........................................................... S. diversibranchium, S. mariavulcanoae
- Dorsal primary branch same girth as ventral primary branch. Dorsal primary branch bifurcates basallyinto two fine secondary branches, each of which bifurcates into finer filaments at different levels in basalhalf of gill (Fig. 235) ................................................................................................................ S. subnigrum
Species descriptions, distribution and biology
BOTULIBRANCHIUM species group
This is a small species group with only three species, all of which occur in Brazil.
Simulium (Inaequalium) botulibranchium Lutz(Figs. 1, 15, 31, 32, 67–69, 107, 120, 134, 148, 149, 168, 169, 183, 194, 205, 206)
A common species in southern Brazil closely related to S. petropoliense.
Simulium botulibranchium Lutz, 1910: 256. LECTOTYPE pupal exuviae, BRAZIL: Rio de Janeiro State, Morro doObservatório; [Without date.], (A. Lutz) (IOC, no. 12.038, Bd 103) [Examined.] [Lectotype designation byD’Andretta & D’Andretta, 1947: 161.]
Female. General body color black. Body length (specimens pinned) 2.0–2.2 mm (mean=2.0 mm, s.d.=0.09,n=4); wing length 1.9–2.2 mm (n=3); wing width 1.0–1.1 mm (mean=1.1 mm, s.d.=0.07, n=4).
Head—dichoptic, with dark red eyes and nudiocular area well developed (Fig. 1). Frons, clypeus andocciput black, with silvery grey pruinosity; frons, clypeus and occiput with semi-recumbent, black hairs.Mouthparts black. Antennae with scape and pedicel yellowish brown, remainder of flagellar segments black.Cibarium with well-developed, sclerotized cornuae and irregular rows of sharp teeth of varying size extendingfrom base of cornuae to central area of cibarium, which is protuberant (Fig. 15).
Thorax—scutum black covered with recumbent, golden hairs interspersed with black hairs. Scutal patternweak and varying depending on light incidence. With anterior illumination scutum black; humeri and lateralmargins weakly pruinose; posterior margin black, with faint pruinosity (Fig. 31). With posterior illumination,thorax black, with 1+1 indistinct median grey pruinose vittae on anterior region of scutum; humeri greypruinose; lateral and posterior margins black (Fig. 32). Scutellum dark brown to black, with recumbent,golden hairs interspersed with long, black bristles. Postnotum black, with grey pruinosity. Pleura black, withsilver pruinosity. Costa of wing with sparse distribution of spines and setae. Subcosta with line of setae exceptapical third bare. Radius with row of setae intermixed with distinct spines, basal section of radius with line ofsetae. Basal tuft of sparse, light brown setae. Leg coloration and proportions as in Figs. 67–69. Foreleg withcoxa, trochanter, femur, apical third and internal surface of tibia, and tarsal segments dark brown; externalsurface of tibia whitish. Mid and hind legs with coxa, trochanter, distal fourth of femur, distal two thirds oftibia, apical fourth of basitarsus and tarsal segments II–IV dark brown; basal third of tibia and three quartersof basitarsus whitish. Claws weakly curved with small basal tooth. Halteres lemon yellow, with dark brownbase.
Abdomen—tergite I black; tergite II black, brownish laterally, with distinct silver pruinosity onanterolateral margin [best seen when specimens viewed laterally]; tergites III–IX velvet black. Tergal platesdeveloped. Sternites greyish; genitalia black. Eighth sternite weakly sclerotized with long, irregularlydistributed setae on posterior margin; gonapophyses subtriangular, half length of eighth sternite at midpoint,membranous, except sclerotized internal margins (Fig. 107). Cerci suboval, covered with distinct, long brown
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.16 · Zootaxa 1649 © 2007 Magnolia Press
setae; paraproct subtriangular twice as long as cercus, highly sclerotized, covered with long setae basally andmacrotrichiae on anterior margin of posterior prominence (Fig. 120). Genital fork weakly sclerotized, withstem slightly expanded apically; lateral arms with well-developed, subtriangular anterior processes andreduced posterior processes, area between lateral arms subquadrate (Fig. 134). Spermatheca suboval, withoutexternal sculpturing and irregularly distributed spicules on internal surface; area of insertion of spermathecalduct membranous.
Male. General body color black. Body length (specimens pinned) 2.1–2.6 mm (mean=2.3 mm, s.d.=0.19,n=5); wing length 1.4–2.1 mm (mean=1.8 mm, s.d.=0.33, n=4), wing width 1.0–1.1 mm (mean= 1.0 mm,s.d.=0.07, n=5).
Head — holoptic, with dark red eyes. Remainder of head coloration as in female.Thorax—scutum black, covered with evenly distributed golden, recumbent hairs. Scutal pattern varies
slightly with light incidence. With light source anterior to specimen scutum black, with 1+1 silver pruinose,submedian cunae on anterior margin; humeri, lateral and posterior margins black (Fig. 148) [In somespecimens the silver pruinose cunae cannot be seen.] With light posterior, thorax black; humeri, lateral andposterior margins of scutum black (Fig. 149). Scutellum dark brown, covered with recumbent, golden hairsinterspersed with long, erect, dark brown setae. Postnotum black, with silvery grey pruinosity. Pleura black,with grey pruinosity. Halteres whitish yellow, with light brown base. Wing setation as in female, except Scbare. Leg coloration as in female, except mid and hind femora and tibiae covered with small, slightly spatulatesetae.
Abdomen—tergites black, basal fringe of long, brown hairs. Silver pruinose ornamentation on anterolateralmargin of tergite II and ventrolateral margins of tergites V–VII. Genitalia black; sternites whitish pruinose;tergal plates developed. Gonocoxite subrectangular; gonostyle subcylindrical, pointed apically, as long asgonocoxite at mid point, with weakly sinuous margins and terminating in single, stout spine; gonocoxite andgonostyle covered with long setae (Figs. 168, 169). Ventral plate crescent-shaped, weakly sclerotized, withmain body well developed centrally and median concavity on anterior margin; basal arms well developed,sclerotized and rounded; main body of ventral plate covered with long hairs (Fig. 183). Median scleritepyriform, as long as wide at mid point, with incision on apical third (Fig. 194). Paramere with developed andsclerotized basal process, and few short spines mesally; membrane between basal arms of paramere with finespicules (Fig. 194).
Pupa. Cocoon length dorsally 2.7–3.4 mm (mean=3.0 mm, s.d.=0.2, n=13); ventrally 3.2–3.8 mm (mean=3.4mm, s.d.=0.19, n=13); pupa length 2.6–3.8 mm (mean=3.2 mm, s.d.=0.41, n=13); gill length 1.8–2.7 mm(mean=2.2 mm, s.d.=0.29, n=13).
Cocoon — slipper shaped, light to dark brown composed of a fine network of coalescent fibers andreinforced rim to anterior aperture.
Gill — light brown, with two, irregular, stout filaments arranged in vertical plane. Gill configuration withmain trunk short, giving rise to two primary branches, one dorsal and one ventral. Dorsal primary branchforwardly directed with small bulbous protrusion basally, one or two ventrally directed short, stout filamentsbeyond mid point on ventral surface and 1–4 terminal short stout filaments apically. Ventral primary branchdirected ventrally with bulbous protrusion near base and apically curved forwards. All filaments swollen,often rounded distally, covered with small spicules on surface, edges smooth to wavy (Figs. 205, 206).
Head — frontoclypeus with 2+2 simple, frontal and 1+1 bifid, facial trichomes; frontoclypeus with groupof platelets mesally, 1+1 groups of approximately eight platelets dorsolaterally and 2–3 platelets in groups oftwo laterally in frontal region; tubercles rounded, sparsely distributed over entire surface.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 17NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
Thorax — 5+5 large, quadrifid trichomes near margin of dorsal cleft, one long, simple trichome on centralregion and two long, simple trichomes near base of gill ventrally; tubercles rounded, densely distributed oncentral and dorsal region of thorax, but absent near base of gill.
Abdomen — tergite I with 2+2 long, sublateral, simple trichomes; tergite II with 4+4 submedian spiniformsetae in row, the outer, fourth seta more spaced than three inner setae, 1+1 spiniform setae anterior to outersetae, and 3+3 small, single setae on lateral margin; tergites III and IV with 4+4 submedian, simple hooks inrow along posterior margin, 1+1 spiniform setae anterior to outer hooks, and 2+2 small, spiniform setae onlateral margin; tergite V with 1+1 submedian, small, simple trichomes and 3+3 sublateral, simple trichomes inrow along posterior margin; tergite VI with 1+1 submedian, simple and 1+1 sublateral, simple, smalltrichomes and spine combs on anterior margin; tergites VII and VIII with 2+2 small, submedian, simpletrichomes on posterior margin and well-developed spine combs resembling teeth on anterior margin; tergiteIX weakly sclerotized, with well-developed spine combs resembling teeth on anterior margin and 1+1 small,apical spines. Abdominal sternite III with 3+3 sublateral, simple trichomes; sternite IV with 2+2 median and2+2 sublateral, simple, spiniform setae on posterior margin; sternite V with 2+2 submedian, adjacent, bifidhooks on posterior margin, the outermost on the right side trifid, remainder bifid, and 2+2 small, simpletrichomes anterior to outer hooks; sternites VI–VII with 2+2 well separated hooks on posterior margin, theinner being bifid and the outer simple, 1+1 or 2+2 simple, small trichomes anterior to outer hooks, 1+1 small,simple trichomes between outer hooks; sternite VIII without visible setae or trichomes; sternite IX weaklysclerotized. Spine combs on anterior margin of sternites III–IX.
Taxonomic discussionLutz (1910) first proposed the name S. botulibranchium for several larvae and pupae that he had collected
at Morro do Observatório, Petrópolis, near Rio de Janeiro, Brazil. However, he only provided themorphological characters to distinguish the pupa of this species. In the same paper, Lutz figured the pupa ofactual S. botulibranchium, but referred to this species in the figure’s legend as S. distinctum (see Lutz, 1910:261, Fig. 1). A typographical error was also made by Lutz (1922), in which the species name was given as S.tulibranchium. D’Andretta & D’Andretta (1947) redescribed the pupa of S. botulibranchium, described thefemale and male for the first time, and designated as lectotype the pupal exuviae “Lámina [=slide] no. 312”used by Lutz (1910) to illustrate this species. We have examined the lectotype of S. botulibranchium, which isdeposited in the IOC. The slide does not bear a label with locality information, date or collector’s name, butD’Andretta & D’Andretta (1947) stated that the pupa was collected in “Morro do Observatório, Petrópolis,Rio de Janeiro by A. Lutz in 1910”. Their statement was probably based on the IOC catalogue of Lutz’Simuliidae collection. The pupal exuviae is in relatively good condition, although the dorsal branch of theright gill is broken apically. We also examined one slide with only male genitalia (Fig. 168) and three pinnedspecimens (two females mounted on the same pin with one male) bearing a pink label as “TIPO” [TYPE].These are not types because Lutz in his original description only described pupae. We are unable to makefurther comments on the identity of this material, as they are not link-reared. Therefore, they have beenidentified with doubt as S. botulibranchium and labelled accordingly (see Material Examined).
Simulium botulibranchium belongs to the botulibranchium species group based on the combination ofcharacters given in Coscarón (1980, 1987) and Coscarón & Coscarón-Arias (2007). The adults of S.botulibranchium are externally similar to other species of the botulibranchium species group from which thisspecies can only be separated by the configuration of the pupal gill filaments. Within this species group, S.botulibranchium is most similar to S. petropoliense in having the primary branches distinctly stout andbulbous with a prominence nearly at the base of the dorsal primary branch (Figs. 205–210). However, S.botulibranchium can be recognized by the dorsal primary branch having shorter and stouter secondarybranches and all gill filaments being covered with small spicules (Figs. 205–206). In S. petropoliense, the
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.18 · Zootaxa 1649 © 2007 Magnolia Press
dorsal primary branch often divides into four, apical, finger-like projections covered by distinct hairs (Figs.207–210).
Descriptions of adults and immature stages of S. botulibranchium are found in Coscarón (1991), Coscarón& Coscarón-Arias (2007), D’Andretta & D’Andretta (1947) and Strieder & Py-Daniel (1999; 2000).
DistributionSimulium botulibranchium is a common species in southern Brazil where it has been recorded in the states
of Rio de Janeiro and São Paulo. Elsewhere, it occurs in Missiones Province, Argentina (Crosskey & Howard,1997; 2004; Coscarón & Coscarón-Arias, 2007; Material Examined).
Biology and medical importanceIn Brazil, the immature stages of S. botulibranchium have been found attached to vegetation in small (20
cm to 1 m wide, 2–3 cm deep), fast flowing streams with a variety of riverbeds (rock and sand) sympatricallywith other simuliid species, e.g. S. petropoliense, S. clavibranchium, S. subclavibranchium [now a synonymof S. subnigrum, see Hernández et al., 2007] and S. diversibranchium (A.J. Shelley, A.P.A. Luna Dias & P.R.Garritano, M.Maia-Herzog, L.M.Hernández, unpublished data). Nothing is known about the feeding habits offemales of S. botulibranchium. They have never been collected biting humans in the Neotropical Region.
Simulium (Inaequalium) petropoliense Coscarón(Figs. 2, 16, 33, 34, 70–72, 108, 121, 135, 150, 151, 170, 195, 207–210)
An uncommon species in Brazil, closely related to S. botulibranchium. Its taxonomy has been recentlyreviewed by Hernández et al. (2006).
Simulium petropoliense Coscarón, 1980: 298. HOLOTYPE & (reared), BRAZIL: Rio de Janeiro State, Petrópolis;12.v.1979 (Coscarón) (MLP). [Examined.]
Female. Coscarón (1980) stated in the original description of S. petropoliense that the females are externallysimilar to females of S. botulibranchium. We redescribe the female of S. petropoliense based on two pharatefemales and an examination of a slide containing one hind leg, one wing, and the genitalia of the holotype.General body color black. Body length (specimens in ethanol) 2.8–3.5 mm (n=2).
Head—dichoptic, with dark red eyes and nudiocular area developed (Fig. 2). Frons, clypeus and occiputblack, with silvery grey pruinosity, covered with semirecumbent, black hairs interspersed with white erecthairs. Mouthparts black. Antennae with scape and pedicel yellowish brown; flagellomeres dark brown.Cibarium with well-developed, sclerotized cornuae and irregular rows of teeth of varying size extending frombase of cornuae to central area of cibarium, which is weakly protuberant (Fig. 16).
Thorax—scutum black, covered with recumbent, pale golden hairs [specimen photographed in ethanol].Scutal pattern variable, depending on light incidence. With anterior illumination scutum black, humeriyellowish, lateral and posterior margins black (Fig. 33). With posterior illumination, thorax black, with faint1+1 median grey pruinose vittae on anterior region of scutum; humeri yellowish; lateral and posterior marginsweakly pruinose (Fig. 34). Scutellum pale brown, devoid of hairs in the single specimen examined. Postnotumdark brown, with grey pruinosity. Pleura black, with silver pruinosity. Costa of wing with sparse distributionof spines and setae. Subcosta with line of setae, except apical third bare. Radius with row of setae intermixedwith distinct spines, basal section of radius with line of setae. Basal tuft of sparse, light brown setae. Legcoloration as in Figs. 70–72 [based on a teneral specimen]. Foreleg with coxa pale brown, trochanter, femur,
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 19NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
tibia apically and basitarsal segments I–IV dark brown; basal two thirds of tibia whitish. Mid leg with coxa,trochanter, apex of femur, apex of tibia, and apex of basitarsus and tarsal segments II–IV dark brown; basaltwo thirds of femur pale brown; basal two thirds of tibia and basitarsus, and tarsal segment II whitish. Hindleg with coxa, trochanter, apical third of femur and tibia, and apex of tarsal segments I–II dark brown; basaltwo thirds of femur pale brown; basal two thirds of tibia and tarsal segments I and II basally whitish. Clawsweakly curved, with small basal tooth. Halteres lemon yellow, with dark brown base.
Abdomen—tergite I pale brown; tergite II dark brown, with silver pruinosity on anterior margins; tergitesII–IX black. Tergal plates weakly developed. Sternites greyish; genitalia black. Eighth sternite weaklysclerotized, with long, irregularly distributed setae on posterior margin; gonapophyses subtriangular, nearlysame length as eighth sternite at midpoint, membranous, except sclerotized internal margins (Fig. 108).Cercus suboval, sclerotized, covered with distinct long brown setae; paraproct subtriangular, sclerotized,longer than cercus, with distinct prominence on posterior margin near junction with cercus; paraproct coveredwith long setae on posterior half and macrotrichia apically (Fig. 121). Genital fork sclerotized, with stemslightly expanded apically; termination of lateral arms with anterior processes well developed, subtriangular(Fig. 135). Spermatheca suboval, without external sculpturing and irregularly distributed spicules on internalsurface; area of insertion of spermathecal duct membranous.
Male. General body color black. Body length (specimens in ethanol) 2.6–3.2 mm (n=2). Wing length 1.98mm (n=1); wing width 0.8 mm (n=1).
Head—holoptic, with dark red eyes. Frons and clypeus with silvery grey pruinosity and covered by dark hairs. Mouthparts black. Antennae with scape and pedicel yellowish brown and flagellomeres black.
Thorax—scutum dark black covered with evenly distributed golden, recumbent hairs [specimenphotographed in ethanol]. Thorax, irrespective of light incidence, black (Fig. 150), but with faint 1+1submedian silver cunae if specimen tilted dorsolaterally (Fig. 151); humeri pale yellow; lateral and posteriormargins black. Scutellum pale brown, covered with recumbent golden hairs interspersed with long, erect, darkbrown setae. Postnotum dark brown, with silvery grey pruinosity. Pleura black, with grey pruinosity. Haltereswhitish yellow, with light brown base. Wing setation as in female, except Sc bare. Leg coloration as in female.Claw without tooth, but with thumb-like basal protuberance. Femur and tibia of hind leg with lanceolate setae. Abdomen—tergites dark brown to black, basal fringe of long, brown hairs. Faint silver pruinoseornamentation present on ventrolateral margins of tergites II–III. Genitalia black; sternites greyish, with faintsilver pruinosity [specimens in ethanol]; tergal plates undeveloped. Gonocoxite subquadrangular; gonostylesubcylindrical and pointed distally, nearly same length as gonocoxite, terminating in single spine; gonocoxiteand gonostyle covered with long setae (Fig. 170). Ventral plate sclerotized, with main body well developed,prominently produced on anterior margin, and wide concavity on central region of posterior margin; basalarms weakly sclerotized and curved inwards; main body of ventral plate covered by long hairs (Fig. 184).Median sclerite pyriform, as long as wide at mid point, with incision on apical third (Fig. 195). Paramere withdeveloped and sclerotized basal process and numerous long and short spines along distal half; membranebetween basal arms of paramere with fine spicules (Fig. 195).
Pupa. Cocoon length dorsally 2.3–2.8 mm (mean=2.5 mm, s.d.= 0.14, n=9); ventrally 2.6–3.2 mm (mean=3.2mm, s.d=0.21, n=7); pupa length 2.0–2.6 mm (mean=2.3 mm, s.d.= 0.20, n=7); gill length 1.1–1.9 mm(mean=1.4 mm, s.d.=0.2, n= 9).
Cocoon—slipper shaped, light to dark brown composed of a fine network of coalescent fibers andreinforced rim to anterior aperture.
Gills—light to dark brown, with filaments arranged in vertical plane. Gill bifurcates basally into swollendorsal and ventral orientated primary filaments that then each form a right angle and point forwards. Dorsalprimary branch terminating in 3–4 finer filaments at variable distance from base. Ventral branch sometimes
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.20 · Zootaxa 1649 © 2007 Magnolia Press
with bulbous protrusion near base, tapering distally and usually unbranched, but occasionally bifid distally(Figs. 207–210).
Head—frontoclypeus with 2+2 long, single, bifid or trifid frontal and 1+1 long, bifid or quadrifid, facialtrichomes (Coscarón, 1980: 300); frontoclypeus with group of platelets mesally, 1+1 groups of approximately13 platelets dorsolaterally and two groups of platelets in groups of 2–3 laterally in frontal region; tuberclesrounded and densely distributed over entire surface.
Thorax—approximately 4+4 long, bifid to quadrifid trichomes near margin of dorsal cleft, one long, bifidtrichome on posterior region mesally, 1+1 long and 1+1 short, simple trichomes on alar region, and 3+3 long,simple trichomes on ventral margin of alar region; tubercles rounded (some pointed near base of gill) anddensely distributed over entire surface of thorax.
Abdomen—tergite I with 1+1 long, sublateral simple trichomes; tergite II with 3+3 submedian, spiniformsetae in row, 3+3 small, simple trichomes in vertical line to spiniform outer setae, and 1+1 spiniform setae onlateral margin; tergites III–IV with 4+4 submedian, simple hooks in row along posterior margin, 1+1 simpletrichomes anterior to outer trichomes, and 1+1 spiniform setae on lateral margin; tergites V–VI withouttrichomes or setae; tergites VII and VIII without visible trichomes, but well-developed spine combsresembling teeth on anterior margin; tergite IX weakly sclerotized, with well-developed spine combsresembling teeth on anterior margin and terminating in 1+1 small, apical spines. Abdominal sternite III with3+3 submedian and 2+2 lateral spiniform setae; sternite IV with one submedian and 2+2 lateral spiniformsetae; sternite V with 2+2 separated bifid or trifid hooks on posterior margin, and 2+2 sublateral and twolateral spiniform setae; sternite VI with 2+2 separated, bifid hooks on posterior margin, 1+1 spiniform setaeanterior to outermost hooks, 1+1 long trichomes between outermost hooks; sternite VII with 2+2 wellseparated bifid or simple hooks on posterior margin, 1+1 long, simple trichomes between and anterior tooutermost hooks; sternite VIII without setae; sternite IX sclerotized.
Taxonomic discussion Simulium petropoliense was described by Coscarón (1980) from a reared female holotype and pupal
paratype collected in Petrópolis, Rio de Janeiro State on 12.v.1979 by this author. The taxonomy of thisspecies has recently been reviewed by Hernández et al. (2006) who also described the male and the larva forthe first time, provided information on the condition of the holotype and its confusion with a paratype of S.souzalopesi in the MLP Simuliidae collection, and compared it with related species in the botulibranchiumspecies group (sensu Crosskey & Howard, 2004).
The most reliable character to recognize S. petropoliense is the configuration of the pupal gill filaments.Simulium petropoliense and S. botulibranchium differ from S. souzalopesi in that in the latter species thepupal gill filaments are bare and there are only six filaments (Coscarón, 1980). In S. petropoliense and S.botulibranchium the gills are stout and bulbous for the main part and consist of a dorsal and a ventral primarybranch that divide from the main stem basally in the vertical plane, each of them forming a right angle ornearly a right angle causing the remainder of the gill to be directed anteriorly. A prominence is present oneach primary branch at or near the right angle bend (Figs. 205–210; Coscarón, 1980). The dorsal primarybranch in the two species then divides into four (S. petropoliense) or up to five (S. botulibranchium)secondary branches of variable length. In S. petropoliense they are finer and longer (Figs. 207–210), but muchstouter and shorter in S. botulibranchium (Fig. 205–206; Coscarón, 1980). The ventral primary branchremains bulbous and simple in S. botulibranchium (Figs. 205–206; Coscarón, 1980), but it can be single oralso be divided into two filamentous secondary branches in S. petropoliense (Figs. 207–210). Anotherdifference between these two species is the morphology of the gill surface. In S. botulibranchium the gill iscovered with minute spicules, whereas in S. petropoliense it is covered with hairs.
Variation in the pupal gill filaments, frontoclypeal trichomes, and setation of the abdominal sternites V–VIII was seen in specimens of S. petropoliense examined by Hernández et al. (2006) from Brazil. Simulium
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 21NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
petropoliense is placed in the botulibranchium species group following the combination of characters given inCoscarón (1980, 1987, 1991) and Coscarón & Coscarón-Arias (2007), especially the females with asubtriangular paraproct with a basal protuberance (Fig. 121), male with an elongate, subcylindrical gonostyle,the same length or longer than the gonocoxite (Fig. 170), and asymmetric pupal gill filaments (Figs. 207–210). We agree with the placement of S. petropoliense in this species group.
Distribution Simulium petropoliense has only been collected in Brazil in the municipality of Petrópolis, Rio de Janeiro
State (Coscarón 1980, 1987, 1991; Coscarón & Coscarón-Arias, 2007; Crosskey & Howard 2004; Strieder &Py-Daniel2000; Material Examined).
Biology and medical importanceCoscarón (1980, 1991) and Strieder & Py-Daniel (1999) stated that the immature stages of Simulium
petropoliense can be collected in open spaces in small trickles with clear water, commonly found attached tovegetation and rocks, sympatric with S. botulibranchium and S. souzalopesi. We have collected larvae andseveral pupae of S. petropoliense attached to rocks in trickles of clear water in a waterfall. It was alsocollected on leaves where the water was running faster. The immature stages were collected in the open partof the waterfall receiving direct sunlight. The feeding habits of S. petropoliense are not known, but femaleshave never been found biting humans in Brazil.
Simulium (Inaequalium) souzalopesi Coscarón(Figs. 3, 17, 35, 36, 73–75, 109, 122, 136, 152, 153, 171, 185, 196, 211)
This is an uncommon species from southern Brazil.
Simulium souzalopesi Coscarón, 1980: 296. HOLOTYPE & (reared), BRAZIL: Rio de Janeiro State; 12.v.1979 (Cos-carón) (MLP) [Examined.]
Female. General body color black. Body length (specimens pinned) 2.3–2.4 mm (n=2); wing length 1.7–1.9mm (n=2); wing width 0.8–1.1 mm (n=2).
Head—dichoptic, with dark red eyes and nudiocular area developed (Fig. 3). Frons, clypeus and occiputblack, with silvery grey pruinosity; frons, clypeus and occiput with semi-recumbent, black hairs. Mouthpartsblack. Antennae with scape and pedicel yellowish brown, remainder of flagellar segments black. Cibariumwith well-developed, sclerotized cornuae and irregular rows of sharp teeth of varying size between cornuaecentrally, anterior medium margin of cibarium with a slight depression (Fig. 17).
Thorax—scutum black, covered with recumbent, golden hairs. Scutal pattern variable depending on lightincidence. With anterior illumination, scutum black, with faint pruinosity on median region of thorax; humeriweakly pruinose; lateral and posterior margins black (Fig. 35). With posterior illumination, thorax black, with1+1 indistinct, median grey pruinose vittae on anterior region of scutum; humeri and lateral margins greypruinose; posterior margin black (Fig. 36). Scutellum dark brown, with recumbent, golden hairs interspersedwith long, black bristles. Postnotum black, with grey pruinosity. Pleura black, with silver pruinosity. Costa ofwing with sparse distribution of spines and setae. Subcosta with line of seven setae on basal third. Radius withrow of setae intermixed with distinct spines, basal section of radius with line of setae. Basal tuft of sparse,light brown setae. Leg coloration and proportions as in Fig. 73–75. Foreleg with coxa, trochanter, base andapex of femur and tibia, and basitarsal segments dark brown; remaining parts of femur and tibia yellowish.Mid leg with coxa, trochanter, base and apex of femur, apex of hind tibia, and apex of tarsal segments I–IV
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.22 · Zootaxa 1649 © 2007 Magnolia Press
dark brown; remaining parts of legs whitish yellow. Hind leg as mid leg except basal part of femur whitishyellow. Hind and mid femora covered with spatulate setae. Claws weakly curved with small basal tooth.Halteres lemon yellow, with dark brown base.
Abdomen—tergite I black, with silver pruinosity on lateral margin; tergites II–IX black. Tergal platesdeveloped. Sternites greyish; genitalia black. Eighth sternite weakly sclerotized with long, irregularlydistributed setae on posterior margin; gonapophyses subtriangular, half length of eighth sternite at midpoint,membranous, except sclerotized internal margins (Fig. 109). Cerci suboval, covered with distinct, long, brownsetae; paraproct subtriangular, nearly twice as long as cercus and weakly rounded apically, covered with longsetae basally and macrotrichia apically (Fig. 122). Genital fork sclerotized; termination of lateral arms withanterior processes well developed, subtriangular; area between lateral arms suboval (Fig. 136). Spermathecaoval, without external sculpturing and irregularly distributed spicules on internal surface; area of insertion ofspermathecal duct membranous.
Male. General body color black. Body length (specimen pinned) 2.5 mm (n=1); wing length 1.7 mm(n=1), wing width 1.0 (n=3).
Head—holoptic, with dark red eyes. Remainder of head coloration as in female.Thorax—scutum dark black covered with evenly distributed golden, recumbent hairs. Scutal pattern varies
with light incidence. With light source anterior to specimen scutum black, with 1+1 sublateral, grey-pruinosecunae on anterior margin; humeri, lateral and posterior margins black (Fig. 152). With light posterior, thoraxblack; humeri, lateral and posterior margins of scutum black. Scutellum dark brown covered with recumbent,golden hairs interspersed with long, erect dark brown setae (Fig. 153). Postnotum dark brown, with silverygrey pruinosity. Pleura black, with grey pruinosity. Halteres whitish yellow, with light brown base. Wingsetation as in female except Sc bare. Leg coloration as in female except coxae, trochanters, femora and mostof tibia dark brown, nearly entire hind of femur pale brown; hind femur and tibiae covered with slightlyspatulate setae.
Abdomen—tergites dark brown to black, with silver pruinosity on anterolateral margins of segments III, IVand VII [when specimen viewed laterally segment II dark brown and silver pruinose laterally], basal fringe oflong, brown hairs. Genitalia black; sternites whitish pruinose; tergal plates developed. Gonocoxitesubrectangular; gonostyle subcylindrical and pointed apically, longer than gonocoxite, weakly curved apicallyand terminating in single, stout spine; gonocoxite and gonostyle covered with long setae (Fig. 171). Ventralplate crescent-shaped, weakly sclerotized, with main body well developed centrally and median concavity onanterior margin; basal arms well developed, lightly sclerotized and rounded; main body of ventral platecovered with long hairs (Fig. 185). Median sclerite not examined. Paramere with developed and sclerotizedbasal process, and numerous long and short spines mesally and distally; membrane between basal arms ofparamere with thin spicules (Fig. 196).
Pupa. Cocoon length dorsally 2.7–3.1 mm (mean= 3.0 mm, s.d.=0.11, n=4); ventrally 2.8 mm (n=1); pupalength 2.7–3.8 mm (n=2); gill length 1.5–1.9 mm (mean=1.8 mm, s.d.=0.22, n=4).
Cocoon—slipper shaped, light brown, with anterior projection; cocoon composed of thin, coalescentnetwork of fibers, with lightly reinforced rim to anterior aperture.
Gill—bifid, primary branches initially orientated in dorsal and ventral planes and then forwardly directed,irregular in shape with dorsal primary branch narrowing apically. Dorsal primary branch bifurcating basallyforming tapering secondary branches; upper branch bifurcating distally and lower branch bifurcating basally.Ventral primary branch short and bulbous, with two short, pointed filaments apically. Variation in this patternoccurs with ventral primary branch having the two terminal filaments of different size, often the anteriorshorter than posterior. Gill surface glabrous (Fig. 211).
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 23NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
Head—frontoclypeus with 2+2 long, simple frontal and 1+1 long, simple, facial trichomes; frontoclypeuswith group of platelets mesally, 1+1 groups of approximately six platelets dorsolaterally and two groups ofplatelets in groups of 2–3 laterally in frontal region; tubercles rounded, densely distributed over entire surface.Thorax—approximately 1–2, long, single trichomes along dorsal cleft, 2–3 single or bifid trichomes centrallynear gill base, one long, single trichome on posterior region mesally, and two small, simple and one long,simple trichomes near ventral margin; tubercles rounded and densely distributed over entire surface of thorax.
Abdomen—[Chaetotaxy similar to S. petropoliense], tergite I with 1+1 long, sublateral, simple trichomes;tergite II with 3+3 submedian, spiniform setae in row, 3+3 small, simple trichomes in vertical line tospiniform outer setae, and 3+3 spiniform setae on lateral margin; tergites III–IV with 4+4 submedian, simplehooks in row along posterior margin and 1+1 simple trichomes anterior to outer trichomes; tergites V–VIwithout trichomes or setae; tergites VII and VIII without visible trichomes, but well-developed spine combsresembling teeth on anterior margin; tergite IX weakly sclerotized, with well-developed spine combsresembling teeth on anterior margin and 1+1 small, apical spines Abdominal sternite III with 3+3 submedianspiniform setae; sternites IV–V with 2+2 adjacent, bifid hooks; sternites V–VI with 2+2 well separated hookson posterior margin, outermost on right side being bifid while remaining hooks are single; sternites VII–VIIIwith visible spine combs, remaining segments and sternite VII–IX damaged in the few specimens examined.
