
V·M·I · 2014-06-13 · Order Number 9205871 Isolation and characterization...
143
INFORMATION TO USERS This manuscript has been reproduced from the microfilm master. UMI films the text directly from the original or copy submitted. Thus, some thesis and dissertation copies are in typewriter face, while others may be from any type of computer printer. The quality of this reproduction is dependent upon the quality of the copy submitted. Broken or indistinct print, colored or poor quality illustrations and photographs, print bleedthrough, substandard margins, and improper alignment can adversely affect reproduction. In the unlikely event that the author did not send UMI a complete manuscript and there are missing pages, these will be noted. Also, if unauthorized copyright material had to be removed, a note will indicate the deletion. Oversize materials (e.g., maps, drawings, charts) are reproduced by sectioning the original, beginning at the upper left-hand corner and continuing from left to right in equal sections with small overlaps. Each original is also photographed in one exposure and is included in reduced form at the back of the book. Photographs included in the original manuscript have been reproduced xerographically in this copy. Higher quality 6" x 9" black and white photographic prints are available for any photographs or illustrations appearing in this copy for an additional charge. Contact UMI directly to order. V·M·I Uruversitv Microfilms International A Bell & Howell Information Company 300 North Zeeb Road. Ann Arbor. MI 48106-1346 USA 313,761·4700 800'521-0600
Transcript of V·M·I · 2014-06-13 · Order Number 9205871 Isolation and characterization...

INFORMATION TO USERS
This manuscript has been reproduced from the microfilm master. UMI
films the text directly from the original or copy submitted. Thus, some
thesis and dissertation copies are in typewriter face, while others may
be from any type of computer printer.
The quality of this reproduction is dependent upon the quality of thecopy submitted. Broken or indistinct print, colored or poor quality
illustrations and photographs, print bleedthrough, substandard margins,
and improper alignment can adverselyaffect reproduction.
In the unlikely event that the author did not send UMI a complete
manuscript and there are missing pages, these will be noted. Also, if
unauthorized copyright material had to be removed, a note will indicate
the deletion.
Oversize materials (e.g., maps, drawings, charts) are reproduced by
sectioning the original, beginning at the upper left-hand corner and
continuing from left to right in equal sections with small overlaps. Each
original is also photographed in one exposure and is included in
reduced form at the back of the book.
Photographs included in the original manuscript have been reproduced
xerographically in this copy. Higher quality 6" x 9" black and white
photographic prints are available for any photographs or illustrations
appearing in this copy for an additional charge. Contact UMI directly
to order.
V·M·IUruversitv Microfilms International
A Bell & Howell Information Company300 North Zeeb Road. Ann Arbor. MI 48106-1346 USA
313,761·4700 800'521-0600


Order Number 9205871
Isolation and characterization of calmodulin-binding heat shockproteins and cDNAs encoding calmodulin-binding proteins incultured tobacco cells
Lu, Yingtang, Ph.D.
University of Hawaii, 1991
V·M·I300 N. Zeeb Rd.Ann Arbor,MI 48106


ISOLATION AND CHARACTERIZATION
OF CALMODULIN-BINDING HEAT SHOCK PROTEINS
AND cDNAS ENCODING CALMODULIN-BINDING PROTEINS
IN CULTURED TOBACCO CELLS
A DISSERTATION SUBMITTED TO THE GRADUATE DIVISION OF THEUNIVERSITY OF HAWAII IN PARTIAL FULFILLMENT OF THE
REQUIREMENTS FOR THE DEGREE OF
DOCTOR OF PHILOSOPHY
IN
BOTANICAL SCIENCES(PLANT PHYSIOLOGY)
AUGUST 1991
BY
YINGTANG LU
Dissertation Commitee:
H. Michael Harrington, ChairpersonThomas L. GermanSamuel S. M. Sun
Marguerite VoliniAlton L. Boynton

ACKNOWLEDEMENTS
I wish to express my sincere thanks to Dr. H. Michael
Harrington for his support and guidance during the
development of this dissertation, and to Dr. Alton L.
Boynton, Dr. Thomas L. German, Dr. Samuel S. M. Sun, Dr.
Marguerite Volini and Dr. Harry Y. Yamamoto for their
valuable comments.
Special thanks to Dr. John I. Stiles in whose lab I was
well trained in molecular biology.
My wife, Shuping Liang's support and understanding
helped to finish this study.
iii

ABSTRACT
Calmodulin-binding heat shock proteins were
characterized in cultured tobacco cells (Nicotinan tabacum
L. cv Wisconsin-38). Analyses of 35S-labeled calmodulin
binding proteins (CaMBPs) purified by affinity
chromatography and SDS-PAGE indicated that heat shock (38°C)
enhanced/induced the synthesis of some CaMBPs while the
synthesis of others was repressed during the heat shock
response (HSR). The synthesis of CaMBPs with apparent
molecular weights of 82, 78, 71, 68, 22, 20, 19.5 and 17 Kd
was stimulated by heat shock.
A procedure for the isolation of eDNA clones encoding
CaMBPs was refined. Twenty five positive eDNA clones were
isolated by screening a tobacco heat shock eDNA expression
library with 35S-CaM as a ligand probe. These clones
produced peptides exhibiting Ca2+-dependent, CaM-binding
activity when assayed by gel overlay analysis. While most
cloned mRNAs such as pTCB40 were unaffected by heat shock,
two clones, pTCB60 and pTCB48 were heat shock-related.
Analysis of Northern blot demonstrated that a 2.1 kb mRNA
recognized by pTCB60 decreased by at least 70% in a 2 hour
heat shock treatment. The expression of pTCB48 mRNA was
induced by heat shock. This translationally active mRNA was
detected after 15 minutes of heat shock and accumulated to
maximum amounts after 1.5 hours. These results suggest that
iv

the ca2+/CaM second messenger system plays a role in tobacco
heat shock response.
The natures of CaM-binding domains were determined for
pTCB48 and pTCB60. DNA sequences of these clones were
determined and several deletion constructs from both the 5'
and 3' ends of the inserts were constructed. The CaM
binding activities of the proteins generated from these
deletion constructs were assayed by gel overlay analysis.
These data combined with secondary structure analyses of
deduced proteins localized the CaM-binding domains in the C
terminus of these proteins. The CaM-binding domain of TCB60
was a basic amphiphilic a-helix similar to that of several
animal and human CaMBPs. No similar structure was found in
the c-terminal region of TCB48 suggesting that an
alternative structure is responsible for the CaM-binding.
v

TABLE OF CONTENTS
ACKNOWLEDGEMENTS .....................................•.. 111
ABSTRACT ••••••.•.••••.•..•...•...........•..••.•....•••.. i v
LIST OF TABLES .•................... ., 0 •••••••• vii
LIST OF FIGURES ••••••••..•.•.•.•. 0 ••••••••••••••••••••• viii
LIST OF ABBREVIATIONS ..••.••.•.•..•••.•............•...... x
CHAPTER I. LITERATURE REVIEW .•.••••....••.......•..•.... 1Introduction 1Heat shock response 2Heat shock proteins 5Functions of heat shock proteins .•...........•....... 9Regulation of heat shock gene expression 14
Transcriptional regUlation 15Translational regulation ................•...... 23
Calcium, calmodulin and calmodulin-binding proteinsin heat shock response 31
CHAPTER II. SIGNIFICANCE AND HYPOTHESIS .•............... 36
CHAPTER III. CHARACTERIZATION OF HEAT SHOCK INDUCEDCALMODULIN-BINDING PROTEINS IN CULTURED TOBACCOCELLS • 0 ••••• 0 ••••••••••••••••• II ••••••••••••••••••••• 38Introduction 38Materials and methods ........•........•........•.... 39Results and discussion .: •......................••... 41
CHAPTER IV. ISOLATION AND CHARACTERIZATION OF eDNA CLONESENCODING TOBACCO CALMODULIN-BINDING PROTEINS ...•.... 50Introduction 50Materials and methods ....•...•.................•.... 52Results and discussion .......•.......•..•........•.. 62
A. Isolation and confirmation of eDNA clonesencoding calmodulin-binding proteins ..... 62
B. Characterization of calmodulin-bindingprotein clone pTCB40 .•..•...••........... 69
C. Characterization of calmodulin-bindingprotein clone pTCB60 and identification ofcalmodulin-binding domain .•..•.......•... 78
D. Characterization of a eDNA clone encoding aheat shock-induced calmodulin-bindingprotein 89
CHAPTER V. CONCLUSION ...................•............. 100
REFERENCES •••••••••••••••••••••••••••••••.•• 0 ••••••••••• 103
vi

Table
1
2
LIST OF TABLES
Page
comparison of radioactive proteins recoveredfrom CaM-sepharose-4B and sepharose-4B ••.. 45
The sizes of the inserts of the cDNA clonesencoding CaMBPs ••.••••••...•...•.••...•... 66
vii

Figure
LIST OF FIGURES
Page
1 CaM-sepharose-4B elution profile ....••.....•..•. 43
2 8DS-PAGE of CaM-sepharose chromatographyfractions 0 ••••••••••• 46
3 8DS-PAGE of 35S-labeled calmodulin ...•.....•.... 55
4 pBluescript SK- vector .•..•.......•.••.•....... 58
5 Autoradiograms of blotted phages screened with35S-labeled calmodulin ...................•. 64
6 Double-digestion of the recombinant pBluescriptphagemids encoding CaMBPs 65
7 Calmodulin gel overlay analysis of the proteingenerated from clone pTCB01 67
8 Calmodulin gel overlay analysis of extractsfrom 25 independent recombinants ..•...•..• 68
9 Northern blot analysis of pTCB40 ....•..•.••.•.. 71
10 DNA sequence and deduced amino acid sequence ofpTCB40 •.•.••••.•.•••.••••.•.••••.••••••••. 72
11 Deletion constructs of pTCB40 74
12 Calmodulin gel overlay analysis of extractsfrom E.coli XL1-Blue cells harboringpTCB40 or deletion constructs .•...•....... 75
13 Predicted secondary structure of TCB40 proteinwith Chou-Fasman method ..•..•.....•....•.. 77
14 Northern blot analysis of pTCB60 79
15 DNA sequence and deduced amino acid sequence ofpTCB60 ...•. ., ......•.•...•................. 81
16 Deletion constructs of pTCB60 .........••....•.. 83
17 Calmodulin gel overlay analysis of extractsfrom E.coli XL1-Blue cells harboringpTCB60 or deletion constructs .•..•........ 84
viii

18 Predicted secondary structure of TCB60 proteinwith Chou-Fasman method ... 0 •••••••••• 0 •••• 85
19 Helical wheel projection for tentative CaM-binding domain of TCB60 protein •...•.•.•.. 86
20 Helical wheel projections for CaM-binding domainfrom chicken myosin light chain kinase .•.. 88
21 Deletion constructs of pTCB48 •.......••.•.•.... 90
22 Calmodulin gel overlay analysis of extractsfrom E.coli XL1-Blue cells harboringpTCB48 or deletion constructs .•........•.. 91
23 Northern blot analysis of pTCB48 .•...•.•....•.. 93
24 Time course of expression of pTCB48 mRNAfor different lengths of heat shocktreatrnent II •••••••••• 94
25 DNA sequence and deduced amino acid sequence ofpTCB48 ••.••.••.••••.•..••••.•..••.•••..••. 96
26 Predicted secondary structure of TCB48 proteinwith Chou-Fasman method •..•.......•....•.. 98
27 A model for the possible functions of CaMBPs inthe HSR ..•.................•......•....•. 102
ix

A260
~OOBSAcDNACaMCaMBPcAMPcpmcvDAGDEPCDNADNAseDTTE.coliEDTAEGTA
ggHSHSEHSFHSPHSRIEFIPTGIP3KbKdMLCKMOPSmRNANP-40oligo(dT)PEGpfuPIP2PKCPLCPMSFpoly(A)PVPRNARNPSDSSDS-PAGETris
LIST OF ABBREVIATIONS
absorbance at 260 nmabsorbance at 600 nmbovine serum albumincomplementary DNAcalmodulincalmodulin-binding proteincyclic AMPcounts per minutecultivardiacylglyceroldiethylpyrocarbonatedeoxyribonucleic aciddeoxyribonuclasedithiothreitolEscherichia coliethylenediamine tetraacetic acidethyleneglycol-bis-(~-aminoethylether) N,N,N',N'tetraacetic acidgramgravityheat shockheat shock elementheat shock factorheat shock proteinheat shock responseisoelectric focusisopropylthiogalactosideinositol 1,4,S-triphosphatekilobasekilodaltonmyosin light chain kinase3-(N-morpholino)propanesulfonic acidmessenger RNAnonidet P-40oligothyrnidylic acidpolyethylene glycerolplaque forming unitsphosphatidylinositol-4,S-bisphosphateprotein kinase Cphospholipase Cphenylmethylsulfonyl fluoridepoly (adenylated)polyvinylpyrrolidoneribonucleic acidribonucleoproteinsodium dodecyl sulfateSDS-polyacrilaminde gel electrophoresisTris[hydroxyrnethyl]aminomethane
x

CHAPTER I
LITERATURE REVIEW
INTRODUCTION
All organisms are subjected to a large number of biotic
and abiotic stresses. Environmental perturbations such as
light, heat, anaerobiosis influence gene expression. Recent
studies on the responses of organisms to stress have focused
on the analysis of gene expression. The heat shock response
(HSR) provides a convenient system for investigating
mechanisms of gene expression in a wide variety of organisms
and is the subject of intense investigation.
The HSR was first investigated in the fruit fly
Drosophila melanogaster by Ritossa (227). When Drosophila
cells are shifted from normal growing temperature (2SoC) to
an increased temperature (37°C), a novel set of heat shock
proteins (HSPs) is synthesized with the rapid shutdown of
most normal protein synthesis. The mRNAs encoding the HSPs
result from de novo transcription and are selectively
translated during heat shock. Thus, the HSR involves both
transcriptional and translational control of gene
expression. A number of lines of evidence support the
contention that the presence of HSPs in cells confers
1

tolerance to sUbsequent, more intense heat stress. However,
little information is available on the possible functions of
HSPs in thermotolerance. Moreover, the mechanisms by which
heat-shock induces/represses genes and how the cell senses
the heat-shock signal and converts the signal into responses
are unclear. Several lines of evidence suggest that
Ca2+/calmodulin mediated processes are involved in the HSR
(56, 141, 151, 192, 271, 299).
HEAT SHOCK RESPONSE
All organisms respond to higher than normal growing
temperatures through profound alterations in gene
expression. The major features of the HSR are the shutdown
of most normal protein synthesis; de novo synthesis of heat
shock mRNAs and heat shock proteins (HSPs), and the
acquisition of thermotolerance to otherwise non-permissive
heat stress (66, 132, 133, 163, 164, 189). In general, the
initiation of HSP synthesis occurs within minutes after the
start of heat shock treatment. The maximum induction of HSP
synthesis requires about 10°C increase above normal growth
temperature for a variety of different organisms. For
example, Drosophila cells are normally grown at 25°C and
HSPs are initially induced when the temperature is raised to
29°C. optimum HSP synthesis occurs at 36-37°C (161). At the
2

optimum temperature, heat shock mRNAs are produced within
four minutes and within an hour several thousand heat shock
specific transcripts are present in each cell (160). The
heat shock specific mRNAs are translated with very high
efficiency (160). At the same time, both the transcription
of previously active genes (20, 95, 124, 269) and the
translation of pre-existing control mRNAs are suppressed
(162). Normal protein synthesis gradually resumes when
cells are returned to normal temperature (163).
The HSR of most plants such as tobacco (111), spinach
(263), wheat (113, 143), soybean (135), barley (57) and
sorghum (57) is generally similar to that of Drosophila and
other systems (163). In soybean seedlings the synthesis of
normal proteins is greatly decreased and HSPs are induced
when sUbjected to heat shock treatment at 40°C for 4 hours
(131). Similar results are also obtained in cultured
tobacco and tomato cells in which heat shock proteins are
induced by heat shock treatment with little constitutive
proteins synthesis (111, 195). Some exceptions are observed
in the tropical crops such as maize (61, 62) and sugarcane
cells (185) where the HSR does not result in complete
shutdown of normal protein synthesis. This may mean that
different regulation mechanisms for the HSR exist in these
tropical plants.
3

The relationship of the HSPs and development of
thermotolerance has been subjected to great attention. Many
lines of evidence indicate that the synthesis of heat shock
proteins is necessary for the development of thermotolerance
(18, 100, 111, 150, 158, 168, 177, 185, 235, 246, 275)
although a few contradictory reports also exist (33, 109,
220). In brief, data show increased survival of cells
exposed to a normally lethal temperature treatment if the
cells are first subjected to a less extreme heat treatment
(heat shock). The development and decay of thermotolerance
closely parallels the rate of HSP accumulation and decay
when cells are returned to normal temperature. Similar
results are obtained in plant systems. In cultured tobacco
cells, treatment of heat shock (38°C for two hours) confers
thermotolerance to otherwise lethal temperature (8 minutes
at 54°C) (111). Tomato and sugarcane cells require the
synthesis of the small heat shock proteins to achieve
thermotolerance (185, 194).
Several lines of direct evidence for the essential role
of HSPs has been reported (152, 226, 235). HSP 70 has been
demonstrated to be necessary for thermotolerance in animal
systems (226). Injection of antibodies against HSP 70 into
rat cells results in the inability to develop
thermotolerance (226). A role of small HSPs in
thermotolerance development is suggested since a
4

Dictyostelium mutant which is deficient for the synthesis of
several of the small heat shock proteins lacks the ability
to survive extreme temperature (235). Recently, genetic
transfection experiments indicate that HSP 27 plays a major
role in thermoresistance. The thermotolerant phenotype can
be conferred to Chinese hamster and mouse cells by
transfection with the human HSP 27 gene (152).
Heat shock proteins are induced not only by heat shock,
but also by a wide variety of agents such as amino acid
analogs (129), arsenite (134), ABA and water-stress (112),
salt stress (111), methomyl (224), metals (69, 283), light
(103) and many other factors (43). Some stresses induce a
complete set of HSPs while others do not, suggesting that
there is a set of stress proteins common to several forms of
stress. Further evidence demonstrates that the expression
of certain heat shock genes including HSP 90 and small HSPs
are developmentally regulated (32, 51, 69, 74, 107),
implying a role of the HSPs in normal cellular growth.
HEAT SHOCK PROTEINS
Heat shock proteins are generally defined as whose
synthesis is sharply and dramatically induced/enhanced at
high temperature. The genes for such proteins have heat
5

shock elements (HSE, see below) which account for strong
induction upon exposure to elevated temperature (163, 276).
The major HSPs can be grouped into two size classes based on
SOS-PAGE. The large HSPs range from 68 to 110 Kd while
small HSPs range from 15 to 30 Kd (35). When proteins
extracted from heat-shocked soybean seedlings are separated
by SOS-polyacrylamide gel electrophoresis, ten HSP bands are
distinguished on one-dimensional gels. A more complex
pattern appears on two-dimensional gels with more than 60
labeled proteins (131). These phenomena are ubiquitous in a
variety of experimental systems. Using cultured tobacco
cells, Harrington and AIm (1988) indicate that many proteins
are induced during heat shock. Apparent molecular weights
of 94, 80, 71, 50, 48, 44, 41, 40, 36, 30, 28, 26, 25, 23,
22 and 20 to 15 (kd) have been reported (111). Two
dimensional gels reveal that there may be as many as 100
polypeptides synthesized during heat shock in tobacco cells
(S. Oharmasiri, personal communication). Of these HSP 94,
80, 71 and small HSPs (20 to 15 Kd) are most prominent.
The HSPs show a remarkable conservation throughout
evolution. The larger HSPs appear to be more highly
conserved than the smaller HSPs. In fact, HSP 70 has been
suggested to be the most conserved protein in nature. A
polyclonal antibody against chicken HSP 70 cross-reacts with
HSP 70 from yeast, dinoflagellates, slime molds, maize,
6

worm, frogs, Drosophila, mice, rats and humans (130).
Analyses of HSP 70 genes from different species indicate
that eukaryotic families of related genes for HSP 70 evolved
from a single bacterial gene (66, 115, 163). Yeast has
eight genes for the HSP 70 family and these are grouped into
four subfamilies (SSA, SSB, SSC and SSD) with homologies
ranging from 96% to 50% with each other (165). Comparison
of the deduced amino acid sequences of cloned HSP 70 genes
from different eukaryotic species reveals a high degree of
homology, ranging between 60% and 70%. (66, 163).
Furthermore, E.coli HSP 70, DnaK gene product, is 48%
identical to the HSP 70 of yeast and Drosophila (12). Plant
HSP 70 genes have been isolated from maize (232),
Arabidopsis (303), Petunia (305) and soybean (189). All
these genes have homologies with each other ranging from 72%
to 86%. with the exception of the soybean gene further
similarities exist in the presence of an intron located in
all of these plant genes (189).
other major HSP families are HSP 110 and HSP 90.
Mammalian cells produce proteins of 110 Kd and 100 Kd which
do not appear to have counterparts in Drosophila (165). For
the HSP 90 family, the genes have been cloned from several
evolutionarily diverse organisms, including Drosophila,
yeast, chickens, mammals and bacteria. Sequence analysis
reveals that HSP 90 is the second-most highly conserved HSP
7

examined to date. The proteins from eukaryotic species have
50% identity and all have greater than 40% identity with the
E.coli protein (13, 94, 165). Using a Drosophila HSP 83
gene fragment as probe, HSP 90 genes have been isolated from
soybean (229), maize (262) and Arabidopsis (60).
Small HSPs have also been the subject of great
interest, especially in plants where they are abundantly
expressed during the HSR. In animal systems, the greatest
proportion of HSP synthesis is represented by the high
molecular mass HSPs of 68 to 110 kd with HSP 70 being the
most abundant species (198). In Drosophila, several low
molecular weight HSPs have been reported (19). In contrast,
as many as 27 small HSPs have been detected in soybean
seedlings (173). Varying numbers of small HSPs have been
also reported in other plants: pea, sunflower, wheat, rice,
maize, millet (173), cotton (42), tomato (195), tobacco
(111) and sugarcane (185). Many genes for these small HSPs
have been isolated from different organisms and sequence
analyses indicate that these small HSPs of different
organisms are clearly related (165, 274). However, small
HSPs show much greater homology within organisms than
between organisms. For example, members of a sUbgroup of
the soybean small HSP family have 90% amino acid identity
with each other but only 20% amino acid identity with the
proteins of Drosophila, Xenopus and Caenorhabditis elegant
8

