
Wang Et Al 2006 Dave Long Paper
11
Anticancer activity of litchi fruit pericarp extract against human breast cancer in vitro and in vivo Xiujie Wang a, ⁎ , Shulan Yuan a , Jing Wang a , Ping Lin a , Guanjian Liu b , Yanrong Lu a , Jie Zhang a , Wendong Wang c , Yuquan Wei d, ⁎ a Divisio n of Experimental Oncology , Nationa l Key Laborato ry of Biothera py, Wes t China Hospital, Sichuan Universit y, Chengdu 610041, Sichuan Province, P .R. China b Department of Clinical Epidemiolo gy, Wes t China Hospital, Sichuan Universit y, Chengdu 610041, Sichuan Province , P .R. China c Department of Pathology of Public Health School, Sichuan University, Chengdu 610041, Sichuan Province, P.R. China d Divisio n of Biother apy , National Key Laborat ory of Biother apy , West China Hospital, Sichuan Universit y, Chengdu 610041, Sichuan Provinc e, P .R. China Received 8 October 2005; revised 23 January 2006; accepted 8 February 2006 Available online 23 March 2006 Abstract Litchi fruit pericarp (LFP) extract contains significant amounts of polyphenolic compounds and exhibits powerful antioxidative activity against fat oxidation in vitro. The purpose of this study is to confirm the anticancer activity of LFP extract on human breast cancer in vitro and in vivo, and to elucidate the mechanism of its activity. Human breast cancer cells were tested in vitro for cytotoxicity, colony formation inhibition, BrdU incorporation, and gene expression profiling after treatment with LFP extract. Seven nude mice bearing human breast infiltrating duct carcinoma orthotopically were tested for its anticancer activity and expression of caspase-3 in vivo by oral administration of 0.3% (0.3 mg/ml) of LFP water- soluble crude ethanolic extract (CEE) for 10 weeks. LFP extract demonstrated a dose- and time-dependent inhibitory effect on cell growth (IC 50 = 80 μg/ml), and it significantly inhibited colony formation and BrdU incorporation of human breast cancer cells. Oligonucleotide microarray analysis identified 41(1.22%) up-regulated and 129 (3.84%) down-regulated genes after LFP water-soluble CEE treatment; the predominantly up-regulated genes were involved in various biological functions including cell cycle regulation and cell proliferation, apoptosis, signal transduction and transcriptional regulation, and extracellular matrix/adhesion molecules; and down-regulated genes were mainly associated with adhesion, invasion, and malignancy of cancer cells. A 40.70% tumor mass volume reduction and significant increase of casepase-3 protein expression were observed in vivo experime nt. The findings in this study suggested that LFP extract might have potential antican cer activity on both ER positive and negative breast cancers, which could be attributed, in part, to its DNA damage effect, proliferating inhibition and apoptosis induction of cancer cells through up-regulation and down-regulation of multiple genes involved in cell cycle regulation and cell proliferation, apoptosis, signal transduction and transcriptional regulation, motility and invasiveness of cancer cells; ADP-ribosyltransferase (NAD+; poly (ADP-ribose) polymerase)-like 1 (ADPRTL1), Cytochrome P450, subfamily I (CYP1A1) and Hyaluronan-mediated motility receptor (HMMR) might be the main molecular targets at which LFP water-soluble CEE acted. © 2006 Elsevier Inc. All rights reserved. Keywords: Litchi fruit pericarp extract; Anticancer activity; Breast cancer; Growth inhibition; Apoptosis Introduction Breast cancer is one of the main life-threatening diseases that a wo man may ha ve to face d ur i ng her li f et im e (Ang elop oul os et al., 2004). The incre asing incid ence of breast neoplasia reported over the last a few decades has led to development of new anticancer drugs, drug combinations, and chemoth era py stra teg ies by meth odical and sci enti fic exp lor atio n of eno rmo us poo l of synthetic, bio log ica l, and Toxicology and Applied Pharmacolog y 215 (2006) 168 – 178 www.elsevier.com/locate/ytaap ⁎ Correspo ndi ng authors. X. Wang is to be con tac ted at Division of Experimen tal Oncology , National Key Laborator y of Biothera py , West China Hospital, Sichuan University, Chengdu 610041, Sichuan Province, P.R. China. Fax: +86 28 851 6402 9. Y . Wei, Division of Biot hera py, Nati ona l Key Laborator y of Biothera py , West China Hospital, Sichuan University , Chengdu 610041, Sichuan Province, P.R. China. Fax: +86 28 85164059. E-mail addresses: [email protected] (X. Wang), [email protected] (Y. Wei). 0041-008X/$ - see front matter © 2006 Elsevier Inc. All rights reserved. doi:10.1016/j.taap.2006.02.004
-
Upload
riga-azoe-alriani -
Category
Documents
-
view
220 -
download
0
Transcript of Wang Et Al 2006 Dave Long Paper

8/8/2019 Wang Et Al 2006 Dave Long Paper
http://slidepdf.com/reader/full/wang-et-al-2006-dave-long-paper 1/11
Anticancer activity of litchi fruit pericarp extract against human breast cancer in vitro and in vivo
Xiujie Wang a,⁎, Shulan Yuan a , Jing Wang a , Ping Lin a , Guanjian Liu b, Yanrong Lu a ,Jie Zhang a , Wendong Wang c, Yuquan Wei d,⁎
a Division of Experimental Oncology, National Key Laboratory of Biotherapy, West China Hospital, Sichuan University,
Chengdu 610041, Sichuan Province, P.R. China b Department of Clinical Epidemiology, West China Hospital, Sichuan University, Chengdu 610041, Sichuan Province, P.R. China
c Department of Pathology of Public Health School, Sichuan University, Chengdu 610041, Sichuan Province, P.R. Chinad Division of Biotherapy, National Key Laboratory of Biotherapy, West China Hospital, Sichuan University, Chengdu 610041, Sichuan Province, P.R. China
Received 8 October 2005; revised 23 January 2006; accepted 8 February 2006Available online 23 March 2006
Abstract
Litchi fruit pericarp (LFP) extract contains significant amounts of polyphenolic compounds and exhibits powerful antioxidative activity against fat oxidation in vitro. The purpose of this study is to confirm the anticancer activity of LFP extract on human breast cancer in vitro and in vivo, andto elucidate the mechanism of its activity. Human breast cancer cells were tested in vitro for cytotoxicity, colony formation inhibition, BrdUincorporation, and gene expression profiling after treatment with LFP extract. Seven nude mice bearing human breast infiltrating duct carcinomaorthotopically were tested for its anticancer activity and expression of caspase-3 in vivo by oral administration of 0.3% (0.3 mg/ml) of LFP water-soluble crude ethanolic extract (CEE) for 10 weeks. LFP extract demonstrated a dose- and time-dependent inhibitory effect on cell growth(IC50 = 80 μg/ml), and it significantly inhibited colony formation and BrdU incorporation of human breast cancer cells. Oligonucleotide
microarray analysis identified 41(1.22%) up-regulated and 129 (3.84%) down-regulated genes after LFP water-soluble CEE treatment; the predominantly up-regulated genes were involved in various biological functions including cell cycle regulation and cell proliferation, apoptosis,signal transduction and transcriptional regulation, and extracellular matrix/adhesion molecules; and down-regulated genes were mainly associatedwith adhesion, invasion, and malignancy of cancer cells. A 40.70% tumor mass volume reduction and significant increase of casepase-3 proteinexpression were observed in vivo experiment. The findings in this study suggested that LFP extract might have potential anticancer activity on
both ER positive and negative breast cancers, which could be attributed, in part, to its DNA damage effect, proliferating inhibition and apoptosisinduction of cancer cells through up-regulation and down-regulation of multiple genes involved in cell cycle regulation and cell proliferation,apoptosis, signal transduction and transcriptional regulation, motility and invasiveness of cancer cells; ADP-ribosyltransferase (NAD+; poly(ADP-ribose) polymerase)-like 1 (ADPRTL1), Cytochrome P450, subfamily I (CYP1A1) and Hyaluronan-mediated motility receptor (HMMR)might be the main molecular targets at which LFP water-soluble CEE acted.© 2006 Elsevier Inc. All rights reserved.
Keywords: Litchi fruit pericarp extract; Anticancer activity; Breast cancer; Growth inhibition; Apoptosis
Introduction
Breast cancer is one of the main life-threatening diseasesthat a woman may have to face during her lifetime(Angelopoulos et al., 2004). The increasing incidence of
breast neoplasia reported over the last a few decades has ledto development of new anticancer drugs, drug combinations,and chemotherapy strategies by methodical and scientificexploration of enormous pool of synthetic, biological, and
Toxicology and Applied Pharmacology 215 (2006) 168–178www.elsevier.com/locate/ytaap
⁎ Corresponding authors. X. Wang is to be contacted at Division of Experimental Oncology, National Key Laboratory of Biotherapy, West ChinaHospital, Sichuan University, Chengdu 610041, Sichuan Province, P.R. China.Fax: +86 28 85164029. Y. Wei, Division of Biotherapy, National KeyLaboratory of Biotherapy, West China Hospital, Sichuan University, Chengdu610041, Sichuan Province, P.R. China. Fax: +86 28 85164059.
