

· Web viewAuxin Binding Protein 1 Reinforces Resistance to Sugarcane Mosaic Virus in Maize...
41
Auxin Binding Protein 1 Reinforces Resistance to Sugarcane Mosaic Virus in Maize Pengfei Leng 1,2,# , Qing Ji 1,# , Torben Asp 3 , Ursula K. Frei 1 , Christina R. Ingvardsen 3 , Yongzhong Xing 4 , Bruno Studer 5 , Margaret Redinbaugh 6 , Mark Jones 6 , Priyanka Gajjar 1 , Sisi Liu 1,7 , Fei Li 1,8 , Guangtang Pan 7 , Mingliang Xu 2,* , Thomas Lübberstedt 1,* 1 Department of Agronomy, Iowa State University, Ames, IA, 50011, USA 2 National Maize Improvement Center, China Agricultural University, Beijing, 100193, China 3 Department of Molecular Biology and Genetics, Aarhus University, Forsøgsvej 1, DK-4200 Slagelse, Denmark 4 National Key Laboratory of Crop Genetic Improvement, Huazhong Agricultural University, Wuhan, 430070, China 5 Institute of Agricultural Sciences, Molecular Plant Breeding, ETH Zurich, Universitaetstrasse 2, 8092 Zurich, Switzerland
Transcript of · Web viewAuxin Binding Protein 1 Reinforces Resistance to Sugarcane Mosaic Virus in Maize...

Auxin Binding Protein 1 Reinforces Resistance to Sugarcane Mosaic
Virus in Maize
Pengfei Leng1,2,#, Qing Ji1,#, Torben Asp3, Ursula K. Frei1, Christina R.
Ingvardsen3, Yongzhong Xing4, Bruno Studer5, Margaret Redinbaugh6, Mark
Jones6, Priyanka Gajjar1, Sisi Liu1,7, Fei Li1,8, Guangtang Pan7, Mingliang Xu2,*,
Thomas Lübberstedt1,*
1Department of Agronomy, Iowa State University, Ames, IA, 50011, USA
2National Maize Improvement Center, China Agricultural University, Beijing, 100193, China
3Department of Molecular Biology and Genetics, Aarhus University, Forsøgsvej 1, DK-
4200 Slagelse, Denmark
4National Key Laboratory of Crop Genetic Improvement, Huazhong Agricultural University,
Wuhan, 430070, China
5Institute of Agricultural Sciences, Molecular Plant Breeding, ETH Zurich, Universitaetstrasse 2,
8092 Zurich, Switzerland
6USDA, Agricultural Research Service, Corn, Soybean and Wheat Quality Research Unit and
Department of Plant Pathology, Ohio State University, Wooster, OH, 44691, USA
7Maize Research Institute, Sichuan Agricultural University, Chengdu, 611130, China
8School of Life Sciences, Guizhou Normal University, Labboratory of Plant Physiology and
Developmental Regulation, Guiyang, 550000, China
#- These authors contributed equally to this work

Methods
Plant materials
Two early-maturing European maize inbred lines, FAP1360A and F7 were used to generate a
series of isogenic lines: F7SS/RR, and F7RR/RR (Xing et al., 2006). As previously described (Xing et
al., 2006), the letters left of the slash refer to the genotype at Scmv2 on chromosome 3 and those
on the right refer to the genotype at Scmv1 on chromosome 6, with R indicating a resistance
allele and S a susceptibility allele. Recombinant F2 plants derived from the cross of F7RR/RR and
F7SS/RR were used for further fine mapping of Scmv2 (Ingvardsen et al., 2010).
Virus inoculation and phenotype scoring
Seedlings were grown in a climate chamber at 22/20 °C day/night and 16/8 hour photoperiod
conditions. Inoculum was prepared and plants were inoculated with the Seehausen isolate of
SCMV as previously described (Kuntze et al., 1997). Plants not showing symptoms one week
post the first inoculation were inoculated a second time. Plant phenotypes were scored every day
until 4 wpi. F7 and FAP1360A were included as susceptible and resistant controls for each assay.
Fine-mapping of the Scmv2 region
The sequence of B73 within the 1.3 Mb candidate region (Ingvardsen et al., 2010) was retrieved
from the NCBI database as reference for marker development. Twelve informative markers were
used to screen the remaining six recombinant F2 offspring (Ingvardsen et al., 2010) in the Scmv2
region. Since all the offsprings were susceptible to SCMV (Kuntze et al., 1997), recombinants
showing the same pattern as FAP1360A were considered as recombinants in relation to the
resistance gene Scmv2.
Isolation of BAC clones covering the Scmv2 region

To identify positive BAC clones covering the Scmv2 region from a FAP1360A BAC library
(Ingvardsen et al., 2005), flanking and co-segregating markers (184B1, DJF004, 184FG018, and
212FG008) were used to screen the FAP1360A BAC library by a three-step PCR strategy (Xu et
al., 2001). Additional primers (Supplementary Table 1) were used within the target region to
test for redundancy of positive BAC clones. Fingerprinting was used to confirm the overlap
between adjacent clones. Plasmid DNA from BAC clones was digested with HindIII, EcoRI and
BamHI (Thermo Scientific, Waltham, MA, USA) separately. After gel electrophoresis at 40 V
overnight at 4 °C, DNA fragments were visualized under UV light. Clones sharing more than
three bands of the same size for each of the three digestions were considered as overlapping. A
minimum tiling path covering the Scmv2 region was constructed after eliminating redundant
BAC clones.
BAC clone sequencing and sequence analysis
Plasmid DNA from BAC clones was extracted using QIAGEN® Large-Construct Kit (QIAGEN,
Valencia, CA, USA) according to the manufacturer’s instructions and quantified using a
NanoDrop (Thermo Scientific, Wilmington, DE, USA). For each clone, 5 μg of DNA was sent to
the DNA facility at University of Iowa for sequencing using 454 GS FLX Titanium (Roche
Diagnostics Corporation, Indianapolis, IN, USA) technology. Vector pIndigoBAC-5 and host
strain DH10B genome sequences were identified using BLAST and removed from the resulting
reads. Sequence analysis on the subsequent dataset was conducted using NextGENe® software
(Softgenetics, College, PA) using the Greedy assembly method (Zhang et al., 2000) and B73
sequence as reference. Traditional Sanger sequencing on ABI3730 (Life technologies, Carlsbad,
CA, USA) was subsequently applied to close gaps resulting from 454 pyrosequencing. Genes

