
virologia veterinaria
386
TEXTBOOK OF VETERINARY VIROLOGY Prof. S. N. Sharma Dr. S. C. Adlakha International Book Distributing Co.
-
Upload
gladys-aguilar -
Category
Documents
-
view
187 -
download
6
description
Libro
Transcript of virologia veterinaria

TEXTBOOK
OF VETERINARY
VIROLOGY
Prof. S. N. Sharma Dr. S. C. Adlakha
International Book Distributing Co.

TEXTBOOK
OF
VETERINARY VIROLOGY


Textbook of Veterinary Virology
Prof S N Sharma Ex Professor of Virology
Department of Veterinary Microbiology Punjab Agricultural University
Ludhiana
Dr S C Adlakha Ex President
National Academy of Veterinary Sciences New Delhi
• International Book Distributing Co. (Publishing Division)

Published by
INTERNATIONAL BOOK DISTRIBUTING CO. (Publishing Division) Khushnuma Complex Basement 7, Meerabai Marg (Behind Jawahar Bhawan) Lucknow 226 001 V.P. (INDIA) Tel.: 91-522-2209542,2209543, 2209544,2209545 Fax: 0522-4045308 E-Mail: [email protected]
First Reprint 2009
ISBN 978-81-8189-274-4
© Publisher All Rights Reserved
No part of this publication may be reproduced, stored in a retrieval system, or transmitted, in any form or by any means, electronic, mechanical, photocopying, recording or otherwise, without the prior written permission of the publisher.
Printed at:
Salasar Imaging Systems C-7/5, Lawrence Road Industrial Area Delhi -110 035 Tel. : 011-27185653, 9810064311

Preface
This book is intended to fulfil the need of veterinary students in general and Post-Graduates in Microbiology in particular. besides the veterinary disease Investigators and Practitioners of veterinary medicine. Virology is one of those branches of science which has experienced a tremendous growth during the last few years especially in the area of Molecular Virology. The resultant information is spread over a number of publications. An attempt has been made to present all the relevant information in a concise manner including the latest advances.
This book is divided into two parts: General Virology and Systematic Virology. There is plethora of literature on general virology, yet the authors have tried to present the basic principles of animal virology in a concise manner with the hope that the reader appreciates the nature of viruses, their pathogenicity. replication etc. In.the second part information on infections of vertebrates has been given with emphasis on the diagnostic and preventive aspects of virus infections of domestic animals and poultry. The organization of chapters is hierarchial and follows the taxonomy of animal viruses. A short family description precedes each chapter. To present the material in a limited number of pages, the authors have given only selected references at the end of each chapter. There are more viruses in domestic animals and birds than those discussed in this book; the viruses of little or no pathogenic importance or viruses encountered as ·contaminants in animal cell culture have been omitted. Greater importance has been given to viruses of economic importance in India and other developing countries of Asia and Africa.
The authors will feel rewarded if this book will meet the requirements of the veterinary profession in the developing countries. The suggestions for improvement of this book in a future edition are welcome.
Authors


Contents
Abbreviations xi PART I
General Virology
1. Structure and Composition 3 2. Classification of Viruses 13 3. Viral Replication 34 4. Cultivation of Viruses 46 5. Viral Genetics 55 6. Viral Pathogenesis 65 7. Persistent Infections 76 8. Viral Immunity 81 9. Epidemiology of Viral Diseases 87
10 .. Viral Tumorogenesis 94 11. Viral Vaccines and Antiviral Agents 102 12. Diagnosis of Viral Diseases 115
PARTll Systematic Virology
D.N.A. Viruses
13. Poxviridae 129 Vaccinia Virus; Cow Pox Virus; Buffalo Pox Virus; Camel Pox Virus; Sheep Pox Virus; Goat Pox Virus; Lumpy Skin Disease; Ecthyma (Ort) Virus; Bovine Papular Stomatitis Virus; Milkers Node Virus; Swine Pox Virus; Myxoma Virus; Fibroma Virus; Fowl Pox Virus,
14. Parvoviridae 150 Bovine Parvovirus; Porcine Parvovirus; Feline Parvovirus; Canine Parvoviruses.

\/iii TeXlbook of Veterinary Virology
15. Papovaviridae 159 Bovine Papillomavirus type 1 and 2; Bovine Papilloma virus type-3; Bovine Papillomavirus type-4; Bovine Papillomavirus type-5; Bovine Papilloma virus type-6; Canine Papillomavirus; Rabbit Papillomavirus; Equine Papillomavirus.
16. Adenoviridae 164 Bovine Adenoviruses; Ovine Adenovirses; Canine Adenoviruses; Infectious Canine Laryngotracheitis Virus; Equine Adenoviruses; Porcine Adeno-viruses; Avian Adenoviruses.
17. Hcrpesviridae 176 Bovine Herpes Virus-I; Bovine Herpes Virus-2; Malignant Catarrhal Fever Virus; Bovine Herpes Virus-3; Hcrpes Virus of Sheep; Herpes Virus of Goats; Equine Herpes Viruses; Pseudorabies Virus; Simian Herpes Virus-I; Canine Herpes Virus; Fowl Hcrpes Virus-I; Duck Herpes Virus-I; Pigcon Hcrpes Virus; Marek's Disease Virus.
18. Unclassified DNA Virus 206 African Swinc Fever Virus.
RNA Viruses
19. Picomaviridae 211 Apthovirus; Enterovirus; Swine Enteroviruses; Porcine Enterovirus-I; Porcine Enterovirus-9; Bovjne Enteroviruses; Avian Encephalomyelitis Virus; Duck Hepatitis Virus; Bovine Rhino-virus-I; Equine Rhinovirus 1 & 2.
20. Calciviridae 231 Vesicular Exanthema Virus; Feline Calcivirus.
21. Togaviridae 234 Alphavirus; Equine Encephalomyelitis Virus; Pestivirus; Bovine Viral Diarrhoea Virus; Border Disease Virus; Swine Fever Virus; Arterivirus; Equine Viral Arteritis.
22. Flaviviridae 247 Japanese B Encephalitis Virus; Wesselsbom Virus; Louping III Virus.

COnlellls
23. Reoviridae 251 Reovirus; Bovine Reovirus (type 1 & 3); Avian Reovirus type 1 to 5; Rotavirus; Bovine Rota-virus; Orbivirus; Blue Tongue Virus; AfricIDl Horse Sickness Virus.
24. Bimaviridae 264 Infectious Bursal Disease Virus.
25. Coronaviridae Bovine Corona Virus; Canine Corona Virus; Feline Infectious Peritonitis Virus; Porcine Corona Viruses; Avian Infcctious Bronchitis Virus.
267
26. Orthomyxoviridae 278 Equine Influenza Virus 1 and 2; Swine Influenza Virus; Avian Influenza Virus; Fowl Plague Virus.
27. Paramyxoviridae 284 Paramyxoviruses; Avian Parnmyxoviruses; New Castle Disease Virus; Mammalian Parninfluenza Viruses; Parainfluenza-l; Parainfluenza-3 Virus in cattle; Parainfluenza-3 Virus in sheep; Parninfluenza-5; Morbillivirus; Canine Distemper Virus; Rinderpest Virus; Peste-Des-Petits Virus; Pneumovirus; Bovine Respiratory Syncytial Vims.
28. Rhabdoviridae 309 Vesicular Stomatitis Virus; Rabies Virus; Bovine Ephemeral Fever Virus; Marburg Vims.
29. Rettoviridae 322 Oncoviruses; Bovine Leukaemia Virus; Feline Leukaemia Virus; Murine Leukaemia Viruses; Avian Leukosis Viruses; Avian Reticuloendo-theliosis Viruses; Murine Mammary Tumour Virus; Bovine Syncytial Virus; Lentiviruses; Equine Infectious Anaemia Virus; Visna/Maedi Virus; Caprine Arthritis-Encephalitis Virus: Jaagsiekte (Ovine Pulmonary Adellomatosis Virus).

x Textbook o/Velerinary Virology
30. Bunyaviridae 347 Rift Valley Fever Virus; Akabane Virus; Nairobi Sheep Disease.
31. Toroviridae 356 Breda Virus; Berne Virus.
32. Unclassified RNA Virus 360 Borna Disease Virus.
33. Unclassified Agents 362 Scrapie.
Index 364

Ads AEV AGID AHS AIBV ALV ASFV BAV BDV BEV BHV BLV BPV BPoV BRV BTV BVD CAEV CAM CCV CDV CE CF CHV CIE CK CM! CPE CPV ere
Abbreviations
adenoviruses avian encephalomyelitis virus agar gel immunodiffusion African horse sickness avian infectious bronchitis virus avian leukosis virus African swine fever virus bovine adenovirus border disease virus bovine ephemeral fever bovine herpesvirus bovine leukosis virus bovine papilloma virus bovine parvovirus bovine rhinovirus bluetongue virus bovine viral diarrhoea caprine arthritis-encephalitis virus chorio-allantoic membrane canine corona virus canine distemper virus contagious ecthyma complement fixation caprine herpesvirus counter immuno electrophoresis chicken kidney cell mediated immunity cytopathic effect canine parvovirus cytotoxic T cells

xii
CIL EAV EBV REV EHV EIAV ELISA EM F FAV FCV FIPV FMD FPLV H HA HI IBD IBH IBR ICH IF IFN ILT IP IPV mv JSRV Kb LD50 LCM MCF MDV MHV MVV N NI l'i1>V OAV PEV
cytotoxic T lymphocytes equine arteritis virus Epstein-Barr virus
Textbook of Veterinary Virology
equine encephalomyelitis virus equine herpes virus equine infectious anaemia virus enzyme-linked immunosorbent assay electron microscope/microscopy fusion protein fowl adenovirus feline calcivirus feline infcctious peritonitis virus foot-and-mouth disease feline panleucopenia virus haemagglutinin haemaggl utination haemagglutination inhibition infectious bursal disease inclusion body hapatitis infectious bovine rhinotracheitis infectious canine hepatitis immunofluorescence interferon infectious laryngotracheitis immunoperoxidase infectious pustular vulvovaginitis Japanese encephalitis virus jaagsiekte retrovirus kilobases 50 percent lethal dose lymphocytic choriomeningitis malignant catarrhal fever Marek's disease virus major histocompatibility complex maedi-visna virus neuraminidase neutralization index Newcastle disease ovine adenovirus porcine enterovirus

Abbreviations xiii
PI parainfluenza PPR peste-des-petits-ruminants PPV porcine parvovirus REV reticuloendotheliosis virus RIA radioimmunoassay RSV respiratory syncytial virus RVF Rift valley fever SN serum neutralization SPY sheep pox virus SV simian virus SVE swine vesicular exanthema TGE transmissible gastroenteritis UV ultraviolet VE vesicular exanthema VN virus neutralization VSV vesicular stomatitis virus

PART!
GENERAL VIROLOGY


Chapter 1
Structure and COlD position
The viral diseases of man and animals have been known for many centuries. The science of virology emerged during the last decade of last century. Ivanovski in 1892 reported that tobacco mosaic virus agent could pass through filters which retained bacteria. In 1898 Beijerinck showed that the tobacco mosaic disease agent differed fundamentally from toxin and it diffused through agar and he used the term 'contagium vivum fluidum' - that it was liquid or soluble. He also reported that only those plants which were growing and whose cells were dividing could be infected. The disease causing agent must be incorporated into the living protoplasm in order to propagate and it cannot multiply outside cells. Loeftler and Frosch in 1898 independently reported that foot and mouth disease of cattle could also be produced by a material passed through the filter which retained bacteria. Twort (1915) and d' Herelle (1917) recognised that bacteria also could be infected by filter passing agents.
Virology is now recognised as a basic biological science and veterinary virology has grown immensely during the past few decades. The subject of virology is divided into four main divisions -
i) Animal viruses - the viruses of man and animals. ii) Insect viruses - the viruses of insects and worms. iii) Bacterial viruses (Bacteriophages). iv) Plant viruses - viruses of plants. The real nature of viruses has been elucidated since 1930. Stanley
(1935) crystallized tobacco mosaic virus. Hershey and Chase (1952) discovered that only DNA of bacteriophage entered its bacterial host

4 Textbook of Veterinary Virology
and only DNA was necessary for infection. Fraenkel-Conrat (1956) proved that RNA of tobacco mosaic virus carried all the information for growth. Since then an enormous upsurge in our knowledge regarding the nature of viruses and its molecular biology has taken place.
Viruses have a very simple structure. The mature virus particle (Virion) consists of a central core of nucleic acid surrounded by protein coat. They vary in size ranging from 300 x 200nm of pox viruses to 20-25 nm of picornaviruses. Viruses can be distinguished from other unicellular microorganisms (Table 1.1). Lwoff and Toumier (1966) described the viruses having following five characters -
1.
2.
3. 4.
5.
6.
i) Possession of only onc type of nucleic acid, either DNA or RNA.
ii) Reproduction solely from nucleic acid, whereas other agents grow from the sum of their constituents and reproduce by division.
iii) Do not undergo binary fission. iv) Lack of genetic information for the synthesis of essential
cellular systems. v) Use of ribosomes of their host cells.
Table 1.1 IMPORTANf PROPERTIES OF UNICELLULAR ORGANISMS AND VIRUSES
Property Bacteria Mycoplasma Chlamydia Rickettsia Viruses
Nucleic DNA DNA and DNA and DNA and Either DNA acid (NA) and RNA RNA RNA or RNA, not
RNA both Nucleic Some DNA acid(NA) and RNA infectious viruses have
infectious NA
Ribol!omes + + + + Action of No No Inhibifs No Inhibits interferon action action growth action replication Metabolic + + ± + activity Binary + + + + fission

Structure and Composition 5
The criteria given above clearly distinguish viruses from other microorganisms; the most important criterion is that viruses contain only one type of nucleic acid. DNA or RNA and are completely dependent on the host cell for their reproduction. Some viruses may persist in their host cells by integration of their genome (DNA) or DNA CQPy of their RNA into the genome of host cell. The viruses are not '<lsceptible to antibiotics that act against specific steps in the metabolic pathways of bacteria.
Physical structure
Morphology: The size of virus particles range from about the size of smallest bacteria (300 nm) to about the size of largest protein molecules (20 nm). The unit of length is nanometre (nm) which is equal to 10-6 millimetres. For recording the size of very small structures Angstrom unit (A 0 or AV) is used. One nanometre is equal to 10 Angstrom units. The viruses occur in many shapes and sizes. The viruses were also known as 'ultrafiIterable viruses' or 'ultramicroscopic' since the viruses could pass the filters which retained bacteria and could not be seen under the light microscope. The viruses were measured by their capacity to pass through earthenware filters. The use of earthenware fillers was replaced by collodion or cellulose acetate membrane filters of gruded pore sizes. The membrane filters are non toxic to cells in culture and do not alter the pH of the medium and they do not adsorb large quantities of virus particles during filtration.
Another procedure for determining the size of viruses is high speed centrifugation. The rate of sedimentation 0f virus particles depend upon its size and the density and' viscosity of the suspending fluid. The relationship of sedimentation and size of virus particles IS governed by Stoke's law. During later half of 1930's and 1940's electron microscope made it possible to study the morphology and size of virus particles. In 1959 negative staining to electron microscopy of viruses transformed the knowledge of viral ultrastructure.
The simplest viruses consist of a single molecule of nucleic acid (DNA or RNA) enclosed within or built into protein coat. the capsid. The capsid and its enclosed nucleic acid constitute the nucleocapsirl (Fig. 1.1). The capsid is composed of morphological uniL~ called capsomeres. which are held together by nonconvalent bonds. The

6 Textbook ojVeterinary Virology
capsomeres consist of one or more molecules of polypeptidcs and are seen in the electron microscope. In some of the viruses there is an envelope of lipoprotein surrounding the nucleocapsid. The envelope is acquired as the virus passes through or buds from host cellular membrane~ and contains components of the host cell.
2 3 5
Fig. 1.1 Schematic Diagram of the Structure of a Virus
The assembly of capsomeres in a virion is defined by the nature of bonds formed between individual capsomeres, which imparts symmetry to the capsid. Two types of viral symmetry have been recognised, the cubical symmetry and helical symmetry; and these constitute an icosahedral capsid and a helical or tubular capsid respectively (Fig. 1.2). Some of the viruses have a combination of symmetries aand various structures. These are called complex viruses.
Fig. 1.2 Schematic Representation of the Structure of a Virus with Cubical (a) and Helical (b) Symmetry.

Structure and Composition 7
1. lcosahedral Symmetry: An icosahedron means a figure of 12 verticles (corners) and 20 faces each of an equilateral triangle. This indicates that the capsid be build of 60 (or multiples of 60) equivalent parts. Electron microscopic studies of icosahedral viruses show that these viruses have a regular surface array of morphologic units and the number is often greater than 60 but not generally a multiple of 60. The icosahedron has axes of 2. 3 and 5 fold rotational symmetry passing through its edges. faces and vertices. respectively (Fig. 1.3). The icosahedron is a strong structure which encloses a maximum volume. The total number of capsomeres can be calculated by the formula 10 (n-l)2+2, in which n is the number of morphologic' subunits between and including those on any five fold axes. The number of capsomeres (M) can also be calculated from the formula M = 10 T +2, in which T is the triangulation number. the number of small triangles formed on the single face of an icosahedron when each adjacent capsomere is connected by a line. In adenovirus Tt = 6 or T = 25. giving a total of 252 capsomeres. In adenovirus particles, capsomeres on the faces and edges bond to six neighbouring capsomeres and are called hexamers; those at the vertices bond to five neighbouring capsomeres are called pentomers. There are 240 hexamers and 12 pen tamers. Each penton has 1 fibre of filament in mammalian adenoviruses while there are 2 filaments in avian adenoviruses.
Capsid J "ucl . Nuel~oeapsid
~.J.....-;..--r- .. ..le acid
Co r~prot~in
Fig. 1.3 The lrosahedron. It h~ a Rotational Axes of Twofold, Threefold and F'ivefold Symmetry.
2. Helical or tubular symmetry: The nIJcleocapsids of ~ever?l RNA viruses have a helical symmetry, the capsomeres ~nd nuc1~lc aCid molecules self-assemble. as helix. The morphologiC subumts are arranged in their hmg axes In the nucleocapsid in a screwlike man.ner with several units per turn ()f the helix. The flcxuous hel.lcal nucleocapsid coil is always inside a glycolipoprotein envelope. poSSibly to give the very tong nuc!eocapsids st;lbility.

8 Textbook o/Veterinary Virology
Viral envelope: The envelope is derived at host cell membranes-plasma membrane, nuclear membrane, endoplasmic reticulum and Golgi complex during maturation by budding. The lipids of viral envelope are acquired from the cell while the proteins are virus coded. One kind of protein is glycoprotein peplomer (peplos means envelope) or spike whi~e the other kind of protein is nonglycosylated and is found on the inside of envelope of virions of several families and is knOW1l as matrix protein. The matrix protein gives rigidity to the viron morphology, e.g. the envelope of rhabdoviruses is closely attached to
the bullet shaped matrix protein that encloses nucleocapsid. In certain other viruses like arena viruses, bunyaviruses and corona viruses there is no matrix protein and are therefore more pleomorphic. The envelope is also possessed by certain icosahedral viruses like African swine fever virus, herpes viruses, toga viruses, flaviviruses and retroviruses. The envelope is immunogenic and is required for infectivity but in some poxviruses which have an envelope the infectivity is not dependent on the envelope.
3. Complex symmetry: Some viruses like poxviruses have a dumbell shaped core surrounded by complex membranes and other icosahedral or helical symmetry. The retroviruses have a tubular nucleocapsid surrounded by an icosahedral capsid enclosed within an envelope.
Chemical structure
The essential components of infectious virus particles are nucleic acid and protein. The enveloped viruses contain lipids and carbohydrates in their glycoprotein peplomeres. The complex viruses like pox viruses also contain lipids. The chemical composition of viruses can only be determined when the viruses are obtained in a pure form as the viruses contain host cell constituents. Most of the virus particles are attached to cell debris having almost similar chemical properties. Therefore, until 1950, without the introduction of more refined methods of purification, the animal viruses were not purified sufficiently for their chemical analysis. The other reasons of delay in purification of animal viruses was the small quantity of virus material available. With the introduction of tissue culture, it is now possible to
obtain sutlicient vIrUS for chemical analysis. In the purificaiton of viruses differential ultracentrifugation leads

Structure and Composition 9
to their considerable purification. The important technique introduced in 1950's is density gradient centrifugation where sucrose gradients ensure finer separation of particles with different sedimentation properties. Anotll.!r method which has proved of great value in purification of viruses is equilibrium sedimentation in caesium chloride and potassium tartrate which separate the particles according to their buoyant density. Density gradients of these salts are prepared and the mixture of virus and host cell debris is centrifuged in a high speed centrifuge. The different particles take positions in the gradient according to their buoyant density.
The viruses can also be separated from contaminating material by using fluorocarbons or other organic solvents; mild detergents to
remove host cell material selectively - especially for removing lipid material and denatured host protein. The enveloped viruses cannot be purified by detergents or lipid solvents because they are disrupted due to the action of these agents. The non enveloped viruses or naked viruses are stable in lipid solvents or even in strong detergents like sodium dodecyl sulphate.
1. Nucleic acid: Any particular virus contains either DNA or RNA which may be either single stranded or double stranded and the genome consists of either one molecule or several molecules. In most of DNA viruses the genome consists of a single molecule while several RNA viruses contain the genome of several molecules. The genome may be of linear or circular configuration. The nucleic acid of certain DNA or RNA viruses is infectious i.e. it can start multiplication cycle if introduced into susceptible cell. In such cases messanger RNA (mRNA) is transcribed from viral DNA in the nucleus by a cellular transcriptase, while in the case of RNA viruses the viral RNA itself acts as mRNA. In other virus families the extracted nucleic acid is not infectious. Among DNA viruses transcription requires viral rather than cellular transcriptase. Among RNA viruses when the viral RNA is of minus (-) sense or is double stranded its transcription to produce positive (+) sense in RNA requires a virion associated transcripitase which is separated from nucleic acid by extraction procedures. In the positive (-t) sense RNA viruses the viral RNA itself acts as its own mRNA. The positive (+) sense RNA of retroviruses is not infectious because replication of RNA occurs only after production of DNA provirus by a virion associated reverse transcriptase.

10 Textbook o/Veterinary Virology
The genome of all DNA viruses consists of a single molecule, which is double stranded except in parvovilUses. The genome may be linear or circular. The papovavirus DNA is a supercoilcd circle ·known as superhelix, when an enzyme nicks one of the strand the DNA molecule becomes a relaxed circle. The hepadnavirus genome is partially double stranded as one of the strand of circular DNA is shorter than the other. In case of linear DNA viruses during replication a temporary circular configuration is adopted. The molecular weight of DNA of different viruses varies from 1 to over 200 x 1 ()6.
The genome of RNA viruses may also be single or double stranded and in some viruses it is as single molecule while in others it is segmented. In arena virus and bimavirus, RNA consists of 2 segments, bunyavirus RNA is of 3 segements, orthomyxovirus RNA consists of 7 or 8 segments, and reovirus RNA is of 10, 11 or 12 segments. All viral RNA's are linear. The RNA of some of the viruses is said to have a positive (+) sense (also known as polarity), that is the RNA has same sense as mRNA. Picomaviruses, calciviruses, togaviruses, flaviviruses, coronaviruses and retroviruses have positive (+) sense genome. If the nucleotide sequence of the genome is complementary to mRNA it is said to have a negative (-) sense. The (-) sense genome is with paramyxoviruses, rhabdoviruses, arena viruses and bunyaviruses. All these viruses have an RNA dependent RNA polymerase (transcriptase) in the virion. In arena viruses and in one genus of bunyaviruses one of the RNA segment is ambisense, i.e. part (+) sense and part (-) sense. The molecular weight varies from 2 to 15 x 106 which is much less than seen in most DNA viruses.
Some viruses contain host cell nucleic acid, e.g. some papoviruses contain host cell DNA and arenaviruses contain cellular ribosomes. Sometimes several copies of viral genome may be enclosed in a single virus particle or the virus particles contain no nucleic acid, known as empty particles or contain incomplete genome known as defective interfering particles.
2. Proteins: The proteins make up 50-70% constituent of the virion. The virus coded proteins are structural, i.e. they form part of the virion, and non structural, i.e. proteins required during the replication cycle of virion. The structural proteins provide a protective coat to the viral genome. The vertebrate viruses contain sC'/eral proteins ranging from 4 distinct species of proteins in foot arid mouth disease VIruses to

Structure and Composition 11
over 100 in ease of poxviruses. There is normally one copy of viral nucleic acid in a virus particle but there arc many copies of each viral protein. Apart from providing protective shell to the viral genome the proteins have other properties as well.
Table 2.1 NATURE OF GENETIC MATERIAL OF VIRUSES
Family
Poxviridae Parvoviridae
Papovaviridae Adenoviridae Herpcsviridae Iridoviridae Hepadnaviridae Picomaviridae Ca1civiridae Togaviridae Flaviviridae Coronaviridae Rhabdoviridae Paramyxoviridae Orthomyxoviridae
Relroviridae Bunyaviridae Reoviridae
B irnav iridae
Structure of Nucleic acid
Linear ds DNA Linear as DNA (-) sense, a hairpin structure at onc end Circular supcrhelical ds DNA Linear ds DNA Linear ds DNA Linear ds DNA Circular ds DNA with ss region Linear ss RNA (+) sense Linear ss RNA (+) sense Linear ss RNA (+) sense Linear ss RNA (+) sense Linear ss RNA (+) sense Linear ss RNA (-) sense Linear ss RNA (-) sense Linear ss RNA (-) sense. Genome segmental, 7 or 8 molecules Linear ss RNA (+) sense, diploid genome ss RNA (-) sense, segmented genome, 3 molecules Linear ds RNA, segmented genome 10, 11, or 12 molecules. Linear ds RNA, segmented genome, 2 molecules
The surface proteins have an affinity for the specific receptors on the surface of susceptible cells and contain anti genic determinants which produce protec~ive antihody in the infected animal. Some of the virus proteins have an enzymic activity, e.g. a protein in (-) stranded RNA viruses acts as transcriptase. Glycoproteins make up the peplomers projecting from thee~velope. There is a second type of envelope protein which is nonglycosylated matrix protein that occurs as a layer at the inner surface of lipId envelope in orthomyxoviruses, paramyxoviruses and rhabdoviruses.

12 Textbook of Veterinary Virology
3. Lipid and carbohydrate: These constituents are found only in the envelope except complex viruses like poxviruses. Lipids and carbohydrates are derived from the host cells. Carbohydrate is the major part of glycoproteins of peplomers. Glycoproteins act as important antigenic determinants to which host immunity is directed.
References
FBNNER, FRANK, 1987. Veterinary Virology. Academic Press, New York.
FRED, BROWN, 1984. The nature of viruses. In Topley and Wilsons Principles of Bacteriology, Virology arul Immunity, Vol. 4. Williams and Wilkins, Baltomore.
LAUFfER, M.A.; BANG, F. B.; MARAMOROSCH, K., AND SMITH, K.M., 1982. Advances in virus research. Academic Press, New York.

Chapter 2
Classification of Viruses
The object of virus classification is to make a systematic ordered arrangement of viruses that have similarities and differences. Earlier efforts to classify viruses arranged them according to host symptoms or type of diseases and tissue affinities. This system had deficiencies e.g. the same virus produces different disease syndrome in different hosts, different strains of same virus can produce different syndromes in the same host and different viruses can produce the same clinical picture.
A classification based on epidemiological data was also tried. Enteric viruses: These viruses are acquired by ingestion and
replicate primarily in the digestive tract. The important enteric viruses include rotaviruses, coronaviruses, enteroviruses and adenoviruses.
Respiratory viruses: These viruses enter the host by inhalation and replicate in the respiratory tract. These viru~s include orthomyxoviruses. rhinoviruses, paramyxoviruses, adenoviruses and coronaviruses.
Arboviruses: Arthropod bovine viruses infect arthropods and ingest vertebrate blood. These viruses replicate in arthropod host and are transmitted to a vertebrate host by bite. These viruses replicate also in verteblate host These include orbiviruses. bunyaviruses, flaviviruses, togaviruses. rhabdoviruses and African swine fever virus.
The viruses classified on epidemiological data comprise viruses belonging to different families with different physical and chemical properties. Therefore, the most important criteria for classification are the physical and chemical characteristics of the virion and its mode of replication. The criteria for classification into different families are-

14 Textbook o/Veterinary Virology
1. the kind of nucleic acid e.g. single or double stranded DNA or RNA and mode of replication; 2. morphology of the virion including its size, shape, nucleocapsid symmetry and presence or absence of nucleocapsid, number of capsomeres and pH sensitivity.
The criterion of subdivision of families is controversial. Most virologists agree that viruses should differ substantially in nucleic acid sequence to be designated as different species but there is yet no agreement on how such differences should be quantitatcd. Monoclonal antibodies are of great value in the differentiation of viruses at species level and below. There are other techniques being used for studying the composition of viral nucelic acid to identify species and to understand minor differences in viral s..rains. The techniques being employed are molecular hybridization, oligonucleotide finger printing, clectrophoresi~ in gels and nucleotide sequence analysis.
The International Committee on Taxonomy of viruses (ICTV) has recommended that the highest taxonomic group is the family, and is named with a suffix-viridae. Subfamilies have a suffix-virinae and genera with a suffix-virus. Latinized names for families, subfamilies and generic names are written in italics and vernacular names derived from them are written in roman letters. It is still customary to use vernacular terms rather than latinized binomials for viral species e.g. Newcastle disease virus.
There are still viruses affet::ting man and animals which are still unclassified but majority of these viruses known to man have been assigned to one or another of the 20 families (Table 2.1). A brief description of each family or vertebrate viruses is given below:
DNA viruses
Poxviridae: Pock means a pustule or ulcer. These are complex large, brick shaped or ovoid virus particles measuring 300-450 x 170-260 nm in diameter. The virions have an envelope containing lipid and tubular and globular protein structures. All have an inner core which contains a single linear molecule of double stranded (ds) DNA; 130-280 Kbp. There are more than 30 structural proteins and several enzymes aSSOCIated with the virus particle apart from 4 percent lipid and 3 percent carbohydrate. The pox viruses replicate in the cytoplasm, mRNA is transcnbed by a virion-associated transcriptase. Mature particles are released from microvilli or by cellular disruption.

Classification o/Viruses 15
The family is divided into two subfamilies-1. Chordopoxvirinae. comprises of vertebrate viruses and is
divided into 6 genera, which include animal pathogens. 2. Entompoxvirinae. comprises of insect viruses. Parvoviridae: Parvoviruses are small about 20 nm in diameter,
have icosahcdral symmetry with 32 capsomeres. The genome is a single stranded (ss) DNA with molecular weight 1.5-2.2 x Ht. The virions are heat stable. The family comprises of 3 genera and members of2 genera affect the vertebrate hosts. Genus parvovirus include animal pathogens, while genus dependovirus includes defective viruses which depend on adenovirus for replication. They occur in birds, animals and human beings but arc not pathogenic. Replication takes place in nucleus.
Papol'Qviridae: Pa stands for papilloma; po for polyoma and va for vacuolating agent. These are noneveloped icosahedral with a diameter of 45-55 nm. The genome is a single cyclic molecule of double stranded DNA with molecular weight 3-5 x 106• The replication takes place in the nucleus. There are two genera papillomavirus and polyomavirus. Most species arc oncogenic.
At/ellm'iridae: Adeno means glands. The virions arc noneveloped with icosahedral symmetry, 70-90 nm in diameter with 252 capsomeres. Vertex capsomeres arc distinct from others and carry 1-2 filamentous projections. The genome is a single linear molecule of double stranded DNA with molecular weight 20-30 x 106•
Adenoviruses replicate in the nucleus. The viruses arc usually associated with respiratory and intestinal infections and sometimes with eye infection. Many viruses arc conditionally oncogenic. There are two genera in this family 1. Mastadenovirus, these arc mammalian pathogens and 2. A viadenovirus, pathogenic for birds. A common antigen is shared by all mammalian strains which differ from corresponding antigen of avian strains.
Herpesviridae: Herpes means creeping. The herpes viruses are enveloped 120-150 nm in diameter with icosahedral symmetry and 162 capfoomeres. The virion consists of a core in which genome is wrapped, the icosah~dral nucleocapsid, a tegument surrounding the capsid and an envelope. The genome is (ds) DNA with a moleuclar weight 80-150 x 106. The multiplication takes place in the nucleus and virion mature by the addition of glycoprotein lipid membrane as the virus passes through

16 Textbook o/Veterinary Virology
the inner lamellae of nuclear membrane into endoplasmic reticulum. The family has been divided into 3 subfamilies 1. Alphaherpesvirinae- which include infectious bovine rhinotracheitis virus, bovine mammallitis virus, B virus, pseudorabies virus, equine rhi~opneumonitis and equine coital exanthema virus, viruses of dogs, cats and chickens. 2. Betaherpesvirinae-includes cytorr.egaloviruses of man and animals. 3. Gammaherpesvirinaeincludes viruses associated with tumors like Marek's disease virus of chickens.
The herpes viruses produce lifelong persistant infections usually in the latent form. The excretion of virus from the host may be continuous or intermittent without disease or episodes of recurrent clinical disease.
Iridoviridae: Irido means shining, iridescent. The viruses in this family are icosahedral enveloped viruses measuring from 130-300 nm in diameter. The genome is a single linear (ds) DNA with a molecular weight of 130-160 x 1()6. The multiplication occurs in the cytoplasm using virion associated transcriptase but nuclear involvement is needed for viral DNA synthesis.
Hepadnaviridae: Hepa means liver and dna. It includes human hepatitis B virus. The viruses are spherical particles with 42 nm in diameter, consist of an icosahedral core which is 27 nm. The genome is circular partially double stranded DNA molecule, which consists of a long and short strand. The multiplication takes place in nucleus of heptocytes.
Families of RNA viruses
Picornaviridae: The name picorna is originally derived from poliovirus, insensitivity to ether, coxsackievirus, orphan virus, rhinovirus and ribonucleic acid omitting one ('r'), but also consistent with pico which means small and rna is ribonucleic acid. The viruses are small noneveloped, icosahedral, 25-30 nm in diameter.The genome is 1 piece linear single stranded (+) sense RNA with a molecular weight about 2.5 x 106• The viruses replicate in the cytoplasm and functional proteins are mainly produced by post-translational cleavage. The family comprises of 4 genera 1. Enterovirus, these are stable at pH 3, include poliovirus and large number of species that affect domestic animals; 2. Rhinovirus, virus is unstable at pH 3; 3. Cardiovirus, comprises the viruses of encephalomyelocarditis of swine and rodents; 4. Aphthovirus comprises of foot and and mouth disease virus.
Calciviridae: Calix means cup. The calciviruses are noneveloped,

Classification of Viruses 17
IcosahedIal with 32 capsomeres and measure about 35-40 nm if. diameter. The genome is ss RNA in 1 piece. The family comprises of one genus- Cl)lcivirus which includes vesicular exanthema of swine.
Togaviridae: Toga means cloak. These are small spherical enveloped viruses, measuring about 60-70 nm in diameter. The single molecular genome is single stranded RNA with a molecular weight 4 x 1()6 a!ld is (+) sense. The viruses multiply in the cytoplasm and mawre by budding from cell membranes. The arthropod borne viruses multiply in arthropods as well as in vertebrates. The family comprises of 4 genera-Alphavirus, Rubivirus, Pestivirus and Arterivirus.
FlavivirUtae: F1avi means yellow. The viruses included in this family were formerly classified in togaviridae but now placed in new family-Flaviviridae, mainly because of different strategy of iephcation but in other respects the flaviviruses resemble the alpha virus genus. These viruses are smaller measuring about 40-50 nm in diameter. The family comprises of only one genus-Flavivirus.
Reoviridae: The family name is a single, respiratory enteric orphan virus. The viruses are nonenveloped, icosahedral wilh a diameter of 60-80 nm with two protein coats. The genome is ds RNA in 10 to 12 pieces with a molecular weight of 10-16 x 1()6. The viruses multiply in the cytoplasm. There are 3 genera in the family causing infections of veterinary importance.
BirnavirUtae: Sigla bi-two and rna. The family comprises of ic.osahedral symmetry and measure about 60 nm in diameter. The genome is ds RNA which is linear and is in two segments. There is only one genus in the family-Birnavirus, which includes the important poultry virus of chickens, infectious bursal disease virus.
4renaviridae: Areana means sand. Arenaviruses acqui:'ed their names because of presence of ribosomes incorporated within pleomorphic enveloped virions measuring about 50-300 nm in diameter. The genome consists of two pieces of (-) sense ss RNA.
Coronaviridae: Corona means crown. The virions are enveloped, pleomorphic with 75-160 nm in diameter. There are widely spaced, pear shaped peplomeres in the lipoprotein envelope. The envelope lacks a matrix protein and encloses a core of undetermined symmetry. The genome is a single molecule of (+) sense ss RNA with a molecular weight 5.5-6.1 x 1()6. The family comprises of only one genus which includes important poUltry and other viruses of domesticated animals.

18 Textbook of Veterinary Virology
Orthomyxoviridae: Myxo means mucus. The viruses are enveloped, pleomorphic particles with 80--120 nm in diameter. The nucleocapsid has a helical symmetry. The envelope has surface projections of two types, a haemagglutinin and a neuraminidase. The genome consists of eight segments of (-) sense ss RNA with a molecular weight of 5 x 106 and is associated with viral transcriptase. The family consists of one genus-influenza virus. Two important species of viruses influenza A and influenza B are included in this family.
Paramyxoviridae: The viruses are pleomorphic, enveloped, usually spherical measuring about 150 nm in diameter with a helical nucleocapsid symmetry. The genome consists of a single molecule of (-) sense ss RNA with a molecular weight of 5-7 x 106. The virion contains a transcriptase. The envelope contains two glycoproteins, haemalglutinin and in some species with neuraminidase activity, and fusion protein. The family is subdivided into 3 gencra. Paramyxovirus, Morbillivirus and Pneumovirus. The family includ~s the viruses of important pathogens of veterinary importancc.
Rhabdoviridae: Rhabdo means rod. The virus particles are enveloped, bullet shaped or bacilliform measuring about 180 x 75 nm. The capsid has a helical symmetry which is closely attached to
lipoprotein envelope, with surface projections. The genome is a single molecule of H sense ss RNA with a molecular weight of 3.5-4.5 x 106 with transcriptase. The virus family comprises of two generaVesiculovirus and Lyssavirus besides ungrouped rhabdoviruses causing disease in animals like bovine ephemeral fever virus.
Retrm'iridae: Re stands for reverse and tr stands for tmnscriptase. This is a large family of enveloped viruses with icosahedral core containing a helical nucleoprotein. There is reverse transcriptase within the virion. The genome is diploid consisting of inverted dimer of (+) sense ss RNA with a molecular weight of 3 x 106 (for one monomer). The ds DNA of copy of genome of the virus is transcribed by the viral reverse transcriptase and is integrated into the cellular DNA as an essential part of replicate cycle. Proviral DNA is found in the DNA of all normal cells of many species of animals and may produce virus under certain circumstances. These are known as endogenous retroviruses. The exogenous retroviruses are transmitted

Classification of Viruses 19
horizontally. The family is subdivided into 3 subfamilies, two of which contain pathogens of veterinary importance.
Bunyaviridae: Buyamwera is a locality in Africa. These are oval or spherical particles measuring about 90-120 om in diameter with a nucleocapsid of helical symmetry. The genome is of 3 segments of circular (-) sense ss RNA with a molecular weight of 3.2 and 0.5 x 1()6. They replicate In-the cytoplasm and bud from Golgi body membranes. Due to segmented genome the viruses readily undergo genetic resortment and may produce antigenic shift. The family comprises of 5 genera.
Other viruses: A family Filoviridae has been proposed. There are other viruses which are yet to be classified e.g. toro viruses , astroviruses, the unclassified virus of Borna disease and mysterious agents, which cause the subacute spongiform encephalopathies which include scrapie.
Filoviridae: The viruses included in this family are Marburg virus and Ebola virus which resemble rhabdoviruses but the viruses are pleomorphic and sometimcs very long. The genome is a single molecule (-) sense ss RAN.
Toroviruses: Torus stands for an object like donut. The viruses are aS30ciated with diarrhoea in horsces and calves. The viruses are enveloped, disk shaped measuring 35 x 170 nm and contain a nucleocapsid, probably of helical symmetry. The genome is a single molecule of (+) sense ss RNA.
Astroviruses: Astro means star. The viruses are star shaped and found in the faeces of calves, lambs and humans. The genome consists of one molecule of ss RNA.
References
BROWN, F. 1986. The classification and nomenclature of viruses. Summary of results of meetings of the International Committee on Taxonomy of viruses in Sendai, Sept. 1984, Intervirology 25, 141.
MATHEWS, R.E.F. 1983. Classification and nomenclature of viruses. Fourth report of the International Committee on Taxonomy of viruses. Intervirology 12,1.

Family
1
DNA VIRUSES: Poxvlrldae Sub-family
Genus
2
Species
3
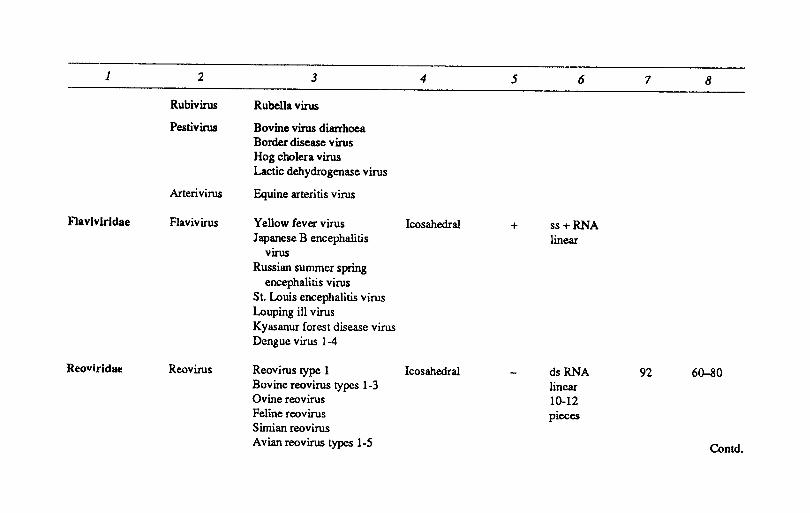
Table 2.2 CUSSIflCATION OF ANIMAL VIRUSES
Symml!lry o/capsid
4
Presence o/DlVelope
5
i. Chordopox- Orthopoxvirus Variola virus Complex + virinae (Vertebrates)
Vaccinia virus Cowpox virus Camel pox virus Ectromelia virus Buffalo pox virus Horse pox virus Monkey pox virus Rabbitpox virus
Capripoxvirus Sheep pox virus Goat pox virus Lumpy skin disease
Leporipoxvirus Myxoma virus Rabbit fibroma virus Squirrel fibroma virus
Nucleic acid
6
dsDNA associated with virion transcriptase
No. 0/ Particle capsomues diameter
(nm)
7 8
300-450 x
170-260

1 2 3 4 5 6 7 8
Parapoxvirus Contagious ecthyma virus (Orfvirus)
Pseudocowpox virus Bovine papular stomatitis
virus
Suipoxvirus Swine pOx virus Yaba monkey tumorpox
virus
Avipoxvirus Fowl pox virus Canary pox virus Pigeon pox virus Sparrow pox virus Starling pox virus Junco pox virus Quail pox virus Turkey pox virus
ii. Entomopox Insect viruses virinae
Parvovlridae Parvovirus Kilham rat virus Icosahcdral ss +Of- 32 18-26 Bovine parvo virus DNA Porcine parvo virus
Contd. Canine parvo virus

1 2 3 4 5 6 7 8
Feline panleukopenia Aleutian mink disease virus Mink enteritis virus Goose parvovirus
Dependovirus Adeno associated virus (AA V) type 1 human
BovineAAV EquineAAV CanineAAV OvineAAV Sirnia., AA V AvianAAV
Densovirus Insect parvoviruses
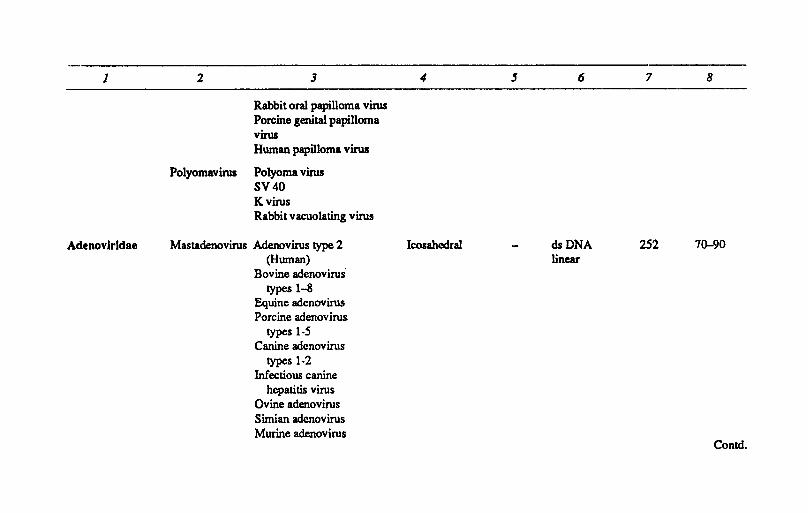
Papovarlridae Papillomavirus Cotton tail rabbit Icosahedral dsDNA 72 45-55 Papilloma virus (Circular) Bovine papilloma virus
type-1-6 Equine papilloma virus Canine oral papilloma virus Sheep papilloma virus Goat papilloma virus Deer papilloma virus Comd.
1 2 3 4 5 6 7 8
Rabbit oral papilloma virus Porcine genital papilloma virus Human papilloma virus
Polyomavirus Polyoma virus SV40 K virus Rabbit vacuolating virus
Adenovirldae Mastadenovirus Adenovirus type 2 Icosahedral dsDNA 252 7~90 (Human) linear
Bovine adenovirus types 1-8
Equine adenovirus Porcine adenovirus
types 1·5 Canine adenovirus
types 1·2 Infectious canine
hepatitis virus Ovine adenovirus Simian adenovirus Murine adenovirus
Contd.
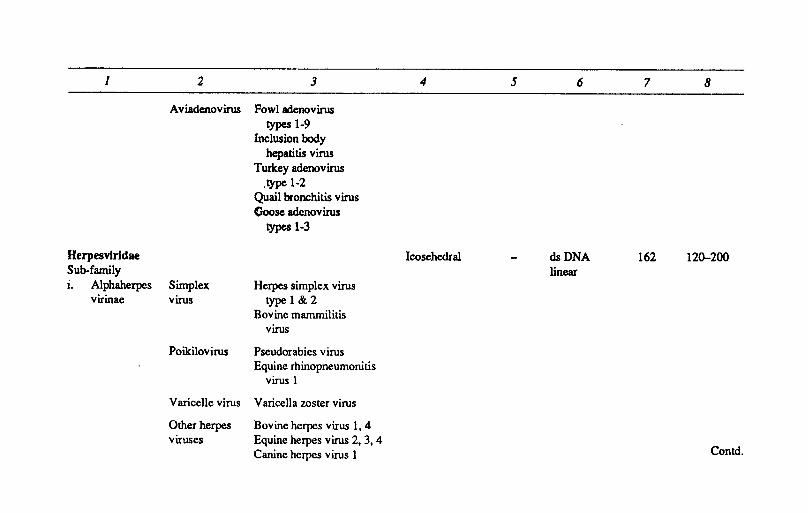
1 2 3 4 5 6 7 8
Aviadenovirus Fowl adenovirus types 1-9
Inclusion body hepatitis virus
Turkey adenovirus .type 1-2
Quail bronchitis virus (;oose adenovirus
types 1-3
Herpesvlridae Icosehedral dsDNA 162 12~200
Sub-family linear 1. Alphaherpes Simplex Herpes simplex virus
virinae virus type I &2 Bovine mammilitis
virus
Poikilovirus Pseudorabies virus Equine rhinopneumonitis
virus 1
Varicelle virus Varicella zoster virus
Other herpes Bovine herpes virus 1,4 viruses Equine herpes virus 2, 3, 4
Canine herpes virus 1 Contd.

1 2 3 4 5 6 7 8
Porcine herpes viruses Feline herpes virus 1 Ovine herpes virus Caprine herpes virus Simian herpes virus Fowl herpes virus Duck herpes virus Pigeon herpes virus
ii. Betaherpes Cytomegalo- Cytomegalovirus of multiple virinae virus animal species
Muromeglo- Cytomegalovirus of mice virus
Ul. Gammaherpes Lymphocrypto Epstein-barr virus virinae virus
Theta Marek's disease virus lymphcrypto Turkey herpes virus virus Ateline herpes virus-3
Saiminine herpes virus 2
Iridoviridae Iridovirus Insects viruses Icosahedral + dsDNA 1892-2172 200-220 Chloriridov irus
ConlrJ.
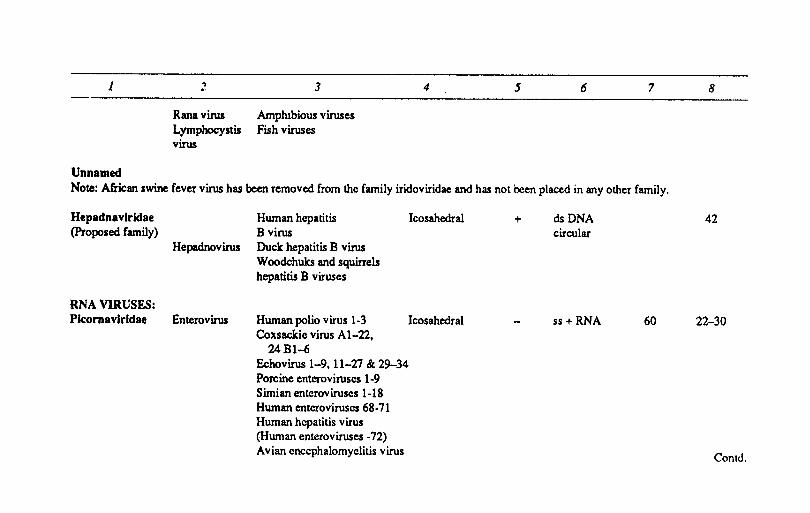
1 ~ 3 4 5 6 7
Rana virus Amphibious viruses Lymphocystis Fish viruses virus
Unnamed Note: African swine fever virus has been removed from the family iridoviridae and has not been placed in any other family.
Hepadnavlrklae (Proposed family)
RNA VIRUSES: Picomavlrldae
Hepadnovirus
Enterovirus
Human hepatitis B virus Duck hepatitis B virus Woodchuks and squirrels hepatitis B viruses
Icosahedral
Human polio virus 1-3 Icosahedral Coxsackie virus AI-22,
24BI-6 Echovirus 1-9, 11-21 & 29-34 Porcine enteroviruses 1-9 Simian enteroviruscs 1-18 Human enteroviruscs 68-11 Human hepatitis virus (Human enteroviruses -12) Avian encephalomyelitis virus
+ dsDNA circular
ss+ RNA 60
8
42
22-30
Contd.
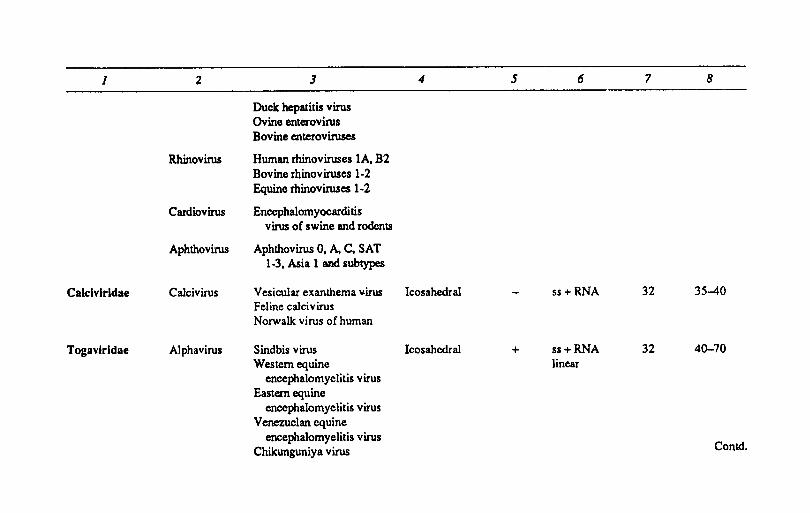
1 2 3 4 5 6 7 8
Duck hepatitis virus Ovine enterovirus Bovine enteroviruses
Rhinovirus Human rhinoviruses lA, B2 Bovine rhinoviruses 1-2 Equine rhinoviruses 1-2
Cardiovirus Encephalomyocarditis virus of swine and rodents
Aphthovirus AphthovirusO, A,C, SAT 1-3. Asia 1 and subtypes
Calclvlrldae Calcivirus Vesicular exanthema ~irus Icosahedral ss + RNA 32 35-40 Feline caJ-civirus Norwalk virus of human
Togav-irldae Alphavirus Sindbis virus Icosahcdral + ss+RNA 32 40-70 Western equine linear
encephalomyelitis virus Eastern equine
encephalomyelitis virus Venezuelan equine
encephalomyelitis virus Conk!. Chikunguniya virus
1 2 3 4 5 6 7 8
Rubivirus Rubella virus
Pestivirus Bovine virus diarrhoea Border disease virus Hog cholera virus Lactic dehydrogenase virus
Arterivirus Equine arteritis virus
Flaviviridae Flavivirus Yellow fever virus Icosahedral + ss+RNA Japanese B encephalitis linear
virus Russian summer spring
encephalitis virus St. Louis encephalitis virus Louping ill virus Kyasanur forest disease virus Dengue virus 1-4
Reovlridae Reovirus Reovirus type 1 Icosahedral dsRNA 92 60-80 Bovine reovirus types 1-3 linear Ovine reovirus 10-12 Feline reovirus pieces Simian reovirus Avian reovirus types 1-5 Contd.

1 2 3 4 5 6 7 8
Orbivirus Blue tongue virus types 1-24 African horse sickness virus 1-9 Ibraki virus Epizootic haemorrhagic
disease virus of deer
Rotavirus Nebraska calf diarrhoeal virus Foal gastroenteritis virus Piglet gastroenteritis virus Simian rotavirus Ovine rotavirus Caprine rotavirus Feline rotavirus Avian rotavirus
Birnavlridae Bimavirus Infectious bursal Icosahedral dsRNA 32 60 disease virus (segmented 2)
Infectious pancreatic necrosis virus of fish
Arenavlrldae Arenavirus Lymphocytic choriomen- Helical + ss RNA Ambiscnse 50-300 ingitis virus (segmented 2)
Lassa virus linear or circular
Contd.

1 2 3 4 5 6 7 8
Coronavlrldae Coronavirus Avian infectious Helical + ss+RNA 75-160 bronchitis virus linear
Bovine coronavirus diarrhoea
Canine coronavirus diarrhoea
Feline infectious peritonitis virus
Equine coronavirus diarrhoea
Porcine haemagglutinating encephlomyelitis virus
Turkey blue comb virus
Orthomyxoviridae Influenza Influenza virus Helical + ss -RNA 80-120 virus Equine influenza virus (segmented
1 and 2 8) Swine influenza virus Influenza virus B Influenza virus C Avian influenza virus Turkey influenza virus Duck influenza virus Unnamed Contd.

1 2 3 4 5 6 7 8
Paramyxoviridae Paramyxovirus New castle disease virus Helical + ss-RNA about Mumps virus 150-300 Bovine parainfluenza 3 (Pleomorphic)
virus Equine parainfluenza 3
virus Canine parainfluenza 2
virus Avian parainfluenza 2
virus
Morbillivirus Measles virus Rinderpest viru~ Canine distemper virus
Pneumovirus Respiratory syncytial virus Bovine and ovine respiratory
syncytial virus Pneumonia virus of mice.
Rhabdoviridae Vesiculovirus Vesicular stomatitis virus Helical + ss -RNA 180x75 Lyssavirus Rabies virus
Mokola virus Two unnamed Bovine ephemeral sub groups fever virus
Contd. Fish rhabdoviruses
1 2 3 4 5 6 7 8
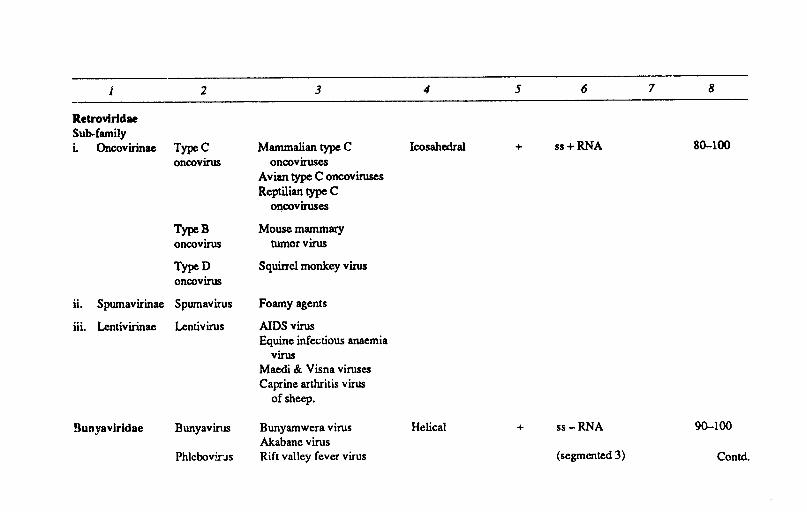
Retrovlrldae Sub-family i. Oncovirinae TypeC Mammalian type C Icosahedral + ss+RNA 80-100
oncovirus oncoviruses Avian type C oncoviruses Reptilian type C
ollcoviruses
TypeB Mouse mammary oncovirus tumorvirus
TypeD Squirrcl monkey virus oncovirus
ii. Spumavirinae Spumavirus Foamy agents
iii. Lentivirinae Lentivirus AIDS virus Equine infectious anaemia
virus Maedi & Visna viruses Caprine arthritis virus
of sheep.
Bunyavlrldae Bunyavirus Bunyamwera virus Helical + ss-RNA 90-100 Akabane virus
Phlcbovins Rift valley fever virus (segmented 3) Contd.
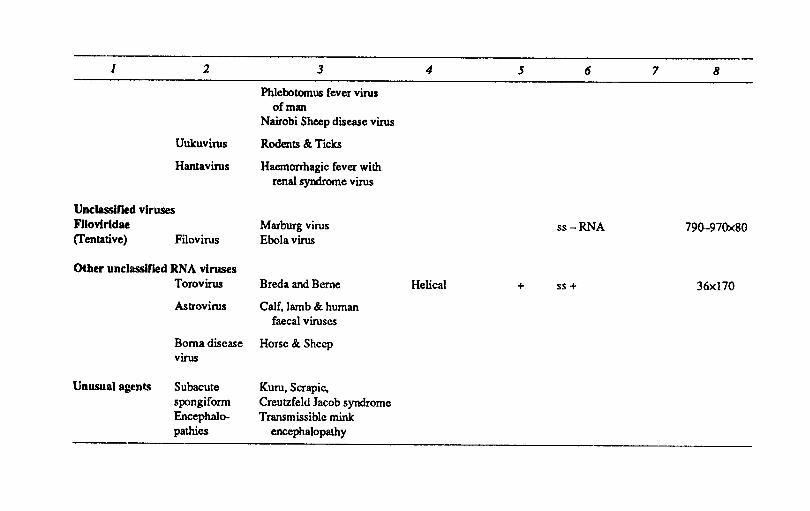
1 2 3 4 5 6 7 8
Phlebotomus fever virus of man
Nairobi Sheep disease virus
Uukuvirus Rodents & Ticks
Hamavirus Haemorrhagic fever with renal syndrome virus
Unclassified viruses Fllovlrldae Marburg virus ss-RNA 79Q,-97Ox80 (Tentative) Filovirus Ebola virus
Other unclassified RNA viruses Torovirus Breda and Berne Helical + ss + 36x170
Astrovirus Calf. lamb & human faecal viruses
Borna disease Horse & Sheep virus
Unusual agents Subacute Kuru. Scrapie. spongiform Creutzfeld Jacob syndrome Encephalo- Transmissible mink pathies encephalopathy
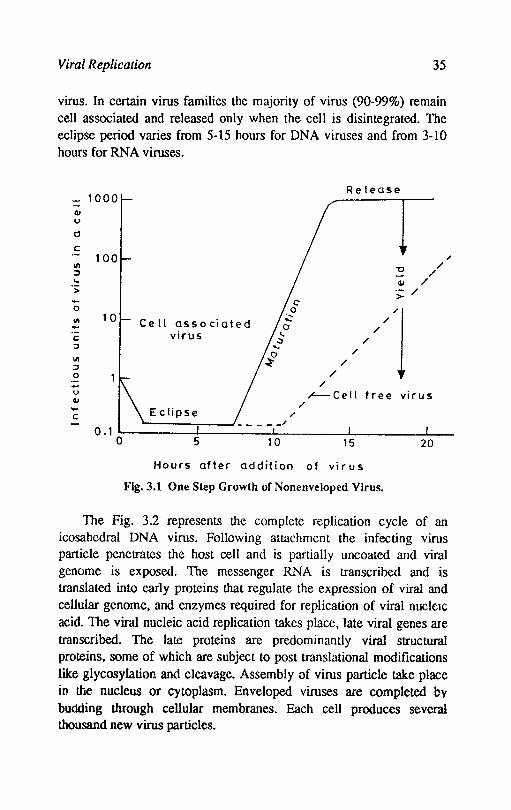
Viral Replication 35
virus. In certain virus families the majority of virus (90-99%) remain cell associated and released only when the cell is disintegrated. The eclipse period varies from 5-15 hours for DNA viruses and from 3-10 hours for RNA viruses.
Release - 1000
I -QI u
0
c 100
III U / :::I / ... QI / > :;: / .... 0 //'1 III 10 Ce II associated -c; virus :I
III :I 0 - / u ~Cell free virus QI /
c Eclipse / - 0.1 0 5 10 15 20
Hours after addition of vi r us
Fig.3.1 One Step Growth of Nonenveloped Virus.
/
The Fig. 3.2 represents the complete replication cycle of an icosahcdral DNA virus. Following attachment the infecting virus particle penetrates the host cell and is partially uncoated and viral genome is exposed. The messenger RNA is transcribed and is translated into early proteins that regulate the expression of viral and cellular genome, and enzymes required for replication of viral nucleIc acid. The viral nucleic acid replication takes place, late viral genes are transcribed. The late proteins are predominantly viral structural proteins, some of which are subject to post translational modifications like glycosylation and cleavage. Assembly of virus particle take place in the· nucleus or cytoplasm. Enveloped viruses are completed by budding through cellular membranes. Each cell produces several thousand new virus particles.

36 Textbook o/Veterinary Virology
Cell membrane
Cytoplasm
Virus
I ~ N,,,.ooaP'id@ +~
/. ~ Neuraminidase
Haemagglutinin
Ribosome
Fig. 3.2 Schematic Representation of Influenza Virus Reproduction 1 and 2. Penteration or Virion to Cell; 3. Release or Ribonucleoprotein (RNP); 4. Entry or RNP into Cell Nucleus; S. Replication or Viral RNA; 6. Viral RNP; 7. Synthesis or Viral Proteins in the Ribosomes; 8. Newly Formed Viral Protein; 9. Formation or Virion; 10. Virion Leaves the Cell by Budding.
Adsorption
The initial interactions between viruses and cells occur randomly, although both are negatively charged and repel each other, this attachment is reversible which is facilitated by cations and initial concentration of infectious virus particles and cells. For firm binding the virus must have surface capsid proteins or envelope spikes which bind to specific receptor sites on cell membrane. ThIS specificity often explains the host range of viruses. In caSe of myxoviruses the binding is via haemagglutinin, an envelope glycoprotein, to glycoprotein or glycolipid cellular receptors with oligosaccharide chains terminating in N-acetylneuraminic acid. In picoma viruses, poliovirus binds only f.(I
human or primate cells, because only the homologous cells carry

Viral Replication 31
receptoIS which the relevant viral capsid protein attachment site can bind. There is some specificity about the binding of virions to particular cellular receptors; several different viruses may utilise the same receptor.
Penetration and uncoating
The recent studies have shown that virions can enter the cells by endocytosis, fusion and translocation. The majority of virions entering the cell fail to set up infection because they are degraded by lysosomal enzymes.
Endocytosis: Adsorbed virus is incoporated into endosomes which are cytoplasmic vacuoles. Following attachment to receptors the virus particles move down into coated pits, coated with clathrin, fold inwards to produce coated vesicles that enter the cytoplasm and fuse with lysosome and form a phagolysosome. In case of enveloped viruses the envelope of endocytosed virion fuses with lysosomal membrane, releasing the viral nucleocapsid into the cytoplasm.
Fusion: The fusion glycoprotein of paramyxoviruses enables the envelope of these viruses to fuse directly with the plasma membrane. This way the nucleocapsid is released directly into the cytoplasm.
Translocation: Certain noneveloped viruses are capable of passing directly through the plasma membrane into the cytoplasm.
The uncoating of those viruses which enter by fusion, their nucleocapsid is discharged directly into the cytoplasm. In case of viruses with helical nucleocapsids, the transcription, begins from viral RNA while it is still associated with nucleoprotien. In icosahedral reoviruses the nucleic acid viral genome expresses while it is still covered with protein membranes, only certain capsid proteins are removed and it is not fully exposed from the core. The pox viruses are uncoated in 2 stages, fIrstly upto core from which half genome is transcribed, then completely following the synthesis of virus coded uncoating protein. In case of picoma viruses, cQnformational changes take place during the process of attachment. This results in the loss of capsid proteins and the virion becomes susceptible to proteases. For viruses which replicate in the nucleus, there is evidence that later stages of uncoating take place in nucleus.
Viral synthesis
The naked viral genome codes for messenger RNA to produce

38 Textbook o/Veterinary Virology
virus proteins on cellular ribosomes. The viral nucleic acid also codes for new viral nucleic acid. The new viral nucleic acid associates with capsid proteins to make nucleocapsid. In enveloped viruses additional viral envelope glycoproteins become associated with host cell membranes.
Messenger RNA production (Transcription): In Case of DNA viruses which replicate in the nucleus, the cellular dependent RNA polymerase II performs the function of transcription. In other viruses, a virus coded and integrated component of the virus particle performs this function, cytoplasmic ds DNA viruses carry a DNA dependent RNA polymerase while ds RNA viruses have a ds RNA dependent RNA polymerase. The (-) sense ss RNA viruses carry a ss RNA dependent RNA polymerase. The viral RNA of (+) sense RNA viruses binds directly to ribosomes and is translated.
a. DNA viruses: Particular part of genome is transcribed in sequence, early genes ftrst and late genes later. The strategy in different DNA viruses is as under:
i) ds DNA, cellular transcriptase: In papovaviruses, adenoviruses and herpesviruses, the viral DNA is transcribed within the nucleus by a cellular-dependent RNA polymerase. In adeno and herpes viruses there are at least2 cycles and in each cycle the structural proteins of the virus particles are made from mRNAs produced in last cycle of transcription. Polycistronic RNA transcripts under cleavage and splicing to produce monocistronic mRNAs.
ii) ds DNA, virion transcriptase: In case of pox viruses and African swine fever virus which replicate in the cytoplasm, carry their own transcriptase. The monocistronic mRNAs are directly transcribed from viral DNA. The transcripts are translated directly into proteins in 3 cycles of transcription.
iiy ss DNA, cellular transcriptase: The (:..) sense ss DNA (parvoviruses) requires the synthesis of a complementary strand to form ds DNA. This transcription takes place in the nucleus and the transcripts are processed to produce mRNA's before being exported to the cytoplasm for translation.
iv) dslss DNA, cellular transcriptase, virion DNA po1ymerases: The ss DNA of the genome of hepadnaviruses is ftrst repaired by virion associated DNA polymerase and then DNA is converted into

Viral Replication 39
supercoiled ds DNA. Transcription of mRNA then takes place by cellular RNA polymerase 11.
b. RNA viruses: The transcription of RNA viruses is more complicated:
i) ss (+) sense RNA: The (+) sense ss RNA is itself infectious. In picoma' and flaviviruses the genome acts as a single polycistronic mRNA which is translated into a single polyprotein which is subsequently cleaved to give individual viral polypeptides. In alphavirus genus of Togaviridae which contains (+) sense ss RNA molecule, only about two thirds of viral RNA at 5' end is translated. In Coronaviruses part of virion RNA acts as mRNA and is translated to produce a RNA polymerase which then synthesizes genome length (-) sense strand. From this overlapping subgenomic RNA is transcribed, of which only the nonoverlapping sequence is translated.
ii) ss RNA (-) sense, virion transcriptase: In paramyxo and rhabdoviruses the (-) sense virion RNA is copied in two distinct ways, the replication mode and transcription mode. Copying in the replication mode produces a full length (+) sense strand which acts as a template for the synthesis of new virion RNA. In transcription mode, 5 subgenomic (+) sense RNA's are produced and each serves as a moncistronic mRNA. In orthomyxoviruses, btlnyaviruses and arenaviruses the genome is segmented. Each segment is transcribed to yield a mRNA which is translated into one or more proteins. In orthomyxoviruses transcription of 8 RNA's occur in the nucleus called as 'cap snatching'. A virion associated endonuclease enters the nucleus and removes a short segment from the capped 5 terminus of cell mRNA, this is transported to the cytoplasm where it binds to the virion RNA and serves as a primer to initiate transcription.
iii) ds RNA, virion transcriptase: In reoviridae and birnaviridae the ds RNA genome is segemented. Each segment is separately transcribed into cytoplasm by virion assoiated RNA dependent RNA polymerase. The ds RNA segments correspond to single gene. Monocistronic mRNA are transcribed from each segment These RNA's complex with a protein before each is copied to produce ds RNA, which serves as a template for further mRNA's transcription.
iv) ss RNA (+)'sense, virion reverse transcriptase: In retroviruses ss RNA (+) sense is transcribed into DNA by a viral RNA-dependent

40 Textbook o/Veterinary Virology
DNA polymerase. the resulting RNA-DNA hybrid is converted into ds DNA and gets integrated in cellular DNA. Transcription of RNA occurs from the integrated DNA via cellular transcriptase. followed by splicing of RNA transcript as well as cleavage of resulting proteins.
Virus. protein synthesis (Translation): Viral proteins are translated from viral mRNAs at ribosomes in the same fashion as cell mRNAs produced their own proteins. In reoviruses. which have been studied in detail. each monocistronic mRNA binds via capped 5' terminus to 40s ribosomal subunit.· which moves along mRNA molecule until stopped at the initiation codon. The 60s ribosomal subunit then binds together with methionyl tRNA and various initiation factors and then the translation proceeds.
The proteins translated from early transcripts of DNA viruses include enzymes and other proteins required for replication of viral .RNA as well as proteins which suppress host cell RNA and protein synthesis. The function of many early proteins of large DNA viruses is still not known. The late proteins are translated from late mRNA. most of which is transcribed from progeny viral nucleic acid molecules. Most'of the late viral proteins are viral structural proteins and are often produced in considerable excess. The regulation of synthesis of protein is mainly at transcription level. With RNA viruses also early and late proteins are made but the control is not generally as rigorous as in DNA viruses and occurs at the level of translation.
The newly synthesized viral proteins migrate to various sites in the cell where they are needed. The mechanism controlling such migration are not known but probably resemble those employed for cellular proteins.
Viral genome replication: Different mechanisms of DNA replication are employed by each family.
The replication of single stranded DNA of parvoviruses takes place by cellular polymerases. They are initially converted into double replicative form due to a reaction primed by 3' terminus of infecting DNA. Further DNA synthesis requires the binding of a virus coded protein to the 5' terminus. The viral ss DNA appears to be produced after nicks at 5' end and repeated rounds of synthesis.
The replication of linear double stranded adenovirus DNA also require specialized terminal structure. The replication is initiated at either end of the double strand. It is primed by an early adenovirus protein. part of which remains covalently attached to the 5' terminus of new DNA strand. Synthesis from 5' to 3' end proceeds continuously
Viral Replication 41
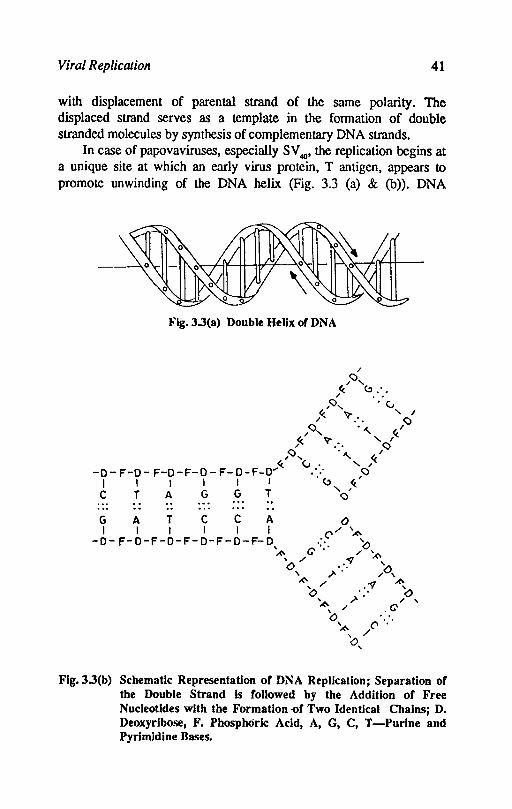
with displacement of parental strand of the same polarity. The displaced strand serves as a template in the formation of double stranded molecules by synthesis of complementary DNA strands.
In case of papovaviruses, especially SV 40' the replication begins at a unique site at which an early virus protein, T antigen, appears to promote unwinding of the DNA helix (Fig. 3.3 (a) & (b)). DNA
Fig.3.3(a) Double Helix or DNA
GAT C C A ° I I I I I I ('/ ""
-0- F-O-F-O-F-O-F-O-F-O ';'. '<:> ~ C' ...... ~ '/ .~ , 0, '" '.' ..... 0,
,('0 / .~ ~ <:>, "'... 0 ~" ~/' , '0 .
'A' (' , / 0,
Flg.3.3(b) Schematic Representation of DNA Replication; Separation of the Double Strand Is followed by the Addition of Free Nucleotldes with the Formation -of Two Identical Chains; D. Deoxyribose, F. Phosphoric Acid, A, G, C, T -Purine and PyrimIdine Bases.

42 Textbook o/Veterinary Virology
synthesis by host polymerases proceeds in both directions. The simultaneous synthesis of both strands takes place in the direction opposite to the movement of other. This is accomplished by repeated initiation and synthesis of short DNA segments complementary to one strand of template, followed by ligation of the segments to produce a complete strand. The replication of DNA is semi-discontinuous, in each direction one strand is produced as a continuous polymer, the other in a discontinuous fashion in segments.
The herpes viruses specify a large number of enzymes involved in DNA synthesis. The replicating DNA initially consists of circles and linear forked forms, which are later replaced by large bodies of tangled DNA.
The replication of poxivirus DNA takes place in the cytoplasm and depends upon virus coded proteins. The replication begins at each end of genome and involves a strand displacement mechanism, with the formation of small DNA fragments covalently linked to RNA primers.
The replication of virion RNA requires first the synthesis of complementary RNA, which then serves as a template for making more virion RNA. In RNA viruses, where RNA is of (-) sense the complementary RNA is of (+) sense and RNA polymerase is the virion associated transcriptase used for transcription of subgenomic RNA. The primary transcripts from (-) sense virion RNA are cleaved to produce mRNA's, some remain uncleaved to serve full length template of virion RNA synthesis.
In case of (+) sense virion RNA, complementary RNA is (-) sense. Several RNA molecules can be transcribed simultaneously from a single complementary RNA template. The resulting structure is known as replicative intermediate, which is partially double stranded with single stranded tails. The replication of picorna and calcivirus RNA is initiated by a protein similar to that of adenovirus DNA. This small protein VPg is covalentIy bound to S' terminus of (+) and (-) RNA strands as well as virion RNA but not to mRNA. In retroviruses replication needs the synthesis of double stranded DNA through reverse transcriptase of virus RNA and this is integrated into cellular chromosome to function in replication. A virion associated reverse transcriptase using the RNA molecule as a primer produces a ss DNA copy. The reverse transcriptase functioning as ribonuclease removes the parental RNA molecule from DNA-RNA hybrid. The (-) sense ss RNA can then occur from integrated (proviral) DNA.

Viral Replication 43
Reoviruses replicate their double stranded RNA by encapsidating mRNA and copying each one of them to form double stranded molecules within the subviral particle. All other viruses replicate their genomes by forming complete transcripts of virion RNA which are then used as templates for RNA replication.
Virion Assembly
a. Icosahedral viruses: The structural proteins of non enveloped icosahedral viruses associaLe spontaneously to form capsomeres, which self assemble to form empty procapsids into which the viral nucleic acid is packed. The packaging of viral nucleic acid in adenovirus mechanism is elucidated. One terminus of viral DNA is characterised by a nucleotide sequence, which enables the DNA to enter the procapsid bound to basic core proteins, after which some of the (;Ore proteins are cleaved to make mature virion.
b. Enveloped viruses: The viruses with helical symmetry of nucleocapsids and few with icosahedral nucleocapsids acquire envelope Lhrough budding cellular membrane. The envelope contains from glycoproteins. The mechanism of glycosylation of envelope proteins is described in brief. In the budding of viruses from cell membranes, the nucleic acid containing subviral particles interact with cytoplasmic domains of glycoproteins to induce the modified membrane to envelope them. The interaction between the helical nucleocapsids of myxo, paramyxo and rhabdoviruses is mediated by additional viral proteins, the M proteins. The icosahedral nucleocapsids interact with their membrane associated glycproteins to yield virus particles containing equal number of virus capsid proteins and each membrane glycoprotein. The cellular glycoproteins are excluded from virus assembly process. In final maturation of viruses containing number of membranes, proteolytic cleavage of specific virus glycoproteins is also required to form the infectious virus particles ego the fusion glycoprotein of parainfluenza viruses and heamagglutinin of influenza viruses.
Replication groups: The following replication groups can be assigned' to the viruses.
i) The viruses with double stranded DNA. They mostly divide iDto the nucleus except pox viruses. All these viruses produce mRNA by DNA transcription, the enzymes used for transcription are packed in

44 Textbook of Veterinary Virology
virus particles like pox viruses or the nuclear polymerases of the cell may be used with or without modification.
ii) This group contains single stranded DNA viruses. The replication involves the formation of double stranded DNA replicative forms. m,RNA is transcribed from one strand of this template. Cell polymerases are required in transcription, which takes place in nucleus.
iii) The retroviruses produce mRNA by transcription of double stranded DNA replicative intermediate. The DNA is produced by reverse trariscriptase of genome RNA and is integrated with cell chromosome. The integrated DNA is transcribed by cell enzymes like other cellular genes.
iv) The double stranded RNA viruses produce mRNA by conservative transcription of double stranded virus RNA using virus specific enzymes. The replication of these viruses is cyptoplasmic and host enzymes are not involved in RNA synthesis.
v) The negative stranded RNA viruses replicate exclusively in the cytoplasm or may involve nucleus as in influenza viruses. The genome may be single RNA molecule or segmented. Messenger RNA is produced by transcription of genome RNA by virus specified enzymes in case of viruses which replicate in the cytoplasm. In influenza virus which involves nucleus for replication, the primers formed to initiate transcription in nucleus are formed by cellular polymerases and both virus and cell polymerases are concerned with influenza mRNA synthesis.
vi) In the (+) stranded RNA viruses the initial expression of genome requires direct translation of the infecting nucleic acid to
produce proteins concerned in RNA replication. Certain members in this group like picorna viruses use only complete virus RNA as mRNA throughout replication while others produce a subgenomic mRNA's to amplify the synthesis of particular gene products.
Selected References
ALBERTS, B.; BRAY, D.; lawIs, I.; RAFP, M.; ROBERTS, K., and WATSON, lD., 1983. Molecular biology of the cell. Garland, New York and London.
BALTIMORE, D., 1971. Expression of animal viral genomes. BacterioI. Rev. 35, 235.
BISHOP, D.H.L. and COMPANS, R.W., eds. 1984. Nonsegmented negative strand

Viral Replication 45
viruses; Paramyxoviruses and Rhabdoviruses. Academic Press, New York.
COMPANS, R.W. and BISHOP, D.H.L. eds. 1984. Segmented negative strand viruses; Arenaviruses, Bunyaviruses and Orthomyxoviruses. Academic Press, New York.
JOKLIK, W.K., 1983. Structure and function of reovirus genome. Microbiol. Rev. 45, 483.
McGEOGH, D.l, 1981. Structural analysis of animal virus genome J. Gen. Virol. 55,1.
SIMONS, K.; GAROFF, H. and HELENUIs, A., 1982. How an animal virus gets into and out of its host cell. Sci. Amer. 246,46.
STRAUSS, E.G. and STRAUSS, J.H., 1983. Replication strategies of single stranded RNA viruses of eukmyotes. Curr. Top Microbiol. Immunol. 105,1.

Chapter 4
Cultivation of viruses
The viruses replicate in living cells only. Some of the viruses have a restricted host range. Most of the viruses can be grown in the cell culture system, embryonated hen's eggs or in the laboratory animals.
Experimental animals
The experimental animals in the case of veterinary viruses may be homologous hosts or heterologous hosts. In human medicine the homologous host cannot be used. Loeffler and Frosch used cattle for earliest studies in viral assay of foot and mouth disease virus. The natural host is still being used for studies ofpathogenesis, immunology, vaccine trials, diagnosis and chemotherapy. The experimental animals should be specific pathogen free. The animals should have no prior immunity to particular virus. The experimental animals are used for following purposes.
i) Virus isolation - For diagnostic purposes the experimental animals are still used e.g. mice in rabies and Louping ill disease diagnosis.
ii) To study pathogenicity and host immune reactions - This is studied in homologous host e.g. pig in swine fever. The cost of using the homologous host is very high and therefore inbred experimental animals are used instead of homologous host, e.g. inbred mice used in African swine fever. The laboratory animals used as models are:
a. Rabbits - The rabbits were used by Pasteur to adapt street virus of rabies. In malignant catarrhal fever virus these animals react in the similar manner as the cattle.

Cultivation of Viruses 47
b. Guinea pigs - Guinea pigs react to foot and mouth disease virus when inoculated intradermally in the foot pad. Primary vesicle is formed on the foot pad and secondary vesicles appear in the mouth following viraemia.
c. Ferrets - Ferrets are used in the study of pathogenesis of distemper virus.
Other laboratory animals are also used in virus study or in the preparation of antisera against different viruses.
iii) To test and develop viral vaecines-Mice, guinea pigs, rabbits are used for attenuation of virus strains as well as for testing vaccines. Foot and mouth disease virus vaccine is initially tested in guinea pigs and finally in cattle and pigs.
iv) To raise manoelonal or polyclonal antibodies-Various routes are employed to inoculate experimental animals with virus infected material. The usual routes are intracerebral, intranasal, intradermal, intramuscular, intravenous and subcutaneous. The route of inoculation largely depends upon the nature of virus, its possible affinity for the tissue, age and species of experimental animal. The experimental animals used in virus work has been replaced to great extent by the use of embryonating chicken and cell culture but still it is a useful method for studying clinical manifestations, pathogenecity, pathogenesis and epidemiology of animal virus diseases.
Embryonated ben's eggs
The embryonating hen's eggs are being used since 1931 when Woodruff and Goodpasture cultivated fowl pox virus on the chorioallantoic membrane. Bumet used chicken embryo for cultivation of viruses very extensively. Nearly all the viruses known at that time could be grown in the chicken embryos by various routes of inoculation. The chicken embroys are still used for isolation and cultivation of many avian and few mammalian viruses. This method is more economical and convenient than animal inoculation. The use of embryonated eggs have number of advantages.
a. It is readily available, cheap and easy to maintain. b. Free from bacteria and many latent viruses. c. Free from specific and non specific factors of defence. d. Se\1sitive to viruses which do not produce infection in adult
birds. The presence of virus of fertile eggs can be detected by changes in
embryos like mortality, deformities, haemorrhages of the embryos,
48 Textbook o/Veterinary Virology
pocks and oedema of chorioallantoic membrane. specific antigens in the fluids like haemagglutinis. complement fixation antigen etc.
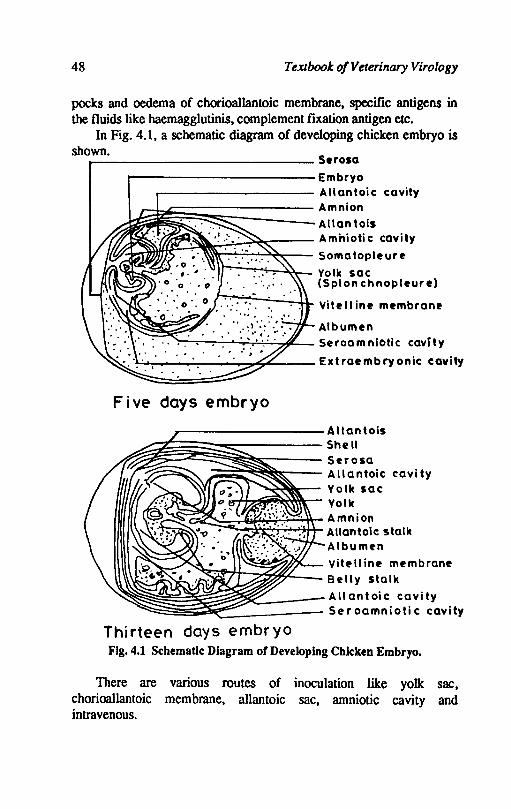
In Fig. 4.1. a schematic diagram of developing chicken embryo is shown. _-------------- S.rosa
~-------------Embryo
------------ Allantoic cavity
~~~~~~~s~==== Amnion ~ Allantois
I'r~f;f~~~~'~i.z:~;:== A mhioti c cavity It Somatopleure I,..,.-........ ;....,...~- Yolk sac
(Splon chnopleure)
Vitell ine membrane
Albumen ~~~.--'~c.r:,.:...:...,4.~ Seroamniotic cavity
.l...:...'--r-__ -_ ....... ....,."':;"O:~ __ Extraembryonic cavity
Five days embryo
_--------- A lIan tois
:::§§i~§~~~~~==== Shell ~ Serosa ;:;~~~;;:--- Allantoic cavity
-......;~~- Yolk sac -;-/-+,..~~~.....- Yo I k
~~~d~~:t:~t Amnion ~ Allantoic stalk
Albumen
~~~,~~ Vitf'lIine membrane Belly stalk
~~~~~~~~~~=====::: A 11 an t oi c ca vi ty Seroamniotic cavity
Thirteen days embryo Fig. 4.1 Schematic Diagram or Developing Chicken Embryo.
There are various routes of inoculation like yolk sac. chorioallantoic membrane. allantoic sac. amniotic cavity and intravenous.