E-mail addresses: [email protected] (X. Wang),[email protected] (Y. Wei).
0041-008X/$ - see front matter © 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.taap.2006.02.004

8/8/2019 Wang Et Al 2006 Dave Long Paper
http://slidepdf.com/reader/full/wang-et-al-2006-dave-long-paper 2/11
natural products (Mukherjee et al., 2001). In light of thecontinuing need for effective anticancer agents, and theassociation of fruit and vegetable consumption with reducedcancer risk, edible plants are increasingly being considered assources of anticancer drugs (Ferguson et al., 2004); there is alarge amount of scientific evidence showing that fruits and
vegetables lower the risk of cancer (Chen et al., 2004), andmedicinal plants constitute the main source of new pharma-ceuticals and healthcare products, including medications for ethnoveterinary medicine (Ivanova et al., 2005). Recently,cancer chemoprevention with strategies using foods andmedicinal herbs has been regarded as one of the most visiblefields for cancer control (Jo et al., 2004); however, whether fruit, vegetable, and antioxidant micronutrient consumption isassociated with a reduction in breast cancer incidence remainsunresolved (Gaudet et al., 2004).
Epidemiological studies suggested that antioxidant supple-ments might reduce the risk of breast cancer recurrence or breast
cancer-related mortality (Fleischauer et al., 2003), and consum-ing food and beverages rich in polyphenols (e.g., catechins,flavones, and antocyanines) is associated with a lower incidenceof cancers ( Naasani et al., 2003). Experimental investigationsdemonstrated that many naturally occurring agents and plant extracts have shown antioxidant and anticancer potential in avariety of bioassay systems and animal models, havingrelevance to human disease (Aziz et al., 2003), e.g., Crudemethanolic extract (CME) from the pericarp of Garcinia
mangostana (family Guttiferae) has antiproliferative, apoptotic,and antioxidative activities against human breast cancer cell linein vitro (Primchanien et al., 2004). The antioxidant andanticancer activity of the extracts from medical plants and
herbs was associated with their components of phenoliccompounds; the major types of phenolic compounds included
phenolic acids, flavonoids, tannins, coumarins, lignans, qui-nones, stilbenes, and curcuminoids (Cai et al., 2004).
Litchi ( Litchi chinensis, Sapindaceae) is a tree that originatesfrom China and is cultivated for its sweet fruits all over theworld in warm climates (Gontier et al., 2000). Pharmacologicalstudies showed the petroleum ether extract of leaves of the plant
Litchi chinensis Gaertn possess anti-inflammatory, analgesic,and antipyretic activity without toxicity (Besra et al., 1996).
Litchi fruit pericarp (LFP) contains significant amounts of polyphenolic compounds, the principal characteristic of the
polyphenolic compounds is their ortho-diphenolic structure,which gives them high oxidability; the major components of fresh LFP extract were condensed tannins (polymeric proantho-cyanidins), epicatechin, and procyanidin A2 (Sarni-Manchadoet al., 2000). The main components of mature and prematureLFP extract were phenolic compound and flavonoids, whichexhibited powerful antioxidative activity against fat oxidation invitro (Zheng et al., 2003).
Based on the main components of LFP extract and itsantioxidant properties (Sarni-Manchado et al., 2000; Zheng et al., 2003), we hypothesized that LFP extract may haveanticancer activities against some cancer cell lines in vitro andanimal models in vivo. However, there are no such reports. Toconfirm this hypothesis, the inhibitory effect of LFP extract on
the growth of human breast cancer cells in vitro and the growthof human breast infiltrating ductal carcinoma (IDC) in vivo, andthe mechanism of its activity were investigated in thisexperimental study.
Materials and methods
Reagents. RPMI-1640 was purchased from GIBCO/BRL Invitrogen(Caithershurg, MD). Fetal bovine serum (FBS) was purchased from HuaxiBiology Institute (Chengdu, People's Republic of China). Trypsin, methylthia-zolyldiphenyl-tetrazolium bromide (MTT), and dimethyl sulfoxide (DMSO)were purchased from the Sino-American Biotechnology Company of Beijing(Beijing, People's Republic of China). 5-Bromo-2-deoxy-uridine (BrdU) was purchased from Roche (Nutley, NJ), and S-P immunohistochemical staining kit (SP9001) was purchased from Beijing Zhongshan Biological Technology Ltd.(Beijing, People's Republic of China). All of other chemicals and reagents wereobtained from Sigma (St. Louis, MO).
Preparation of extracts. Mature Baila litchi ( L. chinensis, Sapindaceae inGuangdong, China) fruit pericarp was collected, dried at room temperaturenaturally, and powdered. The powderedmaterial (100g) was extracted with 95%
ethanol (800 ml, two times) for 48 h at room temperature, blended with magneticforce stirrer continuously. The extracts were filtered and concentrated to removethe solvent at 75 °C for 4 h and freeze dried, and more than 22.0 g of crudeethanolic extract (CEE) was yielded eventually, the percentage yield was higher than 22.0%. Of the extract, 60% was water-soluble, 40% was soluble in ethanoland acetone.
Cell line and culture. Human breast cancer MCF-7 (GDC055) and MDA-MB-231(HTB-26) cell lines were obtained from China Center for Type CultureCollection (Nanjing, People's Republic of China). The cells were maintained inRPMI-1640 supplemented with 10% FBS, penicillin (100 U/ml), andstreptomycin (100 μg/ml) in a humidified atmosphere of 50 μg/ml CO2 at 37 °C.
In vitro assay for cytotoxic activity (MTT assay). The cytotoxicity of LFPwater-soluble CEE on both MCF-7 and MDA-MB-231 cells was determined by the MTT assay (Selvakumaran et al., 2003). Cells (3 × 103/well) were plated in 100 μl of medium/well in 96-well plates (Costar Corning, Rochester, NY). After incubation overnight, LFP water-soluble CEE was added invarious concentrations (20, 40, 80, 160, 320 μg/ml) without cytotoxicity tohuman normal liver cell line L-02 (reported in elsewhere); 5 wells wereincluded in each concentration. After treatment with LFP water-soluble CEEfor 1, 2, 3, 4, and 5 days, 20 μl of 5 mg/ml MTT (pH 4.7) was added per well and cultivated for another 4 h, the supernatant fluid was removed, 100 μlDMSO was added per well and shaken for 15 min. The absorbance at 570 nmwas measured with a microplate reader (Bio-Rad, Richmond, CA), usingwells without cells as blanks. All experiments were performed in triplicate.The effect the water-soluble CEE from LFP on the proliferation of human breast cancer cells was expressed as the % cytoviability, using the followingformula: %cytoviability = A570 of treated cells / A570 of controlcells × 100%.
Clonogenic survival determination. MCF-7 and MDA-MB-231 cells wereassayed for colony-forming ability by replating them in specified numbers(300–400/well) in 6-well plates and treated with 40, 80, and 160 μg/ml of LFPwater-soluble CEE, respectively. After 12 days of incubation, the cells werestained with 0.5% crystal violet in absolute ethanol and colonies with > 50 cellswere counted under dissection microscope.
BrdU incorporation in vitro. Cells were seeded onto glass coverslips at aninitial density of 4.0 × 104 cells/cm2 and allowed to grow for 12 h, and thentreated with 100 μg/ml of LFP water-soluble CEE for 48 h. The cells wereincubated with BrdU (20 μg/ml)for 12 h. At the appropriate time, the cells werefixed in methanol at −20 °C for 1–2 min, allowed to air dry, then stored at −20 °C until all coverslips were ready for processing. The cells were rehydrated
in PBS for 5 min followed by immersion in 2 N HCl for 1 h at room temperature.The cells were incubated in 0.1 M borate buffer (pH 8.5, 0.1 M boric acid,
169 X. Wang et al. / Toxicology and Applied Pharmacology 215 (2006) 168 – 178

8/8/2019 Wang Et Al 2006 Dave Long Paper
http://slidepdf.com/reader/full/wang-et-al-2006-dave-long-paper 3/11
25 mM Na2B4O7, and 75 mM NaCl) twice for 5 min each, followed by threetimes of 10 min washes in PBS. Next, cells were incubated with BrdU mousemonoclonal antibody (11B5, Zymed Laboratories, USA) at a dilution of 1:100,overnight at 4 °C, with biotylated second antibody for 20 min, and withstreptavidin/peroxidase for 30 min at room temperature. Subsequently, thesections were subjected to color reaction with 0.02% 3,3′-diaminobenzidinetetrahydrochloride containing 0.005% H2O2 in PBS (pH 7.4), and were
counterstained with hematoxylin lightly. A percentage of BrdU-labeled cellswere determined by counting several fields of 200 cells (in areas of the slidecontaining the highest labeling cells) (Thor et al., 1999).