within the Scmv2 region were predicted using Softberry
(http://linux1.softberry.com/berry.phtml), and using FGENESH with the organism as monocot.
Cloning of candidate gene sequences from positive BAC clones, RNAi, and deletion
construct
The restriction sites flanking the candidate genes were identified based on the FAP1360A
sequence and used to sub-clone intact Scmv2 candidate genes from the corresponding BAC
clones. In this study, an intact candidate gene was defined as containing around 1.7 kb promoter
sequence, the full coding region, and the 3’ UTR region, totally 7.4 kb. DNA fragments with
intact candidate genes were sub-cloned into the binary vector pTF101.1 (provided by Dr. Kan
Wang, Plant Transformation Facility (PTF) at ISU:
http://agron-www.agron.iastate.edu/ptf/index.aspx) (Figure 1D).
The binary vector pMCG1005 (provided by Dr. Kan Wang, PTF) was used for RNAi constructs.
Part of the predicted transcript for each candidate gene was amplified from FAP1360A by SpeI
and SmaI restriction site-tailed primers (Supplementary Table 1). The amplicon was digested
with SpeI and SmaI to obtain the insert fragment. The final RNAi construct was prepared by two
ligations with the same fragment. SpeI and SmaI digestion was done first to create sense
insertions, followed by AvrII and XcaI digestion to create the antisense insertion
(Supplementary Figure 1).
The deletion construct (pTF-dABP1FAP) was modified from the full length construct of the
ABP1FAP gene by deleting the 524 bp insertion (Figure 1D). All transgenic constructs were
sequenced to verify the insertion and direction prior to transformation into HiII immature zygotic
embryos via Agrobacterium EHA101 mediated transformation at PTF (ISU, Ames, IA, USA)
(Frame et al., 2002).

Transgenic plant assays
T0 transgenic plantlets were transplanted into the Agronomy greenhouse (ISU) at 16/8 hours
(light/dark) growing conditions at a light intensity of 230 µE m -2s-1 and temperature at 28/21 ˚C
(day/night).
For RNAi tests, T0 transgenic plants from RNAi were crossed and then backcrossed to F7RR/RR.
BC1 plants homozygous for the Scmv2 allele from F7RR/RR were selected for further analysis using
a ABP1FAP specific marker 197S06 (Leng et al., 2015). Moreover, BC1 individuals were either
heterozygous or homozygous for the dominant Scmv1 resistance allele from F7RR/RR. For each
transgenic event, at least 36 progeny were planted in a growth chamber for SCMV inoculation.
Inoculation and phenotyping was performed by visual inspection as described above, and
combined with enzyme-linked immunosorbent assays (ELISA). Protein-A sandwich (PAS)-
ELISA assays (Edwards and Cooper, 1985) were conducted using antisera specific for SCMV at
a dilution of 1: 3000. For complementation tests, T0 transgenic plants were crossed and then
backcrossed to F7SS/RR. Only BC1 plants fixed for the F7 allele at Scmv2 locus were selected for
further analyses. A similar approach was used to test the the pTF-dABP1FAP construct. Leaf
samples were harvested at 4 wpi and frozen by liquid nitrogen immediately for RNA expression
analysis, ZmABP1 expression level in transgenic lines were calibrated by F7.
Rapid Amplification of cDNA Ends (RACE) and subcellular localization
Plant leaves from both FAP1360A and F7 were harvested before and at different time points post
SCMV inoculation. Total RNA was extracted using RNeasy Mini Kit (QIAGEN Inc, Valencia,
CA, USA) following manufacturer’s instructions. Genomic DNA was removed by
Deoxyribonuclease I (Invitrogen, Carlsbad, CA). The 5’ and 3’ ends of the ZmABP1 cDNAs

from FAP1360A and F7 were amplified using the SMARTer™ RACE cDNA Amplification Kit
(Clontech, Takara, Japan) according to the manufacturer’s guide.
Primers tailed with XhoI and BamHI (Supplementary Table 1), respectively, were designed to
amplify the ZmABP1 cDNA from both F7 and FAP1360A, and cloned into pEZS-NL XhoI and
BamHI site. The resulting constructs were named pEZS-ABP1F7 and pEZS-ABP1FAP,
respectively. In this study, as ZmABP1 contains a potential ER signal peptide at the C-terminus,
the mCherry tagged ER marker ER-rk CD3-959 was used as reference (Nelson et al., 2007). A
total of 1 µg plasmid DNA of both pEZS-ABP1F7, pEZS-ABP1FAP, and CD3-959 were prepared
using QIAprep Spin Miniprep Kit (QIAGEN, Valencia, CA). Bombardment of onion epidermal
cells was conducted as described (Sanford et al., 1993). CD3-959 was co-bombarded with either
pEZS-NL, pEZS-ABP1F7, and pEZS-ABP1FAP. Onion epidermal peels were pre-cultivated for 4-5
hours on agar plates containing 0.5 mM 2-(N-morpholino) ethanesulfonic acid (MES), and 15
g/L of BD BactoTM agar, pH 5.7). Gold particles (1.0 μm) coated with plasmid were bombarded
into onion epidermal cells using a PDS 1000/He biolistic gun (Bio-Rad, Hercules, CA, USA) at a
helium pressure of 4.5 MPa and 6 cm target distance. The bombarded samples were incubated at
room temperature at darkness for 24 hours and then inspected using a Leica TCS NT confocal
microscope system. Green fluorescence was excited with a 488 nm laser. Leica Confocal
Software (LAS-AF-Lite_2.6.0_7266) was used for post-acquisition image processing.
ABP1F7-GFP and ABP1FAP-GFP were subcloned into the pBCXUN vector via XcmI, the resulting
constructs and CD3-959 were transformed into GV3101 strains and grown in YEB liquid
medium containing 50 mg/mL rifampicin, 15 mg/mL kanamycin, at 28 °C. Agrobacterium
suspensions together with the Agrobacterium p19 strain were mixed at final optical density at
600 nm (OD600) of 0.5. Transient protein expression in N. benthamiana was carried out by