(165, 188). Yeast HSP 26 exhibits 30-50% c-terminal
homology with small HSPs from Drosophila, Xenopus, Human and
c. elegant (35).
HSPs may be modified by phosphorylation, methylation,
ADP-ribosylation and/or glycosylation (89, 195, 196). HSP
89 is modified by phosphorylation (99) and methylation (290,
291). This is also true in tomato where HSP 80 and 70 are
phosphorylated and methylated (195). Moreover, that many
pre-existing proteins are modified during heat shock is
evidenced in tomato (243). Ribosomal protein S6 is rapidly
dephosphorylated when cells are sUbjected to heat shock and
the protein is rephosphorylated after cells return to normal
temperature (243). Further experiments also demonstrated
that several other ribosomal proteins become phosphorylated
during heat shock (244). Nuclear proteins are also modified
during heat shock (45, 46). In light of these discoveries,
it is possible that heat shock may result in induction or
inhibition of specific phosphatases and/or protein kinases.
FUNCTIONS OF HEAT SHOCK PROTEINS
All available evidence indicates that the HSPs protect
cells from damage of heat and other stresses (163, 165).
The determination of the cellular localization of HSPs is of
9

great importance because it is reasonable to suppose that
there is a logical connection between the HSP function and
localization (184). Many methods including cell
fractionation (287, 294), direct autoradiographic analysis
(5, 284) and immunofluorescence (6, 286) have been employed
for HSP localization.
Localization of Drosophila HSP 70 reveals that the
protein is concentrated mainly within the nucleus and
secondarily at the cell membrane after heat shock. During
recovery from heat shock, the protein delocalizes from the
nucleus and is found mainly in the cytoplasm (286).
Furthermore, when the Drosophila HSP 70 gene is introduced
into mammalian cells, the protein shows a very similar
pattern of localization (213). This is consistent with the
hypothesis that macromolecular complexes can be prevented
from unfolding and denaturation by association with HSPs
(247) .
Heat shock causes precipitation of numerous nuclear
proteins, damages the structure of partially assembled
ribosomes and completely blocks nucleolar function (24).
HSP 70 concentrates in nucleoli, binds to the nuclear
matrix, associates with cytoskeleton and pre-ribosomes, and
may protect pre-ribosomes from heat damage (56). Direct
evidence for this comes from the studies on genetic
10

transfected animal cells (214). The Drosophila HSP 70 gene
was placed under the control of adenovirus major late
promotor and introduced into mouse L cells and monkey COS
cells. HSP 70 was mostly found in the nucleus of unstressed
cells but strongly concentrated in nucleoli after heat
shock. When cells containing this chimeric gene were
treated with actinomycin, followed by heat shock and
recovery, nucleolar morphology and ribosome export resumed
much more rapidly than in cells carrying no chimeric gene.
This recovery requires neither RNA or protein synthesis.
Taken together with observations that heat shock disrupts
pre-ribosomal RNPs, these imply that HSP 70 may bind to
damaged RNPs and catalyze their ordered reassembly (214).
HSP 70 and related HSPs are involved in a variety of
cellular processes including DNA replication (165, 190),
post-translational translocation of proteins across
membranes (17, 53, 80), protection of RNA splicing (310),
association with hnRNA (138) and organization of
cytoskeleton (56), and uncoating coated vesicles (52, 280).
These diverse functions have been suggested to be due to the
ability of HSP 70 to prevent or disrupt inappropriate
protein-protein interactions by binding to hydrophobic
regions of proteins. This binding can be reversed with the
aid of ATP hydrolysis (145). The fact that HSP 70 family
can bind and hydrolyze ATP favors this idea (52, 295, 316).
11

using ATP-derived energy, HSP 70 breaks protein-protein
interactions and allows the denatured protein to refold or
reassemble into the normal state. Recent evidence from in
vitro experiments demonstrates that HSP 70 from soybean
seedlings, together with other HSPs, has the ability to
protect the control proteins from heat denaturation (126).
HSP 90 associates with membrane ATPase (39, 68),
steroid hormone receptors (50, 237, 252) and tyrosine kinase
(37, 311). The transforming protein of Rous sarcoma virus,
pp60sre a tyrosine kinase, associates with HSP 90 and a 50 Kd
phosphoprotein immediately after synthesis. When the kinase
is disassociated from HSP 90, the kinase inserts into the
membrane as a phosphoprotein and acts as a kinase (37, 38,
65, 204). These results suggest that HSP 90 may function to
keep its targets in an inactive state. This suggestion has
been supported by the observation that HSP 90 associates
with glucocorticoid receptors preventing binding to DNA.
The inactive complex is maintained until hormone disrupts
the association of HSP 90 to the receptor (16, 44, 165,
239). Recent results have demonstrated that HSP 90
stimulates eIF-2a kinase activity by association with it
(144, 233, 234). While HSP 90 is abundant in most cells,
only a small portion is found to be associated with these
cellular proteins and the bulk of HSP 90 is present as a
12

monomer (146, 153, 163). The significance of the excess
amount of HSP 90 is presently unclear.
structural characterization of small HSPs demonstrates
that these HSPs have structural homology to the a-crystallin
(35, 79, 120). In line with these characteristics, small
HSPs may mediate effects in stressed cells via molecular
aggregation either with themselves or with other related
proteins. Recent evidence has shown that the major portion
of small HSPs is present in large aggregates called heat
shock granules. These are found mainly in the perinuclear
region of heat shocked cells of tomato (194, 200),
Drosophila (7) and vertebrates (8, 59). Untranslated
constitutive or normal rnRNAs are detected to associate with
this cytoplasmic heat shock granule fraction (200),
suggesting that small HSPs may play some role in
conservation of untranslated control mRNAs during heat
shock.
Ubiquitin, a highly conserved protein with a mass of
approximately 8 KD, is found in all eUkaryotic cells (30,
31, 97,248, 256). It is synthesized as polyubiquitin and
found in cells either free or linked via its terminal
glycine residue to a variety of cellular proteins.
Ubiquitin forms conjugated complexes with aberrant or
unstable proteins in an ATP-dependent manner, conferring
13

selective degradation of these proteins (166, 207, 248). It
has been suggested that ubiquitin and other HSPs provide
complementary methods of dealing with the production of
denatured protein aggregates in heat shocked cells (96).
Many other HSPs have also been characterized for their
functions. For example, yeast hsp 48 is enolase (118) and
another HSP is glyceraldehyde-3-phosphate dehydrogenase
(G3PDH) (165). Recently, Ostermann and co-workers
discovered that proteins imported from the cytosol into
mitochondria do not refold spontaneously once translocation
across the mitochondrial membrane is completed (205). A
nuclear-coded, mitochondrial HSP 60 is involved in the
folding of imported proteins in conjunction with ATP (205).
REGULATION OF HEAT SHOCK GENE EXPRESSION
The heat shock response is an ideal system for
investigating molecular mechanisms of gene expression
because of the speed of induction, the magnitude of the
response and ubiquity in a wide variety of organisms. Many
lines of evidence indicate that heat shock gene expression
is under the control at either transcriptional or
translational levels or both, depending on the organism. In
E.coli (190, 307) and in yeast (162, 176), the response is
14

controlled primarily at the level of transcription. In
contrast, the response of Xenopus oocytes is controlled at
the translational level (22). In other systems including
animals, plants and Drosophila, both transcriptional and
translational controls are active. While the translational
repression of most pre-existing normal mRNA during the HSR
is common in many systems, translation of normal mRNA in
sugarcane seems not to be repressed by heat (185). These
differences provide impetus for investigation of the
mechanisms of gene regulation.
TRANSCRIPTIONAL REGULATION
Chromatin structure. Many heat shock genes are quickly
activated in a manner that results in the rapid accumulation
of heat shock mRNAs under inductive conditions. The
products of most heat shock genes are barely detectable or
undetectable under normal conditions. For example,
transcription of the heat shock genes in Drosophila can be
fUlly induced within minutes of temperature elevation (161)
and message levels increase lOOO-fold within an hour after
heat shock induction (285). It has been suggested that heat
shock genes are preassembled into an open chromatin
configuration at normal temperature, thus facilitating
immediate activation under heat shock. This is based on the
15

results from the studies of hypersensitive sites of heat
shock genes in chromatin (49, 64, 83, 279, 300). The genes
for HSP 70, 83, 22, 23, 26, and 28 contain two to five DNase
I hypersensitive sites at their 5' ends at normal
temperature. However, heat shock treatment results in the
changes in the position and number of hypersensitive sites
(64, 279, 300). Evidence obtained from Drosophila HSP 70
and 83 genes indicates that the promoter regions containing
heat shock elements (HSEs) are protected only after
temperature upshift while the TATA-box region is protected
in both heat-shocked cells and control cells (301).
Moreover, the HSEs can be protected against DNase I
digestion by applying an extract from heat shocked nuclei to
unshocked nuclei (302). Similar results are also obtained
for small Drosophila HSP gene promoters (49). Several
regions containing HSEs are hypersensitive in non-heat
shocked cells but protected after heat shock. These results
are interpreted as evidence for binding protection by trans
acting protein factors. What prevents nucleosomes from
covering the HSE region of HSP gene promoters is unclear
although the results above suggest it may be the case. HSP
gene transformation experiments done by Costlow and Lis (64)
may provide a clue for this. After the Drosophila HSP 70
and 83 genes were introduced and integrated into yeast
genome, the DNase I hypersensitivity of the promotor
sequence was preserved. This implies that heat shock gene
16

sequences carry information necessary for specific
hypersensitive structure in chromatin.
Promotor structure. Heat shock gene transcription is
coordinately regulated and highly conserved in all
eukaryotic species examined thus far. In spinach, in vitro
translation of heat shock mRNAs results in the synthesis of
all 35 HSPs. The mRNAs for all 35 HSPs are induced by heat
shock at 32°C, indicating coordinate transcriptional
regulation of all heat shock genes with respect to
temperature while non-coordinate synthesis of HSPs is
recorded (263). Similarly, coordinate heat shock mRNA
expression has been also documented in maize (15).
Additionally, transgenic expression experiments indicate
that regulatory mechanisms of the heat shock gene
transcription are highly conserved in different organisms
(63, 251, 267). A chimeric gene construct which contained
Drosophila HSP 70 promotor and the reporter gene, neomycin
phosphotransferase II (NPT II), was introduced into tobacco
cells. The NPT II gene was expressed in a heat-induced
fashion in tobacco (267), suggesting a similar mechanism
regulating heat shock gene expression across widely
divergent species. This view is further strengthened by the
observation that the cloned Drosophila HSP 70 gene is under
heat shock control in mouse fibroblasts (63). The
conservation of heat shock promotor function has been
17

indicated by the expression of the Drosophila HSP 70
promotor in mammalian (41, 63, 182, 212), amphibian cells
(288) and in regenerated tobacco plants (268). These
results suggest that heat shock gene promotor sequence per
se carries the information required for heat shock
activation and that the transacting factor(s) are conserved.
Sequence analyses of heat shock genes have identified a
short sequence upstream of the TATA-box of Drosophila HSP 70
gene promotor that is essential for heat inducibility (182,
211). This palindromic consensus sequence (CT-GAA--TTC-AG),
called the heat shock element (HSE), has been found within
the first 400 base pairs upstream of every eUkaryotic heat
shock gene sequenced to date (24, 70, 71, 165, 187, 232,
249). within the HSE, eight nucleotides, C--GAA--TTC--G,
are highly conserved and seven of these are required in
order to constitute an individual, functional HSE (23).
Many lines of evidence show that it is HSE that confers
heat-inducibility of heat shock genes (for review, see 24).
This conclusion is strengthened by the findings that one
synthetic HSE is sufficient for heat inducible transcription
of the Herpes simplex virus thymidine kinase gene (212).
Many heat shock gene promoters contain multiple HSEs
and the most proximal one is usually found 15-18 bp
immediately 5' to TATA box (24). As many as seven HSE
18

copies have been reported in Drosophila HSP 26 gene promotor
(83). Analyses of the 5' deletion mutants of the Drosophila
HSP 70 gene in Drosophila cells (3) or germline
transformants (87) indicated that deletion mutants which
retained only the TATA-proximal HSE showed only 1% of the
normal heat-induced expression. Thus, at least two copies
of the HSE are needed for high levels of heat shock induced
expression for Drosophila heat shock genes (87). This is
consistent with the observation that multiple HSEs are
required for maximal expression of heat shock genes in plant
cells (71, 105). These results suggest the cooperative
binding of multiple heat shock factors (HSFs, see below) to
separate HSEs because the binding of HSF to HSE is necessary
for the activation of heat shock genes (278).
The sequences immediately flanking the 14 base pair HSE
have been demonstrated to play a role in the heat shock gene
expression by using HSP 70-LacZ fusion genes containing
variant synthetic regulatory regions in Drosophila (167).
The importance of sequences flanking HSEs has been
reinforced by mutational analyses of the Drosophila HSP 70
gene (4). The employment of high resolution methylation
interference mapping also supports the importance of three
to four bases flanking HSE for optimum binding of HSF (258).
Based on the observations of in vivo assays of expression of
HSP 70 gene containing variant synthetic regulatory regions,
19

it has been suggested that heat shock regulatory elements in
gene promoters are composed of contiguous arrays of a 5 bp
unit sequence, -GAA-, in alternating orientations (167).
When normal upstream regulatory region of HSP 70 gene is
replaced with two copies of -TTC--GAA--TTC--GAA- which are
separated by 11 bp, This gene is expressed at a 6-fold
higher level than HSP 70 gene containing the pair of perfect
14 bp consensus sequences and at a 5-fold higher level than
HSP 70 gene containing two native HSEs separated by 11 bp
(167, 306). This is supported by the evidence that heat
shock factor can form a stable complex in vitro with an
inverted repeat of 5 bp recognition unit, -GAA- (218).
Heat shock factor. Studies of heat shock gene
regulation indicate that a transacting protein, heat shock
factor (HSF) , is involved in the activation of heat shock
genes (1, 24, 49, 83, 208, 301, 302) even an HSF-independent
mechanism for heat shock induction of transcription has been
recently described (139). The HSF is present in some form
prior to heat shock because heat shock genes can be
activated without protein synthesis (9, 312). The binding
of HSF to HSE-containing regions of the promotor upon heat
shock is supported by studies of DNase I hypersensitive
sites (301, 302). This is reinforced by similar recent
observations in mouse embryonal carcinoma (EC) cells (181).
Several correlations suggest the importance of HSF in the
20

induction of HSP genes by using heat shock inducible and
non-inducible EC cell lines. These include the high
spontaneous expression of some HSPs and the constitutive
level of HSF activity at normal temperature, the heat
induction of HSP gene transcription and marked increase in
HSF activity in inducible cells, and the deficiency in heat
activation of HSP genes and the loss of HSF activity in non
inducible cells upon temperature upshift. Moreover, yeast
HSF and the Drosophila HSF can bind to each other's HSEs
with approximately equal affinities (297).
The interaction between HSF and HSE for heat shock gene
activation and the heat activation of chimeric genes with
same heat shock upstream region in transformed cells of
different organisms suggest that the HSF is also
functionally conserved. This is supported by the isolation
and characterization of HSFs in yeast (265, 298) and tomato
(245). Sequence analysis indicates that yeast HSF is
composed of 833 amino acids with a mass of about 93 Kd. A
118 amino acid region between positions 166 and 285 has been
defined as the DNA-binding domain. However, analysis of
this domain did not reveal resemblance to any currently
known secondary structural motifs implicated in DNA
recognition and binding. The HSF may have a novel secondary
structural motif involved in a DNA binding (298).
21

The HSF exists in an inactive form in certain non-heat
shocked cells (9, 191, 312). In human and Drosophila cells,
pre-heat shock HSF is not able to efficiently bind HSE but
acquires strong HSE-binding activity upon temperature
upshift (136, 264, 312, 313). The HSF is interconverted
between active and inactive forms in dynamic response to
heat shock and recovery in the presence of protein synthesis
inhibitor, cycloheximide. It is highly likely that some
posttranslational modification is involved in this
interconversion but the exact nature of this change has not
yet been identified (312). Recent data indicate that human
HSF can be activated to bind to HSE by treatment of pH,
calcium and non-ionic detergent in vitro (186). A role of
possible interaction between HSP 70 and HSF in the
expression of all HSPs has been recently suggested by Craig
and Gross (67).
Heat shock does not modify the HSE-binding activity of
yeast HSF, in contrast. The yeast HSF binds HSE at all
temperatures both in vitro (264, 265) and in vivo (122) and
HSP genes are activated only after heat shock (264). Thus,
the regulation of HSF-mediated activation of HSP genes does
not occur at the DNA-binding step but involves the
sUbsequent ability to activate transcription.
Posttranslational activation of yeast HSF may be due to
heat-induced phosphorylation. The mobilities of protein-HSE
22

complexes from heat shock and control are different and this
difference can be significantly reduced by the treatment of
crude heat shock extracts with phosphatase (264, 265).
RNA polymerase II must be involved in heat shock gene
transcription because it is responsible for mRNA synthesis.
Studies on the Drosophila HSP 70 gene demonstrate that this
enzyme is associated with promotor region (between
nucleotides -12 to +65 relative to the transcription start
site at +1) in the absence of heat induction (101). RNA
polymerase II partially transcribes Drosophila HSP 70 gene
and forms a nascent RNA chain of about 25 nucleotides at
normal temperature. Additional transcription can not
continue without heat induction (236). Thus, heat shock
promotor appears to be ready for transcription with the TATA
factors constitutively bound and RNA polymerase engaged in
transcription, but complete transcription is impossible
until HSF is activated. However, how the heat shock signal
is perceived and transduced and finally triggers the
activation of HSF is still unknown.
TRANSLATIONAL REGULATION
Structure of heat shock mRNA. Inspection of heat shock
mRNA structure reveals an unusually long 5' untranslated
23

leader sequence (longer than 200 base pairs). This region
appears to contain two conserved sequences located in the
middle and at the 5' end. Both are rich in adenosine
residues with little secondary structure (108, 116, 119,
163, 164, 266). The translational preference of Drosophila
HSP mRNAs during heat shock is suggested to be due to this
relatively long 5' untranslated leader sequence (116, 137).
This is reinforced by the observations that chimeric
messages from fused genes which contain HSP 70 gene promotor
and untranslated leader sequences are translated at high
temperature (34, 84). A complementary result was obtained
from the analyses of deletion mutants in the 5' untranslated
leader of HSP 70 gene (178). The gene carrying a large
deletion of 204 nucleotides from the total 242 nucleotide
leader was transcribed very efficiently during heat shock,
but the mRNA was not translated. These results clearly
demonstrate that the 5' untranslational leader sequence is
an important factor in determining selective translation
during heat shock.
The identification of the precise sequence required for
selective translation in the 5' leader sequence has proven
difficult. The length of the 5' leader sequence is not a
determining factor for selective translation (164, 178). A
message from a reconstructed HSP 70 gene carrying inverse
orientation of DNA fragment of nucleotides +2 to +205 in
24

leader sequence is not translated during heat shock (164).
In contrast, messages from mutated HSP 70 and HSP 22 genes,
in which nucleotides from +37 to +205 and from +27 to +242
were deleted respectively, were translated during heat shock
(117). These results imply the importance of a conserved
sequence at the 5' end because it was not deleted in these
deletion mutants. However, deletion of the 5' conserved
sequence or conserved middle sequence, even both conserved
sequences does not affect the translation of messages from
these mutants (164, 178).
A possible explanation for these results is that more
than one specific sequence in leader including the 5'
conserved sequence is required for selective translation.
This view is reinforced by experiments with two mutants
containing large deletions, one including the 51 conserved
sequence and another not. Results indicated that the
message lacking this conserved sequence was not translated
during heat shock but the message containing this conserved
sequence was translated (164, 178). However, comparison
shows that sequences homologous to the conserved motifs in
Drosophila heat shock messages are not found in soybean
(250). Using deletion mutants of soybean heat shock gene
Gmhsp 17.3-B promotor including the leader sequence and a
reporter gene (CAT) encoding sequence, Schoffl et al (1989)
demonstrated that effective translation during heat shock
25

requires sequences in heat shock mRNA leader region. But,
these sequences can be functionally replaced by the 5'
leader sequence of the CaMV 358 promotor (251). These
results suggest that secondary structure, not certain
sequences in the leader region may play an important role in
translational efficiency of heat shock messengers (164,
250). Kozak (1988) demonstrated a dramatic reduction of the
translational efficiency under hypertonic stress if the
leader sequence of the reporter gene (preproinsulin II gene)
mRNA is modified by incorporation of secondary structure
elements (142). It is conceivable that a low potential for
secondary structure formation, as indicated in heat shock
leader sequence, is a prerequisite for Cap-independent
unwinding and initiation of mRNA translation during heat
shock (206).
certain viral RNAs are translated efficiently in the
absence of active Cap binding factors (216, 217) or during
heat shock (77). These data favor the importance of having
little secondary structure in the leader region for
selective translation. This is reinforced by the recent
evidence that heat shock impairs the interaction of the cap
binding protein complex with mRNA 5' cap (148, 149). The
cap-binding protein complex binding to mRNA 5' cap with
several other initiation factors mediates binding of the
small ribosomal subunit to mRNA. Part of the function of
26

these factors appears to be the unwinding of mRNA secondary
structure facilitating ribosome binding. As complex binding
to cap is impaired during heat shock, unwinding must also be
impaired. This will result in most mRNA being translated
poorly. However, mRNAs with little secondary structure,
such as HSP mRNAs, will have a selective advantage.
Cellular Components. One important change that occurs
in cells during heat shock is the rapid disaggregation of
non-heat shock polysomes and formation of heat shock
polysomes (131, 160, 180). This rapid decay of control
polysomes is evidently a typical stress phenomenon. The
cytoskeleton, which is thought to be necessary for efficient
translation under non-stress conditions (156), is disrupted
during temperature upshift (25, 93). However, translation
under heat shock conditions proceeds largely independent of
cytoskeleton on free polysomes (93, 131). This difference
may be important for translational regulation.
One aspect of the characterization of the mechanisms of
translation is the identification of cellular factors
involved in selective translational regulation. Lysates
from control and heat-shocked cells were used for cell-free
translations and the results indicate that heat shock
lysates do not contain factors that inhibit the translation
of control mRNAs in control lysates. Control lysates do
27

contain a factor that stimulates the translation of control
mRNAs in heat shock lysates (240, 254, 255). In some cases
this stimulatory activity was found in the ribosomal pellet
(254, 255), or in the supernatants in others (240). This
discrepancy may be due to different methods of fraction
preparation. These results suggest that selective
translation during heat shock may be controlled, in part, by
heat-induced change of a factor that is required for the
translation of control mRNAs.
Analyses of ribosomal proteins indicate that some
proteins are phosphorylated while other are dephosphorylated
during heat shock (243, 244). Of these, a ribosomal
protein, 86, may be a good candidate as a cellular
determining factor in translational control. This protein
is rapidly dephosphorylated when plant cells are subjected
to heat shock and rephosphorylated after cells are returned
to normal temperature (243, 244). considering its
localization in the neck region of the small ribosomal
sUbunit, which is part of the initiator tRNA binding site
(28), 86 could be involved in selectively translational
control during heat shock. However, prolonged incubation of
tomato cells under heat shock (12 hr at 37°C) results in
restoration of normal protein synthesis while 86 remains
unphosphorylated (243).
28