Cultivation of Viruses 49
The yolk sac method of inoculation is performed on 6-8 days old embryos. A vian encephalomyelitis virus aand avian infectious bronchitis virus can be grown by this route. There is dwarfing of embryos inoculated with avian infectious bronchitis virus.
The allantoic cavity route of inoculation is done in 10-12 days old embryos. Influenza virus and Newcastle disease virus are usually cultivated. The virus titre is high and large quantities of virus can be harvested. The amnio-allantoic fluid shows viml haemagglutination when the virus grows.
The chorio-allantoic membrane method is widely used in the veterinary virology. The herpes and pox viruses are usually grown. The virus produces visible foci on the membrane usually known as 'pocks', oedema or other abnormalities. The age of embryo is 10-12 days.
The amniotic sac method of cultivation is done at 10-14 days old embryos. This method is employed in the primary isolation of influenza virus and mumps virus. The growth of virus can be detected by haemagglutination.
The intravenous route of inoculation is carried out in 13 days old embryos. The bluetongue virus is grown by this route. The growth of virus can be detected in the death of the embryo and other changes.
Cell culture
The cell cultures became commonly used after the introduction of antibiotics and fungistatic substances. The use of laminar flow cabinets is also very helpful in preventing the contamination of cultures as well as the leakage of virus. However, the aseptic precautions are still essential. The finger contamination is possible and this necessitates careful handling of cell culture material. There are antibiotic resistant mycoplasma which pose problem. The cultured cells are used for virus isolation, virus titration, vaccine production and biochemical studies. Since 1949 when Enders, Weller and Robbins reported that poliovirus could be grown in non neural cells with the production of cytopathic changes, large number of unknown viruses were isolated in cell cultures. The advantage of cell culture over chicken embryos or experimental animals are:
i) cell cultures can be produced in large quantities and stored at -70°C,
ii) cell cultures give a clear cytopathic effect,

50 Textbook of Veterinary Viro[o gy
iii) cell cultures can be grown in chemically defined medium which is free from antibodies or other infections,
iv) cell cultures can be radiolabelled to study the details of virus multiplication.
Production of cells cultures
The cells may be grown as explants of tissues or usually in monolayer cultures and occasionally as suspension cultures.
a. Monolayer cultures: For almost all diagnostic and research work monolayer cell c.ultures are used. Primary cultures are grown from cells produced by partial proteolytic digestion of tissues like kidney, testis, thyroid etc. Trypsin is used as proteolytic enzyme for cell dispersion. The embryonic cells are often used because they grow well and are easy to be disaggregated. The trypsin dispersed cells are suspended in growth medium and seeded in glass or plastic containers. The cells adhere to the bottom of these containers, flatten and repeatedly divide until a confluent shcct of cells is formed. Secondary cultures are produced by detachment and dispersal of primary monolayer cells which are seeded in fresh growth medium. This cell dispersion is done by trypsin and EDTA mixture. After several passages the cells normally stop to grow but some cell lines have been developed which can be passaged indefinitely. These continuous cell lines become heteroploid and are not diploid in character. Some continuous cell lines have been derived from neoplastic tissue e.g. Hela cell line.
The cells in monolayer cultures in vitro differentiate into three basic morphological types -
i) Epithelial polyhedral cells derived from ectodermal and endodermal tissues like renal cortex, thyroid, amnion and epidermis,
ii) Fibroblast: Spindle shaped cells derived from mesoderm e.g. connective tissue, embryonic muscle,
iii) Macrophages derived from macrophages and monocytes. They can be grown as secondary cultures with difficulty.
The continuous cell cultures or cell line are mostly used in virus work ·but primary cultures are still used to isolate some viruses because the primary cultures are thought to be more susceptible to infection as they carry the receptors which the cell lines lack.
b. Suspension cultures: This process .is used where large scale production of the virus is required as in the case of vaccine production

Cultivation o/Viruses 51
or for biomedical studies. Some cell lines can he adapted to grow in suspension where medium is continuously stirred and aerated, or is stationary. Suspension cell cultures have also been grown from viral and non-viral tumours like lymphoid tumours of Marek's disease, bovine leukosis and canine mammary tumours. Hybridomas are also a special type of suspension cultures.
c. Organ cultures: Explant cultures are occasionally used for research purposes for cultivation of certain viruses. Cells may be grown in vitro as explants of tissues such as respiratory or intestinal epithelium. Tracheal ring cultures are most sensitive assay for avian infectious bronchitis virus.
The cells are grown in tissue culture medium inside closed containers. These containers are made of clear smooth non toxic glass or plastic and sealed with screw caps or rubber bungs. Tissue culture medium is an isotonic sterile solution of essential inorganic ions, glucose, antibiotics and serum. The pH of the medium is maintained at 6.8 to 7.4. In recent years several growth factors have been identified and certain cell lines can be grown in media that are chemically defined. Defined media have advantages for the isolation of viruses that are likely to be neutralized by antibody present in the normal serum but this can be overcome by using foetal calf serum. The serum free media is especially useful for cultivation of hybridoma cells used for production of monoclonal antibodies.
Recongition of virus growth in cell cultures: The following methods are commonly used for diagnostic work:
i) Cytopathic effect: The majority of viruses produce degenerative changes in cell cultures. These changes can be recognised in unstained preparations. The changes produced are rounding, retraction, syncytium formation, detachment from surfaces. Different viruses produce different cytopathic effects (CPE). Fixation and staining of cell monolayers reveals further diagnostic details like inclusion bodies.
ii) Immunofluorescence or immunoperoxidase: Specific antisera labelled with fluorescent dye or enzyme peroxidase can be used in direct or indirect tests to detect the presence of cytopathic or non cytopathic viruses in cell caltures.
Hi) Haemadsorption: Cultured cells infected with certain viruses like orthomyxoviruses, paramyxo .... iruses and togaviruses which bind from cytoplasmic membrane. acquire the ability to adsorb erythrocytes.

52 Textbook o/Veterinary Virology
This ability is due to incorporation into plasma membrane of newly synthesized viral protein that binds the red blood cells. The haemadsorption can be prevented by prior addition of specific antiserum to virus envelope. The haemadsorption can be demonstrated both by cytocidal and noncytocidal viruses. This property is demonstrated very early (24 hours) when only a small number of cells are infected.
iv) Haemagglutination: The medium of cell cultures infected with some viruses like myxo, paramyxoviruses etc. acquire the property to clump the erythrocytes of certain species of birds or animal. This property is inhibited by specific antiserum (Haemaggl uti nationinhibition).
v) Interference: The replication of one virus in cells usnally inhibits the replication of second virus. Interference operates between leukaemia viruses and sarcoma viruses.
The assay of viral infectivity
The content of infectious virus can be assayed or titrated by infecting cell cultures, chicken embryos of laboratory animals or natural host in serial dilution of virus suspension and observing the evidence of virus replication. The virus assays can be quantitative or quantal.
i) Quantitative assay: The quantitative assay is similar to bacterial counts on agar plates. The viable bacteria multiples and forms a colony, the colony count therefore, represents a direct estimate of the number of bacteria. The plaque assay in monolayers of cultured cells is similar to the colony count in bacteria.
a) Plaque assay: Dulbecco in 1952 introduced this technique for quantitation of animal viruses. A suitable range of dilutions (2 fold or 5 fold) is inoculated on the monolayer cultures for an hour or so to allow the virions to adsorb on the cells. The infected cells are then overlaid with semisolid agar or methylcellulose gel, which localises the viral spread. Then each infective virus particle gives rise to a localized focus of infected cells that becomes visible with the naked eye after few days of infection. This area of cytopathology is known as plaque. ~~ make plaque more distinct, the cell monolayers are usually stained with neutral red or crystal violei dye. The living cells are stained while the plaques appear as clean areas against the coloured background.

Cultivation of Viruses 53
Infection with a single virus particle is responsible for a plaque. By counting the number of plaques by a known volume of appropriate dilution, the calculation can be made of the number of infectious virus particles per milliliter of the original suspension which is expressed as plaque forming units (PFU) per millilitre.
b) Transformation assay: Some oncogenic viruses which do not kill cells but transform them i.e. the cells grow in an unrestrained fashion to produce a heaped up mass of cells in a monolayer. These transformed cells can also grow in a semisolid agar or methylcellulose. The number of microtumours are counted and number of infectious virions of oncogenic virus is calculated.
c) Pock assay: The pox group of viruses are grown on the chorioallantoic membrane of chicken emrbyos. Each infectious unit produces a 'pock' which is visible with the naked eye. The number of pocks are counted in a known volume of particular dilution, a calculation can be made to calculate the number of infectious units per millilitre of original suspension.
ii) Quantal assay: This assay does not measure the exact number of infectious virus particles in the inoculum, but only indicates the presence or absence of infectious virus particles. Serial dilution of virus are inoculated into number of replicate cultures, embryonated eggs or animals. The virus, if present, replicates in appropriate time and destroys the cell culture or kills the embryos or animals. Therefore each host gives a single piece of information, whether the infectious virus particles were present or absent in that particular dilution. The end point of quantal titration is taken to that dilution of virus suspension which infects 50% of inoculated hosts and the titre is expressed as 50% infectious doses like infectious dose 50 (ID,J or tissue culture infective dose 50 (TCIDsJ.
Assay based on other properties of virious
Haemagglutination: Many viruses contain virus coded proteins in their outer coat which are capable of binding to the erythrocytes and producing haemagglutination. Hirst in 1941 reported haemagglutination in influenza virus for the first time. Haemagglutinin in influenza virus is a glycoprotein. About 101 influenza virus particles are required to
produce haemagglutination, which can be read with the naked eye. Thus haemagglutination is an indicator of the presence of large number

54 Textbook of Veterinary Virology
of virus particles but it is not sensitive to know the presence of small number of virus particles. Large number of virus families are known to possess the property of haemagglutination like, Adenoviridae, poxviruses,. Togaviridae, Flaviviridae, Orthomyxoviridae, Paramyxoviridae, Corona-viridae, Bunyaviridae, Rhabdoviridae and Reoviridae.
Electron microscopy: Negative staining with potassium phosphotungstate makes it possible to count the number of virus particles by election microscopy. In election microscope even the non infectious virus particles are counted, therefore, the count of a given virus suspension is always higher than in quantal or quantitative assay in cultured cells or eggs.
Selected References
COOPER, P.D., 1967. The plaque assay in animal viruses. In 'Methods in Virology' (K. Maramorosch and H. Koprowski, editors) Vol. 3, 244-311. Academic Press, New York.
FRESHNEY, R.I., 1983. Culture of animal cells. A manual of basic technique. AIan R. LissIWiley, New York.
STOKER, M.O.P. and MACl'HERSON, I.A., (1967). Transformation assay. In 'Methods in Virology' (K. Maramorosch and H. Koprowski, editors). 3,313-336. Academic Press, New York.

Chapter 5
Viral Genetics
Introduction
Viral genetics is concerned with elucidation of the precise structure of the virus genomes to the extent to which determines the biological properties and disease producing capacity of viruses. Viral genetics also involves delineation of the pattern and origin of virus variation. both in terms of virus evoluation and the temporal changes of antigenicity and pathogenecity of viruses.
Effective genetic studies of animal viruses date from Dulbecco's introduction of plaque assay in vertebrate cells and they are now expanding under the impacts of improved cell culture technique. more versatile and precise biophysical and biochemical techniques for studying viruses and their components. The methods of studying virus genetics were based primarily on the isolation of induced or spontaneous mutants followed by their functional classification and by genetic analysis using the methods of genetic engineering and restriction endonucleases. Genetic engineering helps in molecular cloning of fragments of viral nucleic acid and provide methods of sequencing nucleic acid. It is now possible to determine the precise structure of viral genes and even the entire genome of a virus. The possibility of isolating more than one gene and the possibility of mapping genes through shared genomic sequences has been increased with cosmid technology. The development of cosmids have allowed upto 50 kb of genomic DNA to be cloned in a single cosmid vector. Cosmids are plasmids which contain (lambda) cohesive ends. a

56 Textbook o/Veterinary Virology
replication ongm, an antibiotic resistance marker and a unique recognition site into which genomic DNA can be ligated.
Mutation
Good phenotypic characters which are a prerequisite for genetic studies are' provided by mutations that have phenotypic expression. All the early work involved selection for phenotypic changes that reflected a mutation in the particular gene responsible for that function.
Mutation can be defined as alteration or changes in gene, the functional unit of genome. This changes in the gene is the source of variation, it alters the properties of the virus and also causes phenotypic changes of the virus. However, not all phenotypic changes that occur with viruses are due to mutation. Many instances are known in which the virus takes on certain phenotypic characters following the growth in different'types of cells. Such host controlled variation has been found in many groups of viruses such as orthomyxo and paramyxoviruses.
Mutations of animal viruses can occur spontaneously or can be induced by various chemicals such as nitrous acid, 5-Bromodeoxyuridine (BUDR), hydroxylamine and nitrosoguanidine etc. The frequency of mutation either spontaneous or induced is higher with RNA than with DNA viruses because RNA polymerase and reverse transcriptase (in case of retroviruses) are less accurate. The high frequency of mutation usually causes rapid drift of the genome. As a result an RNA virus strain is always a heterogenous collection of different genotypes, the composition of which depends on the selective conditions under which it is grown.
Induced mutation particularly chemical mutagenesis are important for laboratory studies in viral genetics.
Types of mutation
Mutations can be classified according to the change in the nucleic acid and the resultant mutants are classified by their phenotypic expression. Types of mutations according to the change in the nucleic acid are Silent mutation, Nonsense mutation, Frame shift and Deletion mutations. In the first two types of mutations, the effect on polypeptide. function is not adverse or may be variable but in the other two mutations the polypeptide function is usually lost.
The mutants which are classified by their phenotypic expression

Viral Genetics 57
are conditional lethal, temperature sensitive, host dependance (host range), plaque size, on cytopathic and pock types.
The conditioriallethal mutants are those which cannot grow under certain non permissive (i.e. lack of some enzymes or other requirement essential for the replication of a particular virus) conditions, but can replicate under normal or permissive conditions.
The vast majority of the conditional lethal mutants of animal and human viruses are temperature sensitive (ts) mutants. These ts mutants are unable to multiply in susceptible cells at higher temperature. The ts mutants are extremely valuable because they can be recovered in many genes and they form the basis for most of our present knowledge of animal virus genetics.
The Genome of Animal viruses
The nucleic acid or the genetic material of animal viruses is either DNA or RNA. The coding of genetic information in RNA is unique and is the predominant molecular form of the genome in animal and plant viruses.
Viral nucleic acid may be linear or circular, single stranded (ss) or double stranded (ds), covalently bonded to protein or present as unique or diploid subunits according to the type of virus. On the basis of the functional properties, the genome of RNA viruses may be represented as plus or negative strand or both. The genome of plus strand non segmented RNA viruses is infectious since it can function directly as mRNA for synthesis of entire viral proteins and the polymerase necessary for replication of the viral genome. The genome of negative strand RNA viruses is not infectious because it cannot function as mRNA. The nucleocapsid of negative strand viruses includes an RNA dependent RNA polymerase as integral part of its structure so that mRNA synthesis can be initiated. The genome of retroviruses although possess plus strand and function as mRNA, but is not infectious because it is dependent on preformed virus associated reverse transcriptase for its replication. The genome of cytoplasmic viruses (e.g. pox viruses) is not infectious and several enzyme activities including DNA dependent RNA polymerase are associated with the virion.
The Molecular weights (M.W.) of the genomes of RNA viruses range from 1.5 x 1()6 of the parvovirus to the 169 x 106 of the poxvirus.

58 Textbook of Veterinary Virology
On the other hand the RNA viruses are more uniform and the M.W. of the genome vary within a three fold range only.
The relative, coding capacity of a viral genome'provides a measure of the minimum amount of genetic information present in the viral genome. The ratio of the average nucleotide M.W. to the average amino acid M.W. is taken to be 321:110 or 2.9:1 approximately. As a result of thumb calculation the relation between genome M.W. and gene product M.W. is 10:1 for single strand viruses and 20:1 for double straDd viruses. Although the relative coding capacity assumes that genes are unique and non overlapping, it remains useful ind~x for comparison of the minimum genetic information content of the various virus groups.
Number of gene in viruses
All DNA viruses and several groups or RNA viruses have genomes consisting of a single molecule of nucleic acid but the genomes of some RNA viruses consist of several pieces of RNA. If the gene is defined as the nucleotide sequence that specifies a polypeptide it is possible to estimate the number of genes in different viruses from our knowledge of the amount of viral nucleic acid in the virus particle. In viruses with fragmented genomes each molecule of RNA represents a single gene.
Mapping of the genome of animal viruses
The basic aim of virologists is to characterise the various functions involved in virus multiplication and to identify the portion of the viral genome that encode these functions. Great strides have been made in the mapping of animal virus genomes, that is in identifying the portions of their genome that encode specific proteins.
Viruses with a positive strand continuous genome that is translated into a single polypeptide chain (e.g. polio) gene maps can be obtained biochemically. A wide range of inhibitors of protein synthesis with accurately defind modes of action have been available and are used for precise gene sequence with RNA molecule by measuring the order in which proteins are synthesized. Pactamycin for example specifically inhibits inhibition of protein synthesis but permits completion and release of polypeptides. A radio active amino acid is added to the infected cell culture together with the antibiotic pactamycin, which inhibits initiation of protein synthesis. The label will be incorporated

Viral Genetics 59
more by proteins corresponding to the distal 3' end part of the RNA which has the highest chance of being still untranslated when the label is added. The relative labelling of various proteins gives their location in the uncleaved polypeptide chain.
The gene order for several negative strand virus has been determined by measuring the effect of UV irradiation of virions on the synthesis of the polypeptides specified by the various genes. The method is based on the blocking of the progress of transcription by the UV induced pyrimidine dimers, therefore, a gene at the 5' end on the template strand has the highest sensitivity to the radiation because its transcription is blocked by any dimer alorigwith the whole genome and the synthesis of peptide undergoes the greatest reduction.
The location of genes encoding proteins of different viral genomes can be determined by variety of techniques, among these, the genetic recombination analysis and restriction endonuclease cleavage maps are commonly used.
Genetic recombination between viruses
The relative order and location of genes can be specified by measuring the frequencies with which pairs of mutants in the various genes recombine with one another to yield wild type virus.
Several kinds of genetic recombination takes place when two different virus simultaneously infect a same type of cell. This recombination occurs between the newly synthesized nucleic acid. Different types of genetic recombinations are intramolecular recombination, reassortment, reactivation and marker rescue.
Intramolecular recombination: It involves the exchange. of genetic material between different but closely related viruses. Intramolecular recombination observed with all double stranded (ds) DNA viruses and among RNA viruses particul&rly with foot and mouth disease virus and polio virus. Increased frequency among DNA viruses is presumably due to strand switching by the viral polymerase. In rare cases intramolecular recombination occurs between unrelated viruses e.g. SV 40 and adenovirus. By the process of intramolecular recombination the retrovirus also pick up cellular oncogenes and such oncogenes are incorporated into the proviral DNA and subsequent transcription of such oncogene into viral RNA converted into viral oncogenes (Fig. S.la). The mechanism of recombination between

60 Textbook of Veterinary Virology
nonsegmented RNA genome remains 10 be elucidated. High frequency of recombination is also observed with RNA tumour virus and recombination is a normal consequence of replication of the genome.
Reassortment: Viruses with segmented genome undergo recombination by exchange of complete genome subunits. This form of recombination is known as reassortment. This occurs when a particular cell type is infected with two unrealated viruses. No DNA virus has segmented genome, therefore, the phenomenon of reassortment is confined to RNA viruses. Reassortment is an important source of genetic variability. The major antigenic changes of influenza virus are primarily caused by reassortment between different species (Fig. 5.1b).
Reactivation
This is applied to the condition during which there is production of infectious virus particle when a cell is infected with two or more viruses of the same strain and each of which has suffered a lethal mutation. This type of multiplicity recombination leading to the production of viable recombinants is observed among the members of the poxviridae, orthomyxoviridae and reoviridae families.
Marker rescue
This refers to the genetic recombination between an infectious virus and an inactivated virus of a related but distinguishable genotype.
A special kind of marker rescue called fragment rescue has been extensively used for correlating the functional and physical maps of viral DNA. It involves the introduction of a fragment of DNA containing a specific mutation produced by recombinant DNA technique along with the parental virus or its complete genome into a susceptible cell. By this process the desired mutation can be introduced into a portion of the progeny virions (Fig. 5.2).
Restriction endonuclease cleavage maps
In one method the genes are mapped by crossing parts of mutants in different genes of virus strains that are related and characterized by the DNA of wild type recombinant by restriction endonuclease analysis.
In other method, mRNA from infected cells are hybridized 10
separate restriction endonuclease fragments. They are then dissociated from the DNA, translated in cell free protein synthesizing systems an.1

Fig. 5.1(a) Genetic Recombination-Intramolecular Recombination.
Fig.5.1(b) Genetic Recomblnatlon-Reassortment. 0'1 -
62 Textbook of Veterinary Virology
Parental virus
o D~A I Fragments
;9 \+ .+~ ().
+ / ~
O~ 0 Mutant
+ + + and \..~mm / +/ fragments ~
I ""omb;"aHo" I
o 0 Mutant Parental typt>
Fig. 5.2 Fragment Rescue. Cells are Infected with Specific Restriction Endonuclease Generated Fragments of Parental Virus DNA and Its Mutants only the Fragment Carrying the Parental Allele of the Mutated Gene (m) can give Parental Type Recomblnants.
the proteins that are formed are identified. Since the position of the viral genome of each restriction endonuclease cleavage fragment is known the location of the gene coding for the protein that is translated can be identified.

Viral Genetics 63
Genetic Engineering
Restriction endonucleases: These are the enzymes of bacterial origin which have the property to cleave viral DNA at specific sites. The word restriction was given in 1960 when it was observed that certain bacteriophage failed tq grow in particular species of bacteria.
The failure to replicate inside the bacteria is due to the degradation of the phage DNA by specific endonuclease of the bacteria. Several hundreds of restriction endonucleases have been identified and purified from various bacteria. Each of these restriction endonuclease recognises a unique short (usually 4 to 6 nucleotide base pairs) sequence of nucleotides and cleaves DNA into precise number of fragments.
The DNA fragments produced by a panel of restriction endonuclease can be analysed by gel electrophoresis and analysis of these patterns can be used to identify viral sUbtypes.
Recombinant DNA: The development of recombinant DNA methodology has been facilitated by great improvement in the technique of sequencing DNA. With the help of DNA sequencing technique it is possible to get the exact picture of a DNA molecule and large quantities of selected fragments of viral nucleic acid can be obtained by the use of restriction endonucleases.
In recombinant DNA technique the desired DNA or copy DNA from a virus is selectively cut into fragments by the use of specified restriction endonuclease. Such viral DNA fragments are then inserted into the DNA molecules usually of a bacterial plasmid DNA, and are joined together with the help of enzyme ligase. The plasmid DNA containing the foreign DNA is then placed into the vector bacterial species. Inside the bacterium there is replication of the plasmid DNA and there is production of many copies of the plasmid. Bacterium containing the desired plasmid (i.e. the plasmid containing the foreign DNA) is identitied, cloned and allowed to grow.
Recombinant DNA technique is applicable not only for DNA viruses but also for RNA viruses. With the help of reverse transcriptase it is possible to make DNA copy from either viral RNA or mRNA.
References
FENNER, F.; McAusLAN, B.R.; MlMs, e.A.; SA..\-1BROOK, J. and WHITE O.D., 1974. The biology of animal viruses 2nd Ed. Academic Press, New York.

64 Textbook o/Veterinary Virology
FENNER, F.; BACHMANN, P.A.; OmBs, E.PJ.; MURPHY, F.A.; STUDDERT, M.l. and
WHl1B, O.D., 1987. Veterinary Virology Academic Press, Inc. (London) Ltd.
BROWN, 'F. and Wn.soN, S.O., 1984. Topley and Wilson's Principles of Bacteriology, Virology and Immunity Vol. 4, 7th Ed. Edward Amold Ltd. London.
HUNTER, E., 1978. Current Topics in Microbiology and Immunology. 79:295.
CONRAT, F.H., 'KIMBALL, PAUL, C. and LEVY, JAY, A., 1988. Virology, 2nd Ed. Prentice Hall, Englewood Cliffs, New Jersey.
JOICLIK, W.K., 1980. Principles of Animal Virology. Appleton Century Crofts, New York.

Chapter 6
Viral Pathogenesis
The understanding of pathogenesis of viral infections in host can be best understood by studying virus induced changes in cultured cells. The changes observed io cultured cells can be used to interpret the changes in whole animal.
Viruses may be cytocidal or noncytocidal. The cytocidal or lytic viruses produce morphological changes in cells known as cytopathic effects (CPE). The noncytocidal or nonlytic viruses are non cytopathic and produce little metabolic disturbance. The cytocidal and monocytocidal viruses do not always lead to production of new virus particles. In certian viral infections the viral genome either persists as an episome or is integrated with the host cell genome. In such cases the transformation of host cells takes place. These transformed cells may produce tumours in experimental animals. The morphology of host cells is altered and these transformed cells can be passaged indefinitely. Certain noncytocidal viruses produce persistant infection. The infected cells produce and release virions but cellular metabolism is not affected, the infected cells continue to grow and divide. The various types of interactions between virus and cell are summarised as under:
1) Cytocidal infection: Changes in morphology of infected cells (ePE).
2) Persistent non cytocidal mfection: No CPE, the virus and ceUs continue to grow and divide.
3) Persistent noncytocidal non productive infection: No CPE. Viral genome persists a3 episome or integrated. Virus is not produced normally but by cocultivation with permissive cells, irradiation or with chemical mutagens the virus can be eJCpressed.

66 Textbook of Veterinary Virology
4) Transformation: The cell morphology altered. It is produced by RNA tumour viruses and rarely by DNA tumour viruses.
Cytopathic effect
The' lytic infection produces cytopathic effect (CPE) in cultured cells which is an important diagnostic criteria. Several viruses produce characteristic cytopathic effect. In the infected cells there is shut down of cel1ular proteins, large number of viral macromolecules accumulate, sometimes vlfal proteins are found in crystalline aggregates or inclusions and these distort the cell morphology. The cell damage due to virus infection can be due to the reasons given below:
a. Shut down of cellular protein synthesis: The cytocidal viruses produce proteins early in infection which are responsible for stopping the synthesis of cellular proteins and these proteins in turn affect the synthesis of cellular RNA and DNA. This is incompatible with the survival of cells. Some viruses like picornaviruses, poxviruses and herpesviruses shut down the synthesis of cellular proteins rapidly and these viruses are rapidly cytopathogeic. The adenoviruses shut down is more gradual and with noncytocidal viruses like in retroviruses there is no shut down and no cell death. There are certain viruses (flaviviruses) which are cytocidal-and at the same time do not shut down the cellular protein synthesis indicating that this is not the only mechanism responsible for cytopathic effect.
b. Cytopathic effect of viral proteins: The capsid proteins of certain viruses in high conct'ltrations are toxic to the infected cells. The penton and fibre proteins of adenoviruses are toxic and may be the cause of cytopathic effect. The cytopathic effect is also produced when a large inocula is used to infect the cells.
c. Inclusion bodies: Certain viruses produce inclusion bodies in the infected cells. These inclusions may be intranuclear or intracytoplasmic and may be acidophilic or basophilic. The pox viruses, paramyxoviruses, reoviruses, rabies virus produce intracytoplasmic inclusion bodies while adenoviruses, herpes viruses and parvoviruses produce intranuclear inclusion bodies. Canine distemper and rinderpest viruses may produce both intracytoplasmic and intranuclear inclusion bodIes in the same cell. The inclusion bodies are accumulation of viral structural components. The basophilic inclusion bodies in pox viruses are si.es of viral synthesis. In fowl pox and cowpox viruses the inclusion bodies arc acidophilic and represent accumulations of viral

Viral Pathogenesis 67
proteins. In adeno and reoviruses the inclusion bodies represent crystalline aggregates of virions which distort the cell. While in herpes viruses the inclusion bodies are the result of late degenerative changes and these produce margination of chromatin.
d. Cell fusion: The paramyxoviruses, herpesviruses, some coronaviruses and pox viruses produce syncytia due to the changes produced in the cell membranes which result in fusion of infected cells with uninfected cells. The syncytia are also produced by these viruses in the tissues of infected animals.
In addition to the changes produced in cells due to spccific effects of viral replication the virus infected cells also show non specific changes like cloudy swelling. The cloudy swelling changes the permeability of plasma membrane. The cell destruction is also the result of leakage of lysosom:ll enzymes into the cytoplasm.
Persistent infections
The noncytocidal viruses replicate in the cells but do not kill them. These viruses often produce persistent infections in the cells in which they replicate. The cell metabolism is little affected. In several RNA viruses like arenaviruses, retroviruses and some paramyxoviruses the virions are released by budding from plasma membrane and the persistent infection is produced. The infected cells yield the virus and grow and divide for long periods but slow and progressive changes are produced leading to cell death except retroviruses. In the animals the cell replacement is rapid and slow death of cells due to persistent infection has no effect. The persistently infected cells, however, lose this capacity to carry out specialised functions and antigenic changes are produced in the cell membrane of infected cells. These viruses may also interfere with the secretions of immunoglobulins by lymphocytes and hormones by somatotrophic cells e.g. cells of islets of Langerhans, w~thout kilhng the cells concerned. Rhinovirus infection results in cilial stasis and destruction of cilia subsequently but the cells are often not killed. This lowers the resistance of respiratory tract to secondary bacterial infection.
New antigens in infected cells
New virus specified antigens appear into the plasma membrane of infected cells. The plasma membrane of cells infected with enveloped RNA viruses incorporates viral heamagglutinin which is exhibited by the property of haemadsorptioll (influenza virus, paramyxoviruses and

68 Textbook of Veterinary Virology
toga viruses). The virus specified proteins appear in the plasma membrane in the early stage of infection with many viruses. The virus coded antigens in the plasma membrane constitute a target for the virus specified immune mechanisms which may destroy the cells before significant number of virus particles are produced and thereby slow down the progress of infections. In some cases, host immune response may precipitate immunopathological reaction. In retroviruses infection, transplantation antigens appear on the plasma membrane of the transformed cells.
Cell transformation
The viruses of many DNA and RNA viruses change the growth characteristic of cells. This alteration in cell morphology is called cell transformation. The viruses producing cell transformation are related with oncogenic ability of these viruses in animals. In the DNA virus transformation the cells do not produce infectious viruses and the infection is non productive while the infection with RNA viruses like retroviruses, the transformed cells produce infectious virus and it is productive infection. The virus DNA in transformed cells remains integrated into the host cell DNA or may be episomal as in the case of papilloma and herpcsviruscs. The transformed cells survive and can be passaged indefinitely. These transformed cells have the capacity to induce tumours in the nude mice, which have defective cellular immunity. The transformation by DNA viruses is non productive and certain virus specific antigens are regularly demonstrable. Tumour associated transplantation antigens (TST A) are located in the plasma membrane while tumour (T) antigens are found in the nucleus.
Infection and spread of viruses in the body
The viruses enter the animal body through one of its surfaces, then spread, either locally or through blood or lymphatic system to produce systemic infection. For the maintenance of virus infection in nature the infectious virus must be shed in the environment or taken up by a vector or passed congenitally.
Routes of entry of virus in the animal body
Most viruses enter the host either through skin or mucous membranes of respiratory and alimentary tracts, while a few viruses also enter through urogenital tract and conjunctiva. The arthropod borne viruses enter their host by direct inoculation by the bite of insect.

Viral Pathogenesis 69
Certain viruses produce the disease at the site of their entry without any systemic spread, such as influenza viruses and rota viruses. The viruses which produce disease at distant sites from their entry point, penetrate the mucosal barrier and then spread within the host at the site where viral replication and disease production takes place.
Table 6.1 PORTALS OF ENTRY OF VIRUSES IN THE ANIMALS
Route
Skin (abrasions)
Skin Bite by arthropod Mechanical
-do- Biological
Genital tract
Conjunctiva
Respiratory tract
Intestinal tract
Congenital infection
Viruses
Pox viruses (fowl pox, cowpox, swine pox, contagious pustular dermatitis, bovine papular stomatitis viruses), herpesviruses, picornaviruses (swine vesicular disease virus), Papillomaviruses. Fowlpox virus, swinepox virus, myxoma viruses, Marek's disease, equine infectious anaemia virus, rabies virus. Rift valley fever virus, Nairobi sheep disease virus, Equine encephalitis virus, Japanese encephalitis virus, Louping ill virus, Turkey meningoencephalitis virus, Wesselsbron disease virus. Herpes viruses, Equine arteritis virus, Bovine papilloma virus. Equine herpes virus-I, Infectious bovine rhinotracheitis virus. Herpes viruses, Adenoviruses, Feline panleukopenia virus, Canine parvovirus, Rhinoviruses, Apthoviruses, Feline calcivirus, Parainfluenza viruses, Respiratory syncytial viruses, Influenza viruses, Newcastle disease virus, Canine distemper virus, Rinderpest virus, Lymphocytic choriomeningitis virus, Hog cholera virus, Pseudorabies virus. Marek's disease virus, Bovine malignant catarrhal fever virus. Coronaviruses, Rotaviruses, Astroviruses, Toroviruses, Adenoviruses, Enteroviruses. Avian leukosis viruses, Avian encephalomyelitis virus, Infections bovine rhino tracheitis virus, Bovine virus diarrhoea virus, Bovine leukemia virus, Bluetongue viruses, Equine herpes virus-I, Equine arteritis virus, Pseudorabies virus, Swine parvovirus, Japanese encephaliti~ virus in swine, Hog Cholera virus, Border disease virus, Rift Valley fever virus, Canine herpes virus 1, Feline panleukopenia vim:. Feline leukemia virus.

70 Textbook o/Veterinary Virology
a. Entry through the skin: The skin is a strong barrier to infection. The virus is destroyed by desiccation and by acids and other inhibitors formed by skin microorganisms. The cornified epidermis is impermeable unless broken by cuts, abrasions or punctures. Insect bite is the main method of penetration.The mosquitoes, mites, ticks; sand flies, fleas directly inoculate the viruses into the blood strcnm. Some of the viruses are transmitted mechanically while others are transmitted biologically. The bites of large animals, like dogs, jackals, wolves, bats etc. introduce saliva containing rabies viruses into the tissues. The entry through the skin is purely mechanical and virus does not play any part.
b. Respiratory tract: The respiratory tract is protected by defense mechanism, the scavenging role of alveolar macrophages and mucociliary blanket which removes particles. The initial lodgement of air borne virus depends upon the anatomy of respiratory tra~t as well as on the size of the droplet, which is very important. Large particles are trapped by nasal hairs. Particles of about lO/lm in diameter arc deposited on the nasal epithelium over the turbinate bones and those of 5/lm in diameter reach lung alveoli. Wherever the virus particle lands, it has to make a contact with the surface of epithelium to initiate infection. The contact occurs only by chance ?ecause the virus particles entrapped in mucous and passed upwards to the pharynx or backwards from nose by mucociliary escalator. The factors which affect the chance of contact are thickness, flow rate and viscosity of mucus and gaps in mucociliary blanket. Mucus secretion and ciliary movements may be inhibited by changes in temperature, ion concentration and humidity of the air. Many viruses remain localized to the respiratory tract but some viruses enter via the respiratory tract following systemic spread.
c. Alimentary tract: Several viruses enter the alimentary tract by ingestion. The moving contents of intestine remove the .viruses which do not adhere to intestinal epithelium. The mucus surface is acidic in stomach and alkaline in the intestine and also contains phagocytes and virus inhibitors such as proteolytic enzymes, bile and antibodies. The heavy load of bacteria also may have a protective effect. Those viruses which survive, infect the ahmentary tract, enteroviruses, reoviruses, coronaviruses and rotaviruses are more resistant to bile and acid than rhinoviruses, intluenza viruses and enveloped viruses which do not produce intestinal intection. Some of the enteroviruses like avian and

Viral Pathogenesis 71
porcine encephalomyelitis viruses cause generalized infection rather than the disease of the intestinal tract Parvoviruses produce diarrhoea after reaching the cells of intestinal tract via blood.
d. Urogenital tract: Several important pathogens of animals like infectious bovine rhinotracheitis, equine rhinopneumonitis and porcine papilloma viruses infect this tract. Many viruses do not establish infection presumably due to lack of interation with host cell receptors and frequent flushing with the sterile urine.
e. Conjunctiva: The conjunctiva is protected by the secretions from ~he lachrymal and other glands. Occasionally viruses infect the eye (adenoviruses, herpesviruses and vaccinia virus). Experimentally infection through this route is produced by large number of viruses.
The exact mechanism and determinants that enable viruses to survive and penetrate mucous surfaces is little known as yet.
Virus spread within the host
Viruses may spread through a number of pathways depending on the specific entry point and target organs involved. The viruses may remain localized to the body surfaces through which they enter or they may produce generalized infection due to viracmia and subsequent localization in target organs.
Localized spread of infection: In tissues cell to cell infection takes place with or without extracellular phase. The virus can spread over mucous surfaces by moving mucus and lumen contents. Such spread is not possible over the dry skin but some spread may take place by scratching, rubbing and by fingers etc.
Spread through the lymph and blood system: The viruses which survive the local inflammatory reaction reach the subepithelial tissues. The virions enter the local lymphatics with the association of phagocytes. Viruses enter the blood stream through damaged capillaries at the initial lodgement site or later with lymph. The spread of virus to other tissues takes place if the virus survives humoral and cellular nefence mechanism in lymph and blood systems. The virus should have a capacity to counteract the mononuclear phagocytes and lymphocytes if< the lymph and blood, the fixed macrophages in lymphnodes, spleen and liver. The viruses which lack the property for counteracting this defence mechanism remain confine.d to the lodgement site. The size of virus parii(' It: influences the efficacy of ingestion by reticuloendothelial

72 Textbook of Veterinary Virology
macrophages. The half life of circulating large viruses is less than small viruses. The speed of blood and lymph flew also has a role in the efficacy of ingestion by the reticuloendothelial macrophages. The slower the. flow rate the greater is the chance of uptake. Certain viruses like small pox rinderpest and polio replicate in lymph nodes and when virus particles are discharged in the efferent lymph, they enter the blood to produce generalized infection. The blood borne viruses are protected from humoral defence mechanism by associating with mononuclear lymphocytes, phagocytes, erythrocytes and other cells. When the concentration of virus particles increase in lymph and blood, the chances of breaching of biood-tissue junctions increase e.g. escape in large amounts from the 'primary lodgement site and replication within circulating and flxed phagocytes and lymphocytes. Some viruses such as ectromelia and distemper of dogs may maintain their circulating concentration by replicating in endothelial lining of lymph and blood vessels. The original escape of virus from primary lodgement site into the blood results in low viraemia and this primary viraemia is followed by higher, secondary viraemia due to replication of virus in primary target organ. This secondary viraemia also leads to infection of secondary target organs like brain, placenta and skin. Infection of central nervous system might follow from blood during viraemia through the cerebrospinal fluid by either passing or growing through the choroid plexus or virus might enter the cerebrospinal directly from the blood. Viruses seem to cross the blood brain barrier easily in immature host because of the thinner basement membrane. The blood skin barrier is breached due to local inflammation. The blood borne viruses localize in small vessels at the site of inflammation and pass across capillary endothelium. The strength of maternal blood-foetal junction varies with the type of placenta and stage of pregnancy. Few viruses breach this barrier. The placenta may be infected but foetus may still be protected e.g. blue tongue virus, while some viruses cross this junction and infect the foetus.
Virus transport along nerves: The spread of virus from peripheral sites to the central nervous system occurs along nerve flbres. The exact pathway of virus transport along the nerve is not exactly known The possibility in the transport of virus is sequential i.}fection of Schwann cells, transit along with the tissue spaces betweea nerve flbres and carriage up the axons.

Viral Palhogenesis 73
Virus shedding: The virus shedding occurs via one of the body
openings or surfaces involved in the entry of viruses. To maintain the infection in populations the virus shedding is essential. The shedding of virus from animal takes place via skin, respiratory tract, digestive tract, urino-genital tract and milk etc.
a) Skill: Not many viruses are shed from skin lesions to cause virus transmission. In case of foot and mouth disease, vesicular stomatitis; pox virus and certain herpes virus infections, the virus is shed from the vesicular or pustular lesions. The virus shed via saliva and aerosal is more important in their transmission rather than those shed via skin lesion. The skin is an important source of virus where transmission is by direct contact. The localization of virus in feather follicles is important in virus shedding of Marek's disease virus from infected chicks.
b) Respiratory tract: The viruses causing diseases of respiratory tract are shed in fluid expelled from the respiratory tract. Large droplets fall rapidly and contaminate fomites while small droplets remain airborne. Respiratory viruses are excreted both in nasal and oral secretions. In certain virus infections the shedding of virus continues even during convalescence.
c) Digestil'e tract: The enteric virus are excreted in the faeces. These viruses persist for sometime outside body and are more resistant to environmental conditions. The rotaviruses and enteroviruses are relatively heat st.1ble.
d) Urogenital tract: A number of viruses are excreted in the urine, ego rinderpest, canine hepatitis, foot and mouth disease virus. These viruses replicate in tubular epithelial cells of kidney and excreted in the urine. The arenaviruses are excreted throughout the life of infected rodents. During coitus certain viruses are transmitted from male to female and many viruses are excreted in the semen.
e) Milk: There are several viruses which are excreted in milk like foot and mouth disease virus, mouse mammary tumour virus, some tickborne flaviviruses etc.
f) Blood and organs: During viraemic stage the insect acquire infection and spread a(bcviruses. Equine infectious anaemia and bovine leukaemia virus are transmitted by contaminated needles and other equipment. The pig and dog may be infected by consuming virus containing meat. Hog cholera, African swine fever and vesicular

74 Textbook o/Veterinary Virology
exanthema viruses are often transmitted by feeding the contaminated garbage to the pigs.
Damage to host
The virus damage to host results in mild, severe or fatal illness or oncogenesis. There is some information avaiable as to how the damage occurs.
It has been shown that virus replication can take place in vivo without producing morphological damage but biochemical changes in such cells take place with pathological effects. Hence, although apparently damaged cells attract attention first to explain the pathology of the disease but it would not be wise to ignore any cell type showing evidence of virus replication.
The manifestations of primary effects of some of the viruses may be cell damage e.g. damage of nerve cells by rabies virus leading to neurlogical effects, diarrhoea caused by rotaviruses and corona viruses due to infection of intestinal epithelium. While many manifestations are secondary effect of the original cell damage, inflammation is one such general reaction resulting from complement activation, or liberation of endogenous permeability factors from damaged cells. The haemostatic response lowers circulating blood volume. The enteroviruses bring about fluid disturbances. Fever is another reaction resulting from endogenous pyrogen by viruses.
Inflammatory and vascular disturbances occuring later in virus infection may be due to immunopathological reaction. Irtlymphocytic choriomeningitis virus (LCM) infection of mice and Aleutian disease of mink, immunopathology plays a majol role in disease manifestations.
Damage to the foetus
The foetal damage results from direct infection or indirectly from the effect of virus disease on the mother. The cell damage to foetus, is due to direct action of virus and primary and secondary manifestation are the same as in the adult animal. However, these are amplified by rapid metabolic and developmental changes occuring in the foetus. The mitotic inhibition and chromosomal damage assumes more importance in foetus in comparison to adult animal. The timing of infection during foetal development is very important determining the nature and extent of damage. The rubella virus infection In the first trimester of

Viral Pathogenesis 75
pregnancy, the malformations of foetus is followed directly from virus destruction of certain cells within the affected organs. When organs develop later in pregnancy malformations may be induced later in foetal life. The cerebdlar hypoplasia produced by Kilham's rat virus and feline panleucopenia virus may be the result of cytolytic action of viruses on external germinal layer of cerebellum in laler period of pregnancy.
The virus infection of mother also affect foetal development. Fever produces abortions, stillbirths and foetal malformations. Changes in foetal circulation may have rapid and severe affect on foetus.
Damage to infant
Maternal antibody protects infants against many virus infections and also reduces the severity if the disease occurs. There are c~rtain viruses which adversely affect the young animals. Rotaviruscs prGduce enlentis in young animals more than adults.
References
BLOOM, B.R. and RAGA-ZISMAN, B., 1975. Viral immunology and immunopalhology. Academic Press, London.
BlTRUS, W.H. and ALLlSON, A.C., 1977. Virus infection and the cell surface North Holland and Publishing Co. Amsterdam.
COOMBS, R.R.A and GELL, P.G.H., 1969. Clinical aspects of immunology. Blackwell Scientific Publications, Oxford.
DARl\'ELL, M.B. and KOPROWSKI, H. 1974. Mechanisms of virus disease. Benjamin, Mento Park,. California.
DOHERTY, P.C. 1980. The molecular basis of microbial palhogenicity. Verlag Chemie. Weinheim.
MTh1s, C.A. and WHITE, 0.0. V;ral palhogenesis and immunology Blackwell SCientific Publications Oxford.
NollCDls, AL. and OLDSTONE, M.B.A, 1984. Concepts in viral pathogenesis. Springer-Verlog, New York.
Tyrell, D A.J. 1983. liow do viruses invade mucus Surfaces? Phil. Trans. R. Sco. London, B. 3() 3 75.

Chapter 7
Persistent Infections
Viruses of some families (herpes viruses) have been known to cause infections that persist throughout the life of the infected animal. The episodes of clinical disease might occur at long intervals. Certain viruses produce chronic disease and the virus persists for months or for life and produce pathological effects. These persistent viral infections are important as they are often important from epidemiological point of view. They may be reactivated and cause episodes of disease, or may lead to immuno pathological disease or may lead to neoplasia. For convenience, persistent infections may be divided into three categories.
a) Latent injections: The virus is generally not demonstrable except when reactivated to replicate, such episodes being sometimes associated with recurrence of disease e.g. bovine herpes virus 1 and cytomegalus virus.
b) Chronic injections: The virus is continuously demonstrated and often shed. The disease may be chronic or absent or may develop later with immunopathological basis e.g. African swine fever virus, Aleutian disease virus, Hog Cholera virus etc.
c) Slow injections: Virus slowly Increases in concentration during a long preclinical phase leading to slow progressively lethal disease, e.g. Visna-maedia virus, caprine arthritis-encephalitis virus etc.
Latent infections
In herpes virus infections latency IS followed by recovery. The mechanism of latency has been described in great detail in human infections with herpes simplex. varicella-Zoster and EB virus infections

Persistent Infections 77
but the mechanisms is comparable for animal infections as well. During primary infection with infectious bovine rhinotracheitis virus the virion move to cranial or spinal ganglia alongwith the axons of sensory nerves.The virus persists in ganglian neuron as episomal viral DNA. Some of viral DNA is transcribed to mRNA but none is translated except reactivation, when production of infectious virus takes place. In recurrent bovine rhinotracheitis infections the virus moves down the sensory nerves again till it reaches the nasal mucus membrane or the skin, where further replication occurs in the epithelial cells and virus shedding takes place. This is the mechanism which maintains the virus from generation to generation in the bovine population. The cytomegaloviruses establish latent infections in the salivary gland, baldder epithelium and in monocytes and/or lymphocytes. The virus is shed into the oropharynx and urine from which it is transmitted directly.
Chronic infections
In chromc virus infections there is continuous virus production. There may be no disease, chronic disease or disease occuring as late complication. The following chronic diseases are being discussed below in brief.
Lymphocytic choriomenigitis: Lymphocytic choriomeningitis (LCM) is caused by arenavirus, is an example of persistent infection in mice. The virus is transmitted horizontally and in utero. The mice are normal at birth and appear normal for rest of their lives. Infected mice have persistent viraemia and viruria, every cell of the animal is infected and remains so throughout life. The circulating antibody is not detected but immunological tolerance is not complete. A very low antibody is produced which forms virion-lgG-complement complexes. These complexes are infectious. There is no cell mediated immune response to the virus. The inbred miGe late in life exhibit 'late disease' due to deposition of antigen-antibody complexes in the renal glomeruli of the kidneys.
Foot alld Mouth disease: In cattle recovery from the disease is not complete as the virus is not elimianted. The virus is now known to
cause persistent infection of the pharynx of cattle, sheep, goats, and other ruminants. The persistence of virus is not in all infected animals, only few animals become the carriers. The cattle vaccinated with

78 Textbook o/Veterinary Virology
inactivated vaccines may also become carriers if infected subsequent to vaccination. The recovery of the virus from cattle and buffaloes have been made even after 2 years of infection. The mechanism of persistence is not known and its epidemiological significance is difficult to assess. The transfer of infection from carrier to susceptible cattle is doubtful but transfer of infection from carrier infected African buffalo to cattle is known to occur.
Canine distemper: Canine distemper produces a disease known as 'old dog encephalitis' whic is similar to a disease among human beings known as sclerosing panencephalitis due to measles virus. It is now known that a few dogs after recovery from canine distemper virus continue to harbour the virus in brain cells, where it replicates slowly and eventually produces the disease known as old dog encephalitis. The virus can be readily cultivated from the brains of the affected dogs.
Slow infections
.The term 'slow infections' is used to .describe such infections which have large incubation perioo and cause a slow progressive disease which is invariably a fatal disease. The virus can be recovered from infected animals during incubation period as well as during disease. Lentiviruses as well as certain unclassified agents produce subacute spongiform encephalopathies.
VisnalMaedi: Vlsna/Maedi virus belongs to family retroviridae and subfamily lentivirinae. These viruses cause chronic demyelinating disease of central nervous system, chronic pneumonia in sheep and chronic encephalitis and arthritis in goats. These viruses cause persistent infection, mainly in circulating leukocytes and virus may also exist as DNA provirus. An interesting feature of Visna/Maedi and equine infections anaemia viruses is the antigenic drift in surface proteins during the progress of infection in a single animal.
Subacute spongijorm viral encephalopathies: Subacute spongiform viral encephalopathies are produced by five diseases with similar clinicopathological features and causative agents. The diseases are scrapie of sheep and goats, mink encephalopathy, wasting disease of deer and elk, and kuru and Creutzfeldt-Iakob disease in humans. The basic lesions i;; fl progressive vacuolation in neurons, an extensive astroglial hypertrophy and proliferation, and spongiform change in the gray matter. The scrapie infection of sheep is transmitted from mother

Persistent Infections 79
to lamb. The incubation period is very long, upto 3 years. When the clinical symptoms appear the disease progresses slowly leading to paralysis and death. The mice and hamsters can be infected experimentally. _The incubation period is about 1 year in these experimental animals. The disease is like any infectious disease, the causative agent appears to be the size of small virus but there is absence of any immune response and no effect of interferon. The scrapie agent has a higher degree of resistance than conventional viruses. These biological and physiochemical properties are also shared by agents of other four subacute spongiform encephalopathie~.
Pathogenesis of persistent infections
There are several mechanism which bypass host defenses that eliminate virus in acute infection. These factors are primarily related to the virus on one hand and to the host defenses on the other, althogh the two kinds of factors interact in some instances. The possible mechanisms are detailed below:
Nonimmunogellic agents: The agents producing subacute spongiform encephalopathies are uncharacterized agents which appear to be non immunogenic, which do not induce interferon and also are not susceptible to interferon action. It shows that the host cannot control the replication and pathological effects of these agents.
Integrated genomes: The proviral DNA of retroviruses is integrated and maintained from one generation to the next as part of host genome. Such proviral DNA may be implicated in tumorigenesis. In lentiviral infections provirus is not involved but viral infection causes systemic disease.
Growth in protected sites: During latent phase most of alphaherpes-viruses avoid immune elimination by remaining within the cells of the nervous system, as DNA in ganglion cells during the intervals between disease episodes and within axons prior to acute recurrent disease episode. The beta and gamma herpes viruses persist in lymphocytes and avoid immune elimination.
Antigenic variation: Certain retroviruses avoid hosts immune mecha-nism by antigenic drift. During persistent infection, Visnal Maedi and equine infectious anaemia viruses develop succession of antigenic vatiants. These successive antigenic variants produce clinical signs in cycles. The persistence of influenza virus in populations occur

•
80 Textbook of Veterinary Virology
by similar mechanism operating over a long period in succession in animal hosts.
Modification of host defence mechanisms
The modification of immune response is achieved by several ways. These may be ineffective antibodies, disturbance of function of cells of immune system, avoidance of immune lysis of infected cells and antigenic variation of the virus.
Defective antibody response: The viruses which cause persistent plasma associated viraemia replicate in lymphoid tissue and mactophages and induce production of nonneutralizing antibodies: These antibodies produce immune complexes and cause 'Immune complex disease' and also block immune cytolysis of virus infected target cell by T cells. Persistent infections in congenital infections also cause immunological tolerance.
Defective cell mediated immunity: The persistent infections may be caused by partial suppression of hosts cell mediated immune response as a result of many factors.
Growth in macrophages: In many chronic infections virus appears to grow in macrophages. This causes impairment of humoral and cell mediated immune response and impairment of phagocytic and cytotoxic activities of reticuloendothelial system.
References
GIBBS, E.PJ., 1981. Persistent viral infections offood animals: Their relevance to international movement of livestock and germ plasm. Adv. vet. ScL Comp. Med. 25, 7l.
Hn..Ls, T.J., 1985. Herpes simplex latency. In ''The viruses" Vol. 3 p. 175, Plenum New York.
MARY, B.W.J., 1985. Strategies of virus persistence. Br. Med. Bull. 41,50 MIMs, C.A.; CUZNER, M.L.· and KEU..y, R.E. eds. 1984. 'Viruses and
demyelinating disease'. Acadenuc Press, London. NOTKlNs, A.L. and OLDSTONE, M.B.A., eds. 1984. 'Concepts in viral
pathogenesis' Springer-Verlag, New York. PRUSINER, S.B., 1982. 'New proteinaceous infectious partides cause scrapie'
Science 216,136-144. WITfMANN, G.; GASKELL, R.M. and RZIHA, H.J., eds. 1984. 'Latent herpes
virus infections in Veterinary Medicine' Curr Top. vet. Med. Anim. Sci. 27,1

Chapter 8
Viral Imnlunity
The viruses are a group of organisms which must entcr a host cell to proliferate, since they lack the necessary biochemical machinery to manufacture protein and metabolize sugars. Some viruses also lack the enzymes required for nucleic acid replication and are dependent on host cell for this function also.
The illness produced by virus infections are as varied as the viruses themselves. Illness may be acute, recurrent, latent (i.e. dormant infection where the virus is not readily detectable but may recur) or subclinical i.e. acute or chronic symptomless infection where the virus is demonstrable.
The immune response may range from the apparently non existant, of Kuru, to lifelong immunity or chronic immunopathology (e.g. Hepatitis B).
In this chapter discussion will concentrate on those acute virus infections, which usually evoke obvious immunity, since these are the only ones about which there is reliable immunological data. It must therefore be remembered that apart from an assortment of clinical and clinico immunological observations, we have litttle undcrstanding of immunological mechanism underlying the recurrent or latent or lifelong subclinical virus infections.
Viral infection: A typical viral infection starts with local invasion of an epithelial surface and they after one or more viraemic phases results in infection of target organ. III the early stage of viral infection host response is largely non antigenic. spel.ific and involve!; the production of interferons (IFN's) and activating natural kiiier cells.

82 Textbook of Veterinary Virology
Once virus starts replicating the following immune responses are triggered in the second phase production of humoral antibodies; activation of regulatory T cells. (that can amplify or suppress effcctor functions) and T cell mediated immunity effected by cytotoxic T cells that are able to lyse virus infected cells or T cells that are able to induce delayed type hypersensitivity. All this depends upon the species of the virus and phase of infection. The relevance to protection or immunopathology of the various effector systems of immune response therefore depend on the phase of infection and on the biology of the virus. Viral antigens are largely proteins or giycoproteins. The glycoproteins are often glycosylated by the host cell during the budding process. The internal antigens of the virus are not usually relevant to protective immunity. Antigens which are expressed on surface of the virion may be the potential targets for the immune response and so antigens expressed on the membranes of infected cells.
The response to a viral antigen is almost entirely T cell dependent. Even the antibody response requires T cells help.
Humoral response
Antibody is only capable of directly binding to extracellular viruses. IgG and IgM antibodies are limited in their action to plasma and tissue fluids, whereas secretary IgA may protect epithelial surfaces and, therefore, is particularly important in protccting against viruses which lack the viraemic phase. Antibody in association with complement can cause lysis of cells carrying viral antigens or directly damage enveloped viruses. Antibody dependent cellular cytotoxicity is also very effective mechanism for killing virus infected cells which are recognised by the presence of viral antigens on their membrane.
Effects or antibody
Antibody may upset the virus cell interaction which lead to adsorption, penetration, uncoating and replication, for instance following into phagocytic vacuole some viruses have envelopes which interact with vacuolar membrane and cause the dissolution. The viral nucleic acid is liberated into the cytoplasm. However, the essential interaction with vacuolar membmne can be blocked if the virion is coated with antibody. On the other hand, mice artificially made agammaglobulinaemic, which produce no detectable antibody to

Viral Immunity 83
haemagglutinin, recover and are subsequently immune. Thus T cell cesponses appear to be important.
Antibody dependent cell mediated cytotoxicity
Cells with cytotoxic potential, which also possess Fc receptor for IgG may bind to and lyse target cells coated with antibody to relevant class. These cells act by binding specific antibody on virus infected target cells via their Fc receptors. They can specifically kill the virus infected cells.
Cell-mediated immunity
In general it must be seen that protective immunity to a pathogen is mediated by both cellular and hU'lloral immunity. Also the predominant role of CMI versus humoral immunity can be ascribed to a variety of pathogens but not to the mutual exclusion of one to the other. Infact most immune responses in recovery from infectious agents involve the interplay of both humoral and CM! compartments. Major functions of CMI are to defend against pathogens that live faculative/obligatory within the host cells (viruses, some protozoan, bacteria, fungi). A role for CMI is also suggested in tumour immunity and other immunological phenomenon such as allograft rejection, graft versus host reaction.
The components of CMI consisting immune lymphocytes that act directly i.e. cytotoxic T lymphocyte (CfL or Tc) and immune T lymphoytes primarily TDTH cells, possibly Tc accessary cells (mostly monocytes also release lymphokines that recruit and activate macrophages and Nk c~lls) to provide effective immune functions independent of antibody. Both T cell effector functions (Tc and TDTII) are regulated through TH and Tc circuits. These regulatory T cell effectors (TH and TS) are also important in B cell antibody response.
Cytotoxic T cells
The erc is component of the cellular arm of the immune response whose chief function appears to be the destruction of virus infected cells an important means by which host recovers from infection. The erc are particularly important where the load of infection is high.
Cytotoxic T cell are anti genic specific usually belong to OKT8 subSt~t and are restricted to class I MHC molecules, however virus

84 TeXlbook o/Velerinary Virology
specific killer T cells of OKT4 helper inducers subset have also been detected in long term cultures. Interferon gamma and other ]ymphokines are produced by CTC and T4 after stimulation by virus infected cel1 of influenza virus which activate macrophages, stimulate growth and maturation of proginator cells and antibody forming B cells.
In Tc responses there is requirement for endogeneous antigen synthesis pathway, where the antigens are normally presented in association with MHC-l molecules, perhaps because body cells that are natural infection targets having MHC-l molecule. An exception however in measles virus system for example MHC-II restricted Tc are important in immunity and are induced by endogenous antigen synthesis pathway.
Perhaps the most surprising aspect of CTC immunobiology has been the recent observation that with some viruses the principal target antigens are not the transmembrane viral glycoproteins found abundantly at the cell surface. While some but not all transmembrane proteins can act as CTC target In influenza virus system, thus so far the most compre-hensively studied, internal viral proteins and even non structural proteins act as the major target antigens. The advantage of this may be recognition of early proteins that provides the host with means of destroying infected cells before the completion of a replication cycle and spread of new virus to neighbouring cells.
Delayed type hypersensitivity: In contrast to elL class 11 restricted T cel1 do recognise viral glycoproteins as principal targets. These glycoproteins are usually late proteins and are expressed not only on the infected surface but also on the surface of virion. Thus, they form a logical target for antibodies that act primarily in extracellular defence capacity. In addition Class II restricted cells subserve a protective role by mediating a non specific inflammatory response which in turn effect immunity. The essential feature of TDTH component of CMI is that immunity is mediated primarily through various lymphokines which are released on interaction between immune T cells and antigen. These lymphokines recriut and activate accessary cells primarily macrophages and NK cells. Activated accessary cells nonspecifically can destroy or limit the intracellular pathogen (virus, bacteria, protozoa) and neop]astic cells. Exogenous pathway of antigens presenting cells is required for induction TDTH response. Different lymphokines may be

Viral Immunity 85
produced by different lymphocyte subsets. Some lymphokines such as tumour necrotic factor (TMF), lymphotoxin (L 1) act directly to destroy the target cells. Others act an macrophages to recruit/assist (MCF), immoblise (MIF), activate (MAF/gamma IFN) macrophages and activated macrophages are very potent non specific effectors of CM! response to eliminate viral infected cells.
Amongst various lymphokines released by immune TDTH is transfer factor (TF) which is an interesting lymphokine that has potential both for non infectious form and immunoprohylaxix and immunotherapeutic.
Non-specific immune response
Interferons are family of related' cell regulatory glycoproteins produced by many cell types in response to virus infection, double stranded RNA, endotoxin and a variety of antigens and antigenic stimuli. Interferons released from virus infected cells binds to receptors on neighbouring cells and induce an antiviral state which helps to isolate infective foci. The mechanism may involve inhibition of viral protein or nucleic acid synthesis. IFN also particularly inhibits cell growth (suggesting a possible anti tumour activity) and exerts selective effects on protein synthesis and immune response. Thus interferon may contribute to decrease in cell mediated responses seen early in virus infections.
Immunopathology
The immune response to viruses can cause damage to host via the formation of immune complexes or by direct damage to infectf'.d cells. Complexes can form in fluid phase, or following capping and stripping of virus expressed on cell surfaces. Chronic immune complex glomerulone-phritis can occur in mice infected neonatally with lympho choriomenigitis virus (LCM).
References
SISSOMS J.O. and OOLDSTONE, M.B.A., 1980. Antibody mediated destruction of virus infected cells. Adv. Immumol. i, 311-331.
ZINKER:'lAGEL, R.M. and DoHERTY, P.C., 1979. MlIC restricted cytotoxic T. cells. Studies on biological role of polymorphic major transplantation antigens determining T cell restriction specificity, function and responsiveness. Ad". Immuno!. 27, 51·177 .

86 Textbook o/Veterinary Virology
DENAM, A.M., 1983. Viruses and immurwpathology. In Immunology in Medicine Halbrow E.J. and Reeves W.G. (eds) Gurune & stratton.
Rous B.T. and HAWHOV, D.W., 1984. Cytotoxic T lymphocytes in herpes virus infections. Vet. Immunol. Immunopalhol. 6: 35-36.
NOTKINs, A.L., 1975. Viral immunology and immurwpathology. Academic Press, New York.

Chapter 9
Epidemiology of Viral Diseases
The science of epidemiology is the study of factors affecting the health and disease in an animal population or group of animals. Epidemiology unravels the mechanism of disease distribution, transmission, survival of viruses in animal population and their control. The viruses are intracellular organisms, different families of viruses use different mechanisms for their transmission and survival. Many virus families span both vertebrate and invertebrate animals. At cellular level the viruses depend upon multiple cell structures including the membranes and nucleic acid. The epidemiological studies enable us to:
1. Determine the magnitude of the disease including the locations, time of outbreak, as well as the age, sex and breed of ammals affected in relation to population at risk;
2. Indentify etiologic agents by isolation and serologic studies; 3. Plan control measures such as quarantine, sanitation of the
environment, slaughter of affected and incontact animals besides immunization of susceptible population at risk and
4. Evaluation of effectives of control measures by statistical analysis.
Epidemiologic tools
The tools used by epidemiologist to study viral diseases arc virologic, serologic, molecular and ecologic. In the itlitial stages the only way of studying the transmission of viruses wa<; !hI;: homologous hosts and later on the experimental hosts like monkeys, mice, baby mice, guinea pigs and chicken embryos were found to be .u$Cful. The

88 Textbook of Veterinary Virology
cell culture method proved to be very useful and opened up extensive epidemiologic approaches. The approaches like antigen detection, detection of nucleicacid and electron microscopy have paved the way for epidemiologic study of those viruses which are not easily propagated.
The serology is as important to epidemiologist as the isolation of the virus. The serum antibodies appear within a week after infection with most of viruses. The serologic tests to detect these antibodies are neutralization, complement fixation, haemagglutination-inhibition, gel diffusion and enzyme linked immunosorbent assay.
In the recent past molecular techniques have been developed and are of importance in epidemiological studies. The monoclonal antibodies are being used to distinguish rapidly and unambiguously between closely related viruses. The detection of nucleic acid is another sensitive tool in detection and identification of viruses. One of the potential and sensitive method used for identifying fine differences among virus strain is nucleic acid hybridization. By using these molecular methods the virologists have uncovered the modes of spread and true life cycles of many viruses.
The transmission of viruses depends on the interaction of the virus with host and this interaction is modified by host behaviour and environment. The ecology surrounding each virus is different. The viruses use fascinating, complicated and ingenious mechanisms for their survival, transmission and causation of disease.
Virus host environment interactions
The clinical manifestation of disease due to viral infection is the result of a dynamic interaction among the causal agent, the host and the environment. These factors cannot be singled out in explaining the epidemiology of a virus disease.
Viral agent
The major factors in viral diseases are virulence, transmissibility of persistency of infection.
The virulence i.e. the ability of viral agent to produce disease, is an inherent characteristic of the viral agents. Some virus strains are more pathogenic to a population than others. Transmissibility of the virus is also related to virulence.

Epidemiology of Viral Diseases 89
Host factors
The host factors which play a part in the epidemiology are age, breed, sex and immunity status.
The age is most important factor which plays an important role in viral diseases. In certain viral infections like neonatal diarrhoea due to rotavirus and infectious bovine rhinotracheitis, young calves are more susceptible. The age factor is also relevant to acquisition of immunity through consumption of colostrum etc.
Certain breeds of animals show more susceptibility to some infections. For example, hill cattle are more susceptible to rinderpest in comparison to cattle from plains in India. The sex has little bearing on viral epidemiology.
Immunity is an important factor which determines the outcome of the disease. Protection is gained by active as well as passive immunity besides non-immunologic resistance. In a herd, the immunity wanes at different intervals with the result that the probability of disease introduction also rises and falls. The population immunity is influenced by the introduction of new animals or new borns
Environmental factors
The environment influences both the causal agent as well as the host. The important environmental factors are discussed below.
Population density: Extreme crowding plays an important role in determining the mortality rates due to viral infections especially among young animals. The range cattle have a lower mortality rate than calves in feed lots.
Nutritional level: The direct effect of status of level of nutrition in viral diseases is not clear. The other environmental factors which influence viral epidemiology among animals are managerial practices, immigration etc.
Perpetuation of viruses in nature
The viruses can survive outside the host only for a limited period. For the perpetuation of viruses, repeated infectious cycles in susceptible hosts are essential. It is not always necessary that virus infection will result in clinical disease. The immune status of the host and viral pathogenesis are also important factors in the perpetuation of viruses in nature in case of certain diseases. In certain disea~s like foot and mouth disease, carrier animals also act as a source of spread of infeclion to susceptible livestock.

90 Textbook of Veterinary Virology
Perpetuation by short cycle
Certain viruses are highly contagious but their transmission period is usually short. A new susceptible population is required to maintain the continuity of the virus cycle. Some of the important diseases in this group include,rinderpest, foot-and-mouth diseases, Newcastle disease, canine distemper etc. Due to short life span, the turn over in animals is much higher than that in man. This change in popUlation results in higher availability rate of susceptible animals.
There are certain viruses which are resistant and can survive for a long time in the external environment like fowl pox virus, foot and mouth disease virus.
Many viruses which are responsible for chronic infections, perpetuate in the hosts and are transmitted to new hosts continuously for a long period. Examples are herpes and adeno viruses, rabies among bats etc.
The perpetuation of arthropod borne viruses by insects is the usual mechanism of overwintering. Some of the viruses may perpetuate by congenital transmission like, leukemia, hog cholera, bovine viral diarrhoea etc.
The transmission of viruses
The spread of viruses from infected to healthy animals depends upon the mode of shedding of viruses by the affected animals. The major routes for transmission of viruses are:
1. Contaminated skin or mucous membrane 2. Droplet infections originating from respiratory tract 3. Alimentary tract-faecal or oral transmission 4. Genitourinary tract 5. Vertical 6. Horizontal
1. Contaminated skill or mucous membranes: The continuity of the skin must be broken for viruses to establish infection via skin. The break in skin, or mucous membrane may be accidental or may be inflicted by insects or other animals. The accidental damage to the foot may be due to hard ground (foot and mouth disease) while rough fodder may break the mucous membra!le of the mouth (foot and mouth disease virus, papilloma viruses). Damage to teats by milking or sucking facilitates establishment of certain viruses like orf (contagious

Epidemiology o/Viral Diseases 91
ecthyma) virus in sheep and herpes mammalitis virus in cattle. Bites from rabid dogs and bats transmit rabies virus. The mosquitoes, ticks, culicoides transmit orb iviruses, togaviruses, bunyaviruses, where the viruses by biting insects also take place like fowl pox virus and myxomatosis virus in rabbits.
2. Respiratory tract: The particles of 0.5-0.7 m size are optimal for droplet infection because smaller than this size dry out while larger ones fall out. Primary replication usually takes place in the nasal epithelium but high concentration of virus results in primary inhalation into the bronchi and lungs. Viruses gaining access through this route spread only within respiratory tract like influenza, parainfluenza, pox and respiratory syncytial viruses in cattle and those that disseminates via circulation to other sites like distemper virus in dogs and Marek's disease in pOUltry. There are certain viruses which may be confined to respiratory tract or disseminate according to virulence e.g. Newcastle disease virus.
3. Alimelltary tract: The transmission of viruses by alimentary tract is from fomites or faecal contamination. TIle primary site for these viruses is usually oropharynx epithelium like foot and mouth disease virus. Only those viruses which are resistant to acid pH of stomach as well as the effect of bile proteolytic enzymes can establish a primary infection in the intestine. These viruses are picorna, rota and parvoviruses.
4. Gellitourinary tract: The genital tract is a common route of infection for certain viruses like herpes viruses, sheep pox etc. Viraemia leads to transplacental infection. Immunotolerant excretors are important in perpetuating the virus infection like border disease of sheep and mucosal disease in cattle.
Incubation period and dissemination of virus
The incubation period is defined as the time taken from the entry of the virus to the appearance of pmhogenomic symptoms. During this period the virus replicates and spreads from primary to secondary site of infection. The incubation period is generally measured in days but in slow viruses like VisnaIMaedi the incubation period may be months or years.
The main route of dissemination is via the circulatory system as free or cell-associated virus. The virus liberated by the lysed cells enters via lymphatic or directly into blood circulation to give rise 10 viraemia e.g. fOOL and mouth disease virus, Newcastle disease virus etc.

92 Textbook o/Veterinary Virology
The free virus or infected cells may be ingested by macrophages. The macrophages with the infectious virus transport to the local lymph nodes or other secondary sites, e.g. distemper virus, rinderpest virus, herpes viruses etc. The viruses may replicate in lymphocytes leading to leucopenia.
Mode
Intimate contact
Airborne droplets
Oral transmission
Genito Urinary Vertical
HOrizontal) Vertical Transpla. cental By arthropod vectors like mosqlli-toes Culicoidis etc 3) Mechanical
transmission b) Biological
transmission
Table 9.1 TRANSMISSION OF ANIMAL VIRUSES
Viruses Infective material
Pox viruses Dried, scales, vesicle, fluid etc. Herpes viruses Papova viruses Influenza Nasopharyngeal Rhinoviruses Aphtho viruses Coma viruses Discharges, Saliva Adenoviruses Entero viruses Faeces. Saliva Adenoviruses Wat~ ] Rota viruses Food Contaminated Astro viruses Flies Intestinal corona Herpes viruses Semen and other excretions
MTV LCM
Hog cholera, Blue- Blood tongue
Fowl pox. rabbit Blood, secretions and excrctions myxoma from affected animals Alpha viruses Flavi viruses BunyaviIidae -do-and Reoviruses
(Orbiviruses)

Epidemiology of Viral Diseases 93
References
BROWN, F. and WILSON, G., 1984. Principles of Bacteriology, Virology and Immunity. Vol. 4. Edward Arnold.
FENNER, F.; McAuSLAN, B.R.; MIMS, C.A.; SAMHROOK, J. and WHITE, D.D., 1974. The Biology of animal Viruses. Academic Press, New York.
KAHRS, R.F., 1985. Viral diseases of cattle. Iowa University Press.
MOHANTY, S.B. and DlITfA, S.K.. 1981. Veterinary Virology. Lee and Febiger. Philadelphia.

Chapter 10
Viral Thmorogenesis
Tumor viruses are known to transform cells in vitro and induce tumors experimentally in animals while some tumor viruses naturally produce tumors in animals. The studies on tumor viruses have led to the identification of tumour viruses and cellular homologous of several retrovirus oncogenes that appear to play a role in human cancer. The tumor viruses replicate their genomes and express their genes using cellular enzymes. A large number of DNA viruses naturally infect several animals and human beings and transform cells in culture or induce tumors in experimentally infected animals. These viruses arc DNA tumor viruses and belong to five families given as under:
i) Polyomaviruses: simian virus 40 (SV ~, mouse polyomavirus (PY) and human viruses.
ii) Papilloma viruses: animal papillomaviruses, human papilloma -viruses.
iii) Adenoviruses: simian, bovine and human adenoviruses. iv) Herpesviruscs: primate, equine, chicken, frog, rabbit and
human. v) Hepatitis-B like viruses: human hepatitis B virus, Woodchuck
hepatitis virus; ground squirrel hepatitis virus and duck hepatitis B virus.
The genomes of DNA tumor viruses have a wide range of size Polyoma and Papilloma viruses have a small circular duplex, superhelical genomes of about 5.3 and 7.9 kilobases (Kb).
The genomes of adeno and herpes viruses are larger and consist of linear duplex molecules of 30-38 Kb and 120·240 Kb, respectively. The genome of human hepatitis B like viruses, consist of duplex

Viral Tumorogenesis 95
molecule of about 3.2 Kb that contains a single stranded gap. The members of polyomavirus, adenovirus and herpesvirus exhibit two life styles (a) productive infection of cells that are permissive for virus replication and death of the cell and (b) transformation with a low efficiency of small proportion of cells that are nonpermissive for virus replication. The cells productively infected with polyoma, adeno and herpes viruses express the entire viral genome while the cells transformed by these viruses and papilloma viruses express only a subset of viral genome that include the transforming genes. For studying the productive infection of papilloma viruses there is no suitable cell culture system. The viral genome that encodes viral transforming genes is small. Cell transformation is the result of viral DNA integration into cellular DNA and expression of specific viral genes encoded in the transforming region. No specific sites of integration on the host cell genome or viral genome have been identified in DNA viruses.
The cell transformation by polyoma viruses as well as by adenoviruses is a multistep process, which involves at least two viral coded functions (i) a cell immortalization or 'establishment' function and (ii) transformation function required for the expression of fully transformed state.
Polyomaviruses: Polyomaviruses(SV 40' PY, JCV) transform cultured cells and induce tumors in experimental animals but they do not induce tumors in their native host. The transformation region of SV 40 codes for large and small T antigen. The expression of large T antigen is required for initiation of cell transformation and the continued expression of large T antigen is required for maintenance of transformed phenotype. The expression of small T antigen is not essential for cell transformation but plays an enhancing role under certain conditions. PY codes for large, middle and small T antigen. The expression of middle T antigen is essential for the initiation and maintenance of transformation of primary cultures, both middle T antigen and the N-terminal half of large T antigen are required for transformation.
Popillomaviruses: Papilloma viruses in contrast to polyoma viruses and Ads produce cancer in their host species. Virus replication is regulated by the differentiated state of host cell. The papilloma virus genom~ is maintained in the transformed cells as free replicating episomal viral DNA molecules and not as integrated viral DNA as in

96 Textbook a/Veterinary Virology
the case with cells transformed by polyoma viruses and Ads. Adenoviruses: All human Ads have the capacity to transform
rodent cells in vitro but only members of groups A, B and D induce tumors experimentally in animals. The Ad transforming genes are located in the left 11 percent of viral DNA genome.
Herpes viruses: Alpha, beta and gamma subfamilies of herpesviruses transform cells in culture but only gamma herpersivruses induce tumors experimentally in laboratory animals. Several herpesviruses cause cancer in their original hosts, e.g. Marek's disease of chickens, Lucke's carcinoma of frogs. In human beings there is epidemiological evidence that Epstein-Barr virus(EBV) is responsible in the development of Burkitt's lymphoma(BL) and nasopharyngeal carcinoma(NPC). The genomes of EBV are found in BL and NPC tumors. The cells of BL tumors exhibit a translocation of chromosome 8 to one of the three chromosomes. In each case the human Cmycogene is located at the cytogeI1etic breakpoint. It may be possible that C-mycogene plays a role in multiple carcinogenesis that is initiated by infection of B lymphocytes with EBV.
In comparision to polyoma and adenoviruses, very little is known about the putative transforming genes and transforming proteins encoded by members of herpes virus group. Recent studies identified regions of herpesvirus genome that possess transforming activity. The viral DNA sequence is not detected in the transformed cells. However, the cells transformed by e.quine herpesivrus type 1 have been shown to retain a specific region of the viral genome.
Related funtions among transforming genes of DNA tumor viruses and oncogenes of retroviruses and cancer cells
The recent studies have provided an evidence that cell transformation induced by DNA tumor virus transforming genes, by oncogenes of retroviruses and cellular oncogenes cloned from human cancer cells is a multistep process that involves 2 or 3 biological functions. The first function is the immortalization of primary cells. The immortalization function is expressed by N-terminal portion of the PY and SV 40 large T antigen genes by the AD EIA genes and the myc oncogene. The myc oncogene has been found in several types of human tumors. The second class of functIon is known as transformation function. This function is expressed by PY middle T antigen gene, AD

Viral Tumorogenesis 97
EIB transformation region and ras oncogenes. Transforming proteins are generally found on cellular membrane structures. The third function in oncogenic transformation can be supplied by spontaneous cellular change or by transforming gene function encoded in some DNA tumor virus genomes.
Common pathways of oncogenesis and cell transformation
The cells transformed by tumor viruses possess new properties. The origin of transforming genes of DNA tumor viruses is not known. However, the transforming genes of retroviruses(oncogcnes) appear to have evolved from cellular genes, since cellular homologous of retrovirus oncogenes are found in numerous vertebrate species. In recent observation it has beeen found that the portions of Ad El <\ transfomring genes are structurally related to two retrovirus oncogenes myc and myb. It is interesting observation because myc and EIA gene products are both nuclear proteins that immortalize primary cells.
DNA tumor virus transforming genes and retrovirus oncogenes encode interchangeable and complementary functions. The oncogenic activity of human cancer oncogenes can be complemented by DNA tumor virus transforming genes. This suggests that DNA tumor virus transforming gene encode functions that mimic those of cellular genes.
Tumor virus genes or oncogenes that encode the transformation function have no effect on primary cells but they often transform established cell lines by introduction of viral or cellular gene encoding the immortalization function, spontaneous mutation or exposure to chemical carcinogens. The PY middle T antigen cannot transform primary cells but it can transform established cell lines or cells that have been immortalized previously by PY large T antigen. The Ad EIA function can substitute for the PY large T antigen. Thus the immortalization gene of one DNA tumor virus can wmplement the transformation gene of second DNA tumor viIUs. The immortalization function of DNA tumor virus genes and retrovirus Myc oncogene can complement the transformation function of viral and cellular oncogenes.
Recent studie,s have indicated that same cellular genes may be activated by DNA tumor viruses, by retroviruses and by chemical carcinogens. This suggests that there are limited number of pathways to
oncogenesis and aberrant regulation of smali group of cellular genes

98 Texthooko/Veterinary Virology
may be the basis of malignancy. The analysis of the libraries of CDNA clones prepared from S V 40 transfomed cells indicated that some cell mRNA species are differentially expressed in SV40 transfored cells.
Retroviruses
Relroviruses induce a variety of neoplastic disease in vertebrate hosts. The retroviruses have single stranded RNA viral genome and RNA dependent DNA polymerase (reverse transcriptase). This virus family includes: Type C oncovirus-mammalian type C oncoviruses; Avian type C oncoviruses; Type B oncovirus-mouse mammary tumor virus; Type D oncovirus-squirrel monkey-virus.
Retroviruses have been extensiVely studied and have been molecularly cloned and complete sequence of viral genome is known. These viruses possess a number of unusual properties as compared to most other viruses. Retroviruses typically establish a chronic infection in susceptible host cells. To induce the chronic infection, the viral genome has developed a unique strategy for replication. The virus replicates in host celI through a double stranded viral DNA intermediate which is integrated into host cell DNA. Another feature of the replication of retroviruses is that certain viruses behave as cellular genes and they are carried in the germ line as endogenous proviruses (integrated viral genomes linked to host cell-DNA). Certain other retroviruses are spread horizontally e.g. human T-cell leukemia virus (HLTV) which is associated with an adult form of human lymphoma! leukemia. Some of the retroviruses carry genes which are closely related to cellular genes and these genes endows the virus with the capacity to induce transformation of tissue culture cells and enhance the tumorigenic potential of the virus. These genes are called as transforming genes or oncogenes (V-onc) and their cellular counterparts are known as C-onc. Each one gene has been given a 3 letter name such as Src, Ras etc. The comparison of V -onc gene and Cone gene indicate that virus has transduced the C-onc gene with minor genetic changes and placed it under viral regulatory elements.
Virus properties
Retrovimses are 80-130 nm in diameter and consists of an inner electron dense core (nucleoid) surrounded by an envelope composed of lipid containing unit membrane. The core consists of nucleoprotein

ViralTur.norogenes~ 99
surrounded by icosahedral protein capsid, an outer coat core protein is located between capsid and the envelope. The envelope has glycoporteion protections or spikes with knobs. The infectivity is extremely sensitive to lipid solvents and detergents and at 56"C for 30 min. Virions contain the reverse transcriptase. The viral RNA genome is composed of two positive sense single stranded RNA. The two ~A subunits are bound noncovalently to each other near their 5' ends. A specific cell coded tRNA is essential for virus replication. The presence of two viral RNA copies indicates that retroviruses are diploid.
Retroviruses are divided into 3 subfamilies: (i) oncoviruses, (ii) lentiviruses (lente means slow) and (iii) spumaviruses (spuma means foam).
The oncoviruses have been isolated from almost all vertebrate species. The members of the subfamily are intially grouped according to the host species from which they have been isolated and then by morphogenesis of their virions (Type A, B, C or D). Most of these viruses belong to C-type viruses. The C-type viruses develop as they bud from plasma membrane. The mature-C type particles are 80-100 nm with a centrally placed core inside the envelope. The B-type particles develop their inner core in the cytoplasm and acquire their envelope as they bud from plasma membrane. The mature particles measure about 125 nm and contain an eccentrically located electron dense core. The D-type particles develop in a similar manner as B-type. However, the mature D type particles resemble C type particles and are more closely genetically related to C-type viruses. A type particles are found intracellularly and are devoid of infectivity. These particles lack of a lipid containing outer envelope because they do not bud from the membrane. There are two types of A particles intracisternal and intracytoplasmic. The intracisternal forms are derived from endogenous viral genomes and are apparently defective. Some intracytoplasmic A type particles are precursor forms of B & D type particles.
The retroviruses have a simple structure and genetic organisation. There are four identifiable genes-those that are essential for virus replication and V -onc genes, which enhance the oncogenecity of virus but are not essential. There are only three viral genes whIch are required for viral replication: gag for group specific antigen and encodes core protems, pol(polymerase) encodes for reverse ti1lnscriptase and env which encodes for viral envelope proteins. Their

100 Textbook o/Veterinary Virology
5' to 3' order is gag-pol-env. The retroviruses containing V-onc gene are replication defective: since cells infected with a V -onc containing virus will overgrow those cells not infected by this type of virus. Transfonnation competent defective viruses are always isolated as one component of mixed virus population that also contains a replication competent retrovirus. Certain isolates of RSV are both replication competent and transformation competent. The viruses that contain Vonc genes, most carry only one V-onc, although some possess two. The V -onc gene may be located in different regions within the viral genome.
Most retroviruses demonstrate a high degree of specificity in the range of host cells that they can successfully infect. The transmission of many retroviruses are acquired by horizontial spread. These viruses are known as exogenous. Some retroviruses are transmitted vertically as heritable pro-viruses whose genomes are localised to different chromosomes. These viral genomes are called as endogenous. Under natural conditions, most endogenous viruses have low pathogenicity and are widely distributed. The endogenous viral genomes represent an opportunistic parasitism of the host genome. The endogenous proviruses are usually less oncogenic for the animal in which they are found than are exogenous viruses.
The retroviruses have a high frequency of genetic recombination during coinfection with two closely related viruses. The retroviruses have a capacity to integrate their genomes at multiple sites within the host genome which suggests that these viruses may be capable of inducing mutagenesis of host encoded genes via insertional mutagenesis. In vitro insertional mutagenesis has also been demonstrated by using exogenous infection by Moloney leukemia virus in rat cells.
Palhogenesis: The V -onc viruses induce disease after a long latent period. These viruses usually induce one or several fonns of malignacy but infection with some retroviruses leads to some wasting and neurological disorders. In AL V infected chickens the disease is induced after a latent period of several months despite continued high levels of virus. It suggests that either the target cells are rarely infected by AL V or the turnors arise as a multistep genetic process. The recent studies lent a support to latter possibility. The endogenous viral loci and host genes also have a significant influence on the outcome {If virus

Viral Tunwrogenesis 101
infection. High levels of virus with env glycoprotein are required for tumor formation.
The transformation competent (V-Onc +) retroviruses are found frequently in tumor tissues of many species but are isolated rarely. These viruses are isolated as a mixture with the helper independent virus that has transduced the V -Onc gene. Under natural conditions VOnc + viruses are not usually transmitted to second animal. The V -Onc + retroviruses induce a characteristic range of tunors after a short latent period. The V -onc gene is required for tumor induction but role of other viral sequences is not yet known. The C-onc genes from which V-onc genes are derived are conserved in eUkaryotic cells. Initially C-Onc genes were discovered by finding the normal cells of many species which contained sequences and proteins homologous to V -Onc genes and their protein products. It is speculated that C-Onc genes are normally involved in cell growth, division and differentiation. The coding sequence of almost all C-onc genes are segemented. The recombination mechanism by which retroviruses transduce C-onc gene is not known. It is likely that integration of viral DNA near C-onc gene is involved. Inspite of the fact that C-onc gene and V -onc gene are similar at aminoacid level, C-onc genes carries out some important normal cell functions while V-onc genes are highly oncogenic. There are three possible explanations for the oncogenic property of V -onc genes: (i) increased expression of a normal gene product (ii) mutation within V -onc gene coding sequences and (iii) the contribution of viral protein coding sequences. The importance of mutations within the Vonc protein coding sequences for its oncogenicity has been shown for ras genes.
References
FENNER, FRANK, 1987. Veterinary Virology. Academic Press, London.
TEDDER, R.S.and ROBIN, A., 1990-ln Principlies of Bacteriology, Virology and Immunity VolA, edited by LH collier and Morage. Timbury, Edward Amold, London.
GREEN, MAURICB, 1985. In Fields Virology edited by Bemard N.Fields et al. Raven Press, New York.
LowRY, D.R., 1985.1" Fields Virology edited by Bemard N. Fields et al. Raven Prp-ss New York,

Chapter 11
Virus Vaccines and Antiviral Agents
The reduction of economic losses due 10 viral diseases among the domestic animals in the recent past has been due 10 management practices, hygiene and sanitation and immunization. The most important of these measures is by vaccination and this has brought down the losses to great extent in animals. The sanitary methods of control of virus infections have often been applied with great success e.g. control of rinderpest, swine fever, r~bies, foot and mouth disease in United Kingdom accomplished by Slaughter of affected animals.
The history of the development of vaccines is about 200 years old. The first vaccine was used by Edward Jenner in 1798 against small pox; about a century before the is solation of viruses. The next viral vaccine was developed by Louis Pasteur in 1884 against rabies. The most notable development in this country was goat adapted rinderpest virus vaccine by Edward in 1923. Several outstanding vaccines have been produced in the last 30 years employing cell culture.
Vaccination has played a major role in the control of many virus dilleases among men and animals. The viral vaccines have traditionally been classified into two major categories-live attenuated and inactivated. Immunity 10 viral infections-depends on the development of an immune response to antigens present on the surface of virions or virus lOfected cells. An immune response to the nonsurface antigens of the .virus play a minor role in the immunity to viral infection. The immune response to certain internal proteins of the virus may play a \:ooperative role in the development of effective resistance.