Taxonomic discussionSimulium souzalopesi was described by Coscarón (1980) based on several reared males and females, and
larvae collected in Petrópolis, Rio de Janeiro State. We have examined the holotype and several paratypeswhich are housed in the MLP. The holotype is in good condition, with the pupa glued to a card point attachedto the pin (see Material Examined). In the MLP holdings, one female paratype of S. souzalopesi is labelledas the holotype of S. petropoliense. This problem has now been corrected and explained in Hernández et al.(2006) and the specimen is now correctly labelled as S. souzalopesi.
The adults of this species cannot be separated without examination of the pupal gill filaments. Simuliumsouzalopesi is similar to S. petropoliense and S. botulibranchium in having stout pupal gill filaments.However, S. souzalopesi can be recognized by the division of the dorsal primary branches in the basal third ofthe gill into two secondary branches, both having two filaments each, and gill filaments bare (Fig. 211) (seealso Taxonomic discussion in S. petropoliense and S. botulibranchium). Simulium souzalopesi is placed in thebotulibranchium species group by Crosskey & Howard (2004), who followed Coscarón (1980, 1987, 1991)[see also Coscarón & Coscarón-Arias, 2007]. These authors included S. souzalopesi in this species groupbecause of a subtriangular paraproct having a basal protuberance in the female (Fig. 122), male withsubcylindrical gonostyle, which is as long as the gonocoxite (Fig. 171), and the pupal gill filaments irregularin shape (Fig. 211). We agree with the placement of S. souzalopesi in this species group.
Descriptions of the life stages of S. souzalopesi may be found in Coscarón (1980; 1991), Coscarón &Coscarón-Arias (2007) and Strieder & Py-Daniel (1999; 2000).
DistributionSimulium souzalopesi has only been collected in Brazil near Petrópolis, Rio de Janeiro State (Crosskey &
Howard, 1997; 2004; Coscarón, 1980, 1987, 1991; Coscarón & Coscarón-Arias, 2007; Strieder & Py-Daniel,2000; Material Examined).
Biology and medical importanceCoscarón (1980, 1991) and Strieder & Py-Daniel (1999) stated that the immature stages of Simulium
souzalopesi can be collected in small trickles with clear water, commonly found attached to vegetation androcks, sympatric with S. botulibranchium and S. petropoliense. It has also been collected attached to leaves
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.24 · Zootaxa 1649 © 2007 Magnolia Press
and trailing vegetation at the edge of small rapids in clear waters with rocky riverbeds (unpublished data). Thefeeding habits of adult females are unknown.
INAEQUALE species group
This group consists of 11 species in the Neotropical Region. Two of these species, S. lurybayae and S. parimaense, are here considered as species inquirendae (see Taxonomic Discussion under these species).
Simulium (Inaequalium) clavibranchium Lutz(Figs. 4, 18, 37, 38, 76–78, 110, 123, 137, 154, 155, 172, 186, 197, 212, 213)
This is a common species in the southern part of Brazil.
Simulium clavibranchium Lutz, 1910: 257. LECTOTYPE pupal exuviae. BRAZIL: Rio de Janeiro State, Serra de Petró-polis [Without date or collector.] (IOC no. “12.053, Bd 104”, “225”) New type designation.
Female. General body color black. Body length (specimens pinned) 1.7–2.2 mm (mean=1.9 mm,s.d.=0.19, n=4); wing length 1.7–2.5 mm (mean= 2.0 mm, s.d.=0.16, n=5); wing width 0.8–1.1 mm (mean=0.9 mm, s.d.=0.13, n=4).
Head—dichoptic, with dark red eyes and nudiocular area well developed (Fig. 4). Frons, clypeus andocciput black, with silvery grey pruinosity; frons, clypeus and occiput with semi-recumbent, black hairs.Mouthparts black. Antennae with scape and pedicel yellowish brown; remainder of flagellar segments black.Cibarium with well-developed, sclerotized cornuae and 1+1 groups of sharp teeth of varying size extendingfrom base of cornuae to central area of cibarium, which is weakly protuberant (Fig. 18).
Thorax—scutum black covered with recumbent, golden hairs interspersed with black hairs. Scutal patternweak and variable depending on light incidence. With anterior illumination, scutum black; humeri and lateralmargins weakly pruinose; posterior margin black (Fig. 37). With posterior illumination, thorax black, with1+1 median, grey pruinose vittae on anterior region of scutum; humeri and lateral margins weakly pruinose;posterior margin black (Fig. 38). Scutellum dark brown to black, with recumbent, golden hairs interspersedwith long, black bristles. Postnotum black, with grey pruinosity. Pleura black, with silver pruinosity. Costa ofwing with sparse distribution of spines and setae. Subcosta with line of setae except apical third. Radius withrow of setae intermixed with distinct spines, basal section of radius with line of setae. Basal tuft of sparse,light brown setae. Leg coloration and proportions as in Figs. 76–78. Foreleg with coxa, trochanter, femur andventral surface of tibia brown; apical third of tibia and tarsal segments I–IV dark brown to black; basal twothirds of tibia whitish. Mid and hind legs with coxa, trochanter, femur, apical half of tibia, apical fourth ofbasitarsus, and tarsal segments II–IV dark brown; basal third of tibia and three fourths of tarsal segment Iwhitish. Mid and hind femora and tibiae covered with weakly spatulate setae. Claws weakly curved withsmall basal tooth. Halteres lemon yellow, with dark brown base.
Abdome – tergites II–V velvet black, tergites VI-IX shiny black; tergite II with silver spot on each side.Tergal plates developed. Sternites greyish. Genitalia black. Eighth sternite weakly sclerotized with long,irregularly distributed setae on posterior margin; gonapophyses subtriangular with inner, posterior marginsdistended posteriorly, half length of eighth sternite at midpoint, membranous with microtrichiae andsclerotized internal margins (Fig. 110). Cerci hemispherical, covered with distinct, long brown setae;paraproct subrectangular and slightly pointed ventrally, twice as deep as cercus, weakly sclerotized, coveredwith long setae basally and macrotrichiae on anterior margin (Fig. 123). Genital fork weakly sclerotized withends of lateral arms oblique to transverse axis of abdomen, anterior processes of lateral arms triangular andwell developed, posterior processes partially developed (Fig. 137). Spermatheca oval, without external
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 25NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
sculpturing and irregularly distributed spicules on internal surface; area of insertion of spermathecal ductmembranous.
Male. General body color black. Body length (specimens pinned) 2.0–2.2 mm (n=2); wing length 1.2–1.4mm (n= 2), wing width 0.9–2.2 (n= 2).
Head—holoptic, with dark red eyes. Remainder of head coloration as in female.Thorax—scutum black covered with evenly distributed golden, recumbent hairs. Scutal pattern varies
slightly with light incidence. With light source anterior to specimen scutum black, with indistinct 1+1submedian silver pruinose cunae on anterior margin; humeri, lateral and posterior margins black (Fig. 154) [Insome specimens the silver pruinose cunae are less prominent]. With light posterior, thorax black; humeri,lateral and posterior margins of scutum black (Fig. 155). Scutellum dark brown to black covered withrecumbent, golden hairs interspersed with long, erect dark brown setae. Postnotum black, with silvery greypruinosity. Pleura black, with grey pruinosity. Halteres whitish yellow with light brown base. Wing setationas in female except Sc bare. Leg coloration as in female.
Abdomen—tergites black, basal fringe of long, brown hairs. Silver pruinose ornamentation on posteriormargin of tergite I, anterolateral margin of tergite II and anterior margin of tergites IV–V, nearly entiresurface of tergite VI, and latero-ventral margins of VII–IX. Genitalia black; sternites whitish pruinose; tergalplates developed. Gonocoxite subrectangular; gonostyle subtrapezoidal with apical protrusion terminating insingle, stout spine and nearly same length as gonocoxite at mid point; gonocoxite and gonostyle covered withlong setae (Fig. 172). Ventral plate sclerotized and crescent-shaped with pointed apex and basal arms welldeveloped, and main body covered with long hairs (Fig. 186). Median sclerite pear-shaped, as long as wide atmidpoint, with deep apical incision (Fig. 197). Paramere with developed and sclerotized basal process, andnumerous long and short spines mesally; membrane between basal arms of paramere with thin spicules (Fig.197).
Pupa. Cocoon length dorsally 2.6–3.7 mm (mean=2.9 mm, s.d.=0.27, n=13); ventrally 2.9–3.3 mm(mean=3.1 mm, s.d.=0.16, n= 13); pupa length 3.2–3.7 mm (mean=3.4 mm, s.d.=0.21, n=13); gill length 2.6–4.3 mm (mean=3.3 mm, s.d.=0.49, n=15).
Cocoon—slipper shaped, light to dark brown, composed of a thin network of fibers with coalescent fibersand reinforced rim to anterior aperture.
Gill—light brown, with six long, symmetrical, and forwardly directed filaments arranged in vertical plane.[specimens with five gill filaments are recorded by Coscarón & Wygodzinsky (1984), Coscarón (1991) andStrieder & Py-Daniel (1999; 2000)]. Gill configuration variable, with main trunk short, giving rise basally totwo sets of primary branches: one dorsal and stout and one ventral that is thinner; dorsal primary branchalmost immediately divides into two secondary branches, one dorsal that bifurcates towards gill apex and oneventral that bifurcates apically. Ventral primary branch bifurcates near base of gill and secondary filamentsdirected towards frontal region of pupa (Figs. 212, 213). All filaments slightly pointed distally, with smallspicules on surface, edges crenate; filaments of dorsal set of primary branches of same length but relativelylonger than filaments of ventral primary branch. All filaments slender, abruptly expanded apically, withpointed apex, covered with spinules on surface, edges weakly crenate. All filaments of approximately samelength.
Head—frontoclypeus with 2+2 bifid to trifid frontal and 1+1 trifid facial trichomes; frontoclypeus withgroup of platelets mesally, 1+1 groups of approximately six platelets dorsolaterally and two to three plateletsin groups of two laterally in frontal region; tubercles rounded and densely distributed over entire surface.
Thorax—approximately 5+5 large, trifid to multibranched trichomes near margin of dorsal cleft, a long,simple trichome on central region and four long, simple trichomes near alar region and base of gill; tuberclesrounded and densely distributed over entire surface.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.26 · Zootaxa 1649 © 2007 Magnolia Press
Abdomen—tergite I with 1+1 long, lateral, simple or bifid trichomes; tergite II with 3+3 submedianspiniform setae in a row, 2+2 spiniform setae anterior to the outer setae, and 1+1 single setae on lateralmargin; tergites III and IV with 4+4 submedian, simple hooks in row along posterior margin, 1+1 spiniformsetae anterior to outer hooks, 3+3 sublateral, small, spiniform setae in a row, and 1+1 simple setae on lateralmargin; tergite V with 2+2 submedian, small simple trichomes; tergite VI with 2+2 submedian, simpletrichomes along posterior margin and 1+1 groups of spine combs on anterolateral margins; tergites VII andVIII with 2+2 small, submedian simple trichomes on posterior margin and well-developed spine combsresembling teeth on anterior margin; tergite IX weakly sclerotized, with well-developed spine combsresembling teeth on anterior margin and 1+1 small, apical spines. Abdominal sternite III with 3+3 sublateral,simple trichomes; sternite IV with 2+2 median and 2+2 sublateral, simple spiniform setae on posteriormargin; sternite V with 2+2 submedian, close bifid hooks on posterior margin; sternites VI–VII with 2+2 wellseparated hooks on posterior margin, the inner being bifid or trifid and the outer simple, 1+1 or 2+2 simple,small trichomes anterior to outer hooks, 1+1 small, simple trichomes between outer hooks, and 1+1 simpletrichomes on lateral margins; sternite VIII with no visible setae or trichomes; sternite IX weakly sclerotized.Spine combs visible on anterior margin of sternites IV–IX.
Taxonomic discussion Lutz (1910) described Simulium clavibranchium from pupae collected in Rio de Janeiro State: Petrópolis,
Morro do Observatório and Capivary Station (between Barra Mansa & Angra dos Reis) and São Paulo State,Serra da Bocaina. In this paper, he stated that he was unable to obtain adults from mature pupae, butmentioned that in pharate adults there were no “petaloid” scales on the hind legs, and the extremity of the hindfemur was always darkly colored, as in S. auristriatum or S. distinctum.
We have examined in the Lutz collection at the IOC numerous specimens identified as S. clavibranchiumand we have the following comments to make. There are three slides containing pupal exuviae and apparentlywith Lutz’ original labels “225”, “227” and “233” that agree with our current concept of S. clavibranchium.Only one slide (“no. 225”) bears a label with locality data “Serra de Petrópolis”. The pupal exuviae is in goodcondition and has been remounted in Canada Balsam by an unknown reviser. The two other slides “227 and“233” do not have a locality, date or collector’s name. However, we believe that slide “227” is part of thesyntype series, because Lutz referred in his description to the branching of trichomes and there is a label onthe slide to this effect (Material examined). The absence of labels on slide “233” precludes its recognition asa syntype.
Two other slides with Lutz’ original labels “226” and “232” each containing a female are also present inthe Lutz collection at the IOC. Slide “226” with “Morro do Observatório” in Lutz’ hand contains a dissected,pharate female. Since this was referred to in Lutz (1910) the specimen is considered a syntype. We arediscounting the other slide with Lutz’ label “232” containing a female [possibly not pharate] as a syntype,because there is no other label on the slide. We also examined two females collected from “Petrópolis” in1910 [in Lutz’ hand] with a subsequently added printed, purple label stating “Tipo”. We are not aware whoadded these labels, but these specimens are not part of the syntype series because Lutz only mentioned pharateadults in his description of S. clavibranchium. We have selected the best pupal exuviae [slide number “225”]as the lectotype because the species name is based on this life stage (see Fig. 212). The followings slides havebeen labelled as paralectotypes: “226” and “227” (see Material Examined.). Other specimens of S.clavibranchium exist in the Lutz collection at the IOC, but they have no type status and therefore are notdetailed here (see Material Examined).
Coscarón & Wygodzinsky (1984) listed S. mbarigui described by Coscarón & Wygodzinsky in 1973 as asynonym of S. subnigrum, but it was considered as a junior synonym of S. clavibranchium in a later paper (seeCoscarón, 1987). However, this synonymy was not included in Coscarón (1991), but is again cited as asynonym of S. subnigrum in Coscarón & Coscarón-Arias (2007). We have examined the type material of S.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 27NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
mbarigui and studied the original description and illustrations in Coscarón & Wygodzinsky (1973) and agreethat it falls within the variation found in S. subnigrum (see Hernández et al., 2007 for comments and figureson the holotype).
The morphological variation of S. clavibranchium has been recently discussed by Hernández et al. (2007)who also compared it with S. subclavibranchium and discussed the taxonomic confusion between this speciesand S. subclavibranchium [now a synonym of S. subnigrum] and several other species of Inaequalium in theNeotropical Region. The adults of S. clavibranchium are difficult to identify without examination of the pupalgill filaments. This species is best recognized by the pupal gill filaments enlarged apically (Fig. 212).However, variation in this pattern occurs with specimens having some filaments not apically expanded (Fig.213).
Simulium clavibranchium belongs to the inaequale group based on the combination of characters given inCoscarón & Wygodzinsky (1984) and Coscarón (1991). Description of adults and immature stages of S.clavibranchium may be found in Coscarón (1991), Coscarón & Coscarón-Arias (2007), Coscarón &Wygodzinsky (1984), and Strieder & Py-Daniel (1999, 2000).
DistributionSimulium clavibranchium is a relatively common species in Brazil in the states of Minas Gerais, Rio de
Janeiro, Rio Grande do Sul, Rondônia, São Paulo and Santa Catarina (Coscarón, 1991; Coscarón & Coscarón-Arias, 2007; Crosskey & Howard, 1997; 2004; Strieder & Py-Daniel, 2000; Material Examined).
Biology and medical importance In Brazil, the immature stages of S. clavibranchium have been found attached to vegetation in small to
medium fast flowing, clear waters streams or waterfalls mostly with rocky riverbeds (A.J. Shelley, A.P.A.Luna Dias, P.R.Garritano, M. Maia-Herzog & L.M. Hernández, unpublished data). Nothing is known aboutthe feeding habits of females of S. clavibranchium. They have never been collected biting humans.
Simulium (Inaequalium) diversibranchium Lutz(Figs. 5, 19, 39, 40, 79–81, 111, 124, 138, 156, 157, 173, 187, 198, 214)
Simulium diversibranchium Lutz, 1910: 238. LECTOTYPE pupa, BRAZIL: São Paulo State, Bonito; [Without date orcollector’s name] (IOC, no. 12.097). [Examined.] New lectotype designation.
Simulium missionum Coscarón, 1976: 151. HOLOTYPE % (reared), ARGENTINA: Missiones, Campiña S. de B. deIrigoyen; 21.vii.1972 (Coscarón) (MLP). [Examined.] [Synonymy by Coscarón & Wygodzinsky, 1984: 93.]
Female. General body color black. Body length 2.1 mm (n=1) (dry specimen), 2.5 mm (n=1) (in spirit);wing length 2.1–2.2 mm (n=2), wing width 1.1 mm (n=2).
Head—dichoptic, with dark red eyes and nudiocular area well developed (Fig. 5). Frons, clypeus andocciput black, with silvery grey pruinosity; frons and clypeus with pale, semi-recumbent hairs, slightly longeron occiput. Mouthparts black. Antennae with scape, pedicel yellowish brown; remainder of flagellar segmentsblack. Cibarium with well-developed, sclerotized cornuae and rows of sharp teeth of varying size extendingfrom base of cornuae to central area of cibarium, which is protuberant (Fig. 19).
Thorax—scutum black, covered with golden, recumbent hairs sometimes with greenish reflections(depending on lighting). Scutal pattern varies with direction of illumination. With anterior illumination,scutum, humeri, lateral and posterior margins greyish black (Fig. 39). With posterior illumination, thoraxblack, with 1+1 faintly grey pruinose bands on anterior third of scutum; humeri and lateral margins greypruinose (Fig. 40). Scutellum dark brown, with recumbent golden hairs and several dark brown, long bristlesmainly on posterior margin. Postnotum black, with grey pruinosity. Pleura black, with silver pruinosity. Costaof wing with sparse distribution of spines and setae. Subcosta with row of setae. Radius with row of setae
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.28 · Zootaxa 1649 © 2007 Magnolia Press
intermixed with several distinct spines, basal section of radius with row of setae. Basal tuft of sparse, lightbrown setae. Leg coloration and proportions as in Figs. 79–81. Foreleg with coxa light brown; trochanter,femur, apical third of tibia dark brown, with dorsal surface of tibia whitish and tarsal segments black. Mid legwith coxa, trochanter, femur and apical half of tibia, distal tip of basitarsus and tarsal segments mid brown,basal half of tibia and most of tarsus whitish. Hind leg with coxa, trochanter, femur, apical two thirds of tibia,apical third of basitarsal segment and tarsal segments II–IV dark brown; remainder of tibia and basitarsuswhitish. Claws curved with small basal tooth. Halteres lemon yellow, with dark brown base.
Abdomen—tergites I–IV velvet black, tergites V–IX shiny black. Tergal plates weakly developed in thefew specimens examined. Sternites and genitalia black. Eighth sternite weakly sclerotized, with long,irregularly distributed setae on posterior margin; gonapophyses subtriangular, with inner posterior marginsdistended posteriorly, nearly of same length as sternite at midpoint, membranous with microtrichiae, internalmargins sclerotized (Fig. 111). Cerci hemispherical, covered with distinct, long brown setae; paraproctsubrectangular and rounded ventrally, twice as deep as cercus, weakly sclerotized, covered with long setaebasally and macrotrichiae on anterior margin (Fig. 124). Genital fork weakly sclerotized, with ends of lateralarms parallel to transverse axis of abdomen, anterior processes of lateral arms triangular and well developed,posterior processes partially developed (Fig. 138). Spermatheca suboval, apparently without externalsculpturing, and with irregularly distributed spicules on internal surface; area of insertion of spermathecalduct membranous.
Male. General body color black. Body length (pinned specimens) 1.9–2.3 mm (mean=2.1 mm, n=3); winglength 1.9 mm (n=1), wing width 0.9 mm (n=1).
Head—holoptic, with dark red eyes. Remainder of head coloration as in female.Thorax—scutum black, covered with golden, evenly distributed, recumbent hairs. Scutal pattern varies
slightly with light incidence. With light source anterior to specimen scutum black, with weak 1+1 sublateralwhite, pruinose round areas on anterior margin; humeri, lateral and posterior margins black (Fig. 156). Withlight source posterior thorax black; humeri, lateral and posterior margins of scutum black (Fig. 157).Scutellum dark brown covered with golden, recumbent hairs interspersed with long, erect, dark brown setae.Postnotum black, with silvery grey pruinosity. Pleura black, with grey pruinosity. Halteres whitish yellow,with light brown base. Wing setation as in female except Sc bare. Leg coloration similar to female.
Abdomen—tergites black, basal fringe of long, brown hairs. Silver pruinose ornamentation on anterolateralmargin of tergite II and ventrolateral margins of tergites V–VII. Genitalia black; sternites whitish pruinose;tergal plates developed. Gonocoxite subrectangular; gonostyle subtrapezoidal with apical protrusionterminating in single, stout spine and two thirds length of gonocoxite; gonocoxite and gonostyle covered withlong setae (Fig. 173). Ventral plate sclerotized and crescent-shaped, with pointed apex and basal arms welldeveloped, and main body covered with long hairs (Fig. 187). Median sclerite pyriform, with apical incision(Fig. 198). Paramere with developed and sclerotized basal process, and developed spines; membrane betweenbasal arms of paramere with thin spicules (Fig. 198).
Pupa. Cocoon length dorsally 2.7–3.3 mm (mean = 3.1 mm, s.d.=0.22, n=5); ventrally 3.3–3.8 mm
(mean=3.5 mm, s.d.=0.25, n= 5); pupa length 2.5–3.7 mm (mean= 3.2 mm, s.d.=0.44, n=6); gill length 1.8–3.6 mm (mean=2.7 mm, s.d.=0.65, n=5).
Cocoon—slipper shaped, light to dark brown composed of a thin network of coalescent fibers, withreinforced rim to anterior aperture.
Gill—light brown, with forwardly directed six filaments arranged in vertical plane [specimens with 5 gillfilaments are recorded by Coscarón & Wygodzinsky (1984), Coscarón (1991) and Strieder & Py-Daniel(1999; 2000)]. Gill configuration variable, with main trunk short, giving rise basally to two sets of primarybranches: one dorsal and stout and one ventral that is thinner; dorsal primary branch almost immediately
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 29NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
divides into two secondary branches, one dorsal that bifurcates towards gill apex and one ventral thatbifurcates apically. Ventral primary branch bifurcates near base of gill and secondary filaments often directedtowards frontal region of pupa (Figs. 214) [In some specimens these are weakly forwardly directed]. Allfilaments weakly pointed distally, with small spicules on surface, edges crenate; filaments of dorsal set ofprimary branches of same length but relatively longer than filaments of ventral primary branch.
Head—frontoclypeus with 2+2 trifid or 4-branched frontal and 1+1 4-branched facial trichomes;frontoclypeus with group of platelets mesally, 1+1 groups of 10 platelets dorsolaterally and 2–3 platelets ingroups of two laterally in frontal region; tubercles rounded and densely distributed over entire surface.
Thorax—5+ 5 large, bifid to multiramous trichomes near margin of dorsal cleft; one long, bifid trichomenear gill base and one long, simple trichome on ventral margin; tubercles rounded and densely distributedover entire surface of thorax.
Abdomen—abdominal tergite I with 1+1 long, simple trichomes on anterolateral margin; tergite II with2+2 sublateral, simple trichomes on anterior margin; tergite III with 4+4 submedian, simple hooks in rowalong posterior border, 3+3 simple, small trichomes in row anterior to innermost hooks, 1+1 or 2+1 small,simple trichomes on middle and anterior to outer hooks, and 2+2 lateral, short simple trichomes on lateralmargin; tergite IV with 4+4 submedian, simple hooks in row along posterior border, 1+1 simple, small setaeanterior to most lateral hooks and three sublateral, short trichomes; tergite V with five small, simple setaealong posterior margin; tergites VI–VIII with distinct spine combs, some well developed, resembling teeth onanterior margin; tergite IX with spine combs and 1+1 small apical spines. Abdominal sternite III with 3+3median and 2+2 lateral, small, simple trichomes; sternite IV with 3+3 median and 3+3 sublateral, smallsimple trichomes near posterior margin; sternite V with 2+2 close, submedian bifid hooks on posterior marginand 1+1 small, simple trichomes anterior to outer hooks; sternites VI–VII with 2+2 well separated hooks onposterior margin of which innermost hooks are bifid and outermost simple, 1+1 or 2+2 simple, smalltrichomes anterior to outer hooks, and 1+1 small, simple trichomes between outer hooks; sternite IX weaklysclerotized. Spine combs on anterior margins of sternites III–IX.
Taxonomic discussion Simulium diversibranchium was briefly described from several pupae collected near Bonito [Fazenda do
Bonito], Serra de Bocaina by Lutz in 1910. He also considered this material to be very close to a new species,S. infuscatum, that he described in the same paper. Coscarón & Wygodzinsky (1984) stated that theyexamined several pupae from Serra da Bocaina deposited in IOC that agreed with the morphologicaldescription of the pupa given by Lutz as S. diversibranchium. They also recorded that the adults that emergedfrom these pupae did not agree with the description of S. infuscatum and suggested that the material studied byLutz involved two different species. In addition, they pointed out that the material labelled as S. infuscatum inthe IOC is unidentifiable, but followed D’Andretta & D’Andretta (1949) in treating this name as a synonym ofS. auristriatum Lutz.
We have examined four slides containing several pupal exuviae and eight pinned specimens identified asS. diversibranchium in the Lutz collection at the IOC and have the following observations to make. Most ofthe pupal exuviae mounted on slides with IOC numbers “12.095”, “12.097” and “12.098” from Serra daBocaina all agree with the original description of S. diversibranchium except one pupal exuviae on slide“12.098” and two pupal exuviae on slide “12.100”. These appear to be S. subnigrum and they have beencircled in black ink and labelled accordingly. We have selected and labelled the single pupa collected from“Bonito, Serra de Bocaina” with IOC number “12.097” as the lectotype. We have selected a pupal exuviae aslectotype because Lutz based his description of S. diversibranchium on this life stage. The pupal exuviae is infairly good condition (see Material Examined). The remaining pupal exuviae on slides “12.095”, “12.097”and “12.098” have been labelled as paralectotypes.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.30 · Zootaxa 1649 © 2007 Magnolia Press
There are also eight pinned specimens identified as S. diversibranchium without locality data in the IOC[three males with IOC number “12.511”, and two males and three females with number “12.512”]. Thesespecimens are not part of the syntype series because Lutz did not refer to adults in his description of S.diversibranchium. The specimens are in poor condition and so we are unable to make further comments ontheir identity.
In their revision of the subgenera Psaroniocompsa and Inaequalium Coscarón & Wygodzinsky (1984)synonymized S. missionum with S. diversibranchium without explanation. Simulium missionum was describedby Coscarón (1976) based on a single, reared male and a pupal exuviae collected in Campiñas, 5 km S. de B.Irigoyen, Missiones, Argentina. The description of the pupa referred to the presence of five filaments. Wehave examined the holotype and its pupal exuviae, and a pupa [labelled as paratype], which are housed in theMLP. The holotype is in relatively good condition and has been glued to a card point by the left side. Threelegs, the genitalia and the pupal exuviae have been dissected and they are mounted on a slide. The thorax, bothwings and three legs of the left side remain pinned. The pupa is in relatively good condition although only fivefilaments are present on the slide (see Material Examined). We have studied the morphology of the thoracicpattern, male genitalia, and the pupal gill configuration of S. missionum and agree that they fall within thevariation of S. diversibranchium and accept Coscarón & Wygodzinsky’s (1984) synonymy.
The adult thoracic patterns of S. diversibranchium are very similar to other species in the inaequale-species group. The morphology of the female paraproct in our S. diversibranchium specimens is similar toother species of the inaequale group (e.g., S. inaequale, S. maranguapense, S. margaritatum, S.mariavulcanoae, S. rappae, S. subnigrum and S. travassosi (Figs. 125, 127, 128, 129, 131, 132, 133). Thegonostyle is similar to that of S. clavibranchium and S. mariavulcanoae although Coscarón & Coscarón-Arias, (2007) claim to see small differences in morphology. Their main criterion for distinguishing the speciesis the configuration of the pupal gill filaments. Variation in the number of gill filaments in S.diversibranchium has been recorded by Coscarón (1991), Coscarón & Wygodzinsky (1984) and Strieder &Py-Daniel (1999; 2000), who reported pupal exuviae with five or six gill filaments along the distributionrange of this species. However, all reared specimens we have examined of S. diversibranchium fromArgentina (Missiones) and Brazil have a six-filamented pupa. The ventral filaments of the dorsal set ofprimary branches are very thin and they easily break off giving the impression that the pupa only has fivefilaments, if the specimens are examined in alcohol or at low magnification. The base of these filaments isclearly visible if the specimens are mounted on a slide and examined at higher magnification. The pupa of S.diversibranchium is very similar to that of S. mariavulcanoae with differences in branching heights from thebase (Figs. 214, 218). This branching pattern has been the subject of discussion by various authors (Coscarón& Wygodzinsky, 1984: 97, Fig. 25, C–D; Strieder & Py-Daniel, 2000:71, Fig. 4, G). However, variation inthis pattern occurs in which a more splayed-out ventral primary branch has been recorded in S.mariavulcanoae that resembles that in S. diversibranchium (see Coscarón & Wygodzinsky, 1984; Fig. 25 E).Further studies on these two species are needed to observe pupal gill variation in order to establish whether S.diversibranchium and S. mariavulcanoae are conspecific.
Previous descriptions of adult and immature stages of S. diversibranchium can be found in Coscarón(1991), Coscarón & Coscarón-Arias (2007), Coscarón & Wygodzinsky (1984) and Strieder & Py-Daniel(1999, 2000). Coscarón & Coscarón-Arias (1997) and Strieder & Py-Daniel (2002) in their phylogeneticstudies of Inaequalium placed S. diversibranchium in the inaequale-group, closely related to S.mariavulcanoae.
Distribution Simulium diversibranchium has been recorded in Brazil from the states of Minas Gerais, Rio Grande do
Sul, Rio de Janeiro, São Paulo and Santa Catarina (Coscarón, 1991; Coscarón & Coscarón-Arias, 2007;
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 31NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
Crosskey & Howard, 2004; Strieder, 2004; Strieder et al., 1992; Material Examined). Elsewhere, it has beencollected in Argentina (Missiones).
Biology and medical importance In Brazil, the immature stages of S. diversibranchium have been found attached to leaves and branches in
small (50 cm wide, 10 cm deep), fast running, clear water streams with sandy riverbeds (A.J. Shelley, A.P.A. Luna Dias & M.Maia-Herzog, unpublished data).
Simulium (Inaequalium) inaequale (Paterson & Shannon) (Figs. 6, 20, 41, 42, 82–84, 112, 125, 139, 158, 159, 174, 175, 188, 199, 215)
Eusimulium inaequale Paterson & Shannon, 1927: 738. NEOTYPE & (pinned, not associated with pupal exuviae),ARGENTINA: Cerro San Jávier, Tucumán, 1200 m; 7.iv.1960, (Wygodzinsky) (MLP). [Examined.] [Neotype desig-nation by Hernández et al., 2007: 12.]
Simulium baiensis Pinto, 1932: 685. HOLOTYPE %, BRAZIL: Bahia State, Nazaré; 1931 (A. Lutz) (IOC). [Synonymyby Coscarón, 1991:157.]
Trichodagmia manicata Enderlein, 1934: 281. LECTOTYPE &, PARAGUAY: Hohenau, 250 m; 15.x.1907, [Withoutcollector’s name.] (SMT). [Synonymized with doubt by Coscarón & Wygodzinsky, 1984: 80; confirmed by Hernán-dez et al., 2007: 12.] [Lectotype designation by Werner, 1996b: 296-see also Werner, 1996a.].
Thyrsopelma argentata Enderlein, 1936:125. HOLOTYPE &, PERU: Hoch. [=Alto]; [Without date or collector’s name.](NMHU). [Synonymy by Coscarón & Miranda-Esquivel, 1998: 337.]
Simulium clarki Fairchild, 1940: 703. HOLOTYPE & (reared), PANAMA: Summit, C.Z.; 9.i.1940, [Without collector’sname, but probably collected by Fairchild] (MCZ, no. 25751). [Suggested as doubtful junior synonym of S. clarki(as I. clarki) with S. inaequale (as I. inaequale) by Strieder & Py-Daniel, 1999: 63, 2000: 27.] [Synonymy byHernández et al., 2007: 12.]