The possible role of protein synthesis initiation
factors in selectively translational control during heat
shock has also been studied. Heat shock results in the
activation of a protein kinase which phosphorylates the a
subunit of the eukaryotic initiation factor of eIF-2 (eIF
2a) and phosphorylated eIF-2a is dephosphorylated by a
specific phosphatase (29, 78, 88, 91). Further analyses
indicate that HSP 90 may be involved in the phosphorylation
of eIF-2a because it is contained in a highly purified
preparation of the heme-controlled eIF-2a kinase of rabbit
reticulocytes (144, 233). HSP 90 does not phosphorylate
eIF-2a or inhibit the eIF-2-mediated binding of Met-tRNAf to
40S ribosomal subunits. HSP 90 has been suggested to
increase the kinase activity based on the observation that
enzymatic activity of the eIF-2a kinase is markedly
increased by addition of the purified HSP 90 (144, 233,
234). The phosphorylation of eIF-2a leads to a failure of
guanine nucleotide exchange on the initiation factor and
subsequent inhibition of protein synthesis (78, 234). These
results appear to implicate eIF-2a phosphorylation as a
contributory mechanism in the inhibition of translational
activity immediately following heat shock treatment and
indicates a possible role of HSP 90 in the regulation of
eIF-2a kinase activity (233, 234). The role of eIF-2a
phosphorylation in regulating heat-shock translation is
still controversial because others have not found this
29

phosphorylation (174). Recently, eIF-2a has been indicated
to be a heat shock protein because its synthesis is
stimulated by heat shock (58). This stimulation is even
greater during the recovery period, suggesting that the
induction of eIF-2a in the HSR may be important in restoring
the ability to initiate normal protein synthesis (58).
Many lines of evidence suggest that a specific quantity
of functional HSPs in a given heat treatment is needed for
the recovery of normal protein synthesis during recovery
period (81, 164). Drosophila HSP 70 is always the first
protein to be repressed and the time at which HSP 70 reaches
approximately 50% repression always coincides with the time
at which normal protein synthesis reaches 50% recovery
during recovery period (164). Furthermore, the time when
normal protein synthesis is restored and HSP 70 synthesis is
repressed correlates with the time when HSP 70 moves from
the nucleus to cytoplasm (286). Therefore, HSP 70 may play
a role in recovery of control message translation and in
suppression of heat shock message translation (164, 183).
Results from the distribution of small HSPs suggest a role
in conservation of untranslated control mRNAs during heat
shock. A major portion of these small HSPs is contained in
heat shock granules carrying sequestered control messages
(200). HSPs may also act to increase the instability of
30

heat shock messages during recovery because heat shock
messages are stable in the absence of HSPs, even when cells
are returned to normal temperature (81, 82).
CALCIUM, CaM AND CaM-BINDING PROTEINS IN THE HSR
Calcium and cAMP are two major second messengers for
signal perception and transmission in animal cells (21, 222,
223). However, cAMP is apparently not a second messenger in
plants (175, 221, 222) although there is convincing evidence
for the existence of cAMP in plants (36). Calcium has been
shown to mediate various plant physiological processes
elicited by extracellular signals such as light, hormones
and gravity (222). In plant and animal cells, the free
calcium concentration is submicromolar in the resting state,
rising up to as high as micromolar during excitation (2).
Cells may regulate cytoplasmic calcium in a number of ways,
including membrane permeability, calcium channels, inositol
1,4,5-triphosphate (IP3 ) , calcium-induced calcium release,
Ca2+/H+ and Na+/Caz+ exchange, and Caz+-ATPase (2). Of these
IF3 , the product of phospholipase C-catalyzed breakdown of
phosphatidylinositol-4,5-bisphosphate (PIPz), has been
suggested to be involved in heat-induced increase of
cytosolic Ca 2+ (48, 270). Another product of PIPz breakdown,
diacylglycerol (DAG), stimulates the activity of protein
31

kinase C, multifunctional serine/threonine specific protein
kinases (183, 209, 210).
A number of studies have shown that rapid increase of
cytosolic calcium ([ca2+ ] J is a common feature in a variety
of organisms during heat shock (47, 48, 86, 147, 151, 270).
The resting level of [ca2+ ] j in Drosophila melanogaster
larval salivary gland cells is about 200 nM and increases
approximately 10-fold, to 2 MM during heat treatment (85,
86). This increase of [Ca2+ ] i is very rapid with the
concentration doubling by two minutes and increasing up to
five-fold by five minutes after initiation of heat shock
(270). The close correlation of this change with heat
induced generation of IP3 suggests that the phosphoinositide
pathway may be involved in the modulation of cytosolic
calcium concentration during heat shock (48, 270). Several
groups suggest that heat shock alters cellular Ca 2+ through
Ca2+ influx into cytoplasm from both internal stores and
external medium (47, 48, 270). The rapidity and ubiquity of
heat shock-induced increase in cytosolic calcium
concentration underscore the possible importance of calcium
in the HSR.
Cells treated with the ionophores valinomycin,
dinactin, A23187 or ionomycin synthesize HSPs (9, 225, 293,
304). The long-term depletion of cellular calcium with EGTA
32

in rat hepatoma and Chinese Hamster Ovary cells inhibits the
HSR (147). However, some contradictory results have been
recorded in experiments in which the Ca2+ was maintained at
a low level during heat shock. Calcium-depleted Drosophila
salivary glands and BAPTA-Ieaked Kc cells maintain a low
[Ca2+]i level during heat shock but are competent to
synthesize a complete set of HSPs (85, 86). This suggests
that Ca 2+ is not essential or extremely small amounts of Ca2+
are capable of inducing HSP synthesis.
One type of intracellular calcium target which may be
involved in signal transduction in eukaryotic cells is a
class of calcium-binding proteins represented by calmodulin
(CaM) (for review, see 175, 222). Calmodulin has been
isolated and characterized and CaM genes have been cloned
from many different organisms (14, 125, 166, 193, 314, for
reviews, see 221, 222, 230). Calmodulin is a highly
conserved, heat-stable, acidic protein with four Ca2+
binding domains and is ubiquitous among eukaryotes. The
Ca2+/CaM complex may, directly or indirectly, regulate
activities of many enzymes such as ATPase, NAD kinase, H+
ATPase, quinate:NAD+ oxidoreductase, phospholipase, protein
phosphatases and protein kinases. Many lines of evidence
demonstrate that levels of CaM differ with respect to tissue
type as well as on the physiological and developmental state
of cells (26, 125, 170, 221, 230). The level of CaM
33

apparently increases two to three-fold during heat shock
(Harrington HM, personal communication). A sequence similar
to HSE in the upstream region of Chlamydomonas CaM gene
promotor may provide the molecular basis for such heat
regulation (314).
using CaM inhibitors, Wiegant et al (299) concluded
that CaM antagonists sensitize cells to heat by inhibiting
cytoskeleton rearrangements mediated by ca2+jCaM. The CaM
antagonistic drug W13 potentiates hyperthermic cell killing
but the nonfunctional analog W12 has little influence,
supporting the idea that ca2+jCaM mediated processes are
involved in the HSR (151). This view has been reinforced by
the findings that some HSPs, such as HSP 70 (56, 271), 90
(192) and 100 (141) in animal systems are cytoskeleton
associated, Ca2+jCaM binding proteins. Taken together,
these data underscore the potential importance of Ca2+jCaM
mediated processes in the HSR.
Recently, Landry and co-workers demonstrated that
transcriptional activation of the HSP 68 gene by heat and
cytoplasmic accumulation of mRNA are considerably reduced in
cells incubated prior to heat in EGTA-containing medium and
suggested that the block occurs very early at a
pretranscriptional site (151). These data, together with
several other important observations including a.) that
34

Ca2+/caM stimulates activities of many protein kinases
(253); b.) that phosphorylation may be involved in the
activation of HSF (264); and c.) that many proteins are
modified with phosphorylation/dephosphorylation during heat
shock (45, 46, 233, 234, 243, 244) provide the rationale for
the characterization and identification of Calmodulin
binding proteins (CaMBPs) in the HSR.
35

CHAPTER II
SIGNIFICANCE AND HYPOTHESIS
As discussed in Chapter I, Ca2+jCaM mediated processes
may be involved in the HSR of animal systems but no similar
information is available in plants. This lack of
understanding of the Ca2+jCaM system in plants provides the
rationale for the present studies. The characterization of
heat-induced genes for CaMBPs and the subsequent
identification of functions for these proteins are both
theoretically and practically important. In theory,
identification and characterization of CaMBP genes will
enhance our understanding of how cells perceive
environmental signals and how such stresses affect
physiological and biochemical processes. This research will
also enhance our understanding of molecular mechanisms of
gene regulation in plants. Finally, the elucidation of
molecular mechanisms of the HSR will provide clues which
will be ultimately useful in the development of stress
resistant crops.
The hypothesis to be tested in this research is:
The expression of some genes for CaMBPs is regulated by
heat shock in cultured tobacco cells.
36

This hypothesis will be tested by following specific
objectives:
I. To characterize CaMBPs in cultured tobacco cells
during heat shock;
II. To isolate eDNA clones for CaMBPs;
III. To characterize a eDNA clone encoding a heat
shock-induced CaMBP;
IV. To analyze the transcriptional expression of the
cloned CaMBP genes;
V. To characterize calmodulin-binding domains by
deletion analysis.
37

CHAPTER III
CHARACTERIZATION OF HEAT SHOCK INDUCED
CALMODULIN-BINDING PROTEINS IN CULTURED TOBACCO CELLS
INTRODUCTION
Calcium is an important second messenger which mediates
various plant physiological processes elicited by
extracellular signals such as light, hormones and gravity
(21, 222, 223). One target involved in signal transduction
in eUkaryotic cells is a class of Ca2+-binding proteins
exemplified by calmodulin (CaM). This 17 Kd, heat-stable,
acidic protein contains four EF hand ca2+-binding domains
(221, 230). Calmodulin regulates activities of many enzymes
including ATPase, NAD kinase, phospholipase, quinate:NAD+
oxidoreductase, protein phosphatases and kinases in a Ca2+
dependent manner (221, 222, 230).
A number of studies have shown that rapid increase of
cytosolic calcium is a feature common to variety of
organisms during heat shock (47, 48, 86, 147, 151, 270).
The control cells (not heat shocked) treated with the
ionophores valinomycin, dinactin, A23187 or ionomycin
synthesize HSPs (9, 225, 293, 304). Alternatively, long
term Ca2+-depletion with EGTA inhibits the HSR (147).
38

Recent evidence demonstrates that calcium activates HSF
binding activity to HSE in vitro (186). Taken together,
these findings imply that calcium may be involved in the
HSR. other studies indicate that calcium is not necessary
for HSP synthesis per se (85, 86). A role of CaM in the HSR
may be implied since the anti-CaM drug, W13, potentiates
hyperthermic cell killing while the non-functional analog,
W12, has little influence (151). This view is reinforced by
the findings that some proteins, such as HSP 70 (56, 271),
90 (192) and 100 (141) from animal cells are CaM-binding
proteins (CaMBPs). These results serve to underscore the
potential importance of the ca2+/CaM second messenger system
in the HSR. However, similar information is unavailable for
the plant HSR. Few, if any, studies have systematically
analyzed CaMBPs during the HSR and there are limited data on
CaMBPs in plants. This report details the characterization
of CaM-binding HSPs in cultured tobacco cells.
MATERIALS AND METHODS
Materials. Tobacco cells (Nicotiana tabacum L. cv
Wisconsin-38) were grown at 23°C in the dark as suspension
cultures in Gamborg's B-5 medium (98). Cell cultures were
maintained by transferring 6 ml of mid log phase (7 days
old) culture into 70 ml fresh B-5 medium contained in a 250
39

ml erlenmeyer flask. Cells from mid log phase cultures were
used in all experiments.
Isolation of CaM-binding proteins. Tobacco cells
(5g/15 ml culture) were incubated in shaking water baths for
15 minutes at 23°C for control and 38°C for heat shock
treatment and then 250 ~Ci 35S-Trans Label (lCN) was added.
The cultures were continued for 4 hours at the same
temperatures. The labeled cells (20g) were ground into fine
powder under liquid nitrogen and extracted with buffer A (3
ml/g cell) containing 50 roM Tris/HCI pH 7.5, 3 roM MgCI2 , 5
roM KCI and 0.2 roM EDTA at 1°C. This extract was centrifuged
at 20,000g for 20 minutes. The supernatant was adjusted to
1 roM CaCl2 final concentration and applied to a CaM
sepharose-4B column (5 ml) (292). The column was washed at
flow rate of 0.9 ml/minute at 1°C and 4.5 ml fractions were
collected. The column was step eluted with 25 column
volumes each of buffer B (25 roM Tris/HCI pH 8.0, 3 roM MgCI2,
2 roM KCI and 0.1 roM CaCI2 ) , buffer B plus 0.15 M NaCI,
buffer B plus 0.3 M NaCI. The putative CaMBPs were eluted
in buffer Blacking CaCl2 with the addition of 1 roM EGTA.
The fractions containing peaks of radioactive materials were
exhaustively dialyzed against 20 roM ammonium bicarbonate,
lyophilized and resuspended in 2X Lamelli sample buffer
(145). This procedure did not result in obvious protein
degradation. Experiments in which crude extracts and other
40

fractions from the CaM-sepharose-4B columns were allowed to
stand for up to 10 hours at 1°C revealed little proteolysis
as evidenced by SOS-polyacrylamide gel electrophoresis (SDS
PAGE).
Analysis of labeled proteins. Labeled proteins were
detected on SOS-mini gels as previously described by
Harrington and AIm (111). Samples containing equal amount
of radioactive proteins were loaded on 12.5% SOS
polyacrylamide gels. After electrophoresis, gels were
stained with Coomassie Blue, destained, dried and
autoradiographed at -BOOC to locate labeled proteins.
Incorporation of radioactive amino acids into proteins was
estimated by the method of Mans and Novelli (172). All
radioactivities were determined by liquid scintillation
spectrometry.
RESULTS AND DISCUSSION
calmodulin-sepharose chromatography of tobacco
proteins. Initially, 8.5 x 108 and 5.8 x 108 cpm of
radioactive labeled materials for control and heat shock
treatments respectively were loaded onto CaM-sepharose-4B
columns (5 ml) and eluted with over 25 column volumes (130
ml) of each wash buffer. At the end of each wash the
41

radioactive counts (cpm) stabilized at a low level (95-450
cpmj50 ~l). Results in Figure 1 indicated similar elution
profiles for control and heat shock samples. The wash of
the columns with buffer B plus 0.15 M NaCI resulted in the
elution of a radioactive peak containing 3412 cpmj50 ~l for
control and 1322 cpmj50 ~l for heat shock treatments. A
similar smaller peak was observed when the columns were
washed with buffer B plus 0.3 M NaCI (1249 cpmj50 ~l for
control and 641 cpmj50 ~l for heat shock treatments). The
possibility that CaMBPs were eluted in salt washes (0.15 and
0.3 M NaCI) can not be excluded because the binding of a
known CaM-target, myosin light chain kinase is easily
disrupted by relatively low levels of salt (0.1 M). Elution
of the columns with buffer B plus EGTA produced a large
radioactive peak (9720 cpmj50 ~l for control and 4527 cpmj50
~l for heat shock treatment). The fact that EGTA washes
released labeled materials after high ionic strength salt
wash suggests that these peptides exhibit highly specific,
ca2+-dependent binding activity. These fractions and other
column effluents were analyzed by 5DS-PAGE.
To confirm the binding specificity of these putative
CaMBPs, parallel experiments were run using sepharose-4B
columns. Equal amounts of radioactive proteins (5.8 x 10 8
cpm) of heat shock extracts were loaded into CaM-sepharose
4B and sepharose-4B columns and eluted as described above.
42

otoEQ.o
1e+07 f--------·----------··-----··---· '-1~ Control -- HS~ --I
1000000 ~"'"
100000 I=~n:. ~rrM Ca_i,{'!~;: EGTA Wash
:J ------
f t o. ~5M _~I!~~sh \
10000 L \; '" •
E \.... -,..O.3~W_~ ~~1000 t -. ~...... ~:... 1'\
c\,. ~'\" . . .
100 f ''c~ ~~ Z·.... IF ~-I~ I10 I L_. --L L . L ~
o 28 56 84 112 140
4.5 ml/fraction
Figure 1. CaM-sepharose chromatography elution profile
43

Results summarized in Table 1 indicate that few radioactive
proteins bound to sepharose-4B. If the data are expressed
as the ratio of sepharose-4B/CaM-sepharose-4B, 2.9% of the
counts were released from sepharose-4B by the 0.3 M salt
wash. Only 1.6% of the counts were eluted by EGTA wash.
These results suggest that the peptides released from CaM
sepharose-4B with EGTA washes specifically bind to CaM and
not to the column supports.
Analysis of labeled proteins by SDS-PAGE. Equal
amounts of labeled proteins (30,000 cpm/lane) were separated
on 12.5% SDS-gels. The results in Figure 2 show a
comparison of total labeled proteins extracted with 2X
Lamelli buffer from control (lane A1) and heat shocked cells
(lane B1). These data indicate strong induction of HSPs,
especially HSP 96, 82 and 71 and abundant small HSPs in
agreement with Harrington and AIm (111). Few constitutive
proteins were synthesized during heat shock. The extraction
of cells with Buffer A resulted in the detection of
significantly fewer labeled bands for both treatments (Fig.
2, lanes A2 and B2). For example, HSP 96 was not extracted
by this procedure (Fig. 2, lane B2). Chromatography of
these Buffer A extracts on CaM-sepharose-4B as above
resulted in the elution of specific peptides.
44

Table 1. comparison of radioactive proteins recovered fromCaM-sepharose-4B and sepharose-4B columns
Column material 0.3 M NaCl EGTA(cpm) (cpm)
CaM-sepharose-4B 2.94 ), 105 3.2 X 105
sepharose-4B 7.9 x 103 5.2 X 103
sepharose-4B/CaM-sepharose-4B 2.9% 1.6%
45

.. 2
kD
94-67-
i" ~ :"~"- ,43
30-
20-
14-
A
3 4 5
B678 .. 2345678
r'""1,"
Figure 2. SDS-PAGE of CaM-sepharose chromatographyfractions.
Equal amounts of labeled proteins (30,000cpmjlane) wereseparated on 12.5% SDS-polyacrylamide gels. Labeledproteins were visualized by autoradiography.
A, Proteins were isolated from cells treatt:d at 23°Cfor control. B, Proteins were extracted from cells treatedat 38°C for heat shock treatment.
Lane 1, SOS extraction buffer; lane 2, CaM-sepharoseextraction buffer A; lane 3, Initial non-bound fraction;lane 4, 0.1 roM Ca2+ wash; lane 5, 0.15 M NaCI wash; lane 6,0.3 M NaCI wash; lane 7, 1 roM EGTA wash after 0.15 M NaCIwash and Lane 8, 1 roM EGTA wash after 0.3 M NaCI wash.
46

When the CaM-sepharose-4B wash buffer Ca2+
concentration was decreased to 0.1 roM, a unique HSP 25 (25
Kd) was eluted (Fig. 2, lane B4). There were also other
major bands corresponding to 85, 82 and 71 Kd of HSPs in
this elution. A 41 Kd peptide was present both in control
and heat shock samples (Fig. 2, lanes A4 and B4). Column
elution with salt-containing buffers released additional
labeled peptides with major HSP bands at molecular weights
of 90, 85, 82, 71, 68, 34, 32, 22, 21 Kd (Fig. 2, lane B5
and B6) .
The EGTA washes were used to release peptides
exhibiting Ca2+-dependent binding to CaM-sepharose-4B.
These fractions contained the putative CaMBPs. with the
exception of a HSP 17 (17 Kd), the profiles for heat shock
samples were similar to those obtained with salt wash
samples (Fig. 2, lanes B5, B6, B7 and B8). Our recent data
with gel overlay analyses indicated that the proteins
released by EGTA washes bound to 3sS-labeled calmodulin while
the proteins obtained with salt washes did not, suggesting
specific CaM-binding of the proteins in EGTA wash samples
(Data not shown). These EGTA wash fractions contained
heavily labeled heat shock peptides with molecular weights
of 90, 85, 82, 78, 71, 68, 22, 20, 19.5 and 17 Kd. By
comparison, control EGTA fractions contained few distinctive
47

labeled bands including peptides with molecular weights of
100, 50 and 35 Kd (Fig. 2, lane A7 and AB).
The above results indicate that heat shock induced the
synthesis of CaMBPs/HSPs in cultured tobacco cells. These
findings are consistent with recent discoveries in animal
system that HSP 100, 90 and 70 are CaMBPs (141, 192, 271).
The synthesis of CaMBPs during heat shock suggests that the
Ca2+/CaM second messenger system may playa role in the
plant HSR. However, the identities and functions of the
CaMBPs/HSPs in tobacco are currently unknown. Possible
roles could include CaM-dependent protein kinases and other
enzymes that regulate existing metabolic processes, repair
heat damage or protect cellular structures during heat
shock. This first indication that CaMBPs/HSPs are involved
in tobacco HSR provides the rationale for future
characterizations of Ca2+/CaM-mediated processes during heat
shock in plant cells.
since most CaMBPs are present in extremely small
amount, it would be difficult to purify sufficient amounts
of the protein for biochemical identification and molecular
analysis. An alternative approach to the identification of
CaMBPs is to isolate eDNA clones for CaMBPs by screening an
expression cDNA library with radioactive labeled CaM. The
nucleotide sequences from such clones may provide clues as
48

to identities of CaMBP/HSPs and may suggest biochemical
assays to confirm functions.
49

CHAPTER IV
ISOLATION AND CHARACTERIZATION OF eDNA CLONES
ENCODING TOBACCO CALMODULIN-BINDING PROTEINS
INTRODUCTION
All organisms are sUbjected to a large number of
environmental and biological stresses. One of the important
environmental factors for plants in agriculture is heat
shock. Plants, when subjected to a moderate upshift
temperature, undergo a phenomena called heat shock response
(HSR). The HSR is characterized by the rapid shutdown of
most normal mRNA and protein synthesis; induction of de novo
synthesis of heat shock mRNAs and proteins and the
acquisition of thermotolerance to otherwise lethal high
temperature (66, 132, 163, 189). Much progress has been
made in the understanding of the structure and regulation of
heat shock genes. However, little is known about the
biochemical function of individual HSPs.
Calcium, acting as a second messenger, plays a role in
various plant physiological processes elicited by
extracellular signals such as light, hormones and gravity
(175, 222). As in animal cells, many of the effects of
calcium ions in plant cells are mediated through the calcium
50

responsive element, calmodulin (CaM) (114, 175, 221, 230,
281). Calmodulin is a small, heat-stable, acidic protein
with four EF hand Ca2+-binding domains that has been
identified in all eukaryotic species examined (230). It is
highly conserved in structure and is functionally
interchangeable among different species (76, 230),
suggesting that CaM is an essential protein for normal
growth and development performing similar functions in all
eukaryotes (75, 230). The ca2+/caM complex regulates
activities of many enzymes including Ca-ATPase, NAD kinase,
phospholipases, protein phosphatase and a variety of kinases
(221). To date, few CaM-dependent processes have been well
characterized in plants (222, 230). This lack of
information on the role of CaMBPs has hindered the
development of an integrated view of signal transduction in
plant cells. Furthermore, a number of studies suggests that
ca2+/caM mediated processes may be involved in the HSR (47,
48, 147, 151, 186, 271, 293) even though some data suggest
that calcium is not necessary for HSP synthesis (85, 86).
Therefore, the characterization of CaMBPs will enhance our
understanding of mechanisms of how CaM acts to regulate
physiological, metabolic and molecular processes including
the HSR in plants.
Calmodulin-binding proteins have been extensively
studied in animal systems (175, 203). Characterization of
51

CaM-binding sites of several animal and human CaMBPs have
defined a basic amphiphilic a-helix as the common feature of
CaM-binding domain (27, 73, 157, 169, 289). This structure
is characterized by the presence of basic amino acid
residues on one side of the helix while hydrophobic residues
on the opposite side. Short synthetic peptides containing
this structure can efficiently bind to CaM (73, 157, 169,
289). Moreover, two groups have reported successful
screening of expression cDNA libraries in animal systems
with 125I_ or 3sS-CaM (259,296). Such results provide the
rationale for the isolation of plant CaMBP cDNA clones from
an expression library using CaM as a ligand probe. Here we
report the results of isolation and identification of cDNA
clones for heat shock-induced and constitutive CaMBPs in
tobacco and tentative characterization of the CaM-binding
domain of the deduced protein.
MATERIALS AND METHODS
Tobacco cells (Nicotiana tabacum L. cv Wisconsin-38)
were grown at 23°C as suspension culture in Gamborg's B-5
medium (98). Cells from mid log phases culture (7-dayold)
were incubated for 2 hours at 38°C for heat shock and at
23°C for control treatments. These cells were used in all
experiments. Escherichia coli UT481 harboring pVUC-l for
52

preparation of 35S-CaM was a gift from D. Martin Watterson
(Vanderbilt University) (231).
Preparation of 3SS- CaM • The isolation of 35S-CaM was
done essentially as described by Asselin et al (10). An
overnight culture (200 ~l) of E.coli UT481 harboring pVUC-1
was inoculated into 20 ml of 1% NZ amine, 0.5% yeast
extract, 0.5% NaCI, 0.1% casamino acids, 0.2% MgS04 , pH 7.5,
2.5% glycerol and 25 ~g/ml ampicillin and incubated at 37°C
until the ~ of the culture reached 0.6. The bacteria were
collected by centrifugation (1000g, 5 minutes) and
resuspended in 50 ml of 0.1 M Tris/HCl pH 7.4, 92 roM NaCl,
40 roM KCl, 19 roM NH4Cl, 0.26 roM CaCI 2 , 0.98 roM MgC12 , 0.74 ~M
FeCI3, 0.639 roM KH2P04 , 2.5% glycerol and 25 ~g/ml
ampicillin. The cells were centrifuged as above and
resuspended in 20 ml of the latter medium containing 5 mCi
of 35S-sodium sulfate (1200-1400 Ci/roM, NEN) and incubated
for 3 hours at 37°C. The incubation was continued for an
additional 3 hours after addition of 16 ~l of 0.5 M IPTG.
The bacteria were collected and resuspended in 5 ml of 25 roM
Tris/HCI pH 7.5, 1 roM CaC12 and lysed by sonication (10).
The mixture was heated at 90°C for 1 minute and centrifuged
at 1000g for 5 minutes. The supernatant was then applied to
a 500 ~l column of phenyl-sepharose in a 1.5 ml microfuge
tube and washed with 1 ml of 25 roM Tris/HCl, IroN CaC12, pH
7.5 and 3 ml of 25 roM Tris/HCI, IroN CaC12, 0.2 M KCI, pH 7.5.
53