Viral Vaccines and Antiviral Agents 103
Attenuated or live virus vaccine
Most of the attenuated vaccines have been developed by growing the virus in a host other than the natural host until the virulence for natural host has been reduced to a level which allows it to be used safely in that host. Some naturally occuring virulent viruses have also been used as vaccines. The original vaccine (Vacca means cow) introduced by Jenner in 1798 for control of small pox among human beings, used cow pox virus. In Veterinary practice herpes virus of turkeys is used to protect chickens against Marek's disease, bovine rotavirus by oral routc is used to protect piglets against porcine rotavirus infection, naturally occuring lentogenic strains of newcastle disease virus are used to protect chickens against virulent Newcastle Disease virus. Virulent viruses by unatural route are also used as vaccines. Infectious laryngotracheitis virus via cloaca is used to prot~t chickens, contagious ecthyma virus by scarification of inguinal region of sheep is used to vaccinate sheep against orf virus.
Viruses from one species may undergo replication without producing disease in another species but elicit protection e.g. measles virus when inoculated in young dogs protects them against canine distemper virus.
Most of the live virus attenuated vaccines have been derived empirically by serial passage of virulent virus through heterologous nost, laboratory animal, embryonated eggs or through cell culture. During the process there is progressive loss of virulence for the original host but maintains the capacity to immunize the animals against virulent virus. It is important to fix the degree of attenuation, so that the vaccine virus retains the capacity to replicate sufficiently to produce the desired level of immunity. An alternative method of developing attenuated vaccine strains is to select temperature-sensitive mutants. Temperature sensitive mutant vaccines have been developed for bovine vi-rus diarrhoea, bovine infectious rhinotracheitis etc. but there are prob~ems with reversion to virulence. The cold adapted mutants appear to be more stable but are of low immunogenicity.
The attenuated vaccines multiply in the host and thus produce a long lasting immunity. The attenuation of viruses is achieved in the laboratory by adapting the viIuses to different animal hosts, chicken embryos and cell cultures often after many serial passages. These

104 Textbook of Veterinary Virology
vaccines can be administered to animals parenterally, orally or intranasally. Intranasal and oral vaccine3 also induce the synthesis of IgA by the locallymphoiq ceJIs.
The live vaccines have. certain disadvantages as well. The best attenuated vaccines even cause a small number of clinical cases. There is a great concern regarding the viruses to revert to parent virulent form when it multiplies in the natural host. Stringent tests are therefore, necessary to ensure that the product is innocuous. There is also danger of natural spread to incontacts. The attenuated vaccines can also be contaminated by latent viruses from the cell cultures of chicken embryos used to grow the vaccine virus: live polio vaccines prepared in monkey cells contained SV 40 virus. There is also the problem of storage and transportation at low temperature to maintain potency.
Naturally avirulent viruses may also be used as vaccines. Turkey herpes virus is successfully used for vaccination against Marek's disease. The virus does not produce clinical signs in turkeys or chickens.
Inactivated or killed vaccine
The inactivated vaccines work on different principle from attenuated ones. A sufficient amount of viral antigen is administered in a series of intramuscuI.ar or subcutaneous injections to produce sufficient immunity to prevent disease.
Inactivated viral vaccines consist of tissue suspensions derived from artificially infected animals, chicken embryos or cell cultures. The virions in these vaccines are usually killed by destroying their infectivity by physic,al or chemical agents. Formalin usually in the concentration of 0.05 to 0.25% has been used to inactivate the viruses. Formalin denatures the viral proteins to some extent. The other inactivating agents which inactivate the nucleic acid but not the viral proteins are considered better agents such as p-propiolactone, ethylenimines and ethylene oxide. These agents are now being used in the preparation of viral vaccines. However, formalin is still being preferred because of its easy availability and being cheap. Physical agents like ultraviolet light are also used for virus inactivation. It is important to note that unless all nucleic acid is inactivated the mutants may arise.

Viral Vaccines and Antiviral Agents 105
Table 11.1 COY".PARISON OF INACDVATED AND AITENUATED VIRAL VACONES
Sr. Characteristics Inactivated vaccine.f Attenuated vaccine
No.
1. Safety Safe as all virus activity There is a possibility of is completely destroyed. reversion to virulence in
the inoculated hosts. The virus may spread to other susceptible hosts.
2. Freedom from The vaccines may be The live vaccines may extraneous free from extraneous contain certain adventi-
agents agents. tious viruses. 3. Protection The onset of protection The protection is rapid
is slow, short lived and often high and long the quality of protection lasting. is not good.
4. Dose and route Such vaccines must be Can be administered by of administration inoculated in usually oral or respir atory routes.
large volumes and The volume acquired is by parenteral route only. often small and cost of
administration is low. 5. Stability The vaccines are stable These are not stable at
at room temperature. room temperature
unless in freeze dried form.
6. Quantity of virus The quantity of viral The antigen required is antigen acquired is less and therefore more and su<.:h vaccines cost is less. are costly to produce.
The killed virus vaccines do not multiply in the animals and are safe. The inactivating agents in the vaccines also eliminate the contammating agents. The inactivated vaccines are useful even in pregnant animals or in those situations where live vaccincs cannot be used. There are certain difficulties encountered with these vaccines. The vaccine must contain high concentration of viru~ antigen to produce immunity, such immunity is of short dUffltion and must be boosted. These vaccines do not induce satisfactory levels of IgA and

106 1'exJbook of Veterinary Virology
usually produce circulation antibodies against surface components of virus particles. In certain cases the rabies inactivated vaccine (samples vaccine) may produce severe allergic encephalitis in man. Adjuvants are used to enhance immunologic response to killed vaccines. The adjuvants allow a slower release of antigen and its degradation and also stimulate phagocytosis and other activities of reticuloendothelial system. It is likely that certain adjuvants enhance cell mediated or humoral immune response. Mineral oil. gels like aluminium hydroxide. quail A. aluminium phosphate or alum are widely used in veterinary vaccines. Recently aviridine (a lipoidal amine) has been shown to have good adjuvanticity in foot-and-mouth disease vaccine:Virulent virus by abnormal route is sometimes used as vaccine. Orf virus is used by scarification on the thigh of lambs or infectious laryngotracheitis virus by applying on the cloacal mucosa. This practice is dangerous because virulent virus is shed which can infect non immune animals.
Isolated immunogens
Nucleic acid of virion is infectious and does not play any part in immune response. while proteins in the capsid and envelope are responsible for producing immunity. Instead of using whole virion. only the relevant protein against which the immunity is produced is inoculated. Such vaccines are known as subunit varcines. Recent advances in molecular biology and peptide synthesis provide a basis for producing large quantities of pUrified viral proteins or synthetic peptides for use in vaccination. Two separate approaches to the production of viral antigens are being currently exploited. One involves the production of synthetic peptide representing immunologically important domains of surface antigen. The second approach involves the use of clOned viral DNA inserted into a suitable vector to produce viral protein in a bacteria or virus or a yeast cell.
Synthetic peptides~ Rapid progress in determinin~ RNA and DNA sequence of various viral genes has simplified the selection of peptides with potential antigenic activity. In the recent past. major antigcnic sites on protective proteins of a number of viru~s have been identified. The amino acid sequence of viral protein can be determined. Parts of proteins. oligopeptides can be synthesized chemically. It is possible to synthesize short peptides which correspond to the antigenic determinants to which neutralizmg antibodies bind. There is possibility that relatively invaria~ or even buried sequence which are not normally immunogenic when presented in situ in the virion. may be capable in

Viral Vaccines and Antiviral Agents 107
isolation, of eliciting neutralizing antibodies. This may have the advantage of cross protection against heterologus serotypes.
The feasbility of using synthetic ~ptides for inducing immunity in " animals was desmonstrated for foot-and-mouth disease virus (FMDV).
An icosapeptide corresponding to the major antigenic site of VP1
of FMDV was coupled to keyhole limpet haemocyanin which served as carrier and was inoculated with Freund's adjuvant to guinea pigs. These guinea pigs withstood challenge with virulent FMDV. These immunogenic peptides of FMDV did not protect the cattle.
Although the synthetic peptide approach has a considerable potential, there are obstacles which remain to be resolved. The synthetic peptides are poor immunogenic. The adjuvants and liposomes potentiate the immune response to most of peptides. A single peptide may be insufficient to induce immunity because large surface antigens usually contain several distinct immunologic domains that elicit protective immune response. In certain cases it may not be possible to identify small peptide that are protective.
Cloned viral DNA: The viral genes can be cloned in prokaryotic cells using a plasmid or bacteriophage vector. The'-doned viral DNA can be expressed as viral protein in prokaryotic or eukaryotic cells. VP
1
protein of foot-and-mouth disease virus has been expressed in bacteria, E.coli, accounting for 17% of protein produced in transferred bacteria. When mixed with Freund's incomplete adjuvant, the fusion protein induces resistance in cattle and pigs to challenge with virulent foot-andmouth disease virus. Plasmid vectors can also be engineered to express unfused viral proteins such as rabies virus surface glycoprotein.
Allti-idiotype vaccille: The amino acid sequence of antibody molecule which binds to an antigen is known as its idiotype. The antibodies raised against the idiotype are known as antiidiotype antibodies. This antiidiotype antibody binds to same idiotype. Therefore, the antiidiotype could be used as vaccine. The antiidiotype antibodies raised against hepatitis B virus surface antigen elicit an antiviral antibody response in animals.
Antiviral Drugs
While most of the bacterial diseases of man and animals have been brought under control by the use of antibacterial agents, the same cannot be said about viral diseases since anuviral drugs in general do not have the same degree of efficacy as antibacterial drugs. Antiviral drugs have limited use in man in certain advanced count:-ies. Recent

108 Textbook of Veterinary Virology
laboratory investigations have shown that the growth cycle of many RNA and DNA viruses can be interrupted in cell culture and sometimes even in experimental animals. Despite these successes most of the antiviral drugs have proved to be highly cytotoxic and cannot be safely used againsJ viral diseases in man and animals. Another important reason is the close association of viruses with host cells during replication. Short lived nature of many viral diseases is another obstacle so that by the time the disease is diagnosed the viral multiplication has largely been completed. To be effective, antiviral drugs have to be administered early in the viral infection.
For antiviral therapy, the compound must be capable either of preventing entry of a virus in the host cell or specifically inhibiting some essential steps in virus replication, maturation etc.
Inhibitors of viral entry
Amantadine and its derivates have successfully been used in human influenza and have also been shown effective against swine influenza. The drug apparently prevents the penetration of virus into the host cell although the exact mechanism is not fully known. This compound also may act against the viral transcriptase. Methyl adamantanemethylamine hydrochloride, a compound closely related to amantadine is also effective against influezna A viruses.
Purines and Pyramidine antagonists: The halogenated pyramidines are the group of substances known to inhibit DNA synthesis. The halogenated deoxyuridines namely 5-iodo-2-deoxyuridine (IUDR) and its bromine equivalent (BUDR) are incorporated into the viral DNA in place of thymidine and produce defective (fradulent) non functional DNA molecules. The !UDR acts in the final stages of viral replication. It is possible that !UDR inhibits the action of DNA dependant RNA polymerases and blocks in the formation of mRNA. These drugs are administered locally in human cases of herpetic ulceration of cornea, unfortunately these are too toxic to be given parenterally in man. BUDR inhibits DNA synthesis by blocking the synthesis otthymidilic acid.
Thiosemicarbazones: lsatin-B-thiosemicarbazone (IBT) and paminobenzylaldehyde are potent inhibitors of poxviruses and adenoviruses. Some derivatives are also active against certain enteroviruses. The drug has no action on DNA synthesis and probably acts late in viral replication cycle at the level of translation and interferes in the synthesis of viral mRNA.

Viral Vaccines and Antiviral Agents 109
ArabinojurRosyl nucleosides: Adenine arabinoside is effective against some herpes viruses and to some extent against pox viruses. The drug is effective as a topical agent against herpetic keratitis. Its action is inhibition of DNA synthesis by interfering with the reduction of cytidylic acid to deoxycytidylic acid.
A cyclo vir: Acyclognanosine, commonly known as acyclovir is a guanine derivative with an acyclic side chain. This is one of the newest drugs which is more promising antiherpes viral agents. The drug is phosphorylated to monophosphate by a herpes virus specified thymidine kinase in herpes virus infected cells but this does not occur in uninfected·cells. The monophosphate is converted to triphosphate in herpes virus infection cells to a greater extent than in healthy cells.
Benzimidazoles: Benzimidezole and guanidine interfere with the synthesis of single stranded viral RNA without affecting the cellular RNA synthesis. These drugs prevent the multiplication of many enteroviruses in vitro. The drug has no protective effect in animal models due to rapid emergence of drug resistant mutants.
2-deoxy-D-glucose and glucosamines: The drug inhibits the multiplication of many viruses that mature at the cell membrane like orthomyxo, paramyxo and herpes viruses. The drug acts perhaps by interfering with the synthesis of virus specific glycoproteins.
Antibiotics: Certain antibiotics like rifamycin, tolypomycin and streptovaricin display antiviral activity against some pox viruses and oncoviruses by interfering with maturation and by inhibiting the activity of reverse transcriptase in OACoviruses. Actinornycin-D inhibits DNA transcription and mitomycin-C inhibits protein and DNA synthesis but these antibiotics are very toxic and cannot be used paren terall y .
Interferences
In 1957 Issacs and Lindenmann found that allantoic fluid of eggs that had been exposed to irradiated influenza virus posses an interfering property with the multiplication of infectious homologous virus distinct from the virus itself. The substance responsible for this interference was named as 'interferon'. The nature and mode of action of interferon is complex. The interfervns are host coded proteins and there are three distinct types of interferons, one formed primarily in leukocytes (IFNa), another in fibroblasts (IFN-P) and third formed in stimulated lymphocytes (lFN-y). The norma! cells do not synthesize interferon, but the cells can be induced to do so by a variety of inducing agents.

110 Textbook of Veterinary Virology
Interferon blocks the multiplication of many different RNA and DNA containing viruses both in eggs and in cell culture. The DNA recombinant technology has been effectively used for cloning of the genes for human interferon (IFN) in bacteria, yeasts and mammalian cells.
Characteristics and types of interferon: The interferon is a small soluble protein (mol wt. 25-45,000 d) containing nearly all amino acids. The IFN is relatively non toxic, thermostable and active through a wide range of pH. It is a weak antigen and can be neutralized by antiviral serum. The IFN is inactivated by proteolytic enzymes like trypsin but is not affected by receptor destroying enzyme, RNase, DNase or periodate.
There are three classes of human interferon. IFN-a. and IFN-/3 predominantly produced by leucocytes and fibroblasts in response of viral infection or other inducing agents, while IFN-y is produced by unsensitized lymphoid cells in response to mitogens and by sensitized lymphocytes when stimulated with specific antigen. IFN-/3 and IFN-y species are glycoproteins in nature while IFN-a. are not glycoproteins. IFN-/3 and IFN-yare stable at low pH (pH 2 at 4°C) and retain activity in the presence of sodium dodecyl sulphate (SDS).
There appears to be two highly conserved domains in interferon molecules. One in the aminoterminal half of the molecule and probably contains the site that binds to cell surface receptor, while the other biologic functions.
The interferons are produced by almost all vertebrate species. The best studied interferons are those of humans, rodents and birds. The three classes of human interferons are immunologically quite distinct. The interferon appear to be larger than 17000 dallon molecules, when isolated from the cells. The reasons may be that some interferons are glycoproteins, some naturally occuring interferons may be dimers and under certain conditions, interferons may associate with other proteins. The interferons are host species-specific, the basis for spcdficity probably resides in the specificity of the interferon cell surface receptor interaction. The specificity sometimes is very narrow. All interferons possess antiviral and anticellular activity. The interferons cause interference with the multiplication of viruses and regulate a variety of cellular functions. The induction of antiviral state by interferons differs against various VIruses both with type of cell and type of interferon. The activity of IFN-yis fur more potent than IFN-a. and IFN-/3.

Viral Vaccines and Antiviral Agents 111
The production of interferons: The nonnal cells do not synthesize interferon due to the absence of transcription of interferon genes. However, interferon is transcribed spontaneously in some cells. All such cell lines are lymphoblastoid cell lines, some produce lFN-y but most produce lFN-a.. The interferon produced is homogenous but may be the product of single interferon gene. The amount of interferon produced by interferon-relating cell lines is too small and cannot be utilized for practical purpose, The important interferon inducer is virus infection. Both RNA and DNA containing viruses induce interferon production, but RNA containing viruses are good interferon producers than DNA containing viruses' except pox viruses. The interferon production in the infected cells starts approximately 4 hrs after infection at 37'C and reaches a peak when the viral protein synthesis is at maximum and then declines.
The second type of interferon inducer is ds RNA. All naturally occuring ds RNAs of viruses, replicative forms of ss RNA of animal and bacteriophages, synthetic ds polyribonucleotides like poly I: poly C or poly lC are interferon inducers. The ss RNA or DNA or ds DNA or RNA-DNA hybrid molecules do not induce interferon production. The ds polyribonucleotides which induce interferon production have the properties (i) have stable secondary structure (ii) relatively resistant to ribonuclease (iii) have higher molecular weight and (iv) have minimum effective chain length of 100 residues.
The thrid type of interferon inducers are viruses those are unable to replicate such as viruses in non permissive cells and inactivated viruses. Thele is enormous variation in response to these inducers depending on the nature of cell and of the interferon inducer.
There is another important group of interferon inducers consisting of metabolic activators and inhibitors namely mitogcns, metabolic activators etc.
The cells treated with very little interferon produce more interferon than untreated cells, called as priming. Priming is used extensively to increase the large scale production of lFN-a and IFN-/3.
Biological effects of interferons: Interferons have varied biologic effecis, namely antiviral and cell multiplication-inhibitory activities including antitumor activities, immunomodulation, inhibition of certain synthetic cellulaa' activities and enhancement of others, toxicity enhancement etc. The effects of interferons are exerted at plasma cell membranes Vla combination with specific receptors.

112 Texlbook o/Veterinary Virology
Interferon has an antiviral activity. The viral infections cause sever disease in animals treated with antibodies against interferon than in normal cells. The interferon protects cells from effects of virus infection afjd also renders infecticn abortive. In vaccinia virus infected cells, host protein synthesis is inhibited more quickly in interferon treated cells than .in untreated cells, marked cytotoxic effects are induced in interferon treated cells and cells die. In other virus systems it is not the case. In many cell systems, interferon treated cells are protected against virus infection. Protection by interferon· depends on the concentration of interferon and multiplicity of infection. Large doses of interferon protect against low multiplicity of infection while high multiplicity of infection overcome the protective role of interferon. Under natural conditions in the body, interferon protects many cells from cytopathic effect of virus infection but does not completely eliminates the infection. The interferon may be a factor in establishment and maintenance of persistent infections.
Interferons do not actually inhibit virus multiplication but induce the cells to synthesize proteins that are effectors of antiviral state. The interferons do not induce the antiviral state if messenger RNA or protein synthesis are inhibited. The interferon-induced inhibition of virus multiplication, is due to interference with the ability of parental or early viral mRNA molecules to be translated with the result that the infection is aborted. This has proved in case of vaccinia virus infected with L-cells and reoviruses. In some system it ~ not easy to detennine whether the tmnscription or translation of early viral mRNA is inhibited to greater extent. In the cells infected w.il.h VSV and SV 40 it was observed that transcription rather than translation of early viral mRNA is inhibited. However, under strict precautions it was found that in VSV infected cells the .effect of interferon treatment was on translation of mRNA transcribed from parental viral genome. In case of SV 40 infected cells with SV 40 DNA instead of whole virus, translation of early viral mRNA is inhibited much more than transcription. When interferon was added to the cells late in the lytic cycle of SV 40
multiplication, the rate of mRNA was not inhibited but mRNA translation was inhibited. These results suggested that interferon inhibits the uptake, penetration and uncoating of SV..,. These studies were ,:;arried out in vilro and it was found that cause of inhibition was the presence of an inhib~tory factor and the inhibitory factor is associated with ribosomes. Kerr and his associates demonstrated that

Viral Vaccines and Antiviral Agents 113
inhibitor of translation in extracts of interferon treated cells is the induction by interferon of at least three separate enzymes and a further requirement for ds RNA in their enzymes and a further requirement for ds RNA in their activation. These enzymes are 2-SA synthetase, RNase and protein kinase. Two of these enzymes require ds RNA for their activation. The ds RNA is persumed to be formed as an intermediate or by product in replication of all RNA and DNA viruses. The ds RNA also appears to be essential for induction of IFN.
The enzyme 2-SA synthetase or 2-SA polymerase, catalyses the synthesis of 2-SA from ATP. It is active only when bound to ds RNA. Its products are a family of oligoadenylates.
RNase L: Lengyel and his colleagues observed that in the presence of ds RNA and ATP both reovirus mRNA and cellular mRNA were degraded more rapidly in extracts of IFN treated cells. It was further reported that addition of 2-SA to an in vitro protein synthesizing system reduced the translation of mRNA which is attributable to an endoribonuclease that is activated by 2-SA. The function of 2-SA is to
bind to latent endonuclease and activate the enzyme to destroy both viral and cellular mRNA and rRNA.
Protein Kinase: A second pathway, independent to 2-SA synthetase/RNaseL, is inhibited by interferon in the presence of ds RNA. A ds RNA-dependent protein kinase was found to phosphorylate at least one protein. The protein phosphorylated by the activated kinase was shown to be the subnit of the eukaryotic peptide chain initiation factor, eIF-2. Phosphorylation makes this factor inactive and the peptide chain cannot be elongated.
The inhibition of protein synthesis by IFN may also occur by the inhibition of methylation of viral mRNA. The S' terminal guanosine 'cap' is unmethylated apparently as a result of IFN induced changes in the levels of S-adenosylmethionine and S-adenosylhomocysteine.
Effect of Interferon on Transformation by Tumor viruses: SV AO
DNA tumor virus has been extensively studied. It has been observed that pretreatment of cells with interferon markedly reduced the efficacy of cell transformation. It was found that IFN differentially affected the expression of early viral genes whether viral DNA was integrated or not, thus, SV «I transformed cells can be passaged in presence of infection for many generations with no effort on T antigen synthesis, while interferon inhibits T-antigen formation in lyticaHy infected cells. The lytic SV «I multiplication cycle was remarkable in that viral DNA

114 Textbook of Veterinary Virology
replication and mRNA translation were sensitive to interferon added hours after infection. In other viral multiplication cycles the antiviral state must be established before infection for interferon to cause inhibition. In retroviruses IFN acts before integration, it prevents either the synthesis or the integration of proviral DNA.
Prospects for clinical use of interferon: There is little scope of IFN finding an extensive scope in infections like influenza and herpes virus infections but interferons may be useful in life threatening diseases like rabies, herpes virus encephalitis and to eliminate persistent infections caused by certain group of viruses. The remarkable effect of IFN on immunoregulatory mechanism has focussed attention in IFN as lymphokines. The immunoregulatory mechanism of IFN-y and the cell inhibitory activity of lFN has raised hope that IFN may be useful as anticancer agent.
For therapeutic use, large amount of IFN is required and such large amounts of lFN cannot be prepared in vitro. Now with the latest biotechnological techniques large amounts of cloned IFN will become available. Unfortunately the IFNs cause some undersirable effects like fever, hypertension, myalgia, tachycardia and impaired liver function but such side effects have to be weighed against life threatening diseases.
References
FENNER, FRANK, 1987. Veterinary Virology. Academic Press, New York.
STEWARD, M.W. and HOWARD, C.R., 1987. Synthetic peptides a next generaJion ofvaccines. Immunology today 8,51-58.
EsPOsrro, IosEPH I. and MURPHY, FREDRICK A., 1989. Infectious recombinant vectored virus vaccines. In advances in Veterinary Science and comparative Medicine. Vol. 33,196-249.
MURPHY, BRIAN, R.and CHANOCK, ROBERT, M., 1985. Immunization agai/'lSt viruses. In Virology edited by N.N. Fields et al. Raven Press, New York p. 349-370.
Wn.soN, G.; MILES, A and PARKER. M.T., 1984. Principles of Bacteriology, Virology and Immunity. Edward, Amold.
TIZARD, I.R., 197·7. An introduction to Veterinary Immunology. W.B. Saunder~. Philadelphia.

Chapter 12
Diagnosis of Viral Diseases
The clinical diagnostic virology involves visualisation of the virus particles by electron microscopy. virus isolation. serological diagnosis through qualitative and quantitative detection of specific antibodies and direct assay for viral antigens. A recent criterion has been added which involves the detection of viral specific nucleic acid sequences. The choice of diagnostic method depends on nature of samples. pre-existing reagents and the type of virus to be detected. The diagnosis is important for undertaking suitable control measures.
Collection of material for diagnosis
They key to diagnostic success is the examination of suitable samples. The extra expense involved in collecting the proper material from several animals at the appropriate stage of disease and its despatch to reference laboratory to confirm the diagnosis is negligible when compared with unsuitable .samples collected for disease diagnosis. Tissues should be collected under sterile conditions. Organs which arc source of bacterial contamination should be sampled last e.g. alimentary tract. The site of collection of specimen is related to the clinical signs and knowledge of pathogenesis of the suspected disease. In live animals the material from 'target organs' should be collected.
(i) Nasal or nasopharyngeal swabs-for the v:ruses infecting respiratory tract e.g. ortho and paramyxoviruses. herpes virus and adenoviruses. The swabs are broken off immediately after collection in the transport medium which compnses of buffered saline or cell culture

116 Textbook o/Veterinary Virology
medium containing bovine serum and antibiotics and fungistatic agent to suppress the growth of contaminants.
(ii) Vesicle fluid, scab or epithelium-for foot-and-mouth disease, pox viruses etc.
(iii) Faecal and cloacal swabs-for enteric viruses e.g. enteroviruses. parvoviruses etc.
At postmortem virus may be isolated from target organs which contain high concentration of the virus and are less contaminated.
(i) Lymph nodes in rinderpest or mucosal disease. The lymph nodes are less likely to con~n contaminants.
(ii) Spleen in Newcastle disease. infectious bursal dise.'lse. swine fever. rinderpest etc.
(iii) Liver in canine infectious hepatitis. adenovirus. enterovirus infections etc.
(iv). Foetuses in transplacental infection e.g. equine herpes virus (foetal-liver and lung). border disease.
(v) Central nervous system in rabies.louping ill and pseudorabies. The specimens, after collection. are labelled and despatched to the
laboratory with its history for the provisional diagnosis. The specimens are despatched over ice in thermosOasks or in thermocole containers. If the transit time is longer the ice should be replenished during transit or the material may be transported over dry ice (-70"C). For particularly labile viruses like herpes viruses. coronaviruscs etc .• the- cell culture may be taken to the animal.
Electron Microscopy
The visulatisation of viruses, causing disease, by electron microscope (EM) is very logical. The use of electron microscope has been gradual. EM is used with success in detecting viruscs in faecal contents and in detecting viruses involved in acute gastroenteritis. There are intrinsic faults with EM:
(i) The instrument is expensive and is not available in all the diagnostic laboratories.
(ii) A high concentration of virus particles must be present in lhe samples. It is estimated that a lowest concentration of 10' particles per: millilitre is essential.
(iii) The morphology may not be dislinct lO allow exact viral diagnosis.

Diagnosis o/Viral Diseases 117
The EM has also a unique advantage. The viruses which grow with difficulty and for those viruses in culture for which there are no immunoassays like parvoviruses in the brain, astroviruses, calciviruses, the detection is by EM. The inethod is fastest and is least expensive in case the EM is available. The visualisation after negative staining, such as phosphotungstic acid, of virus particles in field samples is a rapid way of diagnosis. The time required is about 15 minutes. A further step is the use of immunoelectron microscope to identify antigenically the virus strain implicated in the disease. The time required is about 3 hours.
Most of the recently discovered viruses as rotavirus, coronavirus were detected by electron microscopy. The samples which often give positive results are vesicular fluid, cutaneous lesions, faeces, liver and tissue culture supematants. The viruses which are easily observed are parapoxvirus, herpesvirus, papiIlomavirus, rotavirus, coronavirus, parainfluenza virus etc.
Virus isolation
Inspite of recent development of rapid methods for detection of viral infcctions, most laboratory diagnosis of viral disease still depends on isolation and identification of viral agents. It is probably one of the most sensitive method in the diagnosis of viral infections if properly collected material is used. The three major systems for isolation of viruses from clinical samples arc tissue culture, embryonated eggs and experimental animals. Out of these the most sensitive method is tissue culture.
A single monolayer type is not sensitive to the growth of all the viruses. Most diagnostic laboratories choose different tissue cultures depending on the suspected viruses. The choice of cell lines and to use embryonated eggs or laboratory animals like suckling mice depends on the availability of cell lines, cost considerations and the laboratory set up. The major types of tissue cultures used are primary cultures, diploid cultures or continuous cell lines. The primary cuilures acquire new properties which render the cells susceptible to infection by viruses, when in lhe native state they might not have been. The secondary cell cultures are similar in properties to primary cultures. The diploid cell cultures are fibrobJastic in nalure and are sensitive to number of viruses which arc difficult 10 culture from clinical specimens. Established cell

118 Textbook o/Veterinary Virology
lines are derived from either normal tissues or malignant tumors, they are heteroploid cells. These cell lines acquire unique characteristic of growth or sensitivity to viruses. The inoculum of the suspected specimen js inoculated in several tubes containing tissue culture and are incubated either stationary at 36-37°C or roller drum at 33°C. The medium is usually changed after an adsorption period of several hours to one day, to prevent the non specific toxicity. The cultures are periodically examined for signs of developing viral growth. The frequently used sign of virus growth is cytopathic effect (CPE). Most viruses produce characteristic morphologic changes like lysis (necoris), Inclusion formation, cytomegally giant cell(or syncytium), particularly myxo and paramyxoviruses are recongiscd by haemadsorption of guinea pig or other erythrocytes on the monolayer. Rubella virus can be detected by its capacity to interfere with the growth of second challenge virus.
The final identification of virus isolates usually depends on the serologic tests and sometimes on morphologic, biophysical or biologic characterization like acid stability, plaque formation etc.
The embryonated eggs at onc time were widely used in viral diagnosis but due to development of tissue culture techniques, which offer superior sensitivity, the use of chicken embryo has become less. The use of suckling mice and guinea pig is still being continued for enteroviruses, apthoviruses, alphaviruses, flaviviruses, arenaviruses, orbiviruses and rhabdoviruses.
Serological methods
The serologic tests are used to detect either antigen or antibody depending upon whether a standard antiserum or standard antigen is used. The serum samples are heat inactivated to remove complement(C) and non specific inhibitors. The serologic tests commonly used for viruses are as under:
1. Virus neutralization test (VN). 2. Immunofluorescence (IF) and immunoperoxidase(IP) tests. 3. Radioimmunoassay (RIA). 4. Immuno-electron microscopy. 5. Enzyme linked immunosorbent assay (ELISA). 6. Agar gel double immunodiffusion test (AGD). 7. Complement fixation test (CFI).

Diagnosis of Viral Diseases 119
8. Haemagglutination inhibition (HI) test 9. Haemadsorption inhibition test (lID!).
1. Virus neutralization (VN) test: VN tests depend on'the ability of antibodies to neutralise virus infectivity, as shown by inoculation into a sensitive host system in which the virus produces recognisable effects. In some cases addition of homologous or guinea pig complement 10 virus and its antiserum mixture enhances neutralization. The VN test is performed in 2 ways. In alpha procedure a constant concentration of antibody is added to an equal volume of serial ten fold dilution of virus. The alpha procedure is generally used to identify the virus. In beta procedure, constant concentration of virus is added to an equal volume of serial two fold dilutions of antiserum. The beta procedure is generally used 10 measure antibody titre. In both procedures after an incubation of 1-2 hours at 37°C, the mixture is inoculated in the suitable host system. The tests are scored and antibody titre calculated by several methods. A 50% reduction in plaque assay, a 50% protection or a neutralization index of 2 is considered significant. VN tests are normally most specific and sensitive: the antibodies involved arc directed against surface antigens of the virus which arc usually type or subtype specific. The neutralizing antibodies usually persist for long periods in the absence of reinfection. In the serotyping, reciprocal weak neutralization may occur between two serotypes but if monoclonal antibody is used instead of conventional hyperimmune serum, the serotyping becomes very clear and specific since there is no cross reaction.
2. Immunofluorescence (IF) and immunoperoxidase(IP) tests: The IF is used to detect antigen in virus infected tissues, cell cultures or impression smears and to detect viral antibodies. The viral antigens can be visulaized in cells by specific antibody conjugated to a fluorescent dye like fluorescin isothiocynate or rhodamine B. The indirect test is more in use where the dye is conJugated to a species antiglobulin. Thus one conjugate can be used with any antiviral sera of that species. In the direct method the conjugated antibody is applied onto the specimen and incubated at 37°C for 30 min. The specimen gives fluorescence if a specific reaction occurs. The indirect test is also used for antibody screening.
A modification of the test is immunoperoxidase (JP) test when the

120 Textbook o/Veterinary Virology
antibody is conjugated to an enzyme which will change a colourless substrate into a coloured one as in ELISA test. This test has the advantage that only light microscope is needed to detect antibody binding and moreover, the preparations (slides) can be maintained for long periods.
3. Radioimmuno assay (RIA): The detection of antibody to viral antigen is performed by radioirnmunoprecipitation. The test involves detecting virus being fixed to polypropylene microwells and binding antibody by radio labelled antiglobulin. The test is rapid, extremely sensitive and specific but due to short life of some of the chemicals means that reagents have to be repeatedly prepared and tested. RIA also detects antigen antibody binding sites by different test procedures e.g. antibody by immunoprecipitation of radiolabclled antigen with antiglobulin coated beads.
4. Immunoelectron microscopy: Some viruses which are difficult to culture have been diagnosed by electron microscopy, e.g. ro~viruses in faeces of animals. The irnmunoelectron microscopy involves reacting the virus with its antiserum. The virus particles are thus clumped and are readily identifiable by negative staining and can also be concentrated by low speed centrifug~tion. Electron dense ferritin or gold can also be linked to antibody so that virus antigens can be localised in ultrathin sections of the cells.
5. Enzyme linked immuno-sorbent assay (ELlSA): The test was developed from RIA with the additional advantage of the samples being scored for colour either by eye or spectrophotometer. ELISA is used either to detect antigen or antibody. It is a rapid sensitive, specific test and is not hazardous. For detecting antibody, wells of microtiter plate are coated with antigen and are than washed to remove excess antigen. The antiserum is added and is allowed to react with antigen. After an incubation period, the excess unattached antibody is removed and wells are washed again. Then enzyme conjugated antiglobulin is added 10 the well. After further incubation an enzyme substrate is added. The attached anzyme causes a colour change in the substrate which may be measured spectrophotometrically. For antigen detection the wells of microtitre are precoated with antibody and the plate blocked with bovine serum albumin as before. The specimen e.g. tissue

Diagnosis o/Viral Diseases 121
extract, plasma or tissue culture fluid is added and the viral antigen is tied with the antibody. The unbound material is washed, off. An enzyme labelled antiviral antibody is subsequently added and binds only if first solid phase antibody has already captured antigen. The antigen can be linked to the filling of antibody sandwich in a positive test.
6. Agar gel double immunodiffussion test (AGD): This test is used to detect viral antigens and antibodies. The glass slides arc covered with agarose (1 %). On hardening, wells of desired pattern are cut. The antigen and antibody are added in the appropriate wells and incubated. The antj.gen and antiserum diffuse toward each other in the gel resulLing in the fonnation of precipitin lines at the zone of equiValence. If the two antigens are identical both the precipitin lines fuse at their continuous ends. When antigens are cross reacting they fuse but produce a spur like projection and extend towards cross reacting antigen. If both antigens arc unrelated the precipitin lines cross each other. The quantitation of viral antigen or antibody can be done by modification of AGDP known as radial immunodiffusion test. When antigen is added to a well in the gel medium containing specific antiserum, a ring of precipitation appears around the well. The diameter of the ring is dependent on the concentration of antigen. By measuring the diameter of the ring, the concentration of the antigen can be detennined.
7. Complement fixation test (eFT): The fonnation of antigenantibody complexes results in binding and fixing of guinea pig complement. The fixation of complement is tested by adding sheep rbc sensitized with the homologus antiserum or haemolysin. The sensitized rbe will lyse if unfixed complement is present. The CFf can be carried out within a day and it detects antibodies to internal viral proteins as well as to envelope proteins. The antigen in the test can be inactivated if the virus is infectious for the worker in the laboratory. The CF antibodies persist for a short period and their presence indicates that the infection is recent. Some of the sera or antigen are anticomplementary and the test is tedious to perfonn.
8. Haemagglutination inhibition (HI) test: The ability to cause agglutination of erythrocytes of many species of animals and birds is found in many virus families. This property of haemagglutination is

122 Textbook of Veterinary Virology
inhibited by the specific antiserum of the virus. The ill test is used either to type antigen or measure antibody. The test is carried out in plastic plates with multiple wells. The agglutinated rbe produce a widespread covering with serrated edge after setting while non agglutinated cells form a button. If the test is used to type an isolate then standard hyperimmune or monoclonal antisera are used. ill antibodies are rapid, specific and sensitive, the ill antibodies appear during late clinical phase of infection and persist for long periods in animals. In ill tests even inactivated virus can be used. The HI titre of the seriserum is expressed as reciprocal of the highest dilution of antiserum that completely inhibits HA.
9. Haemadsorption-inhibition test: Some virus infected cells adsorb red blood cells of certain species of animals because of virus envelope proteins. This is known as haemadsorption. This can be inhibited specifically by viral antiserum e.g. para influenza viruses. This test is performed in a manner similar to ill test.
Detection of viral nucleic acid
The detection of homologous nucleotide sequences offer a valuable tool for recognition of certain viruses. Advances in molecular biological techniques have been made and many of these techniques are useful in clinical virology.
The use of PAGE (polyacrylamide gel electrophoresis) has proved to be a sensitive method for detection of viruses belonging to the family Reoviridae.
Molecular biological techniques
l. Restriction endonuclease digestion of DNA and analysis of fragment patterns: The restriction e'lzyme like DNA endonculease is capble of recognising specific DNA sequence of 4-6 nucleotide bases. After digestion the DNA molecules are cleaved into different subgenomic fragments of different sizes. The pattern of DNA fragments becomes specific for each DNA species or virus, which are isolated by using gel electrophoresis and ethidium chloride.
2. DNA cloning lechnology: The DNA sequences are copied in an ill vivo system to produce identical clones. By using reversc uaD3Criptase, CDNA can be produced from RNA molecules. The.

Diagnosis of Viral Diseases 123
cloned DNA sequences of interest can be amplified through replication of vector.
3. Nucleic acid hybridisation: The hybridisation can occur with both DNA and RNA. The complementary base pairing of DNA molecules is separated from each other by denaturation which is accomplished by heating or raising the pH. Cooling allows reassociation or renaturation. On introduction of foreign DNA molecule with known sequence(probe), Ienaturation of the known sequence or probe contains a shared sequence complementary with the denatured DNA. This process of nucleic acid hybridization occurs both with DNA and RNA. The DNA duplex can be detected if the probe is labelled with radioisotopes or other markers such as biotin or enzymes. The formation of a duplex of probe and target DNA may not be done complementarily and certain amount of mismatching is allowed. The nucleic acid hybridisation reaction can be carried out in solution since detection of specific DNA in body fluids is the common application. The DNA probes can also be used to detect target DNA immobilised on nitrocellulose membranes or nylon membranes (dot blot, slit blot or Southern blot hybridsation). The target DNA can also be detected in formalin fixed tissues. This method is suitable for detection of oncoviruses associated with cancer.
Recent advances in the automated synthesis of oligonucleotide probes has made chemically synthesised short DNA sequences a practical alternative to cloned gene. Traditionally radioactiv~ labelling of probes (32P or 35S) is now been replaced with non radioactive labels as some of the radioactive labels have short half lives and are risky. The commonly used non radioactive label for DNA probes is biotin which can be detected with conventional avidin-enzyme conjugates or antibiotin antibodies. The DNA-RNA duplex after hybridisation using monoclonal antibodies have also been developed.
4. Nucleic acid sequencing: The ultimate method by which the structure, chemical and physical properties and functions of gene products can be analysed is by determining the nucleotide sequence of gene. It is a powerful method for the analysis of the organisation of specific information in viruses. Two methods are involved in DNA scquencing.

124 Textbook of Veterinary Virology
5. Expression of cloned DNA sequence: The strategy for cloning and expression of DNA sequence involves the insertion of DNA downstream from a host promoter sequence so that RNA polymerase of the host cell can recognise the cloned gene and initiate transcription. In certain cases the native protein encoded by the DNA insert may be unstable in the host and expression of the gene product as part of fusion protein may be desirable. Such fusion proteins usually consist of the gene product of interest together with the gene product of the host
6. Polymerase chain reaction: Polymerase chain reaction (PCR) involves in vitro enzymic amplification of specific targed DNA sequences. After cycles of heat denaturation of target DNA, primer annealing and primer extension, the number of target DNA molecules increase exponentially. The amplified DNA is then analysed and detected. The Characterisation and specificity of PCR product is achieved by using DNA probes in Southern or dot blot hybridization.
Applications of molecular techniques in clinical virology
Detection and idetltijication of viruses: The molecular techniques are most useful for those viruses which are difficult to be diagnosed by conventional methods. The hybridisation techniques and probes specific for number of these viruses have been developed. The PCR has offered a new dimension in diagnostic virology. The use of synthetic nucleotides as probes for detection of viruses in clinical specimens is not required for cloning and purification of viral nucleic aeids for probes.
Epidemiological studies of viral infections: The molecular epidemiology has helped to trace the movement of viruses between host populations. The molecular bioliogical techniques like comparisons of oligonucleotide fmgerprints of viruses form different regions or protein and nucleic acid sequences solves epidemiological pattern resulting from antigenic shift or drift of influenza virus.
Prevention and treatment of viral infections: The cloning and expression of viral specific proteins has led to lQe development of vaccine production. These techniques provided tools for studies of virulence factors associated with virus infections and offered new approaches for engineering vaccines. Specific mutations can be

Diagnosis a/Viral Diseases 125
introduced into viruses leading to the development of attenuated vaccine strains. In addition, isolated genes or portions of genes responsible for neutralization of antigens of viruses can be used for the development of subunit vaccines.
References
HITCHNER, S.B.; DoMERMUTII, C.H.; PURCHASE, H.C. and Wll.LIAMS, I.E., 1975. Isolation and identification of avian pathogens. Arnold Printing Coporation, Ithaca, N.Y. New York.
LENNETIE, E.H. and SCHMIDT, N.I., 1979. Diagnostic procedures for viral, rickettsial and chlamydia! infections, 5th Ed. American Publi-:: Health Association, Washington, D.C.
A summary of serological tests used to detect common infectious diseases of animals. Veterinary Medicine and small Animal Clinician. 7661725-61730, 1981.
BACIIMA.'l, P.A., 1983. New Development in Diagnostic Virology. Current Topics Hicrobiol. Immunol. 104:
GARDNER, P.S. and MCQUllIN, 1., 1980. Rapid virus diagnosis. 2nd Edn. Butterworth, London.
LE1'INETTE, E.H., 1985. Laboratory Diagnosis of viral infections. Dekker, New York.
McNULTY, M.S. and McFERRA."I, J.B., 1984. Recent Advances in virus Diagnosis. Martinus NijhotI, The Hague.
Wll.ER, I. and DOUGAN, C., 1989. DNA probes: A new role in diagnostic microbiology. 1. Applied Bactcrio. 67, 229-238.
WRlGIIT, P.A. and WYNFORD, T.D., 1990. The polymerase chain reaction, limitations in diagnosis and research. J .. Pathol. 162, 99-117.


PART II
SYSTEMATIC VIROLOGY
D.N.A. VIRUSES


Chapter 13
Poxviridae
The name of the family has been derived from 'POC' meaning a vesicular skin disease. The family is divided into six genera as given below and include more than 30 viruses among vertebrates.
Table 13.1 Pox VIRUSES (FAMII.Y POXVIRIDAE)
Genus Species
Orthopoxvirus Vaccinia virus-type species Cow pox virus Camel pox virus Buffalo pox virus Monkey pox virus Rabbit pox virus Horse pox virus Ec_lTomeJia virus
Capripox virus Sheep pox virus-type species Goat pox virus Lumpy skin disease virus
Leporipox virus Myxoma virus-type species Rabbit fibroma virus
Suipox virus
Parapox virus
Hare fibroma virus Squirrel fibroma virus Swine pox virus-type species Yaba monkey tumor pox virus Elephant pox virus Ecthyma virus-type species

130
Genus
Avipox virus
Textbook of Veterinary Virology
Species
Bovine papular stomatitis virus Milker's node virus Fowlpox virus-type species CllI1ary pox virus Pigeon pox virus Turkey pox vims Quailpox virus
The members of this family are largest of all viruses, brick shaped or ovoid virions measuring 220-450 x ]40-266 nm and can be seen under the light microscope. The surface structureo(}f some of the virions is of diagnostic relevance. The virions have an external coat containing lipid and enclosing one or two lateral bodies and a core. The central core or nucleoid contains the genome. The virion contains about 30 structural proteins and several enzymes. The nucleic acid is a single molecule of double stranded DNA of molecular weight in the range between 150 and 240 x 106 daltons. The virus particles are built from more than 30 structural proteins and several viral enzymes, to be used with the nucleic acid synthesis and processing which includes a DNAdependent transcriptase. There are about ten major antigens in the virus particle, one antigen cross reacts with most pox viruses of vertebrates. There is genetic recombination within genus and non genetic reactivation within and among other genera of this family. The multiplication takes place in the cytoplasm in special viroplasm factories and produce B type inclusion bodies. The cytoplasmic accumulations produce A type inclusion bodies. Mature particles are released from microvilIi or by cellular disruption.
The members of some genera are ether resistant while those of other genera are ether sensitive. The pox viruses withstand drying for months and even storage at room temperatures. They are destroyed by moist heat at 600 e within 10 minutes. They are also resistant to many common disinfectants. The members of orthopox viruses agglutinate red blood cells from turkeys and about 50 percent of fowls. The haemagglutinin is lipoprotein in nature and separable from the virus particles. It does not elute from agglutinated red cells and specific antibodies inhibit agglutination.

Poxviridae 131
Most of the pox viruses grow readily in embryonated eggs and produce pocks on the chorioallantoic membrane. They can also be cultivated in cell culture like chicken embryo fibroblasts, bovine kidney, rabbit kidney and HeLa cells.
The viruses of this family are pathogenic for man and many species of animals and birds resulting in the formation of focal lesions which are often proliferative in character. The spread of infection is by respiratory route or through the skin. Some members are also mechanically transmitted by arthropods.
Vaccinia Virus
The name of vaccinia virus is derived from the Latin word 'Vacca' means cow, after it was isolated from cowpox in 1796 by J enner. It has been cultured in laboratories to produce small pox vacine. It is not possible to establish if the virus was a. genuine cow pox virus or a naturally occuring vaccinia virus. The detailed antigenic structure seems to indicate that vaccinia is more closely related to variola than to cow pox. The vaccinia virus is widespread in animals and cause local affections and generalised diesase. The infection is also transmitted from affected animals to human beings. The virulent strains are particularly dangerous to immunosuppressed persons.
Properties of the }'irus: In the genus of orthopoxvirus, all members are interrelated as found by cross protection, neutralization and other serological tests. Ectromelia shows the lowest degree of relationship. The vaccinia virus is labile to chloroforms and ether. The scabs remain infected for several weeks in the tropical environment. The cell free virus is inactivated at 50°C in 1-2 hours.
The virus IS serologically uniform. The virus particle contains about 17 antigens which fall in 3 groups. (i) Heat labile antigens (L) (ii) Heat stable antigens(S) (iii) Nucleoprotein antigens (NP). The NP antigen crossreacts with other poxviruses. The protective antigens are localisd in the virus coat.
Cultivation: Vaccinia virus grows well in the developing chicken embryos by chorioallantoic route of inoculation, producing pock lesions. The pocks are discrete when inoculum is diluted. The virus can also be grown in cell culture like chicken embryo fibroblast (CEF), rabbit kidney, rabbit testes etc and cell lines like HeLa and L cells. Cytopathic effect (CPE) appears after 48 hours of inoculation and

132 Textbook of Veterinary Virology
includes fonnation of giant cells, basophilic and eosinophilic inclusion bodies in the cytoplasm.
Vaccinia virus was initially propagated in calves for vaccine productio~. The virus was also propagated in sheep, goats, rabbits and donkeys.
Epidemiology: During small pox vaccination period among human beings, the infections among animals originated from children after primary vaccination. The pustules after becoming dry, fall as scabs and survive for a long time. The dried scabs get mixed with dust and spread in the fonn of airborne particles.
Pathogenesis: Vaccinia virus infections in animals develop as local, generalized or latent infection. The local infection is most common, lesions develop on restricted areas of mucous membranes, skin and on other organs. A generalised disease develops in immunosuppressed animals, there is massive multiplication of virus in the primary target organs which results in viraemia and lesions develop on the skin, mucous membranes and internal organs. The latent infections are rare and virus occasionally persists in lymphoreticular tissues without being secreted.
The typical pustules which develop on the skin or mucous membranes pass from efforscences into papules, blisters and subsequently into pustules when necrosis of blisters, central region occurs. The pustules dry up and fall as scabs after several days of period. In internal organs a localized proliferation occurs at the site of virus multiplication followed by ballooning degeneration and lysis of affected cells. The intracytoplasmic A & B types of inclusion bodies can be detected.
After an incubation period of 3-7 days the lesions develop in about 5 days. If there is no bacterial infection the pustules dry up and fall off after one week. The lesions are mostly confined to mucous membranes, skin of the muzzle udder and rarely in the genital tract.
Immune reaction: After recovery the animals acquire strong immunity. Bot~ cellular and humoral immunity play a part in protection. The humoral immunity is transferred from mother to newborn through colostrum.
Diagnosis: The quick diagnosis can be done by electron microscope detection of vaccinia virus Particles in samples of skin or mucous membrane pustules. The diagnosis can also be arrived at by

Poxviridae 133
CAM inoculation of chicken embryos for appearance of pock lesions or by inoculation of cell cultures to observe typical CPE or detection of immunofluorescene. The vaccinia virus can be distinguished from cowpox virus by inoculation of chickens by feather follicle methods. The vaccinia virus produces typical localized pock lesions while there is no reaction in case of cowpox virus. The virus can be distinguished from parapox viruses by electron microscopy, pock morphology on the CAM, and CPE in cell cultures and immunofluorescence and other serological methods. The detection of antibodies· in paired serum samples by HI and neutralization also confirms the diagnosis.
Cow Pox Virus
The cowpox is a benign contagious disease of cattle which is distinct form the viruses of vaccinia and milkers nodule. The disease occurs mostly is milking cows and the lesions restricted to particular areas of animal body like muzzle, udder, testes or scrotum on the one hand and systemic disease with pock covering the whole body on the other hand. The disease is transmitted from affected animals to human beings. Like vaccinia virus, cow pox virus has a wide host range in laboratory animals.
Properties of the virus: The morphology and physico-chemical characters of the virus are similar to vaccinia(Table 1). The cowpox DNA is double stranded with a MW of 80 x 106 daItons. The virus particle is brick shaped with a diameter of 250 x 300 nm. Cow pox virus is very closely related to vaccinia and cannot be differentiated by complement fixation test There are reports indicating the presence of distinct biological strains. The virus particle has nucleoprotein (NP) and LS antigen like vaccinia. The haemagglutinin of cowpox virus clumps turkey and fowl red blood cells. Immunologically, cowpox virus is a single serotype. The serological test with monoclonal antibodies are used to differentiate cowpox virus from other orthopox viruses.
Cultivation: The virus grows well on the chorioallantoic membrane of 7-13 day old embryonated eggs. The pocks are characterized by an intense haemorrhagic reaction. The virus can alSO
be grown in cell culture of chicken, human and bovine cells and produces A & B type inclusions. The virus re~dily infects guinea pig, mouse, monkey and many other species including man.

134 Textbook o/Veterinary Virology
Epidemiology: Cow pox is mostly transmitted by direct contact The virus passes into the environment through dried pustules from skin and mucous membranes, secretions from eyes, respiratory tract and digestive tract The virus enters the body through broken skin or via the mucous membranes of upper respiratory and digestive tract. The virus is indirectly transmitted during milking. A large number of domestic and zoo animals are susceptible to cow pox virus. The virus is also transmitted to humans, where it causes local affections in the form of pocks on hands, arms and face. Systemic disease with fever, lymphangitis, lymphadenitis, conjuncti-vitis and disseminated pox exanthema may also develop. In recent years the disease has been reported in human with no contact with cattle. It is, therefore, suspected that reservoirs of cowpox infections have been observed in various carnivorous species (Felidae), cats, leopards, lions, -ocelots, cheetahs, jaguars etc. The infection has also been reported from laboratory rats. The infection with virus similar to cowpox also ocur in wild rats.
The virus is excreted in urine and faeces. It has been concluded by Marennkikova et al (19841 that various species of wild rodents are a hidden natural reservoir for cowpox virus, maintaining the chain of infection and final hosts being ungulates, carnivores and humans.
Palhogenesis: The pathogenesis of cowpox is similar to vaccinia virus infection. The incubation period is 3-7 days. The cattle may show slight fever and anorexia followed by appearance of lesions on teats and udder which pass through all stages of pox infection. True cow pox lesions are 1-2 cm in diameter and are thick, tenacious and yellow brown to red in colour. The lesions are mostly confined to teats and lower parts of udder. In severe cases the lesions may spread to inside of the thighs and rarely to perineum, vulva and mouth. In bulls, the lesions u~ually appear on the scrotum. Recovery results in an active immunity and calves born from immune dams acquire protection through colostrum. In man the diseaSe is self limiting and benign but in unvaccinated adults the disease may be severe. Cytoplasmic inclusion bodies (acidophilic)' are present in the infected tissue. In comparision to vaccinia inclusion bodies, they are large, compact, less, granular and occur only in small numbers.
Immune reaction: The recovered animals have a long lasting immuity, more than vaccinia virus infection. The cellular immunity disappears faster than Immoral immunity. Newborn animals receive maternal antibodies through colostrum.

Roxviridae 135
Diagnosis: The diagnosis is based on the appearance of finn dark red circular lesions with raised edges and depressed centres on the teats and udder. Confinnatory diagnosis can be arrived at by isolation of the virus from lesiogs and carrying out serological tests for identification of virus. The appearance of haemorrhagic pock lesions on the CAM of embryonated eggs is of diagnositc value.
Control: There is no specific method of control of cow pox. The use of live virus vaccines for healthy animals after an outbreak of cowpox in a herd of zoo animals is preferred. Either attenuated strains of cow pox or attenuated vaccinia virus strains should be used For primary vaccination two parenteral injections at an interval of 3-4 weeks are recommended.
Buffalo Pox Virus
It is a mild viral disease of buffaloes with lesions on teats, udder, thighs and in few cases in lips and nostrils. The disease affects buffaloes only. The disease was reported for the first time in India by Sharma (1934). Since then the disease has been reported from many states of this country. The disease has also been reported from Indonesia, Italy, Pakistan, Russia and Egypt.
Properties of the virus: The buffalo pox virus resembles vaccinia in morphology. The size of the mature virus particle varies from 280 to 230 nm x 200 to 250 om. The virus particles are brick shaped with a lipid coat and complex symmetry. The virus is heat labiled at 56°C for 30 minutes the virus loses its infectiyity. The virus is sensitive to chloroform and bile salts but resistant to ether. There is a close antigenic relationship between buffalopox, cowpox and vaccinia viruses. There is no serological relationship between buffalopox, sheep pox, goat pox and camel pox viruses. A common group (NP) antigen reacts with antisera of sheep pox, pig pox and fowl pox viruses. There is complete cross protection between cowpox and buffalo pox. The buffalopox virus contains at least 4 major components, protein soluble antigen (LS), nucleoprotein antigen (NP), haemagglutination.,(HA) and the factors responsible for infectivity. The HA can be inhibited both by buffalo pox and vaccinia virus antiserum. It is immunologically uniform and cross reacts both with vaccina and cowpox as well as with other orthopoxivurses. It is more closely related to cowpox and vaccinia then to other orthopox viruses. However, the biological properties of buffalo pox virus differ from those of cow pox and vaccinia viruses.

136 Textbook o/Veterinary Virology
Cultivation: The virus grows well on the CAM of 12 day old chicken embryos producing large raised or flat, white or grey pocks, without haemorrhage. Suckling mice is susceptible, the virus can also be cultivated in chicken embryo fibroblast and HeLa cells.
Epidemiology: The virus is species specific. Experimentally buffalo calves, rabbits, guinea pigs and suckling mice can be infected. The virus can infect man through contact with infected buffaloes. The mode of transmission and other epidemiological features appear to be the same as in cowpox.
Pathogenesis: The ~sease occurs in epidemic forms in generalised and localised forms. The lesions are seen on the teats, udder, medial aspect of thigh and sometimes on the quarters, lips and around the nostrils. In generalised form, the lesions are seen all over the body. Thickening of teats, stenosis of milk duck and mastitis are common sequelae.
Diagnosis: The diagnosis is based on the clinical symptoms and isolation and characterization of the virus.
COlltrol: No vaccine is available to control the disease.
Camel Pox Virus
Camel pox is a severe disease of camels and mainly affects the young animals. The disease is characterised by generalised exanthema. The disease resembles small pox in man rather than cow pox. Camelpox has been known since Middle Ages. The disease is prevalent in India, Pakistan, Egypt, Iran, Iraq, Kenya, Algeria, the Sahara, Sudan, Somalia and Soviet Union. The disease results in heavy losses in these countries amongst animals aged 2-3 years almost every year. The disease causes high mortality loss in condition and fall in milk and meat production.
Properties of the virus: Camel pox virus is a separate species in orthopox virus genus. The morphological and biological properties are similar to other orthopoxviruses. The virus of camel pox is brick shaped and measures 280 x 100 nm. It is highly resistant to natural environment; the virus is inactivated by dry heat of lOO-110°C in 1 hours. Some strains are sensitive to chloroform but resistant to ether. Different strains of virus isolated from different parlS of world are similar antigenically. The strains isolated from Iran and USSR show haemagglutinating activity while the Egyptian strain did not reveal this

Poxviridae 137
property. The cross neutralization tests showed an immunologic relationship between camel pox, variola a'ld cowpox viruses.
Cultimlion: The camel pox virus grows on CAM of chicken embryos and majority of strains produce small, opaque and white pocks. Some strains also produce haemorrhagic pocks. The virus also grows on a variety of cells like kidney and testis of calf, camel, lamb, pig and chicken embryo fibroblast and BHK cells. The CPE includes production of giant cells.
Epidemiology: Camelpox is transmitted both directly and indirectly. The virus contaminates the environment when pustulates in the skin and mucous membranes become dry and fall off. The secretions of eye and upper respiratory and digestive tract is also important in the spread of virus. Virus in dry scabs and crusts remains infectious for 4-50 months or longer. A few cases of human infections have been reported but these reports are not reliable.
Camel pox virus spreads rapidly in camel populations and affects young animals in particular. There is high mortality rate besides loss in conditon and milk
Palhogenesis: Camel pox virus is pathogenic for chicken embryos and mice by intracerebral inoculation. The incubation period is to-IS days. Benign as well as malignant forms of disease occur. The foals are more susceptible than the adult animals. In young animals the disease appears to be generalised. The lesions appear on the lips, nose, eyelids and other parts of the body. The pustules may also appear in the mUf:ous membrane of mouth and nose, Corneal opacity may also occur. The mortality rate varies from 4 to 7% in young animals.
Diagnosis: The diagnosis depends upon the history, course of disease and clinical signs. Confirmation is by isolation and characterisation of virus based on pock morphology on CAM, pathogenicity of adult mice by intracerebral routes and CPE in cell cultures resulting in gi:mt cell formation etc.
Control: No prophylactic vaccine is available at present. Attenuated vaccinia strains are recommended. For primary vaccination, two doses at an interval of 3-4 weeks of MV A vaccinia strain are recommended. Revaccination at an interval of 2-3 years is sufficient.
Some camel owning tribes still use 'Variolation' for protecting the animals. The dried scabs from affected animals arc collected and dried and kept for about one year. The dried scabs in milk are inoculated -in the lips of animals.
138 Textbook of Veterinary Virology
Table 13.2 DIFFERENTIAL CHARACTERISTICS OF ORTIIOPOXVIRUSES
S. Characteristics No.
1. Isolated from
2. Pock morphology on CAM
3. Ceiling temperature for CAM growth (0C)
4. Inclusion type 5. Antigenic specificity 6. Pathogenicity
Baby mice Chick mortality
v - Variola a
Sheep Pox Virus
Vaccinia Cowpox
Man Cow,man and large felines
Opaque, Haemorr-white flat hagic haemorr-hagic ulcer
41.0 40.0
B BandA
Va'V. V.
+++ +++ +++ t+
V. - Vaccinia
Buffalo Came/pox
Buffalo, Camel man
Large Small opaque, opaque white white hearnorr-hagic ulcer 38.5 38.5
B B Va,Vc Va, Vc
+++ + + ±
Sheep pox (SP) is a highly contagious and often fatal disease of sheep. The lesions in generalized cases are associated with haemorrhagic inflammation of respiratory and gastrointestinal mucosa and high mortality. The earliest record of disease dates back to the 2nd century using serological tests. The animals recovered from infection with one isolate were resistant to challenge with any other isolates, whether derived from sheep or goat. The genome of sheep pox, goatpox and lumpy skin disease of cattle are very closely related. The term 'capripox' is proposed to include the viruses of these pox diseases of ruminants.
Cultivation: The virus can be cultivated in sheep. The reports of cultivation in chick embryo are conflicting. SPY produces CPE in various cell cultures of sheep origin like testes, embryonic kidney and thyroid as well as in goat testes and kidney and bovine testes and

Poxviridae 139
kidney. The virus produces inclusion bodies in the cytoplasm of cells which are basophilic and/or eosinophilic in nature.
Epidemiology: The disease spreads by contact with infected sheep, their aerosals, nasal secretions, saliva or dried scabs. The air borne infection spreads to 20-25 yards. The virus may remain viable in wool for 2 months and on contaminated pens upto 6 months. Mechanical transmission by biting insects may play a part.
Pathogenesis: The incubation period varies from 4-7 days. The disease usually causes systemic reaction, with high temperature and eruptions on the cheeks, nostrils, lips, ears, groins, udder and lips of vulva. The lesions begin as macules with slight oedema of sUITouding skin. The macules develop into papllles and then become pustules. The pustules mayor may not become vesicles. In more severe cases, lesions in buccal, digestive and respiratory mucosa also develop. The mom lity rate may reach upto 80%. When the clinical symptoms subside, the temperature falls and during resolution of pocks, there is itching and shedding of wool and exfoliation of epidermis. The course of the disease is 3-4 weeks. The postmortem examination reveals haemorrhagic inflammation and ulcers in mucous membrane of respiratory and gastro-intestinal tracts. Nodules may be present in lungs and kidneys. There is gelatinous oedema of subcutaneous and intramuscular tissues. The incubation period in natural infection is 4-8 days.
Diagnosis: The clinical diagnosis is based on the history and the symptoms. The confirmation is based on the isolation of virus in susceptible sheep or cell cultures. The detection of antibodies in the serum of recovered animals can be done by neutralization, complement fIxation, gel diffusion and fluorescent antibody tests.
Control: Animals which recover from natural infection develop solid immunity. Ovination had been in use since 18th century. Infected scabs powder in glycerine saline had been in uS'e in this country. Serum virus mixture had an improvement over ovination but there is risk of actual spread of disease. Inactivated vaccines even when incorporated with aluminium hydroxide as adjuvallt do not produce an effIcient immunity. A chinese strain of sheep pox adapted to CAM of chicken embryos is reported to be avirulent and induces a high level of protective immunity. In Russia an avirulent strain (K) grown on CAM of chicken embryos is reported to produce immunity upto 5 months.

140 Textbook of Veterinary Virology
The latest vaccine developed from Kenya isolate having no clear preference for sheep and goats. The isolate is attenuated in lambs testis and BHK cells. The vaccine is equally protective in sheep and goat pox. The trials carried out in Middle Ea~;t have proved the efficacy of new vaccine
Goat Pox Virus
Goat pox is an epizootic disease resembling sheep pox. The disease is characterised by fever, mucopurulent nasa! discharge and generalized cutaneous eruptions. The disease is preva;~nt in North Africa, the Middle East, Parts of Europe, India and the Far East. In India the disease is of economic significance causing significant losses in goat producing regions. Naturally the virus affects the goats but there are reports that some strains may produce disease among sheep also. Human beings may get a mild disease.
Properties of the virus: Not much is known about this virus but many workers agree that the virus is similar to sheep pox. The virus remains viabl~ for many months in the dried state. The virus is ether sensitive. Goat pox and sheep pox viruses share a common antigen as revealed by immunodiffusion and complement fixation tests. Some cross reaction has also been reported between goat pox and contagious pustular, dermatitis virus, cross protection between sheep and goat pox has also been reported by some workers.
Cultivation: The goat pox virus grows readily in cell cultures of lamb kidney, kid kidney, and testis and produce CPE similar to sheep pox. The reports regarding cultivation of goat pox virus on CAM of chicken embryos are conflicting. Few strains may be able to grow on the CAM of the embryonated eggs.
Epidemiology: The virus is host specific but some strains are believed to infect sheep as well. The disease spreads by contact with the infected animals or infected fomites. The disease may also be spread by insects mechanically.
Pathogensis: Following incubation period of 5-14 days small papules appear mostly on the hairless parts of the body like nose, mouth, around the eyes, udder, testes, scrotum, inner aspect of thighs. The lesions develop into vesicles and crusts and subsequently heal in 3-4 weeks. The disease is more common in young animals or in lactating animals. The mortality rate varies from 0 to 50%.

Poxviridae 141
Diagnosis: Diagnosis is based on the course of the disease and clinical symptoms. Confmnatory diagnosis can be arrived at by isolation of virus in cell culture and further characterization. Gel diffusion test is helpful in detecting specific precipitnogens in affected tissues.
Control: Chicken embryo adapted strains are being used in certain countries. Goat pox strains attenuated in cell culture are being developed.
Lumpy Skin Disease
It is an acute febrile disca"se of cattle which is characterised by eruption of skin nodules of varying sizes. There is drop in milk yield and abortions occur among pregnant animals. The morbidity rate varies from 5-45% but mortality rate never exceeds 1 per cent. The disease is known to affect cattle only in different countries of African continent. The natural infection and transmission mode is not known. The virus resembles vaccinia in morphology. The virus can be adapted on the CAM of chicken embryo and in a wide variety of cell cultures. Strains of sheep pox virus adapted in cell culture give good immunity and have proved to be useful in controlling the disease.
Ecthyma (Ort) Virus
Orf or cC>ntagious pustular dermatitis or contagious ecthyma (CE) is highly infectious disease of sheep and goats chamcterised by d~velopment of pustular and scabby lesions on muzzle aRd lips. The disease is prevalent all over the world. The incidence among kids and lambs of 4-6 months is very high. In this country the disease occurs wherever sheep and goats are raised. The disease is transmitted to man, where it results in local infections.
Properties of the virus: The virus particles are elongated oval measuring 250 x 160 nm. The virion consists of thick outer wall and centrally placed core. The negatively stained particles look like a ball of wool. The arrangement of filaments on the outer membrane is different than orthopoxviruses. The tubular subunits are arranged regularly like a single coiled up rope. Lateraly'body surface protein and internal body are smaller. The parapox viruses replicate in their own factories in the cytoplasm where 'virions are found in several developmental stages. The structural elements of orf and vaccinia are very much alike. The virus resists desication and survives for years in

142 Textbook o/Veterinary Virology
dried scabs. The virus is inactivated by chlorofonn but is only slightly sensitive to ether.· Haemagglutination has not been reported. The virus has antigenic cross reaction with other members of parapox genus like pseudocowpox. Some antigenic relationship has been reported between orf virus and sheep and goat pox viruses but orf virus appeared to be nearer to sheep pox rather than goat pox. All orf strains isolated from sheep, goats and man are immunologically uniform. Restriction analysis revealed genetic heterogenicity among virus isolates.
The virus has been reported to grow in chicken embryos. It grows well in bovine and ovine cell cultures. Testis cells are particularly suitable, the CPE includes formation of granular cytoplasmic inclusions consisting of acidophiloc paranuclear mass and irregular basophilic peripheral area The virus also grows in rabbits but other laboratory animals are not susceptible.
Epidemiology: Orf is highly cont<'lgious among sheep and goats. The transmission is by contact, aerogenic, Or virus remaining infectious in the ground in the dried scabs. The virus is indireclty transmitted by way of carcasses or processed meat from latently infected animals or slaughtered during viraemic phase. The virus can also be transmitted to lambs by infected ews at birth or later. Latently infected sheep and goats are virus reservoirs. Contagious ecthyma is of zoonotic importance as human beings also get infected. The virus induces proliferative local lesions on the skin and mucous membranes.
The mortality in sheep and goats is usually not more than 1 % but morbidity reaches 100%. Early weaning; crowding weaken the animals defences and increase the virulence of virus as a result of rapid passages. This gives rise to more severe form of disease with mortality ranging 20-50%.
Pathogenesis: The disease mainly affects lambs and kids and runs a cyclic course. The virus enters the body through skin or mucous membranes. After primary multiplication of virus at portal of entry, viraemia develops and primary organs of affinity become infected. The virus gets generalised in second viraemic phase. Lesions mostly occur on hairless parts of the body, lips, mouth and sometimes udder and interdigital region of the feet. Lesions like papules, pustules and ulcers develop in 34 days. The course of the disease is 3-4 weeks. Cytoplasmic inclusion bodies occur in the affected cells. Papular lesions also develop in human beings.

Poxviridae 143
Diagnosis: Diagnosis can be made on the clinical symptoms and contagious nature of disease. The antigen in the lesions can be detected by gel diffusion and complement fIxation test The rapid and reliable diagnosis can be made by electron microscopy.
Control: The live virus vaccine is available in certain countries. Lambs and kids are vaccinated at 1 month of age and again revaccinated 2-3 months later.
Bovine Papular Stomatitis Virus
Bovine papular stomatitis is a mild generalised viral disease of cattle. The disease is widespread and occurs world-wide. Latent infections are frequent in cattle. The virus belongs to genus parapoxvirus. It is similar to ovine orf virus antigenically and morphologically. The clinical and physical properties resemble those of orfvirus.
Bovine papular stomatitis is a cyclic generalised disease. When the latent infection is activated the disease produces lesions on the mucous membrane of the muzzle palate, oesophagus, rumen and on the udder. The incubation period is 2-5 days. The disease is mild, initial erythyematous changes on mucous membrane of the muzzle develop into erosive and proliferative stomatitis. There is no formation of blisters. The rapid and reliable diagnosis can be made by electron microscope. The virus can be propaged in cell cultures of foetal lung, testes and kidney of cattle or sheep, CPE with gaint cell formatio!1 and granular lysiS is produced. The viral antigen is detected by immuofluorescence. No specific preventive measures are available against this virus.
Milker'S Node Virus
Milker's node is a zoonotic disease transmitted to man by close contact with cows, sheep and goat. The lesions on the udders of cows frequently transmit the disease to milkers. The virus can be propagated in in cell cultures of foetal lungs, kidneys, skin from cows and sheep. A characteristic CPE with giant cell formation and lysis is produced.
Swine Pox Virus
Swine pox is usually a benign disease of young pigs characterised by transient fever and typical pox like lesion on the body. The disease

144 Textbook o/Veterinary Virology
occurs naturally in pigs especially young and suckling piglets. The disease is world wide in distribution.
Properties of the virus: The disease is caused by 1 antigenically different viruses. One is probably vaccinia and the other is true swine pox virus.' The swine pox virus is similar in morphology to other pox viruses measuring 320 X 280 nm. The virus is relatively heat stable and survives'lO-12 days at 37°C. The virus of swine pox is antigenically distinct from vaccinia although by agar gel diffusion test a minor component is shared with vaccinia virus. There is no cross protection between vaccinia and swine pox.
Cultivation: The virus does not grow on the CAM of chicken embryos. It replicates only in cell culture of swine origin like pig kidney, pig testes, embryonic porcine lungs and brains. The CPE consists of nuclear vacuolation and intracytoplasmic inclusion bodies.
Epidemiology: The virus is transmitted by contact rarely as biting insects play a role in mechanical transmission of the virus. The pig louse is of major importance in transmission of the disease where the virus survives for a year. Other insects arc also involved in the mode of transmission.
Palhogenesis: The incubation period in swine pox is 3-7 days. The disease is accompanied by slight fever for 1-7 days. Small papules especially on the ventral abdomen appear. The lesions may also appear on the inner aspect of limbs. The papules develop into vesciles and subsequently scab formation takes place. The healing usually take 2-4 weeks. The mortality rate is very low and rarely rises upto 3 per cent.
The disease produced by vaccinia is similar to the disease produced by swine pox virus.
Diagnosis: The disease is usually diagnosed on clinical grounds. The infestation of pigs with lice is suggestive of swine pox. Virus isolation may be attempted and differentiation of pig pox and vaccinia be done on serological tests. Immunodiffusion test with swinepox antigen gives 2 precipitating lines with swinepox convalesent serum.
Control: Recovered animals develop solid immunity. No vaccination is carried out. The best control is by good hygiene and eradication of lice from the swine.
Myxoma Virus
Myxomavirus causes an infectious disease of rabbits. The virus

Poxviridae 145
causes a fatal disease in domestic, Belgian and Angora rabbits, however, cottontail and jackrabbits are resistant.
Properties of the virus: The virus is rectangular in shape which .is similar to vaccinia and measure about 280 x 230 nm. The negatively stained particles show a beaded appearance and consist of mass of threads or tubules. The virus is sensitive to ether and to a pH less than 4.6. It is susceptible to heat and inactivated at 55°C. It is resistant to sodium deoxycholate and survives for many months in the skin of infected animals. The virus cross reacts with rabbit fibroma, hare fibroma and squirrel fibroma viruses in immunodiffusion test. The virus grows and produces CPE in a vareity of cells like rabbit, guinea pig, squirrel, chicken and human tissues. It produces microscopic plaques in rabbit kidney monolayer cell cultures. The virus produces pocks on the CAM of chicken embryos. The virus can be propagated in the brain of suckling mice without showing clinical signs.
Palhogenecily: Following infection in rabbits there is high fever and conjunctivitis followed by generalized tumor like swelling on the skin. The death may take place in 48 hours with virulent strains. The animals that survive show generalised oedema of head, nose and lips. The mortality varies from 25 to 90%.
Epidemiology: The mosquitoes or the fleas act as mechanical vectors and possibly by direct contact
Diagnosis: A diagnosis can be made from characteristic clinical signs and classical lesions and high mortality. The virus isolation can be done in cell cultures and chicken embryos.
Control: Live attenuated vaccines are used. Shope's fibroma virus, used as a vaccine, causes a local fibroma and cross protects against myxomatosis. The rabbits should be protected from mosquitoes and fleas. The disease is of seasonal nature.
Fibroma Virus
Fibroma virus is immunologically closely related to myxoma virus. The vaccination of rabbits with fibroma virus confers protection against myxomatosis. The natural transmission probably takes place by mosquitoes or other biting insects as in myxomavirus. In natural infections in cotton tail and domestic rabbits, firm spherical loosely attached turnors occur on the skin and are usually found on the foot. In domestic rabbits the growth is rapid and regression takes place 10-12 days after infection and quickly disappears.

146 Textbook o/Veterinary Virology
Fowl Pox Virus
Fowl pox also known as avian diphteria is a serious disease causing heavy economic losses. The disease occurs in tow forms:cutaneous and diphtheritic. The disease mainly affects chicken but other poul'try like turkeys, guinea fowl, peacocks, pigeons, water fowl, pheasants, canaries and sparrows may be affected. The disease is prevalent in all the countries of the world and is widespread in India where outbreaks occur.
In recent past several workers have reported the breakdown of immunity in fowl pox vaccinated flocks, heavy economic losses in terms of mortality, drop in egg production and meat occur due to fowl pox in this country.
Properties of the virus: The elementary bodies or Borrel bodies resemble in morphology to other pox viruses and measure about 330 x 200 nm. The virus is resistant to drying and is inactivated at 50°C in 30 minutes. The virus withstands 1 percent phenol and 1 in 100 formalin for 9 days but is inactivated readily by 1-2 per cent NaOH. The virion is inactivated by chloroform but is resitant to ether: however, some strains have been reported to be moderately sensitive. The avian pox viruses are not related to other mammalian pox viruses although NP antigen is shared by all pox viruses. There is cross neutralization among avian pox viruses. Fowl pox virus is closely related to pigeon pox. Fowl pox virus haemagglutinates fowl rbc and also exhibits haemadsorption. During the .work conducted at Punjab Agricultural Universitx on six strains of fowl pox virus, the strains were compared with SDS-acrylamide gel electrophoresis. The result revealed some antigenic differences both in number of protein bands and total molecular weight. Antigenic separation of the three strains of fowl pox in DEAE-cellulose column also revealed slight differences in their elution profile. Some difference in extracellular and intracellular virus has also been reported.
Cultivation: The virus grows readily on CAM of developing chicken embryos and produces pock lesions. The virus also grows on chicken embryos fibroblasts and produces intracytoplasmic inclusions. The virus has also been adapted in chicken embryo dermis cells.
Epidemiology: The disease affects all ages but is mostly seen in 5-12 months old chickens, although chickens few days or few weeks old also get affected. The virus is very stable and survives in dried scabs

Poxviridae 147
and exudates for many months. The infection occurs through mechanical transmission of the virus to injured skin. The virus does not penetrate the intact skin. Blood sucking mthropods like mosquitoes of the genera culex and aedes transmit the virus from bird to bird. The mosquitoes remain infective for several weeks. Virus may also enter through minor abrasions in the mouth caused by rough feed or through feather follicles and also through injuries to comb, wattles or skin as a result of fighting, pecking, scratching etc. Poultry ticks or flies may also take part in transmission.
Pathogenesis: The virus mainl>: affects chickens of all ages but chickens are more susceptible from 5-12 months of age. Other poultry such as turkeys, guinea fowl etc. may be naturally or experimentally infected. Although avian poxes have been isolated from number of avian species but each avian pox is more pathogenic for its own host. The incubation period is 4-10 days. The characteristic lesions of cutaneous form arc mostly found on the comb, wallies, legs, feet, vent and around the eyes. The lesions first appear as white foci which develop rapidly into nodules. The lesions may coal~sce and become rough and dark brown. The base of the lesions are inflammed and become haemorrhagic after about -2 weeks. After another one week or so scab formation takes place. In the diptheritic form, slightly elevated white opaque nodules develop into mucous membranes of pharynx and larynx resulting respiratory distress of the host.
Diagnosis: Typical lesions in comb, wattles, legs etc. are suggestive of the disease diagnosis. The diptheritic form of disease has to be identified from other disease causing respiratory distress. The confirmatory diagnosis can be arrived at by isolating the virus in chicken emrbyos or cell culture. The neutralization test is not practical in convalescent serum samples because the concentration of neutralising antibodies is low. The specific antibodies can be rev('aled by complement fixation and immunodiffusion tests.
Control: The recovered birds develop a solid immunity. Two types of live virus vaccines are in use, fowl pox and pigeon pox. These vaccines arc prepared from infected CAM of chicken embryos. Fowl pox vaccine prepared in tissue culture is also in use. Fowl pox vaccine contains live attenuated virus of chicken embryos or tissue culture origin and can be given to birds by feather follicle or wing web method. The age at vaccination by cutaneous method is 4 to 8 wecks. The oral

148 Textbook of Veterinary Virology
route of va~cination gives successful results with certain strains like HPl (German) and this can be effectively used even when the chicks are 5 days old.
Pigeon pox vaccine of chicken embryo origin does not provide a long lasting protection though it is comparatively safer. The vaccination Can be done even in younger chicks less than 4 weeks old as well as in laying flocks.
References
BAXBY, DEVRlcK, 1984. Poxviruses. In Principles of bacteriology, Virology and Immunity Vol.4. Edited by F.Brown and Sir Graham, WiIson, Edward Amold, London.
BLACK, D.N., 1986. The capripox virus genome. Revue Scientifique a Technique OIE. 5, 495-501.
BUXTON, A. and FRASER, G., 1977. Animal Microbiology Vol.2. Oxford Blackwell Scientific Publications.
FERNANDES, G., SHARMA, S.N. and TANWANI, S.K., 1981. Studies on tissue culture adapted fowl pox disease vaccine with particular reference to immuno-genicity of intracellular and extracellular virus. Indian Veterinary Journal. 58: 599-604.
Firtal Report 1980-85. Development of suitable vaccine for sheeppox and characterization of its immune response in sheep. Department of Veterinary Microbiology and Public Heath. G.B.Pant University of Agriculture and Technology, Pantnagar.
Firtal Report 1980-83. Immunologic characterization of buffalo pox virus and immunity in buffalopox. Department of Veterinary Microbiology and Public Health. G.B.Pant University of Agriculture and Technology, Pantnagar.
Firtal Report 1983-88. Biochemical and biological studies on fowl pox virus with a view to evolve a suitable vaccine. Department of Veterinary Bacterio-Iogy and Virology, Pubjab Agricultural University, Ludhiana.
KrrcHING, R.P., 1986. The control of sheep and goat pox. Revue Scientifique et Technique OIE. 5,503-511.
LAL, S.M. and SINGH, I.P., 1977. Buffalopox-A review. Tropical Animal Health and Production 2: 107-12.

