
WALNUT ROOTSTOCK TRANSFORMATION AND …walnutresearch.ucdavis.edu/2003/2003_97.pdf · WALNUT...
14
WALNUT ROOTSTOCK TRANSFORMATION AND REGENERATION FROM VEGETATIVE TISSUE David Tricoli, Monica T. Britton, Sandra L. Uratsu, Charles A. Leslie, Gale H. McGranahan, Wesley P. Hackett, Edwin L. Civerolo, and Abhaya M. Dandekar ABSTRACT The primary goal of this proposal is to develop novel regeneration technologies needed to engineer walnut rootstocks for pest and disease resistance. The productivity of the walnut industry of California is dependent the availability of superior rootstocks to combat soil disease and pest problems. Individual selections displaying resistance to either Phytophthora crown and root rots or to root lesion nematode, Pratylenchus vulnus, have been identified and can be clonally propagated but these selections are susceptible to crown gall. Rootstocks with multiple resistances are clearly desirable. Walnut transformation depends on the use of somatic embryos. Somatic embryos are readily obtained from immature, and therefore uncharacterized, seed but not from vegetative or other mature tissues. This precludes inserting any genes into a rootstock exhibiting another superior trait if the trait was identified in a seedling or mature tree. Walnuts have a long generation time and the most promising rootstocks, hybrids of English walnut x black walnut, are sterile or nearly sterile, factors that severely impede developing multiple resistances by breeding. The recent discoveries of genes that stimulate regeneration provide a unique avenue to develop cultivar independent transformation for walnut. Oncogene silencing gene constructs previously developed by us will be introduced into walnut rootstocks that are resistant to Phytophthora and/or root lesion nematode. The crown gall resistant lines will be propagated and introduced into the field in two locations in California for further testing of horticultural traits and disease and pest resistance with the goal of developing a new rootstock variety that will be resistant to major soil based disease and pests of significance for the California walnut industry. PROJECT OBJECTIVES 1. To test the efficacy of the XVE inducer system in walnut somatic embryos and track expression at various stages of the regenerated plant. 2. To determine the capability of the ESR1 and PGA6 genes, under the control of a constitutive promoter and the XVE inducible system, to stimulate the production of transgenic somatic embryos or organogenic shoots for a range of rootstock germplasm. 3. To determine the regeneration capability of vegetative tissue collected from in vitro shoots established from embryos transformed with the ESR1 and PGA6 genes and determine if this capability to regenerate is maintained as the tree ages. 4. To determine the regeneration capability of vegetative tissues collected from in vitro shoots established from greenhouse and field grown elite trees when transformed with the ESR1 and PGA6 genes under the control of the XVE inducer system. 5. To determine the stability of the silencing constructs.
Transcript of WALNUT ROOTSTOCK TRANSFORMATION AND …walnutresearch.ucdavis.edu/2003/2003_97.pdf · WALNUT...

WALNUT ROOTSTOCK TRANSFORMATION AND REGENERATION FROM VEGETATIVE TISSUE David Tricoli, Monica T. Britton, Sandra L. Uratsu, Charles A. Leslie, Gale H. McGranahan, Wesley P. Hackett, Edwin L. Civerolo, and Abhaya M. Dandekar ABSTRACT The primary goal of this proposal is to develop novel regeneration technologies needed to engineer walnut rootstocks for pest and disease resistance. The productivity of the walnut industry of California is dependent the availability of superior rootstocks to combat soil disease and pest problems. Individual selections displaying resistance to either Phytophthora crown and root rots or to root lesion nematode, Pratylenchus vulnus, have been identified and can be clonally propagated but these selections are susceptible to crown gall. Rootstocks with multiple resistances are clearly desirable. Walnut transformation depends on the use of somatic embryos. Somatic embryos are readily obtained from immature, and therefore uncharacterized, seed but not from vegetative or other mature tissues. This precludes inserting any genes into a rootstock exhibiting another superior trait if the trait was identified in a seedling or mature tree. Walnuts have a long generation time and the most promising rootstocks, hybrids of English walnut x black walnut, are sterile or nearly sterile, factors that severely impede developing multiple resistances by breeding. The recent discoveries of genes that stimulate regeneration provide a unique avenue to develop cultivar independent transformation for walnut. Oncogene silencing gene constructs previously developed by us will be introduced into walnut rootstocks that are resistant to Phytophthora and/or root lesion nematode. The crown gall resistant lines will be propagated and introduced into the field in two locations in California for further testing of horticultural traits and disease and pest resistance with the goal of developing a new rootstock variety that will be resistant to major soil based disease and pests of significance for the California walnut industry. PROJECT OBJECTIVES
1. To test the efficacy of the XVE inducer system in walnut somatic embryos and track
expression at various stages of the regenerated plant. 2. To determine the capability of the ESR1 and PGA6 genes, under the control of a constitutive
promoter and the XVE inducible system, to stimulate the production of transgenic somatic embryos or organogenic shoots for a range of rootstock germplasm.