Simulium jundiaiensis D’Andretta & González B., 1964: 108. HOLOTYPE & (reared), BRAZIL: São Paulo State: Jun-diaí, Bairro Caxambú; 1.vii.1956, [Collector’s name not stated in the original description.] (MZUSP, no. 855). [Syn-onymy by Coscarón & Wygodzinsky, 1984: 80.]
Simulium pseudoexiguum Nunes de Mello & Barbosa de Almeida, 1974: 65. HOLOTYPE & (man-biting), BRAZIL:Roraima State, Boca de Mata, próximo à BR-174; xi.1972, (J.A.Nunes de Mello & Eduardo Vieira da Silva) (INPA,no. 5041). [Synonymy by Hernández et al., 2007: 13.]
Female. General body color black. Body length (specimens pinned) 1.7–2.3 mm (mean=2.1 mm,s.d.=0.17, n=12); wing length 1.6–2.2 mm (mean= 1.9 mm, s.d.=0.18, n=12); wing width 0.8–1.1 mm (mean=1.0 mm, s.d.=0.08, n=12).
Head—dichoptic, with dark red eyes and nudiocular area well developed (Fig. 6). Frons, clypeus andocciput black, with silvery grey pruinosity; frons and clypeus with pale, semi-recumbent hairs, slightly longeron occiput. Mouthparts black. Antennae with scape and pedicel yellowish brown; remainder of flagellarsegments black. Cibarium with well-developed, sclerotized cornuae and rows of sharp teeth of varying sizeextending from base of cornuae to central area of cibarium, which is protuberant (Fig. 20).
Thorax—black, covered with golden, recumbent hairs sometimes with greenish reflections (depending onlighting). Scutal pattern not clearly visible, especially in well preserved specimens. With anteriorillumination, scutum black, with greyish pruinosity and median dark line and 1+1 submedian, black bands inform of lyre running from anterior margin to posterior (Fig. 41). With posterior illumination, thorax greypruinose, lateral and posterior margins black (Fig. 42). Irrespective of light direction thin, black median lineextending from anterior to posterior margin. Scutellum dark brown, with recumbent golden hairs interspersedwith dark brown long bristles. Postnotum black, with grey pruinosity. Pleura black, with silver pruinosity.Costa of wing with sparse distribution of spines and setae. Subcosta with line of setae, except apical third,which is bare. Radius with row of setae intermixed with several distinct spines, basal section of radius withline of setae. Basal tuft of sparse, light brown setae. Leg coloration and proportions as in Figs. 82–84. Foreleg
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.32 · Zootaxa 1649 © 2007 Magnolia Press
coxa light brown; trochanter, femur basally and middle of tibia yellowish, remainder of femur and tibia midbrown; tarsal segments dark brown to black. Mid leg with coxa, apical two thirds of tibia, apex of basitarsusand tarsal segments II–IV mid to dark brown; basal third of tibia and two thirds of basitarsus whitish. Hind legwith coxa, trochanter, femur, apical two thirds of tibia, apical third of basitarsus and tarsal segments II–IVdark brown; basal third of tibia and two thirds of basitarsus whitish. Claws weakly curved, with small basaltooth. Halteres lemon yellow, with dark brown base.
Abdomen—tergites I–IV velvet black, V–IX shiny. Tergal plates developed. Sternites and genitalia black.Eighth sternite weakly sclerotized, with long irregularly distributed setae on posterior margin; gonapophysessubtriangular with inner, posterior margins distended posteriorly, nearly of same length as sternite atmidpoint, membranous with microtrichiae, sclerotized internal margins (Fig. 112). Cerci hemispherical,covered with distinct, long brown setae; paraproct subrectangular and rounded ventrally, almost twice as deepas cercus, weakly sclerotized, covered with long setae basally and macrotrichiae on anterior margin (Fig.125). Genital fork weakly sclerotized with ends of lateral arms parallel to transverse axis of abdomen, anteriorprocesses of lateral arms triangular and well developed, posterior processes partially developed (Fig. 139).Spermatheca suboval, without external sculpturing and irregularly distributed spicules on internal surface;area of insertion of spermathecal duct membranous.
Male. General body color dark brown to black. Body length (pinned specimens) 1.8–2.6 mm (mean=2.2mm, s.d.=0.22, n=9); wing length 1.5–2.0 mm (mean=1.9 mm, s.d. = 0.13, n=9), wing width 0.9–1.1 mm(mean= 1.0 mm, s.d.=0.06, n=9).
Head—holoptic, with dark red eyes. Remainder of head coloration as in female.Thorax—scutum dark brown to black covered with golden, recumbent hairs. Scutal pattern varies slightly
with light incidence. With anterior light source scutum black, with 1+1 submedian silver pruinose cunae onanterior third (Fig.158). With posterior light source scutum black (Fig.159). Scutellum dark brown coveredwith recumbent, golden hairs intermixed with long, erect, dark brown setae. Postnotum black, with silverygrey pruinosity. Pleura black, with grey pruinosity. Halter whitish yellow, with light brown base. Wingsetation as in female, except Sc bare. Leg coloration as in female except nearly entire hind femur dark brownand slightly spatulate setae on hind femora and tibiae.
Abdomen—tergites black, basal fringe of long, brown hairs. Silver pruinose ornamentation as follows:whole of tergite II except for median part and ventrolateral margin of tergite VI. Genitalia black; sternitesgreyish pruinose; tergal plates developed. Gonocoxite subquadrangular; gonostyle between subtrapezoidalwith apical protrusion and pyriform terminating in single stout spine and two thirds length of gonocoxite [insome specimens up to three spines can be found]; gonocoxite and gonostyle covered with long setae (Figs.174, 175). Ventral plate sclerotized and crescent-shaped with pointed apex and basal arms well developed,and main body covered with long hairs (Fig. 188). Median sclerite pyriform with apical incision (Fig. 199).Paramere with developed and sclerotized basal process, and developed spines; membrane between basal armsof paramere with thin spicules (Fig. 199).
Pupa. Cocoon length dorsally 2.3–3.4 mm (mean=3.0 mm, s.d.=0.31, n=17); ventrally 2.5–4.2 mm(mean=3.5 mm, s.d.=0.50, n=17); pupa length 2.8–4.3 (mean=3.5 mm, s.d.=0.49, n=17); gill length 2.5–4.9mm (mean=3.6 mm, s.d.=0.53, n=20).
Cocoon—slipper shaped, light to dark brown composed of a thin network of fibers and reinforced rim toanterior aperture.
Gill—light brown, with six forwardly directed filaments arranged in vertical plane. Gill configuration withmain trunk short, giving rise to three sets of primary branches, two external and one internal, all sets ofprimary branches bifurcating near gill base at different heights, with internal often slightly more apical than
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 33NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
external set of filaments (Fig. 215). All filaments weakly rounded distally with small spicules on surface,edges crenate; all filaments of same length.
Head—frontoclypeus with 2+2 bifid (sometimes trifid) frontal and 1+1 bifid, facial trichomes;frontoclypeus with group of platelets mesally, 1+1 groups of approximately 6 platelets dorsolaterally and 2–3platelets in groups of two laterally in frontal region; tubercles rounded and densely distributed over entiresurface.
Thorax—with up to 5+ 5 large, bifid to multiramous trichomes near margin of dorsal cleft, one long,simple trichome on posterior region mesally and three long, simple trichomes near base of gill ventrally;tubercles rounded and densely distributed over entire surface.
Abdomen—abdominal tergite I with 1+1 long, simple or bifid trichomes on anterolateral margin [in somespecimens left side with one long, bifid trichome and right side with one long, simple trichome]; tergite IIwith 4+4 submedian spiniform setae in row, outer setae more spaced than inner setae, and 1+1 spiniform setaeanterior to outer setae; tergites III and IV with 4+4 submedian, simple hooks in row along posterior margin,and 1+1 simple setae anterior to outer hooks; tergite V with 2+2 or 4+4 sublateral, simple setae; tergites VI–VIII with well-developed spine combs resembling teeth on anterior margin; tergite IX weakly sclerotized,with well-developed spine combs resembling teeth on anterior margin and 1+1 small, apical spines.Abdominal sternite III with 1+1 median and 2+2 lateral, small, simple trichomes; sternite IV with 1+1 medianand 2+2 sublateral, small simple trichomes; sternite V with 2+2 submedian, close bifid hooks on posteriormargin and 2+2 small, simple trichomes anterior to outer hooks; sternites VI–VII with 2+2 well separatedhooks on posterior margin, the inner being bifid and the outer simple, 1+1 or 2+2 simple, small trichomesanterior to outer hooks, and 1+1 small, simple trichomes between outer hooks; sternite IX weakly sclerotized.Spine combs on anterior margin of sternites III–IX.
Taxonomic discussionA comprehensive revision of S. inaequale with the designation of a neotype and the creation of two new
synonyms was provided by Hernández et al. (2007). Like other species of the group it can only bedistinguished in reared specimens using the pupal gill characters (Fig. 215).
Simulium inaequale has been placed in the inaequale species group by Crosskey & Howard (2004),Coscarón (1987, 1991) and Coscarón & Coscarón-Arias (2007), which is followed in this paper.
Distribution Simulium inaequale is a common, widespread species in the Neotropical region. It has been recorded in
Argentina (Jujuy, Missiones, Salta, Tucumán), Bolivia, Brazil (Amapá, Bahía, Espírito Santo, Minas Gerais,Mato Grosso, Paraná, Paraíba, Rio Grande do Sul, Rio de Janeiro, São Paulo, Santa Catarina States), CostaRica, Ecuador, Guyana, Panama, Paraguay, Peru & Venezuela (Coscarón & Coscarón-Arias, 2007; Coscarónet al., 1992; Crosskey & Howard, 1997; 2004; Gil-Azevedo & Maia-Herzog, 2004; Material Examined).
Biology and medical importanceThe immature stages of S. inaequale have been found in small streams in Brazil and in 50 m wide rivers in
Guyana [as S. clarki] (Shelley et al., 2004). Coscarón & Wygodzinsky (1984) and Coscarón (1991) collectedpupae in small, clear water streams in Argentina. The females are highly anthropophilic throughout thespecies distribution range biting man voraciously in countries like Guyana (Shelley et al., 2004) and in theRiver Uiramutã, Roraima State Brazil together with S. nigrimanum, S. oyapockense s.l. & S. spinibranchium(A.J. Shelley & A.P.A. Luna Dias, unpublished data). Coscarón (1991) recorded this species biting mules inBolivia. Coscarón & Coscarón-Arias (2007) stated that the female can reach pest levels in Santo Inácio (BahiaState, Brazil). Pepinelli & Trivinho-Strixino (2002) suggested that the females showed a tendency to lay eggson dark substrates.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.34 · Zootaxa 1649 © 2007 Magnolia Press
Simulium (Inaequalium) maranguapense (Pessoa, Ríos-Velásquez & Py-Daniel)(Figs. 8, 22, 88–90, 114, 127, 141, 216)
This is a recently described species of Inaequalium from Ceará State, Brazil. Since material was not availablefor this study we base our description and figures on the original description.
Inaequalium maranguapense Pessoa et al. 2005: 5. HOLOTYPE pharate & and pupal exuviae. BRAZIL, Ceará State,
Maranguape municipality, tributary of Gavião River, near Erundina Hotel, Balneario Cascatinha, Maranguapemountain, approximately 600m, 3º53’27’’S 34º41’08’’W; ii.2005, (Felipe Pessoa & Claudia Rio Velásquez) (INPA,no. 6286).
Female. General body color dark. Wing length (specimens in ethanol) 1.7 mm (n=2); wing width 0.7 mm(n=2).
Head—dichoptic, dark red eyes, frons higher than wide; nudiocular triangular well developed (Fig. 8).Frons, clypeus and occiput black, with silvery grey pruinosity; frons, clypeus and occiput with semi-recumbent, black hairs. Cibarium with four rows of well sclerotized pointed teeth in central area, withoutcentral elevation or pronounced central area (Fig. 22).
Thorax—thoracic pattern and abdomen not described, as the specimens were pharate and have beenrecovered from alcohol. Costa with spines interspersed with setae. Subcosta with hairs. Radius with setae andspines, including setae in basal section. Leg proportions and coloration as in Figs. 88–90. Foreleg with coxa,trochanter femur and basal three fourths of tibia pale, distal quarter of tibia and all tarsal segments dark.Middle leg with coxa, and trochanter dark; basal half of tibia, basal four fifths of first tarsal segment, basalhalf of second tarsal segment pale, distal half of tibia, distal fifth of first tarsal segment, distal half of secondtarsal segment and remainder of tarsus dark. Hind leg with coxa, distal third of femur, distal half of tibia,distal quarter of first tarsal segment and remainder of tarsus dark; trochanter and basal two thirds of femurpale; basal half of tibia and basal three quarters of first tarsal segment pale. Tarsal claw of hind leg curvedwith basal tooth.
Abdomen—eighth sternite with 22–26 long setae irregularly distributed on posterior margin.Gonapophyses subtriangular with inner posterior corner protruding, with microtrichiae only on basal part,internal borders sclerotized (Fig. 114). Cerci rounded, covered with brown setae, paraproct subrectangular;cercus almost twice depth of cercus (Fig. 127). Genital fork sclerotized with ends of lateral arms oblique totransverse axis of abdomen, anterior processes of lateral arms triangular and well developed, posteriorprocesses partially developed (Fig. 141). Spermatheca subspherical, spermathecal ducts and area ofattachment unpigmented.
Male. Unknown.
Pupa. Cocoon length dorsally 1.7–2.4 mm (mean=2.1; n = 9); ventrally 2.6–3.1 mm (mean=2.8; n=9); gilllength 1.3–1.8 mm (mean=1.6, n=8) [Other measurements and averages not given in the original description.]
Cocoon—slipper shaped, light brown, composed of thick, spaced woven threads, without anteriorprojection.
Gill—pale brown, with four forwardly directed filaments, arising individually from cephalothorax; filaments of equal length, broad basally diminishing in girth progressively towards rounded apices; surface of gill filaments covered with small rounded spicules [as tubercles] (Fig. 216).
Head—frontoclypeus with rounded platelets and 1+1 hair-like, long, bifid facial trichomes and 2+2 bifid frontal trichomes as long as facial trichomes.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 35NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
Thorax—5+5 hair-like anterodorsal trichomes with 3–5 branches, 1+1 supralateral and 3+3 lateral simple hair-like trichomes. Platelets moderately abundant, rounded with diameter smaller than point of insertion of thoracic trichomes.
Abdomen—tergite I with 1+1 hair-like setae sublaterally; tergite II with 5+5 stout setae; tergites III and IV with 4+4 anteriorly directed hooks on posterior margin, 1+1 stout setae between outermost hooks, 2+2 stout setae sublaterally; tergite V with 3+3 submedian hair-like setae, tergites VI–VIII with 2+2 submedian, hair-like setae and 1+1 groups of comb teeth. Tergite IX with 1+1 short, slightly curved terminal spurs. Abdominal sternite III with 2+2 simple, lateral setae on anterior margin, and with spine combs in centre of anterior margin. Sternites IV–VIII with 1+1 groups of spine combs in the median part of the anterior margin. Sternite IV with 1+1 submedian simple or bifid small hooks and 2+2 simple, submedian setae on anterior margin amongst groups of spines; sternite V with 2+2 bifid to five branched hooks and 2+2 submedian setae lateral to them; sternites VI–VII with 2+2 simple to trifid submedian hooks and 1+1 simple setae between them.
Taxonomic discussionSimulium maranguapense was described by Pessoa et al. (2005) based on several pharate females, larvae
and pupae collected from a locality in Ceará State, Brazil. The holotype is housed in INPA, but was notavailable for study.
Simulium maranguapense is a member of the subgenus Inaequalium following the combination ofcharacters used by Coscarón & Wygodzinsky (1984), Coscarón (1991) and Strieder & Py-Daniel (1999,2002). The characters that justified its inclusion in this genus given by Pessoa et al. (2005) are wings withsetae on the basal section of R; basal portion of female cibarium with prominent teeth, pupa with thoracictrichomes with three or more branches combined with inflated gill filaments. The females of S.maranguapense are very difficult to distinguish without examination of the pupal gill configuration. Thisspecies is more readily distinguished by the pupal gill filaments with four terminal filaments, which are thickin the proximal part of the gill narrowing towards the apex, and all filaments rising at the same point basally(Fig. 216). Other species with a four-filamented gill in the subgenus Inaequalium are S. rappae [atypicalform] and S. margaritatum, but these can be distinguished by the structure of the gill filaments (Figs. 229–234, 217).
Pessoa et al. (2005) did not include S. maranguapense in any of the species groups within Inaequalium.However, the morphology of the female cercus and paraproct (Fig. 127) indicates that it belongs to theinaequale species group.
Distribution Simulium maranguapense has only been collected in the state of Ceará, Brazil (Pessoa et al., 2005).
Biology and medical importance Pessoa et al. (2005) collected larvae and pupae of S. maranguapense on rocks and fallen leaves, in shaded
parts of the river in areas of light riffles. The stream at the type locality is small during the dry season (width =0.5m, depth = 5 cm) with a rocky bed. The authors stated that this species was associated with larvae andpupae of S. rubrithorax Lutz, S. subnigrum Lutz, and a species similar to S. kabanayense Ramírez-Pérez &Vulcano, and larvae of Trichoptera, Plecoptera and Diptera (Psychodidae and Chironomidae). Nothing isknown about the feeding habits of females in Brazil.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.36 · Zootaxa 1649 © 2007 Magnolia Press
Simulium (Inaequalium) margaritatum Pepinelli, Hamada & Luz(Figs. 9, 23–25, 45–48, 91, 92, 115, 128, 142, 160, 161, 176, 189, 200, 217)
This is a recently described species from Brazil, which is known from streams located in four differentmunicipalities in the central region of the state of Bahía, Brazil. Our description is based on the originaldescription of this species, and one reared female and two pharate males collected in the state of Goiás.
Simulium margaritatum Pepinelli et al. 2006: 47. HOLOTYPE % (reared, pupal exuviae in vial with glycerine). BRA-ZIL, Bahía state, Itait Municipality, Rumo village, Chapadinha, small tributary of the Urubu River (#25), 13º11’S41º10’W; 29.vii.2005, (Hamada, N. & Pepinelli, M.) (INPA).
Female. General body color dark brown to black. Body length 1.8–2.2 mm (n = 2); thorax lateral length0.8 mm (n = 2); thorax dorsal length 0.8–0.9 mm (n = 2). Wing length 2.0 mm (n = 2); wing width 0.6–0.7mm (n = 2).
Head—frons, clypeus and occiput black, with silver pruinosity; nudiocular area well developed (Fig. 9).Antenna with silver pubescence; scape and pedicel light brown, remaining segments dark brown. Mouthpartsdark brown. Cibarium with well-developed, sclerotized cornuae, several rows of teeth between cornuae andmedian area protruding (Figs. 23–25) [in five specimens examined by Pepinelli et al. (2006), one did not havethis central elevation].
Thorax—scutum black covered with recumbent, whitish hairs. [Pepinelli et al. (2006) described the thoraxas black, without a pattern see Figs. 45–46 in this paper]. However, a thoracic pattern can be seen in a singlespecimen we have collected from Goiás State. In this specimen the scutal pattern is variable depending onlight incidence. With anterior illumination scutum greyish pruinose with 1+1 curved, submedian black vittaeand 1+1 sublateral vittae in central two thirds of scutum; humeri pale brown, with faint pruinosity (Fig. 47).With posterior illumination, thoracic pattern similar to that with anterior illumination except indistinct (Fig.48). Scutellum dark brown, with long dark setae and few golden setae. Postnotum with silver pruinosity.Costa of wing with spines and setae. Subcosta with sparse distribution of setae. Radius with spines and setae,basal section of radius with setae. Leg coloration and proportions as in male, except hind leg with basitarsusproximal two thirds whitish and distal third dark brown (see Pepinelli et al., 2006) [The latter authors did notdescribe the female leg coloration but stated that it was similar to the male. The coloration pattern theypresented is as follows: Front leg with coxa, trochanter, femur and tibia brown; tarsal segments I–IV darkbrown. Mid and hind legs with coxae, trochanters, femora, two thirds of tibiae, apical third of basitarsus, andtarsal segments II–IV dark brown; basal third of femora and remainder of basitarsus whitish]. In the femalethat we have examined, which only has two legs, the mid leg is largely pale with the apical third of the tibia,apex of basitarsus and tarsomeres mid brown. The hind leg is pale with part of the coxa, apical fourth offemur, apical half of tibia, apical tip of basitarsus, apical half of tarsal segment II and remainder of tarsalsegments mid brown (Figs. 91, 92). Claw curved with basal tooth.
Abdomen—tergite II with silver pruinosity on anterolateral margins; tergites II–IX shiny. Basal fringe oflong, golden hairs. Sternites greyish brown; genitalia dark brown. Gonapophyses subtriangular with innerposterior corner protruding, with microtrichiae only on basal part, internal borders sclerotized (Fig. 115).Cerci rounded, covered with brown setae, paraproct subrectangular; cercus almost one and a half times aslong as cercus (Fig. 128). Genital fork sclerotized with ends of lateral arms oblique to transverse axis ofabdomen, anterior processes of lateral arms triangular and well developed, posterior processes partiallydeveloped (Fig. 142). Spermatheca globular, with internal spicules arranged in groups; spermathecal ductunpigmented.
Male. General body color dark brown. Body length 2.2 mm (n=1); thorax lateral length 0.83 mm (n=1);thorax dorsal length 0.78 mm (n=1). Wing length 1.8 mm (n=1); wing width 1.3 mm (n=1).
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 37NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
Head—holoptic, with dark red eyes; remainder of head coloration as in female. Thorax—scutum black covered with golden hairs. Scutal pattern varies with light incidence: with anterior
light source, scutum black, with 1+1 anterolateral rounded, well-defined, silver areas (Fig. 160). Withposterior light source, scutum black (Fig. 161). Lateral and posterior margins of scutum pruinose. Scutellumdark brown, with long, black hairs and short, sparse golden hairs. Postnotum with silver pruinosity. Pleuradark brown, with silver pruinosity. Costa of wing with spines and setae. Subcosta bare. Radius with spines,basal section of Radius bare. Claw without basal tooth. Femora and tibiae of middle and hind legs with scale-like setae.
Abdomen—tergites black; tergite II black, with silver pruinosity; tergites IV–VII with dorsolateral marginsilver pruinose; basal fringe of long, thin, black and short golden hairs. Abdominal sternites membranous.Sternites III–VIII with silver pruinosity on median region. Genitalia dark brown; gonocoxite and gonostylecovered with small hairs and long setae; gonocoxite subrectangular; gonostyle same length as gonocoxite,subtrapezoidal with slight apical protrusion and single, stout, subapical spine (Fig. 176). Ventral platesclerotized and crescent-shaped with rounded apex and basal arms partly developed, and main body coveredwith long hairs (Fig. 189). Median sclerite pyriform with apical incision (Fig. 200). Paramere with developedand sclerotized basal process, and numerous stout spines (Fig. 200).
Pupa. Cocoon length dorsally 2.4–2.7 mm (mean=2.5 mm, s.d.= 0.13, n=5), ventrally 3.0–3.3 mm (mean= 3.1 mm, s.d.= 0.15, n=5); pupa lateral length 2.1–2.6 mm (mean=2.3 mm; s.d.=0.22, n=5); gill length 3.2–3.7 mm (mean=3.4 mm, s.d.= 0.22, n=5).
Cocoon—slipper shaped, light brown composed of thick coalesced fibers with short dorsal projection.Gills—four, forwardly directed filaments; main trunk short, giving rise basally to a dorsal and ventral
primary branches that bifurcate basally (Fig. 217). All gill filaments approximately same length, with roundedends.
Head—frontoclypeus with 2 + 2 long, bifid or trifid frontal and 1 + 1 bifid dorsal trichomes; tuberclesrounded and densely distributed.
Thorax—five pairs of long, 3–8 branched trichomes near margin of dorsal cleft; tubercles rounded,sometimes pointed, densely distributed over entire surface; on anterior region the tubercles are larger than onposterior region.
Abdomen—tergite I with 1+1 filiform setae and 2+2 submedian, short, and strong setae.; tergite II with 3+3submedian simple or bifid trichomes and 3+3 short setae on lateral region near external trichome; tergites III–IV with 4+4 submedian simple or bifid hooks; tergites V–IX with spine combs on anterior margin; tergites II–IX with 1–5 submedian simple trichomes and 1–3 simple trichomes on lateral margins; tergite IX weaklysclerotized. Sternites III–IV with spine combs on anterior margin; sternite V with 2+2 close, bifid hooks in arow and 2+2 simple trichomes on lateral margin; sternites VI–VII with 2+2 simple or bifid hooks and 2+2simple, small trichomes near these hooks; sternite VIII without hooks; sternite IX sclerotized. Abdominalsternites III–VIII with spine combs on anterior margin.
Taxonomic discussionSimulium margaritatum has been recently described by Pepinelli et al. (2006) from several males, females,
larvae and pupae collected from the municipalites Itait, Piatã, Palmeiras and Rios de Contas, Bahía State,Brazil. The male holotype is deposited in INPA, but we have been unable to examine it. Nonetheless, we havestudied the original description and illustrations of this species and compared it with a link-reared female andmale, and one pharate male identified as S. margaritatum housed in the BMNH and IOC collections (seeMaterial Examined). We have also compared S. margaritatum with a large series of link-reared specimensof other Inaequalium species deposited in the BMNH, IOC, MLP and INPA collections.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.38 · Zootaxa 1649 © 2007 Magnolia Press
Pepinelli et al. (2006) described the adult thoracic pattern of S. margaritatum from specimens recoveredfrom alcohol. In the description of the female (see their Figs. 1, 2, p. 49) and also in the taxonomic discussionthe authors stated that the female of this species has an unpatterned thorax, which distinguishes it from mostknown females of the inaequale species group. In this respect, they compared the female scutum of S.margaritatum with that of S. petropoliense, and were able to separate both species based on the thoracic hairdensity [in S. petropoliense the thorax is more densely covered with hairs than in S. margaritatum]. We haveexamined a pinned reared female and two pharate males from the State of Goiás (Material Examined) thatagree with the general morphology, number and configuration of the pupal gill filaments of S. margaritatum(Fig. 217). However, the scutum of the female shows a clear a pattern that is variable depending on lightincidence (see morphological description of the female; Figs. 45, 46).
By comparison largely with digital images in the Simuliidae archive at BMNH and identified species in theBMNH, IOC, MLP and INPA collections, the female thoracic pattern of S. margaritatum is relatively similarto that of S. rappae Py-Daniel & Coscarón, but they can be distinguished by a different configuration of thepupal gill filaments (see Taxonomic discussion under S. rappae and Py-Daniel& Coscarón, 1982). The malecannot be distinguished from other Inaequalium species, recourse having to be made to the pupal exuviaewhere the configuration of the pupal gill filaments separates this species from others in the subgenus.
The most reliable character to identify S. margaritatum is the configuration and number of the pupal gillfilaments. In S. margaritatum (Fig. 217) the pupa has four, elongated thin gill filaments, which readilyseparates it from most species in Inaequalium. Other species with four-filamented pupae are S.maranguapense, but this species can be separated by the gill filaments that are swollen basally and distinctlynarrow towards the apex (Fig. 216). Simulium rappae usually has a six-filamented pupa (Figs. 219–228), butwe have examined specimens with four-filamented gills in populations from Goiás and the Federal District ofBrazil (Figs. 229– 234). In the latter case, S. rappae can be readily separated from S. margaritatum becausethe gill filaments of the former species are distinctly swollen over their entire length. Some specimens of S.travassosi D’Andretta & D’Andretta have four-filamented gills (BMNH images archives), but in thesespecimens all of the filaments are very stout. Additionally, the configuration of the pupal gill in S. travassosiis very characteristic in usually having three primary branches that divide at the base of the gill (Figs. 236–239; BMNH image archive; D’Andretta & D’Andretta, 1947).
Pepinelli et al. (2006) observed variation in the female cibarium of S. margaritatum. One of the fivespecimens they examined did not have the central region of the cibarium protuberant, which is typical of mostspecies of Inaequalium (Fig. 25). However, the authors stated that this could be the result of someabnormality, but advocated the necessity to examine more specimens to verify their finding.
Pepinelli et al. (2006) suggested the placement of S. margaritatum in the inaequale species groupfollowing the combination of characters given by Coscarón (1987): males with the gonostylus subtrapezoidal,pupae with filiform gill filaments, larvae with the postgenal cleft narrow and subtriangular, and postgenalbridge 0.7 times as long as the hypostomium. However, they did not assign this species to any species groupbecause the males have a ventral plate which is similar to species in the botulibranchium species group. Wehave studied the ventral plate of S. margaritatum and compared it with digital images of other Inaequaliumspecies in the Simuliidae digital images archive in the BMNH. We found that their figure of the ventral plateis morphologically more similar to species in the inaequale than to the botulibranchium groups and,consequently, we place S. margaritatum in the inaequale species group.
Distribution Simulium margaritatum has only been collected in the central region of Bahia state, Brazil, in the Chapada
Diamantina National Park and surrounding area. We now record its distribution to include Goiás state(Pepinelli et al., 2006; Material Examined).
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 39NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
Biology and medical importance The immature stages of S. margaritatum were collected by Pepinelli et al. (2006) in several small, rocky-
bottomed streams in open areas exposed to sunlight, with pupae and larvae attached to trailing vegetation,fallen leaves, and rocks. The streams varied in width from 0.2 to 1.5 m and were located between 770 to 1683m in altitude. The pupae that we collected in the State of Goiás were found in a small (2 m wide, 20 cm deep),medium flowing stream with a rocky bottom. Nothing is known of the feeding habits of female S.margaritatum.
Simulium (Inaequalium) mariavulcanoae Coscarón & Wygodzinsky (Figs. 10, 26, 49–52, 93, 94, 116, 129, 143, 177, 190, 201, 218)
This is a poorly known species having been collected only from its type locality in southeast Brazil. The fol-lowing description is based on the female holotype, identified material by Coscarón deposited in the MLP, andcomparison with illustrations and descriptions in Coscarón & Wygodzinsky (1984), Coscarón (1991) andStrieder & Py-Daniel (1999, 2000).
Simulium mariavulcanoae Coscarón & Wygodzinsky, 1984: 96. HOLOTYPE &, BRAZIL: São Paulo State,E.B.Boracéia, Salesópolis; 13.x.1947 (M.A.V. d’Andretta) (MZSP, no. 115–212) [Examined.]
Female. General morphology and coloration as in S. diversibranchium. General body color dark brown.
Body length (specimens pinned) 2.1–2.4 mm (n=2); wing length 2.0–2.2 mm (n=2); wing width 1.2 mm(n=2).
Head—dichoptic, with dark red eyes and nudiocular area developed (Fig. 10). Frons, clypeus and occiputblack, with silvery grey pruinosity; clypeus covered with pale, semi-recumbent setae and frons with long,erect, black hairs. Mouthparts dark brown. Antennae with scape and pedicel brownish, remainder of flagellarsegments yellowish. Cibarium with well-developed, sclerotized cornuae and small, irregular teeth in medianarea, which protrudes (Fig. 26).
Thorax—scutum dark brown, with faint white pruinosity and evenly arranged groups of dense, recumbent,golden setae that obscure scutal pattern. Scutal pattern varying slightly with illumination. With anteriorillumination, thorax with very feint 1+1 submedian silvery stripes in anterior third and slight silverypruinosity on median part of anterior margin; humeri greyish; lateral and posterior margins black, with weaksilver tinge (Figs. 49, 51). With posterior illumination, thorax greyish with silvery pruinosity on medianmargin; posterior margin black (Figs. 50, 52). Scutellum dark brown. Postnotum dark brown. Pleura black,with silver pruinosity. Costa of wing with sparse distribution of spines and setae. Subcosta with row of setae.Radius with numerous setae intermixed with distinct spines, basal section of Radius with single row of setae.Basal tuft of long, dark setae. Leg coloration and proportions as in Figs. 93, 94. This is from the holotype andconsists of an incomplete mid leg and a hind leg. Mid and hind leg coloration is similar in that the distal halfof the tibiae, apical tip of the basitarsi with the remaining parts whitish; the second tarsal segments are brown,except in the mid leg where the basal half is whitish; the third and fourth tarsal segments are mid brown; Thetrochanter and femur of the hind leg are mid brown. For further details on coloration and leg proportions thereader is referred to the original description (Coscarón & Wygodzinsky, 1984). Claws curved with small basaltooth. Halteres cream with brown base.