Poxviridae 149
Virus infections of ruminants, 1990. Edited by Z.Dinter and B.Morein. Elsevier
Science Publishers B.V.
Mohanty, S.B. and Dutta, S.K., 1981. Veterinary Virology. Lea and Febiger, Philadelphia.
NAKANo, J.H., 1977. Comparative diagnosis of poxvirus diseases. In comparative diagnosis of viral diseases VoU. Academic Press Inc. New Yorl.
P ANDEY, R.; KAUSIIIK, A.K. and GROVER, Y.P., 1985. Biology of Ortlwpox virus infections of domestic ruminants. In Progress in Veterinary Microbiology and Immunology. Edited by R.Pandey, S.Karger, Basel. Vol1, pp. 199-228.
SHARMA, S.N., Nn..AKANTAN, P.R. and DHANDA, M.R., 1966. A preliminary note on pathogenicity and antigenicity of sheep and goat pox viruses. Indian Veterinary Journal. 43,673-78.
SINGH, LP., PANDEY, B and SRIVASTAVA, R.N., 1979. Sheep pox: A review. Veterinary Bulletin: 49: 145-53.
TANTAWJ, H.H., 1974. Comparative studies of camel pox, sheep pox and vaccinia viruses. Acta Virologica, 15: 347-381.
TAYLOR, W.P., 1985. Clinical and antigenic relationship between isolates of sheep and goat pox viruses. Tropical Animal Health Biology 17: 64-79.
TRlPATHY, D.N. and HANSON, L.E., 1975. Immunity to fowlpox. American Journal of Veterinary Research. 36: 541-44.
TRIPATHY, D.N.; HANSON, L.E.; CARANDFLL, B.A., 1981. Pox viruses of Veterinary Importance diagnosis of infections. In Kurstak, comparative diagnosis of viral disease. Vol. 3A, Academic Press London.

Chapter 14
Parvoviridae
The members of this group are small sized non enveloped viruses measuring 18-26 nm in diameter and consist of an icosahedral capsid and probably 32 capsomeres. The genomes consists of a single molecule of single stranded DNA with a moleeular weight of 1.5-2 x 1()6 daltons. Members of genus parvovirus has a (-) sense DNA. In some genera the single strands of both polarities are encepsidated and form double stranded molecules upon nucleic acid extraction. In addition genus parvovirus the Dependovirus and Densovirus belong to this family and found in the insects. Most of the infectious agent of Veterinary importance belong to genus Parvovirus.
Members of the genus Parvovirus replicate in the nucleus of cycling cells. The replication is dependent either on certain helper functions from the host cell or from helper virus (Dependovirus). Two main overlapping transcription units have been identified, from which 3 major mRNA species are transcribed. DNA replication occurs via a double stranded replicative form initiated by self priming mechanism. Following DNA replication, viral capsids assemble into which ss DNA is packed. Accumulation of both empty and full progeny virions can be found in the nuclei of infected cells. Intranuclear inclusion bodies can occasionally be seen in infected cells.
Parvoviruses are remarkably stable to environmental conditions, hence disinfection of contaminated premises is difficult to achieve. They resist 60o~ for about 1 hour and are stable at pH 3-9. The pathogenesis of parvoviruses is determined by the age of the infected animal. Following the infection of foetus (Pig or cat) or newborn (dog

voviridae 151
or cat). the virus may be pantropic. In older animals a narrower range of cells is affected. Following natural inf~on there is rapid immune response.
Bovine Parvoviruses (BPOV)
In cattle two genera-Parvovirus and Dependovirus are found. Bovine parvoviruses are associated with diarrhoea in calves and some problems like abortions. Members of genus dependovirus are defective but multiply in the presence of adenovirus or herpesvirus. Dependovirus have not been found to be associated with pathogenic conditions.
In 1959 Abinanti and Warfield isolated the virus from the intestine of calves. The virus was later identified (1970) as parvovirus. The strain was called HADEN (Haem ADsorbing ENteric) virus (type-I. A Japanese strain different from HADEN strain is considered to represent type-2.
Properties of the virus: The virus particles are small measuring 18-25 nm in diameter. They are extremely resistant to chemical and physical inactivating agents. The most reliable disinfection is achieved by. cholrax 0.5% or ethylene oxide (10% ethylene oxide and 90% CO
2).
All the isolates agglutinate guinea pig and human type 0 erythrocytes. The isolates from cattle are antigenically related or identical to prototype BPOV-l. One Japanese strain appears to be different and is separated as BPOV-2. Bovine parvoviruses differ antigenically from parvoviruses of man and animals.
Cultivation: The virus can grow in actively dividing cells. Primary cell cultures of bovine foetus spleen, kidney and lungs are used. Foetal calf testicle, lymph nodes and adrenal are equally suitable. The CPE in dividing foetal lungs is distinct and reproducible. After 24 hours of inoculation, the infected cells were swollen, refractile and exhibited a stellate appearance and ultimately showed rounding and detached from glass surface. Intranuclear inclusions with unique morphology are formed and can be seen after staining.
Epidemiology: The evidence of parvovirus infection of cattle by detection of antibodies (neutralising and haemagglutination) has been :reported from USA, England, Australia, Algiers, Japan and Brazil. The cattle are the reservoir and remain as source of infection for calves. Vertical transmission has been proved through detection of significant

152 Textbook o/Veterinary Virology
antibody titres in foetal serum and isolation of parvovirus from the tissues of naturally occuring abortion. This has been confirmed by experimental inoculations of pregnant cows or direct foetal inoculations.
Pathogenesis: The calves excrete the virus after 24-48 hours of experimental infection by oral as well as intravenous route. The virus can be isolated from mucosal scrappings of the duodenum and lower levels of intestine for 6 days. Viraemia associated with leukocytes develop after oral inoculation. Infections ofintestinal cells occur during systemic phase. The infection of cells in the cortex of adrenal glands, the thymus, lymph nodes and heart muscle become pronounced during systemic phase. Calves aged 2-8 months become diarrhoeic with 4-7 days of oral or intranasal infection while calves infected by intravenous route develop more severe diarrhoea.
The virus when inoculated intravenously in pregnant cows or by direct foetal inoculation, can be isolated from blood leucocytes 6 days post infection as well as from placenta and foetus. Foetuses of first trimester of gestation are highly susceptible. The foetuses of 2nd and 3rd trimester can also be infected. If foetuses survived, they developed antibodies and from the calves born alive, virus is isolated irregularly.
The BPOV seroreactor cows commonly experience higher titres of embryonic mortality and require more services per conception than did non reactor cattle.
Most calves with BPOV have catarrhal enteritis. Foetuses aborted during first and early second trimester of gestation are oedematous and have increased amounts of pleural and peritoneal fluid. Intranuclear inclusions are formed in the cells of small intestine, liver, lymph nodes and cerebellum.
Calves with naturally occuring infections show clinical signs in age from 1 week to 12 months. Calves show diarrhoea, the calves which survive diarrhoea develop antibodies but virus can be isolated intermittently from the faeces. The parvoviruses have also been isolated from faeces of clinically normal young cattle. The sick calves show a rise of temperature upto 41.2°C and are listless.
Diagnosis: The virus can be isolated in bovine foetal lung and bovine foetal spleen from faeces or intestinal scrappings. Fluorescent antibody test can be applied for locating viral antigen is tissues from infected animals and in cell cultures inoculated with suspected viral

Parvoviridae 153
material. The ID test can be used for detecting rising titres in the serum of animals.
Control: No vaccine is available at present.
Porcine Parvovirus (PPV)
The porcine parvoviruses are responsible for reproductive failures like abortions, still births, teratological defects, return to service and mummification. The PPV infection is now ubiquitous in pig herds throughout the world. The first isolation of the report of the virus was from swine fever virus vaccine in Germany in 1966.
Properties of the virus: The virion measures 20-28 nm in diameter. It has an icosahedral symmetry and is non enveloped. The virus withstands a temperature of 56°C for 30 minutes and is stable at pH 3-9 for 90 minutes. The virus is resistant to ether and chloroform. The virus agglutinates guinea pig, rat, chicken, rhesus monkey and human red blood cells. The haemaglutinating(HA) activity is dependent on temperature. The highest titres are obtained at 4°C. The different strains have been shown to be antigenically similar by ID and SN techniques.
Cultivation: The virus grows in young actively growing porcine, primary kidney cell cultures. Primary testicular, thyroid, foetal kidney and continuous cell lines have also been used. The virus produces CPE and intranculear inclusions as early as 16 hour post inoculation.
Pathogenecity: The virus appears to be primarily responsible for reproductive disorders in swine. The virus has been isolated from herds with a history of stillbirths, neonatal losses, infertility, abortions, myofibrillar hyperplasia, vaginal discharges from sows and semen with respiratory disorders and loss of condition. The virus has also been isolated from routine kidney cultures of normal piglets and foetuses. The PPV crosses the placenta of pregnant sows and aft~r artificial infection by oral, intramuscular and intravenous route. The sequel of infection depends on the time of gestation. The Pathogenic effects of the foetuses are restricted to gestational age of 45-50 days. Death usually occurs before the onset of immune-competence which is estimated at 70 days of ges~tion. Transplacental infection with virus and viral antibodies also take place. The virus is also suspected to cause infertility and congenital defects. The infection of postnatal animals mostly results in mild clinical or subclinical signs. In experimental cases, pneumonic lesions are also produced.

154 Textbook o/Veterinary Virology
Epidemiology: Majority of pig herds in UK and Australia are infected. PPV is resistant to adverse circumstances and persists in vacated pig houses for about 10 weeks. The highest concentration exists in gi!t houses where breeding continues. The virus spreads through the transfer of animals at the time of active infection. Piglets infected in utero are source of infection for about 9 weeks. S.pecific antibody to PPV has been reported in serum of cattle, sheep, cats, guinea pigs and rats. Serocon-version and faecal excretion of PPV Occurs when the rats are experimentally infected. Infected rats may possible be a source of transmission. The oral route is the usual route of virus transfer. Since the virus has been isolated from semen and vaginal mucous it is probable that veneral route may be important in transfer of virus. Immune tolerance has also been reported and these animals may be responsible for veneral transfer of virus.
Diagnosis: PPV induces high haemagglutination inhibition(HI) and serum neutralizing(SN) antibody thres in infected pigs. The HI test is conveniently applied in the diagnosis of infected herds. Active immunity is characterised by persistent HI titres in excess of 256 that continue for 4 years or even more. Litters infected in utero show HI titre of more than 256 at 10-12 weeks of age while piglets with passive immunity have a lower antibody titre. Dead foetuses of over 70 days have an antibody response to PPV. The heart blood and tissue extracts from mummified foetuses show HI titre. The foetuses less than 70 days show antigen in most tissues by fluorescent antibody or by HA technique. Isolation of virus can be made from tissues of aborted foetus (mesentric lymphnodes, ileum, liver, placenta) in actively growing porcine kidney cells. The observation time is upto 30 days. The autoculture of tissue such as kidney readily reveal evidence of PPV within 1-21 days.
Control: There is no vaccine available.
Feline Parvovirus: Feline Panleucopenia Virus (FPL V)
Feline leucopenia is an extremely contagious and fatal disease of cats characterized by high fever, anorexia, vomition, depression anrl leucopenia. The disease has a world wide distribution and is an important disease of cats in Europe and North America. The disease has also been reported in Panther cubs from this country. The virus was isolated for the first time in 1964 from the spleen of a leopard.

Parvoviridae 155
Properties of the virus: The virions are morphologically similar to other parvoviruses and measure 20-25 nm in diameter. The virus is resistant at 56°C for 30 minutes and to ether, chloroform and trypsin. All the strains of the virus are antigenically homogenous. By virus neutralization test it has been shown that this virus is closely related to mink enteritis virus and canine parvovirus. The virus haemagglutinates porc;ine red blood cells at 4°C.
Cultivation: The virus can be cultivated in rapidly dividing celIs of feline origin. The virus produces weak transitory CPE in feline kidney cell culture and produces type ~A' intranuclear inclusion bodies. The virus does not grow in fertile hen's egg or in laboratory animals. The virus also replicates in cell cultures of lion, tiger and mink tissues.
Pathogenecity: Feline panleucopenia is a contagious disease of young cats but cats of all ages are affected. The incubation period varies from 4-10 days and mortality rate in affected animals exceeds 60-70 percent and may even reach 100 percent. The clinical symptoms range widely from paracute to mild cases. In paracute cases death occurs in 24 hours. In acute cases lhe course of the disease is 5-7 days. The symptoms are high fever, anorexia, marked depression and dehydration. Vomition is common and diarrhoea may occur in 2-4 days. Before the temperature there is an initalleucocytosis followed by decrease of Jymphocytes and polymorphs. The leucopenia becomes pronounced with the progress of disease. The virus can pass the placenta and induce abortion. The small intestine is inflammed, thickened and oedematous. There is necrosis of intestinal mucosa, and mesenteric lymph nodes. The red marrow of long bones becomes semifluid in consistency and cellular elements are considerably decreased. The disease can be transmitted to minks. It is considered that feline panleucopenia, mink enteritis and feline ataxia are caused by the same virus.
Epidemiology: All secretions and excretions contain the virus. Recovered cats act as carriers and shed virus in urine and faeces for about 1 year. Under natural conditions the virus spreads by direct contact with infected animals or contaminated material. The natural portals of entry are respiratory and oral route. The fleas and other biting insects may act as mechanical carriers because the virus remains in the blood of affected animals.

156 Textbook of Veterinary Virology
Diagnosis: The provisional diagnosis can be arrived at the clinical signs, presence of leucopenia and presence of intranuclear inclusions in the intestinal epithelium. Virus can be isolated from spleen and other tissues in feline kidney cells. A rise of antibody titre in paired serum samples can be detected by HI test.
Control: Live attenuated and inactivated vaccines are available. The attenuated vaccines are derived from FPLV or mink enteritis virus. The immunity produced is superior to inactivated vaccine. Two doses of vaccine are given, the flrst dose at the age of 9-10 weeks and second dose 2-6 weeks later. The live vaccine is not recommended for kittens less than 4 weeks and for pregnant animals.
Canine Parvoviruses
Canine parvovirus-l also called as minute canine virus was identifled in the faeces of dogs in 1967 but is not associated' with any major cause of disease in dogs, although it may cause mild diarrhoea. Canine parvovirus-2 which is antigenicaIly distinct from canine parvovirus-l was rccognised in 1978 as a cause of haemorrhagic gastroenteritis simuItanasouly from USA, Australia and UK. Subsequently the disease has become enzootic in dogs through out the world. In India the disease was reported by Balu and Thangaraj in 198], in and around Madras city. The disease has also been reported from other parts of the coun try.
Canine parvovirus (CPV) is a small, non-envelop"ed, icosahedral virus measuring approximately 20 nm .in size. The icosahedral capsid encloses single stranded DNA genome. The virus is relatively stable. It is satable at pH 3-9 and at 56°C for 60 minutes as well as in the presence of lipid solvants. The virus agglutinates pig and rhesus monkey red blood cells at 4°C and 25°C and not at 37°C. The haemagglutination is inhibited by CPV antisera. The virus can be grown in actively dividing dog epithelial ceUs.
The epidemiology of disease is similar to feline panleukopenia. The virus is excreted in high concentrations in faeces. Three age related syndromes have been recognised. Generalised neonalal disease is rare. The panleukopenia but cerebraUar hypoplasia found in cats is absent. Myocarditis is recognised as acute disease in pups characterised by sudden death without any clinical signs. Apart from diarrhoea which can be haemorrhagic, vomition often occurs. In acute cases there is

Parvoviridae 157
sudden death while in less acute cases, respiratory problems occur and pulmonary oedema develops. The morbidity rate is very high upto 100% while mortality rate is upto 10%. The demonstration of virus by haemagglutina-tion and haemagglutination inhibition test of faeces and serum samples confirms the diagnosis.
Vaccination with either attenuated live virus or inactivated vaccines is effective. The problems are erlcoutered in devising effective vaccination schedules because of variable levels of antibodies transferred to pups from mothers.
References
ABINANTI, F.R. and W ARFIFLD, M.S., 1961. Recovery of haemadsorbing virus (HADEN) from the gastrointestinal tract of calves. Virology. 14, 288-289.
ACHERMANN, 0., 1982. Canine parvovirus infection and its prophylaxis. The Blue book Dec. 1982 pp. 1-9.
APPEL., M.J.G.; COOPER, B.J.; GREISEN, H.and CARMICHAEL, L.E., 1978. Status report. Canine Viral enteritis. J.Amer. Vet. Med. Assoc. 173: 1516-1518.
BUXTON, A. and FRASER, G., 1971. Animal Microbiology. Vo!.2. Oxford, Blackwell Scientific Publications.
HOGGAN, M.D., 1971. Small DNA viruses. In comparative Virology edited by K.K. Marmorosch and Kurstak. Academic Press, Inc. London
JEE, H.S. and JOIINSON, R.H., 1976. Porcine parvovirus: A review Veterinary Bulletin 46: 653-660.
KAHRS, R.F., 1985. Viral diseases of eaU le. Kalyani Publishers. Ludhiana.
KEu..Y, W.R., 1978. An enteric disease of dogs resembling feline pan leulwpenia. Aust. Vet. J54: 593-595.
MENGEUNG, W.L., 1975. Porcine parvovirus. In diseases of swine. Edited by H.W.Dunne and A.D.Leman. Ames, Iowa, Iowa State University, Press. 1975
MOHANTY, S.B. and DUTl'A, S.K., 1981. Veterinary Virology, Lea and Febiger, Philadelphia
O'SHBA, J.D. and STUDDERT, M.J., 1978. Growth of an autonomously replicating parvovirus (feline pauleulwpenia): Kinetics and morphogenesis. Archieve Virology 57: 107.

158 Textbook o/Veterinary Virology
STORZ, J., 1990. Bovine parvoviruses. In Virus infections of ruminanJs, edited by D. Zintcr and B.Morein. Elsevier Scinece Publishers B.V. Amsterdam.
STORz, J.; Leary, 1.1.; CARSON, J.H. and BATES, R.C., 1978. Parvoviruses asrociated with diarrhoea in calves. Journal American Veterinary Medical Association. 173: 624-627.
WOODS, G.T., 1975. Bovine parvovirus-l, Bovine syncytial virus and bovine respiratory syncytial virus and their infections. Advances in Veterinary Science and comparative Medicine.

Chapter 15
Papovaviridae
The family includes two Papilloma virus and Polymavirus. The name Papova was derived by connecting ftrst two letters from the names of three viruses-Papillomavirus. Polyomavirus and Vacuolating agent. Papilloma viruses are tumorigenic causing papilloma in different species of animals for centuries but the viral etiology was rccognised in 1907. A possible bovine polymavirus was demonstrated in cultured kidney cells of newborn calf. Polymavirus produces tumors in mice. The vacuolating agent is a simian virus also known as SV40. Papillomaviruses are nonenveloped. icosahedral measuring 50-55 nm in diameter. Polyoma-viruses are smaller measuring about 45 nm in diameter. The capsid is formed of 72 well discernible capsomeres in a skew arrangement The viral core contains a single molecule of infectious circular double stranded DNA with a molecular weight 3-5 x 1()6 daltons. The G+C content is 41 to 49%. They are ether stable and withstand heating for 30 minutes at 56 to 65°C. The papilloma viruses are species speciftc but different types may be associated with various forms of papillomatosis in a single animal species. The subdivision of papillomaviruses into six types is made by serology and by comparing restriction endonuclease cleavage patterns of their DNAs and DNADNA homolgy following hybridization. The replication occurs in the nucleus but viral DNA is not integrated into cellular DNA. For growth of papillomaviruses permissive cell type is lacking but established cell lines respond to infection or transfection with foci of transformation.
Papilloma viruses. which were once thought to be of minor importance in animals and man are now being recognised significant

160 Textbook o/Veterinary Virology
oncogenic factors in some neoplastic entities. Laryngeal papilloma and cancer, as well as carcinomas of the cervix and skin in man, skin cancer in goats, eye cancer in cattle and equine sarcoid have been associated with papillomavirus infection. An internal capsid antigen is common to all papillomaviruses. The bovine papillomavirus (BPY) 1 and 2 can serve as eukaryotic vectors in genetic engineering to insert foreign genetic material (DNA) into cells in culture.
The warts of skin are usually self limiting benign tumors, which occur usually in young aminals. The bovine papillomavirus 1 and 2 (BPY-l and BPY-2), the ovine papillomavirus, the deer fibroma virus and European elk papillomavirus produce fibroblastic tumor in hamsters. The BPY -1 and BPY -2 can also cause fibroblastic tumor in horses. The papillomavirus infects the basal cells of epithelium. Some of cells degenerate while others are stimulated to excessive growth and wart formation. The new virus particles are found in the degenerating cells and therefore there is much infective virus at the surface of the wart. The virus is resistant and contaminates the faeces and animal houses. The wounds on skins, tattoo instruments and hypodermic needles lead to infection in cattle.
Bovine Papillomavirus types 1 and 2
Bovine papillomavirus type 1 and 2 (BPY-l and 2) are world wide in distribution and cause fibroblastic tumors of the dermis in addition to epithelial hyperplasia in cattle less than 2 years of age that are housed in close contact with each other. The tumors commonly known as fibropapillomas are common on the skin of neck, legs, back and abdomen and are of various sizes. The tumors have a cauliflower like appearance and a fibroma base in the dermis. The fibropapiIlomas of the penis and vulva-vaginal mucosa have a smooth surface with less epithelial proliferation. The virus is concentrated in the outer keratinized epithelium and when shed contaminates the fences and animal houses. Experimentally growing fibroblastic tumors of brains in cattle and hamsters and polyploid tumors of urinary bladder can be produced in calves with BPY. In certain parts of world bovine haematuria is assoicated with bracken fern and BPY infection.
Yirus is transmitted between animals by contaminated halters, nose rings, grooming equipment, rubbing posts and other articles contaminated with diseased animals. The virus gains entry through

Papovaviridae 161
abrasions in the skin by direct contact with infected animals etc. The disease is more common in housed animals than in cattle on pastures.
The warts are characterisitic and laboratory diagnosis is seldom necessary. The virus particles can be demonstrated by electron microscopy.
The lesions usually disappear spontaneously. Formalinized suspensi.on of bovine warts provide a vaccine for prophylactic immunization when warts are a problem in a herd The efficacy of the vaccine is difficult to assess.
A situation has been reported in which fibropapillomas of the oesophagus and fumen harbour the genome of BPV -2 but neither the mature virus nor its antigens could be dctected. This suggests epithelial transformation to neoplasia without production of infectious virus.
Uovine Papillomavirus type-3
A typical warts in cattle which lack the dermal fibroma component and tend to persist rather than regress involve adult as well as young animals. The atypical papillomas contain a virus similar in morphology with BPV-I and 2 but immunologically distinct. The vaccine containing formalinized atypical warts does not control the natural incidence of the disease.
Bovine Papillomavirus type-4
On the west coast of Scotland with much growth of bracken fern, papillomas and carcinomas are found on the tongue, pharynx, oesophagus, rumen and intestine of cattle. The tumor extracts produce typical papillomas of the oesophagus, palate and skin. The causative agent has been characterized as BPV type 4. The BPV -4 is found in the papillomas of alimentary tract but alimentary carcinomas had no demonstrable virus or viral antigens. Similar carcinomas of the alimentary tract have been observed in enzootic haematuria areas of Brazil and Colombia.
Bovine Papillomavirus type-5
BPV -5 is the cause of papilomas on the teats of cows.
Bovine Papillomavirus type-6
BPV -6 has been characterized in teat epithelial papillomas. Virus particles resembling BPV have been found associated with
ocular lesions in Australian cattle.

162 Textbook o/Veterinary Virology
Fibropapillomas of muzzle and legs in sheep in England have been reported due to papillomavirus.
Canine PapilIomavirus
This type of wart usually begins on the lips and spreads to buccal mucosa, tongue, palate and pharynx of puppies. The warts resolve within few weeks and do not affect the dogs over 1 year of age. Formalinized wart suspensions may have a prophylactic but no curative value.
Rabbit Papilloma viruses
There are two distinct viruses causing papillomas in rabbits. The oral papilloma virus is similar to that of bovine or other papillomaviruses. The other virus or shope papillomavirus is found in cotton tail rabbits where virus particles are readily detected. Experimental transmission of shope virus to domestic rabbits results in formation of cutaneous tumors with maiked virus where virons are not detected but infectious viral DNA is present.
Equine Papillomavirus
Warts may be seen on the skin around lips and muzzle of young horses between 1-3 years of age. The lesions may perisist upto 18 months. The agent is readily tran~missible to horses. The sarcoids are also produced by the same virus. All ages of horses may be affected and lesions persist for about 8 months.
References
BUXTON, A. and FRASER, G., 1977. Animal Microbiology, Vol. 2. Oxford Blackwell Scientific Publications.
HuCK, R.A., 1969. Bovine papillomatosis: Synonymous warts. Infections verrucae, Veterinary Bulletin 35475-477.
JARRETI, W.F.H.; Mc NEIL, P.E.; LAIRD, H.M.; 0' NEIL, 1.; MURPHY, 1.; CAMPO, M.S. and MOAR, M.H., 1980. Papilloma viruses in benign and malignant tumors of cattle. Cold Spring Harbor Conference on cell Proliferation. 7: 215-222.
LANCASTER, W.D. and OLsON, C., 1982. Animal papillomaviruses. Microbiol. Rev.46, 191-207.
MELNICK, 1.L., 1962. Papovavirus groups. Science 135: 1128.
PFIsTER, H., LINZ, V.; GISSMANN, L.; HUCHllIAUSEN, B.; HOFFMAN, D. and

Papovaviridae 163
ZURHAUSEN, H., 1979. Partial characterisation of new type of bovine papilloma virus. Virology .96, 1-8.
MOHANlY, S.B. and DUTIA, S.K., 1981. Veterinary Virology. Lea and Febiger, Philadelphia.
OLsON, C., 1990. Papillomaviruses. In virus infections of ruminants, edited by D2inter and BMorein, Elsevier Science Publishers B.V.Amsterdam.
RUSSELL, P.H. and EOINGTON, N., 1985. Veterinary viruses. The Burlington Press (Camb). Ltd. Foxton, Cambirdge.

Chapter 16
Adenoviridae
Adenoviruses for the nrst time were isolated in 1953 by Rowe and his colleagues in uninoculated cultures of human adenoid tissue (adcn means glands) of children. Adenovirus have been isolated from man, monkeys, cattle, swine sheep, dogs, mice and birds.
The virion is nonenveloped with icosahedral symmetry, 60-90 nm in diameter. The capsid comprises of well discernable protein subunits (capsomcres) surrounding a core of DNA and internal arginine, rich proteins. The 252 capsomeres of the capsid are arranged into an icosahedron having 20 triangular facets and 12 vertex capsomercs. The 12 capsomeres at the vertices have 5 neighbours called pen tons. Each vertex capsomere carries 1-2 ftlamentous projections. The 240 nonvertex capsomeres have six neighbouring capsomeres and are called hexons. The adenoviral genome is a single linear molecule of double stranded DNA of mol. wt between 20 - 25 x 1()6 daltons. The G+C content is 48-61 %. The terminal nucleotide sequences of each strand are inverted repetitions. There are at"least 10 polypeptides in the virion with molecular weights ranging from 5K to 120K. After uncoating the viral DNA is transcribed into early proteins and late proteins. DNA replication occurs by strand displacement; transcription in the nucleus is followed by splicing into mRNA's. The mRNAs migrate into the cytoplasm, where the structural polypeptides are synthesized. The assembly of virus particles take place in the nucleus and release of matured virus particles take place by disintegration of the damaged cells. The host cells DNA, RNA and protein synthesis is effIciently

Adenoviridae 165
stopped by the infection. Some members of the family haemagglutinate rbe. The virion is resistant to ether and relatively stable below 50°C.
The family has 2 genera:- 1. Mastadenovirus-includes all mammalian' adenoviruses and share a common antigen which is not possessed by other genus 2. A viadenovirus-includes all adenoviruses of birds and these aviadenovirus do not haemagglutinate rbe of rats and other species of animals. The adenoviruses can be cultivated in cell cultures of natural host or· closely related species. The CPE is characterized by rounding or clumping of affected cells resembling 'bunches of grapes'. The CPE is produced late, 7-10 days longer. Since the virus multiplies in nucleus, the nuclear changes include fonnation of basophilic or acidophilic inclusion. The inclusions represent aggregation of proteinaceous material and crystal of mature and immature particles.
Bovine infections with adenoviruses are inapparent but in calves pneumonia, enteritis or pneumoenteritis, conjunctivitis or keratoconjunctivitis are produced either solitary or as membrers of miscellaneous microbial flora. In sheep adenoviruses produce enteric and respiratory disease. Among the dogs the diseases are infectious canine hepatitis and canine laryngotracheitis. In poultry the virus is associated with drop in egg production, inclusion body hepatitis and respiratory problems. Many' adenoviruses are capable of inducing malignant tum.ors in newborn hamsters. The oncogenic activity of adenoviruses is closely 'associated with base composition of nucleic acid. Highly oncogenic viruses have a low, 48-49 percent G+C content and mildly oncogenic types have 50-51 percent of G+C content while nononcogenic types have the highest G+C content, 55-60 percent.
Bovine Adenoviruses (BA V)
Properties of virus: The bovine adenoviruses known so far have been classified into nine serotypes. These serotypes have been divided into two subgroups based on the replication of viruses either in calf kidney or testicle cells. These two subgroups also differ in their antigenic properties. The new bovine isolates are constantly being reported and isolated from apparently healthy cattle as well as among those with clinical illness. Seroconversion or rising titres among healthy cattle have been observed.
Members of subgroup 1 readily replicate in bovine kidney cell

166 Textbook a/Veterinary Virology
cutures. They ar~ heat sensitive and have a complement fixing antigen common to human adenoviruses. The members of subgroup 2 are not easy to isolate and require few blind passages in calf testicular cells and do not hav.e a complement fixing antigen with human adenoviruses. The member of this subgroup invariably require calf testicular cell culture for their replication. Some bovine adenoviruses are highly cytopathogenic and some are less cytopathogenic. Most of the viruses produce intranuclear inclusions in cell culture. Typing of bovine adenoviruses is based first of all on cross neutralization tests. Only few serotypes show haemaggluti-nating activity. Rat erythrocytes are agglutinated by types 1, 2, 4 and 7. Bovine erythrocytes are agglutinated to low titre. The two subgroups are differentiate~ by complement fixation and immunodiffusion tests. It has recently been reported that certain ovine adenovirus isolates cross neutralised with BA V type 2 and infected both sheep and cattle under experimental conditions.
Bovine adenoviruses show remarkable stability outside their hosts. They are resistant to pH 2, pH 11, trypsin (0.25%), heating for 30 min. at 50°C., NaOH at 0.5-2% concentration inactivate the virus and are good disinfectants.
Cultivation: Subgroup 1 viruses replicate in wide range of cultured mammalian cell types, while subgroup 2 viruses grow in calf testicle cells. Single inclusion are seen in subgroup-l and mulLiple inclusions in the nucleus in subgroup-2
Epidemiology: The BA V is ubiquitous among cattle popUlation and it appears that cattle are potential reservoir. Serological evidence suggests a high incidence of infection with certain serotypes being replaced by others from time to time. The prevalence of antibodies is more in cattle which have suffered from respiratory disease. The transmission in calves is by direct contact Actually infected animals may shed the virus in nasal and conjunctival secretions, faeces and urine.
Palhogenesis: The BA V was originally considered to be non pathogenic for cattle but now it has been proved that the viruses are associated with several diseases of cattle. Observations on experimentally and naturally infected cattle indicate tJtat BA V is associated with diarrhoea, enteritis, pneumoenteritis, pneumonia, keratoconjunctivitis and weak calf syndrome. After infection there is

Adenoviridae 167
viraemia. The incubation period by experimental inoculation is 2-3 days but in natural cases it may be longer and possibly about 2 weeks. Infection occurs mostly in calves between 3 weeks to 4 months old. There are no gross or histopathological changes pathogenomonic for disease syndromes caused by BA V. Intranuclear inclusions are found in the organs after infection in the early stages.
Immune reaction: The adenoviruses are potent antigens. About 14 polypeptides are associated with the virion. High titre of antibodies appear after infection. Maternal antibodies provide passing protection against the homologous virus type.
Diagnosis: Isolation of virus from nasal and eye swabs and rectal swabs must be attempted. On post mortem, various organs, kidneys, testicles are homogenised and used for virus isolation. Fourfold increase in SN antibody titres between acute and convalescent serum samples confirms the diagnosis. Viral antigens can be demonstrated by CIEP and direct IF tests.
Control: The progress in immunoprophylaxis against BA V is slow. Bivalent adenovirus vaccines have been developed. The viruses are inactivated Wilh p-propiolactone or formaline and aluminium hydroxide and saponin used as vaccines. The vaccination is carried out in pregnant cow and also in the calves or in the calves alone. The cows are vaccinated twice in the last trimester of pregnancy and their calves are vaccinated at 6-8 weeks of age. Vaccination is repeated in calves 10-12 weeks old. Vaccination of calves alone is also practised in certain parts of Europe.
Ovine Adenoviruses (OA V)
Adenoviruses have been isolated from normal as well as from pneumoenteric diseases of sheep. The OA V are responsible as the causative agent of enteritis and respiratory infections in lambs.
Properties of the virus: The virus particles are resistant to heat The exposure at 56°C for 30 min. does not inactivate the virus although there is a fall in titre of the virus. Divalent cations significantly decrease heat resistance. All straIns are resistant to pH 3, ether and sodium deoxycholate. Sodium hypochlorite and formaline produce complete inactivation.
Sheep adenoviruses have been classified into six serotypes so far. All serotypes agglutinate rat erythrocytes. All the strains share a soluble

168 Textbook of Veterinary Virology
group specific antigen with human adenoviruses. The six serotypes could be distinguished by cross neutralization tests.
Cultivation: Ovine adenoviruses readily replicate in various ovine and other Jl?ammalian cell cultures such as lamb kidney, testis, lung cells, calf kidney and testis cells, pig kidney cells and in MDBK cell line. OA V -6 preferably multiplies in lamb testis cell cultures. The CPE appears 14-24 hours of post infection and comprises of increased refractibility rounding of cells followed by lysis.
Epidemiology: The ovine adenoviruses are largely confined to their host species. However, BA V -2 was observed among lambs causing pneumoenteritis. The presence of adenoviral antibodies in sheep populations have been reported from several countries all over the world including India. Adenoviral pneumoenteritis causes grtat losses in lambs due to mortality, increased food consumption and retarded growth. Virus is shed mainly with the nasal discharge, faeces and urine. Direct contact is most important in the spread of infection. Fattening lambs in units with continuous replacement are most likely to develop the disease because the passage of virus is uninterrupted resulting in permanent chain of infection and enhanced virulence. Crowding, insufficient ventilation, heat stress aggravates the disease outcome. The disease is frequently accompanied by other viruses and bacteria. Simultaneous infection with reovirus and/or parainfluenza-3 virus is common. Pasteurella haemolytica. corynebacteria and Mycoplasma ovipneumonia is also common.
Inapparent infection is common. Where clinical disease occurs morbidity may reach 100%. The disease occurs in 2-12 week old lambs and losses may be 30-40% in suckling lambs and 10-15% in fattening lambs. About 10% of losses occur in acute phase of disease while 90% in second phase when bacterial infections complicate the process.
Pathogenesis: The disease can be reproduced by intranasal and intratracheal inoculation of lambs. The virus replicates in the respiratory and intestinal tract, from 4 day post infection, pathological changes appear in other organs. Lesions are found in nasal mucous membranes. lungs, intestines, lymph nodes, spleen, kidneys and liver. The virus is excreted in the nasal discharge and faeces from 2-3 days of post infection onward. In naturally infected lambs the virus can be isolated in the acute phase of disease. Later lambs become carriers and virus can be isolated occasionally from animals with antibodies. The virus is intcrmitently excreted in faeces.

Adenoviridae 169
In nature the disease starts with dia.rrhoea after an incubation period of 7-8 days. The respiratory symptoms follow 2-3 days of diarrhoea. Sneezing, nasal discharge, conjunctivitis and lacrymation are common. Diarrhoea stops after about 7-8 days but respiratory symptoms persist, turning chronic in most cases.
Lesions of focal· interstitial pneumonia are common. The mucous membrane of small intestine is generally congested and covered with viscous exudate. The retropharyngeal, peribronchiolar and mesentirc lymph nodes are swollen. Greyish white foci are seen in the cortex of kidneys.
Diagnosis: Virus isolation is attempted from nasal secretions faeces and urine. The samples for virus isolation should be collected early in acute period of disease. Lamb kidney, thyroid and testicle cells are suitable for virus isolation. The CPE appears after one or two passages. Viral antigens can be detected in the organs and infected tissue cultures by direct immunofluorescence test. Serologic screening can be done by gel precipitation test, immunofluorescence test and ELISA. Neutralisation can be carried out against different serotypes of ovine as well as bovine adenoviruses.
Control: Inactivated, bivalent adenovirus vaccines are available in Europe. Two doses are given 10 or 42 days following fIrst dose. Regular vaccination has reduced the losses due to pneumoenteritis. Pregnant ewes are vaccinated twice with a 6 week interval. The second dose to be given not later than 3 weeks before lambing. The lambs born out of these sheep are actively vaccinated around 5 weeks of age and given a booster vaccination 10 days later.
Canine Adenoviruses (Infectious canine hepatitis virus (ICHV», Fox encephalitis virus
Infectious canine hepatitis is a febrile disease which mainly affects young dogs and foxes and cause centrilobular necrosis of liver. Experimentally the disease can be produced in guinea pigs, wolves, racoons etc. The ferrets are resistant. The disease is widespread in Europe, North America and other parts of world. Rubarth in 1947, fIrst recognised this virus as a <;linical entity in dogs. The disease was reported from India by Nair (1955) and Mukerjee and Mehrotra (1983).
Properties of the Virus: The virus has a morphology similar to other members of adenoviruses. The virus particle measures 55-80 nm.

170 Textbook o/Veterinary Virology
The virus is resistant to 0.5% phenol for several days and survives for 10-14 weeks at room temperature and for 6-9 months at 4"C. The virus is inactivated by 0.2 per cent formaline within 24 hrs. The virus agglutinates human '0' guinea pig and rat rbe. The virus shares a common complement fixing antigen with human adenoviruses.
Cultivation: The virus is readily propagated in primary dog kidney and testicular cells. The virus can also be propagated in cell cuitures of pig, ferret, racoon and guinea pig origin. The CPE is characterized by rounding, swelling of infected cells and formation of intranuclear inclusions.
Epidemiology: The virus is excreted in the secretions from the respiratory tract as well as in urine and faeces. The excretion in the urine continues for a long time, at least 200 days after the acute phase of illness. The virus is spread by direct contact of healthy animal with infected animal or indirectly from contaminated surroundings and clothings.
PalllOgenesis: The dogs of all ages are susceptible although young dogs are more susceptible. Experimentally, guinea pigs, wolves, racoons and coyotes are susceptible. Depending upon the virulence the incubation period varies from 2-10 days. In severe cases, the dogs suddenly die with acute abdominal pain, diarrhoea and vomiting in 12-14hrs. In less acute cases there is high fever, apathy; diarrhoea, vomiting, transient corneal opacity, enlargement of tonsils and submaxillary lymph nodes and leukopenia. The dog assumes tucked up position. The acute disease is usually fatal within a week but mild forms of the disease also occur.
In foxes, the disease is characterized by acute encephalitis with convulsions followed by paralysis, coma and death.
On postmortem the animal shows subcutaneous oedema, hacmorrhagic exudate in peritoneal cav~ty and intestinal tract. The liver is pale and enlarged. The gall bladder wall is oedematous and vcry thick. The spleen is enlarged and haemorrhagic. Microscopically, the liver cells show necrosis and dilation of sinusoids. The intranuclear inclusions are seen in the liver cells.
Diagnosis: The clinical picture and post mortem findings are helpful in reaching at the diagnosis. The presence of intranuclear inclusions in smears or tissue sections is suggestive of ICHV. Viral isolations can be made in primary kidney cultures from blood, tonsils,

Adenovir idae 171
conjunctival sac, urine and nasal secretion of affected animals. The paired serum samples collected from affected animals are titrated for the presence of rising antibodies by complement-fixing and haemagglutination-inhibition antibodies. The fluorescent antibody test is useful for detection of ICHV antigen in the tissues of affected dogs.
Control: Both live and inactivated vaccines are being used for immunization. The virus was modified by serial passages in dog kidney cultures and .subsequently in pig kidney ceh cultures. The modified virus alone or in combination with attenuated distemper virus is used as a vaccine. Canine immunoglobulins confer passive immunity in pups for about 3 weeks and can be used in exposed animals.
Infectious Canine Laryngotracheitis Virus
A virus isolated by Ditchfield and his associates in 1962 was reported from the cases of respiratory illness in dogs. The virus is antigenically related to ICHV but is distinct antigenically on haemagglutination inhibition and neutralisation tests. The virus is considered to be an antigenice variant of ICHV.
Equine Adenoviruses
Adenovirus infection among foals is worldwide. The viruses casue a disease of upper respiratory tract in foals below 3 months. The affected foals develop cough, dysponea, fever and conjunctivitis. The course of the disease is 10 to 56 days. The foals above 5 months of age develop mild signs.
Porcine Adenoviruses
The role of porcine adenoviruses is not clear. The virus has been isolated from pig intestinal tract, brain and its kidney cell cultures. Experimentally, the virus produces a clinical respiratory illness in pigs. Atleast 4 serotypes of porcine adenovirus have been reported.
Avian Adenoviruses
Avian adcnoviruses can be isolated with ease from both healthy birds and those suffering from a variety of disease syndromes. The. avian adenovirus infection in chickens was recognised in 1954 and appears to be prevalent throughout the world. The virus is prevalent in the poultry flocks in this country as well. The avian adenoviruses(AA V) have also been reported in turkeys and pheasants, quail, ducks and geese apart from the fowl.

172 Textbook o/Veterinary Virology
Properties of the virus: Avian adenoviruses have a structure in common with other members of the group. The virus measures 70-75 nm in diameter. The molecular weight of DNA is 30 x 1()6 daltons and G+C content is 54% which is slightly greater than G+C content of the highly oncogenic human viruses (48-49%) but less than the non oncogenic types (55-69%).
AA V show remarkable resistance to heat. Most of the strains resist 56°C for 3 hours. The stability to heat is greater when they are suspended in monovalent cations compared to divalent cations. The virus particles are also resistant to .ultraviolet irradiation. It is also resistant to ether, chloroform, sodium deoxycholate and trypsin. The virus is stable over a wide range of pH (pH 2-9). Some serotypes agglutinate rats, sheep, and chicken erythrocyt~ but this property is not consistent. The AA V do not share a common complement-fixing antigen with human adenoviruses or other mammalian viruses. On the basis of serum neutralization test the fowl adenoviruses have been typed into 12 serogroups.
Cultivation: Most of the AA V are readily propagated in chicken embryos and cause death and stunting of chicken embryos. Chicken kidney, chicken embryo liver, chicken embryo fibroblasts and duck kidney support the gowth and produce CPE. The virus causes the nucleus to enlarge and become filled with basophilic granules. Later, intranuclear inclusion bodies of compact basophilic granules are formed.
Epidemiology: The AA V are excreted in the faeces and to a lesser extent in naso-oral secretions. The excretion of the virus continues for weeks in the faeces. The virus being resistant to various physical chemical agents, it is likely that virus present in the litter reinfects the birds. Under natural conditions the virus is not highly contagious within a flock. However, if other respiratory viruses are present, the spread of virus within a flock is very quick. The major source of spread could be the egg transmission of the virus. The virus has been isolated by many workers from uninoculated chicken embryos. The AA V are also present in cell cultures produced from embryos or chicks. The wild birds can be infected with FA V (Fowl adenovirus) but it is unlikely that other avian species than domestic fowl plays a major role in the epidemiology of FA V, however they could be important in introducing virus into an uninfected flock kept for producing SPE eggs.

Adenoviridae 173
Pathogenesis: It is clearly known that avian adenoviruses have a widespread distribution throughout the internal organs of the host and mayor may not be associated with a mild disease. The pathogenecity of different strains varies considerably. The association of other microbial agents increase tne severity of the disease outcome. Following initial multiplication at the site of virus entry there is viraemia resulting in virus spread to virtually all organs. The blood brain barrier normally prevents entry of virus into the central nervous system. The main sites of virus replication appear to be in the trachea and caeca.
Fowl ade~oviruses (FA V) have been associated with inclusion body hepatitis, aplastic anaemia, haemorrhages, mild respiratory disease and decreased egg production. Some FA V are oncogenic to newborn hamsters.
Inclusion body hepatitis: Inclusion body hepatitis (IBH) was first recognised in USA in 1963. The disease mainly occurs in broilers and mortality rate varies from 2-10%. At post mortem the liver is swollen, light brown to yellow with haemorrhages, there is marked anaemia, icterus of skin, subcutaneous fat deposits, haemorrhages of various organs and pale inactive bone marrow. Eosinophilic inclusion bodies are present in hcpatocytes. Adenoviruses of different serotypes have been isolated from the cases of IBH. The chicks which suffer from infectious bursal disease become immunosuppressive and lead to IBH outbreaks.
Respiratory disease: There is an association between FA V and respiratory disease of chickens. The infection with other microorganisms might activate latent adenoviruses. Experimentally the disease can be reproduced in very young chicks with mild respiratory disease. The clinical signs are enhanced by simultaneous inoculation of Mycoplasma galliseplicum.
Effect on egg production: Infection of hens with FA V cause a 10% fall in egg production for 3 weeks. In Netherlands haemagglutinating adenovirus of duck origin was isolated in 1976 from a flock experiencing fall in egg production ranging from 30 to 85% and production of high percentage of soft shelled eggs. The dropped egg production started at 29-31 weeks of age which did not return to normal and eggs were smaller and of poor shell strength for a short period of time. The haemagglutinating virus resulted in less production of shell, soft shelled eggs and also drop in production after an incubation period

174 Textbook o/Veterinary Virology
of 7 days. The haemagglutinating virus known as EDS-76 virus or BC-14 virus has bren reported from other parts of world like USA, Australia. The virus has recently been reported from this country from Andhra Pradesh, Orissa and Punjab. The seroprevalence of EDS-76 virus infection in poultry flocks has been reported from different parts of this country.
Oncogenicity 0/ Jowl adenoviruses: The FA V produces tumors in newly born hamsters. The virus also transforms human and hamster cells in vitro. Other avian adenoviruses do not show oncogenicity.
The adenoviruses are also responsible to produce certain pathological conditions in other species of birds like quail bronchitis, haemorrhagic enteritis of turkeys, respiratory tract infections in pheasants etc.
Diagnosis: The clinical signs are not helpful to reach at diagnosis. Virus isolation is done in chicken embryo liver cells or chicken embryo kidney cell cultures. The identification can be done by virus neutralization or fluorescent antibody test. The serum antibodies can be detected by gel diffusion or by virus neutralization test.
Control: Immunization is not possible against FAV' because of several serotypes. Attempts to control EDS·76 virus causing drop in egg production is centred around the use of inactivated vaccine. The killed vaccines are available for control of egg drop syndrome.
References
BELAK, S. and PALFI, V., 1974. An adenovirus isolated from sheep and its relationship to bovine adenovirus. Arch. Ges. Virusforsch. 46: 336-369.
BELAK, S., 1990. Ovine adenoviruses. In virus infections of ruminants, edited by D.Zinter and B.Morein Elsevier Science Publishers B.V.Amsterdam.
BURKI, F., 1990. Bovine adenoviruses.ln virus infection of ruminants edited by D. Zinter and B. Morein Elsevier Science Publisher D. V. Amsterdam.
CABASSO, V.I, 1962. Infectious canine hepatitis virus. Annals N.Y. Academy of Sciences.IOI, 498-514.
CHATIY, M.S., 1985. Studies on egg drop syndrome virus (EDS076) in chicks in Andhra ·Pradesh. Ph.D. Thesis, Andhra Pradesh Agri. Univ. Hyderabad.

Adenoviridae 175
DnmlR, Z., MORElN, B., 1990. Virus infections of rwninants Elsevier Science Publishers, Amsterdam
DUBEY, S.C. and SHARMA, S.N., 1985. Ovine adenovirus pneumoenteritis in lambs in India. Indian Journal of Animal Sciences. 55: 878-879.
GREWAL,G.S.; SHARMA, S.N. and DEKA, B.C., 1981. Inclusion body hepatitis in broiler chickens. Indian 1. Poultry Science 16; 51-56.
McFERRAN, 1.B. and ADAIR, B.M., 1977. Avian adenoviruses-A review. Avian Pathology6: 189-217.
MOHANTY. F.e.; VERMA. K.C.; PRADHAN. H.K. and KUMAR, RAM, 1984. Egg drop syndrome (EDS-76) in India, Seroprevalence of EDS-76 virus infection in poultry flocks. Indian J.Poultry Science19: 15-18.
MOHANTY, S.B. and DUTI'A, S.K., 1981. Veterinary Virology, Lea and Febiger, Philadelphia.
MUKHERJEE, S.C. and MEHR01RA, M.L., 1983. Studies on infectuious canine hepatitis. Indian J.Vet Pathology. 7: 57-59.
NAIR, K.P.C., 1955. Infectious canine hepatitis in Madras State. Indian Veterinary Journa131: 243-244.
RUSSEL. P.H. and EDINGTON, N., 1985. Vaterinary viruses 1st Edition. The Burlington Pr~ss (Cambridge) Ltd.
SHARMA, KRJSHAJ'IA, SHARMA, S.N.; SAJ\filYAL, D.S. and BAXI, K.K., 1984. lsolation and characterization of some avian viruses from ovaries of domestic fowl. Indian 1. Animal Science 54: 977-979.
STAUBER, E.; BENS HAW, H.W.; BORO, C.; MATTSON, D. and FRAl"K, F.W.,1976. Isolation of a serogroup two adenovirus from calf with weak calf syndrome. Canadian I.Comparative Medicine40, 98-103.

Chapter 17
Herpesviridae
The family derives its name from Greek word herpein-to creep. The family is subdivided into 3 families. Alphaherpesvirinae, Betaherpesvirinae and Gammaherpesvirinae. There are about 20 herpesviruses which produce disease among vertebrates.
The mature virus particles are large, 180-200 nm in diameter when enveloped. The capsid is about 100 nm in diameter and consists of 162 capsomeres, 12 pentameric and 150 hexameric arranged in icosahedron form. The nucelocapsid is closely surrounded by another layer consisting of protein termed as integument which carries short projections. In the core a fibrillar protein spool is present onto which the DNA is wrapped. The genome is a linear double stranded DNA with a molecular weight between 80 and 150 x 1()6 daltons. The DNA has random nicks and gaps along the length. The guanine and cytosine (G+C) content varies from 45 to 74 percent in different viruses. The DNA of each herpes virus can be distinguished from ,other viruses of the family by characteristic pattern of cleavage with restriction enzymes. More than 25 structural polypeptides with molecular weights between 4K and 20K have been identified, some of which are glycosylated or phosphorylated. The envelope contains lipids. It is suggested that lipids present in the cell before infection are incorporated into the virus envelope. Polyamines have been detected in herpesviruses. Thymidine kinase in increased amounts is found in cells infected with the virus.
Virus DNA is transcribed in the nucleus, mRNA is translated in the cytoplasm. Virus DNA is also replicated in the nucleus and is spooled

Herpesviridae 177
into preformed immature nucleocapsids. The assembled virus particle leaves the nucleus by budding from the nuclear membrane. The virus particle is modified by inserted viral glycoproteins that forms the envelope. Eosinophilic intranuclear inclusion bodies are the remanants of virus factories. In case of cytomegalus virus infected cells, basophilic cytoplasmic inclusions are also found.
The viruses are heat labile and are inactivated at 50°C within 30 minutes. Most viruses do not show property of haemagglutination. Serologoical cross reactions have been reported in different members of this family on the basis of neutralization, complement fixation, precipitation and cross protection tests. The viruses grow in cell cultures derived from a wide range of animal species. Some ;)f the viruses grow on developing chicken embryo and produce pocks on the chorioallantoic membrane. Generally the lesions produced are proliferative type which later become necrotic. Many members of the family grow in the central nervous system and produce latent infections.
Bovine Herpes Virus-I (BHV-I)
Bovine herpesvirus-l (BHV-I) refers to all isolates which are serologically related to infectious boyine rhinotracheitis virus and infectious pustular vulvovaginitis virus (IBR/IPV). IBR is an acute contagious disease of cattle characterised by fever dysponea rhinitis and other inflammatory changes of upper respiratory tract inclUding abortion and still birth. The virus in found worldwide. IBR was described as a new respiratory tract disease and was isolated by Greig et al. (1958) and Kendrick et al. (1958). The virus is prevalent in USA, Canada, Britain, Germany, Australia, Newzealand and African continent. The existance of infection from India was reported by Mehrotra et al. (1976). The antibodies against this virus are present among the cattle population of many organised farms of this country.
Properties of the virus: The outline core in negatively stained preparations appears to be polygonal measuring about 130-180 nm in diameter. The capsid is made up of 162 capsomeres. The envelope consists of a double membrane which is similar to host cell membrane and is about 200 nm. The genome is double stranded DNA with a molecular weight of 54 x 1()6 daltons and G+C ratio ranging from 71-72%. The virus contain lipid in their envelope which renders it

178 Textbook of Veterinary Virology
susceptible to many disinfectants, especially solvents like ether, acetone, alcohol and chloroform. The virus is extremely susceptible to 0.5% NaOH, 0.01 % HgC~, 1% chlorinated lime, ] % phenol derivates and 1 % quaternary ammonium compounds. These chemicals inactivate the virus within seconds. Formaline (5%) inactivates the virus within one minute. The virus is relatively thermolabile and is inactivated within 21 minutes at 56°C. In the environment the virus survives for 30 days in winter, while in the buildings it survives for 6-13 days in winter and 5-9 days in spring. The virus is stable at temperatures below -65°C but less stable at -20°C if storage exceeds one year. The virus is slowly inactivated at 4°C and at 37°C it survives for about 10 days. It is important to note that virus survives in the semen when stored and can contaminate virus free semen when infected semen is stored in the same container. For virus survival in the environment, humidity is important. The virus is stable between pH 6-9.
A one way serological relationship has been reported between BHV -1 and pseudorabies virus. When the genomes of these viruses were compared, the homology was found to be approximately 8%. One way relationship also exists between BHV-l and goat herpes virus (BHV -6). By complement fixation and gel diffusion tests the virus is reported to show antigenic cross-reactivity with equine herpes virus-I. The BHV-l, Marek's disease virus and Burkitts lymphoma virus share a common antigen. Minor antigenic difference is possibly the reflection of immunological drift. All IBR and IPV isolates are basically similar, however, diverse clinical manifestation suggested strain differences. Recently differences in IBR and IPV virus strains have been shown by restriction endonuclease digestion of their DNA.
Cultivation: The virus multiplies in a wide variety of cell cultures but for isolation, the bovine tissues like bovine kidneys and testes cell cultures are most useful. The infected cells become rounded and refractile and form syncytia. Large intranuclear inclusion bodies are formed in the infected cells. The virus does not grow in embryonated eggs.
Epidemiology: The BHV -1 is' widely distributed among cattle population all over the world but the host range IS limited. Many wild species of animals have been found seropositive but clinical signs have only been obseved in cattle. Ferrets have been found susceptible in USA but not in Europe. The rabbits are being used as experimental

llerpesviridae 179
models. Experimentally neonatal hamsters and skunks are susceptible. Serologically goats and sheep have also been reported positive. The wild ruminants in Africa and in zoos appear to be virus reservoir. Various virus strains are harboured by most animals following infection and after a period of persistence reach a state of latency from which they can emerge from stress. The BHV -1 has recently been isolated from soft shelled ticks (Ornithodorus coriaceus) collected from deer bedding areas in Western USA. Cattle and deer graze together in this region. Both these species were seropositive. The ticks may harbour the virus for a long period of time or the virus may replicate in the ticks or mechanical transmission may take place.
Cattle of all ages are susceptible especially newborn calves without maternal antibodies. The virus is excreted through nasal, ocular secretions and placenta of aborted animals. The virus spreads by contact. The outbreaks usually take place whe;i the new animals excrete the virus but it is not certain whether these animals are responsible for perpetuation of virus. The BHV -1 is capable of establishing latent infection in recovered animals and these animals continue to excrete the virus for a ling time. The tissues in which the virus establishes latency and how the reactivation of virus takes place is not yet known. The virus spread also takes place through contaminated semen, natural service, the teaser bull or herdsman. The semen becomes infected with preputial washings. Antigenic drift is another factor which may contribute the virus to survive in nature but this aspect needs detailed study.
Pathogenes;s: The virus enters the body via the mucous membranes of the upper respiratory tract via mucous membranes of the genital tract, by way of conjunctival epithelium and may be through soft shelled ticks. The virus is transported to other tissues by infected leucocytes. The excretion of virus in an infected animal is usually 10-16 days but persistence for longer periods have been observed, especially in calves when passive immunity is on the verge of detection.
The virus may cause different types of clinical manifestation. Respiratory form: The natural infection varies in severity.
Following an incubation period of 4-6 days, the disease manifests with a sudden rise of temperature. The affected animals show anorexia, depression, nasal discharge, coughing, open mouth breathing and

180 Textbook of~'eterinary Virology
dysponea. The nasal mucosa is severely inflammed. New born calves may die due to necrosis of liver. The morbidity is high and varies from 30-39% while mortality rate does not exceed 3%. The course of illness varies from 7-14 days.
Genital form: The genital form results following introduction of virus to the mucosa of the genital tract by mating or through other external agents. After an incubation period of 48 hours the first pustule appears, after 2-7 days and then, further pustules develop. Pustules may from a yellowish membrane which detach leaving white necrotic material on the mucosa of vulva and vagina. This form of the disease predisposes to further bacterial infection and may involve the uterus. There is no abortion but there is infertility in the affected animals.
Abortion: Abortion is a sequel to respiratory form of disease. The virus presumably reaches the foetus through the infected leucocytes and the lymphatic channels of lymph nodes. Abortions usually occur between 4th and 7th month of gestation. Foetuses are invariably dead when expelled and the placenta may be temporarily retained. Abortions may also take place due to administration of live virus vaccines.
Conjunctivitis and keratoconjunctivitis: Conjunctivitis is usually bilateral and is a common clinical manifestation of typical IBR. There is excessive ocular discharge, the conjunctiva is hypeeramic. In secondary bacterial infection, keratitis and corneal ulceration occur. The BHV is associated with ocular carcinoma but definite experimental proof is lacking that it is the etiological agent.
Mastitis: The virus has also been isolated from cattle suffering from mastitis. Experimentally the virus may produce mastitis.
Encephalitic form: The virus has been isolated from encephalitis in calves. The animals show ataxia, depression followed by periods of excitement characterised by running and terminating into stumbling and falling. The animal develops clonic spasm of legs, neck and lumbar muscles, become blind, develop coma and die within 3-4 days of onset of symptoms. A few animals recover but become blind.
Disease of alimentary system: Diarrhoea may be a clinical sign and often fatal in young calves. The virus has been isolated from adult cattle with enteritis.
The concurrent outbreak of IBR/lPV have not been recorded frequently, however, an outbreak of IBR and IPV has been reported.

fJ e rpesviridae 181
The Virus has also been isolated from naturally occuring outbreak of respiratory disease.
Immune reaction: The antibodies appear following infection and persist for years. Higher titres appear after respiratory tract infection while lower titres appear after genital tract infection. No correlation exists between the levels of immune globulins, virus excretion and severity of clinical sings. When the antibody titres decrease the virus latency is terminated and infectious virus is shed. Immunoglobulin of all the three classes are present in the respiratory and genital tract discharges with higher IgG titres during oesterus.
The cell mediated immunity plays a major role in body defense against IBR/IPV infections.
The maternal antibodies persist upto 3-4 months. The antibodies in nasal secretions of calves are in lower concentrations and rarely persist beyond the third week. The calves are 'recommended to be vaccinated at 3 week of age.
Diagnosis: The onset of clinical signs and course of disease are of diagnostic value. The confirmatory diagnosis can be arrived at by isolation of virus. The choice of material for virus isolation depends on the form of infection. In live animals with respiratory from of IBR, nasal swabs made of gauze should be collected at an early stage of disease. In case of IPV, gauze swabs fixed to long forceps are placed into the vagina, left for at least one minute and rotated before removal. The material kept at 4°C should be transported to the diagnostic laboratory. Liver spleen, lymphnode and brain can also be collected for isolation of virus. The serum sample alongwith other material should also be collected for detection of antibodies, second sample of serum is collected after 2-3 weeks of the collection of first sample. For virus isolation primary bovine tissues are used. In bovine kidney cell culture the CPE is apparent after 24-36 hours and is characterised by rounding, increased granularity and characteristic bunch of grapes. Within 79-96 hours there is complete destruction of cell monolayer. Cowdry type A inclusion bodies are seen. The serum neutralization test can be used to characterize the virus. FAT is useful in detecting virus antigen in tissues.
Control: Numerous attenuated and inactivated vaccines are available all over the world. If the disease is confirmed to small number

182 TeXlbook of Veterinary Virology
of herds, strict control programme offers a best chance of elimination. In case higher percentage of herds are infected a prophylactic programme is the best solution.
The fi,rst attenuated live vaccine was employed by intramuscular route in late fIfties. The vaccine proved to be dangerous in pregnant animals because the vaccine strains were not properly attenuated. The emergency vaccination is recommended when the heard is stricken with the disease and diagnosis is made before all animals have contracted the field virus. The vaccines against BHV-l contain other components as well like parainfluenza-3 virus, bovine viral diarrhoea and adenovirus and sometimes Pasteurella or Haemophilus are also included. The vaccination schedule recommended is that first vaccination be given to calves between 3-4 months of age in case the calves had received antibodies in colostrum. Otherwise, the vaccination in calves may be given at any age. Second vaccination is recommended 4-6 weeks later followed by yearly vaccination. The vaccination prevents severe clinical signs but not infection with the possibility of excretion of virus following stress.
Inactivated vaccines are also used to control the disease. If the inactivated vaccine contain low antigen content, the immunity produced is of low grade, and if these animals pick up infection the virulent virus multiplies and is excreted in large amounts without showing clearcut signs of the disease. These animals become latent carriers and spread the disease.
Bovine Herpes virus-2 (Bovine Mammillitis virus)
It is an ulcerative infection of teats and udder of dairy cows. The virus was first isolated in 1957 from South Africa. The disease has been reported from different parts of Africa, Europe, USA and Australia.
Properties of the Virus: The virus particle is similar in morphology to other herpes viruses. It is sensitive to ether and chloroform. The virus is antigenically indistinguishable from the Allerton virus, the causative agent of lumpy skin disease in Africa but can be distinguished from infectious bovine rhinotracheitis and malignant catarrhal fever viruses. The virus has a serological relationship with herpes simplex virus-l and 2 of man.
Cultivation: The virus does not grow in the emrbyonated chicken eggs. It readily grows in bovine, sheep, pig and cat cells derived from

Ilerpesviridae 183
kidney, testes, thyroid and lymph nodes. The CPE includes large syncytia with basophilic or acidophilic Cowdry type A inclusions in the nucleus. The virus does not produce characteristic CPE on established cell cultures.
Epidemiology: The exact nature of transmission is not known but it has been hypothesized that heifers may harbour a latent infection. The virus within the herd is transmitted during milking by contaminated hands or milking machines. Transmission by flies and other insect vectors has also been suggested.
Pathogenecity: The incubation period is 5-10 days. The virus causes a vesicular skin disease which mainly affects the udders and teats of milking cows and heifers before calving. Variable size of vesicles develop on the teats, udder and perineum. A certain percentage of cows develop mastitis. Suckling calves may develop lesions on the muzzle. The disease is self limiting and occurs in autumn or early winter. The lesions develop as raised circumscribed Plaques and there is thic~ening of teat wall. There is severe local cutanous oedema and erythema of the affected skin. The vesicles sometimes rupture and form necrotic painful ulcers.
Diagnosis: The clinical picture of the disease is suggestive of the diagnosis. Th~ virus can be isolated in cell culture and further characterised. The exudate from the lesions can be used for virus isolation.
Malignant Catarrhal Fever Virus (MCFV)
It is an acute generalized disease of cattle and buffaloes, characterized by high fever, profuse nasal discharge, severe hyperaemia of respiratory and alimentary epithelia, extensive lymphoid hyperplasia, keratocon-junctivitis, encephalitis and rapid loss of condition. The disease is prevalent in most parts of the world. The incidence is generally low and sporadic although the incidence is higher in Africa and several explosive outbreak have been reported from USA and Canada. The disease was reported fIrst time by Parihar et al. (1975) from Punjab. Subsequently several outbreaks have been reported from the same area in Faridkot district of Punjab. Singh et al. (1979) have reported the epidemiological features of this disease in Punjab.
Properties of the virus: The virus is very fragile and rapidly destroyed by putrefaction and heat. The cell free virus is inactivated by

184 Textbook o/Veterinary Virology
lipid solvents and detergents. It is relatively stable in culture media containing serum. The infectivity is lost within 10-15 minutes at 56°C. At -70°C the virus maintains the infectivity titre for several months. The virus is highly cell associated. The highest titres of virus are found in buffy coat of blood, lymphnodes and other tissues of reticuloendothelial tissues. There is no evidence of significant immunological variation between the strains of the virus derived from cattle or wild animals although there are reports that virus strains are not antigenically homogenous.
Cultivation: In wildebeest cattle and buffaloes, the infectivity in tissues is cell associated. The cell free infectious virus is found in ocular and nasal secretions of wildebeest. The rich source of virus are Iymphnodes, thymus, spleen, bone marrow and buffy coat from peripheral blood. The most sensitive cell culture for virus isolation was considered to be primary or secondary monolayers prepared from calf thyroid where it produces syncytia, vacuolization, clumping of nuclei and formation of A type inclusion body in nucleus. Recently it has been found that calf ~estis cells are more satisfactory. The CPE is more easily detected and high yields of cell free virus is obtained. The virus does not grow in chicken embryos. The virus can be transmitted serially in young susceptible calves.
Epidemiology: The disease primarily affects cattle and buffaloes but is being increasingly encountered in farmed deer from which it is a major disease hazard. The most important experimental host is the domestic rabbit.
The majority of cases of MCF occur when there is a close contact between sheep and indicator host. When infected sheep are introduced in cattle sheds, the incubation period estimated from time of introduction of sheep is about 96 to 185 days. Cases of MCF continued to appear upto 3 or 4 months after removal of sheep from close contact with cattle. The reports of multiple cases of MCF have supported the thesis that sheep are probably reservoir hosts of the virus but there are also reports where investigators failed to incriminate sheep. There are possibilities that other species act as inapparent carriers. The wildebeest and wild ruminants also act as reservoirs. The virus crosses the placenta in some reserrvoirs through excretions, particularly nasal discharge ocular secretion and placental excretions. The disease generally is not transmitted by contact among cattle. Vectors like flies, ticks, and lice

Herpesviridae 185
are unlikely to play any role in transmission of MCF. The details of virus transmission are not well understood.
The majority of cases of MCF occur in young adults but animals of all ages, sex and breed are susceptible.
Pathogenesis: The portal of entry of MCF virus is probably upper respiratory tract, nasal mucosa and/or tonsil. Aerosal or intratracheal route of infection in cattle and rabbits is successful. The congenital transmission takes place frequently in reservoir host but has not been reported in cattle. The congenital transmission has been demonstrated only once in cattle.
A long incubation period is characteristic of this disease. It is about 19.5 + 3.7 days experimentally. In practice it is essential to wait for about 2 months before concluding that animal has not been infected by a parenteral inoculation. In cattle viraemia is detectable from 9-17 days after inoculation of virulent blood. The viraemia is assoicated with cellular fractions of the blood and not with the plasma, erythrocytes or platelets. The disease MCF is being considered as immunopathological condition. The basis for this are-{a) The lesions are not attributable to the virus as shown by absence of virus or antigens; (b) The long incubation period and prepatent viraemia suggest a requirement for sensitization to virus or virus associated antigens; (c) The lesions are similar to virus induced autoimmune or lymphoproliferative conditions and (d) Association of lymphocytic infiltration with vascular and epithelial lesions. The MCF pathogenesis appears to be immune deregulation attributable to virus infection causing dysfunction of natural killer cells and uncontrolled proliferation of lymphoblastoid elements in many tissues. Some workers have suggested that lesions of MCF may be due to a type III or Arthus like hypersensitivity reaction or a combination of type III and type IV reactions.
The disease is characterized by high fever, severe inflammatory and degenerative lesions of upper respiratory tract, entire digestive tract and eye. There is severe inflammation of oral and nasal mucosa with diffuse necrosis. The lesions are found on the lips, hard and soft palate and cheeks. There is inflammation of eye, lacrimation, eyelids become swollen and corneal opacity develops. There is enlargement of lymph nodes and they become haemorrhagic. The meningites of central nervous system are oedematous and encephalomyelitis may be present. The disease often terminates in fatal encephalitis in 5-12 days after

186 Textbook of Veterinary Virology
onset of pyrexia. The brain, myocardium, lungs show severe congestion. In some cases there is abomastitis and enteritis. The morbidity rate is very low.
The .leS~ons in the alimentary tract are epithelial necrosis and erosions followed by· hlceration in the pharynx, on the soft palate, oesophagus, on the folds of reticulum and pillars of rumen. In abomasum congestion and haemorrhagic erosions of fundic folds and pyloric region are common. The intestines (Calcum, and upper colon) exihibit congestion. There is congestion, diptheritic membranes, erosions, haemorrhages on the turbinates and septum and sometimes in the trachea. The lymphnodes show enlargement. The spleen is enlarged. The mucosa of urinary bladder and vagina show congestion, necrosis and erosions while the kidneys are swollen. There is. destruction of small lymphocytes in the lymphopoietic tissues, proliferation and infiltration in many organs with large lymphoblastoid cells and angiitis affecting components of the walls of arteries and veins.
Immune reaction: The few caule that survive infection arc resistant to challenge for several years, develop neutralizing antibodies in their serum. The neutralizing antibodies appear in the late stage of disease. There is evidence that the neutralizing antibodies do not avert the fatal outcome of the disease. The role of CMI in MCF is not known properly but the failure of neutralizing qntibodies to protect indicates that CMI plays an important role in immue response. Delayed hypersensitivity has been reported in MCP.
Diagnosis: The diagnosis is based on clinical signs and pathogenic changes. Virological diagnosis is well established and based on the recovery of virus from sick or killed animals and on the demonstration of antibodies in the scrum of animals collected at late stage of disease or during convalescence in the very few surviving animals. The virus is isolated from buffy coat of infected blood in cell culture. The CPE is neutralized by specific antiserum.
COlltrol: All attempts to immunize cattle with attenuated virus have failed. The inactivated virus preparations combined with adjuvants produce high level of neutralizing antibodies in cattle but there is resistance to parenteral challenge by cell free or cell associated virus. In rabbits, the inactivated vaccines provide protection against cell free virus given intravenously but not against virulent spleen

f/crpesviridae 187
suspensions. Few workers have claimed to have isolated strains safe and attenuated for cattle, which confer immunity to challenge but have to be confirmed. In the absence of vaccines, the prevention of MCF depends on the prevention of close contacts between susceptible and reservoir hosts. This means avoidance of housing or herding together of sheep and cattle or deer. In the zoological gardens, wildebeest should be separated from many susceptible species.
The Movar type of Bovine Herpesviruses (DHV-3)
A new serotype of bovine herpesvirus nonpathogenic to calves was isolated in 1967. Subsequently agents similar to this virus were isolated from USA, Africa and Europe. The pathogenic role of these viruses is minor or insignificant. These viruses have been named as BHV-3. Cattle of all ages and breeds are susceptible. The morbidity rate is 20-30% while mortality is very low. Transmission is by direct contact. The virus is assoicated with bovine respiratory disease complex. The disease may be inapparent, mild or acute. The virus induces mild clinical signs of tracheitis in young calves. In assoication with Pasteurella multocida the disease is quite severe. The acute disease is characterised by fever. dyspnea, COUgh, hyperpnea and copious nasal discharge.
Herpesvirus of Sheep (CHV -1)
The virus is associated with plllmoRary adenomatosis of sheep. The disease has a long incubation period of 6-9 months and the disease is characterized by chronic progressive pneumonia with the development of adenomatous in growths of the alveolar walls. The role of CHV -1 virus in pulmonary adenomatosis is not clear as the agent of this disease Uaagsiekte) has now been found to be a retrovirus.
Herpesvirus of Goats (CHV-2)
Herpes viruses have been isolated from kids with severe generalized infection of genital lesions in goats, acute and chronic pneumonia, wartlike lesions of eyelids and proliferative lesions around mouth and hard palate and dermal lesions. Experimentally the virus can cause abortions. The restriction endonuclease DNA fragment pattern is quite different from that of BHV-l virus but the DNA of CHV-2 shared a high degree of base s('.quence homology. The common feature of the

188 Textbook o/Veterinary Virology
herpesvirus about latency and reactivation has been shown by using corticosteriod treatment in goats.
Equine Herpesviruses
Three antigenically different herpes viruses are known to infect horses. Equine herpes virus-l (EHV-l) is the most important and causal agent of an important disease of horses known as equine rhinopneumonitis. The equine herpes virus-2 (EHV -2) mayor may not be associated with any disease while equine herpes virus-3 (EHC-3) is the cause of coital exanthema. Morphologically these viruses are similar to one another and exhibit some genetic homology but they differ in immunological properties.
Equine Herpes Virus-l (Equine Rhinopneumonitis Virus)
The virus produces highly infectious disease of horses characterized by fever, respiratory catarrh and abortion in mares. In nature, only equines are affected. The disease has been reported from USA, Germany and many other countries of Europe, Japan and India. In 1965 Sharma and his associates reported the disease on the basis of histopathological examination. The isolation was reported in 1976 by Jain and others from Hissar. The EHV-l infection is reported to be endemic in various equine farms in this country.
Properties of the Virus: Morphologically the virus resembles the other members of the family and possess DNA containing core enclosed in icosahedral capsid and is further enclosed in an envelope. The virus particles are about 150-198 nm. The virus does not survive for long period outside the host but can be stored at -18°C for a year. The virus is inactivated by heating at 56°C for 10 minutes. The virus haemagglutinates equine red blood cells. Earlier EHV-l strains were regarded antigenically similar but strains isolated from abortion cases are said to be more infective, grow better in cell culture and are released to greater extent in comparison to the strains isolated from respiratory infections. Recent findings revealed by restriction endonuclease analysis of the genomes of the isolates suggest that strains from abortion cases are different from those isolated from respiratory cases.
Cultivation: The virus does not grow directly in the chicken embryos. The virus can be adapted to grow on the CAM of chicken

Herpesviridae 189
embryos by alternate serial passages in hamster kidney cell cultures and CAM of chicken embryo. The virus multiplies in human amnion, HeLa and horse, sheep, pig, cattle, cat and chicken embryonic tissues. The CPE produced is rounding and ballooning of infected cells, multinucleated syncytia and acidophilic 'A' type intranculear inclusions. Plaque formation occurs on cells of several species. Unweaned hamsters and unweaned mice can also be infected. The virus produces a fatal hepatitis. In pregnant hamsters the virus may produce upper respiratory tract infection and abortion.
Epidemiology: The introduction of an infected horse may cause abortion. Under natural conditions virus may be maintained- in the upper respiratory tract of carrier animals. There is a possibility that virus may give rise to latent type of infection and spread by veneral contact as in the case of infectious bovine rhinotracheitis virus. Recent evidence has shown that antigenic difference between EHV -1 strains exist. This may be another important factor in the epidemiology of the disease.
Pathogenicity: The incubation period in natural infection varies from 9-60 days. Acute respiratory disease occurs mainly in foals, weanlings and yearlings. The disease is rarely fatal unless characterised by profuse serous nasal ~ischarge which later becomes mucopurulent. Distinct herpetic lesions develop in the respiratory tract characterised by necrosis of respiratory epithelium and intranuclear inclusions. Pregnant mares may abort mostly at 6-11 months of gestation after 14-120 days of exposure to virus. The natural route of infection is presumed to be via the upper respiratory tract followed by cell associated viraemia. The lesions in early abortions are severe autolysis of foetus while in late abortions there is jaundice, petechiation of mucous membranes, excessive pleural fluid, pulmonary oedema and spleenic enlargement The lesions observed in mares are distention of regionallymphatics.
The neurologic from of disease involves the central nervous system and is assoicated with enzootics of respiratory disease or abortion. The clinical sings are ataxia. weakness and paralysis.
The Tatent infection in EHV -1 has not been demonstrated so far but there is indirect evidence that some horses must be carriers and that latent infection can be reactivated.

190 Textbook o/Veterinary Virology
Diagnosis: The presumptive diagnosis is based on the clinical symptom~ and history of abortion. The presence of intranculear inclusions in the exfoliated epithelial cells of the respiratory tract and in the histopathological sections of lungs, liver, spleen thymus, myocardium, kidney of aborted foetus is of value in arriving at diagnosis. Antibodies in paired sera samples can be detected by virus neutralisation, counter immunoelectrophoresis, indirect fluorescent test on the cryosections of frozen lung and live tissues of aborted foetuses.
Control: The immunity resulting from natural infection in foals suffering from respiratory infection is of short duration, while the immunity against abortion in mares is more durable. Live and inactivated vaccines are available in foreign countries. Live, virulent, hamster adapted vaccine given intransally in horses is a good immunogen but may induce abortion in pregnant animals. Attenuated tissue culture vaccines are available. Two vaccinations are administered for primary immunization and then annual administration of vaccine at 2 months pregnacy and again at 6 months of pregnacy. These vaccines produce satisfactory immunity and are safe for pregnant mares. Inactivated vaccine mixed with oil adjuvant have been developed. These vaccines produce satisfactory immunity and are safe for pregnant animals ..
Equine Herpesvirus-3 (Equine Coital Exanthema)
Equine coital exanthema is an acute, usually rare disease characterised by formation of pustular and ulcerative lesions on the vaginal and vestibular mucosa, on the skin of penis, prepuce and perineal region and occasionally on teats, lips and respiratory mucosa. The causative agent is equine herpesvirus-3. The virus shows no serological cross-reactivity with other equine herpesvirues but shares antigens with equine herpesvirus-I. The virus grows in cells of equine origin and produces large plaques, the virus remains cell associated.
In coital exanthema the genital lesions are extensive but there are no systemic signs. The incubation period is short about 2 days and in uncomplicated cases healing is usually complete in 14 _days. Abortion and infertility is not associated with EHV-3. Affected stallions show decreased libido. The virus can also cause subclinical respiratory infection in yearling horses. The presence of disease at the stud farm disrupts the breeding schedule. The control measures consist of removal of stallions from service and symptomatic treatment.

lIerpesviridae 191
Pseudorabies Virus (Porcine herpes virus-I)
Pseudorabies or Aujeszky's disease or mad itch or infectious bulbar paralysis is primarily a disease of swine but occurs naturally in a wide range of animals including swine, cattle, sheep, dogs, cats, minks, ferrets, foxes and rats. In all animals excepting adult swine the disease is characterised by pururitis, paralysis and fatal termination. Mild illness also occurs in man. The disease is enzootic in many countries of Europe like Bulgaria, Czechoslovakia, Denmark, France, Hungary, Italy, the Netherlands, Poland. The disease is also prevalent in U.K and U.S.A. There are reports of disease outbreaks from China, Algeria, Tunisia and Angola.
Properties of the virus: Morphologically the virus is indistinguishable from that of other viruses of herpes group. It is more thermostable and resistant to pH changes than other herpes viruses. It may survive for 2-7 weeks in infected premises and upto 5 weeks in meat. It surives only for few hours on material contaminated with faeces and urine. It is sensitive to ether and chloroform and is rapidly inactivated by 0.5-1 % NaOH. The recommended disinfectants are 5% phenol, sodium and calcium hypochlorites, 2% formalin, 2% sodium hydroxide, trisodium phosphate and quartemary ammonia. There is no report regarding its property of haemagglutination. All the strains of the virus are reported to be antigenically similar by restriction endonuclease technology, the DNA of different strains was found to be slightly different. The virus is antigenically related to B virus.
Cultivation: The virus grows readily on CAM of embryonated eggs and produce pocks after 3-4 days of infection. Many strains are lethal to embryo. The virus grows in a" wide variety of cells derived from swine, caule, rabbits, dogs, monkeys and chicken embryos. Rabbit kidney is highly susceptible. Cowdry type A intranuclear inclusions and polykaryocytes are seen in infected cells.
Epidemiology: Naturally fatal encephalomyelitis is produced in cattle, sheep, goats, dogs, cats, deer, rabbits, rats and mice. The morbidity rate is high in pigs but pigs in first few weeks of life succumb to pseudorabies but in older pigs the mortality is negligjble. The pig is the principal reservoir and almost all cases of disease in cattle are due to cohabitation with swine. Latent infection occurs in swine and inapparently infected pigs are source of infectious virus.

192 Textbook o/Veterinary Virology
Cattle are considered dead-end-hosts of the virus and there is no evidence of interbovine transmission. The cattle and other ruminants rarely transmit the disease to other animals unless their tissues are eaten by susceptible carnivores. The rats and mice are poor shedders of the virus and do not play a role in the spread of the virus. The carnivores may serve as a source of infection to other species after they have become infected by ingesting infected meat.
Pathogenesis: The virus naturally enters into domestic ruminants via break in the skin caused by bite or other wounds and occasionally by aerosal infection. Experimenta~ly cattle can be infected by any route. In prodromal phase the swine shed the virus in saliva and transmit to cattle by muzzling or biting the flank, vulva or anal region while wandering with them. Peroral infection in cattle may occur via contaminated feed or water. Respiratory transmission ,occurs in poorly ventilated areas from swine to cattle. The virus enters the central nervous system via peripheral nerves causing neuronal damage resulting in local pururitis and encephalomyelitis. The virus has been isolated from nasopharynx, lungs, and vagina of affected call1e. Persistant latent infections capable of reactivation are common in adult swine but have not been reported from cattle.
The clinical signs in affected cattle are encephalomyelitis characterized by intense pruritis leading to self mutilation. The incubation period in natural cases is 4-7 days. The clinical signs are short which last about 10-24 hours. There is a general lack of typical and constant gross lesions. The site of pruritis is generally lacerated. There is accumulation of cerebrospinal fluid and meningeal congestion but lesions in the viscera are not significant The lungs may show congestion, oedema and haemorrhages.
Diagnosis: Typical cases of disease in callle and sheep are easily detected, however, diagnosis in pigs is difficult. Subcutaneous inoculation of specimens from clinical cases (brains, lungs, tonsils and kidney) into rabbits usually produce intense itching and death in 3-5 days. Isolation of virus in cell culture is a fruitiful method. Fluorescent antibody technique can also be adopted to brain tissue of the affected animals. In pigs serologic tests like virus neutralisation are useful to detect the presence of antibodies.
Control: Swine recovered from the disease develop neutralizing antibodies which are transferred to new born piglets through colostrum.

J J erpesviridae 193
Maternal antibodies protect the pigs for about 2 months. Attenuated and inactivated vaccines have been used in pigs with promising results. Experimentally attenuated and inactivated vaccines made for pigs have been used in cattle. The cattle vaccinated with inactivated vaccine develop marginal serologic response but remain resistant to challenge. The disease in cattle and sheep can be prevented by keeping pigs out of contact from these animals. Eradication by slaugher and quarantine of infected pigs after the incidence of disease in swine is reduced to manageable level.
Simian Herpes Virus-! (B virus)
The virus is naturally found in normal Asiatic monkeys especially rhesus and cynomologus species. The virus causes a fatal ascending myelitis of man.
Properties: The virus measures 100 nm in diameter and is morphologically similar to herpes simplex virus. The B virus antiserum neutralises human herpes simplex virus but not vice versa.
Cultivation: The virus grows in the chicken embryos and produce pocks on the choroallantoic membrane. The virus also replicate in cell cultures of rabbit, monkey and human cells.
Epidemiology: The virus spreads by direct contact in the monkeys especially when the animals are crowded together in captivity. The virus sets up a latent infection as is the case with herpes simplex virus in man. It has been shown that about 10 percent of newly caught rhesus monkeys have the antibodies against B virus and this percentage goes upto 60 percent when the animals are crowded in captivity. The saliva and central nervous tissue of clinically healthy animal contain the virus. The virus may also be recovered from the primary monkey kidney cell cultures.
Pathogenesis: The naturally infected monkeys may show herpes like lesions on the dorsal surface of tongue and the lips. The lesions in the monkeys develop commonly when the animals are crowded. Man usually gets infected through the bite of a monkey but one laboratory worker became infected after handling glassware used for tissue culture of monkey kidney. The incubation period in man is 10-20 days. At the site of bite there is local inflammation and the virus reaches the central nervous system by way of peripheral nerves and cause an acute encephalomyelitis and death. There is focal necrosis of lymphnode, adrenals, spleen and liver.