Agrobacterium infiltration (Waadt and Kudla, 2008). The infiltrated plants were incubated for 72
hours post infiltration (hpi) in a climate chamber under controlled conditions. Green fluorescence
was excited with a 488 nm laser and red fluorescence was excited with a 545 nm laser, and
visualized using an Olympus FluoView FV1000 confocal microscope (Olympus Optical Co.,
Tokyo, Japan).
Real-Time PCR analysis
For real time PCR (RT-PCR), leaf samples were collected at 0 h, 1, 2, 7, 14, 21, and 28 dpi from
F7 and F7RR/RR for RNA extraction using a RNeasy Mini Kit (QIAGEN, Valencia, CA, USA).
For each time point, two to three young plantlet leaves were pooled for each sample. Total RNA
was digested by Deoxyribonuclease I (Invitrogen, Carlsbad, CA, USA) and tested using the
SuperScript® One-Step RT-PCR System with Platinum®Taq DNA Polymerase (Invitrogen,
Carlsbad, CA, USA). The first-strand cDNA was synthesized using 0.2 µg RNA followed by
SuperScript® III First-Strand Synthesis SuperMix (Invitrogen, Carlsbad, CA, USA).
Comparative qRT-PCR was conducted in a total volume of 20 µl containing 1 µl reverse-
transcribed product, 0.2 uM gene-specific primers (Supplementary Table 1) and 10 µl Power
SYBR® Green RT-PCR Master Mix (Applied Biosystems®, Warrington, UK), reactions were
loaded into 96-well plates and the real-time PCR reactions were carried out in the MX3000P
Real-Time PCR System (Stratagene, USA) according to the manufacturer’s instructions. Primers
were designed by PRIMER3 (http://gmdd.shgmo.org/primer3/?seqid=47), and checked for
primer efficiency and against primer dimer formation. A rice Actin2 gene was used as internal
reference gene, gene expression levels in relation to reference gene were calculated by the 2(-△△Ct)
method, which quantifies the amount of target sequence as fold change relative to a calibrator
and normalized to Actin2 (Livak et al., 2001). For individual candidate genes, the expression at

different time-points and treatments were calibrated to the expression at time 0 (plants before
inoculation). Each time point was determined with three biological replicates, three technological
replicates per each. The following PCR protocol was used: denaturation at 95 °C for 10 min,
followed by 40 cycles of 95 °C for 15 s, 58 °C for 30 s, and 60 °C for 30 s. All primer sequences
are listed in Supplementary Table 1.
Promoter assay
Three regions of ZmABP1 promoter, 430 bp, 1.281 bp and 1711 bp, were cloned from
FAP1360A, the corresponding 1711 bp region of F7 was obtained by two rounds genome
walking (Takara, Dalian, China). Cis elements identification were conducted using PLACE
(plant cis-acting regulatory DNA elements) analysis (Lescot et al., 2002). The transient assay
reporter constructs were developed with reporter vector pGreenII-0800 for Double Luciferase
Assay and a transient assay with maize leaf protoplasts was used to test relative luciferase (LUC)
activity (Studer et al., 2011). A reporter construct containing the cauliflower mosaic virus
(CaMV) 35S minimal promoter driving expression of the firefly luciferase gene was used to test
control region segments. Transient expression assays using maize mesophyll protoplasts were
performed according to Sheen’s protocol (Sheen et al., 2001) with modifications as follows. 2 ×
106 freshly isolated protoplasts in 200 uL buffer were transformed with 10 ug plasmid by PEG
mediated transformation. Protoplast mixes were incubated at 23 °C for 18 hours post
transfection. The harvested protoplasts were lysed with CCLR (Cell Culture Lysis Reagent,
Promega, WI, USA) and assayed using the Dual-Luciferase Reporter Assay System (Promega,
WI, USA) following manufacturer's instructions. Four to six biological replicates, three technical
replicates per each, were assayed for each construct.
Virus and protoplast isolation

SCMV was isolated and purified from 2 wpi Mo17 infected leaves, viral RNA extraction was
done as described (Dijkstra and de Jager, 1998) with modifications. The maize protoplasts
isolation and transfection were performed according to Sheen’s protocol (Sheen et al., 2001) with
some modifications. A mixture of 10 μg plasmids (pEZS-ABP1F7, and pEZS-ABP1FAP) and 5 μg
purified SCMV cDNA was co-transformed via PEG into 0.5 ml protoplasts (2 × 106/ml), and
pEZS-NL was mock treatment. Total RNA from transfected protoplast system at 0, 4, 8, 12, and
24 hours post transfection were isolated by TRIzol (Invitrogen, Carlsbad, California, USA). Both
ZmABP1 expression and SCMV replication in transfected protoplasts were measured by
quantitative RT-PCR, Actin2 was used as internal reference gene. All primer sequences are listed
in Supplementary Table 1.
Yeast two-hybrid screen, protein interaction, BiFC and co-immunoprecipitation assay
Total RNA was extracted from the resistant inbred line 1145 leaf tissue at 24 hours post SCMV
inoculation and used for cDNA library construction according to the manufacturer’s instructions
(the Make Your Own “Mate & Plate” Library System, Clontech, Mountain View, CA). The
ZmABP1 cDNA was cloned into pGBKT7 NdeI and BamHI site (designated as BD-ABP1) and
transformed into the yeast strain Y2H GOLD (Clontech) as bait. Library screening was
performed according to the Matchmaker Gold yeast two-hybrid system protocol (Clontech).
Colonies were plated on triple dropout synthetic selective medium lacking Trp, Leu, and His and
supplemented (TDO) and then screened on the selective medium synthetic dextrose/-Ade/-Trp/-
Leu/-His/X-α-Gal/Auresbasidin A (QDO/X/A). The full-length cDNAs of all positive prey were
cloned from 1145 and into yeast vector pGADT7 for AD construction, and transformed back into
Y2H to validate the interaction with BD-ABP1. Interaction was considered relevant when clones