3. To determine the regeneration capability of vegetative tissue collected from in vitro shoots established from embryos transformed with the ESR1 and PGA6 genes and determine if this capability to regenerate is maintained as the tree ages.
4. To determine the regeneration capability of vegetative tissues collected from in vitro shoots established from greenhouse and field grown elite trees when transformed with the ESR1 and PGA6 genes under the control of the XVE inducer system.
5. To determine the stability of the silencing constructs.

PROCEDURES Establishing Agrobacterium cultures for transformation: Forty 40 µl of the glycerol stock of Agrobacterium tumefaciens strain EHA 105 containing the binary vector pER10-GFP, pDU03-PGA6 or pDU03-ESR1 (see below) was used to inoculate 20 ml of MGL medium (pH 7.0) containing 10 mg/l rif, 5 mg/l tetracycline and 20 mg/l spectinomycin sulfate, and incubated overnight at 28°C at 250 rpms. Five ml of the overnight growth was removed and transferred to 15 ml of TY medium (pH 5.5) containing 10 mg/l rif, 5mg/l tetracycline and 20 mg/l spectinomycin sulfate, 200 µM acetosyringone. The culture was incubated overnight at 28°C at 250 rpms. The following morning a dilution was made by adding 1.5 ml of the overnight culture to 20 ml of TY medium (pH 5.5), containing 200 µM acetosyringone to an O.D at 600 nm of 0.075 to 0.20. 1. Efficacy of the XVE inducer system in walnut somatic embryos.
Construction of binary vector pER10-GFP: We have constructed the binary vector pER10-GFP to test the utility of the XVE system in walnut (Fig. 1). The base vector pER10 was obtained from Dr. Nam-Hai Chua. This particular binary vector is identical to pER8 described in their publication (Zuo et al., 2000), except that the selectable marker in pER10 is kanamycin resistance whereas in pER8 it is hygromycin resistance. We have introduced the GFP coding sequences into the MCS (multiple cloning site) region of pER10 downstream from the chemically inducible cis acting sequences (LexA) creating the binary vector pER10-GFP (Fig. 1). This binary has been introduced into disarmed strains of Agrobacterium tumefaciens EHA105 to create a functional plant transformation system. Plant transformation with binary vector pER10-GFP a. Tobacco: Nicotiana tabacum c.v. SR1 leaf pieces were inoculated with Agrobacterium containing the plasmid pER10-GFP and cultured on MS medium (Murashige and Skoog 1962) supplemented with 30 g/l sucrose, 80 mg/l adenine sulfate, 2.0 mg/l kinetin, 2.0 mg/l IAA, 8g/l TC agar, pH 5.8 (MSKI) plus 200 µM acetosyringone. Cultures were incubated at 23°C in the dark for 48 to 72 hours. Leaves were then transferred to MSKI medium containing 400 mg/l carbenicillin, and 250 mg/l kanamycin sulfate. Cultures were maintained at 26°C and a 16 hour photoperiod. Tissue was subcultured to fresh medium every 21 days. Kanamycin resistant shoots were harvested and transferred to fresh medium and multiple shoots were produced. Clones from transgenic shoots were transferred to fresh medium with or without 5 µM 17-β-estradiol. After 24 hours leaf tissue was harvested and observed for fluorescence under UV light using a Leica Stereomicroscope and GFP filter. b. Walnut: Somatic embryos of Chandler and Paradox were sub-cultured bi-weekly on agar solidified DKW basal salts containing 20 g/L sucrose, MS vitamins and 1 ml/liter PPM and solidified with 8g/l TC agar (Phtyotechnology Inc.). For transformation, actively growing somatic embryos were harvested and placed in a 100 x 15 mm petri dish with 20 ml of the dilute Agrobacterium culture containing the binary vector pER10-GFP and allowed to soak for 5-10 minutes. Embryos were removed and plated onto DKW basal medium containing 20 g/L sucrose, MS vitamins and 200 µM acetosyringone (WCM). Cultures were incubated at 23°C in

the dark for 48 to 72 hours. Embryos were then transferred to DKW basal medium containing 20 g/L sucrose, MS vitamins, 500 mg/l cefotaxime, 100 mg/l kanamycin sulfate (WIM) with or without 5 µM 17-β-estradiol. Cultures were maintained in the dark at 26°C. Cultures were subcultured onto fresh medium every 7-14 days. Sixty days post transformation, embryos transformed with pER10-GFP were observed under UV light using a Leica Stereomicroscope and GFP filter. GFP expressing embryos were harvested and half were transferred to WIM minus 5 µM 17-β-estradiol and half to WIM plus 5 µM 17-β-estradiol and cultured for further observation. 2. Capability of the ESR1 and PGA6 genes, under the control of the XVE inducible system,
to stimulate the production of transgenic somatic embryos or organogenic shoots for a range of rootstock germplasm.