Abdomen—tergites I–IV dark velvet brown; tergites VI–IX shiny, pale brown mesally and black alongposterior and ventrolateral margins. Sternites and genitalia dark brown to black. Eighth sternite weaklysclerotized with long, irregularly distributed setae on posterior margin; gonapophyses nearly same length assternite at midpoint, subtriangular with inner posterior corner protruding, with microtrichiae only in basal
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.40 · Zootaxa 1649 © 2007 Magnolia Press
part, internal borders sclerotized (Fig. 116). Cerci rounded, covered with brown setae; paraproctsubrectangular, almost twice as deep as cercus (Fig. 129). Genital fork sclerotized with ends of lateral armsparallel to transverse axis of abdomen, anterior processes of lateral arms triangular and well developed,posterior processes partially developed (Fig. 143). Spermatheca suboval, apparently without externalsculpturing or spicules on internal surface; area of insertion of spermathecal membranous.
Male. General morphology and coloration as in S. diversibranchium (see Coscarón & Wygodzinsky,1984). General body color black. [Measurements taken from the original description, pinned specimen] winglength 2.3–2.6mm [no other measurements given].
Head—holoptic, with dark red eyes. Remainder of head coloration as in female.Thorax—scutal pattern (Figs. 156, 157) and general morphology as in S. diversibranchium, except minor
difference in paraprocts.Abdomen—tergites black, basal fringe of long, brown hairs. Silver pruinose ornamentation on anterolateral
margin of tergite II and ventrolateral margins of tergites V–VII. Genitalia black; sternites whitish pruinose;tergal plates developed. Gonocoxite subrectangular; gonostyle subtrapezoidal with slight apical protrusionand single, stout, subapical spine (Fig. 177), and almost same length as gonocoxite. Ventral plate sclerotizedand crescent-shaped with pointed apex and basal arms well developed, and main body covered with long hairs(Fig. 190). Median sclerite pyriform with apical incision (Fig. 201). Paramere with developed and sclerotizedbasal process, and numerous well-developed spines (Fig. 201).
Pupa. Gill length 3.2–3.3 mm (n=3) [Measurements given in original description: gill length 3.2–3.6mm.]. [No other measurements given in original description.]
Cocoon—slipper shaped, light to dark brown composed of thin network of coalescent fibers and reinforcedrim to anterior aperture.
Gill—light brown, with six forwardly directed filaments arranged in vertical plane. Gill configurationvariable as in S. diversibranchium with main trunk short, giving rise basally to two sets of primary branches:one dorsal and stout and one ventral that is thinner; dorsal primary branch almost immediately divides intotwo secondary branches, one dorsal that bifurcates towards gill apex and one ventral that bifurcates apically.Ventral primary branch bifurcates almost immediately (Fig. 218). Variation in the pattern occurs with ventralsecondary branch bifurcating more apically than dorsal secondary branch. All filaments weakly roundeddistally, with small spicules on surface, edges crenate; all filaments of same length.
Head—frontoclypeus with 2+2 long, bifid frontal and 1+1 long, multiramous facial trichomes;frontoclypeus with group of platelets mesally, 1+1 groups of 7 platelets dorsolaterally and one group ofplatelets in pairs laterally in frontal region; tubercles rounded and densely distributed over entire surface.
Thorax—with 4+4 quadrifid to multiramous trichomes near margin of dorsal cleft, and simple, longtrichomes on ventral margin of thorax; tubercles rounded and densely distributed over entire surface.
Abdomen—tergite I with 1+1 long, simple trichomes on anterolateral margin; tergite II with 3+3submedian spiniform setae in row, 3+3 simple, spiniform setae vertical to submedian spiniform setae, and onesmall, sublateral trichome; tergite III with 4+4 submedian, simple hooks in row along posterior border, and1+1 small, simple trichomes anterior to outermost hooks; tergite IV with 4+4 submedian, simple hooks in rowalong posterior margin, 1+1 simple, small setae anterior to most lateral hooks and one small, simple trichomeon lateral margin; tergite V without setae or spine combs; tergites VI–VIII with distinct spine combs somewell developed and resembling teeth on anterior margin; tergite IX with spine combs resembling teeth and1+1 small apical spines. Abdominal sternite III with 3+3 submedian, spiniform setae; sternite IV with 2+2close, median bifid hooks in row along posterior margin; sternite V with 2+2 close, submedian hooks onposterior margin, the outermost hooks single and innermost hook bifid or trifid; sternite VI with 4+4
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 41NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
submedian hooks in row along posterior margin, the outermost simple and innermost trifid; sternites VII–IXwithout setae. Spine combs on anterior margins of sternites III–IX.
Taxonomic discussion Simulium mariavulcanoae was described by Coscarón and Wygodzinsky (1984) based on several reared
females and males collected in the state of São Paulo, Brazil. We have examined the female holotype, whichis deposited in the MZUSP and taken digital images prior to dissecting the cibarium and genitalia that are nowmounted on a slide (see Material Examined). The head and the abdomen are in a small vial with glycerineattached to the pin. The specimen was slightly damaged during dissection and the legs and wings were brokenoff the thorax and glued to a card point. The specimen bears a label in Coscarón’s handwriting “Simuliummariavulcanoae det. Coscarón 1979”. Coscarón & Wygodzinsky (1984) and Strieder & Py-Daniel (2000)stated that paratype material of this species is deposited in the MZUSP, but we were unable to find them in theMZSUP holdings.
The adults of S. mariavulcanoae cannot be distinguished from other species of Inaequalium, especially S.diversibranchium, without examination of the pupal gill filaments. Both species have a six-filamented pupa,but S. mariavulcanoae may be separated by the differences in branching heights from the base filaments [seealso Taxonomic discussion under S. diversibranchium, and Coscarón & Wygodzinsky (1984), Strieder & Py-Daniel (1999)].
The morphological description of the adults and pupal stages of S. mariavulcanoae may be found inCoscarón (1991), Coscarón & Coscarón-Arias (2007) and Strieder & Py-Daniel (1999; 2000). The larva stillremains unknown.
Simulium mariavulcanoae is assigned to the Inaequalium species group based on the combination ofcharacters given in Coscarón (1987, 1991). Coscarón & Wygodzinsky (1984) and Strieder & Py-Daniel(2002) in their phylogenetic study of Inaequalium placed this species in the inaequale-group, closely relatedto S. diversibranchium.
Distribution Simulium mariavulcanoae has only been found at its type locality in the state of São Paulo (Coscarón,
1991; Coscarón & Coscarón-Arias, 2007; Crosskey & Howard, 1997; 2004; Material Examined).
Biology and medical importance Nothing is known about the biology of the immature stages or the female feeding habits of S.
mariavulcanoae in the Neotropical region (Coscarón, 1991; Coscarón & Coscarón-Arias, 2007; Strieder &Py-Daniel, 1999).
Simulium (Inaequalium) rappae Py-Daniel& Coscarón(Figs. 12, 28, 53–62, 98–100, 117, 131, 145, 162, 163, 178, 191, 202, 219–234)
This is a relatively common species in southern Brazil, which shows great variation on the female thoracicpattern and number and configuration of the pupal gill filaments. The description here provided is based onlarge series of link-reared specimens and the examination of type material.
Simulium (Inaequalium) rappae Py-Daniel& Coscarón, 1982: 155. HOLOTYPE & & pupal exuviae, BRAZIL: MinasGerais State, Bocaina de Minas, Estrada entre Repouso da Gávea e a Pedra da Gávea, Córrego do Concreto, 22º19’S44º34’W; 12.i.1980, (V. Py-Daniel & J. Carneiro de Vasconcelos Neto) (INPA, no. 5515-13, not 5513-13 as recordedin paper – pers. comm. Dr V. Py-Daniel to Dr R.W. Crosskey on 11.1983).
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.42 · Zootaxa 1649 © 2007 Magnolia Press
Female. General body color grayish black. Body length (dry specimens) 1.7–2.0 mm (mean=1.85 mm,s.d.=0.09, n=10), (specimens preserved in alcohol) 2.1–2.5 mm (mean=2.3 mm, s.d.=0.17, n=4). Wing length1.4–2.0 mm (mean= 1.7 mm, s.d.=0.14, n=11), wing width 0.7–1.1 mm (mean=0.87mm, s.d.= 0.11, n=11).
Head—dichoptic, with dark red eyes and nudiocular area well developed (Fig. 12). Frons, clypeus andocciput black, with silvery grey pruinosity; frons and clypeus with pale, semi-recumbent hairs slightly longeron occiput. Mouthparts light to dark brown. Antennae with scape, pedicel and most of first flagellomereyellowish brown, remainder of flagellar segments black. Cibarium with well-developed, sclerotized cornuaeand group of about 40–50 sharp teeth of varying size extending from base of cornuae to central area ofcibarium, which is protuberant (Fig. 28).
Thorax—scutum color and pattern variable with locality. In specimens from south-eastern Brazil (Rio deJaneiro & Minas Gerais States—see Material Examined NHM, IOC material) brownish to greyish black, withfaint, silver pruinosity forming an indistinct pattern and densely distributed, recumbent, brass-colored hairs(Figs. 53, 54). Scutal pattern varies with direction of illumination. In specimens from Central Brazil (Figs. 55,57, 59, 61) with anterior illumination, scutum whitish grey pruinose with faint median, 1+1 submedian and1+1 lateral black vittae extending from near anterior margin to posterior margin; lateral and posterior marginsand humeri grey pruinose. With posterior illumination, thorax greyish black, with pyriform silver pruinosearea extending from anterior margin almost to posterior margin; humeri silver pruinose (Figs. 56, 58, 60, 62).Scutellum dark brown, with recumbent brass-colored hairs and several dark brown long bristles mainly onposterior margin. Postnotum dark brown, with grey pruinosity. Pleura brown, with silver pruinosity andpleural membrane with recumbent hairs, brass-colored or black depending on light direction. Costa of wingwith sparse distribution of spines and setae. Subcosta with 6–9 setae in basal half. Radius with single row ofsetae intermixed with several distinct spines, basal section of radius with single line of setae. Basal tuft ofsparse, light brown setae. Leg coloration and proportions as in Figs. 98–100. Foreleg coxa, trochanter, femur,and basal two thirds of tibia yellowish and outer face of tibia whitish with distal articulation dark brown; tarsidark brown to black. Mid leg coxa dark brown, trochanter, femur, basal half of tibia and basal three fourths ofbasitarsus light brown, distal half of tibia, distal fourth of basitarsus and remainder of tarsal segments darkbrown. Hind leg coxa dark brown, trochanter light brown, basal half of femur and tibia and basal two thirds ofbasitarsus whitish, remainder of femur, tibia, basitarsi and remainder of tarsal segments dark brown. Scalelike setae on hind tibia. Claws curved with small basal tooth. Halteres lemon yellow, with brown base.
Abdomen—tergite I light brown, with basal fringe of brass-colored hairs; II–IV velvet black, VI–IX shinyblack. Tergal plates undeveloped in the few specimens examined. Sternites and genitalia black. Eighth sterniteweakly sclerotized with about 50 setae of varying size on posterior margin except for central area;gonapophyses nearly same length as sternite at midpoint, subtriangular with inner posterior corner protruding,with microtrichiae only in basal part, internal borders sclerotized (Fig. 117). Cerci rounded, covered withbrown setae, paraproct subrectangular; paraproct slightly produced below lower margin of cercus (Fig. 131).Genital fork sclerotized with ends of lateral arms parallel to transverse axis of abdomen, anterior processes oflateral arms triangular and well developed, posterior processes partially developed (Fig. 145).
Male. General body color dark brown to black. Body length (pinned specimens) 1.6 mm (n=2), (specimenspreserved in alcohol) 2.0–3.1 mm (mean=2.5 mm, s.d.=0.33, n=7). Wing length 1.4–1.6 mm (mean=1.4 mm,s.d.=0.36, n=6), wing width 0.5–0.9 mm (mean=0.7 mm, s.d.=0.2, n=6).
Head—holoptic, with dark red eyes. Remainder of head coloration as in female.Thorax—scutum dark brown to black, with golden, recumbent hairs. The single specimen from Minas
Gerais State (Material Examined) was dark brown, whereas from Goiás State both dark brown and blackspecimens were recorded. Scutal pattern varies slightly with light incidence. With light source anterior tospecimen scutum dark brown or black, with 1+1 silver, submedian areas on anterior margin and extending forone third length of scutum (Fig. 162); in some specimens these areas are indistinct. With posterior light source
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 43NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
thorax black (Fig. 163). Humeri, lateral and posterior margins of scutum silver pruinose. Scutellum black,with recumbent golden and long, erect dark brown setae. Postnotum black, with silvery grey pruinosity.Pleura black, with grey pruinosity. Halteres whitish yellow, with light brown base. Wing setation as in femaleexcept Sc bare. Leg coloration as in female except entire femur of hind leg dark brown and only slightlyspatulate hairs on hind femur and tibia.
Abdomen—tergites black, basal fringe of long, brown hairs. Silver pruinose ornamentation as follows:whole of tergite II except for median part, most of tergites VI, VII except median part and 1+1 lateral areas ontergite VIII. Genitalia black; sternites silvery grey pruinose; tergal plates well developed; sternal platesundeveloped. Gonocoxite rectangular; gonostyle between subtrapezoidal with apical protrusion and pyriformterminating in single, stout spine and two thirds length of gonocoxite; gonocoxite and gonostyle covered withlong setae (Fig. 178). In some specimens small blunt or pointed spine on lower edge of gonostyle at mid pointor slightly distal to this; both forms seen on same specimen. Ventral plate sclerotized and crescent-shapedwith rounded apex and basal arms well developed, and main body covered with long hairs (Fig. 191). Mediansclerite pyriform with no apical incision (Fig. 202). Paramere with developed, lightly sclerotized basal processand several long and short spines along distal half (Fig. 202).
Pupa. Cocoon length dorsally 2.2–3.0 mm (mean=2.6 mm, s.d.=0.21, n=21); ventrally 2.2–3.7 mm
(mean=3.0 mm, s.d.=0.34, n=19); pupa length 1.75–3.7 mm (mean=2.8 mm, s.d.=0.49, n=19); gill length 1.7–4.3 mm (mean=2.9 mm, s.d.=0.73, n=31).
Cocoon—slipper shaped, light to dark brown composed of a thin network of fibers, with slightly reinforcedrim to anterior aperture.
Gill—light brown commonly with six forwardly directed broad filaments arranged in vertical plane. Gillconfiguration very variable with filaments branching at different heights even within same specimen. Generalpattern as follows: main trunk short, giving rise to two sets of primary branches: the dorsal bifurcating almostimmediately into a dorsal secondary filament that bifurcates in distal half to four fifths of gill and a ventralsecondary filament that bifurcates in basal third to distal two thirds of gill; the ventral bifurcating to formsecondary filaments in basal fifth to three tenths of gill (Figs. 219–222). Considerable variation occurs indistance from gill base of bifurcations and in one specimen branching of secondary filaments of the dorsalprimary branch almost apical (Figs. 223–228). An atypical form with four gill filaments also occurs (Figs.230–234) as well as a form with five filaments (Fig. 229). Variation in filament girth occurs ranging fromstout to swollen, with swelling often more evident distally (Figs. 219–234). Filaments pointed distally, withsmall spicules on surface, edges crenate; ventral primary branch filaments of same length, dorsal primarybranch filaments varying in length between different specimens.
Head—frontoclypeus with 2+2 trifid or 4-branched frontal and 1+1 4-branched facial trichomes, alloriginating from a thin, curved base; frontoclypeus with rounded tubercles densely distributed on frons butless so on ocular and antennal regions.
Thorax—5+5 large, simple to seven-branched trichomes near margin of dorsal cleft; tubercles rounded anddensely distributed on dorsal half of thorax.
Abdomen—tergite I with 1+1 long, simple setae on anterolateral margin; tergite II with 4+4 submediansimple spines along posterior margin and 1+1 simple, setae anterior to outermost spine; tergites III and IVwith 4+4 submedian, simple spines along posterior border, 1+1 simple, short setae anterior to most lateralspines and 1+1 short setae on middle of lateral margin; tergites V–VI with 4+4 simple setae on posteriormargin; tergite IX with strong sclerotized terminal spines; spine comb distribution as follows: band onanterior margins of tergites VII–IX (being more developed on last two segments), except in median area.Sternite III with 2+2 simple, small trichomes sublaterally and simple hook on mid-lateral margin, area ofspine combs on median, anterior margin; sternite IV with 3+3 submedian, small, simple setae, and row ofspine combs on posterior margin; sternite V with 2+2 close, median hooks on posterior margin, of which inner
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.44 · Zootaxa 1649 © 2007 Magnolia Press
is bifid and outer bifid or trifid and two small, simple setae lateral to these, and 1+1 submedian groups ofspine combs anterior to hooks and 2+2 simple setae on postero-lateral margin; sternites VI–VII with 2+2 wellseparated hooks of which inner is trifid and outer simple on posterior margin, 1+1 simple setae between hooksand 1+1 simple setae on anterior margin sublaterally, and 1+1 submedian groups of spine combs anterior tospines; sternite IX weakly sclerotized apically with 1+1 groups of spine combs on anterior margin .
Taxonomic discussion Simulium rappae was originally fully described in all stages by Py-Daniel & Coscarón (1982) from
specimens collected in the states of Minas Gerais, São Paulo and Rio de Janeiro, Brazil. We have examinedthe female holotype and a male paratype, which are housed in INPA. The female has been glued by the leftside to a card point attached to the pin. It also has a plastic microvial with cork lid containing the pupalexuviae. It appears that the microvial contained glycerine that has now dried. However, the pupal exuviaeremains in good condition. The microvial also contains two handwritten labels stating “Holotipo” and anumber “5515-13” (see Material Examined).
The adults of S. rappae cannot be distinguished from other species in the Inaequalium-species groupwithout examination of the gill configuration of associated pupae. The pupa of S. rappae is most similar tothat of S. travassosi in having the gill filaments swollen. However, they can be reliably identified by thedifferent number of gill filaments and configuration, six in S. rappae [sometimes four or five filaments occur](Figs. 219–234) and three in S. travassosi (Figs. 236–239).
Variation in the adults’ thoracic pattern, and the number and configuration of the pupal gill filaments iscommon in S. rappae. The female thoracic pattern sometimes resembles species of Psaroniocompsa andChirostilbia by having a median, 1+1 submedian and 1+1 lateral black vittae on the silver pruinose scutum(Figs. 59–62). In specimens collected from Rio de Janeiro and Minas Gerais states the pupal gill filaments canbe more elongated with the dorsal primary branch bifurcating basally or more apically and terminating inpointed or rounded ends, and sometimes having five gill filaments (Figs. 219–234). An extreme variation hasbeen examined in specimens collected from Rio Saia Velha, Goiás State and the Federal District, in whichfour gill filaments are found, and all branches are distinctly swollen with the dorsal and ventral primarybranches bifurcating at the same level from the middle of the gill (Fig. 230–234). In some cases somefilaments are not fully developed (Fig. 223).
A morphological and cladistical revision of this subgenus (Coscarón, 1987, 1991; Coscarón & Coscarón-
Arias, 1997) divided it into the inaequale and botulibranchium groups. Simulium rappae was placed in theinaequale group based on characteristics of adult genitalia and larval and pupal morphology. However, theseauthors did not mention that the pupal gill is atypical for this group in that it is swollen (like S. travassosi, alsoin this group) (Figs. 236-239). Strieder & Py-Daniel (2000) distinguished pupae of these two species groupsonly on filament number and the presence of thin or swollen filaments, which would place both S. travassosiand S. rappae in the botulibranchium group. We assigned S. rappae to the inaequale species group based onthe short female paraprocts (Fig. 131) and subtrapezoidal gonostyle (Fig. 178).
Distribution The species has only been recorded in Brazil where it appears to be confined to highland areas in central
and southern coastal regions of the states of Goiás, Minas Gerais, Rio de Janeiro and São Paulo (Coscarón,1991; Coscarón & Coscarón-Arias, 2007; Crosskey & Howard, 1997; 2004; Strieder & Py-Daniel, 2002;Material Examined).
Biology and medical importance
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 45NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
Little is known about the biology of this uncommon Brazilian species. It breeds in clear, small, sunlitstreams on submerged vegetation and has not been recorded biting humans (A.J. Shelley & A.P.A. Luna Dias,unpublished data).
Simulium (Inaequalium) subnigrum Lutz(Figs. 13, 29, 63, 64, 101-103, 118, 132, 146, 164, 165, 179, 180, 192, 203, 235)
This species is very common in southern Brazil. It has been recently reviewed by Hernandez et al. (2007),who proposed several new synonymies and recorded a wider its distribution range in Brazil. The provenanceof the syntype material is still in doubt.
Simulium subnigrum Lutz, 1910: 239. SYNTYPES &&%% & pupae, BRAZIL: São Paulo State (Rivers Pacaembú andAnhangabahú), Rio de Janeiro State (Mendes and Petrópolis); [Without date or collector’s name.] (IOC).
Simulium diversifurcatum Lutz, 1910: 258. SYNTYPES pupae, BRAZIL: São Paulo State (Serra de Bocaina, Capivary),Rio de Janeiro State (near Petrópolis); [Without date or collector’s name.] [Synonymy by Coscarón & Wygodzinsky,1984: 90.]
Simulium subclavibranchium Lutz, 1910: 260. LECTOTYPE pupal exuviae, BRAZIL: Rondônia State, estrada de ferroMadeira-Mamoré, [Without date or collector’s name] (IOC 12.372, Bd 120, no. 236) [Examined.] [Synonymy andlectotype designation by Hernández et al., 2007: 16.]
Simulium mbarigui Coscarón & Wygodzinsky, 1973: 142. HOLOTYPE & + pupal exuviae. ARGENTINA: MissionesState, 20km N de Apóstoles, Arroyo s/ruta 1, 18.vii.1972, (Coscarón) (MLP). [Examined.] [Synonymy with S. sub-nigrum by Coscarón & Wygodzinsky, 1984: 90; synonymy with S. clavibranchium by Coscarón, 1987: synonymywith S. subnigrum by Coscarón, 1991: 162).
Simulium nogueirai D’Andretta & González B., 1964: 103. HOLOTYPE & (reared), BRAZIL: São Paulo State:Aldeinha, (E.F.S.); 7.vii.1957 (C. d’Andretta Jr. & M.V.Nogueira) (ICBUSB) [Py-Daniel & Moreira (1989) regardedthe type series S. nogueirai “not lost but misplaced”]. [Synonymy by Hernández et al., 2007: 16.]
Simulium (Simulium) beaupertuyi Ramírez-Pérez, Rassi & Ramírez, 1977: 165. HOLOTYPE &, VENEZUELA: Ter-ritório Federal de Amazonas, Departamento de Atabapo, Sierra de Parima; [Without date or collector’s name.][Holotype said to be deposited in Ramírez Pérez private collection in the original description, but Py-Daniel &Strieder (1999) recorded it in IDVC; Holotype depository unknown- see Note on Ramírez Pérez Simuliidae collec-tion.]. [Synonymy by Hernández et al., 2007: 16.]
Simulium nahimi Py-Daniel, 1984:125. HOLOTYPE & (reared, pupal exuviae in ethanol). BRAZIL: Mato Grosso State,Dardanelos, Aripuanã; 13.xi.1975, [Without collector’s name, but probably collected by Py-Daniel] (INPA, 5125- 6)[Examined.] [Synonymy by Hernández et al., 2007: 16.]
Inaequalium leopoldense Strieder & Py-Daniel, 2000:15. HOLOTYPE larva, BRAZIL: Rio Grande do Sul, SãoLeopoldo, 13.x.1983, (H.G.Konrad) (LEU) [The life stage designated as holotype was not stated in the originaldescription, but it is a larva- pers. comm. M.N.Strieder to L.M. Hernández v.2006.] [Synonymy by Hernández et al.,2007: 16.]
Female. General body color black. Body length (specimens pinned) 1.9-2.1 mm (mean=2.0 mm,s.d.=0.09, n=4); wing length 2.1-2.4 mm (mean=2.2 mm, s.d.= 0.05, n=3); wing width 0.9-1.1 mm (mean=1.0mm, s.d.=0.09, n=4).
Head—dichoptic, with dark red eyes and nudiocular area developed (Fig. 13). Frons, clypeus and occiputblack, with silvery grey pruinosity; frons, clypeus and occiput with semi-recumbent, black hairs. Mouthpartsblack. Antennae with scape and pedicel yellowish brown, remainder of flagellar segments black. Cibariumwith well-developed, sclerotized cornuae and rows of sharp teeth of varying size extending from base ofcornuae to central area of cibarium, which is protuberant (Fig. 29).
Thorax—scutum black covered with recumbent, golden hairs. Scutal pattern variable depending on lightincidence. With anterior illumination scutum black, with patches of greyish pruinosity and 1+1 submedianand 1+1 sublateral indistinct black bands extending from anterior to posterior margin of thorax; humeri,lateral and posterior margin dark brown (Fig. 63). With posterior illumination, thorax greyish pruinose withmedian rectangular black area on anterior border and 1+1 submedian, indistinct black vittae on anterior third
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.46 · Zootaxa 1649 © 2007 Magnolia Press
of scutum; humeri and lateral margins grey pruinose; lateral margins black (Fig. 64). Scutellum dark brown toblack, with recumbent, golden hairs interspersed with long, black bristles. Postnotum black, with greypruinosity. Pleura dark brown, with silver pruinosity. Costa of wing with sparse distribution of spines andsetae. Subcosta with row of setae in basal two thirds. Radius with row of setae intermixed with distinct spines,basal section of radius with row of setae apically and irregularly distributed setae basally. Basal tuft of sparse,light brown setae. Leg coloration and proportions as in Figs. 101-103. Foreleg with coxa and tibia whitish,trochanter, femur, apical tip of tibia, and tarsal segments dark brown. Mid leg with coxa, trochanter, femur,distal half of tibia, apical tip of basitarsus, apical half of second tarsal segment and remainder of tarsusmedium to dark brown; remainder of leg whitish. Hind leg with coxa, trochanter, apical third of femur, apicalhalf of tibia, apical fourth of basitarsus and tarsal segments II-IV dark brown; remainder of leg whitish. Hindclaw with basal tooth. Halteres lemon yellow, with dark brown base.
Abdomen—tergite I dark brown, pale brown mesally; tergites II, V-IX shiny pale yellowish on anteriormargins and dark brown on posterior margins; tergites III-IV velvet dark brown; tergite II silver pruinose onlateral margins. Tergal plates developed. Sternites greyish; genitalia dark brown to black. Eighth sterniteweakly sclerotized with long, irregularly distributed setae on posterior margin; gonapophyses nearly samelength as sternite at midpoint, subtriangular with inner posterior corner protruding, densely covered inmicrotrichiae, internal borders sclerotized (Fig. 118). Cercus rounded, covered with brown setae; paraproctsubrectangular, protruding below cercus by about the length of cercus (Fig. 132). Genital fork sclerotized withends of lateral arms parallel to transverse axis of abdomen, anterior processes of lateral arms triangular andwell developed, posterior processes partially developed (Fig. 146). Spermatheca suboval, without externalsculpturing and irregularly distributed spicules on internal surface; area of insertion of spermathecal ductmembranous.
Male. General body color black. Body length (specimens pinned) 2.0-2.2 mm (n=2); wing length 1.2-1.4mm (n=3), wing width 0.9-2.2 mm (n=2).
Head—holoptic, with dark red eyes. Remainder of head coloration as in female.Thorax—scutum dark brown to black covered with evenly distributed golden, recumbent hairs. Scutal
pattern varies slightly with light incidence. With light source anterior to specimen scutum black, with 1+1sublateral, greyish pruinose rounded areas in anterior half; humeri, lateral and posterior margins black (Fig.164). With light posterior, thorax dark brown; humeri, lateral and posterior margins of scutum dark brown(Fig. 165). Scutellum dark brown to black covered with recumbent, golden hairs interspersed with long, erect,dark brown setae. Postnotum dark brown, with silvery grey pruinosity. Pleura black, with grey pruinosity.Halteres whitish yellow, with light brown base. Wing setation as in female except Sc bare. Leg coloration asin female.
Abdomen—tergites dark brown; basal fringe of long, brown hairs. Silver pruinose ornamentation asfollows: whole of tergite II except for median part and ventrolateral margins of tergites IV-IX. Genitalia darkbrown; sternites whitish pruinose; tergal plates developed. Gonocoxite subrectangular; gonostyle slightlyshorter than gonocoxite, subtrapezoidal with slight apical protrusion and single, stout, subapical spine;gonocoxite and gonostyle covered with long spines (Fig. 179). In some specimens several spines may bepresent on basal margin of gonostyle (Fig. 180). Ventral plate sclerotized and crescent-shaped with pointedapex, basal arms well developed, and main body covered with long hairs (Fig. 192). Median sclerite pyriformwith incision in apical third and wide basally (Fig. 203). Paramere with developed, sclerotized basal processand several long and short spines along distal half and in median part (Fig. 203).
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 47NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
Pupa. Cocoon length dorsally 2.9-3.4 mm (mean=3.1 mm, s.d.=0.18, n=7); ventrally3.0-3.6 mm (mean=3.3 mm, s.d.=0.26, n= 7); pupal length 2.9-3.7 mm (mean= 3.3 mm, s.d.=0.31, n=7); gill length 2.3-4.6 mm(mean= 3.4 mm, s.d.=0.65, n=8).
Cocoon—slipper shaped, light to dark brown composed of thin network of fibers and reinforced rim toanterior aperture.
Gill—light to dark brown, with six forwardly directed filaments arranged in vertical plane. Gillconfiguration highly variable with main trunk short. The most common configuration (Fig. 235) is where thegill divides basally into a dorsal and ventral primary branch; dorsal primary branch divides basally into twosecondary branches each of which bifurcates at same or different levels in basal fourth of gill; ventral primarybranch bifurcates basally. Variation occurs in the level of bifurcation in the gill filaments being more basal ormore apical. All filaments of approximately same length, thin, rounded distally or mamiliform, covered withthin spicules on surface, edges weakly smooth.
Head—frontoclypeus with 2+2 trifid, frontal and 1+1 trifid, facial trichomes; frontoclypeus with group ofplatelets mesally, 1+1 groups of approximately six platelets dorsolaterally and 2-3 platelets in groups of twolaterally in frontal region; tubercles rounded and densely distributed over entire surface.
Thorax—with 5+ 5 large, trifid to quadrifid trichomes near margin of dorsal cleft and one long, simpletrichomes near base of gill ventrally; tubercles rounded and densely distributed over entire surface of thorax.
Abdomen—tergite I with 1+1 long, sublateral simple trichomes; tergite II with 3+3 submedian, simpletrichomes in row, 3+3 simple trichomes near outer trichomes, and 2+2 small, simple lateral setae; tergites IIIand IV with 4+4 submedian, simple hooks in row along posterior margin, and 3+3 small, simple setae anteriorto this row; tergite V with 3+3 submedian, simple setae in row along posterior margin; tergite V with 3+3spiniform, simple trichomes in row along posterior margin; tergite VI with 1+1 submedian and 1+1 sublateral,small simple trichomes near posterior margin and spine combs on anterior margin some of them developedand resembling teeth; tergites VII and VIII apparently without trichomes, but spine combs on anterior margin,some of them resembling teeth; tergite IX weakly sclerotized with well-developed spine combs resemblingteeth on anterior margin and 1+1 apical spines. Abdominal sternite III with 3+3 submedian, small simple and2+2 lateral, small and simple trichomes; sternite IV with 4+4 simple trichomes in row along posterior margin;tergite V with 2+2 submedian trifid hooks, 2+2 small, simple trichomes anterior to trifid hooks and 2+2 small,simple trichomes on lateral margin; sternites VI-VII with 2+2 well separated hooks on posterior margin, innerbeing bifid or trifid and outer simple, 1+1 small, simple trichomes between hooks and 1+1 small trichomes onanterior margin; sternite VIII with single, small setae on lateral margin; sternite IX weakly sclerotized. Spinecombs on anterior margin of sternite VI.
Taxonomic discussion The taxonomy of S. subnigrum has been treated comprehensively by Hernández et al. (2007), who
proposed five new synonyms and reviewed the relationship between S. subclavibranchium, S. clavibranchiumand S. subnigrum. The authors also followed the same approach used by Shelley et al. (1997, 2006) for species inthe amazonicum species group, and considered S. subnigrum as a polymorphic species. The main polymorphiccharacter is the variation in spines on the male gonostyle. The adults of S. subnigrum cannot be separated frommost of the species in the inaequale-species group by the thoracic pattern or the morphology of the adultgenitalia. It can only be recognized by the configuration of the pupal gill filaments in link-reared specimens.