194 Textbook a/Veterinary Virology
Diagnosis: The diagnosis can be arrived at by isolating the virus from saliva and brain of monkeys or brain and spinal cord of infected human beings.
Control: There is no specific treatment for human beings. The vaccinatio" of man with formalised vaccines has been unsuccessful. Hyperimmune B virus antiserum be given after the bite and should also be inoculated at the site of the bite.
Canine Herpes Virus (Canine tracheobronchitis virus)
This is a newly recognised virus of puppies causing an acute fatal generalised viral infection 'of neonatal and infant puppies. The virus was reported in 1865 from United states but now it has been reported from Canada and Europe.
Properties of the virus: The virus has a similar morphology like other herpes viruses. The virus particles are ether sensitive and acid labile. The virus is not antigenically related with other members of the group but a low degree of cross reaction has been reported with herpes simplex virus.
Cultivation: The virus readily grows in dog cells, like dog kidney, lung and uterine cells. The CPE arc produced as early as 16 hours after infection.
Epidemiology: The transmission of virus is not airborne. Infected dogs excrete virus in saliva, nasal secretions and urine. Infection can occur transplacentally or by contact.
Pathogenecity: The disease causes anorexia, laboured breathing and abdominal pain. The mortality may reach upto 80% in puppies upto 1 week of age. In dogs of few weeks age the disease is mild or inapparent. Focal areas of necrosis and haemorrhages are found in the liver, kidney and lungs. The incubation period is 3-8 days.
Diagnosis: The virus can be isolated in dog kidney cells and identified by neutralization test.
Control: No vaccines arc available.
Fowl Herpesvirus-1 (Avian Infectious Laryngotracheitis virus)
Infectious laryngotracheitis (ILT) is highly contagious respiratory disease of poultry characterised by respiratory disease, gasping, expectoration of blood stained exudate and high mortality. All breeds and ages of birds are affected. The disease was reported for the first

Herpesviridae 195
time from USA in 1925. It has now been reported from several countries of the world. The disease was reported by Singh et al. (1964) from this country. The infection is widely prevalent in this country where chickens are raised. The disease mainly occurs in a mild or subclinical form, however, there are isolated reports of acute form as well.
Properties of the virus: The physical, chemical and biological properties of the virus are similar to other members of this group. The double stranded genome has G+C content of 45 to 50%. The viruS is one of the resistant poUltry viruses. It is readily destroyed by 3% cresol and i % NaOH. It is ether sensitive and is readily destroyed by exposure to 55°C. The virus does not have the property of haemagglutination. The virus appears to be antigenically homogenous although some variation in serum neutralization has been reported with certain strains. Variation in pathogenicity of different strains has been reported. Some strains are highly pathogenic while others are non pathogenic or mildly pathogenic.
Cultivation: The virus grows in embryonating chicken eggs and produces opaque pocks with central areas of necrosis. The embryos are stunted and death occurs in 2-12 days after inoculation. The virus also grows in chicken embryo cell cultures, chicken embryo kidney, lung and respiratory epithelium and produces CPE characterized by syncytial formation and cowdry type A inclusion bodies.
Epidemiology: Infectious laryngotracheitis virus is usually introduced into a flock by carrier bird and is transmitted by aerosal and inhalation, less commonly by ingestion. The virus is not transmitted through egg and insect vectors do not play any role. The carrier state in the recovered birds exist and the recovered birds may harbour the virus for more than 2 years. Outbreaks following contact between vaccinated and nonvaccinated brids have been reported.
Pathogenesis: The chickens of all ages are susceptible but the disease is most common in birds of 4-18 months of age. After an incubatin period of 2-8 days there is mild coughing and sneezing which is followed by nasal and ocular" discharge, dyspnea, gasping and coughing and depression. There is haemorrhagic tracheitis and the bird extends its head and takes a prolonged inspiration. The coughing may be associated with expectronation of bloody mucus and frank blood. Morbidity is about 100%. The mortality with virulent strains may be

196 Textbook of Veterinary Virology
50-70% and with strains of low virulence about 20%. The strains of low virulence are associated with conjunctivitis, ocular discharge, swollen infraorbital and nasal sinuses and lowered egg production.
The principal lesions are found in the larynx and trachea and which include haemarrhagic inflammation and oedema. In advanced cases, caseous exudate and diptheritic membrane may be seen in these organs. The inflammation in the bronchi, lungs and air sacs may also take place. Microscopically there may be loss of cilia, cellular infllteration and necrotizing tracheitis. Type A intranuclear inclusions may be present in early stages of disease.
Diagnosis: In acute form of disease, characterisitc repiratory symptoms and high mortality are often diagnosed. The mild cases are difficult to diagnose. Demonstration of type A intranuclear inclusions in tracheal or conjunctival tissue is si.gnificantly diagnostic. The most common method of diagnosis is by virus isolation from clinical material (tracheal exudate, lung suspension) in chicken embryos. Typical pocks, are produced on the CAM after 4-5 days of inoculation. The virus can be characterized by serum neutralization and agar gel diffusion test The fluorescent antibody test can be applied for demonstration of antigen in the tissues.
Control: Recovered birds are reportedly immune for 1 year. The vaccination is recommended in enzootic areas since vaccination results in a carrier state. Both virulent and avirulent vaccines have been used. The rust approach of vaccination was adopted in 1930, the young susceptible birds used to be mixed up with known recovered birds before laying stage. This was followed by inoculation of wild virus in the cloaca. It produced immunity without serious respiratory signs. In 1950s it was known that strains of low virulence occured and these viruses were then introduced for cloacal vaccination. When more attenuated strains grown in cell culture became available, other routes of vaccination like inoculation by infraorbital sinus or by intranasal and eye drop procedures were followed. Mass vaccination by aerosal and drinking water administration have also been developed. Immunization with attenuated vaccines protect birds against clinical disease but does not protect against infection with virulent virus or development of latent carrier status for either the virulent or vaccine viruses.
The eradication of the qisease is achieved by complete depopulation and disinfection of infected premises.

l/erpesviridae
Duck Herpesvirus-l (DHV-l) (Anatid Herpesvirus)
(Duck Plaguevirus, Duck virus enteritis)
197
It is an acute fatal contagious disease of ducks, geese and swans, and is of economic importance of duck producing areas. The disease was reported by Baudet in 1923 from Netherlands. Now it is recognised as a major disease .of ducks throughout Europe, North America, China and India. The disease from this country was reported in 1964 by Mukerji et al fr:om West Bengal. The disease outbreaks have been reported from other areas of this country as well. In addition to domestic ducks, wild ducks, geese, swans are other water birds that are equally susceptible.
Properties of the virus: The virus is similar in morphology and other physical and chemical characters with herpes viruses. The virus is ether and chloroform sensitive and does not haemagglutinate duck, chicken, sheep red blood cells. All the strains of the virus are antigenically homogenous.
Cultivation: The virus grows on the chorioaIlantoic membrane of duck embryos and kills the embryos within 4 days with extensive haemorrhages. The virus can also be adopted to grow in chicken embryos and chicken embryo fibroblast cells. The adapted virus becomes attenuated for ducks.
Epidemiology: Natural infection occurs in domestic and wild ducks, geese, swans and other water fowl. Migratory water fowl may contribute to spread within and between continents. The virus is transmitted by direct or indirect contact through infected water. Transmission does not occur through eggs but arthropods have been suspected to transmit the virus. The virus has been isolated from wild ducks upto one year after infection.
Pathogenesis: The incubation period is 3-7 days. A sudden, high and persistent mortality may be the first sign. Morbidity and mortality vary from 5-100%. Most ducks develop clinical signs and die. There is drop of egg production of about 25 to 40%. In affected birds there is viraemia, photophobia, thirst, diarrhoea, nasal and ocular discharge. Ingested virus causes enteritis and viraemic spread leads to vasuculitis and widespread focal necrosis. Blood is present in the body cavities including gizzard and intestinal lumens and petechial haemorrhages in many tissues. Intranuclear, inclusions are readily demonstrated in hapatocytes, intestinal epithelium and lymphoid tissues.

198 Textbook of Veterinary Virology
Diagnosis: Clinical and gross postmortem findings can be confmned by fmding herpesvirus inclusion bodies or positive immunofluorescnece. The virus can be isolated in duck embryos and identified }>y virus neutralization test
Control: The virus produces adequate immunity after natural infection. The inactivated vaccines do not give satisfactory immunity. A chicken embryo attenuated strains when administered subcutaneously, produce good immunity.
Pigeon Herpesvirus
It causes conjunctivitis: respiratory distress, diarrhoea, dehydration and emaciation with lesions in liver and other organs of pigeons. The virus grows in chicken embryo by CAM route and also in chicken embryo kidney and liver celJs. Pigeons 1-6 months of age are most often affected. Focal necrosis and A type intranuclear inclusions are found in the liver and kidneys.
Marek's Disease Viru~ (MDV)
It is a transmissible virus disease which mainly affects domestic fowl and is characterised by mononuclear infiltration around peripheral nerves and to lesser extent in skin, muscle, iris and internal organs. The disease affects mainly domestic poultry and is common in young birds of 2-5 months old. The first detailed account of the disease was published in Holland in 1914. The specific herpesvirus etiology was established in 1967. The disease occurs worldwide. In this country the disease was reported in 1970 and is prevalent in all parts of this country. Mohanty et al.(1973) reported annual losses to the tune of 40 million rupees to poultry industry of this country. Marek's disease is an important model for the study of tumorigenic potential of herpesviruses.
Properties of the virus: Two types of virus particles, onc is naked virion measuring 35-100 nm and second type of virion particles are enveloped and measure 150-170 nm in size, have been described. The enveloped virion of MDV are of two types, one of which is found in the nucleus of infected cells and is 150-180 nm in diameter and presumed to be noninfectious. The second type found predominantly in cytoplasm and in feather follicle epithelial cells is infectious and measure 250-280. nm. The genome is double stranded DNA with a moleular weight of

Herpesviridae 199
lxlO' daltons. The G+C content is 46 percent. In cultured cells· the virus is not readily demonstrated but preparations from lysed feather follicle epithelium reveal the presence of particles measuring between 275-400 nm in diameter. The virus survives for a long time at 60°C in growth medium containing 10% calf serum and 7.5% dimethyl sulphoxide. All the strains isolated have been found to be antigenically homogenous but some differences have been reported. The virus cross reacts with herpes simplex, Aujeszky's disease virus, IBR and EpsteinBarr virus. The highly oncogenic MDV strains and their attenuated variants have been placed in one group and low o~cogenic or non oncogenic strains in second group based on immunodiffusion tests. The herpes virus of turkey (HVT) forms a third antigenically related group. There is certain DNA homology between HVT and MDV viruses.
Cultivation: The virus can be cultivated in susceptible chickens, chicken embryos and cell cultures. When day old chicks are inoculated with suspected material the lesions can be detected after 2-4 weeks in ganglia, nerves and certain visceral organs. By fluorescent antibody technique many tissues of chicks show the viral antigens. The virus can also be cultivated on the CAM of susceptible chicken embryos or by yolk sac route. The virus produces discrete white pocks on the CAM. The virus grows in chicken kidney (CK) and duck embryo fibroblasts (DEF) and produces characteristic CPE in 6-14 days. The CPE consists of rounded and fusiform refractile cells and polykaryocytes which have inclusions in the nucleus. When cells from infected chicken kidney are grown, characteristic CPE is produced. The tumour, spleen, kidney, buffy coat or whole blood from affected birds is suitable material for virus isolation.
Epidemiology: Most of the birds at the time of maturity have antibodies in MDY although the infection persists and virus is shed in the dander of feather follicles. Congenital infcction does not occur. The disease occurs most commonly in young birds between2-5 months of age. If the chickens are exposed on the day of hatching the disease develops at 3 weeks of age. Natural transmission is air borne through inhalation. The infectious virus is present in oral, nasal and tracheal secretions and in feather follicle epithelium. Genetic resistance, age and viral strains influence the outcome of infection. A small percentage of infected birds develop clinical MD. During rust few days of life, chicks are very susceptible to the virus. The virus matures in the feather

200 Textbook o/Veterinary Virology
collicle and is shed through desquamated cells into the environment. The infected feathers remains infected for 6 weeks or more. Blood or tumor material containing live, intact cells can transmit the disease. Infected whole cell cultures may transmit the disease but not cell free fluid as· the virus is cell associated. Occasionally MD has also been reported from free flying birds like crow and myna.
Pathogenesis: The outcome of infection of chickens by MDV is influenced by virus strain, route of infection, dose, age, sex, immune status and genetic susceptibility of the chickens. Subclinical infection with virus shedding is common. The infection is acquired by inhalation. The epithelial cells of respiratory tract are productively infected and contribute to cell associated viraemia involving macrophages. By sixth day there is productive infection of lymphoid cells in a variety of organs. During the second week after infection there is persistent cell associated viraemia followed by proliferation of T lymphoblastoid cells and after few days cells begin to die. The lesions result from infiltration and proliferation of T lymphocytes which may result in leukemia and inflammatory cell response to the lysis of nonlymphoid cells by the virus. Enlargement of one or more peripheral nerve trunks is one of the most constant gross pathological finding. The nerves are 3 times their normal diameter and show loss of striations, oedematous, grey or yellowish and somewhat translucent in appearance. The celiac, cranial, intercostal, mesentiric, brachial, sciatic and greater splanchnic nerves are mostly involved. The enlargement of the nerve is usually unilateral. Lymphoma-tous lesions affects the gonads, lungs, liver, spleen, kidney and thymus.
The behaviour of MDV with lymphoid and non lymphoid cells differs considerably. In non lymphoid cells infectious virus particles are produced in feather follicle epithelium or non infectious virus particles or viral antigen is produced in other epithelial cells or in cell culture. In both cases the death of cell takes place. In lymphoid cells the transformation of cells take place with extensive proliferation without cell death. In these cells the presence of virus particles or the expression of viral antigen is not detected but in some Iymphoblastoid cell lines a low percentage of cells may produce viru~ particles or viral antigen.
~arek's disease is a progressive disease with variable signs. In neurolymphomatosis or classical disease there is paralysis of one or

H erpesviridae 201
both legs or wings. The early sign shown by the birds is incoordination. one leg held forward and other backwards. There is drooping of wings and lowering of head or neck. If vagus nerve is involved there may be dilation of crop and gasping. The acute marek's disease occurs in explosive outbreaks in large number of birds. The clinical signs are depression followed by ataxia and paralysis of some birds. In ocular lymphomatosis the iris of one or both eyes is gray in colour and the pupil is irregular ecentric. The cutaneous form of disease is recognised by nodular lesions upto 1 cm in diameter seen particularly at feather follicles.
Immune reaction: The infected nonlymphoid cells contain viral antigens while infected lymphoid cells contain Marek's associated tumor specific antigen (MATSA). These antigens can be detected by immunofluorescence and immunodiffusion test. MATSA can be detected by using immune sera prepared from chickens or rabbits immunized with transplantable tumor cells or cultured lymphoblastoid cells but not by antiserum derived from chickens that have recovered from MD. MATSA has been associated with cells transformation. The birds develop both humoral and cell mediated immunity (CM!) after infection with MD. The virus specific antibodies appear 1-3 weeks after infection and persist throughout the life of bird. The presence of antibody does not have a significant protective effect but may decrease the severity of the disease. The resistance to MD is primarily due to CMI response. T cells probably play a double role in MD by providing target cells for transformation by MDV and also by participating in immune mechanism against the development of lymphoid tumors. MDV has an immunosuppressive effect. The progeny of infected birds acquire passive immunity which disappears within 3-4 weeks.
Diagnosis: A tentative diagnosis is arrived at on the basis of symptoms and lesions. In classical form there is unilateral or bilateral paralysis of legs or wings, curling of toes, torticollis, dilatation of iris. In acute forms symptoms are rapid depression, emaciation and death. Most cases show oedematous swelling of vagus and sciatic nerves and of brachial, coeliac and lumber plexus.
Detection of viral antigen by immunofluorescence is the reliable diagnostic procedure.
MDV can be isolated by inoculation of susceptible chickens, cell cultures and embryonated chicken eggs. The specimens of choice are

202 Textbook o/Velerinary Virology
intact viable cens from buffy coat, spleen, tumour tissue or other lymphoid cens. Virus can also be isolated from skin and feather tips which contain cell free virus. The day old susceptible chicks ate inoculated intraperitoneally. After 18-21 days the birds show gross or microscopic lesions. The virus isolation and detection of serum antibody ftom the inoculated chickens should be undertaken. MDV produces characteristic CPE in duck embryo fibroblast (DEF) and chicken kidney (CK) cen culture. Characteristic plaques are produced in 6-14 days and consist of rounded refractile cells and polykaryocytes with cowdry type A inclusions. The virus produces pocks on the CAM of chicken embryos. Identification of virus isolated can be confirmed by immunofluorescent technique.
The antibodies can be demonstrated in the sera of infected or recovered birds by agar gel precipitation test, indirect fluorescent antibody test or passive haemagglutination test.
Control: MD is the first virus induced malignacy to be controlled by vaccination. There are three types of live vaccines available. These are herpes virus of turkeys (HVT). apathogenic strain of MDV and cell culture attenuated MDV. All these vaccines are equally effective. The vaccine virus persists in the vaccinated birds and does not prevent superinfection, replication or shedding of virulent MDV but prevents clinical disease. The resistance is life long in vaccinated birds. The mechanism of protection is due to cell mediated immunity response. The vaccinated chickens probably develop lymphopro1iferative lesions containing Marek's associated tumour specific surface antigen (MATS A) and a CMI response directed against MATSA protects the chickens against subsequent lymphoma formation by the ,:,irulent virus. The most extensively used vaccine is HVT. Chickens are generally vaccinated at hatching and should be reared in isolation. The hygienic procedures help to delay the exposure of the chickens and thus allow immunity to be established.
References
ALLAN, G.P.; YEARVAN, M.R.; TURllUEN, L.W.; BRYANS, J.J. and McCOLLUM, W.H., 1983. Molecular epizootologic studies of equifU! herpes virus-l infections by restriction endonuclease fingerprinting of viral DNA. American Journal of Veterinary Research. 44 (2): 263-27l.
BARAHONA, H.; MELl!NDEZ, L.V. and MELNICK, J.L., 1974. A compendium of

Ilerpesviridae 203
herpesviruses isolaJed from non human primates. Intervirology. 3: 175.
BAUDET, A.E.R.F., 1923. Mortality in ducks in the Netherlands caused by a fiIJrable virus fowl plaque. Tijdschi Diergeneesk. 50: 455-459.
BUXTON, A. and FRASER, G., 1977. Animal Microbiology. Vo1.2. Blackwell Scientific Publications. Oxford.
CAMPBELL, T.M. and STUDDERT, MJ., 1983. Equine herpesvirus type-1 (EHV1). Veterinary Bulletin53: 135-145.
CHURCHll.L, A.E. and BlOGS, P.M., 1967. Agent on Marek's disease in tissue culture. Nature215: 528-30.
DEWNETT~D.P.; BARASA, 1.0. and JOOIINSON, R.H., 1976. Infectious bovine rhinotracheitis virus: studies on the veneral carrier status in range cattle. Research in Veterinary Scicnce20: 77-83.
DEv PRAKASII and RAJYA, B.S., 1970. Avian leucosis complex. I. Demographic studies. Il. Pathoanatomy and serum lactic dehydrogenase level of Marek's disease in natural infection. Indian Journal of Animal SciencesAO: 282-296.
DEw, C. and McFERRAN, lB., 1966. Experimental studies on Aujesky's disease in cattle. Journal of comparative Pathology and Thcraeutics. 76: 379-385.
GmBS, E.P.J. and RWEYEMA.\iU, M.M., 1977. Bovine herpes viruses. Part. I. Bovine erpesvirus 1. Part 11. Bovine herpesviruses 2 and 3. Veterinary Bulletin, 47: 317-43 and 411-425.
GREIG, A.S.; BAUMISTER, G.L.; MITCIIFLL, O. and BARKAR, C.A.V., 1985. Cultivation in tissue culture of an infectious agent from coital exantheme of cattle. A preliminary report. Canadian Journal of Comparative Medicine. 22: 119-122.
GREWAL, GURDEV, SINOIl and SINGII, BALWANT, 1976. Incidence of Marek's disease virus infection in domestic fowl5 of Punjab (India). Avian Diseases. 20: 191-194.
HANSEN, L.E., 1978. Laryngotracheitis. In M.S.Hofstad, B.W.Calnek, C.F. Helmboldt. V.M. Reid and H.W. Yoder, JR (Editors), Diseases of POUltry. Oxford a:ld IBH Publishing Co.New Delhi pp. 607-618.
Infectiuous bovine rhinotracheitislinfectious pustular vulvovaginistis. 1984. Publication of Division of Pathology, Indian Veterinary Research Institute, Izatnagar, U.P.

204 Textbook o/Veterinary Virology
JAIN, N.C.; MANCHANDA, V.P.; GARO, D.N. and SHARMA, V.K., 1976. Isolation and characterisation of Equine herpes virus type-I. Veterinary Record.99: 57.
JAIN, N.C.; SB1lJ, P. and PRASAD, G., 1986. Herpes virus infections. In National symposium on 'Current status of herpes virus infections in man and animals in India' held at at Department of Veterinary Microbiology, Haryana Agricultural University, Hisar.
KAHRS, R.F., 1985. Viral disease of cattle. Kalyani Publishers, Ludiana.
HAY, H.G. and TITI'SLER, R.P., 1925. Tracheolaryngitis in Poultry. Journal of of American Veterinry Medical Association. 67: 229-23l.
MEHROTRA, M.L.; RAJYA, B.S. and KUMAR, S., 1976. Infectous Bovine rhinotracheitis (IBR). Keratoconjuctivitis in calves. Indian fournal of Veterinary Pathology. 1: 70-73.
MEHROTRA, M.L.; KUMAR, S. and RAJYA, B.S., 1981. Note on passive haemagglutination test for detection of infectious bovinerhinotracheitis infectious pustular vulvovaginitis (lBRIPN) virus antibody. Indian Journal of Animal Scineces. 51: 559-569.
Mll.LER, NJ., 1955. Infectious necrotic rhinotracheitiS of cattle. Journal of American Veterinary Medical Association. 126: 463-467.
MOHANTY,. G.C.; AcHARJYo, L.N. and RAJYA, B.S., 1973. Epidemiology of Marek's disease (MD). Stuides on the incidence of M.D. precipitins in some zoo birds. Poultry Scinece 52: 963-966
MOHANTY, S.B., 1990. Pseudorabies virus. In virus infections of ruminants edited by Z. Dinter and B. Morein. EIsevier Science Publishers B.V. Amsterdam.
MOHANTY, S.B. and DUTIA, S.K., 1981. Veterinary Virology. Lea and Febiger, Philitdelophia.
MUKERJI, A.; DAS, M.S.; GHOSH, B.B. and GANGULY, lL., 1963. Duck plague in West Bengal I and /I. Indian Veterinary Journal. 40: 457-462.
NAZARIAN. K.; LINDEHL, T.; KLElN, G. and LEE, L.F., 1973. Deoxyribonucleic acid of MareKs disease virus in virus induced tumors. Journal of Virology. 12: 341-346.
PARIHAR, N;S.; RAJYA, B.S. and Gll.L, B.S., 1975. Occurrence of malignant catarrhalfever in India. Indian Veterinary Journal. 52: 857-859.
PLOWRIGHT, W., 1990. Malignant catarrhal fever virus. In virus Infections of

Herpesviridae 205
ruminants, edited by D. Zinter and B. Morein. Elsevier science Publishers, B.V. Amsterdam.
PuRCHASE, H.G., 1972. Recent advances in thl: knowledge of Marek's disease. Advan~es in veterinary Science and comparative Medicine. 16: 229-53.
SAMBY AL, D.S. and BAXI, K.K., 1979. Isolation of Marek's disease virus from tissues of birds suspected for Marek's disease. Poultry Advisor, 12: 17-19.
SHARMA, G.L.; LALL, IM and BHALLA, N.P., 1965. Histopathological evidence of equine viral abortion in India. Indian Journal of Veterinary Science and Animal Husbandry 35: 18-23.
Singh, G.; Singh, B.; Guta; P.P. and Hothi, D.S., 1979. Epizootiological observations on malignant catarrhal fever and transmission of the disease in buffalo calves. (BubaIus bubalis). Acta Veterinaria Bruno. 48: 95-103.
SINGH, S.B.; SINGH, G.R. and SINGH, C.M., 1964. A preliminary report on the occurrence of infectious laryngotracheitis in poultry in Punjab. Poultry Science. 43: 492-494.
SlRAUB, P.C., 1990. Infectious bovine rhinotracheitis virus. In virus infections of ruminants, edited by D. Zinter and B. Morein. Elsevier Science Publishers B.V. AmsterdaIJ)..
STUDENT, M.I; SIMPSON, T. and Rozilllian, B., 1981. Differentiation of respiratory and abortigenic isolates of equine herpes virus I by restriction endonucleases. Science. 214: 562.
WrITHMAN, G.; GASKELL, R.M. and RZIHA, H.J., 1984. Latent herpes virus infections in Veterinary Medicine. In a seminar in the commission of European Communities held at Tubingen.· FRG. Martinus, Nijhoff Publishers, Boston.

Chapter 18
Unclassified DNA virus
African swine fever Virus (ASFV)
The African swine fever virus which was previously included in family Iridoviridae has now been taken out of this family but not yet been placed under any other family. African swine fever virus produces highly contagious fatal disease in pigs with some similarities to swine fever. The disease causes serious economic losses and is indigenous to African continent. The disease first appeared in 1910 in East Africa and Africa and then it spread to other territories of the continent. ASF was reported in 1957 from Portugal. A number of isolated outbreaks have been reported from Spain, France and Italy. In 1971 the disease was reported from Cuba. The disease has not been reported to exist in this country.
Property of virus: The virus particle measures 200-220 nm in diameter and consists of electron dense centrally placed nucleoid surrounded by a hexagonal outer shell. The virion acquires an additional outer envelope from plasma membrane. The genome consists of single stranded DNA, which codes for about 25 structural proteins and also some non structural proteins. The nucleoprotein core is surrounded by icosahedral sheet and contains 9% phospholipid. The virus is stable at room temperature for severn I weeks. The virus is inactivated in 30 minutes at 56 °C and survives for months and even years in refrigerated meat. The virus survives in chilled carcases for 15 weeks or longer and upto 6 months ID processed meat. The virus is stable over a wide pH range and is inactivated quickly in 2% NaOH.

Unclassified DNA Virus 207
The virus is not inactivated by crystal violet which is used to inactivate swine fever virus. The virus does not possess the property of haemagglutination but infected leucocyte cultures haem adsorb pig rbe. The haemadsorption is inhibited by African Swine fever serum. Different strains do not cross protect and it is probable that several anti genic types exist
The virus was fIrst propagated in the yolk sac of chicken embryos. Most of the strains replicate in the buffy coat cells of swine bone marrow. The virus produces syncytia and nuclear and cytoplasmic inclusions. in infected syncytia and nuclear and cytoplasmic inclusions in infected cell culture. however some strains do not produce CPE.
Epidemiology: Recovered animals develop long resistance to reinfection with homologous virus type but not against heterologous types. The virus from the infected animals is excreted in secretions and transmit the virus by contact or by aerosol route. The wart hogs in Africa act as reservoirs and transmit the infection to domestic pigs by indirect contact. The surviving pigs act as life long carriers. lice and ticks of pigs transmit the infection. Since the virus is resistant to atmospheric conditions and persists in the tissues of the carcases of carrier animals. the feeding of swill containing uncooked pork scraps is responsible for secondary epidemics and virus outbreaks. The outbreak in Portugal. Brazil etc. were due to feeding of untreated swill. The swine fever vaccine is also responsible for spread of outbreaks. In Brazil swine fever vaccine was contaminated with ASFV. In the inactivated crystal violet swine fever vaccine. the virus of ASFV survived and caused outbreaks.
Diagnosis: African swine fever is suspected when there is acute or paracute haemorrhagic disease with high mortality is encountered in SF vaccinated pigs. The provisional diagnosis can be made on clinical and postmortem lesions. A confirmation can be made by inoculating pigs susceptible and immune to ASF with the suspected material. The virus isolation can be attempted in cell culture from blood. splcen lymph nodes of infected animals. The infected leucocyte cultures haemadsorb pigs rbe and this property is not shown by SFV. The test based on this property of ASFV is applied for diagnosis. Complement fixation and gel diffusion tests are applied for detection of ASFV antibodies. Fluorescent antibody is also useful in the diagnosis.
Contrpl: Vaccination with killed as well as with live virus has not

208 Textbook o/Veterinary Virology
been effective. The virus appears to be a poor antigen. Pigs recovered from the disease are immune to homologous strain but not heterologous strains. The effective method of eradication is by slaughter and strict import regulations for meat products as well as live animals.
Reference
COOGlNS, L., 1974. African swine fever virus. Pathogenesis: Progress in medical Virology. 18: 48-63.
DETRAY, D.E., 1964. African swine fever virus. Advances in Veterinary Science 8: 299-333
MALlNQUlST, W.A. and HAY, D., 1960. Haemodsorption and cytopathic effect produced by African Swine fever virus in swine bone marrow and buffy coat cultures. American Journal of Veterinary Research 21, 104-108.
MAINER, F.D., 1975. African swine fever. In Diseases of swine, 4th Edition, Edited by H.W.Dunne and A.D. Lemann, Iowa, Iowa State University, Ames, Iowa.
RuSSELL, P.H. and EDlNGTON, N., 1985. Veterinary viruses. The Burlington Pr~ss (Cambirdge) Ltd. Foxton, Cambridge

PARTII
SYSTEMATIC VIROLOGY
R.N.A. VIRUSES


Chapter 19
Picornaviridae
Picorna name was introduced in 1962 to describe a group of very small RNA viruses. The family contains four genera namely Enterovirus, Rhinovirus, Cardiovirus and Aplhovirus. The viruses included in this family are small icosahedral nonenvcloped measuring 22-30 nm in diameter. The capsid appears smooth and round in outline and made up to 60 capsomeres each consisting of onc molecule of each of the four major polypeptides: VPl, 2,3 each of about 30K and VP4 of about 1OK. The picorna virus capsid contains one moleucle of infectious single stranded RNA of positive polarity with a molecular weight of about 2.5 x 1()6. A convalentIy linked protein VPg is present at 5' terminus of the RNA. Four major polypeptides participate in the construction of the capsid. Purified VPl elicit neutralising antibody, though less immunogenic than intact inactivated virus particle. This discovery has provided a major impetus fOr the development of recombinant DNA and synthetic peptide vaccines for foot and mouth disease.
The most important difference between the physiochemical properties of the virions of 4 major genera are the their pH stability. The apthovirus are unstable below pH 7 the rhinoviruses lose activity below pH 5 and entero and cardioviruses are stable at pH 4. The cardioviruses can be distinguished from enteroviruses by their biophasic pH stability in the presence of O.lM halide ions. Another important difference is the presence of polycytidylic acid tract of unknown function in the genome of aptho and cardioviruses but not in entero and rhinoviruses. If protected by mucus and strong sunlight,

212 Textbook o/Veterinary Virology
picornaviruses are relatively heat stable. Replication is entirely in the cytoplasm in <;lose association with the membranes. Partially double stranded replicative intermediates are formed. both strands of which are used as. templates. The functional proteins are found by posttranslational cleavage from large precursor polyproteins. During replication the metabolism of cell is stopped and cells seen show signs of degeneration. This degeneration of cells facilitates the release of progeny.
APTHOVIRUS
Apthovirus (Foot and Mouth Disease virus): Foot and mouth disease is a highly infectious disease of cattle. sheep. goats and pigs as well as of a number of wild animals like buffalo. deer, wild ungulates and hedgehogs. Man is occasionally infected. The disease is characterised by the formation of vesicles on the mucosa of mouth and sometimes alimentary tract and on the skin of the teats and udder. There is a sudden death in young animals due to heart failure. The disease has occurred at one time or other in all parts of the world. The earliest record of the disease comes from Northern Italy in 1514. The geographical distribution of the disease is world wide. The disease is prevalent in Europe, Asia, Africa and South America. It has not been reported from Newzealand and Iceland. Australia had its last outbreak in 1872. Canada had only one outbreak in 1952. The USA is free since 1929. In India the disease is widespread throughout the country. In England the disease was ftrst recorded in 1839. Subsequent serious outbreaks occured in 1871. 1922-24. 1952 and 1967-68.
Properties of the virus: Loeffler and Frosch (1898) reported the ftrst ftltrable agent responsible for causing foot and mouth disease. It is one of the smallest and earliest recognised animal virus. The intact virus particles of 25 nm and 7nm have been found. The viral genome is single stranded RNA with 8000 bases and the molecular weight of genome is 2.6 x 1()6 daltons. There is no cap at 5' end of RNA but instead a protein called VPg. The capsid has 4 polypeptides- VPl. VP2. VP3 and VP4. Traces of VPO and RNA polymerase is also found. There are 60 copies of 4 major polypeptides in each particle.
The foot and mouth disease virion is resistant to ether. chlorofonn. bile salts and detergents. The virus is acid-~nsitive and inactivated at pH 5.0 but is stable in the presence of magnesium chloride. It is stable

Picornaviridae 213
at pH 7.4-7.6 and at pH 3. The virus is resistant to drying, especially if it occurs quickly in the presence of protein. The virus persists on hay and straw for about 15 weeks and on hides for much longer time. It is stable at 4°C or when kept in frozen state but the virus does not survive in the muscles due to the production of acid during setting of meat. The virus is readily inactivated by heat but some strains survive higher temperature. Most strains lose infectivity within 48 hours at 37°C. It remains viable at 4-7°C for many months. The virus of FMD is more resistant to common disinfectants especially when the virus is mixed with organic material. Phenolic type disinfectants, alcohol, acetone, detergents and other organic solvents have little affect on the virus. Formalin, potassium permagnate, lactic acid, ethylene oxide, sublimate of mercury, 0.4% Beta propiolactone and 1-2% sodium carbonate is effective in field conditions.
The antigens produced during the infectious cycle of foot and mouth disease virus in tissue culture are given in Table 19.1.
Table 19.1
DESCRIPIlON OF MAL'! FMD VIRUS ANTIGENS (CROWI1IER, 1986)
Component
Whole particles
Empty particles
Subunit
VIA
Sedimentation Coefficient in Sucrose gradients
146S
75S
12S
3.58
Description
Contains a molecule of ss RNA (2.6 million m.wt) 60 copies of VP1, VP2, VP3, VP4 m.wts VP1-3 =24000
VP4 =8000 No RNA 60 copies VPO (uncleaved) VP2& VP4 60 copies of VPl, VP3 Pentamer of VPl, 2 and 3 (5 copies of each) Virus infection associated antigen, RNA polymerase associated m.wt. 56,000
All the components are antigenic. The integrity of whole particle

214 Textbook of Veterinary Virology
(l46S) is essential for formulation of successful inactivated vaccines. The subunits are antigenic but do not produce immunogenic response. The virus infection associated antigen (VIA) is produced in infected cells and animals and is antigenic. Most studies have implicated that VPl protein in responsible for antigenic variation. The FMDV was the fIrst virus in which anti genic differences between strains were recognised. Vallee and Carre in 1922 observed that French cattle which had recovered from disease were reinfected by sick animals imported from Germany. The two strains were named as Oise(O) and Allemagne(A) from the areas of origin. In 1926 Waldmann and Trautwein described 3rd type and named as type C. In 1930 there was an evidence that strains isolated from Africa do not fIt in the framework of 0, A and C. Galloway and others in 1948 while working at Pirbright confIrmed the presence of 3 more types and designated them as SAT!, SAT
1 and SAT)JIn 1954 a new strain isolated from Pakistan was
identifIed at Pirbright. The same strain was also confrrmed from material received from India, Thailand and Honkong and named as Asia 1. The 0, A and C types are widespread in world while Asia 1 occurs only in Asiatic countries. The SAT types were confined in African countries but in 1962 SAT! spread through Bahrain to the countries of Middle East to Turkey and USSR and strain Au in the same area in 1964-65. Within each serotype there are subtypes which may be suffIciently different to give very poor protection against other viruses of the same serotype. There appears to be continual antigenic drift in enzootic situations. The subtypes are defIned by crossneutralization tests. If one isolate is neutralized three times more slowly than another by antiserum to one of the two isolates it is defIned as different sUbtype and vaccinal cross-immunity is likely to be poor. The individual subtypes are indicated by numerical subscript. The number of subtypes in each serotype is A, 32; 0, 11; C, 5; Asia-I, 3; SAT!, 7; SAT1, 3; SAT),4.
Cultivation: The virus multiplies in cytoplasm of the cells. After attachment the virus particle enters the cell by phagocytosis where uncoating takes place. The virion RNA itself acts as mRNA. The replication of fresh RNA (virion RNA or mRNA) is via replicative intermediate shortly after the entry of the virus into the cell and it involves the translation of RNA into a polypeptide called polyprotein. This polyprotein is split into primary polypeptides or primary products

Picornaviridae 215
which in turn are split by virus specified proteases into polypeptides. The polypeptides are finally split into VP4• VPz• VP, and VPt• The assembly of polypeptide takes place and RNA is inserted into the particle which is then released by cell lysis. Intermediate products are also released on the lysis of cell.
The virus can be grown in laboratory animals. Guineapigs have long been used as experimental hosts, the route of inoculation being the hind footpads. Generalization takes place and vesicles are found on the other pads and tongue. Skinner(1951) used unweaned mice for cultivation of virus by intramuscular or intraperitoneal route. The virus multiplies in high titres in the heart and skeletal muscles. The virus can also be adapted to grow in older mice giving rise to paralysis. Young hamsters and new born rabbits can be readily infected with the production of paralysis and death. The virus has also been adapted to grow in chicks and chick embryos. The most sensitive assay system is multiple intradermolingual inoculation of cattle. Frenkel(1950) reported the multiplication of virus in surviving fragments of bovine tongue epithelium. The virus also grows in monolayers of bovine. porcine. lamb and goat epithelial cells, secondary bovine thyroid cells and continuous baby hamster kidney cell lines. Virus growth is detected by focal cell degeneration.
Epidemiology: The epidemiology of FMD focusses around its wide geographic distribution, its high transmissbility, the multiplicity of virus types and subtypes. The susceptibility of wide range of domestic as well as wild animals, the variety of methods of spread and the rapidity with which the virus spreads in animals, herds and countries and persistent infection in some species are the factors which complicate the epidemiolo~y of the disease.
There are large areas of the world in which one or more FMDV types are endemic. The disease is enzootic in Asia. Africa, and parts of Europe. The countries contiguous to infected areas or which import animal products from infected areas are at high risk. In countries where the disease in not present, the epizootic develops rapidly from introquction of disease to one farm. The outbreak spreads to many farms very rapidly due to highly infectious nature of virus. aerosol spread and short incubation period, which varies form 12 hours to 14 days. but usually is 2-6 days. The direct contact usually occurs from animal to animal by droplet infection or when virus contacts the

216 Textbook o/Veterinary Virology
mucosa of the mouth, nose, conjunctiva or abraded skin. Infection may be indirect through contact with contaminated objects. The spread of virus may occur during the incubation period or at the time of disease in an animal. When an animal is slaughtered at a later stage of incubation' or in the early stage or development of lesions, the virus is present in the tissues. In muscles the virus is inactivated due to acid formation but it survives in lymph nodes, bone marrow and offal. The susceptible animals when come in contact with such material may develop disease. The virus is excreted in milk in later stages of incubation period or early states of disease. Similarly, semen contains virus just before the lesions appear.
The most important route of infection for cattle, sheep and goats appears to be respiratory tract by inhalation. The cattle is most susceptible as they breathe greater volu~e of air and the infectious dose is small. The air borne transmission is dependent on wind direction and speed and is favoured by low temperature, high humidity and overcast skies. The long distance airborne transmission was only realised in 1967-1968 in England which caused direct and indirect losses estimated to $200,000,000 and approximately 634,000 animals were slaughtered. Most spread over land is within 10 kms but depending upon topography, the spread may be 60 kms or more over land and 250 kIns over sea. The criteria for spread are high virus output and survival, low virus dispersi~n and large number of susceptible livestock. The airborne spread has been found mainly in temparate climate, where there is high humidity (60 percent) and density of livestock population is high. In tropical and subtropical countries airborne spread takes place over shorter distances especially during night.
The virus may survive longer in the pharyngeal region of recovered cattle due to which it is maintained for longer periods of time. Though the spread of virus from such animals to susceptible animals is difficult. The pharyngeal secretions may also contain antibodies which neutralise the virus. It could be possible that new antigenic variants could arise in such immune animals.
In South America the Virus is maintained by continual spread among the susceptible population of domestic animals. While in Africa the wild game population plays a role in spreading the infection to domestic livestock where there is extensive contact between wild game animals and cattle. The role of sheep and goat varies in different parls

Picornaviridae 217
of world. In Middle East sheep and goat form major pathway of infection.
In India three major factors of spread of infection have been delineated, (i) movement of affected animals form one place to another, (ii) intermixing of affected and healthy livestock at common grazing ground and (iii) cattle fairs. The problem of the disease is of very complex in nature. India has approximately 400 million susceptible livestock apart from wild animal species. The presence of three main types, i.e A,O,C and their respective variants was reported by Seetharaman. and Datt (1951). These typing results were based upon the cross immunity tests in guinea pigs. Dhanda et al. (1957) reported a fresh isolation of an atypical strain, which was later confirmed from the World Reference Laboratory belonging to type Asia l. An analysis of reported outbreaks of the disease among cattle and buffaloes during the period 1943 to 1973 has shown an average of 5000 outbreaks involving as many as 2,75,000 animals each year. The accumulated data of 42 years (1943 to 1984) show the following prevalence of different types of FMD virus in India (Anon, 1984-85)
F & M virus serolypes
No.of isolates recorded. Percentage
o
3972 54.8
A
916 12.6
C Asia-1
903 1463 12.5 20.2
The percentage figure given above for type A is inclusive of both A5/AlO and A22 strains. In India only four FMD virus types O,A, C and Asia-l are known to occur. The virus types have a regional pref(1rence. During 15 years period type ° has emerged as dominant type in the 8 states of Jammu and Kashmir, Punjab, Himachal Pradesh, Haryana, Uttar Pradesh, Bihar, Sikkim and Kerala. Type Asia 1 was dominant in Madhya Pradesh, Gujrat and Karnataka. Type A was dominant in Andhra Pradesh and Karnak1ka. Virus type C maintained a low profile, the outbreaks occured more in Tamil Nadu, Kerala and Maharastra. Sporadic occurrence was in Himachal Pradesh, Punjab, Uttar Pradesh, Rajasthnn, Madhya Pradesh and Bihar.
Palhogenesis: The incubation period is usually 2-5 days but may be extend~ upto 2-3 weeks. The most severe disease is seen among cattle and pigs. The virus infects the epithelium of upper respiratory tract or alimentary tract by inspiration or ingestion. The virus multiplies

218 Textbook o/Veterinary Virology
at the site of infection where primary vesicles are fonned in 1-4 days. Mter primary vescile fonnation the virus initates viraemia associated with fever. The virus localises in distant epithelia and secondary vesicles appear. Vesicles occur in stratified squamous epithelia or mucous membrane. The vesicles appear on the mucous membrane of tongue, lips, gums, cheeks, dental pad and on the skin of interdigital space, on the coronary bands of feet, at the bulbs of heels and on teats and udder. The lesions in the mouth cause excessive salivation, smacking of lips and anorexia Vesicles and erosions on teats areassociated with rapid drop in milk yield and may be followed by mastitis. Internally lesions may be found in oesophagus and fore stomachs. Sheep and goats may only show lameness without mouth lesions. In pigs lesions may be seen on snout as well as on the feet. In young animals the virus localizes in the heart and produce myocarditis. The disease is generally mild in adult cattle, but pregnant cows may abort. Although morbidity is high, mortality is low but mortality may be high in young calves and lambs. The importance of FMD lie in heavy economic losses. In India it causes an estimated annual loss of US$ 400 million in milk and US$ 10 million in impaired' working capacity of bullocks.
Immune reaction: Humoral antibodies can be detected after about 4 days of infection. The early antibodies are of IgM type and reach a peak titre between 7-14 days and declines within 30 days. These early antibodies of IgM class neutralize and precipitate both homologus and heterologus virus. The IgG class antibodies appear after 10-14 days and reach peak at about 28 days. These antibodies are more type specific, neutralize and precipitate virus and fix complement. The cattle after recovery remain totally immune for 3-4 months. The partial immunity can persist for 4-5 years. The rate at which immunity declines depends upon age, level of nutrition, physiological sUite and breed of animals. The antibodies are transferred to calves through colostrum. These antibodies in calves interfere with vaccination for 3-6 months and afford protection for about 6 weeks of age. Immunity following vaccination is shorter. A single inoculation of inactivated vaccine protects the animal to challenge for 3-6 months.
Diagnosis: The highly contagious nature of disease and presence of salivation, typical raised vesicles with blanched covering epithelium filled with straw coloured fluid, lameness are usually p'1thogenomonic of FMD. The laboratory tests are always essential to establish type and

Picornaviridae 219
subtype and also to differentiate FMD from other vesicular diseases like vesicular stomatis, vesicular exanthema and swine vesicular exanthma.
Table 19.2 DIFFERENTIAL DIAGNOSIS OF VESICULAR DISEASE
Animal species Route of inoculation FMD VS
Natural infection Cattle + Pig + Sheep & goat + Horse
Experimental Infection Cattle Intradermal in tongue, gums +
and lips Intramuscular +
Pig Intradermal in snout, lips + intramuscular
Sheep & Intradermal + goats Horse Intradermal in tongue
Guinea pig Intradermal in foot pad Unweaned mice Adult chicken Intradermal in tongue Growth in tissue culture Calf thyroid BHK IB-RS-2 Chick embryo cells
+ + +
+ + +
+ + ± +
+
+
+
+
+ + +
+ +
+ +
VE SVE
+ +
+ +
+
+
no lesions (+)
no lesions
(some strains)
+
+
Morphology (electron Microscope) Sphe- Bul- Sphe- Spherical let rical rical
shaped dark 30 nm 175x65 stain-
nm ing areas 35--40 nm
FMD Foot arId mouth disease VS Vesicular stomatitis VE Vesicular exanthema SVE Swine vesicular exanthema
In India vesicular disease except FMD have not been reported. It is
\

220 Textbook o/Veterinary Virology
worth while to summarise the differential diagnosis in a table 19.2. The specimens for diagnosis should be collected from those animals that developed clinical signs recently i.e. when the animals have temperature., nasal discharge and the vesicles appear in the mouth and! or the feet. Samples from at least 2 animals should be collected. The samples include blood in anticoagulant. serum pharyngeal and oesophageal fluid collected with cup Probang. The quantity of epithelium available varies from animal to animal but a piece of 2 x 2 cms is appropriate. From dead animals samples may be collected from lymph nodes, thyroid and heart. The samples are frozen and despatched to the laboratory in frozen state. Where it is not possible to send samples in frozen state, the samples are transported in glycerol buffer pH 7.6.
Virus isolation: Cattle, guinea pigs, suckling mice and cell culture are used for isolation of virus when the concentration of the virus in the
" samples is low. The isolated virus is identified by complement fixation or neutralization test.
Cattle: The cattle are most susceptible but are seldom used for routine investigations because of cost and other difficulties of keeping the animals in strict isolation. The virus suspension is inoculated dermolingually into the dorsum of tongue by making several inoculations. About I ml of inoculum is used for injection. The temperature, vesicles and other sysmptoms usually develop between 24-72 hours of inoculation. The vesicular epithelium is collected for carrying out serological test.
Mice: The unweaned mice(6-8 days old) are highly susceptible to experimental infection and can be used for isolation of virus from field samples instead of cattle. The suspected material is inoculated in all th~ animals of the litter except one animal, which is kept as control. If any animal dies between 1-7 days its carcases are used as source of antigen. The mice after 3-4 days of inoculation show paralytic syptoms and die.
Guinea pigs: Healthy guinea pigs weighing about 400 to 500g body weight are used for inoculation by intradermal route into the planter pads by making tunnels. The pads become red and vesicles appear usually by 3rd day of inoculation. The tissues from the lesions are collected and llSed as source of antigen.
Cell cultures: Bovine thyroid, BHK-21, IB-RS-2 cells are used for isolation of virus. Two or three tubes are inoculated with the suspect

Picornaviridae 221
material and incubated at 37°C. A marked degenerative type of cytopatbic effect is observed within 24-48 hrs. The cell culture fluid is used as antigen. In case cytopathic effect is not visible 2-3 passages more may be required.
Serological tests: The serological tests are applied directly to the field material or the material is passaged in experimental animals or in cell culture and material collected is used as antigen.
Complement fIXation text: It is one of the most commonly used and very important for identifying the virus type from the suspected samples of FMD. The antigen used in this test obtained from original suspected material or from cell culture of tissues from the experimental inoculated animals. The hyperimmune antisera against known types is prepared in guinea pigs.
The complement present in the test combines with homologous antigen-antibody complexes. The amount of complement fixed to the extent of 50% by type specific serum and unknown antigen are assessed. The serum antigen complex giving the highest fix.ation is described as homologus. In this way the unknown antigen is identified as belonging to particular type. The method described by Brooksby (1952) is followed in most of the laboratories. The method is quick and results can be obtained within 2-3 hours. The quantity of complement fixed is estimated visually, semiquantitatively, or quantitatively and the exact amount required for giving 50% fixation is ascertained. The micro-complement fixation test is preferred because it conserves the reagents, is simple to perform and large number of samples can be processed at one time.
Neutralization text: This test is used to detect specific antibodies in the sera of recovered animals or for identification of virus with known hyperimmune sera. The sub types are defined by cross neutralization test. The tests are performed by constant virus and varying dilution of sera or vice versa. In cross neutralization test if one isolate is neutralized three times more slowly than another by antiserum to onc of the 2 isolates it is defined as a different subtype and vaccinal cross immunity is likely to be poor.
Gel diffusion lest: The gel diffusion test in agar can also be used for identifying unknown FMD virus strains. The test can be carried out in slides. The test is not very sensitive.
Radial immuno haemolysis lest: This test was applied by several

222 Textbook a/Veterinary Virology
workers to type and subtype strains of FMD virus and a positive correlation of the results with complement fixation test has been reported. The virus antigen is coupled with sheep erythrocytes in presence of chromium chloride and is used for detection of specific antibodies ill the serum.
Immunofluorescence test: The immunofluorescence test is used for detection of antigen in tongue epithelium, oesopharyngeal fluid and various organs of dead animals suspected to have died of FMD. It gives a quick diagnosis in the face of outbreak where the virus cannot be isolated from the samples or for screening of large number of carrier animals by examining the oesopharyngeal fluid. The test can also be used for typing of FMD virus strains but it is a very lengthy procedure and cannot be used as a rountine test.
Immunoperoxidase test: The enzyme horse redish peroxidase is attached with antibody. The peroxidase labelled antibody is reacted to viral antigen (intracellular or extracellular). The reaction is observed by use of substrate which reacts with peroxidase and produces a colour. The test can also be applied for typing of FMD strains where confirmation of complement fixation test is required. In routine, the test is not applied forvirus typing.
Enzyme linked immunosorbent assay (ELISA). The test is hundred fold more sensitive and is likely to replace complement fixation test in FMD virus diagnosis in future. The test is used for serotyping of FMD viruses using ELISA has a. potential to compare viruses according to their immunogenic properties so that in future, it may be possible to predict the protective qualities of vaccine strains against field isolates.
Recently electrofocussing and finger printing of ribonuclease T] oliogonucleotides have been used in FMD virus diagnosis and typing of strains.
Control: The recovered animals produce VN and CF antibodies and resist reinfection by the same subtype of virus for upto one year or more. A modified disease may result when animals immune to one sUbtype are exposed to a second sUbtype of the original serotype. If exposure is to a second serotype there is no resistance to disease.
Various methods for dealing with FMD outbreaks are used in different countries. The stamping out or slaughter method of eradication has been used in USA and other countries. In our country where the disease is enzootic the vaccination is practised. The widely

Picornaviridae 223
used vaccines are inactivated and include the viral types and subtypes that prevail in the field. Some of the current killed vaccines are made from virus grown in BHK cells, Bovine Kidney and PK-15 cell lines. The adjuvants used to enhance the immune response are aluminium hydroxide and oil _ adjuvant. For- inactivation acetyl ethylineimine or formalin is used. Vaccination is usually repeated after 6-12 months. Live vaccines are used to a limited extent only. Research is in progress to produce a subunit vaccine. The following methods are being investigated.
a. A DNA copy is made of the fragment of RNA which codes for VP-l. This DNA is ligated to an erythromycin resistant phage of E.coli. Selective medium is used to select for E.coli containing the phage vector and some such E.coli produce VP-l. This can be produced in large quantities.
b. Peptide synthesizer is used to make peptide sequences of VP-l, 10-15 amino acids in length. These peptides have been tested and some are protective in guinea pigs. This may be the future hope.
ENTEROVIRUS
Enteroviruses are resistant to ether and chloroform and can be distinguished from rhinoviruses by their resistance to acids. These viruses are readily isolated from faeces and other excretions and rapidly produce CPE in a variety of cell culture.
Swine Enteroviruses
The porcine enteroviruses (PEV) are ubiquitous in nature. There are possibly 11 serotypes. Procine enterovirus I cause polioencephalitis or Teschen disease while PEV9 induces swine vesicular disease. The other serotype do not appear to be pathogenic.
Porcine Enterovirus-l (Teschan virus, Talran disease virus, Polio encephalomyelitis virus)
The domestic and wild pigs are susceptible to Teschen virus. The disease was reported in 1930 from Czechoslovakia. The disease occurs in milder form in Denmark and England. The disease has been reported from several European and African countries. The disease may also be present in USA, Canada and Australia in a milder form.
Properties of the virus: The porcine enteroviruses are larger than foot and mouth disease virus, measuring between 25-30 om in

224 Textbook of Veterinary Virology
diameter. They are non-enveloped with 32 capsomeres. They are ether and chloroform resistant; the change in pH has little effect. The virus can be preserved at 4°C in glycerine saline or -50°C but lose infectivity if preserved by freeze drying.
Cultivation: The virus does not grow in chicken embryo or other laboratory animals. The virus can be cultivated in pig kidney cell cultures.
Epidemiology: The disease occurs mainly in young animals aged 5-8 weeks. Faeces are the main source of contamination but pharyngeal secretions make aerosal spread possible. Carrier animals are thought to exist; the virus persist in environment and can be transmitted by fomites.
Palhogenesis: The disease among pigs may be acute, mild, inapparent or chronic. Most of the outbreaks are sporadic and inapparent while the disease may effect the entire herd sometime. The incubation period ranges from 4 to 20 days. The clinical symptoms are fever, anorexia and depression. There may be tremors and incoordination of hind legs with stiffness, prostration and coma leading to death. Paralysis of hind legs is a dominant symptom. There is diffuse encephalomyelitis with degeneration of neurons and perivascular cuffing. The mortality ranges from 50 to 75 percent.
Diagnosis: Isolation and identification of virus from the brain, the intestinal tract and faeces should be carried out. The FAT may be applied to the sections of brain.
COlltrol: Live attenuated or formalin inactivated vaccines are in use in Europe with some success.
Porcine enterovirus-9 (PEV 9) (Swine Vesicular disease virus)
Swine vesicular disease (SVD) is clinically indistinguishable from FMD, Vesicular stomatitis (VS) and Vesicular Exanthema (VE) of swine. The disease was rust reported from Italy in 1966. It has been reported from Hongkong, Japan, UK and C?ther parts of Europe. The disease is only restricted to pigs.
Properties of the virus: The virus measures 30 nm in diameter. The density of the virus is 1.34 glml with a sedimentation coefficient of 1508. The virus is stable over a wide pH range of 2-12 and protected against inactivation at 50°C by IM MgClz• Minor anti genic variation in different strains have been observed. The virus is antigenically related to coxsackie-B-5 virus of man.

Picornaviridae 225
Cultivation: The virus does not grown in chicken embryos but readily grows in primary PK cells and in PK-15 cell lines. It is highly cytopathic, CPE is detected as early as 6 hours after inoculation and cell destruction is completed in 24 hours. Newborn mice are readily infected by intracerebral or intraperitoneal inoculation but older mice are refractory.
Epidemiology: Transmission of the virus is by direct and indirect contact and ,by feeding pigs with uncooked garbage. Feeding of swill has remained the primary source of outbreak:. The virus is very resistant and therefore persists in the environment and carcases.
Pathogenesis: The incubation period is 2-7 days. The clinical signs in pigs are high temperature (41-42°C) and development of vesicular lesions on snout, coronary bands, bulbs of heels and interdigital spl!ce. Healing is quick and animals recover completely. Subclinical disease can occur. Laboratory workers get the infection and show symptoms of malaise and mild meningoencephalitis.
Diagnosis: Vasicular fluid is a rich source of antigen for CFT and counter immunoclectrophoresis (CIE). The CIE is rapid and very little quantity of antigen is required. Animal susceptibility tests should be used to differentiate from FMD~ VS and VE. Day old mice are susceptible and die within 5-10 days of inoculation but 7 days old mice and guinea pigs are susceptible only to FMDV. Virus isolation can be done in pig kidney cells or PK 15 cell lines.
Control: No vaccine is available. Quarantine and slaughter policies are implemented to control the disease. Prohibition of feeding of uncooked swill must be enforced.
Bovine Enteroviruses
These viruses have been isolated from the intestinal tracts of healthy cattle as well as from the respIratory and reproductive disease. Ther are 7 serotypes known. These viruses grow well in cell cultures of bovine embryonic and adult kidney. They are cytopathic and produce lysis of cells. Some strains possess the property of haemagglutination. The role of these viruses in respiratory and reproductive disease in cattle and nconatal calves is not clear. Some serotypes are reported to produce catarrhal type vaginitis. Respiratory illness can be produced in cortisone treated calves; in colostrum deprived calves the virus produces fever and leukopenia. Experimentally the virus produces

226 Textbook o/Veterinary Virology
abortion and stillbirth in guinea pigs. Dunne et al. 1973 detected BEV antibodies in bovine aborted foetuses. They consider these viruses as possible causative agents in bovine abortion either alone or in combination with bovine parainfluenza 3 virus.
Avian Encephalomyelitis Virus (AEV)
The virus produces an acute disease of young chicks; the infection in mature birds is mild or inapparent The natural outbreaks occur in chickens, pheasants and Japanese quail. The disease was reported in 1930 from USA for the fIrst time. Now the disease occurs in almost all areas of the world where poultry is raised. The virus' is embryo transmitted and disease spreads via egg or young chicks.
Properties of the virus: The virus has properties similar to other enteroviruses. All the strains are anti genically similar. There are 15 serotypes of avian enteroviruses; the AEV is particularly a neuturotropic and exhibit variable pathogenecity. Some strains are more neturotropic.
Cultivation: The virus can be cultivated in baby chicks, chicken embryo kidney, chicken fIbroblasts and cultures of neuroglial cells. The yolk sac route of inoculation in chicken embryos is route of choice.
Epidemiology: The disease is transmitted vertically through infected eggs or horizontally by ingestion of contaminated feed. The virus is shed in the droppings. The transmission of infection can occur in the incubator.
Pathogenesis: The incubation period by experimental inoculation by various routes is 6-10 days. The disease mainly affects young chicks during first 6 weeks of age. The disease is characterized by dullness, progressive ataxia especially of head and neck, loss of condition, paralysis, prostration and death. The mortality is about 25% while morbidity is upto 60%. The adult birds only show a drop in egg production and loose droppings. No gross lesions are found, the principal microscopic lesions are found in the eNS.
Diagnosis: A tentative diagnosis in young chicks can be arrived at by observing ataxia and tremors in large number of young chicks. The diagnosis is diffIcult in adult birds. ConfIrmatory diagnosis can be made by isolation and identifIcation of the virus. The brain from the suspected chicks is inoculated via yolk sac in 5-7 days old chick embryos. The chicks are allowed to hatch and observed for 10 days for

Picor naviridae 227
symptoms. The clinical material can also be inoculated intracerebrally in day old chicks. The FAT is used for detection of antigen in the CNS.
Control: Recovered birds are immune. Attenuated egg adapted vaccines are given in drinking water to chickens 10-20 weeks old. The adult birds can be vaccinated by wing web method as well. Inactivated vaccines have been used in laying flocks in some countries. Immunization of breeding flocks prevents the transmission of virus via egg. The chicks which survive the attack remain unthrifty or become blind.
Duck Hepatitis Virus (DHV) The virus causes a fatal disease of young ducklings, affecting those
under 1 month of age. The disease is prevalent all over the world including India.
Properties of the virus: The virus measures 20-40 nm in diameter with a core of ~ingle stranded RNA. The virus is resistant; it survives for 10 weeks in brooders and 5 weeks in droppings. The virus is resistant to 2% lysol at 37°C for 60 .minutes and 0.1 % formalin for 8 hours at 37°C. It does not possess the property of haemagglutination.
Cultivation: The virus can be cultivated in chicken embryos via allantoic cavity route and kills 60% of embryos after 5-6 days of inoculation. The embryos are stunted and oedematous. After 20-30 serial passages, the virus becomes attenuated for duckling. The virus replicates in chicken embryo tissues without showing any CPE.
Epidemiology: The virus is present in the blood. Transmission is by contact. Recovered ducks shed the virus upto 8 weeks in the faeces. Both vaccinated and affected birds excrete the virus in faeces for months.
Pathogentsis: Duckling upto 3 weeks are naturally affected. The adult ducks, chickens and turkeys are not affected. The incubation period is 18-24 hrs but may range from 1-5 days. The disease takes rapid course and mortality goes upto 90-95%. The ducklings stop moving, fall on their sides, kick spasmodically and die within 3-4 days. The characteristic lesions are found in the liver, which is enlarged, oedematous and mottled with haemorrhages. The spleen and kidneys may be swollen. Microscopically' there is necrosis of hepatic cells and proliferation of bile duct epithelium.
Diagnosis: The history of fatal rapidly spreading illness among ducklings and presence of gross lesions in the liver are helpful in the

228 Textbook of Veterinary Virology
diagnosis of the disease. ·The virus can be isolated from liver or blood of affected birds in the chicken embryos via allantoic cavity roule. The FAT is used for diagnosis.
Control: Recovered birds are immune. An egg adapted attenuated vaccine is used to vaccinate day old ducklings via foot web or intramuscular or intranasal route of inoculation. The vaccine can be given to breeding stock to transfer passive immunity giving two doses at 6 weeks apart.
RHINOVIRUS
The rhinoviruses are small spherical particles measuring 20-30 nm in diameter with a single stranded RNA with molecular weight of 2.8 x 1()6 daltons. Unlike human rhinoviruses of which more than 110 serotypes are recognised, bovine and equine rhinoviruses are in a narrower antigenic group with only 2 serotypes in each group.
Bovine Rhinovirus-l (BRV-l)
Bovine rhinoviruses are known since 1962. They are widely distributed among cattle population throughout the world but their pathogenic potential remains to be elaborated.
Properties of the virus: They are small in size measuring about 30 nm in diameter. They are sensitive to mild acid and resistant to sodium dodecyl sulphate and are not stablized in MgClz at 50°C.
Cultivation: The virus replicates in cell culture of bovine origin. Primary bovine embryo kidney, bovine kidney and bovine tracheal organ cultures are used for propagating the virus. The cytopathogenicity may be meagre on primary isolation but becomes pronounced after adaptation in kidney cells. The virus grows better at 30-33°C rather than at 37°C, this may reflect to their adaptation to the lower temperature of their natural habitat in the nasal passages.
Epidemiology: The disease spreads by close contact. Nasal secretions are probably source of infection. The cattle acts as a reservoir host.
Pathogenesis: The pathogenecity of different strains vary. These viruses have been isolated from cattle with respiratory diseases from USA, England, Germany and Japan. They have been isolated from the epizootics of shipping fever. In Japan these viruses have been isolated from calves suffering from non-fatal but rapidly spreading respiratory

Picornaviridae 229
disease. Experimental infections have been mild and a symptomatic, however, a transient rise in temperature and pneumonia may occur. Gross lesions are mainly confmed to respiratory tract Areas of pulmonary consolidation, collapse and emphysema are present.
Diagnosis: Isolation of virus from cattle with acute respiratory disease alongwith increase of four fold SN antibody titre confIrms the diagnosis.
Control: There is no vaccine available.
Equine Rbinoviruses 1 & 2
Two distinct serotypes are known. The 2 serotypes differ from one another in physical and chemical characters. These viruses grow and produce CPE in equine foetal kidney cell cultures and in rabbit kidney cell lines. These viruses cause a disease of upper respiratory tract. The disease is mild or inapparent in uncomplicated cases. The incubation period is 3-9 days. The affected animals show fever, cough, nasal discharge and pharyngitis. The morbidity is high. Transmission is by direct and indirect contact with nasal secretions of affected horses. Diagnosis is confined by isolation and identiflcation of viruses by VN and CF tests. There is no vaccine currently available.
References
ADLAKHA, S.C., 1985. National overview. India: In Veterinary viral diseases. Edited by Antony J. Dell-Porte, New York,
ANoNYMOUS 1984-1985. Report of the task force on foot and mouth disease. Dept1. of Agriculture and Cooperation Minsitry of Agriculture and Rural Development, Gov1. of India.
BROOKSBY, lB., 1952. The technique of complement [vcation infoot and mouth disease research. Ser. No.12, London Her Majesty's Stationery Office, London.
BUXTON, A. and FRASER, G., 1977. Animal Microbiology Vol. 2. Oxford, Blackwell Scientific Publications.
CROWTHER, lR., 1986. Antigenic structure of foot and mouth disease virus. Rev. Sci. Tech. Off. Int. Epiz.5, 299-314.
DeuA, PORTB, A.l., 1983. Current status of foot and mouth disease vaccines including the use of genetic engineering. Aust. Vet. 190: 129-35.

230 Textbook o/Veterinary Virology
DHANDA, M.R.; GoPALKRISHNAN, V.R. and DHlLLON, M.S., 1957. Studies onfoot and l'IIQuth disease vaccination in IndiaJI. A concentrated crystal violet vaccine. Indian J. Vet. Sci. 27: 127-132.
DIN'rnR, Z. and MORBlN, B., 1990. Virus infections of ruminants. Elsevier Science Publishers B.V.Amsterdam.
DUNNE, H.W. and lEMAN, A.D., 1975. Diseases of swine, 4th Ed. Ames, Iowa, Iowa State University Press.
FRENKEL, H.S., 1950. Research onfoot and mouth disease. 11. The cultivation of the virus in explantations of tongue epithelium of bovinne animals. AmerJ. vet. Res.ll~ 371-373.
GALLOWAY, lA.; HENnPJtSON, W.M. and BROOKSBY, lB., 1948. Strains of the virus of foot and mouth disease recovered from outbreaks in Mexico. Proc. Sco. Exp. BioI. Med. 67, 57-63.
HOFSTAD, M.S.; CALNEK, B.W.; HELMBOLDT, C.F.; REID, W.M. and YODER, JR. H.W., 1975. Diseases of poultry . Oxford Publishing Company, Bombay.
LoEfR.ER and FROSCH., 1889. Cited by HENNJNG W.M., 1955. Foot and Mouth Disease. Page. 895. South Africa, Central News Agency Ltd. Oonderstepoort, South Africa.
MOHANTY, S.B and DU1TA, S.K., 1981. Veteriary Virology, Lea and Febiger, Philadelphia.
MowAT, G.N., 1986. Epidemiology offoot and mouth disease in Europe. Rev. Sci. Tech. Off. Int. Epiz. 5,271-78.
PEREIRA, H.G., 1981. Foot and Mouth disease in virus diseases of Food animals. Edited by E.P.J. Gibbs, VoI.II. New York Academic Press.
PHnJp, J.I.H. and DARBYSHIRE, I.H., 1974. Respiratory viruses of cattle. In advances of Veterinary Science and comparative Medicine. Edited,by C.A. Brandly and C.E. Comellius Vol. 15, New York Academic Press
RusSllLL, P.H. and EDINGTON, N., 1985. Veterinary viruses. The Burlington Press (Camb) Ltd. Foxton, Cambridge.
SEETHRAMAN, C. and DATT, N.S., 1951. Frequency of occurrence of foot and mouth disease virus in India. Indian 1. Vet. Sci. 21, 251~255.
WALDMAN, O. and TRAUTWEIN, K., 1926. Experimentalli unlerauchungen uber Maul-and Klauenzeche virus. Berl.tierarztle Wochebschr 42, 569-571.

Chapter 20
Calciviridae
The members of this family are isometric nonenveloped particles measuring about 35-40 nm in diameter. They have a icosahedral symmetry with 32 capsomeres. The surface of the capsid is composed of a number of dark hollow cup shaped structure from which the generic name is derived They are ether and heat resistant, stable at pH 5 but not at pH 3. The genome is (+) sense ss RNA and is infectious. During replication at least 3 RNA species are transcribed from a (-) sense RNA template. One of these is of genomic size while two are subgenomic size. Out of the two subgenomic size RNA the larger one codes for large capsid polypeptide. Other polypeptides are found in the cytoplasm of infected cells. The posttranslational cleavage of a polyprotein does not occur. Calciviruses replicate in the cytoplasm and are rapidly cytopathogenic. They can be cultivated easily in the embryonic kidney cells from the species of origin. Two members of this family viz. vesicular exanthema virus (VEV) and feline calcivirus (FCV) cause serious animal diseases.
Vesicular Exanthema Virus (VEV)
The virus causes an acute disease of swine characterised by formation of vesicles on the snout, mouth and feet. The only natural host is pig but dogs and horses can be infected experimentally. The disease has only been reported from California coast.
Properties of the virus: The virus particles are spherical in shape and measure between 30-40 nm. The virus contain 20-24% RNA with a sedimentation rate of 207S. The virus is heat resistant and survives at

232 Textbook o/Veterinary Virology
60°C for 1 hour. The virus is also resistant to ether, chloroform, deoxycholate but is readily inactivated by 2 percent NaOH. There are 18 serotypes of the virus known which do not cross protect.
The virus can be easily cultivated in pig kidney, lung, liver, testis but to a lesser extent in cell cultures of other hosts like horses, dogs and cats. The virus does not grow in chicken embryos.
Epidemiology: The disease spreads by contact with infected pigs, unboiled swill and carrier animals. Virus is present in almost all the tissues of infected swine. Small amounts of virus may be excreted in the urine and faeces. During outbreak, transmission occurs by direct contact from pig to pig.
Pathogenicity: YE is a disease of swine. Swine of all ages are susceptible. The mortality rate in pigs is about 5 percent but morbidity in high. There is weight loss in affected animals and abortions. Incubation period is 1-4 days but may be extended to 12 days. The disease is characterized by fever and vesicles in the mouth, snout, teats, udders and on the coronary band of feeL The primary lesions rupture and secondary lesions may develop.
Diagnosis: Since YE closely resembles, FMD, vesicular stomatis, a quick diagnosis is necessary. Animal inoculation test should be carried out to distinguish from other disease with vesicular lesions. Virus isolation form vesiculaP fluid can be made and virus identified by VN and CF tests.
Control: The recovered animals are immune .to the same serotype for at least 6 months. Vaccination is not attempted.
Feline Calcivirus (FCV)
Feline calcivirus is associated with respiratory illness of cats. The virus is moderately stable at pH 5 and is inactivated at 50°C in 30 minutes. There are numerous strains with serologic differences by VN test. All the strains are regarded as serologic variants of one serotype. The virus multiples in feline cell cultures and produce CPE. The incubation period in cat is 2-3 days followed by epithelial necrosis with vesicles or ulceration of external nares and of oral and lingual mucosa. Virulent strains may produce pneumonia. The clinical signs are fever, nasal and ocular discharge, dysponea, sneezing and rales. The most virulent form causes 30% mortality among young animals. The recovered cats develop long lasting immunity but few cats become

Calciviridae 233
oropharyngeal carriers and excrete virus for a long time. Spread of the virus is rapid by aerosal and saliva. For diagnosis the virus is isolated in cell culture and identified by VN test.
Control: Live and inactivated vacines are commercially available. The vaccination may be done by intranasal route in 9-12 week old
animals.
Rererences
GILLESPIE, I.H. and Scorr, F.W., 1973. Feline virus infections. Advances in
Veterinary Science and comparative Medicine. 17,163-224.

Chapter 21
Togaviridae
The viruses of this family are small enveloped viruses. Togaviruses owe their name to the envelope (latin-Toga cloak). Some members of the family are transmitted by arthropods. The non-arthropodbome agents show a predilection for recticulo-endothelial system, cause arteritis and _pass through placenta. The togaviridae family have four genera: (i) Alphavirus, (ii) Rubivirus, (iii) Pestivirus and (iv) Arterivirus. The flaviviruses have been assigned an independent family status because of the differences in properties as given below:
Table 21.1 DIFFERENCES IN PROPERTIES
Togaviridae
1. Spherical virion, enveloped, 60-70 run in diameter.
2. lcosahedral capsid, 28-35 run. 3. Two envelope glycoproteins
contain epitopes for neutralizing antibodies and alphavirus serogroup and subgroup specificity, one capsid protein.
4. Linear (+) sense ss RNA genome, 5' end capped, 3' end polyadenylated, genes for nonstructural proteins located at 5' end of genome.
5. Cytoplasmic replication, budding from plasma membrane.
Flavivi,idae
Spherical virion, envelopd, 40-50 nm in diameter. Probable icosahedral 25-30 run. One envelope glycoprotein contains epitopes for neutralizing antibodies one nonglycosylated envelope protein, one capsid protein. Linear (+) sense ss RNA genome, 5' end capped, 3' end not poly adenylated, genes for structural proteins located at 5' end of genome. Cytoplasmic replication, maturation within cytoplasmic vesicles.

To gaviridae 235
The alphavirus genus has 25 members. The pestivirus has mucosal disease/bovine virus diarrhoea and a closely related border disease virus of sheep and hog cholera virus. The Rubivirus and Arterivirus genera has one virus each, rubella virus and equine arteritis virus respectively.
The virus particles are small enveloped, RNA viruses measuring 40-74 nm in diameter. The nucleocapsid measuring about 25-37 nm in diameter is tightly surrounded by envelope. The nucleocapsid in alphaviruses is icosahedral in symmetry with 32 capsomeres. The envelope has spikes with HA activity. The nucleic acid is a single molecule of positive sense, single stranded RNA with a molecular weight of about 4 x 1()6. The virions contains 3 or 4 polypeptides, one or more of which are glycosylated. The viruses are ether resistant :md show variable resistance to trypsin. Th€? viruses replicate in the cytoplasm and mature by budding of either preassembled or assembling nucleocapsids through the plasma membrane. During replication of aJphaviruses and rubiviruses a sUbgenomic 26SRNA is synthesised which contains information for the virion structural proteins. Arteriviruses produce five sUbgcnomic mRNA as a"nested set.
With recent data on genomic organisation and transcription strategies, it appears that pcstiviruses are more related to flaviviruses than to togaviruses. The coronavirus-like transcription strategy of arteriviruses haS a reason to eliminate from their present taxonomic cluster.
Alphaviruses agglutinate chicken and goose rbe. The alphaviruses are capable of infecting wide range of species. The natural cycle may be in non mammalian hosts while mammals are accidental or dead end hosts. These viruses show a visceral localisation which is not fatal except in the pregnant animals or neonates. These viruses initiate a secondary encepahlitis which is often fatal. Pastiviruses replicate in alimentary tract epithelium and show a predilection for reticuloendothelial system. These viruses pass through the placenta and infect the foetus and produce infertility, abortion, foetal abnormalities.
ALPHA VIRUSES
Equine Encephalomyelits Virus
There are three antigenically distinct viruses which cause encephalitis in horses, other vertebrate animals, birds and man. Eastern

236 Textbook o/Veterinary Virology
Table 21.2 FAMILY: TOOAVIRlDAE
Gerws Virus Disease
i. Alphavirus (Mosquito Eastern equine ence- Encephalitis Horse transmitted) phalitis virus (Man) Encephalitis
Western equine Encephalitis encephalitis virus Horse (Man) Venezuelan equine Encephalitis encephalitis virus and Febrile disease
Horse (human) Getab virus Febrile disease Horse
ii. Pestivirus (non Bovine virus Generalised infection arthropod borne) diarrhoea virus cattle.
Hogcholera virus Generalised infection pig.
iiL Arterivirus (non Equine arteritis virus Generalised infection arthropod borne) and abortion Horse
iv. Rubivirus (non Rubella virus Generalised infection arthropod borne) Man
equine encephalomyelitis (EEEV) and Western equine encephalomyelitis (WEE V) virus were fIrst found in Eastern and Western States of USA. The third virus Venezuelan encephalitis virus (VEEV) was found in Venezuela. These viruses cause encephalitis in horses in America. The natural hosts are equlnes, man, mosquitoes and reservoir hosts like birds, small rodents, leopard frog etc. The disease is known t.o occur for many years in the USA, Southern Canada and South America. In India the sporadic cases characterized by incoordination of movement and paraplegia, have been frequently reported. The encephalomyelitis cases have been suspected to be due to a virus. There is no report of virus isolation from India.
Properties 0/ the virus: The morphology is typical of the family. The virion measures 20-30 nm in diameter and contains a single stranded RNA. They haemagglutinate chicken rbe in a narrow range of pH 6.2 to 6.4. The three distinct serotypes occur with geographical restriction. SUbtypes of EEEV and WEEV have not been identified but minor serologic differences among their strains have been noticed. Four major subtypes have been reported in VEEV. A few antigenically

Togaviridae 237
distinct strains of VEEV subtype 1 are assoicated with equine epizootics, whereas majority of viral subtypes are associated in wild rodents and do not cause disease in horses. These viruses grow well and produce cytopathic effects in tissue culture derived from chick embryo, mouse hamster, monkey, guinea pigs and other species. The virus can also be propagated in embryonating chicken eggs causing haemorrhages and deaths of embryos in 24-48 hours. Equine epizootic strains make small discrete plaques in Vero cells and· cause haemagglutination at pH6, indicative of virulence for equine species. The non epizootic strains us~ally produce large plaques with less distinct border and exhibit HA over a broad pH range. The VN test differentiates, EEEV, WEEV and VEEV and identifies the strains. The CF test detects common antigen shared by all viruses. The HI test also differentiates the 3 viruses.
Epidemiology: The EEEV and VEEV spread naturally in birds and small rodents and produce harmless and symptomless infections. The virus is transmitted chiefly by culicine mosquitoes in which the virus propagates and persists throughout life. The horses are accidental dead end hosts. Contact transmission in horses does not take place. Young cattle and pigs can be naturally infected.
Pathogenicity: The clinical features of WEEV and EEEV are indistinguishable. The affected horses show fever, sommolence, paralysis of lips and pharynx and incoordination of movement. Death occurs in 3-8 days after onset of clinical signs. In VEEV, a fatal fulminating form of disease is seen in which generalized, acute febrile disease predominates. The disease is diphasic. In the frrst stage the virus multiplies in viscera giving rise to viraemia between 2-5 days of illness. In the second phase, virus multiplies in eNS and causes nerve cell degeneration in many parts of brain and spinal cord. The affected areas show mononuclear infiltration and perivascular cuffing. In the Eastern and Western form, the main lesions are found in the brains stem while in VEEE the virus is viscerotropic and gives rise to necrotic foci in spleen and lymph nodes. There are no characteristic gross changes in the organs.
Diagnosis: The diagnosis is arrived at by virus isolation or by serological methods. Detection of rise of antibody titre in animals that survive long is carried out by VN test in mice, egg embryos and cell -culture. The HI test is simple and reliable diagnostic procedure. The HI

238 Textbook o/Veterinary Virology
test recognises group rather than type characters. In EEEV and WEEV brain tissue is the best source of material for virus isolation while in VEEV the blood plasma or serum of febrile animals contains the high concentration of virus and is suitable for virus isolation. Intracerebral inoculation of mice or wet chicks produces viral encephalitis. Cell cultures or chicken embryos are suitable for virus isolation.
Control: Formalin inactivated bivalent or trivalent vaccine are used for immunization. Minimizing contact with vector is important either by spraying or mosquito proof accommodation.
PESTIVIRUS
Bovine viral diarrhoea viruslMucosal disease virus (BVDVIMDV)
Bovine viral diarrhoea is caused by bovine virus diarrhoea virus(BVDV), a Ubiquitous, easily transmitted virus. Cattle are naturally infected but outbreaks have been reported in deer. The disease was fIrst recognised in USA in 1946 and the virus was isolated in 1957. The virus is prevalent worldwide in cattle population. The presence of BVD has long been suspected in this country. Pande & Murty(1960) recognised mucosal disease like syndrome among young cattle and buffaloes. The virus isolation has not been reponed so far from this country but there is serological evidence of presence of this disease. Based on serological studies Nyak(1982) reported abortions in cows at Orissa due to BVDV.
Properties: The morphology is typical of the group but variable sizes are recorded. The virus particles are sensitive to low pH, ether, chloroform and other lipid solvents. The virus is readily inactivated at 56°C. Few serotypes of the virus haemagglutinate rhesus monkey, pig, sheep and chicken red blood cells. Variation in antigenicity and cross protection occur. There are virulent and avirulent strains of the virus. The neutralization and immunofluorescence tests suggest that the non pathogenic strains are more closely related to one another than the cytopathic strains. There is a close antigenic relationship with Border disease and swine fever viruses. The virus grows in cell cultures of bovine and ovine origin. Several strains do not produce CPE and are diffIcult 10 detect by an interference test using cytopathic strains or by FA technique, as well as by enhancement of CPE of Newcastle disease virus in bovine testicular cell cult'lfes. There are reports that certain serotypes can grow in yolk sac of chicken embryo.