The CaM was eluted from the column with 500 ~l of 25 roM
Tris/HCl, 1 roM EDTA pH 7.5. This 35S-CaM was analyzed by
SDS-PAGE and autoradiography. The mobility of 35S-labeled
CaM shifted on the SDS gels in the presence or absence of
CaC12 as expected (Fig. 3, lanes A2, A3, B2 and B3). The
behavior was identical to authentic CaM from chicken gizzard
(Fig. 3). The specific activity of labeled CaM was
approximately 1.5 x 107 cpm/~g.
Isolation of total and polysome RNAs. Isolation of
total RNAs was done as described by McGookin (179).
Filtered tobacco cells (5g) were frozen with liquid nitrogen
and ground into a fine powder in a prechilled mortar and
pestle. Four volumes of 5 M guanidine thiocyanate, 50 roM
Tris/HCl pH 7.5, 10 roM EDTA and 5% ~-mercaptoethanol were
added to the powder and the mixture was homogenized with a
glass homogenizer. Sarkosyl and CsCl were then added to
final concentrations of 4% (W/V) and 15% (W/V) respectively.
After centrifugation at 15,000g for 20 minutes at 4°C, the
supernatant was layered on a 4.5 ml cushion of 5.7 M CSCI,
100 roM EDTA, pH 7.5 and centrifuged at 100,000g for 18 hours
at 20°C. The pellet was resuspended in 200 ~l of TE (10 roM
Tris/HCl, 1 roM EDTA pH 7.5) and precipitated by the addition
of 4% (V:V) 6 M ammonium acetate and 2 volumes of ethanol at
-20°C. The resulting RNA pellet was collected by
54

A IB1 2 3 1 2 3
kD
9467
43
30 --20 ~~,¥;J"i' , .....~:
14 .,..).,.; .....""':.;.
Figure 3. 50S-polyacrylamide gel electrophoresis of 355_labeled calmodulin.
12.5% 50S-polyacrylamide gels containing 5 roM EOTA (A)or 1 roM CaCl2 (B) were used to separate the calmodulin.
Lanes 1 and 2, Coomassie blue stain; lane 3,autoradiography.
Lane 1, Chicken gizzard calmodulin; lanes 2 and 3, 355_labeled calmodulin (1 x 106 cpmjlane) .
55

centrifugation, washed with 70% ethanol, resuspended in H20
and stored at -80 aC until use.
Polysomal RNA was isolated as described by Laroche and
Hopkins (154). Filtered tobacco cells were frozen with
liquid nitrogen and homogenized into fine powder with an
omni Mixer. Ice-cold isolation buffer (3 ml/g cells) (200
roM Tris/HCI pH 8.95, 200 roM KCI, 35 roM MgCI2 , 250 roM
sucrose, 12.5 roM EGTA and 15 roM DTT) was added and the
mixture was homogenized in a Polytron. The mixture was
filtered through miracloth and NP-40 was added to a final
concentration of 1% (V:V). After 10 minutes on ice, the
mixture was centrifuged at 15,500g for 15 minutes. The
supernatant was loaded on a 5 ml sucrose cushion (40 roM
Tris/HCI pH 8.95, 40 roM KCI, 7 roM MgCI2 , 1.5 M sucrose, 5 roM
EGTA and 5 roM DTT) and centrifuged at 180,000g for 1.5 hours
at 4°C. The pellet was rinsed 3 times with 40 roM
Tris/acetate pH 8.5, 20 roM KCI and 1 roM MgCl2 and
resuspended in the same buffer. The resuspension was then
extracted twice with phenol, three times with
phenol/chloroform (1:1) and twice with chloroform. The RNAs
were precipitated with 0.1 volume of 3 M sodium acetate pH
5.2 and 2.5 volume of ethanol at -20°C and centrifuged at
11,000g for 10 minutes at 4°C. The pellet was washed with
70% ethanol, resuspended in H20 and stored at -80°C until
use.
56

construction of expression CDNA library. Poly(A) RNA
was purified from heat shock polysome RNAs by oligo(dT)
cellulose chromatography (11). The RNA sample was heated
for 2 minutes and loaded on a column containing 19
oligo(dT)-cellulose. The column was washed with 10 roM
Tris/acetate pH 7.5, 0.4 M NaCl to remove unbound RNA until
the ~oo was at the baseline. Then, the poly(A) RNA was
collected in a wash of 10 mM Tris/acetate pH 7.5. The RNA
was subjected to the same procedure two more times.
Finally, the poly(A) RNA was precipitated wi~h 0.1 volume of
20% potassium acetate pH 5.5 and 2.5 volumes of ethanol at
20°C. This purified poly(A) RNA was dissolved in H20 and
used (5 ~g) in the construction of a cDNA expression library
with a AZAPII-cDNA synthesis kit (Strategene). The library
was constructed following the manufacturer's instructions.
The synthetic cDNAs with 13 bp EcoRI adaptor (AATTCGGCACGAG)
at the 5' end and 6 bp XhoI site (CTCGAG) at the 3' end were
inserted in the EcoRI/XhoI site of the vector AZAPII which
can be in vivo excised into phagemid pBluescript SK- (Fig.
4) (257). A total of 1.1 x 106 recombinants were obtained.
screening of eDNA library with 3SS-CaM. Escherichia
coli BB4 was infected by recombinant A-ZAP II phages and
grown on NZY (0.5% NaCl, 0.2% MgS04 , 0.5% yeast extract, 1%
NZ amine pH 7.5) plates at the density of 50,000 pfu/plate
for 3 hours at 42°C. Nitrocellulose filters saturated with
57

Xhol KpnlTCGAGGGGGGGCCCGGTACCCAATTCGCCCTATAGTGAGT 3'
CCCCCCCGGGCCATGGGTTAAGCGGGATATCACTCA 5'
pBluescript SKill
2958 Basepairs
MET5' GGAAACAGCTATGACCATGATTACGCCAAGCTCGAAATTA3' CCTTTGTCGATACTGGTACTAATGCGGTTCGAGCTTTAAT
Beta-Galactosidase )Sac.
ACCCTCACTAAAGGGAACAAAAGCTGGAGCTCCACCGCGGTGGTGGGAGTGATTTCCCTTGTTTTCGACCTCGAGGTGGCGCCACC
Xhal EcoRICGGCCGCTCTAGAACTAGTGGATCCCCCGGGCTGCAGGGCCGGCGAGATCTTGATCACCTAGGGGGCCCGACGTCCTTAA
Figure 4. pBluescript SK- vector.
58

10 roM IPTG were layered on the plates and the incubation
continued for an additional 6 hours. The filters were
washed in 100 ml of buffer A (50 roM Tris/HCI pH 7.5, 150 roM
NaCI, 1 roM MgCI2) plus 0.1% BSA for one hour at 37°C. The
filters were then incubated in 50 ml of buffer A plus 1 roM
CaCl2 containing 1 x 106 cpm/ml of 3sS-CaM for 3 hours at room
temperature and washed three times with 100 ml of buffer A
plus 1 roM CaCl2 for 10 minutes each. Dried filters were
autoradiographed at -80°C to locate positive signals. The
positive plaques were further purified by 3 more rounds of
screening as described above.
In vivo excision of positive phages into phagemids.
positive phages were in vivo excised into phagemids as
described by Short et al (257). A mixture of 200 ~l of
E.coli XL1-Blue (~=1.0), 200 ~l of phage (>1 x 105 pfu) and
10 ~l of R408 helper phage was incubated for 15 minutes at
37°C. Five ml of 2xYT medium (1% NaCI, 1% yeast extract and
1.6% tryptone pH 7.4) were added and the incubation was
continued for 3 hours at 37°C. After 20 minutes at 70°C,
the cultures were centrifuged at 4,000g for 5 minutes. An
aliquot of the supernatant (200 ~l) and 200 ~l of E.coli
XL1-Blue were mixed and incubated for 15 minutes at 37°C.
The culture was then distributed on LB plates (1% tryptone,
0.5% yeast extract and 0.5% NaCI pH 7.5) containing 50 ~g/ml
59

ampicillin and incubated overnight at 37°C. Colonies
obtained were used for further analyses.
Gel overlay analysis of positive colonies. A positive
colony was inoculated into LB medium and incubated at 37°C
until the ~ of the culture reached 0.2. IPTG was then
added to 10 roM and the culture growth was continued to ~
of 1.0. The culture was microfuged for 3 minutes and the
pellet was resuspended in 2X Lamelli sample buffer (145).
After 3 minutes in a boiling water bath, the sample was
microfuged for 2 minutes and the supernatant was subjected
to SOS-PAGE as described by Harrington and AIm (111). Gel
overlay analysis was done by the methods of Burgess at al
(40). After electrophoresis the gels were washed three
times in 100 ml of 25% (V:V) isopropanol and 10% (V:V)
acetic acid for one hour each and three times with 100 ml of
buffer B (50 roM Tris/HCI pH 7.6 and 0.2 M NaCI) for one hour
each at room temperature. The gels were then blocked in
buffer B plus 0.1% BSA for 2 hours and then incubated in 30
ml of buffer B plus 0.1 roM CaClz or 5 roM EDTA containing 1 x
105 cpm 3sS-CaM/ml for 14 hours at room temperature. After
three washes with 100 ml of buffer B plus 0.1 roM CaClz or 5
roM EDTA for one hour each at room temperature, the gels were
stained with Coomassie Blue, destained, dried and
autoradiographed at -BOOC to detect CaMBPs.
60

DNA deletion and sequencing. Phagemid ONAs were
purified by CsCl procedure as described by Maniatis et al
(171). Phagemid DNA from clone pTCB40 was double-digested
with SacI/EcoRI for deletion from 5' end of the insert. The
deletion was accomplished with an ExoIII/mung bean deletion
kit (Strategene) essentially as described in the
manufacturer's instructions. Colonies containing
appropriately-sized deletions were selected based on
restriction enzyme digestion and subjected to double
stranded DNA sequencing analysis with a Sequenase version
2.0 DNA sequencing kit (United states Biochemicals).
Sequencing electrophoresis was done on 7 M urea/6%
polyacrylamide gels with constant power (38W) at 50°C.
Northern blot/hybridization analysis. Total tobacco
RNAs (20 ~g) were loaded on a 1.5% formaldehyde agarose gel
and electrophoresed in 20 roM MOPS, 5 roM sodium acetate and 1
roM EDTA pH 7.0. The RNAs were transferred by blotting to
nitrocellulose filters in lOX SSC (1.5 M NaCl, 150 roM sodium
citrate pH 7.0) for overnight at room temperature (277).
The filters were incubated in 50% formamide, 5X Denhardt's
(0.1% Ficoll, 0.1% PVP and 0.1% BSA), 0.1% SOS, 5X SSPE (750
roM NaCl, 50 roM NaH2P04 , 5 roM EDTA pH 7.4) and 100 ~g/ml
salmon sperm DNA for 2 hours at 42°C. The filters were
transferred to 50% formamide, 2X Oenhardt's (0.04% Ficoll,
0.04% PVP and 0.04% BSA), 0.1% SDS, 5X SSPE, 100 ~g/ml
61

salmon sperm DNA containing 3 x 106 cpmjml of 32P-labelled
DNA made from pTCB40 by random primer extension (Random
Primers DNA Labeling System, Bethesda Research Laboratories
Life Technologies, Inc.) for 16 hours at 42°C. After
hybridization the filters were washed twice in 100 ml of 2X
SSC (300 roM NaCl, 30 roM sodium citrate pH 7.0) containing
0.1% SDS for 10 minutes each and twice in 100 ml of O.lX SSC
containing 0.1% SDS for 10 minutes each at room temperature.
A final wash was done in 100 ml of 0.1 x SSC containing 0.1%
SDS for one hour at 55°C. The filters were autoradiographed
at -so°c.
RESULTS AND DISCUSSION
A. Isolation and Confirmation of eDNA Clones Encoding
calmodulin-Binding Proteins
Synthesized cDNA from tobacco polysomaljpoly(A) RNA
was inserted into the bacteriophage expression vector A-ZAP
II and the resultant library was screened for the presence
of plaques containing fusion proteins that bind to
radioactive CaM. Initial screens used I~I-labeled CaM
(lactoperoxidase method); however, the results were negative
and the high background was high. These negative results
may be due to structural damage to CaM caused by the
62

iodination procedure. To overcame these potential problems,
in vivo labeled 35S-labeled CaM encoded by pVUC-1 was used to
screen the library. The results showed distinct positive
spots with very low background (Fig. 5). This method was
very efficient because 25 independent positive clones were
obtained from 1 x 106 screened clones. All these positive
phages were converted into phagemids by in vivo excision as
described in the methods section. The recombinant DNAs from
these clones were isolated and double-digested with EcoRI
and XhoI. These two enzymes were used during construction
of the library and should excise the insert from the vector.
These digested DNAs were analyzed on agarose gel (Fig. 6)
and the sizes of the inserts from these clones are
summarized in Table 2.
In order to confirm that positive clones obtained in
the screening procedure encoded CaMBPs, gel overlay analysis
was carried out on extracted proteins. A polypeptide from
the first clone tested (pTBC01) bound to CaM in the presence
of 0.2 M NaCI and 0.1 mM CaCl2 (Fig. 7, B4). This binding
was readily inhibited by the omission of CaCl2 and addition
of EDTA into the overlay incubation buffer (Fig. 7, lane
C4), suggesting that CaM binding was ca2+-dependent.
SUbsequent experiments demonstrated that all 25 positive
clones synthesized proteins that bound to CaM in a Ca2+
dependent manner (Fig. 8). Some fusion proteins were not
63

•
.. "
....
.o..
..
• o'o.
.... 00.. . ..
..• ... ... :- •
.. ••.... .. II...... ....
/
"'. ..0 •,.''.' ,.... ,.Ii .. ,
..
...",". ..,
..
Figure 5. Autoradiograms of blotted phages screened with"S-labeled calmodulin.
A, Primary screen in which a positive signal (arrow)was evident. B, Screen of purified phages in which allplaques bound calmodulin.
64

Figure 6. DNAs from pBluescript recombinants encodingcalmodulin-binding proteins.
DNAs were double-digested with EcoRI and XhoI andseparated on 1% agarose gel.
Samples for lanes from left to right are pBluescript,pTCB01, 03, 04, 05, 06, 10, 14, 15, 18, 22, 23, 25, 28, 29,31, 38, 40, 46, 48, 51, 55, 56, 58, 59, 60 and molecularweight marker.
65

Table 2. Sizes of the inserts of cDNA clones encodingCaMBPs
Clone # Insert size Clone # Insert size(pTCBxx) (Kb) (pTCBxx) (Kb)
01 3.69 29 5.31
03 2.55 31 1.37
04 2.55 38 1.44
05 2.55 40 0.85
06 1.63 46 1. 64
10 2.45 48 1.48
14 1.20 51 0.80
15 1.11 55 2.55
18 3.39 56 4.38
22 1.06 58 1.39
23 2.73 59 2.11
25 1.44 60 1. 36
28 1.30
66

A B ckD
9467
43
30
20
14
1 234 1 234 1 234
Figure 7. Calmodulin gel overlay analysis of the proteingenerated from clone pTCB01.
Equal proteins (20~g/lane) were separated on 12.5% SDSpolyacrylamide gels and gel overlay analysis was done in thepresence of 0.1 roM caClz (B) or 5 roM EDTA (C) as describedin Materials and Methods.
A, Coomassie blue stain; Band C, Autoradiography.Lane 1, XL1-Blue; lane 2, XL1-Blue harboring
pBluescript; lane 3, XL1-Blue harboring pBluescript withIPTG as inducer and lane 4, XL1-Blue harboring pTCB01 withIPTG as inducer.
67

=
15 16 17 18 19 20 21 22 23 24 25 26 27 28
--
30 -
kD
94 67 -
43 -
20 -
14 -
Figure 8. Calmodulin gel overlay analysis of extracts from25 independent recombinants.
Equal proteins extracted from cultures of E.coli XL1Blue harboring different recombinant phagemids without(lanes 1 and 2) or with (remaining lanes) IPTG as inducerwere separated on 12.5% 50S-polyacrylamide gels and geloverlay analysis was done in the presence of 0.1 roM CaCl2 asdescribed in Materials and Methods.
Lane 1, No phagemid; lanes 2 and 3, pBluescript;Remaining lanes from lane 4 to 28, pTCB01, 03, 04, 05, 06,10, 14, 15, 18, 22, 28, 23, 25, 28, 31, 38, 40, 46, 48, 51,55, 56, 58, 59 and 60 in order.
When gel overlay analysis was done in the presence of 5roM EDTA, no CaM-binding activity was detected (Data notshown) .
68

stable in E.coli (Fig. 8); however, partially degraded
peptides still retained the ability to bind CaM. Several
clones such as pTCB 25, 31, 38 and 48 showed the same
patterns for the DNA restriction maps and gel overlay
analysis (Fig. 6, lanes 13, 16, 17, 20 and Fig. 8, lanes 16,
18, 19, 22). These results suggest that the CaM-binding
domain is functional even when they do not reside in an
intact native protein. Similar results have been
demonstrated by using short synthetic peptides in the
studies of CaM-binding domains (203). This feature provides
the rationale for the characterization of CaM-binding
domains using expression vector and DNA deletion analysis.
The data in Figure 7 also indicated that E.coli contained
two minor peptides which bound to CaM. However, these did
not interfere with the screening procedure used here as
positive clones produced fusion proteins with strong ~S-CaM
signals. While E.coli lacks CaM (55, 259), there is protein
with EF hand Ca2+-binding domains present in this organism
(273). Since the EF hand structure is highly conserved,
this finding may be the result of CaM-binding to protein
targets for the E.coli EF hand protein.
B. Characterization of Calmodulin-Binding Protein Clone
pTCB40
Of 25 clones obtained by the screening procedure, the
mRNA levels of 20 clones were unaffected by heat shock
69

treatment. Clone pTCB40 was selected as representative of
this class of the clones (Fig. 9) and sUbjected to further
analysis.
A polypeptide (TCB40) generated from pTCB40 was
detected by "S-CaM gel overlay analysis (Fig 12, lane A4).
A 4.3 kd N-terminal fragment of ~-galactosidase should fuse
with the peptide encoded by the insert provided that the
translation of TCB40 begins from ~-galactosidase initiation
codon. The size (27 Kd) of TBC40 suggested that the cDNA
insert contained an open reading frame (ORF) sufficient to
encode a sequence of approximately 210 amino acids.
To gain more precise information on the ORF and
possible clues as to the identity of the cloned TCB40, the
sequence of the cDNA insert of pTCB40 was determined (Fig.
10). The 928 bp insert contains an ORF encoding 233 amino
acids sufficient to code for a peptide with a calculated
molecular weight of about 27 Kd. Following the stop codon
TAG at positions 814-816 is an untranslated sequence of 112
bp including an 18 bp poly(A) sequence. The
polyadenylylation signal TAATAT is located at the positions
888-893, 17 bp upstream from poly(A) sequence. Numerous
stop codons are present in the 114 bp 5' untranslated leader
70

2.5 Kb
C fiS
I·I
r!
Figure 9. Northern blot analysis of pTCB40.Equal amounts (20 ~g/lane) of total RNAs isolated from
control (C) and heat shocked (HS) tobacco cells wereseparated on 1.5% formaldehyde agarose gel and transferredby blotting to nitrocellulose. The mRNA was probed with 32p_
labeled DNA made from pTCB40 by random primer extension.
71

1 agagagagagagagagaactagtctcgagttttttttaaattcgaactaatcgtattaat
61 ctgctaaatagtagtattcgattttgataaatatatgaatatttgaaggttccaatgtttM F
121 gaaaaaatggatgagcaactcctggatgcactatgtgaccgtctcaggccagtcctctacE K MOE Q L LOA LCD R L R P V L Y
181 acggagaacagcttcattgtgcgtgaaggtgatccagttgatgagatgctttttataatgTEN S F I V REG D P V 0 E M L F I M
241 cggggcaaactattgaccgtaaccactaatggagggagaacaggcttttttaactctgatR G K L LTV T T N G G R T G F F N S D
301 tatctaaaagctggtgatttttgtggagaagagcttcttacttgggctctggatcctcatY L K A G D F C GEE L L TWA L D P H
361 ctatcaaataacctcccaatttcaacccgaactgtccaagcgctctcagaagttgaagcaL S N N L PIS T R T V Q A L S EVE A
421 tttgctctagtggccgatgatttgaagtttgtagcctctcagtttcgaagacttcatagcF A L V ADD L K F V A S Q F R R L H S
481 aagcaactccgccatacttttaggttttactcgcagcaatggagaacctgggcggcctgcK Q L R H T F R F Y S Q Q W R TWA A C
541 ttcatacaagcagcatggcgccgtcattgtaggaagaagctaqaggagtctcttcgcgaaF I Q A A W R Rile R K K LEE S L R E
601 gaagaaagcagattgcaagatgcattagcgaggggaagtggtagctcgccaagtttgggtE E S R L Q D ~ L A R G S G SSP S L G
661 gctactatctatgcatcgcgatttgctgctaatgcacttcgtgctttgcggcgcaatactA T I Y A S R F A A N A L R A L R R N T
721 tcaaagaaagcaaggatggtagacagatatcacccattttgcttcagaaaccagctgaacS K K A R H V 0 R Y II P F C F R N Q L lJ
781 cagattttacggcagaagataagtaattgtatttagtatgtagtaatgcttggccttcttQ I L R Q K I S N C 1 *
8~1 gtatatgattgattacatgtttctcctgtatactagtttgtttgtagtaatatcttatat
901 ttcaatgctcaaaaaaaaaaaaaaaaaa
Figure 10.pTCB40.
DNA sequence and deduced amino acid sequence of
72

for all three reading frames, suggesting that TCB40 is not a
fusion protein translated from the ATG initiation codon of
~-galactosidase. The predicted molecular weight based on
the DNA sequence is similar to what detected by gel overlay
analysis (Fig. 12, lane A4). These two factors suggest that
translation of the cDNA insert of pTCB40 may use the ATG
initiation codon located at the positions 115-117 even
though no conserved sequence for eukaryotic translation
initiation was found bordering this ATG. The high copy
number of rnRNAs transcribed from the insert and the presence
of a Shine-Dalgarno like sequence, GAAGGT, (Shine-Dalgarno
consensus: GGAGGT) at positions 105-110, suggest efficient
synthesis of TCB40 even though the initiation codon (ATG at
the positions 114-117) is remote from terminal codon TAA
(position 59-61) for the ~-galactosidase fragment.
In order to confirm this hypothesis, the insert of
pTCB40 was sUbjected to deletion and several clones
containing different deletion constructs (Fig. 11) were used
for 35S-CaM gel overlay analysis. The first construct, D-6,
removed 62 bp from the vector resulting in an out-of-frame
mutation for the fusion protein. The second deletion
construct, D64, deleted 63 bp from putative 51 untranslated
leader region in addition to 67 bp from the vector. Both
deletions would not interrupt TCB40 provided that
translation begins from ATG (position 115-117) of the ORF of
73