Construction of the binary vectors pER10-PGA6 and pER10-ESR1: Shown in Fig 2 are the two binary vectors that have been constructed to investigate the effect of the regeneration genes under the control of the XVE inducible promoter. These vectors are essentially the same as have been described in the literature; however, they were reconstructed from a pER8 version to contain selection on kanamycin. The binary vector pER10-PGA6 was described by Zuo et al., (2002) and should trigger a vegetative to embryo transition when stimulated with 17-β-estradiol. The second binary vector, pER10-ESR1, should trigger shoot formation. Plant transformation with binary vectors pER10-PGA6 and pER10-ESR1: We inoculated in vitro generated walnut stems, roots, leaves and embryos with vectors pER10-PGA6 and pER10-ESR1. Tissue was inoculated and maintained as described above. 3. Determine the stability of the silencing constructs Inoculation of Tomatoes: Wild-type and transgenic lines of the tomato cultivar Moneymaker were utilized. Transgenic line 01/6 expresses two self-complementary oncogene constructs of the iaaM and ipt oncogenes of Agrobacterium tumefaciens which induce PTGS of their expression (Escobar et al., 2001a). Tomato seeds for the first set of tests were germinated in a greenhouse and then transferred to growth chambers at various temperatures (15°C, 18°C, 20°C, 27°C) for acclimation prior to inoculation. At one, two, three and four weeks after inoculation, five plants were each transferred between the 15°C and the 27°C growth chambers to assist in the determination of any temperature sensitivity to gene silencing. Tomato seeds for the second set of tests were germinated, grown, and inoculated in growth chambers at 15°C and 25°C. Virulent A. tumefaciens strain 20W-5A or disarmed strain EHA-101 were grown in liquid cultures of 523 medium (Moore et al., 1989) at room temperature to OD600 of 0.4 – 0.5, then spun down and resuspended in water to OD600 of 0.10 – 0.15. Tomato plant stems were then inoculated by syringe at the two lowest nodes as described in Escobar, et al. (2001a). Inoculation of Walnut Microshoots: Microshoots of wild-type and transgenic clones of walnut were utilized. Transgenic line 01/258 is derived from the wild-type somatic embryo line CR1 (walnut cultivar Chandler) (Escobar et al., 2002a). Transgenic line MCG-42 is derived from the wild-type somatic embryo line Px1 (walnut cultivar Paradox) (Escobar et al., 2003).