Oligonucleotide microarray analysis. Human breast cancer MCF-7 cellswere treated with 100 μg/ml of LFP water-soluble CEE for a required time(48 h). Total RNA was extracted from the treated and untreated cells using theQiagen (Valencia City, CA) RNeasy Minikit. cDNA was synthesized from totalRNA using cDNA Synthesis System (Roche), cRNA probes were created fromcDNA with cy3-dUTP using MEGAscript T7 Kit (Ambion, Austin, TX) andhybridized to HO4 ExpressChip (Mergen, San Francisco, CA) containing 3360genes (www.mergen.ltd.com), according to the manufacturer's instructions.After a series of washes, the hybridized slides were scanned with GMS418Array Scanner (Affymatrix, Santa Clara, CA). The signal intensity of each pair of spots was quantified using ImaGene4.0 software (BioDiscovery, El Segundo,
CA) by subtracting the local regional background intensity from each spot; thetwo slides were normalized to each other by using the signal of globalnormalization gene (Glyceraldehyde-3-phosphate dehydrogenase, GAPD) provided by the manufacturer. Fold changes were calculated and expressedrelative to untreated control sample for each pair of spots; the results wereconfirmed by duplicate spots of cRNA.
RT-PCR. To verify the microarray results, the representative up-regulated(ADPRTL1, CYP1A1) and down-regulated (HMMR) genes in response to LFPwater-soluble CEE treatment were confirmed experimentally by semi-quantitative RT-PCR (Morandi et al., 2006).
The primers for each gene were designed with Primers Express software(Applied Biosystems) according to ExpressChip Gene Database of themanufacturer (Mergen, San Francisco, CA). Primer sequences and the lengthof amplified products were as follows:
ADPRTL (accession number: AF057160, product size: 477 bp)Fw 5 ′-gaagaatcagtaggcagtctcg-3 ′ Rew 5′-cctggaaatagaagggtcatcc-3 ′CYP1A1 (accession number: K03191, product size: 486 bp)Fw 5 ′-acctctacaccttcaccctcatc-3 ′ Rew 5′- agtgctccttgaccatcttctg-3′HMMR (accession number: U29343, product size: 390 bp)Fw 5 ′- gatactaccttgcctgcttcag-3′ Rew 5′- cttagccatcatacccctcatc-3′β-actin (accession number: BC013380, product size: 695 bp)Fw 5 ′-caccacaccttctacaatgagc-3 ′ Rew 5′-gtgatctccttctgcatcctgt-3 ′
Total RNA was isolated from the control and treated cells as describedabove. One microgram of total RNA was retro-transcribed to cDNA using 100Units of Rever Tra Ace (Toyobo, Japan), 50 μM Oligo(dT) primer, and 10 mÌdNTP mix.
PCR amplification was performed with Taq PCR MasterMix (Tiangen
Biotech, Beijing, People's Republic of China) using the following conditions:denaturation for 30 s at 94 °C, annealing for 30 s at 56 °C, and elongation for 45 s at 72 °C for 30 cycles.
PCR products were analyzed on 1.5% agarose gel and visualized withethidium bromide. Image were acquired and quantified using the ChemiDocTMXRS (Bio-Rad).
In vivo tumor growth inhibition study. To confirm the anticancer activity of LFP water-soluble CEE on human breast IDC,an in vivoexperiment was carriedout using an ER negative human breast IDC-xenografted animal model (Chen et al., 2003). Thirteen 6-week-old female nude mice (Experimental Animal Center,Sichuan University) were xenotransplanted with human breast infiltrating duct carcinoma F35 orthotopically (Chen et al., 2003). The procedures involvinganimals and their carewere conducted in accordance withinstitutional guidelines
for Laboratory Animal Careof Experimental Animal Center,Sichuan University.At 2 weeks, tumors reached 50 mm3 in volume, the tumor bearing mice were
randomized into two experimental groups. The treatment group animals (n = 7)were fed with 0.3% (0.3 mg/ml) of LFP water-soluble CEE through drink water ad libitum for 10 weeks, the control animals (n = 6) werewithout treatment. Micewere weighted, and tumor volume was assessed by measuring two perpendicular dimensions (long and short) using a caliper and calculated using the formula(a × b2) / 2, where a is the larger and b is the smaller dimension of the tumor (Zhang et al., 2003).
Immunohistochemical analysis of tumors for caspase-3 protein
expression. Tumor samplesfrom micewere fixed in 4% paraformaldehyde for 24 h and processed conventionally. Caspase-3 protein expressions were detectedimmunohitochemically with SP kit (Zymed); briefly, tissue sections were de- paraffinized and rehydrated through graded alcohols. Antigen retrieval was performed by microwave oven heating in 0.1 mM citrate buffer (pH 6). Then,endogenous peroxidase activity was blocked with 0.3% H2O2, and after treatment with normal goat serum, the tissue sections were incubated withcaspase-3 rabbit polyclonal antibody (Borster Biotechnology, Wuhan, People'sRepublic of China) at a dilution of 1:100, overnight at 4 °C, with biotylatedsecond antibody 20 min, and with streptavidin/peroxidase 30 min at roomtemperature. Subsequently, the sections were subjected to color reaction with0.02% 3,3′-diaminobenzidine tetrahydrochloride containing 0.005% H2O2 inPBS (pH 7.4) and were counterstained with hematoxylin lightly. Caspase-3
positive cells were determined by counting several fields of 200 cells, Caspase-3labeling index (LI) was calculated using the formula Caspase-3 labeling index(LI) = Number of activated caspase-3-positive cells / Total number of nuclei × 100% (Duan et al., 2003).
Statistical analysis. The statistical significance of difference between controland LFP-extract-treated groups was determined by one-way ANOVA followed by Tukey test for multiple comparisons. Dunnett's t tests (2-sided) wereemployed, as needed, and result was considered significant at P < 0.05.
Results
Cytotoxic activity of LFP water-soluble CEE against human
breast cancer cells
LFP water-soluble CEE showed a dose- and time-dependent inhibitory effect on the growth of MCF-7 and MDA-MB-231
breast cancer cells ( P < 0.05). IC50 was 80 μg/ml, and themaximal inhibition of cell growth (>80%) was obtained at 320 μg/ml. The result of cytotoxic activity of LFP water-solubleCEE against human breast cancer cells is shown in Fig. 1.
Inhibition of colony formation
Untreated MCF-7 and MDA-MB-231cells produced235 ± 16 and 205 ± 12 colonies, respectively; the colony
numbers of MCF-7 cells were suppressed to 145 ± 20( P < 0.05), 70 ± 13 ( P < 0.01), and 28 ± 9 ( P < 0.01) with40, 80, and 160 μg/ml of LFP water-soluble CEE treatment,respectively; the colony numbers of MDA-MB-231 cells weresuppressed to 118 ± 16 ( P < 0.05), 62 ± 11 ( P < 0.01), and 21 ± 7( P < 0.01) after the treatment of 40, 80, and 160 μg/ml of LFPwater-soluble CEE, respectively; and a dose-dependent colony-forming inhibition effect on both cell lines were observed(Fig. 2).