·1/1
//
I pTCB407/
·68/-6
// D·67/
..68/64
// D647/
·68/115
// D1157/
·68/337
// D3377/
Vector Insert
Figure 11. Deletion constructs of pTCB40.First nucleotide from the 5' end of the insert is
referred as number 1.
74

00
r-
U)
an00
~
M
N
"""03
~
U)
It)
<t~
C¥)
N
,..
I I I I I IQ ~r- MOO "t~ aHl) "l:t M C'I "'"
Figure 12. Calmodulin gel overlay analysis of extracts fromE.coli XL1-Blue cells harboring pTCB40 or different deletionconstru.cts.
Equal proteins extracted from cultures of XL1-Bluecells harboring different phagemids with (lanes 3, 4, 5, 6and 7) or without (lanes 1 and 2) IPTG as inducer wereseparated on 12.5% 50S-polyacrylamide gels and gel overlayanalysis was done in the presence of 0.1 roM CaCl2 (A) or 5roM EOTA (B) as described in Materials and Methods.
Lane 1, No phagemidj lanes 2 and 3, pBluescriptj lane4, pTCB40j lane 5, 0-6j lane 6, 064j lane 7, 0115 and lane8, 0337.
75

pTCB40 insert. The CaMBPs generated from both deletions
were the same size, 27 Kd, as TCB40 (Fig. 12, lanes A4, A5
and A6), demonstrating that the deleted sequences were not
required for the synthesis of TCB40. The third deletion,
Dl15, in which the entire 5' leader sequence was exactly
removed, produced a 29 Kd CaMBP, 2 kd larger than TCB40
(Fig. 12, lane A7). This was predicted since this in-frame
deletion fused the ORF of pTCB40 with the 19 amino acids of
the N-terminus of ~-galactosidase. Taken together, these
results indicate that TCB40 is translated from the ORF of
the insert.
The size of mRNA for pTCB40 detected by Northern blot
was 2.5 Kb (Fig. 9). This estimated size is much larger
than expected based on DNA sequence data (Fig. 10). These
conflicting data may arise from errors in cDNA synthesis
during library construction if some sequence of the mRNA was
not copied into the cDNA due to secondary structure. It is
highly unlikely that such rare events would create CaM
binding domains for the following reasons. First, frame
shifts may introduce numerous stop codons and abort
translation. Second, the special domains such as a basic
amphiphilic a-helix are required for CaM-binding. The
probability of creating such a structure as an artifact is
extremely low.
76

gc,
Figure 13. Predicted secondary structure of TCB40 proteinwith Chou-Fasman method.
77

A fourth deletion construct 0377 lacking 86 amino acids
of the N-terminus of TCB40 retained CaM-binding activity
(Fig. 12, lane A8). This result suggests that CaM-binding
site resides in c-terminal 147 amino acids. since a basic
amphiphilic a-helix is the CaM-binding domain of several
animal CaMBPs (203), a secondary structural analysis of
TCB40 was done with the Chou-Fasman method using the
University of Wisconsin sequence analysis software package
(54). The results indicate that three regions around the
positions 100, 155 and 195 have high probabilities for
forming a-helical structure (Fig. 13). However, there is
insufficient evidence to assign the CaM-binding domain to a
specific region. Furthermore, DNA and protein sequence
database searches did not reveal any significant homologies
between pTCB40 and other reported genes or proteins.
C. Characterization of calmodulin-Binding Protein Clone
pTCB60 and Identification of calmodulin-Binding Domain.
Analysis of Northern blots demonstrated that a 2.1 Kb
mRNA was recognized by a probe made from pTCB60. This mRNA
decreased by at least 70% over a 2 hour heat shock treatment
(Fig. 14). Since no degradation of mRNA during heat shock
has been suggested (162, 200, 269) and our previous results
also supported this idea, the apparent specific degradation
of the mRNA for TCB60 may play a role in the HSR. This
78

2.1 Kb
Figure 14. Northern blot analysis of pTCB60.Equal amounts (20 ~g/lane) of total RNAs isolated from
control and heat shocked tobacco cells were separated on1.5% formaldehyde agarose gel and transferred by blotting tonitrocellulose. The mRNA was probed with 32P-Iabeled DNAmade from pTCB60 by random primer extension.
79

degradation of mRNA would presumably affect a metabolic
activity or affect distribution of free CaM.
Analysis of DNA sequence. The phagemid of pTCB60 was
purified and sequenced. A 1572 bp insert containing an 1184
bp ORF encoding 393 amino acids was found (Fig. 15).
Following the TAA stop codon (positions 1182-1184) is an
untranslated sequence of 388 bp including a poly(A) terminus
of 19 bp. A sequence, ATAAA, at the positions 1514-1518, 35
bp upstream from poly(A) is probably the polyadenylylation
signal. This recombinant phagemid produced a CaM-binding
fusion protein of 435 amino acids with a calculated
molecular weight of 48.5 Kd. This size is somewhat larger
than that detected by 35S-CaM gel overlay analysis of clone
pTCB60 (38 KD) (Fig. 17, lane A4). This may be due to
unstability of the fusion proteins in E.coli cells mentioned
earlier. In order to obtain insight as to identities of
this clone, DNA and protein sequence database searches were
done; however, no significant homology between pTCB60 and
other reported genes and proteins was revealed.
Preliminary identification and characterization of the
caM-binding domain. Recent studies on several animal and
human CaMBPs, including non-erythroid spectrin (157), smooth
muscle MLCK (169), skeletal muscle MLCK (27), the Ca2+ pump
(289), calmodulin kinase II (110) and the 1-subunit of
80

1 aagaaacattatcaccagctctaaatgacgatgtctggagattggagaagattggcaaggE T L SPA L NOD V W R L E K I G K D
61 atggttctttccacaagaggttaaataaggctgcgatatttactgttgaagacttccttaG S F H K R L N K A A 1FT V E 0 F L R
121 ggcttgtggttagagacccgcagaaactacggaatattcttggaagcggtatgtcaaataL V V R 0 P Q K L R NIL G S G M S N K
181 agatgtgggatgctctaatagagcatgcaaagacttgtgtgttgagtgggaagctttatgM W D A LIE H A K T C V L S G K L Y V
241 tctattattctgatgattcaagaaatgtgggtgttgttttcaacaatatttatgagctgaY Y SOD S R N V G V V F N N lYE L N
301 atggccttatagctggtgaacaatactattcggcagattcactttcagacagccagaaggG L I AGE Q Y Y SAD S LSD S Q K V
361 tatatgtcgattcattggtcaagaaagcgtatgataattggaatcaagttgttgaatatgY V 0 S L V K KAY D N W N Q V V E Y D
421 atggcaagtcatttttgaacattaagcaaaatcagaaagcaggctcttcgaggaatgagcG K 5 F L N I K Q N Q K A G S S R N E L
481 ttcctgttgggccagtggattaccccaacaacatggttaatcagcttccacaatcacgacP V G P V 0 Y P N N M V N Q L P Q S R L
541 ttcctgtttccgtccaatctgagcaatcttctatggatcccaacttgctaattggaggttP V S V Q SEQ SSM D P N L L I G G S
601 caggttataatgacagcatagttgctagaatgcctaaccaaagtcagatgatgaattctaG Y N D S I V ARM P N Q S Q M M N S S
661 gttcccgttcacagttcgagagcactccatttgctcctcagcagcaaataaccaacactcS R S Q F EST P F A P Q Q Q I TNT H
721 atcagctccaaagtacgagctacgacaacaatgttggcctcgctcttggtcctccgcaatQ L Q S T S Y D N N V G L A L G P P Q S
781 catcatcattccagacaatgacctcatctctcccacaaaccaatcttaatcctttcgaggS S F Q T M T S S L P Q T N L N P FED
841 actggccacacaacagggacaagggagttgacgagttcttgtctgaggaggaaattcgaaW P H N R D K G V D E F L SEE E I R M
901 tgagaagccacgaaattcttgagaatgacgatatgcaacacttgctgagactcttcagcaR SHE I LEN D D M Q H L L R L F S M
961 tgggaggcggccatggctctgtcaatgtgtctgaagacggatatggtttcccgtctttcaG G G H G S V N V SED G Y G F P S F M
1021 tgccttcaccatctcctacttttggttatgatgaggaccccaaaccttcaggaaaagctgP S P S P T F G Y D E D P K PSG K A V
1081 tcgtgggttggctgaaaattaaggctgcaatgagatggggattctttgtgcggaagaaggV G W L K I K A A M R W G F F V R K K A
1141 cagctgagaggcgggcacagattgtggaactggatgatgaataaaatctcagggctttgcA ERR A Q I VEL D D E *
1201 ggcttaaccgtcgaccatttgaagaactattcacaaggtcgtggtgatgcaggcccgagt1261 aattagctacacaggagtttatttccgagggattagtgaattcgattgttctttctcttt1321 gcgtcaaatagtttttcatgtacagaagcctgtacagtaagttcacttttaatgctctca1381 ttgttaactcaatttgtgctcactctctttatctgtttcaagatgtaatgtattagtctg1441 cagaccaaagggaattatactttcaaagaactcgtggccctcctgtgtgttgaattctgt1501 aggaacttttatgataaacaggatgctaattggcttttgcaatttgagcgttcaaaaaaa1561 aaaaaaaaaaaa
Figure 15.pTCB60.
DNA sequence and deduced amino acid sequence of
81

phosphorylase b-kinase (73), indicate that one class of CaM
binding domain is a basic amphiphilic a-helix. To determine
the caM-binding site of TCB60, several deletion mutants from
both the 5' and 3' ends of the insert of pTBC60 were
constructed (Fig. 16) and assayed for the CaM-binding
activity by gel overlay analysis. The protein generated
from mutant D684, in which N-terminal 227 amino acids
encoded by the insert were deleted, bound to CaM, suggesting
that the c-terminal 166 amino acids contained the CaM
binding domain (Fig. 17, lane A12). Similarly, the protein
produced from deletion R1489 in which 64 bp of the 3'
untranslated region were removed retained the ability to
bind to CaM (Fig. 17, lane A5). Alternatively, Rl129 in
which all of the 3' untranslated region and 17 amino acids
from the c-termius of TCB60 were deleted, including three
positively charged amino acids at the positions 377-379,
lacked CaM-binding activity (Fig. 17, lane A6). These
results indicate that the CaM-binding domain is located in
the c-terminus of the protein.
Secondary structural analysis of TCB60 protein using
Chou-Fasman method in the University of Wisconsin sequence
analysis software package indicates that the c-terminus of
TCB60 has a high probability for forming an a-helix (Fig.
18) (54). The sequence of amino acids from 362 to 380 is
depicted as the helical wheel in Figure 19. In this
82
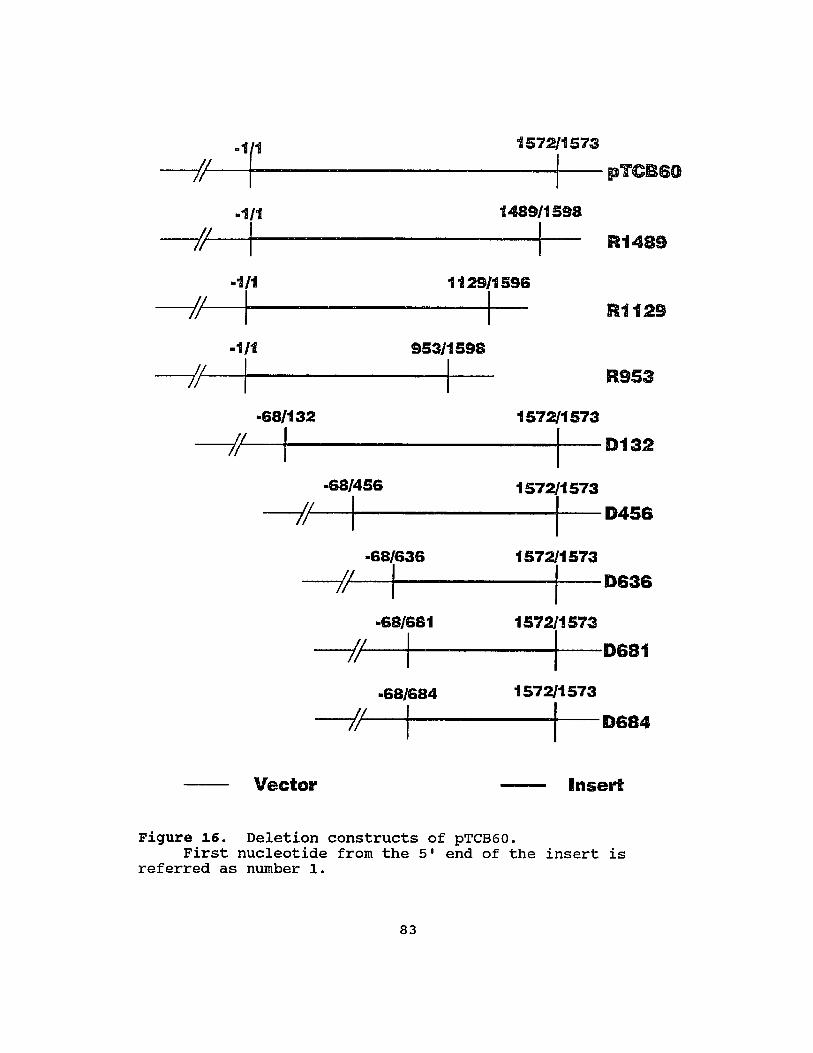
R1489
II .,1(1 1572/1573
Jr ~------------+I--pl'CB60
..1/1 1489/1598
-/I:....--1f--------I-.1/1 1129/1596
-/;~I-1-----1-.1/1
I
.68/456
-/II
·68/681
-/II·68/684
IVector
R1129
R953
Insert
Figure 16. Deletion constructs of pTCB60.First nucleotide from the 5' end of the insert is
referred as number 1.
83

1 2 3 4 5 6 7 8 9 10 11 12kD94 ...67 ...
43 -A
30 -
20 ...
14 ...
--
1 2 3 4 5 6 7 8 9 10 11 12kD94 ...67 ...
43 ...
B30 ...
20 ...
14 ...
Figure 17. Calmodulin gel overlay analysis of extracts fromE.coli XL1-Blue cells harboring pTCB60 or different deletionconstructs.
Equal proteins extracted from cultures of XL1-Bluecells harboring different phagemids without (lanes 1 and 2)or with IPTG (remaining lanes) as inducer were separated on12.5% SOS-polyacrylamide gels and gel overlay analysis wasdone in the presence of 0.1 roM CaCl2 (A) or 5 roM EOTA (B) asdescribed in Materials and Methods.
Lane 1, No phagemidi lanes 2 and 3, pBluescripti lane4, pTCB60i lane 5, R1489i lane 6, Rl129i lane 7, R953i lane8, 0132i lane 9, 0456; lane 10, 0636; lane 11, 0681 and lane12, 0684.
84

SfUitN
J'~
k.L,
Figure 18. Predicted secondary structure of TCB60 proteinwith Chou-Fasrnan method.
85

F
L
G
F
A
+K
G
A
w
M
GWlKiKAAMRWGFFVRKKA
Figure 19. Helical wheel projection for tentativecalmodulin-binding domain of TCB60 protein.
Amino acid sequence is GWLKlKAAMRWGFFVRKKA at positions362-380 of TCB60 protein. The underlined amino acidresidues are deleted from the protein generated from Rl129.
86

projection, six positively charged residues lie on one side
of the helix while the opposite side is predominantly
hydrophobic. This structure is striking in its similarity
to the basic amphiphi1ic a-helices identified as CaM-binding
domain from a variety of animal and human CaMBPs (169, 203,
271). The helical wheel of the CaM-binding domain from
chicken gizzard myosin light chain kinase (MLCK) depicted by
Lukas et al (169) is presented here for comparison (Fig. 20)
(169).
Both hydrophobic and electrostatic interactions between
CaM and the CaM-binding domain have been identified by using
the replacement of certain amino acids in CaM-binding domain
(203). The deletion of the sequence RKK (positions 377-379)
in pTCB60 deletion mutant Rl129 would be predicted to reduce
the electrostatic interaction which may result in the loss
of CaM-binding activity. Indeed the Rl129 mutant protein
failed to bind to CaM (Fig. 17, lane A6). The presence of
tryptophan residues (positions 363 and 372) is also a
feature of the known CaM-binding domains. These residues
appear to be essential for high affinity CaM-binding as
evidenced by amino acid replacement experiments (289). The
bulky tryptophany1 residue hinders the formation of a stable
secondary structure. This provides flexibility of the CaM
binding domain which may be required for optimal packing and
high-affinity binding (289). The results presented here
87

ARRKWQKTGHAVRAIGRL
Figure 20. Helical wheel projection for calmodulin-bindingdomain from chicken myosin light chain kinase (ref. 169).
88

demonstrated that TCB60 contains a basic amphiphilic a-helix
CaM-binding domain as described in animal CaMBPs. This is
the first such evidence for this structure in plant CaMBPs.
D. Characterization of a cDNA Clone Encoding a Heat Shock
Induced calmodulin-Binding Protein.
Clone pTCB48 was further confirmed to code for a CaM
binding protein by gel overlay analysis. Total proteins
isolated from the culture of E.coli XLI-Blue containing this
phagemid were used for gel overlay analysis under more
stringent conditions. Several peptides with molecular
weights of 55, 38, 34 and 24 Kd generated from pTCB48 bound
to CaM in the presence of 0.2 M NaCI and 0.1 roM CaCl2 (Fig.
22, lane A4). caM-binding activity was not detected in the
presence of EDTA (Fig. 22, lane B4), suggesting that the
interaction of CaM with these peptides is Ca2+-dependent.
These CaM-binding peptides were probably expressed from one
ORF of the insert of pTCB48 and resulted due to instability
of fusion proteins in E.coli cells. This was evidenced by
deletion and sequencing analysis (see below).
Analysis of gene expression. Equal RNAs from control
and heat shock treatments were e~ectrophoresed on
formaldehyde agarose gels and probed with a 32P-Iabeled DNA
probe made from pTCB48. Two mRNA bands with molecular
89

-11i 11574/11575
-1M 1 pTCIB48
-77/180 1574/1575
-If-I I D180
-68/747 1574rS75-1M 1>747
-76/835 1574/1575
-If-I I D835
-86/867 1574/1575
-1M I D867
-76/982 1574/1575
-1M I D982
-1/1 1516/1599
-If-I I R1516
-I~~11480/1597
I R1480
-1/1 1184/1600
-1M I R1184
-I)-~r11611600
R1162
-1/1 1087/1600
-1M I R1087
Vector Insert
Figure 21. Deletion constructs of pTCB48.First nucleotide from the 5' end of the insert is
referred as number 1.
90

1 2 3 4 5 6 7 8 9 10 11 12 13 14kD
94 -67 -
A 43 -
30 -
20 -
14 -
----
1 2 3 4 5 6 7 8 9 10 11 12 13 14kD
94 -67 -
B 43 -
30 -
20 -
14 -
Figure 22. Calmodulin gel overlay analysis of extracts fromE.coli XL1-Blue cells harboring pTCB48 or different deletionconstructs.
Equal amounts of proteins extracted from cultures ofXL1-Blue cells harboring different phagemids without (lanes1 and 2) or with IPTG (remaining lanes) as inducer wereseparated on 12.5% 50s-polyacrylamide gels and gel overlayanalysis was done in the presence of 0.1 roM CaC12 (A) or 5roM EOTA (B) as described in Materials and Methods.
Lane 1, No phagemid; lanes 2 and 3, pBluescript; lane4, pTCB48; lane 5, 0180; lane 6, 0747; lane 7, 0835; lane 8,0867; lane 9, 0982; lane 10, R1516; lane 11, R1480; lane 12,Rl184; lane 13, Rl162 and lane 14, R1087.
91

weights of 1.9 and 2.5 Kb were detected in the RNA sample
isolated from heat-shocked cells but not in control extracts
(Fig. 23, lanes 1 and 2). This indicates that these two RNA
species were induced by heat shock. In addition, it is
likely that both RNAs were active in the translation during
heat shock since both were present in heat-shocked polysomal
RNA fraction (Fig. 23, lanes 4 and 5). A time course
analysis of expression indicated that these two mRNAs were
barely detected after a 15 minute heat shock treatment at
38°C (Fig. 24). These RNAs accumulated to maximum amounts
after 1.5 hours of heat shock (Fig. 24).
These two RNAs may result from processing of the
transcript of a single gene. Alternatively, they may result
from two different heat shock-induced genes of very high
homology. If the latter case is true, we suggest that 1.9
kb RNA is transcribed from pTCB48 gene for the following
reasons. First, the much stronger hybridization signal for
the 1.9 Kb RNA on Northern blots suggests a high abundance
of this RNA in the polysomal RNA population as compared with
the 2.5 Kb RNA. The latter band was just above the level of
detection under our conditions (Fig. 23, lanes 4 and 5).
Secondly, our eDNA library was constructed with the poly(A)
RNAs purified from polysomal RNAs. Finally, three
(pTCB25, 31 and 38) of 25 putative eaMBP clones obtained
92

1 2 3 4 5
I 2.5 Kb1.9 Kb
Figure 23. Northern blot analysis of pTCB48.Equal amounts (20 ~g/lane) of total (lanes 1 and 2) and
polysomal (lanes 3, 4 and 5) RNAs isolated from control(lanes 1 and 3) and heat shocked (lane 2, 4 and 5) tobaccocells were separated on 1.5% formaldehyde agarose gel andtransferred by blotting to nitrocellulose. The RNAs wereprobed with 32P-Iabeled DNA made from pTCB48 by random primerextension. Note: Lane 5 was the same as lane 4 but exposedfor longer time.
93

o 15 30 60 90 120
,...•.,(Minutes)
2.5 Kb
1.9 Kb
Figure 24. Time course of expression of pTCB48 mRNA fordifferent lengths of heat treatment.
Equal amounts (20 ~g/lane) of total RNAs isolated fromtobacco cells which have been treated at 38°C for differentlengths were separated on 1.5% formaldehyde agarose gel andtransferred by blotting to nitrocellulose. The RNAs wereprobed with 32P-Iabeled DNA made from pTCB48 by random primerextension.
94

from screened recombinant phages are identical to pTCB48,
further indicating the abundance of this RNA.
Analysis of DNA sequence. The sequence of pTCB48 was
determined (Fig. 25). A 1574 bp insert contains an 1307 bp
ORF which encodes for 434 amino acids (Fig. 25). Following
the TAA stop codon (position 1305-1307) is an untranslated
sequence of 267 bp including a poly(A) terminus of 22 bp. A
polyadenylation signal is the sequence, AATTATTTT, located
at the positions 1511-1519, 33 bp upstream from poly(A)
sequence. The predicted size of the fusion protein produced
from clone pTCB48 based on the sequence data is 53.5 Kd
which is in agreement with the size of the CaMBP (55 Kd)
identified by overlay analysis (Fig. 22, lane A4). No
significant homology between pTCB48 and other reported genes
from DNA and protein database was found.
Tentative identification and features of the CaM
binding domain. Recent studies on animal and human CaMBPs,
including non-erythroid spectrin (157), smooth muscle MLCK
(169), skeletal muscle MLCK (27), the Ca2+ pump (123, 289),
calmodulin kinase II (110) and the ~-subunit of
phosphorylase b-kinase (73), indicate that one type of CaM
binding domain is a basic amphiphilic a-helix (Fig. 20). To
determine the CaM-binding site of TCB48, several deletion
mutants from both the 5' and the 3' ends of pTCB48 insert
95