Microshoots were grown on Driver-Kuniyuki walnut (DKW) medium (Driver et al., 1984) and placed in growth chambers at various temperatures (15°C, 18°C, 20°C, 27°C) for acclimation prior to inoculation. Virulent A. tumefaciens strain 20W-5A or disarmed strain EHA-101 were grown in liquid cultures of 523 medium at room temperature to OD600 of 0.6. Two or three internodes of each microshoot were wounded and inoculated with approximately 0.5 µL of the bacterial suspension as described in Escobar et al. (2002a). The inoculated microshoots were initially cultured on DKW medium without antibiotics. They were transferred within one week to DKW media to which 500 mg/L cefotaxime had been added to prevent bacterial overgrowth. The microshoots were subsequently transferred at one-week intervals to fresh DKW/cefotaxime media during the experimental period. The first set of walnut microshoots were cultured at two temperatures (15°C and 27°C). The second set of walnut microshoots were cultured at four temperatures (15°C, 18°C, 20°C and 27°C). Tomato RNA Analysis: Tomato plants of the second experimental set (15°C and 25°C) were analyzed for the presence of siRNA of the ipt oncogene. Leaf disks were frozen in liquid nitrogen at the growth chambers. Tissue disruption and subsequent homogenization was performed using a Mixing Mill 300 (Qiagen, Valencia, CA). Total RNA was isolated using TRIzol reagent (Invitrogen, Carlsbad, CA), and was subsequently size-fractionated on a RNA/DNA ion exchange column (Qiagen). The low molecular weight RNA fraction was electrophoresed through a 19% polyacrylamide-7 M Urea-0.5X TBE gel using a Mini Protean III vertical electrophoresis apparatus (Bio-Rad, Hercules, CA). The RNA in the gel was electroblotted to a Hybond-NX membrane and UV fixed. The blot was subsequently hybridized with a radiolabeled ipt DNA probe. Image analysis of the autoradiogram was performed on a Typhoon 8600 phosphoimager (Amersham Biosciences, Piscataway, NJ). RESULTS AND CONCLUSIONS The primary objective of this proposal is to develop cultivar independent transformation for walnut by stimulating regeneration to provide a unique avenue to combine resistance to crown gall disease in superior rootstocks that are resistant to Phytophthora and/or to root lesion nematode. This strategy will overcome an important barrier towards the development of elite rootstocks resistant to many disease and pest problems significant in the California horticultural landscape. Efficacy of the XVE inducer system in walnut somatic embryos Tobacco: Leaves harvested from cloned shoots of tobacco plants transformed with the plasmid pER10-GFP and cultured on MS medium supplemented with 30 g/l sucrose, 80 mg/l adenine sulfate, 2.0 mg/l kinetin, 2.0 mg/liter IAA, 8g/l TC agar and 5 µM 17-β-estradiol, pH 5.8 exhibited GFP expression whereas leaves harvested from cloned shoots grown on media lacking 5 µM 17-β-estradiol failed to express GFP (see Fig 3). Walnut: Fourteen weeks post-transformation, kanamycin resistant embryos transformed with pER10-GFP were observed under UV light using a Leica Stereomicroscope and GFP filter. GFP

expression was clearly visible in numerous embryos grown on media with 17-β-estradiol while fluorescence was absent in non-transgenic control embryos or transgenic embryos grown in the absence of GFP (see Fig 4). When GFP expression embryos were harvested and placed on MS medium containing 5 µM 17-β-estradiol they continued to express GFP. However, when GFP expressing embryos were transferred to medium lacking 5 µM 17-β-estradiol the expression of GFP dissipated within 7 days. Twenty-four hours after returning these non-fluorescing embryos to medium containing 17-β-estradiol, GFP expression was again induced (Fig 5). These results demonstrate that the 17-β-estradiol induction system can be used to turn on and off genes in walnut that will be critical to controlling the expression of the regeneration genes. We are in the process of regenerating whole plants from these embryos that will allow us to evaluate the expression pattern of the 17-β-estradiol induction system in whole plants by monitoring for the intensity of GFP expression in various tissues in the presence of 17-β-estradiol. This should provide clues as to which vegetative tissues hold the greatest potential for regeneration since it will be reflective of the pattern of expression of the regeneration genes. Capability the ESR1 and PGA6 genes, under the control of a constitutive promoter and the XVE inducible system, to stimulate the production of transgenic somatic embryos or organogenic shoots for a range of rootstock germplasm. Our previous work using another embryo-inducing transcription factor (LEC2) in tobacco, tomato, alfalfa and lettuce demonstrated that the response generated by these regeneration genes can be greatly affected by the tissue source and the presence or absence of exogenous hormones, particularly 2,4-D. Based on these results we transformed various tissue explants from Paradox walnut on medium that lacked 2,4-D in both the co-culture and induction medium, or media which contained 2,4-D either in the co-culture or induction medium or in both the co-culture and embryo induction medium (Table 1). Inoculated and non-inoculated somatic embryos, stem and root tissues exposed to 2,4-D in the induction medium or in both the co-cultivation and induction media developed brown gelatinous callus approximately 60 days after co-cultivation whereas tissues not exposed to 2,4-D produce nodular or friable callus. Numerous sectors of yellow pro-embryonic callus developed on the gelatinous brown callus generated from somatic embryos inoculated with pER10-PGA6 and grown in the presence of 5 µM 17-β-estradiol (Figures 6 and 7). It is still not clear whether the development of this pro-embryogenic callus is a result of the expression of the PGA6 gene or a 2,4-D effect, although we have yet to observe this