were able to grow on both DDO and stringent QDO medium compared with corresponding
controls. Primers used for the yeast assays are listed in Supplementary Table 1.
For the BiFC assay, full length cDNA of ZmABP1, SCMV- 6K1 and CI were sub-cloned into the
pUC-SPYCE vector via BamHI/XhoI. The full length cDNA of ZmRbCS were sub-cloned into
the pUC-SPYNE vector via EcoRI/SacI. The resultant recombinant constructs pABP1-YFPC and
pRbCS-YFPN were transformed into GV3101 strains. Transient protein expression in N.
Benthamiana was carried out by Agrobacterium infiltration (Waadt and Kudla, 2008). The
yellow fluorescence was excited with a 515 nm laser and visualized using an Olympus FluoView
FV1000 confocal microscope (Olympus Optical Co., Tokyo, Japan).
For co-immunoprecipitation (co-IP) assays, the proteins from transgenic N. Benthamiana leaves
were extracted with extraction buffer (0.5 M Tris-MES, pH8.05, 0.5 M Sucrose, 0.5 M EDTA, 1
M MgCl2, 0.5 × protease inhibitor cocktail and 1 × PMSF) and centrifuged at 14,000 g at 4 °C
for 30 min, the supernatant was collected as the protein extract. Co-IP was performed using anti-
HA mAb-Magnetic Beads (Medical and Biological Laboratories Co., LTD, Japan) according to
the manufacture’s instruction, the antigen-antibody complex was collected, seperated on 12%
SDS/PAGE gels and transferred to PVDF transfer membrane (Amersham Hybond-P; GE
Healthcare, Germany) for western blot analysis, with Anti-HA-tag and Anti-Myc-tag mAb as
Primary antibodies respectively (Medical and Biological Laboratories Co., LTD, Japan) and
HRP Conjugated Goat Anti-Mouse IgG (Easybio, China) as Secondary antibody. Pierce™ ECL
Plus Western Blotting Substrate (Thermo Scientific™, U.S.) for chemiluminescence and imaged
on Tanon 5200 (Tanon Science & Technology Co., Ltd., China).
Statistical analysis

For the transgenic tests, the difference between transgenic and non-transgenic plants were
compared using χ2 tests. For ZmABP1 transcript expression levels in near isoenic lines, three
biological replicates were employed for each, and compared with the CONTRAST procedure of
SAS 9.3 (SAS Institute).
Supplemental Bibliography
Dijkstra, J., and de Jager, C. (1998). Practical plant virology: protocols and exercises. In
Springer Lab Manual, J. Dijkstra, C.P. De Jager, ed. (Berlin, New York Springer-Verlag),
pp. 308-311.
Edwards, M.L., and Cooper, J.I. (1985). Plant-Virus Detection Using a New Form of Indirect
Elisa. J. Virol. Methods 11: 309-319.
Frame, B.R., Shou, H., Chikwamba, R.K., Zhang, Z., Xiang, C., Fonger, T.M., Pegg, S.E.,
Li, B., Nettleton, D.S., Pei, D., et al. (2002). Agrobacterium tumefaciens-mediated
transformation of maize embryos using a standard binary vector system. Plant Physiol. 129:
13-22.
Ingvardsen, C., Xing, Y., Xu, M., and Lübberstedt, T. (2005). Molecular analysis of
Sugarcane mosaic virus resistance in maize. Parasitica 61: 61-66.
Kuntze, L., Fuchs, E., Gruntzig, M., Schulz, B., Klein, D., and Melchinger, A.E. (1997).
Resistance of early-maturing European maize germplasm to sugarcane mosaic virus
(SCMV) and maize dwarf mosaic virus (MDMV). Plant Breeding 116: 499-501.

Leng, P.F., Ji, Q., Tao, Y., Ibrahim, R., Pan, G., Xu, M., and Lübberstedt, T. (2015).
Characterization of Sugarcane Mosaic Virus Scmv1 and Scmv2 Resistance Regions by
Regional Association Analysis in Maize. Plos One 10: e0140617.
Lescot M, Déhais, P., Thijs, G., Marchal, K., Moreau, Y., Van de Peer, Y., Rouzé, P., and
Rombauts, S. (2002). PlantCARE, a database of plant cis-acting regulatory elements and a
portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 30: 325-327.
Livak, K.J., and Schmittgen, T.D. (2001). Analysis of relative gene expression data using real-
time quantitative PCR and the 2−ΔΔCT method. Methods 25: 402-408.
Nelson, B.K., Cai, X., and Nebenfuhr, A. (2007). A multicolored set of in vivo organelle
markers for co-localization studies in Arabidopsis and other plants. Plant J. 51: 1126-1136.
Sanford, J.C., Smith, F.D., and Russell, J.A. (1993). Optimizing the Biolistic Process for
Different Biological Applications. Method Enzymol. 217: 483-509.
Sheen, J. (2001). Signal transduction in maize and Arabidopsis mesophyll protoplasts. Plant
Physiol. 127: 1466-1475.
Studer, A., Zhao, Q., Ross-Ibarra, J., and Doebley, J. (2011). Identification of a functional
transposon insertion in the maize domestication gene tb1. Nat. Genet. 43: 1160-1163.
Waadt, R., and Kudla, J. (2008). In Planta Visualization of Protein Interactions Using
Bimolecular Fluorescence Complementation (BiFC). CSH protocols 2008, pdb.prot4995.
Xu, M., Song, J., Cheng, Z., Jiang, J., and Korban, S.S. (2001). A bacterial artificial
chromosome (BAC) library of Malus floribunda 821 and contig construction for positional
cloning of the apple scab resistance gene Vf. Genome 44: 1104-1113.