1 cagttgcttgtggagctgagtgtgtgttggttttaggttgtctccgttgggcatggaaacV A eGA E C V L V L GeL R W A W K R
61 gatgcacatacaccggcaatgacgacagtgctacgtggccaacagctacctatgaggaatC T Y T G N DDS A T W PTA T Y E E F
121 tcgaaccagttccacgtatctgccgtacaattctagcagtatacgaaccaaatctccgtaE P V P RIC R TIL A V YEP N L R S
181 gccccaagtacccgccgaaggtcaagactaaaccccgattggtcattaaacgggtcacatP K Y P P K V K T K P R L V I K R V T Y
241 atgaacaaacatctggcaatgcacctccgtatttgatcgctattcgtgggttgaatttacE Q T S G NAP P Y L I A I R G L N L L
301 taaacgaaagtgattacaaagttttgttggataataggctgggaaaacagatgtttgatgN E 5 D Y K V L LON R L G K Q M F D G
361 gaggatatgtacatcatgggttattgaaatctgcggt"ttgggttttgaataatgagtctgG Y V H H G L L K S A V W V L NNE S E
421 agactttgaagaagctttqgattgagaatgggaggagttacaagatgatatttgcaggacT L K K L W lEN G R S Y K M I FAG H
481 attctttgggttctggtgtggcgtctttgctgacagtgattgtggcgaatcataaggataS L G S G V A S L LTV I v A N H K 0 R
541 gattagggggaattccaaggagtcttttaaggtgttatgcagttgcaccagcgcggtgtaL G G I P R S L L R C Y A V A PAR C M
601 tgtcactcaacttggctgttaagtatgctgatataatacactctgtggtattgcaggatgS L N L A V K Y A D I I H S V V L Q 0 D
661 atttcttgccaagaacagccaccccacttgaagatatatttaaatccatcttctgtttacF L P R TAT P LED I F K S I F C L P
721 cgtgcttgatatttttggtatgcttgagagataccttcattcctgagggcagaaaactcceLI F L veL R D T F I PEG R K L R
781 gagatccaagaagactttatgcaccaggccgtatgtatcacattgtagaaagaagattctD P R R L YAP G R M Y H I V ERR F C
841 gcagatgtgggaggttcactccagatgttagaactgccatcccagttgacggaagatttgReG R F T P 0 V R T A I P V D G R F E
901 agcatatcgtgttgtcacgcaatgctacagttgatcatggaatcatttggatagaaagagH I V L S RNA T V D H G I I W I ERE
961 aatctgaaaaagtattagcaagacttaaggaggctagtgctgagaccacaactactccccS E K V L A R L K E A S A E T T T T P P
1021 ccaaagtgcagaaaattgaaaggctgaagacattagaaaaagaacacaaggatgcactcgK V Q K I E R L K T L EKE H K 0 ALE
1081 aaagagctgtcagtttaaacataccacacgctgtggac0cag~tgaagaggaatccacgg
R A V S L NIP 1I A V 0 A DEE EST E1141 agagtataactgaggagtcatctcagaaacaggaggaagatgcaatgacaagcaaagcgc
SIT E E S S Q K Q E E DAM T S K A Q1201 agtgcagtgatgcaagaactaactggaatgaagtagttgagaagcttttcaatagagacg
C S 0 ART N W N E V V E K L F N R D E1261 aaagtgggaaattacgattaaagagagatgcaactggtcccgagtaacaactctgttcca
S G K L R L K R D A T G P E *1321 aggtgctttttctctcttcttttttatgatggctgtgtatgttagatgaaagcattatta1381 ctcgtgttccaacaactcataattctttattatccatatcgcatagtacaatatcttctt1441 tttgtatatcaacgaccataatgtataatctgaagccccgtcaatagtacagttggaaat1501 tcttttatgcaattattttaggcagttcctcgtggaatgtgatttgttcatgaaaaaaaa1561 aaaaaaaaaaaaaa
Figure 25.pTCB48.
DNA sequence and deduced amino acid sequence of
96

were constructed (Fig. 21). The CaM-binding activity of the
proteins from these various deletions was assayed by gel
overlay analysis with 35S-labeled CaM. The proteins
generated from all 5' deletion constructs, including D982
which lacked 327 amino acids from the N-terminus encoded by
pTCB48 insert, retained the ability to bind to CaM (Fig. 22,
A5, A6, A7, A8 and A9). These results localized the CaM
binding domain to a sequence of 107 C-terminal amino acids
(from 328-434) of TCB48 (Fig. 25). The fact that D982
synthesized only one peptide (16 Kd) that bound to CaM
indicated that the ORF of pTCB48 was responsible for all
four CaM-binding peptides produced from clone pTCB48 (Fig.
22, A9). Analysis of several 3' deletion constructs
further define the binding domain. The protein produced
from deletion construct Rl184, in which 40 c-terminal amino
acids were removed, lacked CaM-binding activity (Fig. 22,
lane A12). Thus, the CaM-binding domain resides either in
the c-terminal 40 amino acids or overlaps the deletion
point.
A secondary structure analysis of TCB48 protein was
done with Chou-Fasman method in the University of Wisconsin
sequence analysis software package (54). This analysis
suggests that the region from amino acid residue 400 to 434
has a high probability of forming p-sheets and p-turns (Fig.
26). The region from amino acid residue 330 to 400 has a
97

~.
o.:>x
Figure 26. Predicted secondary structure of TCB48 proteinwith Chou-Fasman method.
98

propensity for forming a-helices. However, neither region
apparently contains the classic caM-binding basic
amphiphilic a-helix domain. Even though a-helices are
predicted in the region from amino acid 330 to 400, a total
of 18 negatively charged, acidic residues is dispersed
throughout the sequence. This would prevent the formation
of a strict basic amphiphilic a-helix. Recent experiments
using short synthetic peptides of 25 amino acids based on
the 1-subunit of phosphorylase kinase indicate an
alternative CaM-binding domain structure (73). A peptide of
25 amino acids which binds to CaM forms a ~-turn followed by
~-strand structure (73). The data presented in this study
suggest that some CaM-binding domain other than a basic
amphiphilic a-helix is also present in plant CaMBPs.
99

CHAPTER V
CONCLUSION
The study presented here provides first indication of
the synthesis of CaMBPs as HSPs during plant HSR. Beside
HSP 70, the synthesis of several other CaMBPs with molecular
weights ranging from 17 to 82 Kd was enhanced/induced by
heat shock. These results suggest a role of CaM-mediated
processes in plant HSR.
A procedure for the isolation of CaMBP clones by
screening a cDNA expression library with radioactively
labeled CaM as a ligand probe was highly refined. Twenty
five independent tobacco cDNA clones were isolated using
this screening method and confirmed to encode CaMBPs by gel
overlay analyses. The expression of all these cloned mRNAs
was analyzed with Northern blots. These clones can be
divided into three classes based on the differential
expression during heat shock. The expression of pTCB48 mRNA
was induced while the level of pTCB60 mRNA was reduced by
heat shock. However, the levels of most cloned mRNAs as
represented by pTCB40 were unaffected by heat shock. These
three clones were sequenced and database searches of DNA and
protein sequences did not reveal any significant homology
between these clones and other reported genes or proteins.
100

Thus, these cDNAs apparently represent new sequences
encoding CaMBPs of unknown functions. Based on the present
study and data from literature, a model for the possible
functions of CaMBPs in the HSR is suggested in Figure 27.
This study is the first report for the isolation and
characterization of cDNA clones for CaMBPs in plants.
Furthermore, this research provides the first evidence for
the presence of a basic amphiphilic a-helix as CaM-binding
domain in a plant CaMBP. An alternative structure other
than a basic amphiphilic a-helix is also suggested to be a
caM-binding domain in a plant CaMBP. This structure may be
a ~-sheet following a ~-turn.
Future efforts should focus on the functional
identification of these cloned CaMBPs. The sequence
analyses of the whole coding regions and promoters may
provide insight into the regulation of these proteins during
heat sheck. Finally, further characterization and
identification of the nature of the CaM-binding domains as
well as the interactions with CaM will provide a better
understanding of the mechanisms by which CaM regulates a
variety of protein targets.
101
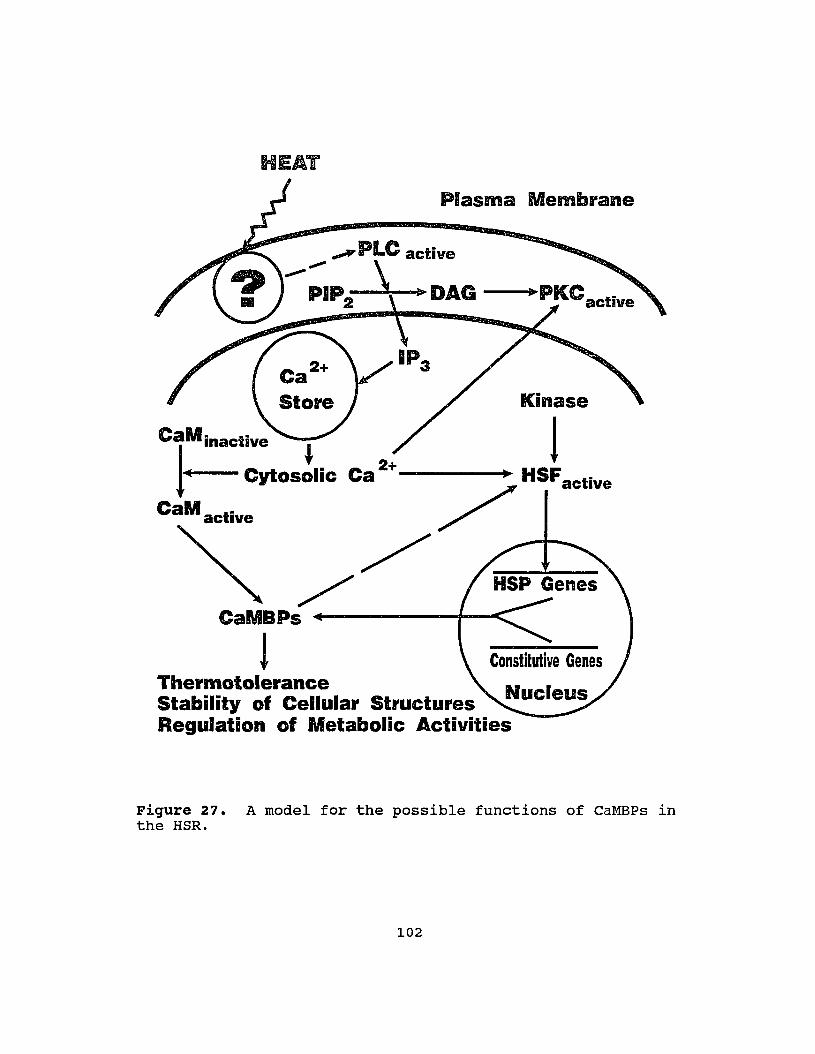
HEAT
1Kinase
Plasma Membrane
",A'PLC active,,~ \
PIP J) DAG ---...PKC2 active
Ca2+- - - - -. HSF
• active-:.>
/CaMBPs
!Thermotolerance N IStability of Cellular Structures uc eusRegulation of Metabolic Activities
CaM inactive
1· cytoS!IiC
CaM active
Figure 27. A model for the possible functions of CaMBPs inthe HSR.
102

REFERENCES
1. Abravaya K, Phillip B, Morimoto RI (1991) Heat shockinduced interactions of heat shock transcription factorand human hsp70 promotor examined by in vivofootprinting. Mol Cell BioI 11:586-592
2. Allan EF, Trewavas AJ (1987) The role of calcium inmetabolic control. IN: Davies DD (eds) TheBiochemistry of plants: A comprehensive treatise. Vol.12. Physiology and metabolism. Acadmic Press, Inc. NewYork. ppl17-149
3. Amin J, Mestril R, Lawson R, Klapper H, Dreano M,Voellmy R (1985) The heat shock consensus sequence isnot sufficient for hsp70 gene expression in Drosophilamelanogaster. Mol Cell BioI 5:197-203
4. Amin J, Mestril R, Schiller P, Dreano M, Voellmy R(1987) organization of the Drosophila melanogasterhsp70 heat-shock regulation unit. Mol Cell BioI 7:10551602
5. Arrigo AP, Fakan S, Tissieres A (1980) Localization ofthe heat shock induced proteins in Drosophilamelanogaster tissue culture cells. Dev BioI 78:86-103
6. Arrigo AP, Ahmad-Zadeh C (1981) Immunofluorescencelocalization of small heat shock protein (hsp23) insalivary gland cells of Drosophila melanogaster. MolGen Genet 184:73-79
7. Arrigo AP (1987) Cellular localization of HSP23 duringDrosophila development and following subsequent heatshock. Dev BioI 122:39-48
8. Arrigo AP, Welch WJ (1987) Characterization andpurification of the small 28,000 dalton mammalian heatshock protein. J. BioI Chern 262:15359-15369
9. Ashburner M, Bonner JJ (1979) The induction of geneactivity in Drosophila by heat shock. Cell 17:241-254
10. Asselin J, Phaneuf S, Watterson DM, Haiech J (1989)Metabolically "S-labeled recombinant calmodulin as aligand for the detection of calmodUlin-bindingproteins. Anal Biochem 178:141-147
103

11. Aviv H, Leder P (1972) Purification of biologicalactive globin messenger RNA by chromatography onoligothymidylic acid-cellulose. Proc Natl Acad Sci USA69:1408-1412
12. Bardwell JCA, Craig EA (1984) Major heat shock gene ofDrosophila and Escherichia coli heat-inducible dnaKgene are homologous. Proc. Natl. Acad. Sci. USA 81:848852
13. Bardwell JCA, Craig WA (1987) Eukaryotic Mr 83,000 heatshock protein has a homologue in Escherichia coli.Proc. Natl Acad Sci USA 84:5177-5188
14. Barnett MJ, Long SR (1990) Nucleotide sequence of analfalfa calmodulin eDNA. Nucleic Acids Res 18:3395-3395
15. Baszczynski CL, Walden DB, Atkinson BG (1985) Maizegenome responses to thermal shifts. IN: Atkinson BG,Walden DB, ed, Changes in eukaryotic gene expression inresponse to environmental stress. Academic Press Inc.,New York pp349-371
16. Baulieu EE, Catelli MG (1989) steroid hormone receptorand heat shock protein Mr 90,000 (HSP90): A functionalinteraction. IN Pardue ML, Feramisco JR, Lindquist S,ed, Stress-induced proteins. Alan R. Liss, Inc., NewYork pp203-219
17. Beckmann RP, Mizzen LA, Welch WJ (1990) Interaction ofHsp70 with newly synthesized protein: Implication forprotein folding and assembly. Science 248:850-854
18. Berger EM, Woodward MP (1983) Small heat shock proteinsin Drosophila may confer thermal tolerance. Exp CellRes 147:437-442
19. Bergh S, Arking R (1984) Developmental profile of theheat shock response in early embryo of Drosophila. JExp Zool 231:379-391
20. Berendes HD (1968) Factors involved in the expressionof gene activity in polytene chromosomes. Chromosoma24:418-437
21. Berridge MJ (1987) Inositol lipids and cellproliferaticn. Biochim Biophys Acta 907:33-45
22. Bienz M, Gurdon JB (1982) The heat shock response inXenopus oocytes is controlled at the translationallevel. Cell 29:811-819
104

23. Bienz M (1985) Transient and developmental activationof heat-shock genes. Trends Biochem Sci 10:157-161
24. Bienz M, Pelham HRB (1987) Mechanisms of heat-shockgene activation in higher eukaryotes. Adv Genet 24:3172
25. Biessmann H, Falkner FG, Saumweber H, Walter MF (1982)Disruption of vimentin cytoskeleton may play a role inheat-shock response. IN Schlesinger MJ, Ashburner M,Tissieres A, ed, Heat shock from bacteria to man. ColdSpring Harbor, New York pp275-281
26. Billigsley ML, Pennypacjer KR, Hoover CG, Brigati DJ,Kincaid RL (1985) A rapid and sensitive method fordetection of calcineurin and calmodulin-bindingproteins using biotinylated calmodulin. Proc Natl AcadSci USA 82:7585-7589
27. Blumenthal DK, Takio K, Edelman AM, Charbonneau H,Titani K, Walsh KA, Krebs EG (1985) Identification ofthe calmodulin-binding domain of skeletal muscle myosinlight chain kinase. Proc Natl Acad Sci USA 82:31873191.
28. Bommer UA, Noll F, Lutsch G, Bielka H (1980)Immunochemical detection of proteins in the smallsubunit of rat liver ribosomes involved in binding ofthe ternary initiation complex. FEBS Lett 111:171-174
29. Bonanou-Tzedaki S, Sohl MK, Arnstein HRV (1981)Regulation of protein synthesis in reticulocyte lysate.Characterization of the inhibitor generated in thepost-ribosomal supernant by heating at 44°C. Eur JBiochem 114:69-77
30. Bond U, Schlesinger MJ (1985) Ubiquitin is a heat shockprotein in chicken embryo fibrolasts. Mol Cell Biol5:949-956
31. Bond U, Schlesinger MJ (1986) The chicken ubiquitingene contains a heat shock promotor and expresses anunstable mRNA in heat-shocked cells. Mol Cell BioI6:4602-4610
32. Bond U, Schlesinger MJ (1987) Heat-shock protein anddevelopment. Adv Genet 24:1-29
33. Bonham-Smith P, Kapoor M, Bewley YD (1987)Establishment of thermotolerance in maize by exposureto stresses other then heat shock does not require heatshock protein synthesis. Plant Physiol 85:575-580
105

34. Bonner JJ, Parks C, Parker-Thornburg J, Mortin MA,Pelham HRB (1984) The use of promoter fusions inDrosophila genetics: isolation of mutants affecting theheat shock response. Cell 37:979-991
35. Bossier P, Fitch IT, Boucherie H, Tuite MF (1989)structure and expression of a yeast gene encoding thesmall heat-shock protein hsp26. Gene 78:323-330
36. Brown EG, Newton RP (1981) Cyclic AMP and higherplants. Phytochem 20:2453-2463
37. Brugge JS, Erikson E, Erikson RL (1981) The specificinteraction of Rous sarcoma virus transforming protein,pp60src, with two cellular proteins. Cell 25:362-372
38. Brugge JS, Yuonemoto W, Darrow D (1983) Interactionbetween the Rous sarcoma virus transforming protein andtwo cellular phosphoproteins: analysis of the turnoverand distribution of this complex. Mol Cell BioI 4:26972704
39. Burdon RH, Cutmore CMM (1982) Human heat shock genemodulation of plasma membrane Na+, K+-ATPase activity.FEBS Lett 140:45-48
40. Burgess WH, Watterson DM, Van Eldik LJ (1984)Identification of calmodulin-binding proteins inchicken embryo fibroblasts. J Cell BioI 99:550-557
41. Burke JF, Ish-Hrowicz D (1982) Expression of Drosophilaheat shock genes is regulated in Rat-1 cells. NucleicAcids Res 10:3821-3830
42. Burke JJ, Hatfield JL, Klein RR, Mullet JE (1983)Accumulation of heat shock proteins in field-growncotton. Plant Physiol 78:394-398
43. Burke JJ, Orzech KA (1988) The heat-shock response inhigher plants: a biochemical model. Plant Cell andEnvri 11:441-444
44. Cadepond F, schweizer-Groyer G, Segard-Maurel I, JibardN, Hollenberg SM, Giguere V, Evans RM, Baulieu EE(1991) Heat shock protein 90 as a critical factor inmaintaining glucocorticostroid receptor in anonfunctional state. J BioI Chern 266:5834-5841
45. Caizergues M, Bouche G, Amalric F, Zalta JP (1980a)Effects of heat shock on nuclear and nucleolar protein
106

phosphorylation in chinese hamster ovary cells. Eur JBiochem 108:399-404
46. Caizergues M, Bouche G, Amalric F, Zalta JP (1980b)Phosphorylated proteins involved in the regulation ofmRNA synthesis in CHO calls recovering from heat shock.FEBS Lett 116:261-264
47. Calderwood SK, stevenson MA, Hahn GM (1987) Heat stressstimulates inositol triphosphate release andphosphorylation of phosphoinositides in CHO and BalbC3T3 cells. J Cell Physiol 130:360-376
48. Calderwood SK, stevenson MA, Hahn GM (1988) Effects ofheat on cell calcium and inositol lipid metabolism. RadRes 113:412-425
49. cartwright IL, Elgin SCR (1986) Nucleosomal instabilityand induction of new upstream protein-DNA associationsaccompany activation of four small heat-shock proteingenes in Drosophila melanogaster. Mol Cell BioI 6:779791
50. Catelli MG, Binart N, Jung-Testas I, Renoir JM, BaulieuEE, Feramisco JR, Welch WJ (1985) The common 90-Kdprotein component of non-transformed "85" steroidreceptors is a heat-shock protein. EMBO J 4:3131-3135
51. Catelli MG, Ramachandran C, Gauthier Y, Legagneux V,Quelard C, Baulieu EE, Shyamala G (1989) Developmentalregulation of murine mammary-gland 90 kDa heat shockproteins. Biochem J 258:895-901
52. Chappell TG, Welch WJ, Schlossmann DM, Palter KB,Schlesinger MJ, Rothman JE (1986) Uncoating ATPase is amember of the 70 kilodalton family of stress proteins.Cell 45:3-13
53. Chirico WJ, Waters MG, Blobel G (1988) 70K heat shockrelated proteins stimulated protein translocation intomicrosomes. Nature 332:805-810
54. Chou PY, Fasman GD (1978) Predication of the secondarystructure of proteins from the amino acid sequence. AdvEnzymol Relat Areas Mol BioI 47:45-148
55. Clark M, Bazari WL, Kayman SC (1980) Isolation andproperties of calmodulin from Dictysotelium discoideum.J Bacteriol 141:397-400
56. Clark BD, Brown IR (1986) A retinal heat shock proteinis associated with elements of the cytoskeleton and
107

bind to calmodulin. Biochem Biophys Res Cornrnun139:974-981
57. Clarke A, critchley C (1990) Synthesis of early heatshock proteins in young leaves of barley and sorghum.Plant Physiol 94:567-576
58. Colbert RA, Hucul JA, Scorsone KA, Young DA (1987) asubunit of eukaryotic translational initiation factor-2is a heat-shock protein. J BioI Chem 262:16763-16766
59. Collier NC, Heuser J, Levy MA, Schlesinger MJ (1988)Ultrastructural and biochemical analysis of the stressgranula in chicken embryo fibroblasts. J Cell BioI106:1131-1139
60. Conner TW, LaFayette PR, Nagao RT, Key JL (1990)Sequence and expression of a HSP83 from Arabidopsisthaliana. Plant physiol 94:1689-1695
61. Cooper P, Ho THD (1983) Heat shock proteins in maize.Plant Physiol 71:215-222
62. Cooper P, Ho THD, Hauptmann RM (1984) Tissuespecificity of the heat shock response in maize. PlantPhysiol 71:431-441
63. Corces V, Pellicer A, Axel R, Meselson M (1981)Integration, transcription and control of a Drosophilaheat shock gene in mouse cells. Proc Natl Acad Sci USA79:7038-7042
64. Costlow N, Lis JT (1984) High-resolution mapping ofDNase I-hypersensitive sites of Drosophila heat-shockgenes in Drosophila melnogater and Saccharomycescerevisiae. Mol Cell BioI 4:1853-1863
65. Courtneidge SA, Bishop JM (1982) Transit of pp60V-SfC to
the plasma membrane. Proc Natl Acad Sci USA 79:71177121
66. Craig EA (1985) The heat shock response. CRC criticalRev Biochem 18:239-280
67. craig EA, Gross CA (1991) Is hsp70 the cellularthermometer? Trends Biochem. Sci. 16:135-140
68. Csermely P, Kahn CR (1991) The 90-kDa heat shockprotein (hsp-90) possesses an ATP binding site andautophosphorylating activity. J BioI Chern 288:4943-4950
108