Simulium subnigrum is placed within the inaequale-species group of Inaequalium based on the combinationof characters given in Coscarón (1987, 1991), Coscarón & Coscarón-Arias (2007), Coscarón & Wygodzinsky(1984), and Strieder & Py-Daniel (1999, 2000, 2002). The most salient characters are the female paraproctextending beyond the cercus by nearly the same length as the cercus, rounded or truncate apically and the pupawith thin gill filaments and its thorax with densely distributed tubercles.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.48 · Zootaxa 1649 © 2007 Magnolia Press
Distribution Based on the synonymies proposed in Hernández et al. (2007), S. subnigrum now has a wider distribution
in the Neotropical Region. In Brazil it has been recorded in the states of Bahía, Ceará, Federal District, Goiás,Minas Gerais, Mato Grosso, Mato Grosso do Sul, Paraná, Rio de Janeiro, Rio Grande do Sul, Santa Catarinaand São Paulo. Elsewhere, this species is found in Argentina (Missiones), Colombia, Paraguay, Trinidad andVenezuela (Coscarón, 1991; Coscarón & Coscarón-Arias, 2007; Crosskey & Howard, 2004; MaterialExamined).
Biology and medical importanceThe immature stages of S. subnigrum are commonly attached to vegetation in small to medium (3-5 m
wide, 20 cm), fast flowing streams with sandy or rocky bottom. Strieder & Py-Daniel (1999) recorded itbreeding in small spillways in dams together with S. inaequale & S. perflavum. In studies on the diversity ofSimuliidae in Rio Grande do Sul, Strieder et al. (2004) collected this species in streams between 220 m and400 m, and above 850m altitude together with S. inaequale. Little is known about the female feeding habits ofthis species. It has never been found biting man along its distribution range, except by Pepinelli et al. (2003)who recorded its anthropophily in their studies in the Parque Estadual Intervales in São Paulo. Lutz (1910)stated that he collected females of S. subnigrum on horses.
Simulium (Inaequalium) travassosi D’Andretta & D’Andretta (Figs. 14, 30, 65, 66, 104-106, 119, 133, 147, 166, 167, 181, 182, 193, 204, 236-239)
Simulium travassosi D’Andretta & D’Andretta, 1947: 161. HOLOTYPE % (reared), BRAZIL: [Without locality, date orcollector] (MZUSP)[Examined.]
Female. General body color black. Body length (specimens pinned) 1.8-2.3 mm (mean=2.0 mm,
s.d.=0.20, n=9); wing length 1.8-2.4 mm (mean=2.0 mm, s.d.=0.18, n=8); wing width 0.7-1.1 mm (mean= 0.9mm, s.d.=0.10, n=8).
Head—dichoptic, with dark red eyes and nudiocular area well developed (Fig. 14). Frons, clypeus andocciput black, with silvery grey pruinosity; frons, clypeus and occiput with semi-recumbent, black hairs.Mouthparts black. Antennae with scape and pedicel yellowish brown, remainder of flagellar segments black.Cibarium with well-developed, sclerotized cornuae and rows of sharp teeth of varying size extending frombase of cornuae to central area of cibarium, which is weakly protuberant (Fig. 30).
Thorax—scutum black covered with recumbent, golden hairs. Scutal pattern variable depending on lightincidence. With anterior illumination scutum black, with faint grey pruinosity on median region of thorax;humeri, lateral and posterior margins black (Fig. 65). With posterior illumination, thorax black, with 1+1median, broad, grey pruinose vittae in anterior third of scutum; humeri grey pruinose; lateral margins black(Fig. 66). Scutellum dark brown to black, with recumbent, golden hairs interspersed with long, black bristles.Postnotum black, with grey pruinosity. Pleura black, with silver pruinosity. Costa of wing with sparsedistribution of spines and setae. Subcosta with row of setae, except apical third. Radius with row of setaeintermixed with distinct spines, basal section of radius with row of setae. Basal tuft of sparse, light brownsetae. Leg coloration and proportions as in Figs. 104-106. Foreleg with coxa, trochanter and external surfaceof femur pale brown; internal surface of femur, basal and apical third of tibia, and tarsal segments dark brown,remainder of tibia whitish. Mid leg with coxa, trochanter, apical half of tibia and basitarsus and tarsalsegments II-IV dark brown; femur pale brown; basal half of tibia and basitarsus whitish. Hind leg with coxa,trochanter, femur, apical half of tibia, apical third of basitarsus, and tarsal segments II-IV dark brown; basalhalf of tibia and two thirds of basitarsus whitish. Hind femur covered with spatulate setae. Claw weaklycurved with small basal tooth. Haltere lemon yellow, with dark brown base.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 49NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
Abdomen—tergites I-II dark brown, with grey pruinosity on anterolateral margin of tergite I and lateralmargins of tergite II; tergites III-V velvet black, VI-IX shiny, dark brown to black. Tergal plates developed.Sternites greyish; genitalia black. Eighth sternite weakly sclerotized with long, irregularly distributed setae onposterior margin; gonapophyses nearly same length as sternite at midpoint, subtriangular with inner posteriorcorner protruding, densely covered in microtrichiae, internal borders sclerotized (Fig. 119). Cercus suboval,covered with distinct, long brown setae; paraproct subrectangular, extending beyond cercus by half depth ofcercus, paraproct with long setae of posterior half and macrotrichiae on anterior margin apically (Fig. 133).Genital fork sclerotized with ends of lateral arms parallel to transverse axis of abdomen, anterior processes oflateral arms triangular and well developed, posterior processes partially developed (Fig. 147). Spermathecasuboval, without external sculpturing and irregularly distributed spicules on internal surface; area of insertionof spermathecal duct membranous.
Male. General body color black. Body length (specimens pinned) 1.8-2.6 mm (mean=2.1 mm, s.d.=0.37,n=3); wing length 1.4-2.0 mm (mean=1.7 mm, s.d.=0.33, n=3), wing width 1.0-1.1 mm (mean=1.0 mm,s.d.=0.08, n=3).
Head—holoptic, with dark red eyes. Remainder of head coloration as in female.Thorax—scutum dark black covered with evenly distributed golden, recumbent hairs. Scutal pattern varies
slightly with light incidence. With light source anterior to specimen scutum black, with 1+1 sublateral, grey-pruinose oval areas and faint 1+1 faint, median pruinose bands on anterior one third of scutum; humeri, lateraland posterior margins black (Fig. 166). With light posterior, thorax black; humeri, lateral and posteriormargins of scutum dark brown (Fig. 167). Scutellum dark brown covered with recumbent, golden hairsinterspersed with long, erect, dark brown setae. Postnotum dark brown, with silvery grey pruinosity. Pleurablack, with grey pruinosity. Haltere whitish-yellow, with light brown base. Wing setation as in female, exceptSc bare. Leg coloration as in female, except nearly entire hind femur pale brown, and slightly spatulate setaeon hind tibia and femur.
Abdomen—tergites dark brown to black, basal fringe of long, brown hairs. Silver pruinose ornamentationas follows: whole of tergite II except for median part and ventrolateral margin of tergites IV-IX. Genitaliablack; sternites whitish pruinose; tergal plates developed. Gonocoxite rectangular; gonostyle half length ofgonocoxite, subtrapezoidal with apical protrusion terminating in single, stout spine; gonocoxite and gonostylecovered with long setae (Fig. 181, 182). Ventral plate sclerotized and crescent-shaped with pointed apex andbasal arms well developed, and main body covered with long hairs (Fig. 193). Median sclerite pyriform withapical incision on apical third (Fig. 204). Paramere with developed, lightly sclerotized basal process andseveral long and short spines along distal half (Fig. 204).
Pupa. Cocoon length dorsally 2.7-3.1 mm (mean=2.9 mm, s.d.=0.13, n=10); ventrally 2.3-3.4 mm(mean=3.0 mm, s.d.=0.30, n= 10); pupa length 3.2-4.2 mm (mean= 3.6 mm, s.d.=0.35, n=10); gill length 1.7-2.3 mm (mean=2.0 mm, s.d.=0.18, n=10).
Cocoon—slipper shaped, light to dark brown composed of thin network of fibers and reinforced rim toanterior aperture.
Gill—light brown commonly with three thickened, forwardly directed filaments arranged in vertical plane.Gill configuration highly variable with main trunk short, commonly giving rise to three primary branches, onedorsal, one median and one ventral all dividing basally (Figs. 236-238). Variation in the gill configurationoccurs with dorsal filament bifurcated apically (Fig. 239); the median and ventral branches can also bedivided apically (see D’Andretta & D’Andretta, 1947, Figs. 86-112, pp. 174-175). All filaments stout, usuallyrounded distally, covered with thin spicules on surface, edges mainly smooth; dorsal and median filamentsoften longer than ventral filament.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.50 · Zootaxa 1649 © 2007 Magnolia Press
Head—frontoclypeus with 2+2 bifid (some trifid) frontal and 1+1 bifid, facial trichomes; frontoclypeuswith group of platelets mesally, 1+1 groups of approximately six platelets dorsolaterally and 2-3 platelets ingroups of two laterally in frontal region; tubercles rounded and densely distributed over entire surface.
Thorax—with 5+ 5 large, bifid to quadrifid trichomes near margin of dorsal cleft, one long, simpletrichome on posterior region mesally and one long, simple trichome near base of gill ventrally; tuberclesrounded and densely distributed over entire surface of thorax.
Abdomen—tergite I with 1+1 long, sublateral, simple trichomes; tergite II with 4+4 submedian spiniformsetae in row, the outer fourth seta more spaced than other three inner setae, 1+1 spiniform setae anterior toouter setae, and 3+three small, single setae on lateral margin; tergites III and IV with 4+4 submedian, simplehooks in row along posterior margin, 1+1 simple setae anterior to outer hooks, and three small, simple setaeon lateral margin; tergite V with 4+4 sublateral, simple setae in row along posterior margin and 1+1 small,single setae on lateral margin; tergite VI with 2+2 submedian, simple trichomes and spine combs on anteriormargin (in some specimens the most inner trichomes are well developed, resembling teeth); tergites VII andVIII with 2+2 small, sublateral simple trichomes on posterior margin and well-developed spine combsresembling teeth on anterior margin; tergite IX weakly sclerotized, with well-developed spine combsresembling teeth on anterior margin and 1+1 small, apical spines. Abdominal sternite III with 3+3 sublateraland 1+1 lateral, simple trichomes; sternite IV with 2+2 median and 2+2 small, sublateral simple trichomes,and 1+1 small, simple trichomes on lateral margin; sternite V with 2+2 submedian, close bifid hooks onposterior margin and 2+2 small, simple trichomes anterior to outer hooks; sternites VI-VII with 2+2 wellseparated hooks on posterior margin, inner being bifid or trifid and outer simple, 1+1 or 2+2 simple, smalltrichomes anterior to outer hooks, 1+1 small, simple trichomes between outer hooks, and 1+1 small, simpletrichomes on lateral margin; sternite IX weakly sclerotized. Spine combs on anterior margin of sternites III-IX.
Taxonomic discussionSimulium travassosi was described by D’Andretta & D’Andretta (1947) based on several females, males,
pupae and larvae collected in various localities in the states of Rio de Janeiro and São Paulo. These authorspointed out the misidentification of the pupa of S. travassosi with S. botulibranchium made by Pinto (1932:688, Pl. 9, Fig. q; 1938:137, Pl. 46, Fig. 5) and also established Angra dos Reis, Rio de Janeiro State as thetype locality for S. travassosi. We have examined the holotype of this species, which is deposited in theMZUSP, and one pinned male and female paratypes housed in the BMNH. The holotype is on three slidescontaining the adult body parts and the pupal exuviae, which are in relatively good condition (see MaterialExamined).
The females of S. travassosi are externally similar to other species of Inaequalium in having a black thoraxwith no distinct pattern (e.g. S. botulibranchium and S. clavibranchium) [light illumination anterior]. Themales are also similar to several other species in the subgenus in having a black thorax with 1+1 sublateral,grey pruinose areas with anterior illumination (e.g. S. inaequale, S. margaritatum, S. rappae and S.subnigrum). Simulium travassosi can only be reliably distinguished from other species of Inaequalium by thenumber and configuration of the pupal gill filaments. The pupal gill filaments are distinctly stout in S.travassosi, a character also seen in S. rappae. However, they can be reliably distinguished by the number ofgill filaments, three in S. travassosi and six in S. rappae (see also Taxonomic discussion under S. rappae).
A full description of adults and immature stages of S. travassosi may be found in Coscarón (1991),Coscarón & Coscarón-Arias (2007), Coscarón & Wygodzinsky (1984) and Strieder & Py-Daniel (1999;2000). Coscarón (1987) and Coscarón & Coscarón-Arias (1987, 2007) placed S. travassosi in the inaequale-species group. However, Strieder & Py-Daniel (2002) in their recent cladistic analysis placed S. travassosi asa basal species in the botulibranchium-species group. We agree with the placement of S. travassosi in theinaequale species group based on the morphology of the male and female genitalia.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 51NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
DistributionSimulium travassosi is a common, widespread species in southern Brazil and northern Argentina. In Brazil
it has been recorded in the states of Goiás, Minas Gerais, Rio de Janeiro, Rio Grande do Sul, São Paulo. InArgentina it has only been found in Missiones Province (Coscarón & Coscarón-Arias, 2007; Crosskey &Howard, 1997; 2004; Material Examined).
Biology and medical importance In Brazil, the immature stages of S. travassosi have been found attached to vegetation in small (20 cm to 1
m wide, 0.5 cm to 30cm deep), medium to fast flowing streams with a variety of river-beds (rocks, sand andmud) sympatrically with other simuliid species, e.g. S. anamariae Vulcano, S. botulibranchium, S.clavibranchium, Lutzsimulium hirticosta (Lutz), L. pernigrum (Lutz), and S. subnigrum (A.J. Shelley, A.P.A.Luna Dias & P.R. Garritano, unpublished data 2004). Nothing is known on the feeding habits of females of S.travassosi.
Species inquirendae
Simulium (Inaequale) lurybayae (Smart) (Figs. 7, 21, 43, 44, 85-87, 113, 126, 140)
This name applies to a poorly known species from Bolivia. Our description is based on the female holotype.
Simulium lurybayae Smart, 1944: 132. HOLOTYPE &, BOLIVIA: La Paz, Luribay; iii.1913, (Dr. Stoecker, S.G.)(ZMUH) [New name for Trichodagmia angustifrons Enderlein, 1934: 288, which was preoccupied by Trichodagmiaangustifrons Enderlein, 1921a: 200 and 1921b: 213 in Nevermannia]. [Examined.]
Female. General body color black. Body length 2.4 mm (pinned); wing length 1.9 mm; wing width 1.1 mm(n=1). Head—dichoptic, with dark red eyes and nudiocular area well developed (Fig. 7). Frons, clypeus andocciput black, with silvery grey pruinosity; clypeus with golden, semi-recumbent hairs; frons and occiput withsemi erect, black hairs. Mouthparts brown dark brown. Antennae with scape pale brown; remaining segmentsmissing. Cibarium with well-developed, sclerotized cornuae and group of sharp teeth of varying sizeextending from base of cornuae to central area of cibarium, which is distinctly protuberant (Fig. 21). Thorax—scutum color black, with recumbent, golden hairs with faint greenish reflection. Scutal patternvaries with direction of illumination. With anterior illumination, scutum black; humeri dark brown, with faintpruinosity; lateral and posterior margins black (Fig. 43). With posterior illumination, thorax black, with 1+1submedian, grey pruinose areas on anterior margin; humeri black, with grey pruinosity; lateral margins black(Fig. 44). Scutellum dark brown, with recumbent, brass-colored hairs and several dark brown, long bristlesmainly on posterior margin. Postnotum black, with grey pruinosity. Pleura brownish black, with silverpruinosity. Costa of wing with sparse distribution of spines and setae. Subcosta with line of setae up to midlength of vein. Radius with single row of setae intermixed with several distinct spines, basal section of radiuswith single line of setae. Basal tuft of sparse, light brown setae. Leg coloration and proportions as in Figs. 85-87. Fore leg coxa, trochanter, femur, apex of tibia mid brown, outer face of tibia whitish; tarsi dark brown.Mid and hind legs with coxae, trochanters; femora, apical two thirds of tibiae, apical third of basitarsus andtarsal segments II-IV dark brown; basal third of tibia and two thirds of basitarsus whitish. Claw of mid legselongated with small basal tooth [claw difficult to see on other legs]. Haltere lemon yellow, with brown base.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.52 · Zootaxa 1649 © 2007 Magnolia Press
Abdomen—tergite I dark brown, with basal fringe of brass-colored hairs; tergites II-IV velvet dark brown,tergite II with silver pruinosity on posterior margin; V-IX shiny dark brown. Tergal plates developed.Sternites and genitalia black. Eighth sternite sclerotized with long setae on posterior margin except for centralarea; gonapophyses well developed, subtriangularmembranous, internal margins lightly sclerotized, withsmall setae distributed over entire surface (Fig. 113). Cerci suboval, covered by long, brown setae; paraproctsubrectangular, nearly twice as long as cercus, sclerotizedwith prominent brown setae basally and smallmacrotrichia apically (Fig. 126). Genital fork thin and sclerotized throughout; lateral arms well developed,with distinct anterior processes (Fig. 140). Spermatheca globular, sclerotized without external sculpturing andsingle spicules on internal surface; area of insertion of spermathecal duct membranous.
Male. Unknown.
Pupa. Unknown.
Taxonomic discussion Simulium angustifrons was described as Trichodagmia angustifrons by Enderlein (1934) from a female
collected in La Paz, Bolivia on 1913 by S.G. Stoecker. This name was preoccupied by Trichodagmiaangustifrons described by Enderlein (1921a, b) in the subgenus Nevermannia. Consequently, Smart (1944)provided the new name S. lurybayae. An incorrect spelling of this species [as Simulium luribayae] is cited byCrosskey & Howard, 1997; 2004.]
Coscarón (1991) stated that S. lurybayae was externally similar to S. subnigrum, but the pilosity on thethorax was more greenish in the holotype of S. lurybayae than in specimens of S. subnigrum. Coscarón &Coscarón-Arias (1997, 2007) did not include S. lurybayae or S. clarki in their phylogenetic analysis ofInaequalium due to the lack of material, but assumed that based on the morphological descriptions of knownInaequalium species the last two species were more related to S. inaequale.
We have examined the single female of S. lurybayae which is housed in the NMHU, and compared it withnumerous link-reared specimens identified as S. clarki [now a synonym of S. inaequale], S. inaequale, and S.subnigrum deposited in the BMNH, IOC and MLP collections. The holotype is in good condition and we havetaken digital images of the specimen prior to dissection of the head, wings, abdomen, legs and genitalia,which are mounted on a slide. The thorax remains pinned. The general morphology of the female (Figs. 43,44), especially the cibarium (Fig. 21) and cercus and paraproct (Fig. 126) falls within the variation found in S.inaequale and S. subnigrum. However, S. lurybayae differs from these two species in its black thorax withouta distinct pattern covered by irregularly distributed, golden hairs with greenish reflections (Figs. 43, 44). Insome specimens of S. inaequale and S. subnigrum faint 1+1 median and 1+1 submedian, pruinose vittae canbe seen when the light source is anterior to the specimens (Figs. 41, 63). The adults of most Inaequaliumspecies are externally morphologically homogenous and they cannot be easily identified without examinationof the pupa gill configuration. In the absence of other life stages in S. lurybayae, we agreed with Coscarón(1991) and maintain this species as species inquirenda within Inaequalium.
Crosskey & Howard (2004) followed Coscarón (1991) in placing S. lurybayae in the botulibranchiumspecies group together with S. botulibranchium, S. petropoliense and S. souzalopesi. However, themorphology of the paraproct is different from those species in the botulibranchium species group. In S.lurybayae, the paraproct is subrectangular, extending ventrally nearly by the same length as the cercus, andwith a small protuberance basally (Fig. 126). This form agrees with the general morphology of most of thespecies in the inaequale species group, where it has now been placed (see also Hernández et al., 2006).
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 53NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
DistributionSimulium lurybayae is only known from its locality, La Paz, Bolivia (Coscarón & Coscarón-Arias, 2007;
Crosskey & Howard, 1997; 2004).
Biology and medical importanceNothing is known about the biology or the feeding habits of the females of S. lurybayae.
Simulium (Inaequalium) parimaense Ramírez Pérez, Yarzábal, Takaoka, Tada & Ramírez(Figs. 11, 27, 95-97, 130, 144)
There is confusion about the provenance of the female holotype of this species and its pupal exuviae. We havebeen unable to obtain type material (see Notes on Ramírez Pérez Simulium collection) and so thedescription of the female here provided is based upon the translation from Spanish of the original descriptiongiven in Ramírez Pérez et al. (1986) [The description of the pupa is not provided in this paper because it doesnot belong to a species of Inaequalium, see Taxonomic discussion.]
Simulium parimaensis Ramírez Pérez et al., 1986: 61. HOLOTYPE & (reared). VENEZUELA, T.F de Amazonas State,Department of Atabapo, Serra de Parima, Rio Aloquitetä, Parima B, 950 m; [Without date or collector’s name.][Holotype said to be deposited in IDVC, but it might be lost—see Note on Ramírez Pérez Simuliidae collection.]
Female. Wing 2 mm [other measurements not given in the original description]. Frons and clypeus black,with grey pruinosity. First two antennal segments and half of third yellow, remaining segments dark withwhite pubescence. Eyes black and nudiocular area developed (Fig. 11). Cibarium with well-developed,sclerotized cornuae and group of sharp teeth of varying size extending from base of cornuae to central area ofcibarium, which is distinctly protuberant (Fig. 27).
Thorax—black covered with irregular, golden hairs. Thorax anteriorly with 1+1 submedian, weaklypruinose spots. Scutellum dark brown, with golden setae and long, black hairs on posterior margin. Postnotumgreyish black, pruinose. Pleura dark brown, with grey pruinosity. Halteres yellow, with base dark. Fore legwith coxa, trochanter and femur brown; tibia white on external surface, and dark at its base and towards apex;tarsal segments black. Mid and hind legs with coxae, trochanters, and femora brown; tibiae brown, exceptbasal third white; tarsal segments I-IV white, except distal third black (Figs. 95-97).
Abdomen—tergal segments I-IV black, remaining segments shiny black, with golden pilosity that turnsblack depending on light incidence; tergite II brown, with silver pruinosity. Paraproct reduced (Fig. 130).Genital fork sclerotized; anterior process developed (Fig. 144).
Male. Unknown.
Pupa. The illustration of the pupa given in the original description is unlike to any species of Inaequalium(Strieder & Py-Daniel, 2000).
Taxonomic discussionRamírez Pérez et al. (1986) described S. parimaense [as S. parimaensis] based on a single female and its
pupal exuviae collected in Amazonas Territory in Venezuela. The female holotype and the pupal exuviae weremounted on a slide, which has been lost (see Note on Ramírez Pérez Simuliidae collection in Material andMethods).
Ramírez Pérez et al. (1986) stated that S. parimaense could be included in the subgenus Inaequaliumbased on the characters given in Coscarón & Wygodzinsky (1984). In the same paper, the latter authors also
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.54 · Zootaxa 1649 © 2007 Magnolia Press
used the female thoracic pattern and the morphology of the cibarium to place the species in the subgenusInaequalium. In addition, they commented that the pupa in this “group” [as “grupo” referring to the subgenusInaequalium] all have six filaments except S. parimaense that has 18 to 20 filaments. The name S. parimaensewas not listed in the review of Coscarón (1987) and world list of Crosskey (1988), and more recently, Cross-key & Howard (1997, 2004) considered it as “unplaced to species group within the subgenus Psaronio-compsa”. It is treated as species inquerenda in Coscarón & Coscarón-Arias (2007).
Strieder & Py-Daniel (2000) have questioned the identity of S. parimaense. They pointed out that thefemale description and illustration of the genitalia and cibarium of S. parimaense given in Ramírez-Pérez etal. (1986) clearly show that it belongs to the subgenus Inaequalium. However, the pupal gill configurationcorresponds to species of Psilopelmia (treated by Strieder & Py-Daniel (2000) as the genus EctemnaspisEnderlein). In addition, Strieder & Py-Daniel (2000) examined pupae and larvae collected from Xitei/Xidea,Roraima in Brazil, approximately 350 kms west of Boa Vista, which is near the type locality of S. parimaense.They concluded that the pupal gill configuration on these specimens conforms with the figures of the pupa ofS. parimaense in Ramírez-Pérez et al. (1986). They also showed that the morphology of the cibarium obtainedfrom the pharate pupae in this material belongs to a species of Ectemnaspis. The authors finally concludedthat what was in the slide preparation of S. parimaense might represent two separate species, and therefore,considered S. parimaense as species inquirenda. In addition, the first author of this paper has examined oneslide containing a single pupa identified as S. parimaense in the MLP collections. The pupa had beencollected in Roraima, Ao. Sta Anita on 27.xi.1987 by R. Pérez et al. However, the number of gill filamentsand pupal gill configuration in this material falls within the variation found in S. rorotaense/S. maroniense(sensu Shelley et al., 2004) of the subgenus Psilopelmia and it has been labelled accordingly.
We have studied the original description and illustrations of the female and the pupa of S. parimaense byRamírez-Pérez et al. (1986) and agree with Strieder & Py-Daniel (2000) that two separate species wereprobably involved in the description of this species, with the female being a species of Inaequalium, and thepupa appearing to be a member of Psilopelmia. The morphology of the female cibarium, cercus and paraproct(Figs. 27, 130, 144) are typical of species of Inaequalium, inaequale species group. Therefore, in the absenceof further material we regard S. parimaense as a species inquirenda.
Distribution Simulium parimaense is only known from its type locality in the Serra da Parima, Venezuela (Crosskey &
Howard, 2004).
Biology and medical importance Nothing is known about the female feeding habits of this species.
Unavailable name
Simulium (Inaequalium) hauseri of Coscarón & Coscarón-Arias, 2007: 283
In the recently published book on the Simuliidae of the Neotropical Region Coscarón & Coscarón-Arias(2007) treated the species S. hauseri within the subgenus Inaequalium and considered it very close to S.inaequale. The authors distinguished S. hauseri from S. inaequale and S. clarki by gill branching being veryclose to the base and fewer tubercles on the frontoclypeus and thorax of the former species. Coscarón & Cos-carón-Arias (2007) cited in their work the reference of Strieder (1998) when referring to the original descrip-tion of S. hauseri. This description was made by Strieder (1998) [as Inaequalium hauseri] in his PhD thesisentitled “Revisão e analise cladística de espécies de Inaequalium (Coscarón & Wygodzinsky, 1984) (Diptera:
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 55NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
Simuliidae)” [Revision and cladistic analysis of species of Inaequalium”]. He recognized this species basedonly on seven pupal exuviae collected in the state of Rio Grande do Sul, Brazil. Subsequent to his thesis,Strieder published several papers about Inaequalium (e.g., Strieder, 1998; Strieder & Py-Daniel, 1999, 2000,2002), but this species name was never cited. A PhD thesis is not considered a published work unless theauthor provides details in accordance with articles 8-9 of the ICZN (International Commission of ZoologicalNomenclature, 1999). These rules were not followed by Strieder in the case of S. hauseri. The first publishedreference to S. hauseri was in Coscarón & Coscarón-Arias (2007), who did not follow the principles of nameavailability ICZN (1999, articles 10-20; see also principles 1-10(a-c)). The name is therefore attributable tothese authors and is unavailable.
Acknowledgements
We would like to thank The Natural History Museum (BMNH) and Instituto Oswaldo Cruz (IOC) forproviding funds for collecting and curating Neotropical Simuliidae over the years, and the second authoracknowledges the Emeritus Fellowship provided by the Leverhulme Trust. We also thank Drs. RogerCrosskey and Ralph Harbach (BMNH) for advice on nomenclatural problems and Dr Klaus Sattler (BMNH)for kindly translating many of the original descriptions written in German by Enderlein. The authors aregrateful to the following colleagues for loans of types and other material from their collections: SixtoCoscarón, MLP, Argentina; Neusa Hamada & Victor Py-Daniel, INPA, Brazil; Milton Norberto Strieder,LEU; Mateus Pepinelli, Laboratório de Entomologia Aquática, Departamento de Hidrobiologia, UniversidadeFederal de São Carlos, São Paulo (UFSCar) [also MZUSP], Brazil; R. Contreras Lichtenberg & Peter Sehnal,NM, Finland; Doreen Werner and Joachim Ziegler, NMHU, Germany; Uwe Kallweit, SMT, Germany;Wayne Mathis & Nancy Adams, USNM, USA; David Grimaldi, AMNH, USA; David Furth & Philip Perkins,MCZ. Luis Hernández would like to thank to Maria E. Grillet (Instituto de Laboratorio de Vectores, Institutode Zoologia Tropical, Universidad Central de Venezuela, Venezuela), Hortencia Frontado, MEPG,Venezuela, and Maria Gloria Basañez (Imperial College, London, UK) for discussions on the status of JaimeRamirez Perez’ Simuliidae Collection. The first author of this paper would like to thank especially Dr. SergioLuz, Instituto Oswaldo Cruz, Fundação Fiocruz, Manaus, Brazil for providing facilities during a visit to thisinstitution, and Dr. Peter Adler for discussions regarding the taxonomy of the subgenus Inaequalium.
References
Coscarón, S. (1976) Notas sobre simúlidos neotropicales VI. Sobre dos especies nuevas de jejenes de la provincia deMisiones, Argentina (Diptera, Insecta). Revista de la Sociedad Entomolágica Argentina, 35, 147–154.
Coscarón, S. (1980) Notas sobre simulidos Neotropicales IX. Sobre un grupo de especies próximo al subgénero Simu-lium (Inaequalium) (Diptera—Insecta). Revista de la Sociedad Entomológica Argentina, 39, 293–301.
Coscarón, S. (1987) El género Simulium Latreille en la región neotropical: análisis de los grupos supraespecíficos, espe-cies que los integran y distribución geográfica (Simuliidae, Diptera). 112 pp. Museu Paraense Emílio Goeldi,Belém, Brazil.
Coscarón, S. (1991) Simuliidae. Fauna de Agua Dulce de la República Argentina 38 (Insecta Diptera) (2). FECIC, Bue-nos Aires, Argentina, 304 pp.
Coscarón, S. & Coscarón-Arias, C.L. (1997) Cladistic analysis of the subgenera Inaequalium, Psaroniocompsa, and Chi-rostilbia of the genus Simulium, with comments on their distribution. Revista de la Sociedad Entomológica Argen-tina, 56, 109–121.
Coscarón & Coscarón-Arias (2007). Neotropical Simuliidae (Diptera: Insecta). In: Adis, J., Arias, J.R., Rueda-Delgado,G. & K.M.Wantzen (Eds.): Aquatic Biodiversity in Latin America (ABLA). Vol. 3. Pensoft, Sofia-Moscow, 685 pp.
Coscarón, S. & Miranda-Esquivel, D.R. (1998) New synonymy to Simulium (Inaequalium) inaequale (Paterson & Shan-non, 1927). Memórias do Instituto Oswaldo Cruz, 93, 337.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.56 · Zootaxa 1649 © 2007 Magnolia Press
Coscarón, S. & Wygodzinsky, P. (1973) Notas sobre simulidos neotropicales. IV. Sobre la presencia de Simuliuminaequale Paterson & Shannon en el nordeste de la Argentina y sudeste del Paraguay, y una especie muy afin(Diptera, Simuliidae). Revista de la Sociedad Entomológica Argentina, 34, 141–149.
Coscarón, S. & Wygodzinsky, P. (1984) Notas sobre Simúlidos Neotropicales. VII. Sobre los subgéneros Psaronio-compsa Enderlein y Inaequalium subgen. nov. Arquivos de Zoologia, 31, 37–103.
Coscarón, S., Lopes de Cerqueira, Schumaker, T.T.S. & La Salvia Filho, V. (1992) Nuevos datos sobre distribución deSimulideos de Brasil y descripción de Simulium (Coscaroniellum) cerradense n.sp. (Diptera, Simuliidae). RevistaBrasileira de Entomologia, 36, 111– 119.
Crosskey, R.W. (1988) An annotated checklist of the world blackflies (Diptera: Simuliidae). Pp. 425–520. In Kim, K.C.& Merritt, R.W., eds., Blackflies: Ecology, population management, and an annotated world list. The PennsylvaniaState University, University Park.
Crosskey, R.W. 1999. First update to the taxonomic and geographical inventory of world blackflies (Diptera: Simuli-idae). The Natural History Museum, London, 10pp.
Crosskey, R.W. 2002. Second update to the taxonomic and geographical inventory of world blackflies (Diptera: Simuli-idae). The Natural History Museum, London, 14pp.
Crosskey, R.W. & Howard, T.M. (2004) A revised taxonomic and geographical inventory of world blackflies (Diptera:Simuliidae). The Natural History Museum, London, 82 pp. [web version: http://www.nhm.ac.uk/entomology/projects/blackflies/Inventory. pdf] (accessed 11 February 2006).
D’Andretta, M.A.V. & D’Andretta, C.D. Jr. (1947) Espécies neotropicais de família Simuliidae Schiner (Diptera, Nema-tocera) III. Simulium botulibranchium Lutz, 1910 e Simulium travassosi n. sp. Papéis Avulsos, Departamento deZoologia, São Paulo, 8, 145–180, + 125 Figs.