Zhang, Z., Schwartz, S., Wagner, L., and Miller, W. (2000). A greedy algorithm for aligning
DNA sequences. J. Comput. Biol. 7: 203-214.

Supplementary Figure 1. Down regulation of ZmABP1 by RNAi resulted in susceptible
plants
(A) A small DNA fragment (421 bp) of ZmABP1 was reversely inserted into the pMCG1005
construct before and after the Rice waxy-a intron1 and driven by an ubiquitin (Ubi) promoter.
The pMCG1005-GSS construct consists of a 250 bp sense/antisense fragment of ZmGSS gene.
(B) The pMCG1005-ABP1 transgenic lines (from transgenic events 1-5 and 27-1) showed
SCMV susceptiblity correlated with interfered ZmABP1 expression. Data are shown as mean ±
standard error (SE). Numbers on top of each column refer to the individual lines used for
comparison, the same as below. Letters on top of each column indicate significant differences (**
P = 0.01).
(C) ELISA assays showed a large amount of SCMV accumulated in symptomatic transgenic
plants supported by OD450 value. Data are shown as mean ± standard error (SE). Letters on top of
each column indicate significant differences (** P = 0.01).

Supplementary Figure 2. Sequence alignment of ZmABP1 alleles among different inbred
lines
A total of five exons (E1-5) were found for all ZmABP1 alleles. A 524 bp insertion (Red triangle)
was discovered in two resistant lines (FAP1360A and Pa405), but not in other resistant inbred
lines. Compared to FAP1360A, F7 has five SNPs, two of which are synonymous SNPs located
in the first exon (E1). No consistent sequence differences were found between susceptibility and
resistance alleles. R, resistant line. S, susceptible line.


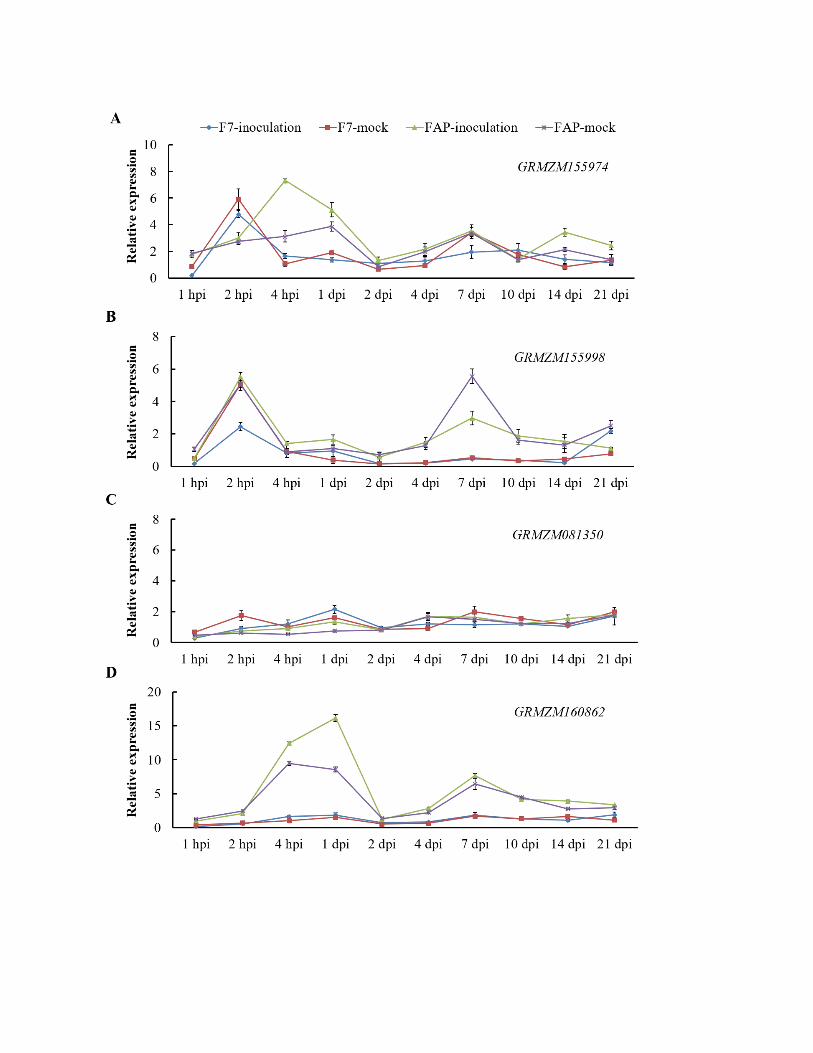
Supplementary Figure 3. Real time RT-PCR analysis of other candidate genes within the
fine mapped Scmv2 region at different time points post SCMV inoculation using F7 and
FAP1360A
(A) Expression of gene (GRMZM155974) encoding Glutathione synthetase was up-regulated
from 2 hours post inoculation (hpi) and higher in F7 than FAP1360A, reached the highest level
at 4 hpi, showed early response to stress.
(B) The gene (GRMZM155998) encoding putative uncharacterized protein also showed an early
up-regulation 2 hpi, and started to decrease from 4 days post inoculation (dpi) while the mock
control was up-regulated at 10 dpi.
(C) The gene (GRMZM081350) encoding hypothetical protein shows no expression difference
between resistant and susceptible genotypes, SCMV inoculated and mock control plants.
(D) The gene (GRMZM160862) encoding unknown protein was immmeadiately induced with
SCMV inoculation and pretty higher in FAP1360A, no significant difference between the mock
and inoculated plants for both F7 and FAP1360A. The data were shown as mean ± SE, obtained
from three technical replicates.