type of callus on non–inoculated tissue exposed to 2,4-D and 5 µM 17-β-estradiol. We have currently transferring these proembryonic masses to media lacking 2,4-D with and without 17-β-estradiol. Similar transformation studies have been initiated with ESR1, but no results are available to date. Stability of the crown gall silencing construct: In previous reports we described the successful application of a biotechnology-based oncogene silencing strategy for the generation of resistance to crown gall disease (Escobar et al., 2000; Escobar et al., 2001b, Escobar et al., 2002b). Arabidopsis, tomato and walnut plants transformed with self-complementary oncogene constructs accumulate iaaM and ipt-homologous small interfering RNAs, display a significant decrease (~90%) in iaaM and ipt mRNA abundance, and show high-level suppression of A. tumefaciens-induced tumorigenesis. The broad-spectrum of

oncogene-silencing mediated resistance has also been experimentally demonstrated through disease challenge assays with a large and diverse group of Agrobacterium isolates. Recent research on silencing of viruses has shown some evidence of a breakdown of gene silencing at temperatures of 15°C (Szittya, et al., 2003). This has important implications for the suppression of crown gall disease through oncogene silencing, particularly for woody perennials, such as walnut, which overwinter at low temperatures. During 2003 two sets of experiments were performed, each utilizing wild-type and transgenic walnut microshoots and tomato plants. Walnut microshoots were acclimated to growth chambers between 15°C and 27°C. Plants were either germinated in the greenhouse and subsequently moved to growth chambers between 15°C and 27°C or germinated in growth chambers at 15°C or 25°C. The microshoots and plants were subsequently inoculated with virulent A. tumefaciens strain 20W-5A or disarmed strain EHA-101. As shown in Tables 2-5, in general, the phenotypes of the inoculated transgenic and wild-type tomato plants and walnut microshoots were consistent with previous experimental data (Escobar, et al., 2001a and 2002a), and showed no breakdown of silencing at 15°C. Nearly all wild-type walnut shoots (lines CR1 and Px1) inoculated with virulent Agrobacteria strain 20W-5A formed galls. None of the transgenic walnut shoots (lines 258 and MCG-42) formed galls. However, there was some callus production. Callus from the inoculated transgenic shoots of the second set was transferred to hormone-free DKW media and showed no subsequent growth, indicating that there was no endogenous hormone production under these conditions. However, gall tissue from wild-type plants proliferated on hormone-free media, demonstrating that the tissue was producing auxins and cytokinins as expected from the inoculation with wild-type Agrobacteria. Although no galls formed on microshoots inoculated with disarmed Agrobacteria strain EHA-101, there was some callus formation, similar to that observed on the transgenic shoots inoculated with 20W-5A. Most wild-type tomato plants inoculated with 20W-5A produced galls. A few transgenic tomato plants at 15°C also developed swelling or small galls. No gall formation was detected on tomatoes inoculated with EHA-101 at any temperature. It should be noted that Botrytis was observed in the 15°C growth chamber used in the first set of inoculations, which did not permit the experiment to be conducted for longer than five weeks. In the second set of tomato inoculations, a different 15°C growth chamber was used, and no Botrytis developed. A blot was produced using RNA isolated from inoculated transgenic tomato plants that had been grown at 27°C (from the first set of inoculations), 15° and 25°C (from the second set of inoculations). DNA probes generated from the A. tumefaciens ipt gene were hybridized to the blot. All samples showed essentially equal amounts of ipt-siRNA, indicating that PTGS was active at both high and low temperatures.

REFERENCES Driver, J.A. and Kuniyuki, A.H. 1984. In vitro propagation of paradox walnut rootstock. Hort. Sci. 19:507-509. Escobar, M.A., Uratsu, S.L., Civerolo, E.L., McGranahan, G.H., Leslie, C.A., and Dandekar, A.M. 2000. Development of a crown gall resistant walnut rootstock. In: Walnut Research Reports, 2000. Walnut Marketing Board, Sacramento. pp. 401-413. Escobar, M.A., Civerolo, E.L., Summerfelt, K.R. and Dandekar, A.M. 2001a. RNAi-mediated oncogene silencing confers resistance to crown gall tumorigenesis. PNAS. 98:13437-13442. Escobar, M.A., Uratsu, S.L., Civerolo, E.L., McGranahan, G.H., Leslie, C.A. and Dandekar, A.M. 2001b. Development of a crown gall-resistant walnut rootstock. In: Walnut Research Reports. Walnut Marketing Board, Sacramento. pp. 355-365. Escobar, M.A., Leslie, C.A., McGranahan, G.H. and Dandekar, A.M. 2002a. Silencing in crown gall disease in walnut (Juglans regia L.). Plant Science. 163:591-597. Escobar, M.A., Uratsu, S.L., Civerolo, E.L., McGranahan, G.H., Leslie, C.A. and Dandekar, A.M. 2002b. Development of a Crown Gall-Resistant Rootstock. In: Walnut Research Reports, 2002. Walnut Marketing Board, Sacramento. Moore, L.W., Kado, C.I. and Bouzar, H. 1988. Laboratory Guide for the Identification of Plant Pathogenic Bacteria. Agrobacterium. 16-36. Murashige, T. and Skoog, F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 15:473-497. Szittya, G., Silhavy, D., Molnár, A., Havelda, Z., Lovas, A., Lakatos, L., Bánfalvi and Burgyán, J. 2003. Low temperature inhibits RNA silencing-mediated defence by the control of siRNA generation. The EMBO Journal. 22(3):633-640. Zuo, J., Niu, Q.-W. and Chua, N.-H. 2000. An estrogen receptor-based transactivator LEXA-VP16-ER mediates highly inducible gene expression in transgenic plants. The Plant Journal. 24:265-273. Zuo, J., Niu, Q.-W., Frugis, G. and Chua, N-H. 2002. The WUSCHEL gene promotes vegetative-to-embryonic transition in Arabidopsis. The Plant Journal. 30(3)349-359.