D’Andretta, M.A. & D, Andretta Jr., C.A. (1949) Espécies neotropicais da família “Simuliidae” Schiner (Diptera, Nema-tocera). V. Rediscrição do “Simulium auristriatum” Lutz, 1910 e “Simulium (Eusimulium) venustum infuscata” Lutz,1909 var. inquirendae. Revista Brasileira de Biologia, 9, 55–66.
D’Andretta, C. & González B., D. (1964) Três novas espécies brasileiras de Simuliidae (Diptera Nematocera). Memóriasdo Instituto Butantan 30, 103–116.
Eaton, D.P., Diaz, L.A., Hans-Filho, G., Santos, V. D., Aoki, V., Friedman, H., Rivitti, E.A., Sampaio, S.A.P., Gottlieb,M.S., Giudice, G. J., Lopez, A. & Cupp, E. W. (1998) Comparison of black fly species (Diptera: Simuliidae) on anAmerindian reservation with a high prevalence of fogo selvagem to neighbouring disease-free sites in the state ofMato Grosso do Sul, Brazil. Journal of Medical Entomology, 35, 120–131.
Enderlein, G. (1921a) Das System der Kriebelmücken (Simuliidae). Deutsche Tierärztliche Wochenschrift, 29, 197–200.[Actual publication date was April 1921.]
Enderlein, G. (1921b) Neue paläarktische Simuliiden. Sitzungsberichte der Gesellschaft naturforschender Freunde zuBerlin, 1921, 212–224. [Actual publication date was June 1921.]
Enderlein, G. (1934) Weiterer Ausbau des Systems der Simuliiden. (Dip.). Deutsche Entomologische Zeitschrift, 1933,273–292.
Enderlein, G. (1936) Simuliologica I. Sitzungsbericht der Gesellschaft naturforschender Freunde ausegegeben zu Berlin,1936, 114–130.
Fairchild, G.B. (1940) Notes on the Simuliidae of Panama (Dipt., Nematocera). Annals of the Entomological Society ofAmerica, 33, 701–719.
Gil-Azevedo, L.H. & Maia-Herzog, M. (2004) Registros novos de ocorrência de seis especies de Simuliidae (Diptera)para o estado do Rio de Janeiro. Biota Neotropica, 4, 1–3.
Hernández, L.M. & Shelley, A.J. (2005) New specific synonymies and taxonomic notes on Neotropical blackflies(Diptera: Simuliidae). Zootaxa, 853, 1–46.
Hernández, L.M., Luna Dias, A.P.A., Maia-Herzog, M. & Shelley, A.J. (2006). Taxonomy of Simulium (Inaequalium)petropoliense Coscarón from Brazil, with first description of the male and larva. Zootaxa, 1275, 1–20.
Hernández, L.M., Shelley, A.J., Luna Dias, A.P.A. & Maia-Herzog, M. (2007). New specific synonymies and taxonomicnotes on Neotropical Blackflies (Diptera: Simuliidae) belonging to the subgenera Chirostilbia Enderlein, Hemic-netha Enderlein, Inaequalium Coscarón & Wygodzinsky, Psaroniocompsa Enderlein and Psilopelmia Enderlein.Zootaxa, 1506, 1–80.
Hernández, L.M. & Shelley, A.J., Luna Dias, A.P.A. & Maia-Herzog, M. (2005) Re-description of the female, male andpupa of Simulium itaunense D’Andretta & González B. (Diptera: Simuliidae). Zootaxa, 896, 1–14.
International Commission on Zoological Nomenclature (1999) International Code of Zoological Nomenclature, 4th Edi-tion, The International Trust for Zoological Nomenclature, xix + 273 pp.
Juñent, S.R. & Coscarón, S. (2001) Biogeographical history of the Neotropical and Nearctic Simuliidae (Diptera).Revista del Museo Argentino de Ciencias Naturales, 3, 119–134.
Lutz, A. (1910) Segunda contribuição para o conhecimento das especies brazileiras do género "Simulium". Memórias doInstituto Oswaldo Cruz, 2, 213–267.
Lutz, A. (1922) Zoologia Médica. Nematoceros hematophagos não pertecendo aos Culicideos. A Folha Médica, 12, 89–
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 57NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
92.Nunes de Mello, J. A. & Barbosa de Almeida, F. (1974) Simuliidae da Amazônia VII. Descrição de Simulium pseudoex-
iguum sp. n. (Diptera, Nematocera). Acta Amazonica, 4, 65–68.Paterson, G. & Shannon, R.C. (1927) Los simúlidos del noroeste Argentino. Revista del Instituto Bacteriológico, 4, 737–
742.Pepinelli, M. & Trivinho-Strixino, S. (2002) Colonizaçao de substratos artificias de diferentes cores por imaturos de Sim-
ulium inaequale Paterson & Shannon, 1927 (Diptera, Simuliidae). Entomologia y Vectores, 9, 349–357.Pepinelli, M., Trivinho-Strixino, T. & Hamada, N. (2003) Imaturos de Simuliidae (Diptera, Nematocera) e caracterização
de seus criadouros no Parque Estadual Intervales, SP, Brasil. Revista Brasileira de Entomologia, 49, 527–530.Pepinelli, M., Hamada, N. & Luz, S.L.B. (2006) Simulium (Inaequalium) margaritatum, a new species of black fly
(Diptera: Simuliidae) from the central region of Bahia State, Brazil. Zootaxa, 1167, 47–60.Pessoa, F.A.C., Ríos-Velásquez, C.M. & Py-Daniel, V. (2005) First survey of Simuliidae (Diptera) from the north of
Ceará state, Brazil, with description of a new species and identification keys for the immature stages. Zootaxa, 1051,1–18.
Pinto, C. (1932) Simuliidae da America Central e do Sul. Reunión de la Sociedad Argentina de Patologia Regional delNorte [1931], 7, 661–763 + I [corrigenda].
Py-Daniel, V. (1984) Simuliidae (Diptera: Culicomorpha) no Brasil. II. Sobre o Simulium nahimi sp. n. Boletim doMuseu Paraense Emilio Goeldi Zoologia, 1, 125–137.
Py-Daniel, V. & Coscarón, S. (1982) Simuliidae (Diptera: Nematocera) no Brasil. 1. Sobre o Simulium rappae sp.n.Revista Brasileira de Biologia, 42, 155–163.
Py-Daniel, V. & Moreira, G.R.P. (1989) Simuliidae (Diptera, Culicomorpha) no Brasil IX. Simulium (Inaequalium)nogueirai D’Andretta & González, 1964. IHERINGIA. Série Zoologia, 69, 85–95.
Py-Daniel, V. & Moreira Sampaio, R.T. (1994a) Jalacingomyia gen.n. (Culicomorpha); a ressurreição de Gymnopaidi-nae; a eliminação do nivel tribal; apresentação de novos caracteres e redescrição dos estágios larval e pupal de Sim-ulium columbaschense (Fabricius, 1787) (Diptera: Simuliidae). Memorias del CAICET, 4, 101–148.
Py-Daniel, V. & Moreira Sampaio, R.T. (1994b) Actualização nomenclatural para Simuliidae Neotropical (Diptera, Culi-comorpha, Simuliidae). Memórias del CAICET, 4, 149–156.
Py-Daniel, V. & Moreira Sampaio, R.T. (1995) Gêneros e espécies de Simuliidae (Diptera: Culicomorpha) assinaladospara o Brasil até 1995. Entomologia y Vectores, 5, 117–121.
Ramírez-Pérez, J. (1983) "Los jejenes de Venezuela". Simposio de oncocercosis americana. Caicet. Puerto Ayacucho,15–17 Octubre, 193, iii + 156 pp.
Ramírez-Pérez, J., Rassi, E. and Ramírez, A. (1977) Cinco especies nuevas de Simulium Latreille, 1802 (Diptera, Simuli-idae) de la región amazónica de Venezuela. Boletin de la Dirección de Malariologia y Saneamiento Ambiental, 17,162–174.
Ramírez-Pérez, J., Yarzábal, L., Takaoka, H., Tada, I. & Ramírez, A. (1986) Simulium parimaensis (Diptera, Simuliidae),nueva especie de jejen en el Territorio Federal Amazonas, Venezuela. Boletin de la Dirección de Malariologia ySaneamiento Ambiental, 26, 61–64.
Shelley, A.J. (1988) Biosystematics and medical importance of the Simulium amazonicum group and the S. exiguumcomplex in Latin America. pp. 203–220, in Service, M.W. (Ed.), Biosystematics of haematophagous insects. xi +63pp. Oxford University Press, Oxford.
Shelley, A.J., Arzube, M. & Couch, C.A. (1989) A biosystematic study of the Simuliidae of the Santiago onchocerciasisfocus of Ecuador. Bulletin of the British Museum Natural History Entomology series, 58, 79–130.
Shelley A.J., Hernández L.M., Davies, JB (2004) Blackflies (Diptera: Simuliidae) of southern Guyana with keys for theidentification of adults and pupae- a review. Memórias do Instituto Oswaldo Cruz, 99, 443–470.
Shelley, A.J., Hernández, L.M. & Penn, M. (2002) A biosystematic revision of the blackflies (Diptera: Simuliidae) ofBelize, Central America. Bulletin of the Natural History Museum, London (Entomology), 71, 135–271.
Shelley, A.J., Hernández, L.M., Maia-Herzog, M. Maia-Herzog, Luna Dias, A.P.A. & Luz, S.B. (2006). An interpretationof the morphological variation in the Simulium amazonicum species group (Diptera: Simuliidae) of Latin America.Zootaxa, 1274, 1–68.
Shelley, A.J., Lowry, C.A., Maia-Herzog, M., Luna Dias, A.P.A. & Moraes, M.A.P. (1997) Biosystematic studies on theSimuliidae (Diptera) of the Amâzonia onchocerciasis focus of Brazil. Bulletin of the Natural History Museum (Ento-mology series), 66, 1–121.
Smart, J. (1944) Notes on Simuliidae (Diptera). II. Proceedings of the Entomological Society of London (B), 13, 131–136.
Strieder, M.N. (2004) Espécies de simulídeos (Diptera, Nematocera) no Rio Grande do Sul, Brasil: distribuição geográ-fica. Entomologia y Vectores, 11, 113–143.
Strieder, M.N. & Py-Daniel, V. (1999) Espécies de Inaequalium (Diptera, Simuliidae): dados bionômicos e chaves parasua identificação. Biociências, 7, 43–72.
Strieder, M.N. & Py-Daniel, V. (2000) Revisão de Inaequalium (Diptera, Simuliidae), com redescrição das formas imat-
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.58 · Zootaxa 1649 © 2007 Magnolia Press
uras e descrição de uma nova espécie. Entomología y Vectores (supplemento 2), 7, 1–90.Strieder, M.N. & Py-Daniel, V. (2002) A cladistic analysis of Inaequalium (Coscarón & Wygodzinsky, 1984), with infor-
mation on geographical distribution (Diptera: Simuliidae). Memórias do Instituto Oswaldo Cruz, 97, 65–72.Strieder, M.N., Corseuil, E. & Py-Daniel, V. 1992. Espécies do genero Simulium (Diptera, Simuliidae) ocorrentes no Rio
Grande do Sul – Brasil, com chaves para sua identificação. Acta Biologica Leopoldensia, 14, 53–73.Strieder, M.N., Santos, J.E. Jr. & Oliveira Pes (2004) Diversidade e distribuíção de Simuliidae (Diptera, Nematocera) no
gradiente longitudinal da bacia do Rios dos Sinos, no Rio Grande do Sul, Brasil. Entomologia y Vectores, 9, 527–540.
Werner, D. (1996a) Die Simuliiden-Typen der Dipteren-Sammlung des Zoologischen Museums Berlin (Diptera: Simuli-idae). Mitteilengun aus dem Zoologischen Museum in Berlin, 72, 221–258.
Werner, D. (1996b) Das Typenmaterial der Zweiflüger des Staatlichen Museums für Tierkunde Dresden (Insecta). TeilIII: Diptera-Simuliidae. Staatliches Museum für Tierkunde Dresden, 57, 295–298.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 59NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
PLATE 1. FIGURES 1–14. Female nudiocular area of: 1: S. botulibranchium; 2: S. petropoliense; 3: S. souzalopesi,
paratype; 4: S. clavibranchium; 5: S. diversibranchium; 6: S. inaequale; 7: S. lurybayae holotype; 8: S. maranguapense
(after Pessoa et al., 2005); 9: S. margaritatum; 10: S. mariavulcanoae holotype; 11: S. parimaense (after Ramírez-Pérez et al.
1986); 12: S. rappae; 13: S. subnigrum; 14: S. travassosi.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.
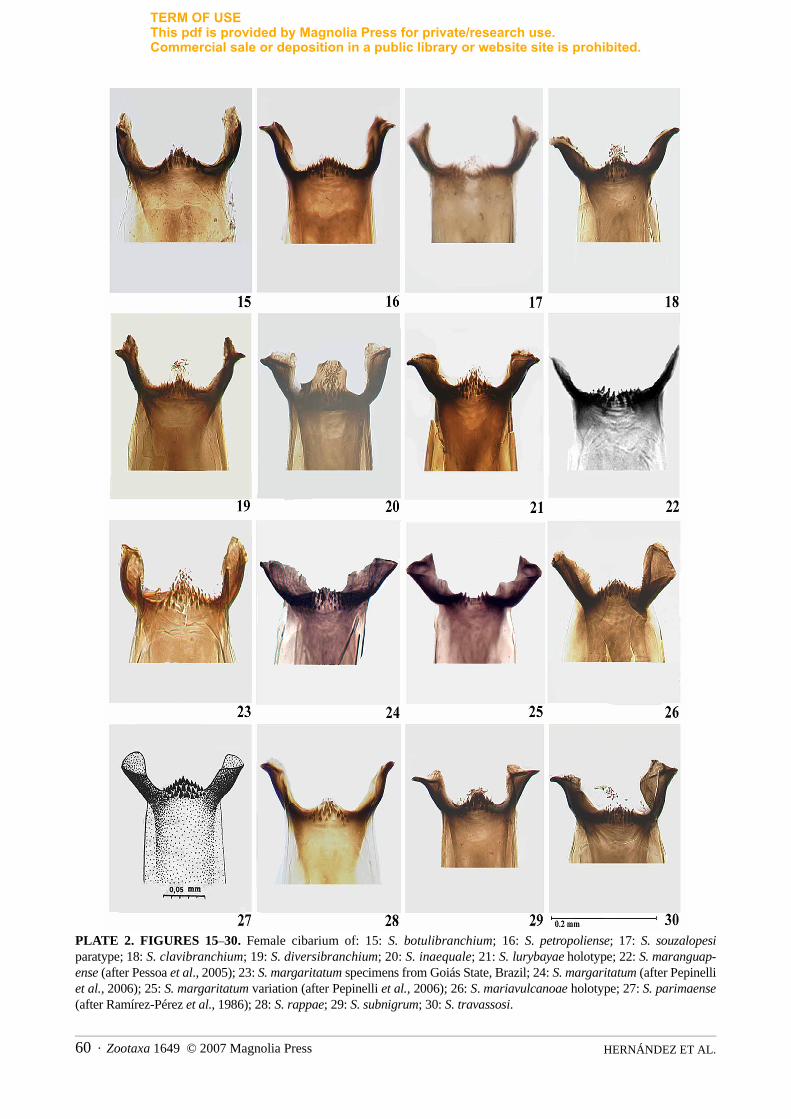
HERNÁNDEZ ET AL.60 · Zootaxa 1649 © 2007 Magnolia Press
PLATE 2. FIGURES 15–30. Female cibarium of: 15: S. botulibranchium; 16: S. petropoliense; 17: S. souzalopesiparatype; 18: S. clavibranchium; 19: S. diversibranchium; 20: S. inaequale; 21: S. lurybayae holotype; 22: S. maranguap-ense (after Pessoa et al., 2005); 23: S. margaritatum specimens from Goiás State, Brazil; 24: S. margaritatum (after Pepinelliet al., 2006); 25: S. margaritatum variation (after Pepinelli et al., 2006); 26: S. mariavulcanoae holotype; 27: S. parimaense(after Ramírez-Pérez et al., 1986); 28: S. rappae; 29: S. subnigrum; 30: S. travassosi.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 61NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
PLATE 3. FIGURES 31–46. Color patterns for female thorax. For each species first figure is with anterior illumination
and second figure with posterior illumination: 31, 32: S. botulibranchium; 33, 34: S. petropoliense (in spirit); 35, 36: S.
souzalopesi holotype; 37, 38: S. clavibranchium; 39, 40: S. diversibranchium; 41, 42, S. inaequale southern Brazil; 43, 44: S.
lurybayae holotype; 45, 46: S. margaritatum (after Pepinelli et al., 2006).
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.62 · Zootaxa 1649 © 2007 Magnolia Press
PLATE 4. FIGURES 47–58. Color patterns for female thorax. For each species first figure is with anterior illumination
and second figure with posterior illumination: 47, 48: S. margaritatum, Goiás State, Brazil; 49, 50: S. mariavulcanoae holo-
type; 51, 52: S. mariavulcanoae; 53, 54: S. rappae holotype; 55, 56: S. rappae; 57, 58: S. rappae variation, Goiás State, Salto
de Itiquira.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 63NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
PLATE 5. FIGURES 59–66. Color patterns for female thorax. For each species first figure is with anterior illumination
and second figure with posterior illumination; 59, 60: S. rappae variation, Goiás State, Salto de Itiquira; 61, 62: S. rappae
Saia Velha, Brazil; 63, 64: S. subnigrum; 65, 66: S. travassosi paratype.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.64 · Zootaxa 1649 © 2007 Magnolia Press
PLATE 6. FIGURES 67–90. Color pattern for female front, mid and hind legs, respectively: 67–69: S. botulibranchium;
70–72: S. petropoliense; 73–75: S. souzalopesi paratypes; 76–78: S. clavibranchium; 79–81: S. diversibranchium; 82–84:
S. inaequale (as S. clarki); 85–87: S. lurybayae holotype; 88–90: S. maranguapense (after Pessoa et al., 2005).
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.
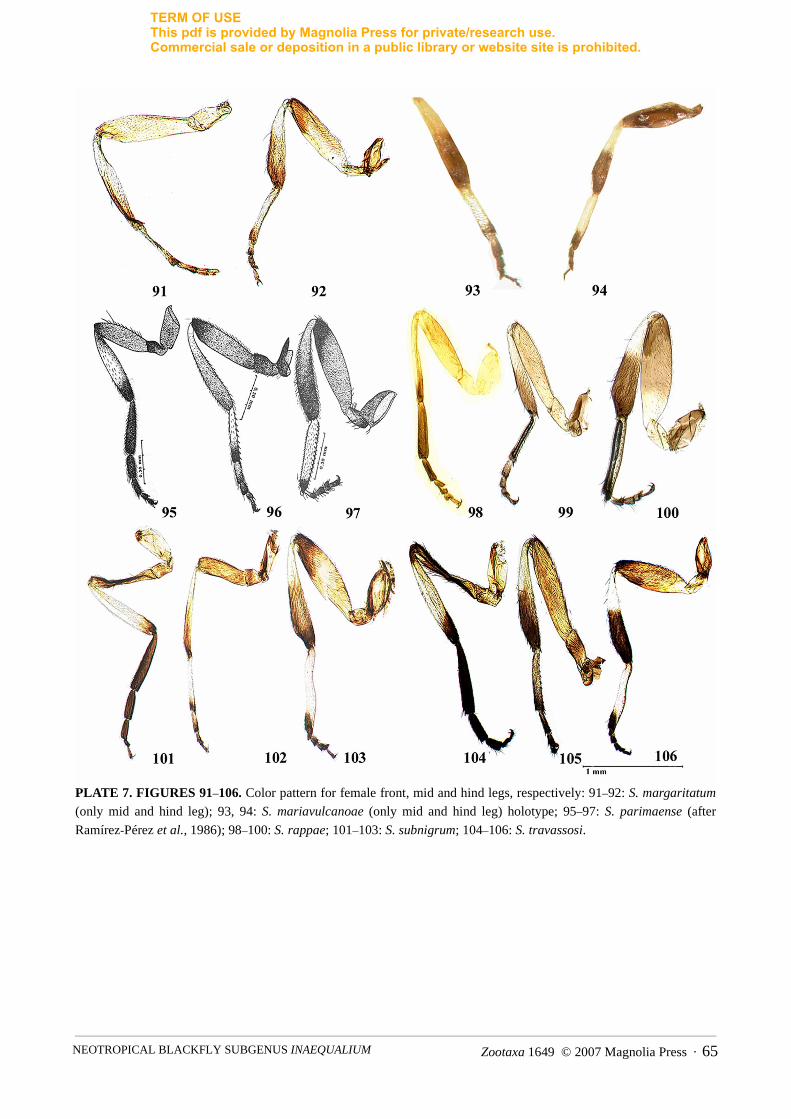
Zootaxa 1649 © 2007 Magnolia Press · 65NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
PLATE 7. FIGURES 91–106. Color pattern for female front, mid and hind legs, respectively: 91–92: S. margaritatum
(only mid and hind leg); 93, 94: S. mariavulcanoae (only mid and hind leg) holotype; 95–97: S. parimaense (after
Ramírez-Pérez et al., 1986); 98–100: S. rappae; 101–103: S. subnigrum; 104–106: S. travassosi.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.66 · Zootaxa 1649 © 2007 Magnolia Press
PLATE 8. FIGURES 107–113. Female gonapophyses of: 107: S. botulibranchium; 108: S. petropoliense holotype; 109:
S. souzalopesi; 110: S. clavibranchium; 111: S. diversibranchium; 112: S. inaequale (as S. clarki paratype); 113: S. lury-
bayae holotype.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 67NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
PLATE 9. FIGURES 114–119. Female gonapophyses of: 114: S. maranguapense (after Pessoa et al., 2005); 115: S.
margaritatum; 116: S. mariavulcanoae holotype; 117: S. rappae; 118: S. subnigrum; 119: S. travassosi.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.68 · Zootaxa 1649 © 2007 Magnolia Press
PLATE 10. FIGURES 120–133. Female cercus and paraproct of: 120: S. botulibranchium; 121: S. petropoliense; 122:S. souzalopesi; 123: S. clavibranchium; 124: S. diversibranchium; 125: S. inaequale (as S. clarki paratype); 126: S. lury-bayae holotype; 127: S. maranguapense (after Pessoa et al., 2005); 128: S. margaritatum; 129: S. mariavulcanoae; 130:S. parimaense (after Ramírez-Pérez et al., 1986); 131: S. rappae; 132: S. subnigrum; 133: S. travassosi.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 69NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
PLATE 11. FIGURES 134–147. Female genital fork of: 134: S. botulibranchium; 135: S. petropoliense; 136: S. souza-lopesi; 137: S. clavibranchium; 138: S. diversibranchium; 139: S. inaequale (as S. clarki paratype); 140: S. lurybayaeholotype; 141: S. maranguapense (after Pessoa et al., 2005); 142: S. margaritatum; 143: S. mariavulcanoae holotype;144: S. parimaense (after Ramírez-Pérez et al., 1986); 145: S. rappae; 146: S. subnigrum; 147: S. travassosi.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.
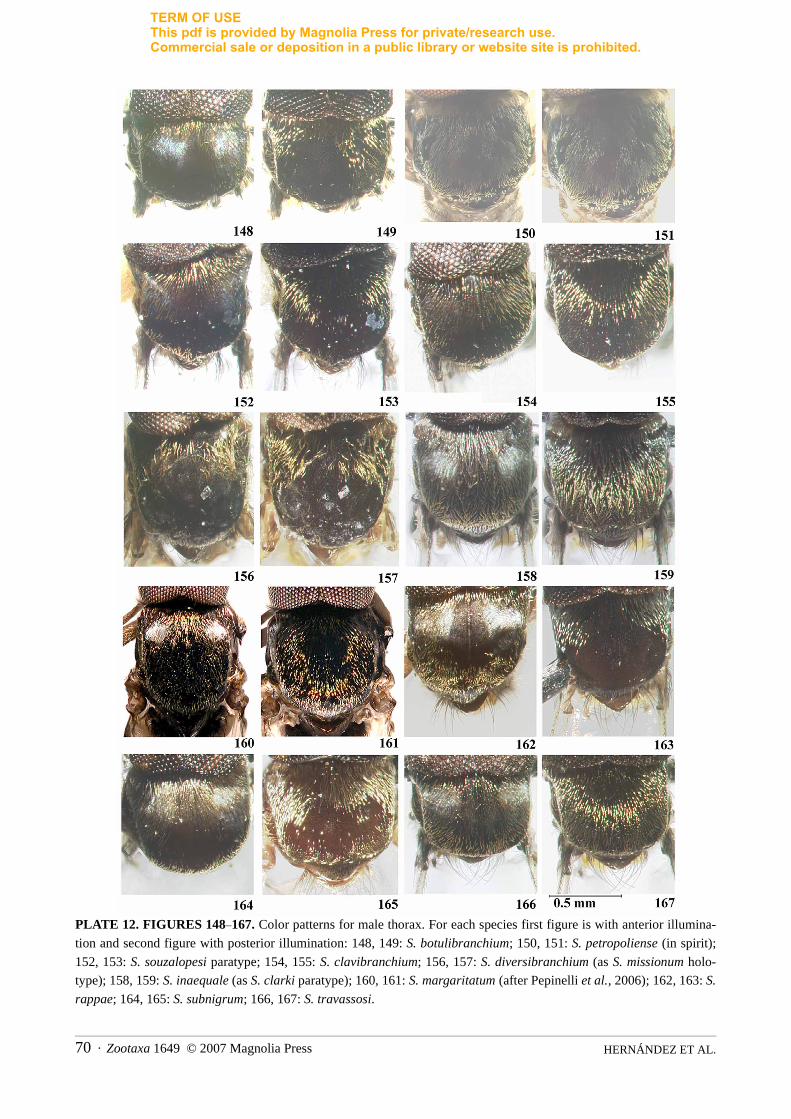
HERNÁNDEZ ET AL.70 · Zootaxa 1649 © 2007 Magnolia Press
PLATE 12. FIGURES 148–167. Color patterns for male thorax. For each species first figure is with anterior illumina-
tion and second figure with posterior illumination: 148, 149: S. botulibranchium; 150, 151: S. petropoliense (in spirit);
152, 153: S. souzalopesi paratype; 154, 155: S. clavibranchium; 156, 157: S. diversibranchium (as S. missionum holo-
type); 158, 159: S. inaequale (as S. clarki paratype); 160, 161: S. margaritatum (after Pepinelli et al., 2006); 162, 163: S.
rappae; 164, 165: S. subnigrum; 166, 167: S. travassosi.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.
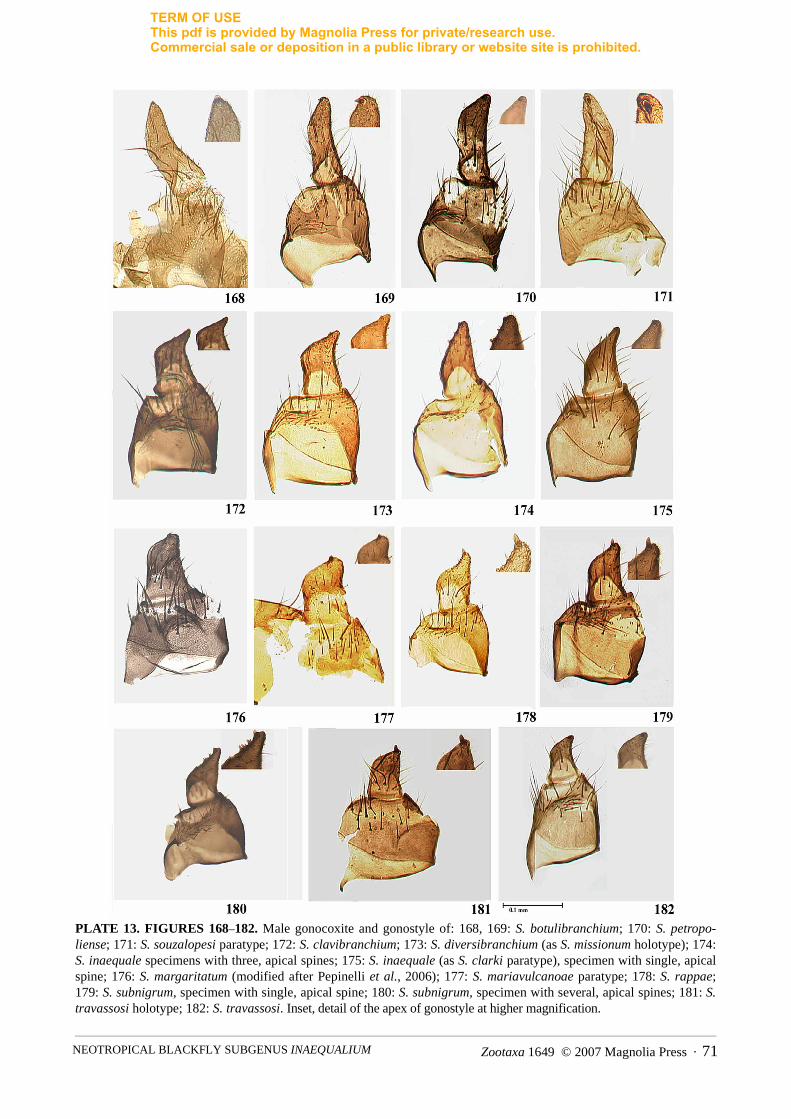
Zootaxa 1649 © 2007 Magnolia Press · 71NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
PLATE 13. FIGURES 168–182. Male gonocoxite and gonostyle of: 168, 169: S. botulibranchium; 170: S. petropo-liense; 171: S. souzalopesi paratype; 172: S. clavibranchium; 173: S. diversibranchium (as S. missionum holotype); 174:S. inaequale specimens with three, apical spines; 175: S. inaequale (as S. clarki paratype), specimen with single, apicalspine; 176: S. margaritatum (modified after Pepinelli et al., 2006); 177: S. mariavulcanoae paratype; 178: S. rappae;179: S. subnigrum, specimen with single, apical spine; 180: S. subnigrum, specimen with several, apical spines; 181: S.travassosi holotype; 182: S. travassosi. Inset, detail of the apex of gonostyle at higher magnification.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.72 · Zootaxa 1649 © 2007 Magnolia Press
PLATE 14. FIGURES 183–193. Male ventral plate of: 183: S. botulibranchium; 184: S. petropoliense; 185: S. souza-
lopesi; 186: S. clavibranchium; 187: S. diversibranchium (as S. missionum holotype); 188: S. inaequale (as S. clarki
paratype); 189: S. margaritatum (modified after Pepinelli et al., 2006); 190: S. mariavulcanoae; 191: S. rappae; 192: S.
subnigrum, 193: S. travassosi.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 73NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
PLATE 15. FIGURES 194–199. Male paramere and median sclerite of: 194: S. botulibranchium; 195: S. petropoliense;
196: S. souzalopesi paratype; 197: S. clavibranchium; 198: S. diversibranchium (as S. missionum holotype); 199: S.
inaequale (as S. clarki paratype).
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.74 · Zootaxa 1649 © 2007 Magnolia Press
PLATE 16. FIGURES 200–204. Male paramere and median sclerite of: 200: S. margaritatum (modified after Pepinelli
et al., 2006); 201: S. mariavulcanoae; 202: S. rappae; 203: S. subnigrum; 204: S. travassosi holotype.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.