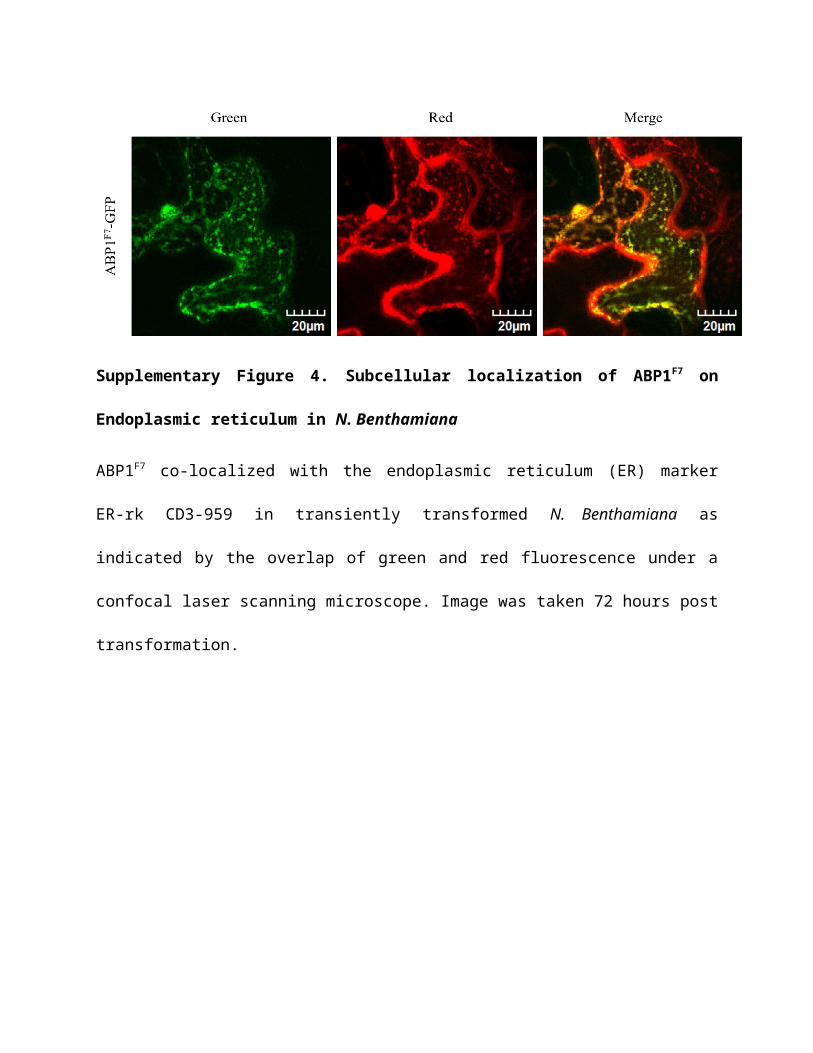
Supplementary Figure 4. Subcellular localization of ABP1F7 on Endoplasmic reticulum in
N. Benthamiana
ABP1F7 co-localized with the endoplasmic reticulum (ER) marker ER-rk CD3-959 in transiently
transformed N. Benthamiana as indicated by the overlap of green and red fluorescence under a
confocal laser scanning microscope. Image was taken 72 hours post transformation.

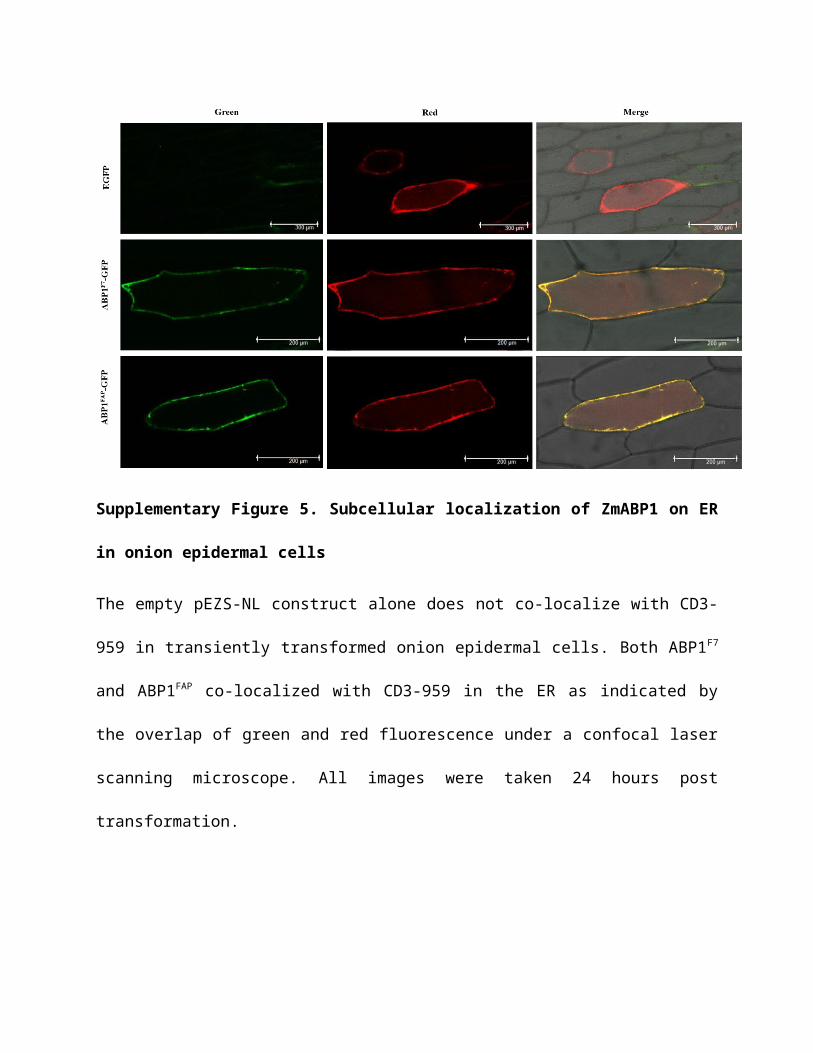
Supplementary Figure 5. Subcellular localization of ZmABP1 on ER in onion epidermal
cells
The empty pEZS-NL construct alone does not co-localize with CD3-959 in transiently
transformed onion epidermal cells. Both ABP1F7 and ABP1FAP co-localized with CD3-959 in the
ER as indicated by the overlap of green and red fluorescence under a confocal laser scanning
microscope. All images were taken 24 hours post transformation.