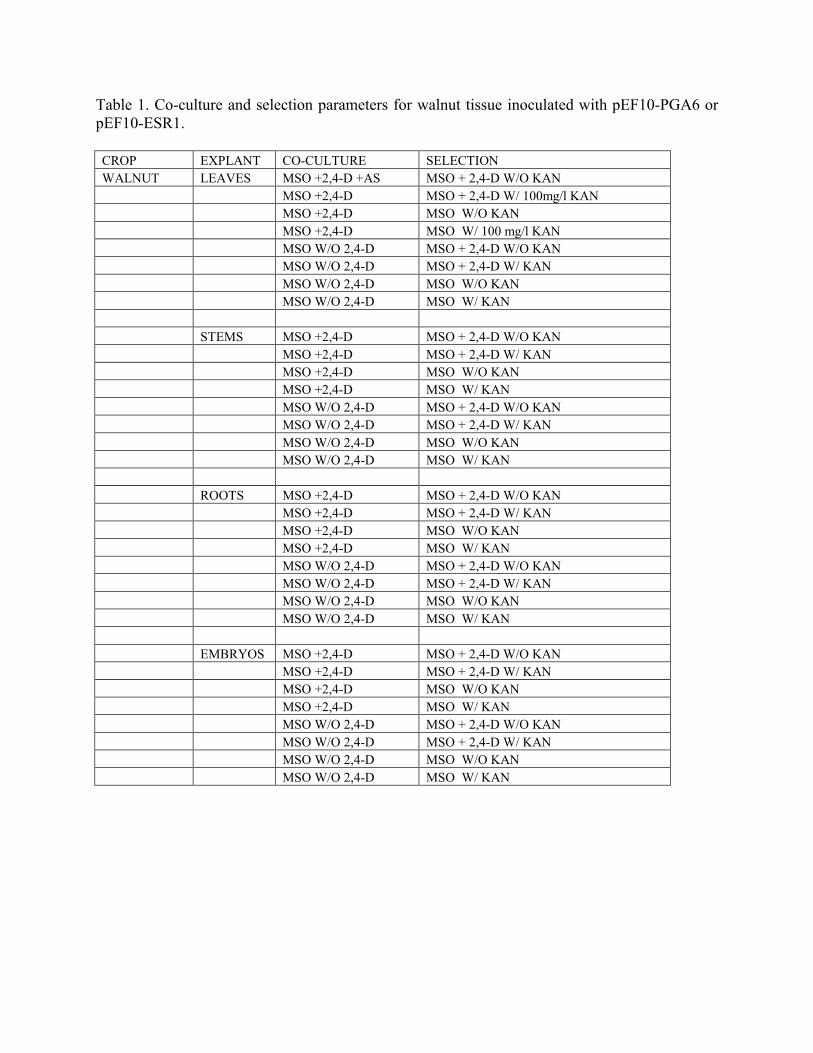
Table 1. Co-culture and selection parameters for walnut tissue inoculated with pEF10-PGA6 or pEF10-ESR1. CROP EXPLANT CO-CULTURE SELECTION WALNUT LEAVES MSO +2,4-D +AS MSO + 2,4-D W/O KAN MSO +2,4-D MSO + 2,4-D W/ 100mg/l KAN MSO +2,4-D MSO W/O KAN MSO +2,4-D MSO W/ 100 mg/l KAN MSO W/O 2,4-D MSO + 2,4-D W/O KAN MSO W/O 2,4-D MSO + 2,4-D W/ KAN MSO W/O 2,4-D MSO W/O KAN MSO W/O 2,4-D MSO W/ KAN STEMS MSO +2,4-D MSO + 2,4-D W/O KAN MSO +2,4-D MSO + 2,4-D W/ KAN MSO +2,4-D MSO W/O KAN MSO +2,4-D MSO W/ KAN MSO W/O 2,4-D MSO + 2,4-D W/O KAN MSO W/O 2,4-D MSO + 2,4-D W/ KAN MSO W/O 2,4-D MSO W/O KAN MSO W/O 2,4-D MSO W/ KAN ROOTS MSO +2,4-D MSO + 2,4-D W/O KAN MSO +2,4-D MSO + 2,4-D W/ KAN MSO +2,4-D MSO W/O KAN MSO +2,4-D MSO W/ KAN MSO W/O 2,4-D MSO + 2,4-D W/O KAN MSO W/O 2,4-D MSO + 2,4-D W/ KAN MSO W/O 2,4-D MSO W/O KAN MSO W/O 2,4-D MSO W/ KAN EMBRYOS MSO +2,4-D MSO + 2,4-D W/O KAN MSO +2,4-D MSO + 2,4-D W/ KAN MSO +2,4-D MSO W/O KAN MSO +2,4-D MSO W/ KAN MSO W/O 2,4-D MSO + 2,4-D W/O KAN MSO W/O 2,4-D MSO + 2,4-D W/ KAN MSO W/O 2,4-D MSO W/O KAN MSO W/O 2,4-D MSO W/ KAN

Table 2. Crown Gall Symptom Formation on Walnut Microshoots Inoculated with Virulent (20W-5A) and Disarmed (EHA-101) Strains of Agrobacterium tumefaciens and Incubated at Various Temperatures. (Walnut clones CR1 and PX1 are wild-type; 258 and MCG-42 are corresponding transgenic clones with crown gall resistance.)