69. Czarnecka E, Edelmen L, Schoffl F, Key JL (1984)Comparative analysis of physical stress responses insoybean seedlings using cloned heat shock cDNAs. PlantMol BioI 3:45-58
70. Czarnecka E, Gurley WB, Nagao RT, Mosquera LA, Key JL(1985) DNA sequence and trnscript mapping of soybeangene encoding a small heat shock protein. Proc NatlAcad sci USA 82:3726-3730
71. Czarnecka E, Key JL, Gurley WB (1989) Regulationdomains of the Gmhsp17.5-E heat shock promotor ofsoybean. Mol Cell BioI 9:3457-3463
72. Dale RMK, Mcclure BA, Houchins JP (1985) A rapidsingle-stranded cloning strategy for producing asequential series of overlapping clones for use in DNAsequence: Application to sequence the cornmitochondrial 18S rDNA. Plasmid 13:31-41
73. Dasgupta M, Honeycutt T, Blumenthal DK (1989) The ~
subunit of skeletal muscle phosphorylase kinasecontains two moncontiguous domains that act in concertto bind calmodulin. J BioI Chern 264:17156-17163
74 Davis R, King ML (1989) The developmental expression ofthe heat-shock response in Xenopus laevis. Development105:213-222
75. Davis TN, Urdea MS, Masiarz FR, Thorner J (1986)Isolation of the yeast calmodulin gene: calmodulin isan essentail protein. Cell 47:423-431
76. Davis TN, Thorner J (1989) Vertebrate and yeastcalmodulin, despite significant sequence divergence,are functionally interchangeable. Proc Natl Acad SciUSA 86:7909-7913
77. Dawson we, Boyd C (1987) TMV protein synthesis is nottranslational regulated by heat shock. Plant Mol BioI8:145-149
78. DeBenedetti A, Baglioni C (1986) Translationalregulation of the synthesis of a major heat shockprotein in Hela cells. J BioI Chern 261:338-342
79. DeJong WW, Liunissen JAM, Leenen PJM, Zweers A,Versteeg M (1988) Dogfish a-crystallin sequences.Comparison with small heat shock protein andSchistosoma egg antigen. J BioI Chern 263:5141-5149
109

80. Deshaies RJ, Koch BA, Werner-Washburne M, Craig EA,Schekman R (1988) A sUbfamily of stress proteinsfacilitates translocation of secretory andmitochondrial precursor polypeptides. Nature 332:800805
81. DiDomenico BJ, Bugaisky GE, Lindquist S (1982a) Theheat-shock self-regulated at both the transcriptionaland posttranslational levels. Cell 31:593-603
82. DiDomenico BJ, Bugaisky GE, Lindquist S (1982b) Heatshock and recovery are mediated by differenttranslational mechanism. Froc Natl Acad Sci USA79:6181-6185
83. Dietz TJ, Cartwright IL, Gilmour DS, Siegfried E,Thomas EH, Elgin SCR (1989) The chromatin structure ofhsp26. IN: Parde ML, Feramisco JR, Lindquist S, ed,Stress-induced proteins. Alan R. Liss, Inc., New Yorkpp15-24
84. DiNocera PP, Dawid lB (1983) Transient expression ofgenes introduced into cultures cells of Drosophila.Proc Natl Acad Sci USA 80:7095-7098
85. Drummond lAS, McClure AA, Poenie M, Tsien RY,steinhardt RA (1986) Large change in intracellular pHand calcium observed during heat shock are notresponsible for the induction of heat shock proteins inDrosophila melanogaster. Mol Cell Biol 6:1767-1775
86. Drummond lAS, Livingstone D, Steinhardt RA (1988) Heatshock protein synthesis and cytoskeletal rearrangementsoccur independently of intracellular free calciumincreases in Drosophila cells and tissues. Rad Res113:402-413
87. Dudler R, Travers AA (1984) Upstream elements necessaryfor optimal function of the hsp70 promotor intransformed flies. Cell 38:391-398
88. Duncan RF, Hershey JWB (1984) Heat shock-inducedtranslational alterations in HeLa cells. J BioI Chem259:11882-11889
89. Duncan RF, Hershey JWB (1989) Protein synthesis andprotein phosphorylation during heat stress, recovery,and adaption. J Cell BioI 109:1467-1481
90. Edmonds M, Vaughn MH, Jr Nakazato H (1971) Polyadenylicacid sequences in the heterogeneous nuclear RNA andrapidly labeled polyribosomal RNA of HeLa cells:
110

Possible evidence for a precursor relationship. ProcNatl Acad sci USA 68:1336-1341
91. Ernst V, Baum EZ, Reddy P (1982) Heat shock, proteinphosphorylation, and the control of translation inrabbit reticulocytes, reticulocyte lysates, and Helacells. IN: Schlesinger MJ, Ashburner M, Tissieres A,ed, Heat shock: from bacteria to man. Cold SpringHarbor Laboratory. Cold spring Harbor, New Yorkpp215-226
92. Falchetto R, Leutenegger S, Bachs 0, Serratosa J,Bloemhard Y, Gazzotti P (1990) Isolation of rat liverspectrin and identification of functional domains.Biochem Biophys Acta 1039:73-80
93. Falkneer FG, Saumweber H, Biessmann H (1981) TwoDrosophila melanogaster proteins related tointermediate filament proteins of vertebrate cells. JCell BioI 91:175-183
94. Farrelly FW, Finkelstein DB (1984) Complete sequence ofthe heat shock-inducible HSP 90 gene of Saccharomycescerevisiae. J BioI Chern 9:5745-5751
95. Findly RC, Pederson T (1981) Regulated transcription ofthe genes for actin and heat shock proteins in culturedDrosophila cells. J Cell BioI 88:323-328
96. Finley D, Crechanover A, Varshavsky A (1984)Thermolability of ubiquitin-activating enzyme from themammalian cell cycle mutant ts85. Cell 37:43-55
97. Finley D, Ozkaynak E, Varshaczky A (1987) The yeastpolyubiquitin gene is essential for resistance to hightemperatures, starvation and other stresses. Cell48:1035-1046
98. Gamborg OL (1970) Effects of amino acids and ammoniumon the growth of plant cells in suspension culture.Plant Physiol 45:372-375
99. Garrels JI (1982) Major phosphoproteins of culturedmuscle cells: kinetics of phosphorylation and inductionby heat shock. IN: Pearson ML, Epstein HF, ed, Muscledevelopment: Molecular and cellular control. Coldspring Harbor Lab. pp43-51
100. Gerner EW, Schneider MJ (1975) Induced thermalresistance in Hela cells. Nature 256:500-502
111

101. Gilmour DS, Lis JL (1986) RNA polymerase II interactswith the promotor region of the noninduced hsp70 genein Drosophila melanogaster cells. Mol Cell BioI 6:39843989
102. Glover CVC (1982) Heat shock induces rapiddephosphorylation of a ribosomal protein in Drosophila.Proc Natl Acad sci USA 79:1781-1785
103. Gromoff EDV, Treier U, Beck CF (1989) Three lightinducible heat shock genes of Chlamydomonasreinhardtii. Mol Cell Biol 9:3911-3918
104. Gubler U, Holfman BJ (1983) A simple and very efficientmethod for generating eDNA library. Gene 25:263-269
105. Gurley WB, Czarnecka E, Nagao RT, Key JL (1986)Upstream sequences required for efficient expression ofa soybean heat shock gene. Mol Cell Biol 6:599-565
106. Gyllensten UB (1989) PCR and DNA sequencing. BioTech.7:700-708
107. Haass C, Klein U, Kloetzel PM (1990) Developmentalregulation of Drosophila melanogaster small heat-shockproteins. J Cell Sci 96:413-418
108. Hackett RW, Lis JT (1983) Localization of the hsp83transcript within a 3292 nucleotide sequence from the63B heat-shock locus of Drosophila melanogaster.Nucleic Acids Res 11:7011-7030
109. Hall BG (1983) Yeast thermotolerance does not requireprotein synthesis. J Bacteriol 156:1363-1365
110. Hanley RM, Means AR, Ono T, Kemp BE, Burgin KE, WaxhamN, Kelly PT (1987) Functional analysis of acomplementary DNA for the 50-kilo0alton subunit ofcalmodulin kinase II. Science 237:293-297
111. Harrington HM, Alm DM (1988) Interaction of heat andsalt shock in cultured tobacco cells. Plant Physiol88:618-625
112. Heikkila JJ, Papp JEF, Schultz GA, Bewley JD (1984)Induction of heat shock protein messenger RNA in maizemesocotyls by water stress, abscisic acid and wounding.Plant physiol 76:270-277
113. Helm KW, Peterson NS, Abornethy RH (1989) Heat shockresponse of germinating embryos of wheat. Plant Physiol90:598-605
112

114. Hepler PK, Wayne RO (1985) Calcium and plantdevelopment. Annu Rev Plant Physiol 36:397-439
115. Heschl MFP, Baillie DL (1989) Characterization of thehdp 70 mUltigene family of Caenorhabditis elegann. DNA8:233-243
116. Holmgran RV, Corces V, Morimoto R, Blackman R,Meselson M (1981) Sequence homologies in the 5' regionof four Drosophila heat-shock genes. Proc Natl Acad SciUSA 78:3775-3778
117. Hultmark D, Klemenz R, Gehring WJ (1986) Translationaland transcriptional control elements in theuntranslational leader of the heat shock gene hsp22.Cell 44:429-438
118. Iida H, Yahara I (1985) Yeast heat-shock protein of Mr
48,000 is an isoprotein of enolase. Nature 315:668-670
119. Ingolia TD, craig EA (1981) Primary sequence of the 5'flanking region of the Drosophila heat shock genes inchromosome subdivision 67B. Nucleic Acids Res9:1627-1642
120. Ingolia TD, craig EA (1982) Four small Drosophila heatshock proteins are related to each other and tomammalian a-crystallin. Proc Natl Acad Sci USA 79:23602364
121. Jackson RJ (1986) The heat-shock response in DrosophilaKc 161 cells: mRNA competition is the main explanationor reduction of normal protein synthesis. Eur J Biochem158:623-634
122. Jakobson BK, Pelham HRB (1988) Constitutive binding ofyeast heat shock factor to DNA in vivo. Mol Cell BioI8:5040-5042
123. James P, Maeda M, Fisher R, Verma AK, Krebs J,Penniston JT, Carafoli E (1988) Identification andprimary structure of a calmodulin binding domain of theCa2+ pump of human erythrocytes. J BioI Chem263:2905-2910.
124. Jamrich M, Greenleaf AL, Bautz EKF (1977) Localizationof RNA polymerase in polytene chromosomes of Drosophilamelanogaster. Proc Natl Acad Sci USA 74:2079-2083
125. Jena PK, Reddy ASN, Poovaiah BW (1989) Molecularcloning and sequencing of a eDNA for plant calmodulin:
113

signal-induced changes in the expression of calmodulin.Proc Natl Acad Sci USA 86:3644-3648
126. Jinn TL, Yeh YC, Chen YM, Lin CY (1989) Stabilizationof soluble proteins in vitro by heat shock proteinenriched ammonium sulfate fraction from soybeanseedling. Plant Cell Physiol 30:463-469
127. Kafatos FC, Johes CW, Efstratiadis A (1979)Determination of nucleic acid sequence homologies andrelative concentrations by a dot hybridizationprocedure. Nuclear Acid Res 7:1541-1552
128. Keene MA, Corees V, Lowenhaupt K, Elgin SCR (1981)DNase I hypersensitive sites in Drosophila chromatin atthe 5' ends of regions of transcription. Proc Natl AcadSci USA 78:143-146
129. Kelley PM, Schlesinger MJ (1978) The effect of aminoacid analogues and heat shock on gene expression inchicken embryo fibroblasts. Cell 15:1277-1286
130. Kelley PM, Schlesinger MJ (1982) Antibodies to twomajor chicken heat shock proteins cross-react withsimilar proteins in widely divergent species. Mol CellBioI 2:267-274
131. Key JL, Lin CY, Chen YM (1981) Heat shock proteins ofhigher plants. Proc Natl Acad Sci USA 78:3526-3530
132. Key JL, Lin CY, Ceglarz E, Sckoffl F (1982) The heatshock response in plants: physiological considerations.IN: Schlesinger MJ, Ashburner M, Tissieres A, ed, Heatshock: from bacteria to man. Cold Spring HarborLaboratory, Cold Spring Harbor, New York pp329-355
133. Key JL, Kimpel JA, Lin CY, Nagao RT, Vierling E,Czarnecka E, Gurley WB, Roberts JK, Mansfield MA,Edelman L (1985) The heat shock response in soybean.IN: Key JL, Kosuge T, ed, Cellular and molecularbiology of plant stress. Alan R. Liss Inc., New Yorkpp161-179
134. Kimpel JA, Key JL (1985) Heat shock in plants. TrendsBiochem Sci 11:353-357
135. Kimpel JA, Nagao RT, Goekjian V, Key JL (1990)Regulation of the heat shock response in soybeanseedlings. Plant Physiol 94:988-995
114

136. Kingston RE, Schuetz TJ, Larin Z (1987) Heat-induciblehuman factor that binds to a human hsp 70 promotor. MolCell BioI 7:1530-1534
137. Klemenz R, Hultmark D, Gehring WJ (1985) Selectivetranslation of heat shock mRNA in Drosophilamelanogaster depends on sequence information in theleader. EMBO J 4:2053-2060
138. Kloetzel PM, Bautz EKF (1983) Heat shock proteins areassociated with hnRNA in Drosophila melanogaster tissueculture cells. EMBO J 2:705-710
139. Kobayashi N, McEntee K (1990) Evidence for a heat shocktranscription factor-independent mechanism forheat shock induction of transcription inSaccharomyces cerevisiae. Proc Natl Acad Sci USA87:6550-6554
140. Koening H, Goldstone AD, Lu CY (1983) ~-adrenergic
stimulation of Ca2+ fluxes, endocytosis, hexosetransport, and amino acid transport in mouse kidneycortex is mediated by polyamine synthesis. Proc NatlAcad Sci USA 80:7210-7214
141. Koyasu S, Nishida E, Miyata Y, Sakai H, Yahara I (1989)Hsp100, a 100-KDa heat shock protein, is a Ca2+ calmodulin-regulated actin-binding protein. J BioI Chern264:15083-15087
142. Kozak M (1988) Leader length and secondary structuremodulated mRNA function under condition of stress. MolCell BioI 8:2737-2744
143. Krishnan M, Nguyen HT, Burke JJ (1989) Heat shockprotein synthesis and thermal tolerance in wheat.Plant Physiol 90:140-145
144. Kudlicki W, Fullilove S, Read R, Kramer G, Hardesty B(1987) Identification of spectric-related peptidesassociated with the reticulocyte heme-controlled asubunit of eukaryotic translational initiation factor 2kinase and a Mr 95,000 peptide that appears to be thecatalytic subunit. J BioI Chern 262:9695-9701
145. Laemmli UK (1970) Cleavage of structural proteinsduring assembly of the head of bacteriophage T4. Nature227:680-685
115

146. Lai BT, Chin NW, stanek AE, Keh W, Lanks KW (1984)Quantitation and intracellular localization of the 85Kheat shock protein by using monoclonal and polyclonalantibodies. Mol Cell BioI 4:2802-2810
147. Lamarche 5, Chretien P, Landry J (1985) Inhibition ofthe heat shock response and synthesis of glucoseregulated proteins in Ca2+-deprived rat hepatoma cells.Biochem Biophys Res Commun 131:868-876
148. Lamphear BJ, Panniers R (1990) Cap binding proteincomplex that restores protein synthesis in heat-shockedEhrlich cell lysate contains highly phosphorylated eIF4E. J BioI Chern 265:5333-5336
149. Lamphear BJ, Panniers R (1991) Heat shock impairs theinteraction of cap-binding protein complex with 5'mRNA cap. J BioI Chern 266:2789-2794
150. Landry J, Chretien P, Bernier 0, Nicole LM, Marceau M(1982) Thermotolerance and heat shock proteins inducedby hyperthermia in rat liver cells. IntI J Radiat OncolBioI Phys 8:59-62
151. Landry J, Crete P, Kamarche 5, Chretien P (1988)Activation of Ca2+-dependent processes during heatshock: Role in cell thermoresistance. Rad Res113:426-436
152. Landry J, P Chretien, H Lambert, E Hickey, LA Weber1989 Heat shock resistance conferred by expression ofthe human HSP27 gene in Rodent cells. J. Cell BioI.109:7-15
153. Lanks KW, Kasambalides EJ (1979) Purification andcharacterization of a major component fromcytoplasmic matrix of cultured murine L cells.Biochem Biophys Acta 578:1-12
154. Laroche A, Hopkins W (1987) Isolation and in vitrotranslation of polysome from mature rye leaves. PlantPhysiol 83:371-376
155. Lehrach H, Diamond 0, Wozney JM, Boedtker H (1977) RNAmolecular weight determinations by gel electrophoresisunder denature conditions, a critical reexamination.Biochem 16:4743-4751
156. Lenk R, Ranson L, Kaufmann Y, Penman S (1977) Acytoskeletal structure with associated polyribosomesobtained from Hela cells. Cell 10:67-68
116

157. Leto TL, Pliasic S, Forget BG, Benz EJ, Marchesi V(1989) Characterization of the calmodulin-binding siteof nonerythroid a-spectrin: Recombinant protein andmodel peptide studies. J BioI Chern 264:5826-5830
158. Levis E, Penman S (1978) 5 1-terminal structure ofpoly(A)+ cytoplasmic messenger RNA and of poly(A) + andpoly(A)- heterogeneous nuclear RNA of cells of thedipteran Drosophila melanogaster. J Mol BioI 120487-515
159. Li GC, Laszlo A (1985) Thermotelerance in mammaliancells: a possible role for the heat shock proteins.IN: Atkinson BG, Waldon DB, ed, Changes in eukaryoticgene expression in response to environmental stress.Academic Press, Inc., New York pp227-254
160. Lindquist S (1980a) Translational efficiency ofheat-induced messages in Drosophila melanogaster cells.J Mol Biol 137:151-158
161. Lindquist S (1980b) Varying patterns of proteinsynthesis during heat shock: implications forregulation. Dev Biol 77:463-479
162. Lindquist S (1981) Regulation of protein synthesisduring heat shock. Nature 293:311-314
163. Lindquist S (1986) The Heat-shock response. Ann RevBiochem 55:151-191
164. Lindquist S (1987) Translational regulation in theheat-shock response of Drosophila cells. IN: J Ilan,ed, Translational regulation of gene expression.Plenam Press, New York, London, pp187-207
165. Lindquist S, craig EA (1988) The heat-shock proteins.Ann Rev Genet 22:631-677
166. Ling V, Zielinski RE (1989) Cloning of eDNA sequenceencoding the calcium-binding protein, calmodulin, frombarley (Hordeum vulgare L.). Plant Physiol 90:714-719
167. Lis JT, Xiao H, Perisic 0 (1989) A structural unit ofheat shock regulatory region. IN: Pardue ML, FeramiscoJR, Lindquist S, ed, stress-induced proteins. Alan R,Liss, Inc., New York, pp73-82
168. Loomis WF, Wheeler SA (1982) The physiological role ofheat-shock proteins in Dictyostelium. IN: SchlesingerMJ, Ashburner M, Tissieres A, ed, (1982) Heat shock
117

from bacteria to man. Cold Spring Harbor, New Yorkpp353-360
169. Lukas TJ, Burgess WE, Prendergast FG, Lau W, WattersonDM (1986) Calmodulin binding domains: Characterizationof phosphorylation and calmodulin binding site frommyosin light chain kinase. Biochem 25:1458-1464
170. MacManus JP, Gillen NF, Korczak B, Nojima H (1989)differential calmodulin gene expression in fetal,adult, and cytoplasmic tissues of rodents. BiochemBiophys Res Commun 159:278-282
171. Maniatis T, Fritsch E, Sambrook J (1982) Molecularcloning. A laboratory Mannul. Cold Spring HarborLaboratory, Cold spring Harbor, New York
172. Mans RJ, Novelli GD 1961 Measurement of incorporationof radioactive amino acids into protein by a filterpaper disk method. Arch Biochem Biophys 94:48-53
173. Mansfield MA, Key Jl (1987) synthesis of the lowmolecular weight heat shock proteins in plants. PlantPhysiol 84;1007-1017
174. Mariano TM, Siekierka J (1986) Inhibition of HeLa cellprotein synthesis under heat shock conditions in theabsence of initiation factor elF-2a phosphorylation.Biochem Biophys Res Commun 138:519-525
175. Marime D (1989) The role of calcium and calmodulinsignal transduction. IN: Second messengers in plantgrowth and development. Alan R. Liss, Inc., pp57-80
176. McAlister L, Finkelstein DB (1980a) Alterations intranslatable ribonucleic acid after heat shock ofSaccharomyces cerevisiae. J Bacteriol 143:603-612
177. McAlister L, Finkelstein DB (1980b) Heat shock proteinsand thermal resistance in yeast. Biochem Biophys ResCommun 93:819-824
178. Mcgarry TJ, Lindquist S (1985) The preferentialtranslation of Drosophila hsp70 mRNA requires sequencesin the untranslated leader. Cell 42:903-911
179. McGookin R (1984) RNA extraction by the guanidinethiocyanate procedure. IN: JM Walker, ed, Methods inmolecular biology. Vol 2. Human Press. Clifton, NewJersey, ppl13-116
118

180. Mckenzie SL, Henikoff S, Meselson M (1975) Localizationof RNA from heat-induced polysome at pUff sites inDrosophila melanogaster. Proc Natl Acad Sci USA72:1117-1121
181. Mezger V, Bensaude 0, Morange M (1989) Unusual levelsof heat shock element-binding activity in embryonalcarcinoma cells. Mol Cell BioI 93888-3896
182. Mirault ME, Southgate R, Delwart E (1982) Regulation ofheat shock genes: A DNA sequence upstream of Drosophilahsp 70 genes is essential for their induction in monkeycells. EMBO J 1:1279-1285
183. Misra UK, Sahyoun N (1987) Protein kinase C binding toisolated nuclei and its activation by aCa2+/phospholipid-independent mechanism. BiochemBiophys Res Commun 145:760-768
184. Mitchell HK, Lipps LS (1978) Heat shock and phenocopyinduction in Drosophila. Cell 15:907-918
185. Moisyadi S, Harrington HM (1989) Characterization ofheat shock response in cultured sugarcane cells: I.Physiology of the heat shock response and heat shockprotein synthesis. Plant Physiol 90:1156-1162
186. Mosser DD, Kotzbauer PT, Sarge KD, Morimoto RI (1990)In vitro activation of heat shock transcription factorDNA-binding by calcium and biochemical caonditions thataffect protein conformation. Proc Natl Acad Sci USA87:3748-3752
187. Nagao RT, Czarnecka E, Gurley WB, SchoffI F, Key JL(1985) Genes for low-molecular-weight heat shockproteins of soybeans: sequence analysis of a mUltigenefamily. Mol Cell BioI 5:3417-3428
188. Nagao RT, Kimpel JA, Vierling E, Key JL (1986) The heatshock response: A comparative analysis. IN: BJ Miflined, Oxford Surveys of Plant Molecular and Cell Biology,Vol 3. Oxford University Press, New York, pp384-438
189. Nagao RT, Key JI (1989) Heat shock protein genes ofplants. IN: Cell Culture and Somatic Cell Genetics ofPlants, Academic Press, Inc., 6:297-328
190. Neidhardt FC, VanBogelen RA, Vaughn V (1984) Thegenetics and regulation of heat-shock proteins. Ann RevGenet 18:295-329
119