Inhibition of cell proliferation
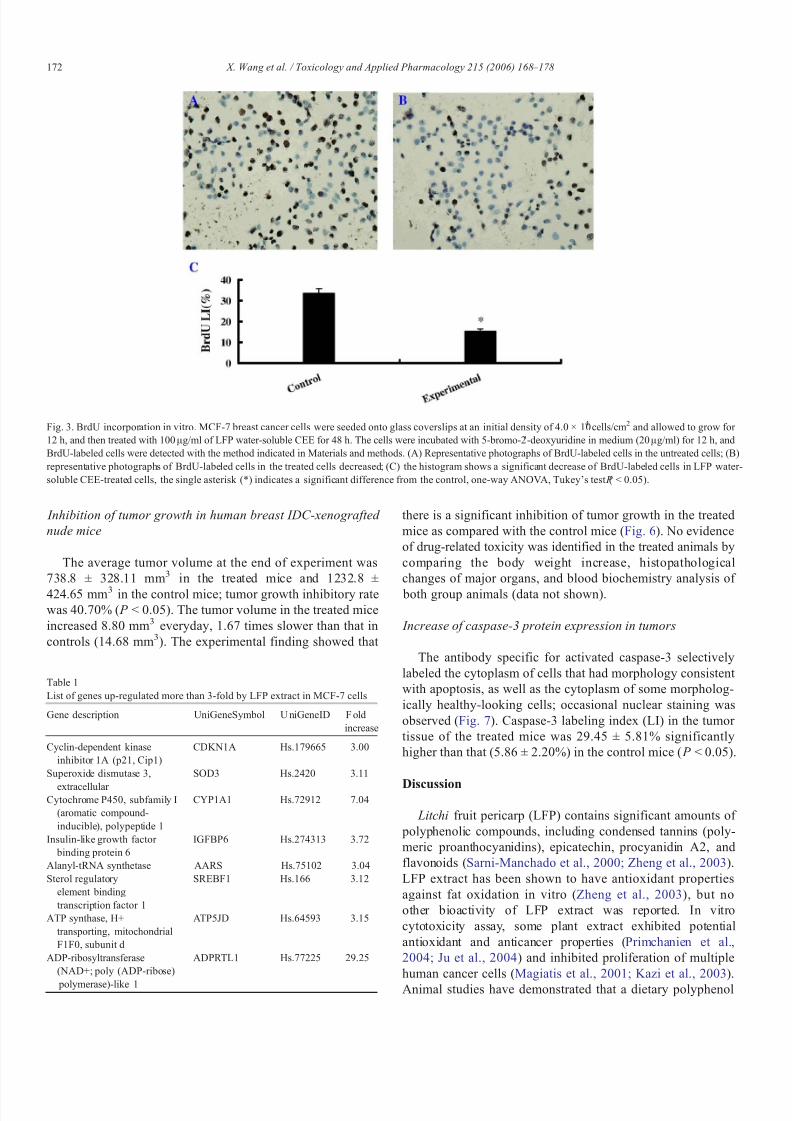
The inhibitory effect of LFP water-soluble CEE on MCF-7cells was further confirmed using BrdU incorporation into the
170 X. Wang et al. / Toxicology and Applied Pharmacology 215 (2006) 168 – 178

8/8/2019 Wang Et Al 2006 Dave Long Paper
http://slidepdf.com/reader/full/wang-et-al-2006-dave-long-paper 4/11
untreated and treated (100 μg/ml) breast cancer cells in vitro(Fig. 3). BrdU-labeled cells in LFP water-soluble CEE-treatedcells were 15.20 ± 1.30%, and lower than that in untreatedcontrols (33.50 ± 2.18%), a statistical significance was found( P < 0.05).
Change of gene expression profile
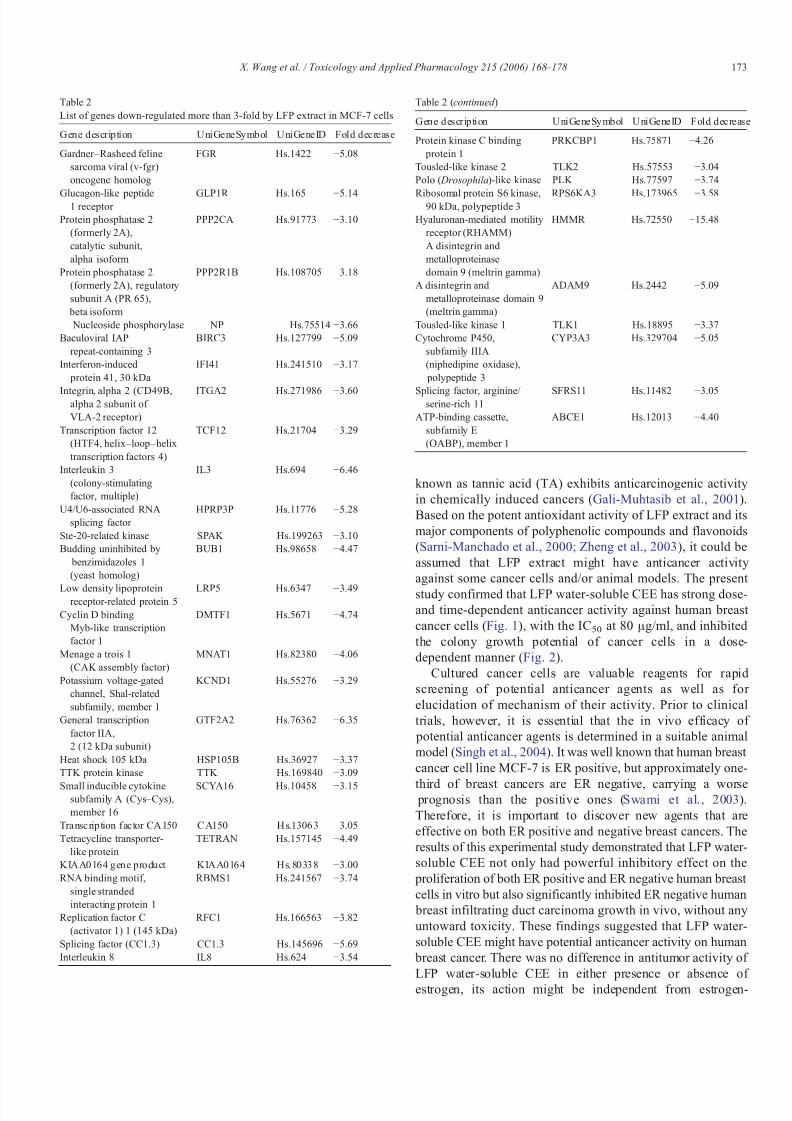
Out of 3360 genes studied, 170 differential genes wereidentified from oligonucleotide microarray in which geneexpression was increased or decreased more than 2-fold incells treated with 100 μg/ml of LFP water-soluble CEE
compared with control cells cultured under identical conditionwithout treatment; 41(1.22%) were up-regulated (Table 1) and129 (3.84%) were down-regulated (Table 2). Eight genesshowed more than 3-fold up-regulation, and 38 showed morethan 3-fold down-regulation, two genes were up-regulated for more than 5-fold, i.e., Cytochrome P450, subfamily I (aromaticcompound-inducible), polypeptide 1 (CYP1A1), and ADP-ribosyltransferase (NAD+; poly (ADP-ribose) polymerase)-like1 (ADPRTL1); their expression level increased by 7.04-foldand 29.25-fold, respectively (Table 1, Fig. 4), and 10 genesincluding Gardner –Rasheed feline sarcoma viral (v-fgr)oncogene homolog (FGR), Glucagon-like peptide 1 receptor (GLP1R), Baculoviral IAP repeat-containing 3 (BIRC3),Interleukin 3 (IL3), U4/U6-associated RNA splicing factor
(HPRP3P), General transcription factor IIA, 2 (GTF2A2),Splicing factor (CC1.3), HMMR, A disintegrin and metallo-
proteinase domain 9 (ADAM9), Cytochrome P450, subfamilyIIIA polypeptide 3 (CYP3A3), were down-regulated for morethan 5-fold (Table 2, Fig. 4). The range of fold regulation variedwidely with ADP-ribosyltransferase (ADPRTL1) exhibiting the
maximum up-regulation (29.25-fold) and Hyaluronan-mediatedmotility receptor (RHAMM) exhibiting the maximum down-regulation (−15.48-fold).
Confirmation of the representative genes by RT-PCR
To assess the reliability of the microarray results, 3representative genes with increased or decreased expressionwere analyzed by semi-quantitative RT-PCR using β-actin asthe reference gene to normalize expression data. A goodqualitative correlation was observed in the results obtained
by the two techniques, with all 3 genes tested showing
similar trends; however, quantitative differences in regulationare present (Fig. 5).
Fig. 2. Inhibition of human breast cancer cell colony formation by LFP water-soluble CEE. Cells were seeded onto 6-well plate at 300/well and weretreated with LFP water-soluble CEE at different concentration; the colonynumber was counted under dissection microscope. (A) A dose-dependent colony formation inhibition on MCF-7 breast cancer cells (ER positive); (B) adose-dependent colony formation inhibition on MDA-MB-231 (ER negative) breast cancer cells. The single asterisk (*) indicates a significant differencefrom the control ( P < 0.05); the double asterisk (**) indicates a verysignificant difference from the control ( P < 0.01), one-way ANOVA, Tukey'stest. Results are mean values ± SD of independent experiments performed intriplicate.
Fig. 1. Inhibition of human breast cancer cell growth by LFP water-soluble CEE.Cells were seeded onto 96-well plate at 3 × 103/well and were treated with LFPwater-soluble CEE at different concentrations, and percentage of cell viabilitywas determined by MTT assay after 1–5 days of treatment, respectively. (A) Adose- and time-dependent growth inhibition of MCF-7 breast cancer cells (ER positive) was observed at concentrations ranging from 20 to 320 μg/ml( P < 0.05); (B) A dose- and time-dependent growth inhibition of MDA-MB-231 breast cancer cells (ER negative) was observed at concentrations ranging from20 to 320 μg/ml ( P < 0.05). Results are mean values ± SD of independent experiments performed in triplicate.