Togaviridae 239
Epidemiology: The host range of BVDV is restricted to domestic and wild ruminants and pigs. These species can be infected in the field as judged from the detection of antibodies specific for B VD as reported by many workers. No reservoir host exist The cattle are persistently infected. Vectors among invertebrate hosts have not been reported so far. The BVDV has a global distribution but from China there is absence of neutralising antibodies in the serum of cattle population against this virus. The spread of virus cannot be properly determined, in most cases it occurs unnoticed. Over 90% of infections are inapparent. The virus may spread from infected to susceptible animals by direct or indirect contact. The calves are infected in utero by transplacental virus transmission.
Pathogenesis: The disease principally affects cattle but also has been reported in deer. A variety of clinical disease pattern is attributed of BVDV. The disease may be acute, mild or chronic. The most common form of BVD is subclinical probably due to widespread SN antibodies to the virus. The bovine viral diarrhoea syndrome is seen predominantly in 6-18 months old cattle as a primary infection, which is via oropharyngeal route. The primary replication of the virus takes place in the oropharynx followed by rapid-uptake of virus in the draining lymph nodes. Viraemia leads to infection of lymphocytes giving rise to leucopenia and spread to other lymphoid tissues particulalry Peyer's patches. There is concurrent replication in the epithelium of alimentary tract which results in discrete erosions and induces diarrhoea. Lesions in the nasal cavity and conjunctiva may also appear. Concurrent respiratory disease occurs.
The majority of animals recover since disease has high morbidity and low mortality. The primary infection of pregnant cows results in transplacental infection and symptoms appear according to the stage of pregnancy. In early pregnancy there may be infertility and embryonic death. In mid pregnancy congenital anomalies like cerebraller hypolasia or abortion may result in congenital infection of apparently normal calves. These animals frequently excrete virus and are source of infection and may also develop mucosal disease. The mucosal disease seems to occur in congenitally infected animal which are immunotolerent and harbour the virus without showing symptoms. The mucosal disease is usually sporadic, progressive and fatal. When these animals get infected with the second strain .the animals develop

240 TexJbook o/Veterinary Virology
extensive erosions throughout the alimentary tract. The outcome may be fatal or chronic progressive disease develops with animals remaining antibody negative. Lesions similar to those produced by rinderpest virus primarily occur in the alimentary tract and lymphatic system. The lesions may also be present in the upper respiratory tract and consist of congestion, haemorrahge, oedema and erosions. The oral lesions are small, shallow and irregular and may be found on the muzzle, tongue, dental pad, oesophagus and pharynx. In rumen, haemorrhages and erosions are often seen while the abomasum is inflammed and oedematous. Oedema, congestion and erosions are seen in the intestin~. There is damage to immunocompetent cells. Lymphnodes and Peyer's patches are enlarged and may be devoid of lymp~oid tissues. In congenitally defective calves, there may be cerebellar hyperpIesia, cataract, retinal degeneration etc.
MVDV usually induces high SN antibody titres that persist for long time. Due to immunological tolerance or due to destruction of immunocompetent cells the virus persists with circulating antibodies. The fatally infected cattle frequently do not produce detectable antibodies against BVDV. It is persumed that it is due to immunologic tolerance as a result of antigenic experience at or before birth. The cause of fatal BVDV infections and the role of immunologic tolerance remained obscure.
Immune reaction: BVDV usually produces life long immunity. The presence of neutralizing antibody in ~he serum reflects immunity. The disease picture of BVD is of immune pathogenetic phenomenon, especially of immunologic tolerence in persistently infected animals to a particular variant of virus. In calves failure to develop antibodies has been attributed to immune tolerance, immune paralysis or immune' suppression.
Diagnosis: It is difficult to diagnose the disease clinically. Paired serum samples indicate increase in neutralizing antibody. A definitive diagnosis is made by virus isolation and identification. Virus isolation can be attempted from nasal secretions, faeces, blood, lymphnodes and intestines. The fluorescent antibody test is very useful in detecting the virus. Agar gel diffusion test using tissues form infected animals is also useful.
Control: Recovery from natural infection confers a desirable immunity. Inactivated and modified live virus vaccines are available in

Togaviridae 241
many countries. The inactivated vaccines are safe for use in pregnant animals. The attenuated vaccine either alone or combined with IBR and PI) provide satisfactory immunity. Colostral antibodies appear in calves and last for 4-6 months.
Border Disease Virus
Border disease or hairy shaker disease is a congenital condition of lambs characterized by excessive hairiness of birth coat, poor growth and nervous abnormalities. The disease has been reported from U.K. Australia and Newzealand.
Properties of the virus: The virus of Border disease (BDV) is morphologically similar to BVDV and HCV and has a size of 45-54 nm with a core of 30 nm. The physical properties are also similar to other members of pestiviruses. The virus have both cytopathic and non cytopathic variants and have been grown in cells or foetal lamb kidney, procine kidney, sheep choroid plexus and bovine testis where it produces CPE. The virus of BDV is inactivated at 56°C for 30 min; lipid solvents, U.V light, desicalion and common disicfectants.
There are strain differences with BDV, in terms of pathogenicity for different breeds of sheep. Moreover, there is varying ability of BDV antibodies to neutralize or protect against different strains of virus both in vitro and in vivo indicating appreciable antigenic differences.
Epidemiology: It is a disease of sheep, although goats, cattle and swine can be experimentally infected. It has been shown experimentally that contact infection of BDV from sheep to cattle and of BVDV from cattle to sheep readily occurs. There is serological evidence of BDV infection in several species of wild deer, the wild ungulates may serve as a reservoir and source of infection for sheep.
The infection is acquired by inhalation or ingestion. The infection in adults is usually subclinical and short lived. In pregnant ewes, the foetus also is infected. The infected foetus may be expelled or born alive at full term .• either immune or as persistently infected animals. The virus is present in the aborted foetus and its membranes and in the secretions and excretions of persistently infected animals. Outbreaks of BD characterized by birth of 'hairy shaker' lambs, clinically- vary greatly in severity.
Pathogenesis: In nonpregnant sheep, infection with BDV is subclinical, although pyrexia and transient leukopenia has been

242 Textbook o/Veterinary Virology
encountered 6-11 days pI. In pregnant animals the virus crosses the placenta. The effect on foetus depend upon the stage of pregnancy.
The virus replicates in the lymphoid tissues of the oropharynx followed ,by viraemia. In adults no clinical signs are exhibited. The virus passes the placenta and cause abortion associated focal necrotising placentitis. Infection may result in foetal developmental abnormalities which are: abnormal skin follicle development giving rise to hair rather than wool in the coat, the lambs appear hairy; hypomyelinogensis-infection of myelin forming cells leading to lack of myelin, resulting in muscle tremors and incoordinate gait which is described as 'Shakers'. There is poor growth rate of foetuses and hydrocephalus occurs in foetuses. Congenitally infected immunotolerant foetuses excrete virus but lack antibodies. Most clinically abnormal animal die before weaning, but in some animals locomotor disorders progressively improve with age. The brain and spinal cord show varying degree of deficiency of myelin and hypercellularity of white matter with many glial cells of abnormal appearance.
Diagnosis: The best material for diagnosis is the newborn affected lamb, alive and unsuckled along with blood samples from the mother and from other animals of the affected flock. If examination of cryostat sections, neuropathology and serology is carried out on the suspected material.
The clinical symptoms, particularly the hairy coat and locomotor abnormalities, are suggestive of the disease. Virus can be isolated from infected foetuses and nasal swabs of excretors in cell culture. Neutralizing antibodies in ewes can be detected. Immunofluorescence, virus neutralization, Cfl, and immunodiffusion tests are used to detect the presence of antibodies.
Control: No commercial vaccines are currently available. The disease may be controlled by eliminating the infected progeny, although symptomless excretors are difficult to detect. The excretion can be detected from seronegative animals.
Swine Fever or Hog Cholera
Swine fever is a highly infectious virus disease of pigs which on occasion can cause high mortality. The natural disease occurs in swine only. The virus is world wide in distribution. It was described for the

Togaviridae 243
flfSt time at Ohio(USA) in 1833. The disease has been eradicated from Australia, Denmark, Newzealand, UK and USA. In this country the disease was recorded for the ftrst time in Punjab in 1961 and subseqently form Maharastra and UP in 1962. During the past few years the outbreaks of swine fever have been recorded from the states ofNagaland, Manipur, Tripura, Meghalaya etc. However, the disease is also occasionally reported from other parts of the country.
Properties: The virus particles are 40-60 nm in diameter. The envelope is easily distrupted to reveal nucleocapsid which has icosahedral symmetry. The virus contains three structural proteins, two of them being envelope glycoproteins and a third non-glycosylated protein. It is a stable virus under laboratory conditions but is inactivated when exposed in the field. It persists for a long time in pork or garbage. The virus is resistant to most of the ordinary disinfectants. Strains of varying virulence and antigenicity have been reported to cause vaccine break down but there appears to be only one serotype. It is antigenically related to bovine virus diarrhoea.
The virus replicates in primary pig kidney, spleen, bone marrow, testicle. lymph node, leucocytes and in PK-15 cell lines. Most of the strains produce little or no CPE. These strains are dctectcd by FA test and by exaltation of virulence of Newcastle disease virus (NDV) method. Mild noncytopathic strains enhance CPE of Newcastle disease virus in pig testicular cells. The virus has been adapted in chicken and duck embryos.
Epidemiology: The virus is found in saliva, blood, nasal and conjunctival secretions, faeces and urine. Transmission occurs by droplet. fomites and infective swill. The virus. survives for several months in fomites at 20°C. Marketing of pigs is a common source of dissemination. The congenitally infected excreting animal is important particularly as carrier sows. It has been mooted that flying insects and birds may act as vectors.
Pathogenesis: Domestic and wild pigs only are susceptible to swine fever. The virus enters the body via oropharyngeal route and primary multiplication takes place in lymphocytes and macrophages of tonsils. The virus has a predilection for RE system and alimentary epithelium. The incubation period of the natural disease is 3-8 days. The morbidity is 95-100% and the mortality is as high. The disease may be acute or chronic. In acute cases the mortality i~ 100%. The

244 Textbook o/Veterinary Virology
affected pigs have high temperature(41.0°C) and are lethergic. This is followed by hyperaemia of skin, particularly of ears, conjunctivitis nasal discharge and vomiting. Sometimes convulsion may be observed. In peracute cases, death takes place as early as 5 days after exposure to virus, while in acute cases death takes place between 10-20 days. In chronic cases the death takes place after 30 days of exposure. The pigs have persistent viraemia and have an imparied antibody response. Transplacental transmission follows a mild infection is which no clinical symptoms are noticed. These mild infections are due to virus strains of low virulence. The virus produces embryonic death, malformation, mummification. still birth. or perinatal death. Life long persistent infections acquired in uterus have been reported.
The replication in lymphoid cells produces lymphadenitis and leucopenia. Peyers patches are extensively affected. In the caecum healing ulcers, sometimes associated with concurrent bacterial infection over lymphoid foci are described as button ulcers. Tertiary dissemination to eNS results in a widespread encephalomyelitis and symptoms of tremors, incordination and paralysis are observed. Gross changes are usually absent in peracute cases. Secondary pneumonia and enteritis may accompany the primary lesions. There are petechial and echymotic haemorrhages on all serous surfaces. There is haemorrhagic lymphadenitis, petechial haemorrhages in kidney, infection in the spleen and ulcers in the colon. The histotological changes are hydropic degeneration and proliferation of endothelial cells which occlude blood vessels.
Diagnosis: The clinical symptoms and post-mortem examination is helpful. Viral antigen can be detected by gel diffusion test using pancreas as a source of antigen or by fluorescent antibody test on the cryostat sections of spleen, lymph nodes and tonsils. The virus isolation can be made from spleen. tonsils lymph nodes and blood in cell culture.
Control: 111ree methods of vaccinations are used. The killed vaccines are safe but are not very effective. The crystal violet vaccine was used in Europe before 1960. Simultaneous vaccination with virulent virus and antiserum is still used in certain countries. Modified live vaccines made by serial passages in rabbits, swine or tissue cultures are used for immunization. The live vaccines are not safe because transplacental infection can occur and the modified virus may spread in the herd, thus complicating diagnosis of the disease. The live attenuated vaccines are recommended in those herds where incidence of the disease is high, strict quarantine and enforcement of garbage

Togaviridae 245
cooking should be enforced. Heterotypic vaccines using BVDV do not provide complete protection against swine fever.
ARTERlVIRUS
Equine Arteritis Virus (EA V)
It is an acute infectious disease of horses characterised by fever, depression, rhinitis and oedema of conjunctiva, trunk, limbs and external genitalia. The disease occurs mainly in USA and Northern Europe. Neutralizing antibodies against EA V has been reported from India recently but requires further confIrmation.
Properties: The virus measures 50-70 nm in diameter. It is sensitive to ether, heat, and low pH. The virus does not grow in embryonating chicken eggs. The virus can be propagated in equine, rabbit and baby hamster kidney cells. The isolates require repeated passage before producing CPE in equine monolayers.
Epulemio!ogy: The virus is transmitted through respiratory tract particularly in foals. The virus persists for a long time in the kidneys of infected horses and disease may spread by the infectious virus in urine. The carrier horses are important source of spread.
Pathogenicity: The oral or aerosal infection results in replication in the lymphoid tissue of nasopharynx and then viraemia occurs. Following incubation period of about 5 days there is pyrexia, anorexia and leucopenia. The predilection sile is medium sized blood vessels; virus replicates in the endothelium of blood vessels resulting in necrotic lesions in the muscularis of blood vessels. This leads to haemorrhages, oedema and sometimes thrombosis in the muscularis of blood vessles. There is conjunctivitis and palpebral oedema which give the name 'Pink eye'. Oedema may also appear in the leg and lower abdomen. The infection of respiratory and alimentary tracts leads to nasal dis_charge, coughing, dyspenea, diarrhoea and colic. In about 50% of pregnant mares, abortions takes place after 10-30 days of infection. Congenital infection can also occur and some animals seem to be consitently infected. The gross lesions are oedema, congestion and haemorrhages of the subcutaneus tissue, lymph nodes, and viscera of peritoneal and pleural cavities. There is degeneration and necrosis of arteries.
Diagnosis: The tentative diagnosis is made from the clinical signs and characteristic lesions in the arteries. The CF test is useful in diagnosing the recent infection. Virus isolation from fIeld cases is often diffIcult. The specimens taken from conjunctival sac and nostril of

246 Textbook o/Veterinary Virology
infected horses and blood, liver and spleen of aborted foetuses is used for virus isolation in cell culture of equine origin.
Control: An attenuated live virus vaccine is available which is protective against abortion but it should 110t be given to mares in late pregnancy.
References
BUXTON, A. and FRASER, G., 1977. Animal Microbiolgoy, Vo1.2, Oxford, Blackwell Scientific Publications.
BARLow, R.M., 1982. Epidemiology. In: R.M. Barlow and D.S.P. Patterson (Editors), Border Disease of sheep: A virus induced Terqtogenic Disorder. Paul, Parey, Berlin Hamburg pp. 70-71.
DUNNE, H.W., 1975. HOG CHOLERA. In Diseases of Swine, 4th Ed. Edited by H.W. Dunn and A.D. Leman, Ames, Iowa. Iowa State University fress.
KAIIRS, R.F., 1985. Viral diseases of cattle, Kalyani Publishers Ludhiana.
MALMQUIST, W.A., 1968. Bovine viral diarrhoea-mucosal diseases: Etiology, pathogenesis and applied immunity. Journal Veterinary Medical Association 152: 763-68.
MOIlANTY, S.B. and Durrs, S.K., 1981. Veterinary Virology. Lea and Febiger, Philadelphia.
MURTHY, D.K. and AoLAKHA, S.C., 1962. Preliminary studies on outbreaks of swine fever. Indian Veterinary Journal. 39: 406-19.
NAYAK, B.D; PANDA, S.N.; MISRA, D.B.; KAR, B.C. and DAS, B.C., 1982. Note on serological evidence of viral abortion in cattle in Orissa. Indian Journal Animal Sciences, 52 102-103.
PANDE, P.G. and MURTY, D.K., 1960. Incidence and pathology of some recently recognised disease-like syndromes amongst cattle and buffaloes in India. 9th Animal Disease conference Bhubneswar, Orissa.
RussELL, P.H. and EOINGTON, 1985. Veterinary viruses. The Burlington Press (Cambridge) Foxten, Cambridge.
S'jCWARO, W.C., CARBREY, E.A and KRESSE, 1.1., 1972. Transplacental hog cholera in immune sows. American Journal of Veterinary Research, 33,791-98.
UPPAL, P.K., SiNGH, B.K and YAOAV, M.P., 1990. An outbreak of abortion in mares due to equine herpes virus with possible concurrent infection of equine arteri/is virus. Virus information Exchange letter. 7, 95.

Chapter 22
Flaviviridae
Flaviviridae is a recently established family which has only one genus-Flaviviruses.
The virions arc enveloped measuring about 40-50 nm in diameter and consist of core particle surrounded by a projection bearing membrane. The virus particle contains a single molecule of single stranded RNA with a moleular weight of 4-4.6 x 1()6. The structural polypeplides conslilue a nucleocapsid protein and a small additional polypeptide as well as one glycosylated envelope protein. Flaviviruses replicate in the cytoplasm and maturation occurs by budding through intracytoplasmic membranes. Subgenomic mRNAs do not occur and posttranslational processing of one large precursor molecule is the mechanism involved.
Yellow fever virus is the type species of this family. The flaviviruses of veterinary importance are: the louping ill virus and WesselsbOrn disease virus, Encephalitis viruses and Dengue viruses.
Genus
Flavivirus
Table 22.1 FAMILY: FLAVIVijUDAE
Virus
Louping ill virus (tick borne) Wesselsbom disease virus (Mosquito borne) Japanese encephalitis virus (Mosquito borne)
Disease
Encephalitis-sheep
Generalised infection, abortion sheep
Encephalitis, abortion pig

248 Textbook o/Veterinary Virology
Japanese B Encephalitis Virus
Japanese B encephalitis virus produces an acute arthropod borne disease among animals and human being. Japanese encephalitisOE) is widespread throughout Asia but does not exist in other parts of the world. Th~re is serological evidence of existence of JE in animals in India. In a serological survey conducted by Mall and Natarajan (1981) in India, the highest percentage of positive sera was reported in horses (66.17%), followed by pigs(62.5%), buffaloes (13.35%) and dogs (9.0%). Cattle, sheep, goat and mule sera were negative. Horses, pigs, man and cattle are susceptible.
Properties of the. virus: The virus is typical of the family. It contains a large amount of lipid and is sensitive to ether and deoxycholate. It is inactivated at 56°C in 30 min. The virus haemagglutinates chicken, pigeon and goose rbc. The virus has a wide natural host range and can infect a variety of laboratory animals like monkeys, hamsters, guinea pigs, rabbits, chickens etc. Suckling mice are highly susceptible to intracerebral inoculation, which produces encephalitis and death. The virus readily grows and produces CPE in various cell cultures. JEV also grows in chicken embryos by yolk sac route of inoculation and causes death of embryos. There is some antigenic cross reaction with other flaviviruses. Some antigenic modifications have been reported. The infected animals produce SN, CF and HI antibodies. The SN and HI antibodies appear in 1-2 weeks and persist for several years while CF antibodies appear later and disappear sooner.
Epidemiology: Japanese B encephalitis virus is probably a primary parasite of wild birds. Night herons is probably a primary parasite of wild birds. Night herons is the principal reservoir host. Pigs act as amplifiers of the infection in nature. Mosquitoes are the natural vectors and the infection maintained mosquito-heron cycle or mosquite-pig cycle. Humans and horses are victoms and appear to be dead-end hosts.
Pathogenecity: The incubation period is 8-10 days. In mild infections in horses, clinical signs are fever, slow movement and sometime jaundice. In several cases in horses the course of infection resembles that of WEE, with facial paralysis, encephalitis following an initial febrile phase. Mortality ranges from 40-70%. The virus is less pathogenic for pigs; most of the pigs recover after viraemia while a very low percentage show encephalitis. Stillbirths in pregnant sows

Flaviviridae 249
occur in high percentage of cases. Persons working with infected horses or pigs may develop fatal encephalitis. Pathologic lesions in the animals are restricted to haemorrhage, oedema and congestion. In the brain, lesions of encephalitis are seen in pigs.
Diagnosis: Tentative diagnosis can be made on the clinical symptoms and histopathological lesions. The- confrrmatory diagnosis can be arrived at by isolation of the virus from infected brain and by serological tests like, CF, HI and VN with the paired serum samples.
Control: Formalised virus grown in chick or mouse brain is effective as vaccine in horses. Contact of animals with mosquitoes should be minimized.
Wesselsborn Virus
A mild but sometimes acute febrile disease giving rise to abortions among sheep and death of newborn lambs and pregnant ewes is enzootic in certain areas of South Africa and Zimbabwe. Humans, adult sheep and other ruminants get inappar~nt infection. The transmission of the virus takes place by culicines. The virus can be readily propagated in chicken embryos by yolk sac route and grows in cell cultures of lambs kidney. Un weaned mice are highly susceptible to intraperitoneal, intramuscular and intracerebral route of inoculation. The adult mice is susceptible to intrflcerebral route only. A formalized vaccine is used to control the disease.
Louping III Airus
The ,;:irus causes ovine encephalomyelitis, a tick borne disease in sheep and cattle. Occasionally the disease has been reported in dogs. Human infection has been reported' among laboratory workers, veterinarians and shepherds. There is evidence that 32 vertebmte species can be infected. Most infections are accompanied by negligible clinical response. The disease is prevalent in Scotland, England and other parts of Europe. Antigenic variants of this' virus with similar epidemiology are: Central Europea'n encephalitis; Russian spring summer encephalitis and Far Eastern Russian encephalitis.
Properties: The virus has a typical flavivirus morphl.!~gy. In glycerine-saline the virus survives at room temperature for 3 months. The virus is inactivated at 60°C in 5 minutes and by 2% phenol and 1% formalin. The virus haem agglutinates chicken (newly hatched) and goose rbe. Th(1 virus can l;>e readily propagated in chicken embryo by

250 Textbook o/Veterinary Virology
CAM or yolk sac route of inoculation. On CAM the virus produces discrete pock marks. The virus can grow in cell culture of various mammalian and avian species and produce cytopathic effec~
Pathogenecity: Two distinct phases follow in sheep, the first phase is charaGterized by fever upto 42°C and viraemia. The animal returns to nonnal and may recover. After about 5 days a second phase of pyrexia may occur associated with incoordination and ataxia followed by progressive paralysis and death. The incubation period is 7 to 18 days. Natural infection can occur in various domestic animals. Gross lesions are not seen in infected sheep. Microscopic lesions are around the Purkinje cells of the cerebellum in the motor nuclei of the brain stem and ventral horns of spinal cord. In man influenza like symptoms occur.
Epidemiology: Tick activity is optimal from April to June and in September, so is the disease. The transmission occurs through tick bite. Transoviral spread of virus occurs. Animal to animal transmission is rare but laboratory workers are infected via respiratory or conjunctival routes ..
Diagnosis: The diagnosis can be confirmed by isolation of virus from the brain and spinal cord in cell cultures or by intra-cerebral inoculation in mice. HI and VN tests are used to detect presence of antibodies and rise of antibody titre in the sera samples of sheep.
Control: Formalized sheep spleen or brain tissue was formerly used which has now been replaced by sheep kidney tissue culture methanol inactivated vaccine. Regular dipping of sheep should be carried out to control the vector.
References
BARNARD, B.J.H., 1990. Virus infections of ruminants In: Z. Dinter and B. Morein (Editors), Wessels-bron disease Virus. Elsvier Science Publishers B.V.pp. 291-294.
MALL, M.P., and NATARAJAN, C., 1981. Paper presented at XV annual convention of Indian Allergy and applied immunology, Indian Veterinary Research Institute, Izatnagar, Nov. 9-13.
REID, H.W., 1990. Virus infections of ruminants. In: Z. Dinter and B. Morein (Editors) Louping ill virus. Elsvier Science Publishers B.V. pp. 279-289.

Chapter 23
Reoviridae
Reo is an acronym derived from 'respiratory enteric orphan'. Reoviridae was once part of picomaviridae but were reclassified on the basis of being larger in size and less cytopathic. The family comprises of the following genera:
Reovirus - Respirarory enteric Orphan viruses Orbiviruses - Orbis means ring shaped appearence Rotavirus - Rota means wheel like arrangement of capsomeres PhYloreovirus - Plant viruses Fijivirus - Plant viruses Only the first three genera are of Veterinary importance. The viruses of this family are spherical particles with diameter of
60-80 nm with icosahedral symmetry and double shelled capsid. The outer shell is digested by chymotrypsin. The viruses do not possess the envelope. The genome of rotaviridae family is segmented and consists of ten (Reovirus, Orbivirus) or eleven (Rotavirus) fragments. The nucleic acid molecules arc linear and double stranded with a mol. wt. between 0.7 and 7 x 106• There are 6-10 polypeptides in the virions including RNA dependent RNA polymerase. The removal of outer shell by proteolytic enzymes is required for activation of RNA dependent RNA polymerase. Replication of reoviruses occurs in the cytoplasm. The infecting virion genome of rotavirus is not completely uncoated. Virion transcriptase synthesize first positive strands and replicase enzyme makes the negative strands, thereby forming the double stranded progeny RNA and excess viral antigens accumulate in larger masses in the cytoplasm, forming the characteristic inclusions.

252 Textbook o/Veterinary Virology
The three genera can be distinguished morphologically. The characteristic of orbiviruses being ring shaped capsomeres of the inner capsid and rotaviruses a wheel like capsid appearance with a wide hub, short spokes and well defined rim. The outer capsid layer of rotaviruses is easily l.ost, which results in incomplete virus particles of 58-62 nm in size.
REOVIRUS
Three mammalian serotypes have been recognised. The viruses of animal origin are indistinguishable form those of human origin. Antibodies against reoviruses have been found in cattle and other animals in many countries. This indicates that animal reoviruses like those in man are ubiquitous in distribution. The rcoviruses have been isolated from respimtory and intestinal tmcts of cattle and are antigenically similar to human serotypes.
Bovine Reoviruses (type-I-type 3)
Antibodies to all the 3 bovine reoviruses have been found in cattle serum. In India reoviruses have been isolated from call1e. Naturally occuring infection in cattle was first reported in 1960. It has been postulated that rcoviruses of cattle are less important in etiology of respiratory disease than the number of other viruses that have been implicated as primary pathogens. In the case of alimentary tract, their role as pathogens is even less defined.
Reoviruses have also been recovered from sheep showing signs of concurrent signs of respiratory and alimentary tract disease but the evidence of pathogenic role is limited. It appears that behaviour of reoviruses as pathogens of sheep and cattle depends upon the secondary agents which precipitate the disease.
Virus properties: When the virus is heated at 55°C in the presence of magnesium ions the virus is not inactivated but its infectivity is enhanced. The infectivity is also enhapced when the virus is treated with the proteolytic enzymes. The virus resists the effect of 1 % phenol for 1 hour but is inactivated by 70% ethanol or 3% formalin at 56°C. It is stable between pH 2.2-8.0 and resists the action of ether and chloroform.
The mammalian reoviruses comprise of three serotypes. All the 3 serotypes occur in cattle, sheep and man and possess two group specific

Reoviridae 253
antigens in common. By immunoelectrophoretic analysis it has been found lhat lhere are 4 group specific and two type specific antigens.
The reoviruses agglutinate human'O' rbe. For virus isolation monkey kidney cells are superior for bovine strains in comparison to cells of bovine origin. Eosinophilic inclusions are produced in the cytoplasm of infected cells.
Epidemiology: The host range of three mammalian serotypes is very wide. They are found in most domesticated and wild animals. The reoviruses have also been isolated from culicine mosquitoes and may act as live vectors or merely mechanical carriers. The bovine reoviruses have been isolated from North America, Europe, Africa and Far East including India. The viruscs spread easily in callle and sheep population. The infection is mild.
Pathogenesis: The rcoviruses initiate infection in the rcspiratory and alimentary tract of both callle and sheep. In experimental infection the latent period is of less than 24 hours. This is followed by viraemia which lasts upto onc week. The virus can be recovcred from nasal discharges faeces and conjunctival sac up to 2 weeks. Most of the virus strains have tropism for respiratory tract, nasal turbinate mucosa, tonsils, trachea, lungs and mediastinal lymph nodcs. The virus may also be demonstrated in the spleen and kidney and throughout alimentary tract and mesentric lymphnodes. Chlamydia may act synergistically wilh reoviruses to induce pneumopathy lhan either agent alone. The clinical response to infection is more pronounced in sheep than in cattle. After an incubation period of 4-6 days the lambs show pyrexia nnd ocular and nasal discharge, there was evidence of pneumonia accompained by diarrhoea. In lambs only reovirus type-l is capablc of producing disease whereas type 2 and 3 are not. The experimentally infected calves with type 1 and 2 show rise of tcmparaturc only. In Natural outbreaks in sheep mild respiratory and enteric signs are noted.
Diagnosis: The diagnosis can be made by virus isolation in monkey kidney cells or pig or lamb kidney. Paired serum samples arc collected for demonstration of HI and SN antibodies.
Control: Limited number of attempts have been made to evaluate vaccines against reoviruses in cattle. Both inactivated and multifactoral vaccines have been tried but results are not encouraging.
The revorises have also been isolated from horses (type 2 and 3), pigs (type 1 and 3), dogs (type 1) and goaLS (1 and 3).

254 Textbook of Veterinary Virology
Avian Reovirus type 1 to 5
Numerous reoviruses have been isolated from chicken with arthritis (tenosynovitis), respiratory signs, diarrhoea retarded growth and healthy chickens. The presence of avian reovirus is world wide. In India' the virus has been isolated from arthritis cases and presence of precipitins to avian reovirus has been reported. The avian reoviruses share a group specific antigen. Five serotypes have been reported. Avian reoviruses differ antigenically from other mammalian reoviruses. They grow readily in yolk sac or CAM of chicken embryos. The chicken embryos die between 3-8 days of infection. Small white. discrete pocks are found on the CAM of chicken embryos. The virus also multiplies in the cell culture of chicken embryos and vero cells. In CK cells syncytia are produced. Avian reovirus type 1 is associated with viral arthritis in chicken. Avian reovirus type 1 is associated with viral arthritis in chicken. It is likely that other serotypes may be involved. The viral arthritis is common among 7 week old chicken. After an incubation period of 1-11 days, the chicks develop inapparent infection in most cases. In the acute cases the birds become lame and may also he stunted. Lamencss becomes more pronounced in chronic cases. The morbidity is 100% in broilcrs and mortality is about 1%. Growth ratar~ation is prominent sign. Gross lesions involve swelling of digital flexor or metatarsal extensor tendons. A small. amount of exudate is found in hock or elbow joint Myocarditis is a constant feature. A carrier state is known to exist. A definite diagnosis can only be made on isolation of virus. An attenuated vaccine has been developed for the control of the disease.
ROTAVIRUS
Rotaviruses is responsible for neonatal diarrhoea in man and farm animals. Virus specific antibodies are widespread in adult animals. In India there is a serological evidence of presence of rotavirus antibodies among cattle and there is a report of isolation of rotavirus from a calf.
Bovine Rotavirus
Properties: The complete virus particle measures about 70-75nm and consist of a central core surrounded by two layers of virus specific polypeptides. The genome consists of 11 double stranded RNA segments. All rotaviruses share a common antigen associated with the

Reoviridae 255
inner capsid layer. Rotaviruses of different animal species can be distinguished by VN tests. Gel electrophoresis of the genome segments have revealed differences in RNA patterns for rota viruses obtained from different species as well for different isolates from the same species. Inspite of large number of virus particles found in the faeces, rotaviruses are difficult to propagate in cell culture. Bovine rotaviruses have been adapted to grow in bovine and simian kidney cultures. Pretreatment of faecal samples with trypsin and pancreatin facilitate isolation and cultivation. The enzymes cleaves the outer capsid and this cleavage renders virus susceptible to uncoating wiLhin the cell. This uncoating is a necessary step in converting n'on infectious virus to infectious virus.
The bovine rotaviruses remain viable for six months in faecal material at room temperature. Sodium hypochlorite (3%), lysoI"(5%), formaline (10%) had little effect on lamb rotavirus in the intestinal contents. Lysol .(5%) and formaline were effective when intestinal contents were exposed for 2 hours.
Epidemiology: The rotavirus infections have been reported from many species of domestic and zoo animals including man. Experimentally human infant rotavirus caused diarrhoea in calves but serverity' of illness was less than caused by infection of bovine rotavirus.
There is no evidence under field conditions Lhat cross-species infection is important but experimentally cross-species infection has been reported.
The high antibody prevalence among cattle and sheep and usual onset of disease between 3-10 days of age suggests widespread distribution of virus, carrrier state and age susceptibility. In different surveys rotavirus has been found to be assoicated wiLh 41-48% of cases of neonatal calf diarrhoea. Animals confined to sheds and barns have a high incidence of disease because of exposure to accumulated viruses and bacteria. The rotavirus diarrhoea is highly contagious. An animal in early stage of diarrhoea excretes IOS - 1()9 virions per ml of faeces. The rotavirus diarrhoea develops a pattern at certain age. In one herd age may be 3-5 days and in another 14 day. No age resistance has been reported. The climatic conditions cause stress in the occurrence of disease. Mortality depends upon the secondary infection. In uncomplicated cases most of the animals recover but mortality is high

256 Textbook of Veterinary Virology
when there are complicating bacterial infections. Certain animals species like dogs and cats may play a role in dissemination of bovine rotavirus. In dogs and cats bovine rotavirus mUltiply asmyptomatically by experimental infection and rotavirus of bovine origin has been isolated from faeces of dogs.
Pathogenesis: The portal of entry is mouth. The primary site of replication of virus is the mature villous epithelial cells of small intestine but in lambs replication has been found to occur in the epithelium of caecum and colon. There is absence of viraemia in rotavirus infection. The virus is shed in the faeces. The titre of virus is very high about 108 _109 per ml in faeces in ftrst few hours of diarrhoea. There is circumstantial evidence that healthy calves and adult cattle act as carriers. Persistant infection has also been suggested. In experimental infection the incubation period varies and is about 12.5 hours to 36 hours. The clinical signs arc anorexia, depression, drooling of thick saliva and diarrhoea. The calves generally suffer at 3-10 days of age. The columnar villous epithelial cells are destroyed by infection and replaced by cuboidal squamous cells.
Immune reaction: In both calves and lambs circulating rotavirus antibody has little effect in preventing enteritis. The passive protection results from the presence of rotavirus immunoglobulin in the lumen of intestine. Susceptible calves are resistant to rotavirus infection just 48 hours of inoculation with attenuated rotavirus.
Diagnosis: Immunofluorescence is used to detect rotavirus antigen in faecal smears or in sections of infected small intestine collected early in the course of infection. Electron microscopic examination of faeces sometimes permits recognition of virus particles. Immunoelectron microscope is more speciftc for identification of virus particles. Virus isolation in cell culture can also be used. The clinical specimen of choice are faeces from early cases of diarrhoea or section of intestine and these yield satisfactory results. Number of other serological tests like ELISA reverse passive haemagglutination are useful for detection of viral antigens in faecal samples.
Control: An attenuated vaccine is available for oral immunization for newborn calves. Inoculation of cows with inactivated vaccine has been reported to reduce the incidence of diarrhoea is new approach for use on pregnant cattle. A combined rotavirus and E.coli antigen as a vaccine is also found to be effective for immunization of pregnant cows and newborn calves.

Reoviridae 257
Rotavirus infection in other animals
Rotavirus gastroenteritis has been reported in foal usually during ftrst 2 weeks of life. The equine rotavirus can infect piglets. A bovine rotavirus vaccine is used in foals to control the disease. Ovine rotavirus is anti genically related to bovine rotavirus. The virus causes gastroenteritis in 1-2 week old lambs. Rotavirus has also been isolated from goal<;. ~ota-like viruses have been observed in faecal samples of pups with diarrhoea but their etiologic role has not been ftrmly established. Naturally occuring rotavirual diarrhoea has been observed in pigs 10-28 days of age. The rotaviruses have also been isolated from turkey and broiler chIcken with and without diarhoea.
ORBIVIRUS
The virus particles of this genus have a double protein shell, the outer being skin-like; the inner shell has 32 ring shaped capsomeres with icosahedral symmetry. The virus consists of 10 segements of double stranded RNA.
Blue Tongue Virus (BTV)
Blue tongue (BT) is an insect borne infection and non-contagious disease of domestic and wild ruminants. It is prImarily a disease of sheep but cases of inapparent infection in cattle and goats take place and these animals may act as reservoir of virus for sheep and wild ruminants. The disease was recognised in Africa in 1652. It is widespread in African continent. The disease was identified outside Africa in Cyprus if! 1943. The disease was subsequently reported from Palestine, Turkey, Syria. The disease has been reported from USA and certain European countries like Portugal and Spain. Recently evidence of the presence of the disease has been reported from Latin America, Caribbean, South East Asia and Australia. In India the disease was first reported by Sapre (1964) from Maharasthra state. Uppal and Vasudeven (1980) reported the occurrence of disease in this country and virus was isolated and identified as serotype 3,4,9,16 and 17 from Himachal Pradesh and serotype 1,4 at Haryana. Subsequently Kulkarni and Kulkarni (1984) reported isolation of virus serotypes 9 and 18 from Maharasthra.
Properties of the virus: The virus particle is spherical and about 69 nm in diameter with 32 capsomeres. The capsid is double layered. The

258 Textbook o/Veterinary Virology
genome is of 10 segments double stranded RNA. The capsid contains fou/ major and three minor polypeptides. The virus particle is resistant to ether, chloroform and deoxycholate and gets inactivated below pH 6. The virio,n is stable at 4°C and -70°C but losses infectivity rapidly when frozen at -20°C. No haemagglutinin has been reported so far. There are 22 serotypes in BTV as revealed by virus neutralization test. Group specific antigens are revealed by complement flxation agar immuno-diffusion and -immunofluorescence tests.
Cultivation: The virus grows readily in fertile eggs after inoculation of embryos 10-11 da,Ys old by intravenous route or 6-8 days old embryos by yolk sac route. The intravenous route is more sensitive. The embryos die by both the methods of inoculation with multiple haemorrhages and are cherry red in colour. The optimium temperature of incubation of chicken embryos is 33.5°C. The virus can also be cultivated in 1-4 days old mice by intra-cerebral route of inoculation. After one or more passages in chicken embryos the virus grows in cell cultures like BHK21 , Vero and L cells with production of CPE. Lamb kidney monolayer cultures are less readily infected and the CPE production is less extensive.
Epidemiology: The blue tongue (B1) disease is an African disease but now it has been reported from almost all the countries of the world. The disease occurs in USA, in the near Middle and Far East, Japan, Indonesia, India and Northern region of Australia. Europe appears to be free at present. BTY has a wide host range including sheep, cattle, goats, deer, most Mrican antelopes and various other artiooacytles. The disease may be fatal and inapparent. Mostly the inapparent infection exists in most species. The infection progresses in cattle midge cycle and once a certain level of infection is attained the infection spills over to sheep. Sheep are apparently involved in a secondary cycle. Midges of the genus culicoides act as biological vectors of BTV. Midges become infected after feading an infected animal and the virus replicates in the salivary glands where it reaches a maximum titre in 6-8 days. The infected midge is infective for life. Transovarial transmission does not take place in midges. Overwintering of BTY is due to survival of infected midges as well as latently infected cattle. At least 22 species of culicoides transmit the disease. The BTY is not contagious. The transmission occurs only when infective midges bite the susceptible host or infective blood or tissue suspension arc

Reoviridae 259
inoculated parenterly. The excretions and secretions from infected animals contain minimal concentration of virus and therefore, regarded as harmless from epidemiological point of view. The semen from infected bulls during viraemic stage may be infective and cows inseminated with such semen become infected. The embryo transfer in cattle appears to be safe from risk of transmitting BTY, even when the embryo donors are viraemic at the time of transfer. However, sheep embryos collected from infected ewes occasionally may give rise to BTY infection in recipient ewes.
Palh~genesis: Following infection the virus replicates in regional lymphnodes and thereafter in other lymphnodes and lymphoreticular tissues and in endothelium, periendothelial cells and pericytes of small blood vessels. This leads to degenerative changes and necrosis leading to vascular occulusion, stasis and exudation. Once replication in target cells take place, virus appears in the blood stream and spreads to entire body. Virus is detected in blood from about 3-6 days PI and viraemia reaches its peak in about 7-8 days and then declines rapidly. After 14 days the virus is not detected in the blood of sheep but in cattle viraemia persists for a longer period. The virus is cell associated involving blood cells. A panleukopenia proceeds the appearance of viraemia. In cattle there is evidence that expression of clinical discase is due to IgE-mediated hypersensitivity reaction induced by previous exposure of BTV oc related viruses, however, it does not happen in sheep.
In sheep the incubation period is 6-7 days. The virus causes viraemia and is easily isolated from the blood during febrile stage. The affected sheep have a marked fever, oedema of muzzle, oedema and hyperaemia of the lips, buccal, nasal mucosa of eyelids. There is nasal discharge which may become mucopurulent. The saliva drops from the lips and muzzle becomes encrusted with discharge from nose and mouth. There is swelling and hyperaemia of the mucosa of mouth and ulceration of tongue, dental pad and lips. The tongue becomes swollen, eynotic and purple blue. Swelling and tenderness of coronary band and sensitive lamina of hoof result in lameness. Pregnant ewes may abort. Mortality in sheep may be high.
Experimental disease in cattle is subclinical. In natural infection, laminitis, stiffness, ulcers in the mouth, nose and muzzle and salivation are seen. In affected pregnant cows the foetus may die and be

260 TeXlbook of Veterinary Virology
reabsorbed, aborted or still born. The BTV has teratogenic properties. Affected newborn lambs or calves are blind ataxic, undersized and have congenital defects.
lmmu",e reaction: In immunocompetent animals the group specific and type specific antibodies appear within 7-10 days of infection. The type specific neutralizing antibodies persist for more than 3 years, while the group specific antibodies persist for 6-18 months only. The protective immunity is generally associated with neutralizing antibodies but sometimes animal resist infection of BTV w.ith no demonstrable neutralizing antibodies. The CMI responses to BTV infection in sheep are also protective and may be less type specific. Lambs born to BTV immune animals are passively immune and remain protected upto 6 months.
Diagnosis: In sheep, tentative diagnosis is made on the clinical signs. Confirmation is made by positive disease transmission to susceptible sheep or by viral isolation from blood, spleen, lymph nodes etc in chicken embryos. The blood should be collected from febrile animals in the early stage in EDTA or citrate. Occasionally isolations can also be made by intracerebral inoculation of newborn mice or inoculation of cell cultures, BHKzI' Vero or Aedes albopicuts cells. The embryos die within 6 days of inoculation with haemorrhages. Infected CAM is used as a source of virus in virus neutralization or complement fixation test, ELISA etc.
Control: Live egg adapted polyvalent vaccines are widely used and are effective. The live vaccines are not recommended to be used because the live virus may cause some abnormalities in foetus. Promising results are being obtained with inactivated vaccines. Import controls is practised in certain countries like Australia.
African Horse Sickness Virus African horse sickness (AHS) is an insect borne virus disease of
solipeds. Horses are more susceptible as compared to donkeys. The disease is prevalent in Africa and Middle East. The epidemic of 1959 spread over a large area reaching on one side upto Turkey and on the other side upto India. In India the frrst outbreak was reported in April, 1960. Approximately about 90% of affected animals died. It is suspected that movement of affected animals in the adjoining areas of Pakistan was responsible for the spread of the disease. Since 1963, no new case of the disease has been reported from India.

Reoviridae 261
Properties of the virus: The virus has an icosahedral symmetry with 32 capsomeres and measures about 50 nm in diameter. The virus is stable between pH 6-10 but is labile at pH 3. The virus is stable at 4°C but is inactivated at 36°C when kept for 40 days. The infected blood can be preserved in oxalate-carbol-glycerine solution for years. The virus haemagglutinates equine rbe. Nine anti genically distinct serotypes of the virus have been identified. A common group specific antigen is shown by complement fixation test in different strains of virus. By virus neutralisation and haemagglutination inhibition the virus strains can be distinguished. Strains within each serotype vary in virulence.
Cultivation: The virus can be cultivated by intracerebral route in mice, rats, guinea pigs and other laboratory rodents. Viscerotropic field strains become neurotropic by serial intracerebral passages. The virus become attenuated for horses without loosing antigenicity. Ferrets can be infected by intravenous route. Neurotropic strains can be grown by CAM route of inoculation in chicken embryos. The small plaque variant is usually more virulent than the large one.
Epidemiology: The AHS is the disease of horses, mules and zebras are less susceptible but donkeys are generally resistant. Contact transmission does not occur among horses. A number of blood sucking insects have bcen circumstantially incriminated in the spread of virus. The culiocoides transmit the disease but exact role has not yet been determined. Trransmission is primarily through the mosquitoes. Lymphnodes, lungs and spleen have the highest concentration of virus.
Pathogenesis: Under natural conditions, the incubation period is less then 9 days while experimentally the incubation period range from 2-21 days. There are four different forms of disease. African horsesickness fever, acute pulmonary form, cardiac form and mixed form. There is no relationship between the virus type involved and the kind of clinical disease produced.
Mixed form is a common form where combination of pulmonary and cardiac form symptoms are observed. The mortality rate varies: sometime it may be 90% and in certain outbreaks it may be 25%. The pathologic lesions depend on the clinical form of the disease. In pulmonary form the changes are oedema of lungs and hydrothorax. In the cardiac form the lesions are oedematous infiltration of subcutaneous, subserous and intramuscular tissues and lymphnodes. Hydropericardium is a constant 'feature.

262 Textbook of Veterinary Virology
Diagnosis: During outbreak of disease tentative diagnosis can be made fonn the clinical symptoms and gross pathologic lesions. The virus isolation is done from the blood collected at an early stage of disease or from the spleen extracts when inoculated intracerebrally into suckling mice, ferrets, embryonating chicken eggs and cell culture. Infected mice die within 5-15 days of inoculation. The virus is identified by virus neutralization test conducted in mice or cell culture system. Antibodies in the convalescent serum are detected by virus neutralization, complement fixation, haemagglutination inhibition and immunodiffusion tests.
Control: Polyvalent vaccines are used for immunization. There is some cross immunity among certain serotypes. Mouse brain adapted or cell culture vaccines are used for animal vaccination. Antigenic drift or vaccinal break-down can occur. Therefore, horses in enzootic areas should be protected from flies during outbreaks.
References
BARBER, T.L. and IOCHlM. M.M., 1985. Bluetongue and related orbiviruses (progress in clinical and Biowgical Research, Vol. 178) Alan R. Liss, New York. 746 pp.
BUXTON, A. and FRASER, G., 1977. Animal Microbiology Vol. 2. Oxford, Blackwell Scientific Publications.
CAMPBEU., C.H. and GRUBMAN, M.I., 1985. Current knowledge on the biochemistry and immunology of bluetongue. In R. Pandey (Editor), Progress in Veterinary Microbiology and Immunology, 1, 58-79.
GIBBS, E.P.l, 1981. Virus diseases offood animals. Vol. IT. Academic Press, London.
GORMAN, B.M.; TAYLOR, I. and WALKER, P.I., 1983. Orbiviruses In: W.K. Joklik (Editor). The Reoviridae.Plenum New YQrk. pp. 287-357.
HOUSE, lA., 1978. Economic impact of rotavirus and other neonatal disease agents of animals. 1 Am. Vet. Med. Assoc. 173: 573-576.
KAHRs, R.F.,1985. Viral diseases of cattle. Iowa State Press.
KULKARNI, 0.0. and KULKARNI, M.N., 1984. Isolation of bluetongue virus from sheep. Indian lournal Comparative microbiology Immunology and Infections Diseases 5, 125.

Reoviridae 263
KUMAII., S., 1976. The African horse sickness. Indian Council of Agricultural Research, New Delhi.
MRSOLAIS, G.; AssAF, R.; MoNTPe1TJ', C. and MAROIS, P., 1978. Diagnosis of viral agenls associated with neOTUJlai calf diarrhoea. Canadian Journal Comparative Medicine. 42, 168-71.
MARtEL, J.L. and PERRIN, B., 1981. Infectious etiology of diarrhoea in newborn calves. Prevalence in France of E.coli Kgg + and rotavirus. Practcical diagnosis techniques. Bull. Group Tech.Vet., 4,45-54.
OLsBN, N.O., 1978. Reovirus infection. In diseases of poultry, 7th ed. By M.S. Hofsted et al. Ames. Iowa. Iowa State University.
PBNsAERT, M.B., 1984. Viral gastroenleritis in suckling pigs. Rev. Sci. Tech. Off. INt. Epiz. 3,809-18.
RusSEl.L, P.H. and EDINGTON, N., 1985. Veterinery Viruses. The Burlington Press (Cambridge ltd) Foxton, Cambridge.
SAPRE, S.N., 1964. An outbreak of bluetongue in goats and sheep in Maharastra State India. Veterinary Review (M&B) 15, 69-71.
TAYLOR, W.P., 1986. The epidemiology of bluetongue. Rev. Sci. Off. Int. Epizootics 5, 351-56.
UPPAL, P.K. and V ASUDEVA1~, B., 1980. Occurrence of bluetongue in India. Indian Journal comparative microbiology, Immunology and Infectious Diseases 1, 18-20.
VERWOERD, D.W.; HUlSMANS, H. and ERASINUS, BJ., 1979. Orbiviruses. In H. Fraenkel-Conrat and R.R. Wagner (Editors), Comprehensive Virology, 14,285-345.

Chapter 24
Birnaviridae
In 1984 a new family was designated Bimaviridae to include infectious bursal disease of chickens and infectious pancreatic necrosis virus of fish.
The viruses in this family are icosahedral having a size of 60 nm in diameter with 32 capsomeres. The virions are relatively heat stable and resistant to ether, chloroform and at pH 3. There are four structural nonglycosylated polypeptides. The genome consists of 2 segments of ds RNA. The viruses replicate in the cytoplasm without depressing cellular RNA or protein synthesis to great extent. The infectious bursal disease virus (IBDV) replicates in both chicken and mammalian cells and produce CPE in 3-4 days.
The family comprises of IBDV, infections pancreatic necrosis virus of fish and viruses affecting insects and molluscous.
Infectious Bursal Disease Virus (IBDV)
Infectious bursal disease virus is no longer considered to be in reovirus group. The virus has one layer of capsid protein and genome has two segments instead of 10-12 in reoviridae.
The virus causes an active infectious disease of young chickens. The virus has two synonyms; avian nephrosis and Gumboro disease after the name of a district in Delaware (USA), where the virus was flfst encountered, in 1950. Now the virus has been reported from all major poUltry producing-areas of USA, Great Britain, Italy, Israel, West Germany, the Netherlands, Japan, Iraq as well as South American countries. The disease is prevalent in India as reported by Mohanty et

Birnaviridae 265
al. (1971) and other workers. The virus infects chickens and turkeys but not ducks. The disease incidence is greatest in 3-6 weeks old chicks but outbreaks have been reported in chicks 11 days old as well as 12 weeks old. The virus destroys immunological organ namely bursa and leads to
breakdown of imunity breading the flock susceptible to many infections and there are also cases of vaccine failures.
Properties: The virus particle measures 50-60 nm in diameter. The genome is double stranded RNA with 2 segments. The virus is comparatively heat resistant. It survives for 5 hours at 56°C. The virus is inactivated by formalin and iodophores but is resistant to ether, chloroform, phenol, thiomersal etc. The virus grows on embryonating chicken eggs. The CAM route of inoculation is more sensitive then yolk sac and allantoic cavity route. Lesions produced in chicken embryo are subcutaneous oedema, haemorrhages, dwarfing, liver necrosis and death. Serial passages in chick embryo reduces virulence for chicks. IBDV also grows in chicken embryo kidney cell cultures and produce CPE in 3-5 days. No CPE is produced in pig, hamster, calf kidney, HeLa and amnion cell lines.
Epidemiology: IBDV affects chicken only. The chickens 3-6 weeks of age are most susceptible but older chickens also catch infection. Laying birds as well as chicks 1-14 days of age do not show disease. The yirus spreads directly by contact. It is highly contagious disease. The specific mode of transmission appeared to be contaminated feed and water. IBDV remains viable for atleast six months in dry litter and in unused houses for more than one year. The virus has also been isolated from insects and mosquitoes which indicates that insects may be involved in the spread of virus.
Pathogenesis: IBD has a short incubation period of 2-3 days. Lesions normally appear in bursa before the onset of clinical signs. Chickens between 3-6 weeks are most susceptible but older chickens have been reported to sOffCJ. The clinical signs are ruffled feathers, anorexia, depression, diarrhoea with soiled vent. The affected birds become dehydrated and die. The mortality is 20-30% but morbidity reaches 100%. The lesions are dehydration, haemorrhages in leg and thigh muscle, hepatic infarction, enlarged kidneys; the bursa is usually enlarged, oedematous and yellowish after 2-3 days of infection. ·It returns to its normal weight by 5th day and continues to atrophy rapidly. It is necrotic in many birds. The histological changes include

266 Textbook o/Veterinary Virology
fecal necrosis and hyperplasia of epithelial and reticular cells of bursa, spleen and thymus. There is suppression of cell mediated immunity by IBOV infection.
Diagnosis: The clinical signs, gross lesions and histopathological changes in. the bursa are characteristic and helpful to arrive at a tentative diagnosis. Other factors like, age, history of birds, clinical signs and mortality pattern are important in arriving at tentative diagnosis. Confirmation of the disease is based on isolation and identification of the virus. The isolation can be made from the kidneys, spleen or bursa of the affected birds in the eggs. The virus can be identified by VN in chicken embryos or cell cultures. The FAT and agar gel precipitation test is applied to affected bursal tissue of the birds.
Control: A solid immunity results in 10-16 weeks of age. Chicken embryo or cell culture vaccines are available in certain countries for oral or ocular administration. These vaccines produce about 1 % mortality and bursal lesions. The attenuated vaccine prepared in Vero cells is giving good results.
References
BESHT, H., 1980. Infeclious bursal disease virus. Curr. Top. Microbiol. Immunol. 90, 107.
FARAGHER, J.T., 1972.lnfeclious bursal disease of chickens. Vet. Bull. 42, 361.
HITCHNER, S.B., 1978.lnfeclious bursal disease. In diseases of poUltry. 7th Ed. Edited by M.S. Hofstad el al. Ames Iowa State Univeristy, Press.
OKOYE, J.O.A., 1984. Infeclious bursal disease. In diseases of Poultry. 7th Ed.
Edited by M.S. Hofstad et a1. Ames Iowa State University, Press.

Chapter 25
Coronaviridae
The characteristic club-shaped surface projections studding the virion envelope (Latin corona-crown wreath) have given the family its name.
The viruses of this family are pleomorphic, enveloped, measure about 100-120 nm in diameter with unique club shaped peplomers projecting from envelope, which measure about 20 nm in length. They are widely spaced then in myxoviruses and resemble a coronet The capsid is of helical symmetry. The genome consists of one molecule infectious RNA of about 7 x 1()6 mol wt. The virions contain three major classes of structural proteins, the nucleocapsid protein, the matrix or envelope protein and the surface or peplomer protein. The capsid protein is phosphorylated.
Coronaviruses replicate in the cytoplasm. There is production of 3' coterminal sUbgenomic RNA with unique sequences extending in S' direction. The virions bud into cisternae acquiring lipid membrane from the cell and subsequently transported to and accumulated in Golgi vesicles. The budding from plasma membrane does not take place. Coronaviruses do not grow well in continuous cell lines until they are adapted in tissue culture. The viruses can be isolated in primary culture of the appropriate species. The CPE may be minimal with some vacuolation.
There are several coronaviruses of veterinary importance such as: Chickens: Avian infections bronchitis virus (IBV) Pigs: Transmissible gastroenteritis (TGE) virus, haemagglutinating
encephalitis virus and epidemic diarrhoea virus.

268 Textbook of Veterinary Virology
Dogs: Canine coronaviruses Cats: Feline infectious peritonitis virus (FlPV) Mice: Mouse hepatitis virus Cattle: Bovine coronavirus causing diarrhoea in calves and
occassioI1ally precipitates in .respiratory tract infections. In adult cattle the bovine coronavirus may be responsible for a disease ca\1ed winter dystentry. All isolates belong to one serotype.
Bovine Coronavirus
Bovine coronavirus produces neonatal calf diarrhoea in calves day to 3 weeks Or" more of age. The virus has also been isolated as a member of mixed infection from trachea and lungs of calves with respiratory disease. A virus similar with morphological, physicochemical and antigenic properties to coronaviruses was identified in faeces of calves suffering from winter dystentry. The young mature dairy cattle aged 2-3 years are infected during winter in crowded barns. The bovine coronavirus is also called the Nebraska calf diarrhoea coronavirus.
Properties of the virus: In negatively stained preparations the virus measures 120 nm in diameter and covered by petal shaped projections of 20 nm long. The virus haem agglutinates and haemadsorbs red blood cells of hamster, mice and rats. The virus is stable at pH 3 and thermostabilized by IM MgCI2• The virus 'is sensitive to ether, chloroform, deoxycholate, and exposure to 50°C for 1 hour. Formalin 0.02% at 37°C completely inactivates the virus in cell culture fluid in 24 hours. The virus can be isolated in BEK cells and vero cells. The CPE is produced after few passages and is characterised by syncytium. Some bovine strains have not been grown in cell culture and are maintained by oral inoculation of calves. Only one serotype has been identified so far.
The virus strains isolated from cases of winter dystenlry in adult cattle have properties similar to bovine corona virus. Morphological, physical and chemical properties, anti genic properties are similar to those of coronaviruses.
Epidemiology: The epidemiology is not completely known. It appears that carrier and infected cows are the reservoir animals and source of infection. The virus is excreted in faeces and ingestion is the source of infection. The virus appears to be ubiquitous in cattle

Coronaviridae 269
population. The occurrence of coronavirus is suspected in India. Morbidity in an outbreak in calves in high but mortality is influenced by the age of the calf, management and type of secondary infections. The winter dysentry in cattle has been reported from USA, Great Britain, Sweedan, Germany, France, Belgium, Israel, Japan, Australia and Newzealand. Animals frequently affected are of 2-3 years of age and are infected during winter in crowded barns especially during post partum. The young calves under 4 months resist the disease or are affected with a milder form. The mortality does not exceed 10% but morbidity is 100%. The economic consequences concern milk production, which is reduced by 25-95% for 1-2 weeks and afterwards it does not come to the intiallevel. The faeces of the effected cattle is the source of infection and virus is transmilted by oral route.
Pathogens is: Portal of entry is the mouth through contact with feed or fomites contaminated with infected faeces. The primary site of viral replication are the mature epithelial cells on the small intestinal villi and surface epithelial cells of the colon.
There is no precise data available of the winter dysentry. It is persumcd that (i) coronavirus isolated from cases of winter dysentry may be of a single serotype causing diarrhoea in cattle regardless of their age, (ii) the campylobacter acts as opportunistic agent due to their invading property and colonize the crypts in the same region, (iii) lipopolysaccharides or endotoxins of bacteria induce a phenomenon of localised and/or disseminated intravascular coagulation, the source of dysentry.
The incubation period in gnotobiotic calves by oral inoculation is 20 hours. Initially the calves are depressed, eat slowly and have a liquid yellowish diarrhoea. The diarrhoea is acute and faeces may contain coagulated milk or mucus. There may be excess salivation, weakness, lethargy and severe dehydration and shock. The diarrhoea may persist for 5-6 days. The diarrhoea probably results from villous atrophy in small intestinal mucosa and consequent maladsorption and loss of enzymatic activity. Severely shortened villi and occasional fusion of adjacent villi are seen in the intestine. Immunodeficiency, failure to ingest colostrum, secondary bacterial infection and multiple viral infections all contribute to the clinical signs observed. Except for distended bowel and small intestine with liquid faeces, no gross lesions are observed. There may be pulmonary congestion and pneumonia. In

270 Textbook o/Veterinary Virology
winter dysentry the incubation period is between 3-7 d.ays. The symptoms start all of a sudden with brief period of temperature (39.5-40.5°C). anorexia and apathy. Sometimes there is cough and nasal discharge. Milk production is reduced drastically. In most cases the diarrhoea· lasts for several days about 2 weeks on an average.
Immune reaction: Circulating coronavirus antibodies do not prevent infection but cause attenuation of infection in neonatal calf diarrhoea. Regular ingestion of colostrum from immune dams can be protective. The resistance to coronavirus infection is primarily due to
presence of colostral and milk antibody or actively produced IgM and! or IgA in the intestinal lumen. .
In winter dysentry. immunity is formed after a herd epizootic. The protection ranges from 6 month to 3 years. The greater susceptibility in young adults may be due to loss of colostral antibodies and increasing stress.
Diagnosis: The diagnosis can be made by demonstration of virus particles in faecal samples collected at early stages of virus diarrhoea. The IF staining of spiral colon sections collected from a calf killed within 4 days of onset of diarrhoea are of diagnostic value. In faecal samples coronaviral antigen can be detected by immunoelectrophoresis. reversed passive HA and ELISA. The virus can be cultivated in primary bovine embryonic kidney cells. BEK-l cell lines. The replication is enhanced by the addition of trypsin.
Control: A live attenuated vaccine is available and is effective if given to newborn calves soon after birth. A combined attenuated rotacorona virus vaccine is also available which can be given to newborn calves as well as to pregnant cows.
Canine Coronav!rus (CCV)
In 1971 CCV was isolated in United States from military dogs with diarrhoea! disease. The significance of CCV infection in dogs was recognised in 1978 when outbreak of corona associated diarrhoea occured in dogs in USA. The infectious range in severity from inapparent to fatal. younger dogs are mpre severely affected. Experimental infection of neonatal dogs causes gastroenteritis with diarrhoea and virus is excreted in faeces for 6-9 days. The corona viral infections occur concurrently with other viruses. parasites or pathogenic bacteria. In natural infections the majority of dogs recover

C oronaviridae 271
in 7-10 days but death in few dogs takes place within 24-36 hours of onset of symptoms. The morbidity rate is variable and mortality rate is low. The CCV produces CPE in several primary and continuous dog cell cultures. Primary dog kidney and thymus cells, thymus cell lines and other dog cell lines support the growth of virus. Infected cells become enlarged and bizzare shaped. At present no vaccine is available. The anti genic differences observed among CCV isolates may complicate the development of a vaccine.
Feline Infectious Peritonitis Virus (FIPV) Feline infectious peritonitis is a disease of wild and domestic cats.
Two strains of virus are reported. One strain causes clinical disease in domestic and wild cats and second one is less virulent and causes diarrhoea in young animals. The virus is similar in morphology to other corona viruses and has similar physical properties. The virus can be propagated in organ cultures of feline tissues. Virus can sometimes be isolated by co-cultivating peritoneal macrophages with susceptible cat cell lines. The virus causes rounding of few foci of cells and syncytia and replicates by intracerebral inoculation in suckling mice. The replication of virus in the brain of suckling mice or in cell culture can be detected by IF test. FIP occurs in cats of all ages. The disease is inapparent in most cases but is fatal once the symptoms appear. The incubation period is at least 4 months. The initial symptoms are anorexia, depression and biphasic fever causing weakness. In effusive form of disease there is abdominal distension due to peritonitis. The cats die within 1-8 weeks after showing the clinical signs. The CNS, eyes and visceral organs are also involved. There is moderate to marked leucocytosis. Hyperproteinemia is a constant feature due to elevation of gamma globulin. The characteristic post mortem lesions is parietal fibrinous peritonitis and/or pleuritis with fluid in body cavities. Inflammation or necrosis of visceral organs occur. Mesenteric and caecal lymph nodes are enlarged. The exudate layer adhering to the peritoneum is composed of fibrin. The mode of the transmission of disease is not known but close contact allows transmission. The disease results from Arthus type of immune complex vasculitis. The tentative diagnosis can be made from clinical signs. Antibodies can be detected by IF test. The virus isola,tion can also be attempted. Presently there is no vaccine availab~ and incidence is sporadic.

272 Textbook of Veterinary Virology
Porcine Coronaviruses
Transmissible gastroenteritis virus(TGEV), Haemagglutinating encephalitis virus (HEV) and Epidemic diarrhoea virus.
Transmlssble gastroent~ritis virus (TGEV)
The virus produces highly contagious disease of swine characterized by profuse diarrhoea, vomiting, dehydration and high mortality in young piglets. Only pigs suffer from natural disease. Mice and dogs can be experimentally infected and subsequently shed virus in their urine. The disease was flrst reported in United States in 1946 and subsequently reported from Canada, Taiwan, Britain, Japan, Soviet Union and many European countries.
Properties of the }'irus: The virus is mostly circular in shape but pleomorphic forms are also present. The size of the virus is between l)()-200 nm. The morphology is typical of the group. The virus survives several weeks in frozen tissues. TGEV is inactivated at 56°C within 30 min., but survives for 3 days at room temperature. The virus is stable at pH 3 and is not affected by 0.5% trypsin. It is inactivated by detergents, ether and other lipid solvents. The virus does not show haemagglutination. All strains belong to one antigenit type.
TGEV can be grown in primary pig kidney, thyroid, testis and salivary gland cell cultures. Some fleld strains may produce transient or negligible CPE in initial passages. Growth in dog kidney cells and chicken embryos has been reported.
Epidemiology: The route of infection is by ingestion and probably by respiratory route. The virus multiplies in the intestine. The virus is excreted in the faeces and this continues upto 8 weeks or longer in recovered pigs. Non lactating sows have mild disease but act as carriers. The persislance of virus in farm buildings, swill and recovered carrier sows play a major role in the spread of the disease.
Pathogenecity: The severity of the disease is related to the age of pigs but swine of all ages are susceptible. The disease is fatal 10 piglets under 1 week of age but mortality halves with each extra week of age. The mortality in pigs over 5 weeks of age is low. The majority of outbreaks occur from mid winter 10 early spring. The incubation period is 18 hours to 3 days. The clinical signs are vomiting, profuse diarrhoea, thirst, loss of body weight with high morbidity and mortality in pigs under 2 weeks of age. In older pigs the diset\ses runs a chronic

Coronaviridae 273
course and often no symptoms are observed except failure to gain weight The post mortem examination of young piglets show the stomach filled with coagulated milk and intestines distended with yellow foamy fluid. There is acute enteritis and severe ulceration. In the terminal stages of the disease the intestinal tract becomes thin and transparent There is extensive atrophy of intestinal villi in jejunum and ileum. Degenerative lesions and haemorrhages may be found in the spleen, kidney, urinary bladder and heart. There may be some congestion and encephalitis in the central nervous system. Excessive villous atrophy of jejunum and ileum and absence of chyle in mesentric lymphatics is characteristic ofTGE.
Diagnosis: A presumptive diagnosis is possible on clinical and epidemiological features of the disease. Villous atrophy is suggestive of TGE. The FA test is applied to demonstrate the presence and viral antigen in colonic lesions and faecal smears. The virus particles can be demonstrated in the faeces or small intestine by electron microscopy or immune electron microscopy. Four fold increase of neutralizing antibodies can be demonstrated in paired serum samples. The virus isolations can be done in cell cultures and identified by neutralization test or FA test.
Control: The control measures are usually inadequate. Available vaccines are unsatisfactory because they fail to elicit adequate lactogenic immunity.
Haemagglutinating-Encephalomyelitis Virus (HEV)
Vomiting/wasting disease
The virus produces an acute highly infectious disease of young pigs under 3 weeks of age. The disease was reported from Canada in 1958 and the virus was isolated in 1962 in Canada from the brains of suckling pigs with fatal encephalomyelitis. HEV is prevalent in Canada, England and USA. All strains of the virus are believed to be anligenically homogenous.
The virus haemagglutinates and haemadsorbs chicken, turkey, sheep, rabbit, mouse and hamster rbe but does not possess neuraminidase. The virus can be cultivated in pig kidney cell cultures and produces syncytia. The vomiting and wasting syndrome appears in pigs 1-3 weeks old. There is vomiting, depression, anorexia and constipation. Some- piglets may have respiratory signs, including

274 Textbook o/Veterinary Virology
sneezing and coughing. The piglets become emaciated and die of starvation; recovered pigs are unprofitable. In the encephalitis form there is depression, anorexia and loss of oondition. Hyperesthesia, incoord\nation and tremor with occasional vomiting for 2-3 days is followed by acute encephalitis, convulsions and death. The mortality rate goes upto 100%. There are no gross lesions on post mortem examination. Mild lesions may be seen in eNS, a presumptive diagnosis can be made from the clinical history. The definite diagnosis is arrived at by isolating the virus from the respiratory tract and brain. The FA techniques can be applied to demonstrate the viral anti~en in tissues.
Avian Infectious Bronchitis Virus (IllV)
Avian infectious bronchitis virus (lBV) causes an acute, highly contagious respiratory disease of chickens. The disease probably occurs in most parts of the world where poultry is raised. The disease was first described in United States in 1931. The presence of disease in India was established in 1969. There is quite high incidence of the disease in this country. The chicken is the only natural host of IBV.
Properties: The virion are approximately spherical in shape and measure about 80-120 nm in diameter. The virus particle has club shaped projections distributed on the surface. These are approximately 20 nm in length. The virion contains a central core of single stranded RNA and is enclosed within a capsid. Approximately 16 polypeptides have been found. Some IBY types after treatment with phospholipase C agglutinate chicken rbe. This is inhibited by IBY antiserum. The virus is sensitive to treatment with ether and other lipid solvents:The thermal stability varies from one strain to another. Newly isolated strains are inactivated at 56°C within 15-30 mins while·chicken embryo adapted strains survive upto 3 hours at this temperature. Infectious bronchitis strains vary in antigenicity. By neutralization test, two antigenic variants have been recognised-Connecticut and Massachusetts. There are reports that other serotypes also occur. By immunodiffusion test three soluble antigens have been recognised. One of the antigens resides in the virus particles while other two are distributed over the surface of virus.
Cultivation: The virus grows well. in 9-12 days old embryo by allantoic cavity inoculation. The yolk sac route is unreliable because of

Coronaviridae 275
the possibility of maternal antibodies in yolk. The virus does not produce pock marks on the CAM but there is characteristic dwarfing and curling of embryos. Serial passages in egg embryos result in decreased pathogenicity, antigenicity and immunogenecity with a progressive increase in embryo lethality. A genotypic modification probably occurs after serial passage3 in chicken embryos. A few strains can also be grown by intracerebral inoculation in unweaned mice, producing ascending prsalysis and death after 3-4 days of inoculation. The virus can be propagated in chicken embryo kidney, lung and liver cells, in CK and Vero cells and chicken ~acheal organ cultures. The virus produces syncytia in affected cell cultures. New isolates or low embryo isolates grow poorly or not at all in cell cultures; tracheal organ cultures are suitable for this purpose.
Epidemiology: The virus is highly infectious and may be shed in the respiratory discharge for 4 weeks and faeces 3 .weeks after infection. Virus may be present in the fertile eggs laid during illness but chicken hatched out of these eggs are not infected. In some cases the virus is shed upto 7 weeks. The virus spreads by droplet infection but indirect transmission may also occur through feed racks, water troughs, clothing and equipment.
Pathogenicity: Chicken is the only natural host. Birds of all ages are susceptible; however, chicks of 1-4 weeks of age are most severely affected. The incubation period is i8-36 hours. The severity of clinical symptoms vary considera!)ly and may go unnoticed in growing and laying birds. Symptoms are severe in younger birds where mortality is between 20-90% particularly with secondary infection by mycoplasma or septicaemic E.coli. The clinical symptoms are depression, coughing, gasping, tracheal roles, watery eyes and nasal discharge. The virus replicates in respiratory and urogenital tract epithelium 1-8 days post infection. Urogenital tract infections results in nepharitis with tubular damage and interstitial infiltration of lymphoid cells and its epithelium cells. The laying flocks have lowered egg production and eggs may be misshapen, rough; soft shelled and of poor internal quality. Necropsy reveals sinusitis, catarrhal tracheitis; bronchitis, congestion and oedema of lungs. The caseous plugs may be found in the lower trachea and the bronchi.
Immune reaction: The virus induces S.N, C.F and precipitating antibodies in infected chickens. IgA type antibodies have also been

276 Textbook of Veterinary Virology
demonstrated The chickens which recover from natural infection are resistant to intratracheal inoculations with homologous strains. Some degree of immunity and antibodies can be demonstrated for at least one year after natural infection. A high level of antibodies appear 3 weeks after exposure.
Diagnosis: A presumptive diagnosis of IB is possible on the basis of clinical signs and highly contagious nature of the disease. Confirmation is based on isolation of IB V in chicken embryos from trachea, bronchi and lungs. The tissue cultures are not sensitive for isolation of the virus but tracheal organ cultures can be used. Antibodies in the infected flocks can be detected by HI tests using phospholipase with treated virus. The FA test on tracheal scrapings provides a rapid diagnosis. A blot-dot hybridisation test has been developed which identifies all isolates of IBV because their RNA will hybridise with DNA probes from the matrix protein gene of a standard strain.
Control: Recovered chicks are immune after natural attack for atleast one year. Local immunity in the respiratory tract is important in protection against the disease. Vaccination with attenuated strains is widely practiced. The vaccination is done with attenuated virus Massachusetts(1924) by aerosal spray, intranasal or intraocular in 1 day old chicks or in the drinking water 2-3 weeks old. Administration by aerosal or spray is more effective as compared to drinking water.
References
BUXTON, A. and FRASER, G., 1977. Animal Microbiology. Vo1.2 Oxford, Blackwell Scientific Publications.
CARMICHAEL, L.E. and BlNN, L.N., 1981. New enteric viruses in dogs. Advances in Veterinary science and comparative Medicine. 25,1-37.
DURHAM, P.J.; SlEVENSON, DJ. and FARQUIIARSON, B.C., 1979. Rotavirus and coronavirus associated diarrhoea in domestic animals. N.z. Vet. 1. 27,3()"32.
GRElG, A.S., 1975. Haemagglutinating encephalomyelitis virus infection. In disease of Swine 4th Ed. Edited by H.W. Dunne and A.D.Leman, Ames, Iowa, Iowa State University, Press.
HOSFSTAD, M.S., 1975. Immune response to infectious bronchitis virus. Am. 1. Vet Res. 36, 520.

Coronaviridae 277
HOFSTAD, M.S., 1978. Avian injections bronchitis virus. In diseases of poultry, 7th ed. Edited by M.S. Hofstad at al. Ames, Iowa, Iowa State University Press
KAIIRS, R.F., 1985. Viral diseases of cattle. Iowa State University Press.
MAC lNToSH, K., 1974. Coronaviruses: a comparative review. Curr. Top. Microbiol. InunW1ol. 63: 85-139.
MEBUS, C.A.;- STAIR, E.L.; RHODES, M.B. and TWICHAus, MJ., 1973. Neonalal calf diarrhoea! Propagation, attenuation and characteristics oJ-a coronavirus like agent. Am. J. VeL Res. 34,145-150.
RUSSELL, P.H. and EDINGTON, N., 1985. Veterinary viruses. The Burlington Press (Cambridge) Ltd.Foxton, Cambridge.
SIDDELL, S.; WEGE, HaMUT and MEULEN,VoLKER,TER; 1983. The biology of corona viruses. J. General Virology.64: 761-776.
TAKAHASHI, E.; lNABA,Y.; SATO, K.; ITo, Y.; KUROGJ, H.; AKAsm, H.; SATODA, K. and OMORJ, T., 1980. Epizootic diarrhoea of adult cattle associated with a coronavirus-like agent. Vet. Microbiol. 5, 151-154.

Chapter 26
Orthomyxoviridae
The virus family orthomyxoviridae consists of one genus made up of influenza A, influenza B and influenza C on the basis of shared antigens by complement fixation and immunodiffusion tests. Type A influenza viruses include all pathogenic veterinary viruses. However, type C has been isolated from pigs in China during abaLloir surveys. The influenza A viruses consist of roughly spherical or filamentous enveloped particles which measure 80-120 nm is diameter. The nucleocapsid is helical and is enclosed within a protein matrix. The protein matrix is enclosed by a lipid membrane which is covered by two types of glycoprotein spikes with which haemagglutinin and neuraminidase activities are assoicated. The virus genome is a single stranded RNA in eight segments with a moleuclar weight of about 4 x 1()6 daltons. Recombination occurs with high frequency within a species but not among species. The influenza viruses are divided into types, A, B and C on the basis of anti genic character of internal nucleoprotein antigen. The influenza A viruses are further divided into sUbtypes on the basis of haemagglutinin (H) and neuraminidase (N) antigens. Influenza viruses are sensitive to heat, drying, detergents and disinfectants and do not survive in external environmenL The viruses can be grown in chicken embryos in amniotic and allantoic cavity of 10 day old fertile eggs. Some isolates grow in cell cultures; the CPE produced is rounding and detachment of cells. Influenza viruses are usually limited to their original host species.
Strain differences within the subtypes take place with altered H & N antigens. The alteration is the res.ult of residual virus infection in

Orthomyxoviridae 279
layer
Lipid layer
----_.--' Fig. 26.1 Schematic Representation or the Structure ofInnuen:l.a Virus.
animals whose antibodies favour the selection of mutants with altered Hand N epitopes. A change of subtype within species is known as antigenic shift The antigenic shift may result from, (i) mutation, (ii) genetic reasortment, (iii) adaptation of influenza viruses from one species 10 another.
Equine Influenza Virus subtypes 1 and 2
The virus causes an acute respiratory illness in horses. The disease is world wide in occurrence. There was a disease outbreak is 1987 in Northern India.
Properties of the virus: The virus of equine influenza has a morphology typical of the orthomyxoviridae family. The virus agglutinates horse, calf, pig, rhesus monkey, fowl, pigeon and human rbe. The viruses belong 10 type A on the basis of their type specific internal soluble antigen as detected by CF test. There are two distinct serotypes. The virus type 1 carries I~N7 antigen while type 2 isolates carry ~. antigens.
The virus can be cultivated in embryonating chicken eggs and in cell cultures of equine, bovine and monkey kidney celIs. The CPE is characterised by an early development of syncytia and formation of multiple eosinophilic intracytoplasmic inclusions. The infected cell cultures haemadsorb guinea pig red blood cells.

280 TeXlbook of Veterinary Virology
Epidemiology: The horses excrete virus upto 10 days via nasal secretions. Spread is by droplet infection. The disease spreads rapidly due to frequently violent cough and partly due to short incubation period.. The infection is transmitted exclusively form horse to horse by aerosal infection.
Pathogenesis: Equine influenza is an acute disease of lower respiratory tract of horses that affects all ages. The serotype 2 is more pathogenic than serotype 1. The incubation period is 1-3 days. The morbidity rate is 95-98% while mortality rate in uncomplicated cases is low. In foals the disease is more severe. The Symptoms are rise in body temperature, loss of appetite, depression, increased respiiatory and pulse rate with nasal discharge and cough. Pneumonia occurs frequently in foals. The typical pathogenic lesion is bronchiolitis in which bronchioles are filled first with serous and then with mucoid discharge.
Diagnosis: The diagnosis can be made from the symptoms and rapid spread of disease. The virus can be isolated in embryonating chicken eggs and in primary cultures of equine or monkey kidney cells from nasopharyngeal swabs during 48-72 hours of illness. The HA test is applied for detection and HI test for identification of virus. A serologic diagnosis can be made by detection of four fold increase in antibody titres between acute and convalescent sera by CF and HI tests.
Control: After infection the immunity to homologous virus may persist upto 2 years. Inactivated virus vaccines containing both serotypes mixed with oil adjuvant are available. Foals at 3-4 months are vaccinated and second vaccination is given after 2-6 weeks followed by a booster at 1 year of age. More frequent vaccination is advocated for animals at risk.
Swine Influenza Virus
The swine influenza is endemic throughout the world. The first outbreak was reported in 1918 from USA when most serious human influenza pandemic of the century was going on. The swine influenza and human influenza of 1918 share the same haemagglutining antigen<Rt). The pig pandemic of influenza might have resulted from the host adaptation by the human HI virus. There are evidences of intercbange of HI viruses betwen turkeys and pigs in France. The zoonotic nature of swine influenza has been confmned when influenaza

Orthomyxoviridae 281
virus in 1976 was simultaneouly isolated from infected pigs and from a man in USA with clinical signs of influenza who had been attending the pigs.
Swine influenza is 'A' type virus. The swine influenza has its own haemagglutinin but its neuraminidase is of human subtype (HI NI). Like all influenza viruses the swine influenza virus undergoes antigenic changes. The virus readily grows in chicken embryos and can be propagated in pig embryo kidney besides cell lines of canine and human origin. The virus haemagglutinates and haemadsorbs the red blood cells of several species.
The disease occurs naturally as a herd disease during autumn, winter and early spring. The incllbation period is 2-7 days. The mortality ·rate is 1-2 per cent but morbidity rate is high. The virus causes a'mild upper respiratory tract disease but the disease is severe in association with secondary invaders like Haemophilus suis or other bacteria. The animals generally recover in 4-6 days but with a considerable loss of weight. The virus is transmitted by droplet infection. The idea that pigs acquire infection by eating earthworms containing virus infected larvae of pig lung worm appears to be questionable. It appears that virus may be latent in some pigs. The disease can be diagnosed by isolating the virus from diseased pigs. The vaccination is not practised.
Avian Influenza Virus
Numerous type A influenza viruses with HI_13 NI_, have been isolated from the faeces of wild and domestic birds. Isolates of the avian influeza virus vary in pathogenicity from rapidly fatal disease of fowl plaque to mild air sac disease and drop in egg production. Acute disease of influenza is termed as fowl plague. The role of avian influenza in public health is undefined. There is a possibility that recombinational events could occur among many of the viru~s
circulating in brids. This mixture could result in emergence of viral hybrids infectious for avian and mammalian species.
Fowl Plague Virus
The virus causes an acute fatal disease in several avian species like chickens, turkeys, pheasants, terns and probably other wild birds. The disease is enzootic in United Arab Republic, North Africa and some

282 Textbook o/Veterinary Virology
countires of Europe. The disease has not been reported from India. Properties of the virus: The disease is usually caused by the
viruses carrying H, or H, antigens. Viruses are thought to be introduced into domestic flocks by wild brids e.g. migrating ducks, pheasants, starlings or terns. These viruses increase in virulence during chicken to chicken or turkey to turkey passage.
The fowl plaque virus can be cultivated in chicken embryos and chicken embryo fibroblasts. Some of the strains replicate in chicken and monkey kidney cells.
Epidemiology: The virus excretion ~curs prior to clinical disease in birds. Virus survives in fomites and in dead birds upto 15 weeks at 4°C. Spread within farm is direct while between farms is indirect via infected fomites. Wild birds are considered to introduce viruses into poultry flocks. The spread in wild birds is aided by water or by clonial testing of terns.
Pathogenesis: The virus enters via nasal epithelium and replicates to high titre in every tissue of body with resultant necrosis and haemorrhages. The incubation period is 3-7 days. The disease has a rapid onset and runs a rapid course. The affected birds are lethargic with nasal discharges, oedema of head and neck, respiratory signs, gasping and diarrhoea. The combs :md wattles become cynotic. The post mortem lesions may be similar to Newcastle disease which include haemorrhages on proventriculis, haemorrhages in serosal surfaces, swollen kidneys and caseous bronchial plugs. The mortality rate varies from 40-100%. The mortality among younger birds is higher.
Diagnosis: The confirmation of diagnosis can be done by isolating the virus in the allantoic cavity of embryonated eggs. HI and VN tests are applied for identification of the virus.
Control: Slaughter and disinfection of premises should be done Inactivated vaccines are not satisfactory, chicken embryo attenuated vaccines have been used with sQme success.
References
EASTERDAY, B.c., 1975. Swine influenza. In Diseases of swine, 4th Ed. Edited by H.W. Dunne and A.D. Leman, Ames, Iowa, Iowa State University Press, 1975
EASlF..RDAY, B.C. and TUMOVA, B. 1978. Avian influenza. In Diseases of poulJ.--y.

Orthomyxoviridae 283
7th Ed. Editl!d by M.S. Hofstad et. a!. Ames, Iowa, Iowa State University Press
MOIIAL'iTY, S.B. and DlTITA, S.K., 1981. Veterinary Virology Philadelphia, Lea and Febiger.
RUSSELL, P.H. and EDINGTON, N., 1985. Veterinary viruses. Cambirdge, Foxton, Burlington Press.

Chapter 27
Paramyxoviridae
The viruses in this family are usually spherical with 100-150 nm in diameter. The filamentons forms of several micrometers in length also occur. There is lipid bilayer which contains matrix proteins. It is trnnsversed by glycoprotein spikes. The envelope has two glycoproteins which comprise separate spikes, the haemagglutinatioin (H) and fusion (F) proteins. The genome is single stranded unsegemented RNA with a molecular wight of 5.8 x 1()6 daltons. The RNA is negative sense. The viruses are relatively heat stable and pH st.'lble. The viruses enter the cell by fusion of their envelope with the cell surface membrane. The neucleocapsid is the functional tempelate for transcription of complementary viral mRNA and for RNA replication. The assembled nucleocapsids are enveloped at the cell surface at sites at which the viral surface proteins (envelope) have been inserted. Virions leave the cell by budding. The viruses grow in chicken embryos and tissue culture. The CPE is characterised by syncytial formation and inclusion bodies, death of the cells and detachment. The paramyxoviridae family contains the following 3 genera:
1. Para myxovirus- Virions are pleomorphic but usually spherical with about 150 nm in diameter. The envelope is derived from cellular lipids and contains viral glycoproteins at the outer surface and nonglycosylated inner protein. The viral envelope bears two types of projections, the larger one (HN) responsible for haemagglutinating and neuraminidase activities while the smaller type (F) causes cell fusion and haemolysis.