191. Nieto-sotelo J, Wiederrecht G, Okuda A, Parker CS(1990) The yeast heat shock transcription factorcontains a transcriptional activation domain whoseactivity is repressed under nonshock condition. Cell62:807-817
192. Nishida E, Koyasu S, Sakai H, Yahara I (1986)Calmodulin-regulated binding of the 90-KDa heat shockprotein to actin filaments. J BioI Chem 261:16033-16036
193. Nojima H (1989) Structural organization of multiple ratcalmodulin genes. J Mol BioI 208:269-282
194. Nover L, Scharf KD, Neumann D (1983) Formation ofcytoplasmic heat-shock granules in tomato cells. MolCell BioI 3:1648-1655
195. Nover L. Scharf KD (1984a) Synthesis, modification andstructural binding of heat-shock proteins in tomatoculture cells. Eur J Biochem 139:303-313
196. Nover L, Scharf KD (1984b) The heat shock proteins. IN:Nover L, ed, Heat shock response of eUkaryotic cells.Springer-Verlag, Berlin, Heidelberg, New York, pp1-7
197. Nover L (1984) Changing translation patterns IN: NoverL, ed, Heat shock response of eUkaryotic cells.springer-Verlag Berlin Heideberg, New York, pp28-34
198. Nover L, Hellmund D, Neumann D, Scharf KD, Serfling E(1984) The heat shock response of eUkaryotic cells.BioI Zentralbl 103:357-435
199. Nover L, Munsche 0, Neumann 0, Ohme K, Scharf KD (1986)Control of ribosome biosynthesis in plant cell culturesunder heat-shock conditions. Ribosomal RNA. Eur JBiochem 160:297-304
200. Nover L, Scharf KD, Neumann D (1989) Cytoplasmic heatshock granules are formed from precursor particles andare associated with a specific set of mro~As. Mol CellBioI 9:1298-1308
201. O'Farrel PH (1975) High resolution two-dimensionalelectrophoresis of proteins. J BioI Chem 250:4007-4021
202. O'Farrel PZ, Goodman HM, O'Farrel PH (1977) highresolution two dimensional electrophoresis of basic aswell as acidic proteins. Cell 12:1133-1142
120

203. O'Neil KT, DeGrado WF (1990) How calmodulin binds itstargets: sequence independent recognition ofamphiphilic a-helices. Trends Biochem Sci 15:59-64
204. Oppermann H, Levinson W, Bishop JM (1981) A cellularprotein that associates with the transforming proteinof Rous sarcoma virus is also a heat-shock protein.Proc Natl Acad Sci USA 78;1067-1071
205. Ostermann J, Horwich AL, Neupert W, Hartl FU (1989)Protein folding in mitochondria requires complexformation with hsp60 and ATP hydrolysis. Nature314;125-130
206. Panniers R, Scorsone KA, Wolfman A, Hucul JA, HenshawEC (1986) Regulation of translation through modulationof energy charge, intercellular calcium level, e1F-2phosphorylation, and eIF-4A activity. IN: Mathews MBed, Current communications in molecular biology.Translational control. Cold Spring Harbor LaboratoryPress, New York, pp52-57
207. Parag HA, Raboy B, Kulka RG (1987) Effect of heat shockon protein degradation in mammalian cells: involvementof the ubiquitin system. EMBO J 6:55-61
208. Parker CS, Topol J (1984) A Drosophila RNA polymeraseII transcription factor binds to the regulatory site ofhsp70 gene. Cell 37:273-283
209. Parker PJ, Bajaj M, Marais R, Mitchll F, Pears C,Stabel S (1989) Protein kinase C as a second messengertarget. Biochem Soc Trans 17:279-280
210. Pelech SL, Vance DE (1989) signal transduction viaphosphatidylcholine cycles. Trends Biochem Sci 14:31-33
211. Pelham HRB (1982) A regulatory upstream promotorelement in Drosophila hsp 70 heat-shock gene. Cell30:517-528
212. Pelham HRB, Bienz M (1982) A synthetic heat-shockpromotor element confers heat-inducibility on theherpes simplex virus thymidine kinase gene. EMBO J.1:1473-1477
213. Pelham H, Lewis M, Lindquist S (1984) Expression of aDrosophila heat shock protein in mammalian cells:transient association with nucleoli after heat shock.Phil Trans R Soc Lond B 307:301-307
121

214. Pelham HRB (1984) Hsp 70 accelerates the recovery ofnucleolar morphology after heat shock. EMBO J 3:30953100
215. Pelham H (1988) Coming in from the cold. Nature332:776-777
216. Pelletier J, Sonenberg N (1988) Internal initiation oftranslation of eUkaryotic mRNA directed by a sequencederived from poliovirus RNA. Nature 334:320-325
217. Pelletier J, Kaplan G, Racaniello VR, Sonenberg N(1988) Cap-independent translation of poliovirus mRNAis conferred by sequence elements within 51 noncodingregion. Mol Cell BioI 8:1103-1112
218. Perisic 0, Xiao H, Lis JT (1989) Stable binding ofDrosophila heat shock factor to head-to-head and tailto-tail repeats of a conserved 5bp recognition unit.Cell 59:797-806
219. Petersen NS, Mitchell HK (1981) Recovery of proteinsynthesis after heat shock: Prior heat treatmentaffects the ability of cells to translate mRNA. ProcNatl Acad Sci USA 78:1708-1711
220. Petko L, Lindquist S (1986) HSP26 is not required forgrowth at high temperature, nor for thermotolerance,spore development and germination. Cell 45:885-894
221. Piazza GJ (1988) Calmodulin in plants. IN: Thompson MPed, Calcium-binding proteins. Vol. I. Characterizationand properties. eRC press, Inc., Boca Raton, Floridapp127-143
222. Poovaiah BW, Reddy ASN (1987) Calcium messenger systemin plants. CRC critical Rev Plant Sci 6:47-103
223. Rasmussen H, Barrett PQ (1984) Calcium mesenger system:an integrated wiew. Physiol Rev 64:938-984
224. Rees CAB, Gullons AM, Walden DB (1989) Heat shockprotein synthesis induced by methomyl in Maize (Zeamays L.) seedling. Plant Physiol 90:1256-1261
225. Resendez Jr E, Attenello JW, Grafsky A, Chang CS, LeeAS (1985) Calcium ionophore A23187 induces expressionof glucose-regulated genes and their heterology fusegene. Mol Cell BioI 5:1212-1219
122

226. Riabowol KT, Mizzen LA, Welch WJ (1988) Heat shock islethal to fibroblasts microinjected with antibodiesagainst hsp 70. Science 242:433-436
227. Ritossa F (1962) A new pUffing pattern induced by heatshock and DNP in Drosophila. Experimenta 18:571-573
228. Rivin CJ, Zimmer EA, Walbot V (1982) Isolation of DNAand DNA recombinants from maize. IN: Sheridan WF, ed,Maize for biological research. A special publication ofthe plant molecular biology association. UniversityPress, University of North Dakota pp161-164
229. Roberts JK, Key JL (1985) Characterization of the genesfor the heat shock 70 KD and 80 KD proteins in soybean.Proc Int Congr Plant Mol BioI 1st p137 (abstract)
230. Roberts DM, Lukes TJ, Watterson DM (1985) Structure,function, and mechanism of action of calmodulin. CRCcritical Rev Plant Sci 4:311-339
231. Roberts DM, Crea R, Malecha M, Alvarado-Urbina G,Chiarello Rh, Watterson DM (1985) Chemical synthesisand expression of a calmodulin gene designed for sitespecific mutagenesis. Biochem 24:5090-5098
232. Rochester DE, winter JA, Shah DM (1986) The structureand expression of maize genes encoding the major heatshock protein, hsp70. EMBO J 5:451-458
233. Rose DW, Wettenhall REH, Kudlicki W, Kramer G, HardestyB (1987) The 90-kilodalton peptide of theheme-regulated elF-2a kinase has sequence similaritywith the 90-kilodalton heat shock protein. Biochem6583-6587
234. Rose DW, Welch WJ, Kramer G, Hardesty B (1989) Possibleinvolvement of the 90-KDa heat shock protein in theregulation of protein synthesis. J BioI Chern 264:62396244
235. Rosen E, Sivertsen A, Firtel RA, Wheeler A, Loomis WF(1985) Heat shock genes of Dictyostelium. IN: AtkinsonBG, Walden DB, ed, Changes in eUkaryotic geneexpression in response to enviromental stress.Academic Press, Inc., New York, pp257-278
236. Rougvie AE, Lis JT (1988) The RNA polymerase IImolecule at the 5' end of the uninduced hsp70 gene ofD. melanogaster is transcriptionally engaged. Cell54:795-804
123

237. Sachs MM, Ho THO (1986) Alteration of gene expressionduring environmental stress in plants. Ann Rev PlantPhysiol 37:363-376
238. Sanchez EH, Toft DO, Schlesinger MJ, Pratt WB (1985)Evidence that the 90-KOa phosphoprotein associated withthe untransformed L-cell glucocorticoid receptor is amurine heat-shock protein. J BioI Chem 260:12398-12401
239. Sanchez ER, Meshinchi S, Tienrungroj W, Schlesinger MJ,Toft DO, Pratt WB (1987) Relationship of the 90-KOamurine heat shock protein to the untransformed andtransformed states of the Hale cell glucocorticoidreceptor. J BioI Chem 262:6986-6991
240. Sander MM, Triemer OF, Olsen AS (1986) Regulation ofprotein aynthesis in heat-shocked Drosophila cells. JBioI Chern 261:2189-2196
241. Sanger F, Nicklen S, Coulsen AR (1977) DNA sequencingwith chain terminating inhibitors. Proc Natl Acad SciUSA 74:5463-5467
242. Sanger F, Coulsen AR (1978) The use of thin acrylamidegels for DNA sequence. FEBS Lett 87:107-110
243. Scharf KO, Nover L (1982) Heat-shock-inducedalterations of ribosomal protein phosphorylation inplant cell cultures. Cell 30:427-437
244. Scharf KD, Nover L (1987) Control of ribosomebiosynthesis in plant cell culture under heat shockconditions. II. Ribosomal proteins. Biochim BiophysActa 909:44-57
245. Scharf KD, Rose S, Zott W, Schoff F, Nover L (1990)Three tomato genes code for heat shock stresstranscription factor with a region of remarkablehomology to the DNA-binding domain of the yeast HSF.EMBO J 9:4495-4501
246. Schlesinger MJ, Ashburner M, Tissieres A (1982) Heatshock from bacteria to man. Cold Spring Harbor, NowYork
247. Schlesinger MJ, Aliperti G, Kelley PM (1982) Theresponse of cells to heat shock. Trends Biochem Sci7:222-225
248. Schlesinger MJ (1990) Heat shock proteins. J BioI Chem265:12111-12114
124

249. Schoffl F, Raschke E, Nagao RT (1984) The DNA sequenceanalysis of soybean heat-shock genes and identificationof possible regulatory promotor elements. EMBO J3:2491-2497
250. Schoffl F, Baumann G, Raschke E (1988) The expressionof heat shock genes-A model for environmental stressresponse. IN: Verma DPS, Goldberg RB, ed, Plant generesearch: Temporal and spatial regulation of plantgenes. Springer-Verlag wien New York pp253-273
251. Schoffl F, Rieping M, Baumann G, Bevan M, Angermuller S(1989) The function of plant heat shock promotorelements in the regulated expression of chimeric genesin transgenic tobacco. Mol Gen Genet 217:246-253
252. Schuh S, Yonemoto W, Brugge J, Bauer VJ, Riehl RM,Sullivan WF, Toft DO (1985) A 90,000-dalton bindingprotein common to both steroid receptors and the Roussarcoma virus transforming protein, pp60v-sn: . J BioIChem 260:14292-14296
253. Schuman H, Lou LI (l989) MultifunctionalCa2+/calmodulin-dependent protein kinase: domainstructure and regulation. Trends Biochem Sci 14:62-66
254. Scott MP, storti RV, Pardue ML, Rich A (1979) Cell-freeprotein synthesis in lysates of Drosophila melanogastercells. Biochem 18:1588-1594
255. Scott MP, Pardue ML (1981) Translational control inlysates of Drosophi~a melanogaster cells. Proc NatlAcad Sci USA 78:3353-3357
256. Shimogawa~a K, Muto S (1989) Heat shock induced changein protein ubiquitination in Chlamydomonas. Plant CellPhysiol 30:9-16
257. Short JM, Fernandez JM, Sorge JA, Huse WD (1988) AZAP:a bacteriophage A expression vector with in vivoexcision properties. Nuclear Acid Res 16:7583-7600
258. Shuey OJ, Parker CS (1986) Binding of Drosophila heatshock gene transcription factor to the hsp70 promotor:evidence for symmetric and dynamic interactions. J BiolChern 261:7934 170.
259. Sikela JM, Hahn WE (1987) screening an expressionlibrary with a ligand probe: Isolation and sequence ofa cDNA corresponding to a brain calmodulin-bindingprotein. Proc Natl Acad Sci USA 84:3038-3042
125

260. Simon JA, sutton CA, Lobell RB, Glaser RL, Lis JT(1985) Determinants of heat-shock-induced chromosomepUffing. Cell 40:805
261. Singh H, Clerc RG, Lebowitz JH (1989) Molecular cloningof sequence-sppecific DNA binding proteins usingrecognition site probes. BioTech 7:252-261
262. Sinibaldi RM, Coldiron P, Dietrich PS (1985) Expressionof putative maize heat shock clones. Proc Int CongrPlant Mol BioI 1st p34 (Abstract)
263. Somers DJ, Cummins WR, Filion WG (1989)Characterization of the heat-shock response in spinach(Spinacia oleracea L.). Biochem cell BioI 67:113-120
264. Sorger PK, Lewis MJ, Pelham HRB (1987) Heat shockfactor is regulated differently in yeast and Helacells. Nature 329:81-84
265. Sorger PK, Pelham HRB (1988) Yeast heat shock factor isan essential DNA-binding protein that exhibitstemperature dependent phosphorylation. Cell 54:855-864
266. Southgate R, Ayme A, Voellmy R (1983) Nucleotidesequence analysis of Drosophila small heat shockcluster at locus 67B. J Mol BioI 165:35-57
267. Spena A, Hain R, Ziervogal U, Saeler H, Schell J (1985)Construction of a heat-inducible gene for plants.Demonstration of heat-inducible activity of theDrosophila hsp70 promotor in plants. EMBO J 4:2739-2743
268. Spena A, Schell J (1987) The expression of a heatinducible chimeric gene in transgenic tobacco plants.Mol Gen Genet 206:436-440
269. Spradling A, Penman S, Pardue ML (1975) Analysis ofDrosophila mRNA by in situ hybridization: Sequencetranscribed in normal and heat shocked cultured cells.Cell 4:395-404
270. Stevenson MA, Calderwood AK, Hahn GH (1986) Rapidincreases in inositol triphosphate and intracellularCa++ after heat shock. Biochim Biophys Res Commun137:826-833
271. Stevenson MA, Calderwood SK (1990) Members of the 70kilodalton heat shock protein family contain a highlyconserved calmodulin-binding domain. Mol Cell BioI10:1234-1238
126

272. Sun S, Buchbinder BU, Hall TC (1975) Cell-freesynthesis of the major storage protein of the bean,Phaseolus vulgaris L. Plant Physiol 56:780-785
273. Swan DG, Hale RS, Dhillon N, Leadlay PF (1987) Abacterial calcium-binding protein homologous tocalmodulin. Nature 329:84-85
274. Takahashi T, Komeda Y (1989) Characteriation of twogenes encoding small heat-shock proteins in Arabidopsisthaliana. MGG 219:365-372
275. Tangay RM (1983) Genetic regulation during heat shockand function of heat-shock proteins: A review. Can JBiochem Cell BioI 61:387-394
276. Tanguay RM (1988) Transcriptional activation of heatshock genes in eukaryotes. Biochem Cell BioI 66:584-593
277. Thomas PS (1980) Hybridization of denatural RNA andsmall DNA fragment transferred to nitrocellulose. ProcNatl Acad Sci USA 77:5201-5205
278. Topol J, Ruden OM, Parker CS (1985) Sequences requiredfor in vitro transcriptional activation of a Drosophilahsp70 gene. Cell 42:527-537
279. Udvardy A, Schedl P (1984) Chromatin organization ofthe 87A7 heat shock locus of Drosophila melanogaster. JMol BioI 172:385-403
280. Ungewickell E (1985) The 70-kd mammalian heat shockproteins are structurally and functionally related tothe uncoating protein that release clathrin triskelionfrom coated vesicles. EMBO J 4:3385-3391
281. Van Eldik LJ, Zendegui JG, Marshak DR, Watterson DM(1982) calcium-binding proteins and the molecular basisof calcium action. Int Rec Cytol 77:1-61
282. Van Eldik LJ, Burgess WH (1983) Analytical subcellulardistribution of calmodulin and calmodulin-bindingproteins in normal and virus-transformed fibroblasts.J BioI Chern 258:4539-4547
283. Vanbogelen RA, Kelley PM, Neidhardt FC (1987)Differential induction of the heat shock, SOS andoxidation stress regulons and accumulation ofnucleotides in Escherichia coli. J Bacteriol 169:26-32
127

284. Velazquez JM, DiDomenico BJ, Lindquist S (1980)Intracellular localization of heat shock proteins inDrosophila. Cell 20:679-689
285. Velasquez JM, Sonoda S, Bugaisky G, Lindquist S (1983)Is the major Drosophila heat shock protein present incells that have not been heat shocked? J Cell BioI96:286-290
286. Velazquez JM, Lindquist S (1984) Hsp 70: nuclearconcentration during environmental stress; cytoplasmicstorage during recovery. Cell 36:655-662
287. Vincent M, Tanguay RM (1982) Different intracellulardistributions of heat-shock and arsenite-inducedproteins in Drosophila Kc cells. J Mol BioI 162:365-378
288. Voellmy R, Rungger 0 (1982) Transcription of aDrosophila heat shock gene is heat-induced in Xenopusoocytes. Proc Natl Acad sci USA 79:1776-1780
289. Vorherr T, James P, Krebs J, Enyedi A, McCormick OJ,Penniston JT, Carafoli E (1990) Interaction ofcalmodulin with the calmodulin binding domain ofthe plasma membrane Ca2+ pump. Biochem 29:355-365
290. Wang C, Gomer RH, Lazarides E (1981) Heat shockproteins are methylated in avian and mammalian cells.Proc Natl Acad sci USA 78:3531-3535
291. Wang C, Lazarides E, O'Connor CM, Clarke ST (1982)Methylation of chicken fibroblast heat shock proteinsat lysyl and arginyl residues. J BioI Chern 257:83568362
292. Watterson OM, Vanaman TC (1976) Affinity chromatographypurification of a cyclic nucleotide phosphodiesteraseusing immobilized modulator protein, a troponin C-likeprotein from brain. Biochem Biophys Res Commun 73:40-46
293. Welch WJ, Garrels JI, Thomas GP, Lin JJC, Ferarnisco JR(1983) Biochemical characterization of the mammalianstress proteins and identification of two stressproteins as glucose- and Ca2+-ionophore-regulatedproteins. J BioI Chem 258:7102-7111
294. Welch WJ, Feramisco JR (1984) Nucleolar localization ofthe 72,000 dalton heat-shock protein in mammaliancells. J BioI Chem 259:4501-4513
128

295. Welch WJ, Feramisco JR (1985) Rapid purification ofmammalian 70,000-dalton stress proteins: Affinity ofthe proteins for nucleotides. Mol Cell BioI 5:1229-1237
296. Widada JS, Ferraz C, Asselin J, Trave G, Colote S,Haiech J, Marti J, Liautard JP (1989) Cloning anddeletion mutagenesis using direct protein-proteininteraction on an expression vector. Identification ofthe calmodulin binding of a-fodrin. J Mol BioI 205:455458
297. Wiederrecht G, Shuey DJ, Kibbe WA, Parker CS (1987) TheSaccharomyces and Drosophila heat shock transcriptionfactors are identical in size and DNA bindingproperties. Cell 48:507-515
298. Wiederrechi G, Seto D, Parker CS (1988) Isolation ofthe gene encoding the s. cerevisiae heat shocktranscription factor. Cell 54:841-853
299. Wiegant FAC, Tuyl M, Linnemans W (1985) Calmodulininhibitors prevent heat-induced cytoskeletalreorganization and potentiate hyperthermic cellkilling. Int J Hyperthermia 1:157-169
300. Wu C (1980) The 5' ends of Drosophila heat shock genein chromatin are hypersensitive to DNase I. Nature286:854-860
301. Wu C (1984a) Two protein-binding sites in chromatinimplicated in the activation of heat-shock genes.Nature 309:229-234
302. Wu C (1984b) Activating protein factor binds in vitroto upstream control sequences in heat-shock genechromatin. Nature 311:81-84
303. Wu CH, Caspar T, Browse J, Lindquist S, Somerville C(1988) Characterization of an HSP70 cognate gene familyin Arabidopsis. Plant Physiol 83:731-740
304. Wu FS, Park YC, Roufa D, Martinosi A (1981) Selectivestimulation of the synthesis of an 80,000-daltonprotein by calcium ionophores. J BioI Chem256:5309-5312
305. winter J, Wright R, Duck N, Gasser C, Fraley R, Ahah D(1987) The inhibition of Petunia hsp70 mRNA processingduring CdC~ stress. Mol Gen Genet 211:315-319
129

306. Xiao H, Lis JT (1988) Germline transformation used todefine key features of heat-shock response elements.Science 239:1139-1142
307. Yamamoto T, Yura T (1980) Temperature-induced synthesisof specific proteins in Escherichia coli: Evidence fortranscription control. J Bacteriol 142:843-851247.
308. Young RA, Davis RW (1983) Efficient isolation of genesby using antibody probes. Proc Natl Acad Sci USA80:1194-1198
309. Yost HJ, Lindquist S (1986) RNA splicing is interruptedby heat shock and is rescued by heat shock proteinsynthesis. Cell 45:185-193
310. Yost HJ, Lindquist S (1991) Heat shock proteins affectRNA processing during the heat shock response ofSaccharomyces cerevisiae. Mol Cell BioI 11:1062-1068
311. Ziemiecki A (1986) Characterization of the monomericand complex-associated forms of the gag-one fusionproteins of three isolates of feline sarcoma virusphosphorylation, kinase activity, acylation, andkinetics of complex formation. Virology 151:265-273
312. Zirnarino V, Wu C (1987) Induction of sequence-specificbinding of Drosophila heat shock activator proteinwithout protein synthesis. Nature 327:727-730
313. Zirnarino V, Tsai C, Wu C (1990) Complex modes of heatshock factor activation. Mol Cell BioI 10:752-759
314. Zimmer WE, Schloss JA, silflow CD, Youngblom J,Watterson DM (1988) Structural organization, DNAsequence, and expression of the calmodulin gene. J BioIChern 262:19370-19383
315. Zimmerman JL, Petri W, Meselson M (1983) Accumulationof a specific subset of D. melanogaster heat shockmRNAs in normal development without heat shock. Cell32:1161-1170
316. Zylicz M, Lebowitz JH, McMachen R, Georgopoulos C(1983) The dnak protein of Escherichia coli possessesan ATPase and autophosphorylating activity and isessential in an in vitro DNA replication system. Proc.Natl Acad Sci USA 80:6431-6435
130


