171 X. Wang et al. / Toxicology and Applied Pharmacology 215 (2006) 168 – 178
8/8/2019 Wang Et Al 2006 Dave Long Paper
http://slidepdf.com/reader/full/wang-et-al-2006-dave-long-paper 5/11
Inhibition of tumor growth in human breast IDC-xenografted
nude mice
The average tumor volume at the end of experiment was738.8 ± 328.11 mm3 in the treated mice and 1232.8 ±
424.65 mm3 in the control mice; tumor growth inhibitory ratewas 40.70% ( P < 0.05). The tumor volume in the treated miceincreased 8.80 mm3 everyday, 1.67 times slower than that incontrols (14.68 mm3). The experimental finding showed that
there is a significant inhibition of tumor growth in the treatedmice as compared with the control mice (Fig. 6). No evidenceof drug-related toxicity was identified in the treated animals bycomparing the body weight increase, histopathologicalchanges of major organs, and blood biochemistry analysis of
both group animals (data not shown).
Increase of caspase-3 protein expression in tumors
The antibody specific for activated caspase-3 selectivelylabeled the cytoplasm of cells that had morphology consistent with apoptosis, as well as the cytoplasm of some morpholog-ically healthy-looking cells; occasional nuclear staining wasobserved (Fig. 7). Caspase-3 labeling index (LI) in the tumor tissue of the treated mice was 29.45 ± 5.81% significantlyhigher than that (5.86 ± 2.20%) in the control mice ( P < 0.05).
Discussion
Litchi fruit pericarp (LFP) contains significant amounts of polyphenolic compounds, including condensed tannins (poly-meric proanthocyanidins), epicatechin, procyanidin A2, andflavonoids (Sarni-Manchado et al., 2000; Zheng et al., 2003).LFP extract has been shown to have antioxidant propertiesagainst fat oxidation in vitro (Zheng et al., 2003), but noother bioactivity of LFP extract was reported. In vitrocytotoxicity assay, some plant extract exhibited potentialantioxidant and anticancer properties (Primchanien et al.,2004; Ju et al., 2004) and inhibited proliferation of multiplehuman cancer cells (Magiatis et al., 2001; Kazi et al., 2003).Animal studies have demonstrated that a dietary polyphenol
Fig. 3. BrdU incorporation in vitro. MCF-7 breast cancer cells were seeded onto glass coverslips at an initial density of 4.0 × 104 cells/cm2 and allowed to grow for 12 h, and then treated with 100 μg/ml of LFP water-soluble CEE for 48 h. The cells were incubated with 5-bromo-2 ′-deoxyuridine in medium (20 μg/ml) for 12 h, andBrdU-labeled cells were detected with the method indicated in Materials and methods. (A) Representative photographs of BrdU-labeled cells in the untreated cells; (B)representative photographs of BrdU-labeled cells in the treated cells decreased; (C) the histogram shows a significant decrease of BrdU-labeled cells in LFP water-soluble CEE-treated cells, the single asterisk (*) indicates a significant difference from the control, one-way ANOVA, Tukey's test ( P < 0.05).
Table 1List of genes up-regulated more than 3-fold by LFP extract in MCF-7 cells
Gene description UniGeneSymbol U niGeneID F oldincrease
Cyclin-dependent kinaseinhibitor 1A (p21, Cip1)
CDKN1A Hs.179665 3.00
Superoxide dismutase 3,
extracellular
SOD3 Hs.2420 3.11
Cytochrome P450, subfamily I(aromatic compound-inducible), polypeptide 1
CYP1A1 Hs.72912 7.04
Insulin-like growth factor binding protein 6
IGFBP6 Hs.274313 3.72
Alanyl-tRNA synthetase AARS Hs.75102 3.04Sterol regulatory
element bindingtranscription factor 1
SREBF1 Hs.166 3.12
ATP synthase, H+transporting, mitochondrialF1F0, subunit d
ATP5JD Hs.64593 3.15
ADP-ribosyltransferase(NAD+; poly (ADP-ribose)
polymerase)-like 1
ADPRTL1 Hs.77225 29.25
172 X. Wang et al. / Toxicology and Applied Pharmacology 215 (2006) 168 – 178
8/8/2019 Wang Et Al 2006 Dave Long Paper
http://slidepdf.com/reader/full/wang-et-al-2006-dave-long-paper 6/11
known as tannic acid (TA) exhibits anticarcinogenic activityin chemically induced cancers (Gali-Muhtasib et al., 2001).Based on the potent antioxidant activity of LFP extract and itsmajor components of polyphenolic compounds and flavonoids(Sarni-Manchado et al., 2000; Zheng et al., 2003), it could beassumed that LFP extract might have anticancer activityagainst some cancer cells and/or animal models. The present
study confirmed that LFP water-soluble CEE has strong dose-and time-dependent anticancer activity against human breast cancer cells (Fig. 1), with the IC50 at 80 μg/ml, and inhibitedthe colony growth potential of cancer cells in a dose-dependent manner (Fig. 2).
Cultured cancer cells are valuable reagents for rapidscreening of potential anticancer agents as well as for elucidation of mechanism of their activity. Prior to clinicaltrials, however, it is essential that the in vivo efficacy of
potential anticancer agents is determined in a suitable animalmodel (Singh et al., 2004). It was well known that human breast cancer cell line MCF-7 is ER positive, but approximately one-
third of breast cancers are ER negative, carrying a worse prognosis than the positive ones (Swami et al., 2003).Therefore, it is important to discover new agents that areeffective on both ER positive and negative breast cancers. Theresults of this experimental study demonstrated that LFP water-soluble CEE not only had powerful inhibitory effect on the
proliferation of both ER positive and ER negative human breast cells in vitro but also significantly inhibited ER negative human
breast infiltrating duct carcinoma growth in vivo, without anyuntoward toxicity. These findings suggested that LFP water-soluble CEE might have potential anticancer activity on human
breast cancer. There was no difference in antitumor activity of LFP water-soluble CEE in either presence or absence of estrogen, its action might be independent from estrogen-
Table 2List of genes down-regulated more than 3-fold by LFP extract in MCF-7 cells
Gene description UniGeneSymbol UniGeneID Fold decrease
Gardner –Rasheed felinesarcoma viral (v-fgr)oncogene homolog
FGR Hs.1422 −5.08
Glucagon-like peptide1 receptor GLP1R Hs.165 −5.14
Protein phosphatase 2(formerly 2A),catalytic subunit,alpha isoform
PPP2CA Hs.91773 −3.10
Protein phosphatase 2(formerly 2A), regulatorysubunit A (PR 65), beta isoform
PPP2R1B Hs.108705 −3.18
Nucleoside phosphorylase NP Hs.75514 −3.66Baculoviral IAP
repeat-containing 3BIRC3 Hs.127799 −5.09
Interferon-induced protein 41, 30 kDa
IFI41 Hs.241510 −3.17
Integrin, alpha 2 (CD49B,alpha 2 subunit of VLA-2 receptor)
ITGA2 Hs.271986 −3.60
Transcription factor 12(HTF4, helix–loop–helixtranscription factors 4)
TCF12 Hs.21704 −3.29
Interleukin 3(colony-stimulatingfactor, multiple)
IL3 Hs.694 −6.46
U4/U6-associated RNAsplicing factor
HPRP3P Hs.11776 −5.28
Ste-20-related kinase SPAK Hs.199263 −3.10Budding uninhibited by
benzimidazoles 1(yeast homolog)
BUB1 Hs.98658 −4.47
Low density lipoproteinreceptor-related protein 5
LRP5 Hs.6347 −3.49
Cyclin D bindingMyb-like transcriptionfactor 1
DMTF1 Hs.5671 −4.74
Menage a trois 1(CAK assembly factor)
MNAT1 Hs.82380 −4.06
Potassium voltage-gatedchannel, Shal-relatedsubfamily, member 1
KCND1 Hs.55276 −3.29
General transcriptionfactor IIA,2 (12 kDa subunit)
GTF2A2 Hs.76362 −6.35
Heat shock 105 kDa HSP105B Hs.36927 −3.37TTK protein kinase TTK Hs.169840 −3.09
Small inducible cytokinesubfamily A (Cys–Cys),member 16
SCYA16 Hs.10458 −3.15
Transcription factor CA150 CA150 Hs.13063 −3.05Tetracycline transporter-
like proteinTETRAN Hs.157145 −4.49
KIAA0164 gene product KIAA0164 Hs.80338 −3.00RNA binding motif,
single strandedinteracting protein 1
RBMS1 Hs.241567 −3.74
Replication factor C(activator 1) 1 (145 kDa)
RFC1 Hs.166563 −3.82
Splicing factor (CC1.3) CC1.3 Hs.145696 −5.69Interleukin 8 IL8 Hs.624 −3.54
Table 2 (continued )
Gene description UniGeneSymbol UniGeneID Fold decrease
Protein kinase C binding protein 1
PRKCBP1 Hs.75871 −4.26
Tousled-like kinase 2 TLK2 Hs.57553 −3.04Polo ( Drosophila)-like kinase PLK Hs.77597 −3.74
Ribosomal protein S6 kinase,90 kDa, polypeptide 3 RPS6KA3 Hs.173965 −3.58
Hyaluronan-mediated motilityreceptor (RHAMM)A disintegrin andmetalloproteinasedomain 9 (meltrin gamma)
HMMR Hs.72550 −15.48
A disintegrin andmetalloproteinase domain 9(meltrin gamma)
ADAM9 Hs.2442 −5.09
Tousled-like kinase 1 TLK1 Hs.18895 −3.37Cytochrome P450,
subfamily IIIA(niphedipine oxidase), polypeptide 3
CYP3A3 Hs.329704 −5.05
Splicing factor, arginine/ serine-rich 11 SFRS11 Hs.11482 −3.05
ATP-binding cassette,subfamily E(OABP), member 1
ABCE1 Hs.12013 −4.40
173 X. Wang et al. / Toxicology and Applied Pharmacology 215 (2006) 168 – 178

8/8/2019 Wang Et Al 2006 Dave Long Paper
http://slidepdf.com/reader/full/wang-et-al-2006-dave-long-paper 7/11
regulatory mechanism, but further more experimental investi-
gation is needed to confirm its antitumor activity against ER positive breast cancer model in vivo.