Temp. A. tumefaciens
strain Walnut clone
Weeks post inoculations
Gall formation Comments
15°C 20W-5A CR1 7 ¾ Callus by 5 weeks; large galls by 7 weeks
258 7 0/4 PX1 7 4/4 2/4 callus by 5 weeks; 4/4
medium galls by 7 weeks MCG-42 7 0/4 15°C EHA-101 CR1 5 0/0 258 5 0/0 2/4 little callus PX1 5 0/0 1/4 little callus MCG-42 5 0/0 27°C 20W-5A CR1 5 4/4 258 5 0/4 2/4 little callus PX1 5 4/4 MCG-42 5 0/4 27°C EHA-101 CR1 5 0/0 258 5 0/0 1/4 little callus PX1 5 0/0 1/4 little callus MCG-42 5 0/0
Table 3. Crown Gall Symptom Formation on Walnut Microshoots Inoculated with Virulent Agrobacterium tumefaciens (strain 20W-5A) and Incubated at Various Temperatures.
Temp. A. tumefaciens
strain Walnut clone
Weeks post inoculations
Gall formation Comments
15°C 20W-5A CR1 10 5/5 258 10 0/22 1/22 some callus or exudate PX1 10 4/4 MCG-42 10 0/10 18°C 20W-5A CR1 5 4/4 258 5 0/5 2/5 some callus PX1 5 4/4 MCG-42 5 0/6 20°C 20W-5A CR1 5 4/4 258 5 0/6 PX1 5 4/4 MCG-42 5 0/6 27°C 20W-5A CR1 4 9/9 258 4 0/24 5/24 some callus PX1 4 5/5 MCG-42 4 0/12

Table 4. Crown Gall Symptom Formation on Tomato Plants Inoculated with Virulent (20W-5A) and Disarmed (EHA-101) Strains of Agrobacterium tumefaciens and Grown at Various Temperatures. (Tomato variety is ‘Moneymaker’; line 01/6 is transgenic ‘Moneymaker’ with crown gall resistance.)