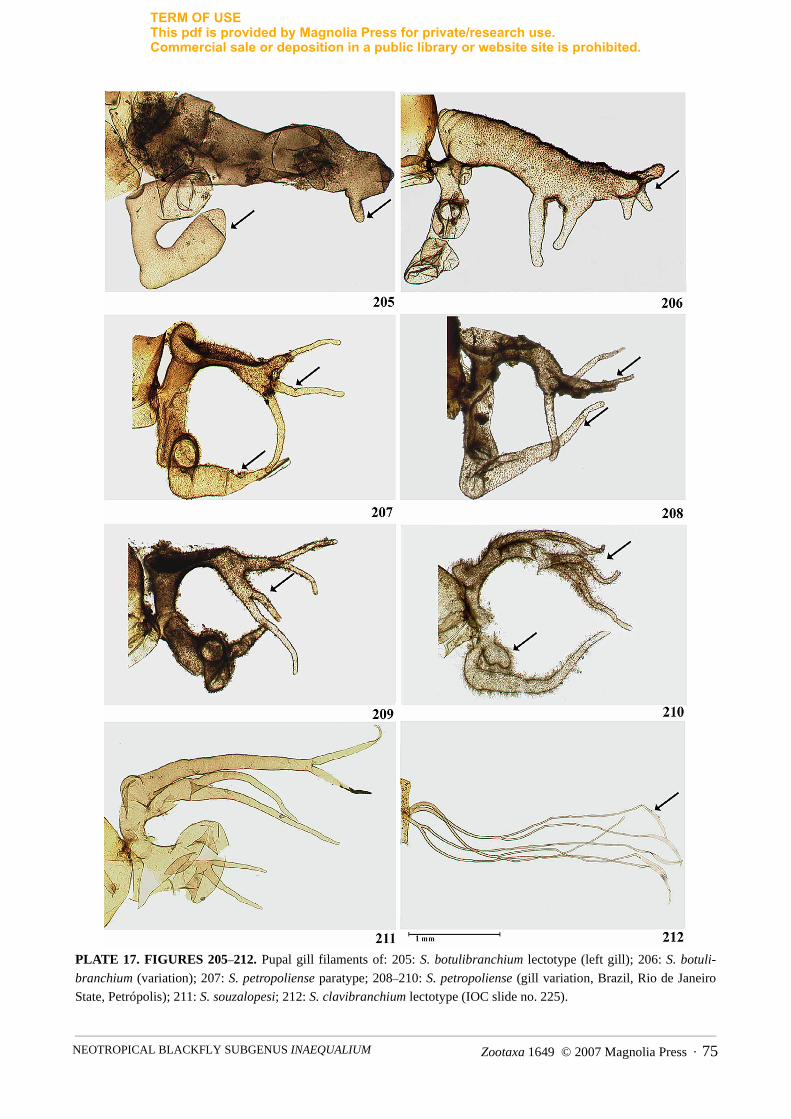
Zootaxa 1649 © 2007 Magnolia Press · 75NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
PLATE 17. FIGURES 205–212. Pupal gill filaments of: 205: S. botulibranchium lectotype (left gill); 206: S. botuli-
branchium (variation); 207: S. petropoliense paratype; 208–210: S. petropoliense (gill variation, Brazil, Rio de Janeiro
State, Petrópolis); 211: S. souzalopesi; 212: S. clavibranchium lectotype (IOC slide no. 225).
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.76 · Zootaxa 1649 © 2007 Magnolia Press
PLATE 18. FIGURES 213– 220. Pupal gill filaments of: 213: S. clavibranchium (gill variation, Brazil, São Paulo State,S. José do Barreiro, Nascente na Fazenda S. Sebastião da Cachoeira); 214: S. diversibranchium; 215: S. inaequale speci-men from Brazil, São Paulo State; 216: S. maranguapense (after Pessoa et al., 2005); 217: S. margaritatum; 218: S. mar-iavulcanoae; 219: S. rappae holotype; 220: S. rappae (common pattern of gill configuration). Arrows indicate differentends of the gill filaments. [Scale bar given for all figures except for figures 215 and 219].
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 77NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
PLATE 19. FIGURES 221–228. Pupal gill filaments of: S. rappae (gill variations)-: 221, 222: Brazil, Goiás State; 223:
Brazil, Goiás, Rio Itiquira, 224: Brazil, Rio de Janeiro State, Parque Nacional Itatiaia, Rio Campo Belo; 225, 226: Brazil,
Goiás state, Pirenópolis, Córrego Caiçara; 227: Brazil, Minas Gerais State, Fazenda Sobradinho; 228, Brazil, Goiás
State, Rio Saia Velha. Arrows indicate different branching, enlargement and end of the gill filaments.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.78 · Zootaxa 1649 © 2007 Magnolia Press
PLATE 20. FIGURES 229–234. Pupal gill filaments of: S. rappae (gill variations):- 229–234: Brazil, Goiás State, Rio
Saia Velha. Fig. 234 shows the configuration of the pupal gill filaments and anterior region of cocoon in spirit. Arrows
indicate different branching, enlargement and end of the gill filaments.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.
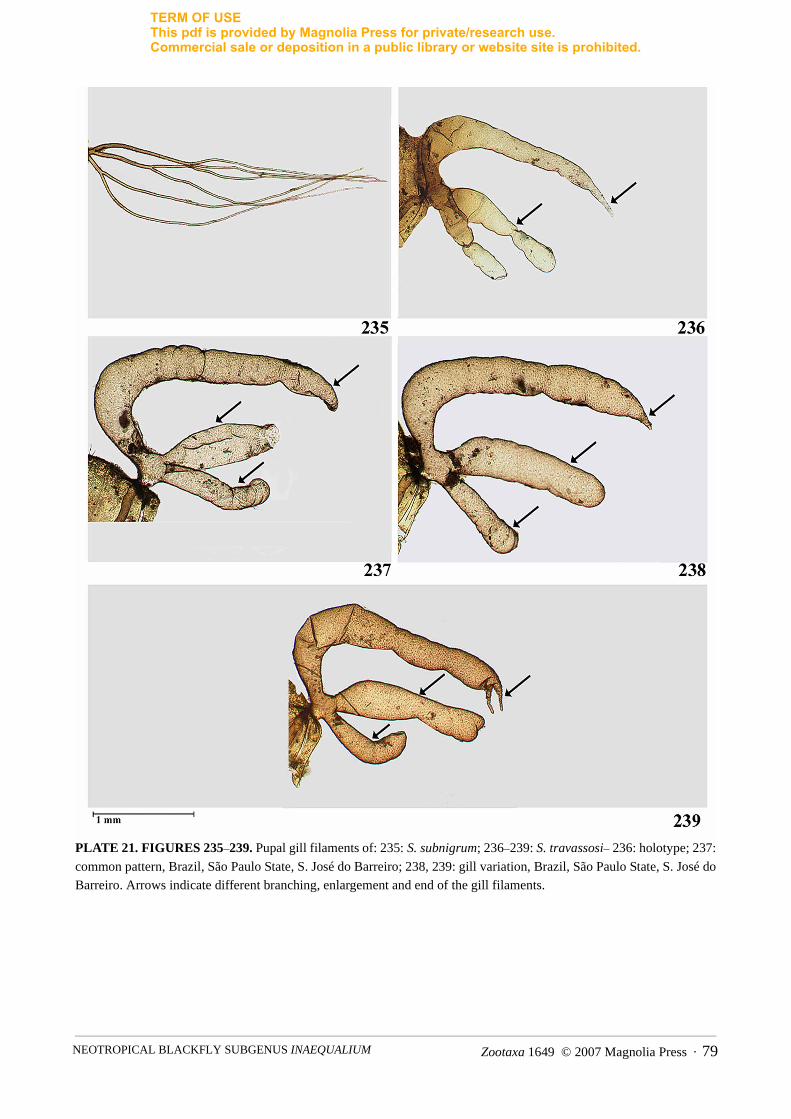
Zootaxa 1649 © 2007 Magnolia Press · 79NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
PLATE 21. FIGURES 235–239. Pupal gill filaments of: 235: S. subnigrum; 236–239: S. travassosi– 236: holotype; 237:
common pattern, Brazil, São Paulo State, S. José do Barreiro; 238, 239: gill variation, Brazil, São Paulo State, S. José do
Barreiro. Arrows indicate different branching, enlargement and end of the gill filaments.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.80 · Zootaxa 1649 © 2007 Magnolia Press
Material Examined
BOTULIBRANCHIUM species group
Simulium (Inaequalium) botulibranchium Lutz, 1910
TYPE MATERIAL
BRAZILRio de Janeiro StateSLIDE
[Without locality, date or collector’s name]—pupal exuviae [LECTOTYPE] (IOC, no. 12.038) [Theslide has several labels: White, handwritten label “ Simulium botulibranchium Lutz”; White “12.038, Bd103”. It also has a white label “ 312”. We have added other labels: A white, handwritten label by L.M.Hernández “Lectotype designated by D’Andretta & D’Andretta 1947:161” and “Photographed parts of gill,cocoon weave, frontoclypeus trichomes L.M. Hernández 2003”.] [The locality information “Rio de JaneiroState, Morro do Observatório, Petrópolis, collector A. Lutz, 1910” has been given by D’Andretta &D’Andretta, 1947” The date might be “4.xii.1910” based on IOC Simuliidae Catalogue.]
OTHER MATERIAL
BRAZILRio de Janeiro StatePINNED
Petrópolis, stream from mountain at km 85 towards Rio, (site 1687, D25), 22º31’S, 44º 14’W;18.iii.2004, (A.J. Shelley, A.P.A. Luna Dias & P.R. Garritano)—2&&%% (BMNH; IOC). Petrópolis;11.ix.1911, [Without collector’ name.]—2&& (on same pin), 1% (IOC, no. 12.476) [The specimens bear apurple label “Tipo”. We have added a white label by “Simulium botulibranchium Lutz? Det. L.M.Hernández”]. Rua de Rio de Janeiro to Jardim de Itaipava, BR040, first cachoeira after site 1694, (site 1695),22º27’29’S43º12’59’’W, 1017 m; 22.iii.2005, (L.M. Hernández & A.P.A. Luna Dias)—1% (reared) (IOC).Rua de Rio de Janeiro to Jardim de Itaipava, cachoeira in front of Posto Brassão, (site 1696a-b); 22.iii.2005,(L.M. Hernández & A.P.A. Luna Dias)—1% (reared) (IOC). Rua de Rio de Janeiro to Jardim de Itaipava,cachoeira in front of Posto Brassão, (site 1697, 5A1-2); 23.ii.2005, (L.M. Hernández & A.P.A. Luna Dias) –1& 1% (reared) (IOC). Petrópolis, stream from mountain at Km 85 towards Rio, (site 1687); 20.iii.2004, (A.J.Shelley, A.P.A. Luna Dias & P.R. Garritano)—1% (reared) IOC). Petrópolis; 11.ix.1911, [Without collector’sname.]—2&&1% (IOC, 12.0476) [The specimen bear a label “Cotipo” [=Cotype.] Petrópolis, Observatório;11.ix.1911, [Without collector’s name.]- 2&&1%, 4 pupal exuviae (IOC, 12.0477) [The specimen bears a label“Cotipo” [=Cotype.] Petrópolis; [Without date or collector’s name.]– 1& (IOC).
SLIDE[Without locality information, date or collector’s name but probably “Petrópolis”.]—1% (only genitalia
and one hind leg) (IOC, no. 12.037, Bd. 103) [The slide bears a handwritten label “ S. botulibranchium %Typo” and a printed, white label “311”. We have added other labels: A white label “Digitally photographedparts(s): hind leg, gonostyle + gonocoxite. Taken by L.M. Hernández BMNH 2003” and a white label“Simulium botulibranchium Lutz? Det. L.M. Hernández”.] Petrópolis, Morro do Observatório; 4.ii.1910,[Without collector’s name.]—1% (reared, 12.041), 2 pupal exuviae (12.042, 12.043) (IOC). Petrópolis, stream
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 81NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
from mountain at km 85 towards Rio, (site 1687, D25), 22º31’S, 44º 14’W; 18.iii.2004, (A.J. Shelley, A.P.A.Luna Dias & P.R. Garritano)—1&1% (reared), 1 pupal exuviae, 2 larvae (BMNH).
SPIRITPetrópolis, stream from mountain at km 85 towards Rio, (site 1687, D25), 22º31’S44º14’W;
18.iii.2004, (A.J. Shelley, A.P.A. Luna Dias & P.R. Garritano)—numerous pupal exuviae (BMNH; IOC). Ruade Rio de Janeiro to Jardim de Itaipava, cachoeira in front of Posto Brasão, (site 1696ab, coll. 4); 22.iii.2005,(L.M. Hernández & A.P.A. Luna Dias)—2 pupal exuviae (BMNH). Rua de Rio de Janeiro to Jardim deItaipava, cachoeira in front of Posto Brassão, (site 1697ab, coll. 5), 33°59’59.1’’N 68°54’19.0’’E, 870m;23.iii.2005, (L.M. Hernández & A.P.A. Luna Dias)—several pupal exuviae, several larvae (BMNH; IOC).
São Paulo StatePINNED
Nth. Serra da Bocaina, (loc 22); 15–18.v.1979, (R.W.Crosskey & A.J. Shelley)—1& (reared, onlythorax; head, wings, legs abdomen and genitalia on slide) (BMNH, BM 1979-258). S. José do Barreiro,Nascente na Faz. S. Sebastião da Cachoeira, (site 1675, D13), 22º41’S, 44º 30’W; 18.iii.2004, (A.J. Shelley,A.P.A. Luna Dias & P.R. Garritano)—1& (IOC). Fazenda Pau d’Alho; 22.vi.1947, (Marino & Pereira)—1%(reared) (AMNH) [Determined by S. Coscarón 79]. Km 52 on SP183, NW, Cruzeiro, Cascade, Serra daMantequeira, (site 462); 18.v.1979, (A.J. Shelley & A.P.A. Luna Dias)—2 && (reared, only thorax; head,wings, legs, abdomen and genitalia on slide) (BMNH, B.M. 1979-580). Obiti; 23.iv.1956, [Withoutcollector’s name]– 1&1% (reared, pupa in spirit) (BMNH; & no.1330, % no, 1331). Rio Pelões, Cubatão;7.iii.2003, (M. Pepinelli)—1& (not associated with pupa) (MZUSP)
SLIDENth. Serra da Bocaina, (loc 22); 15–18.v.1979, (R.W. Crosskey & A.J. Shelley)—1& (reared, only head,
wings, legs, abdomen and genitalia, thorax pinned) (BMNH, BM 1979-258). Km 52 on SP183, NW.Cruzeiro, Cascade, Serra da Mantequeira, (site 462); 18.v.1979, (A.J. Shelley & A.P.A. Luna Dias)—2&&(reared, only, wings, legs, abdomen and genitalia; thorax pinned) (BMNH, B.M. 1979-580).
SPIRITBoraceia; 30.x.1979, (Coscarón)—several pupal exuviae, larvae (BMNH). Obiti; 23.iv.1956, [Without
collector’s name.]– 2 pupal exuviae (&% adults pinned) (BMNH; & no.1330, % no. 1331). S. José do Barreiro,cachoeira going to Arapeí, (site 1673, D11); 17.iii.2004, (A.J. Shelley, A.P.A. Luna Dias & P.R. Garritano)—1 pupal exuviae (IOC). S. José do Barreiro, 3rd un-named stream after waterfall to Arapeí, (site 1672, D10),20º00’S47º48W; 3.viii. 2004, (A.J. Shelley, A.P.A. Luna Dias, P.R. Garritano)—several pupal exuviae(BMNH). S. José do Barreiro, stream at 500 m from site 1676, returning to Arapeí, (site 1677, D15);18.iii.2004, (A.J. Shelley, A.P.A. Luna Dias & P.R. Garritano)—1 pupal exuviae (BMNH).
Minas Gerais StatePINNED
Pedralva, Fzda. Sto Antonio; 15.x.1978, (A.J. Shelley & A.P.A. Luna Dias)—1&1% (reared) (IOC).
WITHOUT COUNTRY OR STATE [BUT PROBABLY BRAZIL]PINNED
[Without locality, date or collector’s name]– 1& (AMNH) [The specimen does not bear a locality label, but has a white printed label “156.29” which agrees with the numbering system used by M.A.V. D’Andretta (also known as M.A. Vulcano) in her studies of the Brazilian Simuliidae].
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.82 · Zootaxa 1649 © 2007 Magnolia Press
Simulium (Inaequalium) petropoliense Coscarón, 1980
All materiel listed in Hernández et al. (2006) was examined for this paper.
Simulium (Inaequalium) souzalopesi Coscarón, 1980
TYPE MATERIAL
BRAZILRio de Janeiro StatePINNED
Petrópolis; 12.v.1979, (Coscarón)—1& (reared, pupa glued to a card point) [HOLOTYPE] (MLP) [Thespecimen bears several labels: White label in Coscarón’s hand with locality information, date and collector’sname; White label “Simulium souzalopesi n. sp. Det. Coscarón; Red label “HOLOTIPO”. We have addedanother label: White label “Digitally photographed Thorax (ant.+post.) abdomen By L.M. Hernández”; Whitelabel “ Simulium souzalopesi Coscarón Determined L.M. Hernández”]. Same date as holotype—1&1%(reared) [PARATYPES, the male as ALLOTYPE; the pinned female has lost its pupal exuviae] (BMNH).
SLIDESame date as the holotype—1&1& (reared, but not associated with pupal exuviae), 1 larva, 3 pupal
exuviae (MLP) [PARATYPES, 2 pupal exuviae without label stating their type status, but we assumed theyare paratypes as all material collected by Coscarón was designated as such (Coscarón, 1980)].
SPIRIT
Same date as the holotype—several pupal exuviae and larvae (MLP) [PARATYPES, the speci-mens does not bear a label stating their type status, but we assumed they are paratypes as all materialcollected by Coscarón was designated as such (see Coscarón, 1980)].
OTHER MATERIAL
BRAZILRio de Janeiro StatePINNED
Rua Itaipava to Teresópolis, Br 040, cachoeira in front of Posto Brasão, (site 10),22º27’29’’S43º12’59’’W, 1017 m; 8.x.2005, (A.J. Shelley & A.P.A. Luna Dias)—1& (reared) (BMNH).
SPIRITRua de Rio de Janeiro to Jardim de Itaipava, cachoeira in front of Posto Brassão, 22º27’29’’S–
43º12’59’’W, 1017 m, (site 1697); 23.iii.2005, (L.M. Hernández & A.P.A. Luna Dias)—1 pupal exuviae(BMNH).
INAEQUALE species groupSimulium (Inaequalium) clavibranchium Lutz, 1910
TYPE MATERIAL
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 83NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
BRAZILRio de Janeiro StateSLIDES
Serra de Petrópolis; [Without date or collector’s name.]—pupal exuviae [LECTOTYPE] (IOC, no. 255)[The slide bears several labels: Handwritten label with locality data; handwritten label with black ink “12.053, Bd 104” and printed at the bottom of the label “Instituto Oswaldo Cruz”; handwritten label:“Simulium clavibranchium Lutz” [there is small sentence at the bottom of the label which is illegible]. Wehave added other labels: Round, with purple edge label “LECTOTYPE”; White label “Digitally photographedright gill, frontoclypeus No. 255 P-1 L.M. Hernandez”; White label “Simulium clavibranchium LutzDetermined A.J. Shelley & L.M. Hernández”.]. Morro do Observatório, Petrópolis; [Without date orcollector’s name.]—1 pharate & (not associated with pupal exuvia) [PARALECTOTYPE] (IOC, no. 255)[The specimen bear several labels: Handwritten label with locality data; White label “226”; Handwritten label“ 12.054, Bd 104” and printed “Instituto Oswaldo Cruz”; Handwritten label “Simulium clavibranchium” andin Portuguese “ex pupa, sem escamas [= from pupa, without scales.]”. We have added a rounded, purple label“PARALECTOTYPE”; White label “Simulium clavibranchium Lutz Determined A.J. Shelley & L.M.Hernández”]. [Without locality data, date or collector’s name.]—1 pupal exuviae, 2 pharate pupal exuviae[PARALECTOTYPE] (IOC, no. 227) [The pupal gill of these specimens re partially broken] [The specimensbear several label: Handwritten label “Simulium clavibranchium”; printed label “ 227”; handwritten label “12.055, BD 104”; a handwritten label in Portuguese “trichomas bif. e ram.” [= bifid and multibranchedtrichomes”.] We have added other labels: Rounded, with blue edges label “PARALECTOTYPE”; White label“Simulium clavibranchium Lutz Determined A.J. Shelley & L.M. Hernández”.
OTHER MATERIAL
Minas Gerais StatePINNED
Pedralva, Fzda. Sto Antonio, (site 442-1); 25.iii.1979, (A.P.A. Luna Dias)—2&& (reared) (IOC).Pedralva, Fzda Sto Antonio, (site 442-1); 30.iii.1981, (A.P.A. Luna Dias—1& (reared) (IOC). Pedralva, FzdaSto Antonio, canal entre canal de turbine e córrego, (site 484-13); 13.x.1979, (A.J. Shelley & A.P.A. LunaDias)—1&1% (reared) (IOC).
Rio Grande do Sul StateSPIRIT
Maquine; 16.iv.1993, (Strieder, M.N.)—2 pupal exuviae (LEU).
Rio de Janeiro StatePINNED
Petrópolis; 1.xii.1910, [Without collector’s name.]—2&& (IOC, no. 479, 481) [Both specimens have apurple label “Tipo”.] [Identified with doubt as S. clavibranchium by L.M. Hernández.]. Petrópolis; 7.iii.1910,[Without collector’s name]—1& (reared) (IOC) [Identified with doubt as S. clavibranchium by L.M.Hernández.] Km 4 Br. 354, (site 477); 22.v.1979, (A.J. Shelley & A.P.A. Luna Dias)—1& (reared) (IOC). P.N.Serra dos Órgãos, Subsede Teresópolis, (site 586); 29.iii.1982, (R. Malaguti)—1& (reared) IOC). Represa dosciganos; 24.v.1984, (R. Malaguti & R.R. Figueredo)—2&& (reared) (BMNH).
SLIDES
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.84 · Zootaxa 1649 © 2007 Magnolia Press
Petrópolis; [Without locality data, date or collector’s name.]—1 % (only abdomen and genitalia) (IOC,no. 12.435, Bd 123).
São Paulo StatePINNED
S. José do Barreiro, Posto do IBDF, Rio Mambucaba, (site 333); 26.iii.1978, (A.J. Shelley & A.P.A.Luna Dias)—3&&1% (reared) (IOC). Km 52, Br 459 rd (462-8); 18.v.1979, (A.J. Shelley & A.P.A. LunaDias)—1% (reared) (IOC) [Identified by Regina Malaguti]. S. José do Barreiro, Fazenda do Bonito, RioMambucaba, (site 340); 4.iv.1978, (A.J. Shelley & A.P.A. Luna Dias)—1& (reared) (IOC). S. José doBarreiro, Fazenda do Bonito, Córrego do Segredinho, (site 344); 5.iv.1978, (A.J. Shelley & A.P.A. LunaDias)—1&1% (reared) (IOC). Serra da Bocaina, Fazenda de Bonito, (site 347-38); 6.iv.1978, (A.J. Shelley &A.P.A. Luna Dias)—3&& (reared). S. José do Barreiro, stream at 200 m do site 1679, (site 1680, D18),22º43’S44º 32’W; 18.iii.2004, (A.J. Shelley, A.P.A. Luna Dias & P.R. Garritano)—1 & (reared) (BMNH). S.José do Barreiro, small stream at 300m do site 1678, (site 1679, D17), 22º43’S44º 32’W; 18.iii.2004, (A.J.Shelley, A.P.A. Luna Dias & P.R. Garritano)—1& (reared) (BMNH). S. José do Barreiro, stream at 400mfrom site 1684, (site 1685, D23); 18.iii.2004, (A.J. Shelley, A.P.A. Luna Dias & P.R. Garritano)—1& (reared)(BMNH). S. José do Barreiro, un-named stream, at 200 m from Córrego at site 1683, (site 1684, D22),22º46’S 44º 31’W; 18.iii.2004, (A.J. Shelley, A.P.A. Luna Dias & P.R. Garritano)—1% (reared) (BMNH). S.José do Barreiro, stream at 400 m from site 1684, (site 1685, D23); 18.iii.2004, (A.J. Shelley, A.P.A. LunaDias & P.R. Garritano)—2&&1% (IOC) (BMNH). C. Bocaina, P.E.Intervales; 1.ii.2002, (Pepinelli, M.)—1&(reared, only thorax, wings and legs; head, genitalia and pupa on slide) (MZUSP, no. C 100). C.Barranquinho, P.E.Intervales; 2.ii.2002, (Pepinelli, M.)—1& (not associated with pupal exuviae) (MZUSP).Cachoeira da Usina, São José do Barreiro; 16.v.2003, (Pepinelli, M.)—1& (MZUSP). Fazenda Canchim, SãoCarlos; 2.iv.2002, (Pepinelli, M.)—1% (reared) (MZUSP). Km 57 on SP183, NW Cruzeiro Cascade da SerraMantequeira, (site 461, 462); 28.v.1979, (A.J. Shelley)—2&&1% (reared) (BMNH). Serra da Bocaina, RioMambucaba, Posto 18DF, (site 333-9); 26.iii.1978, (A.J. Shelley)—1& (reared) (BMNH). Capivari, Est. E.Ribas; 10-20.i.1952, (M.A. Vulcano)—1&(reared) (BMNH). Boracéia, Rio Claro; 25.vii.1979, (Coscarón)—1% (reared) (BMNH). S. José do Barreiro, small stream at 300m do site 1678, (site 1679, D17), 22º43’S44º32’W; 18.iii.2004, (A.J. Shelley, A.P.A. Luna Dias & P.R. Garritano)—1& (reared) (BMNH). José doBarreiro, un-named stream, at 300m from córrego at site 1680, (site 1681, D19), 22º43’S, 44º 32’W;18.iii.2004, (A.J. Shelley, A.P.A. Luna Dias & P.R. Garritano)—1& (reared, BMNH). S. José Barreiro,Cachoeira na Faz. S. Sebastião da Cachoeira, (site 1676, D14), 22º41’S, 44º 30’W; 18.iii.2004, (A.J. Shelley,A.P.A. Luna Dias & P.R. Garritano)—1&2%% (BMNH; IOC). S. José do Barreiro, un-named stream, at 300mfrom córrego at site 1680, (site 1681, D19), 22º43’S, 44º 32’W; 18.iii.2004, (A.J. Shelley, A.P.A. Luna Dias &P.R. Garritano)—1% (IOC). F. de Bonito, S. de Bocaina; i.1915, [Without collector’s name.]—1 pupalexuviae (IOC, no. 12.05[illegible], Bd 104].
SLIDEP.E. Cubatão, C. do Jeep; 25.ii.2002, (M. Pepinelli)—1% (reared) (MZUSP). C. Bocaina,
P.E.Intervales; 1.ii.2002, (Pepinelli, M.)—1& (reared, only head, genitalia and pupa; thorax, wings and legspinned) (MZUSP, no. C. 100). S. José do Barreiro, un-named stream, at 200 m from Córrego at site 1683, (site1684, D22), 22º46’S 44º 31’W; 18.iii.2004, (A.J. Shelley, A.P.A. Luna Dias & P.R. Garritano)—1% (reared)(BMNH). S. José do Barreiro, stream at 500 m do site 1682, (site 1683, D21), 22º46’S44º 31’W; 18.iii.2004,(A.J. Shelley, A.P.A. Luna Dias & P.R. Garritano)—1& (reared) (BMNH).
SPIRIT
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 85NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
S. José do Barreiro, Posto do IBDF, Rio Mambucaba, (site 333); 26.iii.1978, (A.J. Shelley & A.P.A.Luna Dias)—2 %% (reared) (IOC). Serra da Bocaina, Fazenda Bonito, (site 344-7); 5.iv.1978, (A.J. Shelley)—1& (reared) (BMNH). Boracéia, Rio Claro; 25.vii.1979, (Coscarón)—2 pupal exuviae (BMNH) [Identified asS. subclavibranchium.]
Rio Grande do Sul StateSPIRIT
Maquiné; 16.iv.1993, (Strieder, M.N.)—2 pupal exuviae (LEU) [Previously identified as S.subclavibranchium.]
Without State or locality [but most likely Brazil]SLIDE
[Without full locality data or date.]; (A. Lutz)—1 pupal exuviae (AMNH) [The slide has a handwrittenlabel “DS1”.]. [Without full locality data or date.]—1 pupal exuviae (IOC, 12.061, Bd. 105; no. 233), 1 & (notassociated with pupal exuviae) (IOC, 12.060, Bd 104).
Simulium (Inaequalium) diversibranchium Lutz, 1910
TYPE MATERIAL
BRAZILSão Paulo StateSLIDE
Bonito, Serra da Bocaina; [Without date or collector’s name.]—pupal exuviae [LECTOTYPE] (IOC,no. 12.097) [The specimen bears a label “ Remounted por Regina Malaguti” and “N. 214”. We have added awhite, handwritten label by L.M. Hernández “S. diversibranchium Det. A.P.A. Luna Dias & L.M.Hernández”.]. Serra da Bocaina; [Without date or collector’s name.]—1 pupal exuviae [PARALECTOTYPE](IOC, no. 12.095)] [The specimen bears an extra label “N. 211”. We have added a white, handwritten label byL.M. Hernández “S. diversibranchium Det. A.P.A. Luna Dias & L.M. Hernández”.] Serra da Bocaina;[Without date or collector’s name.]—2 pupal exuviae [PARALECTOTYPES] (IOC, no. 12.098) [Thespecimen bears an extra label “N. 307”. We have added a white, handwritten label by L.M. Hernández “S.diversibranchium Det. A.P.A. Luna Dias & L.M. Hernández”.] [The slide has three pupal exuviae, onecorresponding to S. subnigrum. We have added an extra label “one pupal exuviae of S. subnigrum” and “twopupal exuviae of S. diversibranchium” Lutz Det. A.P.A. Luna Dias & L.M. Hernández.]
As Simulium missionum Coscarón, 1976
ARGENTINAPINNED
Missiones, Campiña, 5 km S. de Irigoyen; 24.vii.1972, (Coscarón)—1% (reared; only head, thorax,both wings and three legs of the left side; three legs of the right side, genitalia and pupa on slide)[HOLOTYPE] (MLP) [The specimen bear several labels: White label, hand-written in black ink with localityinformation, date and collector’s name; White label in Coscarón’s hand “Simulium missionum n.sp. det.Coscarón 76 Holotipo”; White label “1”, which is encircled; Red label without further data; White,handwritten, black-bordered label “ Simulium diversibranchium det. Coscarón 1979”. We have added otherlabels: White label “Digitally photographed Th. (ant.+post.) %-1 L.M. Hernández; White label “Simuliummissionum Coscarón, 1976 syn. of S. diversibranchium Lutz Determined L.M. Hernández.]
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.86 · Zootaxa 1649 © 2007 Magnolia Press
SLIDESame data as holotype with new labels added—1% (reared; only three legs of the right-hand side,
genitalia and pupa; head, thorax, both wings and three legs of the left side pinned) [HOLOTYPE] (MLP).Same data as the holotype—1 pupal exuviae [PARATYPE] (MLP).
OTHER MATERIAL
BRAZILRio Grande do Sul StateSPIRIT
Bom Jesus; 2.ix.1984, (V. Machado & S. Member)—2 pupal exuviae, 2 larvae (UNISINOS, M. 180).
São Paulo StatePINNEDS. José do Barreiro, 3rd un-named stream after waterfall to Arapeí, (site 1672, D10), 20º00’S47º48W;
3.viii. 2004, (A.J. Shelley, A.P.A. Luna Dias, P.R. Garritano)—1& (reared, pupa on slide) (BMNH). CasaGrande; 3.x.1979, (Coscarón)—2&& (reared) (MLP) [Identified as S. diversibranchium by Coscarón 1979].Capivari, Est.Emilio Ribas; 10-20.1.1952, (Vulcano)—1& (reared but not associated with pupa) (MLP, no,148-1) [The specimens bear two labels: 1- White, handwritten “missionum %-1”; White, handwritten in black-ink “S. diversibranchium by Coscarón 1979”]. Casa Grande; 25.viii.1979, (Coscarón)—only one pupalexuviae (adult lost) (MLP) [Determined as S. diversibranchium by Coscarón 1979.]
The following pinned material does not have the locality information, date or date or collector’s name onthe labels, but they bear the numbers used by Vulcano in her catalogue and they are most likely to have beencollected from São Paulo, Brazil (S. Coscarón, per. com):
[Without locality information, date or collector’s name]—5&&1% (reared, pupal exuviae on slide)(MLP, nos. 151-11, 21, 23, 30-32) [All determined as S. diversibranchium by Coscarón 1979.][Withoutlocality information, date or collector’s name.]—2&&3%% (reared, pupal exuviae on slide) (MLP, nos. 151-18,41, 47, 51) [Two specimens bears determination label as “S. missionum” and “S. diversibranchium” byCoscarón 1979.]
SLIDES. José do Barreiro, 3rd un-named stream after waterfall to Arapeí, (site 1672, D10), 20º00’S47º48W;
3.viii. 2004, (A.J. Shelley, A.P.A. Luna Dias,.P.R. Garritano)—1& (only pupal exuviae, adult pinned)(BMNH). S. José do Barreiro, 3rd un-named stream after waterfall to Arapeí, (site 1672, D10),20º00’S47º48W; 3.viii. 2004, (A.J. Shelley, A.P.A. Luna Dias, P.R. Garritano)—1& (reared), 2 pupal exuviae(BMNH). Serra da Bocaina, Bonito, (no. 12-512); [Without date or collector’s name.]—1% (only genitalia,adult pinned), 1& (MLP) [Identified as S. diversibranchium by Coscarón 1979.]