The anticancer activity of LFP water-soluble CEE might
result, at least in part, from inhibition of DNA synthesis, proliferation, as well as apoptosis induction of cancer cells.
Fig. 4. Oligonucleotide microarray analysis. Human breast cancer MCF-7 cells were treated with 100 μg/ml of LFP water-soluble CEE for 72 h, total RNA wasextracted from the treated and untreated cells, cRNA probes were created, and hybridized to HO4 ExpressChip, the hybridized slides were scanned with GMS418Array Scanner. (A) Gene expression profiling of the untreated cells; (B) gene expression profiling of LFP water-soluble CEE-treated cells; (C) the histogram shows theup-regulation and down-regulation of more than 5-fold.
Fig. 5. Comparison of microarray results with those determined by RT-PCR. (A) Gel image of 3 representative genes confirmed by RT-PCR. The amplified fragmentswere quantified using β-actin for the normalization. (B) The correlation coefficient is shown in the graph.
174 X. Wang et al. / Toxicology and Applied Pharmacology 215 (2006) 168 – 178

8/8/2019 Wang Et Al 2006 Dave Long Paper
http://slidepdf.com/reader/full/wang-et-al-2006-dave-long-paper 8/11
Inhibition of DNA synthesis and proliferation of cancer cellswere verified by its ability to reduce BrdU incorporation intocancer cells that correlates with decreased cell proliferation
(Milosevic et al., 2002) after treatment with LFP water-soluble
CEE, BrdU-labeled cells in the treated cells reduced signifi-cantly ( P < 0.05), which indicated that LFP water-soluble CEEinhibited proliferation of cancer cells. Apoptosis induction was
additionally determined by increased caspase-3 protein
Fig. 6. Inhibition of tumor growth in nude mice xenograted with human breast IDC by LFP water-soluble CEE. Two weeks after orthotopical inoculation of human breast IDC, the mice were randomly divided into two groups, six mice in control and seven mice in treatment group. In the treated mice, LFP water-soluble CEE(0.3 mg/ml) was given through drink water ad libitumfor 10 weeks. A significant reduction of tumor volume was observed in the treated mice. (A) Tumorbearingnudemice of the untreated group (up row) and the treated group (down row); (B) tumor masses of the untreated group (up row) and the treated group (down row); (C) thehistogram shows that there was a significant difference of tumor volumes between the untreated and treated mice, the single asterisk (*) indicates a significant difference from the control, one-way ANOVA, Tukey's test ( P < 0.05).
Fig. 7. Increase of caspase-3 protein expression by LFP water-soluble CEE. Caspase-3 protein expressions of tumor tissues derived from the untreated and treated micewere detected through immunohistochemical staining. (A) Representative photographs of caspase-3 protein expression of the untreated mouse tumor; (B) the treated
mouse tumor; (C) the histogram shows that there was a significant increase of caspase-3 protein expression of LFP water-soluble CEE-treated mouse tumors, the singleasterisk (*) indicates a significant difference from the control, one-way ANOVA, Tukey's test ( P < 0.05).
175 X. Wang et al. / Toxicology and Applied Pharmacology 215 (2006) 168 – 178

8/8/2019 Wang Et Al 2006 Dave Long Paper
http://slidepdf.com/reader/full/wang-et-al-2006-dave-long-paper 9/11
expression in tumors of the animals treated with LFP water-soluble CEE.
The anticancer mechanism of LFP extract was further investigated by oligonucleotide microarray; it was demonstrat-ed that non-toxic dose of LFP water-soluble CEE affected thegene expression profile of cancer cells by up-regulation of 41
genes (1.22%) and down-regulation of 129 genes (3.84%),involved in various biological functions including cell cycleregulation and cell proliferation, apoptosis, signal transductionand transcriptional regulation, and extracellular matrix/adhesionmolecules, etc. The range of fold regulation varied widely withADPRTL1 exhibiting the maximum up-regulation (29.25-fold)and RHAMM exhibiting the maximum down-regulation(−15.48-fold).
The predominantly up- and down-regulated genes weremainly associated with apoptosis, cell–cell and cell–matrixinteractions, motility, and invasiveness of cancer cells.
CYP1A1, encoding Cytochrome P450, subfamily I, up-
regulation is paralleled with cytotoxicity of aminoflavone insensitive breast tumor cells (Loaiza-Perez et al., 2004a, b),resveratrol-induced differentiation and apoptosis of medullo-
blastoma cells (Liu et al., 2004), antiproliferative activity andapoptosis induction of aminoflavone analogue (AF) of humantumor renal cell carcinoma lines (Loaiza-Perez et al., 2004a, b),and CYP1A1 activation led to cytotoxicity of anticancer agent through DNA damage-induced apoptosis toxicity (Monks et al.,2003). ADPRT gene encodes a zinc-finger DNA-binding
protein, poly (ADP-ribose) polymerase-1 (PARP-1), that modifies various nuclear proteins by poly (ADP-ribosyl)ationand functions as a key enzyme in the base excision repair
pathway; altered ADPRT/PARP-1 enzyme function was
associated with response to oxidative damage (Griesenbeck et al., 1997; Lockett et al., 2004), during almost all forms of apoptosis, PARP is activated by caspases (Hatip-Al-Khatib et al., 2004), and inhibition of ADP-ribosyltransferase significant-ly reduced its ability to induce apoptosis of cancer cells(Hatip-Al-Khatib et al., 2004; Takamura-Enya et al., 2001). BIRC,encoding apoptosis inhibitory protein was associated apoptosisand cell cycles (Takita et al., 2004). Inhibitors of apoptosis(IAPs) antagonize cell death and regulate the cell cycle (Dong et al., 2002); decrease of c-IAP2 expression led to promotion of apoptosis ( Nishihara et al., 2003). In the present study, theexpressions of ADPRTL1 and CYP1A1 in LFP water-soluble
CEE-treated breast cancer cells increased and the expression of BIRC3 decreased significantly; it could be inferred that LFPwater-soluble CEE inhibited proliferation and induced apopto-sis of breast cancer cells mainly through up-regulationexpressions of CYP1A1 and ADPRTL1, and down-regulationof BIRC genes.
For other predominantly down-regulated genes, ADAMencoding a disintegrin and metalloprotease family contributes toregulation of the cell–cell and cell–matrix interactions that arecritical determinants of malignancy; ADAM9 overexpressionenhances cell adhesion and invasion of non-small cell lungcancer cells (Shintani et al., 2004), and is associated with poor differentiation and shortened survival of pancreatic cancer (Grutzmann et al., 2004). HMMR encoding the receptor for
hyaluronan mediated motility (RHAMM), a hyaluronan (HA) binding protein, has been shown to play an important role in themotility and invasiveness of malignant cells (Kuwabara et al.,2004); expression and splicing of RHAMM are important molecular determinants of disease severity in multiple myeloma(Maxwell et al., 2004); and RHAMM blockade might be a
potential therapeutic target for difficult-to-treat neoplasm (Tolget al., 2003). The decreased expressions of ADAM9 (−5.09-fold) and RHAMM (−15.48-fold) in the treated cancer cells(Table 2, Fig. 4) suggested that LFP water-soluble CEE couldinhibit the motility and invasiveness of cancer cells, anddecreased their malignancy (Shintani et al., 2004; Grutzmann et al., 2004; Kuwabara et al., 2004; Maxwell et al., 2004; Tolg et al., 2003).