Supplementary Figure 6. Subcellular localization of ZmABP1 on Plasma membrane in
onion epidermal cells
Confocal images of transiently transformed onion epidermal cells, which express GFP and RFP
fusion proteins. The merged yellow flurorescence indicates both ABP1F7 and ABP1FAP GFP
fusion proteins under control of the 35S promoter co-localize with the plasma membra (PM)
marker PM-rk CD3-1007. All images were taken 24 hours post transformation.

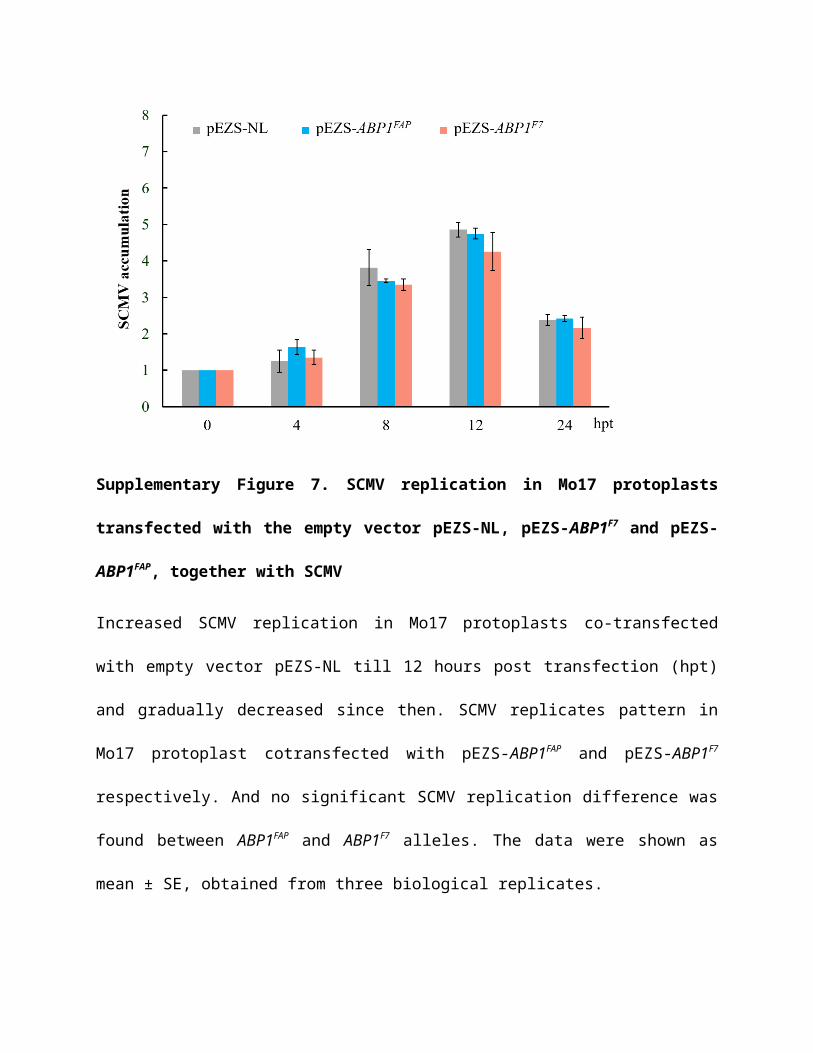
Supplementary Figure 7. SCMV replication in Mo17 protoplasts transfected with the
empty vector pEZS-NL, pEZS-ABP1F7 and pEZS-ABP1FAP, together with SCMV
Increased SCMV replication in Mo17 protoplasts co-transfected with empty vector pEZS-NL till
12 hours post transfection (hpt) and gradually decreased since then. SCMV replicates pattern in
Mo17 protoplast cotransfected with pEZS-ABP1FAP and pEZS-ABP1F7 respectively. And no
significant SCMV replication difference was found between ABP1FAP and ABP1F7 alleles. The
data were shown as mean ± SE, obtained from three biological replicates.

Supplementary Figure 8 Analysis of interaction between ZmABP1 and ZmRbCS by yeast
two-hybrid assay
Interaction of ZmABP1 and ZmRbCS was conducted by a yeast two-hybrid assay in vitro.
Cultures of diploid yeast clones were cultivated for eight hours, and three dilutions (1: 10, 1:
100, and 1: 1000) were spotted on double dropout synthetic selective medium lacking Trp, Leu
(DDO) to monitor proper growth, and on synthetic
dextrose/-Ade/-Trp/-Leu/-His/X-α-Gal/Auresbasidin A (QDO/X/A) medium for interaction
assays. BD-ABP1/pGADT7, vector control; BK-53/AD-T, positive control; BK-Lam/AD-T,
negative control.

Supplementary Figure 9 Analysis of interaction between ZmABP1 and ZmRbCS by BiFC
assay in N. Benthamiana
BiFC assays shows that ZmABP1 and ZmRbCS interacts in vivo. 5-6 weeks old N. benthamiana
leaves were agro-infiltrated with NE-ABP1 and CE-RbCS and co-infiltrated with p19. The right
panel is merged. RbCS-YFPN + 35S-YFPC and ABP1-YFPC + 35S-YFPN, negative control.
Supplementary Figure 10 Detection of interaction between ZmABP1 and ZmRbCS.
Co-immunopreceipitation (co-IP) assay of the interaction between ZmRbCS and ZmABP1
proteins in the leaf tissues of N. benthamiana. Anti-myc antibodies detected RbCS-Myc among
proteins pulled-down with anti-HA antibodies, while anti-HA antibodies detected ABP1-HA
among proteins pulled-down with Anti-Myc antibodies.