The following material on slide does not have the locality information, date or date or collector’s name onthe labels, but they bear the numbers used by Vulcano in her catalogue and they are most likely to have beencollected from São Paulo, Brazil (S. Coscarón, per. com):
[Without locality information, date or collector’s name]–6 pupal exuviae (adults pinned) (MLP, nos.151-11, 21, 23, 30-32) [All determined as S. diversibranchium by Coscarón 1979.] [Without localityinformation, date or collector’s name.]– 6 pupal exuviae (MLP, nos. 151-14, 29-8, 38, 36, 54) [All specimensbears determination label as “S. diversibranchium” by Coscarón 1979]. [Without locality information, date or
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 87NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
collector’s name.]– 2%% (only pupae, adult pinned) (MLP, nos. 151-24, 36) [All specimens bearsdetermination label as “S. diversibranchium” by Coscarón 1979][Without locality information, date orcollector’s name.]– 5&& (reared but not associated with pupal exuviae), 1 & (reared) (MLP, nos. 151-36, 38,52, 54, 141-30) [All specimens with blue bordered labels handwritten by Coscarón “S. diversibranchium det.Coscarón 1979”.]
SPIRITBoraceia; 3.x.1979, (Coscarón)—several pupal exuviae and larvae (MLP) [All determined as S.
diversibranchium by Coscarón 1979.] Casa Grande; 25.viii.1979, (Coscarón)—several pupal exuviae andlarvae (MLP) [All determined as S. diversibranchium by Coscarón 1979.]
Simulium (Inaequalium) inaequale (Peterson & Shannon, 1927)
All the material listed under S. inaequale in Hernández et al. (2007) was examined in this paper. Further material examined is as follows:
BRAZILSao Paulo StateBr 139, km 18, Hotel Fazenda Boa Vista; 15.v.1979, (R.W.Crosskey & A.J. Shelley)—3&&1% (reared)
(BMNH).
SLIDEBr 139, km 18, Hotel Fazenda Boa Vista; 15.v.1979, (R.W.Crosskey & A.J. Shelley)—2 %% (reared)
(BMNH, B.M. 1979-258).
Simulium (Inaequalium) lurybayae Smart, 1944
TYPE MATERIAL
As Simulium lurybayae Smart, 1944 [new name for Trichodagmia angustifrons Enderlein, 1934]
BOLIVIAPINNED
La Paz, Luribay; iii.1913, (Stocker, S.G.)—1& (only thorax; head, wings, legs, abdomen and genitaliaon slide) [HOLOTYPE] (NMUH) [The specimen bears several labels: Green label with locality and collectorinformation; White label handwritten by Enderlein “Trichodagmia angustifrons Type Ender. &”, printed andthe bottom of the label “Dr. Enderlein det. 1933; Orange label “Typus. We have other labels: White label“Digitally photographed parts Thorax ant.+ post.; abdomen; lateral view &-1. Taken by L.M. HernándezBMNH 2004”; White label “Material in slide collection &-Holotype L.M. Hernández 2004”; White label “Simulium lurybayae (End.) Holotype of S. angustifrons End., 1934. Examined by L.M. Hernández 2004”].
SLIDEData as the pinned adult with new labels added—1& (only head, wings, legs, abdomen and genitalia;
thorax pinned) [HOLOTYPE] (NMUH).
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.88 · Zootaxa 1649 © 2007 Magnolia Press
Simulium (Inaequalium) margaritatum Pepinelli, Hamada & Luz
BRAZILGoiás StatePINNED
Córrego sem nome, Est. GO 18 (Brasília-Campos Belos), km 228, (site 183); 22.iv.1976, (A.J.Shelley)—1& (reared, only thorax; head, wing, two legs, abdomen and genitalia on slide) (IOC) [Thespecimen bear several labels: White label with locality and collector’s name, IOC no. 183”; White labellocality information, date and collector’s name and IOC no. 183I. We have added the following labels: Whitelabel “Digitally photographed before dissection L.M. Hernández 2005 &-1”; White label “Pupa on slide &-183”.]
SLIDESame data as pinned specimen—1& (reared, only head, wing, two legs, abdomen and genitalia; thorax
pinned) (IOC). Córrego sem nome, Est. GO 18 (Brasília-Campos Belos), km 228, (site 183); 22.iv.1976, (A.J.Shelley)—1% (pharate) (BMNH).
SPIRITCórrego sem nome, Est. GO 18 (Brasília-Campos Belos), km 228, (site 183); 22.iv.1976, (A.J. Shelley)—
1% (pharate) (BMNH).
Simulium (Inaequalium) mariavulcanoae Coscarón & Wygodzinsky, 1984
TYPE MATERIAL
BRAZILSão Paulo StatePINNEDE.B. Boracéia, Salésopolis; 13.x.1947, (MAV D’Andretta)—1& (only thorax, legs and wings; genitalia and
cibarium on slide) [HOLOTYPE, 115-212] (MZSP) [The legs of the holotype were damaged during dissec-tion; wings and legs were glued to the point card; remainder of head and abdomen are in a vial with glycerineattached to the specimen] [The specimen bears a white, handwritten label by S. Coscarón “ Simulium vulca-noae n.sp. det. Coscarón 79”.]
SLIDESame date as the pinned adult with new labels added—1& (only cibarium and genitalia; remainder pinned)
[HOLOTYPE, 115-212] (MZSP).
OTHER MATERIAL
BRAZIL[Some of the specimens does not have a label staying the country, state or locality, but they must have been
collected in São Paulo State, Brazil as all the material bear a label with D’Andretta (as well as Vulcano)catalogue’s number (Sixto Coscarón, pers. com.]
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 89NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
São Paulo StatePINNED
E.B.Boraceia, Salesópolis; 13.x.1947, (D’Andretta, M.A.V.)—2&& (reared, but one & not associatedwith pupal exuviae; the other & with no. 115-232 has the pupal exuviae on a slide) (MLP). [Without locality,date or collector’s name]—1& (reared, pupal exuviae on slide) (MLP, no. 141-16).
SLIDEE.B.Boraceia, Salesópolis; 13.x.1947, (D’Andretta, M.A.V.)—1 pupal exuviae (adult pinned) (MLP,
no. 112-232) [The slide have a white, label in Coscarón’ hands “Simulium mariavulcanoae Coscarón Det.Coscarón 1979”.]. Salesópolis (Boraceia); 13.x.1947, (L. Travassos Filho)—1 pupal exuviae (MLP, no. 115-16) [The slides have a white, label in Coscarón’ hands “Simulium mariavulcanoae n. sp.”] [Without locality,date or collector’s name]—1& (reared, adult pinned) (MLP, no. 141-16). [Without locality, date or collector’sname.]—1% (reared) (MLP, no. 115-263) [The specimen is in two slides, one slide have a handwritten labelwith blue borders “ Simulium mariavulcanoae Coscarón det. Coscarón 1979 % n.sp.] [Without locality, dateor collector’s name.]—1& (reared but not associated with pupa) (MLP, no.151-14)]. [Without locality, date orcollector’s name.]—2 pupal exuviae (MLP, no. 141-21) [The two slides have a determination label inCoscarón’s hands Simulium mariavulcanoae Coscarón det. Coscarón 1979)]. [Without locality, date orcollector’s name.]—5 pupal exuviae (MLP, nos. 115-263, 136, 141-30, 148-24, 151-47) [The slides havehandwritten determination labels “Simulium vulcanoae Coscarón det. Coscarón 1979” and “Simuliummariavulcanoae Coscarón, 1979”].
Simulium (Inaequalium) rappae Py-Daniel & Coscarón, 1981
TYPE MATERIAL
BRAZILMinas Gerais StatePINNED
[Without locality information, date or collector’s name.]—1& (reared) (INPA, no. 5513) [Thespecimens does not a label with locality information, but it has a label with no. 5515-13 that agree with thenumbering system given in the original description by Py-Daniel & Coscarón (1980) “BRAZIL: Minas GeraisState, Bocaina de Minas, Estrada entre Repouso da Gávea e a Pedra da Gávea, Córrego do Concreto, 22º19’S44º34’W; 12.i.1980, (V. Py-Daniel & J. Carneiro de Vasconcelos Neto)”.] [The specimen bear several labels:1- White label “ 194-33”; 2- Red, handwritten label “ HOLOTIPO S. rappae Py-Daniel & Coscarón”; 3-White printed label “5515-13”. We have added another label: 6- White label “Digitally photographed by L.M.Hernández 2005”]. Same data as holotype—1% (reared) [PARATYPE, As ALLOTYPE] (INPA, no. 194-25,5515-9) [The specimen bear several labels: 1- White label “ 194-25”; 2- Red, handwritten label “ ALLOTIPOS. rappae Py-Daniel& Coscarón”; 3- White printed label “5515-19”.]
OTHER MATERIAL
BRAZILFederal District
PINNED
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.90 · Zootaxa 1649 © 2007 Magnolia Press
Est. de Brasilinha, (site 357k); 28.vi.1979, (A.J. Shelley)—1& (reared) (BMNH, B.M. 1979-580).Córrego Dois Irmãos, (site 247); 14.vi.1976 (A.J.Shelley)—1& (IOC). Rio Palma, 11Km DF5, (site 250);21.vi.1976, (A.J. Shelley)—1 % (IOC).
The following specimen from Federal District has been identified as an extreme variation of S. rappae:
SLIDERio Palma, (site 250); 21.vi.1976, (A.J. Shelley)—1 pupal exuviae (BMNH) [Identified as “Variation of
S. rappae Py-Daniel, L.M. Hernández & A.J. Shelley”.]
Goiás StatePINNED
Pirenópolis, Fazenda Caiçara, Córrego Caiçara; 12.ix.1993, (D. Eaton)—1& (reared, pupal exuviae onslide) (BMNH). R. Itiquira, (site 106C); 29.v.1975, (A.J. Shelley)—1& (reared) (BMNH). Formosa, Salto doItiquira, Cachoeira 34 km from Formosa, (site 1094); 6.v.1994, (A.P.A. Luna Dias & P.R. Garritano)—2&&1% BMNH). Formosa, Salto do Itiquira, Cachoeira 34 km from Formosa, (site 1094); 6.v.1994, (A.P.A.Luna Dias & P.R. Garritano)—2&& ((reared, but not associated with pupal exuviae), 2 pupal exuviae (IOC).Parque Municipal de Itiquira, Salto do Itiquira, R. Itiquira, (site 961); 9.vi.1994, (A.J. Shelley, M.M. Herzog &A.P.A. Luna Dias)—4&& (reared) (IOC). Rio Itiquira, 50m abaixo de Salto Grande, (site 106), 29.v.1975,(A.J. Shelley)—2 &&1% (reared) IOC). Formosa, Parque Municipal do Itiquira Rio Itiquira, (site 961);9.vi.1994, (A.J. Shelley, M. Maia-Herzog, A.P.A. Luna Dias & E. Mokrabe)—4&& (reared) (IOC). FormosaParque Municipal do Itiquira, Rio Itiquira, (site 1638); 2.vi.2003, (A.P.A. Luna Dias & M.C. Alves)—13&&10%% (reared) (IOC).
SLIDER. Itiquira, (site 106C); 29.v.1975, (A.J. Shelley)—3 &&1% (reared), 5%% (reared, but not associated
with pupal exuviae), 4 pupal exuviae (BMNH). Parque Municipal de Itiquira, Salto do Itiquira, R. Itiquira,(site 961); 9.vi.1994, (A.J. Shelley, M.M. Herzog & A.P.A. Luna Dias)—1% (reared) (BMNH), 2 && (reared),2 pupal exuviae (IOC). Formosa, Salto do Itiquira, Cachoeira 34 km from Formosa, (site 1094); 6.v.1994,(A.P.A. Luna Dias & P.R. Garritano)—2 &&3%% (reared) (BMNH), 1& (reared), 2 && (reared, but notassociated with pupal exuviae), 3 pupal exuviae (IOC). Salto do Itiquira, (site 1138); 7.vii.1996, (A.P.A. LunaDias & P.R. Garritano)—1& (reared) (IOC). Pirenópolis, Fazenda Caiçara, Córrego Caiçara; 12.ix.1993, (D.Eaton)—1 pupal exuviae [adult pinned] (BMNH).
SPIRITBelém-Brasilia rd., km. 35, Rio Padre Souza, (site 207); 26.v.1976, (A.J. Shelley)—2&&1% (reared)
(BMNH, B.M.1979-580). Rio Saia Velha, (site 151 E); 13.i.1976, (A.J. Shelley)—1 pupal exuviae (BMNH).
The following specimens from Goiás State has been identified as a extreme variation of S. rappae:
PINNEDRio Saia Velha, (site 151); 13.i.1976, (A.J. Shelley)—1& (reared, only thorax; head, one wing, two
partially broken hind legs, abdomen, genitalia and pupal exuviae on slide) (IOC) [Identified as “Variation ofS. rappae by L.M. Hernández & A.J. Shelley”]. Rio Saia Velha, (site 152); 13.i.1976, (A.J. Shelley)—1&(reared, only thorax; head, one wing and pupal exuviae on slide) (IOC) [Identified as “Variation of S. rappaeby L.M. Hernández & A.J. Shelley”]. Rio Saia Velha, (site 151); 13.i.1976, (A.J. Shelley)—1 pupal exuviae
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 91NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
(in microvial with glycerine; adult lost) (IOC) [Identified as “Variation of S. rappae by L.M. Hernández &A.J. Shelley”].
SLIDESame data as pinned specimen—1& (reared, only head, wing, two partially broken hind legs, abdomen,
genitalia and pupal exuviae; thorax pinned) (IOC) [Identified as “Variation of S. rappae by L.M. Hernández& A.J. Shelley”]. Rio Saia Velha, (site 152); 13.i.1976, (A.J. Shelley)—1& (reared, only head, one wing andpupal exuviae; thorax pinned), 1 pupal exuviae (IOC) [Identified as “Variation of S. rappae by L.M.Hernández & A.J. Shelley”]. Rio Saia Velha, (site 151); 13.i.1976, (A.J. Shelley)—2 pupal exuviae (BMNH).Rio Saia Velha, (site 150); 13.i.1976, (A.J. Shelley)—3 pupal exuviae (BMNH).
Minas Gerais StatePINNED
Km 36-BR 354 rd, Faz Sobradinho, (site 472-14); 22.v.1970, (A.J. Shelley & A.P.A. Luna Dias)—1%(reared, pupal exuviae on slide)
SLIDEKm 36-BR 354 rd, Faz Sobradinho, (site 472-14); 22.v.1970, (A.J. Shelley & A.P.A. Luna Dias)—1%
(only pupal exuviae, adult pinned).
Rio de Janeiro State
PINNEDParque Nacional de Itatiaia, abaixo do Sanatório Militar, Rio Campo Belo, (site 483); 23.v.1979, (A.J.
Shelley & A.P.A. Luna Dias)—1& (reared, only thorax, remainder of female and pupal exuviae), 1&1%(reared, pupal exuviae on) (BMNH).
SLIDE
Parque Nacional de Itatiaia, abaixo do Sanatório Militar, Rio Campo Belo, (site 483); 23.v.1979, (A.J.Shelley & A.P.A. Luna Dias)—1 pupal exuviae, 1& (only abdomen, head, wings & legs) [remainder of femalepinned]; 1&1% (only pupal exuviae) [adult pinned] (BMNH).
São Paulo StateSLIDE
Boraceia, Rio Claro; 25.vii.1979, (Coscarón)—1 pupal exuviae (MLP).
SPIRITSão José do Barreiro, próx. A represa; 30.viii.1978, (Coscarón)—1 pupal exuviae, 1 larva (MLP). Santa
Rosa da Jiteiba, Rio Pardo; 5.[Illegible.].1985, (Coscarón)— 1 pupal exuviae (MLP).
Simulium (Inaequalium) subnigrum Lutz, 1910
All material listed in Hernández et al. (2007) was examined in this paper. Further material examined is asfollows:
BRAZILRio de Janeiro State
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.92 · Zootaxa 1649 © 2007 Magnolia Press
PINNEDRep. Dos Ciganos, (site 18.22B); 11.xii.1984, (M.M. Herzog & M. de Castro)—1% (reared; only head,
wings, legs; abdomen, genitalia, and pupal exuviae on slide) (BMNH). Rep. Dos Ciganos, (site 18.18B);11.xii.1984, (M.M. Herzog & Marua de Castro)—4&& (BMNH), 2 && (IOC). Rep. Dos Ciganos, (site 8.44/A); 11.xii.1984, (M.M. Herzog & Marua de Castro)—2&& (reared) (BMNH). Rep. Dos Ciganos, (site10.50.A, 10.37A, 10.62B, 10.57), 24.vii.1984, (R. Malaguti & A.R. Figueredo)—2&&1% (reared) (BMNH),1& (reared). Rep. Dos Ciganos, (site 16.2A), 19.xi.1984, (R. Malaguti & A.R. Figueredo)—1& (reared)(BMNH). P.N. Tijuca, Rep. Dos Ciganos, (site 623-5, 6); 21.viii.1982, (T.C. Carvalho & T.C.C. Leite)—2 &&(reared) (BMNH). Rep. Dos Ciganos, (site 11.23); 13.viii.1984, (A.J. Shelley & M.M. Herzog)—1% (reared,only head, wings, and legs; remaining of adult on slide) (BMNH).
SLIDEIBOF- Rep. Dos Ciganos; 10.vii., 24.vii.1984, (M.M. Herzog & A.R. Figueroa)–1&1m (reared) (BMNH).
Rep. Dos Ciganos; 11.xii.1984, (M.M. Herzog & Marcia de Costa)–1& (reared) (BMNH). Rep. Dos Ciganos;2.i.1985, (M.M. Herzog & Garritano)–1& (reared). Rep. Dos Ciganos, (site 11.23); 13.viii.1984, (A.J. Shelley& M.M. Herzog)—1% (reared) (BMNH). Rep. Dos Ciganos, (site 18.22B); 11.xii.1984, (M.M. Herzog &Marua de Castro)—1% (reared, only pupa, adult abdomen and genitalia; remaining of adult pinned) (BMNH).
Sao Paulo StatePINNEDSerra de Bocaina, Fazenda Bonito; 6.iv.1978, (A.J. Shelley & A.P.A. Luna Dias)–1& (reared) (BMNH).
PARAGUAYSLIDESMbaracayu, Conndiyae, arroyo en Balsinaticuí, 2850 m east from Horquetami; 20.vii.1996, (C. Nauzille &
L.Lagacne)– 3%%, 2 pupal exuviae (BMNH).
Simulium (Inaequalium) travassosi D’Andretta & D’Andretta, 1947
TYPE MATERIAL
BRAZILSão Paulo StatePINNED
São Vicente; 28.i.1947, (Andre. & Andre.)—1&1%(reared, but pupa in alcohol) [PARATYPES, & no.1.175, % 1.176] (BMNH, B.M. 1952-234) [The female does not have a locality label, but the pupa, which is inalcohol, bear full locality data.]
SLIDE[Without locality, date or collector]—% (reared) [HOLOTYPE, C.89.20] (MZSP) [The holotype is in
three slides bearing different labels: Slide 1- White label, “Holotipo (em parte), a label with modern black ink“1098” and on the slide “c.89.20 1.098”. This slide only contains the head and mouth parts; Slide 2- Labels asSlide 1 except “C.89.20” on adhesive label and not in black ink. The slide only contains the wings, legs,gonostyle and coxites, paramere and median sclerite, both wings with setae and spines broken off, butshowing the insertion spots; Slide 3- White label “1098” and written on the slide in ink “C.89.20”; theremainder of the information is illegible. The slide only containing complete pupal skin and exuviae dissectedout possibly from the male holotype, but it is not labeled as such.]
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 93NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
SPIRITSao Vicente; 28.i.1947, (Andre. & Andre.)—1&1% (only pupal exuviae, adults pinned) [PARATYPES,
& no. 1.175, % 1.176] (BMNH, B.M. 1952-234)
OTHER MATERIAL
ARGENTINAPINNED
Missiones, Oberá Villa Alberdi; 7.ii.1966, (Bachman)—1% (reared) (MLP).
BRAZILGoiás StateSPIRIT
Rio Saia Velha, (site 151); 13,I, 1976, (A.J. Shelley)—1 pupal exuviae (BMNH, B.M. 1979-580).
Minas Gerais StatePINNED
Mantequeira Mts., (loc. 24); 19-22.v.1979, (R.W.Crosskey & A.J.Shelley)—1% (reared) (BMNH, B.M.1979-258). Mantequeira Mts., km 31, Cruzeiro Itajubá, stream, (site 463); 19-22.v.1979, (A.J. Shelley &A.P.A. Luna Dias)—1&1% (reared) (BMNH).
Rio de Janeiro StatePINNED
Ilha Grande, ponto no. 11; 8.vi.1997, (A.P.A. Luna Dias & M.C.Viera)—1& (reared) (IOC). Ilha Grande, ponto A, (site 267); 30.v.1995, (R. Malaguti) [other collector’s name illegible]– 1& (reared) (IOC). Ilha Grande, Praia da Tapera, un-named stream; 8.vi.1997, (A.P.A. Luna Dias)—1& (reared) (IOC). Itaguaí, Sítio Porangaba; 30.v.1995, (R.Calvão)—1& (reared) (IOC).
SPIRITPetrópolis; 12.v.1979, (Coscarón)—2 pupal exuviae (BMNH). Petrópolis, stream from mountain at km
85 towards Rio, (site 1687, D25), 22º31’S44º 14’W; 18.iii.2004, (A.J. Shelley, A.P.A. Luna Dias & P.R.Garritano)—several pupal exuviae (BMNH).
São Paulo StatePINNED
S. José do Barreiro, un-named stream, at 500m from córrego at site 1685, (site 1686, D24), 22º42’S44º31’S; 18.iii.2004, (A.J. Shelley, A.P.A. Luna Dias & P.R. Garritano)—2&&1% (reared). S. José do Barreiro,Nascente na Faz. S. Sebastião da Cachoeira, (site 1675, D13), 22º41’S44º 30’W; 18.iii.2004, (A.J. Shelley,A.P.A. Luna Dias & P.R. Garritano)—1&1% (reared) (BMNH). S. José do Barreiro, un-named stream, at500m from córrego at site 1685, (site 1686, D24), 22º42’S44º 31’W; 18.iii.2004, (A.J. Shelley, A.P.A. LunaDias & P.R. Garritano)—5&&1% (BMNH; IOC). Itú, Fazenda Pau d'Alho; 18.vi.1960, [Without collector’sname.]– 1& (reared) (AMNH) [Identified by S. Coscarón 1979]. S. José do Barreiro, un-named stream at2500m from Fazenda Bonito; 18.iii.2004, (A.J. Shelley, A.P.A. Luna Dias & P.R. Garritano)—2&&1%(reared) (IOC). Ilha Bela, Usina Pontas da Canãs Armaçao; Faz. Cocara; 27.ii1957, (Vulcano & Martinez)—1&1% (reared) (MLP). Ilha Bela, Pontas de Canãs o riacho do lado da casa do prefeito; 14.vi.1957, (Travassos,Vulcano)—1% (reared) (MLP).
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.94 · Zootaxa 1649 © 2007 Magnolia Press
SPIRITS. José do Barreiro, un-named stream, at 500m from córrego at site 1685, (site 1686, D24), 22º42’S44º
31’W; 18.iii.2004, (A.J. Shelley, A.P.A. Luna Dias & P.R. Garritano)—numerous pupal exuviae and severallarvae (BMNH). José do Barreiro, cachoeira going to Arapeí, (site 1673, D11); 17.iii.2004, (A.J. Shelley,A.P.A. Luna Dias & P.R. Garritano)—1 pupal exuviae (BMNH). S. José do Barreiro, stream with waterfalls,
4th stream, (site 1674, D12); 17.iii.2004, (A.J. Shelley, A.P.A. Luna Dias & P.R. Garritano)—2 pupal exuviae(BMNH). S. José do Barreiro, Nascente na Faz. S. Sebastião da Cachoeira, (site 1675, D13), 22º41’S44º30’W; 18.iii.2004, (A.J. Shelley, A.P.A. Luna Dias & P.R. Garritano)—several pupal exuviae (BMNH; IOC).S. José do Barreiro, small stream at 300m do site 1678, (site 1679, D17), 22º43’S44º 32’W; 18.iii.2004, (A.J.Shelley, A.P.A. Luna Dias & P.R. Garritano)—1 pupal exuviae (BMNH). S. José do Barreiro, stream at 500mdo site 1682, (site 1683, D21), 22º46’S44º 31’W; 18.iii.2004, (A.J. Shelley, A.P.A. Luna Dias & P.R.Garritano)—1 pupal exuviae (BMNH). S. José do Barreiro, un-named stream, at 200m from córrego at site1683, (site 1684, D22), 22º46’S44º 31’W; 18.iii.2004, (A.J. Shelley, A.P.A. Luna Dias & P.R. Garritano)—numerous pupal exuviae (BMNH). Petrópolis; 12.v.1979, (Coscarón)—several pupal exuviae (MLP).Boraceia; 25.vii.1979, (Coscarón)—2 pupal exuviae (MLP). Bragança Paulista; 8.v.1949, (Andr. & Andr.)—1pupal exuviae (MLP). Entrada Velha de Cantos; 12.vi.1986, (Coscarón)—1 pupal exuviae (MLP). Boraceia;25.vii.1979, (Coscarón)—1% (reared) (MLP).
The following specimens does not have the locality information, date or collector’s name, but some slidesbear numbers “1-107, 1233, 1222, 1224, 1225, 1511” that might indicate they have been collected in Brazil,probably from São Paulo State (S. Coscarón, pers. com.):
[Without locality, date or collector’s name.] – 3&&2%% (reared) (MLP) [All specimens bears ahandwritten, black-bordered label “Simulium travassosi D’Andretta y D’Andretta det. Coscarón 1979” andnumbers “1-107, 1233, 1222, 1224, 1225”]. [Without locality, date or collector’s name.] – 1& (reared but notassociated with pupal exuviae) (MLP, no. 1511).
Santa Catarina StatePINNED
State boundary on BR 101, waterfall by road, (site 780); 18.xi.1986, (A.J. Shelley & A.P.A. LunaDias)—1pupal exuviae (adult lost) (BMNH).
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1649 © 2007 Magnolia Press · 95NEOTROPICAL BLACKFLY SUBGENUS INAEQUALIUM
Appendix. 1. Diagnosis of the subgenus Inaequalium by various authors.
Coscarón & Wygodzinsky (1984) [see also Coscarón (1987, 1991)]
Female. Body black. Frons and clypeus without opaline iridescence. Antennae, palps and proboscisblack, base of antennae pale brown. Thorax black, with grey pollinosity that with certain lights can have asubmedian grey pollinose band at each side, and on lateral and posterior borders. Pilosity of thorax with silverto yellow greyish tones. Scutellum, metanotum and pleura black; pleura silver pruinose. Halteres paleyellowish grey. Legs light greyish yellowish brown, black at apex of femur III and tarsal segments, exceptbasitarsal segments II and III (only apically) and coxae II and III. Abdomen black, with a pair of silver spotson tergite II, tergites I–IV velvet, tergites V–VIII shiny. Frons weakly convergent. Nudiocular angles withdeep incision. Basal region of cibarium elevated, covered by strongly sclerotized spines [=teeth]. Lutz’ organapproximately half diameter of basal segment with dense tubercles. Legs with petaloid scales. Calcipala welldeveloped, longer than wide. Sc with approximately 20 setae; basal sector of R with variable number of setaearranged in 1–2 lines. Claws with or without basal tooth. Gonapophyses divergent with internal bordersclerotized; eighth sternite with approximately 80 setae; paraprocts slightly shorter than wide, with apexrounded to truncate. Genital fork with median branch [=stem] nearly two times the length of each lateral arms. Male. Thorax velvet black, with silver bands on humeri and lateral and posterior margins, with certainlights grey pollinose iridescence extends to anterior third in form of pair of submedian silver areas. Thoracicpubescence golden in color. Abdomen black, with tergites II, VI and VII and small anterior region of VIIIwith pair of silver spots. Sc with 3–4 setae and basal sector of R with setae arranged in single line. Calcipalalonger than wide. Basimere [=gonocoxite] sub-trapezoidal, approximately as long as wide, and distimere[=gonostyle] slightly shorter, subtriangular and with stout apical spine. Ventral plate subovoid tosubtriangular, wider than long.
Pupa. Cocoon of thick threads and reinforced anterior border. Pupal gill generally with six filaments; ifthey are not filiform they have an irregular form composed of three thick branches. Head and thorax withdense tubercles. Cephalic trichomes with three-four branches and thoracic trichomes with 3–5 branches.Abdominal chaetotaxy very similar to Psaroniocompsa.
Larva. Naked [= without setae]. Cephalic apotome with a diffuse ornamentation, but resembling a centralcircle rounded by a darker area. Postgenal cleft slightly less or the same as the height [=length] ashypostomium. Median teeth of hypostomium slightly lower than lateral teeth. Hypostomial setaeapproximately six on each side. Antennal segments longer than fan stalk or slightly smaller with third segmentlonger than remaining segments. Mandibles with two marginal teeth, first twice as long as second. Cephalicfan with 35–40 rays. Anal ring [=posterior circlet] with approximately 65–75 rows of 12–15 hooks; body,distally, with two lobes [=ventral papilla] in the subterminal ventral region, sometimes indistinct. Anal lobeswith 6–13 branches on each lobe.
Coscarón (1987, 1991)
Coscarón provided similar morphological characters given in Coscarón & Wygodzinsky (1984) torecognize Inaequalium and added a few other characters: pupa with cephalic trichomes having 3–5 branches,thorax with 3-8-branched trichomes, and larva with 2–13 lobules on each lobe. They recognized two speciesgroups:�
inaequale species groupFemale. Paraprocts longer than wide, thick distally. Male. Distimere [= gonostyle] subtrapezoidal to subtriangular much shorter than basimere [=gonocoxite];
ventral plate much wider than high [=long]. Pupa. Gill filaments symmetric and filiform (except travassosi). Larva. Postgenal cleft deep, with short postgenal bridge (approximately 0.7 the height [=length] of
hypostomium).
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

HERNÁNDEZ ET AL.96 · Zootaxa 1649 © 2007 Magnolia Press
botulibranchium species groupFemale. Paraprocts shorter than high, narrow apically. Male. Distimere [=gonostyle] subcylindrical as long as basimere [=gonocoxite]; ventral plate as long as
or longer than wide. Pupa. Gill filaments asymmetric, thick. Larva. Postgenal cleft shallow, with postgenal bridge long (approximately 1.1 height [= length] of
hypostomium).
Py-Daniel et al. (1989)
The authors agreed with Coscarón & Wygodzinsky’s (1984) diagnosis of Inaequalium, but added a fewother characters.
Female. With setae on Sc. Male. Without setae on Sc. Larva. Antennal segments I and II darker than III; 1+1 sclerotized areas lateral to anal sclerite with
anterior region (that includes anterior arms) much longer than posterior.
Strieder & Py-Daniel (1999, 2000)
Adults. Simuliidae of black general coloration, medium size with wing length between 2.3–2.8 mm long;thorax laterally and posteriorly with grey stripe; cibarium of female elevated centrally, with conspicuous,sclerotized spines [=teeth]. Female with short paraprocts much longer than wide. Males basistyle[=gonocoxite] length and width almost equal and longer than the dististyle [= gonostyle]; dististylesubtriangular or subcylindrical with strong pre-apical spine.
Pupa. Dorsal length of cocoon between 2.1–3.8 mm and ventral length of cocoon 2.8–4.5 mm. Cocoontexture, in general, with visible threads and reinforced anterior border without basal or dorsal projection;filamentous gills with six terminal filaments (“inaequale” species group) or thick gills with 2-6 branches(“botulibranchium” species group); cephalic and thoracic trichomes multibranched or simple (I. souzalopesi).Thorax generally with tubercles densely distributed over entire surface or reduced in number (I.botulibranchium and I. souzalopesi). Abdomen membranous, chaetotaxy less variable with regards to numberand place of setae, hooks and denticules; posterior end of abdomen rounded, with terminal hooks small,subtriangular.
Larva. Body length 4.2–7.3 mm. Boyd shape bilobular or claviform. Ventral 1+1 posterior tuberclessubtriangular. Antennal segments generally longer than cephalic fans; third antennal segment usually darkerand longer than first and second antennal segments. Cephalic fan with 6 petaloid, rays. Postgenal cleft deepvarying from subtriangular to reduced in circular shape (I. souzalopesi). Mandibular lateral process (PLM)straight, curved or sinuous, usually reaching or longer the apex of mandibular ventral margin. Anterior thirdof distal comb with bifurcate setae near the base; posterior comb of setae of distal comb smaller than apicalthird of anterior ray; first ray anterior to larval mandible with 2–4 branches.
Coscarón & Coscarón-Arias (2007)
The diagnosis is the same as Coscarón (1987, 1991) except for the following additions: inaequale species group with basal portion of cibarium pyramidal and pointed apically, whereas in
botulibranchium species group basal portion of cibarium varies from low pyramidal median process torelatively depressed medially. Gill in botulibranchium with 2-6 thick, irregular filaments.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.


