Final Temperature/shift/ weeks after inoculation
A. tumefaciens strain
Tomato line
Weeks post inoculations
Gall formation Size of galls
15°C / no shift 20W-5A w.t. 5 3/5 Large 01/6 5 2/5 Small 15°C / 27 15 / 1 w.t. 5 4/5 Large 01/6 5 1/5 Small 15°C / 27 15 / 2 w.t. 5 5/5 Large 01/6 5 0/5 15°C / 27 15 / 3 w.t. 5 5/5 Large 01/6 5 0/5 15°C / 27 15 / 4 w.t. 5 4/5 Large 01/6 5 2/5 Swelling
(no callus) 15°C / no shift EHA-101 w.t. 5 0/5 01/6 5 0/5 15°C / 27 15 / 1 w.t. 5 0/5 01/6 5 0/5 15°C / 27 15 / 2 w.t. 5 0/5 01/6 5 0/5 15°C / 27 15 / 3 w.t. 5 0/5 01/6 5 0/5 15°C / 27 15 / 4 w.t. 5 0/5 01/6 5 0/5 18°C / no shift 20W-5A w.t. 5 5/5 Large 01/6 5 0/5 18°C / no shift EHA-101 w.t. 5 0/5 01/6 5 0/5 20°C / no shift 20W-5A w.t. 5 5/5 Large 01/6 5 0/5 20°C / no shift EHA-101 w.t. 5 0/5 01/6 5 0/5

Table 4. continued 27°C / no shift 20W-5A w.t. 5 5/5 Medium to
large 01/6 5 0/5 27°C / 15 27 / 1 w.t. 5 5/5 Various
sizes 01/6 5 0/5 27°C / 15 27 / 2 w.t. 5 5/5 Various
sizes 01/6 5 0/5 27°C / 15 27 / 3 w.t. 5 4/5 Various
sizes 01/6 5 0/5 27°C / 15 27 / 4 w.t. 5 4/5 Various
sizes 01/6 5 0/5 27°C / no shift EHA-101 w.t. 5 0/5 01/6 5 0/5 27°C / 15 27 / 1 w.t. 5 0/5 01/6 5 0/5 27°C / 15 27 / 2 w.t. 5 0/5 01/6 5 0/5 27°C / 15 27 / 3 w.t. 5 0/5 01/6 5 0/5 27°C / 15 27 / 4 w.t. 5 0/5 01/6 5 0/5
Table 5. Crown Gall Symptom Formation on Tomato Plants Inoculated with Agrobacterium tumefaciens (strain 20W-5A) and Grown at Various Temperatures.
Temp. A. tumefaciens
strain Tomato
line Weeks post inoculations
Gall formation Comments
15°C 20W-5A w.t. 12 18/18 01/6 12 1/17 25°C 20W-5A w.t. 12 10/10 01/6 12 0/15

Figure 3 17-β-estradiol induced GFP expression in transgenic tobacco leaves cultured on MS minimal organics medium supplemented with 500 mg/l cefotaxime, 100 mg/l kanamycin sulfate and 5 µM estradiol (right) or without 17-β-estradiol (left).

Figure 4. 17-β-estradiol induced GFP expression in transgenic walnut somatic embryos cultured on DKW minimal organics medium supplemented with 500 mg/l cefotaxime, 100 mg/l kanamycin sulfate and 5 µM 17-β-estradiol (bright field image to left of Fluorescent image)
Figure 5. 17-β-estradiol induced GFP expression and subsequent loss of expression in transgenic walnut somatic embryos cultured on DKW minimal organics medium supplemented with 500 mg/l cefotaxime, 100 mg/l kanamycin sulfate with 5 µM estradiol and then transferred to the same media with (top row) or without (bottom row) 5 µM 17-β-estradiol at 24h, 72 hrs and 7 days, and then 24 hours after subculturing tissue onto fresh media containing 17-β-estradiol (far right top and bottom) + 17-β-estradiol + 17-β-estradiol + 17-β-estradiol + 17-β-estradiol
24 hrs 72 hrs 7 days 24 hours after return to 17-β-estradiol media - 17-β-estradiol - 17-β-estradiol - 17-β-estradiol + 17-β-estradiol
24 hrs 72 hrs 7 days 24 hours after return to
17-β-estradiol media

Figure 6. Induction of proembryogenic callus on somatic embryos inoculated with pER10-PGA6 and plated on medium containing 1.0 mg/L 2,4-D and 5 µM 17-β-estradiol without kanamycin.
Figure 7. Proembryogenic and embryogenic callus from somatic embryos inoculated with pER10-PGA6 and plated on medium containing 1.0 mg/L 2,4-D and 5 µM 17-β-estradiol and lacking kanamycin.




![Valencia Advanced Production System [APS] Rootstock Trial ...](https://static.fdocuments.net/doc/165x107/62b438cf90c85f32066cb17f/valencia-advanced-production-system-aps-rootstock-trial-.jpg)