The most predominantly up-regulated gene after LFP water-soluble CEE treatment was ADPRTL1 (29.25-fold) which wasassociated with DNA repair, damage, and cell apoptosis(Griesenbeck et al., 1997; Lockett et al., 2004; Hatip-Al-Khatib
et al., 2004; Takamura-Enya et al., 2001), and the most predominantly down-regulated gene HMMR (−15.48-fold) wasassociated with the motility and invasiveness of cancer cells.The findings in this study showed LFP water-soluble CEEresulting in DNA damage, proliferation inhibition and apoptosisof cancer cells through up-regulation of ADPRTL1 expression,and inhibition of the motility and invasiveness of cancer cellsthrough down-regulation of HMMR expression. ADPRTL1 andHMMR might be the main drug target at which LFP water-soluble CEE acted, but this needs to be confirmed in normaland/or other cancer cells.
Although there was qualitative agreement between thecDNA microassay and PCR assay, with 90% of genes tested
showing similar trend reported, false positive results werecommon in cDNA microarray (Swami et al., 2003). Therefore,3 representative genes in response to LFP water-soluble CEEtreatment were confirmed by semi-quantitative RT-PCR. Agood qualitative correlation was observed in the results obtained
by microarray and RT-PCR, but quantitative differences exist.As cDNA microarray tended to have greater interassay variationthan quantitative RT-PCR (Swami et al., 2003), the existence of quantitative differences is inevitable, even reverse correlation is
present (Swami et al., 2003; Morandi et al., 2006). The findingsin this work could show the trend of gene express profile of cancer cells treated with LFP water-soluble CEE.
In conclusion, the potential anticancer activity of LFP extract against human breast cancer was investigated in this experi-mental study, for the first time. LFP extract exhibited a stronginhibitory effect on the proliferation of both ER positive andnegative breast cancer cells in vitro and inhibited growth of ER negative breast cancer in vivo. The results of this experimentalstudy suggested that Litchi fruit pericarp contains someconstituents, which would be useful for anticancer drugdiscovery. The anticancer activity of LFP extract could beattributed, in part to its proliferating inhibition and apoptosisinduction of cancer cells through up-regulation (CYP1A1,ADPRTL1) and down-regulation (BIRC3, ADAM9, HMMR)of multiple genes, which are involved in the cell cycleregulation and cell proliferation, apoptosis, signal transduction
176 X. Wang et al. / Toxicology and Applied Pharmacology 215 (2006) 168 – 178

8/8/2019 Wang Et Al 2006 Dave Long Paper
http://slidepdf.com/reader/full/wang-et-al-2006-dave-long-paper 10/11
and transcriptional regulation, motility, and invasiveness of cancer cells. ADPRTL1, CYP1A1, and HMMR might be themain molecular targets at which LFP water-soluble CEE acted.
References
Angelopoulos, N., Barbounis, V., Livadas, S., Kaltsas, D., Tolis, G., 2004.Effects of estrogen deprivation due to breast cancer treatment. Endocr. Relat.Cancer 11, 523–535.
Aziz, M.H., Kumae, R., Ahmad, N., 2003. Cancer chemoprevention byresveratrol: in vitro and in vivo studies and the underlying mechanisms(review). Int. J. Oncol. 23, 17–28.
Besra, S.E., Sharma, R.M., Gomes, A., 1996. Antiinflammatory effect of petroleum ether extract of leaves of Litchi chinensis Gaertn (Sapindaceae).J. Ethnopharmacol. 54, 1–6.
Cai, Y., Luo, Q., Sun, M., Corke, H., 2004. Antioxidant activity and phenoliccompounds of 112 traditional Chinese medicinal plants associated withanticancer. Life Sci. 74, 2157–2184.
Chen, D., Zhao, Y., Bai, S., Shi, Z., Zhang, J., 2003. Effect of ginsenoside Rg3on the progression of orthotopically xenotransplanted human breast cancer
in nude mice and its mechanism. Sichuan Daxue Xuebao Yixue Ban 34,546–548.
Chen, M.S., Chen, D., Dou, Q.P., 2004. Inhibition of proteasome activity byvarious fruits and vegetables is associated with cancer cell death. In Vivo 18,73–80.
Dong, Z., Nishiyama, J., Yi, X., Venkatachalam, M.A., Denton, M., Gu, S., Li,S., Qiang, M., 2002. Gene promoter of apoptosis inhibitory protein IAP2:identification of enhancer elements and activation by severe hypoxia.Biochem. J. 364, 413–421.
Duan, W.R., Garner, D.S., Williams, S.D., Funckes-Shippy, C.L., Spath, I.S.,Blomme, E.A.G., 2003. Comparison of immunohistochemistry for activatedcaspase-3 and cleaved cytokeratin 18 with the TUNEL method for quantification of apoptosis in histological sections of PC-3 subcutaneousxenografts. J. Pathol. 199, 221–228.
Ferguson, P.J., Kurowska, E., Freeman, D.J., Chambers, A.F., Koropatnick,
D.J., 2004. A flavonoid fraction from cranberry extract inhibits proliferation of human tumor cell lines. J. Nutr. 134, 1529–1535.
Fleischauer, A.T., Simonsen, N., Arab, L., 2003. Antioxidant supplements andrisk of breast cancer recurrence and breast cancer-related mortality among postmenopausal women. Nutr. Cancer 46, 15–22.
Gali-Muhtasib, H.U., Younes, I.H., Karchesy, J.J., el-Sabban, M.E., 2001. Plant tannins inhibit the induction of aberrant crypt foci and colonic tumors by 1,2-dimethylhydrazine in mice. Nutr. Cancer 39, 108–116.
Gaudet, M.M., Britton, J.A., Kabat, G.C., Steck-Scott, S., Eng, S.M.,Teitelbaum, S.L., Terry, M.B., Neugut, A.I., Gammon, M.D., 2004. Fruits,vegetables, and micronutrients in relation to breast cancer modified bymenopause and hormone receptor status. Cancer Epidemiol. Biomarkers.Prev. 13, 1485–1494.
Gontier, E., Boussouel, N., Terrasse, C., Jannoyer, M., Menard, M.,Thomasset, B., Bourgaud, F., 2000. Litchi chinensis fatty acid diversity:
occurrence of the unusual cyclopropanoic fatty acids. Biochem. Soc.Trans. 28, 578–580.
Griesenbeck, J., Oei, S.L., Mayer-Kuckuk, P., Ziegler, M., Buchlow, G.,Schweiger, M., 1997. Protein– protein interaction of the human poly(ADP-ribosyl)transferase depends on the functional state of the enzyme.Biochemistry 36, 7297–7304.
Grutzmann, R., Luttges, J., Sipos, B., Ammerpohl, O., Dobrowolski, F.,Alldinger, I., Kersting, S., Ockert, D., Koch, R., Kalthoff, H., Schackert,H.K., Saeger, H.D., Kloppel, G., Pilarsky, C., 2004. ADAM9 expressionin pancreatic cancer is associated with tumour type and is a prognosticfactor in ductal adenocarcinoma. Br. J. Cancer 90, 1053–1058.
Hatip-Al-Khatib, I., Iwasaki, K., Chung, E.H., Egashiea, N., Mishima, K.,Fujiwara, M., 2004. Inhibition of poly (ADP-ribose) polymerase andcaspase-3, but not caspase-1, prevents apoptosis and improves spatialmemory of rats with twice-repeated cerebral ischemia. Life Sci. 75,
1967–1978.
Ivanova, D., Gerova, D., Chervenkov, T., Yankov, T., 2005. Polyphenols andantioxidant capacity of Bulgarian medicinal plants. J. Ethnopharmacol. 96,145–150.
Jo, E.H., Hong, H.D., Ahn, N.C., Jung, J.W., Yang, S.R., Park, J.S., Kim, S.H.,Lee, Y.S., Kang, K.S., 2004. Modulations of the Bcl-2/Bax family wereinvolved in the chemopreventive effects of licorice root (Glycyrrhiza
uralensis Fisch) in MCF-7 human breast cancer cell. J. Agric. Food Chem.
52, 1715–1719.Ju, E.M., Lee, S.E., Hwang, H.J., Kim, J.H., 2004. Antioxidant and anticancer activity of extract from Betula platyphylla var. japonica. Life Sci. 74,1013–1026.