Supplementary Figure 11. ABP1F7 can also interact with ZmRbCS
(A) diploid yeast clones containing ABP1F7 and ZmRbCS could grow on double dropout
synthetic selective medium lacking Trp, Leu (DDO) and synthetic dextrose/-Ade/-Trp/-Leu/-
His/X-α-Gal/Auresbasidin A (QDO/X/A) medium by yeast two hybid assay.
BD-ABP1/pGADT7, vector control; BK-53/AD-T, positive control; BK-Lam/AD-T, negative
control.
(B) the BiFC assays showing that ABP1F7 can also interact with ZmRbCS in vivo. 5-6 weeks old
N. benthamiana leaves were agro-infiltrated with NE-ABP1F7 and CE-RbCS, co-infiltrated with
p19, and imaged 72 hours post infiltration. The right panel is merged. RbCS-YFPN + 35S-YFPC
and ABP1-YFPC + 35S-YFPN, negative control.
Supplementary Table 1 Primers used in this study
Description Primer name (5'-3')
RNAi,
allele
sequencing
AuxRNAiForward actagtCGTTGATAAGAGAGGAGAGC
Reverse cccgggACTTAGCAGTGTTGGTCTCA
GSHSRNAiForward actagtCAACTTCCGATCCGATGGTG
Reverse cccgggGCACTCATCGCAGGTATTGG
PMCG1005-1Forward AGGCGTCGCATATCTCAT
Reverse GGCTCACCAAACCTTAAACAA
Allele
sequencing 207P5Forward GCAGCTATGGGATTGAA
Reverse GCTCGTCAGAATTCCAAACC
AuxW2F Forward GCCGGATCTAAGCGAACTCG
AuxW3R Reverse TGATTGAGCGCACCAGCAAC
Aux19W1 Forward CATGGCGTTGCTTCATAAGG
Aux19W1 Reverse ACACCCTAGAAACGACTCTC
Transgenic
testing BarForward GTACGGAAGTTGACCGTGCT
Reverse GGATCTACCATGAGCCCAGA
ABP1Forward ATGTGGGCTATCCGGATTTA
Reverse CGGGTGGGTAGTACATGGAT
Sub-cellular
localization FAPGFPForward ctcgagTCATTCCACTCCGACATTCA
Reverse ggatccACGAAGCAGTCCTCATCCCAGA
F7GFP Forward ctcgagTCATTCCACTCCGACAATCA
Reverse ggatccACGAAGCAGTCCTCATCCCAGA

qRT-PCRABP1RT
Forward AATCCCTGTAAACGATCCAC
Reverse GGCATGCTCCAATCATCATA
GSRTForward GCCTGCACCAGGAGAGTTAG
Reverse GCAGGCCTGGGTATTGAAGT
0126RTForward CCACTCCCACAACAACGGAG
Reverse CGAATGCGACCAGGAGAGTG
ZincRTForward TTGCCGCAGTTGTGAGGTAT
Reverse CAATCGTCCACACGCATCAC
Actin2RTForward GGCTGACGCAGAGGACATTC
Reverse AGTGTGACGTGGACGACCAA
Middle type
construction Mid1Forward TAGTGCCATGCTGTAGTTTG
Reverse AGCAAGAAGGAACCATAACA
SCMV
detection SCMVForward GGCGAGACTCAGGAGAATACA
Reverse ACACGCTACACCAGAAGACACT
Protein
interaction
BD-ABP1,
AD-ABP1
Forward catatgGCGCCGGATCTAAGAGA
Reverse ggatccCTAGAGTTCGTCTTTTGC
BD- RbCS,
AD-RbCS
Forward catatgATGGCGCCCACCGTGATGA
Reverse gaattcCTACTCGCTGCCGGGGGGC
CE-ABP1,
NE-ABP1
Forward ggatccATGGCGCCGGATCTAAGAGA
Reverse ctcgagGAGTTCGTCTTTTGCTGCTTCG
CE-6K1 Forward ggatccATGGGTAAATCCAACTTGGAAACA
Reverse atcgatCTGTTGTGTTACAGGAGGC
NE-RbCSForward atcgatATGGCGCCCACCGTGATGA
Reverse cccgggCTCGCTGCCGGGGGGC
Genome
walking
GSP1 Reverse AATGCAAACCGAGCCACTAAGC
GSP2 Reverse CTTAGCAGTGCTGGTCTCAGTGG
GSP3 Reverse AGTTCTCTTAGATCCGGCGCCAT
GSP4 Reverse GCGAAGGTGAAGGCCAAGCG
GSP5 Reverse GGTGATATGGTACAAGTCACAGGG
GSP6 Reverse CCTAGCCACACGGTGAACAGCG
Promoter
activity analysis FAP0.4KForward aagcttCAGTGCACCGTGCAGCACGGTC
Reverse ggatccGGGACTCCCGAGTAAGCATGTCGGG
FAP1.2KForward aagcttGGCGCCATAATCGGGGGTCCAGG
Reverse ggatccTGCCTGCTCCCGACAGCTGCA
FAP1.7KForward aagcttCAGTGCACCGTGCAGCACGGTC
Reverse ggatccTGCCTGCTCCCGACAGCTGCA
F70.4 KForward aagcttGGTCGAGGTGCCGTAAAGCACT
Reverse ggatccGGCGCCGGATCTAAGAGAACTAAGG
F7 1.7KForward aagcttGGTCGAGGTGCCGTAAAGCACT
Reverse ggatccTGCCTGCTCCCGACAGCTGC
Nucleotides that are underlined indicate restriction sites.


