Paramyxoviridae 285
2. Morbilivirus- The envelope contains haemagglutinin but not neuraminidase activity.
3. Pneumovirus- The virions contain neither haemagglutinin nor neuraminidase.
Avian Paramyxoviruses
There are nine serotypes of avian paramyxoviruses of which paramyxovirus 1 or Newcastle disease virus is most important. However, paramyxovirus 3 also causes clinical symptoms and drop in egg production in pOUltry.
Avian Paramyxovirus or Newcastle Disease Virus (NDV)
Newcastle disease is also known as Ranikhet disease in India. Virus causes a world wide disease of birds e.g. chickens turkeys, guinea fowl, pheasant'> and pigeons. Man is susceptible and suffers from self limiting conjunctivitis. Different isolates of virus cause a spectrum of disease ranging from a highly lethal disease to subclinical respiratory tract infection. This depends upon the virulence of the virus strain. The original description of the disease by Kraneveld (1926) and Doyle (1927) related to the virulent form of disease while Beach (1944) reported that a mild strain of the virus causes avian pneumoencephalitis. The disease was recognised for the first time in Batavia (now Djakerta), Indonesia (Kraneveld, 1926). In the same year in autumn the disease was reported by Doyle (1927) at Newcastle upon-Tyne, England. The disease in India was reported in 1927 at Ranikhet Kumaon hills by Edwards (1927). Today the disease is'almost world wide in distribution except in the Scandinavian countries of Denmark, Norway, Sweden and Finland.
Properties of the virus: The mature virus particles are relatively large, 100-250 nm. The envelope of NDV has haemagglutinin (H), neuraminidase (N) and fusion protein (F) spkies. The' virus is inactivated at 55°C within 45 minutes and at 100°C within 1 minute. The sunlight inactivates the virus within 30 min. The infectivity remains unchanged at 4°C for a few days. The virus survives jn frozen poUltry carcases for more than 2 years. NDV is more su.sccptible 10 the action of alkali than acid. A solution of sodium hydroxide (2-5%) is used for cleaning the buildings and for disinfcclion of wooden poUltry crates or similar equipment. All strains of the virus agglutinate red

286 Textbook o/Veterinary Virology
blood cells of a number of mammalian and avian species but chicken, guinea pig and human'O' erythrocytes are most frequently used. After haemagglutination the elution of red cells takes place. The virus also possess haemolysin which haemolyses fowl red blood cells. The property of haemadsorption is also shown by the infected celis with the virus. The specific antiserum inhibits haemagglutination, haemadsorption and haemolysis.
Cultivation: The virus is readily cultivated in chicken embryos. The most common route of inoculation is by "allantoic cavity route. Most of the strains kill embryos between 24-72 hours causing heamorrhagic lesions and encephalitis in chicken embryos. The maximum titre of about 1O-9EIDso per 0.1 ml is obtained belween 24-36 hours of inoculation. The virus also replicates in many cell culture systems like chicken embryo fibroblast, chicken embryo kidney, chicken, rabbit, pig, calf and monkey kidney and BHK-21 and HeLa cells. Polykaryocytes are produced in infected cells which may become chronically infected. The NDV is a potent inducer of interferon.
Epidemiology: Transmission is by droplet, direct contact, fomites or the ingestion of excreted virus. Excretion can occur for at least two months following infection of partially immune birds. The disease spreads between farms or countries because virus can survive for several months in either poultry carcases and eggs in dried material. The virus has been isolated from variety of wild birds. The caged birds have been incriminated in the introduction of exotic strains of NDV. Migratory birds and scavenger birds appear to be of little importance in the spread of ND. It is probable that wild birds act as mechanical carriers. Dried material may be transmitted on dothing or as wind borne dust. There is minor serologi.i:al variation between isolates obtained from different outbreaks as shown by serum neturalizatipn, haemagglutination inhibition and neuraminidase inhibition tests, one of these differences indicate a significant antigenic variation which should have a bearing on practical immunology. Therefore, there is no evidence to date to indicate that the field virus is not immunologically different from vaccine strains of virus.
Pathogenecity: The virus mainly infects chickens and turkeys but is capable of infecting a variety of wild and domestic brids. The average incubation period is 5-6 days but it may vary from 2-15 days. In severe outbreaks the symptoms appe.ar within 3 days. The

Paramyxoviridae 287
pathogenicity is largely detennined by the viral strains and age of chickens. According to pathogenicity three types of strains have been described velogenic (highly virulent), mesogenic (moderately virulent) and lentogenic (avirulent). The classification of three types of strains is based on the three techniques.
1. The 'Mean Death Time' of infected 10 day old embryonated eggs.
2. Intracerebral pathogenicity index in day old chicks. 3. Intravenous pathogenicity index in 6 week old chicks.
A large number of comparative studies have been carried out to find strain differences in relation to virulenc~. Two parameters namely, the capacity to induce cell fusion and to form plaques emerged as factors related to virulence. This led to search systematically for structural and functional differences in fusion glycoproteins between virulent and avirulent strains.
Four distinct form of disease have been described -1. Doyle's (Asiatic) form: This is an acute fatal disease of chickens
of all ages casued by velogenic viscerotropic strains. The clinical signs are hypcrpnea, blood-stained diarrhoea, dehydration, tremors, torticillis and paralysis of legs or wings. Haemorrhagic lesions are prominent in the digestive tract. The mortality is close to 90%.
2. Beach's (Pneumoellcephalitis) form: An acute infection caused by velogenic, neutrotropic strains. The clinical signs are respiratory distress, coughing, gasping, lowered egg production and paralysis. Haemorrhages are absent from digestive tract but lesions arc present in respiratory tract and eNS. The mortality is 10-50% but may be higher upto 90% in young chickens.
3. Beaudelle's form: It is an acute respiratory and occasionally lethal nervous infection of young chickens caused by mesogenic strains. The clinical signs are coughing, anorexia and lowered egg production but mortality is rare.
4. Hitcher's form: A mild inapparent respiratory infection caused by lentogenic strains.
The virus causes mild influenza like signs and conjunctivitis in human beings. In this country the disease produced is by velogenic strains and is of Doyle's form.
Diagnosis: A presumptive diagnosis is possible on the basis of

288 Textbook o/Veterinary Virology
clinical signs in severe fonns of ND but milder fonns may be confused with other respiratory disease. The conftnnatory diagnosis can be arrived at by isolating the virus from the blood, brain, spleen, lungs and other tissues of diseased birds. The virus is easily isolated in 9-12 days old chicken embryos by allantoic cavity route. The chicken embryo ftbroblast and kidney cell cultures have also been used. The harvested material which shows haemagglutination is conftrmed by haemagglutination inhibition or serum neutralization test The sera of recovered birds reveal antibodies in the haemagglutination inhibition test.
Control: In countries where the disease is unknown, the slaughter policy is followed. In Australia the slaughter policy has proved effective. Most countries where disease occurs have adopted a vaccination policy as being the most practical method of control. Live and inactivated vaccines are used.
Inactivated vaccines: The virulent virus strains are used to produce inactivated vaccincs~ The virus is inactivated without destroying its antigenecity. The inactivating agents used are ultraviolet radiation, formalin, chloroform of ~-propiolactone. The inactivating agent not only inactivates NDV but also other viruses pathogenic for poultry. The inactivated vaccine is safe and is not capable of initiating infection or spread of virus. It is suitable for young chicks as well as laying birds and chicks in poor health. The duration of immunity is short and is usually 6 months. The adjuvants like aluminium hydroxide, mineral oil or vegetable oil emulsion are incorporated to prolong immunity. The standard programme of vaccination is ftrst dose at 21 days of age; second one at 8-10 week; third one at 16-20 weeks followed by another at 5 months.
Live Vaccines
Live Lentogenic strains: The known strains F., Hitchner Bp La Sota are used. The optimum dose is 1()6-' to 107EID~ per bird.
Live mesogenic strains: The known strains used arc Roakin, Komorov, Hertfordshire, Mukteswar (R1B). The optimum dose is about 100EID~. The mesogenic strains produce a long lasting immunity but cannot be administered in chicks below 6-8 weekS old. The lentogenic strains are usually administered by instilling a drop of vaccine either in eye or in nostril or as aerosal or through drinking water in day old

Paramyxoviridae 289
chicks. This is followed by a mesogenic strain. The mesogenic strain is given by intramuscular route or by wing web method. In India lentogenic F strain is given in day old chicks by intranasal or intraocular route followed by mesogenic strain Mukteswar (R1B) at 6-8 weeks old by intramuscular or wing web method. The necessity to revaccinate the birds again does not arise a~ the vaccine gives sufficient immunity to birds in productive life. Although the live vaccines especially the mesogenic strains give a long lasting immunity but it also perpetuates the presence of Newcastle disease virus and is considered to be responsible for the higher incidence of chronic respiratory disease and other respiratory disorders activating potential pathogens present in the flock at vaccination time.
Avian Paramyxovirus-3
This virus is prevalent in Europe & USA and causes drop in egg production, respiratory disease and stunting of turkeys. The virus evokes a low level of cross-protection and cross reactive HI antibodies toNDV.
Mammalian Parainnuenza Viruses
The parainOuenza viruses generally replicate. in the upper respiratory tract and produce mild disease. The immune response is poor and reinfection usually follows. The parainfluenza virus (PI) has 5 serotypes.
Parainnueza-l (Sendai Virus)
The virus is associated with the upper respiratory tract infection and results is chronic ill health and breeding problems in most of the mouse colonies. The virus haemagglutinates and haemadsorbs a variety of rbc and possesses a haemolysin active against chickens and guinea pig rbc. The virus can be isolated in a variety of cell cultures and embryonating chicken eggs from the lungs of infected mouse. Experimental mouse to mouse passage of virus increases its virulence which can cause fatal pneumonia in young mice.
Parainnuenza-3 Virus in Cattle
The infection of the Parainflueza-3 (PI-3) is widespread in cattle population throughout the world. About 80-90% of cattle have HI antibody. Repeated subclinical infections occur when the virus is

290 Textbook o/Veterinary Virology
associated with bacteria and stress. PI3 has also been associated with pleural lesions of sheep.
Properties 0/ the virus: The structure, physical and chemical properties pf bovine parainflueza types 3 virus are the same as other members of this genus. The important viral proteins are HN and F proteins. Various parainfluenza virus type 3 (pI3) strains differ in neuraminidase activity, some strains being strong while others weak in neuraminidase activity. PI-3 haemagglutinates and haemadsorbs rbe of certain species of animals including bovine rbc. The PI3 virus strains grow in cells of bovine porcine and human origin. The ePE is characterised by plaque formation, syncytia and eosinophilic inclusion body formation. Variations in cytopathogenicity, plaque morphology and haemagglutinating activity and syncytium activity have been reported with various PI3 virus strains. No anti genic differences have been reported between different strains isolated from different countries. With monoclonal antibodies variation in the structure of viral envelope proteins have been observed in some bovine PI3 virus strains and these differences might be related to viral pathogenicity.
Epidemiology: PI3 virus was first isolated from nasal mucus of cattle suffering from clinical signs of shipping fever in 1959. The distribution of PI3 is now world wide. Most reports of PI3 association with respiratory disease are from young cattle while reports from adult cattle have been much less. The bovine PI3 infections are not always associated with disease. The subclinical infections often take place and are common during winter months. Most calves are infected by respiratory route. The virus multiplies to a high titre in the upper and lower respiratory tract and large quantities of virus arc excreted in nasal mucous, ocular secretions and in droplets. The virus is very stable in aerosals of nasal secretions when the temperature is low. The infection has been demonstrated in the respiratory tract of calves aged few days, removed from dams shortly after their birth and reared in isolation. It is thought- that the source of this early caltbood infection may have occured in utero or shortly after birth. Experimentally it is possible to establish infection of bovine foetus by direct intrafetal inoculation. The possibility of cross-infections between cattle and sheep exists in nature. The cross infection between calves and man is not considered important. The PI3 once establishes infection, spreads quickly in

Paramyxoviridae 291
susceptible young cattle in close contact in poorly ventilated houses. These conditions are frequently encountered in modem intensive calf rearing systems.
Pathogenecity: PI3 virus enters the animal body via the respiratory tract. Experimentally cattle of all ages can be infected. The infection cannot be established with neuraminidase weak strains of the virus. Once the infection is established in the respiratory tract, viraemia occur with transient location of virus outside respiratory tract. Occassionally virus infection has ~n associated with systemic lesions such as splenitis, or enteritis. The virus, however, is a primary respiratory pathogen and produce severe effect on respiratory tract epithelial cells. The glycoproteins of mucus secretions of the respiratory tract are rich in N-actyl-neuraminic acid (NANA) for which paramyxoviruses have affinity. The mucus gJycoproteins thus act as receptors for PI3 virus. After viral adsorption to rcceptor site, viral neuraminidase splits off NANA from the mucous glycoproteins and virus free to attach to the next rcceptor bound NANA. The virus replicates in the epithelial cells of upper and lower respiratory tract and in alveolar macrophages. The replication of PI3 virus r;auses epithelial changes, including hyperplasia and hyperplasia of'type II pneumocytes. This leads to thickening of the respiratory barrier by cuboidal epithelization of alveolar walls. -The amount of damage caused on the respiratory tract by virus depends on the virulence of the infecting strain, the immune status of animal and on environmental stress factors.
Primary infection of seronegative calves results in virus shedding. Calves with maternal antibody also shed virus. The shedding is usually for 8-10 days. In natural infection the virus has been found in the respiratory tract of calves over a period of months. In natural outbreak, the PI3 virus infection is accompanied by other microorganisms including other respiratory viruses, mycoplasma and bacteria. The most important respiratory disease of PI3 virus is enzootic pneumonia in calves and shipping fever. In enzootic pneumonia in calves there is high morbidity and a mortality approaching 10% has been recorded. The survivors have a permanent lung damage with less food conversion efficiency and light weight. In shipping fever there is depression, anorexia, fever rhinitis and nasal discharge, diarrhoea occurs in few animals. Morbidity rate varies from 444% and mortality upto 20%.

292 Textbook o/Veterinary Virology
Immune reactions: The seronegative calves develop humoral and local virus neutralizing antibody to PI3 infection. IgM and IgG are predominatly the serum antibodies while IgA resides mainly in the nasal tract. The serum antibodies appears 6 days post infection and persist at high titre upto 3-5 months. The maternal antibodies in calves persist upto 10 week~ of age. CMI also participates in the immune response. The relative importance of CMI compared with antibody mediated immunity in protection against PI3 disease virus is not known.
Diagnosis: The diagnosis can be established by taking nasal samples for antibody detection and necropsy material for histopathological examination. The nasal swabs should be collected at an early stage of disease. The nasal swabs is an excellent material-for viral isolation as well as for rapid diagnosis by IF because nasal mucus samples provide excellent material for IF. In PI3 infection outbreak a four fold or greater rise of serum antibody titre is detected. The commonly used serological tests are HI, SN, indirect IF and ELISA. In the histopathological examination eosinophilic intracytoplasmic inclusion bodies~ presence of epithelial. syncytia on bronchiolar and alveolar walls is often accompanied by hyperplasia of alveolar epithelium. The viral antigen in lung tissue can be detected by direct or indirect IF.
Control: Overcrowding should be avoided. Both inactivated and modified live vaccines are available. Inactivated vaccine stimulate systemic and local antibody production and reduce virus excretion on challenge. The modified live vaccines are given by intranasal or intramuscular route, also result in the production of systemic and local secretory antibody. Vaccinal virus may be shed from nasal passages. Inactivated vaccines are frequently combined with inactivated IBR vaccine and Pasturella bacterins. Two injections are usually recommended.
Parainfluenza-3 Virus in Sheep
Only one serotype Qf PI3 virus has been recognised. The virus is antigenically related to both bovine and human PI3 but can be distinghished by serological methods. Infections of sheep with PI3 virus are common. The antibodies in sera of sheep are prevalent in several countries throughout the world and percentage of prevalence is

Paramyxoviridae 293
over 70%. The maternal antibodies in lambs prevent the infection. The maternal antibodies wane rapidly and animals become susceptible within a year of age. Transmission of virus between animals is due 10
respiratory excretion. It is not clear as to how the virus is maintained in a flock, it may be due 10 viral persistence. It is known that majority of infection are mild in nature but severe outbreaks have also been recorded. The role of PI3 virus in these outbreaks is confounded by the dominating role of Pasteurella haemolytica. Experimentally it has been observed that virus may initiate the disease process and predisposes 10
Past. haemolytica. The infected lambs killed at different intervals show the presence
of viral antigen in the bronchioles and surrounding alveoli. The lesions are formation of inclusion bodies and bronchiolar epithelial hyperplasia followed by necrosis and desquamation. The development of these lesions is associated with appearance of illness in the affected animals. The clinical signs regress between 6-7 days of PI when the antibodies appear. The epithelial destruction and accumulation of debris in the lower respiratory tract provides a focus in which secondary bacteria localize and proliferate. The PI3 infection predisposes to lambs 10
severe pneumonia caused by several serotypes of P. haemolytica biotype A.
Most PI3 virus infections are subclinical but occasionaly acute outbreaks of disease occur.
Immunity to PI3 virus can be produced by local or parenteral administration of PI3 antigens, which prevent virus replication and development of lesions.
Parainfluenza-5
Parainluenza-5 (PI-5) is associated with Kennel cough. The virus in severe cases is associated with canine distemper virus or Bordetella bronchiseptica. Most dogs have antibodies 10 PI5 without having clinical disease.
MORBILIVIRUS
The genus includes the triad measles, distemper and rinderpest viruses. All these viruses possess 'H' antigens but measles virus haemagglutinates rbc, none of three viruses possess neuraminidase antigens. These viruses share some (F) antigens which may be cross

294 Textbook o/Veterinary Virology
protective. Antigens differ in three viruses. The H and F antigens occur on separate peplomers.
Canine Distemper Virus (CDV) Hard pad disease
The canine distemper is a highly contagious disease of dogs with a heavy death rate in young susceptible dogs. Prior to the development of vaccination this was the major pathogen of dogs. The disease occurs naturally in dogs (Canidae family) mink, ferrets and skunks. The members of family felidae (cat, lions and tigers) are not susceptible. The disease is world wide in distribution. Laidlaw and Dunkin in 1926 confinned the viral aetiology of the disease. Till then, Bordelella bronchiseplica was considered to be the primary aetiological agent.
Properties 0/ the virus: The morphology is similar to other members of the genus. The virus particle measures 90-250 nm in diameter. Thermal inactivation of the virus is rapid. The virus is rapidly destroyed if stored above O°C. But survives for months at -lOoC. It is inactivated by ether, 0.1 % formalin and 1 % lysol. An irregular partial haemagglutination of chicken and guinea pig rbe has been reported in high concentration of egg adapted virus but this property is variable. The CDY forms are triad with measles and rinderpest virus, the last two viruses provide protection against CDV. Currently, only one antigenic strain is recognised ·although it is well established that strains vary in virulence. The pneumo, neutrotropic and epidermal strains (causing Hard Pad) have been recognised. The variation in virulence exists in measles and rinderpest viruses as well. In measles virus, antigenic variation between haemagglutinins of different isolates have been detected by monoclonal antibodies. It is thus likely that antigenic variation also exists between isolates of distemper virus while the immunodominant epitopes remain cross protective.
The virus can be adapted to grow in un weaned mice, baby hamsters and rabbits. Green (1939) by successive passages of virus in ferrets discovered that the virus was reduced in virulence for dogs while the virulence for ferrets was enhanced. This led to the discovery of live modified vaccine against distemper in dogs. The ferret adapted strains can be further adapted in chicken embryos. After about ten passages the virus produces thickening of the CAM. After about ioo passages in chicken embryos, the virus becomes attenuated for dogs and ferrets and can be used as vaccines. The virus replicates in primary

Paramyxoviridae 295
and continuous dog and ferret kidney cells and in chicken embryo fibroblasts. The CPE is typical of the group which includes granular degeneration and vaculation of cells, formation of giant cells and syncytia with cytoplasmic and sometimes nuclear inclusion bodies. Attenuation of the virus occurs after repeated passages.
Epidemiology: Distemper is a highly contagious disease. 'The portal of entry is the respiratory tract. Transmission is both by droplet infection and direct contact. The virus is shed in excretions and secretions. The dog is the principal reservoir of the virus. The virus appears in the urine upto 8th week post inoculation. Mortality is variable according to the virulence of the strain (average is 20%), sometimes mortality reaches 90%. Maternal antibody is transferred by colostrum to the puppies and prevents infection usually upto 8 weeks.
Pathogenesis: The disease in nature occurs in dogs, wolves, foxes, coyotes, ferrets, raccoons; minks, weasels and dingos. It primarily attacks puppies and seldom older dogs, because of natural immunity. The incubation period is 3-8 days. The clinical signs are diphasic fever, coryza conjunctivitis, viraemia, leukopenia, followed by Icukocytosis, severe gastroenteritis, respiratory distress, vesicular and pustular dermatitis and hardened foot pads. Some dogs have nervous manifestations like myalgia, incordination, convulsions and coma. The nervous signs usually appear after the second febrile phase and are generally followed shortly by death. The pattern of distemper is largely dominated by the immune status of dog population in a given locality at one time.
Haemorrhagic enteritis is common in puppies under 8 weeks of age. The epithelium of bladder and pelvis is congested and large intestine has excessive mucus. The lungs are generally congested and pneumonic. The spleen may be enlarged and necrotic areas seen in liver. Perivascular cuffing, neuronal degeneration and demyelination occur in dogs with neurologic signs. Intracytoplasmic and sometimes intranuclear inclusion bodies may be found in the bladder, renal pelvis, epithelial cells of respiratory tract, intestine and brain.
Diagnosis: A provisional diagnosis can be made from the clinical symptoms which include respiratory symptoms, diarrhoea, catarrhal discharges from eyes and nose, hyperkeratosis of foot pad and nervous signs. Intracytoplasmic inclusions may be found in stained conjunctival smears. On postmortem examination inclusion bodies may be seen in

296 TexJbook o/Veterinary Virology
bronchial epithelium. bladder epithelium and lung macrophages. A confmnatory diagnosis is made by isolation of virus from blood. excretions and secretions. lymphnodes. liver. spleen. lungs and brain of dogs with nervous mainfestations. Susceptible dogs and ferrets are used for isolations. Virus isolation in canine or ferret kidneys can be made. Detection of specific distemper antigens using immunofluorescence. complement fixation or gel diffusion test with hyperimmune antidistemper or antirinderpest sera.
Control: In most animals long lasting immunity develops after recovery from natural infection. Maternal antibody is transferred to
puppies via colostrUm and usually prevents infection upto 8 weeks or occasionally upto 12 weeks. The maternal antibodies interfere with active immunisation . .The puppies cannot be immunized until they loose maternal antibodies. Earlier inactivated vaccines were used followed by virulent virus or by simulataneous antiserum and virus or by ferret attenuated live virus. The current vaccines arc attenualCd egg passaged virus and then grown in cell culture. These vaccines evoke antibodies both against F&H antigens and give strong immunity. These vaccines are usually given in puppies more than 12 weeks of age. Combined CDV and infectious canine hepatitis vaccines arc also available. The attenuated measles vaccine has been used in puppies of 3-6 weeks of age. The measles virus stops symptoms of distemper to develop but may not prevent the infection of CDV.
Rinderpest Virus
Rinderpest generally known as cattle plague is one of the most dreaded disease of ruminants (mainly cattle and buffaloes) and of the swine characterized by ulceration of mucous membranes of the mouth. erosion and necrosis of the mucous membranes of the digestive tract with diarrhoea and lachrymal discharges.
Rinderpest is believed to have originated from China in pre-Roman times. The disease was spread to Near East and Europe with the stock brought in by invading forces. The disease was introduced in Africa in 1842 with cattle imported to Egypt from Romania. Towards the end of 19th century. the disease was introduced from India through the horn of Africa and within few years swept through 90% of cattle population south of Sahara. The economic importance of this disease was responsible for establishment of veterinary services by colonial powers

Paramyxoviridae 297
at home and in their colonies, including first Veterinary school at Lyon in 1764 and the international office of Epizootics in Paris. In India the disease must have been prevalent through centuries. In view of serious losses a cattle plague commission was appointed by the Government of India in 1869. On the recommendations of the commission the Imperial Bacteriological Laboratory (now Indian Veterinary Research Institute) was set up in 1889 at Mukteswar, Kumaon.
The disease has been eradicated by slaughter policy in all European countries, Southern Africa, Australia and Brazil. Eradication plans were undertaken in India and in Africa and were successful in bringing the disease under control. Before the eradication plan 'in India there used to be 8000 outbreaks and 2,00,000 deaths in cattle which has come down to 1200 deaths in 1983 and 147 outbreaks. In spite of the concrete efforts the disease still occurs in certain countries of Africa and Asia in cattle, sheep and goats and in wild life. Th~ countries where the disease is still prevalent are India, Mali, Upper Volta, Niger, Benin, Nigeria, Cameroons, Chad, Sudan, Egypt, Ethopia, Uganda, Kenya, Tanzania, Turkey, Syria, Iran, Saudi Arabia and Kuwait.
Properties: The ultramicroscopic nature of rinderpest virus was demonstrated in 1902 by Curasson when cattle were infected with bacteria-free filtrate. The virus particle is spherical or ovoid in morphology and measures 90-250 nm in diamater, some filamentous and elongated forms vary upto 500-100 nm. Most viral particles are bound externally by a membrane with large projections. The virus is relatively fragile. The infectivity of cell culture preparations has a half life of 1-3 hours at 37°C and only few minutes at 56°C. The virus is stable between pH 7.2 and 7.9 but can survive between pH 4 to pH 10.2. The virus preparations are best preserved by lympholization or by addition of 2% dimethyl sulphoxide and stored at 4°C. The virus is sensitive to ether, chloroform and trypsin. Various chemicals like-f3-propiolactone, hydroxylamine, phenol, formalin inactivate the virus. The virus also gets inactivated with prolonged treatment with glycerine. The virus is rapidly inactivated by putrefaction. There are reportS that rinderpest virus haemagglutinates rbc of rodents, rabbits and monkeys. DK strain of rinderpest virus treated with Tween 80 and ether agglutinated red cells from rodents, rabbits and monkeys. The titres were low but reaction was inhibited by hyperimmune rinderpest antiserum. The haemadosorption with rinderpest infected cells has not been reported so far.

298 Texlbook o/Veterinary Virology
Strains of rinderpest virus are considered to be immunologically homogenous. The attenuated vaccine strains like caprinised, lapinised, avianised and tissue culture adapted strains protect the animals against natural outbreaks as well as experimental challenge with any virulent strain of.rinderpest virus. The rinderpest virus is antigenically related to distemper and measles virus. It has been reported that the three viruses have immunologically identical nucleocapsids and share envelope antigens. The serological relationship between peste des petits (PPR) and rinderpest viruses is close but the viruses are not identical. The cross neutralization tesl~ diffe.renliate them. PPR virus protects cattle against rinderpest and rinderpest virus protects sheep and goats against PPR. There are two soluble antigens apart from infectious rinderpest virus particle. The soluble antigens are complement fixing and precipitinogen. The complement fixing antigen survives boiling for 30 mins. Strains of. rinderpest virus vary in their virulence for particular hosts. The strain 01 isolated from sick elands is avirulent for cattle while a strain isolated from sick giraffe kills upto 84% of inoculated cattle.
Cu/timlion: The virus has been adapted to grow in sheep, goats, rabbits, hamsters and mice. The virus has been adapted to grow in chicken embryos by CAM, intravenous and yolk sac route of inoculation, some strains become attenuated for cattle after 20 passages. The virus grows well in primary or continuous cell cultures and produce a CPE. The susceptible cells are from cattle, sheep, goats, chicken embryos, pigs hamsters dog and man. The CPE is characterised by multinucleated giant cells or syncytia with intranuclear inclusions.
Epidemiology: Natural infections occur in even-toed ungulates belonging to the order Artiodectyla. Cattle and buffaloes are most commonly attached and perpetuate the disease. Epizootics in goats and sheep have been reported from Africa, Asia and Europe and regularly reported from India mostly from the South India. Asiatic domestic pigs frequently suffer and die to rinderpest while pigs of European origin do not suffer although the virus multiplies and is excreted. Free living wild life also suffer and losses are highest in buffalo, eland, giraffe, kudu, warthog, wildbeest in Africa; blackbuck, gaur nilgai and sambhar in Asia. It is possible thal low pathogenic strains of rinderpest virus are being maintained in wild East African Ungulates.
The role of vector in the epidemiology of rinderpest is not

Paramyxoviridae 299
important. The virus may survive for sometime in horse flies, house flies, stable flies, tsetse flies, mosquitoes lice, ticks, leeches and vultures.
Animals of all ages are attacked in virgin-soil epizootics while the disease in enzootic area is characterized by age incidence. The adults and very young calves do not suffer as adults are immune either through natural attack of virus or vaccination, young are immune due to colostrum antibody. Innate resistance varies widely within and between species. In India local plain cattle are less susceptible in comparison to exotic breeds, cross breed caule and hill cattle. The Asiatic pigs have severe clinical reaction and often die, while European pigs, undergo inapparent infection. Species differences have also been observed in cohabitating wild life. The morbidity and mortality are profoundly influenced by innate resistance of the infected host and the virulence of strain. The degree of virulence also varies for particular host. ~Jne attenuated vaccine strains for cattle and buffaloes have proved to be virulent' for wild life. In enzootic areas the mortality rate is around 3% but when exotic animals arc exposed to the same strain the mortlity rate exceeds 80 to 90%.
Most of the strains are catholic and spread rapidly between cattle and buffaloes while a few of the strains are remarkably selective. The rinderpest outbreak may not spread to all susceptible animals in aerosals and contact has to be close as infectious droplets arc large and short lived. Air borne transmission is possible at night at high or low humidities. Transmission through ingestion of contaminated food and water is rare but pigs acquire infection by eating contaminated meat scraps.
Palhogenesis: The virus enters the animal body through mucous membranes of upper respiratory tract The primary multipication occurs in the palatal tonsil or in the pharyngeal or mandibular lymph nodes. After primary multiplication the virus enters in the blood circulation where it attaches to mononuclear cells. The viraemia is detectable -1-2 days before the symptoms appear. Secondary multiplication of virus takes place in all the lymphoid tissues, in mucous membranes of the alimentary and respiratory tracts and in the lungs. The virus does nOl multiply in the brain, kidneys or myocardium. The virus titre is at the peak during prodromal fever and remains in the circulation upto erosive mucosal phase and then falls slowly till the antibodies appear, 4-5 day

,"'"
300 Textbook o/Veterinary Virology
after the onset of fever. The duration of viraemia is influenced by the strain of the virus and innate resistance of the host. The virus is shed in all the secretions and excretions of affected animals, with ocular and nasal secrytions and faeces. The virus is shed throughout the fever period and few days after fever regresses. Recovered animals are not considered as carriers as clear evidence of latency has not been found. The persistent infection of rinderpest virus has been established in Vero cells. This has led to controversy over the possible resistance of carriers in rinderpest virus infection.
The classic rinde~est syndrome is characterized by a short sharp fever, erosive stomatitis, gastroenteritis, dehydration and death. The disease progresses through five distinct clinical phases. The incubation period, the prodromal phase, mucosal or erosive phase, diarrhoeic phase and convalesrence.
The incubation period ranges from 3-15 days depending upon the innate resistance of animals. It is longest in animals having high innate resistance. A sudden onset of fever marks the end of incubation period and start of prodromal phase in which the clinical signs are absent, within 48 hours the signs of fever are mainfested by depression and restlessness; dry muzzle staring coat, serous nasal and lachrymal discharges, congested mucous membranes, anorexia and constipation. There is leucopenia. The mucosal phase begins 2-5 days after the onset of prodromal fever with necrotic pinheads on the mucosa of mouth nostrils and urogenital tract. The necrotic foci are readily abraded leaving erosions with red raw floors. As the disease progresses the erosions enlarge and coalesce, profuse salivation follows and lachrymal discharges. the infected animal becomes restless, drinks copiously but does not eat and passes soft faeces. The diarrhoeic phase starts when the fever falls after two to three days after the appearance of mucosal erosions. The faeces are dark and fetid with mucus and streaks of blood. The profuse diarrhoea causes dehydration; emaciation and prostration. Most of the deaths occur in diarrhoeic phase. The onset of convalescence is ill defined. The recovery to full health takes many weeks. Pregnant aniI1lals abort within six weeks of illness.
Abbarrant clinical forms are not uncommon and range from peracute encephalitis and sudden death to transient subacute diarrh,oea. The clinical picture in buffaloes simulate those in cattle. In pigs after an incubation period of 3-5 days the symptoms are manifested by fever,

Paramyxoviridae 301
inappetance and depression. Within 48 hours the animal starts shivering and becomes prostrate. Vesicles appear around the anus, vomiting and epistaxis takes place. Diarrhoea begins within 48 hours of illness and persists until death or 10-12 days in survivors. In sheep and goats the incubation period is 3-5 days. The clinical symptoms are manifested by sharp fever, there is loss of appetite. :Qiarrhoea begins after 2 days of onset of fever. Prostration and subnormal temperature precede death.
Lesions are moslty found in the digestive tract Erosive lesions are found in the mouth the upper respiratory tract and upper part of oesophagus. In abomasum similar type of lesions also occur in ~yer' s patches. There is a massive destruction of lymphocytes especially in lymphnodes. Congestion, necrosis ar.d erosion occur in the ilea-caecal valve, caecocolic junction and rectum.
Immune reaction: The strains of rinderpest virus are immunologically homogenous and there is complete cross protection. The animals that survive the infection are life long protected although in few animal there may be transient multiplication of virus in tonsillarpharyngeal tissues when exposed to virulent virus. The humoral antibodies, IgM appear after 5-7 days of illness. Within 2 weeks IgM decline and IgG class appear. The IgG antibodies reach peak within one week and then the titre fall back to a threshold level and persist. The reexposure results in anamnestic response. IgA antibodies appear in the respiratory mucus in naturally infected cattle or vaccinated by intranasal route. From immune catlle the calves get the maternal antibodies in the colostrum. The passive protection in the calves in maintained from 4-8 months or sometimes longer. The role of CM! in cattle against rinderpest virus is not understood.
The rinderpest virus is immunosuppressive because it destroys T and B lymphocytes. Due to immunosuppression in rinderpest virus infection the latent infections are activated, which complicates the clinical picture and causes diagnositc confusion. The virus suppresses both humoral and cell mediated mechanism but production of memory cells is not impaired. In rabbits. the destruction of lymphoid tissues by virus triggers autoimmunity. Antinuclear and cold haemagglutining antibodies are produced.
Diagnosis: The clinical symptoms, pattern of the disease and post mortem examination generally lead to strong suspicion of rinderpest. The examination of stained impression smears prepared from the

302 Textbook o/Veterinary Virology
epithelia of tonsils and other lymphoid tissue reveals the presence of syncytia and acidophilic ~nclusion bodies. The method is useful but cannot be used as a routine diagnostic procedure. For conftrmatory diagnosis the laboratory procedures employed are 1) isolation of the virus 2) detection of rinderpest speciftc antigen and 3) demonstration of the development of rinderpest antibodies.
For isolation of virus whole blood in EDT A, lymphnodes, spleen and other tissues are employed. These clinical material is obtained preferably during the prodromal or early mucoid phase of the disease. The material is preferably inoculated in susceptible and known immune cattle. The susceptible cattle will react in 2-10 days time. The material is preferably inoculated in bovine kidney cells cultures. If the virus is present, characteristic CPE develops within 3-12 days, the spccificity of the reaction is conftrmed by means of neutralization tests.
The soluble antigens of rinderpest can be detected by complement ftxation and agar gel diffusion tests. The tissue of choice are fresh carcase or visceral lymph nodes or spleen taken at 3rd to 6th day of pyrexia.
The speciftc neutralizing complement fixing antibodies develop in the sera of canle which develop upto 4th day of the disease or later. Paired serum samples are detected for the presence of antibody. The serum neutralization test is carried out in cell culture or in chicken embryos.
Other tests which are more sensitive and rapid have also been developed like single radial haemolysis test, reverse passive haemagglutination test, fluorescent antibody technique, immunoperoxidase test, enzyme immunosorbent assay etc.
Control: Recovered animals develop a long lasting immunity but it may not be life "long. The rinderpest virus is always introduced into disease free terrilOries by animals having mild infection. The rinderpest free countries ensure the freedom from disease by imposing a complete embargo on the importation of live animals and frozen animal products from countries where disease is prevalent. Virgin outbreaks in disease free countries can be controlled by slaughter of infected and in contact ruminants and swine. Countries where the disease is endemic, regular vaccination is carried out to limit the losses.
The vaccine that revolutionized the control of rinderpest was attenuated goat adapted virus. The vaccine was developed at Indian

Paramyxoviridae 303
Veterinary Research Institue, Mukteswar by Edward in 1928. The vaccine is still being used in India and certain African countries. The goat adapted virus is not attenuated enough and produces severe reaction in susceptible breeds like exotic cattle, hill cattle and certain Japanese breeds. The tissue culture adapted virus vaccine developed by Plowright and Ferris is suitable for all breeds of cattle and other species of animals. The immunity produced by attenuated vaccines are of long duration lasting for about 10 years or more. The efficacy of vaccination is dependent on the variable dUJation of the passive protection conferred by the ingestion of antibodies in the clostrum.
Peste-des-Petits-Ruminants Virus
In early 1940's, a fatal disease of goats with high mortality was described as peste-des-petits-ruminants (PPR) in the Ivory coast in Africa. Momet and his colleagues in 1956 rediscovered the disease and observed that PPR was a strain of rinderpest virus since it had a serologial relationship with rinderpest virus. PPR has since been shown to be a distinct entity having antigenic re,lationship with rinderpest, canine distemper and measles viruses. The disease has synonyms like 'goat plague', erosive stomatitis and enteritis of goats, goat catarrhal fever, 'Kata' and stomatitis peneumonitis complex.
The virus looses its infectivity within 30 min at 50°C. The virus is sensitive to lipid solvents, low pH etc. but in lymph nodes the virus is protected from pH changes after death. The virus does not haemagglutinate rbe but PPR antibodies inhibit haemagglutination of monkey rbe by measles virus. The strains isolated from different parts of Africa are homogenous. Cross-protection has been observed, PPR virus protects cattle against rinderpest virus and rinderpest vaccines protect against PPR.
Natural disease is prevalent in goats and much less in sheep. Cattle in contact with sick goats do not suffer, Experimentally deer and pigs support virus mulliplication but their role in epidemiology of disease is not important. The PPR virus is prevalent in African continent south of Sahara and in Arabian peninsula. The disease outbreaks mostly occurs in West Africa but disease has been reported from Sudan. The disease has also been reporte,d from sheep and goats in Saudi Arabia. Morbidity in goats is high while mortality in clinically affected goats range from 70-80%. The animals get the infection from the sick animal when the

304 Textbook ojVeterinary Virology
animals are concentrated during rain etc. The infection is aerosal in nature. The virus after entry in the upper respiratory tract is disseminated before the onset of clinical signs and virus is shed in nasal secretions, tears, saliva and urine.
After an incubation period of 6 days, sheep and less frequently goats develop subacute infection. The illness is manifested by low glade fever, nasal catarrh, mucosal erosion and intermittent diarrhoea. After 10-14 days the animals usually recover. The acute reactions are observed in goats after incubation period of 3-4 days. Fever and diarrhoea appear but the animals look normal but after few hours the animals show depression. Erosions develop in the mucous membranes lining the upper respiratory, upper alimentary and urogenital tracts after 1-2 days after onset of fever. There is profuse salivation with protruded tongue. There is lot of mucous secretion from the nostrils which often block the nostrils. The mucosal erosions coalesce and produce extensive lesions. Diarrhoea and pneumonia then supervenes. Most of the animals die within 10 days. The incubation period is about 2 days in paracute cases. There is profuse nasal catarrh, high fever, dyspnea, anorexia and constipation. The mucous surfaces are congested and eroded. The diarrhoea starts after about 3 days of onset of clinical symptoms and animal die within 4-6 days after iJIness. Paseurella pneumonia invariably supervenes in actue cases. With the onset of fever there is severe leukopenia. On histopathological examination multinucleated giant cells containing intranuclear and intracytoplasmic inclusions, the syncytia in lung parenchyma is conspicuous.
Surviving goats develop immunity and resist infection and immunity is lifelong. The humoral antibodies can be detected by CF, AGID, measles HI and neutralization test. Clostral immunity protects kids and lambs until they are weaned.
The diagnosis of PPR outbreak is confirmed by detection of antigen in the tissues and isolation of the virus. The histopathology is also useful in the diagnosis. The lymph nodes and tonsils collected from animals killed in the early stage of disease or even after death of animals are used in AGID test or CIEP test against PPR hyperimmune serum. Blood collec~ed over EDTA during febrile stage of the disease or lungs, lymph nodes and tonsils from killed animals during febrile stage are useful for virus isolation. The tissue suspensions are inoculated into primary lamb kidney cells or in known susceptible and

Paramyxoviridae 305
immune goats. The animal inoculation test is quicker. A histopathological examination of early mucosal le$ions, lymphoid tissues and lungs reveal multinucleated giant cells with incusions. The convalesent sera of sheep and goats is used to detect antibodies by CIEP, AGID or neutralization tests. It is difficult to distinguish PPR form rinderpest without laboratory tests or animal inoculations. PPR has been reported from India by Shaila et al. (1989). The rinderpest virus infection is also common in sheep and goalS in India. The importance of laboratory diagnosis in these animals becomes all the more important.
Virulent cell culture passage PPR virus when ~dministered
simultaneously with hyperimmune serum gives durable immunity. An attenuated PPR virus strain has been developed but it is not yet available for vaccination purposes. The cell cultured rinderpest virus vaccine gives protection against PPR and is used widely for immunization but there have been some failures as well.
PNEUMOVIRUS
Hovine Respiratory Syncytial Virus (BRSY)
The antibody to bovine respiratory syncytial virus is widespread in bovine population. The epizootics among bovines of acute viral infections have been reported form Europe, Japan, United States, Australia and North Africa.
Properties: The-morphology is that of paramyxovirus. The virus shows great pleomorphism with a diameter of 80-450 nm. The genome codes for ten proteins. The virus docs not possess active haemagglutinin or neuraminidase and antigenically distinct form morbiliviruses. Bovine RSV is antigenicaIIy related to human RSV but it is not serologically identical. The virus is fragile and can be recovered from clinical specimens in cell culture within one hour of collection of the specimen. The virus grows in the cell culture of bovine origin. Monolayers of bovine testis are very sensitive for virus isolation. The CPE produced ;lre numt:rous syncytia and eosinophilic cytoplasmic inclusions. The virus does not grow in embryonating eggs.
Epidemiology: The presence of virus carriers has not been proven but the disease generally. appears in BRSV free country after introduction of infected cattle. The velerinarians and animal handlers play a role in virus transmission. The disease rapidly spreads from farm

306 Textbook of Veterinary Virology
to farm. The animals especially beef cattle are most susceptible between 3-9 months of age, the younger calves of few weeks can also be affected.
Pathogenesis: Experimental infection with BRSV produces a mild infection, The concomirant presence of BVDV as well as sudden fall in atmospheric pressure or a drop of temperature seem to aggravate the disease. The incubation period is 5-7 days. Pyrexia occurs four days after exposure to virus and is associated with the replication of virus throughout the upper respiratory tract Syncytia formation occurs in the epithelial lining and intracytoplasmic inclusions occur in the infected cells. The mild rhinitis and tracheitis is associated with cough but extensive bronchopneumonia occurs only when secondary bacterial infection takes place. The animals recover within 2 weeks of infection. There are reports ihat BRSV infection can occur in the presence of circulating antibodies, however, the protective effect of nasal neutralizing antibodies against infection has been shown.
The mortality goes upto 20%. The lossess are high among the calves. Surviving calves recover after few days of illness.
Diagnosis: A 4 fold increase in CF or VN antibody titre in paired scrum samples is diagnostic of RSV infections. The virus can be isolated from the early cases. The nasal secretions are the source of virus. Monolayers of bovine testis are incubated with the fresh material. 1be isolation in cell culture is difficult because of late appearance of CPE which ranges from 20 days to 50 days. The earliest CPE noted are small areas where four or five cells are balloolJed with shrinkage of cytoplasm followed by syncytia formation and then become opaque and holes appear in the monolayer. The viral antigen can be detected in nasal mucus and lung tissue by direct IF. FLISA is now applied more extensively for detection of viral antigens.
Control: The management of calves is important. Live vaccine administered parenterally are available. The vaccines do not prevent infection but decrease the severity.
References
ADLAKlIA, S.C., 1982. Rinderpest. Indian Council of Agricultural Research, New Delhi
AI.ExANDER, DJ., 1980. Avian paramyxoviruses. Veterinary Bull. 50: 737-52

Paramyxoviridae 307
Au..AN, W.H; LANCASTER, J.E and Tom, B., 1973. Newcastle disease voccines. FAO
BANSAL, R.P., 1986. Rinderpest and its eradication from India. National symposium on·Animal Health at IVRI, Izatnagar June 24 to June 26, 1986.
BANSAL, R.P., 1986. Diagnosis of rinderpest with special reference to application of modern techniques. Indian Vet. Res. Inst. Mukteswar, 55 pp.
BETfS, AP.; JENNINGS, AR.; OMAR, AR; PAGE, Z.E.; SPENCE, J.B. and WALKER, R.G., 1964. Pneumonia in calves by parainJluenza virus type·3. Vet. Rec. 76, 382-384.
BRYSON, D.G.; EVERMANN, J.F.; LIGGIT, D.; FOREYT, W.J. and BREEZE, R.G., 1988. Studies on patfwgenesis and interspecies transmission of respiratory syncytial virus isolated from sheep.Am. J.Vet. Res. 49: 1424-1430.
BUXTON, .A. and FRASER,. G., 1977. Animal Microbiology Vol.2. Oxford, Blackwell Scientific Publications.
CURASSON, G., 1932. Le Peste Bovine. Vigot Freres, Paris, pp.344
DAVIES, D.H; DUNGWORTII, D.L; HUMPIIREYS, S. and JOIINSON, AJ., 1977. Concurrent infection of lambs with parainJluenza virus type 3 and Pasteurella haemolytica. N.Z. Vet. J. 25,263-265.
FRANK, G.H. and MARSIIALL, R.C., 1973. ParainJluema-3 viruses infection of cattle. J.A Vet: Med. Assoc.163: 858-60.
GILLESPIE, J.H. and CJ\R.\UCHAEL, L.E., 1968. Distemper. In Canine Medicine, Edited by E.J. Contocott, Wheaton, III. American Veterinary Publications.
LANCASTER, J.E., 1981. Newcastle disease. In virus disease of Food Animals. Vol.Il. Edited by E.P.J. Gibbs, Academic Press, London.
McFERRAN, J.B. and McNULTY, M.S., 1981. Aids to diagnosis of virological disease. Br.Vet. J.137, 455-463.
MOHANTY, S.B. and DUTIA, S.K., 1981. Veterinary Virology, Lea and Fibger, Philadelphia.
MOHANTY, S.B. ,1978. Bovine respiratory. viruses. Adv. Vet. Sci. Comp. Med. 22,83-109.

308 Textbook o/Velerinary Virology
NAOAl, Y. and TOYODA, T., 1987. MolecularlJiology of Newcastle disease virw. Prog.Vet. Microbiol. Immunol. 5,16-64.
NAWA1lIB, D.R. and Lm.toRDA, A.G., 1983. Towards global eradication of rinderpest. Rev. Sci. Tech. Off. Int. Epiz 2,998-1012.
PLOWRIGHT, W., 1968. Rinderpest virus. Virol. Monogr. 3,25-110.
RusSEU.. P.H. and EDINOTON, N., 1985. Veterinary viruses. The Burlington Press (Cambridge) Led.Foxton. Cambridge.
REISINGER, R.C.; HEnDLBSTON, K.L. and MANI'HER, C.A., 1959. A myxovirus (SF-4) associated with shipping fever of cattle. J. Am. Vet. Med. Assoc. 135, 147-152:
RYDBIlCK, R; LoVEL, A.; ORVELL, C. and NORBY, E., 1987. Anligenic Analysis of human and bovine parainJluenza type 3 strains with monoclonal antibodies. J. Gen. Viral. 68: 2153-2160.
SCOlT, G.R., 1990. Rinderpest virus. In Z. Dinter and B. Morain (Editors). Virus infections of ruminants. Elsevier Science publishers B.V. Amstardam pp. 341-354.
SCOlT, G.R., 1985. Rinderpest in 1980. In Prog. vet.Microbiol and ImmunoI. Vol. 1 pp. 145-74 Frager, Basel.
SCOlT, G.R., 1981 Rinderpest and peste des petits ruminants. In virus diseases of food animals. Edited by E.P-Gibbs Vol.ll. Academic Press, London.
SHAll-A, M.S ; PRUSHOlTAMMA, V.; BHAvASAR, D.; VENUGOPAL, K. and VENKA'IESAN R.A., (1989). Peste des petits ruminants of sheep in India. Vet. Record 125: 602.
TAYLOR, W.P., 1984. The distribution and epidemiology of pestes-des-petits ruminants. Prev. Vet. Med. 2,157-166.
WELLEMENS, G., 1977. Laboratory diagnosis, methods for bovine respiratory syncytial virus. Vet. Sci. Commun.l, 179-189.
BEACH, J.R., 1944. The neutralization in vitro of avian pneumoencephalitis virus by Newcastle disease immune serum. Science 100: 361-362.
DoYLE, T.M., 1927. A hitherto unrecorded disease of fowls due to a fillerpassing virus. 1. comp. Path. 40, 144-169.
EDWARDS, J.T., 1928. A new fowl disease. Ann. Rep.lmp, Inst. Vet. Res. Mukteswar, 1928. p. 14-15.
KRANEVELD, F.C., 1926. A poultry disease in Dutch East Indies (Dutch). Ned. Ind. B
I, Diergeneesk. 38,448-451.

Chapter 28
Rhabdovjridae
This is a remarkable family of animal viruses for its characteristic morphology and for the wide range of forms of life that are infected from plants to mammals by the members of this family.
Animal rhabdoviruses (from Greek rhabdos means rod) are oblong particles with one rounded and one planar end which gives the virus particle the shape of a bullet. The virus particles are of varying sizes, length 130-380 nm and diameter between 50-95 nm. The lipo-protein envelope contains virus specific peplomeres and has projecting glycoprotein spikes which determine cell tropism and virulence. These projections are 5-10 nm in length and 3 nm in diameter. The nucleocapsid measures about 50 nm in diameter and consists of helical structure (20 x 700 nm) rightly coiled into a cylinder. This gives a characteristic striations of the particle in electron microscopic preparations. The genome consists of one molecule of non infectious linear single stranded RNA of negative polarity with a molecular weight between 3.5 to 4.6 x 1()6 daltons. Virion proteins are associated with the membrane and the nucleocapsid. Five major proteins are possessed by the virus particle. Protein G forms the glycoprotein spikes which determine cell tropism and virulence. Protein M unites with host lipid to form envelope. N and NS proteins form structural components for RNA helix, while L protein is associated with transcriptase. There is also a protein in the envelope associated with haemagglutination of goose erythrocytes. Thin ouier !ay~r is usually derived from host cell membrane either by budding out through p!~ma membrane or into cytoplasmic vesicles. The virus particles are sensitive to UV light but

310 TeXJbook o/Veterinary Virology
survive in dark place at lower temperature. The virions are sensitive to lipid solvents, moderately resistant to heat, acid pH and deter~ts. Members of the family grow in wide range of tissues and prod lice rounding and retraction of cells. Viral replication is entirely cytoplasn:tic with formation of inclusion bodies. The virion transcriptase copies the viral RNA into several positive mRNA which are encountered in polysome complexeS. Virion RNA replication involves an RNA nucleoprotein intermediate. Morphogenesis is by budding from intracytoplasmic or peripheral membranes.
The family of rhabdoviruses is d~vided into two antigenically distinct genera. 1. Vesiculovirus which includes vesicular stomatitis virus and is the only virus in this genus. 2. Lyssavirus-Rabies Virus is the only member of vaterinary importance. The other animal pathogens in this family do not have an established group. The important viruses of veterinary importance are Ephemeral fever virus of cattle and Marburg virus of man and monkeys.
Vesicular Stomatis Virus (VS V)
The virus causes a benign contagious vesicular disease in cattle, pigs, horses, deer and occasionally in man. The disease primarily occurs in Western Hemisphere and is enzootic in parts of North, Central and South America. Occassional outbreaks have been reported in Europe.
Properties 0/ the virus: The virus particles are cylindrical, bullet shaped rods with a diameter of 150-180 x 50-70 nm. The envelope is covered by short spikes about 10 nm long. Truncated virus particles are also present in addition to bullet shaped particles. The virus agglutinates goose rbe at 0-4°C. The virus is resistant to physical and chemical agents. At 4°C in soil it is infective for many days. There are two immuno\ogicaIly distinct types, New Jersey and Indiana identified by neutralization and and complement fixation tests.
There are three subtypes of Indiana VSV. Indiana virus is antigenically diverse. New Jersey virus isolates are antigenically similar irrespective of the host species or the geographic location. Hybridization studies conducted on several antigenically similar New Jersey isolates RNA has shown that New Jersey isolates can be differentiated into two groups having less than 25% homology. More genetic difference exists between two antigenically similar New Jersey

Rhabdoviridae 311
groups than three antigenic subtypes of Indiana Virus. Isolates of both Indiana and New Jersey viruses differ in their physical, biological and genetic properties. The virus grows in the chicken embryos by amniotic, allantoic and yolk sac routes and produce pocks on the chorioallantoic membrane. VSV can be propagated in a wide variety of cell cultures showing rapid CPE. The virus can be adapted to mosquitoes and nearly all mammalian species can be infected.
Epidemiology: The epidemiology of VSV is complex, due to two antigenically distinct viruses, Indiana and New Jersey. Indiana virus has further three distinguishable serotypes. These viruses induce vesicular lesions In mouth and on the feet of cattle, horses and swine. With the exception of white tailed deer, there is no evidence of clinical disease occuring in wild mammals. Disease in man has been reported in individuals working in virus laboratories, clinicians and livestock handlers. The antibodics are prevalent in these persons.
The manner in which thc virus is transmitted form one animal to another is controversial. From experimental studies it is known that virus can be transmiLlcd from infected to susceptible animal by contaminating an abmsion in mucosa of gum or tongue, skin of teat orifice or skin bordering the hoof with saliva or any exudate from a lesion. The infection through intact skin is not established. Insects may introduce vesicular stomatitis virus either as mechanical or biological vectors into susceptible sites of the skin at the teat orifice or within the narrow band of skin adjacent to hoof of cattle. Mouth lesions may be produced as a result of generalization and viraemia.
The virus has been isolated from Aedes and Phelbotomus mosquitoes as they support viral replication and transovariaI transmission occurs in these flies. Apart from insect vectors other modes of transmission may occur.
The natural reservoir is still not known but virus probably persists in one or more unknown hosts. Antibodies against this virus are found in wide variety of wild animals like feral swine, white tailed deer and wild turkeys. The virus may be present in pastures and associated with forage. VSV has been isolated from phlebotomus and culex species of mosquitoes. The mode of transmission vary with different circumstances. The virus is transmitted mechanically by milking machines. The abrasions may be required for contact transmission.
Pathogenecity: VSV produces primarily a disease in horses but is

312 Textbook o/Veterinary Virology
also important in cattle and pigs because the disease resembles FMD. After a short incubation period of 1-3 days, the animals become febrile and viraemic. Vesicles on the tongue, dental pad and buccal mucosa appear with 3 and 5 days on coronary band but teat lesions are rare. Milking cows may get teat and udder lesions which may lead to the development of mastitis. The vesicles rupture quickly and erode the surface epithelium. The ruputure of vesciles is associated with anorexia and excessive salivation. In older animals, more extensive leSions develop. The lesions heal if secondary infection is not serious. There is no mortality but morbidity may be 5-10%. A mild influenza like syndrome has been reported in humans.
Immune reaction: Neutralizing and CF antibodies are detectable in 10-14 days PI. The titres rise upto 4 or 5 weeks and then remain at higher levels fot several months and then decline gradually. Recovered cattle are immune for one year.
Diagnosis: For differentiation of VSV from other vesicular diseases and particularly from FMD, animal inoculation as detailed below is useful.
Table 28.1 ANIMAL INOCUlA nON
Animal Ro.uJe of FMD Vesicular Vesicular Swine inoculation stomatitis Exanthema vescular
disease
Cattle Intradermal tongue + + Horse Intradermal + ±
tongue Swine Intramuscular + + + + Guinea Intradermal + + pig foot pad
The virus can be isolated from vesicular fluid or affected tissues in chicken embryos and cell culture; serologic diagnosis can be made by CF, VN and agar gel diffusion test
Control: Vaccines from vesicular stomatitis virus are primarily used in dairy cattle. Live embryo-attenuated vaccines are used in large scales in certain South American countries and a significant reduction

R habdoviridae 313
in field cases has been observed. The same vaccine when used in swines, vesciular lesions developed and there was shedding of virus.
Rabies virus
Rabies is one of the oldest disease known to mankind. The disease existed as early as 500 BC. The infectious nature of saliva from infected dogs was recognised by Zinke in 1804. Pasteur in 1881 demonstrated the neurotropism of the virus. In 1885 Pasteur used antirabic vaccine successfully. Remlinger in 1903 demonstrated the fitrability of rabies virus and in the same year Negri demonstrated acidophilic intracytoplasmic inclusions called Negri bodies -in the
'.
neuron cells of dogs, cat and rabbits experimentally infected with rabies virus. The disease is prevalent throughout the World except for rabies free areas like Britain, Newzealand, Australia, Hawaii and few other islands. Rabies is an acute, bite transmitted fatal encephalomyelitis of all mammals. The birds can be experimentally infected. The most comtnon reservoir hosts are fox, dogs, wolf, jackal, skunk bats etc., occasional hosts are man, cats, catlle and horse. The bats are the only species in which the virus is not pathogenic. In India rabies problem in man and animals is quite serious. It has been generally observed that 3,00,000 people undergo antirabies treatment annually and 15,000 to 20,000 die. The problem among animals is eSlimated to be 100 times more than in man.
Properties of the virus: Morphology is typical of the family. The virion is bullet shaped measuring 140-180 nm x 75-80 nm in diameter. In some preparations particles are cone shaped. The virus consists of an internal helical, structure called ribonucleoprotein which is surrounded by a, closely associated protein layers called the matrix. The ribonucleoprotein and matrix form the core of the virus particle. The core is surrounded by a membranous envelope which is derived from the host cell. The ribonucleoprotein has 30-35 coils in the form of a cylinder. The rabies virus particles contain about 2-3% RNA, 15-25% lipid and a small amount of carbohydrate, the rest being made up of four major proteins. The proteins are designated as G, N, M and ~ with molecular weight of 80,000, 62,000, 40,000 and 25,000 respectively. There is also a minor protein L of molecular weight 150,000. The protein N is the major protein associated with nucleocapsid, while the spikes or the surface projections consist of

314 TeXlbook of Veterinary Virology
glycoprotein G. The exact positions of ~ and M, proteins are not known. The protein L is probably associated with ribonucleoprotein. The N protein is present in large quantities in cytoplasmic inclusions. The surface projections are important in the attachment of the virus to susceptible cells. They also carry the virus specific antigen which elicits the production of neutralising antibody when injected into the animal. The G protein can fix complement and is responsible for hamagglutinating activity. Like most other enveloped viruses, rabies virus is fragile. It is readily inactivated by organic solvents and detergents and media having pH below 4 or pH above 10. It is sensitive to DV light and heat. Temperature above 56°C destroys the infectivity within minutes. Rabies virus is inactivated by most disinfectant but it is partially resistant to phenol.
Although minor antigenic differences between isolates have been reported, rabies virus was regarded a single serological entity embracing classical virus strains. Derriengue (Vampire bat rabies) which causes a paralytic dumb form of rabies in cattle, Oulofato, a nonfatal encephalitis of dogs in West Africa and Arctic rabies, Nigenain horse virus associated with clinical condition known as 'Staggers' and viruses isolated from shrews in Czechslovakia and German Federal Republic are ronsidered to be rabies virus. Shope and his assoicates have described several new viruses which have a distant serologic relatedness to rabies virus and to each other. None of these viruses appears to be of a major epidemiological significance. Recent work with monoclonal antibodies has shown that aminoacid changes in 'G' prote,in alter both antigenicity and virulence. Thus antigenic variation and antigenic drift may be responsible for vaccine failure. Monoclonal antibodies clearly distinguish between strains with different passage histories and of different geographic origin and between rabies viruses circulating in different host populations.
Rabies virus isolates from clinical cases are referred to as 'Street' viruses. The street viruses by intracerebral inoculation in the mammals produce a laboratory or fixed virus. The fixed virus has a diminished capacity of infection by peripheral routes and is consistent in behaviour, when inoculated intracerebrally, usually killing the animals in 5-7 days in contrast to ematic reaction produced by street virus where death takes place between 7-8 days. Origninally dogs and rabbits were the species used to obtain fixed virus. Fixed viruses are virulent

Rhabdoviridae 315
even after peripheral inoculation. The attenuated viruses of LEP (Iow egg passage), HEP (high egg passage) and street Albama Dufferin (SAD) immunize after peripheral inoculation of most species but SAD and LEP strains kill when given intracerebrally. Nowadays, suckling mice or embryonated eggs are commonly used. The virus can be grown in laboratory animals by intracerebral method of inoculation. Strains of virus isolates from natural cases of disease are difficult to cultivate in embryonated eggs and tissue culture. Embryonated chicken eggs 5-6 days old are suitable for cultivation of a number of rabies virus strains. Tissue cultures have been used for the growth of rabies virus but in most cultures the CPE is poor, a few cells are infected and virus yield is low. In human diploid cell strains almost all cells are infected and there is marked CPE. The yield of virus may be increased by incubating the infected baby hamster kidney cells at 35°C for 5-7 days. Cell infected with rabies virus can also be maintained and passaged in culLures for extended periods.
Epidemiology: The rabies virus is prevalent in all continents except Australia, Antractic, Great Britain, the Netherlands, Japan and Jamaica. One can easily distinguish the areas where the rabies is maintained by dogs and where it is maintained by wild animals or vampire bats, like in case of latin America.
The transmission through the broken skin is invariably caused by bite wounds. The virus can also be introduced by inhalation and through mucous membranes of eyes and mouth. In India and many other countries of the world dog has been the major reservoir of rabies. Dog bite cases are maximum in this country (93%). The importance of other species like jackals, wolf, fox, mongoose cannot be properly assessed until the data from the total dog bite cases is segregated. About 2000 human beings die every year from rabies while the number of herbivores dying of rabies infection is not fully known. There is definite evidence of rabies in mongoose. This suggests the possibility of transmission of rabies from mongoose to other rodents like rats. There is no evidence of rabies infection in the various species of bats in India. Vampire bats are the reservoir and usual source of infection for paralytic bovine rabies in latin America. The disease with such bats kill about 50,000 to 1,00,000 cattle each year from bat-transmiued rabics. Vampire and insectivorous bats in South America infect cattle and cause paralytic form of rabies. Asymptomatic salivary gland infection

316 TeXlbook o/Veterinary Virology
occurs in bats and results in a prolonged viraemia. Rabies is frequently enzootic in wild animals and bats. In India about 3-4 lakh people take rabies prophylexis and 2000 people die annually.
The virus is not always present in the saliva of rabid dogs, but in other cases it may be found in saliva 3-7 days before signs appear in dog. In blOOd, urine, milk and other secretions, its prescence is rare.
Pathogenesis: The rabies virus primarily affects dogs, cats and other carnivores but all warm blooded mammals including man are susceptible. The fowl can be experimentally infected. The transmission is usually by bite or an accidental break in the skin, though it can be introduced by inhalation and through the mucous membranes of eyes and mouth. The infection does not take place by feeding the infected carcase. The primary replication occ~s in the muscle fibre at the site of inoculation, the virus then aggregates around the proprio receptor nerve endings of their acetylcholine receptors, and migrates along the exoplasm, the centripetal migration takes place. When the virus enters centrol nervous system, usually the spinal cord, its migration to brain is rapid. Although the ultimate target of the virus is eNS, it also infects salivary glands by centrifugal movement along the exoplasmic route. And thus the virus is secreted in saliva. The incubation period which may vary from 10 days to 12 months depends upon the distance of bite_ from the eNS. Early clinical signs in the prodromal stage include fev,*, slight changes in temperament and dilatation of pupils. The disease is manifested in one of the two main forms.
a Dumb or paralytic: Initially there is change in voice which is howling or bellowing. There is short period of excitement followed by incoordination, paralysis, dropped lower jaw, dehydration, loss of condition and death. In cattle there is rumina 1 tympany, tenesmus and sometimes diarr~oea. Tremors and progressive paralysis follow leading to coma and death. The rabies is not always fatal in bats.
b. Furious: Animals attack without provocation. Incoordination, convulsions, coma and death follow within 3-10 days of showing signs.
The rabies is not always fatal in bats, there is inapparent infection and they are consistently viraemic. Skunks and foxes may recover from infection. Dogs rarely show mild symptoms and J;ccover but remain as excretors of virus.
The average incubation period observed in cattle bitten by vampire bats was 60-70 days, while after skunk or fox bites it was 35-45 days.

R habdoviridae 317
This may be due to amount of virus introduced and site of the bite. Skunks and foxes bite mostly on the head or nose while vampire bats bite mainly neck, ears, fetlocks and vulva. Cattle also excrete virus in saliva when ill, like the dogs. The virus in caule is mainly distributed in salivary glands and brain. The virus has never been isolated from meat. Virus in the milk or rabid cows is absent or is present at low levels, since there are few direct glandular nerve connections to the udder and virus in milk is highly diluted.
The clinical signs of rabies in cattle include a short premonitory period which lasts from few hours to few days, in which there is mild temperature, malasie, anorexia and cessation of milk. This is followed by salivation (92%), aggressiveness (47%), bellowing (69%), paralysis(30%) and straining (12%). Most human cases occur during early period when the veterinarian or the owner suspects choke and searches for some object in the throat of affected cattle. In vampire bat rabies the predominant signs in cattle is paralysis.
On histopathological examination the brain show lesions of encephalitis with neuronal degeneration in the midbrain and medulla. Negri bodies in the neurons of hippocampus and cerabellum are pathogenomonic.
Diagllosis: Early clinical diagnosis of rabies is very critical since there is probability of human exposure. The clinkal signs are helpful but are not confirmatory. If the animal dies within 2-3 days after showing symptoms or if the animal dies in the quarantine or the animal has the symptoms, tissues like brain preserved in glycerine saline should be sent to laboratory for diagnosis. The laboratory diagnosis includes the following:
1. Search for Negri bodies in sections or impression smears of the . cerebellum stained by selle~'s or Giemsa stain.
2. If Negri bodies are not found, weaned mice are inoculated intracerebrally. The mice show symptoms within 6-15 days but the symptoms may be delayed upto 21 days and the mice usually die within 1-2 days after that. The mouse brains are examined for presence of Negri bodies from dead as well as from those which survive 5-6 days of inoculation.
3. The fluorescent antibody test is the preferred 'method of diagnosis and is about 100% correct. Smears from hippocampus medulla and cerebrellum are used.

318 Textbook of Veterinary Virology
Serum neutralization test can be used to detennine the antibody response of vaccinated individuals and antibody titre of antiserum used for post-exposure treatment The serum neutralization test is performed in mice or in susceptible cell culture.
Control: The immunization in dogs and cats is generally prophylactic whereas in man and cattle the vaccination is usually carried out after exposure. Prophylactic vaccination is recommended for veterinarians and other persons with a high risk. Hyperimmune serum or immune gammaglobulin obtained from horses or sheep is administered within 48 hours especially in cases of severe bites. This is followed by vaccination after 24 hours. Various types of vaccines have been used since Pasteur vaccine. Previously inactivated vaccines derived from brain were in use, which are still being used in India and some other developing countries. Heat, phenol, UV irradiation and P propiolactone have also been used as inactivating agents. Since a series of injections of vaccine are given in animals and man, the brain tissue somtimes results in allergic encephalitis. These vaccines in man are now replaced with tissue culture inactivated vaccinc. The commonly used vaccine is prepared in human diploid cells. Efforts arc bcing made to prcpare a sub unit vaccine for human use to over come harmful side effects.
The attenuated vaccines used for animals are low egg passaged (40-50 times) and high egg pass aged (180 times) Flury strain. The low egg passaged (LEP) vaccine is less attcnuated and is given to adult dogs only. The high egg passaged (REP) is highly attenuated and is given to puppies, cats and cattle. The inactivatcd and attenuated vaccines have been prepared in cell cultures from hamster kidney etc. The attenutated tissue culture vaccines provide immunity for 3 years.
In India the dog is considered to be the chief animal reservoir for the disease. It is estimated that there are about 10-30 million of dogs in the country. Most of these are stray dogs. All domesticated dogs must be licensed and immunized. In Latin America efforts have been made to reduce the Vampire bat population by inoculating cattle with an anticoagulant (Warfarin) in dosage harmless to cattle but lethal to blood sucking bats. In Switzerland and Bavaria chicken heads inoculated with live attenuated virus have been dropped in mountainous areas in an attempt to vaccinate the reservoir host-foxes.

Rhabdoviridae 319
Bovine Ephemeral Fever Virus (BEFV)
Ephemeral fever is an acute insect transmitted disease of cattle. Sheep and other domestic animals do not suffer. The disease was fIrst reported from central Africa in 1867. The disease is now enzootic in parts of Africa, most of Asia, including India, Japan and parts of Australia.
Properties of the virus: In electronmicroscopic preparation of BEFV three different shapes are found. The longest virions are bullet shaped with a length of about 183 nm and coiled helical nucleocapsid. Particles are also found which have. a truncated bullet shape, while others appear as blunt cones. The length of short particles varies from 70-140 nm and these are considered to be defective and cause interference with the growth of BEFV in tissue cultures. The genome is a single strJndcd RNA and not double stranded as reported by earlier workers. The virus is inactivated within 10 min at 56°C. Citrated blood from affcctcd cattle remains infective for 8 days at 4°C. The virus can be adapted to grow in the brain of unweened mice or hamster which leads to rapid loss of pathogenicity for calves. BEFV can also be adepted to grow in BHK 21 cells, monkey kidney cell line and bovine embryonic kidney.
Epidemiology: It appears that disease maintains a state of endemicity with sporadic individual cases where ideal ecologic conditions prevail. Periodically, after development of susceptible populations, the infection overflows during arthropod season. The usual source of infection is believed to be arthropod vectors. The virus has been isolated from mosquitoes and midges. The role of wild life as perpetuating or amplifying hosts needs to be explored.
Pathogenicity: The incubation period of BEF is 2-10 days. The affected animals show clinical symptoms characterized by a sudden onset of high fever, muscular shivering, lameness, stiffness of joints and enlargement of peripheral lymphnodes. The fever lasts 2-3 days during which time the animals remain pnostrate. There is hyperpnea and dyspnea, excessive salivation, nasal and ocular discharge, anorexia and sharp fall in milk yield. In most of the cases recovery takes place in 3-5 dyas. The lymph nodes are usually enlarged and oedematous. There is congestion of mucous membranes in the nasal cavities, abomasum and small intestine. Degenerative changes in the synovial membrane and increase in synovial fluid are seen.

320 Textbook o/Veterinary Virology
Diagnosis: The sudden onset of disease lasting 2-3 days with recovery is clinically diagnostic. Virus isolation is accomplished by inoculation of buffy coat of caule undergoing ~cute febrile disease in the suckling mouse brain. Serologic diagnosis can also be made by complement flxation and virus neutralisation tests.
Control: Both mouse attenuated or inactivated vaccines mixed with adjuvant are available in certain countries. The inactivating agent is either P propiolactone or formalin. Both the vaccines evoke a protective response.
Marburg Virus (Varet monkey disease)
The virus produces a febrile infectious illness of man handling infected monkey tissues or cell cultures. African green monkey may be the natural host and rarely exhibit clinical symptoms.
References
BURGESS, G.W., 1971. Bovine ephemeral fever: A review. Veterinary Bulletin 41,887-95.
BUXTON, A. and FRASER, G., 1977. Microbiology Voi.2. Oxford Blackwell Scientific Publications.
COMBS, G.P., 1978. Bovine ephemeral fever. Proc. U.S. Animal Health. Assoc.82, 29-35.
Consultation on the WHO!F AO programme in comparative Virology. Report of the rhabdovirus study group Rome. Sept. 25-27, 1979.
HANSON, R.P., 1952. The natural history and vesicular stomaJitis. Bacterial.Review 16, 179.
HENDERSON, W., 1960. Foot and mouth disease and related vesicular disease. Veterinarian, 26 726-730.
KAHRS, R.F., 1985. Viral diseases of cattle. Iowe State University Press, Iowa
KURVERT, E.; MERIEUX, C.; KOPROWSKI, H. and BooET, K., 1985. Rabies in the Tropics, Springer-Verlag. Berlin.
MOHANTY, S.B. and DurrA, S.K., 1981. Veterinary Virology. Lee and Febiger, Philadelphia.
RusSELl., P.H. and EDINGI'ON, N., 1985, Veterinary Viruses. The Burlington Press(Cambridge) Ltd. Foxton, Cambridge.

Rhabdoviridae 321
'" SPRADBROW. P.B., 1977. Vaccines against bovine ephermeral fever. Aust.Veterinary Journal. 53,351-52.
SCHNEIDfR, L.G., 1983. Rabies Bulletin Europe/Information Surveillance Research, Federal Research Institute for Animal Virus Diseases, Tubingen, Germany.
WHO expert Committee on rabies. Sixth Report WHO Technical Report. Service, No. 523, 1973.
Wno document: Guidelinesfor rabies control VPH? 83-43. 1984.

Chapter 29
Retroviridae
The family retroviridae derives its name from the enzyme reverse transcriptase (RNA-dependent DNA-polymerase) which transcribes a DNA copy from viral RNA genome. The Latin word retro means backwards. The family consists of three subfamilies: oncovirinae, lentivirinae and spumavirinae. Oncovirinae subfamily contains tumorigenic viruses, the oncoviruses. The lentivirinae cause chronic diseases in sheep, goats, horses and human acquired immunodeficiency virus of man. The spumaviruses are not pathogenic.
The subfamily oncovirinae includes three genera namely type C, type B and type D oncovirus groups. Many members of these genera cause neoplastic disease of animals and birds. Type C oncovirus group includes number of import.'mt species such as bovine type C oncovirus, feline leukemia and sarcoma viruses, murine leukemia and sarcoma viruses, avian leukosis and sarcoma viruses and avian reticuloendotheliosis viruses. Type B oncovirus group includes the species such as murine (mouse/guinea pig) mammary tumour virus. Type D oncovirus group do not have species important for domestic animals and birds.
The retrovirions are spherical enveloped particles measuring 80-100 nm in diameter and possessing glycoprotein surface projections of approximately. 8 nm. The particles have a three layered structure. The innermost is the genome-nucleoprotein complex, which includes 30 molecules of reverse transcriptase. This is enclosed within an icosehedral capsid which in turn is surrounded by host cell membrane derived envelope. The genome of retroviruses is an inverted dimer of

Retroviridae 323
linear single stranded RNA with a positive polarity. The monomers are held together at 5' ends by hydrogen bonds probably by base pairing. A cellular tRNA serves as a primer for reverse transcription and is bound to specific primer attachement site about 100 residues from 5' end of the genome. The genetic information is contained in the genes gag (coding for the nonglycosylated internal proteins), pol (coding for -reverse transcriptase) and env (coding for envelope glycoproteins). Some retroviruses incorporate genetic information for nonstructural proteins from the host cell which are important for pathogenesis in oncogenicity. The RNA of ral?idly transforming oncoviruses contain a fourth gene, the viral oncogene (V-onc). The presence of V-onc gene is usually assoicated with deletion of some of env base sequences and this renders the virus as replication defective and depend upon transforming helper virus for the production of infectious progeny. After entry of the virus into the cell and uncoating, replication starts with the reverse transcription of virion RNA into DNA, which is made double stranded. This double stranded DNA is integrated into chromosomal DNA of the host cell at nonspccified sites. The virus can replicate when DNA is integrated. Cellular RNA polymerase II transcribes the integrated provirus into virion RNA, mRNA. Virus is released from the cells by budding.
ONCOVIRUSES Bovine Leukaemia Virus (BL V)
Bovine leukemia virus (BL V) is the causative agent of bovine leukemia (lymphosarcoma, leukemia) a lymphoproliferative disease of catLIe.
Properties of the virus: BLV is classified as a type-C oncovirus in the retroviridae family and shares characteristics ·with leukaemia viruses of other species. The mature virus particle is 90-120 nm diameter in size. The virions possess a major nucleoprotein antigen with molecular weight of 24,000 daltons and an envelope glycoprotein antigen of 51,000 daltons molecular weight. The virus is fragile and looses its infectivity by routine manipulations. It is readily destroyed at 56°C within 30 minutes. The virus is inactivated at pasteurization temperatures. Binary ethylenemine and N-acetylethylcnemine are effective inactivating agents.
The BL V isolates from various parts of the world do not show

324 Textbook of Veterinary Virology
antigenic differences. There is no antigenic relationship between BL V and leukemia virus of other domestic animal species. There is some evidence that BL V shares structural and functional properties and even minor antigenic determinants with human-T cellleukemia virus.
The. virus replicates in some primary cell cultures of bovine or ovine origin and causes syncytia formation in various transformed and non-transformed cell cultures.
Epidemiology: BL V infection among cattle are world wide in distribution but certain countries like the Netherlands, Austria' and British Isles have a low incidence. The cattle are considered to be primary and important host of BLV. Although sheep have been found to be infected and deaths due to lymphosarcoma have taken place, but these instances are rare and are regarded as due to accidental introduction of BL V rather than considered that sheep are significant reservoir of the virus. There is report that water buffalo and a large rodent. capybara in South America has antibodies against BL V but the percentage of positive sera prevalence is well below that of cattle. The information supports the hypothesis that BLV is propagated and maintained primarily in caule population and infections in other species are accidental introductions. The frequency with which the transmission occur may be related to the vectors capable of transmission to callle as well as to other species. Ticks and biting flies have been incriminated but other biting insects may also be involved. The available evidence indicates that arthropods are only transport vectors and not true biological vectors.
There is no evidence of human infection with BLV. However, there is an evidence of distinct antigenic relationship with T-cell leukemia viruses. This results into appearance of antibodies which react with BLV in few human subjects known to be infected with human Tcellleukemia virus.
The economic losses arise from deaths and restrictions on the movement of BLV infected animals or their products (semen, embryo) in international trade. The virus is transmitted horizontally and does not gain enterance to the germ line and there is no vertical transmission but the transmission in utero takes place. The in utero infection perpatuates the virus form one generation to the next. Calves which are not infected in utero but born to infected cows are occasionally infected b)' ingesting of colostrum or milk that contains infected lymphocytes.

R elroviridae 325
Contact virus transmission may occur among young animals. The calves remain protected from contact infection until the colostral antibody wanes, usually 4-6 months of age.
The factors which influence epidemiological pattern have not been identifIed but the prevalence of infection is more among dairy animals than beef cattle. The genetic factors may be responsible for this. The development of tumors in infected animals is influenced by virologic, genetic and immunologic factors but these are not understood as yet. The genetic factors influence the development of tumors because numerous tumor cases have been recorded within certain cow families and sire groups.
Palhogenesis: The susceptible host gets probably BL V infection through infected lymphocytes. Experimental inoculation with infected lymphocytes by parenteral route, particularly intracutaneous inoculation is most successful route of establishing infection. The oral, intranasal and intauterine instillation may establish infection in susceptible host not in all the animals, when large quantities of infectious material is inoculated. It indicates that blood sucking parasites may play a role in the transmission of virus. Once the infected lymphocytes enter the host, a cycle of virus replication starts in the infected lympnocytes. The virus produce infection in susceptible cells, probabl,y of lymphoid or stem cell origin and further replication goes on. The plasma of infected animals may contain virus on occassion but not regularly. The B,L V infection is established in lymphoid tissues in the first few weeks of infection and virus can be isolated from lymphocytes 3-5 weeks of inoculation. No cell free virus is produced in the tissues. Blood is the potent source of infectivity. Saliva, urine, semen, milk may contain infectivity due to their lymphocyte content
The molecular mechanism involved in cell transformation and tumor development is not known. BL V is integrated at various sites in the cell genome, one of the BLV gene products is capable of activating cellular genes and brings about malignant transformation.
The BL V infection is associated with lymphocytosis. About 29-85% animals develop lymphocytosis. The development of lymphocytosis and tumor formation are determined by separate genetic factors. The development of lymphosarcoma is a rare result of BLV infection. There is no tissue predilection for tumor development and the clinical signs of disease may be extremely varied. There may be

326 Textbook ofVelerinary Virology
clinical signs related to organ involvement such as cardiac distress, digestive disturbances or neurological signs. In some cases there may be vague clinical signs like decreased production, inappetance, weight loss, weakness and debility. . Immune Reaction: TIle humoral antibodies appear within 3-9 weeks of infection. The neutralizing antibodies do not lead to elimination of BLV because of the mechanism of virus persistence. The constant presence of viral genome is accompanied by continuous presence of antigen, with the result that antibody titres usually remain high throughout the life span of the infected animal. There is no evidence of CMI response to BL V infection.
Diagnosis: The diagnosis is aimed at detection of BL V antibodies and diagnosis of lymphosarcoma.
For BL V infection serological tests are carried oul. A complete herd of all cattle over 6 months are tested. The RIA, ELISA and VN test have the greatest sensitivity but AGID test is widely used because of its practicality.
The diagnosis of clinical cases (lymphosarcoma) requires a pathological investigation. The tissues may be from necropsy or biopsy. The effect of BL V infection on the haematologic values in cattle is important from diagnostic point of view. Many animals develop persistent lymphocytosis. The examination of blood smears reveal the presence of abnormallymphocytes.
Prevention and Control: No vaccines are available. The management practices such as sanitation and good nursing care are valuable for short period of time. Common use of needles and surgical equipment should be avoided. Insect control, prevention of the introduction of cattle from positive reactor herds and slaughter of infected heards are important.
Feline Leukemia/Sarcoma virus
The feline leukemia/sarcoma virus causes Ieukemia/ lymphosarcoma in cat and is manifested by malignant, proliferation and degenerative changes of haematopoietic tissues.
Properties of the virus: The virus particle resembles C type particle. They are ciruclar or oval in shape with 100-110 nm external diameter. Two kinds 9f viral structural antigens such as glycoprotein envelope antigen which has a MW of 70,000 and a major internal

Relroviridae 327
.protein antigen with a MW of 30,000 are present. Three subgroups (A, B, C) of feline leukemia virus were recognised. All subgroups of feline leukemia and sarcoma virus have an identical major internal antigen. A cell surface antigen called feline oncovirus associated ceU membrane antigen has been detected in lymphoid cells of cats with lymphosarcoma. The feline leukemia/sarcoma virus can be propagated in normal embryonic feline cell cultures inoculated with extracts' of tumour tissues. The virus can also reproduce in cultured cells derived from canine, procine and human sources but fails to replicate~ chicken and bovine cell cultures. Feline leukemia virus can activate defective mouse sarcoma viruses which can infect kitten cells an'd the 'rescued hybrid' experimentally can produce sarcoma in kittens.
Epidemiology: Most infected cats excrete the virus in saliva, urine, faeces, milk and nasal secretions. The virus can be transmitted vertically from infected mother to off spring in utero and also through the milk from infected dams to the kittens. There is a report that the virus is present in the salivary glands and the virus can be transmitted from cat to cat by biting and scratching.
Pathogenesis: Young cats under 4 months of age are comparatively more susceptible and the susceptibility decreases following increase of age. The incubation period of the disease is variable. On experimental infection, virus can be demonstrnted in tissues within 3 to 4 weeks and lymphosarcoma appaears at about 6 to 12 months of age. Early symptoms of anaemia, leukopenia,
. thrombocytopenia develop few weeks or months prior to the development of acute leukemia. Lymphosarcomas commonly originate from lymphnodes, kidney or intestine and from these sites the disease may spread to other organs and to the bone marrow. The thymic form of feline lymphosarcoma is most common and is characterised by the presence of tumor mass in the anterior mediastinum.
Experimentally feline lymphosarcoma can be produced in kittens under 12 hours old by the inoculation of cell-free extracts of spontaneous and induced tumor tissues.
Diagnosis: Provisional diagnosis can be made from the gross lesions, clinical and haematological findings. Confirmatory diagi}osis is achieved by isolation of virus from the plasma of infected animals or tumour tissues, electron microscopical examination of affected tissues and serological tests. Virus specific antigen can be detected in

328 Textbook of Veterinary Virology
ciruclating leukocytes of affected animal by immunoflorescence. Immunodifussion .test is suitable for detecting viral antigen in the tumour mass. Indirect haemagglutination and virus neutralization test can also be used for detection of virus specific antibody.
Control: A live virus vaccine derived from the virulent virus types has been' shown protective against the disease. Immunization of pregnant mothers with inactivated virus provides 3-4 months protective passive immunity to the new born. Kittens should be vaccinated at abOut 3-4 months of age.
Feline Leukemia is an excellent model for the study of viral oncogenesis in man.
Murine Leukemia/Sarcoma Viruses
The murine leukemia (MuLV) and sarcoma (MSV) viruses are assoicated with the production of a vareity of morphologic tumours, leukemias and sarcomas in a number of host species.
Properties of the virus: First MuLV was discovered by Gross in 1951. Subsequently founeen different leukaemogenic RNA murine viruses have been isolated. It is probable that the majority of murine leukemia viruses do not bear any etiological relationship to the tumour in different host species. All MSV exist as a mixture with MuL V and bear the antigenicity of assoicated MuLV. An envelope glycoprotein antigen with a MW at about 70,000 has been recognised and this antigen is common to mouse leukemia/sarcoma viruses, but does not cross react with the glycoprotein antigens of other mammalian or avian . oncoviruses. The MSV and Rous sarcoma virus of chicken and the avian leukosis virus and MuL V are similar in their nueleo protein antigen. The MuL V and MSV possess a major internal antigen with a MW of 30,000. MuLV can mUltiply in cell cultures of mouse and rat without producing CPE. MSV are defective but can cause transformation of cell cultures derived from mouse, rat, hamster or human embryonic tissues.
Palhogenesis: MuL V and MSV are widespread in nature. Viruses can be transmitted vertically as well as horizontally through milk and sperm of infected animals. Most often these viruses remain in latent form and the activation of viruses leading to formation of leukemia/ sarcoma occurs by an unknown mechanism. Different strains of mice show considerable variation in susceptibility to one virus.

Relroviridae 329
Experimentally tumor formation takes place following inoculation of the virus to the newborn mice of high susceptible strain. Most MuL V also produce leukemia in rats. The gross's MuLV may give rise to almost all known types of leukemia including lymphatic stem cell, myeloid and monocytic leukemia, erythroblastosis, lymphosarcoma and reticulum cell sarcoma.
Diagnosis: Diagnosis is based on the gross lesions, demonstration of virus specific antigen by radioimmunoassay, complement fixation (COMUL), immunofluoreScence and immunodiffusion test
Control: Vaccines have been prepared and are in the experimental trials.
Avian Leukosis/Sarcoma viruses
Avian Ieukemia/sarcoma viruses cause a variety of transmissible benign and malignant ncoplasma in chickens and to a lesser extent in other avian species. The neoplastic conditions are lymphoid leukosis, erythroblastosis, myeloblastosis, myelocytomastosis, endothelioma, nephroblastoma, hepatocarcinoma, fibrosarcoma and oteopetrosis. Of these ncoplastic conditions, lymphoid leukosis is most common. Avian leukosis virus predominately affects the haematopoietic cells and the sarcoma virus affects the fibroblastic cells.
Properties of the virus: Avian leukosis/sarcoma viruses are grouped together because they possess may properties in common. Viruses of the avian leukosis/sarcoma group cannot be distinguished on the basis of their ultrastructural characteristi.s such as size, shape etc. The virus has a MW of 10 x 1()6 daltons and 60 to 70s sedimentation coefficient. Avian lcukosis/sarcoma viruses are divided into subgroups (A to G) on the basis of host range in genetically different chick embryo fibroblast cultures, interference ~ith members of the same subgroup and viral envelope glycoprotein antig<!ns. Genetic host range or cellular genetic resistance to infection by viruses of a particular subgroup is controlled by an autosomal locus called the avian tumour virus locus and cells resistant to any viral subgroup are designated as Cl subgroup and cells susceptible to all subgroups are desiggnated as C/o. Interference between leukosis and sarcoma viruses has been well characterised. Chicken embryo fibroblast cell cultures infected with avian Ieukosis virus become resistent to superinfection by sarcoma virus of the same subgroup and this is the basis of the resistance inducing factor (RIF) test.