Kazi, A., Urbizu, D.A., Kuhn, D.J., Acebo, A.L., Jackson, E.R., Greenfelder,G.P., Kumar, N.B., Dou, Q.P., 2003. A natural musaceas plant extract inhibits proteasome activity and induces apoptosis selectively in humantumor and transformed, but not normal and non-transformed, cells. Int. J.Mol. Med. 12, 879–887.
Kuwabara, H., Yoneda, M., Nagai, M., Hayasaki, H., Mori, H., 2004. A new polyclonal antibody that recognizes a human receptor for hyaluronanmediated motility. Cancer Lett. 210, 73–80.
Liu, J., Wang, Q., Wu, D.C., Wang, X.W., Sun, Y., Chen, X.Y., Zhang, K.L., Li,H., 2004. Differential regulation of CYP1A1 and CYP1B1 expression inresveratrol-treated human medulloblastoma cells. Neurosci. Lett. 363,
257–261.Loaiza-Perez, A.I., Kenney, S., Boswell, J., Hollingshead, M., Alley, M.C.,
Hose, C., Ciolino, H.P., Yeh, G.C., Trepel, J.B., Vistica, D.T., Sausville,E.A., 2004a. Aryl hydrocarbon receptor activation of an antitumor aminoflavone: basis of selective toxicity for MCF-7 breast tumor cells.Mol. Cancer Ther. 3, 715–725.
Loaiza-Perez, A.I., Kenney, S., Boswell, J., Hollingshead, M., Hose, C.,Linehan, W.M., Worrell, R., Rubinstein, L., Sausville, E.A., Vistica, D.T.,2004b. Sensitivity of renal cell carcinoma to aminoflavone: role of CYP1A1.J. Urol. 171, 1688–1697.
Lockett, K.L., Hall, M.C., Xu, J., Zheng, S.L., Berwick, M., Chuang, S.C.,Clark, P.E., Cramer, S.D., Lohman, K., Hu, J.J., 2004. The ADPRT V762Agenetic variant contributes to prostate cancer susceptibility and deficient enzyme function. Cancer Res. 64, 6344–6348.
Magiatis, P., Pratsinis, H., Kalpoutzakis, E., Konstantinidou, A., Davaris, P.,
Skaltsounis, A.L., 2001. Hydrolyzable tannins, the active constituents of three Greek Cytinus taxa against several tumor cell lines. Biol. Pharm. Bull.24, 707–709.
Maxwell, C.A., Rasmussen, E., Zhan, F., Keats, J.J., Adamia, S., Strachan, E.,Crainie, M., Walker, R., Belch, A.R., Pilarski, L.M., Barlogie, B.,Shaughnessy Jr., J., Reiman, T., 2004. RHAMM expression and isoform balance predict aggressive disease and poor survival in multiple myeloma.Blood 104, 1151–1158.
Milosevic, J., Kanazir, S., Medic-Mijacevic, L., Pejanovic, V., Stokic, Z.,Konjevic, G., Rakic, L., Ruzdijic, S., 2002. Sulfinosine-induced cell growthinhibition and apoptosis in human lung carcinomas in vitro. Invest. NewDrugs 20, 229–240.
Monks, A., Harris, E., Hose, C., Connelly, J., Sausville, E.A., 2003. Genotoxic profiling of MCF-7 breast cancer cell line elucidates gene expressionmodifications underlying toxicity of the anticancer drug 2-(4-amino-3-
methylphenyl)-5-fluorobenzothiazole. Mol. Pharmacol. 63, 766–772.Morandi, E., Zingaretti, C., Chiozzotto, D., Severini, C., Semeria, A., Horn, W.,
Vaccari, M., Serra, R., Silingardi, P., Colacci, A., 2006. A cDNA-microarrayanalysis of camptothecin resistance in glioblastoma cell lines. Cancer Lett.231, 74–86.
Mukherjee, A.K., Basu, S., Sarkar, N., Ghosh, A.C., 2001. Advances in cancer therapy with plant based natural products. Curr. Med. Chem. 8, 1467–1486.
Naasani, I., Oh-Hashi, F., Oh-Hara, T., Feng, W.Y., Johnston, J., Chan, K.,Tsuruo, T., 2003. Blocking telomerase by dietary polyphenols is a major mechanism for limiting the growth of human cancer cells in vitro and invivo. Cancer Res. 63, 824–830.
Nishihara, H., Kizaka-Kondoh, S., Insel, P.A., Eckmann, L., 2003. Inhibition of apoptosis in normal and transformed intestinal epithelial cells by cAMPthrough induction of inhibitor of apoptosis protein (IAP)-2. Proc. Natl.Acad. Sci. U.S.A. 100, 8921–8926.
Primchanien, M., Nuttavut, K., Sineenart, K., Omboon, L., Narongchai, P.,
177 X. Wang et al. / Toxicology and Applied Pharmacology 215 (2006) 168 – 178

8/8/2019 Wang Et Al 2006 Dave Long Paper
http://slidepdf.com/reader/full/wang-et-al-2006-dave-long-paper 11/11
Neelobol, N., 2004. Antiproliferation, antioxidation and induction of apoptosis by Garcinia mangostana (mangosteen) on SKBR3 human breast cancer cell line. J. Ethnopharmacol. 92, 161–166.
Sarni-Manchado, P., Le Roux, E., Le Guerneve, C., Lozano, Y., Cheynier, V.,2000. Phenolic composition of litchi fruit pericarp. Agric. Food Chem. 48,5995–6002.
Selvakumaran, M., Pisarcik, D.A., Bao, R., Yeung, A.T., Hamilton, T.C., 2003.
Enhanced cisplatin cytotoxicity by disturbing the nucleotide excision repair pathway in ovarian cancer cell lines. Cancer Res. 63, 1311–1316.Shintani, Y., Higashiyama, S., Ohta, M., Hirabayashi, H., Yamamoto,
S., Yoshimasu, T., Matsuda, H., Matsuura, N., 2004. Overexpressionof ADAM9 in non-small cell lung cancer correlates with brain metastasis.Cancer Res. 64, 4190–4196.
Singh, A.V., Xiao, D., Lew, K.L., Dhir, R., Singh, S.V., 2004. Sulforaphaneinduces caspase-mediated apoptosis in cultured PC-3 human prostate cancer cells and retards growth of PC-3 xenografts in vivo. Carcinogenesis 25,83–90.
Swami, S., Raghavachari, N., Muller, U.R., Bao, Y.P., Feldman, D., 2003.Vitamin D growth inhibition of breast cancer cells: gene expression patternsassessed by cDNA microarray. Breast Cancer Res. Treat. 80, 49–62.
Takamura-Enya, T., Watanabe, M., Totsuka, Y., Kanazawa, T., Matsushima-Hibiya, Y., Koyama, K., Sugimura, T., Wakabayashi, K., 2001. Mono(ADP-
ribosyl)ation of 2′-deoxyguanosine residue in DNA by an apoptosis-inducing protein, pierisin-1, from cabbage butterfly. Proc. Natl. Acad. Sci.U.S.A. 98, 12414–12419.
Takita, J., Ishii, M., Tsutsumi, S., Tanaka, Y., Kato, K., Toyoda, Y., Hanada, R.,Yamamoto, K., Hayashi, Y., Aburatani, H., 2004. Gene expression profilingand identification of novel prognostic marker genes in neuroblastoma.Genes Chromosomes Cancer 40, 120–132.
Thor, A.D., Liu, S., Moore II, D.H., Edgerton, S.M., 1999. Comparisonof mitotic index, in vitro bromodeoxyuridine labeling, and MIB-1assays to quantitate proliferation in breast cancer. J. Clin. Oncol. 17,470–477.
Tolg, C., Poon, R., Fodde, R., Turley, E.A., Alman, B.A., 2003. Genetic deletionof receptor for hyaluronan-mediated motility (Rhamm) attenuates theformation of aggressive fibromatosis (desmoid tumor). Oncogene 22,6873–6882.
Zhang, D.Y., Wu, J., Ye, F., Xue, L., Jiang, S., Yi, J., Zhang, W., Wei, H., Sung,M., Wang, W., Li, X., 2003. Inhibition of cancer cell proliferation and prostaglandin E2 synthesis by Scutellaria Baicalensis. Cancer Res. 63,4037–4043.
Zheng, G., Yi, Z., Zhang, J., Zhong, D., 2003. Studies on the antioxidative effect of extract from mature and premature litchi pericarp. Nat. Prod. Res. Dev.15, 341–344.
178 X. Wang et al. / Toxicology and Applied Pharmacology 215 (2006) 168 – 178


