330 Textbook o/Veterinary Virology
The avian leukosis/sarcoma viruses can be inactivated by heating at 56°C for 30 minutes. Inactivation can also be achieved by formaldehyde and by common disinfectants such as 0.5% phenol. Viability of virus preparations can be maintained for long periods at -70°C or in 50% buffered glycerol saline.
The, oncogenic leukosis viruses are exogenous in nature but the endogenous leukosis viruses are non oncogenic. All chicken cells can carry an endogenous viral genome required to produce a complete leukosis virus. Most of the avian Leukosis viruses on the other hand infect genetically susceptible chicken cell cultures and produce virus continuously. 'The avian leukosis/sarcoma viruses can be grown in embryonated chicken eggs of susceptible lines. Avian sarcoma viruses produce pocks on the CAM of embryonated eggs and are capable of producing tumors in embryos when inoculated by the intravenous and yolk sac routes of inoculation. Avian leukosis virus when inoculated by intravenous route to chicken embryos cause death of chicks 1 to 2 weeks after hatching.
Epidemiology: The natural host for avian leukosis/sarcoma viruses is the chicken. Experimentally, wide range of hosts may be infected. The viruses can be transmitted vertically form parent to off spring or horizontally form bird to bird. The transmission of the viruses through egg is of utmost importance in the perpetuation of virus infection. Chicks that are developed from congenitally infected embryos are immunologically tolerant and have large quantities of infectious virus in their blood and tissues. Large amouts of virus are also shed in their droppings and saliva.
Palhogenesis: The incubation period of the disease in natural conditions is not clearly known. Natural disease usually occur after 14 weeks of age and the incidence is highest at about sexual maturity. Experimentally when day old chicks were inoculated intra abdominally with leukosis virus, the lymphoid leukosis develops within 5 to 8 months and those that receive sarcoma virus intramuscularly or intradermally the sarcoma develops with in 3 to 35 days. The clineal signs of lymphoid leukosis are not specific. There may be paleness of the comb and cyanosis, inappetance, emaciation and weakness occur. Abdomen is often enlarged due to the presence of tumours. Enlargement of liver, bursa of fabricious and kidney can often be detected on palpation. It is suggested that the lymphoid leukosis is a

Retroviridae 331
malignancy of the bursa dependent lymphoid system. There is also evidence that the surgical or hormonal removal of the bursa from chickens reduces the incidence of malignancy from leukosis virus. Similarly hormonal atrophy of bursa resulted in similar effect. Grossly, however, visible tumours in natural conditions can be seen in liver, spleen and bursa of fabricious. Many other organs such as kidney, lungs, gonads, heart, bone marrow and mesentary may also be affected. The tumors are soft, smooth and glistening and the cut surface had greyish or creamy white appearance. Granular or miliary form ·of leukosis is usually seen in liver and distributed throughout the parenchyma. Microscopically tumours consist of aggregates of large immature lymphoid cells. In erythroblastosis the prominant clinical signs are weekness, cynosis of the comb, anaemia and the presence of erythroblasts in the blood. There may be cherry red discolouration .. md diffuse enlargement of the liver and spleen due to accumulation of erythroblasts in the blood sinusoids and capillaries. In myeloblastotis signs are similar to that of erythroblastosis and massive accumulation of myeloblast can be seen both intravascularly and extravascularly. In osteopetrosis thickening of long bones of the limbs is a common feature.
Diagnosis: Provisional diagnosis can be made from the history, clinical signs and the histopathological lesions. Confirmatory dagnosis tests is achieved through isolation of the virus and perological tests. Virus can be isolated from plasma, serum or tumour tissues of infected birds by inoculating into cell cultures. Virus specific antigen can be detected by radio immunossay, complement fixation test (COFAL), immunoflourescence and the EpSA. Presence of virus in infected cell cultures can be detected by reverse tianscriptaseassay. Virus specific antibodies can be detected in serum and in egg yolk of infected chicken by VN test using sarcoma virus pseudotypes. Differential diagnosis should be made from Marek's disease, a herpes virus disease which usually occurs in chickens at lower age. Le. less than 14 weeks old.
Prevention and control: No vaccines are available. Control is only by flock managment and genetic selection of chickens for resistance to avian leukosis/Sarcoma viruses.
Avian Reticuloendotheliosis Viruses (REV)
This group of virus causes tumours of the reticuloendothelial

332 Textbook of Veterinary Virology
system. This group of virus comprise of turkey REV, chicken syncytial virus, duck infectious anaemia virus and spleen necrosis virus. All these viruses are grouped together and termed as strains of REV.
Properties of the virus: The virus belongs to the subfamily oncovirinae and genus oncovirus type C of the Retroviridae family. The mature virus particles measure 85 to 110 nm in diameter, possess a diffuse, relatively dense nucleoid and a limiting membrane and can be distinguished from leukosis viruses. The virus particles consist of 2 distinct nucleoid component and the major internal antigen had a M.W. of 30,000 daltons and the M.W. of envelope glycoprotein antigen is 73,000 daltons. The v'iruses are thermolabile, viral activity is destroyed at 56°C in 30 minutes and after exposure to 25 to 50% ether. Cell free virus retained activity at -70°C for 4 months and at 4°C for 12 hrs. Virus replicates in cultures of chicken, quail and duck embryo fibroblasts and can persist for long periods without marked changes in the infected cell. Chicken emrbyos are also infected when inoculated by yolk sac and CAM routes.
Pathogenesis: Natural infection is only reported in turkey flocks. Experimentally, chicken, turkey, Japanese quail and ducks are infected. Turkey poults are most susceptible. The viruses may induce a variety of syndrome such as spleen necrosis, anaemia, lymph nerve lesions etc. The affected birds manifest enlarged livers and spleens and infiltration of subcapsular nodules. There may be splenomegaly, liver may be yellowish brown or pale. Distended gall bladder is characteristic. Nodular lesions may be seen in other visceral organs including gonads, heart, kidneys and pancreas. Enlargement of peripheral nerves particularly the cervical portion of the vagus nerve can be seen. Histologically most characteristics findings are rapid proliferation of mononuclear cells of the reticulo endothelial system around blood vessals and in lymphoid follicles. Mitotic figures are common. Proliferation is marked in liver and spleen. Sometimes there may be diffuse infiltration of lymphocytes and plasma cells in peripheral nerves.
Diagnosis: Provisional diagnosis can be made from gross and microscopic changes. Confirmatory diagnosis can be obtained by isolation-of the virus from cell free tumour homogenates, spleen, whole blood or plasma in susceptible cells culture or in chicken embryos by yolk sac route. Virus specific antigen can be detected by IF and VN test

Retroviridae 333
in infected cell cuture. Experimentally disease can be reproduced by inoculations of day old chicks, turkey poults and Japanese quail. Differential diagnosis should be made from lymphoid leukosis and Marek's disease.
Murine Mammary Tumour Virus (MMTV)
Murine mammary tum~)Ur virus (MMfV) produces mammary carcinoma in certain strains of mice and in guinea pigs.
'Bittner(1936) observed that newborn mice of high incidence strains showed low incidence when nursed on low incidence foster mothers and low incidence strains showed a high incidence when fostered on high incidence mice. He regarded this phenomenon as a milk borne factor in this disease, but susbsequently causative virus has been discovered.
The MMfV is transmitted horizontally to new borns, Some strains of MMTV are also transmitted vertically also. The MMTV strains showed variability in inducing tumors. Hormonal factors or host genetic factors play important role in the production of the disease.
The MMfV particles in milk or extracts of infected mouse tissue can be inactivated by trypsin and by heating at 56°C for 30 minutes. The virus cannot be propagated by conventional tissue culture methods. Several strains of MMTV have been recognised and are antigenically related.
The MMTV produce carcinoma of the mammary gland after a latent period of 6 to 12 months, high concentration of the virus is found in the milk and is transmitted to the newborns through the milk: Virus is also present in the sperm of male mice.
No antibody against the virus has bren found in infected mice serum but sera from guincapigs, rabbits and rats immunized with MMfV when mixed with the virus and injected into susceptible mice results in the failure of the production of carcinoma.
SPUMA VIRUSES
Bovine Syncytial Virus (BSV)
Bovine syncytial virus (BSV) is associated with lymphosarcoma of cattle and was first isolated in 1969 in the United States. The virus has been isolated in different countries not only from the diseased cattle but also from apparently normal cattle. It is believed that approximately

334 Textbook o/Veterinary Virology
25% of nonna! caule remain as carriers of BSV. Properties of the virus: The virus belongs to the subfamily
spumavirinae under the family Retroviridae. Electromicroscopy revealed that the double mambrane bound virions had approximately 125 nm in diameter. The virus is extremely cell assoicated. It is sensitive to chlorofonn, heat and freezing. The viral replication can be inhibited by actinomycin D and 5-bromouridine. The virus can be propagated with production of syncytia in a number of cell systems such as bovine embryonic spleen cells, rabbit cornea cells, cell lines of vero and BHK-21 cells. Bovine embryonic cells appear to be most sensitive.
Pathogellesis: The virus can be recovered from the buffy coat cells of diseased and apparently normal cattle. Therefore blood transfusion or use of contaminated needles etc. may be possible source of infection. The virus has been recovered from milk of infected cattle. The role of the virus in disease production is not clearly understood. However virus has been recovered from the spleen, lungs, lymph nodes and uterus and foetuses of infected animals. Virus has also been isolated from tissues of cattle suffering from lymphosarcoma.
Diagnosis: Virus· can be isolated from the buffy coat cells and from other infected tissues by inoculating into the sensitive cell systems. Virus specific antibodies can be detected by immunodiffusion and 1FT.
Control: Young calves acquired colostral immunity for a period of 3 to 4 months. No vaccines are available.
LENTIVIRUSES
Equine Infectious Anaemia Virus (EIA V)
Equine infectious anaemia virus (EIA V) is responsible for the production of a progressive disease characterised by severe anaemia in horses. The disease casued by EIA V is also known as Swamp fever. The disease was first recognised in France in 1843, and its viral etiology was detected in 1904 by Vallee and Carre. Disease has been reported from the different parts of the world. In India the disease was first reported by Uppal, Yadav and Ahmed in 1987 from the National Research Centre on Equines, Hisar, India. Horses are the only animals, affected. However, the outbreaks have been recorded in donkeys and mules. Human beings are also susceptible.

Retroviridae 335
Properties of the virus: The virus belongs to the family Retroviridae and subfamily Lentivirinae. Nucleic acid of the virus is RNA and the virions possess RNA dependent DNA polymerase. Virions possess surface knobs and a tubularly shaped core. The virion has a MW of 4.8 x IOS dalton and measures 90 to 140 nm in diameter and the sedimentation coefficient is 110 to 120s. The virus·is relatively
. resistant to heating, freezing and drying. It is inactivated by heating at 58°C for 30 minutes and by a variety of chemical disinfectants such as 2% phenol, 0.022% mercury chloride, 4% formalin, sodium hydroxide and sodium hypochloride. The virus particles have a major group specific antigen and also bear surface antigens which may vary. The virus has a number of strains or variants that are immunologically distinct by their surface glycoprotein antigens. The virus· can be propagated in equine cell cultures and in horse leukocyte cultl!res.
Epidemiology: The disease has a seasonal incidence and is not prevalent during the late summer and autumn. The commonest vectors are the blood sucking insects. Mosquitoes such as Aedes, Anopheles and Psorophora and biting flies of the family Tabanidae and Stomaxys calcitrans play important role in natural transmission of the disease. Mechanical transmission occur following the use of contaminated surgical instruments and syringe needles. Vertical transmission also occurs but not all foals from affected mares acquire the disease. Although horses, asses and mules are only naturally susceptible animals, pigs may become affected occasionally. Several workers reported transmission to sheep, rabbits, mice and rats without the production of any symptoms.
Pathogenesis: The incubation period ofthe disease varies from 1-3 weeks but it may be longer. The disease may occur in acute, subacute, chronic or inapparent forms.
In acute form there is sudden rise in temperature, which is intermittent beside depression, anorexia, thirst, serous, ocular and nasal discharges, progressive weakness and debility. Congestion and petechial haemorrhages are found in the visible mucous membranes. Slight albuminuria, anaemia and general weakness may be noticed. Oedema is most prominant in the subcutis of the ventral wall. The acute illness lasts for 3-5 days and if "it does not terminate fatally than it passes to subacute or chronic form. In the subacute form there may be relapses of temperature, oedematous swelling in the lower trunk or

336 Textbook o/Veterinary Virology
limbs and horses may die unexpectedly. Swelling and pigmentation of the liver, enlargement of the spleen, lymphnodes, kidney and hyperplasia of bone marrow are found.
In chronic form there is debility and anaemia. Hypertrophy of the spleen and pone marrow are characteristic pathologic lesions.
Sometimes there may be inapparent infections and horses may remain as symptomless carriers for long periods.
The anaemia that appears intermittently in this disease is possibly due to the destruction of RBC by an immunologically mediated mechanism. It has been observed that RBC of infected horses are coated with antiviral antibodies and compliment component. This binding to the cell surface results in increased osmotic fragility, shortened half life and erythrophagocytosis. Plasma haemoglobin increases and serum hepatoglobin levels decrease in infected animals. This finding suggests that the haemolysis is responsible for the development of anaemia. Another less important factor in the genesis of anaemia is the depression of bone marrow during acute episodes. Glomerulitis was also observed in infectious anaemia and this was thought to be due to deposition of virus antibody complexes. Similar immunologic factors are likely to be associated with the development of lesions in other organs.
Diagnosis: Provisional diagnosis of the disease can be made from the history, clinical signs and lesions. Confirmatory diagnosis is based on the isolation of virus from the serum whole blood or leukocytes of infected horses in equine leukocytes culture, demonstration of virus specific antibodies using immunodiffusion (Coggins test) and complement fixation tests and by transmission experiments. Transmission experiment is carried out by inoculating 20 ml of whole blood or serum from infected animal into young (12 months) healthy foals. Recently it has also been observed that the FAT is useful for detection of virus specific antigen in peripheral leukocytes cultures of infected animals.
Control and Prevention: There is no satisfactory vaccine against the disease. The control of the disease depends on the early detection of infected animals, their separation and slaughter besides close observation of the incontact animals. Regular quarantine practices and control of the flies and mosquitoes are also important for the control of the disease.

Retroviridae 337
VisnaIMaedi Virus
Visna and Maedi are two disease manifestations due to a virus belonging to lentivirus subgroup of retroviruses. Maedi is a word derived from Icelandic word which means dyspnea and is used for slow progressive interstitial pneumonia. While the tenn visna means wasting is used for slow progressive inflammatory disease of the central nervous system resulting in paralysis.
Properties of the virus: The maedi-visna virus (MVV) has a typical morphology of retrovirus. It is about 100 nm in diameter contains about 30-40 nm nucleoid. The virus is readily inactivated by ether, chloroform, ethanol, formaldehyde and trypsin. It is stable at -50°C and between pH range of 5.1 and 10. The structural proteins are similar to lentiviruses except certain differences in the glycoproteins. A major virus component (40% of the virion mass) consists of a single protein of 25-30,000 daltons and is a constituent of the virus core; a glycoprotein of 125000 daltons is associated with surface knobs. These two proteins give two precipitation bands in immunodiffusion tests. The envelope of MVV contains neuraminic acid. The virus does not show haemagglutination property but inhibits haemagglutination by influenza virus. Maedi and Visna viruses are antigenically similar. Antigenic drift of the virus has been observed in infected animals. More than 1 strain of virus can exist in the same animal.
The virus replicates in sheep cell cultures with the production of syncytia and cellular degeneration. Virus can also be propagated in BHK-21 cells and primary cell cultures of bovine, porcine, canine and human choroid plexus cells. The virus does not grow in embryonated chicken eggs.
Epidemiology: MVV infection are found in sheep only. The virus can be transmitted to goats by parenteral infection but it is not clear if the infection of sheep can be transmitted to goats under natural conditions. It is not possible to infect laboratory animals. There are no known reservoir hosts or vectors. There is a widespread geographic distribution of MVV. The disease is prevalent in USA, Netherlands, South Mrica, France, Germany, Belgium, Spain, Italy, Greece, Hungary, Bulgaria, Switzerland, Israel, Soviety Union, Canada, Kenya, Peru, India besides Iceland. There is indication that genetic differences in the virus and/or in sheep population may determine which manifestation of the infection is dominant.

338 Textbook of Veterinary Virology
Pathogenesis: Naturally the virus spreads laterally probably by respiratory route. Where the disease is enzootic the important route of infection is from ewe to lamb through clostrum. The transmission of infection in utero is rare under natural conditions. The virus is found in lungs, CNS, spleen lymphnodes, salivary glands, white blood cells and mammary gland. There is no evidence of transmission through semen. In experimental infection there is severe restiriction of virus replication in various tissues. Many cells contain viral genome in DNA provirus form. In smaller fraction of cells the DNA is transcribed into RNA. The production of lesions in visna appears to be immunologically mediated, perhaps by a CM! response to virus antigen on infected cells. The pulmonary lesions in maedi may also be due to some immunopathological process taking place in the lungs. The infected animals never become free from the virus and remain a constant threat to any healthy contact animal.
The clinical symptoms of maedi is failure to thrive. The affected animals show respiratory distress when led to pastures and lag behind the healthy animals. Later there is increased respiration at rest. Coughing is not prominent and no discharge from nostrils. The animals become weak inspite of good appetite. The clinical course lasts for months or even more then a year. The clinical maedi is rarely seen in animals younger then 3-4 years of age.
The clinical symtoms of visna are dominated by nervous symtoms. The animals show lameness of one or both hind legs, weight loss inspite of good appetite. Paralysis of hind legs progresses but the front legs are atfected at final stage of disease. There is slight tremor of head and of the muscles around the face and occasionally blindness. The course of disease runs for several months or even years and ends in prostration and death. Maedi and visna may coexist in the same flock or even in the same individual. Recently arthritis has been observed in flocks in USA with progressive pneumonia. Chronic indurative mastitis is also associated with MVV.
In maedi the lesions primarily affect the lungs and maediastinal lymph nodes. The lungs increase two to three times of their normal size and do not collapse when the thoracic cavity is opened and exhibit a dark greyish blue or greyish brown in colour rather than pink colour. The lungs are compact to touch like a rubber sponge; The tmcheobronchial and mediastinal lymphnodes are enlarged.

Relroviridae 339
Microscopically lesions consist of diffuse thickening of interalveolar septa which encroaches upon alveoli. The thickening of septa is caused by infiltration of lymphocytes, monocytes and/or macrophages with few plasma cells.
In Visna the cardinal features are chronic inflammation of brain and spinal cord, lesions are mainly distributed around the ventricular system affecting both grey and white matter. Subependymal and perivascular infiltrates of lymphocytes, monocytes and/or macrophages with plasma celis are typical. The inflammatory changes are sometimes severe with liquefaction necrosis. Destruction of myelin together with the axons is seen in necrotic foci.
Immune reaction: The infected sheep develop MVV antibodies which can be detected by VN test. The VN test has several drawbacks because it is time consuming and gives false negative results due to antigenic variation of virus. The neutralizing antibodies appear 2-3 months after experimental infection of sheep. The CF antibodies appear 3-4 months'after allaining a plateau. More positive animals are detected with CF test rather than VN test. The glycoprotein and core antigen can be detected by AGID. About 90% of the sheep infected with MVV show a positive reaction. The ELISA, indirect IF are equally effective is detecting MVV specific antibodies.
The CMI response is irregular and vary with time. There is evidence from experimentally infected sheep that CNS lesions are immunologically mediated in the disease. The pathogenesis of primary demyelination indicate immune attack on infected oligodendrocytes may be responsible for the primary demyelination.
Diagnosis: In living animals the useful test for diagnosis are aimed at detecting the antibodies against the virus. ELISA and AGID tests are used as herd tests, since individual sera may give false negative results. At autopsy the histopathology of lungs and brains reveals typical lesions. The virus isolation is the final test for detection of infection. The best method of virus isolation are the cultures in plasma clot and cocultivation of cells from infected animals with susceptible cell strains such as cultivated plexus choroideus or testis cells. The CPE produced is rounding of cells, giant cell formation, degeneration and lysis. Some strains are not strongly cytolytic and do not produce higher titres. The spleen, lungs, mediastinal lymph nodes and plexl\s choroideus are organs of choice for virus isolation.

340 Textbook o/Veterinary Virology
Control: No vaccine is available. The only method to control the disease is slaughter of infected animals. Attempts to clean up the infected herd by removing lambs at birth from ewes and raising them artificially in isolation have been successful in Netherlands and USA. In Iceland the disease has been eradicated by stamping out policy.
Caprine Arthritis-Encephalitis Virus
Caprine arthritis-encephalitis virus (CAEV) produces a disease complex among domestic goats of all ages. The disease is characterized by paresis leading to paralysis among young goats while in adults there is chronic persistent arthritis and mastitis. Rarely the adult goats also develop progressive fatal pneumonia and encephalitis. The disease is widely disseminated in goat herds in North America, Europe and Australia.
Properties of the virus: The CAEV is a nononcogenic member of lcntiviruses. The polypeptides of the virion are similar to MVV. Antigenically the virus is related to MVV: The virus can be cultivated in early subcultures of outgrowths of synovial membranes of goats. Replication of virus produces CPE, characterised by cell fusion and is the major CPE produced by the virus.
Epidemiology: The epidemiology of CAE is still being evaluated. The studies on epidemiology have bcen complicated because a single virus causes three different diseases in different age groups of the host. These are (i) rapidly progressive lcukoencephalitis is new born and young animals (ii) chronic mastitis and arthritis and masti~s in adult animals (iii) a sporadic slowly progressive pneumonia encephalitis in adult goats. The n~urological disease in kids occur in age below 4 months and incidence may go upto 20% while the arthritic disease may have an incidence of 10-20%
Another complication in the evaluation of CAEV epidemiology is its close relation with MVV in sheep. The question is whether goat disease could be caused by sheep agent or vice versa. There is recent demonstration that arthritis and mastitis also occur as a complication of MVY of sheep. It keeps the issue of cross species infection alive.
The main method of dissemination of virus of CAE may be via feeding of infected clostrum. Transplacental spread of infection to foetus by maternal blood does not appear to be major method of virus spread.

Retroviridae 341
The infection in goats persists often for life and u'1ese animals become virus shedders either via colostrum or via respiratory secretion. The infection is subclinical and the virus spread is insidious. The bottle feeding of infected kids as well as physical contact due to handling of kids also fosters virus spread.
Pathogenesis: CAEV and MVV leads to persistent infection. These viruses can sequester themselves in host cells by integrating their provirial DNA into- host cell DNA and these elude immunologic elimination. The viruses replicate in macrophages and these viruses do not usually induce virus neutralizing antibodies. Due to these reasons the virus replication can thus continue independent of any control by humoral immune system.
Infection of sheep and goats with their lentiviruses results in persistent systemic infection during which the virus infects and replicates in the cells of monocyte-macrophage series. The clinical cases show low grade systemic infection i.e. non productive infection in monocytes. The macrophages in organs with lesions may be the sites for virus replication. Newborn goats are susceptible to infection of CNS with CAEV but isolation of virus form CNS of persistently infected adult animals is rare except with encephalitic disease. This also applies to animals with chronic arthritis and/or pneumonia. Experimental infection also corroborates the rarity of CNS involvement. Intracerabral inoculation with virus results in virus replication in brain with acute leukoencephalitis. Viral antigens produced by these cells elicit CMJ response which cause the immunopathological consequences.
CAE causes at least 3 disease syndrome in nature. The most frequently encountered is arthrititic disease in adult goats. The arthritis is insidious in onset and progresses slowly over a period of months to years. Joints, bursae and tendon sheaths are targets of disease but the common and severe localisation is in the carpal (knee) joints. The hock and stifle joints are involed to a lesser extent. Disease is common in animals of 2-9 years of age and longer the disease duration the greater is tissue damage. The affected animals are weak and have long coarse poor hair. The affected animals are afebrile and maintain good appetite. The histopathological lesions are characterized by proliferation of synovial membrane with development of villous projections into the lumen of joint Later there is necrosis of collagen structures.
In addition to arthritis the lactating animals have a chronic

342 Textbook o/Veterinary Virology
inflammatory lesions in the mammary glands. These lesions consist of marked lymphoid hyperplasia. The infammatory cells are of the same type as in arthritis lesions like lymphocytes, plasma cells and macrophages.
Rapidly progressive neurological disease of young goats of 2-4 months old show this syndrome. The earliest signs are posterior paresis and ataxia or weakness in hind quarters. It progresses and includes front legs. The kids become recumbent and die. Goats with encephalomyelitis have rough hair coats and have muscle atrophy.
Immune reaction: Humoral and CMI response to the virus play important role in infection but does not produce any benefit to the host.
The cellular immune response in sheep inf~cted with MVV and goats infected with CAEV is virus specific. The CMI response correlates with the onset of encephalitis, pneumonia and arthritis and persists as long as inflammation persists. The sensitized lymphocytes are important in causation of the inflammatory disease and cannot be answered directly. Indirectly it has been proved that virus does not cause primary cytopathic effect in tissues as it does in cell cultures.
Although CAEV and MVV have similar pathogenic mechanism and share major antigenic determinants, the CAEV is distinct from MVV. In competitive hybridization assays of radiolabelled RNA of these two viruses show only 20% homology with each other.
Diagnosis: As the animals are persistently infected after exposure to virus, the demonstration of antibodies indicates infection. AGID is the test of choice. ELISA is also being employed. Virus isolation can be done from 10-20 ml of blood collected over anticoagulant. The buffy coat cells are separated and cocultivated with normal goat synovial membrane cultures. The cocuIture is maintained for 2-3 weeks at 37°C and examined for CPE.
Control: The hyperimmunization of animals with live or inactivated virus preparation fails to protect these animals. Control measures rest on the identification of infection and elimination of the positive animals.
Jaagsiekte (Ovine Pulmona~y Adenomatsis) Virus
Jaagsiekte was reported about one hundred years ago in South Africa but the viral etiology was known recently and identified as retrovirus. It does not seem to be related to either the known

Retroviridae 343
lentiviruses or oncoviruses. The rust indication of its being retrovirus was the observation of particles possessing type C morphology in jaagsiekte lungs and biochemical evidence for the presence of particles with reverse transcriptase activity in lung extracts as well as demonstration of morphologically typical retroviruses in cell cutures of adenomatous lungs.
Properties of the virus: The jaagsiekte retrovirus (JSRV) is about 104 nm in diameter. The mature virus particles are electron dense with a close fitting envelope. A slightly eccentric nucleoid can sometimes be seen under electron microscope. The overall morphology is typical of a retrovirus but can be distinguished in finer details. Morphologically JSRV seems to be closely related to mouse mammary tumor virus (MMTV). The JSRV does not show any cross relation with MVV related lentiviruses.
All attempts to grow JSRV in a variety of cell cultures have not been successful.
Epidemiology: The domestic sheep is the main host affected by JSRV but there are reports regarding the disease among goats as well but these reports are not authentic. Experimentally the infection can be transferred to newborn kids. The infection cannot be established in laboratory animals except successful transplantation of cultured tumor cells into nude mice. Jaagsiekte is widely prevalent. The disease is endemic in most European, African, Middle East and South American countries. Sporadic cases have also been diagnosed from USA and Canada.
The incidence of flock varies from 5-20%. The infection i's airborne and close contact facilitates the spread of the disease. The genetic predisposition exists. Certain breeds of sheep in Iceland were found to be highly susceptible than others. The disease in its natural form is a slow virus infection as symptoms are rarel y seen in animals younger than 3-2 years. Once animal shows symptoms, the prOgressive proliferation of cancerous cells leads to death. 100 martality of the d~ease cannot be regarded as 100% as certain individuals have the ability to contain tumorous lesions.
Pathogenesis: The disease can be experimentally uaosmitted by co-habit3tion, by droplet infection, aerosal spray and by inoculation of extracts of adenomatous lungs or virus by intrapulmonary, intrapleural, intratracheal or intranasal routes. Replication of virus takes place in the

344 Textbook ojVelerinary Virology
type IT alveolar epithelial cells, the virus does replicate in other organs or blood. The infection remains localized in the lung and there is no viraemia. The type 11 alveolar epithelial cells and other nonciliated bronchial epithelial cells are transformed to neoplastio cells which proliferate and eventually fill the alveolar spaces. Normal lung tissue is replaced by solid tissue. These tumor ceIls are also surfactantproducing secretory cells. The typical jaagsiekte lungs are therefore oedematous in appearance with copious amount of clear viscous fluid produced which accumulates in the air passages.
It is not yet known that. JSRV is present as a provirus in the genome of sheep cells. There is no evidence of vertical transmission or intra-uterine infection.
The incubation period in natural infection varies from months to years, while in experimental infection the incubation period can be reduced to weeks or even days. The natural infection occurs in young animals. When lesions develop, symptoms are seen and they mainly are various manifestations of dyspnea. There is increased respiration with jerky expiratory movement Ultimately animals show acute respiratory distress. Coughing and moist rales ~e observed. Viscous fluid runs from the nose. Loss of.appetite and emaciation are terminal signs.
At autopsy the lungs are found double in size or 3-4 times the normal weight Initially white coloured nodules are found which expand to form greyish white areas of consolidation. The microscopic lesions consist of an alveolous lined by cuboidal or columnar epithelial cells. These cells proliferate, filling the lumen of the alveolous and forming acinar or papilliform masses.
Immune reaction: There is evidence of acquired resistance to jaagsiekte. No detectable circulating antibodies develop and there is no evidence of cellular immunity. The virus derived from lung fluid is complexed with immunoglobulins predominantly of IgA type. The lung exudate is also rich in IgA and IgG. It is possible that immune response to jaagsiekte virus is mainly of local nature and mediated by IgA. It may be due to absence of viraemia in JSRV infection as the replication of virus is localised in lung epithelium.
Diagnosis: Clinical signs like dyspnea in absence of fever and clear viscous lung exudate is indicative of jaagsiekte. Histopathological examination is method of choice.

Retroviridae 345
Control: Control measures depend upon strict isolation and elimination ofanimals showing symptoms.
Rererences
BUXTON, A. and FRASER, G., 1977. Animal Virology Vol. 2. BlackweIl Scientific Publications, Oxford, London, Edinburgh.
CHEBVERS, W.P. and McGuire, T.C., 1988. The lenliviruses: maedivisna, caprine arthritis-encephalitis and equine infectious anaemia. Adv. Vet. ResJ4: 189-215.
CRAWFORD, T .B.; ADAMS, D.S.; CHEEVERS, W.P. and CORK, L.C., 1980. Chronic arthritis in goats caused by a retrovirus. Science, 207, 997-999.
FENNER, I.F., 1980. Bovine lymphosarcoma. Adv. vet Sci. Corn. Med. 24, 1-68.
HAASE, A.T., 1986. Pathogenesis of lenlivirus infections. Nature, 322, 130-136.
HOFSTAD, M.S., CALNEK, B.W. HELMBOLDT, C.F., RElD, W.M. YADER, H.W. IR. 1972. Diseases of Poultry. 6th Ed. Iowa State University, Press, Ames.
KAHRS, R.F., 1985. Viral diseases of cattle. Kalyani Publishers, New Delhi.
LuCAs, M.H. and ROBERTS, D.H., 1982. Transmission of bovine leulwsis virus (BLV) Curr. Top. Vet. Med. Vet. Sci.15, 264-266.
MELNICK, I.L., 1982. Taxonomy and Nomenclature of viruses. Progress Medical Virology. 28: 208-21.
MOHANTI, S.B. and DUITA, S.K., 1981. Veterinary Virology, Lea and Febiger, Philadelphia.
NARYAN, 0.; Cl£'v!E.'ITS, 1.E.; SlRANDBERG, I.D.; CORK, L.C. and GRIFFIN, D.E., 1980. Biological characterisation of the virus causing leukoencephalitis and arthritis in goats. I.Gen. Virol. 50, 69-70.
NARYANA, O. and CLEMENTS, 1.E., 1989. Biology and pathogensis of lentiviruses.1. Gen. Viral. 70: 1617-1639.
NOTKlNs, A.L. and OLDSTONE, M.B.A., 1984. Concepts in viral pathogenesis. Springer-Verlag.New York, Berlin Heidelberg Tokoyo.
SlRAUB, O.C. 1981. Enzootic bovine leulwsis. In E.P.I. Gibbs (Editors) Virus Diseases of Food Animals. Vol. I. International perspectives. Academic Press, New York, NY-pp 693-718.
TUSTIN, R.C., 1969. Ovinejaagsielae, 1.S. Afr. Vet. Med. Assoc.40: 3-23.

346 TeXlboolc o!Veterinary Virology
UPPAL, P.K. YADAV, M.P. and AHMm, S.N .. 1981. OCCllTrence of equiM infectious aNlD1Jia in India. Virus information Exchange News letler 4, 45-46.
VERWOI1JtD, D.W; PAYNI!, A.L.; MYER, M.M. and YORK, D.F., 1983. IsolatiOfl and. prelimituJry characterization of jaagsielcJe retrovirus (JSRV). Ondersteport, 1. VeL Res. 50,309-336.
VERWOARD, D.W.; TumN, R.e. and PAYNI!, A.L., 1985. Jaagsiekte: An infections pulmonary adenomatosis of sheep. In O. Olsen Karkowka and J.R. Blakeslee (Editor) Comparative Pathology of viral diseases. eRe Pr~s, Boca Raton, F.L. pp.53-76.
Virus, Information Exchange Newsletter for South East Asia and the Western pacifIC. 1987. Vo1.4 No. 2. Australia.

Chapter 30
Bunyaviridae
Bunyamwera is the name of a place in Uganda where type species, Bunyamwera virus was isolated. Bunyaviruses, with the exception of hantaviruses, are transmitted by arthropods. The family contains more than 200 viruses, in five genera: Bunyavirus, Phelbovirus, Nairovirus, Unkuvirus and Hantavirus. The genera of Bunyavirus and Nairovirus are organised into various serogroups. The genera of Phelbovirus, Unkuvirus and Hantavirus at present have a single serogroup. The viruses is this family are medium sized, spherical or oval measuring 90-lOO nm in diameter. They possess a membrane envelope with projections. The projections are glycoproteins, designated Gt and Gz• The genome consists of three molecules of a single stranded noninfectious, RNA large (1.), medium (M) and small (S), which are surrounded by a nucleocapsid protein (N). A minor large protein(L) is probably a transcriptase. Most of the genome is (-) sense buUt has been shown with some viruses (phelbovirus) that the 5' end of the S segment is (+) sense, the term ambisense is used to describe this genome. Virions contain four major proteins: a transcriptase(L), nucleoprotein (N) and two glycoproteins (G t and G~).
The virions are sensitive to lipid solvents and detergents. The viruses replicate in the cytoplasm and mature by budding into smooth surfaced vesicles in Golgi region or nearby. The viruses readily grow in many kinds of cell cultures e.g. Vero cells, BHK-21 cells and mosquito cells. Most of the viruses grow to high titre is suckling mouse brain. The virus gains entry into host cell by fu!;ion of viral envelope with cell

348 Textbook of Veterinary Virology
membrane. The replication takes place in the cytoplasm. After penetration the virion transcriptase is activated and transcribes subgenomic mRNA from each of virion RNA and a second round of transcription takes place. The lRNA segment codes for virion transcriptase and M segment codes for glycoproteins (G1 and G1). The sRNA in case of Bunyavirus codes for nucleoprotein and a nonstructural protein. In case of Phelbovirus, sRNA also codes for two proteins but it employes an ambisence transcription strategy. The viruses mature by budding in Golgi vesicles and are released by fusion of the vesicle membrane with plasma membrane and exocytosis or by cytolysis.
Rift Valley Fever Virus (RVFV)
Rift valley fever (RVF) is a mosquito borne disease that cause devastating epidemics among sheep, goats, cattle and human beings. The disease is characterized by its epizootic nature, short incubation period, fever and typical focal to diffuse necrosis of the liver. The disease was first reported from Kenya in 1912 and the virus was isolated in 1931. RVF is now reported form most of sub-Saharan Africa and the Nile Valley. RVF has not been reported so far outside African continent.
Properties of the virus: The virions are spherical in shape bounded by host cell derived membrane with virus coded glycoprotein spikes, the diameter of the virion is 85-100 nm. The morphogenesis of Bunyaviridae is consistent with helical symmetry of the nucleoprotein core. The virions are formed by budding through smooth endoplasmic reticulum with many virions in Golgi cisternae which communicate with extracelluar environment.
All isolates have been found to be identical by using serologic tests. The difference in strains can be detected by RNA fingerprinting. RVFV is sensitive to lipid solvents, detergents and low pH. The virus retains its infectivity at 4°C in the presence of protein and at neutral or alkaline pH for about 4 months and for 6 months in 0.5% phenol. In serum the infectivity is retained upto 3 hours at 56°C. Formalin inactivates the virus even at low concentration. In human RVFV vaccine the inactivation is done at 0.3% final concentration for 72 hours at 37°C. Betapropiolactone 0.1 % also is used to inactivate the virus. RVFV contaminated surfaceS are sterilized effectively with bleaching powder, sodium or calcium hypocholorite.

Bunyaviridae 349
Cultivation: The virus readily grows in the chicken embryos via yolk sac or by chorioallantoic membrane route. It can also be propagated in mouse brains, in foetal rhesus monkey lung cells and in kidney cultures of lamb and vero cell line. Acidophilic intranuclear inclusions are found in the infected cells.
Epidemiology: In Africa the disease is limited to domestic ruminants i.e. sheep and cattle. Goats are variable than sheep or cattle with respect to severe disease. The disease progression and severity are inversely proportional to age. The adult sheep and cattle suffer 10-30% mortality while the animals less than 7 days old may have fatality rate of 100%. Outbreaks are characterized by abortion and rapidly fatal neonatal disease. Equines and camels develop antibodies in the absence of disease. The disease is spread by mosquitoes the natural host reservoirs are unidentified. Mosquitoes of many species of the genera Aedes, Anopheles, Culex, Eretrnapodites and Mansonia transmit RVFV under field conditions.
Blood and serum from RVFV infected animals are important infectious sources for human disease.
Palhogellesis: Arthropod transmission is the main mode of disease spread, although infections following cOnlac~ with infected tissues or inhalation of aerosals may occur. The liver is the primary site of replication. In pregnant animals there is a predilection of virus for placentomes via haematogenous route. The rapid foetal death is due to direct viral infection rather than generalized placentitis. There is rapid progress from mild hepatocellular changes to massive necrosis. Haemorrhagic diathesis, reduced prothrombin levels and prolonged clotting time has been observed in experimentally infected animals.
The clinical signs observed are related to species and age of animals involved. The younger animals are not most susceptible. The incubation period in sheep and cattle is 4-6 days but is shorter in experimentally infected animals. The disease appears as paracute, acute, mild or sometimes inapparent. The paracute form may be seen in new born and very young lambs which die without showing clinical symptoms. The acute form is encountered in young lambs, kids and calves while mild form is seen in adult sheep and goats. The adult cattle may experience the mild or inapparent form. The most prominent clinical manifestation is severe icterus and abortion amongst pregnant animals. In young lambs the disease is characterised by prostration and

350 Textbook of Veterinary Virology
death. The animals are listless, off feed, pass bloody wine, and exhibit diarrhoea, dysponea and muscular tremors. There is an elevated temperature upto about 106°F. Abortion is a dominant feature in adult cattle apart from elevated temperature, buccal erosions, diarrhoea, necrosis qf skin over udder and scrotum and cessation of milk production. In human beings there is high fever, icterus, nausea, epistaxis, headache and muscle pains. Recovery usually takes about 10-14 days. The disease may tenninate fatally in babies infected through the milk of infected mothers.
The most striking features at necropsy is extensive jaundice because of focal hepatic necoris. There may be oedema, congestion or hamemorrhange of gall bladder as well as scattered hamorrhages throughout the body. The kidney are of mottled appearance and show petechiae on the surface. The gestrointestinal tract mucosa is hyperaemic, haemorrhagic and lymph nodes are generally enlargcd and soft Histopathologic examination confIrms the haemorrhagic nature of lesions. There is focal and diffuse necrosis of the liver with acidophilic inclusion bodies in the liver ceils.
Immune reaction: After an incubation period of 1-4 days and a viraemic phase of 1-4 days, humoral antibodies appear and risc mpidly. The antibodies can be measured by neutmlization, haemagglutination inhibition, complement fixation, ELISA and immunofluorescence tests. The antibodies persist in low titre after 1-2 years Post infection. The maternal antibodies protect the lambs upto 3 months. The immunogenicity of live attenuated vaccine is superior to inactivated vaccine. High titre antibodies appear after inoculation with live vaccine.
In human beings it is speculated that the encephalitic and ocular clinical RVF syndrome is due to immunopathological component.
Diagnosis: High abortion rate in cows and ewes, increased mortality among lambs and calves less than 7 days of age, extensive liver lesions, mosquito season with high mosquito population and human febrile disease are suggestive of RVF outbreak:. Earlier the disease had a limited geographical area i.e. sub-Saharan Africa but now the disease has spread over to Egypt and most countries in Africa. When the disease in suspected, virus isolation should be attempted in mouse brain and cell culture. Blood samples and serum collected at the height of fever, specimens of liver, placenta and foetus are the

Bunyaviridae 351
specimens to be used for virus isolation. Samples from liver, brain, kidney, heart and spleen, are collected for serological diagnosis. Histopathological examination of liver reveals characteristic necorisis of hepatocytes associated with semi demarcated foci of primary coagulative necrosis. In acute hepatic lesions, haemorrhages are widespread with no inflammatory response. Eosinophilic intranuclear inclusion bodies may be seen in degenerating hepatocytes.
Control: Immunization of susceptible animals is the most effective mechanism of control. Two types of vaccines are used. The modified live virus 'vaccine is stable w~en properly lyophilized and is highly effective. It gives a long lasting immunity. The protection is induced after 6-7 days of administration and the off spring of the vaccinated animals remain protected upto 5 months of age. The vaccine is not recommended for pregnant animals as the vaccine is abortogenic and teratogenic. Formaline inactivated vaccines are effective when multiple inoculations are done. The immunity produced is not long lasting and therefore yearly boosting is essential. The vaccine can be administered in pregnant animals.
Akabane Virus
Akabane virus infection during early pregnancy causes abortion, premature delivery, stillbirths and a foetal anomaly in cattle, sheep and goats that is known as arthrogryposis hydranencephaly (AH) syndrome. The virus causes disease in Japanese cattle. The syndrome in cattle, sheep and goats has also been reported in Australia and Israel. The virus was fIrst isolated from mosquitoes in Japan in 1959 (MaLSuyama et aI, 1960; Oya et ai, 1961) - Akabane is the name of the village where the virus was fIrst isolated.
Properties of the virus: The virus particles are enveloped roughly spherical, variable in size, 70-130 nm in diameter. Most of the virions have a ragged closely adherent envelope with 9 nm peplomers. It is inactivated by ether, chloroform and sodium deoxycholate. The virus is readily inactivated ~t pH 3 and trypsin. It is also heat labile loosing infectivity about 0.3 log per hour at 37°C. The virion contains a single stranded RNA genome of three size classes (L, M and S). There appear to be five proteins in virus infected cells L, Gp Gz' N and one small nonstructural protein. The virus shows haemagglutination and the haemaggluting activity is structurally associated with the virus particle.

352 Textbook o/Veterinary Virology
The haemagglutination is dependent upon the pH as well as NaCI molarity of the diluent Duck, goose and pigeon erythrocytes are agglutinated. Cattle, sheep, guinea pigs, day old chicken and human erythrocytes are not agglutinated. Haemagglutination is specifically inhibited by antisera. The virus also shows haemolytic activity. It lyses pigeon erythrocyte at 37°C.
Cultivation: The virus replicates in the cytoplasm. For primary isolation the brain of suckling mouse (1-2 day old) is the most sensitive system of viral isolation. The virus can also be cultivated in primary cell cultures and celIlines with the production of CPE. The virus can be propagated in the chicken embryo by yolk sac inoculation. Inoculated embryos show dwarfism, cerebral defects, hydroencaphalus, deformed legs and toes and arthrogrypsis.
Epidemiology: The result of virus isolation and serological tests carried out in different parts of the world indicate a wide distribution of Akabane virus among cattle and other domestic animals in many southeast Asian countries, the Arabian peninsula. the Middle East and Africa. Apart from cattle, other species found to have antibody are horses, goats, sheep, pigs and monkeys. The biting midge Culicoides brevitarsis a principal vector of virus in Australia. The virus has becn isolated from Aedes vexans. Culex tritaeniorhynchus. and Culicoides oxystoma in Japan, and form Anopheles /unestus in Kenya. In Japan the geographical distribution and seasonal occurrence of the disease and active transmission in summer suggest the involvement of vectors. However, the information on the vector(s) is lacking and mechanism of transmission and survival of virus is not yet know.
Pathogenesis: Congenital AH syndrome is caused by intrauterine infection of foetuses with Akabane virus in pregnant cattle, sheep and goats. The intrauterine infection of foetus can cause abortion, still birth, premature birth and deformities but no clinical abnormalities have been recognised in dams during pregnancy. Arthrogrypotic calves are often born dead, the musculature of limbs and spinal column is atrophied and one or more limbs are fixed. The site of inilial infection is not known, viraemia seems to be a constant feature. The virus does not produce a persistent infection of the foetus. The histopathological examination of infected bovine foetus show encephalomyelitis and polymyositis of skeletal muscles. Immuno fluorescence demonstrates the viral antigen in skeletal muscles and brain tissue. The lesions in· the

Bunyaviridae 353
foetus result in abortion, premature birth, or stillbirth. The foetuses which survive the infection gradually develop brain lesions and reduction in the number of motor neurons in the spinal anterior born. Arthrogryposis may ensue for the damage in the CNS.
An epizootic in cattle may be fIrst noticed by increased incidence of abortions and premature births. The calves born with hydranencephly may survive for several months but they never thrive. The calves are mature but underweight at birth and may show blindness, nystagmus, deafness, slow suckling, paralysis and incoordination.
Immune reaction: The infected animals develop neutralizing, HI and haemolysis-inhibiting antibodies between 7-14 days post infection. The neutralizing antibodies persist for about 2 years. Foetal calves, lambs and kids develop antibodies in uterus betwccn 70-96 days of gestation in cattle and foetal lambs and foetal kids betwccn 30-155 days of gestation.
Diagnosis: Akabane virus infection is suspected when congenital AH syndrome appears sporadically or endemically alongwith stillbirths or abortion. Serum from aborted foetuses, still born calves and dams can be tested for SN antibodies. The virus can be isolated from foetuses and still born calves, from blood of cows by intracerebral inoculation of suckling mice.
Control: There are two approaches of control. The vector control and vaccination. The vector control measures are not practical as the knowledge concerning vector is still inadequate. A formalin inactivated aluminium phosphate gel adsorbed vaccine has been developed in hamster lung cultures. Two doses at an interval of 4 weeks are recommended in cattle. The vaccination prevents development of viraemia and infection of foetus is pregnant cattle and goats after challenge with virulent virus.
An attenuated strain has also been developed by serial passages in hamster lung cultures at 30°C. The vaccine has been found to be innocuous in calves but in pregnant ewes it causes intrauterine infection and viraemia.
Nairobi sheep Disease
It is an acute infectious sheep and goat disease transmitted by ticks. The disease is prevalent in African continent. The virus withstands a

354 Textbook o/Veterinary Virology
temperature of 500C for 1 hr and is resistant in blood and serum at 4°C. The virus can be propag·ated in sheep and goat kidney cell cultures and in hamster cell lines in which CPE is produced. Intracytoplasmic inclusions are seen in infected cell cultures. The virus grows well in young mice by intracerebral inoculation. The incubation period of the disease is 4-15 days. The disease is manifested by fever, mucopurulent nasal discharge and increased respiration. There is haemorrhagic gastroenteritis and congestion of lymphnodes. Abortion may take place in pregnant ewes. The mortality ranges between 30-80%. Goats are susceptible to this disease but cattle are not infected. The disease is transmitted by tick and is maintained by tick-sheep or tick-goat cycle. The disease can be diagnosed by isolation of virus from spleen and infected blood in cell culture a,!d mice. Mouse brain adapted inactivated vaccine is available. A modified live virus attenuated by serial mouse brain passage is also available.
References
BUXTON, A. and FRASER, G., 1977. Animal Microbiology, Vol.2, Oxford. Blackwell Scientific Publications.
DAVIES, F.G., 1978. Nairobi sheep disease in Kenya. The isolation of virus from sheep and goats, ticks and possible maintenance hosts. Journal of Hygiene Cambridge, 81, 259-66.
DINiER, Z. and MOREIN, B., 1990. Virus infections of ruminants. Elsevier Science Publishers B.V. Amsterdam.
EASn:.RDAY, B.C., 1965. Rift valley fever, In advances in Veterinary sciences. Edited by C.A. Brrandly and C.L. Coruelius Vol. 10, New York, Academic Press.
GWBS, B.PJ., 1981. Virus Diseases of food animals. Academic Press, New York.
KUROOI, H.; lNABA, Y.; TAKAsHASHI, E.; SATO, K.; <>MORI, T.; MIURA Y.; COTO, Y.; FUJIWARA, Y; HATANO, Y; KODAMA., FUKUYAMA, S.; SASAKI, N. and
MATIJMOTO, M., 1976. Epizootic congenital arthrogryposishydranencephaly syndrome in cattle. Insolation of Akabane virus from infected foetuses. Archives of Virology. 51: 57-74.
LUPTON, H.W. and PETERS, CJ., 1984. Rift valley fever. Proc. U.S. Animal Health Assoc. 87, 279-290.

Bunyaviridae 355
MATSUYAMA, T.; OVA, A.; OoATA, T.; KOBAYASHI, I.; NAKAMURA, T.;TAKAHASHI, M. and Kitaoka., M., 1960. Isolation of arboviruses from mosquitoes collected at livestock pens in Gumma Prefecture in 1959. Japn J. Med. Sci. Bio1.l3, 191-198.
MATIJMOTO, M. and INABA, Y., 1980. Akabane disease and Akabane virus. Kitasato Arch. Exp. Med. 53, 1-21.
MATIJMOTO, M. and INABA, Y., 1980. Akabane disease and Akabane virus. Kitasato Arch. Exp. Med. 53, 1-21.
MOHANlY, S.B. and DUITA, S.K., 1981. Veterinary virology, Lea and Febiger, Philadelphia.
OVA, A., OKuBO,T., OaATA, T., KOBAYASHI, 1. and MATSuyAMA, T., 1960. Akabane, a new arbovirus isolated in Japan. Jpn. J. Med. Sci. Bio1.14, 101-108.
POR1ERFIELD, J.S. and DELLA-PORTA, A.I., 1981. Bunyaviridae: Infections and diagnosis. In E.Kurstak and C.Kurstak (Editors) comparative diagnosis of viral disease. Academic Press, New York, Vol. 4 pp. 479-504.
RUSSELL, P.H. and EDINGTON, N., 1985. The Burlington Press (Cambridge) Ltd. Foxton, Cambirdge.
The Rift Valley fever, Office International Epizootics Technical Series No.l, 1981.

Chapter 31
Toroviridae
Toroviridae is a newly created family for a group of antigenically related viruses demonstrated in horses, cattle and men. Two new viruses detected in fecal material from horse and cattle in 1982 and 1983 with unique morphological (Latin torus-a doughnut shaped ring), biochemical and serological characters. Toroviruses are enteric viruses recognised in three different species, namely horse, cattle and human beings. Evidence of infection has been obtained in pig, sheep, goat, in lagomorphs and rodents.
Toroviruses are enveloped RNA viruses with peplomer bearing envelope and an elongated tubular nucleocapsid of helical symmetry. The nucleocapsid may be bent giving kidney or disk shaped morphology to the virus particle or the neuclocapsid may be straight resulting in rod shaped virion. The genome is single stranded RNA with positive polarity. The moleular weight is about 8 x 106
• There are two major proteins in the virus particle, a phosphorylated nucleocapsid polypeptide of 20K and a 22K protein the main constituent of envelope. Additional proteins of 37K and 75-100K range are also present in the virus particle. Toroviruses replicate in the cytoplasm where four subgenomic mRNAs are formed. The replication is dependent on some nuclear function of the host cell.
Breda Virus
In 1972 a virus was isolated from a rectal swab of horse with diarrhoea in Berne (Switerland). In 1982 Woode et al. described the isolation of virus from epizootic of neonatal calf diarrhoea in Breda,

Toroviridae 357
Iowa. The resemblence of 'Berne virus' and 'Breda virus' particles was described some 10 years later. In 1984 particles resembling Berne virus (BEY) and Breda virus (BRV) in stoqls of adults and children with diarrhoea, which reacted with antibodies against BEV and BRV were described.
Properties of the virus: In negatively stained preparations of intestinal contents, faeces or tissue fluids, the virus particles are either elongated or kidney shaped with a diameter of 105-140 x 12-40nm or spherical forms measures 82 nm in diameter; with a pcplomer bearing envelope. The peplomers may be short (7-9 nm) or long (20 nm). Many particles have short stubby peplomers which make the virus easily distinguishable form coronavirus. The jejunal epithelial cells infected with BRV show brick shaped or elongated particles with dimensions 80-100 x 35-42 nm.
The BRV has two serotypes BRVl and BRV2. This virus is anligenically related to BEV. BRVl and BRV2 carry common antigens measurable by IF and ELISA. HI has shown clear differences between BRVl and BRV2. BRV possess the haemagglutinating activity. Mouse and rat cells are the only cells agglutinated. The serotype 1 does not elute from erythrocytes while serotypes 2 does. The BRV looses infectivity if faeces are stored at 4°C for 2-3 weeks. It is probable that the virus is readily destroyed by disinfectants and heat. The virus does not grow in cell culture.
Epidemiology: Breda was isolated from acute enteritis of calf aged 5 days. About 56% of calves in the herd developed diarrhoea in ftrst 20 days -of life. Fifteen percent of calves died. The natural spread of virus takes place by fecal/oral route. The subclinical excretion of rotavirus by sows at parturition has been conftrmed. The bovine dams are considered to be source of neonatal virus infections. There are no reservoir hosts or vectors involved in the spread of virus. The incubation period is as short as 24 hours, experimentally. The serological survey carried out in parts of USA, the Netherlands and Germany by ELISA have shown antibodies against BRV in calf and cow sera to the extent of 85 to 94%.
Pathogenesis: The virus fed to colostrum deprived calves or gonotobiotic calves aged 1 hour to 24 days infects the intestine and reaches faeces within 24-72 hours. Diarrhoea or change in appearance of faeces conicides with the virus or change in appeearance of faeces

358 Textbook o/Velerinary Virology
conicides with the virus excretion. After recovery the virus can be excreted for atleast 4 months. In colostrum fed calves from immune dams the infection cannot be prevented but symptoms are mild. Viraemia has not been reported.
The calves develop anorexia and depression followed within few hours by greenish yellow or yellow diarrhoea. Some calves may show shivering. The signs of dehydration reddening and loss of tone of intestine are noticed. Villus atrophy takes place due to infection of crypt and villus epithelial cells. There is focal necoris and moderate inflammation of the small intestine.
Immune reaction: Following primary infection with BRV, the animals develop specific IgM and IgG antibodies. The peak titre is attained 1-2 and 3-4 weeks. The IgA antibodies can also be detected within 2 weeks. The passively derived antibodies do not protect calves from infection.
Diagnosis: BRV does not grow in vitro. The EM detection of virus using 3-4% potassium phosphotungstate, pH 7.0, is used to identify ~ virus. The BRV can be distinguished from coronavirus by !EM. Viral haemagglutinin titres of infected calves fecal samples vary from 20,000 to 50,000 while the titre in normal calves is 16-32. ELISA test is used to survey serum antibodies in the herds.
Control: There is no specific control measure available as no vaccine has been developed so far.
Berne Virus
In 1972 a virus was isolated from rectal swab of horse with diarrhoea in Berne (Switzerland). Berne virus (BEV) has been cultured in vitro, in horse kidney cultures. It can also be cultured in other cells of equine origin. In the presence .of actinomycin D and aminitin the BEV replication is drastically reduced when drugs are added during the first 8 hours of infection, indicating that BEV replication depends upon some nuclear funclion of the host cell. The BEV agglutinates the erythrocytes of human '0' group, rabbit and guinea pig but not fJf rat
and mouse. The virus shows cross neutralization with Breda virus. There is wide distribution of Berne virus in Swiss horse population and antibodies also occur in sera of cattle, sheep, goats, pigs. laboratory rabbits and two species of wild mice.

Toroviridtu 359
The pathogenic role of BEY has not been proved. It cannot be said that BEY was responsible for diarrhoea from the horse it was isolated.
References
HOR2JNBX, M.C.; FLEWETI', T.H.; SAIF, L.I.; SPAAN. I.M.; WEISS, M. and WOOD, O.N., 1987. A new family of vertebrate viruses. Toroviridae Intervirology.27, 17-24.
Wmss, M.; and HORZINEK, M.C., 1987. Theproposedfamily Toroviridae Agents of enteric infections. Arch. Virol. 92, 1-15.
Woode. O.N.; Reed, D.E.; RUJUlels, P.L.; Herrig, M.A and Hill, IT., 1982. Studies with an unclassified virus isolated from dimrhoeic calves. Veteminary Microbiology 7: 221-224.
WOOD£. O.N.; MOHAMMED, K.A.; SAIF, L.J.; WINAND, N.I; QUESADA, M. Kaso, N.E. and POIll.ENZ, IF., 1983. Diagnostic methods for the newly discovered 'Breda' group of calf enteritis inducing viruses. In: Proceedings of the third sympsosium. World Association of Veterinary Laboratory Diagnosticians pp.533-538.

Chapter 32
Unclassified RNA Virus
Borna Disease Virus
Boma disca<;e virus is a slow viral disease of horses that affects central nervous system. The disease is prevalent in Germany and othcr parts of Europe. The name of disease has been derived from the city of Boma, Gcrmany where large number of horses died in 1899.
The detailed morphology of virus has not been worked out but the virus is smaller than 100 nm with an envelope. The virus is heat and acid labile. The virus particles contain RNA. There is only one antigenic type of boma disease virus. The virus multiplies in many varieties of cell cultures from many species of animals but monkey kidney cell lines (MS) are most suitable. The CPE does not appear but the presence of virus can be detected by IF or by acridine orange staining. Intranuclear inclusion bodies appear in some cells after several weeks of incubation. The virus remains closely associated with cellular structures and persists through several subpassages in cell lines without losing its infectivity. The virus can also be propagated on the chorioallantoic membrane of 11 days old chicken embryos at 35°C .
. Boma disease is an infectious encephalomyelitis of horses but natural infection also occurs in sheep. Experimentally most domestic and laboratory animals can be infected. Hamsters and young rats are most suitable experimental animals. The incubation period of natural disease is about 30 days but experimentally the disease in young rabbits appear between 15-20 days. The affected horses show slight fever,

Unclassified RNA Virus 361
anorexia, excessive salivation, lassitude and constipation. As the clinical picture develops there is drowsiness followed by restlessness, biting, kicking, excitability and convulsions. Paralytic signs appear in early stage of disease and frequently take the form of generalized paresis. The course of disease is about 1-3 weeks and mostly the animals do not recover. The mortality rate is about 90%.
The virus is present in saliva and nasal secretions during acute illness of horses. In the surviving horses the virus can be recovered for about 6 months after recovery. There are no gross lesions at necropsy but typical encephalitis affecting brain stem is observed on histopathological examination of brain. The'intranuclear inclusions can be seen in the neurons. The spread of infection results from contamination of food and water, by virus shed in the saliva and nasal secretions during acute illness of horses. The virus is also excreted, in the milk and urine. The outbreaks appear in early spring and reach a peak in warm damp weather and disappear in the autumn. It suggests that arthropod vectors play an important role in the spread of infection. The virus has also been isolated from the brains of herons and other wild birds, as well as from ticks of the genera Hyalomma, dermacentor and ornithodoros.
The presence of intranuclear inclusion bodies in the nerve cells of the brain is diagnostic. The virus can also be detected in infected cell cultures by IF test. The antibodies in the serum and central nervous system can be detected by IF test.
The recovered animals are immune but the duration of immunity is not known. Infected rabbit or horse brain tissue inactivated with phenol or phenol glycerol is used for vaccination. A single dose of vaccine is administered subcutaneously provides protection for about a year.
Reference
MAYR, A. and DANNER, K., 1978. Borna-a slow virus disease. Comp. Immunol. Microbiol. Infect. Dis.l, 3-14.

Chapter 33
Unclassified Agents
Scrapie
Scrapie is an infectious and chronic degenerative condition of thc CNS of sheep and goats caused by a small and uncharacterized agent. The disease is endemic in UK but has been exported to Canada, Australia, USA and India by British shccp.
Properties of the virus: The agent is small and is filterable through a 40-50 nm filter. It is not known whcther the agent has a small nucleic acid or no nucleic acid. It is suggested to be a small hydrophobic complex of protein with an outer layer of protective glycolipid and inner nucleic acid or a self replicating protease whose enzymic activity produces more of the agent from precursors found in normal brain. The agent is extremely resistant to protein and nucleic acid denaturants; e.g. heat, formalin, propiolactone, ultraviolet light, nucleases, strong acids and alkalies. The agent kills mice several months after inoculation and
has also been transmitted to rats and primates but no laboratory worker has so far picked up the infection. Cells from the brains of infected sheep and mice have been grown in culture and the replication of the agent has corresponded to the replication of host cells. A continuous cell line has been established from scrapie infected mouse brain, which produces the scrapie agent continuously. The scrapie agent has also been adapted to hamsters.
Pathogenesis: Sheep and goat can be infected by all routes of inoculation with infected brain tissue. The incubation period in natural cases is 2-3 years but experimentally it is 6-9 months. The disease is

Unclassified Agents 363
usually seen in sheep and goats, 3 years of age and older. Following inoculation the agent is first recoverable from lymphoid tissues and 2-3 months later from the medullary region of brain. In the mid brain the neurons become vacuolated and then shrink and this is accompanied by the formation of myeloid plaques and proliferation of astrocytes. The sheep first appear hyperexcitable with an erect bead and high stepping gait Later on there are muscle tremors, intense pururitis or stuper and finally ataxia develops. Deaths are most frequent in animals at 2-5 years of age and 10% of the stock may die in endemic area.
The remarkable feature of scrapic agent is the lack of humoral and CM! responses in the infected animals.
No anitbody or inflammation is produced in the affected animals. Infection is congenital either via ingestion of amniotic fluid or close contact Contaminated vaccines may also spread the disease.
Diagnosis: This is' based on clinical symptoms. A definitive diagnosis can be made by inoculating normal susceptible sheep or goalS with suspected material and observing the development of clinical disease as well as the characteristic histopathological lesions in the central nervous system. There is spongiform degeneration and severe vacuolation of neuronal cytoplasm. Hypertrophy of astroglial cells is also characteristic.
Control: There is no vaccine available. The only method available is to slaughter all the affected animals and animals those had contact with the affected animals during past 3-4 years.
References
CARP, R.I.; MERz, P.A.; KAsESAK, R.J.; MERG, G.S. and WIsnieioski, H.M., 1985. Nature of the scrapie agent. Current status and facts and hypotheses J. Gen. Virol. 66, 1357-1368.
KIMBERLlN, R.H., 1976. Slow virus disease of Animals and Man (Research Monographs Frontiers of Biology 44) North Holland/Elsevier Amsterdarn/New York, 264 pp
PRUSINER, S.B. and HADLOW, WJ., 1979. Slow Transmissible disease of the Nervous system. Academic Press, New York Voll,472 pp and Vol2, 52 pp.
TYRELL, D.A.J., 1979. Aspects of slow and Persistent virus Infections (New Perspectives in clinical Microbiolgoy 2). Martinus Nijhoff, The Hague, 286 pp.

Inilex
Abinanti, 151 Abortion, 180 Acyclovir, 109 Adcnoviridae virus, 15, 23-24,
164-74 avian adenoviruses, 171-74 bovine adcnoviruses, 165-67 canine adcnoviruses, 169-71 control of, 167, 169, 171, 174 cultivation of, 166, 168, 170, 172 diagnosis, 167, 169-71, 174 effect on egg production, 173-74 epidemiology, 166, 168, 170, 172 equine adenoviruses, 171 irnmunereaction, 167 inclusion body hepatitis, 173 infectious canine lary-
ngotracheitis virus, 171 ovine adenoviruses, 167-69 pathogenesis, 166-70, 173-74 porcine adenoviruses, 171 properties, 165-70, 172 respiratory disease, 173
Adenoviruses, 94, 96,165-74_ African horse sickness virus
(AHSV), 260-62 African swine fever virus (ASFV),
206-08 Agar gel double inununo diffusion
test, 118, 121 Alimentary system, disease of,
180-81
Alphaviruses, 235-38 Anatid herpesvirus, 197-98 Animals,
entry ofviruses in, 68-71 viruses, 20-33,90-92
c1assifaction, 20-33 transmission of, 90-92
Antibiotics, 109 Antigens, in infected cells, 67-68 Anti-idiotype vaccine, 107 Antiviral drugs, 107-08 Arabinofumosyl nucleosides, 109 Arboviruses, 12-13 Arcnaviridae, 17, 29 Arterivirus, 245-46 Astroviruses,19 Attenuated virus vaccine, 102-05 Avian adenoviruses, 171-74 Avian encephalomyelitis virus
(AEV),226-27 Avian infectious bronchitis virus
(lBV),274-76 Avian infectious laryngotracheitis
virus, 194-96 Avian influenza virus, 281 Avian leukosis, 329-33 Avian paramyxovirus-3, 289 Avian paramyxoviruses, 285-89 Avian reovirus, 254 Avian reticu1aendotheliosis
viruses, 331-33

Index
B-virus, 193-94 Bacteria, properties of, 4-5 Baudet, A.E.R.F.,197 Beach, I.R .• 285 Beijerinck, 3 Benzimidazols.109 Berne virus. 358-59 Birnaviridae viruses. 17, 29,
264-66 Bittner, 333 Blue tongue virus (BTY), 257-60 Border disease virus (BOY), 241-42 Borna disease virus, 360-61 Bovine adenoviruses (BAY), 165-67 Bovine coronavirus, 268-70 Bovine enteroviruses, 225-26 Bovine ephemeral fever virus,
319-20 Bovine herpes virus-l (BHY-l),
177-82 Bovine herpes virus-2 (BHY -2),
182-83 Bovine herpes virus-3 (BHY-3), 187 .Bovine leukemia virus (BL Y),
323-26 Bovine papillomavirus, 160-62 Bovine papular stomatitis virus, 143 Bovine parvoviruses (BPOY),
151-53 Bovine reoviruses, 252-53 Bovine respiratory syncytical
virus (BRSY), 305-06 Bovine rhinovirus-I, 228-29 Bovine rotavirus, 254-57 Bovine syncytial virus (BSY).
333-34 Bovine viral diarrhoea virus
(BybY),238-41 Breda virus, 356-58 Brooksby, J.B., 221 Buffalo pox virus, 135-36 Bungaviridae virus, 19,32-33.
347-54
akabane virus, 351-53 control, 351, 353 cultivation, 349, 352 diagnosis, 350-351. 353 epidemiology, 349, 352 immune reaction, 350, 353 Nairobi sheep disease, 353-54 pathogenesis, 349-50, 352-53 properties, 348, 351-52 rift valley fever virus, 348-51
Bumet,47
Calciviridae, 16-17,27,231-33 Camel pox virus, 136-37 Canine adenoviruses, 169-71
·365
Canine coronaviruses (CCY), :UO-71 Canine distemper virus (CDV),
78,294-96 Canine papillomavirus, 162 Canine, parvov:ruses, 156-57 Caprine arthritis-encephalitis
virus (CAEV), 340-42 Carbohydrate, 12 Cell cultures, 49-54
production of, 50-51 virus growth in, 51-53
Cell mediated cytotoxicity, 83 Cell mediated immunity, 80, 83 Cell transformation, 68 Cellular protein, 66 Chase,3 Chicken embryo, 48 Chlamydia properties, 4-5 Chronic infections, 77-78 Clinical virology, 124-25 Cloned viral DNA, 107 Complement fIXation test (CFf).
118,121 Coronaviridae viruses, 17, 30,
267-76 bovine coronavirus. 268-70 canine coronavirus, 27~71 control, 270. 273, 276

366
cultivation. 274-75 diagnosis, 270, 273, 276 epidemiology, 268-69, 272, 275 faline infectious peritonitis
virus, 271 iminune reaction. 270, 275-76 pathogensis, 269-70, 272-73,
275 porcine coronaviruses, 272-76 properties, 268,272,274
Cow-pox virus, 133-35 Creutzfeldt Iakob diSease, 78 Crowther, I.R., 213 Cytopathic effect, 51, 65-67
DNA cloning technology, 122-23 DNA replication, 41 DNA tumor viruses, 94-101 DNA viruses, 14-16,20-26,129-208
adenoviridae, IS, 23-24, 164-74 herpesviridae, 15-!6, 24-25,
176-202 papovaviridae, 15, 22-23, 159-62 parvoviridae, 15,21-22, 150-57 poxviridae, 14-15,20-21, 12948 unclassified, 206-08
Datt, N.S., 217 Dhanda, M.R., 217 d'herelle, 3 Diagnosis of viral diseases, see,
under Viral diseases Doyle, T.M., 285 Duck hepatitis virus (DHV), 227 -28 Duck herpesvirus-I, 197-98 Duck plaguevirus, 197-98 Duck virus enteritis, 197-98 Dulbecco, 52, 55 Dunne, H.W., 226
Ecthyma (Ort) virus, 14143 Edwards, I.T., 102,285,303 Electron microscopy, 54
TeXlbook of V clerinary Virology
Enders,49 Enteric viruses, 13 Enveloped viruses, 43 Enzyme linked immunosorbent
assay (EUSA), 118, 120-21 Epithelial polyhedral cell, 50 Equine adcnoviruses, 171 Equine arteritis virus (EA V), 24546 Equine encephalomyelits virus,
235-38 Equine herpes virus-I, 188-90 Equine herpes virus-2, 188 Equine herpes virus-3, 190 Equine infectious anaemia virus
(EIA V), 334-35 Equine influenza virus, 279-80 Equine papilloma virus, 162 Equine rhino pneumonits virus,
188-90 Equine rhinovirus, 229 Experimental animals, 4647
Feline calcivirus (FCV), 232-33 Feline infectious peritonitis virus
(FIPV).271 Feline lentemia virus, 326-28 Feline panleucopenia virus (FPLV),
154-56 Feline parvovirus, 154-56 Fibroblast, 50 Fibroma virus, 145 Filoviridae. 19, 33, 356-59 Aaviviridae, 17,28,247-50 Foetus, damage to, 74-75 Foot and mouth disease virus
(FM DV). 77-78, 107,213-23 Foot and mouth disease virus
antigenes, 213 Fowl herpesvirus-I, 194-96 Fowl plague virus, 281-82 Fox encephalitis virus, 169-7] Fraenkel-Conrat, 4

Ind~
Frenkel, H.S., 215 Frosch, 3, 46
Genome, of animal viruses, 57-59 mapping of 58-59
Glucosamines, 109 Goat pox virus, 140-41 Goodpasture.47 Greig. A.S., 177
Haemadsorption inhibition test (HO!), 119, 122
Haemagglutinatingencephalomyelitis virus (HEY),
273-74 Haemagglutination, 52-54 Haemagglutination inhibition (HI)
test, 119, 121-22 Helical symmetry, 7-8 Hen's egg, embryonating, 47-49 Hepadnaviridae, 16,26 Hepatitis B-viruss. 94. 96 Herpesviridae viruses. 15-16. 24-25,
177-202 bovine herpes virus-I, 177-82 bovine herpes virus-2, 182-83 clinical manifestation. 179-81 control, 181-82, 186-87. 190.
192-94,196,198,202 cultivation, 178. 182-84, 188-89,
191. 193-95. 197. 199 diagnosis. 181. 183, 186, 190,
192. 194, 196, 198 epidemiology. 178-79. 183-85.
189, 191-95, 197, 199-200 equine herpesviruses, 188-98 immune reaction. 181. 186,
201-02 malignant catarrhal fever virus,
183-87 Marek's disease virus, 198-202 pathogenesis. 179, 183,
185-86, 192-97,200-01 properties, 117-78, 182-84,
188, 191, 193-95, 197-99 Herpesvirus of goats (CHY-2),
187-88
367
Herpesvirus of sheep (CHY -1), 187 Hershey.3 Hirst, 53 Hog cholera, 242-45
Icosahedral symmtery, 7 Icosahedral virus, 43 Immunity. viral. 81-85 Immuno-electron microscopy, 118,
120 Immunofluorescence. 51 Immunofluoresce'}cil and
immunoperoxidase te~ts, 118-20 Immunopathology, 85 Immunoperoxidase. 51.118-20 Inactivated virus vaccine. 102,
104-06 Inclusion body hepatitis. 173 Infant. damage to. 75 Infestions,
chronic, 77-78 defence mechanism. 80 latent, 76-77 pathogenesis of. 79-80 persistent. 67. 76-80 slow, 78-79 spread of viruses and, 68-72 viral. 81-82. 124-25
Infectious bursal disease virus (!BOY). 264-66
Infectious canine hepatitis virus (ICHY).169-71
Infectious canine laryngotracheitis virus, 171
Influenza virus, 36. 279-82 Interferon, 110-14 Intrnational Committee on

368
Taxonomy of Viruses (lCfV), 14 Intramolecular recombination of
viruses, 59-61 Iridoviridae virus, 16, 25-26 Issacs, 109 ' Ivanovski,3
Jaagsiekte (Ovine pulmonary adenomatsis) virus, 342-45
Japanese B encephalitis virus, 248-49
Jenner, Edward, 102-03
Kendrick, 177 Kilham,75 Killed vaccine, 104-06 Kranoveld, F-C., 285 Kulkami, 0.0., 257 Kulkami, M.N., 257
Latent infections, 76-77 Lentiviruss, 334-45 Lindenman, 109 Lipid, 12 Live virus vaccine, 103-04 Loeffer 3, 46 Louping III virus, 249-50 Lumpy skin disease, 141 Lwoff,4 Lympho choriomenigitis virus
(LCM), 77, 85
Macrophages, 50, 80 Maedivirus,78,337-40 Malignant catarrhal fever virus
(MCFV), 183-87 Mammalian parainfluenza virus,
289-93 Marburg virus, 320 Marek's associated turnor specific
antigen (MATSA), 201-02 Marek's disease virus, 51, 73,
103,178,198-202
Te~book o/Veterinary Virology
Marennkikova, 134 Mastitis, 180 Matsuyama, T., 351 Mehrotra, M.L., 169, 177 Messenger RNA production, 38-40 Milker's node virus, 143 Mohanty, S.B., 198,264 Monolayer culures, 50 Morbilivirus, 293-305 Mornet, 303 Mucosal disease virus (MDV),
238-41 Mukerjee, A., 169 Murine leukemia, 328-29 Murine mammary tumour virus
(MMTV),333 Murty, D.K., 238 Mutation, of animal viruses, 56-57 Mycoplasma, properties of, 4-5
Nair,169 New castle disease virus (NO V),
285-89 Nonenveloped virus, 34··35 Nucleic acid, 9-10 Nucleic acid hybridisation, 122 Nucleic acid sequency, 123 Nyak, B.O., 238
C>ncogenes, 96-98 C>ncoviruses, 323-33 Organ cultures, 51 Orthomyxoviridae virus, 18,30,
278-82 Ovine adenoviruses (OA V).
167-69 Oya, A_,351
Pande, P.G., 238 Papillomaviruses, 94-96 Papovaviridae viruses, 15, 22-23,
159-62 Parainfluenza-l virus, 289

Index
Parainfluenza-3 virus. 289-93 Parainfluenza-5 virus. 293 Paramyxovirus. 284-93 Parihar. N.S .• 183 Paramyxoviridae viruses. 18,31,
284-306 control, 288-89, 292, 296, 302-03,
306 cultivation, 286, 298 diagnosis, 292, 295-96, 301-02,
306 epidemiology, 286, 290-91, 295,
298-99,305-06 Unrnunereaction,292,301 morbilivirus, 285, 293-305 paramyxovirus, 284-93 pathogenesis, 286-88, 291, 295,
299-301,306 pn~ovirus,285,305-06
properties, 285, 290, 294-95, 292-98,305
Parvoviridae virus, 15,21-22, 150-57 bovine parvoviruses, 151-53 canine parvoviruscs, 156-57 control, 153-54, 156 cultivation, 151, 153, 155 diagnosis, 152-54, 156 epidemiology, 151-52, 154-55 feline panleucopenia virus, 154-56 pathogenesis, 152-54, 156 porcrine parvovirus, 153-54 properties of. 151, 153, 155
Pasteur, Louis, 46,102,313 Pathogenesis of viral infections,
65-75 cytocidal infection, 65-67 non cytocidal infection, 65, 67-68 non cytocidal non productive
infection, 65, 67-68 transformation, 66. 68
Persis.tent infections, 67, 76-80
Peste-des-petits-ruminants virus, 303-05
Pestivirus, 23845
369
Picomaviridae virus, 26-27, 213-29 apthovirus, 212-23 control. 222-25, 228-29 cultivation, 214. 224-,28 diagnosis, 218-22, 224-29 enterovirus. 223-28 epidemiology, 215-17, 224-28 irnmunereaction.218 pathogenesis, 217, 224-28 properties of, 212-14,223-24,
226-28 rhinovirus, 228-29
Pigeon herpesvirus, 198 Plaque assay, 52-53 Pneurnovirus,305-06 Pock assay, 53 Polio encephalomyelitis virus,
223-24 Polymerase chain reaction (PCR),
124 Polyomaviruscs, 94-95 Porcine adenoviruscs, 171 Porcine enterovirus-l , 223-24 Porcine enterovirus-9 (PEY -9).
224-25 Porcine herpes virus-I, 191-93 Porcine parvovirus (PPY), 153-54 Pox viruses, see, Poxviridae viruses Poxviridae viruses, 14-15,20-21,
12948 avipox virus, 130, 14648 capripox virus, 129,13841 control, 135-37, 13941, 14345.
14748 cultivation, 131-33, 136-40. 142,
144, 146 diagnosis, 132-33, 135-37.
13940, 142, 144-47 epidemiology, 132, 134, 136-37,
13940, 142, 144-47

370
family, 129-30 immune reaction, 132, 134,
136-37 1eporipox virus, 129, 144-45 orthopoxvirus, 129-38 parapox virus, 129, 144-43 pathogenesis, 132, 134, 136-37,
139-40,143-45,147 properties, 131, 133, 135-36, 138,
140-42, 144-46 suipox virus, 129, 143-44
Proteins, 10-11 Pseudorabies virus, 191-93 Purines antagonists, 108 Pyrarnirdine antagonists, 108
Quantal assay, 53
RNA viruses, bimaviridae. 17, 29,264-66 bunyaviridae, 19.33,347-54 calciviridae. 16-17,27.231-33 coronaviridae, 17,30,267-76 flaviviridae. 17.28,234.247-50 orthomyxoviridae, 18,30.278-82 pararnyxoviridae. 18.31,284-306 picomaviridae, 26-27,211-29 reoviridae. 17,28-29.251-62 retroviridae, 19,32-33.322-45 rhabdoviridae, 18,32.309-20 togaviridae, 17,27-28,234-46 toroviridae, 19.33,356-59 unclassified. 360-61
RNaseL,113 Rabbit papillomaviruses. 162 Rabies virus, 313-18 Radio immunoassay (RIA), 118. 120 Reassortment, 60-61 Remlinger, 313 Reoviridae viruses, 17, 28-29.
251-62
Textbook ofVelerinary Virology
control, 253, 256, 262 cultivation, 258-59, 261 diagnosis,253,256, 260 epidemiology, 253, 255-56, 261 Fiji virus, 251 immune reaction, 260 orbiviruses, 251. 257-62 pathogensis,253,256, 259-61 phytoreovirus, 251 ~perties,252-55,257-58,261 rovirus, 251-54 rotavirus, 251. 254-57
Reovirus, 252-54 Replication groups, 43-44 Replication of viruses, 34-44 Respi!atory viruses 13 Restriction endonuclease digestion
of DNA, 122 Retroviridae viruses, 18, 32, 322-45
control, 326, 328-29, 331, 334, 336, 340. 342, 345
diagnosis. 326-29, 331-33, 334, 336, 339, 342, 344
epidemiology, 324-25, 327, 330, 335,337,340-41,343
immune reaction. 326,339,342, 344
lentiviruses, 334-45 oncoviruses,323-33 pathogenesis, 325-32, 334-36.
338-39,341-44 properties, 323-24. 326-30,
332,334-35,337,340,343 spumaviruses, 333-34
Retroviruses, 96-98 Rhabdoviridae virus, 18,31,309-21
bovine ephemeral fever virus, 319-20
control, 312-13, 318, 320 diagnosis,312,317-18,320 epidemiology, 311,315-16,319 immunereaction,312

Index
marburg virus. 320 pathogenesis. 311-12. 316-17. 319 properties, 310-11. 313-15. 319 rabies virus. 313-18 vesicular stomatis virus, 310-13
Rhinovirus. 228-29 Rickettsia, properties of. 4-5 Rift valley fever virus (RVFV),
348-,51 Rinderpest virus, 296-303 Robbins,49 Rotaviruses. 254-57 Rowe, 164 Rubarth, 169
Sapre, SN., 257 Sarcoma viruses, 326-31 Sorapie, 362-63 Seet.haraman, C .. 217 Sendaivirus,289 Shaila, M.S., 305 Shedding ofvirus, 73-74 Sheep pox virus, 13840 Simian herpes virus-I, 193-94 Singh, 95, 183 Skinner, 215 Slow infection, 78-79 Spwnavirus, 333-34 Stanley, 3 Stoke's law,S Subacute spongiformviral
encephalopathies, 78-79 Suspension cultures, 50-51 Swine enteroviruses, 223-25 Swine fever, 24245 Swine influenza virus, 280-81 Swine pox virus, 143-44 Swine vesicular disease virus,
224-25
Talf" dis~e, 223-24 Teschan virus 223-24
Thiosemicarbazones, 108 Togaviridae viruses, 17,27-28,
234-46 alphaviruses, 235-38 arterivirus, 245-46
371
control, 23~, 240-42, 244-46 diagnosis, 237, 240, 242. 24446 epidemiology, 237, 239, 241,
243,245 family of, 236 flaviviridae and, 234 bmununereaction,240 pathogenesis, 237, 23940 pestivirus,23845 properties of, 236-38, 241, 243.
245 Toroviruses,19 Toumier,4 Transformation assay, 53 Transmissble gastroenteritis virus
(l'GEV), 272-73 Trautwein, K., 214 Tubular symmetry, 7-8 Tumor viruses,
effect of interferon on transformation by, 113-14
families, 94-96 oncogenes, 96-98 retroviruses, 96-101 transformation of genes, 96-98
Twort, 3
Unicellular organism, properties of, 4-5
Vaccinia virus, 131-33 Varet monkey disease, 320 Vasudeven, D.W., 257 Vesicular disease diagnosis, 219 Vesicular exanthema virus
(VEV),231-32 Vesicular stomatis virus (VS V),
310-13

372
Viral disease, diagnosis of, 115-25
material for, 115-16 molecular biological
techniques, 122-25 serological methods, 118-22 viral nucleic acid detection, 122 virus isolation 117-18 visualisation by election
microscope, 116-17 epidemiology of, 87-92, 124
dissemination of virus, 91-92 environmental factors, 89 host agent, 89 incubation period of vi~s,
91-92 perpetuation of viruses, 89-90 tools, 87-88 transmission of viruses, 90-92 viral agent 88
Viral envelope, 8 Viral genetics, 55-63
genes in viruses, 58 genome, 57-59 marker rescue, 60, 62 mutation, 56-57 reactivation, 60 restricted endonuclease cleavage
maps, 60, 62-63 viruses recombination, 59-60, 63
Viral genome replication, 40-43 Viral immunity, 81-85
antibody effect, 82-83 cell mediated immunity, 83 cytotoxic T cells 83-85 humoral response, 82 infection, 81-82
Viral infectivity assay, 52-53 Viral protein, cytopathic effect of, 66 Viral replication, 34-44 Viral synthesis, 37-38 Viral vaccines,
antiviral drugs, 107-08
TexJbook o/Veterinary Virology
attenuated, 102-05 categories, 102-06 inactivated, 102, 104-06 inhibitors, 108-09 interferences, 109-14
biological effects,1l1-13' clinical use, 114 prod~tion of, 111 tumor virus and,113-14 types of, 110
isolated irnmunogenes, 106-07 Virion assembly, 43 Viroiogy,
divisions of, 3 see Viruses
Virus neutralization test (VN), 118-19
Virus protein synthesis, 40 Viruses,
chemical structure, 8-12 classification, 13-33 cultivation, 44-54 DNA viruses, 14-16,20-26,
129-208 detection and identification,
124-25 diseases, 82-92, 115-25 dissemination, 91-92 entry to animal body, 88-72 genes, 58 genetics, 11-12,55-63 immunity,81.85 incubation period, 91-92 morphology, 5-8 pathogenesis, 65-75 perpetuation of, 89-90 physical structure, 5-8 properties, 4-5 RNA viruses, 16-19,26-33,
211-361 replication, 34-44 see also specific vi.,.ses

Index
shedding, 73-74 sized,4-5 sprelld of, 68-72 transmission of, 90-92 tumorogenesis !Lid, 94-101 vaccines, 102-14
Visna virus, 78, 337-40
Waldman, D., 214 Warfield, 151 WeBer, 49 Wesselsbom virus, 249 Woodruff,47
Zinke, 313
373


















